Область техники
Изобретение относится к области биотехнологии и молекулярной биологии, в частности генной инженерии, а именно к рекомбинантному вирусу везикулярного стоматита, экспрессирующему химерный белок IL12 - GMCSF.
Уровень техники
Высокий терапевтический потенциал онколитических вирусов раскрылся в последние 2 года в ходе исследований эффекта их применения в комбинации с различными ингибиторами контрольных точек иммунного ответа (immune checkpoint inhibitors) при лечении резистентных, неоперабельных или рецидивирующих форм рака [Russell L. et al. Oncolytic viruses: priming time for cancer immunotherapy // BioDrugs. - 2019. T. 33. №.5. C. 485-501]. Например, эффективность терапии значительно возрастала при использовании препарата knlygic® (talimogene laherparepvec) в комбинации с пембролизумабом, ниволумабом и ипилимумабом [Dummer R, Hoeller С, Gruter IP, Michielin О. Combining talimogene laherparepvec with immunotherapies in melanoma and other solid tumors. Cancer Immunol Immunother. 2017 Jun; 66(6):683-695. doi: 10.1007/s00262-017-1967-1]. В настоящее время разработка онколитических вирусов является одним из приоритетных направлений иммунотерапии опухолей. Ведется более 100 клинических исследований ранних фаз [Pearl Т.М. etal. Oncolytic virus-based cytokine expression to improve immune activity in brain and solid tumors // Molecular Therapy-Oncolytics. - 2019. - T. 13. - C. 14-21; Zheng M. et al. Oncolytic viruses for cancer therapy: barriers and recent advances // Molecular Therapy-Oncolytics. - 2019. T. 15. - C. 234-247].
Вирус везикулярного стоматита (VSV, везикуловирус, везикуловирус Индианы) является перспективным для создания на его основе безопасного и эффективного онколитика, в особенности в комбинации с иммунотерапевтическими подходами. Низкая патогенность (вирус поражает крупный домашний скот, а в случае заражения человека вирус чаще всего не вызывает никаких симптомов), репликация в цитоплазме (без интеграции в геном), возможность получения высокого вирусного титра при наработке VSV в культурах клеток млекопитающих (ВНК, НЕК) и простота выделения, легкость генетической модификации, отсутствие в популяции нейтрализующих антител сделали VSV идеальным кандидатом для получения вирусных вакцин и онколитических вирусов [Geisbert Т.W., Feldmann Н. Recombinant vesicular stomatitis virus-based vaccines against Ebola and Marburg virus infections // The Journal of infectious diseases. 2011. T. 204. №. suppl 3. C. S1075-S1081; Zemp F., Rajwani J., Mahoney D. J. Rhabdoviruses as vaccine platforms for infectious disease and cancer // Biotechnology and Genetic Engineering Reviews. - 2018. - T. 34. - №.1. - C. 122-138.; Velazquez-Salinas L. et al. Oncolytic recombinant vesicular stomatitis virus (VSV) is nonpathogenic and nontransmissible in pigs, a natural host of VSV // Human Gene Therapy Clinical Development. 2017. T. 28. №.2. C. 108-115]. Одним из способов модификации VSV является встраивание генов иммуностимулирующих факторов, что может помочь преодолеть основные механизмы устойчивости опухолевых клеток к иммуноопосредованному уничтожению, значительно увеличив эффективность иммунотерапии злокачественных новообразований.
В последние годы одной из исследуемых стратегий стала вставка в онколитические вирусы иммуностимулирующих трансгенов (например, GM-SCF [Leveille S, Goulet ML, Lichty BD, et al. Vesicular Stomatitis Virus Oncolytic Treatment Interferes with Tumor-Associated Dendritic Cell Functions and Abrogates Tumor Antigen Presentation. J Virol. 2011; 85(23): 12160-12169; Ramsburg E, Publicover J, Buonocore L, et al. A vesicular stomatitis virus recombinant expressing granulocyte-macrophage colony-stimulating factor induces enhanced T-cell responses and is highly attenuated for replication in animals. J Virol. 2005; 79(24): 15043-15053]) для ускорения созревания дендритных клеток и презентации антигена. Кроме того, были предприняты попытки увеличения созревания и дифференцировки Т-клеток с использованием иммуномодулирующих трансгенов (например, IL-12, [Shin EJ, Wanna GB, Choi В, et al. Interleukin-12 expression enhances vesicular stomatitis virus oncolytic therapy in murine squamous cell carcinoma. Laryngoscope. 2007 Feb; 117(2):210-214]). Однако необходима дальнейшая работа с такими подходами для достижения устойчивости ответа и минимизации нецелевых эффектов, таких как трансгены, которые могут влиять как на здоровые, так и на раковые клетки [Munis AM, Bentley ЕМ & Takeuchi Y. A tool with many applications: vesicular stomatitis virus in research and medicine, Expert Opinion on Biological Therapy, 2020; 20:10, 1187-1201].
Перспективными мишенями для направленной модификации VSV являются IL12 и GM-CSF. ИЛ-12 это цитокин с плейотропной (многофункциональной) активностью. ИЛ-12 стимулирует дифференцировку Thl-клеток, повышает активность Т- и NK-клеток, ингибирует и/или перепрограммирует иммуносупрессивные клетки, например, опухоль-ассоциированные макрофаги и миелоидные супрессорные клетки (миелоидные супрессоры). Кроме того, ИЛ-12 стимулирует выработку интерферона гамма (IFNy), который, в свою очередь, оказывает цитостатическое/цитотоксическое/антиангиогенное действие и повышает уровень экспрессии главного комплекса гистосовместимости классов I и II (ГКГС классов I и II) на раковых клеток, что способствует улучшению их распознавания лимфоцитами [Toda М, Martuza RL, Kojima Н, Rabkin SD. In situ cancer vaccination: an IL-12 defective vector/replication-competent herpes simplex virus combination induces local and systemic antitumor activity. J Immunol. 1998 May 1; 160(9):4457-64. PMID: 9574551.; Nguyen KG, Vrabel MR, Mantooth SM, et al. Localized Interleukin-12 for Cancer Immunotherapy. Front Immunol. 2020 Oct 15; 11:575597. doi: 10.3389/fimmu.2020.575597. PMID: 33178203; PMCID: РМС7593768].
GM-CSF является фактором роста, который стимулирует дифференцировку, пролиферацию и миграцию миелоидных клеток. Потенциальный положительный эффект от экспрессии GM-CSF в составе вируса простого герпеса человека был продемонстрирован в многочисленных клинических исследованиях онколитика T-VEC (талимоген, лагерпарепвек), который имеет статус первого онколитического вируса, одобренного Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA).
Включение генов IL12 и GM-CSF в состав VSV для ко-экспрессии в ходе репликации вируса в опухоли может привести к активации и пролиферации дендритных клеток и макрофагов, индукции экспрессии IL-12 and INFy внутри опухоли, активации и амплификация Т-клеток и NK- клеток и их инфильтрации в ТМЕ и индукции экспрессии провоспалительных цитокинов (IFN-g, TNF и др.).
Однако следует учитывать, что, в отличие от многих других вирусов, на репликацию везикуловируса и соответственно онколитическую способность, а также на уровень экспрессии добавляемого трансгена и соответственно на суммарный терапевтический эффект могут влиять позиция трансгена в геноме вируса, состав трансгена, а также способы модификации генома VSV [Wertz GW, Moudy R, Ball LA. Adding genes to the RNA genome of vesicular stomatitis virus: positional effects on stability of expression. J Virol. 2002 Aug; 76(15):7642-50. doi: 10.1128/jvi.76.15.7642-7650.2002]. Это связано с особенностью репликативного цикла VSV. Например, одним из способов аттенюации вируса является изменение последовательности генов в геноме VSV [Flanagan ЕВ, Zamparo JM, Ball LA, Rodriguez LL, Wertz GW. Rearrangement of the genes of vesicular stomatitis virus eliminates clinical disease in the natural host: new strategy for vaccine development. J Virol. 2001 Jul; 75(13):6107-14. doi: 10.1128/JVI.75.13.6107-6114.2001; Wertz GW, Perepelitsa VP, Ball LA. Gene rearrangement attenuates expression and lethality of a nonsegmented negative strand RNA virus. Proc Natl Acad Sci USA. 1998 Mar31; 95(7):3501-6. doi: 10.1073/pnas.95.7.3501].
Одним из способов преодоления возможного негативного влияния добавленного гена может быть создание химерных конструкций, в которых одна рамка считывания кодирует два полипептида, связанные через линкер. Химерный белок IL12-GM-CSF и его противоопухолевый эффект описан в уровне техники [RU2263118C2], однако неизвестно, как встраивание гена, кодирующего этот химерный белок, отразится на биологических и терапевтических свойствах VSV. В частности, неизвестно, насколько добавление GM-CSF влияет на активность IL12, будет ли активность, например, IL12 сопоставима с активностью белка без добавления GM-CSF в составе экспрессионного вектора и в составе генома VSV и как будет влиять длина линкера между генами IL12 и GM-CSF на активность и уровень синтеза белка и т.д.
Из патента RU2263118C2 (Лексиген Фармасьютикэлс Корп) известен слитый белок p40-GM-CSF, мышиный и человеческий, который применяется для активации иммунного ответа при лечении опухолевых заболеваний в виде белка, слитого с антителом или Fc-фрагментом антитела. Последовательность гена, кодирующего слитый белок по патенту RU2263118C2, приведена в Seq Id No: 16.
Из работы Шин с соавторами известен онколитический вирусный вектор VSV (WSV-IL12) для экспрессии гена мышиного интерлейкина 12 (IL12), который эффективно экспрессирует IL12 и значительно улучшает лечение карциномы головы и шеи у мышей. Этот цитокин-несущий онколитический сравнивали с аналогичным фузогенным вирусом (rVSV-F) не содержащим цитокин при лечении мышиной плоскоклеточной карциномы SCC VII (SCC). Оба вируса продемонстрировали одинаковую инфекционную эффективность, репликацию вируса и цитотоксичность in vitro и in vivo. В модели ортотопического дна ротовой полости SCC VII у иммунокомпетентных мышей С3Н/HeJ множественные внутриопухолевые инъекции каждого вируса вызывали значительное уменьшение объема опухоли по сравнению с инъекциями только физиологического раствора. Опухоли, обработанные rVSV-IL12, показали поразительное уменьшение объема опухоли по сравнению с опухолями, обработанными rVSV-F и физиологическим раствором (Р<0,005). Это поразительное уменьшение объема опухоли привело к существенному улучшению выживаемости у животных, получавших rVSV-IL12. Никакой связанной с лечением токсичности не наблюдалось ни в одной из групп [Shin EJ, Wanna GB, Choi В, et al. mterleukin-12 expression enhances vesicular stomatitis virus oncolytic therapy in murine squamous cell carcinoma. Laryngoscope. 2007 Feb; 117(2):210-4. doi: 10.1097/01.mlg.0000246194.66295.d8].
