[0001] Настоящее изобретение относится к новым селективным в отношении вида эндолизинам фагов и их применению для лечения бактериального вагиноза (BV). В настоящем изобретении предложены рекомбинантные эндолизины, то есть эндолизины с заменой домена. Изобретение также относится к указанным эндолизинам для применения в лечении заболеваний или расстройств, таких как бактериальные инфекции, в частности BV. Изобретение также относится к полинуклеотидам. кодирующим указанные эндолизины. Указанные полинуклеотиды также можно применять для лечения таких заболеваний или расстройств. В настоящем изобретении также предложена фармацевтическая композиция, содержащая эндолизин по настоящему изобретению для применения в лечении таких заболеваний или расстройств. Указанные эндолизины, полинуклеотиды и фармацевтическую композицию можно вводить местно, в частности местно во влагалище.
[0002] Бактериальный вагиноз (BV), который также упоминается в литературе как бактериальный вагинит, неспецифический вагиноз и неспецифический вагинит, является наиболее распространенной вагинальной инфекцией во всем мире и связан с существенными неблагоприятными последствиями, включая преждевременные схватки и роды, послеродовой эндометрит и повышенный риск заражения HIV (вирус иммунодефицита человека). Это дисбиоз влагалища, при котором комменсальные Lactobacilli вытесняются полимикробной биопленкой, рН увеличивается от естественных 3,5-4,5 до 5,5 и образуется жидкость с неприятным запахом. Как сообщается, уровень распространенности варьирует от 10 до 40% в зависимости от исследуемой популяции. Однако неоптимальные способы диагностирования и высокий процент бессимптомных пациентов затрудняют установление истинной распространенности BV. Gardnerella vaginalis (G. vaginalis) представляет собой вид бактерий, ассоциированный с BV.
[0003] Этиопатогенез BV остается плохо изученным. Чаще всего его определяют как патологическое состояние, характеризующееся потерей нормальной микрофлоры влагалища, в частности видов Lactobacillus, продуцирующих Н2О2, и одновременным чрезмерным ростом анаэробных бактерий, включая G. vaginalis, виды Mobiluncus и Mycoplasma hominis. Однако последние данные предполагают основную роль G. vaginalis как специфического и передающегося половым путем этиологического агента при BV (Muzny et al., 2016, J. of Infect. Dis. 214 Suppl. 1., S1).
[0004] В 1950-х годах в половых путях у женщин с BV наблюдались многочисленные мелкие плеоморфные грамвариабельные палочки. Этот организм, который сначала назвали Haemophilus vaginalis и неоднократно переименовывали по мере того, как появлялось все больше информации о его характеристиках, теперь классифицируется как G. vaginalis, и до 2018 года его считали единственным представителем рода Gardnerella. Однако в начале 2019 года показали, что род Gardnerella на самом деле включает не менее 13 видов, и наиболее часто встречающиеся из них переименовали в G. vaginalis sensu stricto, G. leopoldii, G. piotii и G. swidsinskii (Vaneechoutte et al., 2019 Int. J. Syst. Evol. Biol. 898661).
[0005] Особенностью бактерий рода Gardnerella является то, что они являются грамвариабельными, то есть они не образуют внешнюю мембрану, которая определяет грамотрицательные виды. Клеточная стенка обычно очень тонкая, а содержание в ней пептид о гликанов составляет лишь 10% или менее, поэтому кристаллический фиолетовый краситель, используемый для окраски по Граму, не всегда дает темно-фиолетовый цвет, типичный для грамположительных видов. Вместо этого клетки Gardnerella при окрашивании по Граму могут выглядеть как грамположительными, так и грамотрицательными. Филогенетический анализ на основе 16S рРНК (рибосомальная РНК) помещает Gardnerella в группу грамположительных Bifidobacteriales.
[0006] При BV поверхность эпителия покрыта плотным скоплением G. vaginalis в биопленке, которая часто не поддается лечению. Биопленки представляют собой адгезирующие сообщества микроорганизмов, удерживаемые вместе полимерным матриксом, состоящим из полисахаридов, белков и/или нуклеиновых кислот. Четкий паттерн экспрессии генов, а также физическая структура биопленок повышают устойчивость бактерий ко многим отрицательным стимулам, включая химические дезинфицирующие средства, экстремальные значения рН, иммунную защиту хозяина и антибиотики. Стандартом лечения BV являются антибиотики метронидазол и клиндамицин, которые, однако, часто не могут уничтожить биопленку, так что частота рецидивов в течение 6 месяцев достигает 60%. Кроме того, лечение антибиотиками наносит удар по микробному влагалища, несмотря на то, что оно оставляет некоторые остатки жизнеспособной биопленки, что открывает эту экологическую нишу для других патогенов, например грибов. Таким образом, частым эффектом лечения BV является кандидоз. Также предпринимались попытки лечить BV пробиотиками, в частности полезными Lactobacilli, которые, как предполагается, заново заселяют влагалище. Однако, несколько клинических испытаний не показали положительных результатов.
[0007] Таким образом, существует большая потребность в новых способах и композициях для лечения инфекций, вызванных G. vaginalis, и в частности, BV. например, путем избирательного уничтожения бактериальных клеток рода Gardnerella, предпочтительно без вреда для полезных Lactobacilli, пока они заново заселяют влагалище. Таким образом, техническая задача, лежащая в основе настоящего изобретения, заключается в предоставлении новых средств и способов лечения BV. Предложенные воплощения, охарактеризованные в формуле изобретения, решают техническую задачу.
[0008] Настоящее изобретение основано на получении новых рекомбинантных эндолизинов профагов Gardnerella с неожиданными свойствами и структурой, делающих их особенно подходящими для различных применений и способов, в частности для лечения, деконтаминации или обнаружения бактериальных инфекций и расстройств, в частности связанных с Gardnerella.
[0009] В первом аспекте изобретения предложен эндолизин, содержащий или состоящий из
(1) N-концевого каталитического домена или его функционального варианта,
(2) С-концевой связывающейся с клеточной стенкой области или ее функционального варианта, где С-концевая связывающаяся с клеточной стенкой область содержит или состоит из по меньшей мере одного связывающегося с клеточной стенкой домена, и
(3) линкерной области между N-концевым каталитическим доменом и С-концевой связывающейся с клеточной стенкой областью,
где эндолизин обладает уничтожающей активностью в отношении Gardnerella.
[0010] В одном аспекте изобретения N-концевой каталитический домен имеет происхождение из первого естественного эндолизина, линкерная область и С-концевая связывающаяся с клеточной стенкой область имеют происхождение из второго естественного эндолизина, и где первый и второй естественные эндолизины закодированы разными геномами разных профагов. Таким образом, в изобретении предложен рекомбинантный эндолизин, содержащий или состоящий из
(1) N-концевого каталитического домена или его функционального варианта,
(2) С-концевой связывающейся с клеточной стенкой области или ее функционального варианта, где С-концевая связывающаяся с клеточной стенкой область содержит или состоит из по меньшей мере одного связывающегося с клеточной стенкой домена, и
(3) линкерной области между N-концевым каталитическим доменом и С-концевой связывающейся с клеточной стенкой областью,
где N-концевой каталитический домен имеет происхождение из первого естественного эндолизина, линкерная область и С-концевая связывающаяся с клеточной стенкой область имеют происхождение из второго естественного эндолизина, и первый и второй естественные эндолизины закодированы разными геномами разных профагов, и
где указанный рекомбинантный эндолизин обладает уничтожающей активностью в отношении Gardnerella.
