Область техники
Настоящее изобретение относится к микроорганизму, продуцирующему L-тирозин, включающему регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с ней, а также способу получения L-тирозина с использованием микроорганизма.
Предшествующий уровень техники
L-тирозин представляет собой одну из аминокислот и используется в качестве важного материала для фармацевтического сырья, пищевых добавок, кормов для животных, биологически активных добавок и т.д. Для получения L-тирозина и других полезных материалов проводятся различные исследования по разработке микроорганизмов с использованием высокоэффективного производства и технологий процессов ферментации.
Процесс продуцирования L-тирозина микроорганизмами начинается с 3-дезокси-D-аробино-гептулозонат-7-фосфата (DAHP), полученного в результате реакции полимеризации фосфоенолпирувата (PEP), который является промежуточным звеном гликолиза, с эритрозо-4-фосфатом (Е4Р), который является промежуточным звеном пентозофосфатного пути. Затем DAHP биосинтезируется из хоризмата в префенат по общему ароматическому биосинтетическому пути и, наконец, преобразуется в L-тирозин по биосинтетическому пути L-тирозина. Во время этого процесса хоризмат может быть превращен в L-триптофан, а префенат может быть превращен в L-тирозин или L-фенилаланин. Таким образом, когда общий путь биосинтеза ароматических аминокислот усилен так, чтобы увеличить количество продуцируемого L-тирозина, можно ожидать, что продуцирование L-триптофана и L-фенилаланина при этом также увеличится. То есть при продуцировании L-тирозина, одновременно продуцируются фенилаланин и триптофан в качестве побочных продуктов, и, соответственно, необходимы различные исследования, такие как рекомбинация генов, очистка и т.д. Между тем, известно, что продуцирование L-триптофана регулируется репрессорами и аттенюаторами в соответствии с концентрацией L-триптофана, вырабатываемого микроорганизмами (Патент Кореи №10-0792095 B1).
При этих условиях, авторы настоящего изобретения приложили значительные усилия для разработки микроорганизма, способного продуцировать с высокой эффективностью L-тирозин, при этом они подтвердили, что когда гены, продуцирующие L-фенилаланин, регулируются промоторами оперона, продуцирующего L-триптофан, выход продукции L-тирозина увеличивается до более высокого уровня, тем самым осуществив настоящее изобретение.
Техническая задача
Одной из целей настоящего изобретения является создание микроорганизма, продуцирующего L-тирозин, включающего регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью.
Другой целью настоящего изобретения является разработка способа получения L-тирозина, включающего культивирование микроорганизма в среде; и извлечение L-тирозина из культивируемого микроорганизма или среды.
Еще одной целью настоящего изобретения является создание экспрессионной кассеты, включающей регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью.
Еще одной целью настоящего изобретения является разработка способа для регулирования активности префенатдегидрогеназы с использованием регуляторной области оперона trp и гена, кодирующего префенатдегидрогеназы, функционально связанного с этой обалстью.
Еще одна цель настоящего изобретения заключается в использовании L-тирозин-продуцирующего микроорганизма для получения L-тирозина.
Еще одна цель настоящего изобретения заключается в использовании экспрессионной кассеты для получения L-тирозина.
Еще одна цель настоящего изобретения заключается в использовании композиции для получения L-тирозина.
Полезные эффекты изобретения
L-тирозин-продуцирующий микроорганизм по настоящему изобретению, который включает ген, кодирующий префенатдегидрогеназу и регуляторную область оперона trp, минимизирует накопление L-фенилаланина, не оказывая влияния на рост клеток и может с высокой эффективностью продуцировать L-тирозин Лучший вариант осуществления изобретения Настоящее изобретение подробно описано ниже. Между тем, соответствующие описания и воплощения, раскрытые в настоящем описании, могут также применяться к другим описаниям и воплощениям. То есть все комбинации различных элементов, представленные в настоящем раскрытии, относятся к области настоящего изобретения. Кроме того, объем настоящего раскрытия не может рассматриваться как ограниченный конкретным описанием, представленным ниже.
Для достижения вышеуказанных целей один из аспектов настоящего изобретения предлагает микроорганизм, продуцирующий L-тирозин, включающий регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью.
Используемый здесь термин "L-тирозин" представляет собой одну из 20 α-аминокислот и классифицируется как гидрофильная аминокислота или ароматическая аминокислота. Тирозин является коммерчески важной аминокислотой, используемой в качестве предшественника фармацевтических препаратов, флавоноидов, алкалоидов и т.д.
Используемый здесь термин "оперон триптофана (оперон Trp)" относится к группе генов, кодирующих ферменты, участвующие в синтезе триптофана из хоризмовой кислоты (хоризмат), и включает структурные гены и регуляторные области экспрессии (или регуляторные области). В частности, оперон триптофана может быть опероном триптофана, полученным из микроорганизма рода Corynebacterium или опероном триптофана, полученным из микроорганизма рода Escherichia, но может включать оперон триптофана различного происхождения без ограничений при условии, что он может регулировать ген pheA в соответствии с настоящим изобретением. В частности, микроорганизм рода Corynebacterium может представлять собой Corynebacterium glutamicum, а микроорганизм рода Escherichia может представлять собой Е. coli, но микроорганизмы этим не ограничиваются. Кроме того, оперон триптофана может иметь известную нуклеотидную последовательность, и специалисты в данной области могут легко получить нуклеотидную последовательность оперона триптофана из базы данных, такой как NCBI или Kegg, и т.д. Обычный оперон триптофана активно транскрибируется, чтобы продуцировать достаточное количество триптофана, необходимое клеткам, но когда в клетках присутствует достаточное количество триптофана, репрессор связывается с триптофаном и инактивирует оперон триптофана, тем самым ингибируя транскрипцию. В настоящем раскрытии изобретения вышеуказанный термин может использоваться взаимозаменяемо с термином триптофановый оперон, trp оперон, и т.д.
Используемый здесь термин "регуляторная область оперона trp" относится к области, которая находится против хода транскрипции относительно структурных генов, составляющих оперон trp, и может регулировать экспрессию структурных генов. Структурные гены, образующие оперон trp в микроорганизме рода Corynebacterium, могут быть составлены из генов trpE, trpG, trpD, trpC, trpB и trpA, а структурные гены, образующие оперон trp в микроорганизме рода Escherichia могут быть составлены из генов trpE, trpD, trpC, trpB, и trpA. Регуляторная область оперона trp может находиться выше trpE в положении 5' структурных генов оперона trp.В частности, она может включать регулятор trp (trpR), промотор (промотор trp), оператор (trp оператор), лидерный пептид оперона trp (trp L), и аттенюатор trp, исключая структурные гены, которые могут составлять оперон trp.Более конкретно, она может включать промотор (промотор trp), оператор (оператор trp), лидерный пептид trp (trp L) и аттенюатор trp (аттенюатор trp). В настоящем описании регуляторная область оперона trp может включать, без ограничения, любую регуляторную область, расположенную против хода транскрипции относительно гена pheA, кодирующего префенатдегидрогеназу, и может регулировать экспрессию гена pheA. В настоящем раскрытии регуляторная область оперона trp может использоваться взаимозаменяемо с регуляторным фактором оперона trp, промотором trpE, промотором оперона trp, регуляторной областью trpE, регуляторным фактором trpE и регуляторной последовательностью trpE.
Например, регуляторная область оперона trp может включать нуклеотидную последовательность SEQ ID NO: 1, но этим не ограничивается. Последовательность SEQ ID NO: 1 может быть подтверждена известной базой данных GenBank of NCBI.
В частности, регуляторная область оперона trp может состоять из нуклеотидной последовательности SEQ ID NO: 1 или нуклеотидной последовательности, имеющей гомологию или идентичность, составляющую по меньшей мере 70%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, или 99% или более с SEQ ID NO: 1. Кроме того, очевидно, что регуляторная область с делецией, модификацией, заменой или добавлением части последовательности также входит в объем настоящего изобретения при условии, что его нуклеотидная последовательность имеет такую гомологию или идентичность и выполняет функцию, соответствующую регуляторной области.
Используемые здесь термины "гомология" и "идентичность " относятся к степени соответствия между двумя заданными аминокислотными последовательностями или нуклеотидными последовательностями и могут быть выражены в процентах. Термины "гомология" и "идентичность" часто используются взаимозаменяемо друг с другом.
Гомология последовательностей или идентичность консервативных полинуклеотидов или полипептидов может быть определена стандартным алгоритмом выравнивания и может использоваться вместе с установленным по умолчанию штрафом за пропуск в последовательности, обнаруженным используемой программой. Как правило, ожидается, что гомологичные или идентичные последовательности гибридизуются со всеми или по меньшей мере с примерно 50%, примерно 60%, примерно 70%, примерно 80%, примерно 85% или примерно 90% всей длины последовательностей при умеренных или жестких условиях. Полинуклеотиды, которые содержат вырожденные кодоны вместо кодонов, также рассматриваются при гибридизации полинуклеотидов.
Гомология или идентичность полипептидных или полинуклеотидных последовательностей может быть определена, например, с помощью алгоритма BLAST по литературе [см: Karlin and Altschul, Pro. Natl. Acad. Sci. USA, 90, 5873(1993)], или FASTA по Pearson (см: Methods Enzymol., 183, 63, 1990). На основе алгоритма BLAST была разработана программа, называемая BLASTN или BLASTX (см.: www.ncbi.nlm.nih.gov). Кроме того, независимо от того, имеют ли какие-либо аминокислотные или полинуклеотидные последовательности гомологию, сходство или идентичность друг с другом, это может быть идентифицировано путем сравнения последовательностей в эксперименте саузерн-блот гибридизация при жестких условиях, как определено, и соответствующие условия гибридизации, определенные в рамках данной области техники, могут быть установлены методом, хорошо известным специалистам в данной области, (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology)
Используемый здесь термин "префенатдегидрогеназа" является одним из белков, необходимых для биосинтеза L-фенилаланина. Ген, кодирующий белок, может представлять собой, например, ген pheA, но не ограничивается этим. Белок также называют как бифункциональная хоризматмутаза/префенатдегидрогеназа. Поскольку префенатдегидрогеназа представляет собой фермент в пути, вовлеченном в получение L-фенилаланина из хоризмата или префената, и является ферментом на стадии конкуренции с биосинтетическим путем тирозина, его обычно выбирают в качестве целевого гена для инактивации продукции штаммов, продуцирующих тирозин. Однако когда префенатдегидрогеназа инактивирована, возникает проблема, заключающаяся в значительном снижении продуктивности, так как он влияет на рост штаммов.
В настоящем изобретении микроорганизмы, которые были генетически модифицированы таким образом, чтобы ген pheA регулировался регуляторной областью оперона trp, в частности, микроорганизмы, в которых регуляторную область оперона trp подставляли к области промотора для регуляции pheA, минимизируют накопление фенилаланина, не влияя при этом на рост клеток и улучшая продуцированием тирозина.
