[0001] Настоящая заявка испрашивает приоритет по предварительной заявке США № 62/484,217, поданной 11 апреля 2017 г., озаглавленной «ПОЛИСПЕЦИФИЧЕСКИЕ ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ДВОЙНОЙ ЭФФЕКТОРНОЙ ФУНКЦИЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ», содержание которой включено в качестве ссылки в полном объеме.
Включение в качестве ссылки списка последовательностей
[0002] Настоящая заявка подается вместе со списком последовательностей в электронном формате. Список последовательностей предоставляется в виде файла 744952000140SeqList.TXT, созданного 11 апреля 2018 года, размер которого составляет 174179 байт. Информация в электронном формате Списка последовательностей включена посредством ссылки в полном объеме.
ОБЛАСТЬ РАСКРЫТИЯ
[0003] Изобретение в целом относится к полиспецифическим полипептидам, обладающим ограниченным связыванием с CD3. В некоторых вариантах осуществления полиспецифичные полипептиды содержат расщепляемые линкеры, которые при расщеплении приводят к двойным эффекторным функциям. Также предложены способы получения и применения этих полиспецифических полипептидов при различных терапевтических, диагностических и профилактических показаниях.
УРОВЕНЬ ТЕХНИКИ
[0004] Терапевтические антитела, которые вызывают истощение клеток-мишеней, обычно зависят от эффекторных функций, опосредованных через взаимодействие с Fc-гамма-рецепторами (FcγR) и белками комплемента. Эффекторные клетки, экспрессирующие FcγR, представляют собой преимущественно клетки врожденной иммунной системы. Т-клетки не являются прямыми эффекторными клетками, вовлеченными в опосредованное антителами истощение клеток-мишеней.
[0005] CD3 (кластер дифференцировки 3) Т-клеточный корецептор представляет собой мультимерный белок, состоящий из четырех отдельных полипептидных цепей, называемых цепями ε, γ, δ и ξ. Комплекс CD3 служит сигнальным модулем Т-клеточного рецептора, который нековалентно связывается с антигенсвязывающими а/b-цепями Т-клеточного рецептора (TCR).
[0006] Поскольку прямое рекрутирование CD3 приводит к активации Т-клеток, он является желательной мишенью при различных терапевтических и/или диагностических показаниях. Соответственно, существует потребность в антителах и терапевтических средствах, которые нацелены на путь CD3/TCR.
СУЩНОСТЬ РАСКРЫТИЯ
[0007] Настоящее раскрытие относится к полиспецифическим полипептидным конструкциям, которые демонстрируют ограниченное связывание с CD3. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция состоит из первого компонента, содержащего Fc-область иммуноглобулина, и второго компонента, содержащего CD3-связывающую область, где первый и второй компоненты связаны линкером или функционально связаны, где Fc-область расположена на N-конце относительно CD3-связывающей области; и один или оба из первого и второго компонентов содержат антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA). В некоторых вариантах осуществления полиспецифическая полипептидная конструкция в неактивном состоянии состоит из первого компонента и второго компонента, где первый и второй компоненты функционально связаны, причем каждый из первого и второго компонентов содержит антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA), где первый компонент содержит Fc-область, где второй компонент содержит CD3-связывающую область, и где первый и второй компоненты связаны расщепляемым линкером. В некоторых вариантах осуществления CD3-связывающая область связывается с CD3 (CD3ε).
[0008] В некоторых вариантах осуществления антигенсвязывающий домен расположен на аминоконце относительно Fc-области и/или на карбокси-конце относительно CD3-связывающей области полиспецифической полипептидной конструкции. В некоторых вариантах осуществления первый компонент содержит первый антигенсвязывающий домен, а второй компонент содержит второй антигенсвязывающий домен, где каждый из антигенсвязывающих доменов связывается с опухолеспецифическим антигеном (ТАА). В некоторых случаях первый антигенсвязывающий домен расположен на аминоконце полиспецифической конструкции, а второй антигенсвязывающий домен расположен на карбокси-конце полиспецифической конструкции. В некоторых вариантах осуществления первый антигенсвязывающий домен расположен на аминоконце относительно Fc-области и/или на карбокси-конце относительно CD3-связывающей области полиспецифической полипептидной конструкции.
[0009] В настоящем описании предложена полиспецифическая полипептидная конструкция, которая содержит по порядку от N-конца к C-концу: первый антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA); Fc-область иммуноглобулина; линкер; CD3-связывающую область, которая связывается с CD3 (CD3ε и второй антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA). Также предложена полиспецифическая полипептидная конструкция, которая содержит по порядку от N-конца к C-концу: Fc-область иммуноглобулина; линкер; CD3-связывающую область, которая связывается с CD3 (CD3ε и антигенсвязывающий домен, который связывается с опухолесипецифическим антигеном (TAA). Предложена полиспецифическая полипептидная конструкция, которая содержит по порядку от N-конца к C-концу: антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA); Fc-область иммуноглобулина; линкер; и CD3-связывающую область, которая связывается с CD3 (CD3ε).
[0010] Среди вариантов осуществления настоящего раскрытия находятся полиспецифические полипептидные конструкции, которые связываются по меньшей мере с CD3 и со вторым антигеном, таким как опухолеспецифический антиген (TAA). Полиспецифические полипептидные конструкции, представленные в настоящем документе, включают по меньшей мере первый компонент, который включает одну или более копий антигенсвязывающего домена, который связывается с антигеном, связанным с Fc-областью иммуноглобулина, второй компонент, который включает одну или более копий, по меньшей мере, связывающего домена, который связывается с CD3 (называемый здесь как анти-CD3-связывающий домен или CD3-связывающая область, которые используются в настоящем описании взаимозаменяемо), и линкер, такой как расщепляемый линкер, который соединяет первый компонент и второй компонент.
[0011] Расположение Fc-области на N-конце относительно CD3-связывающей области уменьшает или предотвращает способность CD3-связывающей области связываться с CD3. В некоторых вариантах осуществления в нерасщепленном/неактивном состоянии первый компонент (компонент #1) и второй компонент (компонент #2) полиспецифических полипептидных конструкций связаны, и связывание с CD3 невозможно, если антигенсвязывающий домен(ы) не связан со своим родственным антигеном. Это является предпочтительным, поскольку предотвращает системное связывание CD3-связывающей области с Т-клетками и фокусирует ее на сайте экспрессии антигена. Это выгодно, поскольку устраняет основной связывающий карман периферических Т-клеток, обеспечивая более благоприятное распределение и локализацию в месте экспрессии антигена, например, в опухолевых клетках или в микроокружении опухоли. В некоторых случаях связывание и/или рекрутирование CD3 умножается или усиливается путем включения расщепляемого линкера, соединяющего компонент #1 и компонент #2, в котором после расщепления расщепляемого линкера, такого как протеолиз, увеличивается связывание в результате доступности CD3-связывающей области.
[0012] В неактивном, т.е. нерасщепленном состоянии, компонент #1 и компонент #2 полиспецифических полипептидных конструкций функционально связаны и не связываются или иным образом не рекрутируют CD3 кроме случаев, когда антигенсвязывающий домен(ы) связан с его родственным антигеном. В некоторых вариантах осуществления настоящего изобретения нерасщепленные полиспецифические полипептидные конструкции способны взаимодействовать с FcγR и опосредовать врожденные иммунные эффекторные функций, например, антителозависимую клеточную цитотоксичность (ADCC) и антителозависимый клеточный фагоцитоз (ADCP). В некоторых вариантах осуществления нерасщепленные полиспецифические полипептидные конструкции способны взаимодействовать с белками комплемента, а именно C1q, и опосредовать зависимую от комплемента цитотоксичность.
[0013] Полиспецифические полипептидные конструкции по настоящему изобретению, как правило, имеют более чем один антигенсвязывающий домен(домены). В предложенных аспектах, в которых полиспецифические полипептидные конструкции содержат расщепляемый линкер, после того как линкер, соединяющий первый и второй компоненты, расщепляется, например, протеазой, каждый компонент сохраняет по меньшей мере один антигенсвязывающий домен. Первый компонент (то есть компонент #1) содержит, по меньшей мере, Fc-область и антигенсвязывающий домен. Второй компонент (то есть компонент #2) содержит, по меньшей мере, анти-CD3-связывающий домен и антигенсвязывающий домен.
[0014] Расщепление, например, путем протеолиза, внутри расщепляемого линкера физически разделяет компонент #1 и компонент #2, каждый из которых имеет терапевтическое применение, хотя и зависит от разных эффекторных клеток. Компонент #1 содержит по меньшей мере один антигенсвязывающий домен и Fc-область. В некоторых вариантах осуществления компонент #1 способен вызывать эффекторные функции врожденного иммунитета, например ADCC, высвобождение цитокинов, дегрануляцию и/или фагоцитоз. Компонент #2 содержит, по меньшей мере, домен CD3-связывающей области, и антигенсвязывающий домен, первый из которых способен связываться с CD3 (при отделении от компонента #1). Компонент # 2 способен формировать иммунный синапс между антиген-экспрессирующей клеткой и Т-клеткой. Такое совместное взаимодействие обеспечивает антиген-зависимую активацию Т-клеток, цитотоксичность, высвобождение цитокинов, дегрануляцию и пролиферацию. В расщепленном/активированном состоянии компонент #2 не является функционально связанным с Fc-областью компонента #1, и, таким образом, компонент # 2 не взаимодействует с FcRn и имеет повышенный клиренс в сыворотке, если локализован на сайте без антиген-экспрессирующей клетки. Это является предпочтительным, поскольку ограничивает системное воздействие активированного анти-CD3-связывающего домена и фокусируется непосредственно в антиген-экспрессирующих тканях, например в опухолевых клетках или в микроокружении опухоли.
[0015] В некоторых вариантах осуществления полиспецифический полипептид находится в неактивном состоянии, то есть нерасщепленном состоянии, и связывание CD3-связывающей области с CD3 ингибируется или по существу снижается, когда полиспецифическая полипептидная конструкция находится в нерасщепленном состоянии по сравнению с расщепленным состоянием. В некоторых вариантах осуществления полиспецифический полипептид находится в активированном состоянии, и первый и второй компоненты не являются функционально связанными. В некоторых вариантах осуществления полиспецифический полипептид находится в активированном состоянии, то есть в расщепленном состоянии, и второй компонент связывается с эпсилон-цепью CD3 (CD3ε) и опухолеспецифическим антигеном (TAA).
[0016] В некоторых аспектах антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов выбран из антитела или антигенсвязывающего фрагмента, родственного партнера по связыванию, Антикалина (модифицированного липокалина), Дарпина, Финомера, Центрина (сконструированный домен фибронетикина III), домена с цистиновым узлом, Аффилина, Аффитела или сконструированного домена CH3. В некоторых вариантах осуществления природный родственный партнер по связыванию содержит внеклеточный домен или его связывающий фрагмент нативного родственного партнера по связыванию TAA или его вариант, который проявляет активность связывания с TAA.
[0017] В некоторых аспектах антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов содержит внеклеточный домен или связывающий фрагмент нативного родственного партнера по связыванию TAA или его вариант, который проявляет активность связывания с TAA.
[0018] В некоторых вариантах осуществления первый компонент включает одну или более копий антигенсвязывающего домена. В некоторых вариантах осуществления первый компонент содержит по меньшей мере два антигенсвязывающих домена, как например, два антигенсвязывающих домена. В некоторых вариантах осуществления по меньшей мере два антигенсвязывающих домена первого компонента связываются с одним и тем же TAA. В некоторых случаях, по меньшей мере, два антигенсвязывающих домена первого компонента связываются с другим эпитопом того же TAA. В некоторых случаях, по меньшей мере, два антигенсвязывающих домена первого компонента связываются с другим эпитопом того же TAA. В некоторых вариантах осуществления по меньшей мере два антигенсвязывающих домена первого компонента связываются с другим TAA.
[0019] В некоторых вариантах осуществления антигенсвязывающий домен первого компонента, который в некоторых случаях является первым антигенсвязывающим доменом, включает одну или более копий антитела или его антигенсвязывающего фрагмента. В некоторых вариантах осуществления антигенсвязывающий домен первого компонента, такого как первый антигенсвязывающий домен, включает одну или более копий антитела или его антигенсвязывающего фрагмента, выбранного из группы, состоящей из Fab-фрагмента, F(ab')2-фрагмента, Fv-фрагмента, scFv, scAb, dAb, однодоменного антитела с тяжелой цепью и однодоменного антитела с легкой цепью. В некоторых вариантах осуществления первый антигенсвязывающий домен включает одну или более копий одного или более фрагментов однодоменного антитела (sdAb), например, VHH, VNAR, сконструированных доменов VH или VK. VHH могут быть получены только из антител верблюжьей тяжелой цепи. VNAR могут быть получены только из антител тяжелой цепи хрящевой рыбы. Различные способы были реализованы для генерации мономерных sdAb из традиционно гетеродимерных доменов VH и VK, включая конструирование интерфейса и выбор определенных семейств зародышевой линии.
[0020] В некоторых вариантах осуществления антигенсвязывающий домен первого компонента, такой как первый антигенсвязывающий домен, связывается с антигеном, таким как опухолеспецифический антиген (TAA). В некоторых вариантах осуществления TAA выбран из группы, состоящей из 1-92-LFA-3, 5T4, альфа-4 интегрина, альфа-V интегрина, альфа4бета1 интегрина, альфа4бета7 интегрина, AGR2, анти-Lewis-Y, рецептора апелина J, APRIL, B7-H3, B7-H4, BAFF, BTLA, компонента комплемента C5, C-242, CA9, CA19-9, (Lewis а), карбоангидразы 9, CD2, CD3, CD6, CD9, CD11a, CD19, CD20, CD22, CD24, CD25, CD27, CD28, CD30, CD33, CD38, CD40, CD40L, CD41, CD44, CD44v6, CD47, CD51, CD52, CD56, CD64, CD70, CD71, CD74, CD80, CD81, CD86, CD95, CD117, CD123, CD125, CD132, (IL-2RG), CD133, CD137, CD138, CD166, CD172A, CD248, CDH6, CEACAM5 (CEA), CEACAM6 (NCA-90), CLAUDIN-3, CLAUDIN-4, cMet, коллагена, Cripto, CSFR, CSFR-1, CTLA-4, CTGF, CXCL10, CXCL13, CXCR1, CXCR2, CXCR4, CYR61, DL44, DLK1, DLL3, DLL4, DPP-4, DSG1, EDA, EDB, EGFR, EGFRviii, рецептора эндотелина B (ETBR), ENPP3, EpCAM, EPHA2, EPHB2, ERBB3, F-протеина RSV, FAP, FGF-2, FGF8, FGFR1, FGFR2, FGFR3, FGFR4, FLT-3, рецептора альфа фолата (FRα), GAL3ST1, G-CSF, G-CSFR, GD2, GITR, GLUT1, GLUT4, GM-CSF, GM-CSFR, рецепторов GP IIb/IIIa, Gp130, GPIIB/IIIA, GPNMB, GRP78, HER2/neu, HER3, HER4, HGF, hGH, HVEM, гиалуронидазы, ICOS, IFNальфа, IFNбета, IFNгамма, IgE, рецептора IgE (FceRI), IGF, IGF1R, IL1B, IL1R, IL2, IL11, IL12, IL12p40, IL-12R, IL-12Rbeta1, IL13, IL13R, IL15, IL17, IL18, IL21, IL23, IL23R, IL27/IL27R (wsx1), IL29, IL-31R, IL31/IL31R, IL2R, IL4, IL4R, IL6, IL6R, рецептора инсулина, Jagged лигандов, Jagged 1, Jagged 2, KISS1-R, LAG-3, LIF-R, Lewis X, LIGHT, LRP4, LRRC26, Ly6G6D, LyPD1, MCSP, мезотелина, MRP4, MUC1, муцина-16 (MUC16, CA-125), Na/K-АТФаза, NGF, никастрина, рецепторов Notch, Notch 1, Notch 2, Notch 3, Notch 4, NOV, OSM-R, OX-40, PAR2, PDGF-AA, PDGF-BB, PDGFRальфа, PDGFRбета, PD-1, PD-L1, PD-L2, фосфатидилсерина, P1GF, PSCA, PSMA, PSGR, RAAG12, RAGE, SLC44A4, фосфат сфингозина 1, STEAP1, STEAP2, TAG-72, TAPA1, TE M-8, TGFбета, TIGIT, TIM-3, TLR2, TLR4, TLR6, TLR7, TLR8, TLR9, TMEM31, TNFальфа, TNFR, TNFRS12A, TRAIL-R1, TRAIL-R2, трансферрина, рецептора трансферрина, TRK-A, TRK -B, uPAR, VAP1, VCAM-1, VEGF, VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGFR1, VEGFR2, VEGFR3, VISTA, WISP-1, WISP-2 и WISP-3.
[0021] В некоторых вариантах осуществления Fc-область представляет собой гомодимерную Fc-область. В некоторых вариантах осуществления Fc-область представляет собой гетеродимерную Fc-область.
[0022] В некоторых вариантах осуществления Fc-область иммуноглобулина первого компонента представляет собой изотип IgG, выбранный из группы, состоящей из изотипа IgG1, изотипа IgG2, изотипа IgG3 и подкласса IgG4. В некоторых примерах Fc-область представляет собой Fc-область человеческого IgG1, человеческого IgG2, человеческого IgG3 или человеческого IgG4 или представляет собой их иммунологически активный фрагмент. В некоторых вариантах осуществления Fc-область включает полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 1, или последовательность аминокислот, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% идентичности последовательности с SEQ ID NO: 1. В некоторых случаях Fc-область включает полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 2, или последовательность аминокислот, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% идентичности последовательности с SEQ ID NO: 2. В некоторых из любых таких вариантов осуществления Fc-область включает полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 4, или последовательность аминокислот, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94% 95%, 96%, 97%, 98%, 99% идентичности последовательности с SEQ ID NO: 4. В некоторых примерах Fc-область включает полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 5, или последовательность аминокислот, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% идентичности последовательности с SEQ ID NO: 5.
[0023] В некоторых вариантах осуществления Fc-область иммуноглобулина представляет собой полипептид, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-6. В некоторых вариантах осуществления Fc-область иммуноглобулина представляет собой полипептид, содержащий аминокислотную последовательность, которая имеет, по меньшей мере, 50%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1-6.
[0024] В некоторых вариантах осуществления Fc-область иммуноглобулина представляет собой полипептид, содержащий аминокислотную последовательность, которая получена из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-6. В некоторых вариантах осуществления Fc-область иммуноглобулина представляет собой полипептид, содержащий аминокислотную последовательность, которая получена из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-6, включая одну или более модификации. В некоторых вариантах осуществления Fc-область иммуноглобулина представляет собой полипептид, содержащий аминокислотную последовательность, которая получена из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-6, включающей одну или модификации для предотвращения гликозилирования, для изменения взаимодействия Fc-рецептора, для уменьшения связывания с Fc-рецептором, для усиления взаимодействия с CD32A, для снижения связывания белка C1q комплемента, для увеличения периода полужизни, для усиления связывания с FcRn, для изменения антителозависимой клеточной цитотоксичности (ADCC) и/или комплементзависимой цитотоксичности (CDC), чтобы вызвать гетеродимеризацию, предотвратить димеризацию, стабилизировать гомодимеризацию в интерфейсе CH3:CH3 и их комбинации.
[0025] В некоторых вариантах осуществления Fc представляет собой гетеродимерную Fc-область. В некоторых случаях один или оба полипептида Fc гетеродимерной Fc-области включают, по меньшей мере, одну модификацию для индукции гетеродимеризации по сравнению с полипептидом гомодимерной Fc-области, необязательно по сравнению с полипептидом Fc, представленным в SEQ ID NO: 1, или его иммунологически активным фрагментом. В некоторых вариантах осуществления каждый из полипептидов Fc гетеродимерной Fc-области независимо содержит по меньшей мере одну аминокислотную модификацию. В некоторых случаях каждый из полипептидов Fc гетеродимерной Fc-области содержит модификацию «выступ-во-впадину» или содержит мутацию заряда для увеличения электростатической комплементарности полипептидов. В некоторых примерах модификация аминокислоты представляет собой модификацию «выступ-во-впадину».
[0026] В некоторых вариантах осуществления первый полипептид Fc гетеродимерной Fc-области содержит модификацию, выбранную из Thr366Ser, Leu368Ala, Tyr407Val и их комбинаций, и второй полипептид Fc гетеродимерной Fc-области содержит модификацию T366W, и где необязательно первый и второй полипептиды Fc дополнительно содержат модификацию нецистеинового остатка до остатка цистеина, где модификация первого полипептида находится в одном из положений Ser354 и Y349, а модификация второго полипептида Fc находится в другом положении Ser354 и Y349.
[0027] В некоторых примерах аминокислотная модификация является мутацией заряда для увеличения электростатической комплементарности полипептидов. В некоторых вариантах осуществления первый и/или второй полипептиды Fc содержат модификацию в комплементарных положениях, где модификация представляет собой замену аминокислотой, имеющей заряд, противоположный комплементарной аминокислоте другого полипептида. В некоторых вариантах осуществления первый или второй полипептид содержат модификацию в комплементарных положениях, где модификация представляет собой замену аминокислотой, имеющей заряд, противоположный комплементарной аминокислоте другого полипептида. В некоторых вариантах осуществления, по меньшей мере, первый или второй полипептиды Fc, каждый, содержат модификацию в комплементарном положении, где модификация представляет собой замену аминокислотой, имеющей заряд, противоположный комплементарной аминокислоте другого полипептида. В некоторых вариантах осуществления каждый первый и второй полипептиды Fc содержат модификацию в комплементарных положениях, где модификация представляет собой замену аминокислотой, имеющей заряд, противоположный комплементарной аминокислоте другого полипептида.
[0028] В некоторых вариантах осуществления один из первого или второго полипептида Fc гетеродимерной Fc-области дополнительно содержит модификацию по остатку Ile253. В некоторых случаях модификация представляет собой Ile253Arg. В некоторых вариантах осуществления один из первого или второго полипептида Fc гетеродимерной Fc-области дополнительно содержит модификацию по остатку His435. В некоторых случаях модификация представляет собой His435Arg. В некоторых вариантах осуществления Fc-область содержит полипептид, в котором отсутствует Lys447.
[0029] В некоторых вариантах осуществления модификации в Fc-области уменьшают связывание с Fc-гамма-рецептором, в то же время оказывая минимальное влияние на связывание с неонатальным Fc-рецептором (FcRn). В некоторых вариантах осуществления мутированный или модифицированный полипептид Fc включает следующие мутации: Met252Tyr и Met428Leu или Met252Tyr и Met428Val (M252Y, M428L или M252Y, M428V) с использованием системы нумерации Kabat.
[0030] В некоторых вариантах осуществления Fc-область содержит полипептид, содержащий по меньшей мере одну модификацию для усиления связывания FcRn. В некоторых примерах модификация находится в положении, выбранном из группы, состоящей из Met252, Ser254, Thr256, Met428, Asn434 и их комбинаций. В некоторых случаях модификация находится в положении, выбранном из группы, состоящей из Met252Y, Ser254T, Thr256E, Met428L, Met428V, Asn434S и их комбинаций. В некоторых конкретных вариантах осуществления модификация находится в положении Met252 и в положении Met428. В некоторых случаях модификация представляет собой Met252Y и Met428L. В некоторых случаях модификация представляет собой Met252Y и Met428V.
[0031] В некоторых вариантах осуществления первый полипептид гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 82, 86, 94 или 96, и второй полипептид гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 83, 87, 90, 92, 98 или 100.
[0032] В некоторых вариантах осуществления Fc-область содержит полипептид, содержащий по меньшей мере одну аминокислотную модификацию, которая снижает эффекторную функцию и/или уменьшает связывание с эффекторной молекулой, выбранной из Fc-гамма-рецептора или C1q. В некоторых примерах одна или более модификаций аминокислот представляют собой делецию одного или более из Glu233, Leu234 или Leu235. В некоторых аспектах первый полипептид гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 84, 88, 95 или 97, и второй полипептид гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 85, 89, 91, 93, 99 или 101.
[0033] В некоторых вариантах осуществления Fc-область содержит полипептид, содержащий по меньшей мере одну модификацию для усиления связывания FcγR. В некоторых случаях модификация представляет собой модификацию по Ser239 или Ile332. В некоторых вариантах осуществления гликозилирование Fc-области модифицировано для усиления FcγR связывания по сравнению с немодифицированной Fc-областью. В некоторых примерах Fc-область не содержит или имеет пониженное содержание фукозы.
[0034] В некоторых вариантах осуществления CD3-связывающая область представляет собой антитело против CD3 или антигенсвязывающий фрагмент. В некоторых вариантах осуществления антитело против CD3 или антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL). В некоторых из любых таких вариантов осуществления CD3-связывающая область является одновалентной.
[0035] В некоторых вариантах осуществления антитело против CD3 или антигенсвязывающий фрагмент не являются одноцепочечным антителом, необязательно, не являются одноцепочечным вариабельным фрагментом (scFv). В некоторых вариантах осуществления Fc представляет собой гетеродимерную Fc-область, и VH и VL, которые содержатся в антителе против CD3 или антигенсвязывающем фрагменте, связаны с противоположными полипептидами гетеродимерной Fc-области. В некоторых вариантах осуществления CD3-связывающая область не способна или по существу не способна связывать или рекрутировать CD3, если по меньшей мере один из антигенсвязывающего домена не связан с его TAA. В некоторых аспектах CD3-связывающая область не способна или по существу не способна связывать или рекрутировать CD3, если по меньшей мере, два антигенсвязывающих домена не связаны с их TAA.
[0036] В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит линкер, который представляет собой полипептидный линкер. В некоторых вариантах осуществления линкер представляет собой полипептид длиной до 25 аминокислот. В некоторых случаях линкер представляет собой полипептид, содержащий или содержащий примерно от 2 до 24 аминокислот, от 2 до 20 аминокислот, от 2 до 18 аминокислот, от 2 до 14 аминокислот, от 2 до 12 аминокислот, от 2 до 10 аминокислот, от 2 до 8 аминокислот, от 2 до 6 аминокислот, от 6 до 24 аминокислот, от 6 до 20 аминокислот, от 6 до 18 аминокислот, от 6 до 14 аминокислот, от 6 до 12 аминокислот, от 6 до 10 аминокислот, от 6 до 8 аминокислот, от 8 до 24 аминокислот, от 8 до 20 аминокислот, от 8 до 18 аминокислот, от 8 до 14 аминокислот, от 8 до 12 аминокислот, от 8 до 10 аминокислот, от 10 до 24 аминокислот, от 10 до 20 аминокислот кислоты, от 10 до 18 аминокислот, от 10 до 14 аминокислот, от 10 до 12 аминокислот, от 12 до 24 аминокислот, от 12 до 20 аминокислот, от 12 до 18 аминокислот, от 12 до 14 аминокислот, от 14 до 24 аминокислот, от 14 до 20 аминокислот, от 14 до 18 аминокислот, от 18 до 24 аминокислот, от 18 до 20 аминокислот или от 20 до 24 аминокислот. В некоторых вариантах осуществления линкер представляет собой полипептид длиной 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот. В некоторых случаях линкер является расщепляемым линкером.
[0037] В некоторых вариантах осуществления первый антигенсвязывающий домен и полипептид Fc иммуноглобулина функционально связаны через аминокислотные линкеры. В некоторых вариантах осуществления эти внутрикомпонентные линкеры состоят преимущественно из аминокислот глицина и серина, обозначаемых здесь как GS-линкеры. GS-линкеры слитых белков по настоящему изобретению могут иметь различную длину, например 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 аминокислот в длину.
[0038] В некоторых вариантах осуществления GS-линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из GGSGGS, т.е. (GGS)2 (SEQ ID NO: 10); GGSGGSGGS, т.е. (GGS)3 (SEQ ID NO: 11); GGSGGSGGSGGS, т.е. (GGS)4 (SEQ ID NO: 12); и GGSGGSGGSGGSGGS, т.е. (GGS)5 (SEQ ID NO: 13).
[0039] В некоторых вариантах осуществления второй компонент также включает одну или более копий анти-CD3-связывающего домена. В некоторых вариантах осуществления анти-CD3-связывающий домен включает одну или более копий антитела или его антигенсвязывающего фрагмента. В некоторых вариантах осуществления анти-CD3-связывающий домен включает одну или более копий антитела или его антигенсвязывающего фрагмента, выбранного из группы, состоящей из Fab-фрагмента, F(ab')2-фрагмента, Fv-фрагмента, scFv, scAb, dAb, однодоменного антитела тяжелой цепи и однодоменного антитела легкой цепи. В некоторых вариантах осуществления анти-CD3-связывающий домен содержит Fv-фрагмент антитела, который связывает CD3ε (называемый здесь как анти-CD3ε Fv-фрагмент). В некоторых вариантах осуществления анти-CD3ε Fv-фрагмент антитела содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 32-81. В некоторых вариантах осуществления анти-CD3ε Fv-фрагмент антитела включает в себя аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32-81. В некоторых вариантах осуществления анти-CD3ε Fv-фрагмент антитела включает в себя комбинацию аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32-62, и аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 63-81. В некоторых вариантах осуществления анти-CD3ε Fv-фрагмент антитела включает в себя комбинацию аминокислотной последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32-62, и аминокислотной последовательности, которая, по меньшей мере, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из аминокислотной последовательности SEQ ID NO: 63-81.
[0040] В некоторых вариантах осуществления анти-CD3ε Fv-фрагмент антитела представляет собой стабилизированный дисульфидом анти-CD3-связывающий Fv-фрагмент (dsFv).
[0041] В некоторых вариантах осуществления второй компонент также включает одну или более копий антигенсвязывающего домена. В некоторых вариантах осуществления второй компонент содержит по меньшей мере два антигенсвязывающих домена, таких как два антигенсвязывающих домена. В некоторых вариантах осуществления по меньшей мере два антигенсвязывающих домена второго компонента связываются с одним и тем же TAA. В некоторых случаях, по меньшей мере, два антигенсвязывающих домена второго компонента связываются с другим эпитопом того же TAA. В некоторых случаях, по меньшей мере, два антигенсвязывающих домена второго компонента связываются с другим эпитопом того же TAA. В некоторых вариантах осуществления по меньшей мере два антигенсвязывающих домена второго компонента связываются с другим TAA.
[0042] В некоторых вариантах осуществления первый компонент содержит первый антигенсвязывающий домен, а антигенсвязывающий домен второго компонента представляет собой второй антигенсвязывающий домен. В некоторых вариантах осуществления второй антигенсвязывающий домен второго компонента связывается с тем же антигеном, что и первый антигенсвязывающий домен первого компонента. В некоторых вариантах осуществления второй антигенсвязывающий домен второго компонента связывается с другим эпитопом на том же антигене, что и первый антигенсвязывающий домен первого компонента. В некоторых вариантах осуществления второй антигенсвязывающий домен второго компонента связывается с эпитопом на том же антигене, что и первый антигенсвязывающий домен первого компонента.
[0043] В некоторых вариантах осуществления антигенсвязывающий домен второго компонента, такой как второй антигенсвязывающий домен, включает одну или более копий антитела или его антигенсвязывающего фрагмента. В некоторых вариантах осуществления второй антигенсвязывающий домен включает одну или более копий антитела или его антигенсвязывающего фрагмента, выбранного из группы, состоящей из Fab-фрагмента, F(ab')2-фрагмента,Fv-фрагмента, scFv, scAb, dAb, однодоменного антитела тяжелой цепи и однодоменного антитела легкой цепи. В некоторых вариантах осуществления второй антигенсвязывающий домен включает одну или более копий одного или более фрагментов однодоменного антитела (sdAb), например VHH, VNAR, сконструированных доменов VH или VK. VHH могут быть получены только из антител верблюжьей тяжелой цепи. VNAR могут быть получены только из антител тяжелой цепи хрящевой рыбы. Различные способы были реализованы для генерации мономерных sdAb из традиционно гетеродимерных доменов VH и VK, включая конструирование интерфейса и выбор определенных семейств зародышевой линии.
[0044] В некоторых вариантах осуществления антигенсвязывающий домен второго компонента, такой как второй антигенсвязывающий домен, связывается с антигеном, таким как опухолеспецифический антиген (TAA). В некоторых вариантах осуществления TAA выбран из группы, состоящей из 1-92-LFA-3, 5T4, альфа-4 интегрина, альфа-V интегрина, альфа4бета1 интегрина, альфа4бета7 интегрина, AGR2, анти-Lewis-Y, рецептора апелина J, APRIL, B7-H3, B7-H4, BAFF, BTLA, компонента комплемента C5, C-242, CA9, CA19-9, (Lewis а), карбоангидразы 9, CD2, CD3, CD6, CD9, CD11a, CD19, CD20, CD22, CD24, CD25, CD27, CD28, CD30, CD33, CD38, CD40, CD40L, CD41, CD44, CD44v6, CD47, CD51, CD52, CD56, CD64, CD70, CD71, CD74, CD80, CD81, CD86, CD95, CD117, CD123, CD125, CD132, (IL-2RG), CD133, CD137, CD138, CD166, CD172A, CD248, CDH6, CEACAM5 (CEA), CEACAM6 (NCA-90), CLAUDIN-3, CLAUDIN-4, cMet, коллагена, Cripto, CSFR, CSFR-1, CTLA-4, CTGF, CXCL10, CXCL13, CXCR1, CXCR2, CXCR4, CYR61, DL44, DLK1, DLL3, DLL4, DPP-4, DSG1, EDA, EDB, EGFR, EGFRviii, рецептора эндотелина B (ETBR), ENPP3, EpCAM, EPHA2, EPHB2, ERBB3, F-протеина RSV, FAP, FGF-2, FGF8, FGFR1, FGFR2, FGFR3, FGFR4, FLT-3, рецептора фолата альфа (FRα), GAL3ST1, G-CSF, G-CSFR, GD2, GITR, GLUT1, GLUT4, GM-CSF, GM-CSFR, рецепторов GP IIb/IIIa, Gp130, GPIIB/IIIA, GPNMB, GRP78, HER2/neu, HER3, HER4, HGF, hGH, HVEM, гиалуронидазы, ICOS, IFNальфа, IFNбета, IFNгамма, IgE, рецептора IgE (FceRI), IGF, IGF1R, IL1B, IL1R, IL2, IL11, IL12, IL12p40, IL-12R, IL-12Rbeta1, IL13, IL13R, IL15, IL17, IL18, IL21, IL23, IL23R, IL27/IL27R (wsx1), IL29, IL-31R, IL31/IL31R, IL2R, IL4, IL4R, IL6, IL6R, рецептора инсулина, Jagged лигандов, Jagged 1, Jagged 2, KISS1-R, LAG-3, LIF-R, Lewis X, LIGHT, LRP4, LRRC26, Ly6G6D, LyPD1, MCSP, мезотелина, MRP4, MUC1, муцина-16 (MUC16, CA-125), Na/K-АТФаза, NGF, никастрина, рецепторов Notch, Notch 1, Notch 2, Notch 3, Notch 4, NOV, OSM-R, OX-40, PAR2, PDGF-AA, PDGF-BB, PDGFRальфа, PDGFRбета, PD-1, PD-L1, PD-L2, фосфатидилсерина, P1GF, PSCA, PSMA, PSGR, RAAG12, RAGE, SLC44A4, фосфат сфингозина 1, STEAP1, STEAP2, TAG-72, TAPA1, TE M-8, TGFбета, TIGIT, TIM-3, TLR2, TLR4, TLR6, TLR7, TLR8, TLR9, TMEM31, TNFальфа, TNFR, TNFRS12A, TRAIL-R1, TRAIL-R2, трансферрина, рецептора трансферрина, TRK-A, TRK -B, uPAR, VAP1, VCAM-1, VEGF, VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGFR1, VEGFR2, VEGFR3, VISTA, WISP-1, WISP-2 и WISP-3.
[0045] В некоторых вариантах осуществления антигенсвязывающий домен второго компонента, такой как второй антигенсвязывающий домен, и анти-CD3-связывающий домен функционально связаны через аминокислотные линкеры. В некоторых вариантах осуществления эти внутрикомпонентные линкеры состоят преимущественно из аминокислот глицина и серина, обозначаемых здесь как GS-линкеры. GS-линкеры слитых белков по настоящему изобретению могут иметь различную длину, например 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 аминокислот в длину.
[0046] В некоторых вариантах осуществления GS-линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из GGSGGS, т.е. (GGS)2 (SEQ ID NO: 10); GGSGGSGGS, т.е. (GGS)3 (SEQ ID NO: 11); GGSGGSGGSGGS, т.е. (GGS)4 (SEQ ID NO: 12); и GGSGGSGGSGGSGGS, т.е. (GGS)5 (SEQ ID NO: 13).
[0047] В настоящем описании предложена полиспецифическая полипептидная конструкция, причем полиспецифическая полипептидная конструкция содержит первый компонент, содержащий гетеродимерную Fc-область, и второй компонент, содержащий антитело против CD3 или антигенсвязывающий фрагмент, содержащий вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где: VH и VL, которые содержит антитело против CD3 или антигенсвязывающий фрагмент, связаны с противоположными полипептидами гетеродимерной Fc; первый и второй компоненты связаны расщепляемым линкером, где гетеродимерная Fc-область расположена на N-конце относительно антитела против CD3; и один или оба из первого и второго компонентов содержат антигенсвязывающий домен, который связывается с опухолесипецифическим антигеном (TAA).
[0048] В некоторых вариантах осуществления связывание CD3-связывающей области с CD3 существенно снижается, когда полиспецифическая полипептидная конструкция находится в нерасщепленном состоянии по сравнению с расщепленным состоянием. В некоторых вариантах осуществления в расщепленном состоянии первый и второй компоненты не связаны.
[0049] В некоторых вариантах осуществления расщепляемый линкер представляет собой полипептид. В некоторых вариантах осуществления расщепляемый линкер представляет собой полипептид, который является субстратом для протеазы. В некоторых вариантах осуществления протеаза продуцируется иммунной эффекторной клеткой, опухолью или клетками, присутствующими в микроокружении опухоли. В некоторых вариантах осуществления протеаза продуцируется опухолью, которая находится вблизи клеток, которые экспрессируют CD3ε, и/или продуцируется опухолью, которая локализована совместно с клетками, которые экспрессируют CD3ε в ткани, и где протеаза расщепляет расщепляемый линкер в полиспецифической полипептидной конструкции, когда полиспецифическая полипептидная конструкция подвергается воздействию протеазы. В некоторых вариантах осуществления протеаза продуцируется опухолью, которая находится вблизи клеток, которые экспрессируют один или более опухолеспецифических антигенов (TAA), и/или продуцируется опухолью, которая локализована совместно с клетками, которые экспрессирут TAA-мишень в ткани, и где протеаза расщепляет расщепляемый линкер в полиспецифической полипептидной конструкции, когда полиспецифическая полипептидная конструкция подвергается воздействию протеазы. В некоторых вариантах осуществления протеаза продуцируется иммунной эффекторной клеткой. В некоторых вариантах осуществления протеаза продуцируется иммунной эффекторной клеткой, которая находится вблизи клеток, которые экспрессируют TAA. В некоторых примерах протеаза продуцируется иммунной эффекторной клеткой, а иммунная эффекторная клетка представляет собой активированную Т-клетку, клетку-натуральный киллер (NK) или Т-клетку NK. В некоторых вариантах осуществления протеаза расщепляет расщепляемый линкер в полиспецифической полипептидной конструкции, когда полиспецифическая полипептидная конструкция подвергается воздействию протеазы. В некоторых вариантах осуществления протеаза продуцируется иммунной эффекторной клеткой, которая находится вблизи с клетками, которые экспрессируют TAA, и где протеаза расщепляет расщепляемый линкер в полиспецифической полипептидной конструкции, когда полиспецифическая полипептидная конструкция подвергается воздействию протеазы.
[0050] В некоторых вариантах осуществления расщепляемый линкер представляет собой полипептид длиной до 50 аминокислот. В некоторых вариантах осуществления расщепляемый линкер представляет собой полипептид длиной до 25 аминокислот. В некоторых вариантах осуществления расщепляемый линкер представляет собой полипептид длиной до 15 аминокислот.
[0051] В некоторых вариантах осуществления расщепляемый линкер представляет собой субстрат для протеазы, выбранной из протеаз, описанных в настоящем документе. В некоторых вариантах осуществления расщепляемый линкер представляет собой субстрат для протеазы, выбранной из группы, состоящей из uPA, легумаина, матриптазы (также называемой в настоящем документе MT-SP1 или MTSP1), ADAM17, BMP-1, TMPRSS3, TMPRSS4, MMP-9, ММР-12, ММР-13, ММР-14 и любой их комбинации. В некоторых вариантах осуществления расщепляемый линкер представляет собой субстрат для протеазы, выбранной из группы, состоящей из uPA, легумаина и матриптазы. В некоторых вариантах осуществления протеаза выбрана из матриптазы, матриксной металлопротеазы (ММР), гранзима B и их комбинаций.
[0052] В некоторых вариантах осуществления протеаза представляет собой гранзим B. В некоторых примерах расщепляемый линкер содержит аминокислотную последовательность общей формулы P4 P3 P2 P1 ↓ P1' (SEQ ID NO: 150), где P4 представляет собой аминокислоту I, L, Y, M, F, V или A; P3 представляет собой аминокислоту A, G, S, V, E, D, Q, N или Y; P2 представляет собой аминокислоту H, P, A, V, G, S или T; P1 представляет собой аминокислоту D или E; и P1' представляет собой аминокислоту I, L, Y, M, F, V, T, S, G или A. В некоторых вариантах осуществления расщепляемый линкер содержит аминокислотную последовательность общей формулы P4 P3 P2 P1 ↓ P1' (SEQ ID NO: 151), где P4 представляет собой аминокислоту I или L; P3 представляет собой аминокислоту E; P2 представляет собой аминокислоту P или A; P1 представляет собой аминокислоту D; и P1' представляет собой аминокислоту I, V, T, S или G. В некоторых примерах расщепляемый линкер содержит аминокислотную последовательность IEPDI (SEQ ID NO: 136), LEPDG (SEQ ID NO: 152, LEADT (SEQ ID) NO: 137), IEPDG (SEQ ID NO: 138), IEPDV (SEQ ID NO: 139), IEPDS (SEQ ID NO: 140), IEPDT (SEQ ID NO: 141) или LEADG (SEQ ID NO: 153). В некоторых случаях расщепляемый линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 22, 105-112, 136-141, 148, 150-153.
[0053] В некоторых вариантах осуществления протеаза представляет собой матриптазу. В некоторых случаях расщепляемый линкер содержит последовательность P1QAR↓ (A/V) (SEQ ID NO: 154), где P1 представляет собой любую аминокислоту; или расщепляемый линкер содержит последовательность RQAR (A/V) (SEQ ID NO: 155). В некоторых примерах расщепляемый линкер содержит последовательность RQARV (SEQ ID NO: 156). В некоторых случаях расщепляемый линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 23, 154-156.
[0054] В некоторых вариантах осуществления протеаза представляет собой ММР. В некоторых примерах MMP представляет собой MMP-2. В некоторых вариантах осуществления расщепляемый линкер содержит общую формулу P3 P2 P1 ↓ P1' (SEQ ID NO: 157), где P3 представляет собой P, V или A; P2 представляет собой Q или D; P1 представляет собой A или N; и P1' представляет собой L, I или M. В некоторых случаях расщепляемый линкер содержит общую формулу P3 P2 P1 ↓ P1' (SEQ ID NO: 158), где представляет собой P; P2 представляет собой Q или D; P1 представляет собой A или N; и P1' представляет собой L или I. В некоторых вариантах осуществления расщепляемый линкер содержит последовательность PAGL (SEQ ID NO: 24). В некоторых вариантах осуществления расщепляемый линкер представляет собой субстрат для матричной металлопротеазы (ММР).
[0055] В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит, по меньшей мере, (i) первый полипептид, содержащий первый полипептид Fc гетеродимерной Fc-области, линкер и домен VH антитела против CD3 или антигенсвязывающего фрагмента; и (ii) второй полипептид, содержащий второй полипептид Fc гетеродимерной Fc-области, линкер и домен VL антитела против CD3 или антигенсвязывающего фрагмента, где один или оба из первого и второго полипептида содержат, по меньшей мере, один антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA). В некоторых случаях только один из первого или второго полипептида содержит по меньшей мере один антигенсвязывающий домен, который связывается с TAA.
[0056] В некоторых из любых представленных вариантов осуществления антигенсвязывающий домен(ы) приводит к моновалентному, двухвалентному, трехвалентному или четырехвалентному связыванию с TAA. В некоторых вариантах осуществления один или более антигенсвязывающих доменов, которые независимо связываются с TAA, выбирают из sdAb, scFv или Fab. В некоторых вариантах осуществления один или более антигенсвязывающих доменов, которые связываются с TAA, представляет собой одноцепочечную молекулу, такую как фрагмент одноцепочечного антитела, содержащий VH и VL, например sdAb или scFv. В некоторых вариантах осуществления, по меньшей мере, один из антигенсвязывающих доменов представляет собой Fab, содержащий первую цепь, содержащую VH-CH1 (Fd), и вторую цепь, содержащую VL-CL.
[0057] В некоторых вариантах осуществления, по меньшей мере, один антигенсвязывающий домен расположен на аминоконце относительно Fc-области и/или расположен на карбоксиконце относительно CD3-связывающей области одного из первого или второго полипептида полиспецифической полипептидной конструкции. В некоторых случаях, по меньшей мере, один антигенсвязывающий домен расположен на аминоконце относительно Fc-области полиспецифической конструкции, а второй антигенсвязывающий домен расположен на карбоксиконце относительно CD3-связывающей области полиспецифической конструкции.
[0058] В некоторых вариантах осуществления, по меньшей мере, один из антигенсвязывающих доменов представляет собой Fab. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит: (i) первый полипептид, содержащий первый полипептид Fc гетеродимерной Fc-области, линкер и домен VH антитела против CD3 или антигенсвязывающего фрагмента; (ii) второй полипептид, содержащий второй полипептид Fc гетеродимерной Fc-области, линкер и домен VL антитела против CD3 или антигенсвязывающего фрагмента, и (iii) третий полипептид, содержащий VH-CH1 (Fd) или VL-CL Fab-фрагмента антитела, который связывается с опухолеспецифическим антигеном, где первый и/или второй полипептид дополнительно содержит другой из VH-CH1 (Fd) или VL-CL Fab-фрагмента антитела. В некоторых случаях только один из первого или второго полипептида содержит другой из VH-CH1 (Fd) или VL-CL Fab-фрагмента антитела. В некоторых вариантах осуществления оба, первый или второй полипептид, содержат другой из VH-CH1 (Fd) или VL-CL Fab-фрагмента антитела. В некоторых случаях другой из VH-CH1 (Fd) или VL-CL Fab-фрагмента антитела расположен на аминоконце относительно Fc-области и/или на карбоксиконце относительно CD3-связывающей области одного из первого или второго полипептида полиспецифической полипептидной конструкции. В некоторых вариантах осуществления другой VH-CH1 (Fd) или VL-CL Fab-фрагмента антитела расположен на аминоконце относительно Fc-области первого полипептида или второго полипептида и на карбоксиконце относительно CD3-связывающей области другого первого или второго полипептида.
[0059] В некоторых примерах антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов связывается с опухолевым антигеном, выбранным из 1-92-LFA-3, 5T4, альфа-4 интегрина, альфа-V интегрина, альфа4бета1 интегрина, альфа4бета7 интегрина, AGR2, анти-Lewis-Y, рецептора апелина J, APRIL, B7-H3, B7-H4, BAFF, BTLA, компонента комплемента C5, C-242, CA9, CA19-9, (Lewis а), карбоангидразы 9, CD2, CD3, CD6, CD9, CD11a, CD19, CD20, CD22, CD24, CD25, CD27, CD28, CD30, CD33, CD38, CD40, CD40L, CD41, CD44, CD44v6, CD47, CD51, CD52, CD56, CD64, CD70, CD71, CD74, CD80, CD81, CD86, CD95, CD117, CD123, CD125, CD132, (IL-2RG), CD133, CD137, CD138, CD166, CD172A, CD248, CDH6, CEACAM5 (CEA), CEACAM6 (NCA-90), CLAUDIN-3, CLAUDIN-4, cMet, коллагена, Cripto, CSFR, CSFR-1, CTLA-4, CTGF, CXCL10, CXCL13, CXCR1, CXCR2, CXCR4, CYR61, DL44, DLK1, DLL3, DLL4, DPP-4, DSG1, EDA, EDB, EGFR, EGFRviii, рецептора эндотелина B (ETBR), ENPP3, EpCAM, EPHA2, EPHB2, ERBB3, F-протеина RSV, FAP, FGF-2, FGF8, FGFR1, FGFR2, FGFR3, FGFR4, FLT-3, рецептора фолата альфа (FRα), GAL3ST1, G-CSF, G-CSFR, GD2, GITR, GLUT1, GLUT4, GM-CSF, GM-CSFR, рецепторов GP IIb/IIIa, Gp130, GPIIB/IIIA, GPNMB, GRP78, HER2/neu, HER3, HER4, HGF, hGH, HVEM, гиалуронидазы, ICOS, IFNальфа, IFNбета, IFNгамма, IgE, рецептора IgE (FceRI), IGF, IGF1R, IL1B, IL1R, IL2, IL11, IL12, IL12p40, IL-12R, IL-12Rbeta1, IL13, IL13R, IL15, IL17, IL18, IL21, IL23, IL23R, IL27/IL27R (wsx1), IL29, IL-31R, IL31/IL31R, IL2R, IL4, IL4R, IL6, IL6R, рецептора инсулина, Jagged лигандов, Jagged 1, Jagged 2, KISS1-R, LAG-3, LIF-R, Lewis X, LIGHT, LRP4, LRRC26, Ly6G6D, LyPD1, MCSP, мезотелина, MRP4, MUC1, муцина-16 (MUC16, CA-125), Na/K-АТФаза, NGF, никастрина, рецепторов Notch, Notch 1, Notch 2, Notch 3, Notch 4, NOV, OSM-R, OX-40, PAR2, PDGF-AA, PDGF-BB, PDGFRalpha, PDGFRбета, PD-1, PD-L1, PD-L2, фосфатидилсерина, P1GF, PSCA, PSMA, PSGR, RAAG12, RAGE, SLC44A4, фосфат сфингозина 1, STEAP1, STEAP2, TAG-72, TAPA1, TE M-8, TGFбета, TIGIT, TIM-3, TLR2, TLR4, TLR6, TLR7, TLR8, TLR9, TMEM31, TNFальфа, TNFR, TNFRS12A, TRAIL-R1, TRAIL-R2, трансферрина, рецептора трансферрина, TRK-A, TRK -B, uPAR, VAP1, VCAM-1, VEGF, VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGFR1, VEGFR2, VEGFR3, VISTA, WISP-1, WISP-2 и WISP-3.
[0060] В некоторых вариантах осуществления полиспецифический антигенсвязывающий домен содержит, по меньшей мере, первый антигенсвязывающий домен и второй антигенсвязывающий домен, где первый антигенсвязывающий домен и второй антигенсвязывающий домен связываются с одним и тем же TAA. В некоторых случаях первый антигенсвязывающий домен и второй антигенсвязывающий домен связываются с другим эпитопом того же TAA. В некоторых случаях первый антигенсвязывающий домен и второй антигенсвязывающий домен связываются с одним и тем же эпитопом того же TAA. В некоторых вариантах осуществления полиспецифический антигенсвязывающий домен содержит, по меньшей мере, первый антигенсвязывающий домен и второй антигенсвязывающий домен, где первый антигенсвязывающий домен и второй антигенсвязывающий домен связываются с разными TAA.
[0061] В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит первый связывающий пептид (LP1) между первым антигенсвязывающим доменом и полипептидной Fc-областью иммуноглобулина (Fc-область). В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит второй связывающий пептид (LP2) между анти-CD3-связывающим доменом (CD3-связывающей областью) и вторым антигенсвязывающим доменом. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит первый связывающий пептид (LP1) между первым антигенсвязывающим доменом и полипептидной Fc-областью иммуноглобулина (Fc-область) и второй связывающий пептид (LP2) между анти-CD3-связывающим доменом (CD3-связывающая область) и второй антигенсвязывающий домен.
[0062] В некоторых вариантах осуществления полиспецифическая полипептидная конструкция в нерасщепленном состоянии имеет структурное расположение от N-конца к C-концу следующим образом: первый антигенсвязывающий домен-LP1-линкерная область полипептида Fc-области иммуноглобулина (Fc-область)-линкер (такой как расщепляемый линкер)-анти-CD3-связывающий домен-LP2-второй антигенсвязывающий домен. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция в нерасщепленном состоянии имеет структурное расположение от N-конца к C-концу следующим образом: второй антигенсвязывающий домен-LP2-анти-CD3-связывающий домен (CD3-связывающая область)-линкер (такой как расщепляемый линкер)- линкерная область полипептида Fc иммуноглобулина - LP1 - первый антигенсвязывающий домен. В некоторых примерах линкер является расщепляемым линкером. В некоторых вариантах осуществления два связывающих пептида не являются идентичными друг другу. В некоторых случаях LP1 или LP2 независимо представляют собой пептид длиной от 1 до 20 аминокислот. В некоторых примерах LP1 или LP2 независимо включают пептид, который представляет собой или содержит любой линкер Gly-Ser, представленный в SEQ ID NO: 10-13, 119, 135, 147, 149.
[0063] В некоторых вариантах осуществления полиспецифическая конструкция представляет собой конструкцию, имеющую любое из структурных расположений, представленных на ФИГ. 1. В некоторых вариантах осуществления полиспецифическая конструкция представляет собой биспецифическую конструкцию, имеющую структурное расположение, показанное на ФИГ. 2. В некоторых вариантах осуществления биспецифическая конструкция имеет следующее структурное расположение от N-конца к C-конца. N-конец биспецифической конструкции включает в себя первый антигенсвязывающий домен, который связывается с опухолесипецифическим антигеном (TAA). Первый связывающий домен связывается с первым эпитопом на TAA- мишени. Соединенная с первым антигенсвязывающим доменом центральная полипептидная Fc-область иммуноглобулина, которая регулирует взаимодействия FcγR и/или взаимодействие FcRn. В некоторых вариантах осуществления центральная полипептидная Fc-область иммуноглобулина является гетеродимерной. Полипептидная Fc-область иммуноглобулина связана с расщепляемым линкером, который содержит один или более сайтов протеолитического расщепления, расположенных в С-концевом положении относительно полипептидной Fc-области иммуноглобулина. В некоторых вариантах осуществления один или более сайтов протеолитического расщепления представляет собой субстрат для матриптазы, матричной металлопротеазы (ММР) или гранзима В. Расщепляемый линкер присоединен к анти-CD3-связывающей последовательности, расположенной на С-конце Fc-области, и в некоторых случаях на дистальном конце второго компонента.
[0064] В некоторых вариантах осуществления антитело против CD3 или его антигенсвязывающий фрагмент представляет собой Fv-фрагмент антитела. В некоторых вариантах осуществления Fv-фрагмент антитела содержит стабилизированный дисульфидом анти-CD3-связывающий Fv-фрагмент (dsFv). В некоторых вариантах осуществления анти-CD3-связывающая последовательность представляет собой Fv-фрагмент антитела, который сконструирован так, чтобы включать дисульфидную связь между вариабельной областью тяжелой цепи (VH) и вариабельной областью легкой цепи (VL) с получением тем самым стабилизированного дисульфидом анти-CD3-связывающего Fv-фрагмента (dsFv). В некоторых вариантах осуществления домены VH и VL, которые содержит анти-CD3 Fv, функционально связаны с противоположными членами гетеродимерной Fc-области. В этих вариантах осуществления анти-CD3 Fv связывается с CD3 моновалентным образом. Анти-CD3 dsFv не связывается с CD3, когда расщепляемые линкеры интактны, т.е. находятся в нерасщепленном или неактивном состоянии. С-конец биспецифической конструкции включает второй антигенсвязывающий домен, который связывается с ТАА. В некоторых вариантах осуществления второй антигенсвязывающий домен связывается с тем же TAA, что и первый антигенсвязывающий домен, расположенный внутри первого компонента. В некоторых вариантах осуществления второй антигенсвязывающий домен связывается со вторым эпитопом на TAA, где второй эпитоп неконкурирует с первым эпитопом на TAA. В некоторых вариантах осуществления второй антигенсвязывающий домен связывается с TAA, отличным от первого антигенсвязывающего домена.
[0065] В некоторых вариантах осуществления каждый из первого антигенсвязывающего домена и второго антигенсвязывающего домена биспецифической конструкции включает одну или более копий антитела или его антигенсвязывающего фрагмента. В некоторых вариантах осуществления каждый из первого антигенсвязывающего домена и второго антигенсвязывающего домена биспецифической конструкции включает одну или более копий антитела или его антигенсвязывающего фрагмента, выбранного из группы, состоящей из Fab-фрагмента, F(ab')2-фрагмента, Fv-фрагмента, scFv, scAb, dAb, однодоменного антитела с тяжелой цепью и однодоменного антитела с легкой цепью. В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов представляет собой антитело или его антигенсвязывающий фрагмент, выбранный из группы, состоящей из Fab-фрагмента, F(ab')2-фрагмента, Fv-фрагмента, scFv, scAb, dAb, однодоменного антитела с тяжелой цепью и однодоменного антитела с легкой цепью. В некоторых вариантах осуществления каждый из первого антигенсвязывающего домена и второго антигенсвязывающего домена биспецифической конструкции включает одну или более копий одного или более фрагментов однодоменного антитела (sdAb), например VHH, VNAR, сконструированных доменов VH или VK. VHH могут быть получены из природных антител, содержащих только верблюжьи тяжелые цепи, из генетически модифицированных грызунов, которые продуцируют только антитела с тяжелой цепью, или из наивных/синтетических библиотек верблюжьих антител или гуманизированных верблюжьих однодоменных антител. VNAR могут быть получены только из антител с тяжелой цепью хрящевой рыбы. Различные способы были реализованы для генерации мономерных sdAb из традиционно гетеродимерных доменов VH и VK, включая конструирование интерфейса и выбор определенных семейств зародышевой линии.
[0066] В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент представляет собой sdAb. В некоторых случаях sdAb является человеческим или гуманизированным sdAb. В некоторых аспектах sdAb представляет собой VHH, VNAR, сконструированный домен VH или сконструированный домен VK. В некоторых примерах антитело или его антигенсвязывающий фрагмент представляет собой scFv. В некоторых случаях антитело или его антигенсвязывающий фрагмент представляет собой Fab.
[0067] В некоторых из любых представленных вариантов осуществления антитело против CD3 или его антигенсвязывающий фрагмент содержит VH CDR1, содержащую аминокислотную последовательность TYAMN (SEQ ID NO: 16); VH CDR2, содержащую аминокислотную последовательность RIRSKYNNYATYYADSVKD (SEQ ID NO: 17); VH CDR3, содержащую аминокислотную последовательность HGNFGNSYVSWFAY (SEQ ID NO: 18), VL CDR1, содержащую аминокислотную последовательность RSSTGAVTTSNYAN (SEQ ID NO: 19); VL CDR2, содержащую аминокислотную последовательность GTNKRAP (SEQ ID NO: 20); и VL CDR3, содержащую аминокислотную последовательность ALWYSNLWV (SEQ ID NO: 21).
[0068] В некоторых вариантах осуществления анти-CD3 dsFv содержит: VH, имеющую аминокислотную последовательность любой из SEQ ID NO: 14, 44 и 32-62, или последовательность, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с любой из SEQ ID NO: 14, 44 и 32-62; и VL, имеющую аминокислотную последовательность любой из SEQ ID NO: 15, 72 и 63-81 или последовательность, которая имеет, по меньшей мере, 90%, 91%, 92%, 93%, 94%, 95%, 96% 97%, 98% или 99% идентичности последовательности с любой из SEQ ID NO: 14, 44 и 32-62. В некоторых случаях анти-CD3 dsFv содержит аминокислотную последовательность SEQ ID NO: 14 и аминокислотную последовательность SEQ ID NO: 15. В некоторых случаях анти-CD3 dsFv содержит аминокислотную последовательность SEQ ID NO: 44 и аминокислотную последовательность SEQ ID NO: 72.
[0069] В некоторых вариантах осуществления Fc-область иммуноглобулина первого компонента представляет собой изотип IgG, выбранный из группы, состоящей из изотипа IgG1, изотипа IgG2, изотипа IgG3 и подкласса IgG4. В некоторых вариантах осуществления Fc-область иммуноглобулина представляет собой полипептид, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-6. В некоторых вариантах осуществления Fc-область иммуноглобулина представляет собой полипептид, содержащий аминокислотную последовательность, которая имеет по меньшей мере 50%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1-6.
[0070] В некоторых вариантах осуществления Fc-область иммуноглобулина представляет собой полипептид, содержащий аминокислотную последовательность, которая получена из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-6. В некоторых вариантах осуществления Fc-область иммуноглобулина представляет собой полипептид, содержащий аминокислотную последовательность, которая получена из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-6, содержащей одну или модификации. В некоторых вариантах осуществления Fc-область иммуноглобулина представляет собой полипептид, содержащий аминокислотную последовательность, которая получена из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-6, включающей одну или модификации, для предотвращения гликозилирования, для изменения взаимодействия рецептора Fc для уменьшения связывания с Fc-рецептором, для усиления взаимодействия с CD32A, для снижения связывания белка C1q комплемента, для увеличения периода полужизни, для усиления связывания с FcRn, для изменения антителозависимой клеточной цитотоксичности (ADCC) и/или комплементзависимой цитотоксичности (CDC), чтобы вызвать гетеродимеризацию, предотвратить димеризацию, стабилизировать гомодимеризацию на границе раздела CH3:CH3 и их комбинации. В некоторых вариантах осуществления модификации в Fc-области уменьшают связывание с Fc-гамма-рецепторами, в то время как оказывают минимальное влияние на связывание с неонатальным Fc-рецептором (FcRn). В некоторых вариантах осуществления мутированный или модифицированный полипептид Fc включает следующие мутации: Met252Tyr и Met428Leu или Met252Tyr и Met428Val (M252Y, M428L или M252Y, M428V) с использованием системы нумерации Kabat.
[0071] В некоторых вариантах осуществления первый антигенсвязывающий домен и полипептид Fc иммуноглобулина функционально связаны через аминокислотные линкеры. В некоторых вариантах осуществления эти внутрикомпонентные линкеры состоят преимущественно из аминокислот глицина и серина, обозначаемых здесь как GS-линкеры. GS-линкеры слитых белков по настоящему изобретению могут иметь различную длину, например 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 аминокислот в длину.
[0072] В некоторых вариантах осуществления GS-линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из GGSGGS, т.е. (GGS)2 (SEQ ID NO: 10); GGSGGSGGS, т.е. (GGS)3 (SEQ ID NO: 11); GGSGGSGGSGGS, т.е. (GGS)4 (SEQ ID NO: 12); и GGSGGSGGSGGSGGS, т.е. (GGS)5 (SEQ ID NO: 13).
[0073] В некоторых вариантах осуществления анти-CD3ε dsFv-фрагмент антитела включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 32-81. В некоторых вариантах осуществления анти-CD3ε dsFv-фрагмент антитела включает аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32-81. В некоторых вариантах осуществления анти-CD3ε Fv-фрагмент антитела включает в себя комбинацию аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32-62, и аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 63-81. В некоторых вариантах осуществления анти-CD3ε Fv-фрагмент антитела включает в себя комбинацию аминокислотной последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32-62, и аминокислотной последовательности, которая, по меньшей мере, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из аминокислотной последовательности SEQ ID NO: 63-81.
[0074] В некоторых вариантах осуществления второй антигенсвязывающий домен и анти-CD3-связывающий домен функционально связаны через аминокислотные линкеры. В некоторых вариантах осуществления эти внутрикомпонентные линкеры состоят преимущественно из аминокислот глицина и серина, обозначаемых здесь как GS-линкеры. GS-линкеры слитых белков по настоящему изобретению могут иметь различную длину, например 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 аминокислот в длину.
[0075] В некоторых вариантах осуществления GS-линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из GGSGGS, т.е. (GGS)2 (SEQ ID NO: 10); GGSGGSGGS, т.е. (GGS)3 (SEQ ID NO: 11); GGSGGSGGSGGS, т.е. (GGS)4 (SEQ ID NO: 12); и GGSGGSGGSGGSGGS, т.е. (GGS)5 (SEQ ID NO: 13).
[0076] В некоторых вариантах осуществления расщепляемый линкер представляет собой полипептид. В некоторых вариантах осуществления расщепляемый линкер представляет собой полипептид, который является субстратом для протеазы. В некоторых вариантах осуществления протеаза продуцируется опухолью, которая находится вблизи клеток, которые экспрессируют CD3ε, и/или продуцируется опухолью, которая локализована совместно с клетками, которые экспрессируют CD3ε в ткани, и где протеаза расщепляет расщепляемый линкер в полиспецифической полипептидной конструкции, когда полиспецифическая полипептидная конструкция подвергается воздействию протеазы. В некоторых вариантах осуществления протеаза продуцируется опухолью, которая находится вблизи клеток, которые экспрессируют один или более опухолеспецифических антигенов (TAA), и/или продуцируется опухолью, которая локализована совместно с клетками, которые экспрессирут TAA-мишень в ткани, и где протеаза расщепляет расщепляемый линкер в полиспецифической полипептидной конструкции, когда полиспецифическая полипептидная конструкция подвергается воздействию протеазы.
[0077] В некоторых вариантах осуществления расщепляемый линкер представляет собой полипептид длиной до 50 аминокислот. В некоторых вариантах осуществления расщепляемый линкер представляет собой полипептид длиной до 25 аминокислот. В некоторых вариантах осуществления расщепляемый линкер представляет собой полипептид длиной до 15 аминокислот. В некоторых вариантах осуществления расщепляемый линкер представляет собой субстрат для протеазы, выбранной из протеаз, описанных в настоящем документе. В некоторых вариантах осуществления расщепляемый линкер представляет собой субстрат для протеазы, выбранной из группы, состоящей из uPA, легумаина, матриптазы (также называемой в настоящем документе MT-SP1 или MTSP1), ADAM17, BMP-1, TMPRSS3, TMPRSS4, MMP-9, ММР-12, ММР-13, ММР-14 и любой их комбинации. В некоторых вариантах осуществления расщепляемый линкер представляет собой субстрат для протеазы, выбранной из группы, состоящей из uPA, легумаина и матриптазы. В некоторых вариантах осуществления расщепляемый линкер представляет собой субстрат для матричной металлопротеазы (ММР).
[0078] В некоторых вариантах осуществления полиспецифическая конструкция также включает агент, конъюгированный с полиспецифической конструкцией. В некоторых вариантах осуществления агент представляет собой терапевтический агент. В некоторых вариантах осуществления агент представляет собой детектируемый фрагмент. В некоторых вариантах осуществления детектируемый фрагмент представляет собой диагностический агент. В некоторых вариантах осуществления агент конъюгирован с полиспецифической конструкцией через линкер. В некоторых вариантах осуществления линкер представляет собой расщепляемый линкер. В некоторых вариантах осуществления линкер представляет собой нерасщепляемый линкер.
[0079] В некоторых вариантах осуществления описанная в настоящем документе анти-полиспецифическая конструкция используется в сочетании с одним или более дополнительными агентами или комбинацией дополнительных агентов. Подходящие дополнительные агенты включают современные фармацевтические и/или хирургические методы лечения для назначенного применения, как например при злокачественном новообразовании. Например, полиспецифическая конструкция может быть использована в сочетании с дополнительным химиотерапевтическим или противоопухолевым агентом.
[0080] В некоторых вариантах осуществления полиспецифическая конструкция и дополнительный агент включены в одну терапевтическую композицию, и полиспецифическая конструкция и дополнительный агент вводятся одновременно. Альтернативно, полиспецифическая конструкция и дополнительный агент отделены друг от друга, например, каждый включен в терапевтическую композицию, и полиспецифическая полипептидная конструкция и дополнительный агент вводятся одновременно, или полиспецифическая конструкция и дополнительный агент вводятся в разное время во время схемы лечения. Например, полиспецифическая конструкция вводится до введения дополнительного агента, полиспецифическая конструкция вводится после введения дополнительного агента, или полиспецифическая конструкция и дополнительный агент вводятся поочередно. Как описано в настоящем документе, полиспецифическая конструкция и дополнительный агент вводятся в однократных дозах или в многократных дозах.
[0081] В некоторых вариантах осуществления полиспецифическая конструкция естественным образом содержит одну или более дисульфидных связей. В некоторых вариантах осуществления полиспецифическая конструкция может быть сконструирована так, чтобы включать одну или более дисульфидных связей.
[0082] Раскрытие также относится к выделенной молекуле нуклеиновой кислоты или полинуклеотиду, кодирующему, по меньшей мере, часть полиспецифической конструкции, описанной в настоящем документе, и/или к одной или более молекулам нуклеиновой кислоты, кодирующим полиспецифическую конструкцию, описанную в настоящем документе, таким как, например, по меньшей мере, первая нуклеиновая кислота, кодирующая по меньшей мере, часть первого компонента полиспецифической конструкции, и вторая нуклеиновая кислота, кодирующая, по меньшей мере, часть второго компонента полиспецифической конструкции, а также к векторам, которые включают эти выделенные последовательности нуклеиновой кислоты.
[0083] Среди представленных вариантов осуществления находится полинуклеотид(ы), кодирующий любую из представленных полиспецифических полипептидных конструкций. Также предложен полинуклеотид, кодирующий полипептидную цепь любой из представленных полиспецифических полипептидных конструкций. Далее предложен полинуклеотид, содержащий первую последовательность нуклеиновой кислоты, кодирующую первый полипептид любой из представленных полиспецифических конструкций, и вторую последовательность нуклеиновой кислоты, кодирующую второй полипептид полиспецифической конструкции, где первая и вторая последовательности нуклеиновой кислоты разделены участком внутренней посадки рибосомы (IRES) или нуклеиновой кислотой, кодирующей саморасщепляющийся пептид или пептид, который вызывает пропуск рибосомы. В некоторых случаях первая последовательность нуклеиновой кислоты и вторая последовательность нуклеиновой кислоты функционально связаны с одним и тем же промотором. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит третью полипептидную цепь, а полинуклеотид дополнительно содержит третью нуклеиновую кислоту, кодирующую третий полипептид полиспецифической конструкции. В некоторых вариантах осуществления третья нуклеиновая кислота отделена от первого и/или второго полипептида участком внутренней посадки рибосомы (IRES) или нуклеиновой кислотой, кодирующей саморасщепляющийся пептид или пептид, который вызывает пропуск рибосомы, и/или третья последовательность нуклеиновой кислоты функционально связана с тем же промотором, что и первая и/или вторая последовательность нуклеиновой кислоты. В некоторых примерах нуклеиновая кислота, кодирующая саморасщепляющийся пептид или пептид, который вызывает пропуск рибосомы, выбирается из T2A, P2A, E2A или F2A (SEQ ID NO: 159-164, или кодируется последовательностью, представленной в SEQ ID NO: 165).
[0084] В настоящем описании предложен вектор, содержащий любой из предоставленных полинуклеотидов. В некоторых вариантах осуществления вектор представляет собой экспрессирующий вектор. В некоторых примерах вектор представляет собой вирусный вектор или эукариотический вектор, необязательно, где эукариотический вектор представляет собой вектор млекопитающего.
[0085] Предложена клетка, содержащая любой из предоставленных полинуклеотидов или векторов. В некоторых случаях клетка является рекомбинантной или выделенной. В некоторых примерах клетка является клеткой млекопитающего. В некоторых примерах клетка является клеткой HEK293 или CHO.