В работе Рамсбург с соавторами описан рекомбинантный вирус везикулярного стоматита (VSV-GMCSF1), экспрессирующий высокий уровень GM-CSF с первой позиции в вирусном геноме, который сильно ослаблен с точки зрения вирусной диссеминации и патогенеза после интраназальной доставки мышам. Первичный ответ Т-клеток CD8 и нейтрализующих антител на VSV-GMCSF1 эквивалентен ответу, который вызывает контрольный вирус VSV-EGFP1, несущий GFP, в то время как ответ CD8 Т-клеток памяти был усилены у мышей, инфицированных VSV-GMCSF1 [Ramsburg Е, Publicover J, Buonocore L, et al. A vesicular stomatitis virus recombinant expressing granulocyte-macrophage colony-stimulating factor induces enhanced T-cell responses and is highly attenuated for replication in animals. J Virol. 2005 Dec; 79(24): 15043-53. doi: 10.1128/JVI. 79.24.15043-15053.2005]
В работе Бергман с соавторами описан рекомбинантный реплицирующийся VSV (rrVSV), нацеленный на клетки рака молочной железы, экспрессирующий мышиный GM-CSF. При лечении перитонеальных опухолевых имплантатов клеток D2F2/E2, линии клеток опухоли молочной железы мыши BALB/c нацеленный rrVSV устраняет перитонеальные имплантаты опухоли, экспрессирующей Her2/neu, и вызывает противоопухолевый Т-клеточный иммунологический ответ [Bergman I, Griffin JA, Gao Y, et al. Treatment of implanted mammary tumors with recombinant vesicular stomatitis virus targeted to Her2/neu. Int J Cancer. 2007 Jul 15; 121(2):425-30. doi: 10.1002/ijc.22680].
В работе Лемей с соавторами описан рекомбинантный вирус везикулярного стоматита VSV-A51, в который клонирован ген, кодирующий GM-CSF, чтобы усилить иммуностимулирующие свойства онколитического вируса (VSV-A51-GMCSF). VSV-A51-GMCSF предотвращал приживление опухоли B16-F10 у более чем 95% протестированных мышей, однако он не продемонстрировал повышенной эффективности на подкожной модели B16-F10 по сравнению с вирусом VSV-A51-GFP [Lemay CG, Rintoul JL, Kus A, et al. Harnessing oncolytic virus-mediated antitumor immunity in an infected cell vaccine. Mol Ther. 2012 Sep; 20(9): 1791-9. doi: 10.1038/mt.2012.128].
Для создания эффективного онколитика на основе VSV необходимо, с одной стороны, обеспечить безопасность терапии, например, модификацией М гена в геноме VSV или иным способом, а с другой стороны - усилить онколитические свойства вируса и иммуностимулирующий эффект посредством комбинации с одним или более иммуностимулирующими факторами или путем ко-экспрессии генов иммуностимулирующих факторов, встроенных в геном VSV. Следует отметить, что инициация исследований, использующих онколитические вирусы с более чем одним дополнительным геном в геноме говорит о перспективности такого подхода [https://replimune.com/pipeline/, дата обращения 26.12.2022]. Для преодоления опухолевого ингибирующего микроокружения целесообразно задействовать одновременно несколько механизмов иммунного ответа. При этом желательно минимизировать количество генов в геноме VSV для сохранения репликативной активности вируса, что можно сделать с помощью использования химерных конструкций - например IL12-GM-CSF. Модификация VSV должна сохранять цитопатический эффект вируса, а закодированная последовательность химерного гена должна при этом обеспечивать синтез функционального химерного белка. При создании рекомбинантного VSV, который модифицирован иммуностимулирующими факторами, необходимо доказывать эффективность онколитика не только на мышиных моделях с использованием последовательностей генов мыши, но и работоспособность конструкции на клетках человека, чтобы гарантировать эффективность противораковой виротерапии.
Технической задачей, на решение которой направлено настоящее изобретение, является решение как минимум одной из вышеуказанных в уровне техники проблем.
Сущность изобретения
Технической решением является использование описанного признаками в пунктах формулы изобретения.
Одной из возможных технических задач, на решение которой может быть направлено настоящее изобретение, являлось создание средства для иммунотерапии опухолей, в частности создание онколитического вируса, дополнительно экспрессирующего в клетках опухоли цитокины.
Задача может решаться тем, что создан рекомбинантный вирус везикулярного стоматита, экспрессирующий химерный белок IL12-GMCSF, при этом IL12 и GMCSF в химерном белке соединены посредством аминокислотного линкера.
В варианте осуществления изобретения химерный белок IL12-GMCSF содержит аминокислотную последовательность SEQ Id No.: 6. В другом варианте осуществления изобретения химерный белок IL12-GMCSF содержит аминокислотную последовательность SEQ Id No.: 7.
Технический результат может достигаться тем, что рекомбинантный вирус везикулярного стоматита содержит в геноме последовательность SEQ Id No.: 10. В еще одном варианте осуществления изобретения рекомбинантный вирус везикулярного стоматита содержит в геноме последовательность SEQ Id No.: 11
Фигуры
Изобретение иллюстрируются следующими графическими материалами:
На Фиг. 1 представлена структурная карта плазмиды, содержащей последовательность SEQ Id No: 8, для транзиентной (временной) экспрессии генов в клетках млекопитающих.
На Фиг. 2 представлена структурная карта плазмиды, содержащей последовательность SEQ Id No: 9, для транзиентной (временной) экспрессии генов в клетках млекопитающих.
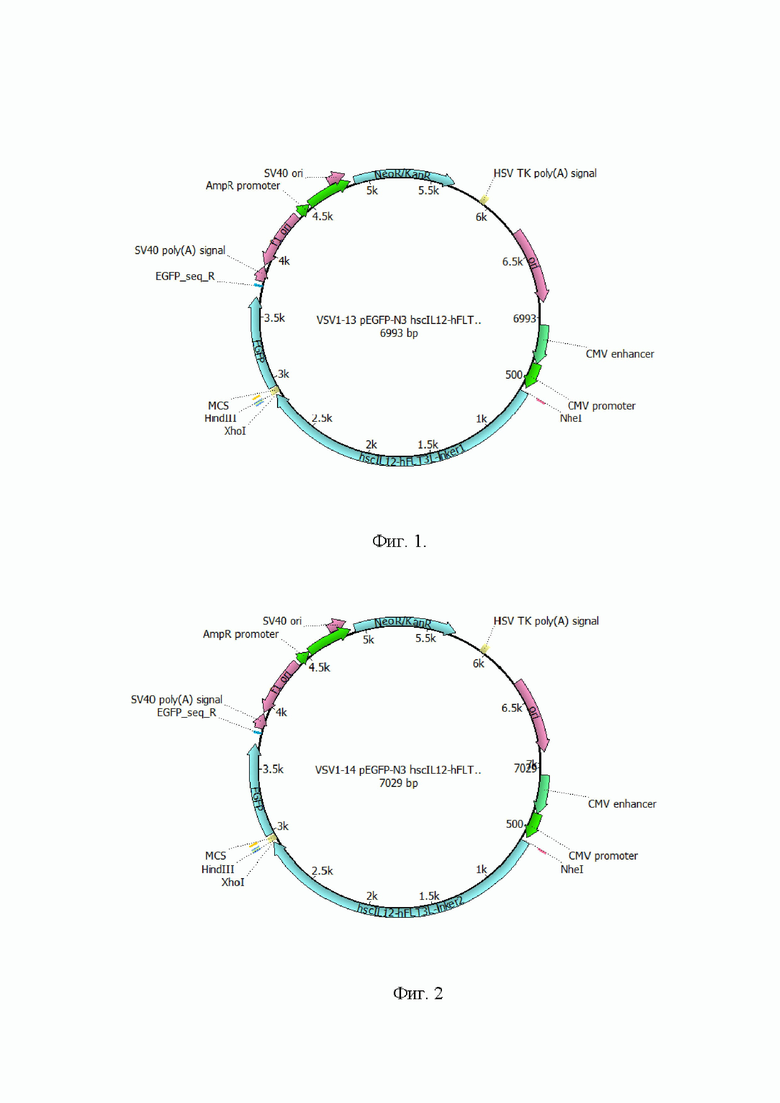
На Фиг. 3 представлена структурная карта плазмиды, содержащей последовательность SEQ Id No: 10, для транзиентной (временной) экспрессии генов в клетках млекопитающих.
На Фиг. 4 представлена структурная карта плазмиды, содержащей последовательность SEQ Id No: 11, для транзиентной (временной) экспрессии генов в клетках млекопитающих.
На Фиг. 5 представлена структурная карта плазмиды pEGFP-N3 для транзиентной (временной) экспрессии гена GFP в клетках млекопитающих.
На Фиг. 6 представлена микрофотография монослоя клеток HEK293TN, трансфицированных плазмидой pEGFP (положительный контроль).
На Фиг. 7 представлена структурная карта плазмиды pEGFP-N3-mscIL12-mGM-CSF.
На Фиг. 8 представлена структурная карта плазмиды pVSV-G-GFP-dM51.
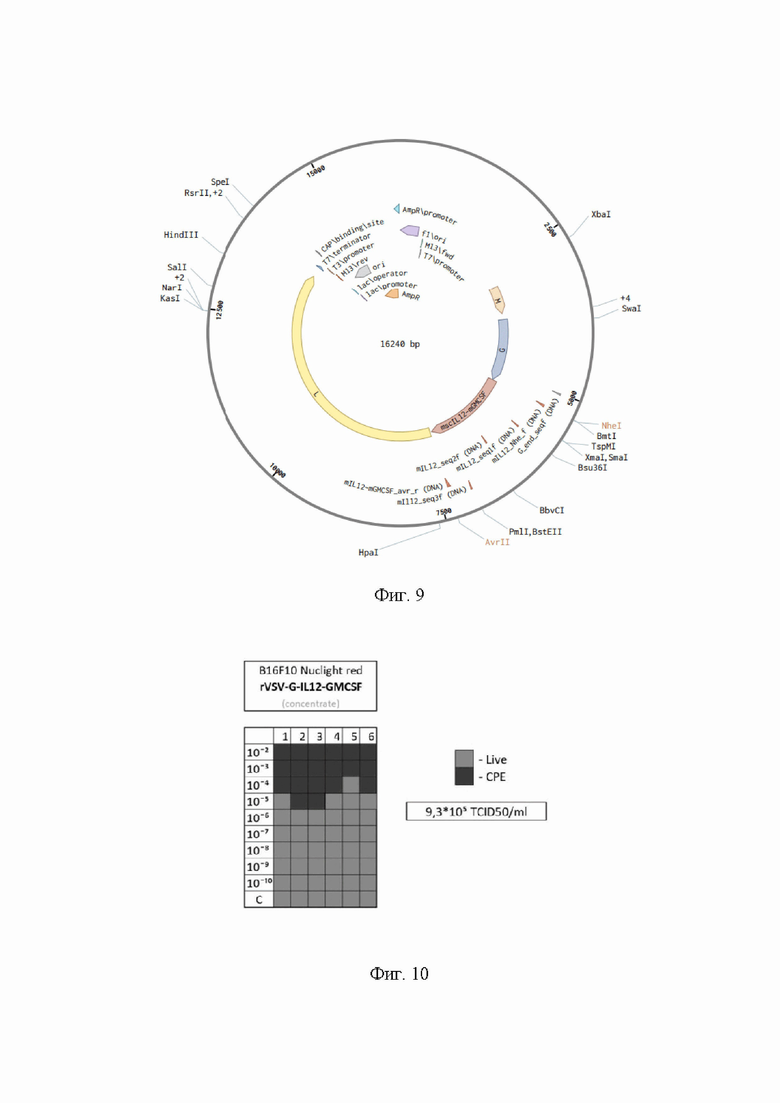
На Фиг. 9 представлена структурная карта плазмиды pVSV-G-dM51-mIL12-mGMCSF, содержащей последовательность SEQ ID NO: 15.