Кроме того, ген pheA, который представляет собой ген, кодирующий префенатдегидрогеназу, может иметь делецию в части или всей регуляторной последовательности, находящейся против хода транскрипции по отношению к структурному гену pheA. Для осуществления настоящего изобретения, выше расположенная область структурного гена pheA может быть заменена регуляторной областью оперона trp, или регуляторная область оперона trp может быть вставлена выше области структурного гена pheA, чтобы включить структурный ген pheA, который может регулироваться экспрессией белка, кодируемого геном pheA с помощью регуляторной области оперона trp. Используемый здесь термин "ген pheA" может использоваться взаимозаменяемо с "геном, кодирующим префенатдегидрогеназу" и "структурным геном pheA ".
Используемый здесь термин "микроорганизм, продуцирующий L-тирозин" относится к микроорганизму, естественным образом обладающему способностью продуцировать L-тирозин или микроорганизму, который приобрел L-тирозин-продуцирующую способность от своего родительского штамма, не имеющего L-тирозин-продуцирующей способности. В частности, микроорганизм может представлять собой микроорганизм, продуцирующий L-тирозин, включающий регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью, но не ограничивающийся этим.
Более конкретно, "микроорганизм, продуцирующий L-тирозин" включает все микроорганизмы дикого типа и естественно или искусственно генетически модифицированные микроорганизмы, и может представлять собой микроорганизм, обладающий генетической модификацией или повышенной активностью для необходимого продуцирования L-тирозина, может представлять собой микроорганизм, в котором конкретный механизм ослаблен или усилен механизмом из-за введения чужеродного гена, или усиления или инактивации активности эндогенного гена. Для осуществления настоящего изобретения, микроорганизм, продуцирующий L-тирозин, включает регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью, и обладает повышенной способностью продуцировать L-тирозин, а также может быть генетически модифицированным микроорганизмом или рекомбинантным микроорганизмом, но не ограничивается этим.
Более конкретно, микроорганизм, имеющий L-тирозин-продуцирующую способность по настоящему изобретению, относится к рекомбинантному микроорганизму, имеющему повышенную L-тирозин-продуцирующую способность посредством включения регуляторной области оперона trp и гена, кодирующего префенатдегидрогеназу, функционально связанного с этой областью, по настоящему изобретению, или в результате трансформации вектором, включающим регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с ней.
В настоящем раскрытии "микроорганизм, включающий ген" может означать "генетически модифицированный микроорганизм", "микроорганизм,
модифицированный для экспрессии гена", "рекомбинантный микроорганизм, экспрессирующий ген" или "рекомбинантный микроорганизм, обладающий активностью белка, кодируемого геном", но не ограничивается этим.
Например, это может быть 1) микроорганизм, включающий регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью, по настоящему изобретению, 2) микроорганизм, модифицированный для экспрессии регуляторной области оперона trp и гена, кодирующего префенатдегидрогеназу, функционально связанного с этой областью, 3) рекомбинантный микроорганизм, экспрессирующий регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью, и 4) рекомбинантьный микроорганизм, который трансформирован вектором, включающим регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью, для повышения L-тирозин-продуцирующей способности, но не ограничивается этим.
"Микроорганизм, обладающий повышенной способностью продуцировать L-тирозин", может относиться к микроорганизму, обладающему повышенной способностью продуцировать L-тирозин по сравнению с исходным штаммом или немодифицированным микроорганизмом до трансформации. "Немодифицированный микроорганизм" может быть нативным штаммом или микроорганизмом, который не был трансформирован вектором, включающим гены, кодирующие полинуклеотид и целевой белок.
В качестве микроорганизма для осуществления настоящего изобретения могут быть все микроорганизмы, способные продуцировать L-тирозин путем включения регуляторной области оперона trp и гена, кодирующего префенатдегидрогеназу, функционально связанного с ней. В настоящем изобретении термин "микроорганизм, способный продуцировать L-тирозин" может быть взаимозаменяемо использован с "микроорганизмом, продуцирующим L-тирозин" и "микроорганизмом, обладающим способностью продуцировать L-тирозин".
Микроорганизм может быть, например, микроорганизмом, принадлежащим к роду Enterbacter, микроорганизмом, принадлежащим к роду Escherichia, микроорганизмом, принадлежащим к роду Erwinia, микроорганизмом, принадлежащим к роду Serratia, микроорганизмом, принадлежащим к роду Providencia, микроорганизмом, принадлежащим к роду Corynebacterium, и микроорганизмом, принадлежащим к роду Brevibacterium, но он не ограничивается этим.
В частности, микроорганизм может быть микроорганизмом, принадлежащим к роду Corynebacterium. В настоящем раскрытии "микроорганизм, принадлежащий к роду Corynebacterium, может быть конкретно микроорганизмом Corynebacterium glutamicum, Corynebacterium ammoniagenes, Brevibacterium lactofermentum, Brevibacterium flavum, Corynebacterium thermoaminogenes, Corynebacterium efficiens, Corynebacterium stationis, и т.д., но он не ограничивается этим.
Другой аспект настоящего изобретения предлагает способ получения L-тирозина, включающий культивирование микроорганизма в среде; и извлечение L-тирозина из культивируемого микроорганизма или среды.
В вышеописанном способе стадия культивирования микроорганизма может быть осуществлена известным периодическим культивированием, непрерывным культивированием, культивированием с подпиткой и т.д., но этим не ограничивается. В частности, в зависимости от условий культивирования, рН культуры может быть скорректирован до подходящего значения (например, рН от 5 до 9, в частности рН от 6 до 8, и более конкретно рН 7,0) с использованием основного соединения (например, гидроксида натрия, гидроксида калия или аммиака) или кислого соединения (например, фосфорной кислоты или серной кислоты), но не ограничивается этим. Кроме того, кислород или кислородсодержащую газовую смесь можно вводить в культуру для поддержания аэробного состояния. Температура культивирования может поддерживаться от 20°С до 45°С, в частности от 25°С до 40°С, и культивирование может проводиться в течение от 10 до 160 часов, но культура не ограничивается вышеуказанным. Аминокислота, продуцируемая культурой, может секретироваться в среду или оставаться в клетках.
Кроме того, в качестве источника углерода для используемой питательной среды можно использовать сахара и углеводы (например, глюкоза, сахароза, лактоза, фруктоза, мальтоза, патока, крахмал и целлюлоза), масла и жиры (например, соевое масло, масло семян подсолнечника, арахисовое масло и кокосовое масло), жирные кислоты (например, пальмитиновая кислота, стеариновая кислота и линолевая кислота), спирты (например, глицерин и этанол) и органические кислоты (например, уксусная кислота) по отдельности или в комбинации, но источники углерода не ограничиваются этим. В качестве источника азота можно использовать азотсодержащие органические соединения (например, пептон, дрожжевой экстракт, мясная подливка, солодовый экстракт, кукурузный экстракт, соевая мука и мочевина) или неорганические соединения (например, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония) по отдельности или в комбинации, но источник азота этим не ограничивается. В качестве источника фосфора можно использовать дигидрофосфат калия, гидроортофосфат калия и их соответствующие натрийсодержащие соли и т.д., по отдельности или в комбинации, но источник фосфора этим не ограничивается. Кроме того, среда может содержать необходимые вещества, способствующие росту, такие как соли других металлов (например, сульфат магния или сульфат железа), аминокислоты и витамины.
В способе извлечения аминокислоты, полученной на этапе культивирования настоящего изобретения, целевые аминокислоты могут быть собраны из культивируемого раствора с использованием соответствующего способа, известного в данной области техники, согласно способу культивирования. Например, могут быть использованы центрифугирование, фильтрация, анионообменная хроматография, кристаллизация, ВЭЖХ и т.д., и целевые аминокислоты могут быть извлечены из среды или микроорганизма с использованием соответствующего метода, известного в данной области техники.
Кроме того, стадия извлечения может дополнительно включать процесс очистки и может быть выполнена с использованием соответствующего метода, известного в данной области техники. Таким образом, извлеченные аминокислоты могут находиться в очищенном состоянии или в виде раствора микробиологической ферментации, содержащего аминокислоты (Introduction to Biotechnology and Genetic Engineering, A. J. Nair., 2008). Кроме того, до и после стадии культивирования, а также до и после стадии выделения, целевые аминокислоты могут быть эффективно извлечены путем дополнительного выполнения соответствующего способа, известного в данной области техники.
Еще один аспект настоящего изобретения предлагает экспрессионную кассету, включающую регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью.
В настоящем изобретении термин "экспрессионная кассета" может представлять собой единицу генетической конструкции, включающую существенный регуляторный элемент, функционально связанный с геном, таким образом, что введенный ген экспрессируется, когда он присутствует в клетках индивидуума. Экспрессионная кассета может быть, например, в форме экспрессионного вектора, но не ограничивается этим, и может включать все генетические конструкции, имеющие наименьшую единицу, способную экспрессировать целевой ген, подлежащий введению.
Экспрессионная кассета может быть получена и очищена стандартной технологией рекомбинантной ДНК. Любой тип экспрессионной кассеты может использоваться без особых ограничений до тех пор, пока он может функционировать для экспрессии необходимого гена в различных клетках-хозяевах, таких как прокариотические клетки и эукариотические клетки, и до тех пор, пока он может функционировать для получения необходимого белка. Экспрессионная кассета может включать промотор, стартовый кодон, ген, кодирующий целевой белок и терминирующий кодон. Кроме того, он может, соответственно, включать ДНК, кодирующую сигнальный пептид, последовательность энхансера, нетранслируемые области на 5' и 3' концах целевого гена, область селектируемого маркера или реплицируемый участок и т.д. Кроме того, экспрессионная кассета может включать моноцистронный вектор, включающий полинуклеотид, кодирующий один белок или полицистронный вектор, включающий полинуклеотид, кодирующий два или более рекомбинантных белка, но не ограничивается этим.
Используемый здесь термин "вектор" может относиться к конструкции ДНК, содержащей нуклеотидную последовательность полинуклеотида, кодирующего целевой белок, который функционально связан с подходящей регуляторной последовательностью, так что целевой белок может быть экспрессирован в соответствующем хозяине. Регуляторная последовательность может включать промотор, способный инициировать транскрипцию, последовательность любого оператора для регулирования транскрипции, последовательность, кодирующую соответствующий домен связывания мРНК с рибосомой, и последовательность, регулирующую прекращение транскрипции и трансляции. После трансформации в подходящую клетку хозяина вектор может реплицироваться и функционировать независимо от генома хозяина и может быть интегрирован в сам геном хозяина.
Вектор, используемый в настоящем изобретении, не ограничен особым образом, при условии, что он может экспрессироваться в клетке хозяина, и таким образом, может быть использован любой вектор, известный в данной области. Примеры традиционно используемых векторов могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора могут использоваться pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и т.д.; и в качестве плазмидного вектора могут быть использованы векторы на основе pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pET и т.д.. В частности, могут использоваться векторы pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и т.д., но не ограничиваются этим.