[0086] Раскрытие относится к способам получения полиспецифической конструкции путем культивирования клетки в условиях, которые приводят к экспрессии полиспецифической конструкции, где клетка содержит такую молекулу(молекулы) нуклеиновой кислоты. В некоторых вариантах осуществления клетка содержит такой вектор.
[0087] В настоящем описании предложен способ получения полиспецифической полипептидной конструкции, включающий введение в клетку любого из предложенных полинуклеотидов или векторов и культивирование клетки в условиях, которые приводят к экспрессии полиспецифической конструкции с получением полиспецифической полипептидной конструкции. Также предложен способ получения полиспецифической полипептидной конструкции, включающий культивирование любой из предложенных клеток в условиях, в которых полиспецифический полипептид экспрессируется или продуцируется клеткой. В некоторых случаях клетка является клеткой млекопитающего. В некоторых примерах клетка является клеткой HEK293 или CHO. В некоторых вариантах осуществления способ дополнительно включает выделение или очистку полиспецифической полипептидной конструкции из клетки. В некоторых случаях полиспецифическая полипептидная конструкция представляет собой гетеродимер.
[0088] В настоящем описании предложена полиспецифическая полипептидная конструкция, полученная любым из предложенных способов.
[0089] В настоящем описании предложен способ стимулирования или индукции иммунного ответа, включающий контакт клетки-мишени и Т-клетки с любой из предложенных полиспецифических полипептидных конструкций или фармацевтических композиций, причем указанная клетка-мишень экспрессирует опухолеспецифический антиген, распознаваемый полиспецифической полипептидной конструкцией. В некоторых вариантах осуществления клетка-мишень представляет собой опухолевую клетку, экспрессирующую опухолеспецифический антиген (TAA).
[0090] В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит расщепляемый линкер, который функционирует в качестве субстрата для протеазы, и индукция или стимулирование иммунного ответа усиливаются в присутствии протеазы. В некоторых случаях протеаза продуцируется иммунной эффекторной клеткой, опухолью или клетками, присутствующими в микроокружении опухоли.
[0091] В некоторых вариантах осуществления протеаза продуцируется иммунной эффекторной клеткой, и иммунная эффекторная клетка представляет собой активированную Т-клетку, клетку-натуральный киллер (NK) или Т-клетку NK. В некоторых случаях иммунная эффекторная клетка находится в непосредственной близости от клеток, которые экспрессируют антиген. В некоторых вариантах осуществления протеаза продуцируется опухолью, которая находится вблизи клеток, которые экспрессируют TAA в ткани, и/или продуцируется опухолью, которая локализована совместно с TAA в ткани, и где протеаза расщепляет расщепляемый линкер в полиспецифической полипептидной конструкции, когда полиспецифическая полипептидная конструкция подвергается воздействию протеазы. В некоторых примерах протеаза выбрана из матриптазы, матриксной металлопротеазы (ММР), гранзима В и их комбинаций. В некоторых случаях протеазой является гранзим B.
[0092] В некоторых вариантах осуществления контактирование осуществляется ex vivo или in vitro. В некоторых вариантах осуществления контактирование осуществляется in vivo у пациента.
[0093] Предложен способ стимулирования или индукции иммунного ответа у пациента, включающий введение пациенту, нуждающемуся в этом, терапевтически эффективного количества любого из предложенных полиспецифических конъюгатов или фармацевтических композиций. В некоторых случаях способ повышает клеточно-опосредованный иммунитет. В некоторых вариантах осуществления способ увеличивает активность Т-клеток. В некоторых вариантах осуществления способ увеличивает активность цитолитических Т-клеток (CTL). В некоторых примерах иммунный ответ повышается против опухоли или злокачественного новообразования. В некоторых вариантах осуществления с помощью способа подвергают лечению заболевание или состояние у пациента.
[0094] Настоящее раскрытие также относится к способам лечения, предотвращения, задержки прогрессирования или иного облегчения симптома одной или более патологий или облегчения симптома, связанного с такими патологиями, путем введения полиспецифической полипептидной конструкции раскрытия пациенту, для которого такое лечение или профилактика желательны. В настоящем описании предложен способ лечения заболевания или состояния у пациента, причем способ включает введение пациенту, нуждающемуся в этом, терапевтически эффективного количества любого из предложенных полиспецифических конъюгатов или фармацевтических композиций. В некоторых вариантах осуществления заболевание или состояние представляет собой опухоль или злокачественное новообразование.
[0095] В некоторых вариантах осуществления любого из представленных способов пациент, такой как пациент, подлежащий лечению, представляет собой, например, человека или другое млекопитающее. В некоторых вариантах осуществления любого из предложенных способов пациент представляет собой человека. В некоторых вариантах осуществления пациент представляет собой млекопитающее, не являющееся человеком, такое как примат, не являющийся человеком, домашнее животное (например, кошка, собака, лошадь), сельскохозяйственное животное, рабочее животное или животное зоопарка. В некоторых вариантах осуществления пациентом является грызун.
[0096] Полиспецифическая полипептидная конструкция по изобретению, используемая в любом из вариантов осуществления этих способов и применений, может быть введена на любой стадии заболевания. Например, такую полиспецифическую полипептидную конструкцию можно вводить пациенту, имеющему злокачественное новообразование любой стадии, от ранней до метастатической. Термины субъект и пациент используются в настоящем описании взаимозаменяемо.
[0097] Полиспецифическая полипептидная конструкция по настоящему изобретению, используемая в любом из вариантов осуществления этих способов и применений, может быть использована в схеме лечения, включающей неоадъювантную терапию.
[0098] Полиспецифическая полипептидная конструкция по изобретению, используемая в любом из вариантов осуществления этих способов и применений, может вводиться либо отдельно, либо в комбинации с одним или более дополнительными агентами, включая низкомолекулярные ингибиторы, другие методы лечения на основе антител, полипептидную или пептидную терапию, терапии на основе нуклеиновых кислот и/или других биологических препаратов. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция вводится в комбинации с одним или более дополнительными агентами, такими как, в качестве неограничивающего примера, химиотерапевтический агент, такой как алкилирующий агент, антиметаболит, антимикротрубочковый агент, ингибитор топоизомеразы, цитотоксический антибиотик и любой другой агент, повреждающий нуклеиновую кислоту. В некоторых вариантах осуществления дополнительный агент представляет собой таксан, такой как паклитаксел (например, Abraxane®). В некоторых вариантах осуществления дополнительный агент представляет собой антиметаболит, такой как гемцитабин. В некоторых вариантах осуществления дополнительный агент представляет собой алкилирующий агент, такой как химиотерапивтеческо средство на основе платины, такое как карбоплатин или цисплатин. В некоторых вариантах осуществления дополнительный агент представляет собой целевой агент, такой как ингибитор киназы, например сорафениб или эрлотиниб. В некоторых вариантах осуществления дополнительный агент представляет собой целевой агент, такой как другое антитело, например, моноклональное антитело (например, бевацизумаб), биспецифическое антитело или полиспецифическое антитело. В некоторых вариантах осуществления дополнительный агент представляет собой ингибитор протеасомы, такой как бортезомиб или карфилзомиб. В некоторых вариантах осуществления дополнительный агент представляет собой иммуномодулятор, такой как ленолидоминд или IL-2. В некоторых вариантах осуществления дополнительный агент представляет собой излучение. В некоторых вариантах осуществления дополнительный агент представляет собой агент, который считается стандартным для специалистов в данной области. В некоторых вариантах осуществления дополнительный агент представляет собой химиотерапевтический агент, хорошо известный специалистам в данной области. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция и дополнительный агент(ы) включены в одну композицию. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция и дополнительный агент(ы) вводятся в виде двух или более отдельных композиций. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция и дополнительный агент(ы) вводятся одновременно. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция и дополнительный агент(ы) вводятся последовательно.
[0099] В некоторых вариантах осуществления дополнительный агент(ы) представляет собой химиотерапевтический агент, такой как химиотерапевтический агент, выбранный из группы, состоящей из доцетаксела, паклитаксела, абраксана (то есть альбумин-конъюгированного паклитаксела), доксорубицина, оксалиплатина, карбоплатина, цисплатина, иринотекана, и гемцитабина.
[0100] В некоторых вариантах осуществления дополнительный агент(ы) представляет собой ингибитор контрольной точки, ингибитор киназы, агент, нацеленный на ингибиторы в микроокружении опухоли, и/или Т-клеточный или NK-агонист. В некоторых вариантах осуществления дополнительный агент(ы) представляет собой лучевую терапию, отдельно или в комбинации с другим дополнительным агентом(ами), таким как химиотерапевтический или противоопухолевый агент. В некоторых вариантах осуществления дополнительный агент(ы) представляет собой вакцину, онковирус, и/или DC-активирующий агент, такой как, в качестве не ограничивающего примера, агонист toll-подобного рецептора (TLR) и/или α-CD40. В некоторых вариантах осуществления дополнительный агент(ы) представляет собой антитело, нацеленное на опухоль, предназначенное для уничтожения опухоли через ADCC или путем прямого конъюгирования с токсином (например, конъюгат лекарственное средство-антитело (ADC).
[0101] В некоторых вариантах осуществления ингибитор контрольной точки представляет собой ингибитор мишени, выбранный из группы, состоящей из CTLA-4, LAG-3, PD-1, PDL1, TIGIT, TIM-3, B7H3, B7H4 и Vista. В некоторых вариантах осуществления ингибитор киназы выбирают из группы, состоящей из ингибиторов B-RAFi, MEKi и Btk, таких как ибрутиниб. В некоторых вариантах осуществления ингибитор киназы представляет собой кризотиниб. В некоторых вариантах осуществления ингибитор микроокружения опухоли выбран из группы, состоящей из ингибитора IDO, ингибитора α-CSF1R, ингибитора α- CCR4, TGF-бета, клетки-супрессора миелоидного происхождения или T-регуляторной клетки. В некоторых вариантах осуществления агонист выбран из группы, состоящей из OX40, GITR, CD137, CD28, ICOS, CD27 и HVEM. В некоторых вариантах осуществления ингибитор контрольной точки представляет собой антитело, которое связывается с мишенью, выбранной из CTLA-4, PD-1 и/или PD-L1. В некоторых вариантах осуществления ингибитор контрольной точки представляет собой антитело против CTLA4, антитело против PD-1 и антитело против PD-L1 и/или их комбинации. В некоторых вариантах осуществления ингибитор контрольной точки представляет собой антитело против CTLA4, такое как, например, Yervoy™. В некоторых вариантах осуществления ингибитор контрольной точки представляет собой антитело против PD-1, такое как, например, Opdivo™ и/или Keytruda™.
[0102] В некоторых вариантах осуществления ингибитор представляет собой ингибитор CTLA-4. В некоторых вариантах осуществления ингибитор представляет собой ингибитор LAG-3. В некоторых вариантах осуществления ингибитор представляет собой ингибитор PD-1. В некоторых вариантах осуществления ингибитор представляет собой ингибитор PDL1. В некоторых вариантах осуществления ингибитор представляет собой ингибитор TIGIT. В некоторых вариантах осуществления ингибитор представляет собой ингибитор TIM-3. В некоторых вариантах осуществления ингибитор представляет собой ингибитор B7H3. В некоторых вариантах осуществления ингибитор представляет собой ингибитор B7H4. В некоторых вариантах осуществления ингибитор представляет собой ингибитор Vista. В некоторых вариантах осуществления ингибитор представляет собой ингибитор B-RAFi. В некоторых вариантах осуществления ингибитор представляет собой ингибитор MEKi. В некоторых вариантах осуществления ингибитор представляет собой ингибитор Btk. В некоторых вариантах осуществления ингибитор представляет собой ибрутиниб. В некоторых вариантах осуществления ингибитор представляет собой кризотиниб. В некоторых вариантах осуществления ингибитор представляет собой ингибитор IDO. В некоторых вариантах осуществления, ингибитор представляет собой ингибитор α -CSF1R. В некоторых вариантах осуществления ингибитор представляет собой ингибитор α-CCR4. В некоторых вариантах осуществления ингибитор представляет собой TGF-бета. В некоторых вариантах осуществления ингибитор представляет собой клетку-супрессор миелоидного происхождения. В некоторых вариантах осуществления ингибитор представляет собой регуляторную T-клетку.
[0103] В некоторых вариантах осуществления агонист представляет собой OX40. В некоторых вариантах осуществления агонист представляет собой GITR. В некоторых вариантах осуществления агонист представляет собой CD137. В некоторых вариантах осуществления агонист представляет собой CD28. В некоторых вариантах осуществления агонист представляет собой ICOS. В некоторых вариантах осуществления агонист представляет собой CD27. В некоторых вариантах осуществления агонист представляет собой HVEM.
[0104] В некоторых вариантах осуществления полиспецифическая полипептидная конструкция вводится во время и/или после лечения в комбинации с одним или более дополнительными агентами, такими как, например, химиотерапевтический агент, противовоспалительный агент и/или иммуносупрессивный агент. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция и дополнительный агент включены в состав одной терапевтической композиции, и полиспецифическая полипептидная конструкция и дополнительный агент вводятся одновременно. Альтернативно, полиспецифическая полипептидная конструкция и дополнительный агент отделены друг от друга, например, каждый включен в отдельную терапевтическую композицию, и полиспецифическая полипептидная конструкция и дополнительный агент вводятся одновременно, или полиспецифическая полипептидная конструкция и дополнительный агент вводятся в разное время во время схемы лечения. Например, полиспецифическая полипептидная конструкция вводится до введения дополнительного агента, полиспецифическая полипептидная конструкция вводится после введения дополнительного агента, или полиспецифическая полипептидная конструкция и дополнительный агент вводятся поочередно. Как описано в настоящем документе, полиспецифическая полипептидная конструкция и дополнительный агент вводятся в однократных дозах или в многократных дозах.
[0105] В некоторых вариантах осуществления полиспецифическая полипептидная конструкция и дополнительный агент(ы) вводятся одновременно. Например, полиспецифическая полипептидная конструкция и дополнительный агент(ы) могут быть включены в одну композицию или введены в виде двух или более отдельных композиций. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция и дополнительный агент(ы) вводятся последовательно, или полиспецифическая полипептидная конструкция и дополнительный агент вводятся в разное время во время схемы лечения.
[0106] В дополнение к элементам, описанным выше, полиспецифическая полипептидная конструкция может содержать дополнительные элементы, такие как, например, аминокислотная последовательность N- или C-конца полиспецифической полипептидной конструкции. Например, полиспецифическая полипептидная конструкция может включать нацеливающий фрагмент для облегчения доставки в интересующую клетку или ткань. Полиспецифическая полипептидная конструкция может быть конъюгирована с агентом, таким как терапевтический агент, детектируемый фрагмент или диагностический агент. Примеры агентов раскрыты в настоящем описании.
[0107] Полиспецифическая полипептидная конструкция также может включать любой из конъюгированных агентов, линкеров и других компонентов, описанных в настоящем документе в сочетании с полиспецифической полипептидной конструкцией по настоящему изобретению.
[0108] Изобретение также относится к иммуноконъюгатам, содержащим полиспецифическую полипептидную конструкцию, конъюгированным с цитотоксическим агентом, таким как токсин (например, ферментативно активный токсин бактериального, грибного, растительного или животного происхождения, или их фрагменты) или радиоактивный изотоп (т.е. радиоконъюгат). Подходящие цитотоксические агенты для применения при нацеливании на больные Т-клетки, такие как Т-клетки с происхождением из лимфомы, включают, например, доластатины и их производные (например, ауристатин E, AFP, MMAD, MMAF, MMAE). В некоторых вариантах осуществления агент представляет собой доластатин. В некоторых вариантах осуществления агент представляет собой ауристатин или его производное. В некоторых вариантах осуществления агент представляет собой майтансиноид или производное майтансиноида. В некоторых вариантах осуществления агент представляет собой DM1 или DM4. В некоторых вариантах осуществления агент представляет собой дуокармицин или его производное. В некоторых вариантах осуществления агент представляет собой калихеамицин или его производное. В некоторых вариантах осуществления агент представляет собой пирролобензодиазепин.
[0109] В некоторых вариантах осуществления линкер между полиспецифической полипептидной конструкцией и цитотоксическим агентом является расщепляемым. В некоторых вариантах осуществления линкер является нерасщепляемым. В некоторых вариантах осуществления присутствуют два или более линкеров. Два или более линкеров являются одинаковыми, например, расщепляемыми или нерасщепляемыми, или два или более линкеров являются разными, например, по меньшей мере, один расщепляемый и, по меньшей мере, один нерасщепляемый.
[0110] Полиспецифические полипептидные конструкции и их конъюгаты полезны в способах лечения различных расстройств и/или заболеваний. Неограничивающие примеры заболевания включают: все виды злокачественных новообразований (рак молочной железы, легких, колоректальный, предстательной железы, меланомы, рак головы и шеи, поджелудочной железы и т. д.), ревматоидный артрит, болезнь Крона, SLE, сердечно-сосудистые повреждения, ишемия и т. д., например, показания могут включать лейкозы, включая Т-клеточный острый лимфобластный лейкоз (T-ALL), лимфобластные заболевания, включая множественную миелому, и солидные опухоли, включая рак легкого, колоректальный рак, рак предстательной железы, поджелудочной железы и молочной железы, включая тройной негативный рак молочной железы. Например, показания включают заболевание костей или метастазирование при злокачественном новообразовании, независимо от первичного происхождения опухоли; рак молочной железы, включая в качестве неограничивающего примера, ER/PR+рак молочной железы, Her2+рак молочной железы, тройной негативный рак молочной железы; колоректальный рак; рак эндометрия; рак желудка; глиобластому; рак головы и шеи, как например, рак пищевода; рак легкого, как, например, немелкоклеточный рак легкого; множественную миелому, рак яичников; рак поджелудочной железы; рак предстательной железы; саркому, как например, остеосаркому; рак почки, как например, в качестве неограничивающего примера, почечно-клеточную карциному; и/или рак кожи, как например, в качестве неограничивающего примера, плоскоклеточный рак, базально-клеточную карциному или меланому. В некоторых вариантах осуществления злокачественное новообразование представляет собой плоскоклеточный рак. В некоторых вариантах осуществления злокачественное новообразование представляет собой плоскоклеточную карциному кожи. В некоторых вариантах осуществления злокачественное новообразование представляет собой плоскоклеточную карциному пищевода. В некоторых вариантах осуществления злокачественное новообразование представляет собой плоскоклеточную карциному головы и шеи. В некоторых вариантах осуществления злокачественное новообразование представляет собой плоскоклеточную карциному легкого.
[0111] Также предложена фармацевтическая композиция, содержащая любую из полиспецифических полипептидных конструкций, представленных в настоящем документе, и фармацевтически приемлемый носитель. В некоторых случаях фармацевтическая композиция является стерильной. Фармацевтические композиции по настоящему изобретению могут включать полиспецифическую полипептидную конструкцию по настоящему изобретению и носитель. Эти фармацевтические композиции могут быть включены в наборы, такие как, например, диагностические наборы.
[0112] Специалисту в данной области будет понятно, что антитела по изобретению имеют множество применений. Например, белки по настоящему изобретению используются в качестве терапевтических агентов для различных расстройств. Антитела по изобретению также используются в качестве реагентов в диагностических наборах или в качестве диагностических инструментов, или эти антитела могут использоваться в конкурентных анализах для получения терапевтических реагентов.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0113] ФИГ. 1 представляет собой схему основных компонентов полиспецифических полипептидных конструкций по настоящему изобретению, обладающих ограниченным связыванием с CD3. Антигенсвязывающий домен(ы) расположен на амино- и/или карбоксиконце. Fc-область, такая как гетеродимерная Fc-область, расположена на N-конце относительно CD3-связывающей области. Такое расположение Fc в непосредственной близости от CD3-связывающей области препятствует связыванию CD3.
[0114] ФИГ. 2 представляет собой иллюстрацию, изображающую примерную структуру полиспецифической молекулы раскрытия, содержащей расщепляемый линкер и имеющей двойные эффекторные функции, где протеолитическое расщепление расщепляемого линкера приводит к активации полиспецифической полипептидной конструкции с образованием двух компонентов, каждый из которых обладает биологической активностью.
[0115] ФИГ. 3 представляет собой схему различных FRα-нацеленных CD3-ограниченных конструкций, состоящих из двух полипептидов, Цепи 1 и Цепи 2. Верхняя панель представляет примерное изображение расщепляемой полиспецифической полипептидной конструкции, имеющей расщепляемый линкер, содержащий сайт или сайты узнавания субстрата протеазы, например, для одного или более из MTSP1, MMP и/или гранзима B. Цепь 1 содержит FRα sdAb (антигенсвязывающий домен), связанный с гетеродимерной «впадиной» Fc, связанный через расщепляемый протеазой линкер (cx1547: только гранзим B, cx309: MTSP1, MMP и гранзим B) с анти-CD3 VL-доменом, связанным со вторым FRα sdAb, Цепь 2 содержит FRα sdAb, связанный с комплементарным гетеродимерным «выступом» Fc, связанным через тот же протеазный линкер, что и выше, с анти-CD3 VH-доменом, связанным со вторым sdAb FRα. Нижняя панель ФИГ. 3 изображает конфигурацию, подобную верхней панели, за исключением того, что линкер представляет собой нерасщепляемый линкер (в диапазоне от 3 аминокислот в cx1356 до 18 аминокислот в cx681). При совместной экспрессии CD3-связывающий домен правильно собирается через ассоциацию VL:VH на «впадине» и «выступе», соответственно.
[0116] ФИГ. 4А-4С изображают конструкции, которые были сгенерированы для сравнения влияния линкера на ограничение связывания CD3 в сгенерированных конструкциях. ФИГ. 4А изображает пример полиспецифической полипептидной конструкции, содержащей один и тот же расщепляемый линкер в каждой полипептидной цепи, для присоединения каждого полипептида Fc гетеродимерной Fc-области к домену CD3-связывающей области (показана иллюстративная конструкция cx1762). В формате, показанном на ФИГ. 4А, конструкция изображена в своем нерасщепленном состоянии. Альтернативный вариант конструкции показан на ФИГ. 4B, в котором используется только один расщепляемый линкер для связывания Fc-области с CD3-связывающим доменом, обозначенным как наполовину расщепленный (показана примерная конструкция cx3238). ФИГ. 4C изображает структуру, которая представляет С-концевую часть формата конструкций на ФИГ. 4А и 4В, когда они полностью расщеплены (показана примерная конструкция cx2190). Конструкции, представляющие продукт протеолитического расщепления, могут быть получены рекомбинантно путем совместной экспрессии различных изображенных цепей. FRα-нацеленное sdAb расположено в С-концевом положении в каждой конструкции.
[0117] ФИГ. 5А-5Е изображают репрезентативные ограниченные по CD3 рекрутеры, нацеленные на EGFR и с двойным нацеливанием на EGFR/cMET. На ФИГ. 5А, cx2513 содержит EGFR-нацеленное sdAb, расположенное на С-конце каждой цепи гетеродимера и таким образом отображает двухвалентное связывание с EGFR. На ФИГ. 5B, cx3030 содержит sdAb, нацеленные на EGFR, расположенные как на N, так и на С-концах каждой цепи гетеродимера, и, таким образом, обнаруживает четырехвалентное связывание с EGFR. ФИГ. 5C, cx2973 содержит sdAb, нацеленное на cMET, расположенное на N-конце, и sdAb, нацеленное на EGFR, расположенное на C-концах каждой цепи гетеродимера, и, таким образом, демонстрирует двухвалентное связывание с каждым cMET и EGFR. На ФИГ. 5D, cx2979 содержит sdAb, нацеленное на cMET, расположенное на N-концах одной цепи гетеродимера, и sdAb, нацеленное на EGFR, расположенное на C-концах каждой цепи гетеродимера, и, таким образом, обнаруживает одновалентное связывание cMET и двухвалентное связывание с EGFR. На ФИГ. 5E, cx2977 содержит sdAb, нацеленное на cMET, расположенное на N-конце, и sdAb, нацеленное на EGFR, расположенное на C-конце одной или другой цепи гетеродимера, и, таким образом, обнаруживает одновалентное связывание с каждым, cMET и EGFR.
[0118] ФИГ. 6А и 6В представляют собой схемы цепей компонентов для сборки примерных B3H3-нацеленных ограниченных CD3-рекрутеров. B7H3-связывающие домены, используемые в этих репрезентативных конструкциях, включали sdAb, scFv или FAB. Обычно конструкции, содержащие sdAb и scFv, состояли из двух гетеродимерных цепей, тогда как конструкции, содержащие FAB, включали третью цепь для родственной легкой цепи (VL-CL).
[0119] ФИГ. 7 изображает 5T4-нацеленные ограниченные CD3-рекрутеры. Ядро молекулы было сгенерировано так, чтобы оно содержало гетеродимерную Fc-область, за которой следовали расщепляемые линкеры и стабилизированный дисульфидом анти-CD3 Fv. TAA-связывающая часть этих молекул была размещена на N- или C-концах любой гетеродимерной Fc-цепи. В верхнем ряду TAA-связывающей единицей является Fab, состоящий из Fd (VH-CH1) на N-конце полипептида-выступа и С-конца полипептида-впадины. В случае, когда Fab является связывающей единицей, третья цепь (легкая цепь - VL-CL) экспрессируется для связывания с Fd. В среднем и нижнем ряду TAA-связывающие единицы представляют собой однодоменные антитела, которые были расположены на полипептиде-выступе на N- и C-конце. В среднем ряду TAA-связывающие sdAb сгенерированной конструкции, одинаковы, а в нижнем ряду TAA-связывающие sdAb сгенерированной конструкции представляют собой разные последовательности с разными эпитопами.
[0120] ФИГ. 8 представляет собой схему репрезентативной CD20-нацеленной ограниченной CD3-рекрутирующей конструкции, cx3309, где CD20-связывающие домены представляют собой scFv, полученные из CD20-антитела GA101.
[0121] ФИГ. 9 представляет собой схему репрезентативной DLL3-нацеленной ограниченной CD3-рекрутирующей конструкции, cx3308, где DLL3-связывающими доменами являются scFv. Эта примерная конструкция состоит из двух цепей, каждая из которых содержит комплементарный компонент гетеродимерной Fc-области, связанный с одним компонентом CD3-связывающего домена, и DLL3-связывающий scFv. В собранном виде конструкция является двухвалентной по отношению к DLL3 и имеет CD3-связывающий домен, расположенный на С-конце относительно гетеродимера Fc.
[0122] ФИГ. 10А представляет собой изображение SDS-PAGE репрезентативной FRα-нацеленной ограниченной CD3-рекрутирующей конструкции, cx1547, восстанавливающие (R) и невосстанавливающие условия (NR). Ожидаемая молекулярная масса 135 кДа. ФИГ. 10B и 10C представляют собой графики хроматограммы из эксклюзионного анализа cx1547, демонстрирующие, что это один вид с определенной молекулярной массой 137,9 кДа. ФИГ. 10C представляет собой увеличенный вид вокруг основного пика, показанного на ФИГ. 10B.
[0123] ФИГ. 11А и 11В представляют собой пару графиков, демонстрирующих способность к связыванию примерной полиспецифической полипептидной конструкции по изобретению, обозначенной здесь как cx309, для связывания человеческой Т-клетки в нерасщепленном или протеолитически расщепленном состоянии. Матриптаза и ММР-2 были использованы для расщепления cx309 на ФИГ. 11А и ФИГ. 11B, соответственно.
[0124] ФИГ. 12A-12D изображают связывание клеток с помощью репрезентативных FRα-нацеленных ограниченных CD3-рекрутирующих конструкций, cx1356, cx681 и cx1547. ФИГ. 12А и ФИГ. 12C демонстрируют связывание с клетками Ovcar5 (FRα-положительная клеточная линия рака яичника). ФИГ. 12B и ФИГ. 12D изображают отсутствие связывания с Т-клетками.ФИГ. 12А и ФИГ. 12B изображают гистограммы нормализованных количеств клеток по сравнению с флуоресценцией при 100 нМ каждой конструкции. Полное титрование каждой конструкции на клетках различных типов показано на ФИГ. 12C и ФИГ. 12D. Только контроль вторичного антитела против человеческого АРС показан закрашенной черной кривой, в то время как связывание положительного контроля с анти-CD3 показано незакрашенной кривой, а cx1356, cx681 и cx1547 показаны серыми заштрихованными кривыми на ФИГ. 12А и ФИГ. 12B.
[0125] ФИГ. 13А-13В изображают связывание клеток с помощью репрезентативной EFGR-нацеленной ограниченной CD3-рекрутирующей конструкции, cx3030. ФИГ. 13А демонстрирует связывание с EGFR-положительной клеточной линией Colo-205 при 100 нМ. ФИГ. 13B демонстрирует отсутствие связывания с Т-клетками при 100 нМ. Связывание отображается в виде гистограмм нормализованных клеток по сравнению с флуоресценцией. Только контроль вторичного антитела против человеческого АРС показан черным закрашиванием, тогда как связывание положительного контроля с анти-CD3 показано как незакрашенное, а cx3030 показано серой штриховкой.
[0126] ФИГ. 14A-14D изображает связывание с B7H3-положительными A375 (ФИГ. 14A и 14B) и отсутствие связывания с CD3 на Т-клетках (ФИГ. 14C и 14D) с помощью B7H3-нацеленных ограниченных CD3-рекрутеров. Альтернативный формат DART-Fc с нацеливанием на B7H3 и CD3, показал сильное связывае с B7H3 и CD3 на Т-клетках. Здесь использовались различные антигенсвязывающие домены B7H3, включая cx3095 sdAb, cx3313 FAB и cx3314 scFv. ScFv и FAB содержат те же последовательности VH и VL анти-B7H3, которые используются в формате DART-Fc. ФИГ. 14А и 14С демонстрируют сравнительные гистограммы концентрации 100 нМ для каждой конструкции. Только контроль вторичного антитела против человеческого АРС показан закрашенным черным, а различные B7H3-нацеленные CD3-рекрутирующие конструкции показаны белыми незакрашенными. ФИГ. 14B и 14D демонстрируеют титрование связывания различными конструкциями с BH73 и CD3, соответственно.
[0127] ФИГ. 15А-15В изображают связывание клеток с помощью репрезентативных 5T4-нацеленных ограниченных CD3-рекрутирующих конструкций, cx3262 и cx3315. ФИГ. 15А демонстрирует связывание с 5Т4-положительной клеточной линией, Ovcar-5 при 400 нМ. ФИГ. 15B демонстрирует отсутствие связывания с Т-клетками при 400 нМ. Связывание отображается в виде гистограмм нормализованных клеток по сравнению с флуоресценцией. Только контроль вторичного антитела против человеческого АРС показан черным закрашиванием, тогда как связывание положительного контроля с анти-CD3 показано как незакрашенное, а cx3262 показано серой штриховкой.
[0128] ФИГ. 16A-16D изображает связывание клеток с помощью репрезентативной CD20-нацеленной ограниченной CD3-рекрутирующей конструкции, cx3309. ФИГ. 16А и ФИГ. 16C демонстрируют связывание с клетками Ramos (CD20-положительная клеточная линия). ФИГ. 16B и ФИГ. 16D демонстрируют отсутствие связывания с Т-клетками. ФИГ. 16A-16B отображают гистограммы нормализованных количеств клеток по сравнению с флуоресценцией при 100 нМ каждой конструкции. Полное титрование каждой конструкции на клетках различных типов показано на ФИГ. 16C-16D. Контроль только вторичного антитела против человеческого АРС показан черным закрашиванием, тогда как связывание положительного контроля с анти-CD3 показано как незакрашенное, а cx3309 - серой штриховкой.
[0129] ФИГ. 17 представляет собой график, демонстрирующий способность расщепленных или нерасщепленных cx309 активировать CD3 NFAT репортерную клеточную линию Jurkat (Promega, США) в присутствии или в отсутствии FRα-экспрессирующих клеток, Ovcar5.
[0130] ФИГ. 18 изображает антиген-зависимую активацию Т-клеток с помощью cx1547. Различные клеточные линии, являющиеся либо FRα- положительными (T47D, IGROV1, NCI-H2342, Ovcar-5, Skov-3 и A2780) или отрицательными (NCI-H460), были совместно инкубированы с репортерной клеточной линией Jurkat CD3 NFAT-GFP и флуоресценцию измеряли через 6 часов. Это демонстрирует способность ограниченных CD3-конструкций активировать Т-клетки антигенным образом.
[0131] ФИГ. 19А-19D изображает усиление способности активации Т-клеток ограниченными CD3-рекрутерами, если бы происходил протеолиз в линкере между Fc-доменом и CD3-связывающим доменом. Здесь показана кинетика активации Т-клеток, опосредованной 20 нМ cx1762, cx3238 или cx2190 в присутствии FRα-положительных клеток Ovcar-5 (фиг. 19A) или FRα-отрицательных клеток CCRF-CEM (ФИГ. 19B). Здесь также показана эффективность активации Т-клеток, опосредованная cx1762, cx3238 или cx2190, в присутствии FRα-положительных клеток Ovcar-5 (ФИГ. 19C) или FR-отрицательных клеток CCRF-CEM (фИГ. 19D). Репортерные клетки Jurkat CD3 NFAT-GFP использовали для мониторинга передачи сигналов CD3 в течение 24 часов с использованием сканера Incucyte ZOOM. В частности, активация Т-клеток зависит от экспрессии антигена на линии клеток-мишеней и значительно усиливается удалением домена Fc, расположенного на N-конце относительно CD3-связывающего домена, на одной или обеих сторонах CD3-связывающих доменов VH:VL.
[0132] ФИГ. 20A-20D представляет собой серию графиков, демонстрирующих антиген-зависимую способность активации Т-клеток различных EGFR-нацеленных и EGFR/cMET-нацеленных ограниченных CD3-рекрутеров. Примечательно, что способность активации Т-клеток повышается с увеличением валентности или дополнительной специфичности связывания антигена-мишени. Кинетика активации Т-клеток, опосредованная различными конструкциями на антиген-положительных клетках А431, изображена на ФИГ. 20А, или на антиген-отрицательных клетках CCRF-CEM. ФИГ. 20C. Эффективность активации Т-клеток различными конструкциями на антиген-положительных клетках А431 показана на ФИГ. 20B или антиген-отрицательных клетках CCRF-CEM на ФИГ. 20D. Здесь была использована репортерная клеточная линия Jurkat CD3 NFAT-GFP.
[0133] ФИГ. 21A-21B изображают способность опосредовать активацию специфических к антигену-мишени Т-клеток с помощью репрезентативной B7H3-нацеленной ограниченной CD3-рекрутирующей конструкции, cx3095, и альтернативного формата DART-Fc, нацеленного на B7H3 и CD3. Репортерные клетки Jurkat CD3 NFAT-GFP использовали для оценки активации Т-клеток в присутствии B7H3-положительной клеточной линии, A375 (ФИГ. 21A), и B7H3-отрицательной клеточной линии, Raji (ФИГ. 21B).
[0134] ФИГ. 22A-22F изображают способность опосредовать активацию специфических к антигену-мишени Т-клеток с помощью репрезентативных B7H3-нацеленных ограниченных CD3-рекрутирующих конструкций и альтернативного формата DART-Fc, нацеленного на B7H3 и CD3. Примечательно, что ограниченные CD3-рекрутирующие конструкции используют любое из sdAb, scFv или FAB, нацеленных на B7H3. Репортерные клетки Jurkat CD3 NFAT-GFP использовали для оценки активации Т-клеток в присутствии B7H3-положительной клеточной линии, A375 (ФИГ. 22A, 22C, 22E), и B7H3-отрицательной клеточной линии, CCRF (ФИГ. 22B, 22D, 22F). Показана кинетика активации T-клеток, опосредованная 50 нМ (ФИГ. 22A и 22B) или 2 нМ (ФИГ. 22C или 22D) каждой конструкции. Также показана эффективность активации Т-клеток, опосредованной каждой конструкцией на антиген-положительных (ФИГ. 22E) и отрицательных (ФИГ. 22F) клеточных линиях.
[0135] ФИГ. 23A-23B представляет собой серию графиков, показывающих способность активации T-клеток 5T4-нацеленных ограниченных CD3-рекрутирующих конструкций. В этом примере показано, как двухвалентное биэпитопное нацеливание TAA может повысить активность ограниченного CD3-рекрутера по сравнению с двухвалентным моноэпитопным белком на TAA-положительных клетках (OVCAR5). Ни одна из этих конструкций не индуцировала активацию Т-клеток в присутствии TAA-отрицательных клеток (CCRF).
[0136] ФИГ. 24 представляет собой график, показывающий способность индуцировать антиген-зависимую активацию Т-клеток с помощью репрезентативной 5T4-нацеленной ограниченной CD3-рекрутирующей конструкции, cx3315. Репортерную клеточную линию Jurkat CD3 NFAT-GFP использовали для мониторинга активации Т-клеток с помощью cx3315 в присутствии 5T4-положительной клеточной линии (OVCAR5) и 5T4-отрицательной клеточной линии (CCRF-CEM).
[0137] ФИГ. 25 изображает способность индуцировать антиген-зависимую активацию Т-клеток с помощью репрезентативной CD20-нацеленной ограниченной CD3-рекрутирующей конструкции, cx3309. Репортерную клеточную линию Jurkat CD3 NFAT-GFP использовали для мониторинга активации Т-клеток cx3309 в присутствии CD20-положительной клеточной линии: Ramos, и CD20-отрицательной клеточной линии: CCRF-CEM.
[0138] ФИГ. 26 представляет собой график, показывающий способность индуцировать активацию Т-клеток с помощью репрезентативной DLL3-нацеленной ограниченной CD3-рекрутирующей конструкции, cx3308. Репортерную клеточную линию Jurkat CD3 NFAT-GFP использовали для мониторинга активации Т-клеток cx3309 в присутствии клеток SHP-77, которые являются DLL3-положительными. Это демонстрирует, что фрагменты scFv могут использоваться для нацеливания на TAA в ограниченном формате CD3 и эффективно активируют T-клетки при связывании с клеточной линией, положительной по родственному TAA.
[0139] ФИГ. 27А-27F изображают влияние длины линкера на способность активировать Т-клетки в присутствии FRα-положительных клеток - IGROV1 (ФИГ. 27A, 27C, 27E) или FRα-отрицательных NCI-H460 (ФИГ. 27B, 27D, 27F). ФИГ. 27А-27В демонстрируют кинетику активации Т-клеток с помощью 2 нМ различных конструкций на антиген-положительных и отрицательных клетках, соответственно. ФИГ. 27C-27D демонстрируют величину способности активации Т-клеток с помощью 2 нМ различных конструкций на антиген-положительных и отрицательных клетках, соответственно. ФИГ. 27E-27F демонстрируют эффективность способности активации Т-клеток различных конструкций с различной длиной линкера на антиген-положительных и отрицательных клетках, соответственно. Репортерная клеточная линия Jurkat CD3 NFAT-GFP была использована для оценки активации Т-клеток. Ограниченные по CD3 белки эффективно рекрутируют и кластеризуют CD3 на Т-клетках, когда связываются со вторым антигеном на клетках-мишенях.
[0140] ФИГ. 28A-28C изображают FRα-зависимую T-клеточно-опосредованную цитотоксичность cx1547. ФИГ. 28А демонстрируют, что cx1547 не индуцирует Т-клеточно-опосредованную цитотоксичность антиген-отрицательной клеточной линии (NCI-H460). ФИГ. 28B демонстрирует, что cx1547 индуцирует T-клеточно-опосредованную цитотоксичность антиген-положительной клеточной линии (Ovcar5). ФИГ. 28C демонстрирует кинетику Т-клеточно-опосредованной цитотоксичности в отношении клеток OVCAR5, индуцированную cx1547 при 3 нМ. cx1547 индуцировала Т-клеточно-опосредованную цитотоксичность только в антиген-положительных клеточных линиях. Цитотоксичность контролировали с использованием флуорогенного субстрата каспазы-3/7 дифференцированно меченных клеток-мишеней на Incucyte ZOOM. В этом анализе соотношение эффекторных клеток и клеток-мишеней (E:T) оценивалось в 20:1 и 10:1.
[0141] ФИГ. 29A-29F изображают кинетику Т-клеточно-опосредованной цитотоксичности, обусловленной репрезентативными B7H3-нацеленными ограниченными CD3-рекрутирующими конструкциями, и альтернативный формат DART-Fc с нацеливанием на B7H3 и CD3. Диапазон титрования от 50 нМ до 80 пМ CD3-рекрутирующих конструкций, на B7H3-положительной клеточной линии A375 показан на ФИГ. 29А-29Е. ФИГ. 29F изображает измерение при 50 нМ каждой конструкции на клетках A549, в которых экспрессия B7H3 была подвергнута нокдауну. Примечательно, что все конструкции демонстрируют B7H3-зависимую T-клеточно-опосредованную цитотоксичность.
[0142] ФИГ. 30 изображает величину индуцированной T-клеточно-опосредованной цитотоксичности с помощью 2,5 нМ B7H3-нацеленных ограниченных CD3-рекрутирующих конструкций и формата DART-Fc B7H3 x CD3 на антиген-положительных (A375) и отрицательных (A549-B7H3 нокдаун) клеточных линиях.
[0143] ФИГ. 31A-31F изображают сравнительную эффективность двух форматов, FRa-нацеленных CD3-рекрутеров, при индукции Т-клеточно-опосредованной цитотоксичности в отношении FRα-положительных клеток Ovcar-5 (ФИГ. 31A-31E) и FRα-отрицательных клеток NCI-H60 (ФИГ. 31F). cx2190 является репрезентативным C-концевым продуктом, который получают при протеолитической обработке гранзимом B cx1762. В частности, cx2190 демонстрирует более высокую эффективность по сравнению с cx1792, демонстрируя значительное усиление связывания CD3, опосредованного протеолизом в области линкера между Fc- и CD3-связывающими доменами. Кинетика Т-клеточно-опосредованной цитотоксичности в отношении FRα-положительных клеток с помощью 20 нМ, 32 пМ и 6 пМ изображена на ФИГ. 31А, 31В и 31С, соответственно. ФИГ. 31D и ФИГ. 31E демонстрируют активность двух форматов FRα CD3-рекрутеров через 24 и 40 часов соответственно. График F. демонстрирует, что никакая существенная цитотоксичность не опосредована какой-либо конструкцией в отсутствие экспрессии FRα на клетке-мишени.
[0144] ФИГ. 32 изображает Т-клеточно-опосредованную цитотоксичность, опосредованную репрезентативной 5T4-нацеленной ограниченной CD3-рекрутирующей конструкицей, cx3315. cx3315 индуцировала специфическую цитотоксичность Т-клеток в отношении клеточной линии, экспрессирующей 5t4, Ovcar-5, но не в отношении 5T4-отрицательной клеточной линии, CCRF-CEM. В этом анализе использовали 20 нМ cx3315.
[0145] ФИГ. 33 представляет собой график, демонстрирующий активацию Т-клеток после 20-часового совместного культивирования Т-клеток и клеток Ovcar5 в присутствии расщепленной или нерасщепленной cx309. Только расщепленная cx309 была способна опосредовать активацию FRα-зависимой активации Т-клеток посредством связывания CD3. Активацию Т-клеток контролировали с помощью анализа проточной цитометрии CD25% популяций CD4 и CD8.
[0146] ФИГ. 34A-34H изображают способность активировать CD4 (ФИГ. 34A и 34E) и CD8 (ФИГ. 34C и 34G) Т-клетки в зависимости от мишени с помощью репрезентативных B7H3-нацеленных ограниченных CD3-рекрутирующих конструкций и альтернативного формата DART-Fc с нацеливанием на B7H3 и CD3. Т-клетки инкубировали с B7H3-положительной клеточной линией A375 (фиг. 34A, 34C, 34E, 34G) или с клеточной линией A549, подвергнутой нокдауну по B7H3 (ФИГ. 34B, 34D, 34F, 34H), и маркеры активации CD25 и CD71 оценивали с помощью проточной цитометрии. Эти данные демонстрируют B7H3-зависимую способность активации Т-клеток используемых конструкций.
[0147] ФИГ. 35 изображает способность B7H3-нацеленной ограниченной CD3-рекрутирующей конструкции, cx3095, опосредовать антиген-зависимое продуцирование INFγ. Продуцирование цитокинов определяли с помощью ИФА INFγ. A375 использовали в качестве B7H3-положительной клеточной линии, тогда как клеточную линию A549 с нокдауном B7H3 использовали в качестве отрицательной клеточной линии.
[0148] ФИГ. 36A-36B изображают способность типичной FRα-нацеленной ограниченной CD3-рекрутирующей конструкции, cx1547, индуцировать FRα-зависимый IFNγ (ФИГ. 36A) и IL-2 (ФИГ. 36B) из PBMC человека. Продуцирование цитокинов измеряли с использованием анализа захвата цитокинов FluoroSpot. IGROV-1 и NCI-H460 использовали в качестве FRα-положительных и отрицательных клеточных линий, соответственно.
[0149] ФИГ. 37 изображает способность B7H3-нацеленной ограниченной CD3-рекрутирующей конструкции, cx3095, опосредовать антиген-зависимое продуцирование INFγ. Продуцирование цитокинов контролировали с использованием анализа FluoroSpot. Клеточные линии A375 и CCRF-CEM использовали в качестве B7H3-положительных и отрицательных клеточных линий, соответственно.
[0150] ФИГ. 38A-38D изображают способность FRα-нацеленной ограниченной CD3-конструкции активировать Т-клетки, присутствующие в диссоциированном образце первичной опухоли яичника человека, и вызывать цитотоксичность. ФИГ. 38A изображает блок-схему относительной распространенности опухолевых клеток (EpCAM+) и инфильтрирующих лимфоцитов (CD45+) в образце диссоциированной опухоли яичника. ФИГ. 38B демонстрирует жизнеспособность (CellTiterGlo) адгезивных опухолевых клеток после обработки обычным антителом FRα или cx1547 после 6-дневной инкубации. ФИГ. 38C демонстрирует продуцирование INFγ после обработки антителом FRα или cx1547 после 6-дневной инкубации. ФИГ. 38D демонстрирует репрезентативные изображения оставшихся адгезивных опухолевых клеток после 6-дневной обработки без антител (слева), обычным антителом против FR (в центре) или cx1547 (справа).
ПОДРОБНОЕ ОПИСАНИЕ
[0151] Настоящее раскрытие относится к ограниченным рекрутирующим Т-клетки слитым белкам в форме полиспецифических полипептидных конструкций, которые связываются по меньшей мере с CD3 и со вторым антигеном. Полиспецифические полипептидные конструкции, представленные в настоящем описании, включают, по меньшей мере, первый компонент, который включает одну или более копий антигенсвязывающего домена, который связывается с антигеном, связанным с Fc-областью иммуноглобулина, второй компонент, который включает одну или более копий, по меньшей мере, связывающего домена, который связывается с CD3 (называемый здесь как анти-CD3-связывающий домен или CD3-связывающая область, которые используются в настоящем описании взаимозаменяемо), и линкер, такой как полипептидный линкер, который соединяет первый компонент и второй компонент. В некоторых вариантах осуществления антиген представляет собой опухолеспецифический антиген (TAA). В некоторых вариантах осуществления линкер представляет собой расщепляемый линкер.
[0152] Представленные полиспецифичные полипептидные конструкции включают конфигурацию, в которой первый компонент, содержащий Fc-область, является N-концевым относительно второго компонента, содержащего CD3-связывающую область. В таком варианте осуществления первый и второй компоненты соединены через линкер, который является C-концевым относительно конца Fc-области. В некоторых вариантах осуществления антигенсвязывающий домен(ы) расположен на аминоконцевой (N-конец) области полиспецифической полипептидной конструкции. В некоторых вариантах осуществления антигенсвязывающий домен(ы) расположен на карбоксиконцевой (C-конец) области полиспецифической полипептидной конструкции. В некоторых вариантах осуществления антигенсвязывающий домен(ы) расположен как на N-, так и на C-концевых областях полиспецифической полипептидной конструкции. Различные конфигурации полиспецифической полипептидной конструкции, представленные в настоящем документе, показаны на ФИГ. 1.
[0153] Представленные полиспецифические полипептидные конструкции проявляют ограниченную активность рекрутирования Т-клеток, потому что такие конструкции по существу связываются с CD3 только после того, как антиген связывается через антигенсвязывающий домен. Это иллюстрируется Примерами и Фигурами, представленными в настоящем документе, которые демонстрируют способность ограниченных CD3-рекрутирующих белков эффективно связывать TAA-положительные клетки, при этом практически не связываясь с Т-клетками. Это уникальное свойство позволяет ограниченным CD3-рекрутирующим белкам распространяться в сайты, где присутствует TAA, без связывания с периферическими Т-клетками. Этот формат отличается от других полиспецифических конструкций, включающих CD3, тем, что связывание конститутивного CD3 запрещено или исключено, что обеспечивает значительное преимущество за счет исключения связывания периферических Т-клеток и обеспечения превосходного распределения в сайт(ы), где присутствует антиген, распознаваемый антигенсвязывающим доменом. Например, как показано в Примерах, формат ограниченного CD3-рекрутирования обеспечивает эффективность, аналогичную формату DART-Fc (например, опубликованная заявка PCT. WO2017/030926), однако связывание с периферическими Т-клетками значительно ослаблено. Кроме того, другие CD3-рекрутирующие конструкции опосредуют антиген-зависимую активацию Т-клеток, однако представленные в настоящем описании полиспецифические полипептидные конструкции опосредуют как антиген-зависимое связывание Т-клеток, так и их активацию.
[0154] Ограниченная активность рекрутирования Т-клеток представленных полиспецифических полипептидных конструкций обусловлена, в некоторых аспектах, расположением Fc-области на N-конце относительно CD3-связывающей области. В некоторых вариантах осуществления такое расположение уменьшает, ослабляет, демпфирует и/или предотвращает связывание CD3 с CD3-связывающей областью. В отсутствие связывания антигена антигенсвязывающим доменом представленные в настоящем описании полиспецифические полипептидные конструкции демонстрируют сниженную или устраненную CD3-связывающую способность и способность активации Т-клеток. В некоторых вариантах осуществления в присутствии антигенсвязывающего события, опосредованного антигенсвязывающим доменом(доменами) полиспецифических полипептидных конструкций, способность связываться с CD3 CD3-связывающей области значительно повышается. В некоторых вариантах осуществления в присутствии антигенсвязывающего события, опосредованного антигенсвязывающими доменами(доменами) полиспецифических полипептидных конструкций, способность активировать Т-клетки значительно повышается. Рекрутирование его родственного антигена антигенсвязывающим доменом (ами) внутри полиспецифической полипептидной конструкции приводит к последующему рекрутированию Т-клеток и опосредует антиген-зависимую активацию Т-клеток, такую как цитотоксичность, высвобождение цитокинов, дегрануляция и пролиферация. В некоторых вариантах осуществления представленные полиспецифические полипептидные конструкции могут быть использованы для усиления иммунного ответа, такого как усиление активности Т-клеток, включая цитолитическую (или цитотоксическую) активность Т-клеток. Модуляция иммунного ответа может, в некоторых аспектах, способствовать лечению заболевания или состояния у пациента.
[0155] В некоторых вариантах осуществления один или более антигенсвязывающих доменов связываются с антигеном на опухолевой клетке или на клетке микроокружения опухоли. В некоторых аспектах представленные полиспецифические полипептидные конструкции могут быть использованы для усиления иммунных ответов, таких как активность Т-клеток, например цитотоксическая активность, против опухоли или злокачественного новообразования. В некоторых вариантах осуществления представленные полиспецифические полипептидные конструкции можно использовать для лечения опухоли или злокачественного новообразования у пациента.
[0156] Полиспецифические полипептидные конструкции по настоящему изобретению гарантируют, что не будет связывания Т-клеток через CD3 в периферической крови, поскольку CD3-связывающая область этих конструкций ограничена или заблокирована иным образом и/или ингибирована присутствием Fc-области. Таким образом, полиспецифические полипептидные конструкции по настоящему изобретению обеспечивают ряд преимуществ. В некоторых аспектах эти конструкции ограничивают эффект поглощения, вызванный связыванием всех Т-клеток. В некоторых аспектах эти конструкции снижают системную токсичность.
[0157] В некоторых вариантах осуществления представленные полиспецифические полипептидные конструкции по настоящему изобретению обеспечивают контролируемое биораспределение в желаемом сайте у пациента, таком как, например, сайт экспрессии опухолеспецифического антигена (TAA). Сайты экспрессии TAA включают, например, опухоль и микроокружение опухоли.
[0158] В некоторых вариантах осуществления полиспецифические полипептидные конструкции по настоящему изобретению проявляют специфичность в отношении CD3 и одного или более других антигенов. В некоторых вариантах осуществления полиспецифические полипептидные конструкции могут содержать более чем один антигенсвязывающий домен, способный связываться с одним или более TAA, как например, с 2, 3 или 4 антигенсвязывающими доменами, см., например, ФИГ. 1. В некоторых вариантах осуществления один или более антигенсвязывающих доменов связываются с одним и тем же антигеном. В некоторых вариантах осуществления полиспецифические полипептидные конструкции включают более чем один антигенсвязывающий домен, который связывается с различными эпитопами на одном и том же антигене. В некоторых вариантах осуществления полиспецифические полипептидные конструкции включают в себя более чем один антигенсвязывающий домен, который связывается с одним или более различных антигенов. В некоторых вариантах осуществления полиспецифические полипептидные конструкции включают более чем один антигенсвязывающий домен, который связывается с различными эпитопами на одних и тех же антигенах, а также включают дополнительные антигенсвязывающие домены, которые связываются с одним или более различными антигенами. В некоторых аспектах представленные полиспецифические полипептидные конструкции представляют собой биспецифические полипептидные конструкции, так что они способны связываться с CD3 и другим антигеном, таким как TAA, посредством связывания антигенсвязывающего домена полиспецифической полипептидной конструкции. В некоторых примерах представленные полиспецифические полипептидные конструкции представляют собой биспецифические полипептидные конструкции, которые обеспечивают четырехвалентное связывание одного или более TAA посредством использования первого антигенсвязывающего домена и второго антигенсвязывающего домена. Например, в некоторых вариантах осуществления биспецифические полипептидные конструкции включают первое антигенсвязывающее однодоменное антитело (sdAb) и второе антигенсвязывающее sdAb, как показано на ФИГ. 1 и 2.
[0159] В некоторых вариантах осуществления полиспецифические полипептидные конструкции, представленные в настоящем описании, существуют в двух состояниях с точки зрения способности связываться с CD3 и впоследствии активировать Т-клетки: (1) «неактивное» состояние, т.е. нерасщепленное состояние, возникает, когда нет связывания какого-либо или всех антигенсвязывающих доменов, так что связывание CD3 ограничено и взаимодействие с Т-клетками исключено, и (2) «активное» состояние возникает при связывании антигена с каким-либо или со всеми антигенсвязывающими доменами, так что CD3-связывающая область способна связываться с CD3 и разрешено взаимодействие Т-клеток.
[0160] В некоторых вариантах осуществления Fc-область связана с CD3-связывающим доменом через линкер или линкеры. В некоторых вариантах осуществления Fc-область связана с CD3-связывающей областью через нерасщепляемый линкер или линкеры. В некоторых вариантах осуществления Fc-область связана с CD3-связывающей областью через расщепляемый линкер или иным образом лабильный линкер или линкеры.
[0161] В некоторых вариантах осуществления Fc-область и CD3-связывающая область связаны расщепляемым линкером. В некоторых аспектах усиленное связывание CD3 происходит после расщепления расщепляемого линкера. В некоторых таких аспектах «активное» состояние может быть дополнительно усилено посредством нескольких механизмов, в том числе посредством расщепления линкера, соединяющего CD3-связывающую область и Fc-область. В некоторых вариантах осуществления расщепляемый линкер представляет собой линкер, который содержит сайт распознавания субстрата для протеазы. В некоторых вариантах осуществления, в которых Fc-область и CD3-связывающая область связаны расщепляемым линкером, усиленное связывание CD3 может происходить после расщепления внутри линкера (линкеров).
[0162] В некоторых аспектах полиспецифические полипептидные конструкции по настоящему изобретению обеспечивают терапевтическую эффективность в отсутствие протеолиза.
[0163] В некоторых вариантах осуществления Fc-область представляет собой гомодимерную Fc-область. В некоторых вариантах осуществления Fc-область представляет собой гетеродимерную Fc-область. В некоторых вариантах осуществления Fc-область представляет собой мономерную Fc-область. В некоторых вариантах осуществления Fc-область полиспецифических полипептидных конструкций способна взаимодействовать с FcγR и опосредовать функции врожденного иммунного эффектора, например, антителозависимую клеточную токсичность (ADCC) и антителозависимый клеточный фагоцитоз (ADCP). В некоторых вариантах осуществления Fc-область полиспецифических полипептидных конструкций способна взаимодействовать с белками комплемента, а именно C1q, и опосредовать комплемент-зависимую цитотоксичность. Таким образом, в некоторых аспектах полиспецифические полипептидные конструкции по настоящему изобретению допускают множественные иммунные эффекторные механизмы, включая врожденные иммунные эффекторы и Т-клетки.
[0164] В некоторых вариантах осуществления, где Fc-область и CD3-связывающая область функционально связаны расщепляемым линкером, расщепление линкера (линкеров) между Fc-областью и CD3-связывающей областью может разделить полиспецифические полипептидные конструкции на первый и второй компонент. В зависимости от состава полиспецифической полипептидной конструкции первый и второй компоненты могут иметь разные функциональные возможности. В некоторых вариантах осуществления Fc-область представляет собой область, которая проявляет одну или более эффекторных функций, таких как функции ADCC, CDC или ADCP. В таких примерах полиспецифические полипептидные конструкции по изобретению могут быть использованы для получения самоусиливающейся системы. Например, полиспецифические конструкции могут быть использованы следующим образом: ADCC, опосредованная NK-клеткой после нацеливания TAA и связывания CD16 Fc-области, приводит к высвобождению гранзима B, который способен к внеклеточному протеолизу и расщеплению линкеров между первым и вторым компонентами полиспецифических полипептидных конструкций.
[0165] В некоторых вариантах осуществления линкер представляет собой расщепляемый линкер. Полиспецифические полипептидные конструкции обеспечивают терапевтическую составляющую «два в одном», имеющую двойные эффекторные функции, причем протеолитическая активация полиспецифических полипептидных конструкций продуцирует два компонента, каждый из которых обладает биологической активностью. Полиспецифические полипептидные конструкции по настоящему изобретению способны обеспечивать Fc-опосредованные эффекторные функции, такие как, например, ADCC (например, высвобождение гранзима B NK-клетками), ADCP и/или CDC.
[0166] Предполагается, что конструкции, ограничивающие CD3, пригодны для использования с любым TAA-связывающим доменом, что позволяет лучше терапевтически воздействовать на опухоль или микроокружение опухоли, избегая взаимодействий с периферическими T-клетками и опосредуя мощную TAA-зависимую цитотоксичность T-клеток. Включение расщепляемого протеазой линкера между Fc и компонентами CD3-связывающего домена позволяет усиливать способность активации Т-клеток, обеспечивая полное экспонирование CD3-связывающего домена. В зависимости от конкретного включенного линкера стадия амплификация может быть опосредована опухолеспецифическими протеазами или гранзимами, высвобождаемыми после активации антиген-зависимых Т-клеток. Если включен линкер, расщепляемый опухолевой протеазой, амплификация опосредуется опухолью или микроокружением опухоли. Принимая во внимание, что если включен расщепляемый гранзимом В линкер, амплификация может быть самоопосредована Т-клетками после антиген-зависимой активации. Кроме того, в случаях, когда в конструкцию включен Fc с возможной функцией эффектора, амплификация может быть опосредована гранзимами, высвобождаемыми из NK-клетки, что происходит по механизму ADCC.
[0167] В некоторых вариантах осуществления протеаза представляет собой протеазу, которая продуцируется в микроокружении опухоли и/или при активации Т-клеток, индуцированной первоначальным связыванием CD3-связывающей области с CD3 в микроокружении опухоли посредством связывания антигенсвязывающего домена (доменов) с ТАА. В некоторых вариантах осуществления протеаза представляет собой гранзим B. В некоторых аспектах полиспецифические полипептидные конструкции по изобретению используют способность протеазы в микроокружении опухоли и/или гранзим B расщеплять линкер в полиспецифической полипептидной конструкции в положении ниже полипептида Fc иммуноглобулина, таким образом генерируя два терапевтически активных белка, в некоторых случаях, с отличным рекрутированием эффекторных клеток. В некоторых аспектах после расщепления расщепляемого линкера расщепленная первая часть или компонент сохраняет функции Fc-эффектора и двухвалентное нацеливание первого антигена, такого как, например, TAA, через первый антигенсвязывающий домен, и вторая часть или компонент сохраняет способность к рекрутированию Т-клеток, поскольку отделение CD3-связывающей области от Fc-области обеспечивает связывание CD3. Расщепленная вторая часть или компонент также, в некоторых случаях, сохраняет способность связываться с TAA, которая может соответствовать двухвалентному связыванию через второй антигенсвязывающий домен.
[0168] В некоторых вариантах осуществления вторая часть или компонент содержит CD3-связывающую область, которая является одновалентной по отношению к CD3, так что активация T-клетки не будет происходить, если не присутствует TAA. В некоторых аспектах, когда поливалентная полипептидная конструкция содержит расщепляемый линкер, расщепленная вторая часть или компонент обеспечивает ТАА-зависимую Т-клеточно-опосредованную цитотоксичность. В некоторых случаях расщепленная вторая часть или компонент обеспечивает отсутствие взаимодействия FcRn. Кроме того, расщепленная вторая часть или компонент будет достаточно малой по размеру, например, всего ~ 50 кДа, что обеспечит быстрый клиренс, если по какой-либо причине отщепленная вторая часть или компонент распределяется за пределами опухолевого участка после расщепления, и/или если она аберрантно расщепляется за пределами области опухоли.
[0169] В некоторых вариантах осуществления изобретения полиспецифические полипептидные конструкции по настоящему изобретению допускают одновременную цитотоксичность, опосредованную Т-клетками и NK-клетками. В некоторых случаях такая активность может проявляться в полиспецифической полипептидной конструкции, в которой содержится первый антигенсвязывающий домен, например, первый анти-ТАА-антигенсвязывающий домен, и второй антигенсвязывающий домен, например, второй анти-ТАА-антигенсвязывающее домен, который может нацеливаться на отдельные и/или неконкурентные эпитопы на данном TAA.
[0170] В некоторых аспектах полиспецифические полипептидные конструкции по настоящему изобретению обеспечивают ряд преимуществ по сравнению с современными биспецифическими терапевтическими средствами. Полиспецифические полипептидные конструкции по настоящему изобретению меньше, чем обычное терапевтическое антитело, например, 150 кДа против 125 кДа, что обеспечит лучшее нацеливание, например проникновение в опухоль. Во-первых, размер всей полиспецифической полипептидной конструкции обеспечивает длительный период полужизни для нерасщепленной конструкции, и после расщепления конструкции расщепленная вторая часть или компонент будет достаточно малым для обеспечения короткого периода полужизни. В некоторых аспектах полиспецифические полипептидные конструкции по изобретению демонстрируют пониженную системную токсичность или токсичность любой области вне опухоли и/или микроокружения опухоли, поскольку связывание CD3 с CD3-связывающей областью зависит от рекрутирования TAA до того, как произойдет рекрутирование CD3. В некоторых случаях включение расщепляемого линкера, специфичного к протеазе опухолевой среды, снижает связывание CD3 полиспецифическими конструкциями до протеолитической активации и рекрутирования TAA, тем самым умножая или усиливая рекрутирование CD3.
[0171] Полиспецифические полипептидные конструкции по изобретению предназначены для обеспечения того, чтобы протеаза, которая расщепляет расщепляемый линкер, не нуждалась в том, чтобы быть односторонне-опухолевой (например, не требовала различной экспрессии только в месте опухоли и/или в окружении опухоли). Скорее, эти полиспецифические полипептидные конструкции требуют только, чтобы протеаза присутствовала в том же месте, что и TAA. Валентность этих конструкций будет стимулировать биораспределение и удержание в опухоли и/или в микроокружении опухоли.
[0172] Все публикации и патентные документы, процитированные в настоящем описании, включены сюда посредством ссылки, как если бы каждая такая публикация или документ были специально и индивидуально указаны для включения в настоящее описание посредством ссылки. Цитирование публикаций и патентных документов не предназначено для признания того, что что-либо относится к предшествующему уровню техники, и не представляет собой какое-либо признание в отношении содержания или даты того же самого. Изобретение теперь описано посредством письменного описания, и специалисты в данной области техники поймут, что изобретение может быть применено на практике во множестве вариантов осуществления и что вышеприведенное описание и примеры ниже предназначены для иллюстрации, а не для ограничения формулы изобретения, следующей далее.
I.Определения
[173] Если не указано иное, научные и технические термины, используемые в связи с настоящим раскрытием, должны иметь значения, которые обычно понимаются специалистами в данной области техники. Термин, относящийся к объекту в «единственном числе» относится к одному или более объектам. Например, соединение относится к одному или более соединениям. Таким образом, термины, указанные в единственном числе, «один или более» и «по меньшей мере, один» могут использоваться взаимозаменяемо. Кроме того, если иное не требуется в контексте, термины в единственном числе включают множественное число, а термины во множественном числе включают единственное число. Как правило, номенклатуры, используемые в связи с методами и технологиями клеточной и тканевой культуры, молекулярной биологии и химии и гибридизации белков и олиго- или полинуклеотидов, описанных в настоящем документе, являются хорошо известными и широко используемыми в данной области. Стандартные методы используются для рекомбинантной ДНК, синтеза олигонуклеотидов, а также для культуры и трансформации тканей (например, электропорация, липофекция). Ферментативные реакции и методики очистки выполняются в соответствии со спецификациями производителя или так, как обычно осуществляются в данной области техники или как описано в настоящем документе. Вышеуказанные методики и процедуры обычно выполняются в соответствии с обычными способами, хорошо известными в данной области техники, и как описано в различных общих и более конкретных ссылках, которые цитируются и обсуждаются в настоящем описании. См. например, Sambrook et al.Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989)). Номенклатуры, используемые в связи с описанными в настоящем документе лабораторными процедурами и методами аналитической химии, синтетической органической химии и медицинской и фармацевтической химии, хорошо известны и широко используются в данной области. Стандартные методы используются для химического синтеза, химического анализа, фармацевтической подготовки, рецептуры и доставки и лечения пациентов.
[0174] Используемые в соответствии с настоящим раскрытием, следующие термины, если не указано иное, должны иметь следующие значения:
[0175] Используемый в настоящем описании термин «антитело» относится к молекулам иммуноглобулина и антигенсвязывающим частям молекул иммуноглобулина (Ig), то есть молекулам, которые содержат антигенсвязывающий сайт, который специфически связывается (иммунореагирует) с антигеном. Под «специфическим связыванием» или «иммунореактивностью с» или «иммуноспецифическим связыванием» подразумевается, что антитело реагирует с одной или более антигенными детерминантами желаемого антигена и не реагирует с другими полипептидами или связывается с гораздо более низкой аффинностью (Kd > 10-6). Антитела включают, но не ограничиваются ими, поликлональные, моноклональные, химерные, полностью человеческие, доменные антитела, одноцепочечные фрагменты, Fab и F(ab')2, Fvs, scFv и библиотеку экспрессии Fab.
[0176] Известно, что основная структурная единица антитела содержит тетрамер. Каждый тетрамер состоит из двух идентичных пар полипептидных цепей, каждая пара имеет одну «легкую» (примерно 25 кДа) и одну «тяжелую» цепь (примерно 50-70 кДа). Аминоконцевая часть каждой цепи включает вариабельную область примерно от 100 до 110 или более аминокислот, в первую очередь ответственную за распознавание антигена. Карбоксиконцевая часть каждой цепи определяет константную область, в первую очередь ответственную за эффекторную функцию. В общем, молекулы антител человеческого происхождения относятся к любому из классов IgG, IgM, IgA, IgE и IgD, которые отличаются друг от друга природой тяжелой цепи, присутствующей в молекуле. Некоторые классы также имеют подклассы, такие как IgG1, IgG2, IgG3, IgG4 и другие. Кроме того, у человека легкая цепь может представлять собой каппа-цепь или лямбда-цепь.
[0177] Используемый в настоящем описании термин «моноклональное антитело» (mAb) или «композиция моноклонального антитела» относится к популяции молекул антитела, которые содержат только один молекулярный вид молекулы антитела, состоящий из уникального продукта гена легкой цепи и уникального продукта гена тяжелой цепи. В частности, определяющие комплементарность области (CDR) моноклонального антитела идентичны во всех молекулах популяции. MAb содержат антигенсвязывающий сайт, способный иммунореагировать с конкретным эпитопом антигена, характеризующимся уникальной аффинностью связывания с ним.
[0178] Термин «антигенсвязывающий сайт» или «связывающая часть» относится к части молекулы иммуноглобулина, которая участвует в связывании антигена. Антигенсвязывающий сайт образован аминокислотными остатками N-концевых вариабельных («V») областей тяжелой («Н») и легкой («L») цепей. Три сильно расходящихся участка в V-областях тяжелой и легкой цепей называются «гипервариабельными областями», которые расположены между более консервативными фланкирующими участками, известными как «каркасные области» или «FR». Таким образом, термин «FR» относится к аминокислотным последовательностям, которые в природе присутствуют в иммуноглобулинах между гипервариабельными областями и рядом с ними. В молекуле антитела три гипервариабельные области легкой цепи и три гипервариабельные области тяжелой цепи располагаются относительно друг друга в трехмерном пространстве, с образованием антигенсвязывающей поверхности. Антигенсвязывающая поверхность комплементарна трехмерной поверхности связанного антигена, и три гипервариабельные области каждой из тяжелых и легких цепей обозначаются как «области, определяющие комплементарность» или «CDR». Присвоение аминокислот каждому домену соответствует определениям Kabat Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md. (1987 and 1991)), или Chothia & Lesk J. Mol. Biol. 196:901-917 (1987), Chothia et al. Nature 342:878-883 (1989).
[0179] Используемый в настоящем описании термин «эпитоп» включает любую конкретную часть антигена, на которую нацелено антитело, фрагмент антитела или другой связывающий домен. Термин «эпитоп» включает любую область белка, на которую направлено специфическое связывание. Термин «эпитоп» включает любую белковую детерминанту, способную специфически связываться с иммуноглобулином или Т-клеточным рецептором. Эпитопные детерминанты обычно состоят из химически активных поверхностных групп молекул, таких как боковые цепи аминокислот или сахаров, и обычно имеют специфические трехмерные структурные характеристики, а также специфические характеристики заряда. Например, антитела могут быть получены против N-концевых, центральных или С-концевых пептидов полипептида. Кроме того, антитела могут быть получены против линейных или прерывистых эпитопов полипептида. Считается, что антитело специфически связывается с антигеном, когда константа диссоциации составляет ≤ 1 мкМ; например, в некоторых вариантах осуществления ≤100 нМ, а в некоторых вариантах осуществления ≤10 нМ и не проявляет связывания с другими белками, как близкородственными, так и отдаленными.
[0180] Используемые в настоящем документе термины «специфическое связывание», «иммунологическое связывание» и «свойства иммунологического связывания» относятся к нековалентным взаимодействиям того типа, который происходит между молекулой иммуноглобулина и антигеном, для которого иммуноглобулин является специфическим. Сила или аффинность иммунологических связывающих взаимодействий могут быть выражены через константу диссоциации (Kd) взаимодействия, где меньшая Kd представляет большую аффинность. Иммунологические свойства связывания выбранных полипептидов могут быть определены количественно с использованием способов, хорошо известных в данной области. Один такой способ включает измерение скоростей образования и диссоциации антигенсвязывающего сайта/антигенного комплекса, причем эти скорости зависят от концентраций партнеров комплекса, аффинности взаимодействия и геометрических параметров, которые в равной степени влияют на скорость в обоих направлениях. Таким образом, как «скорость ассоциации» (Kon), так и «скорость диссоциации» (Koff) могут быть определены путем расчета концентраций и фактических скоростей ассоциации и диссоциации. (См. Nature 361: 186-87 (1993)). Отношение Koff/Kon позволяет аннулировать все параметры, не связанные с аффинностью, и равно константе диссоциации Kd. (См., в общем, Davies et al.(1990) Annual Rev Biochem 59: 439-473). Считается, что антитело по настоящему изобретению специфически связываются с EGFR, когда константа связывания (Кd)≤1 µМ, например, в некоторых вариантах осуществления≤100 нМ, в некоторых вариантах осуществления≤10 нМ, а в некоторых вариантах осуществления≤100-100 пМ, согласно измерению с помощью таких анализов, как анализы связывания меченого лиганда или аналогичных анализов, известных специалистам в данной области.