На Фиг. 10 представлена визуализация ЦПЭ VSV-mIL12-mGMCSF и конечный титр вируса в кондиционированной культуральной жидкости.
На Фиг. 11 представлены результаты подтверждения наличия химерного гена, встроенного в геном VSV-mIL12-mGMCSF с помощью электрофореза в ПААГ и ПЦР.
А - фотография ПААГ с образцом неконцентрированного и концентрированного VSV-G-GFP (положительный контроль ПААГ)
Б фотография геля с нанесенным концентратом VSV-mIL12-mGMCSF (справа от маркера) и кондиционированной культуральной жидкостью клеток (правее)
В - фотография агарозного геля с нанесенными ПЦР-продуктами образцов VSV-mIL12-mGMCSF.
На Фиг. 12 представлена диаграмма значений оптической плотности в лунках культурального планшета. Blank фон, DMEM отрицательный контроль (среда DMEM), VSV-GFP - везикуловирус без трансгена, IL12 - положительный контроль (препарат белка IL12), 12 well кондиционированная среда после первой трансфекции в 12-луночном планшете, Т25 кондиционированная среда после второй трансфекции в культуральном флаконе Т25, Т150 - кондиционированная среда после второй трансфекции в культуральном флаконе Т150, ×1 - без разведения, ×0,5 - разведение в 2 раза, хОД - разведение в 10 раз.
На Фиг. 13 представлены фотографии мышей до (слева) и после (справа) введения VSV-mIL12-mGM-CSF. Место введения отмечено кругом. Наблюдается лизис опухоли, вызванной инъекцией B16F10 клеток.
Детальное описание изобретения
Если не указано иначе, предполагается, что все термины, обозначения и другие научные термины, используемые в данной заявке, имеют значения, которые обычно понимают специалисты в области, к которой относится настоящее изобретение. В некоторых случаях определения терминов с общепринятыми значениями приведены в данной заявке для ясности и/или для быстрой справки и понимания, и включение таких определений в настоящее описание не должно истолковываться как наличие существенного отличия значения термина от обычно подразумеваемого в данной области.
Кроме того, если по контексту не требуется иное, термины в единственном числе включают в себя термины во множественном числе, и термины во множественном числе включают в себя термины в единственном числе. Как правило, используемая классификация и методы культивирования клеток, молекулярной биологии, иммунологии, микробиологии, генетики, аналитической химии, химии органического синтеза, медицинской и фармацевтической химии, а также гибридизации и химии белка и нуклеиновых кислот, описанные в настоящем документе, хорошо известны специалистам и широко применяются в данной области. Ферментативные реакции и способы очистки осуществляют в соответствии с инструкциями производителя, как это обычно осуществляется в данной области, или как описано в настоящем документе.
В одном из вариантов осуществления настоящего изобретения предлагается вирус везикулярного стоматита, экспрессирующий химерный белок IL12-GMCSF. Термин «экспрессирующий» обозначает, что вирусный геном содержит генетическую конструкцию, которая при инфицировании упомянутым вирусом клеток служит матрицей для трансляции целевого полипептидного, или белкового, продукта. Генетическая конструкция сконструирована и внедрена в геном вируса таким образом, чтобы, с одной стороны, не препятствовать процессу репликации (амплификации) вируса, с другой стороны эффективно экспрессироваться в инфицированной клетке. При этом IL12 и GMCSF в химерном белке соединены посредством аминокислотного линкера.
В варианте осуществления изобретения химерный белок IL12-GMCSF содержит аминокислотную последовательность SEQ Id No.: 6. В другом варианте осуществления изобретения химерный белок IL12-GMCSF содержит аминокислотную последовательность SEQ Id No.: 7. Химерные белки, содержащие последовательность SEQ Id No.: 6 или SEQ Id No.: 7, синтезируются в клетках, инфицированных везикуловирусами по настоящему изобретению. При этом химерные белки проявляют свою функциональность, демонстрируя способность активировать компетентные клетки in vitro. Экспериментально показано, что тип и длина линкера не влияют на функциональную активность hIL12 в составе химерного белка с GM-CSF, при этом и уровень экспрессии, и активность hIL12 в клеточном тесте в составе химеры с GM-CSF сопоставимы с уровнем положительных контролей hIL12.
Технический результат может достигаться тем, что рекомбинантный вирус везикулярного стоматита содержит в геноме последовательность SEQ Id No.: 10. В еще одном варианте осуществления изобретения рекомбинантный вирус везикулярного стоматита содержит в геноме последовательность SEQ Id No.: 11. rVSV, содержащие в геноме последовательность SEQ Id No.: 10 или SEQ Id No.: 11, демонстрируют стабильную экспрессию химерного белка, обладающего цитокиновой активностью.
Также, in vivo показано, что рекомбинантный вирус везикулярного стоматита по настоящему изобретению обладает выраженной противоопухолевой активностью в животной модели опухолевого ксенотрансплантата. Онколитическая способность рекомбинантного вируса везикулярного стоматита по настоящему изобретению превышала онколитическую способность аналогичного вируса, не экспрессирующего химерный IL12-GMCSF, в 1000 раз.
Сущность и промышленная применимость изобретения поясняются следующими примерами:
Пример 1. Получение плазмид с химерными генами.
При получении новых плазмид и синтезе конструкций использовали следующие методы, материалы и оборудование. Для выравнивания нуклеотидных последовательностей применяли программу UniPro UGENE. Трансляцию нуклеотидных последовательностей в аминокислотную проводили при помощи инструмента EMBOSS Transeq [https://www.ebi.ac.uk/Tools/st/emboss_transeq/, дата обращения 21.12.2022]. Последовательности генов для модификации генома VSV синтезировали при помощи сервиса TopGenetech (Канада). Клонирование последовательностей, дополнительные модификации проводили с помощью ПЦР и синтезированных in house олигонуклеотидов. При работе с плазмидной ДНК и получение новых плазмид использовали стандартные протоколы генетической инженерии, перечисленные далее. Для трансформации и амплификации плазмид использовали бактериальные штаммы Е. coli (DH5-alpha, NEB® Stable, XLIO-Gold®). Генетическую трансформацию, гидролиз эндонуклеазами рестрикции, лигирование и гель-электрофорез проводили в стандартных условиях [Green, М. and Sambrook, J. (2012) Molecular Cloning: A Laboratory Manual. 4th Edition, Vol.II, Cold Spring Harbor Laboratory Press, New York]. Плазмидную ДНК из бактериальных клеток Е. coli и линеаризованные фрагменты ДНК из агарозных гелей выделяли коммерческими наборами «Plasmid Miniprep», «Оеапир Mini» (ЗАО Евроген, Россия) и «Plasmid mini/midi Kits» (QIAGEN Sciences Inc, США). Плазмиды проверяли с помощью рестрикционного анализа и последующего электрофореза в агарозном геле и методом секвенирования по Сэнгеру. Для внесения мутации в последовательности генов использовали методику инвертированной ПЦР (reverse PCR). Полученный ПЦР-продукт вырезали из геля, обрабатывали Т4 полинуклеотид киназой, и самолигировали фрагмент на самого себя. Полученную лигазную смесь использовали для трансформации клеток Е. coli. Для ПЦР-амплификации фрагментов использовали полимеразу Q5 (NEB). Реакционную смесь готовили согласно рекомендациям производителя. Продукты реакции визуализировали и разделяли электрофоретически в 1% агарозном геле. Полученные ПЦР-фрагменты вырезали из геля, выделили и очистили с помощью набора Cleanup Standard (Евроген). Для анализа колоний-трансформантов проводили ПЦР-скрининг на колониях с использованием праймеров для секвенирования и набора Screen mix (Евроген). Реакционную смесь готовили согласно рекомендациям производителя. Продукты реакции визуализировали и разделяли электрофоретически в 1% агарозном геле. Секвенирование по Сэнгеру осуществляли на амплификаторе ABI 3500 Genetic Analyzer (Applied Biosystems, США) при стандартных условиях и с использованием реагентов, рекомендованных производителем Applied Biosystems. Олигонуклеотидные последовательности проверяли OligoAnalyzer™ Tool [https://www.idtdna.com/pages/tools/oligoanalyzer, дата обращения 21.12.2022]. Длину продукта оценивали с помощью выравнивания на геномную ДНК GRch38/hg38 с помощью Genome Browser [https://genome.ucsc.edu/, дата обращения 21.12.2022].
Для проверки влияния типа линкера и сохранения активности белков в составе химеры, в которой первым компонентом является IL12 создали 4 плазмиды, содержание химеры hIL12-hFlt3L и hIL12-hGM-CSF, где FLT3L - это лиганд fms-подобной тирозинкиназы 3, основного фактора роста дендритных клеток.
Для сборки химерного гена hIL12-hFlt3L и hIL12-hGM-CSF использовали последовательности синтезированных генов hFlt3L (SEQ Id No: 1). hIL12 (SEQ Id No: 2), hGM-CSF (SEQ Id No: 3).
В целевых конструкциях использовали классический глицин-сериновый линкер:
- Длинный GGGGS GGGGS GGGGSGGGGS GS (22 АК)
- Короткий GGGGS GGGGS (10 АК)
В соответствии с литературными данными более длинный стандартный глицин-сериновый линкер теоретически должен обеспечивать большую подвижность молекул химеры друг относительно друга и соответственно большую биологическую активность химерного белка [Chen X, Zaro JL, Shen WC. Fusion protein linkers: property, design and functionality. Adv Drug Deliv Rev. 2013 Oct; 65(10): 1357-69. doi: 10.1016/j.addr.2012.09.039]. Для оценки влияния длины линкера был также использован более короткий вариант глицин серинового линкера.