Векторы, которые могут быть использованы в настоящем изобретении, не ограничены особым образом, и можно использовать известный экспрессионный вектор. Кроме того, ген или полинуклеотид, кодирующий целевой белок, может быть вставлен в хромосому с помощью вектора для внутриклеточной хромосомной вставки. Вставка полинуклеотида в хромосому может быть выполнена любым способом, известным в данной области техники, например, путем гомологичной рекомбинации, но не ограничивается этим. Кроме того, вектор может дополнительно включать селективный маркер для подтверждения вставки в хромосому. Селективный маркер используют для отбора клеток, которые были трансформированы вектором, т.е. для определения того, был ли введен целевой полинуклеотид, при этом могут быть использованы маркеры, которые показывают лекарственную устойчивость, ауксотрофию, устойчивость к цитотоксическим агентам или представляют отобранные фенотипы, такие как экспрессия поверхностных белков. В условиях, когда селективные агенты обрабатывают, только клетки, способные экспрессировать селективные маркеры, могут выжить или экспрессировать другие фенотипические признаки, и таким образом могут быть отобраны трансформированные клетки.
Используемый здесь термин "трансформация" относится к введению вектора, включающего полинуклеотид, кодирующий целевой полипептид, в клетку-хозяина таким образом, что белок, кодируемый полинуклеотидом, экспрессируется в клетке-хозяине. Для того, чтобы трансформированный полинуклеотид был экспрессирован в клетке-хозяине, он может быть либо вставлен в хромосому клетки-хозяина и расположен в ней, либо расположен экстрахромосомно, и оба случая могут быть включены. Кроме того, полинуклеотид включает ДНК и РНК, которые кодируют целевой белок. Полинуклеотид может быть введен в любой форме при условии, что он может быть введен в клетку-хозяина и экспрессирован в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генетическую конструкцию, включающую все элементы, необходимые для собственной экспрессии. Метод трансформации включает любой способ введения нуклеиновой кислоты в клетку и может быть осуществлен путем выбора подходящего стандартного метода, известного в данной области техники, в зависимости от клетки-хозяина. Например, трансформация может быть выполнена использованием электропорации, преципитации фосфатом кальция (CaPO4), преципитации хлоридом кальция (CaCl2), микроинъекции, метода полиэтиленгликоля (ПЭГ), метода ДЕАЕ-декстрана, метода катионных липосом, метода с ацетатом лития-ДМСО, и т.д., но этим метод не ограничивается.
Кроме того, используемый здесь термин "функциональная связь" относится к функциональной связи между вышеуказанной полинуклеотидной последовательностью с промоторной последовательностью, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего целевой белок по настоящему изобретению, или с регуляторной областью. Действующая связь может быть получена методом рекомбинации генов, известным в данной области техники, а сайт-специфическое расщепление ДНК и лигирование может быть выполнено с использованием известных лиазы, лигазы и т.д., но этим не ограничивается.
Еще один аспект настоящего изобретения предлагает композицию для получения L-тирозина, включающую регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью.
Композиция для получения L-тирозина включает регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью, и может дополнительно иметь, без ограничения, конфигурацию, способную управлять геном или регуляторной областью оперона. Ген, кодирующий префенатдегидрогеназу, и регуляторная область оперона trp могут быть в форме, включенной в вектор таким образом, что ген, функционально связанный и введенный в клетку хозяина, может быть экспрессирован. Экспрессия гена pheA, который является геном, кодирующим префенатдегидрогеназу, может регулироваться регуляторной областью оперона trp.
Еще один аспект настоящего изобретения предлагает способ регулирования активности префенатдегидрогеназы с использованием регуляторной области оперона trp и гена, кодирующего префенатдегидрогеназу, функционально связанного с этой областью.
Еще один аспект настоящего изобретения предлагает использование микроорганизма, продуцирующего L-тирозин, для получения L-тирозина.
Еще один аспект настоящего изобретения предлагает применение экспрессионной кассеты для получения L-тирозина.
Еще один аспект настоящего изобретения предлагает применение композиции для получения L-тирозина.
Способ осуществления изобретения
Далее настоящее изобретение будет описано более подробно со ссылкой на следующие примеры. Однако эти Примеры приведены только в иллюстративных целях, и область применения изобретения не ограничивается этими Примерами.
Пример 1: Конструирование штамма, продуцирующего L-тирозин
Хотя Corynebacterium glutamicum дикого типа способен продуцировать L-тирозин, он не продуцирует L-тирозин в достаточном количестве для высвобождения в культуральную среду. В соответствии с целью настоящего изобретения, для идентификации генетического признака, повышающего способность к продуцированию L-тирозина, был использован штамм с повышенной способностью к продуцированию L-тирозина, а не штамм дикого типа. Таким образом, штамм, продуцирующий L-тирозин, был сконструирован путем усиления генов, необходимых для получения L-тирозина, на основе штамма Corynebacterium glutamicum АТСС 13869.
Во-первых, для увеличения поступления эритрозо-4-фосфата (Е4Р) в качестве предшественника L-тирозина, гены tkt были сверхэкспрессированы. В то же время aroP, ген-импортер ароматических аминокислот, который вводит L-тирозин в клетки, был удален.
Для генетической манипуляции сначала были получены нижележащие и вышележащие области гена aroP, в которые ген tkt следовало вставить путем замены. В частности, фрагмент гена в нижележащей области гена aroP был получен с использованием праймеров SEQ ID NO: 2 и SEQ ID NO: 3, а фрагмент гена в вышележащей области гена aroP был получен с использованием праймеров SEQ ID NO: 4 и SEQ ID NO: 5 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы, посредством ПЦР. Полимераза Solg™ Pfu-X DNA была использована в качестве полимеразы, и ПЦР проводили в условиях ПЦР-амплификации: с денатурацией при 95°С в течение 5 минут, с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжига при 60°С в течение 30 секунд и полимеризации при 72°С в течение 60 секунд, и последующей полимеризацией при 72°С в течение 5 минут.
Кроме того, для получения гена tkt, включающего промотор tkt, фрагмент гена tkt, включающий промотор tkt, был получен с использованием праймеров SEQ ID NO: 6 и SEQ ID NO: 7 на основе хромосомной ДНК Corynebacterium glutamicum АТСС 13869 в качестве матрицы посредством ПЦР. Полимеразу Solg™ Pfu-X DNA использовали в качестве полимеразы, и ПЦР проводили в условиях ПЦР-амплификации: с денатурацией при 95°С в течение 5 минут, с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжига при 60°С в течение 30 секунд и полимеризации при 72°С в течение 150 секунд, и последующей полимеризацией при 72°С в течение 5 минут.
Амплифицированные вышележащие и нижележащие области промотора aroP, фрагмент гена tkt, включающий промотор tkt и вектор pDZ (Патент Кореи №10-0924065) для хромосомной трансформации расщепляли рестриктазой SmaI и клонировали с использованием метода сборки Гибсона (DG Gibson et at, NATURE METHODS, VOL.6 NO.5, MAY 2009, NEBuilder HiFi DNA Assembly Master Mix) с целью получения рекомбинантной плазмиды, которая называется pDZ-ΔaroP::Pn-tkt. Клонирование проводили путем смешивания реагента для сборки Гибсона и каждого из фрагментов гена в рассчитанном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
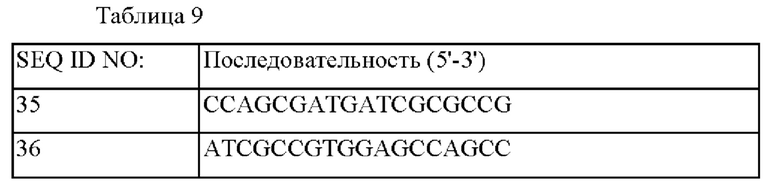
Последовательности праймеров, используемых для построения каждого из векторов, показаны в таблице 1 ниже.

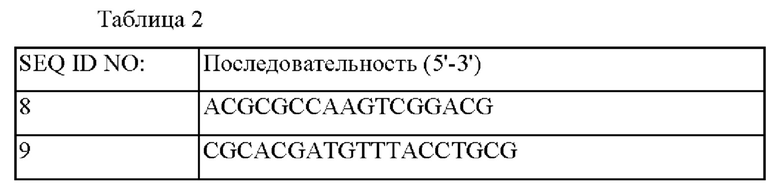
Сконструированный вектор pDZ-ΔaroP::Pn-tkt трансформировали в штамм Corynebacterium glutamicum АТСС 13869 методом электропорации и затем подвергали вторичному кроссинговеру для получения штамма, в который был вставлен ген tkt, включающий промотор tkt, при одновременном удалении гена aroP. Соответствующие генетические манипуляции подтверждали с помощью секвенирования генома и метода ПЦР, с использованием праймеров SEQ ID NO: 8 и SEQ ID NO: 9, который может соответственно амплифицировать внешнюю область вышележащей области и нижележащей области гомологичной рекомбинации, куда был вставлен соответствующий ген, а полученный штамм был назван СМ06-0001.
Для усиления пути L-тирозина, ген tyrA, регулируемый обратной связью посредством L-тирозина, которой обладает Corynebacterium glutamicum, заменяли вариантом tyrA, не регулируемым обратной связью, полученным из Е. coli, включая сильный промотор gapA. Известно, что в белке tyrA, полученном из Е. coli, обратная связь ослабевает, когда метионин в положении 53 мутирован в изолейцин, а аланин в положении 354 мутирован в валин, и используется эта форма белка (SEQ ID NO: 10) (Appl. Microbiol. Biotechnol. 75, 103-110 (2007)).
Для генетической манипуляции сначала были получены вышележащие и нижележащие области гена tyrA, в которые ген tyrA следовало вставить путем замены. В частности, фрагмент гена в вышележащей области гена tyrA был получен с использованием праймеров SEQ ID NO: 11 и SEQ ID NO: 12, а фрагмент гена в нижележащей области гена tyrA был получен с использованием праймеров SEQ ID NO: 13 и SEQ ID NO: 14 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы, посредством ПЦР. Полимеразу Solg™ Pfu-X DNA использовали в качестве полимеразы, и ПЦР выполняли при следующих условиях амплификации: денатурация при 95°С в течение 5 минут, с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжига при 60°С в течение 30 секунд и полимеризации при 72°С в течение 60 секунд, с последующей полимеризацией при 72°С в течение 5 минут.
Кроме того, для того, чтобы получить вариант гена tyrA, происходящий из Е. coli, включающий промотор gapA, получали фрагмент промотора gapA с использованием праймеров SEQ ID NO: 15 и SEQ ID NO: 16 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869, используемой в качестве матрицы, посредством ПЦР, а фрагмент гена tyrA, происходящего из Е. coli, получали с использованием праймеров SEQ-ID NO: 17 и SEQ ID NO: 18 на основе синтетической ДНК варианта tyrA, в качестве матрицы, посредством ПЦР
Полимеразу SolgTM Pfu-X DNA использовали в качестве полимеразы, и ПЦР выполняли при следующих условиях амплификации: денатурация при 95°С в течение 5 минут, с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжига при 60°С в течение 30 секунд и полимеризации при 72°С в течение 60 секунд с последующей полимеризацией при 72°С в течение 5 минут
Амплифицированные вышележащие и нижележащие области гена tyrA, фрагмент варианта Е. coli-полученного гена tyrA, включающего промотор gapA, и вектор pDZ для хромосомной трансформации расщепляли рестриктазой SmaI, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, которая была названа pDZ-ΔtyrA::PgapA-tyrAm. Клонирование проводили путем смешивания реагента для сборки Гибсона и каждого из фрагментов гена в рассчитанном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Последовательности праймеров, используемых для построения каждого из векторов, показаны в Таблице 3 ниже.