[0181] Используемый в настоящем описании термин «выделенный полинуклеотид» означает полинуклеотид геномного, кДНК или синтетического происхождения или некоторую их комбинацию, который в силу своего происхождения как «выделенный полинуклеотид» (1) не связан со всем или частью полинуклеотида, в котором «выделенный полинуклеотид» обнаружен в природе, (2) функционально связан с полинуклеотидом, с которым он не связан в природе, или (3) не встречается в природе как часть большей последовательности. Полинуклеотиды в соответствии с настоящим изобретением включают молекулы нуклеиновой кислоты, кодирующие молекулы иммуноглобулина тяжелой цепи, представленные в настоящем описании, и молекулы нуклеиновой кислоты, кодирующие молекулы иммуноглобулина легкой цепи, представленные в настоящем описании.
[0182] Термин «выделенный белок», упомянутый в настоящем описании, означает белок с происхождением из кДНК, рекомбинантной РНК или синтетического происхождения или некоторую их комбинацию, который в силу своего происхождения или источника происхождения как «выделенный белок» (1) не связан с белками, встречающимися в природе, (2) не содержит других белков из того же источника, например, не содержит мышиных белков, (3) экспрессируется клеткой другого вида или (4) не встречается в природе.
[0183] Термин «полипептид» используется в настоящем описании как общий термин для обозначения нативного белка, фрагментов или аналогов полипептидной последовательности. Следовательно, нативные белковые фрагменты и аналоги являются видами полипептидов. Полипептиды в соответствии с настоящим изобретением включают молекулы иммуноглобулина тяжелой цепи, представленные в настоящем описании, и молекулы иммуноглобулина легкой цепи, представленные в настоящем описании, а также молекулы антитела, образованные комбинациями, содержащими молекулы иммуноглобулина тяжелой цепи с молекулами иммуноглобулина легкой цепи, такими как молекулы иммуноглобулина с легкой цепью каппа, и наоборот, а также их фрагменты и аналоги.
[0184] Используемый в настоящем описании термин «встречающийся в природе» применительно к объекту относится к тому факту, что объект можно обнаружить в природе. Например, полипептидная или полинуклеотидная последовательность, которая присутствует в организме (включая вирусы), которая может быть выделена из источника в природе и которая не была преднамеренно изменена человеком в лаборатории или иным образом, и которая встречается в природе.
[0185] Используемый в настоящем описании термин «функционально связанный» относится к положениям компонентов, которые описаны таким образом, что позволяют им функционировать по назначению. Контрольная последовательность, «функционально связанная» с кодирующей последовательностью, лигируется таким образом, что экспрессия кодирующей последовательности достигается в условиях, совместимых с контрольными последовательностями.
[0186] Используемый в настоящем описании термин «контрольная последовательность» относится к полинуклеотидным последовательностям, которые необходимы для осуществления экспрессии и процессирования кодирующих последовательностей, с которыми они лигированы. Природа таких контрольных последовательностей различается в зависимости от организма хозяина у прокариот, такие контрольные последовательности обычно включают промотор, сайт связывания рибосомы и последовательность терминации транскрипции, у эукариот, как правило, такие контрольные последовательности включают промоторы и последовательность терминации транскрипции. Предполагается, что термин «контрольные последовательности» включает, как минимум, все компоненты, присутствие которых является существенным для экспрессии и процессирования, и может также включать дополнительные компоненты, присутствие которых является выгодным, например, лидерные последовательности и последовательности партнеров по слиянию. Термин «полинуклеотид» в контексте настоящего описания означает нуклеотиды длиной по меньшей мере 10 оснований, либо рибонуклеотиды, либо дезоксинуклеотиды, либо модифицированные формы нуклеотидов любого типа. Термин включает одноцепочечные и двухцепочечные формы ДНК.
[0187] Термин «олигонуклеотид», упомянутый в настоящем описании, включает встречающиеся в природе и модифицированные нуклеотиды, связанные вместе встречающимися в природе и не встречающимися в природе олигонуклеотидными связями. Олигонуклеотиды представляют собой подмножество полинуклеотиднов, обычно содержащие длину 200 оснований или менее. В некоторых вариантах осуществления длина олигонуклеотидов составляет от 10 до 60 оснований, например, в некоторых вариантах осуществления длина составляет 12, 13, 14, 15, 16, 17, 18, 19 или от 20 до 40 оснований. Олигонуклеотиды обычно являются одноцепочечными, например, для зондов, хотя олигонуклеотиды могут быть двухцепочечными, например, для использования в конструировании мутанта гена. Олигонуклеотиды по изобретению представляют собой смысловые или антисмысловые олигонуклеотиды.
[0188] Термин «встречающиеся в природе нуклеотиды», упомянутый в настоящем описании, включает дезоксирибонуклеотиды и рибонуклеотиды. Термин «модифицированные нуклеотиды», упомянутый в настоящем описании, включает нуклеотиды с модифицированными или замещенными сахарными группами и тому подобное. Термин «олигонуклеотидные связи», упомянутый в настоящем описании, включает олигонуклеотидные связи, такие как фосфоротиоатные, фосфородитиоатные, фосфороселеноатные, фосфородиселеноатные, фосфороанилотиоатные, фосфораниладатные, фосфороамидатные и тому подобное. См. например, LaPlanche et al.Nucl. Acids Res. 14: 9081 (1986); Stec et al. J. Am. Chem. Soc. 106:6077 (1984), Stein et al. Nucl. Acids Res. 16: 3209 (1988), Zon et al. Anti Cancer Drug Design 6:539 (1991); Zon et al. Oligonucleotides and Analogues: A Practical Approach, pp. 87-108 (F. Eckstein, Ed., Oxford University Press, Oxford England (1991)); Stec et al. Патент США № 5151510; Uhlmann and Peyman Chemical Reviews 90: 543 (1990). При желании олигонуклеотид может включать метку для детектирования.
[0189] Используемые в настоящем описании двадцать обычных аминокислот и их аббревиатуры следуют общепринятому использованию. См. Immunology - A Synthesis (2nd Edition, ES Golub и DR Gren, Eds., Sinauer Associates, Sunderland7 Mass. (1991)). Стереоизомеры (например, D-аминокислоты) из двадцати стандартных аминокислот, неприродные аминокислоты, такие как α-,α-дизамещенные аминокислоты, N-алкиламинокислоты, молочная кислота и другие нестандартные аминокислоты, также могут быть подходящими компонентами для полипептидов настоящего раскрытия. Примеры нестандартных аминокислот включают: 4-гидроксипролин, γ-карбоксиглутамат, ε-N, N,N-триметиллизин, ε-N-ацетиллизин, O-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, σ-N-метиларгинин и другие подобные аминокислоты и иминокислоты (например, 4-гидроксипролин). В обозначении полипептида, используемом в настоящем описании, левое направление является аминоконцевым направлением, а правое направление является карбоксиконцевым направлением в соответствии со стандартным использованием и соглашением.
[0190] Аналогично, если не указано иное, левый конец одноцепочечных полинуклеотидных последовательностей является 5'-концом, а левый конец двухцепочечных полинуклеотидных последовательностей называется 5'-направлением. Направление 5'-3' добавления образующихся РНК-транскриптов называется областями последовательности направления транскрипции на цепи ДНК, имеющей ту же последовательность, что и РНК, и те, которые находятся в 5'-области относительно 5'-конца РНК-транскрипта, называются «последовательностями, расположенными выше», а области последовательности на цепи ДНК, имеющие ту же последовательность, что и РНК, и те, которые находятся в 3'-области относительно 3'-конца транскрипта РНК, называются «последовательностями, расположенными ниже».
[0191] Применительно к полипептидам термин «существенная идентичность» означает, что две пептидные последовательности, когда они оптимально выровнены, например, с помощью программ GAP или BESTFIT с использованием весов пробелов по умолчанию, имеют, по меньшей мере, 80 процентов идентичности последовательностей, например, в некоторых вариантах осуществления по меньшей мере, 90 процентов идентичности последовательности, в некоторых вариантах осуществления, по меньшей мере, 95 процентов идентичности последовательности, а в некоторых вариантах осуществления, по меньшей мере, 99 процентов идентичности последовательности.
[0192] В некоторых вариантах осуществления положения остатков, которые не являются идентичными, отличаются консервативными аминокислотными заменами.
[0193] Как обсуждается в настоящем описании, небольшие вариации в аминокислотных последовательностях антител или молекул иммуноглобулина рассматриваются как охваченные настоящим раскрытием, при условии, что вариации в аминокислотной последовательности сохраняются, по меньшей мере, на 75%, например, в некоторых вариантах, по меньшей мере, 80%, 90%, 95%, а в некоторых вариантах 99%. В частности, предусматриваются консервативные замены аминокислот. Консервативными заменами являются те, которые происходят в семействе аминокислот, которые являются родственными по их боковым цепям. Генетически кодируемые аминокислоты обычно делятся на семейства: (1) кислые аминокислоты представляют собой аспартат, глутамат; (2) основные аминокислоты представляют собой лизин, аргинин, гистидин; (3) неполярные аминокислоты представляют собой аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан, и (4) незаряженные полярные аминокислоты представляют собой глицин, аспарагин, глутамин, цистеин, серин, треонин, тирозин. Гидрофильные аминокислоты включают аргинин, аспарагин, аспартат, глутамин, глутамат, гистидин, лизин, серин и треонин. Гидрофобные аминокислоты включают аланин, цистеин, изолейцин, лейцин, метионин, фенилаланин, пролин, триптофан, тирозин и валин. Другие семейства аминокислот включают (i) серин и треонин, которые являются алифатически-гидрокси-семейством; (ii) аспарагин и глутамин, которые являются амидсодержащим семейством; (iii) аланин, валин, лейцин и изолейцин, которые являются алифатическим семейством; и (iv) фенилаланин, триптофан и тирозин, которые являются ароматическим семейством. Например, разумно ожидать, что отдельная замена лейцина на изолейцин или валин, аспартата на глутамат, треонина на серин или аналогичная замена аминокислоты структурно родственной аминокислотой не будет иметь основного влияния на связывание или свойства получаемой молекулы, особенно если замена не включает аминокислоту в каркасном сайте. Приводит ли замена аминокислоты к функциональному пептиду, может быть легко определено путем анализа специфической активности производного полипептида. Анализы подробно описаны в настоящем документе. Фрагменты или аналоги антител или молекул иммуноглобулина могут быть легко получены специалистами в данной области. В некоторых вариантах осуществления амино- и карбоксиконцы фрагментов или аналогов встречаются вблизи границ функциональных доменов. Структурные и функциональные домены могут быть идентифицированы путем сравнения данных нуклеотидных и/или аминокислотных последовательностей с общедоступными или частными базами данных последовательностей. Компьютерные методы сравнения используются для идентификации мотивов последовательности или предсказанных доменов конформации белка, которые встречаются в других белках с известной структурой и/или функцией. Методы идентификации последовательностей белков, которые складываются в известную трехмерную структуру, известны. Bowie et al. Science 253: 164 (1991). Таким образом, вышеприведенные примеры демонстрируют, что специалисты в данной области техники могут распознавать мотивы последовательности и структурные конформации, которые могут использоваться для определения структурных и функциональных доменов в соответствии с раскрытием.
[0194] В некоторых вариантах осуществления аминокислотные замены представляют собой такие, которые: (1) уменьшают чувствительность к протеолизу, (2) уменьшают чувствительность к окислению, (3) изменяют аффинность связывания для образования белковых комплексов, (4) изменяют аффинность связывания и (4) обеспечивают или модифицируют другие физико-химические или функциональные свойства таких аналогов. Аналоги могут включать различные мутеины последовательности, отличной от встречающейся в природе пептидной последовательности. Например, единичные или множественные аминокислотные замены (например, консервативные аминокислотные замены) могут быть сделаны в последовательности, встречающейся в природе, например, в части полипептида вне домена (доменов), образующего межмолекулярные контакты. Консервативная аминокислотная замена не должна существенно изменять структурные характеристики родительской последовательности (например, замещающая аминокислота не должна стремиться разрушить спираль, которая встречается в родительской последовательности, или нарушить другие типы вторичной структуры, которая характеризует родительскую последовательность). Примеры известных в данной области полипептидных вторичных и третичных структур описаны в Proteins, Structures and Molecular Principles (Creighton, Ed., W. H. Freeman and Company, New York (1984)); Introduction to Protein Structure (C. Branden and J. Tooze, eds., Garland Publishing, New York, N.Y. (1991)); and Thornton et at. Nature 354:105 (1991).
[0195] Используемый в настоящем описании термин «полипептидный фрагмент» относится к полипептиду, который имеет аминоконцевую и/или карбоксиконцевую делецию и/или одну или более внутренних делеций, но где оставшаяся аминокислотная последовательность идентична соответствующим положениям в последовательности, встречающейся в природе, выведенной, например, из полной последовательности кДНК. Фрагменты обычно имеют длину, по меньшей мере, 5, 6, 8 или 10 аминокислот, например, в некоторых вариантах осуществления, длину, по меньшей мере, 14 аминокислот, в некоторых вариантах осуществления, длину, по меньшей мере, 20 аминокислот, обычно, длину, по меньшей мере, 50 аминокислот, и в некоторых вариантах осуществления, по меньшей мере, 70 аминокислот. Используемый в настоящем описании термин «аналог» относится к полипептидам, которые состоят из сегмента, по меньшей мере, из 25 аминокислот, который имеет существенную идентичность части выведенной аминокислотной последовательности и который специфически связывается с EGFR в подходящих условиях связывания. Обычно полипептидные аналоги содержат консервативную аминокислотную замену (или добавление или делецию) относительно встречающейся в природе последовательности. Аналоги обычно имеют длину, по меньшей мере, 20 аминокислот, например, в некоторых вариантах осуществления, длину, по меньшей мере, 50 аминокислот или более, и часто могут быть такими же длинными, как и полноразмерный природный полипептид.
[0196] Термин «агент» используется в настоящем описании для обозначения химического соединения, смеси химических соединений, биологической макромолекулы или экстракта, изготовленного из биологических материалов.
[0197] Используемый в настоящем описании термин «метка» или «меченный» относится к включению детектируемого маркера, например, путем включения радиоактивно меченной аминокислоты или присоединения к полипептиду биотинильных фрагментов, которые могут детектироваться с помощью маркированного авидина (например, стрептавидина, содержащего флуоресцентный маркер, или ферментативной активности, которая может детектироваться оптическими или калориметрическими методами). В определенных ситуациях метка или маркер также могут быть терапевтическими. Различные способы мечения полипептидов и гликопротеинов известны в данной области и могут быть использованы. Примеры меток для полипептидов включают, но не ограничиваются ими, следующие: радиоизотопы или радионуклиды (например, 3H, 14C, 15N, 35S, 90Y, 99Tc, 111In, 125I, 131I), флуоресцентные метки (например, флуорофор, родамин, лантаноидные люминофоры), ферментные метки (например, пероксидаза хрена, β-галактозидаза, люцифераза, щелочная фосфатаза), хемилюминесцентные, биотинильные группы, заранее определенные полипептидные эпитопы, распознаваемые вторичным репортером (например, последовательности пар лейциновой застежки, сайты связывания для вторичных антител, домены, связывающие металлы, фрагменты эпитопов). В некоторых вариантах осуществления метки прикрепляются с помощью спейсерных плечей различной длины, чтобы уменьшить потенциальное стерическое препятствие. Используемый в настоящем описании термин «фармацевтический агент или лекарственное средство» относится к химическому соединению или композиции, способным вызывать желаемый терапевтический эффект при правильном введении пациенту.
[0198] Используемый в настоящем описании термин «по существу чистый» означает, что целевое соединение представляет собой преобладающее присутствующее соединение (т.е. на молярной основе оно является более распространенным, чем любое другое отдельное соединение в композиции), и по существу очищенная фракция представляет собой композицию, в которой целевое соединение содержит по меньшей мере примерно 50 процентов (в молярном отношении) всех присутствующих высокомолекулярных соединений.
[0199] Как правило, по существу чистая композиция будет содержать более чем примерно 80 процентов всех высокомолекулярных соединений, присутствующих в композиции, например, в некоторых вариантах осуществления более чем примерно 85%, 90%, 95% и 99%. В некоторых вариантах осуществления целевое соединение очищено до существенной гомогенности (загрязняющие вещества не могут детектироваться в композиции обычными способами детектирования), где композиция состоит по существу из одного высокомолекулярно соединения.
[0200] Термин «пациент» включает человека и объекты ветеринарии.
[0201] Другие химические термины в настоящем описании используются в соответствии с общепринятым применением в данной области техники, примером чего является Словарь химических терминов McGraw-Hill (Parker S., Ed., McGraw-Hill, San Francisco (1985)).
II. Полиспецифические полипептидные конструкции
[0202] Настоящее изобретение относится к полиспецифической полипептидной конструкции, содержащей первый компонент, содержащий Fc-область иммуноглобулина, и второй компонент, содержащий CD3-связывающую область, где первый и второй компоненты связаны линкером, где Fc-область расположена на N-конце относительно CD3-связывающей области; и один или оба из первого и второго компонентов содержат антигенсвязывающий домен, который связывается с опухолесипецифическим антигеном (TAA).
[0203] В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит по порядку от N-конца к C-концу: Fc-область иммуноглобулина; линкер; CD3-связывающую область, которая связывается с CD3 (CD3ε и антигенсвязывающий домен, который связывается с опухолесипецифическим антигеном (TAA). В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит по порядку от N-конца к C-концу: антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA); Fc-область иммуноглобулина; линкер; и CD3-связывающую область, которая связывается с CD3 (CD3ε). В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит, по меньшей мере, первый антигенсвязывающий домен, который связывается с TAA, и второй антигенсвязывающий домен, который связывается с TAA. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит по порядку от N-конца к C-концу: первый антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA); Fc-область иммуноглобулина; линкер; CD3-связывающую область, которая связывается с CD3 (CD3ε и второй антигенсвязывающий домен, который связывается с опухолесипецифическим антигеном (TAA).
[0204] Каждый из компонентов полиспецифических полипептидных конструкций по изобретению более подробно описан ниже.
1.Анти-CD3-связывающие домены:
[0205] Полиспецифические полипептидные конструкции по настоящему изобретению включают одну или более копий анти-CD3-связывающего домена. Анти-CD3-связывающие домены по изобретению активируют Т-клетки посредством рекрутирования CD3ε на Т-клетки. Анти-CD3-связывающие домены по настоящему изобретению выступают агонистами, стимулируют, активируют и/или иным образом усиливают CD3-опосредованную активацию Т-клеток. Биологические активности CD3 включают, например, активацию T-клеток и другие сигналы посредством взаимодействия между CD3 и антигенсвязывающими субъединицами T-клеточного рецептора (TCR). Например, анти-CD3-связывающие домены по изобретению полностью или частично активируют Т-клетки посредством рекрутирования CD3ε на Т-клетки путем частичной или полной модуляции, например, выступая агонистом, стимулируя, активируя или иным образом усиливая опосредованную CD3 активацию Т-клеток.
[0206] В предпочтительных вариантах осуществления анти-CD3-связывающие домены по изобретению специфически связываются с эпсилон-цепью CD3, также известной как CD3ε. Анти-CD3ε-связывающие домены по изобретению активируют Т-клетки посредством рекрутирования CD3ε на Т-клетки. Анти-CD3ε-связывающие домены по изобретению включают моноклональные антитела, такие как, например, моноклональные антитела млекопитающих, моноклональные антитела приматов, полностью человеческие моноклональные антитела, а также гуманизированные моноклональные антитела и химерные антитела, а также их антигенсвязывающие фрагменты. В некоторых вариантах осуществления анти- CD3ε-связывающий домен включает одну или более копий антитела или его антигенсвязывающего фрагмента.
[0207] В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает одну или более копий антитела или его антигенсвязывающего фрагмента, выбранного из группы, состоящей из Fab-фрагмента, F(ab')2-фрагмента, Fv-фрагмента, scFv, scAb, dAb, однодоменного антитела тяжелой цепи и однодоменного антитела легкой цепи. В некоторых вариантах осуществления анти-CD3-связывающий домен содержит Fv-фрагмент антитела, который связывается с CD3ε (называемый в настоящем описании как анти-CD3ε Fv-фрагмент). В некоторых вариантах осуществления анти-CD3ε Fv-фрагмент антитела является стабилизированный дисульфидом анти-CD3-связывающий Fv-фрагмент (dsFv). В некоторых вариантах осуществления анти-CD3-связывающий домен является одновалентным для связывания CD3.
[0208] В некоторых вариантах осуществления его анти-CD3ε-связывающий домен включает вариабельную область тяжелой цепи (Hv), содержащую аминокислотную последовательность SEQ ID NO: 14. В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает вариабельную область легкой цепи (Lv), содержащую аминокислотную последовательность SEQ ID NO: 15. В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает вариабельную область тяжелой цепи (Hv), содержащую аминокислотную последовательность SEQ ID NO: 14, и вариабельную область легкой цепи (Lv), содержащую аминокислотную последовательность SEQ ID NO: 15. В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает вариабельную область тяжелой цепи (Hv), содержащую аминокислотную последовательность SEQ ID NO: 44. В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает вариабельную область легкой цепи (Lv), содержащую аминокислотную последовательность SEQ ID NO: 72. В некоторых вариантах осуществления их анти-CD3ε-связывающий домен включает вариабельную область тяжелой цепи (Hv), содержащую аминокислотную последовательность SEQ ID NO: 44, и вариабельную область легкой цепи (Lv), содержащего аминокислотную последовательность SEQ ID NO: 72. В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает комбинацию аминокислотной последовательности вариабельной области тяжелой цепи и аминокислотной последовательности вариабельной области легкой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 32-81. В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает комбинацию аминокислотной последовательности вариабельной области тяжелой цепи, выбранной из группы, состоящей из SEQ ID NO: 32-62, и аминокислотной последовательности вариабельной области легкой цепи, содержащей аминокислотную последовательность, выбранный из группы SEQ ID NO: 63-81.
[0209] В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает вариабельную область тяжелой цепи (Hv), содержащую аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 14.В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает вариабельную область легкой цепи (Lv), содержащую аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 15. В некоторых вариантах осуществления их анти-CD3ε-связывающий домен включает вариабельную область тяжелой цепи (Hv), содержащую аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 14, и вариабельную область легкой цепи (Lv), содержащую аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93% 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 15.
[0210] В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает вариабельную область тяжелой цепи (Hv), содержащую аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 44. В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает вариабельную область легкой цепи (Lv), содержащую аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 72. В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает вариабельную область тяжелой цепи (Hv), содержащую аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 44, и вариабельную область легкой цепи (Lv), содержащую аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93% 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 72.
[0211] В некоторых вариантах осуществления анти-CD3ε Fv-фрагмент антитела содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 32-81. В некоторых вариантах осуществления анти-CD3ε Fv-фрагмент антитела включает аминокислотную последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32-81. В некоторых вариантах осуществления анти-CD3ε Fv-фрагмент антитела включает комбинацию аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32-62, и аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 63-81. В некоторых вариантах осуществления анти-CD3ε Fv-фрагмент антитела включает комбинацию аминокислотной последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32-62, и аминокислотной последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 63-81.
[0212] В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает комбинацию последовательности VH CDR1, последовательности VH CDR2, и последовательности VH CDR3, где по меньшей мере одна из последовательности VH CDR1, последовательности VH CDR2 и последовательности VH CDR3 выбрана из последовательности VH CDR1, которая включает по меньшей мере аминокислотную последовательность TYAMN (SEQ ID NO: 16); последовательности VH CDR2, которая включает, по меньшей мере, аминокислотную последовательность RIRSKYNNYATYYADSVKD (SEQ ID NO: 17); и последовательности VH CDR3, которая включает, по меньшей мере, аминокислотную последовательность HGNFGNSYVSWFAY (SEQ ID NO: 18).
[0213] В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает комбинацию последовательности VL CDR1, последовательность VL CDR2 и последовательность VL CDR3, где по меньшей мере одна из последовательности VL CDR1, последовательность VL CDR2 и последовательности VL CDR3 выбрана из последовательности VL CDR1, которая включает, по меньшей мере, аминокислотную последовательность RSSTGAVTTSNYAN (SEQ ID NO: 19); последовательности VL CDR2, которая включает по меньшей мере аминокислотную последовательность GTNKRAP (SEQ ID NO: 20); и последовательности VL CDR3, которая включает, по меньшей мере, аминокислотную последовательность ALWYSNLWV (SEQ ID NO: 21).
[0214] В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает последовательность VH CDR1, которая включает, по меньшей мере, аминокислотную последовательность TYAMN (SEQ ID NO: 16); последовательность VH CDR2, которая включает, по меньшей мере, аминокислотную последовательность RIRSKYNNYATYYADSVKD (SEQ ID NO: 17); последовательность VH CDR3, которая включает, по меньшей мере, аминокислотную последовательность HGNFGNSYVSWFAY (SEQ ID NO: 18), последовательность VL CDR1, которая включает, по меньшей мере, аминокислотную последовательность RSSTGAVTTSNYAN (SEQ ID NO: 19); последовательность VL CDR2, которая включает по меньшей мере аминокислотную последовательность GTNKRAP (SEQ ID NO: 20); и последовательность VL CDR3, которая включает, по меньшей мере, аминокислотную последовательность ALWYSNLWV (SEQ ID NO: 21).
[0215] В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает комбинацию последовательности VH CDR1, последовательности VH CDR2, и последовательности VH CDR3, где по меньшей мере одна из последовательности VH CDR1, последовательности VH CDR2 и последовательности VH CDR3 выбрана из последовательности VH CDR1, которая включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности TYAMN (SEQ ID NO: 16); последовательности VH CDR2, которая включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности RIRSKYNNYATYYADSVKD (SEQ ID NO: 17); и последовательности VH CDR3, которая включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности HGNFGNSYVSWFAY (SEQ ID NO: 18).
[0216] В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает комбинацию последовательности VL CDR1, последовательности VL CDR2 и последовательности VL CDR3, где по меньшей мере одна из последовательности VL CDR1, последовательности VL CDR2 и последовательности VL CDR3, выбрана из последовательности VL CDR1, которая включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности RSSTGAVTTSNYAN (SEQ ID NO: 19); последовательности VL CDR2, которая включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности GTNKRAP (SEQ ID NO: 20); и последовательности VL CDR3, которая включает последовательность, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности ALWYSNLWV (SEQ ID NO: 21).
[0217] В некоторых вариантах осуществления анти-CD3ε-связывающий домен включает последовательность VH CDR1, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности TYAMN (SEQ ID NO: 16); последовательность VH CD2, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности RIRSKYNNYATYYADSVKD (SEQ ID NO: 17); последовательность VH CDR3, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности HGNFGNSYVSWFAY (SEQ ID NO: 18) последовательность VL CDR1, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности RSSTGAVTTSNYAN (SEQ ID NO: 19); последовательность VL CDR2, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности GTNKRAP (SEQ ID NO: 20); и последовательность VL CDR3, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности ALWYSNLWV (SEQ ID NO: 21).
[0218] В некоторых вариантах осуществления анти-CD3ε-связывающий домен их представляет собой Fv-фрагмент, который включает в себя комбинацию аминокислотной последовательности вариабельной области тяжелой цепи и аминокислотной последовательности вариабельной области легкой цепи. В некоторых вариантах осуществления анти-CD3ε-связывающий домен представляет собой Fv-фрагмент, который включает в себя комбинацию из аминокислотной последовательности вариабельной области тяжелой цепи и аминокислотной последовательности вариабельной области легкой цепи, содержащей аминокислотную последовательность, которые по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичны аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32-81. В некоторых вариантах осуществления анти-CD3ε-связывающий домен их представляет собой Fv-фрагмент, который включает в себя комбинацию аминокислотной последовательности вариабельной области тяжелой цепи и аминокислотной последовательности вариабельной области легкой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 32-81. В некоторых вариантах осуществления их анти-CD3ε-связывающий домен представляет собой Fv-фрагмент, который включает комбинацию аминокислотной последовательности вариабельной области тяжелой цепи, выбранной из группы SEQ ID NO: 32-62, и аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 63-81. В некоторых вариантах осуществления анти-CD3ε-связывающий домен представляет собой Fv-фрагмент, который включает в себя комбинацию аминокислотной последовательности вариабельной области тяжелой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32-62, и аминокислотной последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 63-81.
2. Fc-полипептиды иммуноглобулинов:
[0219] Первый компонент полиспецифических полипептидных конструкций по изобретению включает Fc-область иммуноглобулина. В некоторых вариантах осуществления Fc-область иммуноглобулина представляет собой изотип IgG, выбранный из группы, состоящей из изотипа IgG1, изотипа IgG2, изотипа IgG3 и подкласса IgG4. В некоторых вариантах осуществления Fc-область представляет собой Fc человека. В некоторых вариантах осуществления Fc-область иммуноглобулина представляет собой полипептид, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-6. В некоторых вариантах осуществления Fc-область иммуноглобулина содержит цепь Fc, которая представляет собой иммунологически активный фрагмент любой из SEQ ID NO: 1-6. В некоторых вариантах осуществления Fc-область иммуноглобулина содержит полипептидную цепь Fc, которая по меньшей мере на 50%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности любой из SEQ ID NO: 1-6, или ее иммунологически активный фрагмент.
[0220] В некоторых вариантах осуществления полиспецифическая полипептидная конструкция представляет собой димер, образованный полипептидами, каждый из которых содержит Fc. В некоторых конкретных вариантах осуществления идентичные или по существу идентичные полипептиды будут димеризованы для создания гомодимера. В некоторых вариантах осуществления димер представляет собой гомодимер, в котором два полипептида полиспецифической полипептидной конструкции являются одинаковыми. В других случаях Fc-область образована Fc-доменами, которые подвергнуты мутации или модифицированы так, чтобы способствовать гетеродимеризации, в которой различные полипептиды могут быть димеризованы с получением гетеродимера. Таким образом, в некоторых вариантах осуществления димер представляет собой гетеродимер, в котором две полипептидные цепи полиспецифической полипептидной конструкции различны. Примеры модификаций, способствующих гетеродимеризации, известны, включая любые, как описано ниже.
[0221] В целом, Fc-область отвечает за эффекторные функции, такие как комплемент-зависимая цитотоксичность (CDC) и антитело-зависимая клеточная цитотоксичность (ADCC), в дополнение к антигенсвязывающей способности, которая является основной функцией иммуноглобулинов. Кроме того, последовательность FcRn, присутствующая в Fc-области, играет роль регуляции уровня IgG в сыворотке путем увеличения периода полужизни in vivo путем конъюгации с рецептором FcRn in vivo. В некоторых вариантах осуществления такие функции могут быть изменены, например, уменьшены или усилены в Fc для использования с представленными полиспецифическими полипептидными конструкциями.
[0222] В некоторых вариантах осуществления Fc-область представленных полиспецифических полипептидных конструкций проявляет одну или более эффекторных функций. В некоторых случаях Fc-область способна обеспечивать Fc-опосредованные эффекторные функции, такие как, например, ADCC (например, высвобождение гранзима B NK-клетками), ADCP и/или CDC. Таким образом, в некоторых вариантах осуществления, в которых полиспецифические полипептидные конструкции содержат расщепляемый линкер, расщепление линкера может продуцировать два компонента, каждый из которых обладает биологической активностью: CD3-связывающая область, которая способна связываться и рекрутировать CD3 на Т-клетки, и Fc-область, связанная с TAA-антигенсвязывающим доменом, который может проявлять специфическую для мишени эффекторную функцию.
[0223] В некоторых вариантах осуществления Fc-область включает полипептид Fc, который повергнут мутации или модифицирован для изменения одной или более эффекторных функций. Известны различные примеры изменяющих мутаций полипептидов Fc, как например, для снижения эффекторной функции, включая любые, как описано ниже. В некоторых вариантах осуществления ссылка на аминокислотные замены в Fc-области осуществляется посредством нумерации EU по Kabat (также называемой нумерацией Kabat), если не описано со ссылкой на конкретную SEQ ID NO. Нумерация EU известна и соответствует последней обновленной научной шкале IMGT (IMGT®, международная информационная система ImMunoGeneTics®, http://www.imgt.org/IMGTScientificChart/Numbering/Hu_IGHGnber.html (создана 17 мая 2001 г., последнее обновление: 10 января 2013 г.) и индекс EU, как сообщается у Kabat, EA et al. Sequences of Proteins of Immunological interest. 5th ed. US Department of Health and Human Services, NIH publication No. 91-3242 (1991).
[0224] В некоторых вариантах осуществления представленные полиспецифические полипептидные конструкции, которые содержат Fc-область, которая демонстрирует ослабленные эффекторные функции, могут быть желательными кандидатами для применений, в которых требуется ограниченное связывание CD3, но некоторые эффекторные функции (такие как CDC и ADCC) являются ненужными или вредными. Анализы цитотоксичности in vitro и/или in vivo могут проводиться для подтверждения снижения/истощения активности CDC и/или ADCC. Например, анализы связывания Fc-рецептора (FcR) могут проводиться для того, чтобы убедиться, что полиспецифические полипептидные конструкции и/или их расщепленные компоненты не связываются с FcγR (следовательно, вероятно, не обладают активностью ADCC), но сохраняют способность связывания FcRn. Первичные клетки, опосредующие ADCC, NK-клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Неограничивающие примеры анализов in vitro для оценки активности ADCC интересующей молекулы описаны в патенте США No. 5500362 (см., например, Hellstrom, I. et al. Proc. Nat'l Acad. Sci. USA 83: 7059-7063 (1986) и Hellstrom, I et al., Учеб. Nat'l Acad. Sci. USA 82: 1499-1502 (1985); Патент США 5821337 (см. Bruggemann, M. et al., J. Exp. Med. 166: 1351-1361 (1987)). Альтернативно, могут быть использованы методы нерадиоактивного анализа (см., например, анализ нерадиоактивной цитотоксичности ACTI™ для проточной цитометрии (CellTechnology, Inc. Mountain View, Калифорния; и анализ нерадиоактивной цитотоксичности CytoTox 96™ (Promega, Madison Wis.). Пригодные эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (РВМС) и клетки-натуральные киллеры (NK). Альтернативно или дополнительно, активность ADCC интересующей молекулы может быть оценена in vivo, например, на модели животного, такой как раскрыта в Clynes et al. Proc. Nat'l Acad. Sci. USA 86, 10029-10033 (1989)). Также можно проводить анализы связывания C1q, чтобы подтвердить, что полиспецифическая полипептидная конструкция или ее расщепленные компоненты не способны связывать C1q и, следовательно, не обладают активностью CDC. См., например, ИФА для связывания C1q и C3c в WO 2006/029879 и WO 2005/100402. Для оценки активации комплемента может быть проведен анализ CDC (см., например, Gazzano-Santoro et al., J. Immunol. Methods 202: 163 (1996); Cragg, MS et al., Blood 101: 1045-1052 (2003); и Cragg, MS и MJ Glennie, Blood, 103: 2738-2743 (2004). Определение связывания FcRn и определения клиренса/периода полужизни in vivo также можно проводить с использованием способов, известных в данной области (см., например, Petkova, SB et al., Int'l.Immunol. 18 (12): 1759-1769 (2006).
[0225] В некоторых вариантах осуществления Fc-область иммуноглобулина или ее иммунологически активный фрагмент представляет собой изотип IgG. Например, Fc-область иммуноглобулина слитого белка имеет изотип IgG1 человека, имеющий аминокислотную последовательность:
PAPELLGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL HQDWLNGKEY KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSRDELT KNQVSLTCLV KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLYSK LTVDKSRWQQ GNVFSCSVMH EALHNHYTQK SLSLSPGK (SEQ ID NO: 1)
[0226] В некоторых вариантах осуществления Fc-область иммуноглобулина или ее иммунологически активный фрагмент содержит полипептидную последовательность человеческого IgG1, которая по меньшей мере на 50%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 1.
[0227] В некоторых вариантах осуществления Fc-область человеческого IgG1 модифицирована для изменения антителозависимой клеточной цитотоксичности (ADCC) и/или комплементзависимой цитотоксичности (CDC), например, модификации аминокислот, описанные в Natsume et al., 2008 Cancer Res, 68(10): 3863-72; Idusogie et al., 2001 J Immunol, 166(4): 2571-5; Moore et al., 2010 mAbs, 2(2): 181-189; Lazar et al., 2006 PNAS, 103(11): 4005-4010, Shields et al., 2001 JBC, 276(9): 6591-6604; Stavenhagen et al., 2007 Cancer Res, 67(18): 8882-8890; Stavenhagen et al., 2008 Advan. Enzyme Regul., 48: 152-164; Alegre et al., 1992, J. Immunol, 148: 3461-3468; Рассмотрено в Kaneko and Niwa, 2011 Biodrugs, 25 (1): 1-11, содержание каждой из которых настоящим включено в качестве ссылки в полном объеме.
[0228] В некоторых вариантах осуществления Fc-область, такая как Fc-область человеческого IgG1, модифицирована для усиления активности ADCC или активности CDC. Примеры мутаций, которые усиливают ADCC, включают модификации в Ser239 и Ile332, например Ser239Asp и Ile332Glu (S239D, I332E). Примеры мутаций, которые усиливают CDC, включают модификации в Lys326 и Glu333. В некоторых вариантах осуществления Fc-область модифицируется в одном или обоих из этих положений, например, Lys326Ala и/или Glu333Ala (K326A и E333A) с использованием системы нумерации Kabat.
[0229] В некоторых вариантах осуществления слитые белки Fc-области человеческого IgG1 по настоящему изобретению лишены или имеют пониженное содержание фукозы, присоединенной к N-связанной гликановой цепи у N297. Существует множество способов предотвращения фукозилирования, включая, но не ограничиваясь этим, продуцирование в клеточной линии с дефицитом FUT8; добавление ингибиторов в среду для культивирования клеток млекопитающих, например кастаноспермина; и метаболическое конструирование продуцирующей клеточной линии. В некоторых вариантах осуществления Fc-область человеческого IgG1 модифицирована по аминокислоте Asn297 (обведено в рамку, нумерация Kabat) для предотвращения гликозилирования слитого белка, например, Asn297Ala (N297A) или Asn297Asp (N297D).
[0230] В некоторых вариантах осуществления Fc-область слитого белка изменяется в одном или более из следующих положений для уменьшения связывания с Fc-рецептором: Leu 234 (L234), Leu235 (L235), Asp265 (D265), Asp270 (D270), Ser298 (S298), Asn297 (N297), Asn325 (N325) или Ala327 (A327). Например, Leu234Ala (L234A), Leu235Ala (L235A), Asp265Asn (D265N), Asp270Asn (D270N), Ser298Asn (S298N), Asn297Ala (N297A), Asn325Glu (N325E) или Ala327Ser (A327S). В некоторых вариантах осуществления Fc-область слитого белка модифицирована по аминокислоте Leu235 (обведено в рамку SEQ ID NO: 1 выше, нумерация Kabat) для изменения взаимодействий с рецептором Fc, например, Leu235Glu (L235E) или Leu235Ala (L235A). В некоторых вариантах осуществления Fc-область слитого белка модифицирована по аминокислоте Leu234 (обведено в рамку SEQ ID NO: 1 выше, нумерация по Kabat), чтобы изменить взаимодействия с рецептором Fc, например Leu234Ala (L234A). В некоторых вариантах осуществления Fc-область слитого белка изменена как в аминокислоте 234, так и в 235, например, Leu234Ala и Leu235Ala (L234A/L235A) или Leu234Val и Leu235Ala (L234V/L235A). В предпочтительных вариантах осуществления модификации в Fc-области уменьшают связывание с Fc-гамма-рецепторами, в то время как оказывают минимальное влияние на связывание с неонатальным Fc-рецептором (FcRn).
[0231] В некоторых вариантах осуществления Fc-область человеческого IgG модифицирована для усиления связывания FcRn. Примерами мутаций Fc, которые усиливают связывание с FcRn, представляют собой Met252Tyr, Ser254Thr, Thr256Glu (M252Y, S254T, T256E соответственно) (нумерация Kabat, Dall'Acqua et al. 2006, J. Biol Chem Vol. 281 (33) 23514-23524), Met428Leu и Asn434Ser (M428L, N434S) (Zalevsky et al., 2010 Nature Biotech, Vol. 28 (2) 157-159) (EU индекс Kabat et al 1991 Sequences of Proteins of Immunological Interest.). В некоторых вариантах осуществления мутированный или модифицированный полипептид Fc включает следующие мутации: Met252Tyr и Met428Leu или Met252Tyr и Met428Val (M252Y, M428L или M252Y, M428V) с использованием системы нумерации Kabat.
[0232] В некоторых вариантах осуществления в Fc-области слитого белка отсутствует аминокислота в одном или более из следующих положений для снижения связывания с Fc-рецептором: Glu233 (E233), Leu234 (L234) или Leu235 (L235). В этих вариантах осуществления делеция Fc этих трех аминокислот снижает связывание белка комплемента C1q.
[0233] PAPGGPSVFL FPPKPKDTLM ISRTPEVTCV VVDVSHEDPE VKFNWYVDGV EVHNAKTKPR EEQYNSTYRV VSVLTVLHQD WLNGKEYKCK VSNKALPAPI EKTISKAKGQ PREPQVYTLP PSRDELTKNQ VSLTCLVKGF YPSDIAVEWE SNGQPENNYK TTPPVLDSDG SFFLYSKLTV DKSRWQQGNV FSCSVMHEAL HNHYTQKSLS LSPGK (SEQ ID NO: 2)
[0234] В некоторых вариантах осуществления Fc-область слитого белка изменена в Gly236 (обведено в рамку в SEQ ID NO: 1 выше) для уменьшения связывания с Fc-рецептором. Например, где Gly236 делетирован из слитого белка. В некоторых вариантах осуществления Fc-область человеческого IgG1 модифицируется в аминокислоте Gly236 для усиления взаимодействия с CD32A, например, Gly236Ala (G236A).
[0235] В некоторых вариантах осуществления в Fc-области человеческого IgG1 отсутствует Lys447 (EU индекс Kabat et al 1991 Sequences of Proteins of Immunological Interest).
[0236] В некоторых вариантах осуществления его слитый или иммунологически активный фрагмент содержит полипептидную последовательность человеческого IgG2, которая по меньшей мере на 50%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%., 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 2.
[0237] В некоторых вариантах осуществления Fc-область иммуноглобулина или иммунологически активного фрагмента слитого белка имеет изотип IgG2 человека, имеющий аминокислотную последовательность:
PAPPVAGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVQFNWYVD GVEVHNAKTK PREEQFNSTF RVVSVLTVVH QDWLNGKEYK CKVSNKGLPA PIEKTISKTK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDISVE WESNGQPENN YKTTPPMLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGK (SEQ ID NO: 3)
[0238] В некоторых вариантах осуществления его слитый или иммунологически активный фрагмент содержит полипептидную последовательность человеческого IgG2, которая по меньшей мере на 50%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%., 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 3.
[0239] В некоторых вариантах осуществления Fc-область человеческого IgG2 модифицирована по аминокислоте Asn297 (обведено в рамку, чтобы предотвратить гликозилирование антитела, например, Asn297Ala (N297A) или Asn297Asp (N297D). В некоторых вариантах осуществления в Fc-области человеческого IgG2 отсутствует Lys447 (EU индекс Kabat et al 1991 Sequences of Proteins of Immunological Interest).
[0240] В некоторых вариантах осуществления Fc-область иммуноглобулина или иммунологически активный фрагмент слитого белка имеет изотип IgG3 человека, имеющий аминокислотную последовательность:
PAPELLGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE DPEVQFKWYV DGVEVHNAKT KPREEQYNST FRVVSVLTVL HQDWLNGKEY KCKVSNKALP APIEKTISKT KGQPREPQVY TLPPSREEMT KNQVSLTCLV KGFYPSDIAV EWESSGQPEN NYNTTPPMLD SDGSFFLYSK LTVDKSRWQQ GNIFSCSVMH EALHNRFTQK SLSLSPGK (SEQ ID NO: 4)
[0241] В некоторых вариантах осуществления антитело или его иммунологически активный фрагмент содержат полипептидную последовательность человеческого IgG3, которая по меньшей мере на 50%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%., 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 4.
[0242] В некоторых вариантах осуществления Fc-область человеческого IgG3 модифицирована по аминокислоте Asn297 (обведено в рамку, нумерация по Kabat) для предотвращения гликозилирования антитела, например Asn297Ala (N297A) или Asn297Asp (N297D). В некоторых вариантах осуществления Fc-область человеческого IgG3 модифицирована в аминокислоте 435 для увеличения периода полужизни, например, Arg435His (R435H). В некоторых вариантах осуществления в Fc-области человеческого IgG3 отсутствует Lys447 (EU индекс Kabat et al 1991 Sequences of Proteins of Immunological Interest).
[0243] В некоторых вариантах осуществления Fc-область иммуноглобулина или иммунологически активного фрагмента слитого белка имеет изотип IgG4 человека, имеющий аминокислотную последовательность:
PAPEFLGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSQE DPEVQFNWYV DGVEVHNAKT KPREEQFNST YRVVSVLTVL HQDWLNGKEY KCKVSNKGLP SSIEKTISKA KGQPREPQVY TLPPSQEEMT KNQVSLTCLV KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLYSR LTVDKSRWQE GNVFSCSVMH EALHNHYTQK SLSLSLGK (SEQ ID NO: 5)
[0244] В некоторых вариантах осуществления антитело или его иммунологически активный фрагмент содержат полипептидную последовательность человеческого IgG4, которая по меньшей мере на 50%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%., 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 5.
[0245] В некоторых вариантах осуществления Fc-область иммуноглобулина или иммунологически активного фрагмента слитого белка имеет изотип IgG4 человека, имеющий аминокислотную последовательность:
PAPELLGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSQE DPEVQFNWYV DGVEVHNAKT KPREEQFNST YRVVSVLTVL HQDWLNGKEY KCKVSNKGLP SSIEKTISKA KGQPREPQVY TLPPSQEEMT KNQVSLTCLV KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLYSR LTVDKSRWQE GNVFSCSVMH EALHNHYTQK SLSLSLGK (SEQ ID NO: 6)
[0246] В некоторых вариантах осуществления антитело или его иммунологически активный фрагмент содержит полипептидную последовательность человеческого IgG4, которая по меньшей мере на 50%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%., 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 6.
[0247] В других вариантах осуществления Fc-область человеческого IgG4 модифицируют по аминокислоте 235 для изменения взаимодействия с Fc-рецептором, например, Leu235Glu (L235E). В некоторых вариантах осуществления Fc-область человеческого IgG4 модифицирована по аминокислоте Asn297 (обведено в рамку, нумерация по Kabat) для предотвращения гликозилирования антитела, например, Asn297Ala (N297A) или Asn297Asp (N297D). В некоторых вариантах осуществления в Fc-области человеческого IgG4 отсутствует Lys447 (EU индекс Kabat et al 1991 Sequences of Proteins of Immunological Interest).
[0248] В некоторых вариантах осуществления Fc-область человеческого IgG модифицируют для стабилизации гомодимеризации в интерфейсе СН3:СН3 путем введения двух дисульфидных связей путем замены Ser354 на Cys (S354C) и Tyr349 на Cys (Y349C) (S354C/Y349C).
[0249] В некоторых вариантах осуществления Fc-область человеческого IgG модифицируют для индуцирования гетеродимеризации. Известны различные способы стимулирования гетеродимеризации комплементарных полипептидов Fc, см., например, Ridgway et al., Protein Eng. 9: 617-621 (1996); Merchant et al., Nat. Biotechnol.16(7): 677-81 (1998); Moore et al. (2011) MAbs, 3: 546-57; Von Kreudenstein et al. MAbs, (2013) 5: 646-54; Gunasekaran et al. (2010) J. Biol. Chem., 285: 19637-46; Leaver-Fay et al. (2016) Structure, 24:641-51; Ha et al. (2016) Frontiers in Immunology, 7:1; Davis et al. (2010) Protein Eng Des Sel, 23:195-202; published international PCT Appl. № WO 1998/050431, WO 2009/089004, WO2011143545, WO 2014/067011, WO 2012/058768, WO2018027025; опубликованная патентная заявка США № US20140363426, US20150307628, US20180016354, US20150239991; и патенты США №№ US 5731168, US7183076, US9701759, US9605084 и US9650446. Методы, способствующие гетеродимеризации цепей Fc, включают мутагенез Fc-области, например, путем включения набора мутаций «выступ-во-впадину» или включения мутаций для воздействия на электростатическое взаимодействие Fc, чтобы способствовать взаимодействиям притяжения между различными полипептидными цепями. Например, в некоторых вариантах осуществления полипептиды Fc гетеродимера включают мутацию для изменения полярности заряда в интерфейсе димера Fc, так что коэкспрессия электростатически согласованных цепей Fc поддерживает благоприятные взаимодействия притяжения, тем самым способствуя образованию желаемого гетеродимера Fc, тогда как неблагоприятные взаимодействия отталкивающего заряда подавляют образование нежелательных гомодимеров Fc (Guneskaran et al. (2010) JBC, 285: 19637-19646). При совместной экспрессии в клетке, связь между цепями возможна, но цепи по существу не самоассоциируются из-за отталкивания заряда. Другие стратегии получения гетеродимерной Fc-области включают смешивание сегментов домена CH3 IgG и IgA человека для создания комплементарного гетеродимера CH3, который называется SEED Fc.
[0250] В некоторых вариантах осуществления для содействия гетеродимеризации оба полипептида гетеродимера Fc содержат парные или комплементарные модификации аминокислот. Типичные парные аминокислотные модификации полипептидов слитого Fc приведены в Таблице 1.
[0251] В некоторых вариантах осуществления модификации включают введение выпуклости (выступа) в первый полипептид Fc и полости (впадины) во второй полипептид Fc, так что выпуклость располагается в полости, чтобы способствовать комплексообразованию первого и второго Fc-содержащих полипептидов. Аминокислоты, предназначенные для замены и/или модификации для создания выпуклостей или полостей в полипептиде, обычно представляют собой аминокислоты в интерфейсе, которые взаимодействуют или контактируют с одной или более аминокислотами в интерфейсе второго полипептида.
[0252] В некоторых вариантах осуществления первый полипептид Fc, который модифицирован так, чтобы содержать аминокислоты выпуклости (впадины), включает замену нативной или исходной аминокислоты аминокислотой, которая имеет по меньшей мере одну боковую цепь, которая выступает из интерфейса первого полипептида Fc и поэтому может быть расположен в компенсаторной полости (впадине) в соседнем интерфейсе второго полипептида. Чаще всего замещающая аминокислота представляет собой аминокислоту, которая имеет больший объем боковой цепи, чем исходный аминокислотный остаток. Специалист в данной области техники знает, как определить и/или оценить свойства аминокислотных остатков, чтобы идентифицировать те, которые являются идеальными заменяющими аминокислотами для создания выпуклости. В некоторых вариантах осуществления замещающие остатки для образования выпуклости представляют собой встречающиеся в природе аминокислотные остатки и включают, например, аргинин (R), фенилаланин (F), тирозин (Y) или триптофан (W). В некоторых примерах исходный остаток, идентифицированный для замены, представляет собой аминокислотный остаток с небольшой боковой цепью, такой как, например, аланин, аспарагин, аспарагиновая кислота, глицин, серин, треонин или валин.
[0253] В некоторых вариантах осуществления второй полипептид Fc, который модифицирован так, чтобы он содержал полость (впадину), представляет собой полипептид, который включает замену нативной или исходной аминокислоты аминокислотой, которая имеет по меньшей мере одну боковую цепь, которая углублена из интерфейса второго полипептида и, таким образом, способна приспосабливать соответствующую выпуклость из интерфейса первого полипептида. Чаще всего замещающей аминокислотой является та, которая имеет меньший объем боковой цепи, чем исходный аминокислотный остаток. Специалист в данной области знает, как определить и/или оценить свойства аминокислотных остатков, чтобы идентифицировать те, которые являются идеальными замещающими остатками для образования полости. Обычно замещающие остатки для образования полости представляют собой встречающиеся в природе аминокислоты и включают, например, аланин (A), серин (S), треонин (T) и валин (V). В некоторых примерах исходная аминокислота, идентифицированная для замены, представляет собой аминокислоту, которая имеет крупную боковую цепь, такую как, например, тирозин, аргинин, фенилаланин или триптофан.
[0254] Интерфейс CH3 человеческого IgG1, например, включает шестнадцать остатков в каждом домене, расположенном на четырех антипараллельных β-цепях, которые погружены на 1090 Å2 с каждой поверхности (см., например, Deisenhofer et al.(1981) Biochemistry, 20: 2361-2370; Miller et al., (1990) J. Mol. Biol., 216, 965-973; Ridgway et al., (1996) Prot. Engin., 9: 617-621; Патент США № 5731168). Модификации домена CH3 для создания выпуклостей или полостей описаны, например, в патенте США No. 5731168; Международных патентных заявках WO 98/50431 и WO 2005/063816; и Ridgway et al., (1996) Prot.Engin., 9: 617-621. В некоторых примерах модификации домена СН3 для создания выпуклостей или полостей обычно нацелены на остатки, расположенные на двух центральных антипараллельных β-цепях. Цель состоит в том, чтобы свести к минимуму риск того, что создаваемые выпуклости могут быть приспособлены выступом в окружающий растворитель, а не с помощью компенсаторной полости в домене CH3 партнера.
[0255] Например, в некоторых вариантах осуществления гетеродимер Fc включает полипептид, имеющий аминокислотную модификацию в домене CH3 у Thr366, который при замене на более объемную аминокислоту, например, Try (T366W), способен преимущественно связываться со вторым доменом CH3, имеющим аминокислотные модификации для менее объемных аминокислот в положениях Thr366, Leu368 и Tyr407, например Ser, Ala и Val, соответственно, (T366S/L368A/Y407V). Гетеродимеризацию с помощью модификаций СН3 можно дополнительно стабилизировать путем введения дисульфидной связи, например, путем замены Ser354 на Cys (S354C) и Tyr349 на Cys (Y349C) на противоположных доменах СН3 (Рассмотрено в Carter, 2001 Journal of Immunological Methods, 248: 7-15).
[0256] Полученные в результате полиспецифические полипептидные конструкции могут быть очищены любым подходящим способом, таким как, например, аффинная хроматография на колонках с Протеином А или Протеином G. Если две молекулы нуклеиновой кислоты, кодирующие разные полипептиды, трансформируются в клетки, происходит образование гомо- и гетеродимеров. Условия для экспрессии можно регулировать так, чтобы образование гетеродимера было предпочтительным по сравнению с образованием гомодимера.
[0257] Известны способы восстановления гетеродимеров из гомодимеров на основе дифференциальной аффинности гетеродимеров к аффинному реагенту. В некоторых аспектах такие методы включают конструирование гетеродимера так, чтобы одна из полипептидных цепей Fc не связывалась с аффинным реагентом Протеина А. В некоторых случаях одна из полипептидных цепей может содержать одну или более аминокислотных замен для устранения или снижения аффинности к реагенту Протеина А в одном из полипептидов гетеродимера Fc, см., например, WO2017134440, WO2010151792, Jendeberg et al. (Jendeberg et al., (1997) J. Immunol. Methods, 201(1): 25-34. В некоторых из этих вариантов осуществления Fc-область может быть модифицирована в сайте связывания Протеина A на одном члене гетеродимера, чтобы предотвратить связывание Протеина A и тем самым обеспечить более эффективную очистку гетеродимерного слитого белка. Примером модификации в этом сайте связывания является Ile253, например Ile253Arg (I253R). В некоторых вариантах осуществления модификация может быть H435R или H435R/Y436F. В некоторых вариантах осуществления полипептид Fc гетеродимера Fc может содержать модификацию, позволяющую ему связывать Протеин А, но не Протеин G (pA+/pG-). Типичные модификации аминокислот pA+/pG- включают Fc, содержащий серин в положении 428, серин в положении 434 и необязательно гистидин в положении 436, относительно человеческого IgG1 или содержащий эти остатки в соответствующих положениях в человеческом IgG2, 3 или 4. В некоторых аспектах такие модификации аминокислот в одном полипептиде Fc IgG в положениях 428, 434 и, необязательно, 436 снижают или предотвращают связывание Протеина G, усиливая очистку белка.
[0258] В некоторых вариантах осуществления любая из таких модификаций для придания дифференциальной аффинности аффинному реагенту может сочетаться с любой одной или более другими модификациями аминокислот, описанными выше. Например, модификация I253R может объединяться либо с модификациями T366S/L368A/Y407V, либо с модификациями T366W. Модифицированный Fc T366S/L368A/Y407V способен образовывать гомодимеры, поскольку нет стерической окклюзии в интерфейсе димеризации, как в случае модифицированного Fc T336W. Следовательно, в некоторых вариантах осуществления модификация I253R объединяется с модифицированным Fc T366S/L368A/Y407V для предотвращения очистки любого гомодимерного Fc, который мог образоваться. Аналогичные модификации могут быть использованы путем объединения T366S/L368A/Y407V и H453R.
[0259] В некоторых вариантах осуществления Fc-области гетеродимерной молекулы дополнительно могут содержать одну или более других мутаций Fc, таких как любая описанная выше. В некоторых вариантах осуществления гетеродимерная молекула содержит Fc-область с мутацией, которая снижает эффекторную функцию.
[0260] В некоторых вариантах осуществления один полипептид Fc гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 82, 86, 94 или 96, а другой полипептид Fc гетеродимерной Fc-области содержит последовательность аминокислот, представленных в любой из SEQ ID NO: 83, 87, 90, 92, 98 или 100. В некоторых вариантах осуществления один полипептид Fc гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 84, 88, 95 или 97, а другой полипептид Fc гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 85, 89, 91, 93, 99 или 101.
[0261] В некоторых вариантах осуществления Fc-область человеческого IgG модифицирована для предотвращения димеризации. В этих вариантах осуществления слитые белки по настоящему изобретению являются мономерными. Например, модификация остатка Thr366 до заряженного остатка, например Thr366Lys, Thr366Arg, Thr366Asp или Thr366Glu (T366K, T366R, T366D или T366E, соответственно), предотвращает димеризацию CH3-CH3.
[0262] В некоторых вариантах осуществления Fc-область слитого белка изменяется в одном или более из следующих положений для уменьшения связывания с Fc-рецептором: Leu 234 (L234), Leu235 (L235), Asp265 (D265), Asp270 (D270), Ser298 (S298), Asn297 (N297), Asn325 (N325) или Ala327 (A327). Например, Leu 234Ala (L234A), Leu235Ala (L235A), Asp265Asn (D265N), Asp270Asn (D270N), Ser298Asn (S298N), Asn297Ala (N297A), Asn325Glu (N325E) или Ala7S. В предпочтительных вариантах осуществления модификации в Fc-области уменьшают связывание с Fc-гамма-рецепторами, в то время как оказывают минимальное влияние на связывание с неонатальным Fc-рецептором (FcRn).
[0263] В некоторых вариантах осуществления слитый белок содержит полипептид, полученный из шарнирной области иммуноглобулина. Шарнирная область может быть выбрана из любого подкласса IgG человека. Например, слитый белок может содержать модифицированный шарнир IgG1, имеющий последовательность EPKSSDKTHTCPPC (SEQ ID NO: 7), где Cys220, который образует дисульфид с C-концевым цистеином легкой цепи, подвергают мутации до серина, например, Cys220Ser (C220S). В других вариантах осуществления слитый белок содержит укороченный шарнир, имеющий последовательность DKTHTCPPC (SEQ ID NO: 8).
[0264] В некоторых вариантах осуществления слитый белок имеет модифицированный шарнир из IgG4, который модифицирован для предотвращения или уменьшения обмена цепей, например, Ser228Pro (S228P), имеющий последовательность ESKYGPPCPPC (SEQ ID NO: 9). В некоторых вариантах слитый белок содержит линкерные полипептиды. В других вариантах слитый белок содержит линкерные и шарнирные полипептиды.
3. Линкеры
[0265] Представленные полиспецифические полипептидные конструкции содержат линкер, который присоединяет или связывает первый компонент, содержащий Fc-область иммуноглобулина, и второй компонент, содержащий CD3-связывающая область. В некоторых вариантах осуществления линкер расположен в конце С-концевой области Fc-области, так что Fc-область является N-концевой по отношению к CD3-связывающей области. Поскольку представленные полиспецифические полипептидные конструкции являются мультимерами, такими как димеры, представленные конструкции включают линкер, соединяющий первый полипептид Fc и первый домен (например, VH) CD3-связывающей области первого полипептида и второй полипептид Fc и второй домен (например, VL) CD3-связывающей области второго полипептида. Обычно линкеры, присутствующие в первом и втором полипептидах полиспецифической полипептидной конструкции, являются одинаковыми. Таким образом, в некоторых вариантах осуществления каждый домен CD3-связывающего домена связан через линкер, такой как тот же линкер, с противоположными полипептидами Fc, такими как гетеродимерный Fc.
[0266] Известны различные полипептидные линкеры для использования в слитых белках (см., например, Chen et al.(2013) Adv. Drug. Deliv. 65: 1357-1369; и международную публикацию РСТ № WO 2014/099997, WO2000/24884; Патент США № 5258988; Патент США № 5525491; Патент США № 5525491, Патент США № 6132992).
[0267] В некоторых вариантах осуществления линкер выбирают таким образом, чтобы, когда CD3-связывающая область присоединялась к Fc-области полиспецифического полипептидного конъюгата, CD3-связывающая область была ограничена и не способна или по существу не способна связывать или рекрутировать CD3 на поверхность клетки, например, Т-клетки, при контакте полиспецифической полипептидной конструкции с клеткой. Различные анализы могут быть использованы для оценки связывания или рекрутирования CD3 полиспецифической полипептидной конструкцией, включая анализы для оценки связывания Т-клеток, активации NFAT с использованием репортерной системы, активности цитолитических Т-клеток, продуцирование цитокинов и/или экспрессию маркеров активации Т-клеток. Примеры анализов показаны в представленных Примерах. Как правило, линкер также представляет собой линкер, который обеспечивает корректное сворачивание полипептидной конструкции, не обладает зарядом, который не согласуется с активностью или функцией связанных полипептидов или не образует связи или другие взаимодействия с аминокислотными остатками в одном или более из доменов, которые будут препятствовать или изменять активность связанных полипептидов. В некоторых вариантах осуществления линкер представляет собой полипептидный линкер. Полипептидный линкер может быть гибким линкером или жестким линкером или их комбинацией. В некоторых аспектах линкер представляет собой короткий, средний или длинный линкер. В некоторых вариантах осуществления линкер имеет длину до 40 аминокислот. В некоторых вариантах осуществления линкер имеет длину до 25 аминокислот. В некоторых вариантах осуществления линкер имеет длину, составляющую по меньшей мере 2 аминокислоты, или по меньшей мере примерно 2 аминокислоты. В некоторых аспектах подходящая длина составляет, например, по меньшей мере, один аминокислотный остаток и, как правило, менее чем примерно 40 аминокислотных остатков, как например, 2-25 аминокислотных остатков, 5-20 аминокислотных остатков, 5-15 аминокислотных остатков, 8-12 аминокислот. В некоторых вариантах осуществления длина линкера составляет или составляет примерно от 2 до 24 аминокислот, от 2 до 20 аминокислот, от 2 до 18 аминокислот, от 2 до 14 аминокислот, от 2 до 12 аминокислот, от 2 до 10 аминокислот, от 2 до 8 аминокислот, от 2 до 6 аминокислот, от 6 до 24 аминокислот, от 6 до 20 аминокислот, от 6 до 18 аминокислот, от 6 до 14 аминокислот, от 6 до 12 аминокислот, от 6 до 10 аминокислот, от 6 до 8 аминокислот, от 8 до 24 аминокислот, от 8 до 20 аминокислот, от 8 до 18 аминокислот, от 8 до 14 аминокислот, от 8 до 12 аминокислот, от 8 до 10 аминокислот, от 10 до 24 аминокислот, от 10 до 20 аминокислот, от 10 до 18 аминокислот, от 10 до 14 аминокислот, от 10 до 12 аминокислот, от 12 до 24 аминокислот, от 12 до 20 аминокислот, от 12 до 18 аминокислот, от 12 до 14 аминокислот, от 14 до 24 аминокислот, от 14 до 20 аминокислот, от 14 до 18 аминокислот, от 18 до 24 аминокислот, от 18 до 20 аминокислот или от 20 до 24 аминокислот. В некоторых вариантах осуществления линкер имеет длину 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот.
[0268] В некоторых аспектах, чем длиннее линкер, тем больше связывание CD3, когда полиспецифический полипептидный конъюгат связан с его антигеном, например TAA. Таким образом, в некоторых аспектах линкер имеет длину более чем 12 аминокислот, например, длину более чем 13, 14, 15, 16, 17 или 18 аминокислот. В некоторых вариантах осуществления линкер имеет длину от 12 до 40 аминокислот, от 12 до 30 аминокислот, от 12 до 24 аминокислот, от 12 до 18 кислот, от 12 до 15 аминокислот, от 15 до 40 аминокислот, от 15 до 30 аминокислот, От 15 до 24 аминокислот, от 15 до 18 аминокислот, от 18 до 40 аминокислот, от 18 до 30 аминокислот, от 18 до 24 аминокислот, от 24 до 40 аминокислот, от 24 до 30 аминокислот или от 30 до 40 аминокислот.
[0269] Линкеры могут быть природными, синтетическими или комбинацией обоих. Особенно подходящие линкерные полипептиды преимущественно включают аминокислотные остатки, выбранные из глицина (Gly), серина (Ser), аланина (Ala) и треонина (Thr). Например, линкер может содержать по меньшей мере 75% (в расчете на общее количество остатков, присутствующих в пептидном линкере), например по меньшей мере 80%, по меньшей мере 85% или по меньшей мере 90% аминокислотных остатков выбранных из Gly, Ser, Ala и Thr. Линкер также может состоять только из остатков Gly, Ser, Ala и/или Thr. В некоторых вариантах осуществления линкер содержит 1-25 остатков глицина, 5-20 остатков глицина, 5-15 остатков глицина или 8-12 остатков глицина. В некоторых аспектах подходящие пептидные линкеры обычно содержат, по меньшей мере, 50% остатков глицина, как например, по меньшей мере, 75% остатков глицина. В некоторых вариантах осуществления пептидный линкер содержит только остатки глицина. В некоторых вариантах осуществления пептидный линкер содержит только остатки глицина и серина.
[0270] В некоторых вариантах осуществления эти линкеры состоят преимущественно из аминокислот глицина и серина, обозначаемых здесь как GS-линкеры. В некоторых вариантах осуществления линкер содержит (GGS) n, где n равно от 1 до 10, например от 1 до 5, например от 1 до 3, например, GGS (GGS)n (SEQ ID NO: 171), где n равно 0 до 10. В конкретных вариантах осуществления линкер содержит последовательность (GGGGS)n (SEQ ID NO: 173), где n равно от 1 до 10 или n равно от 1 до 5, например от 1 до 3. В дополнительных вариантах осуществления линкер содержит (GGGGGS)n (SEQ ID NO: 172), где n равно от 1 до 4, например от 1 до 3. Линкер может включать комбинации любого из вышеперечисленного, например, могут быть объединены повторы 2, 3, 4 или 5 линкеров GS, GGS, GGGGS и/или GGGGGS. В некоторых вариантах осуществления такой линкер имеет длину 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18 или 19 аминокислот.
[0271] В некоторых вариантах осуществления линкер представляет собой (в однобуквенном аминокислотном коде): GGS, GGGGS (SEQ ID NO: 149) или GGGGGS (SEQ ID NO: 135). В некоторых вариантах осуществления GS-линкер содержит аминокислотную последовательность GGSGGS, т.е. (GGS)2 (SEQ ID NO: 10); GGSGGSGGS, т.е. (GGS)3 (SEQ ID NO: 11); GGSGGSGGSGGS, т.е. (GGS)4 (SEQ ID NO: 12); GGSGGSGGSGGSGGS, то есть (GGS)5 (SEQ ID NO: 13); GGGGGSGGGGGSGGGGGS, то есть (G5S)3 (SEQ ID NO: 119), GGSGGGGSGGGGSGGGGS (SEQ ID NO: 147) и GGGGSGGGGSGGGGS (SEQ ID NO: 170). В некоторых вариантах осуществления линкер представляет собой GGGG (SEQ ID NO: 103). В некоторых из приведенных выше примеров серин можно заменить аланином (например, (Gly4Ala) или (Gly3Ala)).
[0272] В некоторых вариантах осуществления линкер включает пептидный линкер, имеющий аминокислотную последовательность GlyxXaa-Glyy-Xaa-Glyz (SEQ ID NO: 174), где каждый Xaa независимо выбран из аланина (Ala), валина (Val), лейцина (Leu), изолейцина (Ile), метионина (Met), фенилаланина (Phe), триптофана (Trp), пролина (Pro), глицина (Gly), серина (Ser), треонина (Thr), цистеина (Cys), тирозина (Tyr), аспарагина (Asn), глутамина (Gln), лизина (Lys), аргинина (Arg), гистидина (His), аспартата (Asp) и глутамата (Glu) и где каждый x, y и z это целое число в диапазоне от 1-5. В некоторых вариантах осуществления каждый Xaa независимо выбран из группы, состоящей из Ser, Ala и Thr. В конкретном варианте каждый из x, y и z равен 3 (тем самым получая пептидный линкер, имеющий аминокислотную последовательность Gly-Gly-Gly-Xaa-Gly-Gly-Gly-Xaa-Gly-Gly-Gly (SEQ ID NO: 175), где каждый Xaa выбран, как указано выше.
[0273] В некоторых вариантах осуществления линкер представляет собой обогащенный серином линкер на основе повторения мотива (SSSSG)y (SEQ ID NO: 185), где y равен по меньшей мере 1, хотя y может быть равен 2, 3, 4, 5, 6, 7, 8 и 9.
[0274] В некоторых случаях может быть желательно обеспечить некоторую жесткость пептидного линкера. Это может быть достигнуто путем включения пролиновых остатков в аминокислотную последовательность пептидного линкера. Таким образом, в некоторых вариантах осуществления линкер содержит по меньшей мере один пролиновый остаток в аминокислотной последовательности пептидного линкера. Например, пептидный линкер может иметь аминокислотную последовательность, в которой, по меньшей мере, 25% (например, по меньшей мере, 50% или, по меньшей мере, 75%) аминокислотных остатков представляют собой остатки пролина. В одном конкретном варианте осуществления пептидный линкер содержит только остатки пролина.
[0275] В некоторых аспектах пептидный линкер содержит по меньшей мере один остаток цистеина, как например, один остаток цистеина. Например, в некоторых вариантах осуществления линкер содержит по меньшей мере один остаток цистеина и аминокислотные остатки, выбранные из группы, состоящей из Gly, Ser, Ala и Thr. В некоторых таких вариантах осуществления линкер содержит остатки глицина и остатки цистеина, как например, только остатки глицина и остатки цистеина. Как правило, только один остаток цистеина будет включен в пептидный линкер. Один пример конкретного линкера, содержащего остаток цистеина, включает пептидный линкер, имеющий аминокислотную последовательность Glym-Cys-Glyn, где n и m, каждый, представляет собой целое число от 1 до 12, например от 3 до 9, от 4 до 8 или от 4 до 7. В конкретном варианте такой пептидный линкер имеет аминокислотную последовательность GGGGG-C-GGGGG (SEQ ID NO: 177).