Целевые аминокислотные последовательности химерных белков (SEQ Id No: 4) и (SEQ Id No: 5) содержали короткий и длинный линкеры между полипептидом IL12 и Flt3L соответственно.
Целевые аминокислотные последовательности химерных белков (SEQ Id No: 6) и (SEQ Id No: 7) содержали короткий и длинный линкеры между полипептидом субъединицы IL12 и GM-CSF соответственно.
А. Сборку и клонирование последовательности, кодирующей hIL12-hFlt3L с коротким линкером hscIL12-(GS)short-Flt3L проводили по следующей схеме.
1) Проводили ПЦР амплификацию фрагмента гена hIL12 с помощью специфических праймеров:
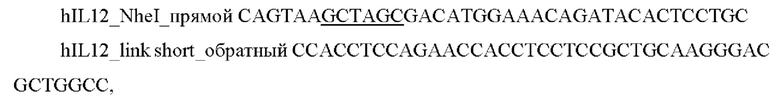
и выделяли фрагмент размером 1600 п.о.
2) Проводили ПЦР амплификацию фрагмента гена hFlt3L с помощью специфических праймеров:
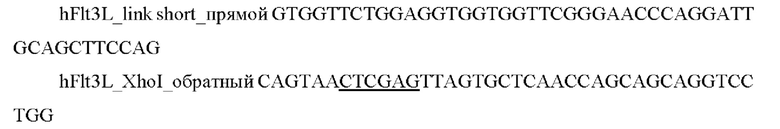
и выделяли фрагмент размером 630 п.о.
3) Проводили отжиг амплифицированных фрагментов друг на друга с последующей ПЦР амплификацией последовательности hscIL12-(GS)short-hFlt3L с использованием праймеров hILl2_Nhel прямой и hFlt3L_XhoI_обратный.
4) Проводили встраивание ПЦР продукта (~2200 п.о.) в вектор pEGFP-N3 по сайтам Nhel/Xhol по стандартным методикам рестриктазно-лигазного клонирования.
5) Проводили анализ полученной конструкции с помощью электрофореза в агарозном геле и секвенированием. Карта полученного вектора представлена на Фиг. 1. Результаты секвенирования подтвердили наличие в векторе целевой последовательности, кодирующей hIL12-(GS)short-hFlt3L (SEQ Id No: 8).
Б. Сборку и клонирование последовательности, кодирующей hscIL12-hFlt3L с длинным линкером hIL12-(GS)long-hFlt3L проводили по следующей схеме.
1) Проводили ПЦР амплификацию фрагмента гена hIL12 с помощью специфических праймеров:
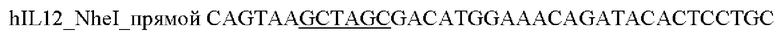
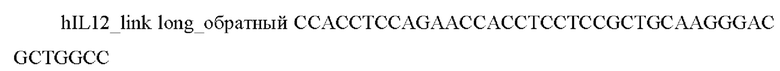
и выделяли фрагмент размером 1600 п.о.
2) Проводили ПЦР амплификацию фрагмента гена hFlt3L с помощью специфических праймеров:
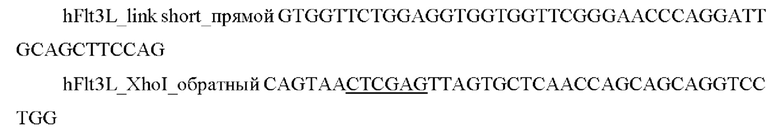
и выделяли фрагмент размером 630 п.о.
3) Проводили сборку фрагмента, содержащего линкер, с использованием коротких олигонуклеотидов (отжиг)

Выделяли целевой фрагмент с последовательностью:
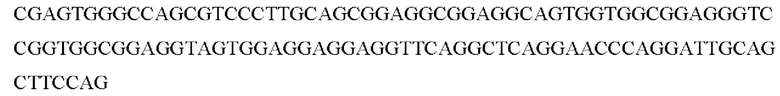
4) Проводили отжиг наработанных трех фрагментов друг на друга (п. 1-3) с последующей ПЦР амплификацей последовательности hscIL12-(GS)long-hFkt3L с использованием праймеров hTL12_NheT прямой и hFlt3L_XhoI_обратный.
5) Проводили встраивание ПЦР продукта (~2350 п.о.) в вектор pEGFP-N3 по сайтам Nhel/Xhol по стандартным методикам рестриктазно-лигазного клонирования.
6) Проводили анализ полученной конструкции с помощью электрофореза в агарозном геле и секвенированием. Карта полученного вектора представлена на Фиг. 2. Результаты секвенирования подтвердили наличие в векторе целевой последовательности, кодирующей hIL12-(GS)long-hFlt3L (SEQ Id No: 9)
В. Сборку и клонирование последовательности, кодирующей hIL12-hGM-CSF с коротким линкером hscIL12-(GS)short-hGM-CSF проводили по следующей схеме.
1) Проводили ПЦР амплификацию фрагмента гена hIL12 с помощью специфических праймеров:
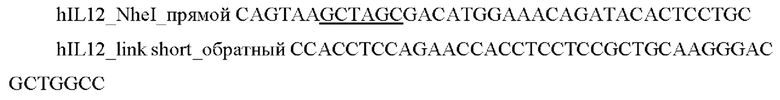
и выделяли фрагмент размером 1600 п.о.
2) Проводили ПЦР амплификацию фрагмента гена hGM-CSF с помощью специфических праймеров:
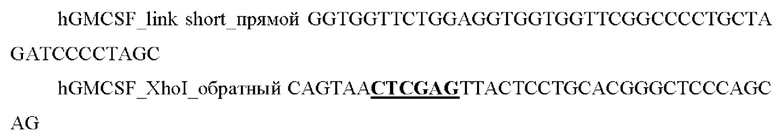
и выделяли фрагмент размером 400 п.о.
3) Проводили отжиг наработанных фрагментов друг на друга с последующей ПЦР амплификацей последовательности hscIL12-(GS)short-hGM-CSF с использованием праймеров ML12 Nhel прямой и hGMCSF_XhoI_обратный.
4) Проводили встраивание ПЦР продукта (~2000 п.о.) в вектор pEGFP-N3 по сайтам Nhel/Xhol по стандартным методикам рестриктазно-лигазного клонирования.
5) Проводили анализ полученной конструкции с помощью электрофореза в агарозном геле и секвенированием. Карта полученного вектора представлена на Фиг. 3. Результаты секвенирования подтвердили наличие в векторе целевой последовательности, кодирующей hIL12-(GS)short-hGM-CSF (SEQ Id No: 10)
Г. Сборку и клонирование последовательности, кодирующей hIL12-hGM-CSF с длинным линкером hIL12-(GS)long-hGM-CSF проводили по следующей схеме.
1) Проводили ПЦР амплификацию фрагмента гена hIL12 с помощью специфических праймеров:
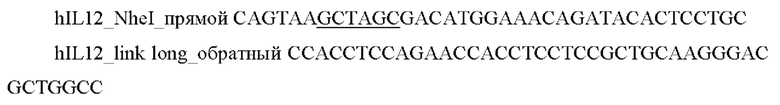
и выделяли фрагмент размером 1600 п.о.
2) Проводили ПЦР амплификацию фрагмента гена hGM-CSF с помощью специфических праймеров:
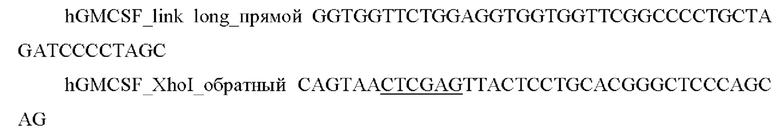
и выделяли фрагмент размером 400 п.о.
3) Проводили сборку фрагмента, содержащего линкер, с использованием коротких олигонуклеотидов (отжиг)

Выделяли целевой фрагмент с последовательностью:
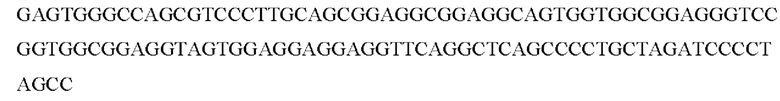
4) Проводили отжиг наработанных трех фрагментов друг на друга (п. 1-3) с последующей ПЦР амплификацей последовательности hscIL12-(GS)long-hGM-CSF с использованием праймеров hlLl2_Nhel прямой и hGMCSF_XhoI_обратный.
5) Проводили клонирование ПЦР продукта (~2150 п.о.) в вектор pEGFP-N3 по сайтам Nhel/Xhol по стандартным методикам рестриктазно-лигазного клонирования.
6) Проводили анализ полученной конструкции с помощью электрофореза в агарозном геле и секвенированием. Карта полученного вектора представлена на Фиг. 4. Результаты секвенирования подтвердили наличие в векторе целевой последовательности, кодирующей hscIL12-(GS)long-hGM-CSF (SEQ Id No: 11)
Пример 2. Проведение трансфекции адгезионной клеточной культуры HEK293TN
Для анализа уровня экспрессии химерных белков и их активности проводили трансфекцию клеточной культуры HEK293TN с помощью липофектамина (Lipofectamin 3000 Reagent).
В первый день подготавливали клеточную культуру HEK293TN (адгезионная) к трансфекции. Для проведения трансфекции адгезионную клеточную культуру HEK293TN субкультивировали в посевной дозе 1,25⋅104-3,75⋅104 клеток/см2 в полной ростовой среде в течение 3-4 суток до достижения уровня конфлюэнтности 70-90%. Смену среды в культуральных флаконах осуществляли раз в 2-3 дня. Избегали образования монослоя клеток в культуральных флаконах выше 90%. Флакон с клеточной культурой инкубировали в СО2-инкубаторе при температуре (37±1)°С в увлажненном воздухе, содержащем 5% диоксида углерода. Перед проведением трансфекции готовили клеточную суспензию HEK293TN, для этого использовали клетки с одного культурального флакона площадью 25 см2. Клетки HEK293TN, засеянные за 3-4 дня до постановки анализа во флаконы 25 см2 с начальной концентрацией 1,25⋅104-3,75⋅104 клеток/см2 в полной ростовой среде и инкубированные при температуре (37±1)°С в увлажненном воздухе с содержанием углерода диоксида 5%, оценивали визуально под микроскопом. Отбирали с помощью серологической пипетки среду в стерильную центрифужную пробирку вместимостью 50 мл. В культуральный флакон вносили 3 мл раствора PBS, промывали монослой клеток, аккуратно покачивая флакон, находящийся в горизонтальном положении, в течение 3-5 с. Отбирали PBS серологической пипеткой и отбрасывали раствор. Прибавляли в культуральный флакон 1 мл 0,25% раствора трипсина. Помещали культуральный флакон в СО2-инкубатор при температуре (37±1)°С на 2-3 мин. Контролировали отслоение клеток микроскопированием. Останавливали реакцию трипсинизации путем прибавления во флакон ростовой (культуральной) среды, в соотношении (1:4) - 4 мл. Перемешивали клеточную суспензию пипетированием 5 10 раз, не допуская вспенивания. Переносили суспензию в стерильную центрифужную пробирку. Пробирку центрифугировали в течение 5 минут при 200 g. Надосадочную жидкость отбирали стерильной пипеткой и отбрасывали. Прибавляли к клеточному осадку 3-5 мл ростовой (культуральной) среды. Ресуспендировали осадок клеток с помощью стерильной серологической пипетки до получения гомогенной суспензии, отбирали 50 мкл.