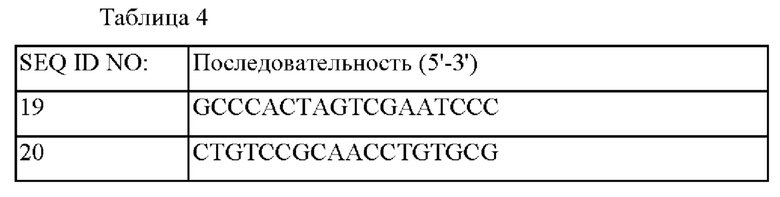
Сконструированный вектор pDZ-ΔtyrA::PgapA-tyrAm был трансформирован в штамм СМ06-0001 методом электропорации, а затем подвергнут вторичному кроссинговеру для получения штамма, в который был вставлен вариант Е. со//-полученного гена tyrA, включающего промотор gapA, при одновременном удалении гена tyrA. Соответствующие генетические манипуляции были подтверждены секвенированием генома и методом ПЦР с использованием праймеров SEQ ID NO: 19 и SEQ ID NO: 20, который может, соответственно, амплифицировать внешнюю область вышележащей области и нижележащей области гомологичной рекомбинации, куда был вставлен соответствующий ген, полученный штамм называется СМ06-0002.
Для увеличения выработки L-тирозина, ген aroG, вовлеченный в первую стадию общего пути биосинтеза ароматических аминокислот, был усилен добавлением сильного промотора к полученному из Е. coli варианту aroG, отвечающему за ослабление регулирования по принципу обратной связи. Известно, что в Е. coli-полученном белке aroG, обратная связь ослабевает (feedback is released) когда пролин в положении 150 заменяется лейцином, и когда используется эта форма белка (SEQ ID NO: 68) (Appl. Environ. Microbiol. 63, 761-762 (1997)).
Для генетических манипуляций были получены нижележащие и вышележащие области, в которые должен был быть дополнительно вставлен ген aroG. В частности, фрагмент гена в вышележащей области гена BBD29 14470 был получен с использованием праймеров SEQ ID NO: 21 и SEQ ID NO: 22, а фрагмент гена в нижележащей области гена BBD29 14470 был получен с использованием праймеров SEQ ID NO: 23 и SEQ ID NO: 24 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869, используемой в качестве матрицы, посредством ПЦР. Полимеразу SolgTM Pfu-X DNA использовали в качестве полимеразы, и ПЦР выполняли при следующих условиях амплификации: денатурация при 95°С в течение 5 минут, с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжига при 60°С в течение 30 секунд и полимеризации при 72°С в течение 60 секунд с последующей полимеризацией при 72°С в течение 5 минут
Амплифицированные вышележащие и нижележащие области, в которые дополнительно вставляют вариант aroG и вектор pDZ для хромосомной трансформации, расщепляли с помощью рестриктазы SmaI, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, которая была названа pDZ-Δ BBD29_14470. Клонирование проводили путем смешивания реагента для сборки Гибсона и каждого из фрагментов гена в рассчитанном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Кроме того, для получения происходящего из E.coli варианта гена aroG, включающего промотор gapA, получали фрагмент промотора gapA с использованием праймеров SEQ ID NO: 15 и SEQ ID NO: 26 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869, используемой в качестве матрицы, посредством ПЦР, а вариант фрагмента происходящего из Е. coli гена aroG получали с использованием праймеров SEQ ID NO: 27 и SEQ ID NO: 28 на основе синтетической ДНК происходящего из Е. coli варианта aroG, ослабляющего обратную связь, в качестве матрицы, посредством ПЦР. Полимеразу SolgTM Pfu-X DNA использовали в качестве полимеразы и ПЦР выполняли при следующих условиях амплификации: денатурация при 95°С в течение 5 минут, с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжига при 60°С в течение 30 секунд и полимеризации при 72°С в течение 60 секунд с последующей полимеризацией при 72°С в течение 5 минут
Амплифицированный фрагмент варианта гена aroG, включающего промотор gapA, и вектор pDZ-ΔBBD29 14470 для хромосомной трансформации расщепляли рестриктазой SeaI и клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, которая была названа pDZ-ΔBBD29_14470::PgapA-aroGm. Клонирование проводили путем смешивания реагента для сборки Гибсона и каждого из фрагментов гена в рассчитанном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Последовательности праймеров, используемые для конструирования каждого из векторов, показаны в Таблице 5 ниже.

Сконструированным вектором pDZ-ΔBBD29 14470::PgapA-aroGm
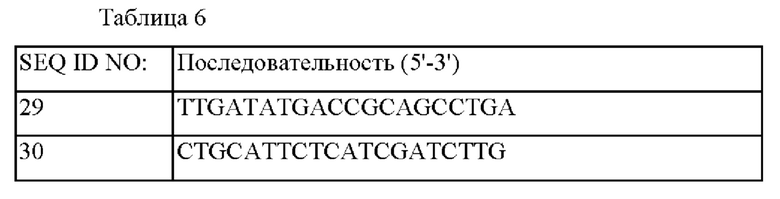
трансформировали штамм СМ06-0002 методом электропорации и затем подвергали вторичному кроссинговеру для получения штамма, в который был вставлен ослабляющий обратную связь Е. coli-полученный вариант гена aroG, включающий промотор gapA. Соответствующие генетические манипуляции были подтверждены посредством секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 29 и SEQ ID NO: 30, который может соответственно амплифицировать внешнюю область вышележащей области и нижележащей области гомологичной рекомбинации, куда был вставлен соответствующий ген, полученный штамм называется СМ06-0003.
Пример 2: Оценка продуцирующей способности штаммов, продуцирующих L-Тирозин
Для подтверждения способности штаммов, сконструированных в примере 1, к продуцированию L-тирозина, штаммы культивировали и оценивали следующим образом. Каждый из штаммов инокулировали в 250-мл колбу с угловой перегородкой, содержащую 25 мл следующей посевной среды и культивировали при встряхивании при 200 об/мин при 30°С в течение 20 часов. Затем 1 мл раствора для посева инокулировали в 250-мл колбу с угловой перегородкой, содержащую 25 мл среды для продуцирования и культивировали при встряхивании при 200 об/мин при 30°С в течение 24 часов. После культивирования количество продуцированного L-тирозина, L-фенилаланина и L-триптофана измеряли с помощью ВЭЖХ.
Посевная среда (рН 7,0)
20 г глюкозы, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 4 г KH2PO4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина HCl, 2000 мкг пантотената кальция и 2000 мкг никотинамида (на литр дистиллированной воды)
Среда для продуцирования (рН 7,0)
30 г глюкозы, 15 г (NH4)2SO4, 1,2 г MgS04.7H2O, 1 г KH2PO4, 5 г дрожжевого экстракта, 900 мкг биотина, 4500 мкг тиамина HCl, 4500 мкг пантотената кальция и 30 г СаСО3 (на литр дистиллированной воды).
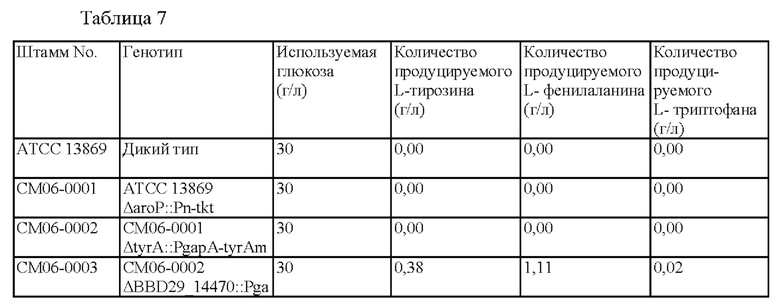

Результаты продуцирования L-тирозина, L-фенилаланина и L-триптофана в культурах дикого типа Corynebacterium glutamicum АТСС 13869, СМ06-0001, СМ06-0002 и СМ06-0003 приведены в Таблице 7.
L-тирозин не продуцировался в штамме СМ06-0001, в котором повышено поступление Е4Р, предшественника, полученного путем удаления гена импортера ароматических аминокислот из штамма дикого типа, и L-тирозин также не был обнаружен в СМ06-0002, в котором ингибирование обратной связи tyrA было дополнительно ослаблено в СМ06-0001.
Продуцирование L-тирозина и L-фенилаланина было подтверждено в штамме СМ06-0003, в котором общий путь синтеза ароматических соединений усиливался в результате ослабления ингибирования обратной связи aroG в штамме СМ06-0002. Кроме того, продуцирование L-тирозина и L-фенилаланина значительно увеличилось по сравнению с предыдущими штаммами, тогда как не было резкого изменения в продуцировании L-триптофана.
Пример 3: Делеция гена pheA у штаммов, продуцирующих L-тирозин
Было подтверждено, что при усилении общего пути синтеза ароматических соединений в Примере 2 L-фенилаланин продуцировался в наибольшем количестве, за ним следовало продуцирование L-тирозина, а L-триптофан продуцировался в небольшом количестве. Следовательно, можно ожидать, что продуцирование L-тирозина и L-триптофана увеличится, если путь синтеза L-фенилаланина будет удален.
Чтобы удалить путь синтеза L-фенилаланина, был удален ген pheA, участвующий в первой стадии ответвления от префеновой кислоты. Для такого удаления гена, сначала были получены вышележащие и нижележащие области гена pheA. В частности, фрагмент гена в вышележащей области гена pheA был получен с использованием праймеров SEQ ID NO: 31 и SEQ ID NO: 32, и фрагмент гена в нижележащей области гена pheA был получен с использованием праймеров SEQ ID NO: 33 и SEQ ID NO: 34 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869, используемой в качестве матрицы, посредством ПЦР. Полимеразу SolgTM Pfu-X DNA использовали в качестве полимеразы, и ПЦР выполняли при следующих условиях амплификации: денатурация при 95°С в течение 5 минут, с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжига при 60°С в течение 30 секунд и полимеризации при 72°С в течение 60 секунд с последующей полимеризацией при 72°С в течение 5 минут
Амплифицированные вышележащие и нижележащие области гена pheA и вектор pDZ для хромосомной трансформации расщепляли с помощью рестриктазы SmaI и клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, которая была названа pDZ-ΔpheA. Клонирование проводили путем смешивания реагента для сборки Гибсона и каждого из фрагментов гена в рассчитанном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
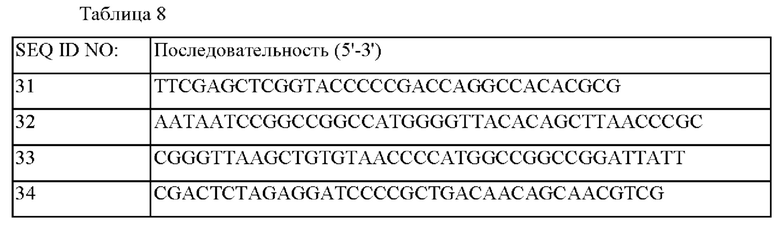
Последовательности праймеров, используемые для построения каждого из векторов, показаны в Таблице 8 ниже.