[0276] В некоторых вариантах осуществления линкер слитого белка представляет собой структурированный или ограниченный линкер. В конкретных вариантах осуществления структурированный линкер содержит последовательность (AP)n или (EAAAK)n (SEQ ID NO: 178), где n равно от 2 до 20, предпочтительно от 4 до 10, включая, но не ограничиваясь этим, AS-(AP)n-GT (SEQ ID NO: 179) или AS-(EAAAK)n-GT (SEQ ID NO: 180), где n равно от 2 до 20, как например 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15. В других вариантах осуществления линкер содержит последовательности (GGGGA)n (SEQ ID NO: 181), (PGGGS)n (SEQ ID NO: 182), (AGGGS)n (SEQ ID NO: 183) или GGS-(EGKSSGSGSESKST)n-GGS (SEQ ID NO: 184, где n равно от 2 до 20. В некоторых вариантах осуществления линкер представляет собой SSSASASSA (SEQ ID NO: 186), GSPGSPG (SEQ ID NO: 187) или ATTTGSSPGPT (SEQ ID NO: 176). В некоторых вариантах осуществления такие линкеры в силу своей структуры могут быть более устойчивыми к протеолитической деградации, тем самым предлагая преимущество при инъекции in vivo.
[0277] В некоторых вариантах осуществления линкер не является расщепляемым линкером, также называемым нерасщепляемым линкером. В некоторых вариантах осуществления линкер не расщепляется протеазой. В некоторых вариантах осуществления линкер, который не является расщепляемым линкером или который не расщепляется протеазой, представляет собой линкер, который обычно стабилен для доставки in vivo или рекомбинантной продукции. В некоторых аспектах линкер, который не расщепляется протеазой, включает линкер, который не содержит по меньшей мере одну пептидную связь, которая предпочтительно лежит в пределах расщепляемой пептидной последовательности или сайта узнавания протеазы. В конкретных вариантах осуществления нерасщепляемый линкер не является целевым субстратом для протеазы, так что он не расщепляется преимущественно или специфически протеазой по сравнению с линкером, который содержит сайт распознавания субстрата для той же протеазы.
[0278] В некоторых вариантах осуществления линкер представляет собой расщепляемый линкер. В некоторых аспектах расщепляемый линкер представляет собой линкер, который включает последовательность, которая является субстратом для протеазы, благодаря наличию по меньшей мере одной связи, которая может быть разорвана в физиологических условиях. В некоторых случаях расщепляемый линкер подвержен расщеплению или чувствителен к расщеплению в определенных условиях, которые существуют in vivo, например, после воздействия внеклеточной протеазы, в том числе присутствующей в клеточных средах in vivo. В некоторых случаях протеаза может присутствовать в конкретном физиологическом микроокружении, таком как микроокружение опухоли, тем самым ограничивая участки, в которых может происходить расщепление.
[0279] Протеаза обычно проявляет специфичность или предпочтительность расщепления конкретного целевого субстрата по сравнению с другим нецелевым субстратом. Такая степень специфичности может быть определена на основе константы скорости расщепления последовательности, например линкера, которая является мерой предпочтения протеазы для ее субстрата и эффективности фермента. Любой метод определения скорости увеличения расщепления во времени в присутствии различных концентраций субстрата может быть использован для расчета константы специфичности. Например, субстрат связан с флуорогенным фрагментом, который высвобождается при расщеплении протеазой. Определяя скорость расщепления при различных концентрациях протеазы, можно определить константу специфичности расщепления (kcat/Km) для конкретной протеазы в отношении конкретного линкера. В некоторых вариантах осуществления расщепляемый линкер представляет собой линкер, который способен специфически расщепляться протеазой со скоростью, по меньшей мере, примерно 1 × 104 М-1с-1 или, по меньшей мере, 5 × 104 М-1с, по меньшей мере, 10 × 104 М-1с, по меньшей мере, 10 × 105 М-1с или более.
Расщепляемый линкер
[0280] В некоторых вариантах осуществления полиспецифические полипептидные конструкции по изобретению включают расщепляемый линкер, который соединяет первый и второй компоненты. В некоторых вариантах осуществления расщепляемый линкер включает аминокислотную последовательность, которая может служить субстратом для протеазы, обычно внеклеточной протеазы. Например, расщепляемый линкер может включать последовательность расщепления, содержащую по меньшей мере одну пептидную связь, которая предпочтительно лежит в пределах пептидной последовательности, расщепляемой протеазой. Подходящие протеазы включают, например, матричные металлопротеазы (ММР), цистеиновые протеазы, сериновые протеазы и активаторы плазмина, которые образуются или активируются интенсивно при заболеваниях, таких как ревматоидный артрит или злокачественное новообразование, что приводит к чрезмерной деградации ткани, воспалениям и метастазированию. В конкретных вариантах осуществления протеаза представляет собой протеазу, которая продуцируется опухолью, активированной иммунной эффекторной клеткой (например, Т-клеткой или NK-клеткой) или клеткой в микроокружении опухоли. В некоторых вариантах осуществления протеаза представляет собой гранзим B, матриптазу или MMP, такую как MMP-2.
[0281] Расщепляемый линкер может быть выбран на основе протеазы, которая продуцируется опухолью, которая находится вблизи клеток, которые экспрессируют мишень, и/или продуцируется опухолью, которая локализована в ткани с желаемой мишенью полиспецифических полипептидных конструкций. В литературе имеются сообщения о повышенных уровнях протеаз, имеющих известные субстраты в ряде злокачественных новообразований, например, в солидных опухолях. См., например, La Rocca et al. (2004) British J. of Cancer 90 (7): 1414-1421.
[0282] В некоторых вариантах осуществления расщепляемый линкер, который соединяет первый и второй компоненты полиспецифической полипептидной конструкции, расщепляется протеазой, продуцируемой иммунной эффекторной клеткой, которая активируется одним из компонентов. Например, полиспецифические полипептидные конструкции, которые включают активированную или усиленную Fc-область IgG, способны вызывать ADCC при взаимодействии с антигеном-мишенью. Центральным для ADCC является высвобождение гранзима B и перфорина из эффекторных клеток, а именно NK-клеток и цитотоксических T-клеток. После высвобождения гранзим В попадает в клетку-мишень перфорин-зависимым образом, где он опосредует апоптоз. Важно отметить, что гранзим B активен во внеклеточном синапсе между эффекторной клеткой и клеткой-мишенью. В некоторых вариантах осуществления расщепляемый линкер, который соединяет первый и второй компоненты полиспецифической полипептидной конструкции, расщепляется гранзимом B. Гранзим B высвобождается во время активации эффекторных клеток, опосредованной одним из компонентов полиспецифической полипептидной конструкции. В некоторых вариантах осуществления гранзим B и другие протеазы могут продуцироваться иммунными эффекторными клетками, включая активированные T-клетки или NK-клетки. В некоторых вариантах осуществления активация Т-клеток путем рекрутирования CD3 при связывании ТАА полиспецифической полипептидной конструкцией может высвобождать такие протеазы, которые затем могут расщеплять специфический расщепляемый линкер, тем самым усиливая или увеличивая активность CD3-связывающей молекулы для рекрутирования CD3. В некоторых вариантах осуществления расщепление может усиливать или увеличивать активность, достигаемую полиспецифической конструкцией при связывании с TAA в нерасщепленном состоянии.
[0283] Типичные субстраты включают, но не ограничиваются ими, субстраты, расщепляемые одним или более из следующих ферментов или протеаз: ADAMS, ADAMTS, например ADAM8; ADAM9; ADAM10; Адам-12; ADAM15; ADAM17/ТАХЭ; ADAMDEC1; ADAMTS1; ADAMTS4; ADAMTS5; аспартатпротеазы, например, BACE или ренин; аспарагиновые катепсины, например катепсин D или катепсин E; Каспазы, например каспаза 1, каспаза 2, каспаза 3, каспаза 4, каспаза 5, каспаза 6, каспаза 7, каспаза 8, каспаза 9, каспаза 10 или каспаза 14; цистеиновые катепсины, например, катепсин B, катепсин C, катепсин K, катепсин L, катепсин S, катепсин V/L2, катепсин X/Z/P; Цистеинпротеиназы, например, крузипаин; Легумаин; отубаин-2; KLK, например, KLK4, KLK5, KLK6, KLK7, KLK8, KLK10, KLK11, KLK13 или KLK14; Металлопротеиназы, например, меприн; неприлизин; PSMA; БМП-1; MMPs, например, MMP1, MMP2, MMP3, MMP7, MMP8, MMP9, MMP10, MMP11, MMP12, MMP13, MMP14, MMP15, MMP16, MMP17, MMP19, MMP20, MMP23, MMP24, MMP26 или MMP27, сериновые протеазы, например, активированный протеин C, катепсин A, катепсин G, химаза, протеаза фактора свертывания крови (например, FVIIa, FIXa, FXa, FXIa, FXIIa), эластаза, гранзим B, гуанидинобензоатаза, HtrA1, нейтрофильная эластаза человека, лактоферрин, NSA/3Aps, Maraps, Maraps, NaraA3, NSA/3, Maraps PACE4, плазмин, PSA, tPA, тромбин, триптаза, uPA; Трансмембранные сериновые протеазы типа II (TTSP), например, DESC1, DPP-4, FAP, гепсин, матриптаза-2, матриптаза, TMPRSS2, TMPRSS3 или TMPRSS4; и любая их комбинация.
[0284] В некоторых вариантах осуществления расщепляемый линкер расщепляется несколькими протеазами, например, 2 или более протеазами, 3 или более протеазами, 4 или более протеазами и так далее.
[0285] В некоторых вариантах осуществления расщепляемый линкер выбран для использования со специфической протеазой, например протеазой, которая, как известно, продуцируется опухолью, которая находится вблизи клеток, которые экспрессируют мишень, и/или продуцируется опухолью, которая совместно локализована с мишенью полиспецифической полипептидной конструкции.
[0286] В некоторых вариантах осуществления расщепляемый линкер содержит сайт распознавания субстрата или сайт расщепления для конкретной протеазы, который представляет собой последовательность, распознаваемую активным сайтом протеазы, которая расщепляется протеазой. Как правило, например, для сериновой протеазы последовательность расщепления состоит из аминокислот P1-P4 и P1'-P4' в субстрате, где расщепление происходит после положения P1. Как правило, последовательность расщепления для сериновой протеазы составляет шесть остатков в длину, чтобы соответствовать расширенной специфичности субстрата многих протеаз, но может быть длиннее или короче в зависимости от протеазы. Как правило, расщепляемый линкер включает в себя расщепляющуюся последовательность связи P1-P1', которая распознается протеазой. В некоторых аспектах расщепляемый линкер сконструирован для введения пептидной связи, способной расщепляться специфической протеазой, например, путем введения последовательности сайта узнавания субстрата или последовательности расщепления протеазы.
[0287] В некоторых вариантах осуществления расщепляемый линкер включает комбинацию двух или более последовательностей субстрата. В некоторых вариантах осуществления каждая последовательность субстрата расщепляется одной и той же протеазой. В некоторых вариантах осуществления по меньшей мере две субстратные последовательности расщепляются разными протеазами. В некоторых вариантах осуществления расщепляемый линкер содержит аминокислоту, которая является субстратом для гранзима В. В некоторых вариантах осуществления расщепляемый гранзимом В линкер содержит аминокислотную последовательность, имеющую общую формулу P4 P3 P2 P1 ↓ P1' (SEQ ID NO: 150), где P4 представляет собой аминокислоту I, L, Y, M, F, V или A; P3 представляет собой аминокислоту A, G, S, V, E, D, Q, N или Y; P2 представляет собой аминокислоту H, P, A, V, G, S или T; P1 представляет собой аминокислоту D или E; и P1' представляет собой аминокислоту I, L, Y, M, F, V, T, S, G или A. В некоторых вариантах осуществления расщепляемый гранзимом B линкер содержит аминокислотную последовательность, имеющую общую формулу P4 P3 P2 P1 ↓ P1' (SEQ ID NO: 151), где P4 представляет собой аминокислоту I или L; P3 является аминокислотой E; P2 представляет собой аминокислоту P или A; P1 является аминокислотой D; и P1' представляет собой аминокислоту I, V, T, S или G.
[0288] В некоторых вариантах осуществления субстрат для гранзима B содержит аминокислотную последовательность LEAD (SEQ ID NO: 22), LEPG (SEQ ID NO: 142) или LEAE (SEQ ID NO: 143). В некоторых вариантах осуществления расщепляемый линкер содержит аминокислотную последовательность IEPDI (SEQ ID NO: 136), LEPDG (SEQ ID NO: 152, LEADT (SEQ ID NO: 137), IEPDG (SEQ ID NO: 138), IEPDV (SEQ ID NO: 139), IEPDS (SEQ ID NO: 140), IEPDT (SEQ ID NO: 141), IEPDP (SEQ ID NO: 144), LEPDG (SEQ ID NO: 152) или LEADG (SEQ ID NO: 153).
[0289] В некоторых вариантах осуществления расщепляемый линкер содержит аминокислоту, которая является субстратом для матриптазы. В некоторых вариантах осуществления расщепляемый линкер содержит последовательность P4QAR↓ (A/V) (SEQ ID NO: 154), где P4 представляет собой любую аминокислоту. В некоторых вариантах осуществления расщепляемый линкер содержит последовательность RQAR (A/V) (SEQ ID NO: 155). В некоторых вариантах осуществления субстрат для матриптазы содержит аминокислотную последовательность RQAR (SEQ ID NO: 23). В некоторых вариантах осуществления расщепляемый линкер содержит аминокислотную последовательность RQARV (SEQ ID NO: 156).
[0290] В некоторых вариантах осуществления расщепляемый линкер содержит аминокислоту, которая является субстратом для одной или более матриксных металлопротеаз (MMP). В некоторых вариантах осуществления MMP представляет собой MMP-2. В некоторых вариантах осуществления расщепляемый линкер содержит общую формулу P3 P2 P1↓ P1' (SEQ ID NO: 157), где P3 представляет собой P, V или A; P2 является Q или D; P1 является A или N; и P1' представляет собой L, I или M. В некоторых вариантах осуществления расщепляемый линкер содержит общую формулу P3 P2 P1 ↓ P1' (SEQ ID NO: 158), где P3 представляет собой P; P2 представляет собой Q или D; P1 представляет собой A или N; и P1' представляет собой L или I. В некоторых вариантах осуществления субстрат для MMP содержит аминокислотную последовательность PAGL (SEQ ID NO: 24).
[0291] В некоторых вариантах осуществления расщепляемый линкер содержит комбинацию аминокислотной последовательности, которая является субстратом для гранзима B, и аминокислотной последовательности, которая является субстратом для матриптазы. В некоторых вариантах осуществления расщепляемый линкер содержит комбинацию аминокислотной последовательности LEAD (SEQ ID NO: 22) и аминокислотной последовательности RQAR (SEQ ID NO: 23).
[0292] В некоторых вариантах осуществления расщепляемый линкер содержит комбинацию аминокислотной последовательности, которая является субстратом для гранзима B, и аминокислотной последовательности, которая является субстратом для MMP. В некоторых вариантах осуществления расщепляемый линкер содержит комбинацию аминокислотной последовательности LEAD (SEQ ID NO: 22) и аминокислотной последовательности PAGL (SEQ ID NO: 24).
[0293] В некоторых вариантах осуществления расщепляемый линкер содержит комбинацию аминокислотной последовательности, которая является субстратом для матриптазы, и аминокислотной последовательности, которая является субстратом для ММР. В некоторых вариантах осуществления расщепляемый линкер содержит комбинацию аминокислотной последовательности RQAR (SEQ ID NO: 23) и аминокислотной последовательности PAGL (SEQ ID NO: 24).
[0294] В некоторых вариантах осуществления расщепляемый линкер содержит комбинацию аминокислотной последовательности, которая является субстратом для гранзима B, аминокислотной последовательности, которая является субстратом для матриптазы, и аминокислотной последовательности, которая является субстратом для ММР. В некоторых вариантах осуществления расщепляемый линкер содержит комбинацию аминокислотной последовательности, которая является субстратом для гранзима B, и аминокислотной последовательности, которая является субстратом для MMP. В некоторых вариантах осуществления расщепляемый линкер содержит комбинацию аминокислотной последовательности LEAD (SEQ ID NO: 22), аминокислотной последовательности RQAR (SEQ ID NO: 23) и аминокислотной последовательности PAGL (SEQ ID NO: 24).
[0295] Расщепляемый линкер может включать любые известные линкеры. Примеры расщепляемых линкеров описаны в Be'liveau et al. (2009) FEBS Journal, 276; Опубликованная заявка США № US20160194399; US20150079088; US20170204139; US20160289324; US20160122425; US20150087810; US20170081397; Патент США № US9644016.
[0296] В некоторых вариантах осуществления расщепляемый линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из TGLEADGSPAGLGRQARVG (SEQ ID NO: 25); TGLEADGSRQARVGPAGLG (SEQ ID NO: 26); TGSPAGLEADGSRQARVGS (SEQ ID NO: 27); TGPAGLGLEADGSRQARVG (SEQ ID NO: 28); TGRQARVGLEADGSPAGLG (SEQ ID NO: 29); TGSRQARVGPAGLEADGS (SEQ ID NO: 30); и TGPAGLGSRQARVGLEADGS (SEQ ID NO: 31); GPAGLGLEPDGSRQARVG (SEQ ID NO: 104); GGSGGGGIEPDIGGSGGS (SEQ ID NO: 105); GGSGGGGLEADTGGSGGS (SEQ ID NO: 106); GSIEPDIGS (SEQ ID NO: 107); GSLEADTGS (SEQ ID NO: 108); GGSGGGGIEPDGGGSGGS (SEQ ID NO: 109); GGSGGGGIEPDVGGSGGS (SEQ ID NO: 110); GGSGGGGIEPDSGGSGGS (SEQ ID NO: 111); GGSGGGGIEPDTGGSGGS (SEQ ID NO: 112); GGGSLEPDGSGS (SEQ ID NO: 113); и GPAGLGLEADGSRQARVG (SEQ ID NO: 114), GGEGGGGSGGSGGGS (SEQ ID NO: 115); GSSAGSEAGGSGQAGVGS (SEQ ID NO: 116); GGSGGGGLEAEGSGGGGS (SEQ ID NO: 117); GGSGGGGIEPDPGGSGGS (SEQ ID NO: 118); TGGSGGGGIEPDIGGSGGS (SEQ ID NO: 148).
4. Антигенсвязывающие домены:
[0297] Полиспецифические полипептидные конструкции по настоящему изобретению включают, по меньшей мере, один антигенсвязывающий домен, такой как, по меньшей мере, первый антигенсвязывающий домен и второй антигенсвязывающий домен. В некоторых аспектах антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов выбран из антитела или антигенсвязывающего фрагмента, родственного партнера по связыванию, антикалина (модифицированного липокалина), дарпина, финомера, центрина (сконструированный домен фибронетикина III), домена с цистиновым узлом, Аффилина, Аффитела или сконструированного домена CH3. В некоторых вариантах осуществления природный родственный партнер по связыванию содержит внеклеточный домен или его связывающий фрагмент нативного родственного партнера по связыванию TAA или его вариант, который проявляет активность связывания с TAA.
[0298] В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов, такой как первый антигенсвязывающий домен и второй антигенсвязывающие домены, включает в себя одну или более копий антитела или его антигенсвязывающего фрагмента. В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов, такой как первый антигенсвязывающий домен и второй антигенсвязывающие домены, включает в себя одну или более копий антитела или его антигенсвязывающего фрагмента, выбранного из группа, состоящей из Fab-фрагмента, F(ab')2-фрагмента, Fv-фрагмента, scFv, scAb, dAb, однодоменного антитела тяжелой цепи и однодоменного антитела легкой цепи. В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов, такой как первый антигенсвязывающий домен и второй антигенсвязывающие домены, включает в себя один или более фрагментов однодоменного антитела (sdAb), например VHH, VNAR, сконструированные VH или VK домены. VHH могут быть получены из природных антител, содержащих только верблюжьи тяжелые цепи, из генетически модифицированных грызунов, которые продуцируют только антитела из тяжелых цепей, или из наивных/синтетических библиотек верблюжьих антител или гуманизированных верблюжьих однодоменных антител. VNAR могут быть получены только из антител тяжелой цепи хрящевой рыбы. Различные способы были реализованы для генерации мономерных sdAb из традиционно гетеродимерных доменов VH и VK, включая конструирование интерфейса и выбор определенных семейств зародышевой линии. В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов, такой как первый антигенсвязывающий домен и/или второй антигенсвязывающий домен полиспецифических полипептидных конструкций содержит последовательности VH и VL, собранные в виде FAB или scFv. В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов, такой как первый антигенсвязывающий домен и/или второй антигенсвязывающий домен полиспецифических полипептидных конструкций содержит связывающие домены в виде однодоменных антител (sdAb).
[0299] В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов представляет собой или включает внеклеточный домен или его связывающий фрагмент нативного родственного партнера по связыванию TAA или его вариант, который проявляет активность связывания с TAA.
[0300] В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов, такой как первый антигенсвязывающий домен и второй антигенсвязывающий домен, связываются с одним и тем же антигеном. В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов, такой как первый антигенсвязывающий домен и второй антигенсвязывающий домен, связываются с другим антигеном. В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов, такой как первый антигенсвязывающий домен и второй антигенсвязывающий домен, связываются с одним и тем же опухолеспецифическим антигеном (TAA). В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов, такой как первый антигенсвязывающий домен и второй антигенсвязывающий домен, связываются с разными TAA. В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов, такой как первый антигенсвязывающий домен и второй антигенсвязывающий домен, связываются с другим эпитопом на одном и том же TAA. В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов, такой как первый антигенсвязывающий домен и второй антигенсвязывающий домен, связываются с одним и тем же эпитопом на одном и том же TAA.
[0301] В некоторых вариантах осуществления антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов, которые связываются с TAA, приводит к моновалентному, двухвалентному, трехвалентному или четырехвалентному связыванию с TAA.
[0302] В некоторых вариантах осуществления TAA выбран из группы, состоящей из 1-92-LFA-3, 5T4, альфа-4 интегрина, альфа-V интегрина, альфа4бета1 интегрина, альфа4бета7 интегрина, AGR2, анти-Lewis-Y, рецептора апелина J, APRIL, B7-H3, B7-H4, BAFF, BTLA, компонента комплемента C5, C-242, CA9, CA19-9, (Lewis а), карбоангидразы 9, CD2, CD3, CD6, CD9, CD11a, CD19, CD20, CD22, CD24, CD25, CD27, CD28, CD30, CD33, CD38, CD40, CD40L, CD41, CD44, CD44v6, CD47, CD51, CD52, CD56, CD64, CD70, CD71, CD74, CD80, CD81, CD86, CD95, CD117, CD123, CD125, CD132, (IL-2RG), CD133, CD137, CD138, CD166, CD172A, CD248, CDH6, CEACAM5 (CEA), CEACAM6 (NCA-90), CLAUDIN-3, CLAUDIN-4, cMet, коллагена, Cripto, CSFR, CSFR-1, CTLA-4, CTGF, CXCL10, CXCL13, CXCR1, CXCR2, CXCR4, CYR61, DL44, DLK1, DLL3, DLL4, DPP-4, DSG1, EDA, EDB, EGFR, EGFRviii, рецептора эндотелина B (ETBR), ENPP3, EpCAM, EPHA2, EPHB2, ERBB3, F-протеина RSV, FAP, FGF-2, FGF8, FGFR1, FGFR2, FGFR3, FGFR4, FLT-3, рецептора альфа фолата (FR α), GAL3ST1, G-CSF, G-CSFR, GD2, GITR, GLUT1, GLUT4, GM-CSF, GM-CSFR, рецепторов GP IIb/IIIa, Gp130, GPIIB/IIIA, GPNMB, GRP78, HER2/neu, HER3, HER4, HGF, hGH, HVEM, гиалуронидазы, ICOS, IFNальфа, IFNбета, IFNгамма, IgE, рецептора IgE (FceRI), IGF, IGF1R, IL1B, IL1R, IL2, IL11, IL12, IL12p40, IL-12R, IL-12Rbeta1, IL13, IL13R, IL15, IL17, IL18, IL21, IL23, IL23R, IL27/IL27R (wsx1), IL29, IL-31R, IL31/IL31R, IL2R, IL4, IL4R, IL6, IL6R, рецептора инсулина, Jagged лигандов, Jagged 1, Jagged 2, KISS1-R, LAG-3, LIF-R, Lewis X, LIGHT, LRP4, LRRC26, Ly6G6D, LyPD1, MCSP, мезотелина, MRP4, MUC1, муцина-16 (MUC16, CA-125), Na/K-АТФазы, NGF, никастрина, рецепторов Notch, Notch 1, Notch 2, Notch 3, Notch 4, NOV, OSM-R, OX-40, PAR2, PDGF-AA, PDGF-BB, PDGFRальфа, PDGFRбета, PD-1, PD-L1, PD-L2, фосфатидилсерина, P1GF, PSCA, PSMA, PSGR, RAAG12, RAGE, SLC44A4, фосфат сфингозина 1, STEAP1, STEAP2, TAG-72, TAPA1, TE M-8, TGFбета, TIGIT, TIM-3, TLR2, TLR4, TLR6, TLR7, TLR8, TLR9, TMEM31, TNFальфа, TNFR, TNFRS12A, TRAIL-R1, TRAIL-R2, трансферрина, рецептора трансферрина, TRK-A, TRK-B, uPAR, VAP1, VCAM-1, VEGF, VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGFR1, VEGFR2, VEGFR3, VISTA, WISP-1, WISP-2 и WISP-3.
[0303] В некоторых вариантах осуществления, по меньшей мере, один антигенсвязывающий домен, или независимо каждый антигенсвязывающий домен связывается с опухолеспецифическим антигеном (ТАА) фолатного рецептора альфа (FRα). Например, антигенсвязывающий домен содержит связывающий домен в виде sdAb, который связывает FRα. Примеры FRα-связывающих sdAb представлены в SEQ ID NO: 120, 121, и 122.
[0304] В некоторых вариантах осуществления по меньшей мере один антигенсвязывающий домен или независимо каждый антигенсвязывающий домен связывается с опухолеспецифическим антигеном (TAA) cMET. Например, антигенсвязывающий домен содержит связывающий домен в виде sdAb, который связывается с cMET. Пример cMET-связывающего sdAb представлен в SEQ ID NO: 123 (патент США № 9346884).
[0305] В некоторых вариантах осуществления по меньшей мере один антигенсвязывающий домен или независимо каждый антигенсвязывающий домен связывается с опухолесипецифическим антигеном (TAA) B7H3. Например, антигенсвязывающий домен содержит связывающий домен в виде scFv, который связывается с B7H3. Пример BFH-связывающего scFv представлен в SEQ ID NO: 124. В некоторых вариантах осуществления антигенсвязывающий домен представляет собой или содержит Fab-фрагмент антитела, содержащий VH-CH1 (Fd) и LC. Пример B7H3 Fd представлен в SEQ ID NO: 127, а пример B7H3 LC представлен в SEQ ID NO: 128 (публикация PCT № WO2017/030926).
[0306] В некоторых вариантах осуществления по меньшей мере один антигенсвязывающий домен или независимо каждый антигенсвязывающий домен связывается с опухолеспецифическим антигеном (TAA) CD20. Например, антигенсвязывающий домен содержит связывающий домен в виде scFv, который связывается с CD20. Примеры CD20-связывающих scFv представлены в SEQ ID NO: 125, 189 и 190 (публикация США № US 2005/0123546).
[0307] В некоторых вариантах осуществления по меньшей мере один антигенсвязывающий домен или независимо каждый антигенсвязывающий домен связывается с опухолеспецифическим антигеном (TAA) DLL3. Например, антигенсвязывающий домен содержит связывающий домен в виде scFv, который связывается с DLL3. Примеры DLL3-связывающих scFv представлены в SEQ ID NO: 126 и 189 (публикация США № US 2017/0037130). В некоторых вариантах осуществления антигенсвязывающий домен представляет собой или содержит Fab-фрагмент антитела, содержащий Fd и LC, который связывается с DLL3. Пример DLL3 Fd представлен в SEQ ID NO: 133, а пример DLL3 LC представлен в SEQ ID NO: 134 (патенте США № 8044178).
[0308] В некоторых вариантах осуществления по меньшей мере один антигенсвязывающий домен или независимо каждый антигенсвязывающий домен связывается с опухолесипецифическим антигеном (TAA) 5T4. Пример 5T4 Fd представлен в SEQ ID NO: 129, а пример 5T4 LC представлен в SEQ ID NO: 130. В некоторых вариантах осуществления связывающий домен антитела содержит VH-CH1 (Fd) или VL-CL, как представлено в SEQ ID NO: 167 и 168 (патент США № 8044178).
[0309] В некоторых вариантах осуществления по меньшей мере один антигенсвязывающий домен или независимо каждый антигенсвязывающий домен связывается с опухолеспецифическим антигеном (TAA) gpNMB. В некоторых вариантах осуществления антигенсвязывающий домен представляет собой или содержит Fab-фрагмент, содержащий цепь Fd и LC. Пример gpNMB Fd представлен в SEQ ID NO: 131, а пример gpNMB LC представлен в SEQ ID NO: 132.
[0310] В некоторых вариантах осуществления антигенсвязывающий домен прямо или косвенно связан через линкер с Fc-областью и/или с CD3-связывающей областью. В некоторых вариантах осуществления связь осуществляется через линкер. В некоторых вариантах осуществления линкер представляет собой связывающий пептид (LP), который может включать в себя любой гибкий или жесткий линкер, как описано в разделе II.3, хотя обычно пептиды, связывающие антигенсвязывающий домен или домены, не являются расщепляемыми линкерами.
[0311] В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит первый связывающий пептид (LP1) между первым антигенсвязывающим доменом и Fc-областью. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит второй связывающий пептид (LP2) между CD3-связывающей областью и вторым антигенсвязывающим доменом. В некоторых вариантах осуществления полиспецифическая полипептидная конструкция содержит первый связывающий пептид (LP1) между первым антигенсвязывающим доменом и Fc-областью и второй связывающий пептид (LP2) между CD3-связывающей областью и вторым антигенсвязывающим доменом. В некоторых аспектах полиспецифическая полипептидная конструкция имеет структурное расположение от N-конца к C-конца следующим образом: первый антигенсвязывающий домен-LP1-Fc-область-линкер-CD3-связывающая область-LP2-второй антигенсвязывающий домен. В некоторых вариантах осуществления два связывающих пептида не являются идентичными друг другу.
[0312] В некоторых вариантах осуществления LP1 или LP2 независимо представляют собой пептид длиной примерно от 1 до 20 аминокислот.В некоторых вариантах осуществления LP1 или LP2 независимо представляют собой пептид, который представляет собой или содержит любой линкер Gly-Ser, как представлено в SEQ ID NO: 10-13, 119, 135, 147, 149 или GGS.
III. Фармацевтическая композиция
[0313] В настоящем описании представлены композиции любой из представленных полиспецифических полипептидных конструкций. Понятно, что введение терапевтических сущностей в соответствии с раскрытием будет осуществляться с подходящими носителями, вспомогательными веществами и другими агентами, которые включены в составы для обеспечения улучшенного переноса, доставки, переносимости и тому подобного. Множество подходящих составов можно найти в формулярах, известных всем химикам-фармацевтам: Remington's Pharmaceutical Sciences (15-е изд., Mack Publishing Company, Easton, PA (1975)), в частности глава 87, Blaug, Seymour. Эти составы включают, например, порошки, пасты, мази, желе, воски, масла, липиды, содержащие липиды (катионные или анионные) везикулы (такие как LIPOFECTIN™), конъюгаты ДНК, безводные абсорбционные пасты, эмульсии масло-в-воде и вода-в-масле, эмульсии карбовакс (полиэтиленгликоли различной молекулярной массы), полутвердые гели и полутвердые смеси, содержащие карбовакс. Любая из вышеперечисленных смесей может быть пригодной для лечения и терапии в соответствии с настоящим раскрытием при условии, что активный ингредиент в составе не инактивируется составом, а состав является физиологически совместимым и переносимым при введении. См. также Baldrick P. «Pharmaceutical excipient development: the need for preclinical guidance.» Regul. Toxicol Pharmacol. 32(2):210-8 (2000), Wang W. «Lyophilization and development of solid protein pharmaceuticals», Int. J. Pharm. 203(1-2):1-60 (2000), Charman WN «Lipids, lipophilic drugs, and oral drug delivery-some emerging concepts», J Pharm Sci.89 (8): 967-78 (2000), Powell et al. «Compendium of excipients for parenteral formulations» PDA J Pharm Sci Technol. 52:238-311 (1998) и ссылки на них для получения дополнительной информации, относящейся к составам, наполнителям и носителям, хорошо известным химикам-фармацевтам.
[0314] В некоторых вариантах осуществления полиспецифические полипептидные конструкции, конъюгированные полиспецифические полипептидные конструкции и их композиции, именуемые в настоящем описании в совокупности терапевтическими средствами и их производными, фрагментами, аналогами и гомологами, могут быть включены в фармацевтические композиции, подходящие для введения. Принципы и соображения, связанные с приготовлением таких композиций, а также рекомендации по выбору компонентов представлены, например, в Remington's Pharmaceutical Sciences: The Science And Practice of Pharmacy, 19th ed. (Alfonso R. Gennaro, et al., editors) Mack Pub. Co., Easton, Pa.: 1995; Drug Absorption Enhancement: Concepts, Possibilities, Limitations, And Trends, Harwood Academic Publishers, Langhorne, Pa., 1994; и Peptide And Protein Drug Delivery (Advances In Parenteral Sciences, Vol. 4), 1991, M. Dekker, New York.
[0315] Такие композиции обычно включают полиспецифическую полипептидную конструкцию или ее конъюгированный и фармацевтически приемлемый носитель. Когда полиспецифическая полипептидная конструкция включает фрагмент антитела, можно использовать наименьший фрагмент антитела, который специфически связывается с белком-мишенью. Например, на основе последовательностей вариабельной области антитела могут быть сконструированы пептидные молекулы, которые сохраняют способность антитела связывать последовательность белка-мишени. Такие пептиды могут быть синтезированы химически и/или получены с помощью технологии рекомбинантных ДНК. (См., например, Marasco et al., Proc. Natl.Acad. Sci. USA, 90: 7889-7893 (1993)).
[0316] Используемый в настоящем описании термин «фармацевтически приемлемый носитель» предназначен для включения любых и всех растворителей, дисперсионных сред, покрытий, антибактериальных и противогрибковых агентов, изотонических и задерживающих абсорбцию агентов и тому подобного, совместимых с фармацевтическим введением. Подходящие носители описаны в самом последнем издании Remington's Pharmaceutical Sciences, стандартном справочном тексте в данной области, который включен в настоящий документ посредством ссылки. Подходящие примеры таких носителей или разбавителей включают, но не ограничиваются ими, воду, физиологический раствор, растворы Рингера, раствор декстрозы и 5% сывороточного альбумина человека. Липосомы и неводные носители, такие как жирные масла, также могут быть использованы. Использование таких сред и агентов для фармацевтически активных веществ хорошо известно в данной области. За исключением случаев, когда какой-либо обычный носитель или агент несовместим с активным соединением, предполагается их применение в композициях.
[0317] Составы, используемые для введения in vivo, должны быть стерильными. Это легко достигается путем фильтрации через стерильные фильтрующие мембраны.
[0318] Фармацевтическая композиция по изобретению приготовлена так, чтобы она была совместима с предполагаемым путем введения. Примеры путей введения включают парентеральное, например, внутривенное, внутрикожное, подкожное, пероральное (например, ингаляционное), трансдермальное (т.е. местное), трансмукозальное и ректальное введение. Растворы или суспензии, используемые для парентерального, внутрикожного или подкожного применения, могут включать следующие компоненты: стерильный разбавитель, такой как вода для инъекций, физиологический раствор, жирные масла, полиэтиленгликоли, глицерин, пропиленгликоль или другие синтетические растворители; антибактериальные агенты, такие как бензиловый спирт или метилпарабены; антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия; хелатообразующие агенты, такие как этилендиаминтетрауксусная кислота (ЭДТА); буферы, такие как ацетаты, цитраты или фосфаты, и агенты для регуляции тоничности, такие как хлорид натрия или декстроза. РН можно регулировать с помощью кислот или оснований, таких как соляная кислота или гидроксид натрия. Парентеральный препарат может быть заключен в ампулы, одноразовые шприцы или флаконы с множественными дозами, сделанные из стекла или пластика.
[0319] Фармацевтические композиции, подходящие для инъекционного применения, включают стерильные водные растворы (где растворимы в воде) или дисперсии и стерильные порошки для немедленного приготовления стерильных инъекционных растворов или дисперсии. Для внутривенного введения подходящие носители включают физиологический раствор, бактериостатическую воду, Cremophor ELTM (BASF, Parsippany, NJ) или фосфатнобуферный солевой раствор (PBS). Во всех случаях композиция должна быть стерильной и должна быть жидкой до такой степени, чтобы ее можно было легко вводить через шприц. Она должна быть стабильной в условиях производства и хранения и должна быть защищена от загрязняющего действия микроорганизмов, таких как бактерии и грибки. Носителем может быть растворитель или дисперсионная среда, содержащая, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и тому подобное) и их подходящие смеси. Надлежащая текучесть может поддерживаться, например, путем использования покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем использования поверхностно-активных веществ. Предотвращение действия микроорганизмов может быть достигнуто с помощью различных антибактериальных и противогрибковых агентов, например, парабенов, хлорбутанола, фенола, аскорбиновой кислоты, тимеросала и тому подобного. Во многих случаях будет целесообразно включать в композицию изотонические агенты, например, сахара, полиспирты, такие как манит, сорбит, хлорид натрия. Длительная абсорбция инъецируемых композиций может быть достигнута включением в композицию агента, который задерживает абсорбцию, например моностеарата алюминия и желатина.
[0320] Стерильные растворы для инъекций могут быть получены путем включения активного соединения в необходимом количестве в подходящем растворителе с одним или комбинацией ингредиентов, перечисленных выше, по мере необходимости, с последующей стерилизацией фильтрованием. Обычно дисперсии готовят путем включения активного соединения в стерильный носитель, который содержит основную дисперсионную среду и требуемые другие ингредиенты из перечисленных выше. В случае стерильных порошков для приготовления стерильных растворов для инъекций, способами приготовления являются вакуумная сушка и сублимационная сушка (лиофилизация), которые дают порошок активного ингредиента плюс любой дополнительный искомый ингредиент из его предварительно стерильно отфильтрованного раствора. Если композиция лиофилизирована, стерилизация с использованием этого способа может проводиться либо до, либо после лиофилизации и восстановления. Композиция для парентерального введения может храниться в лиофилизированной форме или в растворе. Кроме того, парентеральные композиции обычно помещают в контейнер, имеющий стерильное отверстие для доступа, например, пакет для внутривенного раствора или флакон, имеющий пробку, прокалываемую иглой для подкожных инъекций.
[0321] В некоторых вариантах осуществления фармацевтическую композицию вводят пациенту любым путем, в том числе перорально, трансдермально, путем ингаляции, внутривенно, внутриартериально, внутримышечно, путем прямого нанесения на участок раны, нанесения на хирургический участок, внутрибрюшинно, через суппозиторий, подкожно, внутрикожно, чрескожно, путем распыления, внутриплеврально, интравентрикулярно, внутрисуставно, интраокулярно или внутриспинально.
[0322] Пероральные композиции обычно включают инертный разбавитель или пищевой носитель. Они могут быть заключены в желатиновые капсулы или спрессованы в таблетки. С целью перорального терапевтического введения активное соединение может быть включено с вспомогательнымт веществами и использоваться в форме таблеток, пастилок или капсул. Пероральные композиции также могут быть приготовлены с использованием жидкого носителя для использования в качестве жидкости для полоскания рта, где соединение в жидком носителе наносится перорально и путем полоскания и откашливается или проглатывается. Фармацевтически совместимые связующие агенты и/или адъювантные вещества могут быть включены в состав композиции. Таблетки, пилюли, капсулы, пастилки и тому подобное могут содержать любой из следующих ингредиентов или соединений аналогичной природы: связующее вещество, такое как микрокристаллическая целлюлоза, трагакантовая камедь или желатин; вспомогательное вещество, такое как крахмал или лактоза, дезинтегрирующий агент, такой как альгиновая кислота, примогель или кукурузный крахмал; смазывающее вещество, такое как стеарат магния или стеротес; глидант, такой как коллоидный диоксид кремния; подсластитель, такой как сахароза или сахарин; или ароматизатор, такой как мята перечная, метилсалицилат или апельсиновый ароматизатор.
[0323] Для введения путем ингаляции полиспецифическая полипептидная конструкция доставляется в форме аэрозольного спрея из находящегося под давлением контейнера или дозатора, который содержит подходящий пропеллент, например газ, такой как диоксид углерода, или распылитель.
[0324] Системное введение также может осуществляться трансмукозальным или трансдермальным способом. Для трансмукозального или трансдермального введения в препарате используются пенетранты, подходящие для проникновения через барьер. Такие пенетранты обычно известны в данной области и включают, например, для трансмукозального введения, детергенты, соли желчных кислот и производные фузидиевой кислоты. Введение через слизистую оболочку может быть достигнуто путем использования назальных спреев или суппозиториев. Для трансдермального введения активные соединения готовят в виде мазей, мазей, гелей или кремов, общеизвестных в данной области.
[0325] Соединения также могут быть получены в форме суппозиториев (например, с обычными основами для суппозиториев, такими как масло какао и другие глицериды) или удерживающих клизм для ректальной доставки.
[0326] В одном варианте осуществления терапевтические средства получают с носителями, которые будут защищать соединение от быстрого выведения из организма, такими как составы с замедленным/контролируемым высвобождением, включая имплантаты и микроинкапсулированные системы доставки. Могут быть использованы биоразлагаемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Способы приготовления таких составов будут очевидны для специалистов в данной области.
[0327] Например, терапевтические средства могут быть заключены в микрокапсулы, приготовленные, например, методами коацервации или межфазной полимеризацией, например, гидроксиметилцеллюлозой или желатин-микрокапсулами и поли-(метилметакрилатными) микрокапсулами, соответственно, в коллоидных системах доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсиях.
[0328] В некоторых вариантах осуществления фармацевтическая композиция содержит фармацевтически приемлемое вспомогательное вещество, например наполнитель, связующее, покрытие, консервант, смазывающее вещество, ароматизатор, подсластитель, краситель, растворитель, буферный агент, хелатирующий агент или стабилизатор. Примеры фармацевтически приемлемых наполнителей включают целлюлозу, двухосновный фосфат кальция, карбонат кальция, микрокристаллическую целлюлозу, сахарозу, лактозу, глюкозу, маннит, сорбит, мальтол, прежелатинизированный крахмал, кукурузный крахмал или картофельный крахмал. Примеры фармацевтически приемлемых связующих включают поливинилпирролидон, крахмал, лактозу, ксилит, сорбит, мальтит, желатин, сахарозу, полиэтиленгликоль, метилцеллюлозу или целлюлозу. Примеры фармацевтически приемлемых покрытий включают гидроксипропилметилцеллюлозу (HPMC), шеллак, кукурузный белок, зеин или желатин. Примеры фармацевтически приемлемых дезинтегрантов включают поливинилпирролидон, карбоксиметилцеллюлозу или крахмалгликолят натрия. Примеры фармацевтически приемлемых смазывающих веществ включают полиэтиленгликоль, стеарат магния или стеариновую кислоту. Примеры фармацевтически приемлемых консервантов включают метилпарабены, этилпарабены, пропилпарабен, бензойную кислоту или сорбиновую кислоту. Примеры фармацевтически приемлемых подсластителей включают сахарозу, сахарин, аспартам или сорбит. Примеры фармацевтически приемлемых буферных агентов включают карбонаты, цитраты, глюконаты, ацетаты, фосфаты или тартраты.
[0329] Могут быть приготовлены препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих антитело, причем такие матрицы имеют форму профилированных изделий, например пленки или микрокапсулы. В некоторых вариантах осуществления фармацевтическая композиция дополнительно содержит агент для контролируемого или замедленного высвобождения продукта, такой как инъецируемые микросферы, биоразлагаемые частицы, полимерные соединения (полимолочная кислота, полигликолевая кислота), гранулы или липосомы. Примеры матриц замедленного высвобождения включают сложные полиэфиры, гидрогели (например, поли (2-гидроксиэтилметакрилат) или поли (виниловый спирт)), полилактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и γ-этил- L-глутамата, неразлагаемый этилен-винилацетат, разлагаемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOTTM (инъецируемые микросферы, состоящие из сополимера молочной кислоты-гликолевой кислоты и ацетата лейпролида) и поли-D-(-)-3-гидрокси-масляной кислоты. Хотя такие полимеры, как этилен-винилацетат и молочная кислота-гликолевая кислота, позволяют высвобождать молекулы в течение более 100 дней, некоторые гидрогели высвобождают белки в течение более коротких периодов времени.
[0330] Материалы также могут быть получены из коммерческих источников от Alza Corporation и Nova Pharmaceuticals, Inc. Липосомные суспензии (включая липосомы, нацеленные на инфицированные клетки с моноклональными антителами к вирусным антигенам) и также могут быть использованы в качестве фармацевтически приемлемых носителей. Они могут быть получены в соответствии со способами, известными специалистам в данной области, например, как описано в патенте США № 4522811.
[0331] Особенно выгодно готовить пероральные или парентеральные композиции в единичной дозированной форме для простоты введения и единообразия дозировки. Используемая в настоящем описании единичная лекарственная форма относится к физически дискретным единицам, подходящим в качестве единичных доз для пациента, подлежащего лечению; каждая единица содержит заданное количество активного соединения, рассчитанное для получения желаемого терапевтического эффекта в сочетании с необходимым фармацевтическим носителем. Спецификация для единичных дозированных форм раскрытия продиктована и напрямую зависит от уникальных характеристик активного соединения и конкретного терапевтического эффекта, который должен быть достигнут, а также от ограничений, присущих методу приготовления такого активного соединения для лечения пациентов.
[0332] Кроме того, представлены наборы, содержащие фармацевтические композиции (или изделия), описанные в настоящем документе. Фармацевтические композиции могут быть включены в контейнер, упаковку или дозатор вместе с инструкциями для введения. Описанные в настоящем документе наборы могут также включать в себя другие материалы, желательные с коммерческой и пользовательской точек зрения, включая другие буферы, разбавители, фильтры, иглы, шприцы и вкладыши в упаковку с инструкциями для выполнения любых способов, описанных в настоящем документе.
[0333] Композиция может также содержать более чем одну полиспецифическую полипептидную конструкцию, если это необходимо для конкретного показания, подлежащего лечению, например, с комплементарными активностями, которые не оказывают неблагоприятного влияния друг на друга. В некоторых вариантах осуществления или, кроме того, композиция может содержать агент, который усиливает ее функцию, такой как, например, цитотоксический агент, цитокин, химиотерапевтический агент или агент, ингибирующий рост. Такие молекулы подходящим образом присутствуют в комбинации в количествах, которые эффективны для предполагаемой цели.
[0334] В некоторых вариантах осуществления дозировка фармацевтической композиции представляет собой однократную дозу или многократную дозу. В некоторых вариантах осуществления дозы вводятся пациенту один раз в день, два раза в день, три раза в день или четыре или более раз в день. В некоторых вариантах осуществления в неделю вводятся примерно 1 или более (например, примерно 2 или более, примерно 3 или более, примерно 4 или более, примерно 5 или более, примерно 6 или более или примерно 7 или более) доз. В некоторых вариантах осуществления несколько доз вводят в течение дней, недель, месяцев или лет. В некоторых вариантах осуществления курс лечения составляет примерно 1 или более доз (например, примерно 2 или более доз, примерно 3 или более доз, примерно 4 или более доз, примерно 5 или более доз, примерно 7 или более доз, примерно 10 или более больше доз, примерно 15 или более доз, примерно 25 или более доз, примерно 40 или более доз, примерно 50 или более доз или примерно 100 или более доз).
[0335] В некоторых вариантах осуществления фармацевтическую композицию вводят пациенту. Обычно дозировки и пути введения фармацевтической композиции определяют в соответствии с размером и состоянием пациента в соответствии со стандартной фармацевтической практикой. Например, терапевтически эффективная доза может первоначально оцениваться либо в анализах на клеточных культурах, либо в моделях на животных, таких как мыши, крысы, кролики, собаки, свиньи или обезьяны. Животная модель может также использоваться для определения соответствующего диапазона концентраций и пути введения. Такая информация затем может быть использована для определения полезных доз и путей введения человеку. Точная дозировка будет определяться в свете факторов, связанных с пациентом, нуждающимся в лечении. Дозировка и введение подбираются для обеспечения достаточных уровней активного соединения или для поддержания желаемого эффекта. Факторы, которые могут быть приняты во внимание, включают серьезность болезненного состояния, общее состояние здоровья пациента, возраст, вес и пол пациента, время и частоту введения, комбинацию (комбинации) лекарственных средств, чувствительность к реакции и реакцию к терапии. Специалист в области медицины может легко определить оптимальную дозировку и схему лечения для конкретного пациента путем мониторинга пациента на наличие признаков заболевания и соответствующей корректировки лечения.
IV. Способы применения и терапевтическое введение
[0336] Также представлены способы использования и применения полиспецифических полипептидных конструкций. Такие способы и применения включают терапевтические способы и применения, например, включающие введение молекул или композиций, содержащих их, пациенту, имеющему заболевание, состояние или расстройство, такое как опухоль или злокачественное новообразование. В некоторых вариантах осуществления молекулу и/или композицию вводят в эффективном количестве для эффективного лечения заболевания или расстройства. Использование включает применение полиспецифических полипептидных конструкций в таких способах и способах лечения, а также при получении лекарственного средства для осуществления таких терапевтических способов. В некоторых вариантах осуществления способы осуществляются путем введения полиспецифических полипептидных конструкций или композиций, содержащих их, пациенту, имеющему или предположительно имеющему заболевание или состояние. В некоторых вариантах осуществления с помощью способов, таким образом, лечат заболевание или состояние или расстройство у пациента.
[0337] В одном варианте осуществления полиспецифическая полипептидная конструкция по изобретению может быть использована в качестве терапевтических агентов. Такие агенты обычно используются для диагностики, прогнозирования, мониторинга, лечения, облегчения и/или предотвращения заболевания или патологии у пациента. Терапевтическая схема осуществляется путем идентификации пациента, например пациента-человека или другого млекопитающего, страдающего (или подверженного риску развития) расстройства, с использованием стандартных методов. Полиспецифическая полипептидная конструкция вводится пациенту. Полиспецифическая полипептидная конструкция вводится пациенту и, как правило, оказывает эффект благодаря ее связыванию с мишенью (мишенями).
[0338] В некоторых вариантах осуществления в настоящем описании представлен способ модуляции иммунного ответа у пациента путем введения терапевтически эффективного количества любого из представленных полиспецифических конъюгатов или фармацевтических композиций. В некоторых вариантах осуществления способ модуляции иммунного ответа увеличивает или усиливает иммунный ответ у пациента. Например, увеличение или усиление ответа может быть увеличением клеточного иммунитета. В некоторых примерах способ повышает активность T-клеток, например активность цитолитических T-клеток (CTL). В некоторых вариантах осуществления модулированный (например, повышенный) иммунный ответ направлен против опухоли или злокачественного новообразования.
[0339] Введение полиспецифической полипептидной конструкции может активировать врожденные иммунные клетки через рекрутирование FcγR через Fc-область полипептида полиспецифической конструкции. Введение полиспецифической полипептидной конструкции может быть агонистическим, стимулировать, активировать и/или усиливать эффекторные функции врожденных иммунных клеток, включая ADCC, высвобождение цитокинов, дегрануляцию и/или ADCP. Введение полиспецифической полипептидной конструкции может активировать Т-клетки после того, как линкер(ы), соединяющий первый и второй компонент, расщепляется с помощью протеазы, таким образом, позволяя анти-CD3-связывающей части связываться с CD3ε на Т-клетках. Введение полиспецифической полипептидной конструкции может быть агонистическим, стимулировать, активировать и/или усиливать CD3-опосредованную активацию Т-клеток, цитотоксичность, высвобождение цитокинов и/или пролиферацию.
[0340] В некоторых вариантах осуществления представленные способы предназначены для лечения заболевания или состояния у пациента путем введения терапевтически эффективного количества любого из предоставленных полиспецифических конъюгатов или фармацевтических композиций. В некоторых вариантах осуществления заболевание или состояние представляет собой опухоль или злокачественное новообразование. Как правило, облегчение или лечение заболевания или расстройства включает уменьшение одного или более симптомов или медицинских проблем, связанных с заболеванием или расстройством. Например, в случае злокачественного новообразования терапевтически эффективное количество лекарственного средства может достигать одного или комбинации следующего: уменьшать количество опухолевых клеток; уменьшить размер опухоли; ингибировать (то есть уменьшать до некоторой степени и/или останавливать) инфильтрацию опухолевых клеток в периферические органы; ингибировать метастазирование опухоли; ингибировать до некоторой степени рост опухоли; и/или ослабить до некоторой степени один или более симптомов, связанных со злокачественным новообразованием. В некоторых вариантах осуществления композицию настоящего раскрытия можно использовать для предотвращения возникновения или повторного возникновения заболевания или расстройства у пациента, например человека или другого млекопитающего, такого как примат, не являющийся человеком, домашнее животное (например, кошка, собака, лошадь), сельскохозяйственное животное, рабочее животное или животное в зоопарке. Термины субъект и пациент используются в настоящем описании взаимозаменяемо.
[0341] В некоторых вариантах осуществления фармацевтическая композиция может быть использована для ингибирования роста опухолевых клеток млекопитающих (таких как опухолевые клетки человека). Способ лечения злокачественного новообразования может включать введение эффективного количества любой из фармацевтических композиций, описанных в настоящем документе, пациенту, имеющему злокачественное новообразование. Эффективное количество фармацевтической композиции может быть введено для ингибирования, остановки или обращения вспять прогрессирования злокачественного новообразования. Опухолевые клетки человека можно подвергать обработке in vivo или ex vivo. При лечении пациента-человека ex vivo ткани или жидкости, содержащие опухолевые клетки, обрабатывают вне организма, а затем ткани или жидкости снова вводят обратно пациенту. В некоторых вариантах осуществления злокачественное новообразование подвергается лечению у пациента-человека in vivo путем введения терапевтической композиции пациенту.
[0342] Неограничивающие примеры заболевания включают: все виды злокачественных новообразований (рак молочной железы, легких, колоректальный, предстательной железы, меланомы, рак головы и шеи, поджелудочной железы и т. д.), ревматоидный артрит, болезнь Крона, SLE, сердечно-сосудистые повреждения, ишемия и т. д., например, показания могут включать лейкозы, включая Т-клеточный острый лимфобластный лейкоз (T-ALL), лимфобластные заболевания, включая множественную миелому, и солидные опухоли, включая рак легкого, колоректальный рак, рак предстательной железы, поджелудочной железы и молочной железы, включая тройной негативный рак молочной железы. Например, показания включают заболевание костей или метастазирование при злокачественном новообразовании, независимо от первичного происхождения опухоли; рак молочной железы, включая в качестве неограничивающего примера, ER/PR+ рак молочной железы, Her2+ рак молочной железы, тройной негативный рак молочной железы; колоректальный рак; рак эндометрия; рак желудка; глиобластому; рак головы и шеи, как например, рак пищевода; рак легкого, как, например, немелкоклеточный рак легкого; множественную миелому, рак яичников; рак поджелудочной железы; рак предстательной железы; саркому, как например, остеосаркому; рак почки, как например, в качестве неограничивающего примера, почечно-клеточный рак; и/или рак кожи, как например, в качестве неограничивающего примера, плоскоклеточный рак, базально-клеточный рак или меланому. В некоторых вариантах осуществления злокачественное новообразование представляет собой плоскоклеточный рак. В некоторых вариантах осуществления злокачественное новообразование представляет собой плоскоклеточный рак кожи. В некоторых вариантах осуществления злокачественное новообразование представляет собой плоскоклеточный рак пищевода. В некоторых вариантах осуществления злокачественное новообразование представляет собой плоскоклеточный рак головы и шеи. В некоторых вариантах осуществления злокачественное новообразование представляет собой плоскоклеточную карциному легкого.
[0343] Терапевтически эффективное количество полиспецифической полипептидной конструкции по изобретению, как правило, относится к количеству, необходимому для достижения терапевтической цели. Как уже отмечалось выше, это может быть обязательным взаимодействие между полиспецифической полипептидной конструкции и ее антигеном-мишенью (мишенями), которые, в некоторых случаях, выступают агонистами, стимулируют, активируют и/или увеличивают FcγR-опосредованную активацию врожденной иммунной клетки или CD3- опосредованную активацию Т-клеток. Количество, требуемое для введения, будет, кроме того, зависеть от аффинности связывания полиспецифической полипептидной конструкции с ее специфическим антигеном (антигенами), а также будет зависеть от скорости, с которой вводимая полиспецифическая полипептидная конструкция истощается из свободного объема другого пациента, которому она вводится. Общие пределы для терапевтически эффективного дозирования полиспецифической полипептидной конструкции могут, в качестве неограничивающего примера, составлять примерно от 0,01 мкг/кг массы тела до примерно 10 мг/кг массы тела. В некоторых вариантах осуществления терапевтически эффективное дозирование полиспецифической полипептидной конструкции по изобретению может составлять в качестве неограничивающего примера примерно от 0,01 мг/кг массы тела примерно до 5-10 мг/кг массы тела. Общие частоты дозирования могут варьироваться, например, от двух раз в день до одного раза в неделю.
[0344] Эффективность лечения определяется в сочетании с любым известным методом диагностики или лечения конкретного расстройства. Способы скрининга полиспецифической полипептидной конструкции, которая обладает желаемой специфичностью, включают, но не ограничиваются этим, иммуноферментный анализ (ИФА) и другие иммунологически опосредованные методики, известные в данной области. Известно множество способов определения того, достаточно ли введение предложенных полиспецифических полипептидных конструкций модулирует иммунологическую активность путем устранения, секвестрации или инактивации иммунных клеток, опосредующих или способных опосредовать нежелательный иммунный ответ; путем индукции, генерирования или включения иммунных клеток, которые опосредуют или способны опосредовать защитный иммунный ответ; изменения физических или функциональных свойств иммунных клеток; или путем комбинации этих эффектов. Примеры измерений модуляции иммунологической активности включают, но не ограничиваются ими, исследование наличия или отсутствия популяций иммунных клеток (с использованием проточной цитометрии, иммуногистохимии, гистологии, электронной микроскопии, полимеразной цепной реакции (ПЦР)); измерение функциональной способности иммунных клеток, в том числе способности или устойчивости к пролиферации или делению в ответ на сигнал (например, с использованием анализов пролиферации Т-клеток и анализа pepscan на основе включения 3H-тимидина после стимуляции антителом против CD3, антителом против Т-клеточного рецептора, антителом против CD28, ионофорами кальция, антигенпрезентирующими клетками PMA (форбол-12-миристат-13-ацетат), с нагрузкой в виде пептидного или белкового антигена; анализы пролиферации B-клеток); измерение способности уничтожать или лизировать другие клетки (например, анализ цитотоксических Т-клеток); измерения цитокинов, хемокинов, молекул клеточной поверхности, антител и других продуктов клеток (например, с помощью проточной цитометрии, иммуноферментных анализов, вестерн-блот-анализа, анализа белковых микрочипов, анализа иммунопреципитации); измерение биохимических маркеров активации иммунных клеток или сигнальных путей в иммунных клетках (например, вестерн-блот и иммунопреципитационный анализ тирозина, серина или треонина, фосфорилирование, расщепление полипептидов и образование или диссоциация белковых комплексов; анализ массива белков; транскрипция ДНК, профилирование с использованием массивов ДНК или вычитающей гибридизации); измерения клеточной смерти с помощью апоптоза, некроза или других механизмов (например, окрашивание аннексином V, анализы TUNEL, гель-электрофорез для измерения электрофоретического расщепления ДНК, гистология; флуорогенные анализы каспаз, вестерн-блот-анализ субстратов каспаз); измерение генов, белков и других молекул, продуцируемых иммунными клетками (например, Нозерн-блот-анализ, полимеразная цепная реакция, микроматрицы ДНК, белковые микрочипы, 2-мерный гель-электрофорез, вестерн-блот-анализ, иммуноферментные анализы, связанные с ферментами, проточная цитометрия); и измерение клинических симптомов или результатов, таких как улучшение аутоиммунных, нейродегенеративных и других заболеваний, включающих собственные белки или собственные полипептиды (клинические показатели, требования к использованию дополнительной терапии, функциональное состояние, визуальные осмотры), например, путем измерения частоты рецидивов или тяжести заболевания.
[0345] Полиспецифическая полипептидная конструкция также полезна в различных диагностических и профилактических составах. В одном варианте осуществления полиспецифическая полипептидная конструкция вводится пациентам, которые подвергаются риску развития одного или более из вышеупомянутых расстройств. Предрасположенность пациента или органа к одному или более расстройствам может быть определена с использованием генотипических, серологических или биохимических маркеров.
[0346] В другом варианте осуществления изобретения полиспецифическая полипептидная конструкция вводится людям, у которых диагностированы клинические показания, связанные с одним или более из вышеупомянутых расстройств. После постановки диагноза вводят полиспецифическую полипептидную конструкцию, чтобы смягчить или обратить вспять эффекты клинического показания.
Комбинированные терапии
[0347] В некоторых вариантах осуществления полиспецифические полипептидные конструкции, конъюгированные полиспецифические полипептидные конструкции и их композиции, именуемые в настоящем документе как терапевтические средства, вводят в сочетании с одним или более дополнительными агентами или с комбинацией дополнительных агентов. Подходящие дополнительные агенты включают современные фармацевтические и/или хирургические методы лечения для предполагаемого применения. Например, терапевтическое средство можно использовать в сочетании с дополнительным химиотерапевтическим или противоопухолевым агентом. Например, терапевтическое средство и дополнительный агент включают в состав одной терапевтической композиции, и терапевтическое средство и дополнительный агент вводят одновременно. В некоторых вариантах осуществления терапевтическое средство и дополнительный агент представлены отдельно друг от друга, например, каждый включен в отдельную терапевтическую композицию, и терапевтическое средство и дополнительный агент вводят одновременно, или терапевтическое средство и дополнительный агент вводят в разное время в течение схемы лечения. Например, терапевтическое средство вводят до введения дополнительного агента, терапевтическое средство вводят после введения дополнительного агента, или терапевтическое средство и дополнительный агент вводят попеременно. Как описано в настоящем документе, терапевтическое средство и дополнительный агент вводят в однократных или многократных дозах. В некоторых вариантах осуществления дополнительный агент связан или иным образом присоединен к терапевтическому средству. Подходящие дополнительные агенты выбирают в соответствии с целью предполагаемого применения (т.е. уничтожение, предотвращение пролиферации клеток, гормонотерапия или генная терапия). Такие агенты могут включать, но не ограничиваются ими, например, фармацевтические агенты, токсины, фрагменты токсинов, алкилирующие агенты, ферменты, антибиотики, антиметаболиты, антипролиферативные агенты, гормоны, нейротрансмиттеры, ДНК, РНК, киРНК, олигонуклеотиды, антисмысловые РНК, аптамеры, диагностические средства, рентгеноконтрастные красители, радиоактивные изотопы, флуорогенные соединения, магнитные метки, наночастицы, маркерные соединения, лектины, соединения, которые изменяют проницаемость клеточной мембраны, фотохимические соединения, низкомолекулярные соединения, липосомы, мицеллы, векторы генной терапии, вирусные векторы и тому подобное. Наконец, могут быть использованы комбинации агентов или комбинации различных классов агентов.
[0348] В одном варианте осуществления полиспецифические полипептидные конструкции вводят в комбинированной терапии, то есть в комбинации с другими агентами, например терапевтическими агентами, которые полезны для лечения патологических состояний или расстройств, таких как аутоиммунные расстройства и воспалительные заболевания. Термин «в комбинации» в данном контексте означает, что агенты вводятся по существу одновременно, либо одновременно, либо последовательно. При введении последовательно, в начале введения второго соединения, первое из двух соединений все еще можно обнаружить в эффективных концентрациях в месте лечения.
[0349] Например, комбинированная терапия может включать одну или более полиспецифических полипептидных конструкций по настоящему изобретению, включенных в один состав и/или совместно вводимых с одним или более дополнительными терапевтическими агентами, например, одним или более ингибиторами цитокинов и факторов роста, иммунодепрессантами, противовоспалительными агентами, ингибиторами метаболизма, ингибиторами ферментов и/или цитотоксическими или цитостатическими агентами, как более подробно описано ниже. Кроме того, одну или более полиспецифических полипептидных конструкций, описанных в настоящем документе, можно использовать в комбинации с двумя или более терапевтическими агентами, описанными в настоящем документе. Такая комбинированная терапия может преимущественно использовать более низкие дозы вводимых терапевтических агентов, таким образом избегая возможных токсических эффектов или осложнений, связанных с различными монотерапиями.
[0350] В других вариантах осуществления одну или более полиспецифических полипептидных конструкций по настоящему изобретению можно включать в один состав и/или совместно вводить с одним или более противовоспалительными лекарственными средствами, иммунодепрессантами или метаболическими или ферментативными ингибиторами. Неограничивающие примеры лекарственных средств или ингибиторов, которые можно использовать в комбинации с антителами, описанными в настоящем документе, включают, но не ограничиваются ими, один или более из: нестероидных противовоспалительных лекарств (NSAID), например ибупрофен, тенидап, напроксен, мелоксикам, пироксикам, диклофенак и индометацин; сульфасалазин; кортикостероиды, такие как преднизолон; подавляющий цитокины противовоспалительный препарат(ы) (CSAID); ингибиторы биосинтеза нуклеотидов, например ингибиторы биосинтеза пурина, антагонисты фолата (например, метотрексат (N-[4-[[(2,4-диамино-6-птеридинил)метил]метиламино]бензоил]-L-глутаминовая кислота); и ингибиторы биосинтеза пиримидина, например ингибиторы дигидрооротатдегидрогеназы (DHODH). Подходящие терапевтические агенты для применения в комбинации с антителами по изобретению включают NSAID, CSAID, ингибиторы (DHODH) (например, лефлуномид) и антагонисты фолата (например, метотрексат).
[0351] Примеры дополнительных ингибиторов включают один или более из следующих: кортикостероиды (пероральное, ингаляционное и местное введение); иммунодепрессанты, например циклоспорин, такролимус (FK-506); и ингибиторы mTOR, например, сиролимус (рапамицин-RAPAMUNETM или производные рапамицина, например растворимые производные рапамицина (например, сложноэфирные производные рапамицина, например, CCI-779); агенты, которые препятствуют передаче сигналов провоспалительными цитокинами, такими как TNFα или IL-1 (например, ингибиторы IRAK, NIK, IKK, p38 или MAP киназы); ингибиторы COX2, например целекоксиб, рофекоксиб и их варианты; ингибиторы фосфодиэстеразы, например R973401 (ингибитор фосфодиэстеразы типа IV); ингибиторы фосфолипазы, например ингибиторы цитозольной фосфолазы 2 (cPLA2) (например, аналоги трифторметилкетона); ингибиторы фактора роста эндотелиальных клеток сосудов или рецептора фактора роста, например ингибитор VEGF и/или ингибитор VEGF-R, и ингибиторы ангиогенеза. Подходящими терапевтическими агентами для применения в комбинации с антителами по изобретению являются иммунодепрессанты, например, циклоспорин, такролимус (FK-506); ингибиторы mTOR, например производные сиролимуса (рапамицина) или рапамицина, например растворимые производные рапамицина (например, сложные эфиры производных рапамицина, например CCI-779); Ингибиторы СОХ2, например целекоксиб и его варианты; и ингибиторы фосфолипазы, например ингибиторы цитозольной фосфолипазы 2 (cPLA2), например аналоги трифторметилкетона. Дополнительные примеры терапевтических агентов, которые можно комбинировать с полиспецифической полипептидной конструкцией, включают один или более из следующих: 6-меркаптопурины (6-МР); азатиопринсульфасалазин; месалазина; олсалазин; хлорохин/гидроксихлорохин (PLAQUENIL®); пеницилламин; ауротиорналат (внутримышечно и перорально); азатиоприн; колхицин; агонисты бета-2 адренорецепторов (сальбутамол, тербуталин, сальметерал); ксантины (теофиллин, арнинофиллин); хромогликат; недокромил; кетотифен; ипратропий и окситропий; микофенолят мофетил; агонисты аденозина; антитромботические агенты; ингибиторы комплемента; и адренергические агенты.
V. Примерные варианты осуществления
[0352] Среди предложенных вариантов осуществления:
1. Полиспецифическая полипептидная конструкция, при этом полиспецифическая полипептидная конструкция содержит первый компонент, содержащий Fc-область иммуноглобулина, и второй компонент, содержащий CD3-связывающая область, где:
первый и второй компоненты связаны линкером, где Fc-область расположена на N-конце относительно CD3-связывающей области; и
один или оба из первого и второго компонентов содержат антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA).
2. Полиспецифическая полипептидная конструкция согласно варианту осуществления 1, отличающаяся тем, что CD3-связывающая область связывается с CD3 (CD3ε).
3. Полиспецифическая конструкция согласно варианту осуществления 1 или варианту осуществления 2, отличающаяся тем, что антигенсвязывающий домен расположен на аминоконце относительно Fc-области и/или на карбоксиконце относительно CD3-связывающей области полиспецифической полипептидной конструкции.
4. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-3, отличающаяся тем, что первый компонент содержит первый антигенсвязывающий домен, а второй компонент содержит второй антигенсвязывающий домен, где каждый из антигенсвязывающих доменов связывается с опухолеспецифическим антигеном (ТАА).
5. Полиспецифическая полипептидная конструкция по варианту осуществления 4, отличающаяся тем, что первый антигенсвязывающий домен расположен на аминоконце относительно Fc-области полиспецифической конструкции, а второй антигенсвязывающий домен расположен на карбоксиконце относительно CD3-связывающей области полиспецифической конструкции.
6. Полиспецифическая полипептидная конструкция, где полиспецифическая конструкция содержит по порядку от N-конца к C-концу:
первый антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA);
Fc-область иммуноглобулина;
линкер;
CD3-связывающую область, которая связывается с CD3 (CD3ε) и
второй антигенсвязывающий домен, который связывается с опухолесипецифическим антигеном (TAA).
7. Полиспецифическая полипептидная конструкция, где полиспецифическая конструкция содержит по порядку от N-конца к C-концу:
Fc-область иммуноглобулина;
линкер;
CD3-связывающую область, которая связывается с CD3 (CD3ε) и
антигенсвязывающий домен, который связывается с опухолесипецифическим антигеном (TAA).
8. Полиспецифическая полипептидная конструкция, где полиспецифическая конструкция содержит по порядку от N-конца к C-концу:
антигенсвязывающий домен, который связывается с опухолесипецифическим антигеном (TAA);
Fc-область иммуноглобулина;
линкер; и
CD3-связывающую область, которая связывается с CD3 (CD3ε).
9. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-8, отличающаяся тем, что Fc-область представляет собой гомодимерную Fc-область.
10. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-9, отличающаяся тем, что Fc-область представляет собой Fc-область человеческого IgG1, человеческого IgG2, человеческого IgG3 или человеческого IgG4 или представляет собой их иммунологически активный фрагмент.
11. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-10, отличающаяся тем, что Fc-область содержит полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 1, или последовательность аминокислот, которая имеет по меньшей мере 90%, 91%, 92% 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичности последовательности с SEQ ID NO: 1.
12. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-10, отличающаяся тем, что Fc-область содержит полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 2, или последовательность аминокислот, которая имеет по меньшей мере 90%, 91%, 92% 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичности последовательности с SEQ ID NO: 2;
Fc-область включает полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 4, или последовательность аминокислот, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичности последовательности с SEQ ID NO: 4; или
Fc-область содержит полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 5, или последовательность аминокислот, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичности последовательности с SEQ ID NO: 5.
13. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-6, 9 и 12, отличающаяся тем, что Fc-область представляет собой гетеродимерную Fc-область.
14. Полиспецифическая полипептидная конструкция по варианту осуществления 13, отличающаяся тем, что один или оба полипептида Fc гетеродимерной Fc-области содержат по меньшей мере одну модификацию для индукции гетеродимеризации по сравнению с полипептидом гомодимерной Fc-области, необязательно по сравнению с полипептидом Fc, представленным в SEQ ID NO: 1 или его иммунологически активным фрагментом.
15. Полиспецифическая полипептидная конструкция по варианту осуществления 14, отличающаяся тем, что каждый из полипептидов Fc гетеродимерной Fc-области независимо содержит по меньшей мере одну аминокислотную модификацию.
16. Полиспецифическая полипептидная конструкция по варианту осуществления 15, отличающаяся тем, что каждый из полипептидов Fc гетеродимерной Fc-области содержит модификацию «выступ-во-впадину» или содержит мутацию заряда для увеличения электростатической комплементарности полипептидов.
17. Полиспецифическая полипептидная конструкция по варианту осуществления 16, отличающаяся тем, что аминокислотная модификация представляет собой модификацию «выступ-во-впадину».
18. Полиспецифический слитый полипептид по любому из вариантов осуществления 13-17, отличающаяся тем, что первый полипептид Fc гетеродимерной Fc-области содержит модификацию, выбранную из Thr366Ser, Leu368Ala, Tyr407Val и их комбинаций, и второй полипептид Fc гетеродимерной Fc-области содержит модификацию T366W.
19. Полиспецифический слитый полипептид по варианту осуществления 18, отличающийся тем, что первый и второй полипептиды Fc дополнительно содержат модификацию нецистеинового остатка до остатка цистеина, где модификация первого полипептида находится в одном из положений Ser354 и Y349, и модификация второго полипептида Fc находится в другом положении Ser354 и Y349.
20. Полиспецифическая полипептидная конструкция по варианту осуществления 16, отличающаяся тем, что аминокислотная модификация представляет собой мутацию заряда для увеличения электростатической комплементарности полипептидов.
21. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 13-16 и 20, отличающаяся тем, что первый и/или второй полипептиды Fc или каждый из первого и второго полипептидов Fc содержат модификацию в комплементарных положениях, где модификация представляет собой замену аминокислотой, имеющей противоположный заряд к комплементарной аминокислоте другого полипептида.
22. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 14-21, отличающаяся тем, что один из первого или второго полипептида Fc гетеродимерной Fc-области дополнительно содержит модификацию по остатку Ile253.
23. Полиспецифическая полипептидная конструкция по варианту осуществления 22, отличающаяся тем, что модификация представляет собой Ile253Arg.
24. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 14-23, отличающаяся тем, что один из первого или второго полипептида Fc гетеродимерной Fc-области дополнительно содержит модификацию по остатку His435.
25. Полиспецифическая полипептидная конструкция по варианту осуществления 24, отличающаяся тем, что модификация представляет собой His435Arg.
26. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-25, отличающаяся тем, что Fc-область содержит полипептид, в котором отсутствует Lys447.
27. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-26, отличающаяся тем, что Fc-область содержит полипептид, содержащий по меньшей мере одну модификацию для усиления связывания FcRn.
28. Полиспецифический слитый полипептид по варианту осуществления 27, отличающаяся тем, что модификация находится в положении, выбранном из группы, состоящей из Met252, Ser254, Thr256, Met428, Asn434 и их комбинаций.
29. Полиспецифический слитый полипептид по варианту осуществления 28, отличающаяся тем, что модификация находится в положении, выбранном из группы, состоящей из Met252Y, Ser254T, Thr256E, Met428L, Met428V, Asn434S и их комбинаций.
30. Полиспецифический слитый полипептид по варианту осуществления 28, отличающийся тем, что модификация находится в положении Met252 и в положении Met428.
31. Полиспецифический слитый полипептид по варианту осуществления 30, отличающийся тем, что модификация представляет собой Met252Y и Met428L.
32. Полиспецифический слитый полипептид по варианту осуществления 30, отличающийся тем, что модификация представляет собой Met252Y и Met428V.
33. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 13-32, отличающаяся тем, что первый полипептид гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 82, 86, 94 или 96, и второй полипептид гетеродимерной Fc-области включает последовательность аминокислот, представленную в любой из SEQ ID NO: 83, 87, 90, 92, 98 или 100.
34. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-33, отличающаяся тем, что Fc-область содержит полипептид, содержащий по меньшей мере одну аминокислотную модификацию, которая снижает эффекторную функцию и/или уменьшает связывание с эффекторной молекулой, выбранной из Fc-гамма-рецептора или C1q.
35. Полиспецифическая полипептидная конструкция по варианту осуществления 34, отличающаяся тем, что одна или более аминокислотных модификаций представляют собой делецию одного или более из Glu233, Leu234 или Leu235.
36. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 13-32, 34 и 35, отличающаяся тем, что первый полипептид гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 84, 88, 95 или 97, и второй полипептид гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 85, 89, 91, 93, 99 или 101.
37. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-32, отличающаяся тем, что Fc-область содержит полипептид, содержащий по меньшей мере одну модификацию для усиления связывания FcγR.
38. Полиспецифическая полипептидная конструкция по варианту осуществления 37, отличающаяся тем, что модификация представляет собой модификацию по Ser239 или Ile332.
39. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-32 и 37, отличающаяся тем, что гликозилирование Fc-области модифицируют для усиления связывания FcγR по сравнению с немодифицированной Fc-областью.
40. Полиспецифическая полипептидная конструкция по варианту осуществления 39, отличающаяся тем, что Fc-область не содержит или имеет пониженное содержание фукозы.
41. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-40, отличающаяся тем, что CD3-связывающая область представляет собой антитело против CD3 или антигенсвязывающий фрагмент.
42. Полиспецифическая полипептидная конструкция по варианту осуществления 41, отличающаяся тем, что антитело против CD3 или антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL).
43. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-42, отличающаяся тем, что CD3-связывающая область является одновалентной.
44. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 41-43, отличающаяся тем, что антитело против CD3 или антигенсвязывающий фрагмент не является одноцепочечным антителом, необязательно не является одноцепочечным вариабельным фрагментом (scFv).
45. Полиспецифическая полипептидная конструкция по варианту осуществления 42 или по варианту осуществления 44, отличающаяся тем, что Fc представляет собой гетеродимерную Fc-область, а VH и VL, которые содержат антитело против CD3 или антигенсвязывающий фрагмент, связаны с противоположными полипептидами гетеродимерной Fc-области.
46. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-45, отличающаяся тем, что CD3-связывающая область не способна или по существу не способна связывать или рекрутировать CD3, если по меньшей мере один из антигенсвязывающих доменов не связан с его TAA.
47. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-46, отличающаяся тем, что CD3-связывающая область не способна или по существу не способна связывать или рекрутровать CD3, если по меньшей мере два антигенсвязывающих домена не связаны с их ТАА.
48. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-47, отличающаяся тем, что линкер представляет собой полипептидный линкер.
49. Полиспецифическая полипептидная конструкция согласно варианту осуществления 48, отличающаяся тем, что линкер представляет собой полипептид длиной до 25 аминокислот.
50. Полиспецифическая полипептидная конструкция по варианту осуществления 48 или варианту осуществления 49, отличающаяся тем, что линкер представляет собой полипептид длиной, составляющей или примерно составляющей от 2 до 24 аминокислот, от 2 до 20 аминокислот, от 2 до 18 аминокислот, от 2 до 14 аминокислот, от 2 до 12 аминокислот кислоты, от 2 до 10 аминокислот, от 2 до 8 аминокислот, от 2 до 6 аминокислот, от 6 до 24 аминокислот, от 6 до 20 аминокислот, от 6 до 18 аминокислот, от 6 до 14 аминокислот, от 6 до 12 аминокислот, От 6 до 10 аминокислот, от 6 до 8 аминокислот, от 8 до 24 аминокислот, от 8 до 20 аминокислот, от 8 до 18 аминокислот, от 8 до 14 аминокислот, от 8 до 12 аминокислот, от 8 до 10 аминокислот, от 10 до 24 аминокислоты, от 10 до 20 аминокислот, от 10 до 18 аминокислот, от 10 до 14 аминокислот, от 10 до 12 аминокислот, от 12 до 24 аминокислот, от 12 до 20 аминокислот, от 12 до 18 аминокислот, от 12 до 14 аминокислот, от 14 до 24 аминокислот, от 14 до 20 аминокислот, от 14 до 18 аминокислот, от 18 до 24 аминокислот, от 18 до 20 аминокислот или от 20 до 24 аминокислот.
51. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 48-50, отличающаяся тем, что линкер представляет собой полипептид, длина которого составляет 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот.
52. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-51, отличающаяся тем, что линкер представляет собой расщепляемый линкер.
53. Полиспецифическая полипептидная конструкция, причем полиспецифическая полипептидная конструкция, которая содержит первый компонент, содержащий гетеродимерную Fc-область, и второй компонент, содержащий антитело против CD3 или антигенсвязывающий фрагмент, содержащий вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL)), где:
VH и VL, которые содержит антитело против CD3 или антигенсвязывающий фрагмент, связаны с противоположными полипептидами гетеродимерной Fc-области;
первый и второй компоненты связаны расщепляемым линкером, где гетеродимерная Fc-область расположена на N-конце относительно антитела против CD3; и
один или оба из первого и второго компонентов содержат антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA).
54. Полиспецифическая полипептидная конструкция по варианту осуществления 52 или по варианту осуществления 53, отличающаяся тем, что связывание CD3-связывающей области с CD3 существенно снижается, когда полиспецифическая полипептидная конструкция находится в нерасщепленном состоянии по сравнению с расщепленным состоянием.
55. Полиспецифический полипептид по любому из вариантов осуществления 52-54, отличающийся тем, что в расщепленном состоянии первый и второй компоненты не связаны.
56. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 52-55, отличающаяся тем, что расщепляемый линкер представляет собой полипептид, который функционирует в качестве субстрата для протеазы.
57. Полиспецифическая полипептидная конструкция по варианту осуществления 56, отличающаяся тем, что протеаза продуцируется иммунной эффекторной клеткой, опухолью или клетками, присутствующими в микроокружении опухоли.
58. Полиспецифическая полипептидная конструкция по варианту осуществления 57, отличающаяся тем, что протеаза продуцируется иммунной эффекторной клеткой, а иммунная эффекторная клетка представляет собой активированную Т-клетку, клетку-натуральный киллер (NK) или Т-клетку NK.
59. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 56-58, отличающаяся тем, что протеаза выбрана из матриптазы, матриксной металлопротеазы (ММР), гранзима В и их комбинаций.
60. Полиспецифическая полипептидная конструкция по варианту осуществления 59, отличающаяся тем, что протеазой является гранзим B.
61. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 52-60, отличающаяся тем, что расщепляемый линкер содержит аминокислотную последовательность общей формулы P4 P3 P2 P1 ↓ P1' (SEQ ID NO: 150), где P4 представляет собой аминокислоту I, L, Y, M, F, V или A; P3 представляет собой аминокислоту A, G, S, V, E, D, Q, N или Y; P2 представляет собой аминокислоту H, P, A, V, G, S или T; P1 представляет собой аминокислоту D или E; и P1' представляет собой аминокислоту I, L, Y, M, F, V, T, S, G или A.
62. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 52-61, отличающаяся тем, что расщепляемый линкер содержит аминокислотную последовательность общей формулы P4 P3 P2 P1 ↓ P1' (SEQ ID NO: 151), где P4 представляет собой аминокислоту I или L; P3 представляет собой аминокислоту E; P2 представляет собой аминокислоту P или A; P1 представляет собой аминокислоту D; и P1' представляет собой аминокислоту I, V, T, S или G.
63. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 52-62, отличающаяся тем, что расщепляемый линкер содержит аминокислотную последовательность IEPDI (SEQ ID NO: 136), LEPDG (SEQ ID NO: 152, LEADT (SEQ ID NO: 137), IEPDG (SEQ ID NO: 138), IEPDV (SEQ ID NO: 139), IEPDS (SEQ ID NO: 140), IEPDT (SEQ ID NO: 141) или LEADG (SEQ ID NO: 153).
64. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 52-63, отличающаяся тем, что расщепляемый линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 22, 105-112, 136-141, 148, 150-153.
65. Полиспецифическая полипептидная конструкция по варианту осуществления 59, отличающаяся тем, что протеаза представляет собой матриптазу.
66. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 52-65, отличающаяся тем, что:
расщепляемый линкер содержит последовательность P1QAR↓ (A/V) (SEQ ID NO: 154), где P1 представляет собой любую аминокислоту; или
расщепляемый линкер содержит последовательность RQAR (A/V) (SEQ ID NO: 155).
67. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 52-66, отличающаяся тем, что расщепляемый линкер содержит последовательность RQARV (SEQ ID NO: 156).
68. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 52-67, отличающаяся тем, что расщепляемый линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 23, 154-156.
69. Полиспецифическая полипептидная конструкция по варианту осуществления 59, отличающаяся тем, что протеаза представляет собой ММР.
70. Полиспецифическая полипептидная конструкция по варианту осуществления 69, отличающаяся тем, что ММР представляет собой ММР-2.
71. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 52-70, отличающаяся тем, что расщепляемый линкер имеет общую формулу P3 P2 P1 ↓ P1' (SEQ ID NO: 157), где P3 представляет собой P, V или A; P2 представляет собой Q или D; P1 представляет собой A или N; и P1' представляет собой L, I или M.
72. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 52-71, отличающаяся тем, что расщепляемый линкер содержит общую формулу P3 P2 P1 ↓ P1' (SEQ ID NO: 158), где P3 представляет собой P; P2 представляет собой Q или D; P1 представляет собой A или N; и P1' представляет собой L или I.
73. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 52-72, отличающаяся тем, что расщепляемый линкер содержит последовательность PAGL (SEQ ID NO: 24).
74. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 52-73, отличающаяся тем, что расщепляемый линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 22-31, 104-114, 117-118, 136-144, 148, 150 -158.
75. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 45-74, отличающаяся тем, что полиспецифическая полипептидная конструкция содержит, по меньшей мере, (i) первый полипептид, содержащий первый полипептид Fc гетеродимерной Fc-области, линкер и домен VH антитела против CD3 или антигенсвязывающий фрагмент; и (ii) второй полипептид, содержащий второй полипептид Fc гетеродимерной Fc-области, линкер и домен VL антитела против CD3 или антигенсвязывающего фрагмента, где один или оба из первого и второго полипептида содержат, по меньшей мере, один антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA).
76. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-75, отличающаяся тем, что один или более антигенсвязывающих доменов, которые связываются с ТАА, приводит к одновалентному, двухвалентному, трехвалентному или четырехвалентному связыванию с ТАА.
77. Полиспецифическая полипептидная конструкция по варианту осуществления 75, отличающаяся тем, что только один из первого или второго полипептида содержит по меньшей мере один антигенсвязывающий домен, который связывается с ТАА.
78. Полиспецифическая полипептидная конструкция по варианту осуществления 75 или по варианту осуществления 77, отличающаяся тем, что по меньшей мере один антигенсвязывающий домен расположен на аминоконце относительно Fc-области и/или расположен на карбоксиконце относительно CD3-связывающей области одного из первого или второго полипептида полиспецифической полипептидной конструкции.
79. Полиспецифическая полипептидная конструкция по варианту осуществления 75 или по варианту осуществления 77, отличающаяся тем, что по меньшей мере один антигенсвязывающий домен расположен на аминоконце относительно Fc-области полиспецифической конструкции, а второй антигенсвязывающий домен расположен на карбоксиконце относительно CD3-связывающей области полиспецифической конструкции.
80. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-79, отличающаяся тем, что антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов содержит внеклеточный домен или его связывающий фрагмент нативного родственного партнера по связыванию TAA или его вариант, который проявляет активность связывания с ТАА.
81. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-79, отличающаяся тем, что антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов представляет собой антитело или его антигенсвязывающий фрагмент, выбранный из группы, состоящей из Fab-фрагмента, F (ab)'2-фрагмента, Fv-фрагмента, scFv, scAb, dAb, однодоменного антитела с тяжелой цепью и однодоменного антитела с легкой цепью.
82. Полиспецифическая полипептидная конструкция по варианту осуществления 81, отличающаяся тем, что антитело или его антигенсвязывающий фрагмент представляет собой Fv, scFv, Fab, однодоменное антитело (sdAb), VNAR или VHH.
83. Полиспецифическая полипептидная конструкция по варианту осуществления 81 или по варианту осуществления 82, отличающаяся тем, что антитело или его антигенсвязывающий фрагмент представляет собой sdAb.
84. Полиспецифическая полипептидная конструкция по варианту осуществления 83, отличающаяся тем, что sdAb представляет собой человеческое или гуманизированное sdAb.
85. Полиспецифическая полипептидная конструкция по варианту осуществления 83 или по варианту осуществления 84, отличающаяся тем, что sdAb представляет собой VHH, VNAR, сконструированный домен VH или сконструированный домен VK.
86. Полиспецифическая полипептидная конструкция по варианту осуществления 81 или по варианту осуществления 82, отличающаяся тем, что антитело или его антигенсвязывающий фрагмент представляет собой scFv.
87. Полиспецифическая полипептидная конструкция по варианту осуществления 81 или по варианту осуществления 82, отличающаяся тем, что антитело или его антигенсвязывающий фрагмент представляет собой Fab.
88. Полиспецифическая полипептидная конструкция по варианту осуществления 87, отличающаяся тем, что полиспецифическая полипептидная конструкция содержит:
(i) первый полипептид, содержащий первый полипептид Fc гетеродимерной Fc-области, линкер и домен VH антитела против CD3 или антигенсвязывающего фрагмента;
(ii) второй полипептид, содержащий второй полипептид Fc гетеродимерной Fc-области, линкер и домен VL антитела против CD3 или антигенсвязывающего фрагмента, и (iii) третий полипептид, содержащий VH-CH1 (Fd) или VL-CL Fab-фрагмента антитела, который связывается с опухолеспецифическим антигеном, где первый и/или второй полипептид дополнительно содержит другой из VH-CH1 (Fd) или VL-CL Fab-фрагмента антитела.
89. Полиспецифическая полипептидная конструкция по варианту осуществления 88, отличающаяся тем, что только один из первого или второго полипептида содержит другой из VH-CH1 (Fd) или VL-CL Fab-фрагмента антитела.
90. Полиспецифическая полипептидная конструкция по варианту осуществления 89, отличающаяся тем, что как первый, так и второй полипептид содержат другой из VH-CH1 (Fd) или VL-CL Fab-фрагмента антитела.
91. полиспецифическая полипептидная конструкция по варианту осуществления 89 или варианту осуществления 90, отличающаяся тем, что другой из VH-CH1 (Fd) или VL-CL Fab-фрагмента антитела расположен на аминоконце относительно Fc-области и/или на карбоксиконце относительно к CD3-связывающей области одного из первого или второго полипептида полиспецифической полипептидной конструкции.
92. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 89-91, отличающаяся тем, что другой из VH-CH1 (Fd) или VL-CL Fab-фрагмента антитела расположен на аминоконце относительно Fc-области первого полипептида или второго полипептида и на карбоксиконце относительно CD3-связывающей области другого из первого или второго полипептида.
93. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-92, отличающаяся тем, что антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов связывается с опухолевым антигеном, выбранным из 1-92-LFA-3, 5T4, альфа-4 интегрина, альфа-V интегрина, альфа4бета1 интегрина, альфа4бета7 интегрина, AGR2, анти-Lewis-Y, рецептора апелина J, APRIL, B7-H3, B7-H4, BAFF, BTLA, компонента комплемента C5, C-242, CA9, CA19-9, (Lewis а), карбоангидразы 9, CD2, CD3, CD6, CD9, CD11a, CD19, CD20, CD22, CD24, CD25, CD27, CD28, CD30, CD33, CD38, CD40, CD40L, CD41, CD44, CD44v6, CD47, CD51, CD52, CD56, CD64, CD70, CD71, CD74, CD80, CD81, CD86, CD95, CD117, CD123, CD125, CD132, (IL-2RG), CD133, CD137, CD138, CD166, CD172A, CD248, CDH6, CEACAM5 (CEA), CEACAM6 (NCA-90), CLAUDIN-3, CLAUDIN-4, cMet, коллагена, Cripto, CSFR, CSFR-1, CTLA-4, CTGF, CXCL10, CXCL13, CXCR1, CXCR2, CXCR4, CYR61, DL44, DLK1, DLL3, DLL4, DPP-4, DSG1, EDA, EDB, EGFR, EGFRviii, рецептора эндотелина B (ETBR), ENPP3, EpCAM, EPHA2, EPHB2, ERBB3, F-протеина RSV, FAP, FGF-2, FGF8, FGFR1, FGFR2, FGFR3, FGFR4, FLT-3, рецептора альфа фолата (FR α), GAL3ST1, G-CSF, G-CSFR, GD2, GITR, GLUT1, GLUT4, GM-CSF, GM-CSFR, рецепторов GP IIb/IIIa, Gp130, GPIIB/IIIA, GPNMB, GRP78, HER2/neu, HER3, HER4, HGF, hGH, HVEM, гиалуронидазы, ICOS, IFNальфа, IFNбета, IFNгамма, IgE, рецептора IgE (FceRI), IGF, IGF1R, IL1B, IL1R, IL2, IL11, IL12, IL12p40, IL-12R, IL-12Rbeta1, IL13, IL13R, IL15, IL17, IL18, IL21, IL23, IL23R, IL27/IL27R (wsx1), IL29, IL-31R, IL31/IL31R, IL2R, IL4, IL4R, IL6, IL6R, рецептора инсулина, Jagged лигандов, Jagged 1, Jagged 2, KISS1-R, LAG-3, LIF-R, Lewis X, LIGHT, LRP4, LRRC26, Ly6G6D, LyPD1, MCSP, мезотелина, MRP4, MUC1, муцина-16 (MUC16, CA-125), Na/K-АТФазы, NGF, никастрина, рецепторов Notch, Notch 1, Notch 2, Notch 3, Notch 4, NOV, OSM-R, OX-40, PAR2, PDGF-AA, PDGF-BB, PDGFRальфа, PDGFRбета, PD-1, PD-L1, PD-L2, фосфатидилсерина, P1GF, PSCA, PSMA, PSGR, RAAG12, RAGE, SLC44A4, фосфат сфингозина 1, STEAP1, STEAP2, TAG-72, TAPA1, TE M-8, TGFбета, TIGIT, TIM-3, TLR2, TLR4, TLR6, TLR7, TLR8, TLR9, TMEM31, TNFальфа, TNFR, TNFRS12A, TRAIL-R1, TRAIL-R2, трансферрина, рецептора трансферрина, TRK-A, TRK-B, uPAR, VAP1, VCAM-1, VEGF, VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGFR1, VEGFR2, VEGFR3, VISTA, WISP-1, WISP-2 и WISP-3.
94. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-93, отличающаяся тем, что полиспецифический антигенсвязывающий домен содержит, по меньшей мере, первый антигенсвязывающий домен и второй антигенсвязывающий домен, где первый антигенсвязывающий домен и второй антигенсвязывающий домен связываются с одним и тем же TAA.
95. Полиспецифическая полипептидная конструкция по варианту осуществления 94, отличающаяся тем, что первый антигенсвязывающий домен и второй антигенсвязывающий домен связываются с другим эпитопом того же TAA.
96. Полиспецифическая полипептидная конструкция по варианту осуществления 94, отличающаяся тем, что первый антигенсвязывающий домен и второй антигенсвязывающий домен связываются с одним и тем же эпитопом одного и того же TAA.
97. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-96, отличающаяся тем, что полиспецифический антигенсвязывающий домен содержит, по меньшей мере, первый антигенсвязывающий домен и второй антигенсвязывающий домен, где первый антигенсвязывающий домен и второй антигенсвязывающий домен связываются с разными TAA.
98. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 5-97, отличающаяся тем, что полиспецифическая полипептидная конструкция содержит первый связывающий пептид (LP1) между первым антигенсвязывающим доменом и Fc-областью.
99. полиспецифическая полипептидная конструкция по любому из вариантов осуществления 5-98, отличающаяся тем, что полиспецифическая полипептидная конструкция содержит второй связывающий пептид (LP2) между CD3-связывающей областью и вторым антигенсвязывающим доменом.
100. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 5-99, отличающаяся тем, что полиспецифическая полипептидная конструкция содержит первый связывающий пептид (LP1) между первым антигенсвязывающим доменом и Fc-областью и второй связывающий пептид (LP2) между CD3-связывающей областью и второй антигенсвязывающий домен, и где полиспецифическая полипептидная конструкция имеет структурное расположение от N-конца к C-концу следующим образом: первый антигенсвязывающий домен-LP1-Fc-область-линкер-CD3-связывающая область-LP2-второй антигенсвязывающий домен.
101. Полиспецифическая полипептидная конструкция по варианту осуществления 100, отличающаяся тем, что линкер представляет собой расщепляемый линкер.
102. Полиспецифическая полипептидная конструкция по варианту осуществления 100 и варианту осуществления 101, отличающаяся тем, что два связывающих пептида не являются идентичными друг другу.
103. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 98-102, отличающаяся тем, что LP1 или LP2 независимо представляют собой пептид длиной примерно от 1 до 20 аминокислот.
104. Полиспецифический полипептид по варианту осуществления 103, отличающийся тем, что LP1 или LP2 независимо содержат пептид, который представляет собой или содержит любой линкер Gly-Ser, как представлено в SEQ ID NO: 10-13, 119, 135, 147, 149 или GGS.
105. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 41-104, отличающаяся тем, что антитело против CD3 или антигенсвязывающий фрагмент представляет собой Fv-фрагмент антитела.
106. Полиспецифическая полипептидная конструкция по варианту осуществления 105, отличающаяся тем, что Fv-фрагмент антитела содержит стабилизированный дисульфидом анти-CD3-связывающий Fv-фрагмент (dsFv).
107. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 41-106, отличающаяся тем, что антитело против CD3 или его антигенсвязывающий фрагмент содержит VH CDR1, содержащую аминокислотную последовательность TYAMN (SEQ ID NO: 16); VH CDR2, содержащую аминокислотную последовательность RIRSKYNNYATYYADSVKD (SEQ ID NO: 17); VH CDR3, содержащую аминокислотную последовательность HGNFGNSYVSWFAY (SEQ ID NO: 18), VL CDR1, содержащую аминокислотную последовательность RSSTGAVTTSNYAN (SEQ ID NO: 19); VL CDR2, содержащую аминокислотную последовательность GTNKRAP (SEQ ID NO: 20); и VL CDR3, содержащую аминокислотную последовательность ALWYSNLWV (SEQ ID NO: 21).
108. Полиспецифическая полипептидная конструкция по варианту осуществления 106 или воплощению 107, отличающаяся тем, что анти-CD3 dsFv содержит:
VH, имеющую аминокислотную последовательность любой из SEQ ID NO: 14 и 32-62, или последовательность, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с любой из SEQ ID NO: 14 и 32-62; и
VL, имеющую аминокислотную последовательность любой из SEQ ID NO: 15 и 63-81, или последовательность, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с любой из SEQ ID NO: 14 и 32-62.
109. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 106-108, отличающаяся тем, что анти-CD3 dsFv содержит аминокислотную последовательность SEQ ID NO: 14 и аминокислотную последовательность SEQ ID NO: 15.
110. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 102-104, отличающаяся тем, что анти-CD3 dsFv содержит аминокислотную последовательность SEQ ID NO: 44 и аминокислотную последовательность SEQ ID NO: 72.
111. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-109, отличающаяся тем, что полиспецифическая полипептидная конструкция конъюгирована с агентом.
112. Полиспецифическая полипептидная конструкция по варианту осуществления 111, отличающаяся тем, что агент представляет собой терапевтический агент, противоопухолевый агент, токсин или его фрагмент, детектируемый фрагмент или диагностический агент.
113. Полиспецифическая полипептидная конструкция по варианту осуществления 112, отличающаяся тем, что агент конъюгирован с полиспецифической полипептидной конструкцией через линкер.
114. Полинуклеотид(ы), кодирующий полиспецифические полипептидные конструкции по любому из вариантов осуществления 1-113.
115. Полинуклеотид, кодирующий полипептидную цепь любой из полиспецифических полипептидных конструкций по любому из вариантов осуществления 1-113.
116. Полинуклеотид, содержащий первую последовательность нуклеиновой кислоты, кодирующую первый полипептид полиспецифической конструкции по любому из вариантов осуществления 1-115, и вторую последовательность нуклеиновой кислоты, кодирующую второй полипептид полиспецифической конструкции, где первая и вторая последовательности нуклеиновой кислоты разделены внутренним сайтом посадки рибосомы (IRES) или нуклеиновой кислотой, кодирующей саморасщепляющийся пептид или пептид, который вызывает пропуск рибосомы.
117. Полинуклеотид по варианту осуществления 116, отличающийся тем, что первая последовательность нуклеиновой кислоты и вторая последовательность нуклеиновой кислоты функционально связаны с одним и тем же промотором.
118. Полинуклеотид по варианту осуществления 116 или по варианту осуществления 117, отличающийся тем, что полиспецифическая полипептидная конструкция содержит третью полипептидную цепь, а полинуклеотид дополнительно содержит третью нуклеиновую кислоту, кодирующую третий полипептид полиспецифической конструкции.
119. Полинуклеотид по варианту осуществления 118, отличающийся тем, что третья нуклеиновая кислота отделена от первого и/или второго полипептида внутренним сайтом посадки рибосомы (IRES), или нуклеиновой кислотой, кодирующей саморасщепляющийся пептид или пептид, который вызывает пропуск рибосомы, и/или третья последовательность нуклеиновой кислоты функционально связана с тем же промотором, что и первая и/или вторая последовательность нуклеиновой кислоты.
120. Полинуклеотид по любому из вариантов осуществления 116-119, отличающийся тем, что нуклеиновая кислота, кодирующая саморасщепляющийся пептид или пептид, вызывающий пропуск рибосомы, выбрана из T2A, P2A, E2A или F2A.
121. Вектор, содержащий полинуклеотид по любому из вариантов осуществления 114-120.
122. Вектор по варианту осуществления 121, который является экспрессирующим вектором.
123. Вектор по варианту осуществления 121 или 122, который представляет собой вирусный вектор или эукариотический вектор, необязательно, где эукариотический вектор представляет собой вектор млекопитающего.
124. Клетка, содержащая полинуклеотид или полинуклеотиды по любому из вариантов осуществления 114-120, или вектор или векторы по любому из вариантов осуществления 121-123.
125. Клетка по варианту осуществления 124, отличающаяся тем, что клетка является рекомбинантной или выделенной.
126. Клетка по варианту осуществления 125, отличающаяся тем, что клетка представляет собой клетку млекопитающего.
127. Клетка по варианту осуществления 126, отличающаяся тем, что клетка представляет собой клетку HEK293 или CHO.
128. Способ получения полиспецифической полипептидной конструкции, включающий введение в клетку полинуклеотида или полинуклеотидов по любому из вариантов осуществления 114-120 или вектора или векторов по любому из вариантов осуществления 121-123 и культивирование клетки в условиях с получением полиспецифической полипептидной конструкции.
129. Способ получения полиспецифической полипептидной конструкции, включающий культивирование клетки по любому из вариантов осуществления 124-127 в условиях, при которых полиспецифический полипептид продуцируется клеткой.
130. Клетка по варианту осуществления 128 или 129, отличающаяся тем, что клетка представляет собой клетку млекопитающего.
131. Клетка по варианту осуществления 130, отличающаяся тем, что клетка представляет собой клетку HEK293 или CHO.
132. Способ по варианту осуществления 128 или варианту осуществления 129, дополнительно включающий выделение или очистку полиспецифической полипептидной конструкции из клетки.
133. Способ по любому из вариантов осуществления 128-132, отличающийся тем, что полиспецифическая полипептидная конструкция представляет собой гетеродимер.
134. Полиспецифическая полипептидная конструкция, полученная способом по любому из вариантов осуществления 128-133.
135. Фармацевтическая композиция, содержащая полиспецифическую полипептидную конструкцию по любому из вариантов осуществления 1-113 или по варианту осуществления 134 и фармацевтически приемлемый носитель.
136. Фармацевтическая композиция по варианту осуществления 135, которая является стерильной.
137. Способ стимулирования или индукции иммунного ответа, включающий контактирование клетки-мишени и Т-клетки с полиспецифической полипептидной конструкцией по любому из вариантов осуществления 1-113 или по варианту осуществления 134 или фармацевтической композицией по варианту осуществления 109 или по варианту осуществления 110, причем указанная клетка-мишень, экспрессирующая опухолеспецифический антиген, распознаваемый полиспецифической полипептидной конструкцией.
138. Способ по варианту осуществления 137, отличающийся тем, что клетка-мишень представляет собой опухолевую клетку, экспрессирующую опухолеспецифический антиген (TAA).
139. Способ по варианту осуществления 138 или по варианту осуществления 138, отличающийся тем, что полиспецифическая полипептидная конструкция содержит расщепляемый линкер, который функционирует в качестве субстрата для протеазы, и индукция или стимуляция иммунного ответа усиливается в присутствии протеазы.
140. Способ по варианту осуществления 139, отличающийся тем, что протеаза продуцируется иммунной эффекторной клеткой, опухолью или клетками, присутствующими в микроокружении опухоли.
141. Способ по варианту осуществления 139 или по варианту осуществления 140, отличающийся тем, что протеаза продуцируется иммунной эффекторной клеткой, а иммунная эффекторная клетка представляет собой активированную Т-клетку, клетку-натуральный киллер (NK) или NK Т-клетку.
142. Способ по варианту осуществления 141, отличающийся тем, что иммунная эффекторная клетка находится вблизи клеток, которые экспрессируют антиген.
143. Способ по любому из вариантов осуществления 137-142, отличающийся тем, что протеаза продуцируется опухолью, которая находится вблизи клеток, которые экспрессируют TAA в ткани, и/или продуцируется опухолью, которая локализована совместно с TAA в ткани, и где протеаза расщепляет расщепляемый линкер в полиспецифической полипептидной конструкции, когда полиспецифическая полипептидная конструкция подвергается воздействию протеазы.
144. Способ по любому из вариантов осуществления 137-143, отличающийся тем, что протеазу выбирают из матриптазы, матриксной металлопротеазы (ММР), гранзима В и их комбинаций.
145. Способ по варианту осуществления 144, отличающийся тем, что протеаза представляет собой гранзим B.
146. Способ по любому из вариантов осуществления 137-145, отличающийся тем, что контактирование осуществляют ex vivo или in vitro.
147. Способ по любому из вариантов осуществления 137-145, отличающийся тем, что контактирование осуществляют in vivo у пациента.
148. Способ стимулирования или индукции иммунного ответа у пациента, включающий введение пациенту, нуждающемуся в этом, терапевтически эффективного количества полиспецифического конъюгата по любому из вариантов осуществления 1-113 или по варианту осуществления 134 или фармацевтической композиции по варианту осуществления 109 или по варианту осуществления 110.
149. Способ по варианту осуществления 137-147 и 148, который повышает клеточный иммунитет.
150. Способ по любому из вариантов осуществления 137-147, 148 и 149, который повышает активность Т-клеток.
151. Способ по любому из вариантов осуществления 137-147, 148-150, который повышает цитолитическую активность T-клеток (CTL).
152. Способ по любому из вариантов осуществления 137-147, 148-151, отличающийся тем, что иммунный ответ повышается против опухоли или злокачественного новообразования.
153. Способ по любому из вариантов осуществления 137-147, 148-152, отличающийся тем, что способом лечат заболевание или состояние у пациента.
154. Способ лечения заболевания или состояния у пациента, включающий введение пациенту, нуждающемуся в этом, терапевтически эффективного количества полиспецифического конъюгата по любому из вариантов осуществления 1-113 или фармацевтической композиции по варианту осуществления 135 или по варианту осуществления 136.
155. Способ по варианту осуществления 153 или по варианту осуществления 154, отличающийся тем, что заболевание или состояние представляет собой опухоль или злокачественное новообразование.
156. Способ по любому из вариантов осуществления 147, 148-155, отличающийся тем, что указанным пациентом является человек.
157. Полиспецифическая полипептидная конструкция, которая в неактивном состоянии содержит первый компонент и второй компонент, где первый и второй компоненты функционально связаны, причем каждый из первого и второго компонентов содержит антигенсвязывающий домен, который связывается с опухолесипецифическим антигеном (TAA), где первый компонент содержит Fc-область, где второй компонент содержит CD3-связывающую область, и где первый и второй компоненты связаны расщепляемым линкером.
158. Полиспецифический полипептид по варианту осуществления 157, отличающийся тем, что в неактивном состоянии связывание CD3-связывающей области с CD3 ингибируется или существенно снижается.
159. Полиспецифический полипептид по варианту осуществления 157, отличающийся тем, что в активированном состоянии первый и второй компоненты не являются функционально связанными.
160. Полиспецифический полипептид по варианту осуществления 157, отличающийся тем, что в активированном состоянии второй компонент связывает эпсилон-цепь CD3 (CD3ε) и опухолеспецифический антиген (TAA).
161. Полиспецифическая полипептидная конструкция, которая в активированном состоянии связывает эпсилон-цепь CD3 (CD3ε) и опухолеспецифический антиген (TAA), причем полиспецифическая полипептидная конструкция содержит:
первый антигенсвязывающий домен, который связывается с первым эпитопом на TAA;
антитело или его антигенсвязывающий фрагмент, которые связываются с CD3ε;
полипептид Fc-области иммуноглобулина;
второй антигенсвязывающий домен, который связывается со вторым эпитопом на TAA; и
расщепляемый линкер, связанный с полипептидной Fc-областью иммуноглобулина и вторым антигенсвязывающим доменом, где расщепляемый линкер представляет собой полипептид, который функционирует в качестве субстрата для протеазы.
162. Полиспецифическая полипептидная конструкция по варианту осуществления 161, отличающаяся тем, что протеаза продуцируется иммунной эффекторной клеткой.
163. Полиспецифическая полипептидная конструкция по варианту осуществления 162, отличающаяся тем, что иммунная эффекторная клетка находится вблизи клеток, которые экспрессируют TAA.
164. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 161-163, отличающаяся тем, что протеаза расщепляет расщепляемый линкер в полиспецифической полипептидной конструкции, когда полиспецифическая полипептидная конструкция подвергается воздействию протеазы.
165. Полиспецифическая полипептидная конструкция по варианту осуществления 161, отличающаяся тем, что протеаза продуцируется опухолью, которая находится вблизи клеток, которые экспрессируют TAA в ткани, и/или продуцируется опухолью, которая локализована совместно с TAA в ткани, и где протеаза расщепляет расщепляемый линкер в полиспецифической полипептидной конструкции, когда полиспецифическая полипептидная конструкция подвергается воздействию протеазы.
166. Полиспецифическая полипептидная конструкция по варианту осуществления 161, отличающаяся тем, что расщепляемый линкер представляет собой полипептид длиной до 25 аминокислот.
167. Полиспецифическая полипептидная конструкция по варианту осуществления 161, отличающаяся тем, что расщепляемый линкер представляет собой субстрат для матриптазы, матриксной металлопротеазы (ММР), гранзима В и их комбинаций.
168. Полиспецифическая полипептидная конструкция по варианту осуществления 161, отличающаяся тем, что расщепляемый линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 24-31.
169. Полиспецифическая полипептидная конструкция по варианту осуществления 161, отличающаяся тем, что каждый из первого антигенсвязывающего домена и второго антигенсвязывающего домена представляет собой антитело или его антигенсвязывающий фрагмент, выбранный из группы, состоящей из Fab-фрагмента, F(ab')2-фрагмента, Fv-фрагмента, scFv, scAb, dAb, однодоменного антитела с тяжелой цепью и однодоменного антитела с легкой цепью.
170. Полиспецифический слитый полипептид по варианту осуществления 169, отличающийся тем, что антитело или его антигенсвязывающий фрагмент представляет собой Fv, scFv, Fab, однодоменное антитело (sdAb), VNAR или VHH.
171. Полиспецифический слитый полипептид по варианту осуществления 169, отличающийся тем, что антитело или его антигенсвязывающий фрагмент представляет собой sdAb.
172. Полиспецифический слитый полипептид по варианту осуществления 171, отличающийся тем, что sdAb представляет собой человеческое или гуманизированное sdAb.
173. Полиспецифический слитый полипептид по варианту осуществления 171, отличающийся тем, что sdAb представляет собой VHH, VNAR, сконструированный домен VH или сконструированный домен VK.
174. Полиспецифический слитый полипептид по варианту осуществления 161, отличающийся тем, что полипептид Fc-области иммуноглобулина содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-2.
175. Полиспецифический слитый полипептид по варианту осуществления 174, отличающийся тем, что полипептид Fc-области иммуноглобулина содержит, по меньшей мере, одну модификацию для усиления связывания FcγR.
176. Полиспецифический слитый полипептид по варианту осуществления 175, отличающийся тем, что модификация представляет собой модификацию по Ser239 или Ile332.
177. Полиспецифический слитый полипептид по варианту осуществления 174, отличающийся тем, что гликозилирование Fc-области модифицируют для усиления связывания FcγR по сравнению с немодифицированной Fc-областью.
178. Полиспецифический слитый полипептид по варианту осуществления 177, отличающийся тем, что гликозилированная Fc иммуноглобулина не содержит или имеет пониженное содержание фукозы.
179. Полиспецифический слитый полипептид по варианту осуществления 174, отличающийся тем, что полипептид Fc-области иммуноглобулина содержит, по меньшей мере, одну модификацию для индукции гетеродимеризации.
180. Полиспецифический слитый полипептид по варианту осуществления 179, отличающийся тем, что модификация находится в положении, выбранном из группы, состоящей из Thr366, Leu368 и Tyr407 и их комбинаций.
181. Полиспецифический слитый полипептид по варианту осуществления 180, отличающийся тем, что модификация выбрана из группы, состоящей из Thr366Ser, Leu368Ala, Tyr407Val и их комбинаций.
182. Полиспецифический слитый полипептид по варианту осуществления 181, дополнительно включающий модификацию нецистеинового остатка в цистеиновый остаток в положении, выбранном из группы, состоящей из Ser354, Y349 и их комбинаций.
183. Полиспецифический слитый полипептид по варианту осуществления 181, дополнительно содержащий модификацию по остатку Ile235.
184. Полиспецифический слитый полипептид по варианту осуществления 183, отличающийся тем, что модификация представляет собой Ile235Arg.
185. Полиспецифический слитый полипептид по варианту осуществления 174, отличающийся тем, что полипептид Fc-области иммуноглобулина содержит модификацию по остатку Ile235.
186. Полиспецифический слитый полипептид по варианту осуществления 185, отличающийся тем, что модификация представляет собой Ile235Arg.
187. Полиспецифический слитый полипептид по варианту осуществления 174, отличающийся тем, что полипептид Fc-области иммуноглобулина содержит по меньшей мере одну модификацию для усиления связывания FcRn.
188. Полиспецифический слитый полипептид по варианту осуществления 187, отличающийся тем, что модификация находится в положении, выбранном из группы, состоящей из Met252, Ser254, Thr256, Met428, Asn434 и их комбинаций.
189. Полиспецифический слитый полипептид по варианту осуществления 188, отличающийся тем, что модификация находится в положении, выбранном из группы, состоящей из Met252Y, Ser254T, Thr256E, Met428L, Met428V, Asn434S и их комбинаций.
190. Полиспецифический слитый полипептид по варианту осуществления 187, отличающийся тем, что модификация находится в положении Met252 и в положении Met428.
191. Полиспецифический слитый полипептид по варианту осуществления 190, отличающийся тем, что модификация представляет собой Met252Y и M428L.
192. Полиспецифический слитый полипептид по варианту осуществления 190, отличающийся тем, что модификация представляет собой Met252Y и M428V.
193. Полиспецифическая полипептидная конструкция по варианту осуществления 161, отличающаяся тем, что полиспецифическая полипептидная конструкция содержит первый связывающий пептид (LP1) между первым антигенсвязывающим доменом и полипептидом Fc-области иммуноглобулина.
194. Полиспецифическая полипептидная конструкция по варианту осуществления 161, отличающаяся тем, что полиспецифическая полипептидная конструкция содержит второй связывающий пептид (LP2) между анти-CD3-связывающим доменом и вторым антигенсвязывающим доменом.
195. Полиспецифическая полипептидная конструкция по варианту осуществления 161, отличающаяся тем, что полиспецифическая полипептидная конструкция содержит первый связывающий пептид (LP1) между первым антигенсвязывающим доменом и полипептидом Fc-области иммуноглобулина и второй связывающий пептид (LP2) между анти-CD3-связывающим доменом и вторым антигенсвязывающим доменом, и где полиспецифическая полипептидная конструкция в нерасщепленном состоянии имеет структурное расположение от N-конца к C-концу следующим образом: первый антигенсвязывающий домен-LP1-полипептид Fc-области полипептида иммуноглобулина-расщепляемый линкер-анти-CD3-связывающий домен-LP2-второй антигенсвязывающий домен.
196. Полиспецифическая полипептидная конструкция по варианту осуществления 195, отличающаяся тем, что два связывающих пептида не являются идентичными друг другу.
197. Полиспецифическая полипептидная конструкция по варианту осуществления 195, отличающаяся тем, что каждый из LP1 и LP2 представляет собой пептид длиной примерно 1-20 аминокислот.
198. Полиспецифическая полипептидная конструкция по варианту осуществления 161, отличающаяся тем, что антитело или его антигенсвязывающий фрагмент, который связывается с CD3ε, представляет собой Fv-фрагмент антитела.
199. Полиспецифическая полипептидная конструкция по варианту осуществления 198, отличающаяся тем, что Fv-фрагмент антитела содержит стабилизированный дисульфидом анти-CD3-связывающий Fv-фрагмент (dsFv).
200. Полиспецифическая полипептидная конструкция по варианту осуществления 199, отличающаяся тем, что анти-CD3 dsFv включает VH CDR1, содержащую аминокислотную последовательность TYAMN (SEQ ID NO: 16); VH CD2, содержащую аминокислотную последовательность RIRSKYNNYATYYADSVKD (SEQ ID NO: 17); VH CDR3, содержащую аминокислотную последовательность HGNFGNSYVSWFAY (SEQ ID NO: 18), VL CDR1, содержащую аминокислотную последовательность RSSTGAVTTSNYAN (SEQ ID NO: 19); VL CDR2, содержащую аминокислотную последовательность GTNKRAP (SEQ ID NO: 20); и VL CDR3, содержащую аминокислотную последовательность ALWYSNLWV (SEQ ID NO: 21).
201. Полиспецифическая полипептидная конструкция по варианту осуществления 198, отличающаяся тем, что VH и VL, которые содержит анти-CD3-связывающий Fv, связаны с противоположными сторонами гетеродимерной Fc-области.
202. Полиспецифическая полипептидная конструкция по варианту осуществления 200, отличающаяся тем, что анти-CD3 dsFv содержит аминокислотную последовательность SEQ ID NO: 14.
203. Полиспецифическая полипептидная конструкция по варианту осуществления 200, отличающаяся тем, что анти-CD3 dsFv содержит аминокислотную последовательность SEQ ID NO: 15.
204. Полиспецифическая полипептидная конструкция по варианту осуществления 200, отличающаяся тем, что анти-CD3 dsFv содержит аминокислотную последовательность SEQ ID NO: 14 и аминокислотную последовательность SEQ ID NO: 15.
205. Полиспецифическая полипептидная конструкция по варианту осуществления 204, отличающаяся тем, что анти-CD3 dsFv включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 32-81.
206. Полиспецифическая полипептидная конструкция по варианту осуществления 204, отличающаяся тем, что анти-CD3 dsFv содержит комбинацию аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32-62, и аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 63 -81.
207. Полиспецифическая полипептидная конструкция по любому из вариантов осуществления 1-206, отличающаяся тем, что полиспецифическая полипептидная конструкция конъюгирована с агентом.
208. Полиспецифическая полипептидная конструкция по варианту осуществления 207, отличающаяся тем, что агент представляет собой терапевтический агент, противоопухолевый агент, токсин или его фрагмент, детектируемый фрагмент или диагностический агент.
209. Полиспецифическая полипептидная конструкция по варианту осуществления 207, отличающаяся тем, что агент конъюгирован с полиспецифической полипептидной конструкцией через линкер.
210. Фармацевтическая композиция, содержащая полиспецифическую полипептидную конструкцию любого из вариантов осуществления 1-209 и носитель.
211. Способ лечения или облегчения симптома клинического признака, связанного с расстройством у пациента, включающий введение полиспецифической полипептидной конструкции по любому из вариантов осуществления 157-209 или фармацевтической композиции по варианту осуществления 210 пациенту, нуждающемуся в этом, в количестве, достаточном для смягчения симптома клинического признака, связанного с расстройством.
212. Способ по варианту осуществления 211, отличающийся тем, что указанным пациентом является человек.
213. Способ по варианту осуществления 211, отличающийся тем, что расстройство представляет собой злокачественное новообразование.
VI. ПРИМЕРЫ
[0353] Следующие примеры включены только в иллюстративных целях и не предназначены для ограничения объема изобретения.
ПРИМЕР 1. Способ получения белков с ограниченным связыванием CD3
[0354] В примере 1 описано получение и экспрессия полиспецифических полипептидных конструкций, содержащих CD3-связывающая область, которая демонстрирует ограниченное связывание CD3. Полиспецифические конструкции были сгенерированы в различных конфигурациях, как показано на ФИГ. 1, ФИГ. 2, ФИГ. 3, ФИГ.4А-4С, ФИГ. 5А-5Е, ФИГ.6А-6В, ФИГ. 7, ФИГ. 8 и ФИГ. 9, чтобы содержать гетеродимерную Fc-область иммуноглобулина, связанного линкером (например, расщепляемым линкером) с CD3-связывающей областью, и один или более антигенсвязывающих доменов, которые связываются с опухолеспецифическим антигеном (TAA), расположенным на аминоконце относительно Fc области и/или карбоксиконце относительно CD3-связывающей области полиспецифической полипептидной конструкции. Были получены иллюстративные репрезентативные конструкции с различными антигенсвязывающими доменами TAA и линкерами.
[0355] Полинуклеотиды, кодирующие по меньшей мере первую полипептидную цепь и вторую полипептидную цепь гетеродимерной полиспецифической полипептидной конструкции, были получены и клонированы в плазмиду для экспрессии. Первая полипептидная цепь обычно включает по порядку от N-конца к C-концу: полипептид Fc-впадины (например, представленный в SEQ ID NO: 83); расщепляемый линкер, такой как линкер, содержащий один или более сайтов распознавания субстрата для протеазы; и вариабельный домен легкой цепи (VL) анти-CD3 dsFv антитела (например, представленный в SEQ ID NO: 72). Вторая полипептидная цепь обычно включает по порядку, от N-конца к C-концу: полипептид Fc-впадины (например, представленный в SEQ ID NO: 82); тот же расщепляемый линкер, что и первая полипептидная цепь; и вариабельный домен тяжелой цепи dsFv антитела против CD3 (например, представленный в SEQ ID NO: 44). За исключением отмеченного, иллюстративный расщепляемый линкер GGSGGGGIEPDIGGSGGS (SEQ ID NO: 105), содержащий сайт распознавания субстрата для гранзима B, использовали в иллюстративных конструкциях. Подобный линкер, TGGSGGGGIEPDIGGSGGS (SEQ ID NO: 148), использовали в иллюстративной конструкции cx1547, содержащей FRα. Для иллюстративной конструкции cx309 был использован иллюстративный расщепляемый линкер GSPAGLEADGSRQARVGS (SEQ ID NO: 166). Подобные конструкции могут быть сгенерированы с использованием других гетеродимерных конфигураций Fc, в том числе других конфигураций «выступ-во-впадину», таких как любая из описанных; другие линкеры, включая другие расщепляемые линкеры, в частности полипептидные линкеры, которые включают сайт узнавания субстрата для протеазы, такой как гранзим B, матриптаза и/или MMP; и другие CD3-связывающие области, включая другие антитела против CD3, включая dsFv или другие одновалентные фрагменты; или другие антигенсвязывающие фрагменты TAA, такие как формата scFv, sdAb или Fab.
[0356] В некоторых случаях были сгенерированы подобные конструкции, за исключением того, что они содержали нерасщепляемый линкер. Нерасщепляемый линкер включал линкеры размером от 3 до 18 аминокислот. Примерами нерасщепляемого линкера, используемого в иллюстративных сгенерированных молекулах, были GGS (например, содержится в иллюстративной конструкции cx1356), GGSGGS (SEQ ID NO: 10, содержится в иллюстративной конструкции cx1357), GGSGGSGGS (SEQ ID NO: 11, содержится в иллюстративной конструкции cx1358), GGSGGSGGSGGS (SEQ ID NO: 12, содержится в иллюстративной конструкции cx1359), GGSGGSGGSGGSGGS (SEQ ID NO: 13, содержится в иллюстративной конструкции cx1360) и GGGGGSGGGGGGSGGGGGSGSGGGSG (SEG IDGGGGGGGGGGGGGGGGGGGGGGGGGUSSWG)) GGSGGSGGSGGS (SEQ ID NO: 12, содержится в иллюстративной конструкции cx1359), GGSGGSGGSGGSGGS (SEQ ID NO: 13, содержится в иллюстративной конструкции cx1360) и GGGGGSGGGGGSGGGGGS (SEQ ID NO:119) или GGSGGGGSGGGGSGGGGS (SEQ ID NO: 147, содержится в иллюстративной конструкции cx681).
[0357] Одна или обе полипептидные цепи дополнительно кодируют один или более антигенсвязывающих доменов ТАА, расположенных на аминоконце относительно домена Fc и/или карбоксиконце относительно CD3-связывающей области в различных конфигурациях. Когда TAA был представлен в виде одноцепочечного фрагмента, например sdAb или scFv, антигенсвязывающий домен TAA был связан на N-конце с одной или обеими полипептидными цепями гетеродимера Fc (например, впадины и/или выступа) пептидным линкером например, PGGGG (SEQ ID NO: 102) и/или был связан на С-конце с одним или обоими доменами (например, VH и/или VL) CD3-связывающей области пептидным линкером, например, GGGG (SEQ ID NO: 103). Могут быть использованы другие подобные пептидные линкеры. Когда TAA был представлен в виде Fab, как например, в иллюстративной конструкции, обозначенной cx3313 или cx3315, то в плазмиду клонировали дополнительный полинуклеотид, кодирующий легкую цепь Fab. В этом примере кодируемые полипептиды включали одну или более полипептидных цепей, в которых VH-CH1 (Fd) Fab был связан на N-конце одной или обеих полипептидных цепей гетеродимера Fc и/или связан на С-конце одного или обоих доменов CD3-связывающей области, аналогично тому, как указанно выше. Третий полинуклеотид кодировал VL-CL Fab. Любой антигенсвязывающий домен, который связывается с TAA, можно использовать в представленных полиспецифических полипептидных конструкциях.
[0358] Иллюстративные сгенерированные белки содержали антигенсвязывающий домен, который связывает один из следующих опухолеспецифических антигенов: альфа-рецептор фолата (FRα), B7H3, EGFR, 5T4, CD20 и DLL3. Сгенерированные белки были либо составлены из последовательностей VH и VL, собранных в виде Fab или scFv, либо были сгенерированы так, чтобы содержать связывающие домены в виде однодоменных антител (sdAb). Различные TAA-связывающие домены, когда они используются в настоящем описании, включая sdAb, scFv и Fab, в том числе из следующих источников: cMet sdAb, как представлено в SEQ ID NO: 123 (патент США №93446884); B7H3 Fab и scFv, как представлено в SEQ ID NO: 127 и 128, и scFv, как представлено в SEQ ID NO: 124 (публикация PCT № WO2017030926); 5T4 scFv, как представлено в SEQ ID NO: 129, 130, 167 и 168 (патент США № US 8044178); DLL3 scFv, как представлено в SEQ ID NO: 189 (публикация США № US 2017/0037130); CD20 GA101, как представлено в SEQ ID NO: 125, 189 и 190 (публикация США № US 2005/0123546).Были получены полиспецифические полипептидные конструкции, содержащие 1, 2, 3 или 4 антигенсвязывающих домена ТАА, например, для обеспечения одновалентного, двухвалентного, трехвалентного или четырехвалентного связывания. В некоторых случаях антигенсвязывающие домены TAA были одинаковыми. В некоторых случаях антигенсвязывающие домены TAA были разными, так что сгенерированные полиспецифические полипептидные конструкции проявляли специфичность по меньшей мере к двум различным TAA, к различным эпитопам одного и того же TAA или одинаковым эпитопам одного и того же TAA. Например, были получены примерные полиспецифические полипептидные конструкции с двумя мишенями, содержащие антигенсвязывающий домен со специфичностью для EGFR и cMet (см., например, ФИГ. 5C и иллюстративные конструкции, обозначенные cx2973, cx2979 и cx2977). Кроме того, были получены типичные полиспецифические полипептидные конструкции, содержащие разные антигенсвязывающие домены TAA, специфичные для одного и того же эпитопа B7H3 или для двух разных эпитопов B7H3 (например, ФИГ. 6A и иллюстративные конструкции, обозначенные cx2846 и cx3094 соответственно) или для того же эпитопа 5T4 (например, ФИГ. 7, примерная конструкция cx3252).
[0359] Полинуклеотиды были получены для кодирования полипептидных цепей иллюстративных полиспецифических полипептидных конструкций, обозначенных как cx1547, cx309, cx1356, cx1357, cx1358, cx1359, cx1360, cx681 и cx1762 (каждая нацелена на FRα); cx2513 и cx3030 (каждая нацелена на EGFR); cx2973, cx2979, cx2977 (каждая нацелена на cMET и EGFR); cx3095, cx2846, cx3094, cx3314 и cx3313 (каждая нацелена на B7H3); cx3315, cx3265 и cx3262 (каждая нацелена на 5T4); cx3309 (нацелена на CD20); и cx3308 (нацелена на DLL3), включая конструкции, изображенные на ФИГ. 3, 4А (верхняя панель)-4С, 5А-5Е, 6А-6В, 7, 8 и 9. Конструкции были сконструированы так, чтобы содержать расщепляемый или нерасщепляемый линкер.
[0360] Отдельные плазмиды, кодирующие каждую цепь гетеродимерного белка с ограниченным связыванием с CD3, транзиторно трансфецировали в эквимолярном соотношении в клетки млекопитающих (или HEK293, или CHO) с использованием полиэтиленимина. Рекомбинантный белок, секретируемый в супернатант, собирали через 3-7 дней и очищали с помощью хроматографии на протеине А с последующей препаративной эксклюзионной хроматографией (SEC) или проточной хроматографией гидрофобного взаимодействия (HIC). Гетеродимерный белок был селективно очищен благодаря мутации, встроенной в одну цепь гетеродимерной Fc-области в положении I253R или H435R (обычно в Fc-впадины), так что он не связывал протеин A. Вторую стадию хроматографии на SEC (AKTA с Superdex-200 смолой) или FT-HIC (AKTA с бутил/фенилсефарозой) использовали для удаления нежелательных перекрестно спаренных частиц, содержащих два гетеродимерных Fc, которые были более гидрофобными и в два раза превышали ожидаемую молекулярную массу.
[0361] Метод способствовал получению гетеродимерных полиспецифических полипептидных конструкций, содержащих корректно спаренные виды гетеродимерной Fc-области и стабилизированный дисульфидом анти-CD3 Fv, как описано (анти-CD3 VH с мутацией G44C, как представлено в SEQ ID NO: 44, и VL с мутацией G100C, как представлено в SEQ ID NO: 72). Очищенный гетеродимерный белок с ограниченным связыванием с CD3 был стабильным и не накапливал перекрестно спаренные виды при длительной инкубации при 4ºC или повышенной концентрации белка.
[0362] На ФИГ.10А показано изображение SDS-PAGE FRα-нацеленной конструкции с ограниченным рекрутированием CD3, cx1547, восстанавливающие условия (R) и невосстанавливающие (NR) (ожидаемая молекулярная масса 135 кДа). На ФИГ.10B и 10C показана хроматограмма из эксклюзионного анализа cx1547, демонстрирующая, что наблюдали один вид с определенной молекулярной массой 137,9 кДа.
ПРИМЕР 2. Связывание с опухолевыми клетками и первичными Т-клетками с помощью проточной цитометрии
[0363] В этом примере описаны исследования, оценивающие связывание иллюстративных конструкций с Т-клетками или опухолевыми клетками. Эти исследования проводили в отдельных культурах, содержащих либо только Т-клетки, либо только опухолевые клетки в изоляции друг от друга.
1. Связывание с первичными Т-клетками в расщепленном и нерасщепленном состоянии
[0364] Связывание иллюстративной полиспецифической полипептидной конструкции по изобретению, обозначенной здесь как cx309, с CD3 на поверхности первичных Т-клеток оценивали после протеолитического расщепления его расщепляемого линкера GSPAGLEADGSRQARVGS (SEQ ID NO: 27), который содержит сайты распознавания субстрата для матриптазы, гранзима B и MMP-2. Опухолеспецифические связывающие домены cx309 связываются с рецептором фолата альфа (FRα), который не экспрессируется на первичных Т-клетках.
[0365] Первичные Т-клетки были отрицательно обогащены из РВМС, выделенными из лейкопаков здорового донора человека. Там, где отмечено, cx309 предварительно расщепляли перед добавлением к клеткам, с расщеплением, подтвержденным SDS-PAGE. В частности, cx309 подвергали воздействию матриптазы (ФИГ. 11A) или матричной металлопротеазы 2 (MMP-2) (ФИГ. 11B) (расщепленная) или не подвергали воздействию протеазы (нерасщепленная). Конструкцию cx309, расщепленную или нерасщепленную, титровали на клетки или добавляли в одной насыщающей концентрации. Связанную cx309 детектировали с помощью конъюгированных с флуорофором вторичных антител, специфичных для человеческого Fc или гуманизированного vhh, и связывание измеряли с помощью проточной цитометрии. Клетки, инкубированные со вторичным антителом, служили только в качестве отрицательных контролей.
[0366] Как показано на ФИГ. 11А и 11В, cx309, связанный с Т-клетками после расщепления конструкции, либо матриптазой, либо ММР-2 в линкере между Fc и CD3-связывающим доменом до начала анализа связывания. Никакого связывания с Т-клетками у нерасщепленной конструкцией не наблюдалось, показывая, что cx309 была способна связываться с Т-клетками в расщепленном, то есть активном состоянии, но демонстририовала недетектируемое связывание Т-клеток в нерасщепленной форме.
2. Сравнение связывания с Т-клетками против антиген-экспрессирующих опухолевых клеток
[0367] Дополнительные репрезентативные FRα-нацеленные конструкции с ограниченным рекрутированием CD3 с различными линкерами между Fc и компонентом CD3-связывающих доменов оценивали на связывание с Т-клетками, как описано выше, и с клетками, экспрессирующими FRα (Ovcar-5). Для исследований использовали 100 нМ каждой конструкции, cx1356, cx681 или cx1547. Было обнаружено, что дополнительные репрезентативные FRα-нацеленные конструкции с ограниченным рекрутированием CD3 с различными линкерами между Fc и компонентом CD3-связывающих доменов, связывают клетки, экспрессирующие FRα (Ovcar-5) (ФИГ. 12A и 12C), но они не способны связываться с Т-клетками (ФИГ. 12B и 12D).
[0368] В аналогичном исследовании было обнаружено, что репрезентативная EGFR-нацеленная конструкция с ограниченным рекрутированием CD3, сгенерированная, по существу, как описано в Примере 1, cx3030, связывается с клетками, экспрессирующими EGFR (Colo-205) (ФИГ. 13A), но не обладает способностью связываться с T- клетками (ФИГ. 13B). Использовали 400 нМ каждой конструкции. Это наблюдение дополнительно демонстрирует, что, будучи изолированными, конструкции с ограниченным рекрутированием CD3 демонстрируют ослабленное связывание Т-клеток.
[0369] Как показано на ФИГ. 14A-14D, аналогичные результаты наблюдали для репрезентативных B3H3-нацеленных ограниченных CD3-рекрутеров, cx3313, cx3314 и cx3095.Cx3313 и cx3314 были получены, по существу, как описано в Примере 1, и состояли из тех же последовательностей VH и VL B7H3, которые собраны как Fab или scFv, соответственно, как описано в Примере 1. cx3095 содержал B7H3-связывающие домены, которые представляли собой однодоменное антитело. Как показано на ФИГ. 14A и 14B, было обнаружено, что репрезентативные B3H3-нацеленные ограниченные CD3-рекрутеры связываются с клетками A375, экспрессирующими B7H3. Однако, как показано на ФИГ. 14C и 14D, одни и те же конструкции не могли связываться с Т-клетками по отдельности. В этих исследованиях связывание репрезентативных B7H3-нацеленных ограниченных CD3-рекрутеров, cx3313, cx3314 и cx3095 сравнивали с двойным аффинным повторно нацеливающим антителом (DART) - мономерного формата Fc, нацеленным на B7H3 и CD3 (DART-Fc B7H3xCD3; см., например, WO2017030926A1). DART-Fc B7H3xCD3 содержал последовательность B7H3, как представлено в SEQ ID NO: 169, 145 или 146. Следует отметить, что только формат DART-Fc позволяет связываться с Т-клетками в отсутствие взаимодействия с B7H3 (ФИГ. 14C и 14D), тогда как все форматы демонстрируют связывание с B7H3-экспрессирующими клетками (ФИГ. 14А и 14В).
[0370] В дальнейшем исследовании аналогичное ограниченное связывание наблюдалось для репрезентативных 5T4-нацеленных конструкций с ограниченным рекрутированием CD3, полученных, по существу, как описано в Примере 1, cx3262 и cx3315. 5T4-связывающие домены cx3262 были сгенерированы как однодоменные антитела, тогда как связывающий домен 5T4 cx3315 был получен с scFv, полученным, как представлено в SEQ ID NO: 167 и 168 (см. Патент США № US 8044178). Было обнаружено, что обе репрезентативные 5T4-нацеленные конструкции с ограниченным рекрутированием CD3 связываются с 5T4-экспрессирующими клетки (Ovcar-5) (ФИГ. 15A), но при этом они не способны связываться с T-клетками (ФИГ. 15B). Использовали 400 нМ каждой конструкции. Это наблюдение дополнительно демонстрирует, что конструкции с ограниченным рекрутированием CD3, описанные в настоящем документе, демонстрируют ослабленное связывание Т-клеток, по отдельности.
[0371] Как показано на ФИГ. 16A-16D, репрезентативная CD20-нацеленная конструкция с ограниченным рекрутированием CD3, cx3490, как было обнаружено, связывается с клетками, экспрессирующими CD20 (Ramos), но не обладает способностью связываться с T-клетками. Использовали 100 нМ конструкции cx3490. CD20-связывающие домены cx3490 представляли собой scFv, полученные из VH и VL антитела против CD20 GA101, как описано в Примере 1.
ПРИМЕР 3. Оценка CD3 репортерной T-клеточной активации с использованием репортерного анализа
[0372] В этом примере описана оценка способности различных конструкций активировать CD3 NFAT репортерную клеточную линию Jurkat в совместных культурах с клетками, экспрессирующими антиген-мишень. Эти анализы были использованы для демонстрации того, что, хотя связывание Т-клеток через CD3-связывающий домен ограничено или ингибируется на изолированных Т-клетках (как показано в Примере 2), как только представленные в настоящем описании полиспецифические полипептиды связываются с родственным антигеном, они способны рекрутировать Т-клетки и опосредовать активацию Т-клеток.
1. Репортерный анализ с использованием люциферазы
[0373] Антиген-нацеленные конструкции с ограниченным рекрутированием CD3 титровали на совместные культуры клеток-мишеней и генетически сконструированных клеток Jurkat, которые экспрессируют NFAT-управляемый люциферазный репортер (Promega, США). В этом анализе рекрутирование CD3 приводит к передаче сигналов NFAT и продукции внутриклеточной люциферазы. Аналитические планшеты инкубировали при 37 °С в течение приблизительно 6 часов и затем уравновешивали до комнатной температуры. Реагент Bio-Glo добавляли в лунки для образцов и люминесценцию супернатантов измеряли с использованием устройства для считывания микропланшетов SpectraMax L.
[0374] Как показано на ФИГ. 17, репрезентативная FR3α-нацеленная конструкция с ограниченным рекрутированием CD3, cx309, продемонстрировала значительно повышенную способность активировать Т-клетки при расщеплении по линкеру между Fc и CD3-связывающим доменом. Здесь cx309 предварительно расщепляли матриптазой перед началом анализа. Примечательно, что активация Т-клеток наблюдалась только в присутствии FRα-положительной клеточной линии OVCAR-5. Некоторая активация Т-клеток наблюдалась с нерасщепленной cx309. Этот результат согласуется со способностью конструкций с ограниченным рекрутированием CD3 проявлять связывание CD3 при связывании с его антигеном с повышенным рекрутированием CD3 после протеолитического расщепления.
2. GFP репортерный анализ
[375] Антиген-нацеленные конструкции с ограниченным рекрутированием CD3 титровали на совместные культуры клеток-мишеней и сконструированных клеток Jurkat, которые экспрессируют NFAT-управляемый зеленый флуоресцентный белок (GFP). Рекрутирование CD3 приводит к передаче сигналов NFAT и выработке зеленой флуоресценции. Для репортерных анализов с использованием адгезивных клеток-мишеней, клетки-мишени высевали, им давали возможность осесть при комнатной температуре для равномерного распределения и инкубировали в течение нескольких часов при 37 °C, чтобы обеспечить адгезию до добавления репортерных клеток и антиген-нацеленных конструкций с ограниченным рекрутированием CD3. Аналитические планшеты последовательно визуализировали с использованием системы IncuCyte ZOOM, и активацию репортерных клеток CD3 определяли путем измерения общей интегральной интенсивности зеленого объекта.
[0376] На ФИГ. 18 и 19A-19D показаны конструкции, нацеленные на рецептор фолата (FRα); на ФИГ. 20А-20D показаны конструкции, нацеленные на EGFR или EGFR и cMET; На ФИГ. 21А-21В и 22А-22F показаны конструкции, нацеленные на B7H3; На ФИГ. 23А-23В и 24 показаны конструкции, нацеленные на 5Т4; на ФИГ. 25 показана конструкция, нацеленная на CD20; и на ФИГ. 26 показана конструкция, нацеленная на DLL3. Как показано на всех фигурах, каждая из протестированных репрезентативных конструкций была способна индуцировать антиген-зависимую активацию Т-клеток в присутствии клеток, экспрессирующих мишень, как показано повышенным сигналом GFP. На ФИГ. 22A-F показано, что степень активации аналогична или, в некоторых случаях, выше, чем у молекулы-компаратор DART-Fc: B7H3xCD3, описанной в Примере 2.