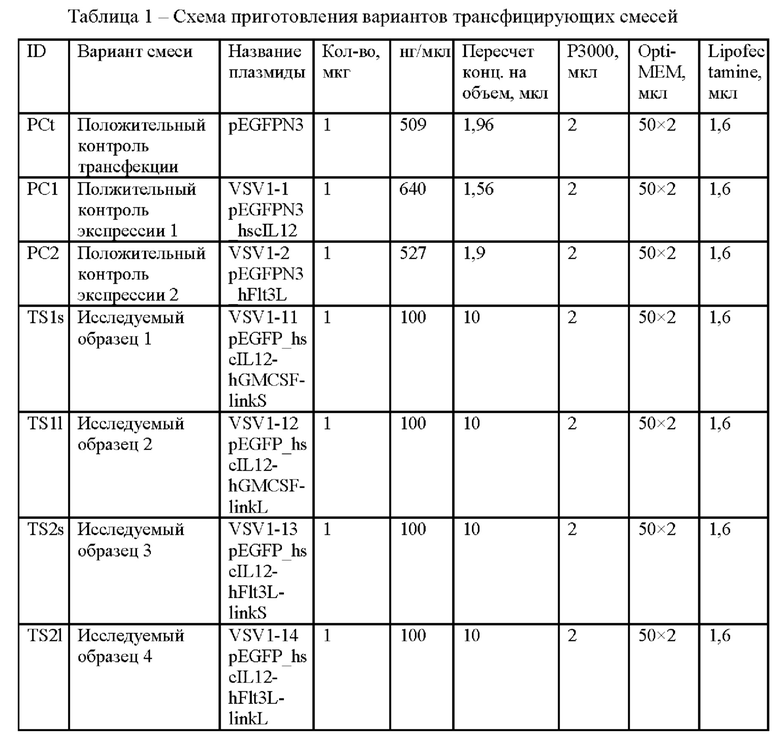
Далее проводили трансфекцию клеток по протоколу, рекомендованному производителем, следующими плазмидами:
Для оценки влияния длины линкера на экспрессию и функциональную активность проводили трансфекцию клеток плазмидами:
- pEGFP-hscIL12-hFLT3LlinkS (короткий линкер)
- pEGFP-hscIL12-hFLT3LlinkL (длинный линкер)
- pEGFP-hFLT3L в качестве положительного контроля экспрессии
Для оценки экспрессии и функциональной активности химеры hIL-12-hGM-CSF проводили трансфекцию клеток плазмидами:
- pEGFP-N3-hscIL12-hGM-CSFlinkS (короткий линкер)
- pEGFP-N3-hscIL12-hGM-CSFlinkL (длинный линкер)
- pEGFP_hIL12 - в качестве положительного контроля экспрессии.
- В качестве отрицательного контроля использовали нетрансфицированные клетки.
- В качестве положительного контроля трансфекции проводили трансфекцию вектором pEGFPN3 (Фиг. 5)
К клеткам HEK293TN с 1 мл среды DMEM+2mM L-Gln+2%FBS добавляли трансфицирующие растворы либо негативный контроль трансфекции по каплям. Перемешивали планшет с помощью покачиваний в горизонтальном положении крест-накрест, переносили в инкубатор для культивирования при температуре (37±1)°С в увлажненном воздухе с содержанием углерода диоксида 5%.
Схема использованных вариантов трансфицирующих смесей представлена в Таблице 1.
Через 24 часа после проведения трансфекции для оценки эффективности трансфекции проводили микроскопирование лунок культурального планшета с помощью флуоресцентного микроскопа. Пример фотографии клеточного монослоя в лунках с положительным контролем эффективности трансфекции представлен на Фиг. 6.
При заданных условиях трансфекции через 24 часа подтверждали эффективность трансфекции тем, что наблюдали с помощью флуоресцентного микроскопа наличие флуоресценции белка GFP в лунке положительного контроля трансфекции и отсутствие схожего уровня флуоресценции в лунках со всеми исследуемыми образцами.
Кондиционированную культуральную жидкость трансфицированной клеточной культуры HEK293TN отбирали и, при необходимости, закладывали на хранение при температуре 80°С или сразу использовали для анализа методом ИФА и на репортерной клеточной линии.
Пример 3. Определение наличия IL-12 человека в кондиционированной культуральной жидкости клеточной линии HEK293TN.
Определение наличия hFLT3L и hIL-12 человека в кондиционированной культуральной жидкости клеточной линии HEK293TN через 24 часа после трансфекции химерными плазмидами pEGFP-hscIL12-hFLT3L и pEGFP-hscIL12-GMCSF с разными вариантами длины линкеров в сравнении с pEGFP-hFLT3L и pEGFP-hIL12 и дополнительными положительными контролями ИФА (стандарт hFLT3L и стандарт hIL12) проводили по методикам, рекомендованным производителями наборов RayBio Human Flt3L ELISA kit (RayBiotech, cat# ELH-FLT3L) и «Human IL-12 p70 Uncoated ELISA Kit» (Invivogen, cat# 88-7126-22) соответственно.
Целью экспериментов было
А) измерить оптическую плотность (OD, от англ. optical density) исследуемых образцов кондиционированной культуральной жидкости, отобранных через 24 часа после трансфекции клеточной культуры плазмидами с химерными белками и с разными вариантами длины линкеров pEGFP-hscIL12-hFLT3L, pEGFP-hscIL12-GMCSF;
Б) сравнить OD со средним значением OD негативного контроля трансфекции (NC), со средним значением OD положительного контроля ИФА (стандарт hFLT3L, стандарт hIL12,) и средним OD положительного контроля экспресии pEGFP-hFLT3L и pEGFP-hIL12.
Результаты дополнительных экспериментов определения hFLT3L:
- определено наличие hFLT3L человека в образцах кондиционированной культуральной жидкости положительного контроля экспрессии hFLT3L через 24 часа после трансфекции плазмидой pEGFP hFLT3L клеточной культуры HEK293TN, сопоставимое с положительным контролем hFLT3L (относительная разница составляет 9,81%);
- определено наличие hFLT3L человека в образцах кондиционированной культуральной жидкости через 24 часа после трансфекции химерными плазмидами pEGFP-hscIL12-hFLT3LlinkS и pEGFP-hscIL12-hFLT3LlinkL в количестве большем, чем в отрицательном контроле трансфекции: относительная разница составляет 33,46% и 34,92% соответственно, но меньшем, чем в положительном контроле hFLT3L: относительная разница составляет 50,27% и 51,36% соответственно;
- относительная разница по сигналу OD для исследуемых образцов химерных плазмид с разной длиной линкера (коротким и длинным) различается несущественно и составляет 2,23%.
Таким образом, в составе двух типов химерных белков ген hFLT3L экспрессируется, включение Flt3 в состав химеры с hIL12 приводит к более низким значениям уровня экспрессии (относительная разница с положительным контролем трансфекции ~50%), но при этом тип линкера не влияет на уровень экспрессии hFLT3L в составе химеры с hIL12.
С другой стороны, проверили уровень экспрессии hIL-12 в составе двух типов химер:
- определено наличие hIL12 в образцах кондиционированной культуральной жидкости положительного контроля экспрессии РС1 через 24 часа после трансфекции плазмидой pEGFP hscIL12 клеточной культуры HEK293TN;
- определено наличие hIL12 в образцах кондиционированной культуральной жидкости через 24 часа после трансфекции химерными плазмидами pEGFP-hscIL12-hFLT3LlinkS HpEGFP-hscIL12-hFLT3LlinkLB количестве большем, чем в отрицательном контроле трансфекции, но меньшем, чем в положительном контроле экспрессии - относительная разница по средним значениям OD составляет 62,18% и 68,84% от уровня положительного контроля соответственно;
- относительная разница по сигналу OD для исследуемых образцов химерных плазмид между собой pEGFP-hscIL12-hFLT3LlinkS и pEGFP-hscIL12-hFLT3LlinkL составляет 21,39%.
- также определено наличие hIL12 в образцах кондиционированной культуральной жидкости через 24 часа после трансфекции химерными плазмидами pEGFP-hscIL12-GMCSFlinkS и pEGFP-hscIL12-GMCSFlinkL в количестве сопоставимом с положительным контролем экспрессии РС1, относительная разница с которым составляет 0,71% и 2,52% соответственно. При этом относительная разница по сигналу OD для исследуемых образцов кондиционированной культуральной жидкости химерных плазмид pEGFP-hscIL12-GMCSFlinkS и pEGFP-hscIL12-GMCSFlinkL между собой составляет 1,76%, то есть различия незначительны.
Таким образом, в составе двух типов химерных белков ген hIL12 экспрессируется, однако уровень его экспрессии по ИФА зависит от второго гена в составе химеры: добавление GM-CSF не сказывается на уровне экспрессии hLL12, в то время как при включении Flt3L в состав химеры уровень экспрессии демонстрирует более низкие значения (относительная разница ~60-70%).
При этом тип линкера не влияет на уровень экспрессии IL12 в составе химеры с GM-CSF, в то время как в составе с Flt3L использование длинного линкера приводит к более низкому уровню экспрессии IL12 (относительная разница ~21%.).
Пример 4. Оценка активности IL-12 в кондиционированной культуральной жидкости после трансфекции клеточной линии HEK293TN
Активность hIL-12 в кондиционированной культуральной жидкости клеточной линии HEK293TN через 24 часа после трансфекции химерными плазмидами pEGFP-hscIL12-hFLT3L и pEGFP-hscIL12-GMCSF с разными вариантами длины линкеров в сравнении с pEGFP-hscIL12 и положительным контролем (стандарт hIL12) проверяли с использованием репортерной клеточной линии HEK-Blue™ IL-12 (InvivoGen).
Конечной целью эксперимента было измерение OD исследуемых образцов отобранных через 24 часа после трансфекции, сравнение со средним значением OD клеточного контроля, сравнение со средним значением OD положительного контроля (hIL12 1000, 100, 10 нг/мл) с помощью расчета относительной разницы по средним значениям OD Δ, %.