Сконструированный вектор pDZ-ΔpheA трансформировали в штамм СМ06-0003 методом электропорации и затем подвергли вторичному кроссинговеру для получения штамма, в котором удален pheA. Соответствующие генетические манипуляции были подтверждены посредством секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 35 и SEQ ID NO: 36, который, соответственно, может амплифицировать внешнюю область вышележащей области и нижележащей области гомологичной рекомбинации, откуда соответствующий ген был удален, полученный штамм называется СМ06-0004.
Пример 4: Оценка продуцирующей способности штаммов, продуцирующих L-тирозин, у которых удален ген pheA
Для подтверждения способности штаммов, сконструированных в Примере 3, к продуцированию L-тирозина, штаммы культивировали с использованием метода и композиций среды, описанных в Примере 2.
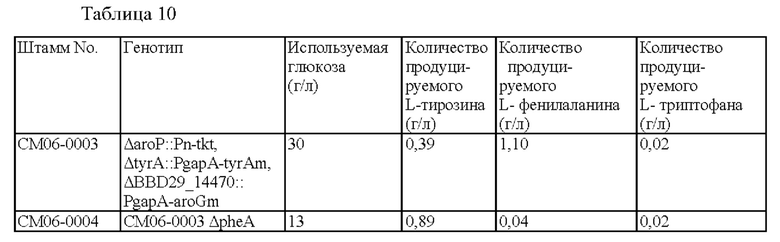
Результаты продуцирования L-тирозина, L-фенилаланина и L-триптофана в культурах штаммов, продуцирующих L-тирозин, СМ06-0003 и СМ06-0004, приведены в Таблице 10 выше. Штамм СМ06-0004, в котором удален pheA, продуцирует L-тирозин в качестве основного продукта со значительным снижением продукции L-фенилаланина. Как и в Примере 2, существенных изменений в продуцировании L-триптофана не наблюдали.
Однако, поскольку штамм СМ06-0004 не может продуцировать L-фенилаланин, он может расти только при наличии L-фенилаланина, содержащегося в дрожжевом экстракте среды, и демонстрирует только около 1/3 потребления сахара по сравнению со штаммом, в котором ген pheA не удален. То есть было подтверждено, что удаление гена pheA у микроорганизмов оказывает большое влияние на рост штаммов, при этом штаммы не могут быть использованы для получения целевого продукта.
Пример 5: Конструирование штаммов, продуцирующих L-тирозин, в которых заменен промотор гена pheA
Из результатов Примеров 2 и 4, можно предсказать, что концентрация L-триптофана точным образом регулируется так, чтобы не мешать росту клеток, при этом L-триптофан поддерживается в низкой концентрации в течение всего времени культивирования. Из литературы также известно, что продуцирование L-триптофана одновременно регулируется с помощью аттенюатора и промотороа в соответствии с концентрацией L-триптофана (Appl. Environ Microbiol 59 791, 1993).
Следовательно, ген pheA, продуцирующий L-фенилаланин, подвергается регуляторному механизму L-триптофана, так что регулирование гена pheA может осуществляться в соответствии с концентрацией L-триптофана.
В частности, в качестве контроля были также сконструированы и подвергнуты сравнению штаммы с введенным промотором ilvB, который позволяет регулировать pheA концентрацией L-изолейцина, а также промотором leuA и промотором leuC, которые позволяют регулировать pheA концентрацией L-лейцина, в дополнение к регуляторной области trpE, которая позволяет регулировать pheA концентрацией L-триптофана.
Для того чтобы ген pheA мог регулироваться регуляторной областью гена trpE, были получены вышележащая область, в которую следует вставить ген, регуляторная область trpE и нижележащая область, в которую следует вставить ген. В частности, фрагмент гена в вышележащей области, в которую следует вставить ген, был получен с использованием праймеров SEQ ID NO: 37 и SEQ ID NO: 38; фрагмент гена в регуляторной области trpE был получен с использованием праймеров SEQ ID NO: 39 и SEQ ID NO: 40; и фрагмент гена в нижележащей области, в которую следует вставить ген, был получен с использованием праймеров SEQ ID NO: 41 и SEQ ID NO: 42 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869, используемой в качестве матрицы, посредством ПЦР. Полимеразу SolgTM Pfu-X DNA использовали в качестве полимеразы, и ПЦР выполняли при следующих условиях амплификации: денатурация при 95°С в течение 5 минут, с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжига при 60°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд с последующей полимеризацией при 72°С в течение 5 минут
Амплифицированные вышележащие и нижележащие области, в которые должен быть вставлен ген, регуляторная область trpE и вектор pDZ для хромосомной трансформации расщепляли с помощью рестриктазы Smal, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, которая была названа pDZ-ΔPpheA::PtrpE. Клонирование проводили путем смешивания реагента для сборки Гибсона и каждого из фрагментов гена в рассчитанном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Для достижения регулирования гена pheA промотором гена ilvB, были получены вышележащая область, в которую должен быть вставлен ген, область промотора гена ilvB и нижележащая область, в которую должен быть вставлен ген. В частности, фрагмент гена в вышележащей области, в которую должен быть вставлен ген, был получен с использованием праймеров SEQ ID NO: 37 и SEQ ID NO: 43, фрагмент гена в промоторной области гена ilvB был получен с использованием праймеров SEQ ID NO: 44 и SEQ ID NO: 45, и фрагмент гена в нижележащей области, в которую должен быть вставлен ген, был получен с использованием праймеров SEQ ID NO: 46 и SEQ ID NO: 42 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869, используемой в качестве матрицы, посредством ПЦР. Полимеразу SolgTM Pfu-X DNA использовали в качестве полимеразы, и ПЦР выполняли при следующих условиях амплификации: денатурация при 95°С в течение 5 минут, с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжига при 60°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд с последующей полимеризацией при 72°С в течение 5 минут
Амплифицированные вышележащие и нижележащие области, в которые должен быть вставлен ген, область промотора гена ilvB и вектор pDZ для хромосомной трансформации расщепляли с помощью рестриктазы SmaI, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, которая была названа pDZ-ΔPpheA::PilvB. Клонирование проводили путем смешивания реагента для сборки Гибсона и каждого из фрагментов гена в рассчитанном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Для того, чтобы ген pheA мог регулироваться промотором гена leuA, были получены вышележащая область, в которую ген должен быть вставлен, область промотора leuA, и нижележащая область, в которую ген должен быть вставлен. В частности, фрагмент гена в вышележащей области, в которую ген должен быть вставлен, был получен с использованием праймеров SEQ ID NO: 37 и SEQ ID NO: 47, фрагмент гена в области промотора leuA был получен с использованием праймеров SEQ ID NO: 48 и SEQ ID NO: 49, и фрагмент гена в нижележащей области, в которую ген должен быть вставлен, был получен с использованием праймеров SEQ ID NO: 50 и SEQ ID NO: 42 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869, используемой в качестве матрицы, посредством ПЦР. Полимеразу SolgTM Pfu-X DNA использовали в качестве полимеразы, и ПЦР выполняли при следующих условиях амплификации денатурация при 95°С в течение 5 минут, с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжига при 60°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд с последующей полимеризацией при 72°С в течение 5 минут
Амплифицированные вышележащие и нижележащие области, в которые ген должен быть вставлен, область промотора leuA и вектор pDZ для хромосомной трансформации расщепляли с помощью рестриктазы SmaI, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, которая была названа pDZ-ΔPpheA::PleuA. Клонирование проводили путем смешивания реагента для сборки Гибсона и каждого из фрагментов гена в рассчитанном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Для того, чтобы ген pheA мог регулироваться промотором leuC, были получены вышележащая область, в которую должен быть вставлен ген, область промотора leuC и нижележащая область, в которую должен быть вставлен ген. В частности, фрагмент гена в вышележащей области, в которую ген должен быть вставлен, был получен с использованием праймеров SEQ ID NO: 37 и SEQ ID NO: 51, фрагмент гена в области промотора leuC был получен с использованием праймеров SEQ ID NO: 52 и SEQ ID NO: 53, а фрагмент гена в нижележащей области, в которую ген должен быть вставлен, был получен с использованием праймеров SEQ ID NO: 54 и SEQ ID NO: 42 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869, используемой в качестве матрицы, посредством ПЦР. Полимеразу SolgTM Pfu-X DNA использовали в качестве полимеразы, и ПЦР выполняли при следующих условиях амплификации: денатурация при 95°С в течение 5 минут, с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжига при 60°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд с последующей полимеризацией при 72°С в течение 5 минут
Амплифицированные вышележащие и нижележащие области, в которуые ген должен быть вставлен, область промотора гена leuC и вектор pDZ для хромосомной трансформации расщепляли с помощью рестриктазы SmaI и клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, которая была названа pDZ-ΔPpheA::PleuC. Клонирование проводили путем смешивания реагента для сборки Гибсона и каждого из фрагментов гена в рассчитанном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Сконструированные векторы pDZ-ΔPpheA::PtrpE, pDZ-ΔPpheA::PilvB, pDZ-ΔPpheA::PleuA, и pDZ-ΔPpheA::PleuC трансформировали в штамм СМ06-0003 методом электропорации, а затем подвергали вторичному кроссинговеру для получения штаммов, в которых ген pheA мог регулироваться регуляторной областью trpE или промотором ilvB, leuA или leuC. Соответствующие генетические манипуляции были подтверждены посредством секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 55 и SEQ ID NO: 56, который может соответственно амплифицировать внешнюю область вышележащей области и нижележащей области гомологичной рекомбинации, в которую была вставлена соответствующая регуляторная область или промотор. Полученный штамм, в который была вставлена регуляторная область trpE выше гена pheA, был назван СМ06-0005, полученный штамм, в который был вставлен промотор ilvB, был назван СМ06-0006, полученный штамм, в который был вставлен промотор leuA, был назван СМ06-0007, а полученный штамм, в который был вставлен промотор leuC, был назван СМ06-0008.
Последовательности праймеров, используемые для построения каждого из векторов, показаны в Таблице 11 ниже.

Пример 6: Оценка продуцирующей способности штаммов с заменой
промотора гена pheA штаммов, продуцирующих L-тирозин
Для подтверждения L-тирозин-продуцирующей способности штаммов, сконструированных в Примере 5, штаммы культивировали с использованием метода и композиций среды, описанных в Примере 2.