[0377] Примечательно, что ФИГ. 19A-19D демонстрируют улучшенную способность активации T-клеток, которая достигается, когда удаляется Fc-часть конструкций с ограниченным рекрутированием CD3, что позволяет неограниченное взаимодействие T-клеток. Fc этих конструкций с ограниченным рекрутированием CD3 может быть удален посредством протеолиза, если протеолитически расщепляемые линкеры включены между Fc и CD3-связывающим доменом. cx3238 представляет собой полностью отщепленную C-концевую часть и была получена путем коэкспрессии двух плазмид, кодирующих 1) анти-CD3 VH, связанный с С-концевым FRα-связывающим sdAb и 2) анти-CD3 VL, связанный с C -концевым FRα-связывающим sdAb. Этот результат согласуется с позиционированием домена Fc как ограничения способности CD3-связывающей области связываться с CD3.
ПРИМЕР 4. Оценка длины линкера по активности
[0378] Влияние линкеров различной длины между Fc и компонентными доменами (VH и VL), которые составляют CD3-связывающую область, на способность активировать Т-клетки тестировали с использованием репортерного анализа Jurkat, описанного в Примере 3. В этих анализах использовали FRα-нацеленные конструкции с ограниченным рекрутированием CD3, полученные, как описано в Примере 1, содержащие линкеры на основе GlySer различной длины, как указано в Таблице E1.
[0379] Как показано на ФИГ. 27A-27F, длина линкера и активирующая способность Т-клеток были положительно коррелированы. Было показано, что способность активации Т-клеток напрямую связана с длиной линкера, что указывает на то, что более короткие линкеры в большей степени ограничивают связывание CD3. Важно, что рекрутирование T-клеток в конструкции зависит от TAA-связывания, так как эти конструкции не демонстрируют способность связывать T-клетки в изоляции (например, в форме раствора, когда он не связан с TAA-мишенью), как показано выше в Примере 2 (ФИГ. 12A-12D). Вместе эти конструкции демонстрировали ограниченное или ингибированное связывание с CD3, но были способны активировать Т-клетки зависимым от мишени способом.
ПРИМЕР 5. Оценка функциональной активности
[0380] Этот пример описывает оценку и характеристику тестируемых конструкций с ограниченным рекрутированием CD3 в первичных Т-клетках человека в анализах in vitro.
1. Т-клеточно-опосредованая цитотоксичность
[0381] Клетки-мишени флуоресцентно метили CytoID красным. Для анализов цитотоксичности с использованием адгезивных клеток-мишеней клетки-мишени высевали, им давали возможность отстояться при комнатной температуре для равномерного распределения и инкубировали в течение нескольких часов при 37 °C, чтобы обеспечить адгезию до добавления других компонентов анализа. Первичные Т-клетки были отрицательно обогащены из РВМС, выделенных из лейкопаков здорового донора человека, и добавлены при соотношении Т-клеток к мишени 10:1-40:1. Добавляли реагент с зеленой каспазой-3/7, который флуоресцентно метил ядерную ДНК клеток, подвергающихся апоптозу. Антитела титровали на совместной культуре, и аналитические планшеты последовательно визуализировали с использованием системы IncuCyte ZOOM. Смерть клеток-мишеней определяли путем измерения общей площади красного/зеленого перекрытия объекта.
[0382] Как показано на ФИГ. 28A- 28C, cx1547, FRα-нацеленная конструкция с ограниченным рекрутированием CD3, индуцирующая сильную опосредованную T-клетками цитотоксичность антиген-положительных, но не антиген-отрицательных клеточных линий, что согласуется со способностью потенциально индуцировать антиген-зависимую активацию T-клеток. Аналогично, B7H3-нацеленные конструкции с ограниченным рекрутированием CD3 индуцировали сильную опосредованную Т-клетками цитотоксичность антиген-положительных, но не антиген-отрицательных клеточных линий, как показано на ФИГ. 29A-29F и 30. Эти конструкции демонстрировали аналогичные возможности в альтернативном формате, DART-Fc B7H3xCD3. Эти наблюдения подтверждают, что антиген-нацеленный формат с ограничением CD3, представленный в настоящем документе, по сравнению с другими форматами, связывающими CD3, известными в данной области, лишен или демонстрирует пониженное связывание Т-клеток отдельно, в то же время сохраняя сильную способность индуцировать антиген-зависимую цитотоксичность Т-клеток.
[0383] Как показано на ФИГ. 31A-31F, конструкция, представляющая C-концевой протеолитический продукт, содержащий только CD3-связывающий домен, функционально связанный с TAA-связывающими доменами, обозначенный cx2190 (см. ФИГ. 4C), демонстрирует повышенную способность индуцировать антиген-зависимую Т-клеточную цитотоксичность по сравнению с нерасщепленной FRα-нацеленной конструкцией с ограниченным рекрутированием CD3, обозначенной cx1762 (см. ФИГ. 4A).
[0384] Как показано на ФИГ. 32 - репрезентативная 5T4-нацеленная конструкция с ограниченным рекрутированием CD3, cx3315, индуцировала специфическую цитотоксичность Т-клеток в отношении линии клеток, экспрессирующих 5t4, Ovcar-5, но не в отношении линии 5T4-отрицательных клеток, CCRF-CEM.
2. Активация Т-клеток
[0385] Для оценки активации Т-клеток суспензионные клетки из анализов цитотоксичности, опосредованной Т-клетками, собирали и окрашивали на живые/ мертвые и конъюгированные с флуорофором анти-CD4, анти-CD8, анти-CD25 и/или анти-CD71 антитела. Клетки анализировали с использованием спектрального анализатора SONY SA3800 и определяли активацию CD4+ или CD8+ T-клеток путем измерения уровней экспрессии CD25 или CD71 или процента CD25- или CD71-положительных.
[0386] На ФИГ. 33 показана активация Т-клеток, измеренная путем инкубации расщепленных cx309 и нерасщепленных, то есть неактивных конструкций cx309 в течение 20 часов в совместной культуре T-клеток и клеток Ovcar5. Как показано, только расщепленная cx309 была способна опосредовать FRα-зависимую активацию Т-клеток посредством связывания CD3. Активацию Т-клеток контролировали с помощью анализа проточной цитометрии CD25% популяций CD4 и CD8.
[0387] Кроме того, была также протестирована конструкция, содержащая В7Н3 ТАА. Как показано на ФИГ. 34A-34H, B7H3-нацеленная конструкция с ограниченным рекрутированием CD3 также опосредовала B7H3-зависимую активацию Т-клеток через связывание CD3. Аналогичные возможности активации T-клеток конструкциями с ограниченным рекрутированием CD3, и форматом DART-Fc наблюдались, несмотря на значительные различия в связывании T-клеток этими двумя форматами (ФИГ. 14A-14D).
[0388] Таким образом, результаты продемонстрировали, что протестированные нацеленные на антиген конструкции с ограниченным рекрутированием CD3 индуцировали сильную антиген-зависимую активацию как CD4, так и CD8 Т-клеток.
3. Продукция Т-клеточных цитокинов (ИФА)
[0389] Супернатанты из анализов цитотоксичности, опосредованной Т-клетками, анализировали на содержание IFNγ с помощью сэндвич-ИФА (BioLegend, США). Следовали инструкциям производителя и получали стандартную кривую, из которой значения концентрации цитокинов в образцах супернатанта были интерполированы. Образцам, у которых значения поглощения были ниже нижнего предела обнаружения, была назначена концентрация цитокинов, равная половине концентрации самой низкой стандартной концентрации. На ФИГ. 35 показано, что репрезентативная B7H3-нацеленная конструкция с ограниченным рекрутированием CD3, как было установлено, вызывает продуцирование IFNγ Т-клетками антиген-зависимым образом.
4. Продуцирование Т-клеточных цитокинов (FluoroSpot)
[0390] Мембраны FluoroSpot покрывали антителами захвата к IFNγ и IL-2 в течение ночи при 4 °C. Мембраны промывали PBS и титровали антитела, добавляли клетки-мишени и PBMC или T-клетки, отрицательно обогащенные из PBMC. Для совместной культуры клеток-мишеней: РВМС клетки высевали в соотношении 1:20. Для совместной культуры клеток-мишеней: Т-клеток клетки высевали в соотношении 1:10. Аналитические планшеты инкубировали в течение ~ 24 ч при 37 °С и мембраны готовили в соответствии с инструкциями производителя (CTL). Мембраны визуализировали с использованием универсального анализатора CTL-ImmunoSpot S6. Количество пятен цитокинов измеряли с использованием одинаковых настроек времени воздействия и интенсивности в аналитических лунках. На ФИГ. 36A-36B и 37 изображена способность антиген-нацеленной конструкции с ограниченным рекрутированием CD3 вызывать продуцирование цитокинов из Т-клеток зависимым от FRα или B7H3 способом, соответственно.
5. Уничтожение диссоциированных опухолевых клеток
[0391] Образец диссоциированных опухолевых клеток яичника (Conversant) окрашивали Zombie-красным и флуорофор-конъюгированными анти-CD45 и анти-EpCAM-антителами для идентификации опухолевых клеток (CD45-/EpCAM +) и опухолевых инфильтрирующих иммунных клеток (CD45 +/EpCAM-) с помощью проточной цитометрии. Неокрашенные клетки высевали в 96-луночный культуральный планшет и добавляли репрезентативную FRα-нацеленную конструкцию с ограниченным рекрутированием CD3 и рекомбинантный человеческий IL-2 до конечной концентрации 20 нМ и 10 нг/мл. После культивирования при 37 °С в течение 6 дней аликвоты супернатанта собирали для анализа содержания IFNγ с помощью сэндвич-ИФА (описано выше) и оставшиеся супернатанты, содержащие неприкрепленные клетки, удаляли. Прикрепленные клетки осторожно промывали PBS для удаления остаточных суспензионных клеток и дебриса, к клеткам добавляли среду и анализировали лунки для анализа с использованием системы IncuCyte ZOOM для визуализации конфлуентности опухолевых клеток. Равный объем реагента жизнеспособности CellTiter-Glo добавляли в лунки для образцов, и люминесценцию измеряли с использованием устройства для считывания микропланшетов SpectraMax L. На ФИГ. 38A-38D показана способность антиген-нацеленной конструкции с ограниченным рекрутированием CD3 активировать Т-клетки, которые ранее проникли в образец опухоли, и опосредовать цитотоксичность и элиминацию опухолевых клеток.
Другие варианты осуществления
[0392] Хотя изобретение было описано в сочетании с его подробным описанием, вышеприведенное описание предназначено для иллюстрации, а не для ограничения объема изобретения, который определяется объемом прилагаемой формулы изобретения. Другие аспекты, преимущества и модификации находятся в пределах объема прилагаемой формулы изобретения.
ТАБЛИЦА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
X2=A, G, S, V, E, D, Q, N, или Y; (P3=A, G, S, V, E, D, Q, N, или Y)
X3=H, P, A, V, G, S, или T; (P2=H, P, A, V, G, S, или T)
X4=D или E; (P1=D или E)
X5=I, L, Y, M, F, V, T, S, G или A (P1' = I, L, Y, M, F, V, T, S, G или A)
(P3=E)
X3=P или A; (P2=P или A)
X5=I, V, T, S, или G (P1' = I, V, T, S, или G)
X1=любая аминокислота; (P1 представляет собой любую аминокислоту)
X5=A или V
X5=A или V
X1=P, V или A; (P3=P, V или A)
X2=Q или D; (P2=Q или D)
X3=A или N; (P1=A или N)
X4=L, I или M (P1' = L, I или M)
(P3=P) X2=Q или D; (P2=Q или D)
X3=A или N; (P1 представляет собой A или N)
X4=L или I (P1' представляет собой L или I)
TAVAWYQQKP GQSPKLLIYW ASTRLTGVPD RFTGSGSGTD FTLTISNVQS
EDLADYFCQQ YSSYPYTFGG GTKLEIK
ASYRYSGVPS RFSGSGSGTD FTLTISSLQP EDFATYYCQQ YNNYPFTFGQ
GTKLEIKGGG SGGGGEVQLV ESGGGLVQPG GSLRLSCAAS GFTFSTYAMN
WVRQAPGKGL EWVGRIRSKY NNYATYYADS VKDRFTISRD DSKNSLYLQM NSLKTEDTAV YYCVRHGNFG NSYVSWFAYW GQGTLVTVSS GGCGGGEVAA LEKEVAALEK EVAALEKEVA ALEKGGGDKT HTCPPCPAPE AAGGPSVFLF PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP
REPQVYTLPP SREEMTKNQV SLWCLVKGFY PSDIAVEWES NGQPENNYKT
TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF SCSVMHEALH NHYTQKSLSL
SPGK
где n=0-10
где n=1-4
где n=1-5
Xaa независимо выбирают из A, V, L, I, M, F, W, P, G, S, T, C, Y, N, Q, K, R, H, D, или E
x, y, и z каждое представляет собой целое число в диапазоне от 1 до 5
Xaa независимо выбирают из A, V, L, I, M, F, W, P, G, S, T, C, Y, N, Q, K, R, H, D, или E
n=2-20
n=2-20
n=2-20
n=2-20
n=2-20
n=2-20
n=2-20
n=1-9
VYYSGTTNYN PSLKSRVTIS VDTSKNQFSL KLSSVTAADT AVYYCASIAV
TGFYFDYWGQ GTLVTVSSGG GGSGGGGSGG GGSEIVLTQS PGTLSLSPGE
RVTLSCRASQ RVNNNYLAWY QQRPGQAPRL LIYGASSRAT GIPDRFSGSG
SGTDFTLTIS RLEPEDFAVY YCQQYDRSPL TFGGGTKLEI K
IFPGDGDTDY NGKFKGRVTI TADKSTSTAY MELSSLRSED TAVYYCARNV
FDGYWLVYWG QGTLVTVSS
LLIYQMSNLV SGVPDRFSGS GSGTDFTLKI SRVEAEDVGV YYCAQNLELP
YTFGGGTKVE IKRTV
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> INHIBRX LP
ECKELMAN, Brendan P.
KAPLAN, Michael D.
WILLIS, Katelyn M.
DEVERAUX, Quinn
TIMMER, John C.
<120> ПОЛИСПЕЦИФИЧЕСКИЕ ПОЛИПЕПТИДНЫЕ КОНСТРУКЦИИ, ОБЛАДАЮЩИЕ ОГРАНИЧЕННЫМ
СВЯЗЫВАНИЕМ С CD3, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ
<130> 744952000140
<150> 62/484,217
<151> 2017-04-11
<140> Not Yet Assigned
<141> Concurrently herewith
<160> 190
<170> PatentIn version 3.5
<210> 1
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG1 Fc
<400> 1
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
1 5 10 15
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
20 25 30
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
35 40 45
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
50 55 60
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
65 70 75 80
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
85 90 95
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
100 105 110
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
115 120 125
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
130 135 140
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
145 150 155 160
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
165 170 175
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
180 185 190
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
195 200 205
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 2
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG2 Fc
<400> 2
Pro Ala Pro Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
1 5 10 15
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
20 25 30
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
35 40 45
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
50 55 60
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
65 70 75 80
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
85 90 95
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
100 105 110
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
115 120 125
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
130 135 140
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
145 150 155 160
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
165 170 175
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
180 185 190
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
195 200 205
Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 3
<211> 217
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG2 Fc
<400> 3
Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 4
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG3 Fc
<400> 4
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
1 5 10 15
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
20 25 30
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp
35 40 45
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
50 55 60
Glu Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu
65 70 75 80
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
85 90 95
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly
100 105 110
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
115 120 125
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
130 135 140
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn
145 150 155 160
Asn Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe
165 170 175
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
180 185 190
Ile Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr
195 200 205
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 5
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG4 Fc
<400> 5
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
1 5 10 15
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
20 25 30
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
35 40 45
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
50 55 60
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
65 70 75 80
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
85 90 95
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
100 105 110
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
115 120 125
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
130 135 140
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
145 150 155 160
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
165 170 175
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
180 185 190
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
195 200 205
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 6
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG4 Fc
<400> 6
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
1 5 10 15
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
20 25 30
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
35 40 45
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
50 55 60
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
65 70 75 80
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
85 90 95
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
100 105 110
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
115 120 125
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
130 135 140
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
145 150 155 160
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
165 170 175
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
180 185 190
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
195 200 205
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 7
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Шарнир
<400> 7
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys
1 5 10
<210> 8
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Шарнир
<400> 8
Asp Lys Thr His Thr Cys Pro Pro Cys
1 5
<210> 9
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Шарнир
<400> 9
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
1 5 10
<210> 10
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> (GGS)2
<400> 10
Gly Gly Ser Gly Gly Ser
1 5
<210> 11
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> (GGS)3
<400> 11
Gly Gly Ser Gly Gly Ser Gly Gly Ser
1 5
<210> 12
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> (GGS)4
<400> 12
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
1 5 10
<210> 13
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> (GGS)5
<400> 13
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
1 5 10 15
<210> 14
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 Hv
<400> 14
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Lys Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Gln Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala
115 120 125
<210> 15
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 Lv
<400> 15
Gln Ala Val Val Thr Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Glu
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Thr Glu Asp Glu Ala Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 16
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH CDR1
<400> 16
Thr Tyr Ala Met Asn
1 5
<210> 17
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH CDR2
<400> 17
Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser
1 5 10 15
Val Lys Asp
<210> 18
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH CDR3
<400> 18
His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe Ala Tyr
1 5 10
<210> 19
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL CDR1
<400> 19
Arg Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn
1 5 10
<210> 20
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL CDR2
<400> 20
Gly Thr Asn Lys Arg Ala Pro
1 5
<210> 21
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL CDR3
<400> 21
Ala Leu Trp Tyr Ser Asn Leu Trp Val
1 5
<210> 22
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> субстрат Гранзима B
<400> 22
Leu Glu Ala Asp
1
<210> 23
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> субстрат Гранзима B
<400> 23
Arg Gln Ala Arg
1
<210> 24
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> субстрат MMP
<400> 24
Pro Ala Gly Leu
1
<210> 25
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 25
Thr Gly Leu Glu Ala Asp Gly Ser Pro Ala Gly Leu Gly Arg Gln Ala
1 5 10 15
Arg Val Gly
<210> 26
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 26
Thr Gly Leu Glu Ala Asp Gly Ser Arg Gln Ala Arg Val Gly Pro Ala
1 5 10 15
Gly Leu Gly
<210> 27
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 27
Thr Gly Ser Pro Ala Gly Leu Glu Ala Asp Gly Ser Arg Gln Ala Arg
1 5 10 15
Val Gly Ser
<210> 28
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 28
Thr Gly Pro Ala Gly Leu Gly Leu Glu Ala Asp Gly Ser Arg Gln Ala
1 5 10 15
Arg Val Gly
<210> 29
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 29
Thr Gly Arg Gln Ala Arg Val Gly Leu Glu Ala Asp Gly Ser Pro Ala
1 5 10 15
Gly Leu Gly
<210> 30
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 30
Thr Gly Ser Arg Gln Ala Arg Val Gly Pro Ala Gly Leu Glu Ala Asp
1 5 10 15
Gly Ser
<210> 31
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 31
Thr Gly Pro Ala Gly Leu Gly Ser Arg Gln Ala Arg Val Gly Leu Glu
1 5 10 15
Ala Asp Gly Ser
20
<210> 32
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH1
<400> 32
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 33
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH2
<400> 33
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 34
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH3
<400> 34
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 35
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH4
<400> 35
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 36
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH5
<400> 36
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 37
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH6
<400> 37
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 38
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH7
<400> 38
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asp Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 39
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH8
<400> 39
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
115 120
<210> 40
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH9
<400> 40
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 41
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH10
<400> 41
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Tyr Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 42
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH11
<400> 42
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Lys Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Gln Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 43
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH12
<400> 43
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Lys Pro
115 120 125
<210> 44
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH13
<400> 44
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Lys Pro
115 120 125
<210> 45
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH14
<400> 45
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Cys Gly Thr Leu Val Thr Val Lys Pro
115 120 125
<210> 46
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH15
<400> 46
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 47
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH16
<400> 47
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 48
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH17
<400> 48
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 49
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH18
<400> 49
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 50
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH19
<400> 50
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 51
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH20
<400> 51
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 52
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH21
<400> 52
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 53
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH22
<400> 53
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 54
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH23
<400> 54
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 55
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH24
<400> 55
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 56
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH25
<400> 56
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 57
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH26
<400> 57
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 58
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH27
<400> 58
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asp Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 59
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH28
<400> 59
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
115 120
<210> 60
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH29
<400> 60
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 61
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH30
<400> 61
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Tyr Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 62
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VH31
<400> 62
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Lys Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Gln Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 63
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL1
<400> 63
Gln Ala Val Val Thr Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Glu
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Thr Glu Asp Glu Ala Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 64
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL2
<400> 64
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Lys Ser Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Ile Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
His Trp Val Phe Gly Cys Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 65
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL3
<400> 65
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Trp Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 66
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL4
<400> 66
Gln Ala Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Thr Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 67
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL5
<400> 67
Gln Ala Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Thr Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 68
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL6
<400> 68
Gln Ala Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Thr Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 69
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL7
<400> 69
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 70
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL8
<400> 70
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 71
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL9
<400> 71
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Ile Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
His Trp Val Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 72
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL10
<400> 72
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Ile Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
His Trp Val Phe Gly Cys Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 73
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL11
<400> 73
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Cys Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Ile Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
His Trp Val Phe Gly Glu Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 74
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL12
<400> 74
Gln Ala Val Val Thr Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Glu
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Thr Glu Asp Glu Ala Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu
100 105
<210> 75
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL13
<400> 75
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Lys Ser Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Ile Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
His Trp Val Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 76
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL14
<400> 76
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Trp Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu
100 105
<210> 77
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL15
<400> 77
Gln Ala Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Thr Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 78
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL16
<400> 78
Gln Ala Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Thr Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu
100 105
<210> 79
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL17
<400> 79
Gln Ala Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Thr Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu
100 105
<210> 80
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL18
<400> 80
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu
100 105
<210> 81
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> анти-CD3 VL19
<400> 81
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu
100 105
<210> 82
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> Выступ Fc
<400> 82
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Thr
225
<210> 83
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> Впадина Fc
<400> 83
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Arg Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Thr
225
<210> 84
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<223> Выступ Fc
<400> 84