Ход эксперимента: использовали следующий состав среды роста клеток: DMEM с содержанием глюкозы 4,5 г/л, 2 мМ L-глутамина, 10% FBS, 100 мкг/мл Normocin.
Для разморозки и культивирования репортерной клеточной культуры HEK-Blue™ IL-12 криопробирку с клетками адгезионной клеточной культуры HEK-Blue™ IL-12, извлеченную из криохранилища с жидким азотом, помещали в водяную баню с температурой (37±1)°С на 1-2 минуты. В стерильную пластиковую коническую центрифужную пробирку вместимостью 15 мл вносили 9 мл предварительно нагретой до (37±1)°С среды роста, переносили в нее содержимое криопробирки с клетками. Центрифугировали пробирку при 200 g в течение 5 мин. Сливали надосадочную жидкость, прибавляли 5 мл предварительно нагретой до (37±1)°С среды роста, ресуспендировали осадок с помощью стерильной серологической пипетки. Переносили полученную клеточную суспензию в культуральный флакон площадью 25 см2, добавляли в него 5-7 мл среды роста. Культуральный флакон с клеточной культурой культивировали в течение 2-3 суток в СО2-инкубаторе при температуре (37±1)°С в увлажненном воздухе с содержанием диоксида углерода 5% до достижения конфлюэнтности монослоя 70-80%, смену среды осуществляли 2 раза в неделю.
Пересев клеток осуществляли при достижении конфлюэнтности не менее 70-80%, не допуская образование монослоя клеток со 100% конфлюэнтностью. Для пересева из культурального флакона отбирали с помощью серологической пипетки культуральную среду в стерильную центрифужную пробирку вместимостью 50 мл, в культуральный флакон вносили 3 мл раствора Версена или PBS, промывали монослой клеток аккуратно покачивая флакон, находящийся в горизонтальном положении, в течение 3-5 с. Помещали культуральный флакон в СО2-инкубатор при температуре (37±1)°С на 2 3 мин. Контролировали отслоение клеток микроскопированием. Перемешивали клеточную суспензию пипетированием 5-10 раз, не допуская вспенивания. Переносили суспензию в стерильную центрифужную пробирку. Пробирку центрифугировали в течение 5 минут при 200 g. Надосадочную жидкость отбирали стерильной пипеткой и отбрасывали. Прибавляли к клеточному осадку 3-5 мл среды роста. Ресуспендировали осадок клеток с помощью стерильной серологической пипетки до получения гомогенной суспензии, прибавляли к клеточной суспензии необходимый объем среды роста, предварительно нагретой до (37±1) С, для получения посевной дозы 2,4-105 клеток/см2 или 1,2-106 клеток/мл. Вносили 5 мл клеточной суспензии в культуральный флакон площадью 25 см2. Флакон с клеточной культурой инкубировали в СО2-инкубаторе при температуре (37±1)°С в увлажненном воздухе, содержащем 5% диоксида углерода. Пересев клеточной культуры производили через каждые 2 3 дня при достижении уровня конфлюэнтности монослоя 70-80%.
Первые два пересева проводили без использования селективных антибиотиков. Затем прибавляли антибиотик и добавки к клеточной культуре до конечной концентрации 100 мкг/мл Normocin, 1×HEK-Blue™ Selection.
Для приготовления Test Medium использовали среду для роста клеток DMEM с содержанием 4,5 г/л глюкозы, 10%, эмбриональной бычьей сыворотки и 2 мМ L-глутамина.
При подготовке суспензии клеток HEK-Blue™ I клетки, пересеянные за 2 дня до постановки анализа во флаконы 25 см2 с начальной концентрацией 2,4-105 клеток/см2 или 1,2⋅106 клеток/мл и инкубированные при температуре (37±1)°С в увлажненном воздухе с содержанием СО2 5%, оценивали визуально под микроскопом. Отбирали с помощью серологической пипетки среду в стерильную центрифужную пробирку вместимостью 50 мл, в культуральный флакон вносили 3 мл раствора PBS, промывали монослой клеток аккуратно покачивая флакон. Помещали культуральный флакон в СО2-инкубатор при температуре (37±1)°С на 2-3 мин. Контролировали отслоение клеток микроскопированием. Перемешивали клеточную суспензию пипетированием 5-10 раз, не допуская вспенивания. Переносили суспензию в стерильную центрифужную пробирку. Пробирку центрифугировали в течение 5 минут при 200 g. Надосадочную жидкость отбирали стерильной пипеткой и отбрасывали. Прибавляли к клеточному осадку 3-5 мл Test Medium. Ресуспендировали осадок клеток с помощью стерильной серологической пипетки до получения гомогенной суспензии, отбирали 50 мкл суспензии, смешивали с эквивалентным объемом трипанового синего 0,4%, определяли концентрацию и жизнеспособность клеток в камере Горяева в процентах. Для анализа использовали суспензию с жизнеспособностью клеток не менее 90%. Готовили суспензию клеток с концентрацией (2,8±0,1)⋅105 клеток/мл в Test Medium из расчета 11 мл суспензии для заполнения одного культурального планшета.
При проведении анализа содержания IL-12 в кондиционированной культуральной жидкости для приготовления положительного контроля IL-12k 10 мкг человеческого IL-12 во флакон прибавляли 50 мкл раствора 0,1% стерильного BSA. Конечная концентрация стокового раствора IL-12 N1 - 0,2 мг/мл. Положительный контроль IL-12 использовали в рабочей концентрации 10 нг/мл, 100 нг/мл, 1 мкг/мл.
Для приготовления негативных контролей активации клеточной линии 1, 2 - в стерильной микропробирке разводили TNFa в Test Medium до концентрации 100 и 10 нг/мл.
В качестве неспецифичных контролей 1 и 2 использовали образцы кондиционированной культуральной жидкости, не содержащие IL12.
Для приготовления исследуемых образцов испытуемые образцы кондиционированной культуральной жидкости после трансфекции размораживали и разводили в Test Medium в 10 раз.
В 96-луночный плоскодонный культуральный планшет (планшет А) вносили исследуемые и контрольные образцы в тройной повторности по 20 мкл на лунку. Готовую клеточную суспензию HEK-Blue™ IL-12 с концентрацией (2,8±0,1)-105 клеток/мл с жизнеспособностью не менее 90% вносили по 180 мкл в лунки B2-G10 культурального планшета. Культуральный планшет переносили на шейкер для планшетов и перемешивали со скоростью 600 об/мин при комнатной температуре в течение 1-2 мин. После этого культуральный планшет помещали в СО2-инкубатор, и инкубировали при температуре (37±1)°С в увлажненном воздухе с 5% СО2 в течение 16-20 ч.
В новый 96-луночный плоскодонный культуральный планшет В вносили по 180 мкл реактива QUANTI-Blue (Invitrogen, США). Из инкубированного в течение 16-20 ч культурального планшета с клетками отбирали по 20 мкл кондиционированной культуральной жидкости и вносили их в планшет В. Культуральный планшет В помещали в СО2-инкубатор и инкубировали при температуре (37±1)°С в увлажненном воздухе с 5% СО2 в течение 15 мин - 6 ч.
Оптическую плотность в лунках культурального планшета В измеряли при помощи прибора для считывания оптической плотности в OD при длине волны 620-655 нм.
С помощью программного обеспечения Excel вычисляли среднее значение по трем повторам OD для контрольных и исследуемых образцов, RSD OD %, вычисляли отношение оптической плотности исследуемых и контрольных образцов.
Полученные результаты:
А) Содержание функционального hIL12 в образце кондиционированной культуральной жидкости после трансфекции химерной плазмидой pEGFP-hscIL12-hFLT3LlinkS было сопоставимо с положительным контролем hIL12 (в концентрации 10 нг/мл) и было больше, чем в кондиционированной культуральной жидкости после трансфекции химерной плазмидой pEGFP-hscIL12-hFLT3LlinkL. Оказалось, что замена длинного линкера на более короткий в случае химеры hIL12-hFLT3L позволяет повысить количество активного IL12 в более чем 4 раза и приблизить его уровень к уровню положительного контроля, относительная разница конструкций с коротким и длинным линкером при сравнении с положительным контролем составляет - 7,95 и 37,5% соответственно). Эти данные хорошо согласуются с данными ИФА (см. Пример 3), по которым уровень экспрессии ML12 в составе конструкции hIL12-hFLT3 в случае с длинным линкером был на 20% ниже, чем с коротким линкером.
Б) Функциональный hIL12 содержится в образце кондиционированной культуральной жидкости клеток линии HEK293TN через 24 часа после трансфекции химерными плазмидами с разными вариантами длины линкеров pEGFP-hscIL12-GMCSFlinkS и pEGFP-hscIL12-GMCSFlinkL в количестве сопоставимом с положительным контролем hIL12 в концентрации 100 нг/мл, относительная разница незначительна и составляет 4,91% и 2,58%. Функциональный hIL12 содержится в образцах кондиционированной культуральной жидкости клеток линии HEK293TN через 24 часа после трансфекции химерными плазмидами pEGFP-hscIL12-GMCSFlinkS и pEGFP-hscIL12-GMCSFlinkL, что сопоставимо с образцом препарата монобелка IL12 (стандарт IL12), относительная разница составляет 0,81% и 1,62% соответственно. Результаты экспериментов полностью согласуются с данными ИФА: тип линкера не влияет на функциональную активность hIL12 в составе химерного белка с GM-CSF, при этом и уровень экспрессии, и активность hIL12 в клеточном тесте в составе химеры с GM-CSF сопоставимы с уровнем положительных контролей hIL12.
При этом конструкции hIL12-GM-CSF не только обеспечивают сохранение функциональной активности hIL12, но и демонстрируют уровень экспрессии hIL12 на уровне положительного контроля ЫЫ2 в концентрации 100 нг/мл, что в 10 раз выше, чем в случае использование химеры hIL12-hFlt3L.
Пример 5. Создание конструкции mI112-mGM-CSF для наработки и проверки VSV.
Для проверки активности химеры IL12-GM-CSF в составе рекомбинантного VSV и последующих экспериментов создали генетическую конструкцию (коровый вектор), содержащий последовательности генов mIL12 и mGM-CSF мыши, для возможности в дальнейшем проводить проверку эффективности и безопасности вектора на сингенных мышиных моделях, например, мышах Balb/c или C57BL/6 с опухолевым трансплантатом мышиных опухолевых клеток, например B16F10 - культура меланомы или других.
Для сборки химерного гена mIL12-mGM-CSF использовали последовательности синтезированных генов mIL12 (SEQ Id No: 12), mGM-CSF (SEQ Id No: 13).
Целевая аминокислотная последовательность mIL12-mGM-CSF представлена (SEQ Id No: 14).