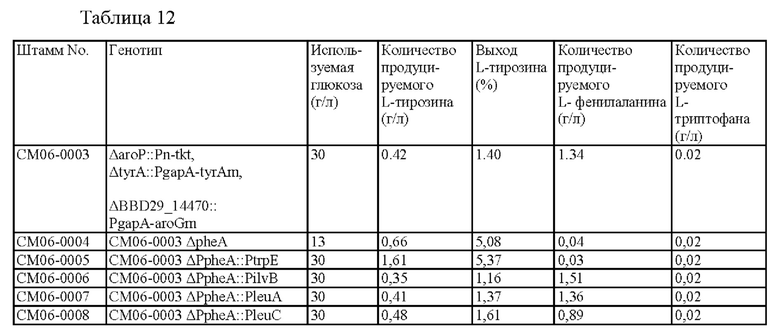
Результаты продуцирования L-тирозина, L-фенилаланина и L-триптофана в культурах штаммов, продуцирующих L-тирозин, СМ06-0003, СМ06-0004, СМ06-0005, СМ06-0006, СМ06-0007 и СМ06-0008 показаны в Таблице 12 выше.
Для штамма СМ06-0004, в котором удален pheA, L-тирозин был получен с выходом 5,08%. Таким образом, в случае штамма СМ06-0005, в котором путь L-фенилаланина мог регулироваться концентрацией L-триптофана, он продуцировал L-тирозин с продуцированием L-фенилаланина в небольшом количестве, и при этом можно было ожидать более высокого выхода L-тирозина по сравнению со штаммом СМ06-0003, в котором путь L-фенилаланина не регулировался, и можно было ожидать более низкого его выхода по сравнению со штаммом СМ06-0004, в котором был удален путь L-фенилаланина. Однако, вопреки ожиданиям, штамм СМ06-0005 показал улучшенную продуцирующую способность по сравнению со штаммом СМ06-0004, продуцируя L-тирозин с выходом 5,37%. Продуцирование L-тирозина штаммом СМ06-0005 увеличилось на 283% по сравнению с исходным штаммом СМ06-0003 и на 144% по сравнению со штаммом СМ06-0004. Приведенные выше результаты подтверждают, что когда pheA может регулироваться концентрацией L-триптофана, продуцирование L-тирозина может значительно увеличиться до неожиданного уровня. Кроме того, в качестве контроля, аналогичного применению L-триптофан-регуляторного механизма на ген pheA, продуцирование L-тирозина было значительно увеличено по сравнению с СМ06-0006 штаммом, в котором pheA мог регулироваться концентрацией L-изолейцина, и штаммами СМ06-0007 и СМ06-0008, в которых регулирование pheA осуществлялось концентрацией L-лейцина. Таким образом, было подтверждено, что введение механизма регулирования L-триптофана на pheA оказывает больший синергетический эффект на продуцирование L-тирозина по сравнению с удалением pheA или введением других механизмов регулирования.
Пример 7: Конструирование штаммов, продуцирующих L-тирозин, с системой, отличной от фосфотрансферазной (PTS), у которых удален ген pheA, и штаммов, у которых заменен промотор
Поскольку продуцирование L-тирозина начинается с PEP и Е4Р в качестве предшественников, использование системы, отличной от фосфотрансферазной системы (PTS), может обеспечить увеличение поставок PEP, и, следовательно, возможно высокое продуцирование L-тирозина (Nature biotechnol 14 620, 1996). Поэтому, ptsG, ген PTS штамма, был удален, и был введен Zymomonas mobilis АТСС10988-полученный glf ген, не относящийся к PTS.
Для удаления ptsG и вставки glf были получены вышележащие и нижележащие области, в которые следовало вставить glf, полученный из Zymomonas mobilis. В частности, фрагмент гена в вышележащей области гена ptsG был получен с использованием праймеров SEQ ID NO: 57 и SEQ ID NO: 58, а фрагмент гена в нижележащей области гена ptsG был получен с использованием праймеров SEQ ID NO: 59 и SEQ ID NO: 60 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869, используемой в качестве матрицы, посредством ПЦР. Полимеразу SolgTM Pfu-X DNA использовали в качестве полимеразы, и ПЦР выполняли при следующих условиях амплификации денатурация при 95 С в течение 5 минут, с последующими 30 циклами денатурации при 95 С в течение 30 секунд, отжига при 60 С в течение 30 секунд и полимеризации при 72 С в течение 60 секунд с последующей полимеризацией при 72 С в течение 5 минут
Кроме того, для того, чтобы получить ген glf включающий хорошо известный промотор cj7 (SEQ ID NO: 61, Патент Кореи No. 10-0620092), фрагмент промотора cj7 был получен с использованием праймеров SEQ ID NO: 62 и SEQ ID NO: 63 на основании синтетической ДНК промотора cj7 в качестве матрицы, посредством ПЦР, и фрагмент гена glf был получен с использованием праймеров SEQ ID NO: 64 и SEQ ID NO: 65 на основании хромосомной ДНК Zymomonas mobilis АТСС10988 в качестве матрицы посредством ПЦР. Полимеразу SolgTM Pfu-X DNA использовали в качестве полимеразы, и ПЦР выполняли при следующих условиях амплификации денатурация при 95 С в течение 5 минут, с последующими 30 циклами денатурации при 95 С в течение 30 секунд, отжига при 60 С в течение 30 секунд и полимеризации при 72 С в течение 60 секунд с последующей полимеризацией при 72 С в течение 5 минут
Амплифицированные вышележащие и нижележащие области гена ptsG, фрагмент гена glf включающий промотор cj7, и вектор pDZ для хромосомной трансформации расщепляли рестриктазой SeaI и клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, которая была названа pDZ-ΔptsG::pcj7-glf. Клонирование проводили путем смешивания реагента для сборки Гибсона и каждого из фрагментов гена в рассчитанном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Последовательности праймеров, используемые в Примере 7, показаны в Таблице 13 ниже.
Сконструированный вектор pDZ-ΔptsG::pcj7-glf трансформировали в штамм СМ06-0003 методом электропорации и затем подвергали вторичному кроссинговеру для получения штамма, в который был вставлен Zymomonas mobilis-полученный ген glf включающий промотор cj7. Соответствующие генетические манипуляции были подтверждены посредством секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 66 и SEQ ID NO: 67, который может соответственно амплифицировать внешнюю область вышележащей области и нижележащей области гомологичной рекомбинации, куда был вставлен соответствующий ген. Полученный штамм называется СМ06-0009.
Вектор pDZ-ΔpheA, сконструированный в Примере 3, трансформировали в штамм СМ06-0009, созданный выше, методом электропорации и затем подвергали вторичному кроссинговеру для получения штамма, в котором был удален ген pheA. Соответствующие генетические манипуляции были подтверждены посредством секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 35 и SEQ ID NO: 36, который может, соответственно, амплифицировать внешнюю область вышележащей области и нижележащей области гомологичной рекомбинации, откуда соответствующий ген был удален. Полученный штамм называется СМ06-0011.
Каждый из векторов pDZ-ΔPpheA::PtrpE, pDZ-ΔPpheA::PilvB, pDZ-ΔPpheA::PleuA, and pDZ-ΔPpheA::PleuC, сконструированных в Примере 5, был трансформирован в штамм СМ06-0009, сконструированный выше, методом электропорации и затем подвергнут вторичному кроссинговеру для получения штаммов, в которые регуляторная область или промотор вставлены выше гена pheA. Соответствующие генетические манипуляции были подтверждены посредством секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 55 и SEQ ID NO: 56, который может, соответственно, амплифицировать внешнюю область вышележащей области и нижележащей области гомологичной рекомбинации, куда вставлена соответствующая регуляторная область или промотор. Полученный штамм, в который регуляторная область trpE вставлена выше гена pheA, называется СМ06-0010, полученный штамм, в который вставлен промотор ilvB, называется СМ06-0012, полученный штамм, в который вставлен промотор leuA, называется СМ06-0013, и полученный штамм, в который вставлен промотор leuC называется СМ06-0014.
Пример 8: Оценка продуцирующей способности штаммов, продуцирующих L-тирозин, с системой, отличной от фосфотрансферазной (PTS), у которых удален ген pheA, и штаммов, у которых заменен промотор
Для подтверждения способности штаммов, сконструированных в Пример 7, к продуцированию L-тирозина, штаммы культивировали с использованием метода и композиции среды, описанных в Примере 2.
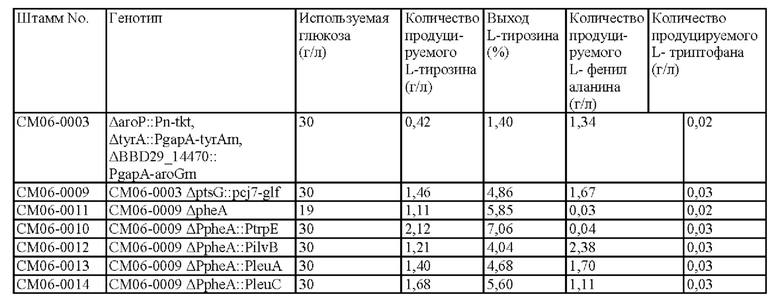
Результаты продуцирования L-тирозина, L-фенилаланина и L-триптофана в культурах штаммов, продуцирующих L-тирозин, СМ06-0003, СМ06-0009, СМ06 - ООП, СМ06-0010, СМ06-0012, СМ06-0013 и СМ06-0014, показаны в таблице 15 выше.
Как и ожидалось, было подтверждено, что продуцирование L-тирозина и L-фенилаланина было увеличено в штамме СМ06-0009, не являющимся штаммом с PTS, способным усиливать подачу PEP, по сравнению со штаммом СМ06-0003, который является штаммом с PTS. Аналогично, в этом случае не наблюдалось значительного увеличения продуцирования L-триптофана.
В случае штамма СМ06-0011, в котором был удален pheA, на основе штамма, не являющимся штаммом с PTS, было подтверждено, что, хотя продуцирование L-тирозина было увеличено по сравнению с исходным штаммом с выходом 5,85%, конечное количество продуцируемого L-тирозина было уменьшено по сравнению с родительским штаммом, а продуктивность была снижена из-за очень низкого потребления сахара. Как и в Примере 6, можно было ожидать, что штамм СМ06-0010 может продуцировать L-тирозин с более низким выходом. Однако на самом деле СМ06-0010 продуцировал L-тирозин с выходом 7,06%, который был увеличен на 20,7% по сравнению со штаммом СМ06-0011. Кроме того, количество L-тирозина, продуцируемого штаммом СМ06-0010 было увеличено на 45% по сравнению с родительским штаммом СМ06-0009 и на 91% по сравнению со штаммом СМ06-0011. Кроме того, было также подтверждено, что продуцирование L-тирозина было значительно увеличена как по сравнению со штаммом СМ06-00012, у которого концентрация L-изолейцина могла регулировать pheA, так и со штаммами СМ06-00013 и СМ06-00014, в которых концентрация L-лейцина могла регулировать pheA, тем самым подтверждая, что введение механизма регулирования L-триптофаном в pheA оказывает больший синергетический эффект на продуцирование L-тирозина по сравнению с удалением pheA или введением других механизмов регулирования.