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Gly Gly Pro Ser
1 5 10 15
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
20 25 30
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
35 40 45
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
50 55 60
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
65 70 75 80
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
85 90 95
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
100 105 110
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
115 120 125
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
130 135 140
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
145 150 155 160
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
165 170 175
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
180 185 190
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
195 200 205
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Thr
210 215 220
<210> 85
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<223> Впадина Fc
<400> 85
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Gly Gly Pro Ser
1 5 10 15
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Arg Ser Arg
20 25 30
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
35 40 45
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
50 55 60
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
65 70 75 80
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
85 90 95
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
100 105 110
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
115 120 125
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
130 135 140
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
145 150 155 160
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
165 170 175
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
180 185 190
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
195 200 205
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Thr
210 215 220
<210> 86
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> Выступ Fc
<400> 86
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly
225
<210> 87
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> Впадина Fc
<400> 87
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Arg Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly
225
<210> 88
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<223> Выступ Fc
<400> 88
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Gly Gly Pro Ser
1 5 10 15
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
20 25 30
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
35 40 45
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
50 55 60
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
65 70 75 80
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
85 90 95
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
100 105 110
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
115 120 125
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
130 135 140
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
145 150 155 160
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
165 170 175
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
180 185 190
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
195 200 205
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
210 215 220
<210> 89
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<223> Впадина Fc
<400> 89
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Gly Gly Pro Ser
1 5 10 15
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Arg Ser Arg
20 25 30
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
35 40 45
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
50 55 60
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
65 70 75 80
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
85 90 95
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
100 105 110
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
115 120 125
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
130 135 140
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
145 150 155 160
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
165 170 175
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
180 185 190
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
195 200 205
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
210 215 220
<210> 90
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> Впадина Fc
<400> 90
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Thr
225
<210> 91
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<223> Впадина Fc
<400> 91
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Gly Gly Pro Ser
1 5 10 15
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
20 25 30
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
35 40 45
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
50 55 60
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
65 70 75 80
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
85 90 95
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
100 105 110
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
115 120 125
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
130 135 140
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
145 150 155 160
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
165 170 175
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
180 185 190
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
195 200 205
Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Thr
210 215 220
<210> 92
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> Впадина Fc
<400> 92
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly
225
<210> 93
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<223> Впадина Fc
<400> 93
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Gly Gly Pro Ser
1 5 10 15
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
20 25 30
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
35 40 45
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
50 55 60
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
65 70 75 80
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
85 90 95
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
100 105 110
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
115 120 125
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
130 135 140
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
145 150 155 160
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
165 170 175
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
180 185 190
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
195 200 205
Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
210 215 220
<210> 94
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> Выступ Fc
<400> 94
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Val
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Thr
225
<210> 95
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<223> Выступ Fc
<400> 95
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Gly Gly Pro Ser
1 5 10 15
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Ser Arg
20 25 30
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
35 40 45
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
50 55 60
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
65 70 75 80
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
85 90 95
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
100 105 110
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
115 120 125
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
130 135 140
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
145 150 155 160
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
165 170 175
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
180 185 190
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Val His Glu Ala
195 200 205
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Thr
210 215 220
<210> 96
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> Выступ Fc
<400> 96
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Val
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly
225
<210> 97
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<223> Выступ Fc
<400> 97
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Gly Gly Pro Ser
1 5 10 15
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Ser Arg
20 25 30
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
35 40 45
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
50 55 60
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
65 70 75 80
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
85 90 95
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
100 105 110
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
115 120 125
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
130 135 140
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
145 150 155 160
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
165 170 175
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
180 185 190
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Val His Glu Ala
195 200 205
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
210 215 220
<210> 98
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> Впадина Fc
<400> 98
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Val
195 200 205
His Glu Ala Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Thr
225
<210> 99
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<223> Впадина Fc
<400> 99
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Gly Gly Pro Ser
1 5 10 15
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Ser Arg
20 25 30
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
35 40 45
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
50 55 60
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
65 70 75 80
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
85 90 95
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
100 105 110
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
115 120 125
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
130 135 140
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
145 150 155 160
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
165 170 175
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
180 185 190
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Val His Glu Ala
195 200 205
Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Thr
210 215 220
<210> 100
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> Впадина Fc
<400> 100
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Val
195 200 205
His Glu Ala Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly
225
<210> 101
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<223> Впадина Fc
<400> 101
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Gly Gly Pro Ser
1 5 10 15
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Ser Arg
20 25 30
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
35 40 45
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
50 55 60
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
65 70 75 80
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
85 90 95
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
100 105 110
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
115 120 125
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
130 135 140
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
145 150 155 160
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
165 170 175
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
180 185 190
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Val His Glu Ala
195 200 205
Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
210 215 220
<210> 102
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> пептидный линкер
<400> 102
Pro Gly Gly Gly Gly
1 5
<210> 103
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> пептидный линкер
<400> 103
Gly Gly Gly Gly
1
<210> 104
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 104
Gly Pro Ala Gly Leu Gly Leu Glu Pro Asp Gly Ser Arg Gln Ala Arg
1 5 10 15
Val Gly
<210> 105
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 105
Gly Gly Ser Gly Gly Gly Gly Ile Glu Pro Asp Ile Gly Gly Ser Gly
1 5 10 15
Gly Ser
<210> 106
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 106
Gly Gly Ser Gly Gly Gly Gly Leu Glu Ala Asp Thr Gly Gly Ser Gly
1 5 10 15
Gly Ser
<210> 107
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 107
Gly Ser Ile Glu Pro Asp Ile Gly Ser
1 5
<210> 108
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 108
Gly Ser Leu Glu Ala Asp Thr Gly Ser
1 5
<210> 109
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 109
Gly Gly Ser Gly Gly Gly Gly Ile Glu Pro Asp Gly Gly Gly Ser Gly
1 5 10 15
Gly Ser
<210> 110
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 110
Gly Gly Ser Gly Gly Gly Gly Ile Glu Pro Asp Val Gly Gly Ser Gly
1 5 10 15
Gly Ser
<210> 111
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 111
Gly Gly Ser Gly Gly Gly Gly Ile Glu Pro Asp Ser Gly Gly Ser Gly
1 5 10 15
Gly Ser
<210> 112
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 112
Gly Gly Ser Gly Gly Gly Gly Ile Glu Pro Asp Thr Gly Gly Ser Gly
1 5 10 15
Gly Ser
<210> 113
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 113
Gly Gly Gly Ser Leu Glu Pro Asp Gly Ser Gly Ser
1 5 10
<210> 114
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 114
Gly Pro Ala Gly Leu Gly Leu Glu Ala Asp Gly Ser Arg Gln Ala Arg
1 5 10 15
Val Gly
<210> 115
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 115
Gly Gly Glu Gly Gly Gly Gly Ser Gly Gly Ser Gly Gly Gly Ser
1 5 10 15
<210> 116
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 116
Gly Ser Ser Ala Gly Ser Glu Ala Gly Gly Ser Gly Gln Ala Gly Val
1 5 10 15
Gly Ser
<210> 117
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 117
Gly Gly Ser Gly Gly Gly Gly Leu Glu Ala Glu Gly Ser Gly Gly Gly
1 5 10 15
Gly Ser
<210> 118
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 118
Gly Gly Ser Gly Gly Gly Gly Ile Glu Pro Asp Pro Gly Gly Ser Gly
1 5 10 15
Gly Ser
<210> 119
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 119
Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5 10 15
Gly Ser
<210> 120
<211> 129
<212> Белок
<213> Искусственная последовательность
<220>
<223> FR альфа sdAb
<400> 120
Gln Leu Gln Leu Gln Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Leu Asp Asn Tyr
20 25 30
Ala Ile Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Cys Ile Ser Ser Ser Asp Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asn Asn Ala Lys Gly Thr Val Tyr
65 70 75 80
Leu Leu Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Glu Leu Val Pro Ala Cys Thr Tyr Ser Asn Gly Arg Gly Pro
100 105 110
Leu Asp Gly Met Asp Tyr Trp Gly Lys Gly Thr Gln Val Thr Val Lys
115 120 125
Pro
<210> 121
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> FR альфа sdAb
<400> 121
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Glu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Ile Phe Ser Ile Asp
20 25 30
Ala Thr Ala Trp Tyr Arg Gln Ala Pro Gly Lys Gln Arg Glu Leu Val
35 40 45
Ala Ile Ile Thr Ser Ser Gly Ser Thr Asn Tyr Pro Glu Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Asn
85 90 95
Ala Ile Thr Arg Tyr Gly Gly Ser Thr Tyr Asp Phe Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Lys Pro
115
<210> 122
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> FR альфа sdAb
<400> 122
Glu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Glu
1 5 10 15
Thr Phe Gly Val Val Phe Thr Leu Gly Trp Tyr Arg Gln Ala Pro Gly
20 25 30
Lys Gly Arg Glu Phe Val Ala Arg Val Thr Gly Thr Asp Thr Val Asp
35 40 45
Tyr Ala Glu Ser Val Lys Gly Arg Phe Thr Ile Ser Ser Asp Phe Ala
50 55 60
Arg Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
65 70 75 80
Ala Val Tyr Tyr Cys Asn Thr Gly Ala Tyr Trp Gly Gln Gly Thr Leu
85 90 95
Val Thr Val Lys Pro
100
<210> 123
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> cMET sdAb
<400> 123
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Leu Asp Tyr Tyr
20 25 30
Ala Ile Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Leu Cys Ile Asp Ala Ser Asp Asp Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Gly Val Tyr Tyr Cys
85 90 95
Ala Thr Pro Ile Gly Leu Ser Ser Ser Cys Leu Leu Glu Tyr Asp Tyr
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Lys Pro
115 120 125
<210> 124
<211> 245
<212> Белок
<213> Искусственная последовательность
<220>
<223> B7H3 scFv
<400> 124
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Asp Ser Ser Ala Ile Tyr Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Asp Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Arg Glu Asn Ile Tyr Tyr Gly Ser Arg Leu Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Leu Thr Gln
130 135 140
Ser Pro Ser Phe Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr
145 150 155 160
Cys Lys Ala Ser Gln Asn Val Asp Thr Asn Val Ala Trp Tyr Gln Gln
165 170 175
Lys Pro Gly Lys Ala Pro Lys Ala Leu Ile Tyr Ser Ala Ser Tyr Arg
180 185 190
Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
195 200 205
Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr
210 215 220
Tyr Cys Gln Gln Tyr Asn Asn Tyr Pro Phe Thr Phe Gly Gln Gly Thr
225 230 235 240
Lys Leu Glu Ile Lys
245
<210> 125
<211> 247
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD20 scFv
<400> 125
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Ser
20 25 30
Trp Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Phe Pro Gly Asp Gly Asp Thr Asp Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Val Phe Asp Gly Tyr Trp Leu Val Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
115 120 125
Gly Thr Gly Gly Gly Gly Ser Asp Ile Val Met Thr Gln Thr Pro Leu
130 135 140
Ser Leu Pro Val Thr Pro Gly Glu Pro Ala Ser Ile Ser Cys Arg Ser
145 150 155 160
Ser Lys Ser Leu Leu His Ser Asn Gly Ile Thr Tyr Leu Tyr Trp Tyr
165 170 175
Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile Tyr Gln Met Ser
180 185 190
Asn Leu Val Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly
195 200 205
Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly
210 215 220
Val Tyr Tyr Cys Ala Gln Asn Leu Glu Leu Pro Tyr Thr Phe Gly Gly
225 230 235 240
Gly Thr Lys Val Glu Ile Lys
245
<210> 126
<211> 241
<212> Белок
<213> Искусственная последовательность
<220>
<223> DLL3 scFv
<400> 126
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Val Tyr Tyr Ser Gly Thr Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Ser Ile Ala Val Thr Gly Phe Tyr Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu
130 135 140
Ser Leu Ser Pro Gly Glu Arg Val Thr Leu Ser Cys Arg Ala Ser Gln
145 150 155 160
Arg Val Asn Asn Asn Tyr Leu Ala Trp Tyr Gln Gln Arg Pro Gly Gln
165 170 175
Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile
180 185 190
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
195 200 205
Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln
210 215 220
Tyr Asp Arg Ser Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
225 230 235 240
Lys
<210> 127
<211> 225
<212> Белок
<213> Искусственная последовательность
<220>
<223> B7H3 Fd
<400> 127
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Asp Ser Ser Ala Ile Tyr Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Asp Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Arg Glu Asn Ile Tyr Tyr Gly Ser Arg Leu Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys
225
<210> 128
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> B7H3 LC
<400> 128
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asn Val Asp Thr Asn
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Ala Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Asn Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 129
<211> 225
<212> Белок
<213> Искусственная последовательность
<220>
<223> 5T4 Fd
<400> 129
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Lys Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Gln Ser Met
65 70 75 80
Leu Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Arg Gln Trp Asp Tyr Asp Val Arg Ala Met Asn Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys
225
<210> 130
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> 5T4 LC
<400> 130
Asp Ile Val Met Thr Gln Ser His Ile Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asp Val Asp Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg Leu Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Ser Ser Tyr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 131
<211> 222
<212> Белок
<213> Искусственная последовательность
<220>
<223> gpNMB Fd
<400> 131
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Phe
20 25 30
Asn Tyr Tyr Trp Ser Trp Ile Arg His His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Tyr Ser Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Thr Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Gly Tyr Asn Trp Asn Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210> 132
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> gpNMB LC
<400> 132
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asp Asn Asn
20 25 30
Leu Val Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Asn Asn Trp Pro Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 133
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<223> DLL3 Fd
<400> 133
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Ala Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Ser
65 70 75 80
Leu Gln Ile Ile Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Ile Gly Asp Ser Ser Pro Ser Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210> 134
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> DLL3 LC
<400> 134
Ser Ile Val Met Thr Gln Thr Pro Lys Phe Leu Leu Val Ser Ala Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Ser Val Ser Asn Asp
20 25 30
Val Val Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Ala Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Ser Phe Thr Ile Ser Thr Val Gln Ala
65 70 75 80
Glu Asp Leu Ala Val Tyr Phe Cys Gln Gln Asp Tyr Thr Ser Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Arg Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 135
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 135
Gly Gly Gly Gly Gly Ser
1 5
<210> 136
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 136
Ile Glu Pro Asp Ile
1 5
<210> 137
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 137
Leu Glu Ala Asp Thr
1 5
<210> 138
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 138
Ile Glu Pro Asp Gly
1 5
<210> 139
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 139
Ile Glu Pro Asp Val
1 5
<210> 140
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 140
Ile Glu Pro Asp Ser
1 5
<210> 141
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 141
Ile Glu Pro Asp Thr
1 5
<210> 142
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 142
Leu Glu Pro Asp
1
<210> 143
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 143
Leu Glu Ala Glu
1
<210> 144
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 144
Ile Glu Pro Asp Pro
1 5
<210> 145
<211> 274
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вторая полипептидная цепь B7-H3 x CD3 биспецифичного диатела DART-A
<400> 145
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Trp Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Ser Phe Gly Met His Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Tyr Ile Ser Ser Asp Ser Ser Ala Ile
165 170 175
Tyr Tyr Ala Asp Thr Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
180 185 190
Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Asp Glu Asp
195 200 205
Thr Ala Val Tyr Tyr Cys Gly Arg Gly Arg Glu Asn Ile Tyr Tyr Gly
210 215 220
Ser Arg Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
225 230 235 240
Gly Gly Cys Gly Gly Gly Lys Val Ala Ala Leu Lys Glu Lys Val Ala
245 250 255
Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu
260 265 270
Lys Glu
<210> 146
<211> 227
<212> Белок
<213> Искусственная последовательность
<220>
<223> Третья полипептидная цепь B7-H3 x CD3 биспецифичного диатела DART-A
<400> 146
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 147
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 147
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
1 5 10 15
Gly Ser
<210> 148
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 148
Thr Gly Gly Ser Gly Gly Gly Gly Ile Glu Pro Asp Ile Gly Gly Ser
1 5 10 15
Gly Gly Ser
<210> 149
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 149
Gly Gly Gly Gly Ser
1 5
<210> 150
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер консенсус
<220>
<221> ВАРИАНТ
<222> (1)..(1)
<223> X1= I, L, Y, M, F, V, или A
<220>
<221> ВАРИАНТ
<222> (2)..(2)
<223> X2 = A, G, S, V, E, D, Q, N, или Y
<220>
<221> ВАРИАНТ
<222> (3)..(3)
<223> X3 = H, P, A, V, G, S, или T
<220>
<221> ВАРИАНТ
<222> (4)..(4)
<223> X4 = D или E
<220>
<221> ВАРИАНТ
<222> (5)..(5)
<223> X5 = I, L, Y, M, F, V, T, S, G или A
<400> 150
Xaa Xaa Xaa Xaa Xaa
1 5
<210> 151
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер консенсус
<220>
<221> ВАРИАНТ
<222> (1)..(1)
<223> X1 = I или L
<220>
<221> ВАРИАНТ
<222> (3)..(3)
<223> X3 = P или A
<220>
<221> ВАРИАНТ
<222> (5)..(5)
<223> X5 = I, V, T, S, или G
<400> 151
Xaa Glu Xaa Asp Xaa
1 5
<210> 152
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 152
Leu Glu Pro Asp Gly
1 5
<210> 153
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 153
Leu Glu Ala Asp Gly
1 5
<210> 154
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер консенсус
<220>
<221> ВАРИАНТ
<222> (1)..(1)
<223> X1 = любая аминокислота
<220>
<221> ВАРИАНТ
<222> (5)..(5)
<223> X5 = A или V
<400> 154
Xaa Gln Ala Arg Xaa
1 5
<210> 155
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер консенсус
<220>
<221> ВАРИАНТ
<222> (5)..(5)
<223> X5 = A или V
<400> 155
Arg Gln Ala Arg Xaa
1 5
<210> 156
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 156
Arg Gln Ala Arg Val
1 5
<210> 157
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер консенсус
<220>
<221> ВАРИАНТ
<222> (1)..(1)
<223> X1 = P, V или A
<220>
<221> ВАРИАНТ
<222> (2)..(2)
<223> X2 = Q или D
<220>
<221> ВАРИАНТ
<222> (3)..(3)
<223> X3 = A или N
<220>
<221> ВАРИАНТ
<222> (4)..(4)
<223> X4 = L, I или M
<400> 157
Xaa Xaa Xaa Xaa
1
<210> 158
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер консенсус
<220>
<221> ВАРИАНТ
<222> (2)..(2)
<223> X2 = Q или D
<220>
<221> ВАРИАНТ
<222> (3)..(3)
<223> X3 = A или N
<220>
<221> ВАРИАНТ
<222> (4)..(4)
<223> X4 = L или I
<400> 158
Pro Xaa Xaa Xaa
1
<210> 159
<211> 22
<212> Белок
<213> Искусственная последовательность
<220>
<223> P2A
<400> 159
Gly Ser Gly Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val
1 5 10 15
Glu Glu Asn Pro Gly Pro
20
<210> 160
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> P2A
<400> 160
Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val Glu Glu Asn
1 5 10 15
Pro Gly Pro
<210> 161
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> E2A
<400> 161
Gln Cys Thr Asn Tyr Ala Leu Leu Lys Leu Ala Gly Asp Val Glu Ser
1 5 10 15
Asn Pro Gly Pro
20
<210> 162
<211> 22
<212> Белок
<213> Искусственная последовательность
<220>
<223> F2A
<400> 162
Val Lys Gln Thr Leu Asn Phe Asp Leu Leu Lys Leu Ala Gly Asp Val
1 5 10 15
Glu Ser Asn Pro Gly Pro
20
<210> 163
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> T2A
<400> 163
Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro
1 5 10 15
Gly Pro
<210> 164
<211> 24
<212> Белок
<213> Искусственная последовательность
<220>
<223> T2A
<400> 164
Leu Glu Gly Gly Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp
1 5 10 15
Val Glu Glu Asn Pro Gly Pro Arg
20
<210> 165
<211> 66
<212> DNA
<213> Искусственная последовательность
<220>
<223> P2A
<400> 165
ggatctggag caacaaactt ctcactactc aaacaagcag gtgacgtgga ggagaatccc 60
ggaccc 66
<210> 166
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 166
Gly Ser Pro Ala Gly Leu Glu Ala Asp Gly Ser Arg Gln Ala Arg Val
1 5 10 15
Gly Ser
<210> 167
<211> 122
<212> Белок
<213> Mus musculus
<220>
<221> Прочие_особенности
<223> анти-5T4 VH
<220>
<221> Прочие_особенности
<222> (97)..(97)
<223> Xaa может быть любой природной аминокислотой
<400> 167
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Lys Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Gln Ser Met
65 70 75 80
Leu Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Xaa Cys Val Arg Gln Trp Asp Tyr Asp Val Arg Ala Met Asn Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 168
<211> 107
<212> Белок
<213> Mus musculus
<220>
<221> Прочие_особенности
<223> анти-5T4 VL
<400> 168
Asp Ile Val Met Thr Gln Ser His Ile Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asp Val Asp Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg Leu Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Ser Ser Tyr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 169
<211> 504
<212> Белок
<213> Искусственная последовательность
<220>
<223> Первая полипептидная цепь B7-H3 x CD3 биспецифичного диатела DART-A
<400> 169
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asn Val Asp Thr Asn
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Ala Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Asn Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
115 120 125
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
130 135 140
Ser Thr Tyr Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
145 150 155 160
Glu Trp Val Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr
165 170 175
Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser
180 185 190
Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr
195 200 205
Ala Val Tyr Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val
210 215 220
Ser Trp Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
225 230 235 240
Gly Gly Cys Gly Gly Gly Glu Val Ala Ala Leu Glu Lys Glu Val Ala
245 250 255
Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu
260 265 270
Glu Lys Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
275 280 285
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
290 295 300
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
305 310 315 320
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