Сборку и клонирование последовательности, кодирующей mIL12-mGM-CSF проводили по следующей схеме.
1) Клонировали последовательности генов mIL12 и mGM-CSF в вектор pEGFP по сайтам NheFXhoI аналогично схеме из примера 1 с использованием специфичных к мышиным последовательностям праймеров.
Получили промежуточный вектор pEGFP-N3-mscIL12-mGM-CSF (Фиг. 7).
2) Проводили ПЦР амплификацию фрагмента гена mIL12-mGM-CSF с матрицы pEGFP-N3-mscIL12-mGM-CSF помощью специфических праймеров
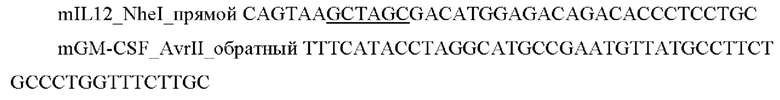
3) Проводили клонирование ПЦР продукта (~2050 п.о.) в вектор pVSV-G-GFP-dM51 (Фиг. 8) по сайтам Nhel/Avrll по стандартным методикам рестриктазно-лигазного клонирования.
4) Проводили анализ полученной конструкции с помощью электрофореза в агарозном геле и секвенированием. Карта полученного вектора представлена на Фиг. 9. Результаты секвенирования подтвердили наличие в векторе целевой последовательности, кодирующей mIL12-mGM-CSF (SEQ Id No: 15).
Пример 6. Наработка образца VSV-mIL12-mGMCSF.
Первый этап сборки VSV-mIL12-mGMCSF включал трансфекцию клеток полученной ранее линии HEK293-Т7, экспрессирующей Т7 полимеразу, 4 мя вспомогательными плазмидами, которые экспрессируют кДНК VSV N, VSV Р, VSV G, VSV L белков и коровой плазмидой (pVSV-G-dM51-mIL12-mGMCSF) в соотношении, представленном ниже:

За день до трансфекции во второй половине дня вносили по 0,9 мл суспензии HEK293-Т7 клеток в лунки 12-луночного планшета из расчета 350000 клеток на лунку. На следующий день при достижении конфлюэнтности 75-90% за два часа до трансфекции меняли питательную среду на среду с более низким содержанием фетальной бычьей сыворотки (FBS, 2%). Приготовление трансфекционных смесей производили последовательным добавлением по отдельности в 2 центрифужные пробирки объемом 1,5 мл плазмид pCAG-VSVN, pCAG-VSVP, pCAG-VSVL, pCAGGS-G, pVSV-G-dM51-mIL12-mGMCSF (в количестве, указанном в таблице с суммарной массой 10,5 мкг на лунку) и PEI (65 мкг для соотношения 5:1). Далее в каждую пробирку вносили бессывороточную среду Opti-Mem из расчета 100 мкл на каждую лунку. После объединения смесей плазмид и PEI перемешивали смесь на вортексе и инкубировали при комнатной температуре в течение 13 минут. Готовые трансфицирующие смеси наносили по каплям на клетки в лунках с 0,9 мл питательной средой с низким содержанием FBS (2%). После этого планшеты переносили в инкубатор для культивирования. Режим культивирования: температура 37,0°С, влажность 90%, содержание СО2 5%. Утром следующего дня после трансфекции меняли среду на среду DMEM с высоким содержанием глюкозы (4,5 г/л) и низким содержанием FBS (2%). Через 72 часа после трансфекции переносили кондиционированную культуральную жидкость с клетками в стерильные центрифужные пробирки и, при необходимости, хранили при 80°С или использовали сразу.
Перед вторым этапом наработки вирусных частиц (амплификация вирусных частиц, полученных на первом этапе), при необходимости, проводили разморозку кондиционированной культуральной жидкости с клетками и центрифугировали при максимальных оборотах в течение 10 минут для избавления от дебриса клеток. После центрифугирования отбирали супернатант для последующего инфицирования. Перед этим супернатант разбавляли два раза средой DMEM без сыворотки. Для второго и последующих этапов наработки использовали клетки линии BHK21. За день до трансдукции (инфицирования) вносили по 0,9 мл суспензии BHK21 в лунки 12-луночного планшета из расчета 250000 клеток на лунку. На следующий день при достижении конфлюэнтности 55-75% во всех лунках снимали питательную среду, вносили по 500 мкл разбавленного 2 раза супернатанта после 1 этапа сборки вирусных частиц и инкубировали 2 часа, добавляли по 500 мкл DMEM с 4% FBS. Через 72 ч культивирования снимали клетки с кондиционированной культуральной жидкостью и, при необходимости, хранили при 80°С или использовали сразу.
Перед третьим этапом наработки вирусных частиц (амплификация вирусных частиц, полученных на втором этапе наработки) также, при необходимости, проводили разморозку кондиционированной культуральной жидкости с клетками и центрифугирование при максимальных оборотах в течение 10 минут для избавления от дебриса клеток. Далее также как перед вторым этапом наработки супернатант разбавляли два раза DMEM без сыворотки. Для этого этапа за день до трансдукции (инфицирования) вносили по 5,0 мл суспензии BHK21 клеток в культуральные флаконы Т25 с площадью поверхности 25 см2 из расчета 1,5 млн клеток на один флакон. На следующий день при достижении конфлюэнтности 55-75% удаляли питательную среду, вносили по 2,5 мл разбавленного 2 раза супернатанта после 2 этапа наработки вирусных частиц и инкубировали 2 часа, добавляли по 2,5 мл DMEM с 4% FBS. Через 72 ч культивирования снимали клетки с кондиционированной культуральной жидкостью и, при необходимости, хранили при -80°С или использовали сразу. На этом этапе отбирали образцы для подтверждения наличия вирусных частиц посредством методов электрофореза в полиакриламидном геле и ПЦР.
После подтверждения наличия вирусных частиц проводили 4 этап наработки в культуральных флаконах Т150 с площадью поверхности 150 см2. Для этого за день до трансдукции (инфицирования) вносили по 30,0 мл суспензии BHK21 в культуральные флаконы с площадью поверхности 150 см2 из расчета 9,0 млн клеток на один флакон. На следующий день при достижении конфлюэнтности 55 75% снимали питательную среду, вносили по 15 мл разбавленного 2 раза супернатанта после 3 этапа наработки вирусных частиц и инкубировали 2 часа, добавляли по 15 мл DMEM с 4% FBS. Через 72 ч культивирования снимали клетки с кондиционированной культуральной жидкостью, отбирали образцы для подтверждения наличия вирусных частиц посредством методов электрофореза в полиакрил амид ном геле и ПЦР и, при необходимости, хранили при -80°С.
Пример 7. Определение титра и анализ VSV-mIL12-mGMCSF
Оценку титра вируса проводили по цитопатическому эффекту на клетки BHK21. Расчет титра (TCID50/мл) осуществляли по методу Рида-Менча [Lei С, Yang J, Hu J, Sun X. On the Calculation of TCID50 for Quantitation of Virus mfectivity. Virol Sin. 2021 Feb; 36(l): 141-144. doi: 10.1007/sl2250-020-00230-5; Reed LJ, Muench H. A simple method of estimating fifty per cent endpoints. Am J Hygiene. 1938; 27(3):493-497. doi: 10.1093/oxfordj ournals. aj e. a118408].
Клетки BHK21 для анализа засевали на 96-луночный планшет: 10 рядов по 6 лунок в каждом (всего 60 лунок). В каждую из лунок по 25'000 клеток в 100 мкл среды DMEM+5% FBS с расчетом через 24 часа перед проведением эксперимента получить конфлюэнтность около 60-70%. 10 рядов соответствуют 9 рядам разведения вируса 10-1-10-9 и 1 контрольному ряду. В каждом ряду по 6 повторностей. Далее готовили серию разведений VSV-mIL12-mGMCSF в среде DMEM без FBS 10-1-10-9: каждый образец получали из предыдущего разведением в 10 раз. Заражение проводили, используя 50 мкл образца соответствующего разведения на лунку. Оставляли планшет инкубироваться при 37°С в течение 2 часов. Через 2 часа в каждую лунку добавляли по 150 мкл заранее приготовленной среды DMEM+2,7% FBS (для достижения конечной концентрации сыворотки 2%) и оставляли планшет инкубироваться при 37°С в течение 72 часов. Через 72 часа под микроскопом фиксировали проявление цитопатического эффекта вируса в каждой лунке. Далее вычисляли TCIDso/ml, используя стандартный протокол. Картину цитопатического эффекта (ЦПЭ) оформляли в виде схемы (Фиг. 10):
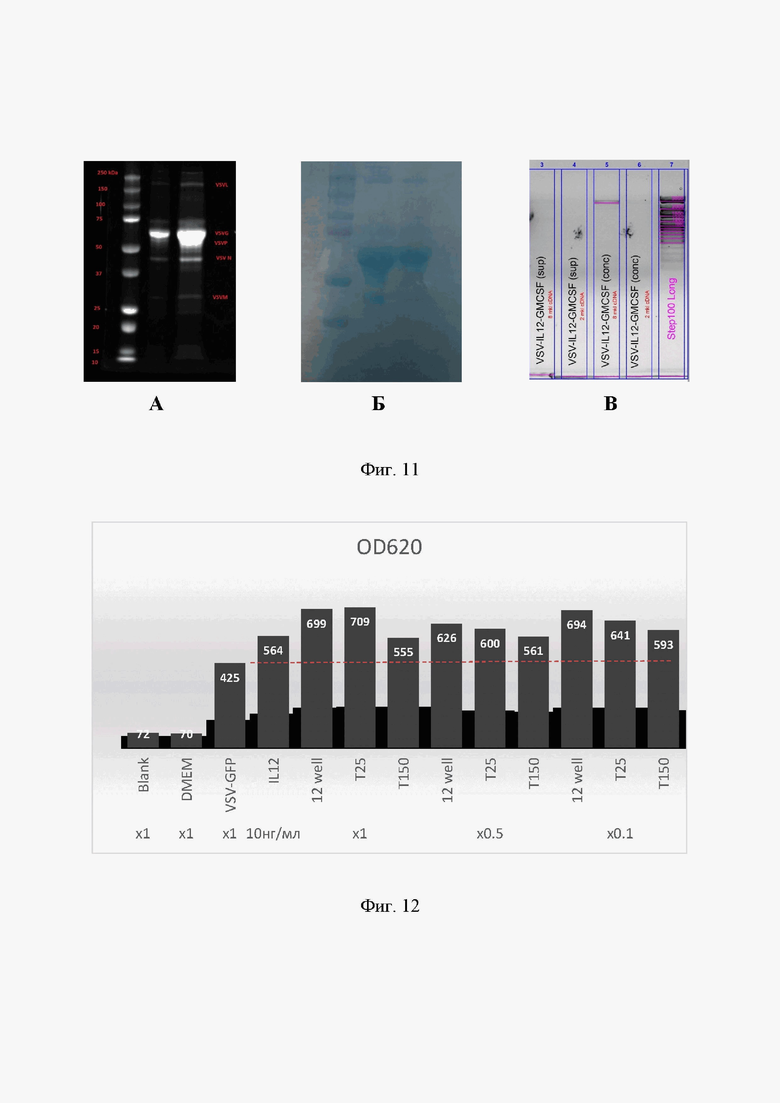
Кондиционированную культуральную жидкость клеток BHK21 после наработки VSV концентрировали с помощью концентраторов (300 кДа, Vivaspin 2) по протоколу производителя и анализировали с помощью электрофореза в полиакрил амид ном геле по стандартному протоколу [Laemmli UK. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature, 1970; V. 227, P. 680-685]. После окрашивания наблюдали полосы, молекулярная масса которых соответствуют пяти белкам VSV (Фиг. 11А, Б).