Пример 9. Скрининг микроорганизмов, устойчивых к аналогам L-тирозина
В этом Примере, чтобы ослабить ингибирование по принципу обратной связи L-тирозина на основе Corynebacterium glutamicum АТСС 13869 в качестве родительского штамма, был проведен эксперимент по отбору штаммов, устойчивых к гидроксамату L-тирозина, который является аналогом L-тирозина, с использованием метода искусственной мутации.
В частности, мутация была индуцирована методом искусственного мутагенеза с использованием N-метил-N'-нитро-N-нитрозогуанидина (далее именуемого NTG). Штамм АТСС 13869 культивировали в посевной среде в течение 18 часов и инокулировали в 4 мл посевной среды, а затем культивировали до тех пор, пока OD660 не достигал примерно 1,0. После центрифугирования культуральной среды для выделения клеток, которые дважды промывали 50 mM Трис-малатным буфером (рН 6,5) и суспендировали в конечном объеме, составляющем 4 мл этого же буфера. Раствор NTG (2 мг/мл в 0,05 Μ Tris-малатного буфера (рН 6,5)) добавляли к суспензии клеток до конечной концентрации 150 мг/л, и полученному продукту давали постоять при комнатной температуре в течение 20 минут, а затем центрифугировали для сбора клеток, и клетки дважды промывали тем же буфером для удаления NTG. Окончательно промытые клетки суспендировали в 4 мл 20%-го раствора глицерина и хранили при -70°С до использования. NTG-обработанные штаммы высевали в минимальной среде, содержащей 0,5 г/л гидроксамат L-тирозина и благодаря этому процессу было получено 100 штаммов, обладающих устойчивостью к гидроксамат L-тирозину.
Пример 10. Оценка способности штаммов, устойчивых к гидроксамату L-тирозина, продуцировать L-тирозин
Была подтверждена L-тирозин-продуцирующая способность устойчивых к гидроксамату L-тирозина 100 штаммов, полученных в Примере 9. Каждый из 100 штаммов, полученных в Примере 9, инокулировали в 250-мл колбу с угловой перегородкой, содержащую 25 мл посевной среды, а затем культивировали при встряхивании при 200 об/мин при 30°С в течение 20 часов. Затем 1 мл раствора посевной культуры инокулировали в 250-мл колбе с угловой перегородкой, содержащей 24 мл среды для продуцирования и культивировали при встряхивании при 200 об/мин при 30°С в течение 48 часов.
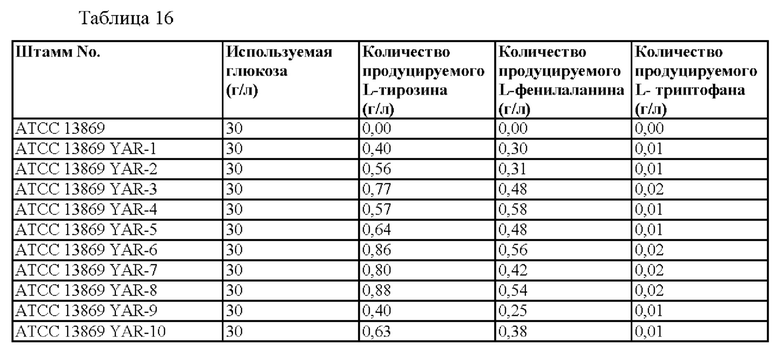
После культивирования количество продуцируемых аминокислот измеряли методом ВЭЖХ. Среди 100 протестированных штаммов концентрация аминокислот в культивируемой среде для 10 лучших штаммов, которые показали отличную способность продуцировать L-тирозин, показана в таблице 16. 10 штаммов-кандидатов, идентифицированных с помощью вышеупомянутого процесса, были названы как АТСС 13869 YAR-1 и АТСС 13869 YAR-10 соответственно.
Среди них были отобраны штаммы АТСС 13869 YAR-6 и АТСС 13869 YAR-8, обладающие самой высокой способностью продуцировать L-тирозин.
Пример 11. Замена промотора гена pheA у штаммов, обладающих устойчивостью к гидроксамату L-тирозина
Ген pheA штаммов АТСС 13869 YAR-6 и YAR-8, отобранных в Пример 10, может регулироваться регуляторной областью оперона trp.С этой целью, вектор pDZ-ΔPpheA::PtrpE, сконструированный в Примере 5, трансформировали в каждый из штаммов АТСС 13869 YAR-6 и YAR-8 методом электропорации и затем подвергали вторичному кроссинговеру для получения штаммов, в которых ген pheA мог регулироваться регуляторной областью trpE. Соответствующие генетические манипуляции были подтверждены посредством секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 55 и SEQ ID NO: 56, который может, соответственно, амплифицировать внешнюю область вышележащей области и нижележащей области гомологичной рекомбинации, куда была вставлена соответствующая регуляторная область. Полученные штаммы АТСС 13869 YAR-6 и YAR-8, в которые была вставлена регуляторная область trpE выше гена pheA, были названы, как YAR-6P и YAR-8P, соответственно.
Пример 12. Оценка продуцирующей способности устойчивых к гидроксамату L-тирозина штаммов АТСС 13869, в которых был заменен промотор гена pheA
Для подтверждения способности штаммов, сконструированных в Примере 11, к продуцированию L-тирозина, штаммы культивировали с использованием метода и композиции среды, описанных в Примере 2.

Результаты продуцирования L-тирозина, L-фенилаланина и L-триптофана в культурах штаммов АТСС 13869, устойчивых к гидроксамат L-тирозину, и штаммов, в которых заменен промотор гена pheA, показаны в Таблице 17 выше.
Как и в Примерах 6 и 8, где ген pheA может регулироваться регуляторной областью trpE, можно ожидать снижения продуцирования L-фенилаланина и увеличения продуцирования L-тирозина. Фактические экспериментальные результаты подтвердили, что в штаммах АТСС 13869 YAR-6P и АТСС 13869 YAR-8P значительно снижается продуцирование L-фенилаланина в качестве побочного продукта и увеличивается продуцирование L-тирозина в качестве целевого продукта, причем на более высоком уровне по сравнению с сокращением продуцирования L-фенилаланина. Основываясь на этих результатах, было подтверждено, что при использовании механизма регулирования гена pheA L-триптофаном количество продуцируемого L-тирозина может быть значительно повышено до непредсказуемого уровня без снижения нормы потребления сахара, и комбинация регуляторной области оперона trp и гена pheA может привести к значительному синергетическому эффекту на продуцирование L-тирозина.
Приведенные выше результаты подтвердили, что продуцирование L-тирозина было значительно увеличено до неожиданного уровня, при регулировании гена pheA концентрацией L-триптофана. В конкретном примере было подтверждено, что продуцирование L-тирозина было значительно увеличено до неожиданного уровня даже у случайно мутировавших штаммов, а также у штаммов, которые были модифицированы для повышения способности продуцировать L-тирозин.
Штамм СМ06-0010 был депонирован в Корейском культурном центре микроорганизмов (KCCM), Международном органе по депонированию, в соответствии с Будапештским договором 15 апреля 2019 года, с регистрационным номером KCCM12487P.
Исходя из вышесказанного, специалист в области техники, к которой относится настоящее изобретение, сможет понять, что настоящее изобретение может быть воплощено в других конкретных формах без изменения технических концепций или существенных характеристик настоящего изобретения. В этой связи примеры воплощений, раскрытые в настоящем документе, предназначены только для иллюстративных целей и не должны истолковываться как ограничивающие рамки настоящего изобретения. Напротив, настоящее изобретение предназначено для охвата не только примерных воплощений, но также различных альтернатив, модификаций, эквивалентов и других воплощений, которые могут быть включены в сущность и объем настоящего изобретения, как определено в прилагаемой формуле изобретения.
--->
<110> CJ CheilJedang Corporation
<120> L-TYROSINE-PRODUCING MICROORGANISM AND METHOD FOR PRODUCING
L-TYROSINE USING THE SAME
<130> OPA20084
<150> KR 10-2019-0071797
<151> 2019-06-17
<160> 68
<170> KoPatentIn 3.0
<210> 1
<211> 294
<212> DNA
<213> Unknown
<220>
<223>регуляторнаяобластьоперона ttp
<400> 1
taattgagac aagcttccca ctatgtgata aagtcccatt ttgtgaataa ctcttgtctc 60
agtcaaagca cccagtggtg gtggcgcgct aactaagcga gcctgacacc tcaagttgtt 120
ttcactttga tgaatttttt aaggctcgta cttcgttcga cgaagaagcg ggccttttgt 180
ggtttttagc ccacaaccgg caagccctgg atcgaatgaa gctcgcagcg agtaattatt 240
tgatgtttcc cagaaaggct tcagccccac aatgatttcc tcggtaggtg cccc 294
<210> 2
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 2
tcgagctcgg taccctggga acttgtcgac gctat 35
<210> 3
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 3
tgttcggcaa gcattgtggt gtgggcaatg atcac 35
<210> 4
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 4
attaacggtt aaagtactca ttgtgaggtg gcggg 35
<210> 5
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 5
ctctagagga tccccggagc tgctgtccaa cgtgg 35
<210> 6
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 6
ccacaccaca atgcttgccg aacatttttc ttttc 35
<210> 7
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 7
cacaatgagt actttaaccg ttaatggagt ccttg 35
<210> 8
<211> 17
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 8
acgcgccaag tcggacg 17
<210> 9
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 9
cgcacgatgt ttacctgcg 19
<210> 10
<211> 1122
<212> DNA
<213> Unknown
<220>
<223>ген tyrA
<400> 10
atggttgctg aattgaccgc attacgcgat caaattgatg aagtcgataa agcgctgctg 60
aatttattag cgaagcgtct ggaactggtt gctgaagtgg gcgaggtgaa aagccgcttt 120
ggactgccta tttatgttcc ggagcgcgag gcatctattt tggcctcgcg tcgtgcagag 180
gcggaagctc tgggtgtacc gccagatctg attgaggatg ttttgcgtcg ggtgatgcgt 240
gaatcttact ccagtgaaaa cgacaaagga tttaaaacac tttgtccgtc actgcgtccg 300
gtggttatcg tcggcggtgg cggtcagatg ggacgcctgt tcgagaagat gctgaccctc 360
tcgggttatc aggtgcggat tctggagcaa catgactggg atcgagcggc tgatattgtt 420
gccgatgccg gaatggtgat tgttagtgtg ccaatccacg ttactgagca agttattggc 480
aaattaccgc ctttaccgaa agattgtatt ctggtcgatc tggcatcagt gaaaaatggg 540
ccattacagg ccatgctggt ggcgcatgat ggtccggtgc tggggctaca cccgatgttc 600
ggtccggaca gcggtagcct ggcaaagcaa gttgtggtct ggtgtgatgg acgtaaaccg 660
gaagcatacc aatggtttct ggagcaaatt caggtctggg gcgctcggct gcatcgtatt 720
agcgccgtcg agcacgatca gaatatggcg tttattcagg cactgcgcca ctttgctact 780
tttgcttacg ggctgcacct ggcagaagaa aatgttcagc ttgagcaact tctggcgctc 840
tcttcgccga tttaccgcct tgagctggcg atggtcgggc gactgtttgc tcaggatccg 900
cagctttatg ccgacatcat tatgtcgtca gagcgtaatc tggcgttaat caaacgttac 960
tataagcgtt tcggcgaggc gattgagttg ctggagcagg gcgataagca ggcgtttatt 1020
gacagtttcc gcaaggtgga gcactggttc ggcgattacg tgcagcgttt tcagagtgaa 1080
agccgcgtgt tattgcgtca ggcgaatgac aatcgccagt aa 1122
<210> 11
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 11
ttcgagctcg gtaccctatc aaaaccgagt tcttcc 36
<210> 12
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 12
gtcgttttta ggcctcctga caagtgtggc acatac 36
<210> 13
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 13
tgacaatcgc cagtaatttt atcggctgat gattct 36
<210> 14
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 14
actctagagg atccccaacg cgattgcatt cggctc 36
<210> 15