325 330 335
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
340 345 350
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
355 360 365
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
370 375 380
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
385 390 395 400
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
405 410 415
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
420 425 430
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
435 440 445
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
450 455 460
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
465 470 475 480
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
485 490 495
Ser Leu Ser Leu Ser Pro Gly Lys
500
<210> 170
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 170
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 171
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> Повтор
<222> (4)..(6)
<223> повторяется 0 - 10 раз
<400> 171
Gly Gly Ser Gly Gly Ser
1 5
<210> 172
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> повтор
<222> (1)..(6)
<223> повторяется 1 - 4 раза
<400> 172
Gly Gly Gly Gly Gly Ser
1 5
<210> 173
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> повтор
<222> (1)..(5)
<223> повторяется 1 - 5 раз
<400> 173
Gly Gly Gly Gly Ser
1 5
<210> 174
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> повтор
<222> (1)..(1)
<223> повторяется 1 - 5 раз
<220>
<221> ВАРИАНТ
<222> (2)..(2)
<223> X2= A, V, L, I, M,F, W, P, G, S, T, C, Y, N,Q, K, R, H, D, или E
<220>
<221> повтор
<222> (3)..(3)
<223> повторяется 1 - 5 раз
<220>
<221> ВАРИАНТ
<222> (4)..(4)
<223> X4 = A, V, L, I, M,F, W, P, G, S, T, C, Y, N,Q, K, R, H, D, или E
<220>
<221> повтор
<222> (5)..(5)
<223> повторяется 1 - 5 раз
<400> 174
Gly Xaa Gly Xaa Gly
1 5
<210> 175
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> ВАРИАНТ
<222> (4)..(4)
<223> X4 = A, V, L, I, M,F, W, P, G, S, T, C, Y, N,Q, K, R, H, D, или E
<220>
<221> ВАРИАНТ
<222> (8)..(8)
<223> X8 = A, V, L, I, M,F, W, P, G, S, T, C, Y, N,Q, K, R, H, D, или E
<400> 175
Gly Gly Gly Xaa Gly Gly Gly Xaa Gly Gly Gly
1 5 10
<210> 176
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 176
Ala Thr Thr Thr Gly Ser Ser Pro Gly Pro Thr
1 5 10
<210> 177
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 177
Gly Gly Gly Gly Gly Cys Gly Gly Gly Gly Gly
1 5 10
<210> 178
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> повтор
<222> (1)..(5)
<223> повторяется 2 - 20 раз
<400> 178
Glu Ala Ala Ala Lys
1 5
<210> 179
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> повтор
<222> (3)..(4)
<223> повторяется 2 - 20 раз
<400> 179
Ala Ser Ala Pro Gly Thr
1 5
<210> 180
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> повтор
<222> (3)..(7)
<223> повторяется 2 - 20 раз
<400> 180
Ala Ser Glu Ala Ala Ala Lys Gly Thr
1 5
<210> 181
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> повтор
<222> (1)..(5)
<223> повторяется 2 - 20 раз
<400> 181
Gly Gly Gly Gly Ala
1 5
<210> 182
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> повтор
<222> (1)..(5)
<223> повторяется 2 - 20 раз
<400> 182
Pro Gly Gly Gly Ser
1 5
<210> 183
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> повтор
<222> (1)..(5)
<223> повторяется 2 - 20 раз
<400> 183
Ala Gly Gly Gly Ser
1 5
<210> 184
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> повтор
<222> (4)..(17)
<223> повторяется 2 - 20 раз
<400> 184
Gly Gly Ser Glu Gly Lys Ser Ser Gly Ser Gly Ser Glu Ser Lys Ser
1 5 10 15
Thr Gly Gly Ser
20
<210> 185
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> повтор
<222> (1)..(5)
<223> повторяется 1 - 9 раз
<400> 185
Ser Ser Ser Ser Gly
1 5
<210> 186
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 186
Ser Ser Ser Ala Ser Ala Ser Ser Ala
1 5
<210> 187
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 187
Gly Ser Pro Gly Ser Pro Gly
1 5
<210> 188
<211> 241
<212> Белок
<213> Искусственная последовательность
<220>
<223> DLL3 scFv
<400> 188
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Val Tyr Tyr Ser Gly Thr Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Ser Ile Ala Val Thr Gly Phe Tyr Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu
130 135 140
Ser Leu Ser Pro Gly Glu Arg Val Thr Leu Ser Cys Arg Ala Ser Gln
145 150 155 160
Arg Val Asn Asn Asn Tyr Leu Ala Trp Tyr Gln Gln Arg Pro Gly Gln
165 170 175
Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile
180 185 190
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
195 200 205
Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln
210 215 220
Tyr Asp Arg Ser Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
225 230 235 240
Lys
<210> 189
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD20 VH
<400> 189
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Ser
20 25 30
Trp Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Phe Pro Gly Asp Gly Asp Thr Asp Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Val Phe Asp Gly Tyr Trp Leu Val Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 190
<211> 115
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD20 VL
<400> 190
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Thr Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Gln Met Ser Asn Leu Val Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val
115
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2788531C2 |
| АКТИВИРУЕМЫЕ ПРОТЕАЗОЙ СВЯЗЫВАЮЩИЕ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИЕ МОЛЕКУЛЫ | 2017 |
|
RU2830288C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2022 |
|
RU2824285C2 |
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2826991C2 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, ОТНОСЯЩИЕСЯ К НИМ СПОСОБЫ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2822200C2 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| АНТИТЕЛА К CD38 И КОМБИНАЦИИ С АНТИТЕЛАМИ К CD3 И CD28 | 2018 |
|
RU2812910C2 |
| ТРИСПЕЦИФИЧЕСКИЕ И/ИЛИ ТРИВАЛЕНТНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ С ПРИМЕНЕНИЕМ КРОССОВЕРНОГО ФОРМАТА С ДВОЙНЫМ ВАРИАБЕЛЬНЫМ ДОМЕНОМ (CODV) ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИИ, ВЫЗВАННОЙ HIV | 2020 |
|
RU2820164C2 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, СВЯЗЫВАЮЩИЕ CD38, CD28 И CD3, А ТАКЖЕ СПОСОБЫ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ВИРУСНОЙ ИНФЕКЦИИ | 2019 |
|
RU2820351C2 |
| ПОЛИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, НАЦЕЛЕННЫЕ НА CAIX, ANO1, МЕЗОТЕЛИН, TROP2, СEA ИЛИ КЛАУДИН-18.2 | 2018 |
|
RU2792671C2 |
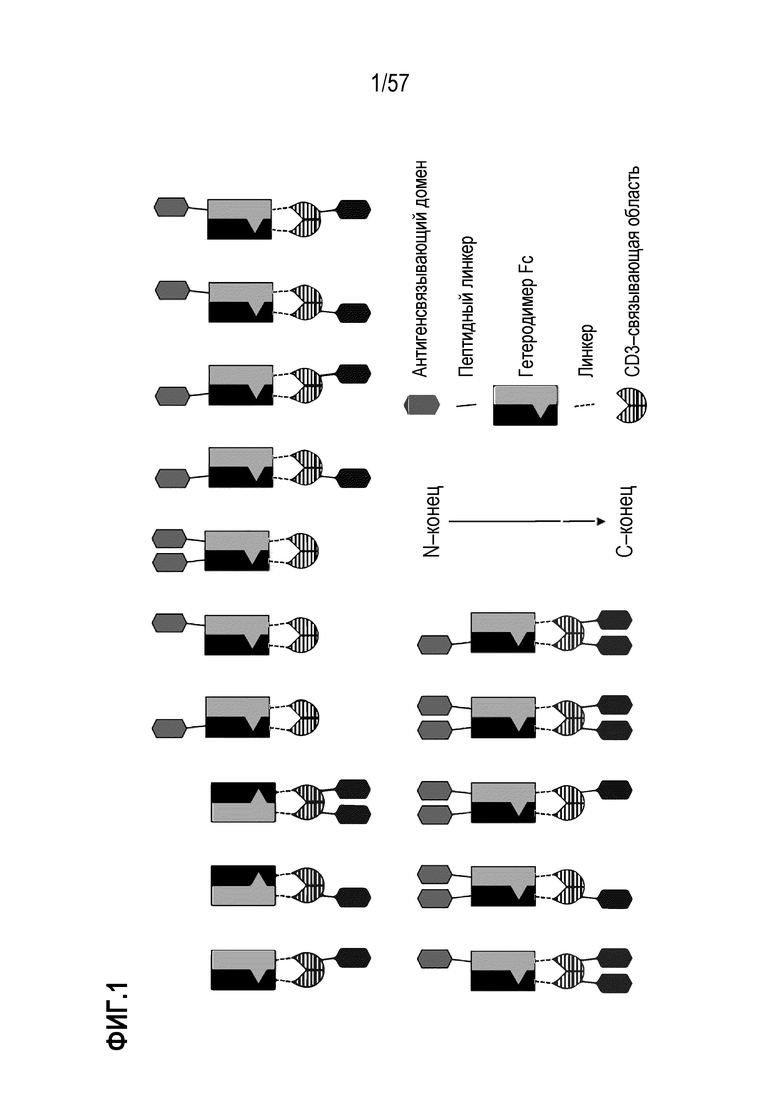
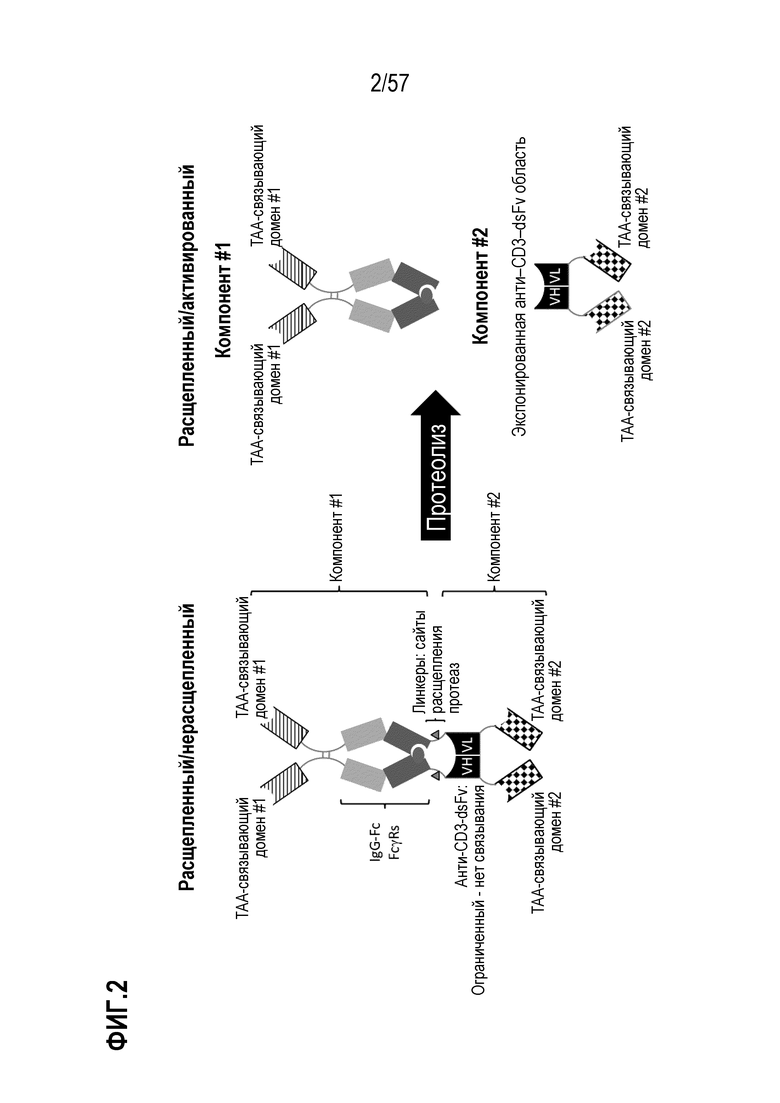
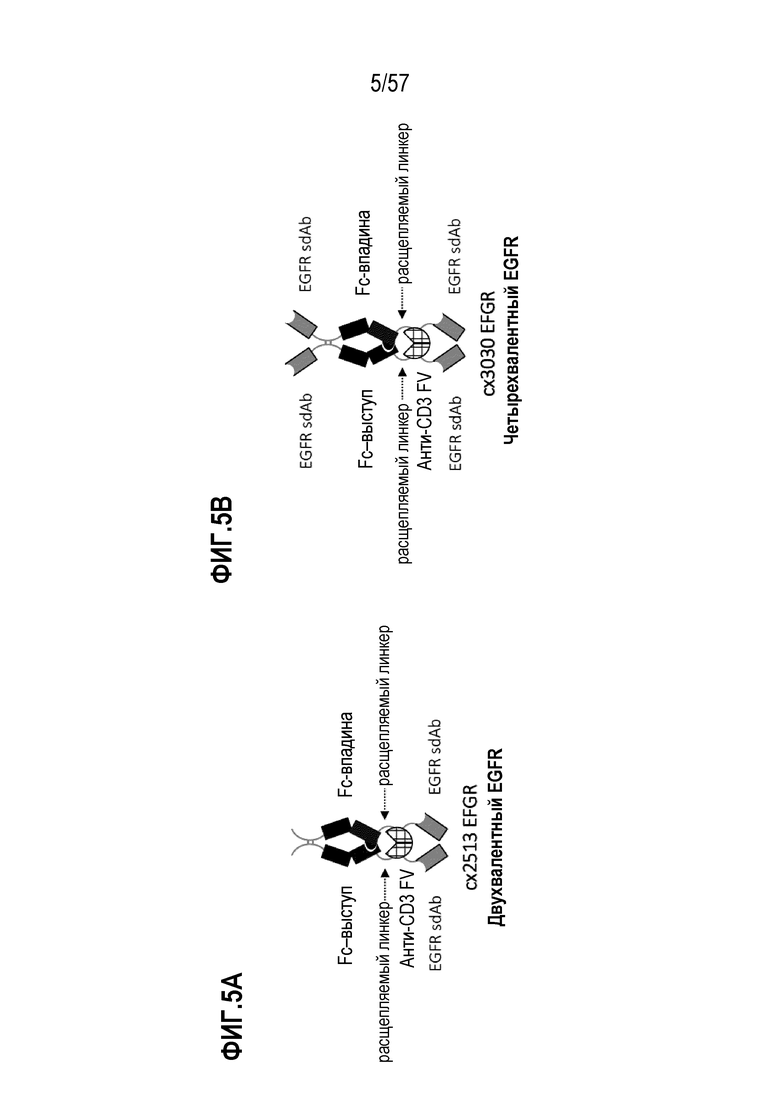
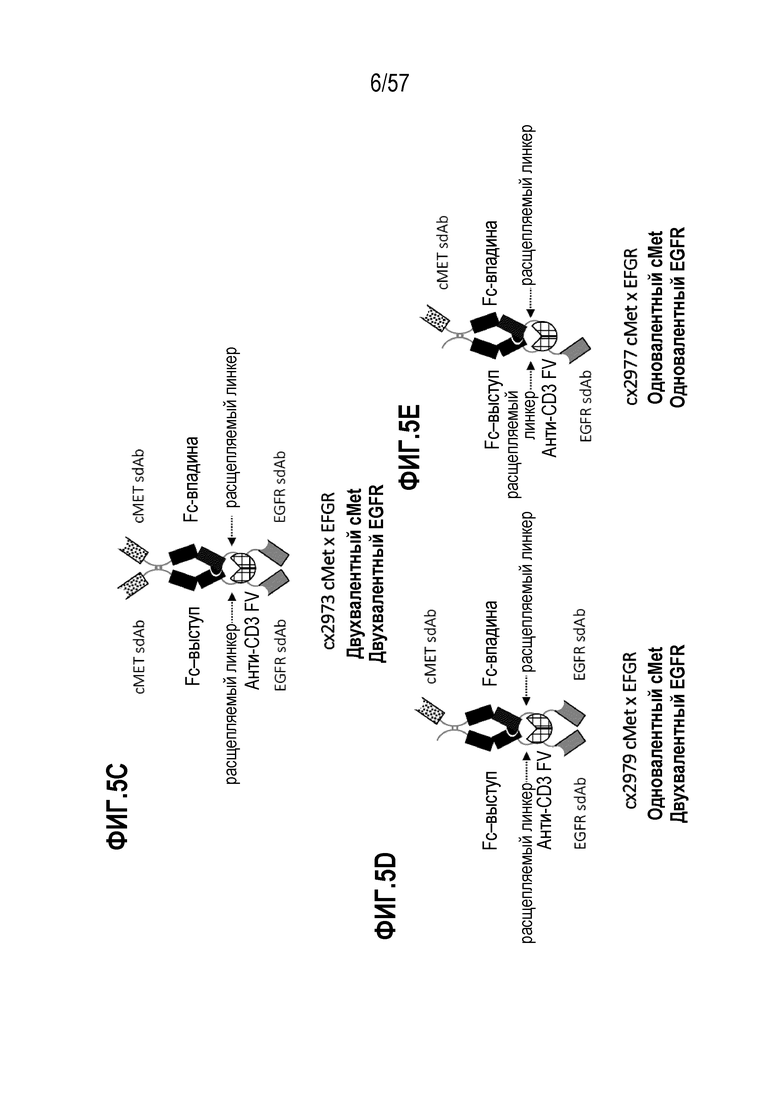
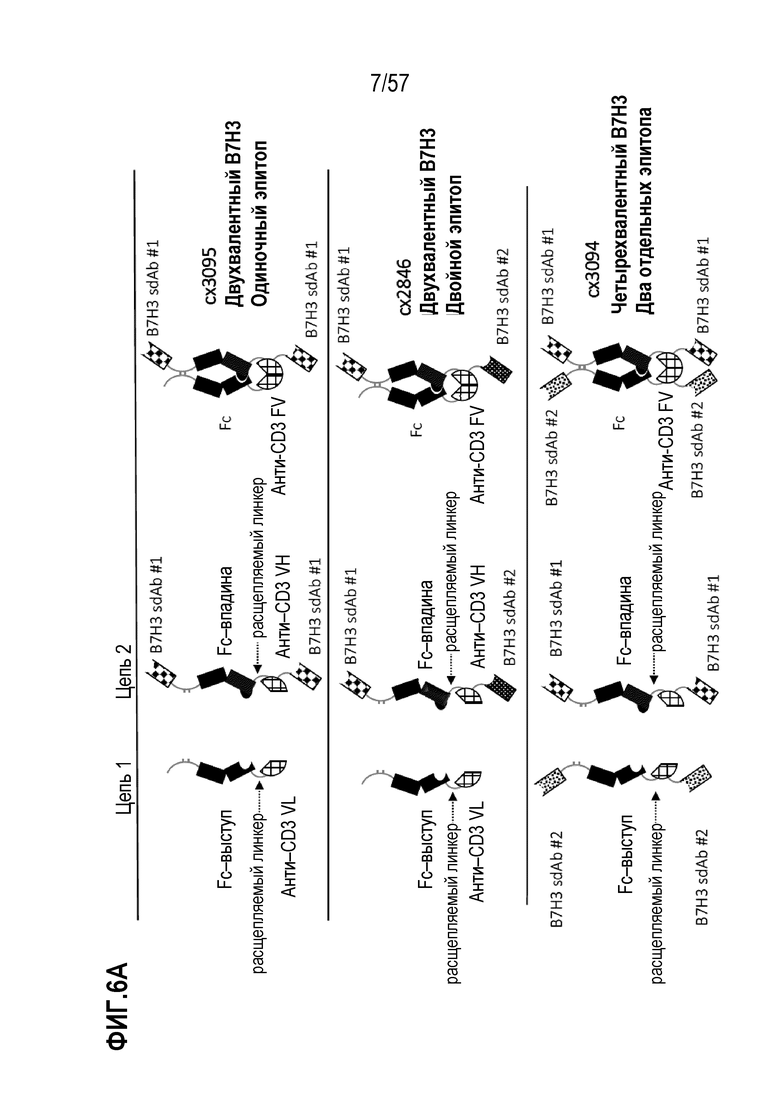
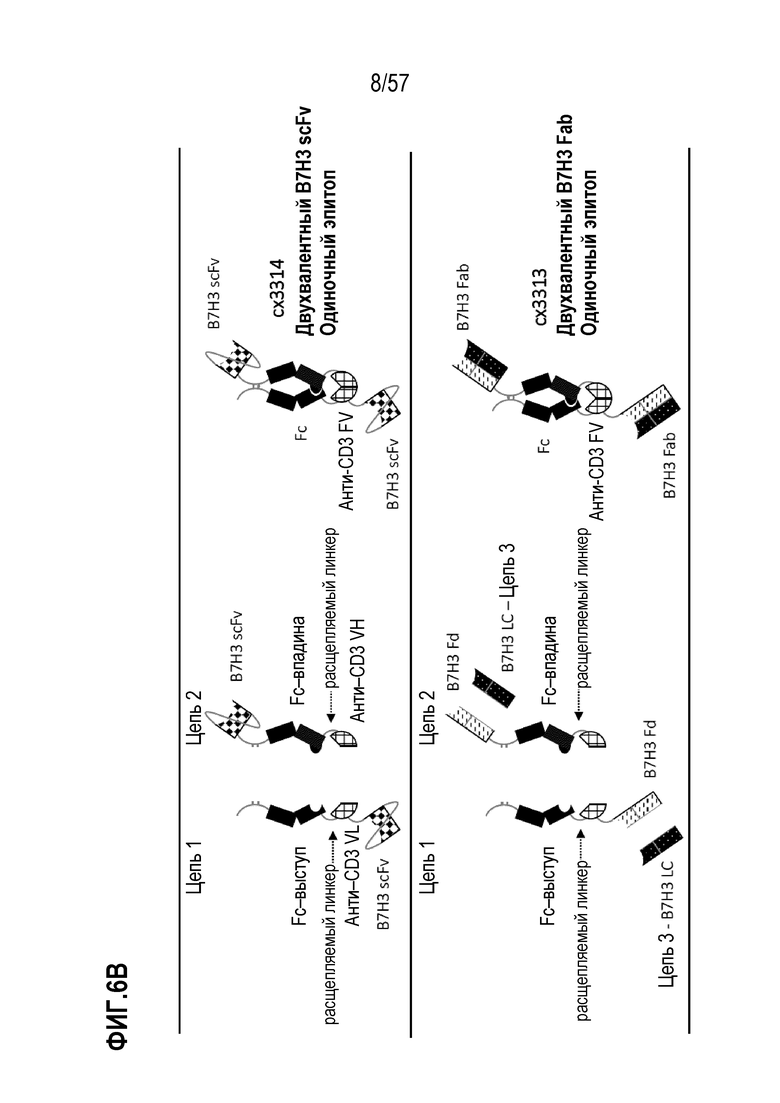
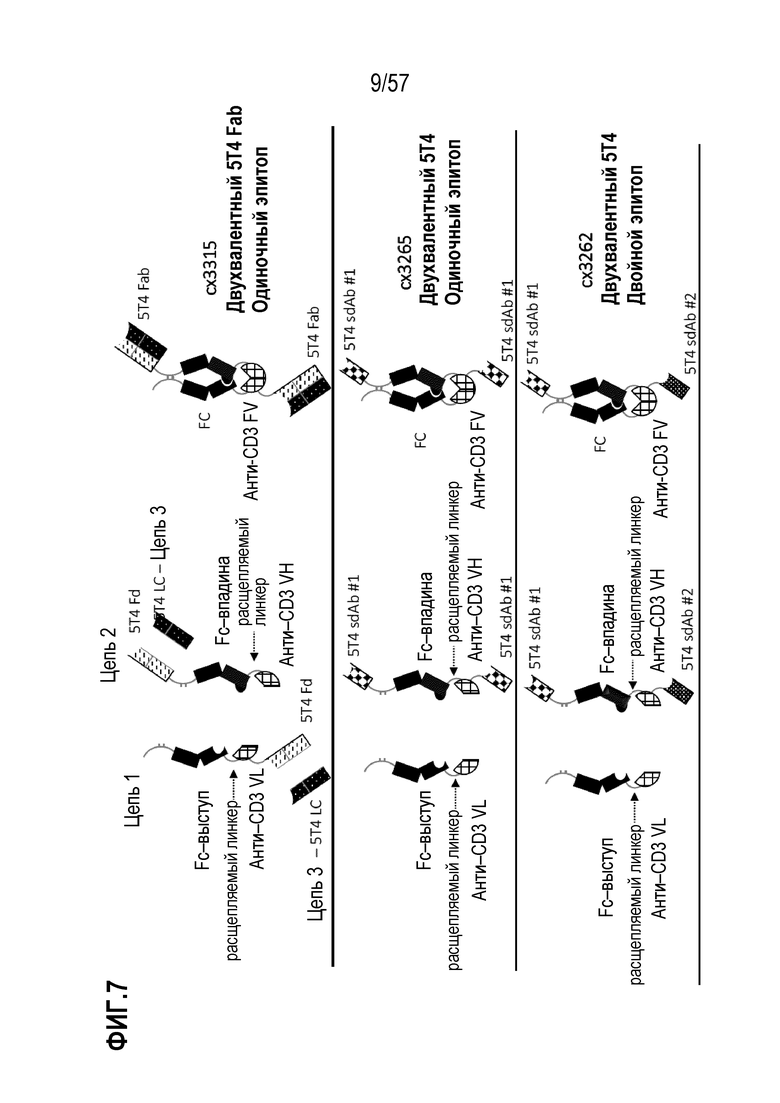
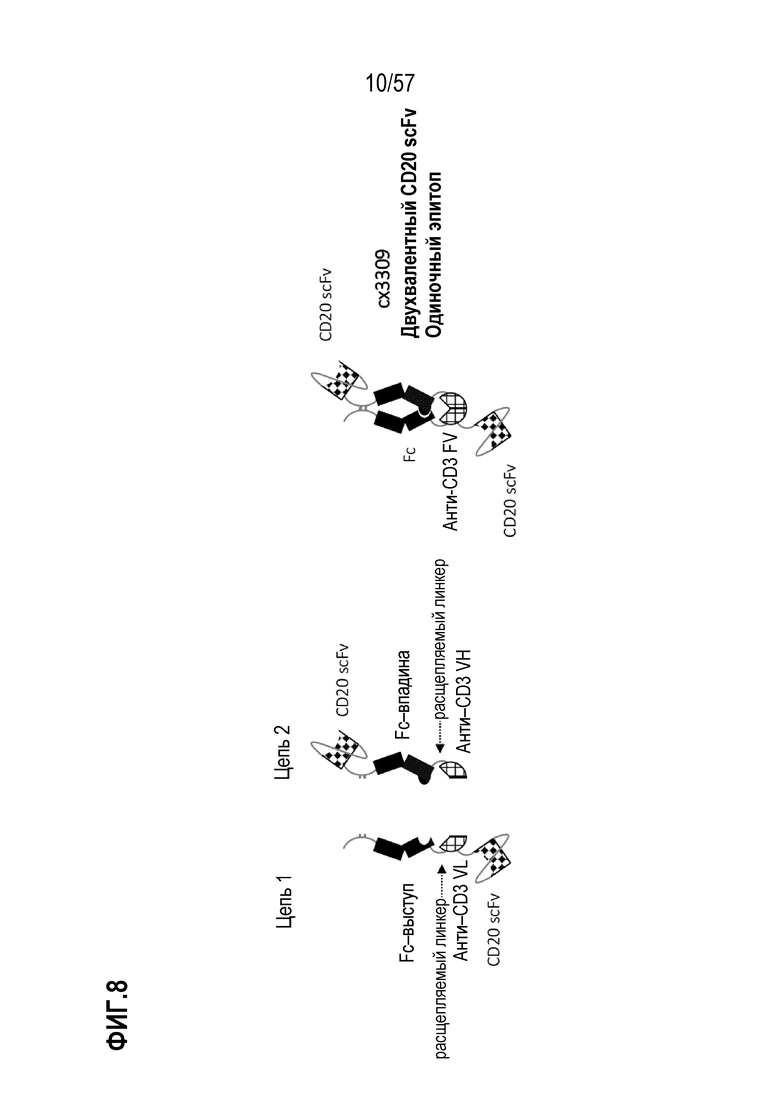
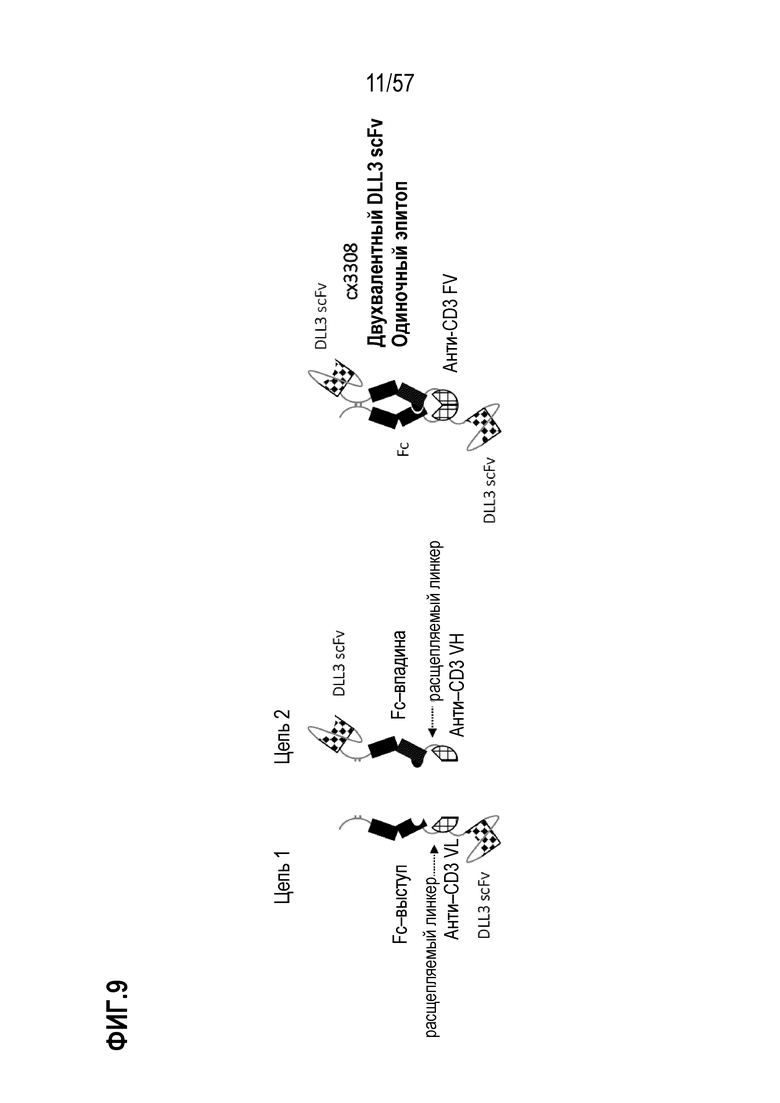

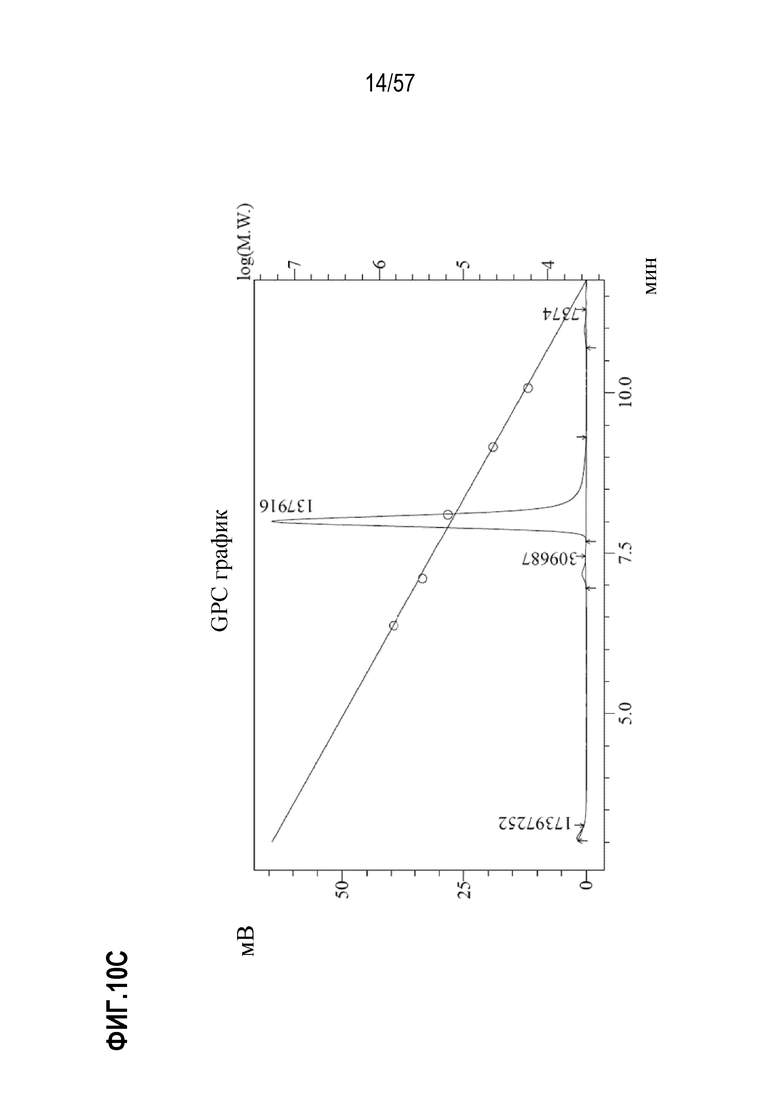
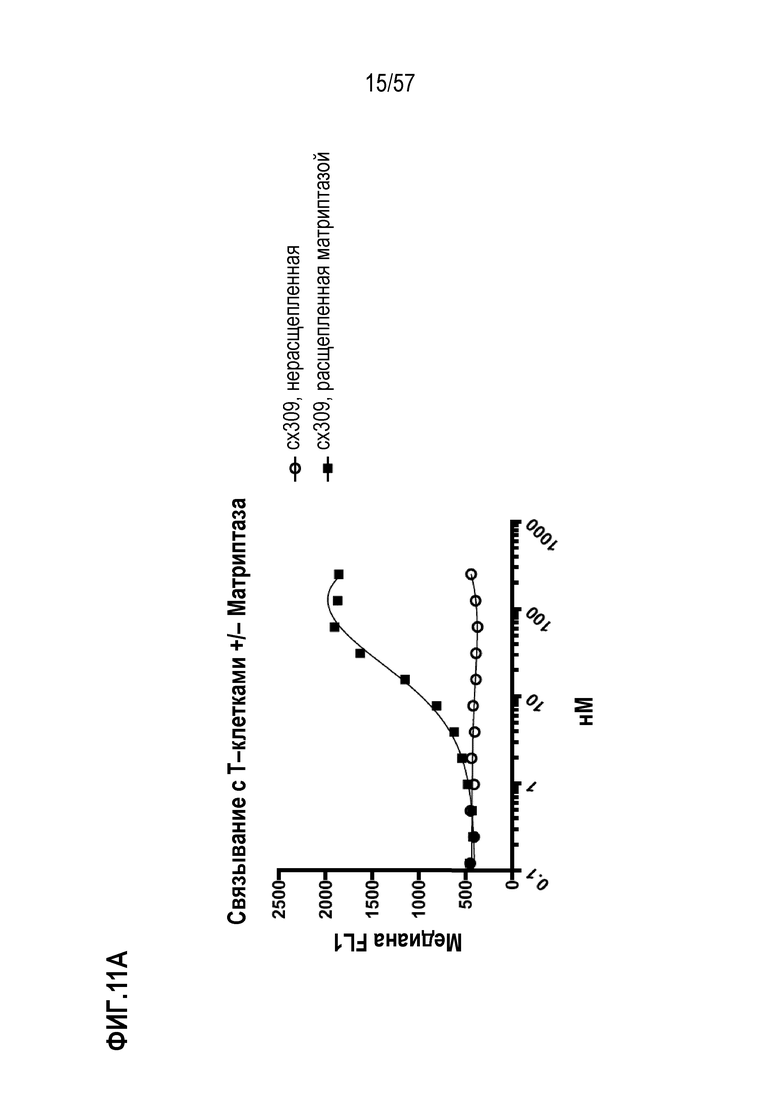
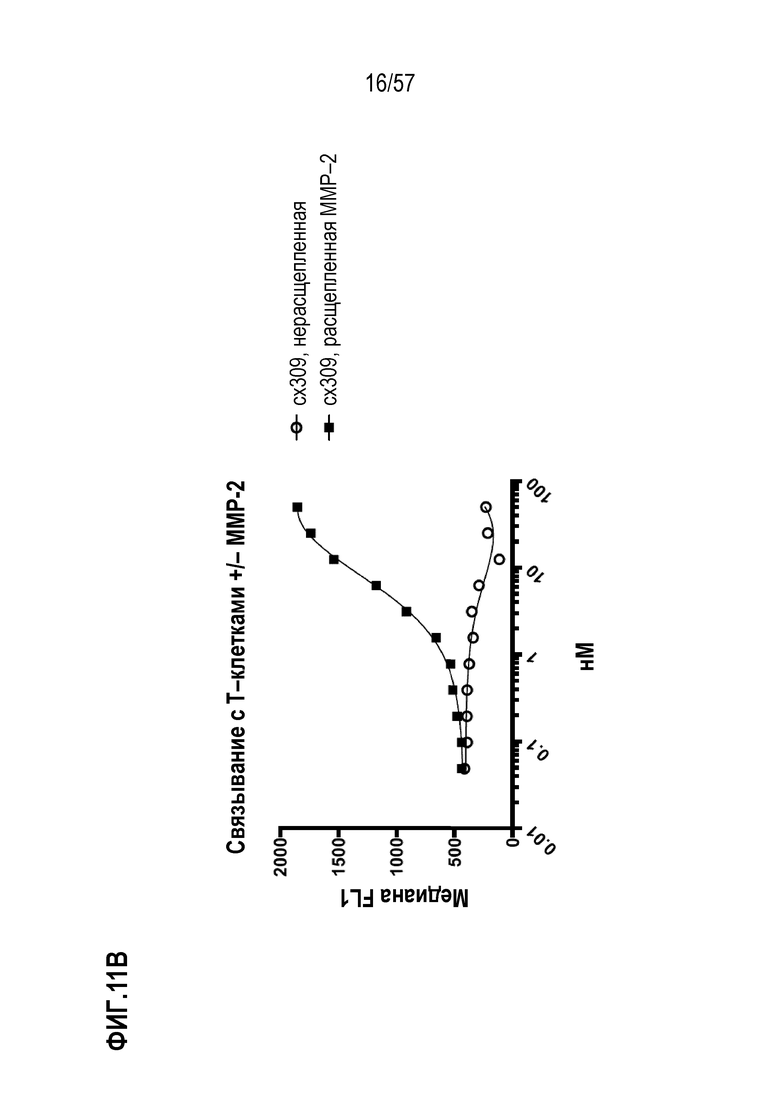
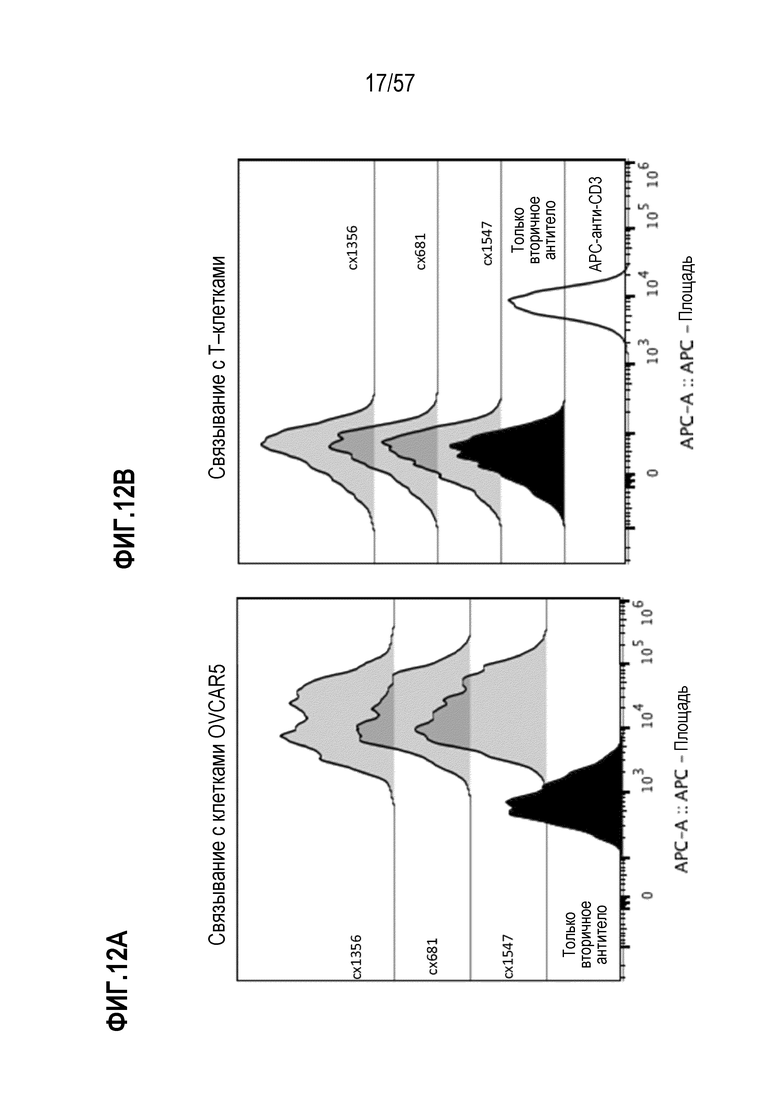
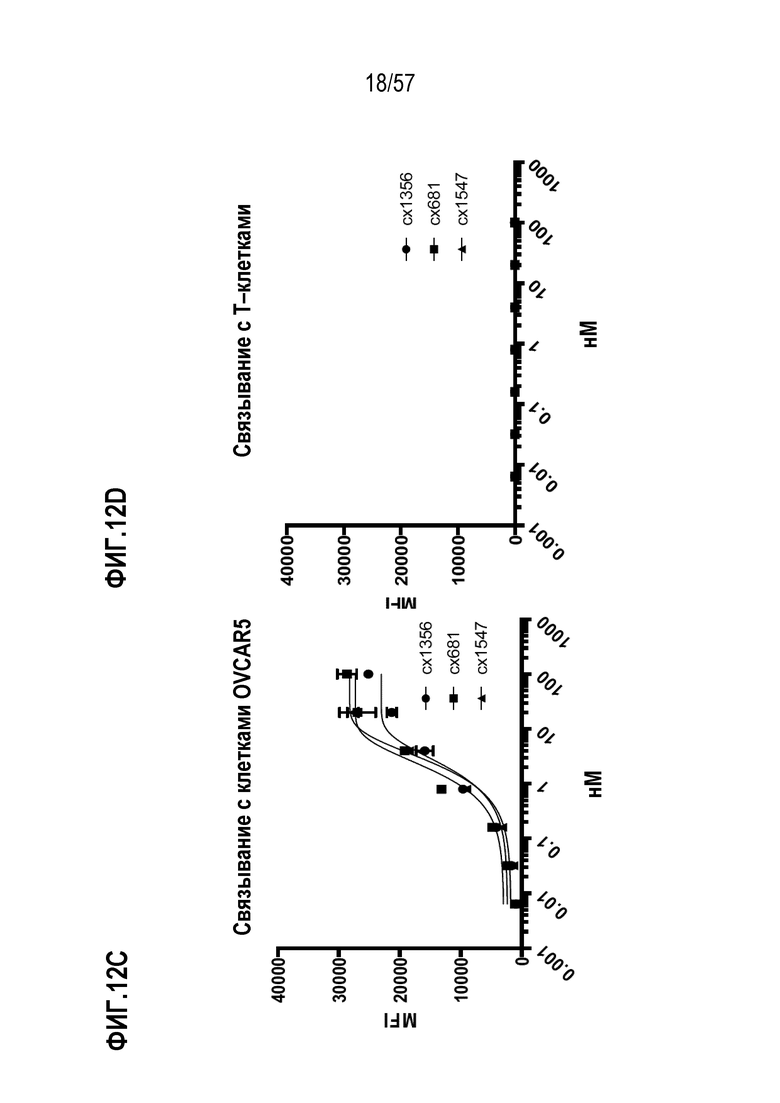
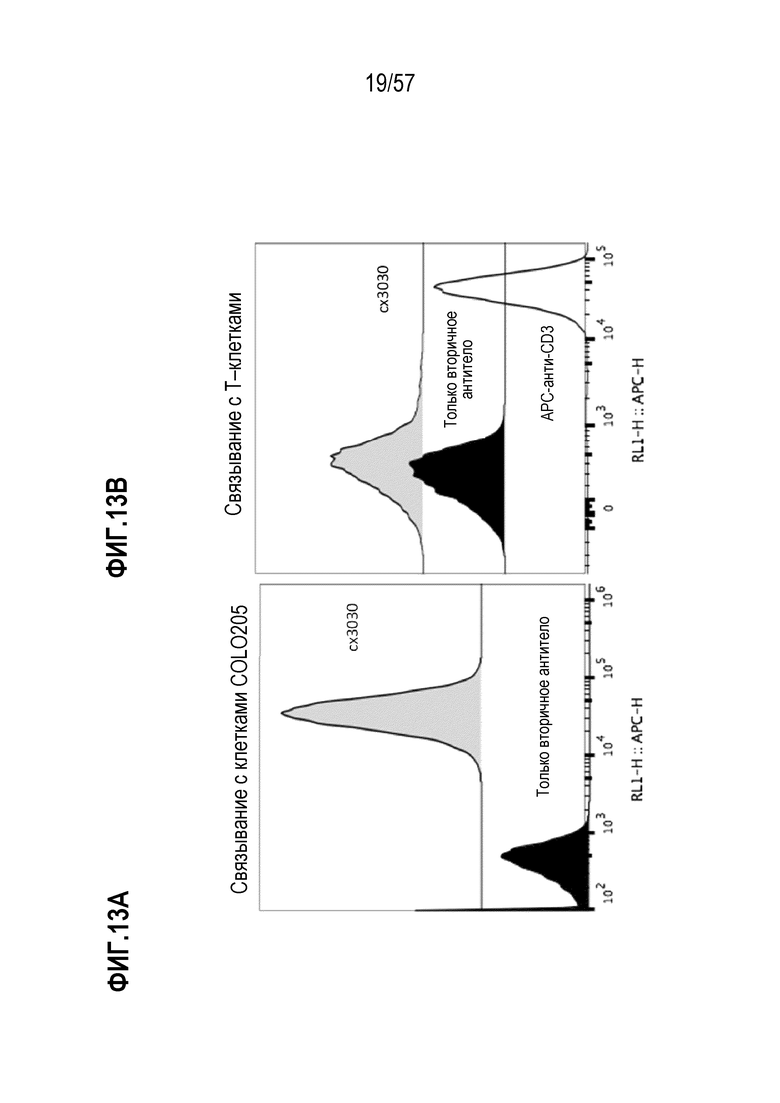
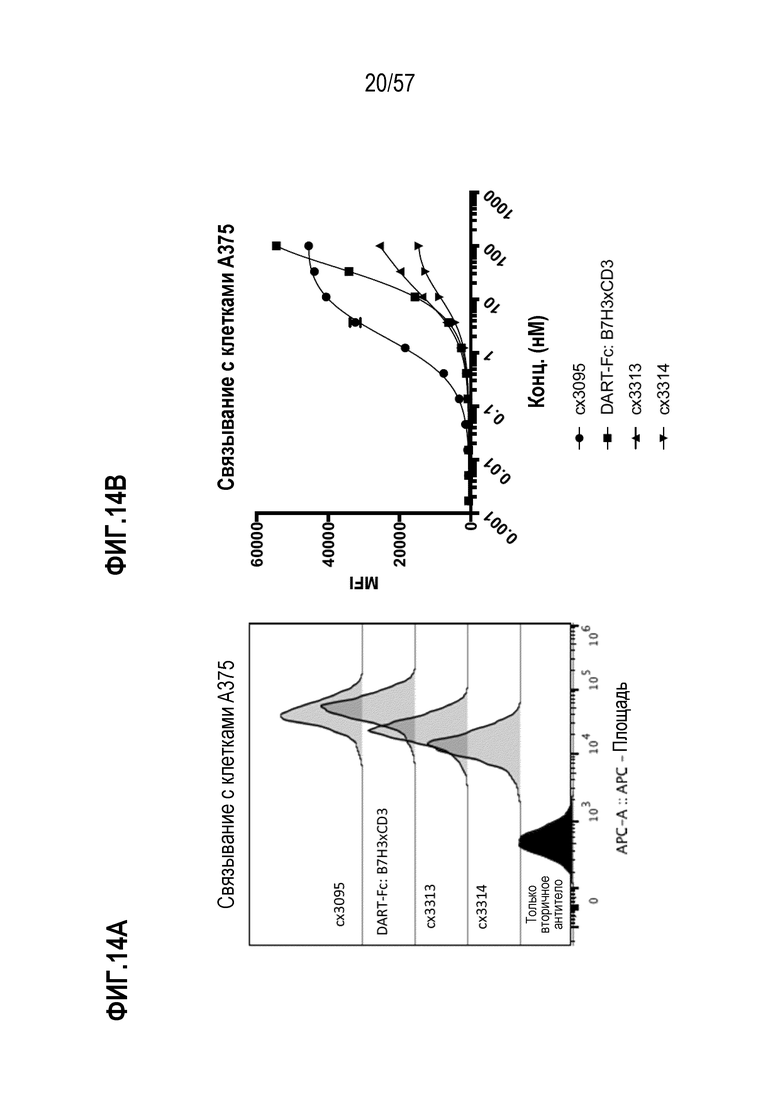
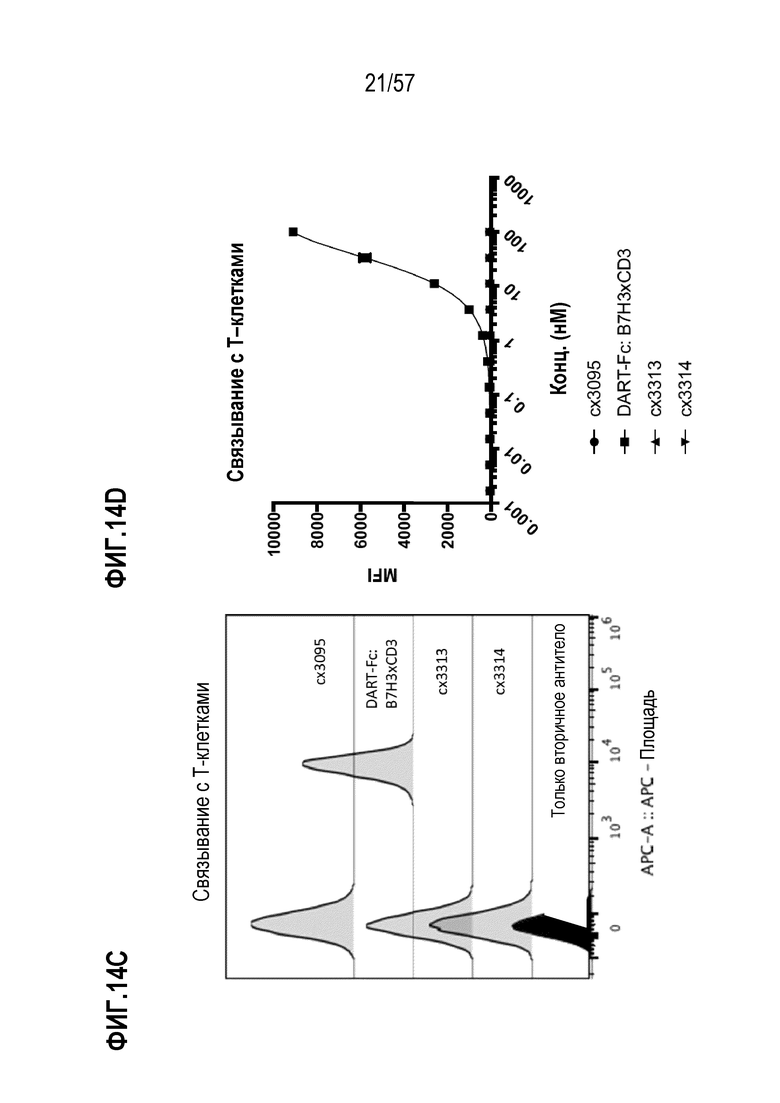
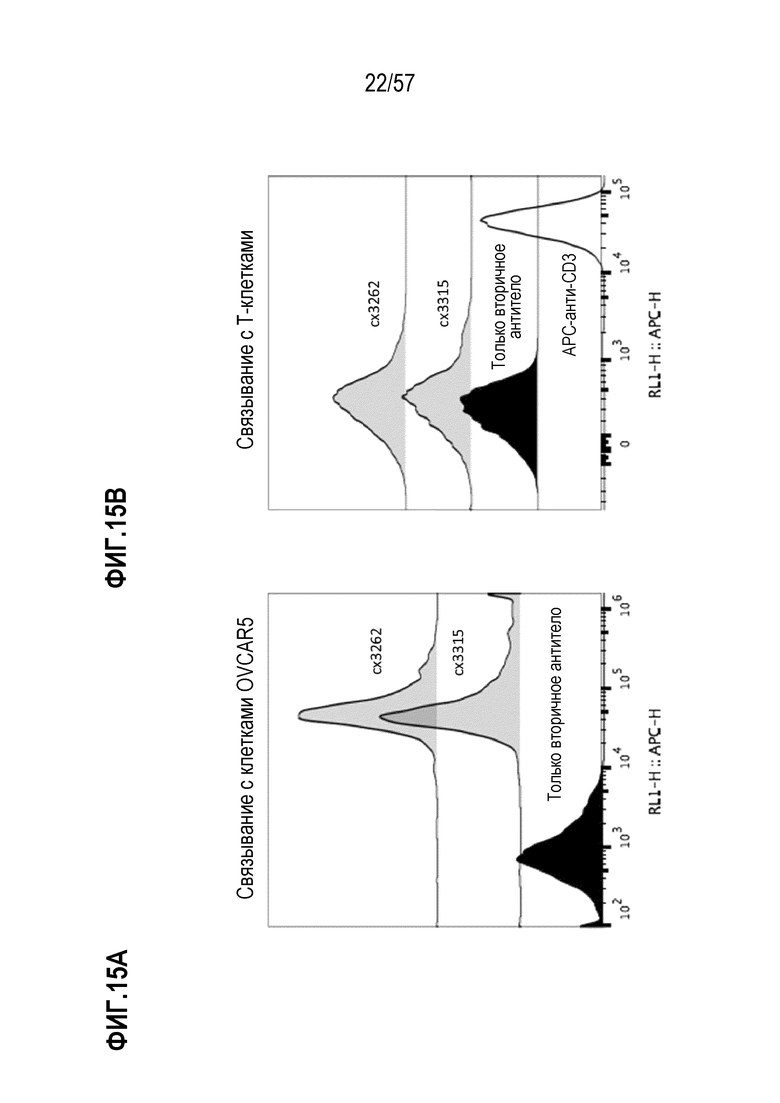
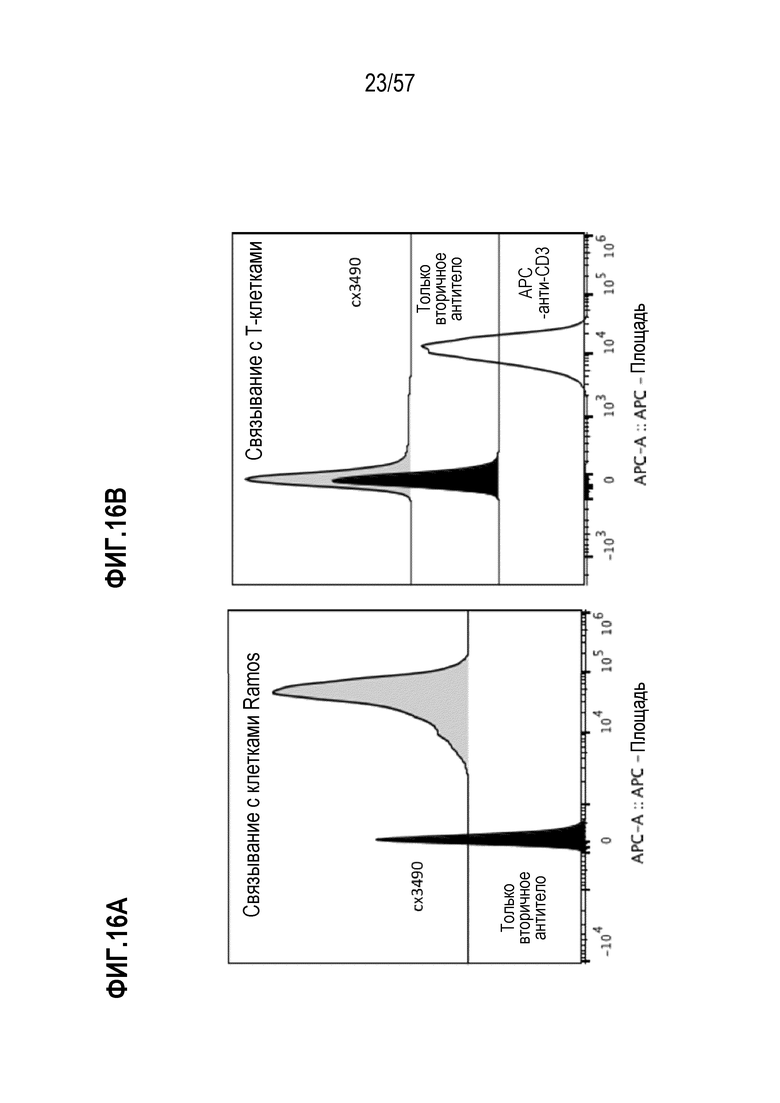
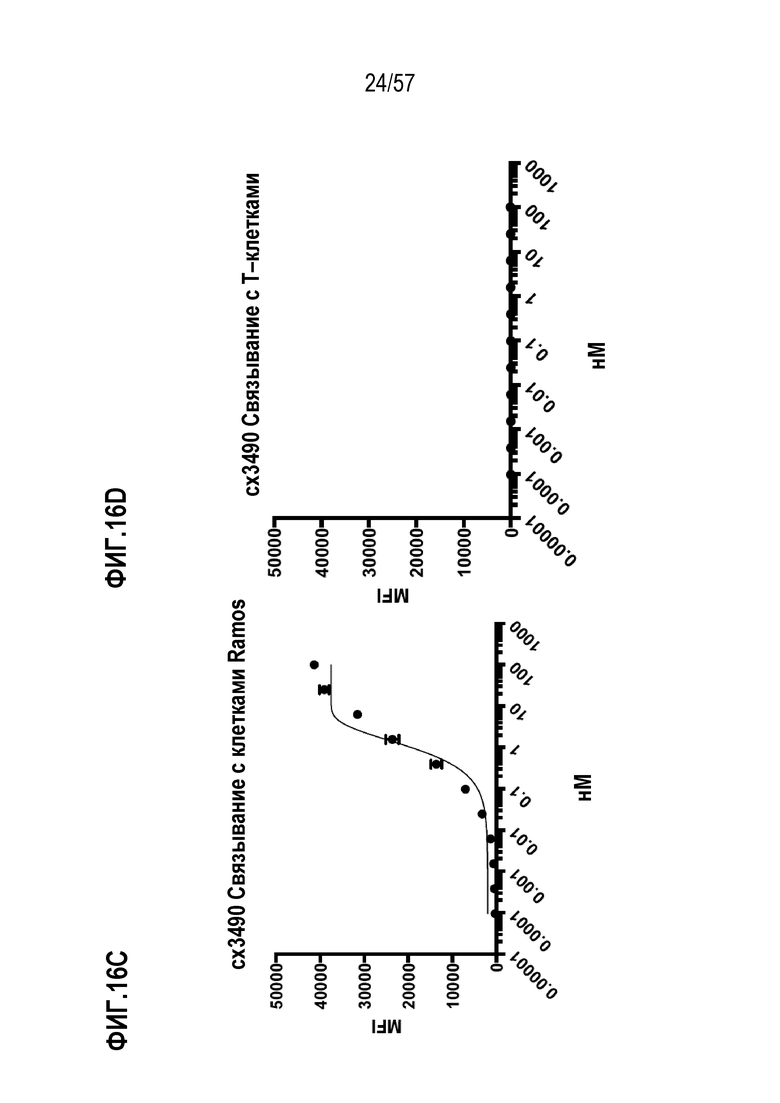
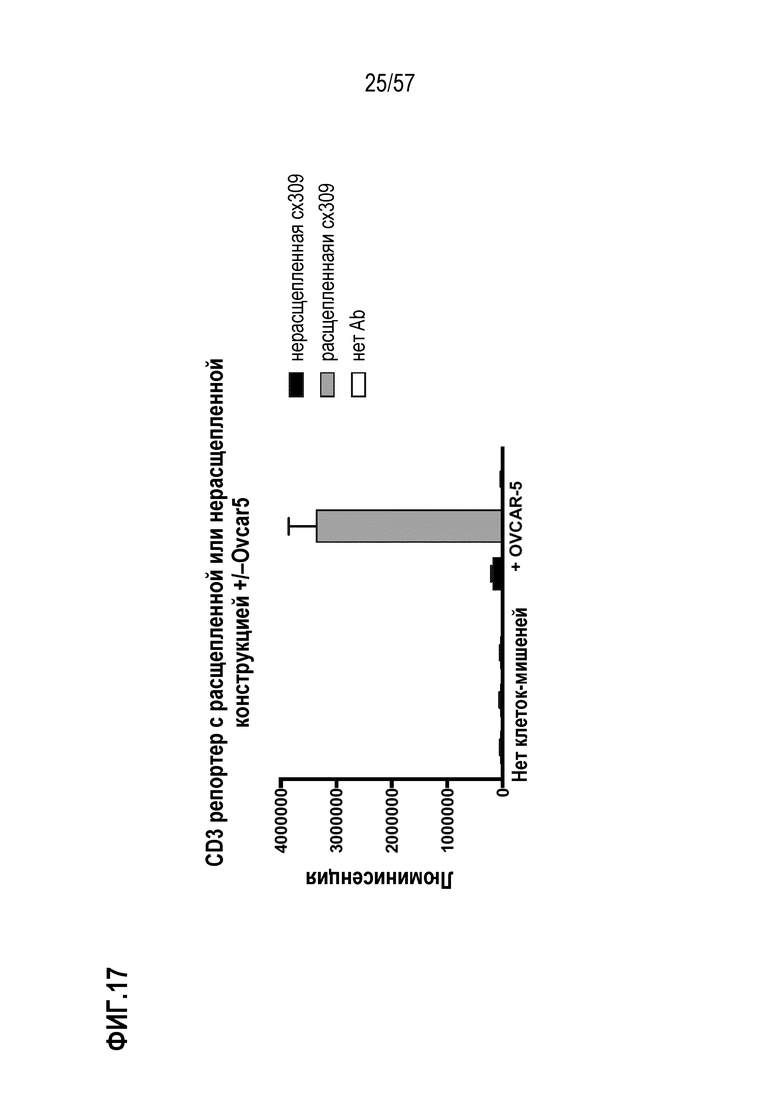
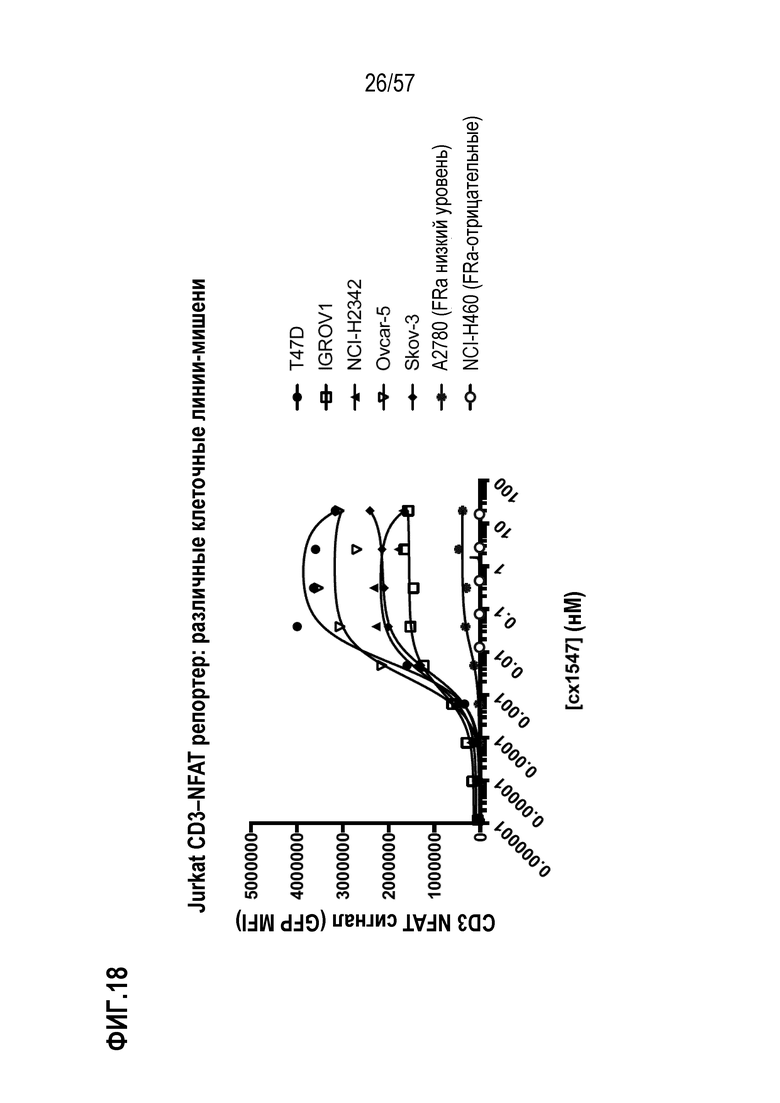
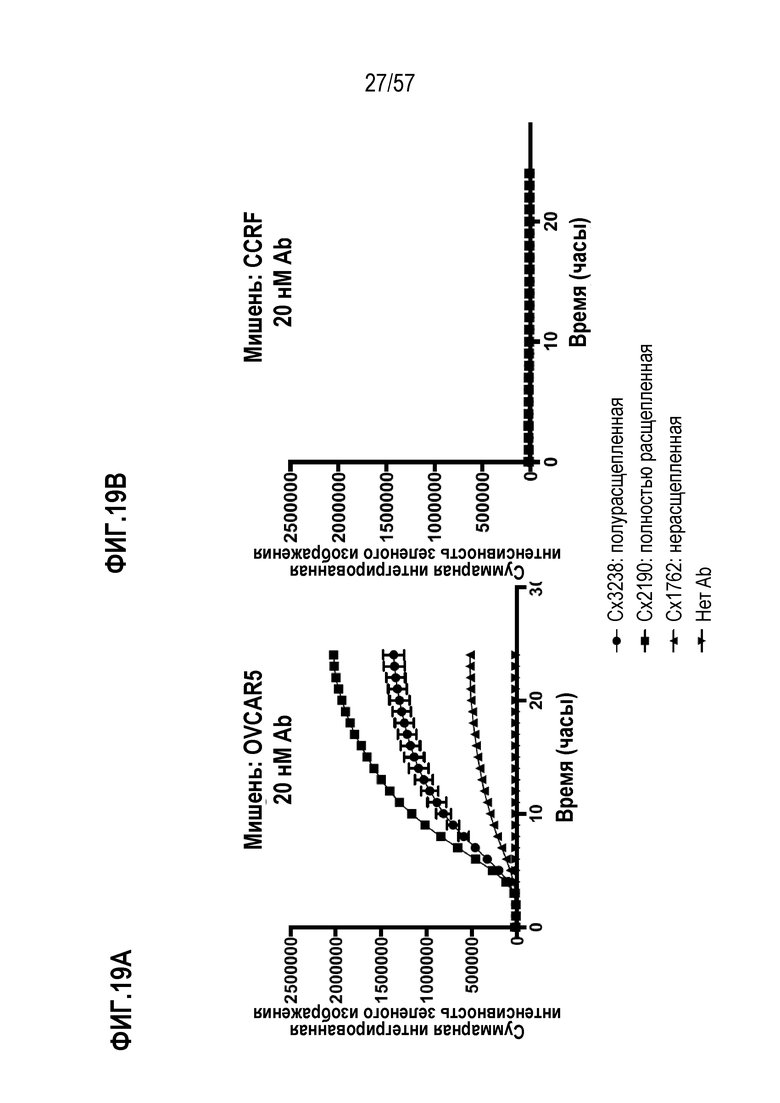
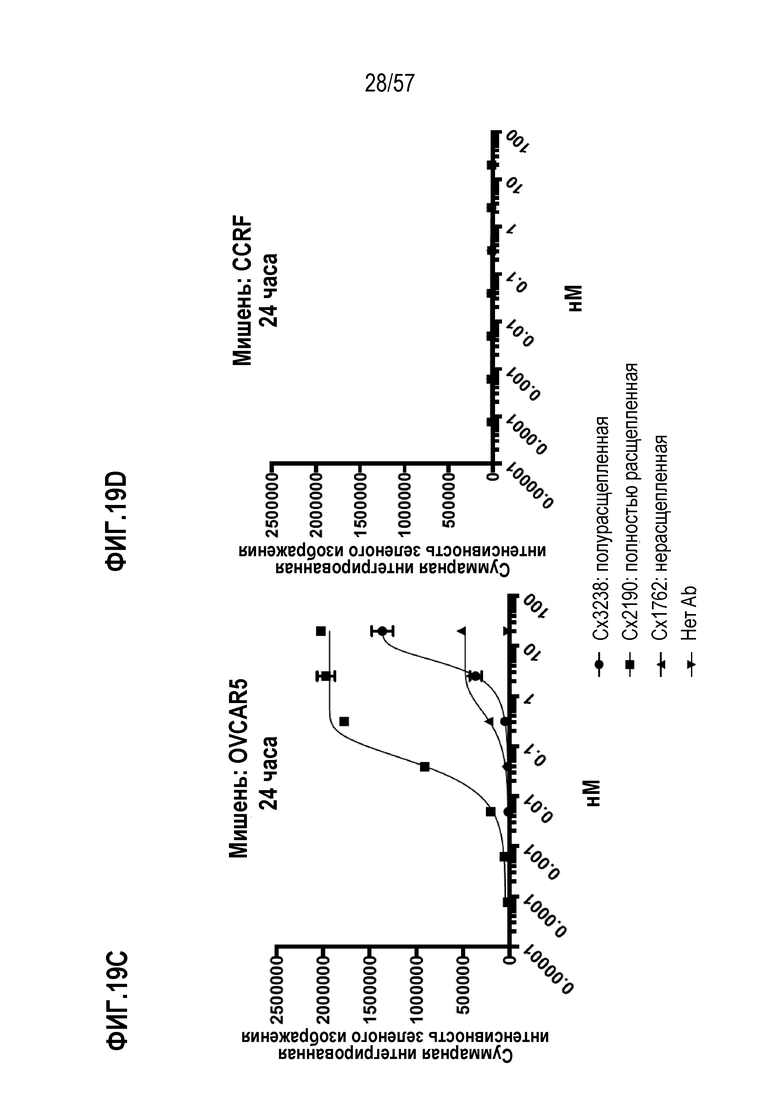
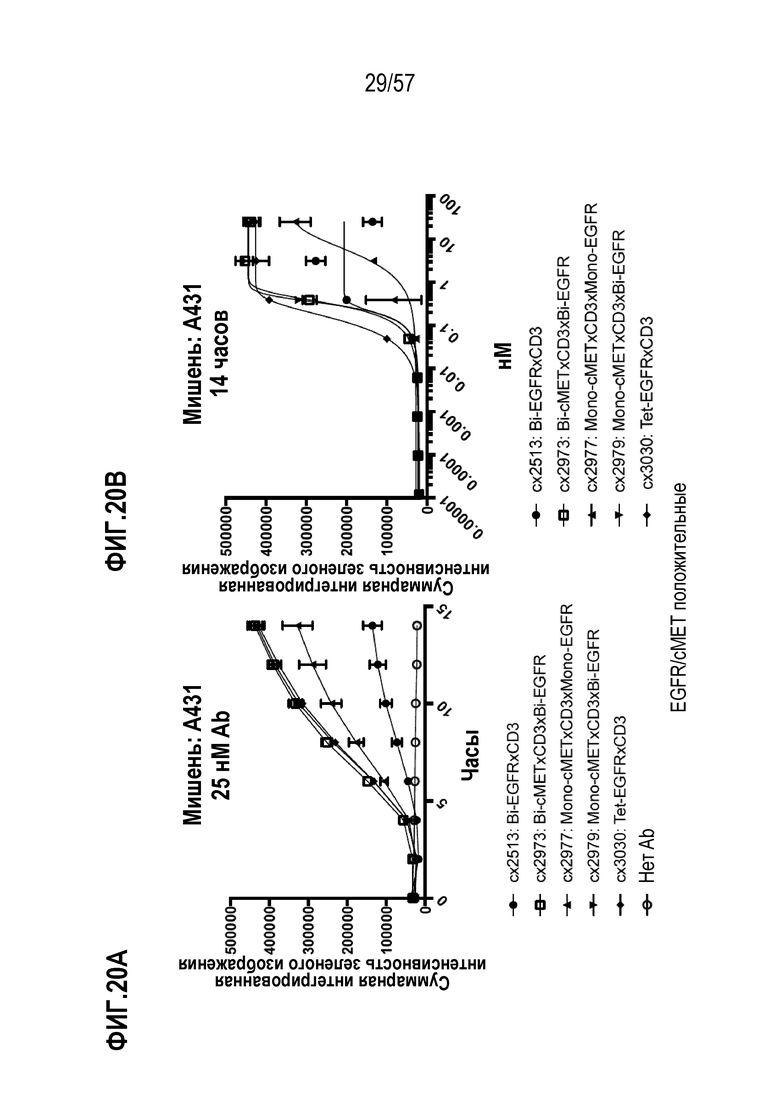
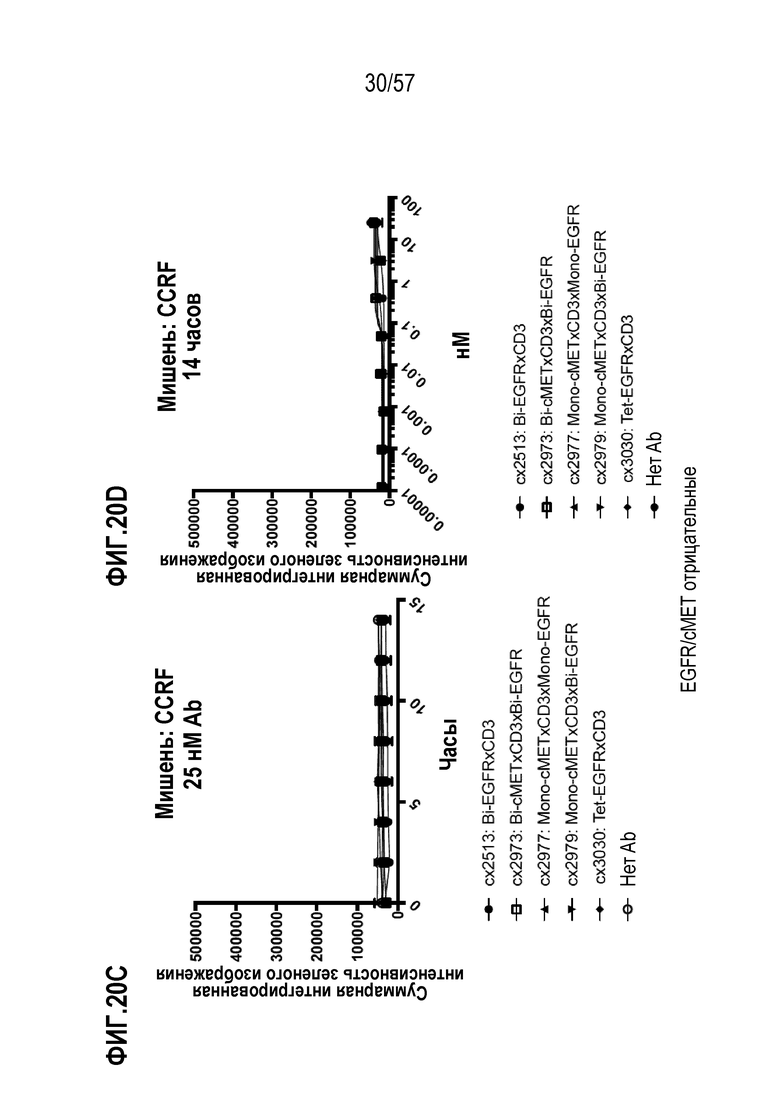
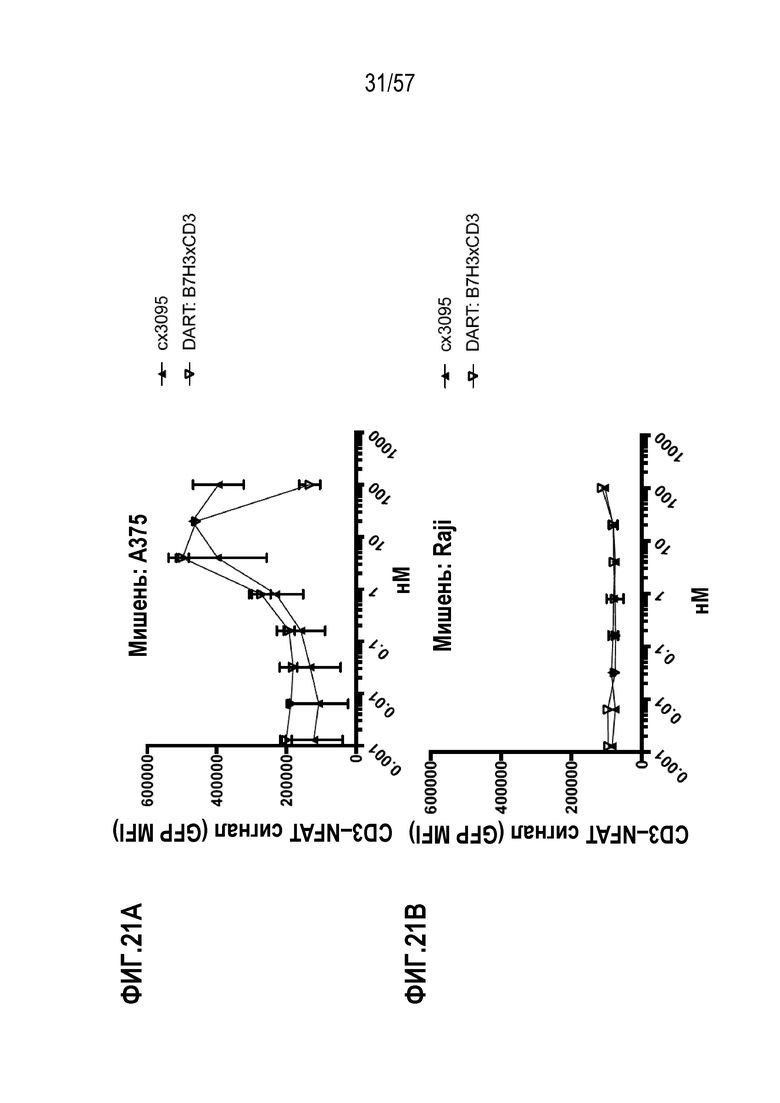
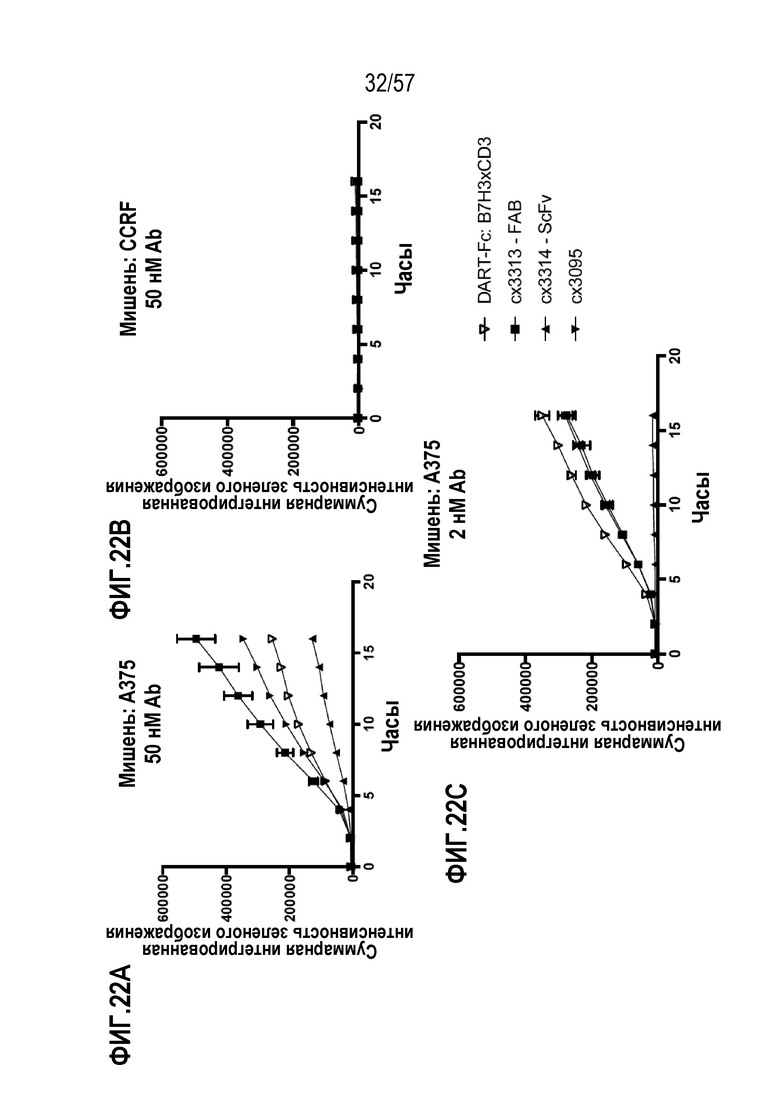
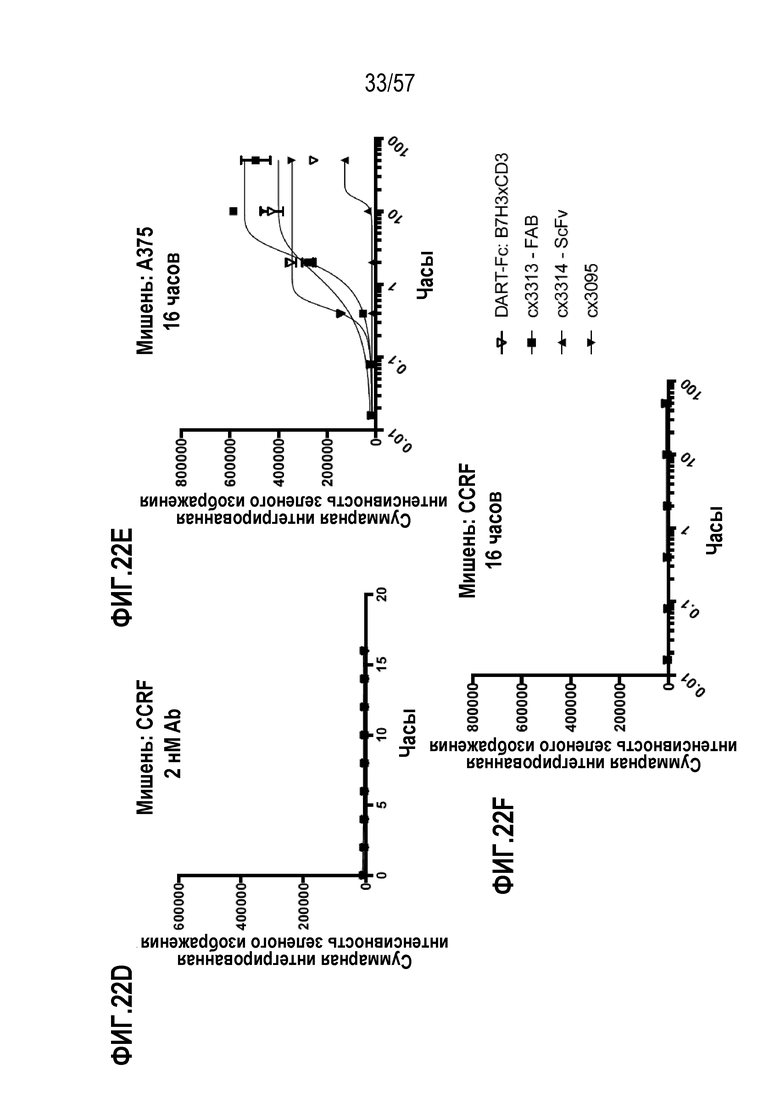
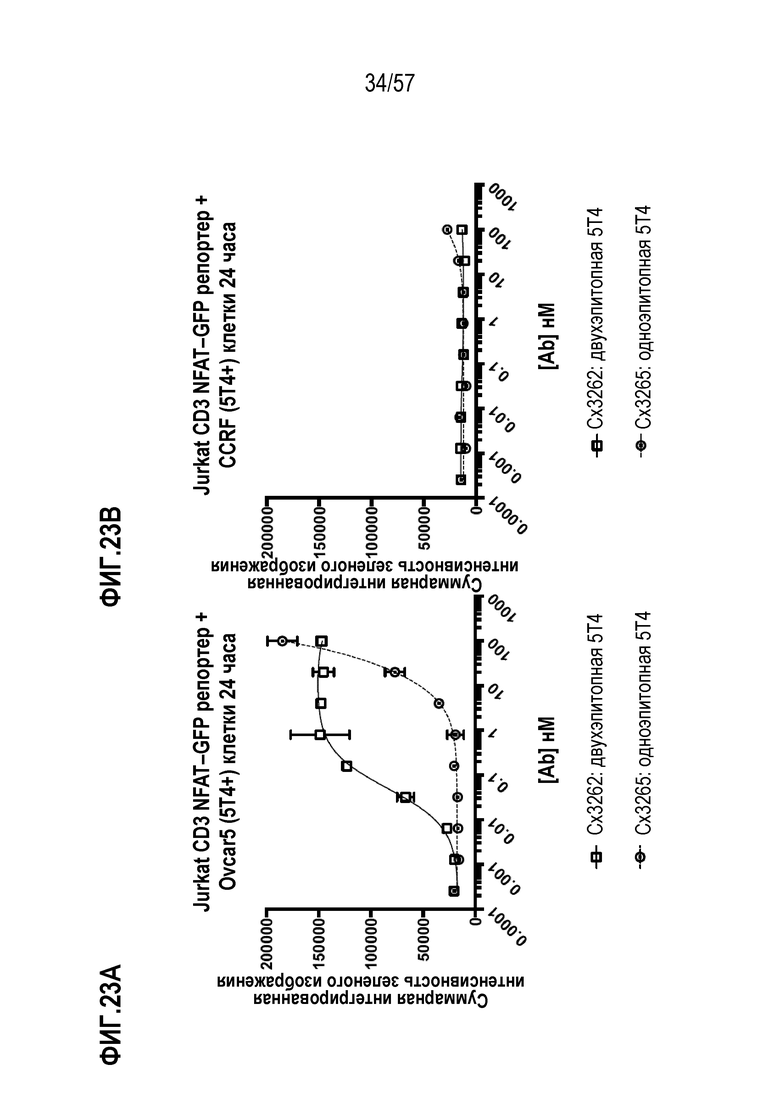
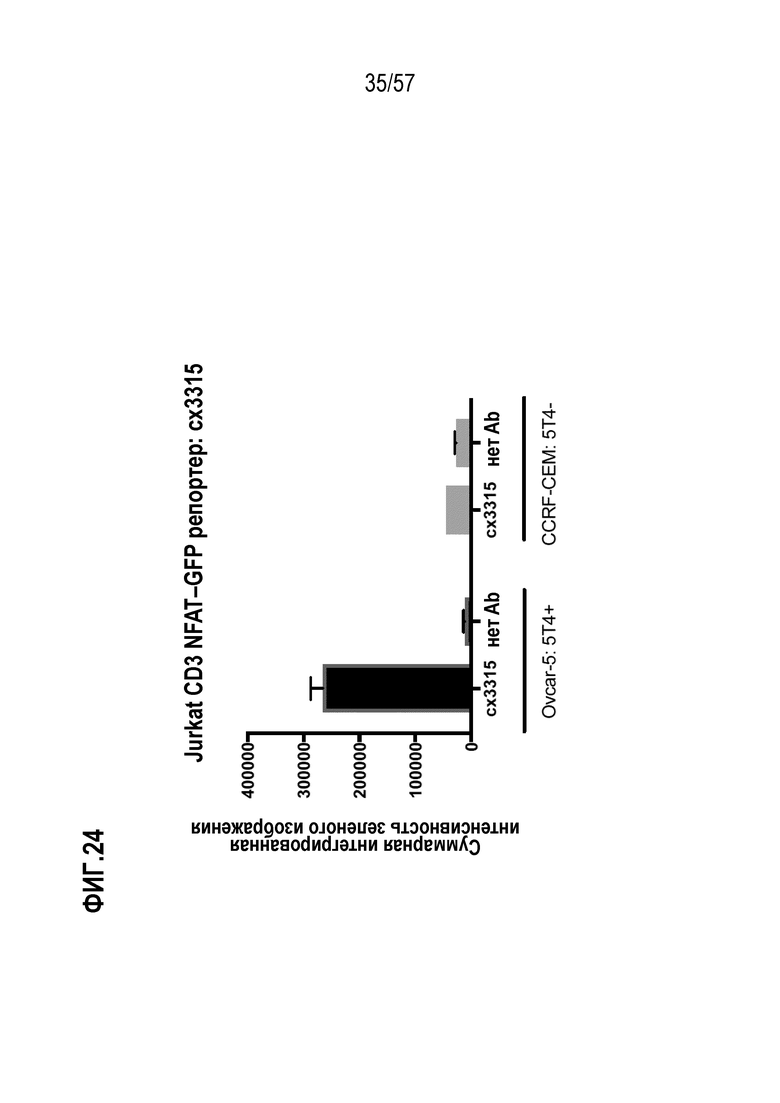
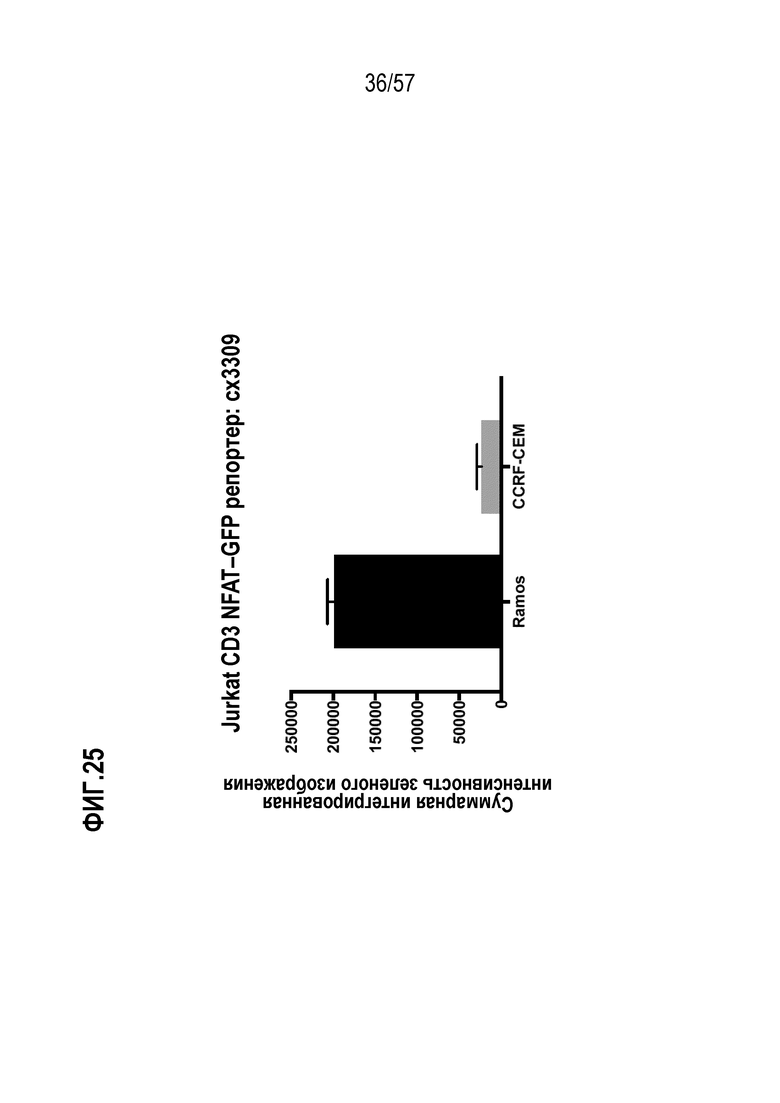
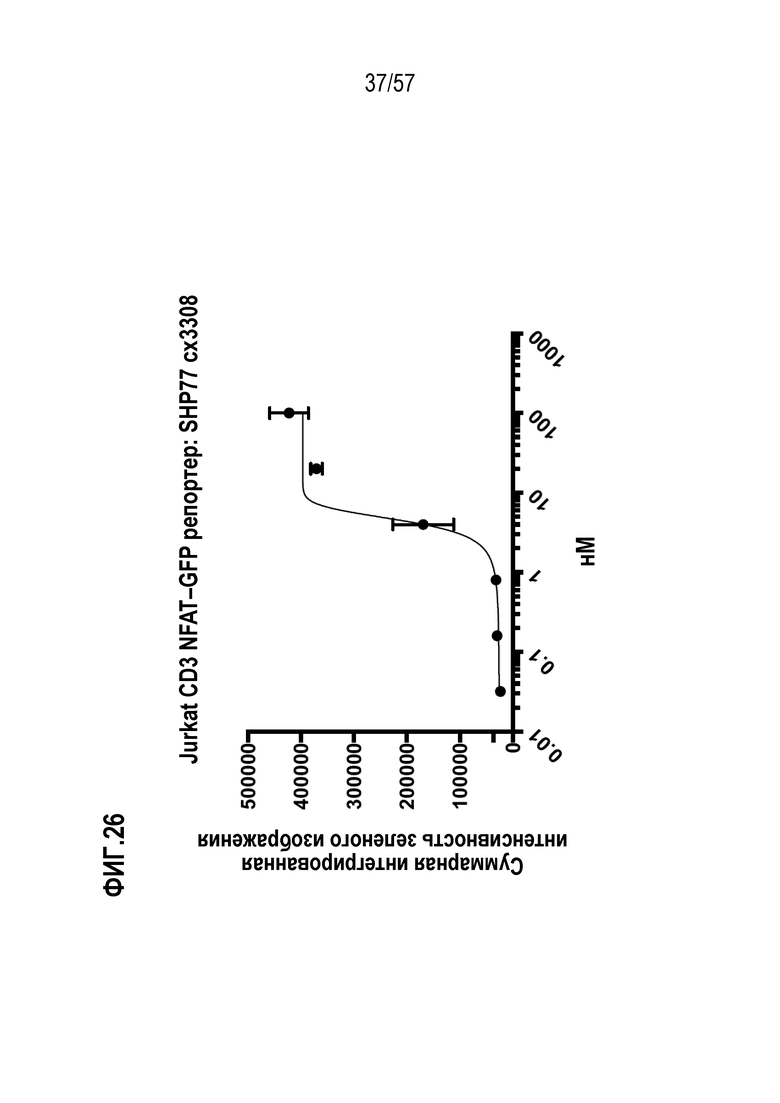
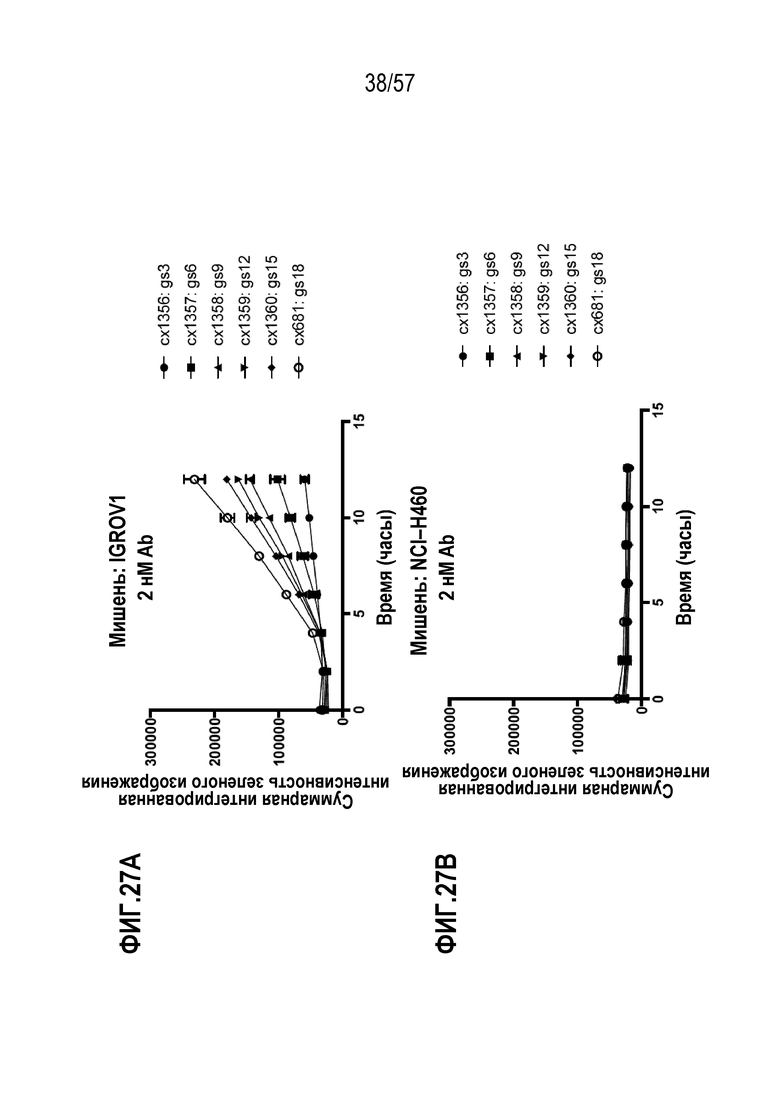
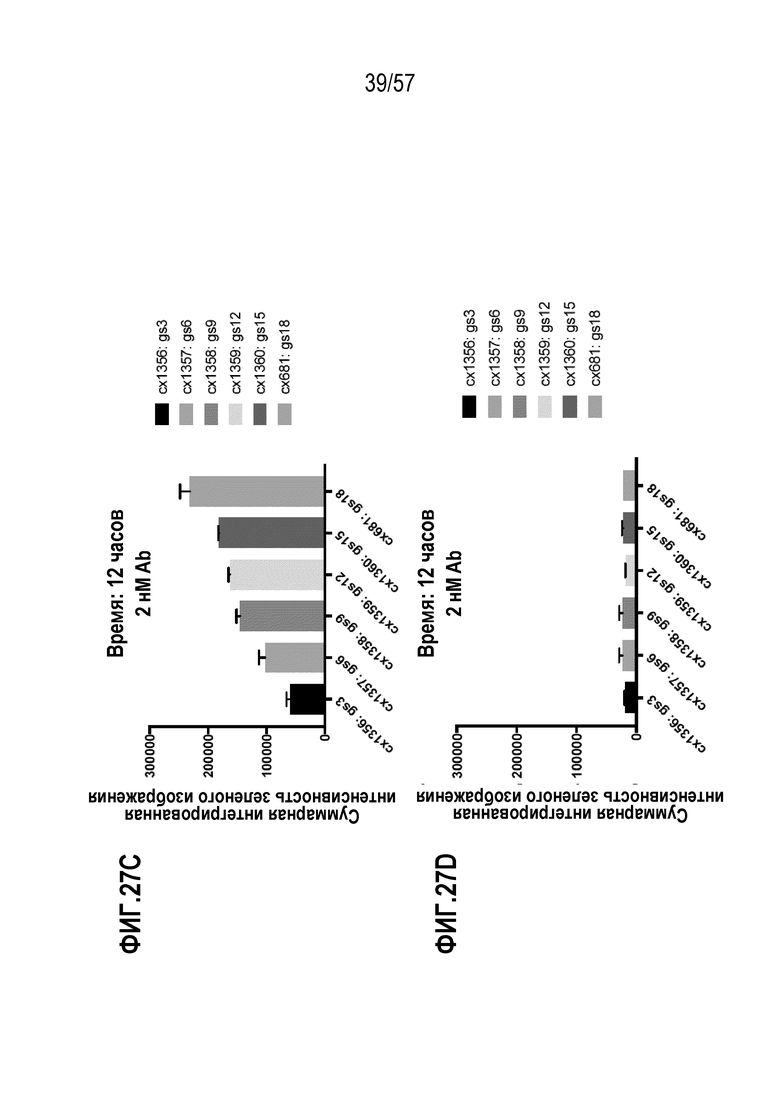
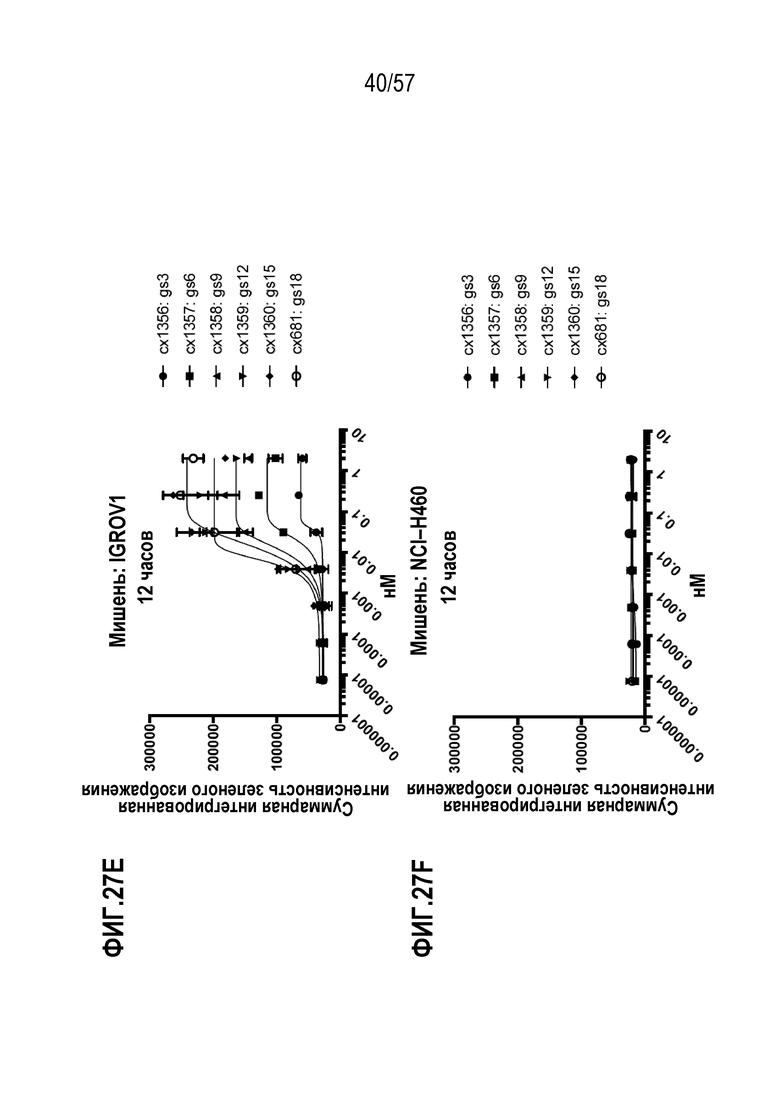
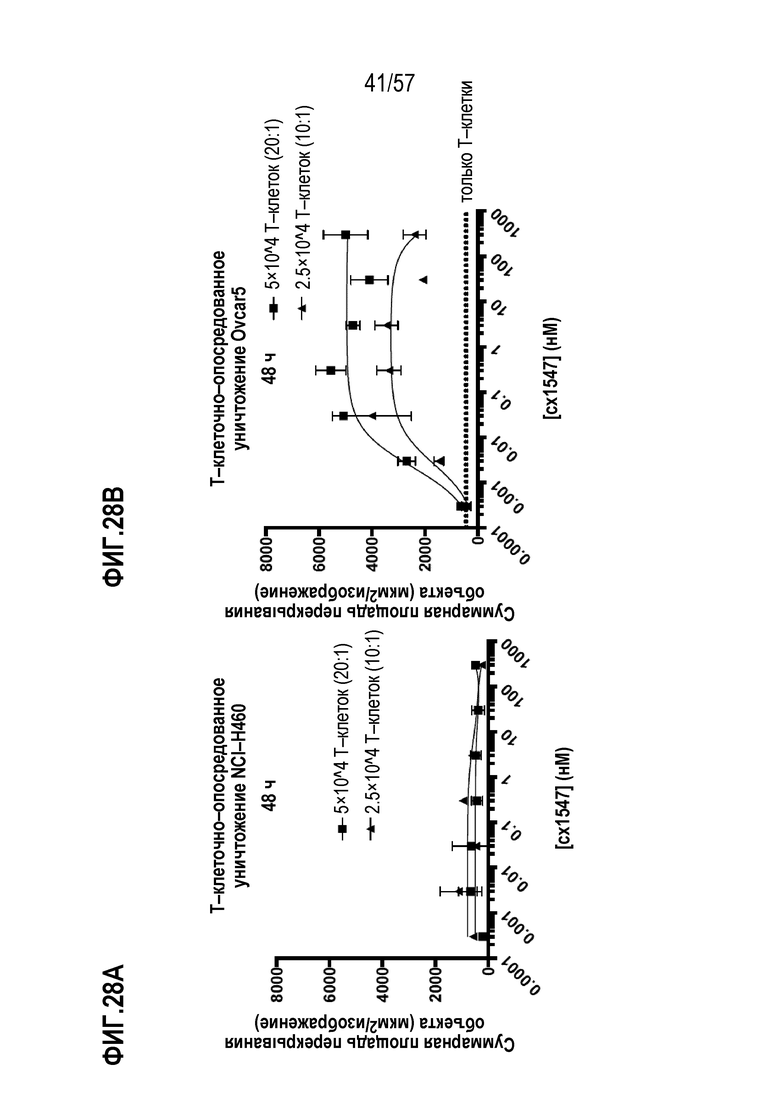
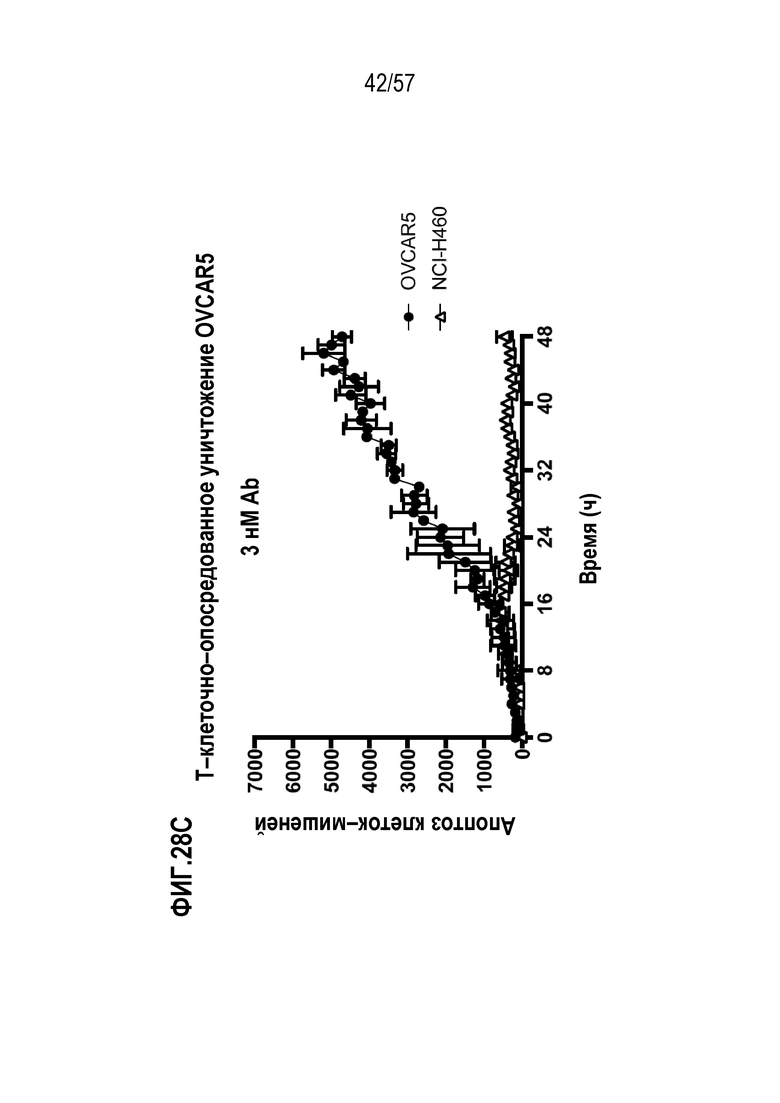
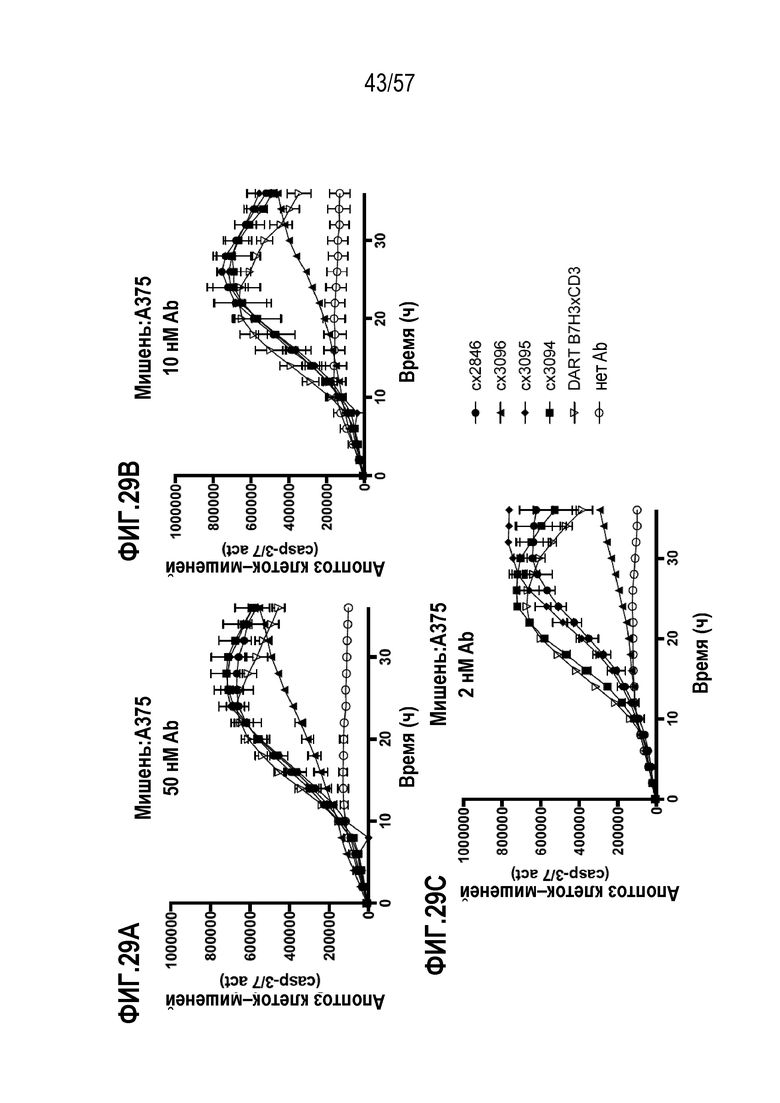
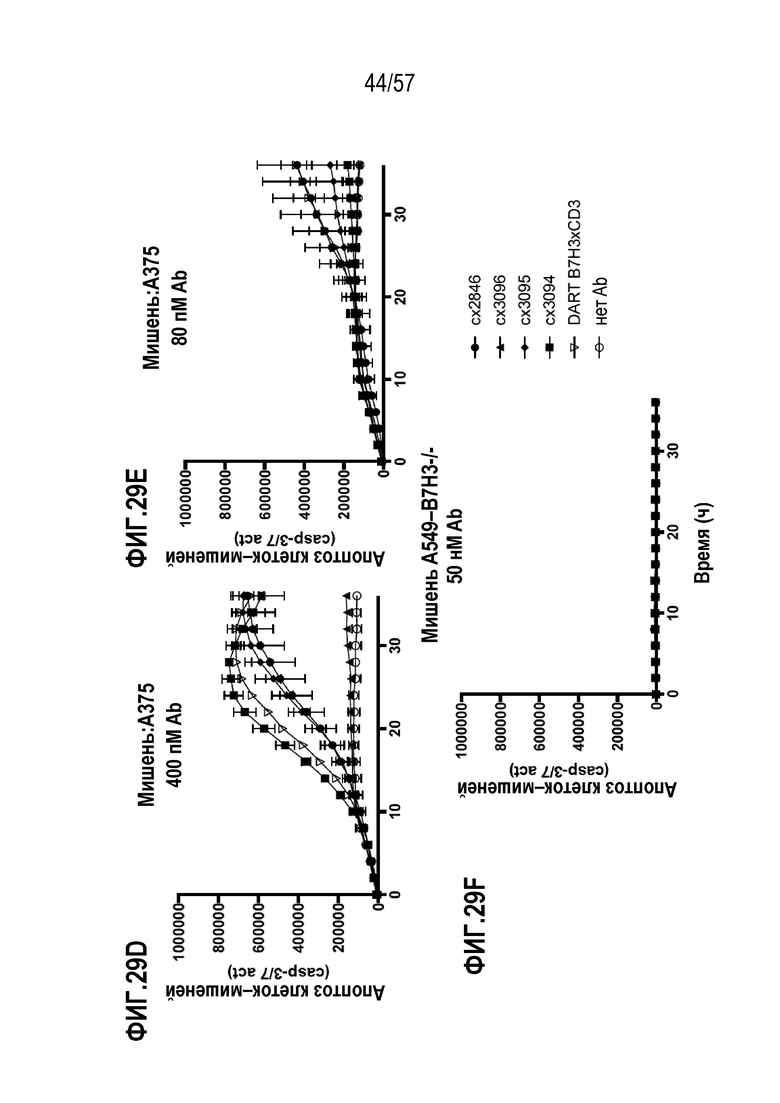
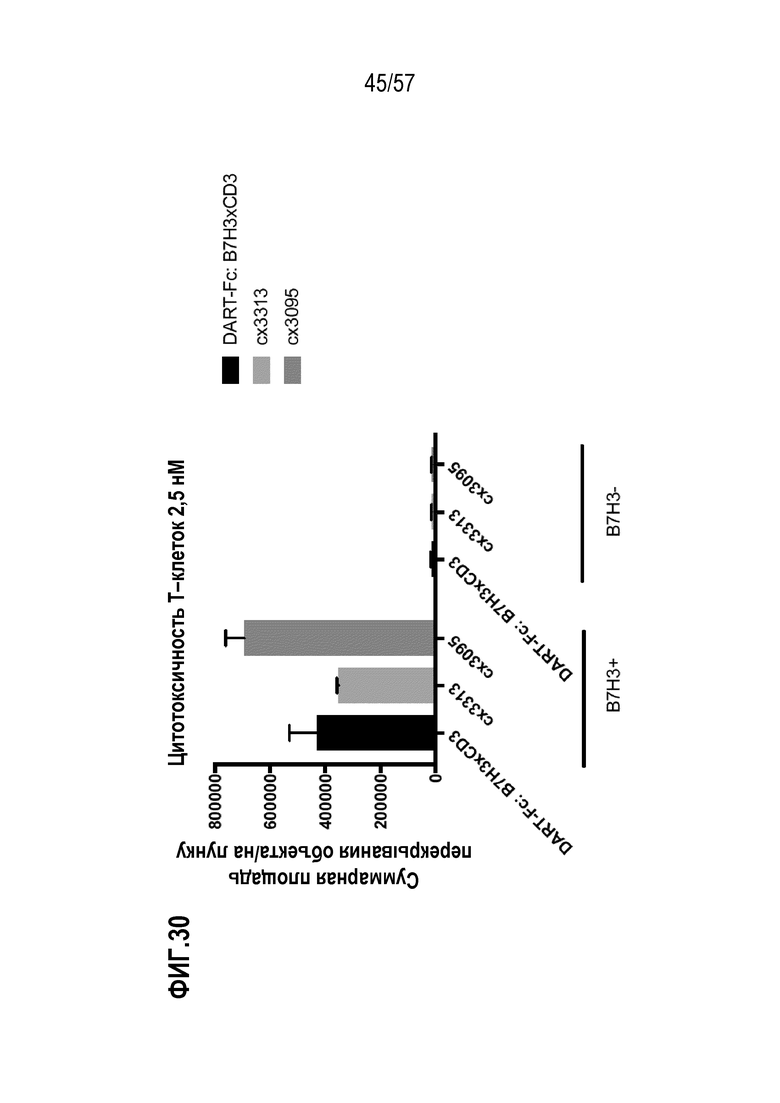
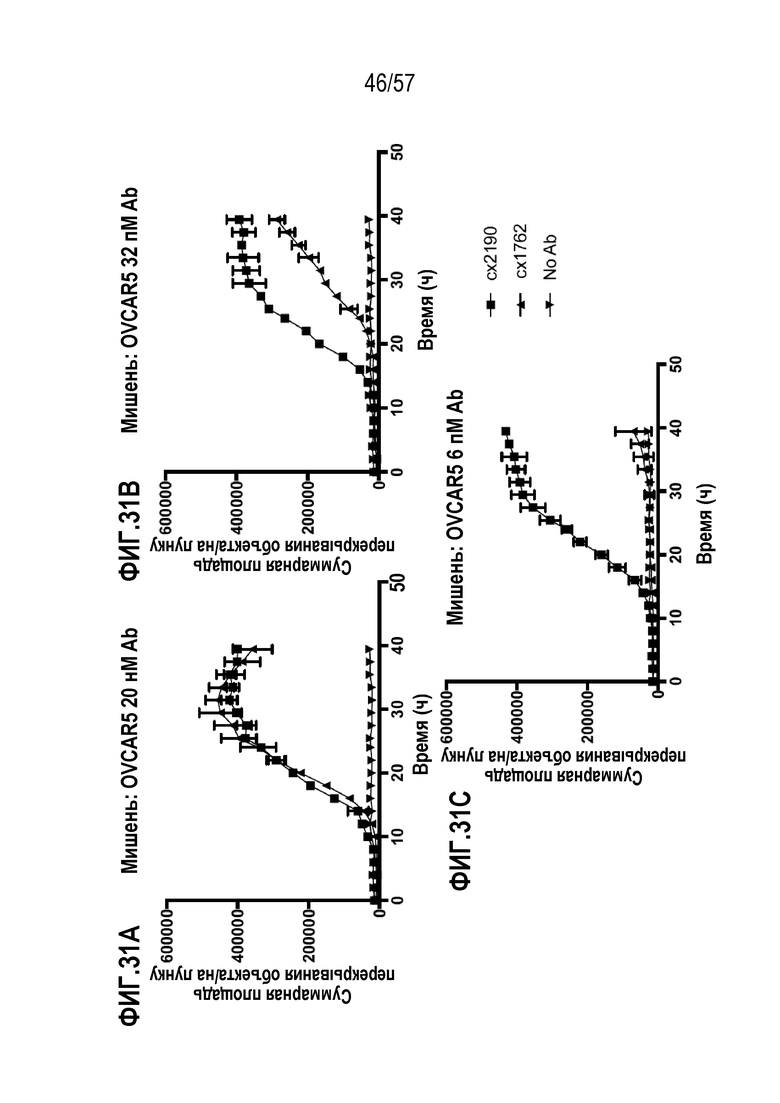
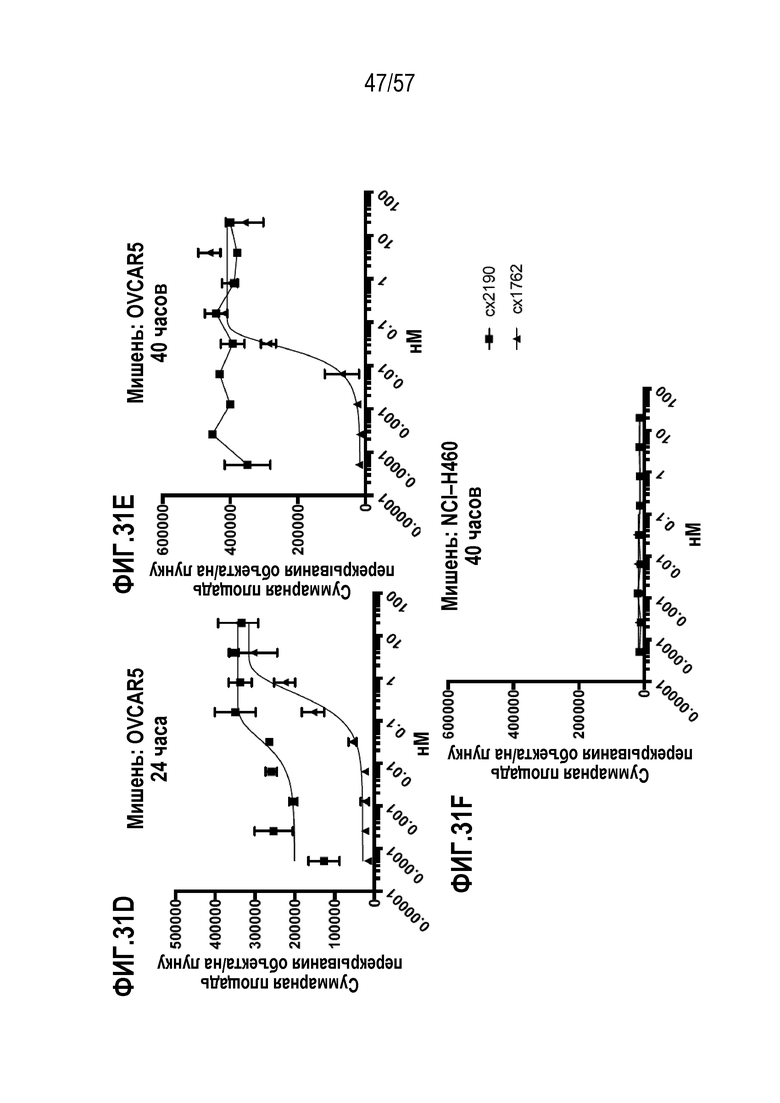
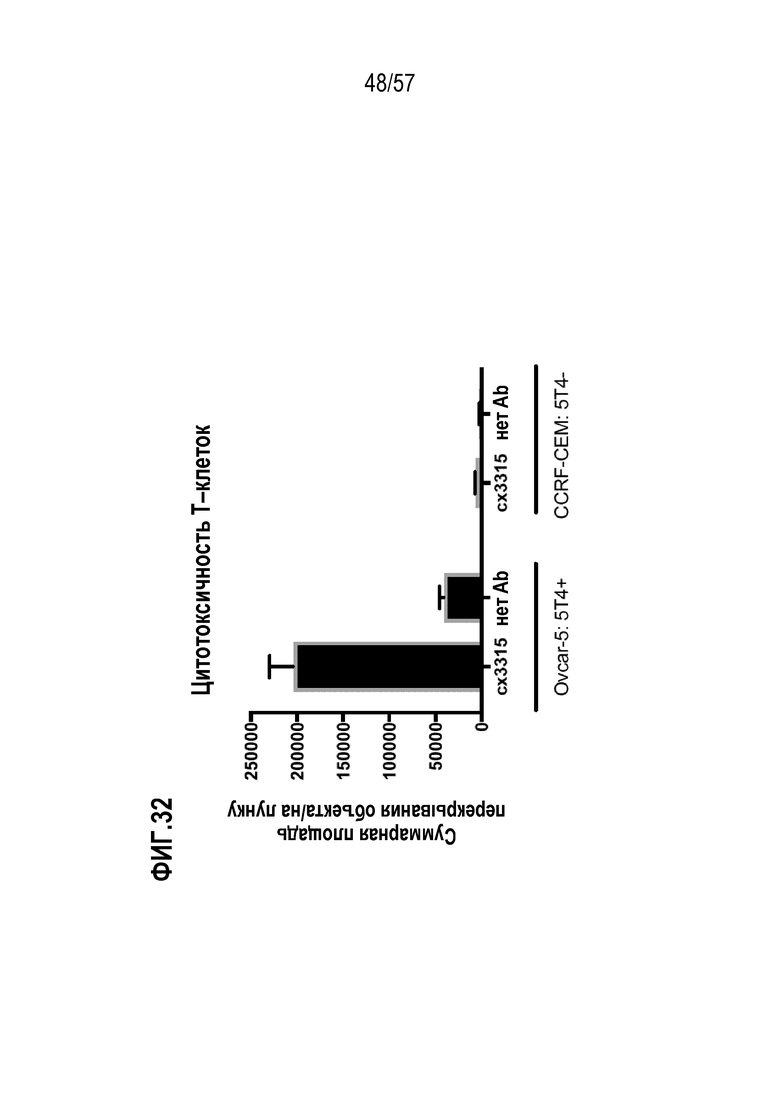
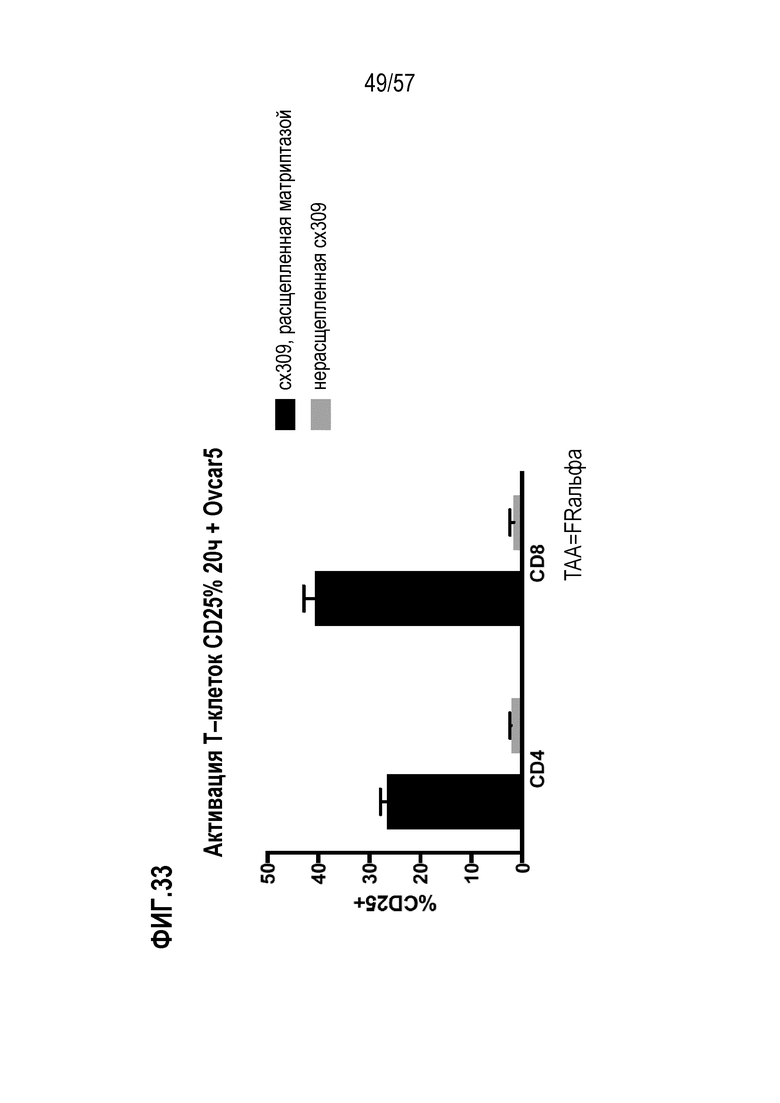
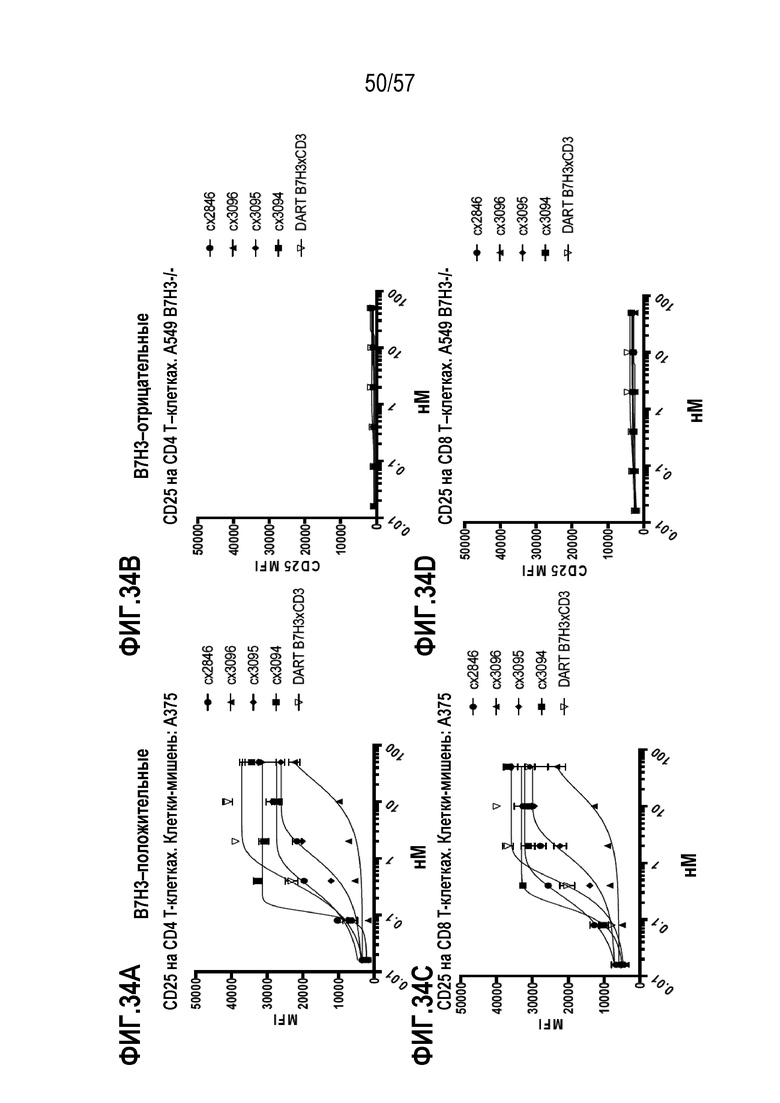
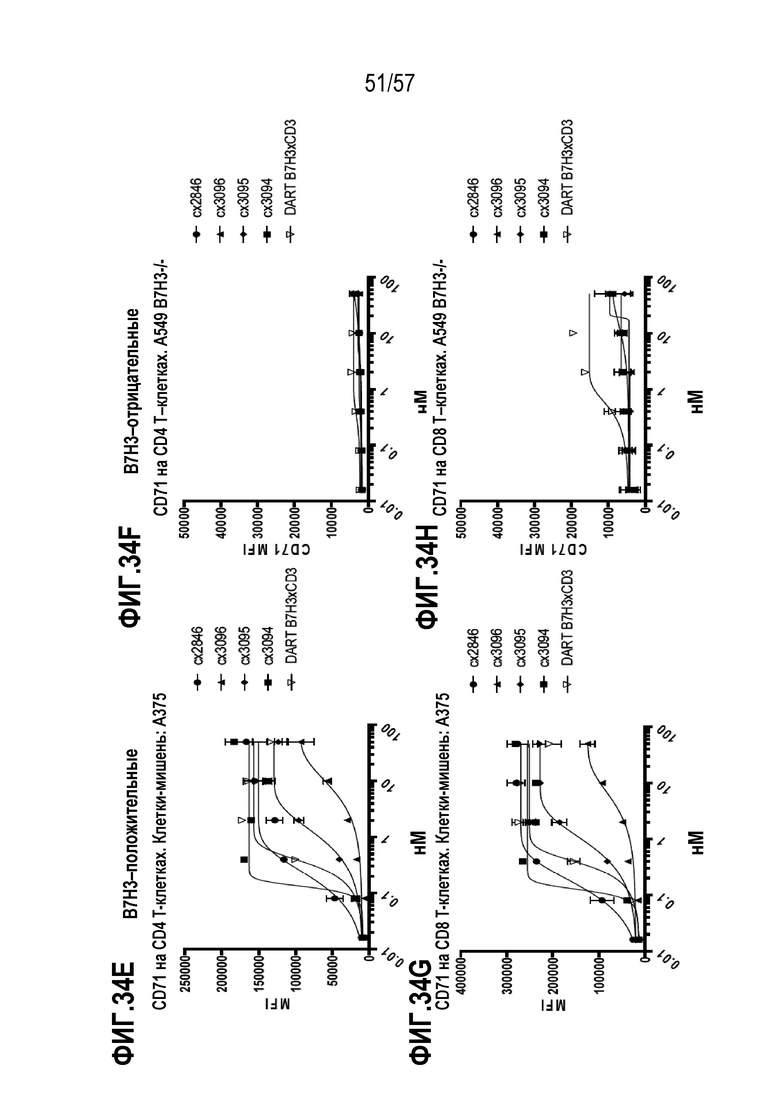
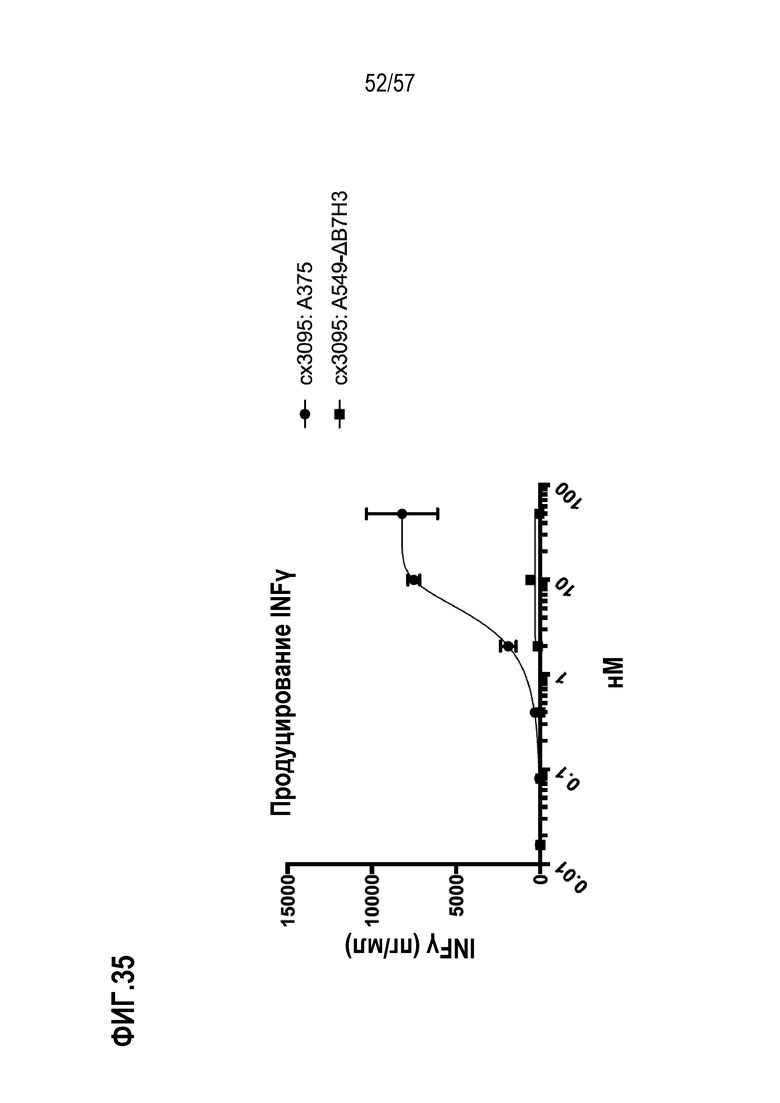
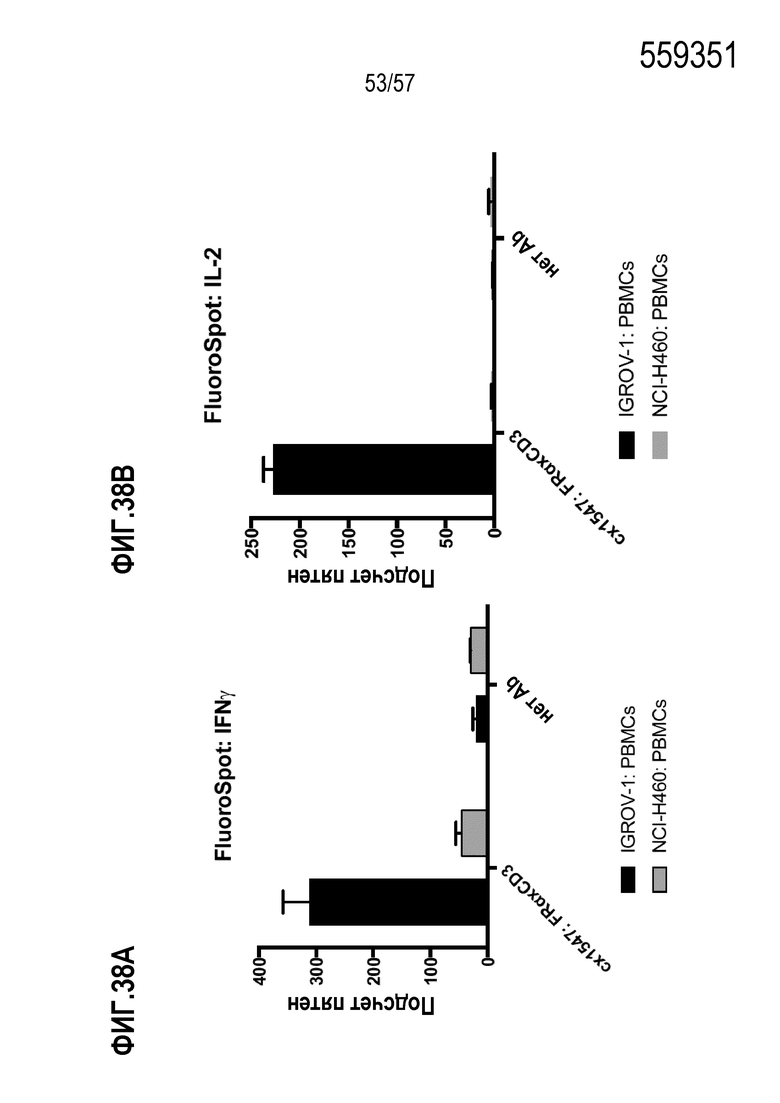
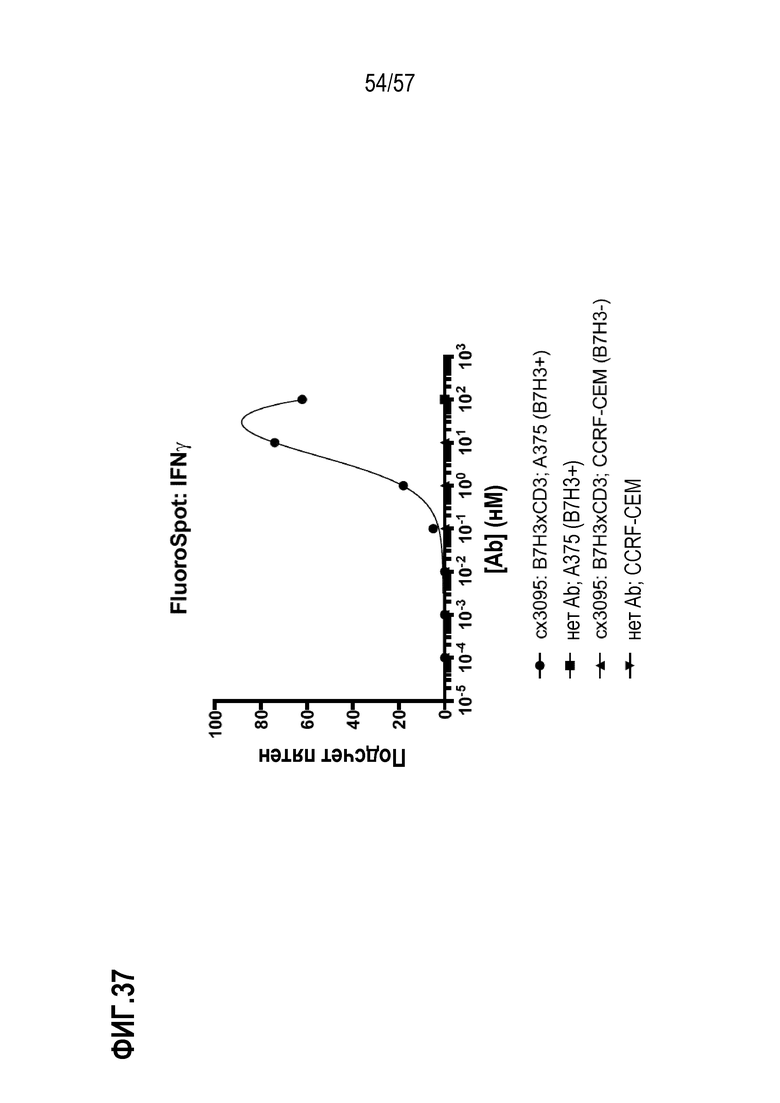
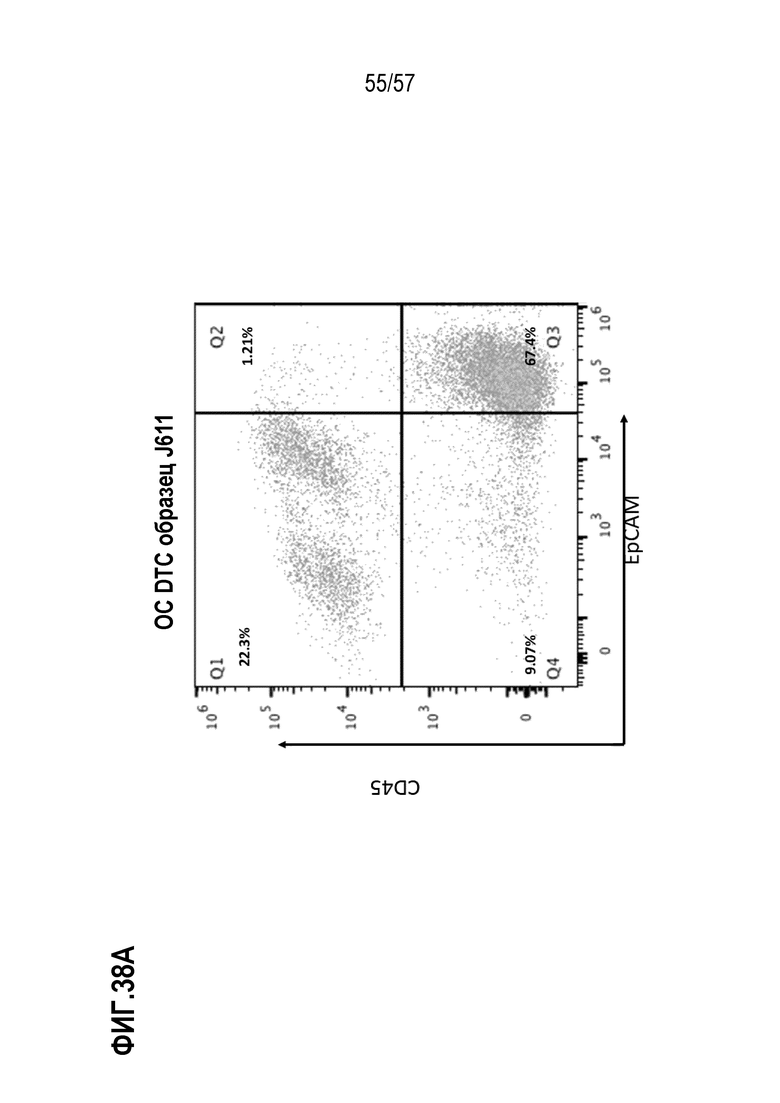
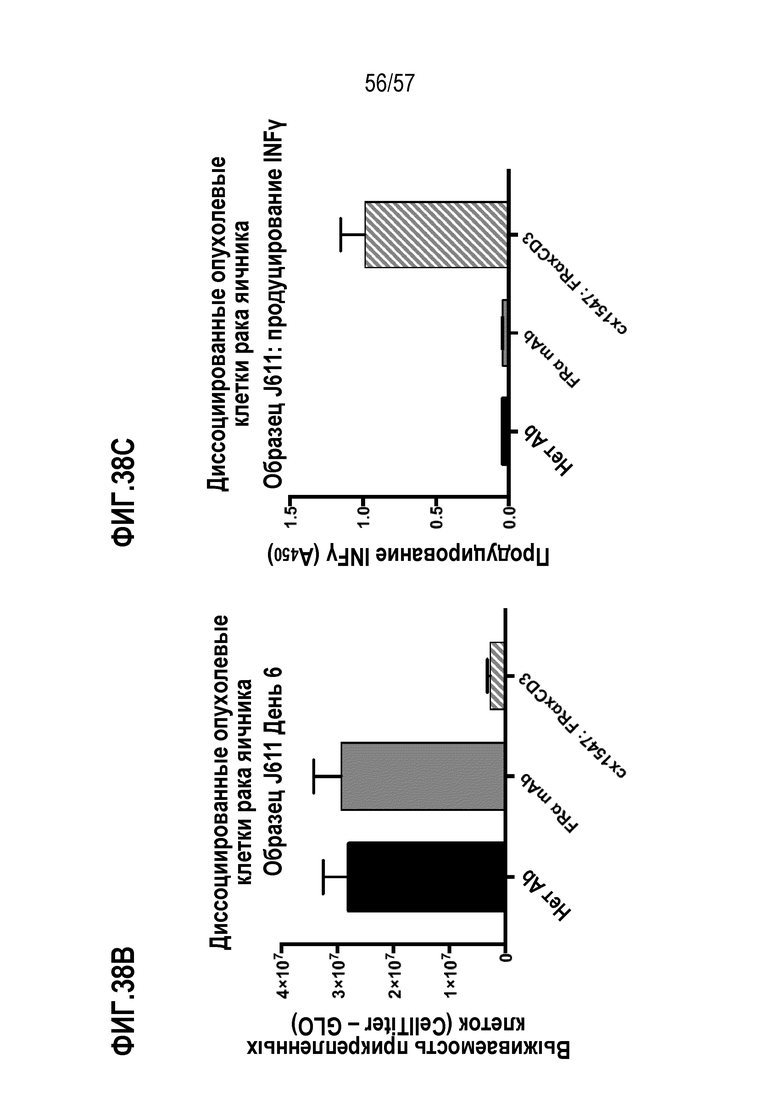
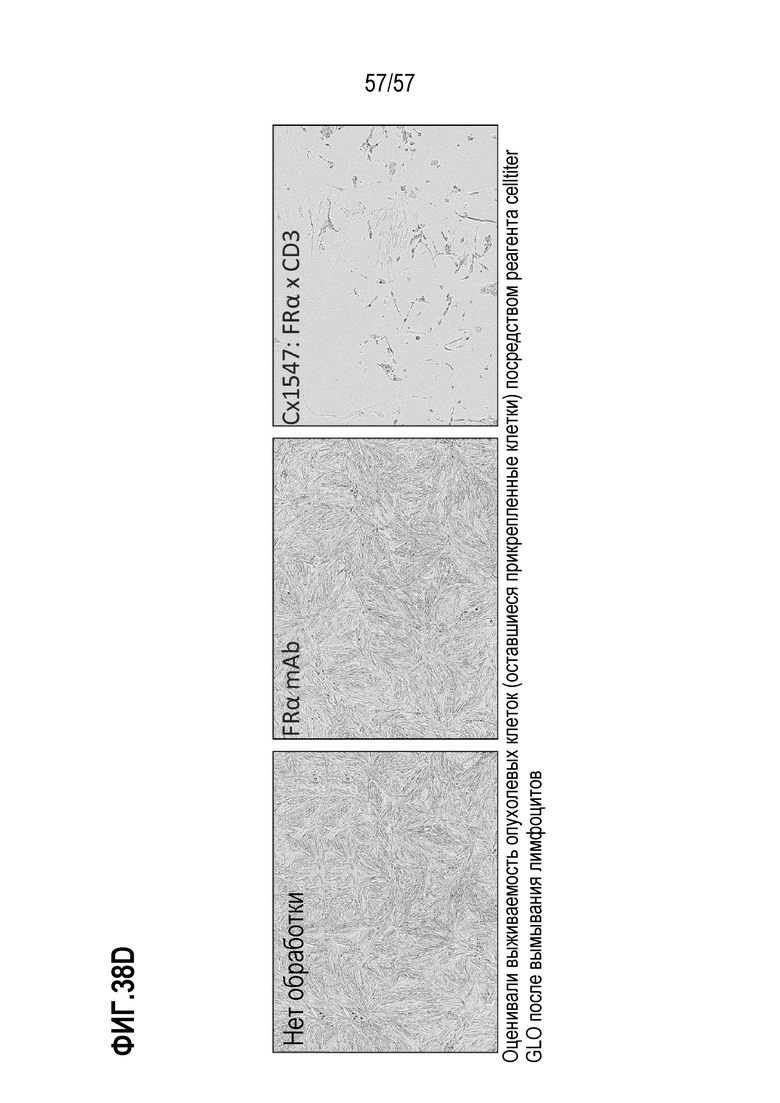
Группа изобретений относится к биотехнологии и иммунологии. Предложена полиспецифическая полипептидная конструкция для связывания с CD3 и опухолеспецифическим антигеном (TAA), обладающая ограниченным связыванием с CD3. Также предложены полинуклеотид, кодирующий полиспецифическую полипептидную конструкцию, экспрессионный вектор, клетка, фармацевтическая композиция и способы получения и применения полиспецифических полипептидов для стимулирования или индукции иммунного ответа у пациента, имеющего опухоль или злокачественное новообразование или лечения опухоли или злокачественного новообразования у пациента. Предложенные конструкции не демонстрируют способность связывать T-клетки, когда они не связаны с TAA-мишенью, в то же время сохраняя сильную способность индуцировать антиген-зависимую цитотоксичность Т-клеток. 11 н. и 44 з.п. ф-лы, 38 ил., 3 табл., 5 пр.
1. Полиспецифическая полипептидная конструкция для связывания с CD3 и опухолеспецифическим антигеном (TAA), содержащая первый компонент, содержащий Fc–область иммуноглобулина, и второй компонент, содержащий CD3-связывающую область, где:
CD3-связывающая область представляет собой стабилизированный дисульфидом анти-CD3-связывающий Fv-фрагмент (dsFv), содержащий вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH имеет аминокислотную последовательность SEQ ID NO: 44 или последовательность, которая демонстрирует по меньшей мере 90% идентичности последовательности SEQ ID NO: 44, а VL имеет аминокислотную последовательность SEQ ID NO: 72 или последовательность, которая демонстрирует по меньшей мере 90% идентичности последовательности SEQ ID NO: 72;
Fc-область представляет собой гетеродимерную Fc-область, а VH и VL, содержащие анти-CD3 dsFv, связаны с противоположными полипептидами гетеродимерной Fc-области;
один или оба полипептида Fc гетеродимерной Fc-области содержат по меньшей мере одну модификацию, способствующую гетеродимеризации, по сравнению с полипептидом гомодимерной Fc-области;
первый и второй компоненты связаны первым полипептидным линкером, который составляет 2-20 аминокислот в длину, который состоит по меньшей мере на 50% из глицина, где Fc-область расположена на амино-конце относительно CD3-связывающей области; где
первый компонент содержит по меньшей мере один антигенсвязывающий домен и связывает опухолеассоциированный антиген (ТАА), причем каждый из указанного по меньшей мере одного антигенсвязывающего домена первого компонента связан с амино-конца с Fc-областью вторым линкером, представляющим собой полипептид длиной 2-6 аминокислот, состоящим по меньшей мере на 50% из остатков глицина; и/или
второй компонент содержит по меньшей мере один антигенсвязывающий домен и связывается с одним и тем же TAA, причем каждый из указанного по меньшей мере одного антигенсвязывающего домена второго компонента связан с карбокси-конца с CD3-связывающей областью третьим линкером, который представляет собой полипептид длиной 2-6 аминокислот, состоящий по меньшей мере на 50% из остатков глицина, и
где каждый из антигенсвязывающих доменов представляет собой верблюжий VHH или гуманизированный верблюжий VHH,
где указанная CD3-связывающая область не способна связывать CD3, если только антигенсвязывающий домен не связан с его ТАА.
2. Полиспецифическая полипептидная конструкция по п.1, отличающаяся тем, что CD3-связывающая область связывается с CD3ε.
3. Полиспецифическая конструкция по п.1 или 2, отличающаяся тем, что первый компонент содержит по меньшей мере один антигенсвязывающий домен, расположенный на амино-конце относительно Fc-области и второй компонент содержит по меньшей мере один антигенсвязывающий домен, расположенный на карбокси-конце относительно CD3-связывающей области полиспецифической полипептидной конструкции.
4. Полиспецифическая полипептидная конструкция по любому из пп.1-3, отличающаяся тем, что Fc-область представляет собой Fc-область человеческого IgG1, человеческого IgG2, человеческого IgG3 или человеческого IgG4 или представляет собой их иммунологически активный фрагмент.
5. Полиспецифическая полипептидная конструкция по любому из пп.1-4, отличающаяся тем, что Fc-область содержит:
полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 1, или последовательность аминокислот, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичности последовательности с SEQ ID NO: 1;
полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 2, или последовательность аминокислот, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичности последовательности с SEQ ID NO: 2;
полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 4, или последовательность аминокислот, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичности последовательности с SEQ ID NO: 4; или
полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 5, или последовательность аминокислот, которая имеет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичности последовательности с SEQ ID NO: 5.
6. Полиспецифическая полипептидная конструкция по любому из пп.1-5, отличающаяся тем, что один или оба полипептида Fc гетеродимерной Fc-области содержат по меньшей мере одну модификацию, способствующую гетеродимеризации, по сравнению с полипептидом Fc, представленным в SEQ ID NO: 1, или его иммунологически активным фрагментом.
7. Полиспецифическая полипептидная конструкция по любому из пп.1-6, отличающаяся тем, что каждый из полипептидов Fc гетеродимерной Fc-области содержит модификацию «выступ-во-впадину» или содержит мутацию заряда для увеличения электростатической комплементарности полипептидов.
8. Полиспецифическая полипептидная конструкция по п.6 или 7, отличающаяся тем, что первый полипептид Fc гетеродимерной Fc-области содержит модификацию, выбранную из Thr366Ser, Leu368Ala, Tyr407Val и их комбинаций, и второй полипептид Fc гетеродимерной Fc-области содержит модификацию T366W, и где необязательно первый и второй полипептиды Fc дополнительно содержат модификацию нецистеинового остатка до цистеинового остатка, где модификация первого полипептида находится в одном из положений Ser354 и Y349, а модификация второго полипептида Fc находится в другом положении из Ser354 и Y349.
9. Полиспецифическая полипептидная конструкция по п.8, отличающаяся тем, что аминокислотная модификация представляет собой мутацию заряда для повышения электростатической комплементарности полипептидов.
10. Полиспецифическая полипептидная конструкция по любому из пп.1-9, отличающаяся тем, что Fc-область содержит полипептид, содержащий модификацию Ile253Arg или His435Arg.
11. Полиспецифическая полипептидная конструкция по любому из пп.1-10, отличающаяся тем, что Fc-область содержит полипептид, содержащий по меньшей мере одну модификацию для усиления связывания FcRn.
12. Полиспецифическая полипептидная конструкция по п.11, отличающаяся тем, что модификация находится в положении, выбранном из группы, состоящей из Met252Y, Ser254T, Thr256E, Met428L, Met428V, Asn434S и их комбинаций.
13. Полиспецифическая полипептидная конструкция по любому из пп.1-12, отличающаяся тем, что первый полипептид гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 82, 86, 94 или 96, и второй полипептид гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 83, 87, 90, 92, 98 или 100.
14. Полиспецифическая полипептидная конструкция по любому из пп.1-13, отличающаяся тем, что Fc-область включает полипептид, содержащий по меньшей мере одну аминокислотную модификацию, которая уменьшает эффекторную функцию и/или уменьшает связывание с Fc-гамма-рецептором или C1q.
15. Полиспецифическая полипептидная конструкция по п.14, отличающаяся тем, что одна или более аминокислотных модификаций представляют собой делецию одного или более из Glu233, Leu234 или Leu235.
16. Полиспецифическая полипептидная конструкция по любому из пп.1-12, 14 и 15, отличающаяся тем, что первый полипептид гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 84, 88, 95 или 97, и второй полипептид гетеродимерной Fc-области содержит последовательность аминокислот, представленную в любой из SEQ ID NO: 85, 89, 91, 93, 99 или 101.
17. Полиспецифическая полипептидная конструкция по любому из пп.1-16, отличающаяся тем, что CD3-связывающая область является одновалентной.
18. Полиспецифическая полипептидная конструкция по любому из пп.1-17, отличающаяся тем, что линкер представляет собой расщепляемый линкер.
19. Полиспецифическая полипептидная конструкция по п.18, отличающаяся тем, что связывание CD3-связывающей области с CD3 существенно снижается, когда полиспецифическая полипептидная конструкция находится в нерасщепленном состоянии по сравнению с расщепленным состоянием.
20. Полиспецифическая полипептидная конструкция по п.18 или 19, отличающаяся тем, что расщепляемый линкер представляет собой полипептид, который функционирует в качестве субстрата для протеазы.
21. Полиспецифическая полипептидная конструкция по п.20, отличающаяся тем, что протеаза продуцируется иммунной эффекторной клеткой, опухолью или клетками, присутствующими в микроокружении опухоли.
22. Полиспецифическая полипептидная конструкция по п.20 или 21, отличающаяся тем, что протеаза выбрана из матриптазы, матриксной металлопротеазы (ММР), гранзима В и их комбинаций.
23. Полиспецифическая полипептидная конструкция по п.22, отличающаяся тем, что протеаза представляет собой гранзим B.
24. Полиспецифическая полипептидная конструкция по любому из пп.18-23, отличающаяся тем, что расщепляемый линкер содержит аминокислотную последовательность общей формулы P4 P3 P2 P1 ↓ P1' (SEQ ID NO: 150), где P4 представляет собой аминокислоту I, L, Y, M, F, V или A; P3 представляет собой аминокислоту A, G, S, V, E, D, Q, N или Y; P2 представляет собой аминокислоту H, P, A, V, G, S или T; P1 представляет собой аминокислоту D или E; и P1' представляет собой аминокислоту I, L, Y, M, F, V, T, S, G или A, необязательно аминокислотную последовательность общей формулы P4 P3 P2 P1 ↓ P1' (SEQ ID NO: 151), где P4 представляет собой аминокислоту I или L; P3 представляет собой аминокислоту E; P2 представляет собой аминокислоту P или A; P1 представляет собой аминокислоту D; и Р1' представляет собой аминокислоту I, V, T, S или G.
25. Полиспецифическая полипептидная конструкция по пп.18-24, отличающаяся тем, что расщепляемый линкер содержит аминокислотную последовательность IEPDI (SEQ ID NO: 136), LEPDG (SEQ ID NO: 152), LEADT (SEQ ID NO: 137), IEPDG (SEQ ID NO: 138), IEPDV (SEQ ID NO: 139), IEPDS (SEQ ID NO: 140), IEPDT (SEQ ID NO: 141) или LEADG (SEQ ID NO: 153).
26. Полиспецифическая полипептидная конструкция по любому из пп.18-25, отличающаяся тем, что расщепляемый линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 22, 105-112, 136-141, 148, 150-153.
27. Полиспецифическая полипептидная конструкция по п.22, отличающаяся тем, что протеаза представляет собой матриптазу.
28. Полиспецифическая полипептидная конструкция по любому из пп.18-27, отличающаяся тем, что расщепляемый линкер содержит последовательность P4QAR↓ (A/V) (SEQ ID NO: 154), где P4 представляет собой любую аминокислоту; необязательно последовательность RQAR (A/V) (SEQ ID NO: 155) или последовательность RQARV (SEQ ID NO: 156).
29. Полиспецифическая полипептидная конструкция по любому из пп.18-28, отличающаяся тем, что расщепляемый линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 23, 154-156.
30. Полиспецифическая полипептидная конструкция по п.22, отличающаяся тем, что протеаза представляет собой ММР; необязательно MMP-2.
31. Полиспецифическая полипептидная конструкция по любому из пп.18-30, отличающаяся тем, что расщепляемый линкер имеет общую формулу P3 P2 P1 ↓ P1' (SEQ ID NO: 157), где P3 представляет собой P, V или A; P2 представляет собой Q или D; P1 представляет собой A или N; и P1' представляет собой L, I или M, необязательно, общую формулу P3 P2 P1 ↓ P1' (SEQ ID NO: 158), где P3 представляет собой P; P2 представляет собой Q или D; P1 представляет собой A или N; и P1' представляет собой L или I, необязательно последовательность PAGL (SEQ ID NO: 24).
32. Полиспецифическая полипептидная конструкция по любому из пп.18-31, отличающаяся тем, что расщепляемый линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 22-31, 104-114, 117-118, 136-144, 148, 150-158.
33. Полиспецифическая полипептидная конструкция по пп.1-32, отличающаяся тем, что полиспецифическая полипептидная конструкция содержит по меньшей мере (i) первый полипептид, содержащий первый полипептид Fc гетеродимерной Fc-области, первый полипептидный линкер и домен VH CD3-связывающей области; и (ii) второй полипептид, содержащий второй полипептид Fc гетеродимерной Fc-области, второй полипептидный линкер и домен VL CD3-связывающей области, где один или оба из первого и второго полипептида содержат по меньшей мере один антигенсвязывающий домен, который связывается с опухолеспецифическим антигеном (TAA).
34. Полиспецифическая полипептидная конструкция по любому из пп.1-33, отличающаяся тем, что один или более антигенсвязывающих доменов, которые связываются с TAA, приводит к одновалентному, двухвалентному, трехвалентному или четырехвалентному связыванию с TAA.
35. Полиспецифическая полипептидная конструкция по п.33 или 34, отличающаяся тем, что
первый антигенсвязывающий домен расположен на амино-конце относительно Fc-области и/или на карбокси-конце относительно CD3-связывающей области первого или второго полипептида полиспецифической полипептидной конструкции; или
первый антигенсвязывающий домен расположен на амино-конце относительно Fc-области, и второй антигенсвязывающий домен расположен на карбокси-конце относительно CD3-связывающей области.
36. Полиспецифическая полипептидная конструкция по любому из пп.1-35, отличающаяся тем, что антигенсвязывающий домен или независимо каждый из антигенсвязывающих доменов связывается с TAA, выбранным из 1–92–LFA–3, 5T4, альфа–4 интегрина, альфа–V интегрина, альфа4бета1 интегрина, альфа4бета7 интегрина, AGR2, анти–Lewis–Y, рецептора апелина J, APRIL, B7–H3, B7–H4, BAFF, BTLA, компонента комплемента C5, C–242, CA9, CA19–9, (Lewis а), карбоангидразы 9, CD2, CD3, CD6, CD9, CD11a, CD19, CD20, CD22, CD24, CD25, CD27, CD28, CD30, CD33, CD38, CD40, CD40L, CD41, CD44, CD44v6, CD47, CD51, CD52, CD56, CD64, CD70, CD71, CD74, CD80, CD81, CD86, CD95, CD117, CD123, CD125, CD132, (IL–2RG), CD133, CD137, CD138, CD166, CD172A, CD248, CDH6, CEACAM5 (CEA), CEACAM6 (NCA–90), CLAUDIN–3, CLAUDIN–4, cMet, коллагена, Cripto, CSFR, CSFR–1, CTLA–4, CTGF, CXCL10, CXCL13, CXCR1, CXCR2, CXCR4, CYR61, DL44, DLK1, DLL3, DLL4, DPP–4, DSG1, EDA, EDB, EGFR, EGFRviii, рецептора эндотелина B (ETBR), ENPP3, EpCAM, EPHA2, EPHB2, ERBB3, F–протеина RSV, FAP, FGF–2, FGF8, FGFR1, FGFR2, FGFR3, FGFR4, FLT–3, рецептора альфа фолата (FR α), GAL3ST1, G–CSF, G–CSFR, GD2, GITR, GLUT1, GLUT4, GM–CSF, GM–CSFR, рецепторов GP IIb/IIIa, Gp130, GPIIB/IIIA, GPNMB, GRP78, HER2/neu, HER3, HER4, HGF, hGH, HVEM, гиалуронидазы, ICOS, IFNальфа, IFNбета, IFNгамма, IgE, рецептора IgE (FceRI), IGF, IGF1R, IL1B, IL1R, IL2, IL11, IL12, IL12p40, IL–12R, IL–12Rbeta1, IL13, IL13R, IL15, IL17, IL18, IL21, IL23, IL23R, IL27/IL27R (wsx1), IL29, IL–31R, IL31/IL31R, IL2R, IL4, IL4R, IL6, IL6R, рецептора инсулина, Jagged лигандов, Jagged 1, Jagged 2, KISS1–R, LAG–3, LIF–R, Lewis X, LIGHT, LRP4, LRRC26, Ly6G6D, LyPD1, MCSP, мезотелина, MRP4, MUC1, муцина–16 (MUC16, CA–125), Na/K–АТФазы, NGF, никастрина, рецепторов Notch, Notch 1, Notch 2, Notch 3, Notch 4, NOV, OSM–R, OX–40, PAR2, PDGF–AA, PDGF–BB, PDGFRальфа, PDGFRбета, PD–1, PD–L1, PD–L2, фосфатидилсерина, P1GF, PSCA, PSMA, PSGR, RAAG12, RAGE, SLC44A4, фосфат сфингозина 1, STEAP1, STEAP2, TAG–72, TAPA1, TEM–8, TGFбета, TIGIT, TIM–3, TLR2, TLR4, TLR6, TLR7, TLR8, TLR9, TMEM31, TNFальфа, TNFR, TNFRS12A, TRAIL–R1, TRAIL–R2, трансферрина, рецептора трансферрина, TRK–A, TRK–B, uPAR, VAP1, VCAM–1, VEGF, VEGF–A, VEGF–B, VEGF–C, VEGF–D, VEGFR1, VEGFR2, VEGFR3, VISTA, WISP–1, WISP–2 и WISP–3.
37. Полиспецифическая полипептидная конструкция по любому из пп.1-36, отличающаяся тем, что полиспецифическая полипептидная конструкция содержит по меньшей мере первый антигенсвязывающий домен и второй антигенсвязывающий домен, где первый антигенсвязывающий домен и второй антигенсвязывающий домен связываются с одним и тем же TAA.
38. Полиспецифическая полипептидная конструкция по любому из пп.1-37, отличающаяся тем, что полиспецифическая полипептидная конструкция содержит по меньшей мере первый антигенсвязывающий домен и второй антигенсвязывающий домен, где первый антигенсвязывающий домен и второй антигенсвязывающий домен связываются с разными TAA.
39. Полиспецифическая полипептидная конструкция по любому из пп.1-38, отличающаяся тем, что dsFv антитела против CD3 содержит:
VH, имеющую аминокислотную последовательность, которая имеет по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с SEQ ID NO: 44; и
VL, имеющую аминокислотную последовательность, которая имеет по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с SEQ ID NO: 72.
40. Полиспецифическая полипептидная конструкция по любому из пп.1-39, отличающаяся тем, что анти-CD3 dsFv содержит аминокислотную последовательность SEQ ID NO: 44 и аминокислотную последовательность SEQ ID NO: 72.
41. Полиспецифическая полипептидная конструкция по любому из пп.1-40, отличающаяся тем, что полиспецифическая полипептидная конструкция конъюгирована с агентом, необязательно, где агент представляет собой терапевтический агент, противоопухолевый агент, токсин или его фрагмент, детектируемый фрагмент или диагностический агент.
42. Полинуклеотид, кодирующий полиспецифическую полипептидную конструкцию по любому из пп.1-41.
43. Полинуклеотид для экспрессии полиспецифической полипептидной конструкции по любому из пп.1-41, где:
указанная полиспецифическая полипептидная конструкция содержит первый полипептид и второй полипептид;
указанный полинуклеотид содержит первую последовательность нуклеиновой кислоты, кодирующую указанный первый полипептид полиспецифической пептидной конструкции по любому из пп.1-41, и вторую последовательность нуклеиновой кислоты, кодирующую указанный второй полипептид полиспецифической полипептидной конструкции по любому из пп.1-41, где первая и вторая последовательности нуклеиновых кислот разделены участком внутренней посадки рибосомы (IRES) или нуклеиновой кислотой, кодирующей саморасщепляющийся пептид или пептид, который вызывает пропуск рибосомы.
44. Экспрессионный вектор, содержащий полинуклеотид по п.42 или 43.
45. Клетка для получения полиспецифической полипептидной конструкции, содержащая полинуклеотид по п.42 или 43 или экспрессионный вектор по п.44.
46. Способ получения полиспецифической полипептидной конструкции по любому из пп.1-41, включающий введение в клетку полинуклеотида по п.42 или 43 или экспрессионного вектора по п.44 и культивирование клетки в условиях для получения полиспецифической полипептидной конструкции.
47. Способ получения полиспецифической полипептидной конструкции по любому из пп.1-41, включающий культивирование клетки по п.45 в условиях, при которых полиспецифический полипептид продуцируется клеткой.
48. Фармацевтическая композиция для стимулирования или индукции иммунного ответа, содержащая полиспецифическую полипептидную конструкцию по любому из пп.1-41 и фармацевтически приемлемый носитель.
49. Способ стимулирования или индукции иммунного ответа, включающий контактирование клетки-мишени и Т-клетки с полиспецифической полипептидной конструкцией по любому из пп.1-41 или фармацевтической композицией по п.48, где указанная клетка-мишень представляет собой опухолевую клетку, экспрессирующую TAA, распознаваемый указанной полиспецифической полипептидной конструкцией.
50. Способ по п.49, отличающийся тем, что полиспецифическая полипептидная конструкция содержит расщепляемый линкер, который функционирует в качестве субстрата для протеазы, и индукция или стимулирование иммунного ответа усиливается в присутствии протеазы.
51. Способ по п.49 или 50, отличающийся тем, что протеаза выбрана из матриптазы, матриксной металлопротеазы (ММР), гранзима В и их комбинаций.
52. Способ по любому из пп.49-51, отличающийся тем, что контактирование осуществляется у пациента ex vivo, in vitro или in vivo.
53. Способ стимулирования или индукции иммунного ответа у пациента, имеющего опухоль или злокачественное новообразование, причем способ включает введение пациенту терапевтически эффективного количества полиспецифической полипептидной конструкции по любому из пп.1-41 или фармацевтической композиции по п.48, где клетки указанных опухоли или злокачественного новообразования экспрессируют опухолеспецифический антиген (TAA), распознаваемый указанной полиспецифической полипептидной конструкцией.
54. Способ лечения опухоли или злокачественного новообразования у пациента, где способ включает введение пациенту, нуждающемуся в этом, терапевтически эффективного количества полиспецифической полипептидной конструкции по любому из пп.1-41 или фармацевтической композиции по п.48, где клетки указанных опухоли или злокачественного новообразования экспрессируют опухолеспецифический антиген (TAA), распознаваемый указанной полиспецифической полипептидной конструкцией.
55. Способ по любому из пп.52-54, отличающийся тем, что указанным пациентом является человек.
| Токарный резец | 1924 |
|
SU2016A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Токарный резец | 1924 |
|
SU2016A1 |
| HOYOS et al | |||
| Парный рычажный домкрат | 1919 |
|
SU209A1 |
| Токарный резец | 1924 |
|
SU2016A1 |