Дополнительно проверяли наличие в геноме VSV гена mIL12-mGMCSF, используя для этого набор универсальных праймеров
3'-конец гена G «vsvG end f»: CGGTCTCAAAATCGTGGACTTCC;
5'-конец гена£ «vsvL_start_r»: CATTCAAGACGCTGCTTCGCAAC
Для этого 500 мкл кондиционированной культуральной жидкости с клеток-продуцентов (BHK21) обрабатывали протеинкиназой-К для высвобождения вирусной РНК. Далее выделяли РНК коммерческим набором «Лира+» (Биолабмикс, РФ). После выделения обрабатывали ДНКазой для ликвидирования возможных остатков сторонней клеточной ДНК. Далее проводили обратную транскрипцию, используя специфичные праймеры, фланкирующие трансген mIL12-mGMCSF.
После получения кДНК проводили ПЦР с использованием этих же праймеров при следующих условиях: Taq-полимераза, денатурация 95°С 30 сек, отжиг 54°С 20 сек, элонгация 72°С 90 сек. Ампликоны анализировали на электрофорезе в агарозном геле. Наблюдали наличие специфичного ПЦР-продукта, соответствующего размеру ампликонат1Ы2-тОМС8Р (Фиг. 11В)
Пример 8. Оценка активности IL12 в кондиционированной культуральной жидкости после амплификации VSV-mIL12-mGMCSF в культуре BHK21.
Активность hIL12 в кондиционированной культуральной жидкости BHK21 на каждом этапе наработки VSV-mIL12-mGMCSF: из 12-луночных планшетов, культуральных флаконов Т25 и Т150, - проверяли с использованием репортерной клеточной линии HEK-Blue™ IL-12 (InvivoGen) по протоколу, описанному ранее в Примере 4. В результате эксперимента выяснили, что в кондиционированной культуральной жидкости содержится функциональный IL12, который вызывает активацию специфичного клеточного репортера на уровне положительного контроля (Фиг. 12).
Пример 9. Оценка эффективности VSV-mIL12-mGMCSF in vivo.
В эксперимент брали мышей линии Balb/c возрастом 6 недель. Подкожно в левый бок вводили 100 мкл суспензии, содержащей 10 млн/мл клеток B16F10. В течение недели наблюдали образование опухоли темного цвета, ясно различимой на белом фоне. Когда опухоль достигла линейных размеров 0,5 см, интратуморально проводили инъекцию 50 мкл концентратов VSV-G-GFP (1,0⋅108 TCID50) и VSV-mIL12-mGM-CSF (7⋅104 TCID50). Через 7 дней наблюдали полное исчезновение опухоли у мышей, которые получили инъекцию VSV-mIL12-mGM-CSF, в то время как лизис опухоли препаратом VSV-G-GFP проходил медленнее (разница в 1 неделю) (Фиг. 13). При этом следует отметить, что препарат VSV-mIL12-mGM-CSF обладал терапевтическим эффектом в 1000-кратно меньшей дозе из расчета титров по TCID50.
Все публикации, патенты и заявки на патенты включены в настоящий документ посредством ссылки. Хотя в вышеприведенном описании это изобретение было описано в отношении некоторых предпочтительных вариантов его осуществления, и многие детали были изложены в целях иллюстрации, для специалистов в данной области техники будет очевидно, что изобретение допускает дополнительные варианты осуществления и что некоторые детали, описанные в данном документе, могут значительно изменяться без отклонения от сущности изобретения.
Использование терминов в единственном числе в контексте описания изобретения должно толковаться как охватывающее как единственное, так и множественное число, если иное не указано в данном документе или явно не противоречит контексту. Термины «состоящий из», «имеющий», «включающий» и «содержащий» следует толковать как неограничивающие термины, т.е. означающие «включая, но не ограничиваясь», если не указано иное. Перечисление диапазонов значений в данном документе просто предназначено для использования в качестве сокращенного способа индивидуальной ссылки на каждое отдельное значение, попадающее в этот диапазон, если здесь не указано иное, и каждое отдельное значение включено в спецификацию, как если бы оно было отдельно изложено в данном документе. Все способы, описанные в данном документе, могут выполняться в любом подходящем порядке, если иное не указано в данном документе или иным образом явно не противоречит контексту. Использование любых и всех примеров или иллюстративного языка (например, «такой как»), представленных в данном документе, предназначено просто для лучшего описания изобретения и не налагает ограничения на объем изобретения, если иное не заявлено. Никакие формулировки в описании не следует истолковывать как указывающие на какой-либо не заявленный элемент как существенный для практического применения изобретения.
Здесь описаны варианты осуществления этого изобретения, включая лучший из известных изобретателям способа осуществления изобретения. Разновидности этих вариантов осуществления могут стать очевидными для специалистов в данной области техники после прочтения предшествующего описания. Авторы ожидают, что квалифицированные специалисты будут использовать такие варианты в зависимости от обстоятельств, и авторы предполагают, что изобретение будет реализовано на практике иначе, чем конкретно описано в данном документе. Соответственно, это изобретение включает в себя все модификации и эквиваленты признаков, изложенных в прилагаемой формуле изобретения, как это разрешено действующим законодательством. Более того, любая комбинация вышеописанных признаков во всех их возможных вариациях охватывается изобретением, если иное не указано в данном документе или иным образом явно не противоречит контексту.
Заявитель просит рассмотреть представленные материалы заявки «Рекомбинантный вирус везикулярного стоматита, экспрессирующий химерный белок IL12-GMCSF» на предмет выдачи патента на изобретение.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения рекомбинантного вируса везикулярного стоматита | 2022 |
|
RU2807751C1 |
| УНИВЕРСАЛЬНЫЕ РАКОВОСПЕЦИФИЧНЫЕ ПРОМОТОРЫ И ИХ ИСПОЛЬЗОВАНИЕ В ПРОТИВООПУХОЛЕВОЙ ТЕРАПИИ | 2013 |
|
RU2539764C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ ХИМЕРНОЕ АНТИТЕЛО ПРОТИВ ФАКТОРА НЕКРОЗА ОПУХОЛИ-АЛЬФА ЧЕЛОВЕКА, ЛИНИЯ ЭУКАРИОТИЧЕСКИХ КЛЕТОК-ПРОДУЦЕНТ ХИМЕРНОГО АНТИТЕЛА И СПОСОБ ПОЛУЧЕНИЯ ХИМЕРНОГО АНТИТЕЛА | 2013 |
|
RU2555533C9 |
| ПЕПТИД MPI БЕЗ СИГНАЛЬНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ, ДНК, ЕГО КОДИРУЮЩАЯ, ВЕКТОР, СПОСОБ ОПРЕДЕЛЕНИЯ СПОСОБНОСТИ К СЕКРЕТИРОВАНИЮ ТЕСТИРУЕМОГО ПЕПТИДА, СПОСОБ ВЫДЕЛЕНИЯ КДНК | 1998 |
|
RU2230751C2 |
| СПОСОБ ПОЛУЧЕНИЯ ОТЛИЧНОГО ОТ АДЕНОВИРУСА ВИРУСА-МИШЕНИ ИЛИ БЕЛКОВ-МИШЕНЕЙ, ЭКСПРЕССИРУЮЩАЯ КЛЕТКА И КЛЕТКА-ХОЗЯИН И СПОСОБЫ ИХ ПОЛУЧЕНИЯ, ПРИМЕНЕНИЕ ЭКСПРЕССИРУЮЩЕЙ КЛЕТКИ | 2006 |
|
RU2449015C2 |
| Гистоны и биодеградируемые липиды как средство для доставки нуклеиновых кислот в клетки эукариот | 2015 |
|
RU2637371C2 |
| МОНОМОЛЕКУЛЯРНЫЙ ХИМЕРНЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР К РАКОВОМУ АНТИГЕНУ СА125 | 2018 |
|
RU2747095C2 |
| МОДИФИЦИРОВАННЫЙ БЕЛОК ИНТЕРЛЕЙКИНА-7 И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2016 |
|
RU2708160C2 |
| АДЕНОВИРУСНЫЕ ВЕКТОРЫ И СПОСОБЫ И ПРИМЕНЕНИЯ, СВЯЗАННЫЕ С НИМИ | 2009 |
|
RU2520823C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA1FI6-ht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IGA1 | 2016 |
|
RU2656142C1 |

Группа изобретений относится к биотехнологии. Представлены: рекомбинантный вирус везикулярного стоматита, экспрессирующий химерный белок IL12-GMCSF и его применение в качестве препарата с противоопухолевой активностью. Заявленный вирус обладает выраженной онколитической способностью и может быть использован для лечения онкологических заболеваний. 2 н. и 4 з.п. ф-лы, 13 ил., 2 табл., 9 пр.
1. Рекомбинантный вирус везикулярного стоматита для экспрессии химерного белка IL12-GMCSF, содержащий нуклеиновую кислоту, кодирующую химерный белок IL12-GMCSF, в котором IL12 и GMCSF соединены посредством аминокислотного линкера.
2. Вирус по п. 1, характеризующийся тем, что экспрессирует белок с последовательностью SEQ Id No.: 6.
3. Вирус по п. 1, характеризующийся тем, что экспрессирует белок с последовательностью SEQ Id No.: 7.
4. Вирус по п. 2, характеризующийся тем, что содержит в геноме последовательность SEQ Id No.: 10.
5. Вирус по п. 2, характеризующийся тем, что содержит в геноме последовательность SEQ Id No.: 11.
6. Применение вируса по любому из пп. 1–5 в качестве препарата с противоопухолевой активностью.
| MUNIS AM, et.al | |||
| A tool with many applications: vesicular stomatitis virus in research and medicine, Expert Opinion on Biological Therapy, 2020 с.1187-1200 | |||
| SHIN EJ, WANNA GB, CHOI B, et al | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Laryngoscope | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |