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 15
gtgccacact tgtcaggagg cctaaaaacg accgag 36
<210> 16
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 16
tcaattcagc aaccatgttg tgtctcctct aaagat 36
<210> 17
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 17
ttagaggaga cacaacatgg ttgctgaatt gaccgc 36
<210> 18
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 18
tcatcagccg ataaaattac tggcgattgt cattcg 36
<210> 19
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 19
gcccactagt cgaatccc 18
<210> 20
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 20
ctgtccgcaa cctgtgcg 18
<210> 21
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 21
tcgagctcgg taccccccgg cggtatcgag gtagt 35
<210> 22
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 22
gacaagttta gtactttaat cacccgcggg gaccc 35
<210> 23
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 23
ggtgattaaa gtactaaact tgtcccgagg gtgag 35
<210> 24
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 24
ctctagagga tcccctatca gtcacttccc tgaga 35
<210> 25
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 25
gcgggtgatt aaagtgaggc ctaaaaacga ccgag 35
<210> 26
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 26
gttctgataa ttcatgttgt gtctcctcta aagat 35
<210> 27
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 27
atgaattatc agaacgacga 20
<210> 28
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 28
cgggacaagt ttagtttacc cgcgacgcgc tttta 35
<210> 29
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 29
ttgatatgac cgcagcctga 20
<210> 30
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 30
ctgcattctc atcgatcttg 20
<210> 31
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 31
ttcgagctcg gtacccccga ccaggccaca cgcg 34
<210> 32
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 32
aataatccgg ccggccatgg ggttacacag cttaacccgc 40
<210> 33
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 33
cgggttaagc tgtgtaaccc catggccggc cggattatt 39
<210> 34
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 34
cgactctaga ggatccccgc tgacaacagc aacgtcg 37
<210> 35
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 35
ccagcgatga tcgcgccg 18
<210> 36
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 36
atcgccgtgg agccagcc 18
<210> 37
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 37
ttcgagctcg gtacccggag gggtttccac ctcg 34
<210> 38
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 38
tgggaagctt gtctcaatta tgtctgttgc tcaattagcg 40
<210> 39
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 39
ctaattgagc aacagacata attgagacaa gcttccca 38
<210> 40
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 40
aattggtgcg tcgctcatgg ggcacctacc gaggaa 36
<210> 41
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 41
ttcctcggta ggtgccccat gagcgacgca ccaattgttg 40
<210> 42
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 42
cgactctaga ggatcccccc gaagagttcg gctgcg 36
<210> 43
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 43
cagcgctaat cttggctctg tctgttgctc aattagcg 38
<210> 44
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 44
ctaattgagc aacagacaga gccaagatta gcgctgaa 38
<210> 45
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 45
aattggtgcg tcgctcatcc gctcaggggc ggcgg 35
<210> 46
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 46
tccgccgccc ctgagcggat gagcgacgca ccaattgttg 40
<210> 47
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 47
ggggggaact ttggagtttg tctgttgctc aattagcg 38
<210> 48
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 48
ctaattgagc aacagacaaa ctccaaagtt ccccccc 37
<210> 49
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 49
aattggtgcg tcgctcattg tgttcaacct tcttaaaaag 40
<210> 50
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 50
ttaagaaggt tgaacacaat gagcgacgca ccaattgttg 40
<210> 51
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 51
aaatgtaaga ttcaaagatg tctgttgctc aattagcg 38
<210> 52
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 52
ctaattgagc aacagacatc tttgaatctt acatttcata g 41
<210> 53
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 53
aattggtgcg tcgctcatgg aactcaccgt ccttac 36
<210> 54
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 54
gtaaggacgg tgagttccat gagcgacgca ccaattgttg 40
<210> 55
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 55
ccagcgatga tcgcgccg 18
<210> 56
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 56
atcgccgtgg agccagcc 18
<210> 57
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 57
ttcgagctcg gtacccgagg gctcactgac gttga 35
<210> 58
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 58
cgctgggatg tttctaccgg attcgattcc tcag 34
<210> 59
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 59
cgctcccaga agtaggctca aaccttgccc ataac 35
<210> 60
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 60
ctctagagga tccccctccc ccaaaccacg ctttt 35
<210> 61
<211> 318
<212> DNA
<213> Unknown
<220>
<223>промотор cj7
<400> 61
agaaacatcc cagcgctact aatagggagc gttgaccttc cttccacgga ccggtaatcg 60
gagtgcctaa aaccgcatgc ggcttaggct ccaagatagg ttctgcgcgg ccgggtaatg 120
catcttcttt agcaacaagt tgaggggtag gtgcaaataa gaacgacata gaaatcgtct 180
cctttctgtt tttaatcaac atacaccacc acctaaaaat tccccgacca gcaagttcac 240
agtattcggg cacaatatcg ttgccaaaat attgtttcgg aatatcatgg gatacgtacc 300
caacgaaagg aaacactc 318
<210> 62
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 62
ggaatcgaat ccggtagaaa catcccagcg ctact 35
<210> 63
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 63
tactttcaga actcatgagt gtttcctttc gttgg 35
<210> 64
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 64
acgaaaggaa acactcatga gttctgaaag tagtc 35
<210> 65
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 65
gggcaaggtt tgagcctact tctgggagcg ccaca 35
<210> 66
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 66
acattaagtg gtaggcgctg a 21
<210> 67
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 67
cataacaggg cagaacaaac 20
<210> 68
<211> 1053
<212> DNA
<213> Unknown
<220>
<223>ген aroG
<400> 68
atgaattatc agaacgacga tttacgcatc aaagaaatca aagagttact tcctcctgtc 60
gcattgctgg aaaaattccc cgctactgaa aatgccgcga atacggttgc ccatgcccga 120
aaagcgatcc ataagatcct gaaaggtaat gatgatcgcc tgttggttgt gattggccca 180
tgctcaattc atgatcctgt cgcggcaaaa gagtatgcca ctcgcttgct ggcgctgcgt 240
gaagagctga aagatgagct ggaaatcgta atgcgcgtct attttgaaaa gccgcgtacc 300
acggtgggct ggaaagggct gattaacgat ccgcatatgg ataatagctt ccagatcaac 360
gacggtctgc gtatagcccg taaattgctg cttgatatta acgacagcgg tctgccagcg 420
gcaggtgagt ttctcgatat gatcacccca caatatctcg ctgacctgat gagctggggc 480
gcaattggcg cacgtaccac cgaatcgcag gtgcaccgcg aactggcatc agggctttct 540
tgtccggtcg gcttcaaaaa tggcaccgac ggtacgatta aagtggctat cgatgccatt 600
aatgccgccg gtgcgccgca ctgcttcctg tccgtaacga aatgggggca ttcggcgatt 660
gtgaatacca gcggtaacgg cgattgccat atcattctgc gcggcggtaa agagcctaac 720
tacagcgcga agcacgttgc tgaagtgaaa gaagggctga acaaagcagg cctgccagca 780
caggtgatga tcgatttcag ccatgctaac tcgtccaaac aattcaaaaa gcagatggat 840
gtttgtgctg acgtttgcca gcagattgcc ggtggcgaaa aggccattat tggcgtgatg 900
gtggaaagcc atctggtgga aggcaatcag agcctcgaga gcggggagcc gctggcctac 960
ggtaagagca tcaccgatgc ctgcatcggc tgggaagata ccgatgctct gttacgtcaa 1020
ctggcgaatg cagtaaaagc gcgtcgcggg taa 1053
<---
Изобретение относится к микроорганизму, продуцирующему L-тирозин, и получению L-тирозина с использованием этого микроорганизма. Предложен микроорганизм рода Corynebacterium, продуцирующий L-тирозин, содержащий регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью, где регуляторная область оперона trp содержит промотор (промотор trp). Также предложены способ получения L-тирозина с использованием указанного микроорганизма, экспрессионная кассета, содержащая регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью, где регуляторная область оперона trp содержит промотор trp, а также композиция для получения L-тирозина, содержащая регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью, экспрессионную кассету, содержащую то же самое, и микроорганизм рода Corynebacterium, содержащий то же самое, где регуляторная область оперона trp содержит промотор trp. Изобретение обеспечивает увеличение выхода продукции L-тирозина. 4 н. и 4 з.п. ф-лы, 17 табл., 12 пр.
1. Микроорганизм рода Corynebacterium, продуцирующий L-тирозин, содержащий регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью, где регуляторная область оперона trp содержит промотор (промотор trp).
2. Микроорганизм по п. 1, где регуляторная область оперона trp расположена против хода транскрипции по отношению к гену, кодирующему префенатдегидрогеназу.
3. Микроорганизм по п. 1, где ген, кодирующий префенатдегидрогеназу, представляет собой ген pheA.
4. Микроорганизм по п. 1, где регуляторная область оперона trp дополнительно содержит одно или более, выбранное из группы, состоящей из регулятора trp, оператора trp, лидерного пептида trp и аттенюатора trp.
5. Микроорганизм по п. 1, где регуляторная область оперона trp состоит из нуклеотидной последовательности SEQ ID NO: 1.
6. Способ получения L-тирозина, включающий культивирование микроорганизма по любому из пп. 1-5 в среде и выделение L-тирозина из культивируемого микроорганизма или среды.
7. Экспрессионная кассета, содержащая регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью, где регуляторная область оперона trp содержит промотор (промотор trp).
8. Композиция для получения L-тирозина, содержащая регуляторную область оперона trp и ген, кодирующий префенатдегидрогеназу, функционально связанный с этой областью, экспрессионную кассету, содержащую то же самое, и микроорганизм рода Corynebacterium, содержащий то же самое, где регуляторная область оперона trp содержит промотор (промотор trp).
| GOWRISHANKAR J | |||
| ET AL | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| J Bacteriol | |||
| Устройство для видения на расстоянии | 1915 |
|
SU1982A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| УСТРОЙСТВО ДЛЯ ИЗМЕРЕНИЯ АЭРОДИНАМИЧЕСКОЙ КАСАТЕЛЬНОЙ СИЛЫ НА ЦИЛИНДРЕ | 1965 |
|
SU216333A1 |
| US 7700328 B2, | |||