[0011] Особенностью Gardnerella является то, что это грамвариабельный вид: он не образует внешнюю мембрану, которая определяет истинные грамотрицательные виды. Его клеточная стенка обычно очень тонкая, а содержание в ней пептидогликанов составляет лишь 10% или менее. Это указывает, что разрушающий пептидогликан фермент, такой как белки эндолизины, не может эффективно лизировать бактериальные клеточные стенки Gardnerella. Однако в контексте настоящего изобретения были обнаружены новые рекомбинантные эндолизины, обладающие предпочтительным свойством, заключающимся в том, что они эффективно уничтожают виды Gardnerella, и, следовательно, их можно применять в качестве новой терапии для лечения BV.
[0012] Здоровое влагалище заселяют 3 вида. Lactobacilli: L. crispatus, L. gasseri и L. jemenii. Они поддерживают кислые значения рН 3,5-4,5, производя молочную кислоту, и защитную окислительную среду, производя Н2О2. Выздоровление после BV связано с заселением влагалища заново указанными Lactobacilli. Однако недостатком антибиотиков (которые обычно применяют при лечении BV) является то, что они мешают процессу заселения влагалища заново Lactobacilli. Напротив, преимуществом новых рекомбинантных эндолизинов по изобретению является то, что они обладают избирательной уничтожающей активностью в отношении рода Gardnerella и не наносят вред Lactobacilli. Кроме того, в прилагающихся Примерах показано, что все протестированные штаммы Gardnerella обладают низкой чувствительностью к метронидазолу и клиндамицину, которые обычно применяют в лечении BV. Это может объяснить высокую частоту рецидивирования BV. Это также подтверждает, что эндолизины по изобретению превосходят антибиотики в лечении BV. Соответственно, лечение BV эндолизинами по настоящему изобретению имеет большие преимущества по сравнению с доступными в настоящее время способами лечения, такими как лечение антибиотиками метронидазолом и клиндамицином.
[0013] В данном документе термин «из первого естественного эндолизина» означает, что соответствующая часть (то есть N-концевой каталитический домен) идентична первому естественному эндолизину или является функциональным вариантом первого естественного эндолизина. В данном документе функциональный вариант представляет собой полипептид, который идентичен по меньшей мере на 80% (предпочтительно идентичен по меньшей мере на 85%, более предпочтительно идентичен по меньшей мере на 90%, еще более предпочтительно идентичен по меньшей мере на 95%, еще более предпочтительно идентичен по меньшей мере на 96%, еще более предпочтительно идентичен по меньшей мере на 97%, еще более предпочтительно идентичен по меньшей мере на 98%, еще более предпочтительно идентичен по меньшей мере на 99%, еще более предпочтительно идентичен по меньшей мере на 99,5% и наиболее предпочтительно идентичен по меньшей мере на 99,7%) аминокислотой последовательности соответствующей части (то есть N-концевому каталитическому домену) первого естественного эндолизина, результатом чего является функциональный эндолизин, где функция включает уничтожающую активность в отношении Gardnerella. Аминокислотные последовательности нескольких эндолизинов естественного происхождения приведены ниже и обобщены в Таблице 7.
[0014] Аналогично данному термину, «из второго естественного эндолизина» означает, что соответствующая часть (то есть линкерная область и С-концевая связывающаяся с клеточной стенкой область) идентична второму естественному эндолизину или является функциональным вариантом второго естественного эндолизина, то есть эндолизина, который отличается от первого естественного эндолизина. В данном документе функциональный вариант представляет собой полипептид, который идентичен по меньшей мере на 80% (предпочтительно идентичен по меньшей мере на 85%, более предпочтительно идентичен по меньшей мере на 90%, еще более предпочтительно идентичен по меньшей мере на 95%, еще более предпочтительно идентичен по меньшей мере на 96%, еще более предпочтительно идентичен по меньшей мере на 97%, еще более предпочтительно идентичен по меньшей мере на 98%, еще более предпочтительно идентичен по меньшей мере на 99%, еще более предпочтительно идентичен по меньшей мере на 99,5% и наиболее предпочтительно идентичен по меньшей мере на 99,7%) аминокислотой последовательности соответствующей части (то есть линкерной области и С-концевой связывающейся с клеточной стенкой области) второго естественного эндолизина, результатом чего является функциональный эндолизин, где функция включает уничтожающую активность в отношении Gardnerella.
[0015] В данном документе N-концевой каталитический домен также обозначают как «Н-домен». Например, термин «Н2» относится к Н-домену естественного эндолизина (EL) 2. «С-концевая связывающаяся с клеточной стенкой область» относится к одному или более чем одному связывающемуся с клеточной стенкой домену. Линкер и связывающиеся с клеточной стенкой домены вместе представляют так называемую «В-область». Например, В10 относится к В-области естественного EL10. Аналогично, B11_N относится к N-концевому связывающемуся с клеточной стенкой домену естественного EL11, В12_С относится к С-концевому связывающемуся с клеточной стенкой домену естественного EL12 и так далее.
[0016] В изобретении также предложен эндолизин, содержащий или состоящий из
(1) N-концевого каталитического домена, состоящего из полипептида, содержащего или состоящего из аминокислотной последовательности любой из SEQ ID NO: 1-5, 7 или 10-12 или любого ее функционального варианта, идентичного по меньшей мере на 80% аминокислотной последовательности любой из SEQ ID NO: 1-5, 7 или 10-12;
(2) С-концевой связывающейся с клеточной стенкой области, содержащей или состоящей из по меньшей мере одного связывающегося с клеточной стенкой домена, независимо выбранного из группы, состоящей из полипептидов, содержащих или состоящих из аминокислотной последовательности любой из SEQ ID NO: 15-24 и 26-33, соответственно, и любого их функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности любой из SEQ ID NO: 15-24 и 26-33, соответственно; и
(3) линкерной области между N-концевым каталитическим доменом и С-концевой связывающейся с клеточной стенкой областью,
где указанный эндолизин обладает уничтожающей активностью в отношении Gardnerella.
[0017] Как показано в прилагаемых Примерах, наиболее активным N-концевым каталитическим доменом (также обозначаемым как «Н-домен») является Н2 (SEQ ID NO: 2), за которым следуют Н7 (SEQ ID NO: 7), H10 (SEQ ID NO: 10) и H5 (SEQ ID NO: 5).
[0018] Таким образом, в предпочтительном аспекте настоящего изобретения N-концевой каталитический домен состоит из полипептида, содержащего или состоящего из аминокислотной последовательности любой из SEQ ID NO: 2, 7, 10 и 5 или любого их функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности любой из SEQ ID NO: 2, 7, 10 и 5;
где эндолизин является функциональным, где функция включает способность лизировать клеточную стенку Gardnerella.
[0019] Таким образом, в предпочтительном аспекте настоящего изобретения N-концевой каталитический домен состоит из полипептида, содержащего или состоящего из аминокислотной последовательности SEQ ID NO: 5 или любого его функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 5; или более предпочтительно содержащего или состоящего из аминокислотной последовательности SEQ ID NO: 10 или любого его функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 10; или еще более предпочтительно содержащего или состоящего из аминокислотной последовательности SEQ ID NO: 7 или любого его функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 7; или еще более предпочтительно содержащего или состоящего из аминокислотной последовательности SEQ ID NO: 2 или любого его функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 2;
где эндолизин является функциональным, где функция включает способность лизировать клеточную стенку Gardnerella.
[0020] В прилагаемых Примерах также показано, что из В-областей наиболее активной является В10 (содержащая связывающиеся с клеточной стенкой домены с SEQ ID NO: 28 и 29), за ней следует В11 (содержащая связывающиеся с клеточной стенкой домены с SEQ ID NO: 30 и 31), В12 (содержащая связывающиеся с клеточной стенкой домены с SEQ ID NO: 32 и 33) и В3 (содержащая связывающиеся с клеточной стенкой домены с SEQ ID NO: 19 и 20).
[0021] Таким образом, в предпочтительном аспекте настоящего изобретения связывающийся(еся) с клеточной стенкой домен(ы) выбран(ы) из группы, состоящей из полипептидов, содержащих или состоящих из аминокислотной последовательности любой из SEQ ID NO: 19, 20 и 28-33 и любого их функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности любой из SEQ ID NO: 19, 20 и 28-33;
где эндолизин является функциональным, где функция включает способность лизировать клеточную стенку Gardnerella.
[0022] Эндолизин по настоящему изобретению предпочтительно содержит два связывающихся с клеточной стенкой домена. В одном аспекте настоящего изобретения связывающиеся с клеточной стенкой домены (В-домены) эндолизинов по изобретению состоят из полипептида, содержащего или состоящего из аминокислотной последовательности любой из SEQ ID NO: 19, 20 и 28-33 и любого их функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности любой из SEQ ID NO: 19, 20 и 28-33;
где эндолизин является функциональным, где функция включает способность лизировать клеточную стенку Gardnerella.
В предпочтительном аспекте настоящего изобретения эндолизин содержит первый связывающийся с клеточной стенкой домен и второй связывающийся с клеточной стенкой домен, где указанный первый связывающийся с клеточной стенкой домен выбран из группы, состоящей из SEQ ID NO: 15, 17, 19, 21, 23, 26, 28, 30 и 32, а второй связывающийся с клеточной стенкой домен выбран из группы, состоящей из SEQ ID NO: 16, 18, 20, 22, 24, 27, 29, 31 и 33. Предпочтительно, указанный первый связывающийся с клеточной стенкой домен расположен в N-концевом направлении от указанного второго связывающегося с клеточной стенкой домена.
[0023] В более предпочтительном аспекте настоящего изобретения эндолизин содержит два связывающихся с клеточной стенкой домена (В-домена) естественного эндолизина EL10 (SEQ ID NO: 28 и 29), естественного эндолизина EL11 (SEQ ID NO: 30 и 31), естественного эндолизина EL12 (SEQ ID NO: 32 и 33) или естественного эндолизина EL3 (SEQ ID NO: 19 и 20), еще более предпочтительно естественного эндолизина EL10 (SEQ ID NO: 28 и 29); или их функционального варианта. Указанный функциональный вариант также может представлять собой набор из двух В-доменов, идентичных по меньшей мере на 80% (предпочтительно идентичных по меньшей мере на 85%, более предпочтительно идентичных по меньшей мере на 90%, еще более предпочтительно идентичных по меньшей мере на 95%, еще более предпочтительно идентичных по меньшей мере на 96%, еще более предпочтительно идентичных по меньшей мере на 97%, еще более предпочтительно идентичных по меньшей мере на 98%, еще более предпочтительно идентичных по меньшей мере на 99%, еще более предпочтительно идентичных по меньшей мере на 99,5% и наиболее предпочтительно идентичных по меньшей мере на 99,7%) аминокислотным последовательностям двух В-доменов естественного эндолизина EL10 (SEQ ID NO: 28 и 29), естественного эндолизина EL11 (SEQ ID NO: 30 и 31), естественного эндолизина EL12 (SEQ ID NO: 32 и 33) или естественного эндолизина EL3 (SEQ ID NO: 19 и 20), еще более предпочтительно естественного эндолизина EL10 (SEQ ID NO: 28 и 29);
где эндолизин является функциональным, где функция включает способность лизировать клеточную стенку Gardnerella.
[0024] В одном аспекте настоящего изобретения связывающийся(еся) с клеточной стенкой домен(ы) (В-домен(ы)) содержит(ат) или состоит(ят) из аминокислотной последовательности SEQ ID NO: 19 и/или 20 или любого их функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 19 и/или 20; более предпочтительно содержит или состоит из аминокислотной последовательности SEQ ID NO: 32 и/или 33 или любого их функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 32 и/или 33; еще более предпочтительно содержит или состоит из аминокислотной последовательности SEQ ID NO: 30 и/или 31 или любого их функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 30 и/или 31; еще более предпочтительно содержит или состоит из аминокислотной последовательности SEQ ID NO: 28 и/или 29 или любого их функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 28 и/или 29;
где эндолизин является функциональным, где функция включает способность лизировать клеточную стенку Gardnerella.
[0025] Предпочтительно, чтобы последовательность VNELL или VNKLL, более предпочтительно VNELL, располагалась на С-конце В-домена. В случае присутствия множества В-доменов в В-области также предпочтительно, чтобы последовательность VNELL или VNKLL, более предпочтительно VNELL, располагалась на С-конце каждого В-домена.
[0026] Неожиданным и удивительным образом в контексте настоящего изобретения было обнаружено, что несколько рекомбинантных эндолизинов обладают более сильной активностью, чем естественные эндолизины, особенно если рассматривать все 4 протестированных штамма Gardnerella (то есть Gardnerella vaginalis sensu strict, Gardnerella leopoldii, Gardnerella piotii и Gardnerella swidsinskii). В частности, каждый из Н2В10, Н2В11, Н2В12 и Н7В3 оказался более активным по сравнению со всеми протестированными естественными эндолизинами. Таким образом, рекомбинантные эндолизины по настоящему изобретению проявляют значительно более высокую активность, чем естественные эндолизины.
[0027] Таким образом, предпочтительно, чтобы «уничтожающая активность в отношении Gardnerella» рекомбинантного эндолизина по изобретению была увеличена по сравнению с уничтожающей активностью естественных эндолизинов, например естественных эндолизинов EL1 -EL12 (имеющих аминокислотные последовательности, показанные в Таблице 7).
[0028] В соответствии со значительно высокой активностью эндолизинов Н2В10, Н2В11, Н2В12 и Н7В3, эти эндолизины (и их функциональные варианты) являются предпочтительными в настоящем изобретении. Таким образом, эндолизин по настоящему изобретению предпочтительно имеет:
(1) N-концевой каталитический домен, состоящий из полипептида, содержащего или состоящего из аминокислотной последовательности SEQ ID NO: 2 или 7 или любого его функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 2;
(2) С-концевую связывающуюся с клеточной стенкой область, содержащую или состоящую из по меньшей мере одного (предпочтительно двух) связывающегося(ихся) с клеточной стенкой домена(ов), независимо выбранного(ых) из группы, состоящей из полипептидов, содержащих или состоящих из аминокислотной последовательности любой из SEQ ID NO: 19, 20 и 28-33, соответственно, и любого их функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 19 - 20 и 28-33, соответственно; и
(3) линкерную область между N-концевым каталитическим доменом и С-концевой связывающейся с клеточной стенкой областью, состоящую из полипептида, содержащего или состоящего из аминокислотной последовательности X1X2GLNGX3X4NGGS, где X1 представляет собой N или K, предпочтительно N, Х2 представляет собой А, Х3 представляет собой Y и Х4 представляет собой K или Q,
где указанный эндолизин обладает уничтожающей активностью в отношении Gardnerella. В отношении линкерной области указано, что, как упоминается ниже, линкерная область может также состоять из полипептида, содержащего или состоящего из аминокислотной последовательности (ХХХ)n, где каждый X может независимо представлять собой G, А или S, предпочтительно где аминокислотная последовательность (ХХХ)n представляет собой (GGS)n, где n соответствует числу повторов последовательности XXX, предпочтительно где n представляет собой 2, 3, 4, 5 или 6.
[0029] В прилагаемых Примерах показано, что рекомбинантный эндолизин Н2В10 имеет наиболее высокую активность. Следовательно, в настоящем изобретении наиболее предпочтительно, чтобы эндолизин по настоящему изобретению представлял собой Н2В10 (или его функциональный вариант). Таким образом, эндолизин по настоящему изобретению наиболее предпочтительно имеет:
(1) N-концевой каталитический домен, состоящий из полипептида, содержащего или состоящего из аминокислотной последовательности SEQ ID NO: 2, или любого его функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичный по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 2;
(2) С-концевую связывающуюся с клеточной стенкой область, содержащую или состоящую из двух связывающихся с клеточной стенкой доменов, независимо выбранных из группы, состоящей из полипептидов, содержащих или состоящих из аминокислотной последовательности SEQ ID NO: 28 или 29, или любого их функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 28 или 29, и
(3) линкерной области между N-концевым каталитическим доменом и С-концевой связывающейся с клеточной стенкой областью, состоящую из полипептида, содержащего или состоящего из аминокислотной последовательности X1X2GLNGX3X4NGGS, где X1 представляет собой N, Х2 представляет собой А, Х3 представляет собой Y и Х4 представляет собой K,
где указанный эндолизин обладает уничтожающей активностью в отношении Gardnerella. Как упоминалось выше, предпочтительно, чтобы «уничтожающая активность в отношении Gardnerella» рекомбинантного эндолизина по изобретению была увеличена по сравнению с уничтожающей активностью естественных эндолизинов, например естественных эндолизинов EL1-EL12 (имеющих аминокислотные последовательности, показанные в Таблице 7).
В отношении линкерной области указано, что, как упоминается ниже, линкерная область может также состоять из полипептида, содержащего или состоящего из аминокислотной последовательности (ХХХ)n, где каждый X может независимо представлять собой G, А или S, предпочтительно где аминокислотная последовательность (ХХХ)n представляет собой (GGS)n, где n соответствует числу повторов последовательности XXX, предпочтительно где n представляет собой 2, 3, 4, 5 или 6.
[0030] В рекомбинантном эндолизине по настоящему изобретению С-концевая связывающаяся с клеточной стенкой область может содержать или состоит из одного, двух или трех связывающихся с клеточной стенкой доменов. Указанные один, два или три связывающихся с клеточной стенкой домена могут быть независимо выбраны из группы, состоящей из полипептидов, содержащих или состоящих из аминокислотной последовательности из SEQ ID NO: 15-24 и 26-33, соответственно, и любого их варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности из SEQ ID NO: 15-24 и 26-33, соответственно, где указанные полипептиды являются функциональными, где функция включает способность связываться с клеточной стенкой Gardnerella. Предпочтительно, чтобы С-концевая связывающаяся с клеточной стенкой область состояла из двух связывающихся с клеточной стенкой доменов. Предпочтительные С-концевые области связывания с клеточной стенкой определены в данном документе выше и ниже.
[0031] Эндолизин по настоящему изобретению предпочтительно не содержит Н-домен или В-область естественного эндолизина EL6. Аминокислотные последовательности Н-домена и В-области естественного эндолизина EL6 приведены в Таблице 7.
[0032] Линкерная область может состоять из полипептида, имеющего длину от 6 до 18 аминокислот, предпочтительно длину от 9 до 15 аминокислот, еще более предпочтительно длину 12 аминокислот. Предпочтительно, линкерная область может состоять из полипептида, содержащего или состоящего из аминокислотной последовательности (1) (ХХХ)n, где каждый X может независимо представлять собой G, А или S, предпочтительно где аминокислотная последовательность (ХХХ)n представляет собой (GGS)n, где n соответствует числу повторов последовательности XXX, предпочтительно где n представляет собой 2, 3, 4, 5 или 6, или (2) X1X2GLNGX3X4NGGS, где X1 представляет собой N или K, Х2 представляет собой А или V, Х3 представляет собой Y или С и Х4 представляет собой K или Q. Как описано выше, в одном аспекте N-концевой каталитический домен эндолизина по настоящему изобретению идентичен первому естественному эндолизину или имеет происхождение из первого естественного эндолизина, линкерная область и С-концевая связывающаяся с клеточной стенкой область идентичны второму естественному эндолизину или имеют происхождение из второго естественного эндолизина, и первый и второй естественные эндолизины закодированы разными геномами из разных профагов.
[0033] Рекомбинантный эндолизин по настоящему изобретению обладает уничтожающей активностью в отношении Gardnerella. Например, эндолизин по настоящему изобретению может обладать уничтожающей активностью в отношении Gardnerella vaginalis sensu stricto, Gardnerella leopoldii, Gardnerella piotii и/или Gardnerella swidsinskii, предпочтительно всех из них. Уничтожающая активность эндолизинов по изобретению в отношении Gardnerella, описанная выше, предпочтительно представляет собой избирательную уничтожающую активность в отношении рода Gardnerella. В данном документе «избирательная уничтожающая активность в отношении рода Gardnerella» означает, что эндолизин по настоящему изобретению не обладает уничтожающей активностью в отношении бактерий в целом. Предпочтительно, эндолизин по настоящему изобретению обладает уничтожающей активностью в отношении Gardnerella, но не в отношении Lactobacilli. В частности, предпочтительно, чтобы указаный эндолизин не обладал уничтожающей активностью в отношении Lactobacilli crispatus, Lactobacilli gasseri и/или Lactobacilli jensenii. Более предпочтительно, чтобы указаный эндолизин не обладал уничтожающей активностью в отношении всех этих Lactobacilli, то есть Lactobacilli crispatus, Lactobacilli gasseri и Lactobacilli jensenii.
[0034] Изобретение также относится к молекуле полинуклеотида, кодирующей эндолизин, как описано выше. Молекула нуклеиновой кислоты может представлять собой ДНК, например, кДНК (комплементарная ДНК) или РНК. В данном документе термины «полинуклеотид» или «молекула полинуклеотида» используются как синонимы с термином «молекула нуклеиновой кислоты» или подобными ему.
[0035] Изобретение также относится к вектору, содержащему указанную молекулу полинуклеотида по изобретению. В одном воплощении вектор представляет собой экспрессирующий вектор. Можно использовать любой подходящий вектор, известный в данной области, такой как вектор серии рЕТ и все векторы на основе Т7. Например, вектор может представлять собой плазмиду. Так, в одном аспекте настоящее изобретение относится к плазмиде, содержащей полинуклеотид по изобретению. Специалистам в данной области будет понятно, что выбор экспрессирующего вектора может определяться выбором клетки-хозяина.
[0036] В настоящем изобретении также предложена клетка-хозяин, содержащая молекулу полинуклеотида по изобретению или вектор/плазмиду по изобретению. В одном воплощении клетка-хозяин представялет собой микробную клетку, например бактериальную клетку. Предпочтительно, клетка-хозяин является непатогенной. Наиболее предпочтительно, клетка-хозяин представляет собой Е. coll. Таким образом, один аспект изобретения относится к бактериальной клетке-хозяину, содержащей плазмиду по изобретению, предпочтительно где бактериальная клетка-хозяин представляет собой клетку Е. coli.
[0037] Настоящее изобретение также охватывает способ получения эндолизина по изобретению, включающий культивирование популяции клеток-хозяев, содержащих молекулу полинуклеотида по изобретению или вектор/плазмиду по настоящему изобретению в условиях, в которых экспрессируется эндолизин, и выделение из них эндолизина.
[0038] В еще одном аспекте изобретения предложена фармацевтическая композиция, содержащая
(а) эндолизин по изобретению;
(б) молекулу полинуклеотида по изобретению;
(в) вектор/плазмиду по изобретению;
(г) хозяина по изобретению; и/или
(д) бактериофаг, способный экспрессировать эндолизин по изобретению,
и фармацевтически приемлемый носитель, разбавитель или эксципиент.
Например, фармацевтическая композиция по настоящему изобретению может содержать эндолизин по изобретению, молекулу полинуклеотида по изобретению и фармацевтически приемлемый носитель и/или разбавитель.
[0039] Еще один аспект изобретения относится к
(а) эндолизину по изобретению;
(б) молекуле полинуклеотида по изобретению;
(в) вектору/плазмиде по изобретению;
(г) хозяину по изобретению;
(д) бактериофагу, способному экспрессировать эндолизин по изобретению; и/или
(е) фармацевтической композиции по изобретению
для применения в лечении заболевания или нарушения. Например, в изобретении предложен эндолизин по изобретению, молекула полинуклеотида по изобретению или фармацевтическая композиция по изобретению для применения при лечении заболевания или нарушения.
Указанное заболевание или нарушение может представлять собой бактериальную инфекцию, предпочтительно бактериальный вагиноз. Например, бактериальный вагиноз может быть вызван Gardnerella vaginalis sensu stricto, Gardnerella leopoldii, Gardnerella piotii и/или Gardnerella swidsinskii.
[0040] В одном аспекте настоящего изобретения рекомбинантный эндолизин по изобретению, молекулу полинуклеотида по изобретению или фармацевтическую композицию по изобретению следует вводить местно, предпочтительно местно во влагалище субъекта. Так, в одном аспекте настоящего изобретения рекомбинантный эндолизин по изобретению, полинуклеотид по изобретению или фармацевтическую композицию по изобретению следует вводить во влагалище субъекта.
[0041] Приложенные Примеры показывают, что активность рекомбинантных эндолизинов по настоящему изобретению особенно высока при рН, составляющем приблизительно 5. Таким образом, один аспект настоящего изобретения относится к рекомбинантному эндолизину по изобретению, молекуле полинуклеотида по изобретению или к фармацевтической композиции по изобретению, где указанный рекомбинантный эндолизин, полинуклеотид или фармацевтическую композицию следует вводить совместно с соединением или композицией, доводящими рН влагалища до 4,0-6,0, предпочтительно до 4,5-5,5, более предпочтительно до приблизительно 5. Подходящие соединения или композиции, которые доводят рН влагалища, включают фосфат, молочную кислоту (например, естественное подкисляющее вещество, секретируемое Lactobacilli для создания кислой среды) или другие органические кислоты, например карбоксизамещенные полимеры, но не ограничены ими.
[0042] Еще один аспект изобретения относится к
(а) эндолизину по изобретению;
(б) молекуле полинуклеотида по изобретению;
(в) вектору/плазмиде по изобретению;
(г) хозяину по изобретению;
(д) бактериофагу, способному экспрессировать эндолизин по изобретению; и/или
(е) фармацевтической композиции по изобретению
для применения в качестве лекарственного средства.
[0043] Еще один аспект изобретения относится к применению
(а) эндолизина по изобретению;
(б) молекулы полинуклеотида по изобретению;
(в) вектора/плазмиды по изобретению;
(г) хозяина по изобретению;
(д) бактериофага, способного экспрессировать полипептид по изобретению; и/или
(е) фармацевтической композиции по изобретению
в изготовлении лекарственного средства для лечения бактериальных инфекций и расстройств.
[0044] В еще одном аспекте изобретения предложен способ лечения бактериальных инфекций и расстройств, таких как BV, включающий введение нуждающемуся в этом субъекту терапевтически эффективного количества
(а) эндолизина по изобретению;
(б) молекулы полинуклеотида по изобретению;
(в) вектора/плазмиды по изобретению;
(г) хозяина по изобретению;
(д) бактериофага, способного экспрессировать полипептид по изобретению; и/или
(е) фармацевтической композиции по изобретению.
В некоторых воплощениях терапевтически эффективное количество представляет собой дозу от 10 до 100 мкг эндолизина, возможно для введения несколько раз в сутки.
[0045] В еще одном аспекте изобретения предложен набор, содержащий эндолизин, описанный в данном документе, и инструкции по применению, в частности, для лечения заболевания или нарушения, предпочтительно BV, как указано выше. Указанный набор может также содержать соединение или композицию, которые доводят рН влагалища до 4,0-6,0, предпочтительно до 4,5-5,5, более предпочтительно до приблизительно 5. Определения и предпочтительные аспекты, указанные в данном документе выше и ниже в отношении эндолизина по настоящему изобретению, применяются с соответствующими изменениями также к молекуле полинуклеотида, вектору/плазмиде, клетке-хозяину, фармацевтической композиции, способу лечения и набору по настоящему изобретению.
В еще одном аспекте изобретения предложен способ in vitro диагностирования заболевания или состояния, которое можно лечить эндолизином по настоящему изобретению, где указанный способ включает стадии:
(1) приведения полученного от субъекта образца в контакт с полипептидом, содержащим или состоящим из С-концевой связывающейся с клеточной стенкой области эндолизина по настоящему изобретению и возможно N-концевого каталитического домена эндолизина по настоящему изобретению, где образец содержит микробные клетки, и где С-концевая связывающаяся с клеточной стенкой область указанного эндолизина возможно является меченой;
(2) тестирования, связывается ли полипептид с микробными клетками образца и/или лизирует ли их; и
(3) установления того, что указанное заболевание или состояние можно лечить эндолизином по настоящему изобретению, если полипептид связывается с микробными клетками образца и/или лизирует их.
Микробные клетки могут быть клетками Gardnerella, предпочтительно клетками G. vaginalis sensu stricto, G. leopoldii, G. piotii, G. swidsinskii или другими видами рода Gardnerella.
Другие признаки и преимущества настоящего изобретения станут очевидны исходя из следующего подробного описания.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0046] На Фиг. 1 показано выравнивание последовательностей естественных эндолизинов профага Gardnerella по настоящему изобретению (множественное выравнивание последовательностей CLUSTAL O(1.2.4)). Большинство эндолизинов имеет 306 остатков, за исключением двух, которые имеют 251 остаток.
[0047] На Фиг. 2 показано филогенетическое древо естественных эндолизинов профага Gardnerella по настоящему изобретению. Среди эндолизинов нет идентичных пар, несмотря на их высокую гомологию.
[0048] На Фиг. 3 показана доменная структура эндолизинов профагов Gardnerella по настоящему изобретению, установленная при помощи InterPro (Mitchell et al., 2019, Nucleic Acids Res. 47, D351-D360). N-концевая часть эндолизинов из 196 остатков идентифицирована как каталитический домен благодаря ее гомологии с гидролазами гликозидов, семейством 25. За каталитическим доменом следует линкерная область и два домена, которые идентифицированы как два связывающих клетки домена вследствие их гомологии с С-концевым доменом лизоцима Cpl-7 (домен CW_7). Согласно номенклатуре настоящей заявки, каталитический домен представляет собой гидролазу или «Н-домен», в то время как линкерная область и связывающиеся с клеточной стенкой домены вместе представляют собой связывающую или «В-область».
[0049] На Фиг. 4А-4С показаны три анализа ферментативной активности, где ферментативную активность естественных эндолизинов профагов Gardnerella по настоящему изобретению измеряли путем выявления изменения мутности суспензии клеток Gardnerella. На Фиг. 4А ферментативную активность эндолизинов измеряли путем выявления изменения мутности суспензии G. leopoldii штамма Gv_10 при рН 6,0. На Фиг. 4В ферментативную активность эндолизинов измеряли путем выявления изменения мутности суспензии G. piotii штамма Gv_17 при рН 7,0. На Фиг. 4С ферментативную активность эндолизинов измеряли путем выявления изменения мутности суспензии G. swidsinskii штамма Gv_23 при рН 7,4. Воздействие осуществляли в среде с доведенным до соответствующего значения рН, в фотометрической кювете против буфера. Затем оценивали изменение мутности путем измерения оптической плотности (OD) при 600 нм. В результате падение мутности было более выраженным для групп, находившихся под воздействием эндолизина, а не буфера, что указывает на ферментативную активность.
[0050] На Фиг. 5 показано снижение количества жизнеспособных колониеобразующих единиц (CFU) в исследовании, где сравнивали не испытывавшие воздействия клетки G. vaginalis sensu stricto штамма Gv_9, которые инкубировали в среде с имидазолом или без имидазола при различных значениях рН. 5×107 CFU/мл клеток инкубировали в условиях, указанных под графиком, в течение 5 часов при 37°С в анаэробных условиях, после чего определяли количество выживших CFU/мл путем количественного посева. Результаты показывают, что выживаемость G. vaginalis Gv_9 в протестированных условиях в высокой степени зависит от отсутствия имидазола и низкого рН.
[0051] На Фиг. 6 показано снижение количества жизнеспособных колониеобразующих единиц (CFU) в исследовании, где сравнивали клетки G. vaginalis sensu stricto штамма Gv_9, находившиеся под воздействием раствора элюата, содержащего рекомбинантный эндолизин Н10В1 и 250 мМ имидазола, при различных значениях рН, или контроля, содержащего 250 мМ имидазола, при различных значениях рН. 5×107 CFU/мл клеток инкубировали в условиях, указанных под графиком, в течение 5 часов при 37°С в анаэробных условиях, после чего определяли количество выживших CFU/мл путем количественного посева. Столбцы, обозначенные контролем с имидазолом, отображают те же данные, что и на Фиг. 5. Результаты показывают, что ферментативная активность Н10В1, взятого в качестве примера для всех эндолизинов по изобретению, выше при низких значениях рН, причем наиболее сильная активность наблюдалась при рН 5,5 и рН 5,0.
[0052] На Фиг. 7A-7D показано снижение количества жизнеспособных колониеобразующих единиц (CFU) в четырех исследованиях, где измеряли уничтожающую активность естественных и рекомбинантных эндолизинов профага Gardnerella по настоящему изобретению в отношении четырех основных видов Gardnerella. На Фиг. 7А, 7В, 7С и 7D уничтожающую активность эндолизинов измеряли путем выявления жизнеспособных CFU в суспензиях G. vaginalis sensu stricto штамма Gv_9, G. leopoldii штамма Gv_11, G. piotii штамма Gv_17 и G. swidsinskii штамма Gv 23, соответственно. 90 мкл 5e7 CFU/мл указанного штамма инкубировали в течение 5 часов при рН 5,0 в анаэробных условиях с 10 мкл раствора эндолизина (концентрацию доводили до 0,2 мг/мл, где это возможно, см. Таблицу 4). По логарифмической оси Y обозначено количество выживших клеток. Пунктирная линия указывает предел обнаружения (LOD), полученный при посеве 2 мкл реакционной смеси (500 CFU/мл). Результаты показывают, что эндолизины по настоящему изобретению обладают способностью лизировать четыре основных вида Gardnerella. Результаты также указывают на то, что некоторые рекомбинантные эндолизины по изобретению обладают более высокой уничтожающей активностью, чем естественные эндолизины по изобретению.
[0053] На Фиг. 8 показано филогенетическое древо (аминокислотный уровень) Н-доменов, построенное с помощью Clustal Omega (Sievers et at, 2011 Mol. Syst. Biol. 7, 539).
[0054] На Фиг. 9 и Фиг. 10 показано филогенетическое древо (аминокислотный уровень) В-областей, построенное с помощью Clustal Omega (Sievers et al., 2011 Mol. Syst. Biol. 7, 539).
[0055] На Фиг. 11 показано выравнивание последовательностей связывающихся с клеточной стенкой доменов (также обозначаемых как В-домены) в составе В-областей естественных эндолизинов по изобретению, построенное при помощи Clustal Omega (Sievers et al., 2011 Mol. Syst. Biol. 7, 539). Для каждой B-области N-концевой связывающийся с клеточной стенкой домен обозначен суффиксом _N (Вх N), а С-концевой связывающийся с клеточной стенкой домен обозначен суффиксом _С (Вх_С). Например, В3_С обозначает вторые (С-концевые) В-домены В3.
[0056] На Фиг. 12 показано филогенетическое древо отдельных В-областей, построенное с помощью Clustal Omega (Sievers et al., 2011 Mol. Syst. Biol. 7, 539).
[0057] На Фиг. 13 показано снижение количества жизнеспособных колониеобразующих единиц (CFU) в трех исследованиях, где измеряли уничтожающую активность рекомбинантных эндолизинов профагов Gardnerella по настоящему изобретению в отношении трех наиболее часто встречающихся видов полезных Lactobacilli при рН 5,0 при анаэробных условиях. Результаты показывают, что эндолизины по изобретению не эффективны против полезных штаммов Lactobacilli.
[0058] На Фиг. 14 показано определение MIC (минимальной ингибирующей концентрации) методом микроразведений в бульоне, где измеряли влияние метронидазола и клиндамицина (полученных от компании Ratiopharm в виде раствора для инъекций, 300 мг/2 мл) на рост в суспензии четырех основных видов Gardnerella. 2,5×107 CFU/мл суспензий Gardnerella инкубировали с антибиотиками в концентрациях, указанных на оси х каждого графика, и инкубировали при 37°С в течение 48 ч. Для определения минимальной ингибирующей концентрации (MIC) оценивали рост клеток путем измерения оптической плотности при 610 нм (OD (610)) до и после инкубации. Результаты показывают, что все штаммы Gardnerella являются устойчивыми как к метронидазолу, так и к клиндамицину (полученным от компании Ratiopharm в виде раствора для инъекций, 300 мг/2 мл), демонстрируя MIC от 64 до <128 мкг/мл и 16 мкг/мл, соответственно (Фиг. 14).
На Фиг. 15 показано определение MIC методом микроразведений в бульоне, где исследовали влияние метронидазола и гидрохлорида клиндамицина (полученных от компании Sigma Aldrich) на суспензии Gardnerella 1×105 - 1×106 CFU/мл. На этот раз результаты показывают, что MIC метронидазола в отношении всех протестированных штаммов Gardnerella составляла от 8 до 128 мкг/мл, а у гидрохлорида клиндамицина в порошке (полученного от Sigma Aldrich (C5269-10MG)) наблюдали MIC от 0,25 до 5 мкг/мл.
На Фиг. 16 показано определение MIC методом микроразведений в бульоне, где исследовали влияние Н2В10, репрезентативного для заявленных здесь эндолизинов с замененными доменами, на рост трех основных видов Gardnerella. 1×105 - 1×106 CFU/мл суспензий Gardnerella инкубировали с Н2В10 в концентрациях, указанных на оси х каждого графика, и инкубировали при 37°С в течение 48 ч в анаэробных условиях. Для определения минимальной ингибирующей концентрации (MIC) оценивали рост клеток путем измерения OD(610) до и после инкубации. Полученные значения MIC от 1 до 4 мкг/мл указывали, что все штаммы Gardnerella в высокой степени чувствительны к эндолизину Н2В10 с замененными доменами.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
[0059] Термин «лизины» относится к ферментам, лизирующим клеточную стенку, кодируемым бактериофагами (эндолизины) или бактериями (автолизины), которые обладают способностью гидролизовать клеточную стенку бактерий-мишеней при экзогенном добавлении (лизис извне). Этот новый класс антибактериальных агентов имеет важные преимущества перед классическими антибиотиками, например новый механизм действия; узкий спектр чувствительных бактерий; быстрое уничтожение бактерий как в стационарной, так и в экспоненциальной фазе роста; активность на слизистых оболочках и бактериальных биопленках; низкая вероятность развития устойчивости; а также снижение воздействия на нормальную микробиоту. Эти уникальные характеристики повысили интерес к биотехнологическому и фармакологическому использованию лизинов и их недавнему включению в число лучших современных альтернатив для борьбы с устойчивостью к антибиотикам. Лизины из грамположительных бактерий и их фагов обычно содержат по меньшей мере один каталитический домен и один более доменов, связывающихся с клеточной стенкой. В отличие от этого, многие лизины, продуцируемые грамотрицательными видами или их фагами, содержат только каталитический домен, хотя сообщали также о модульных эндолизинах. Каталитические единицы определяют тип расщепляемой связи пептидогликана (PG), тогда как домен(ы), связывающийся(еся) с клеточной стенкой, в значительной степени определяет(ют) литический спектр благодаря специфическому распознаванию элементов клеточной стенки, распределенных специфическим для рода или вида/штамма образом.
[0060] В контексте настоящего описания термин «естественный эндолизин» относится к эндолизину, кодируемому последовательностью профага в бактериальном геноме, в частности в геноме клеток Gardnerella. В контексте настоящего описания термин «естественный эндолизин», следовательно, относится к эндолизину, в котором не произошло замены доменов. Естественный эндолизин может быть немодифицированным, что означает, что аминокислотная последовательность эндолизина соответствует нативной последовательности. В качестве альтернативы, естественный эндолизин может быть модифицирован, что означает, что аминокислотная последовательность эндолизина содержит по меньшей мере одну мутацию по сравнению с нативной последовательностью. Аминокислотные последовательности естественных эндолизинов Е1-Е14 показаны в Таблице 7 ниже. Также представлен пример известной последовательности 1,4-бета-N-ацетилмур амид азы (естественный эндолизин EL1) под номером WP_014554482 (версия WP_0145 54482.1 от 27 мая 2013 г.) в базе NCBI.
[0061] В контексте настоящего описания термин «рекомбинантный эндолизин» относится к эндолизину, в котором произведена замена доменов. В контексте настоящего описания термин «эндолизин с заменными доменами» относится к эндолизину, который имеет N-концевой каталитический домен из первого естественного эндолизина и по меньшей мере один связывающийся с клеточной стенкой домен из второго естественного эндолизина, где первый и второй естественный эндолизин закодированы разными геномами разных профагов. Рекомбинантный эндолизин по изобретению может содержать N-концевой каталитический домен из первого естественного эндолизина и два связывающихся с клеточной стенкой домена из второго естественного эндолизина, где первый и второй естественный эндолизины закодированы разными геномами разных профагов, или состоять из них. В качестве альтернативы, рекомбинантный эндолизин по изобретению может содержать N-концевой каталитический домен из первого естественного эндолизина, первый (N-концевой) связывающийся с клеточной стенкой домен из второго естественного эндолизина и второй (С-концевой) связывающийся с клеточной стенкой домен из третьего естественного эндолизина, где первый и второй естественный эндолизин закодированы различными геномами из разных профагов, и где третий естественный эндолизин возможно закодирован другим геномом другого профага, нежели первый и второй естественный эндолизин, или состоять из них. Рекомбинантный эндолизин может быть немодифицированным, что означает, что аминокислотная последовательность эндолизина соответствует нативной последовательности соответствующих доменов, составляющих эндолизин. В качестве альтернативы, рекомбинантный эндолизин может быть модифицированным, что означает, что аминокислотная последовательность эндолизина содержит по меньшей мере одну мутацию относительно нативной последовательности соответствующих доменов, составляющих эндолизин. В соответствии с этим определением специалист в данной области легко поймет, что эндолизины «с замененными доменами», или «рекомбинантные» эндолизины, описанные в данном документе, являются эндолизинами, не встречающимися в природе. То есть рекомбинантный эндолизин по настоящему изобретению был модифицирован вручную человеком, что по определению исключает естественные эндолизины, то есть такие, которые могут естественным образом встречаться в природе. В прилагаемых примерах представлен(ы) подходящий(ие) способ(ы) получения искусственного эндолизина по изобретению.
[0062] Термины «каталитический домен» или «ферментативный домен» относятся к части белковой цепи, которая содержит область, в которой происходит катализируемая химическая реакция.
В контексте настоящего описания термин «Н-домен» относится к части эндолизина по изобретению, содержащей каталитический домен.
[0063] В контексте настоящего описания термин «В-область» относится к части эндолизина по изобретению, содержащей или состоящей из полипептида, обладающего связывающей клеточную стенку активностью. В предпочтительном воплощении В-область содержит или состоит из линкерной области и одного, двух или трех доменов, связывающихя с клеточной стенкой, или «В-доменов».
[0064] В контексте настоящего изобретения термин «В-домен» относится к домену, связывающемуся с клеточной стенкой, входящему в состав В-области.
[0065] В контексте настоящего изобретения термин «домен CW 7» относится к связывающемуся с клеточной стенкой домену белка Cpl-7, то есть эндолизина, кодируемого бактериофагом Ср-7 Streptococcus pneumoniae (см. Bustamante et al., 2010 J. Biol. Chem. 285, 33184-33196, 2012 PLoS One 7, e46654). Кратко, белок Cpl-7 имеет связывающуюся с клеточной стенкой С-концевую область, состоящую из 3 последовательно расположенных доменов CW_7. Все домены CW 7 представляют собой одинаковые аминокислотные последовательности длиной 38 аминокислот, обозначаемые как «мотив CW_7» и определяемые при помощи hiterpro (Mitchell et al. 2019, Nucleic Acids Res. 47, D351-D360) как состоящие из аминокислотной последовательности TVANEVIQGLWGNGQERYDSLANAGYDPQAVQDKVNEXL, где X представляет собой I в мотивах CW_7 No: 1 (аминокислоты 207-245) и No: 2 (аминокислоты 255-293) и где X представляет собой L в мотиве CW 7 No: 3 (аминокислоты 303-341). В белке Cpl-7 имеются короткие линкеры из 9 остатков между мотивами CW 7 No: 1 и No: 2, а также между мотивами CW_No: 2 и No: 3, так что общая длина повтора составляет 47 остатков. Для сравнения, повторы естественных эндолизинов по настоящему изобретению имеют длину 49 остатков.
[0066] Термины «Минимальная ингибирующая концентрация», или «MIC» относятся к самой низкой концентрации химического вещества, обычно лекарственного средства, которая предотвращает видимый рост бактерий. В настоящей заявке MIC определяли как минимальную концентрацию антибиотика, при которой рост через 48 часов не поддавался выявлению путем измерения OD.
[0067] Термины «Минимальная бактерицидная концентрация», или «МВС» относятся к самой низкой концентрации антибактериального агента, необходимой для уничтожения конкретной бактерии. Обычно измеряют МВС90, то есть концентрацию антибиотика, уничтожающую 90% клеток в течение определенного времени, в то время как в настоящей заявке МВС определяют как минимальную концентрацию, обеспечивающую полную эрадикацию суспензии 2,5×107 CFU/мл. В то время как MIC представляет собой самую низкую концентрацию антибактериального агента, необходимая для подавления видимого роста, МВС представляет собой минимальную концентрацию антибактериального агента, которая приводит к гибели всех бактериальных клеток в суспензии.
[0068] Термины «пептид», «полипептид», «белок» и варианты этих терминов относятся к пептиду, олигопептиду, олигомеру или белку, в том числе слитому белку, соответственно, содержащим по меньшей мере две аминокислоты, соединенные друг с другом нормальной или модифицированной пептидной связью, как, например, в случае изостерических пептидов. Эти термины также включают в себя «пептидомиметики», которые определяют как аналоги пептидов, содержащие непептидные структурные элементы, которые способны имитировать биологическое(ие) действие(я) родительского естественного пептида или быть их антагонистом. У пептидомиметика отсутствуют классические характеристики пептидов, такие как расщепляющиеся под действием ферментов пептидные связи. Пептид или полипептид может состоять из аминокислот, отличных от 20 аминокислот, определяемых генетическим кодом. Он может состоять из L-аминокислот и/или D-аминокислот. Пептид или полипептид могут в равной степени состоять из аминокислот, модифицированных естественными процессами, такими как процессы посттрансляционного созревания, или химическими процессами, которые хорошо известны специалисту в данной области. Такие модификации полностью подробно описано в литературе. Эти модификации могут появляться в любом месте полипептида: в пептидном скелете, в аминокислотной цепи или даже на карбокси- или амино-концах. Пептид или полипептид может стать разветвленным после убиквитинирования или быть циклическим с разветвлением или без разветвления. Этот тип модификации может быть результатом естественных или искусственных посттрансляционных процессов, хорошо известных специалисту в данной области. Например, модификации пептида или полипептида могут включать ацетилирование, ацилирование, ADP(аденозиндифосфат)-рибозилирование, амидирование, ковалентное присоединение нуклеотида или производного нуклеотида, ковалентное присоединение липида или производного липида, ковалентное присоединение фосфатидилинозитола, ковалентное или нековалентное поперечное сшивание, циклизацию, образование дисульфидной связи, деметилирование, гликозилирование, включая пегилирование, гидроксилирование, йодирование, метилирование, миристоилирование, окисление, протеолитические процессы, фосфорилирование, пренилирование, рацемизацию, сенелоилирование, сульфатирование, добавление аминокислот, такое как аргинилирование, или убиквитинирование. Такие модификации очень подробно описаны в литературе и хорошо известны специалистам в данной области техники.
[0069] Используемый в данном документе термин «бактериальные инфекции и нарушения» относится к инфекциям и нарушениям, вызванным бактериями, в частности, инфекциям и нарушениям, вызванным по меньшей мере одним штаммом рода Gardnerella, выбранным из группы, состоящей из Gardnerella vaginalis sensu strict, Gardnerella leopoldii, Gardnerella piotii и Gardnerella swidsinskii, и других видов Gardnerella. Бактериальные инфекции и нарушения включают бактериальный вагиноз (BV), но не ограничиваются им.
[0070] В данном документе термин «уничтожающая активность» эндолизина в отношении конкретных бактерий представляет собой уменьшение количества жизнеспособных бактериальных клеток, вызванное лизирующей активностью указанного эндолизина. Уничтожающая активность эндолизина в отношении указанных бактерий может быть полной, что означает, что 100% бактериальных клеток были лизированы, или частичной, что означает, что по меньшей мере приблизительно 80%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 99%, или по меньшей мере приблизительно 99,9% бактериальных клеток были лизированы. Уничтожающую активность можно определять, измеряя уменьшение оптической плотности суспензии бактериальных клеток при 610-620 нм и/или уменьшение количества колониеобразующих единиц (CFU) на миллилитр суспензии бактериальных клеток после воздействия исследуемого эндолизина.
[0071] В данном документе термин «связывающая способность» эндолизина по отношению к клеточной стенке конкретных бактерий относится к способности указанного эндолизина специфически взаимодействовать с клеточной стенкой указанных бактерий и прикрепляться к ней. Способность эндолизина связываться с клеточной стенкой бактерий можно определять способами, известными в данной области техники.
[0072] Используемые в данном документе термины «лечение» и «лечить» и им подобные обычно означают достижение желаемого фармакологического и физиологического эффекта. Эффект может быть профилактическим в плане предотвращения или частичного предотвращения заболевания, симптома или состояния и/или может быть терапевтическим в плане частичного или полного излечения заболевания, состояния, симптома или неблагоприятного эффекта, приписываемого заболеванию. Термин «лечение», используемый в данном документе, охватывает любое лечение заболевания у млекопитающего, в частности человека, и включает: (а) предотвращение возникновения заболевания у субъекта, который может быть предрасположен к заболеванию, но у которого еще не диагностировано его наличие; (б) ингибирование заболевания, то есть остановку его развития; или облегчение заболевания, то есть вызывание обратного развития заболевания и/или его симптомов или состояний, например улучшение или устранение повреждений. В частности, лечение бактериальных инфекций включает предотвращение, уменьшение или даже эрадикацию инфекции, например путем уничтожения бактерий и, таким образом, контроля, уменьшения или подавления пролиферации бактерий, а также уменьшения количества жизнеспособных бактериальных клеток. При этом предпочтительно, чтобы заболевание, например BV, подвергалось терапевтическому лечению, что выражалось бы в частичном или полном излечении заболевания или симптомов.
[0073] Термин «субъект», используемый в данном документе, относится к млекопитающим. Например, млекопитающие, рассматриваемые в настоящем изобретении, включают человека, приматов, домашних животных, таких как крупный рогатый скот, овцы, свиньи, лошади, лабораторные мыши и тому подобные. Предпочтительно, чтобы субъектом являлся человек.
[0074] Используемый в данном документе термин «эффективное количество» относится к количеству по меньшей мере одного эндолизина согласно изобретению, его композиции или фармацевтического состава, которое вызывает искомый биологический или медицинский ответ в ткани, системе, животном или человеке. В одном воплощении эффективное количество представляет собой «терапевтически эффективное количество» для облегчения симптомов заболевания или состояния, лечение которого осуществляют. В другом воплощении эффективное количество представляет собой «профилактически эффективное количество» для профилактики симптомов предотвращаемого заболевания или состояния. В данном документе термин также включает количество активного полипептида, достаточное для уменьшения прогрессирования заболевания, в частности для уменьшения или ингибирования расстройства или инфекции и, таким образом, достижения искомого ответа (то есть «эффективное для ингибирования количество»).
[0075] Термин «эффективность» лечения согласно изобретению можно измерять на основании изменений течения заболевания в ответ на применение или способ по изобретению. Эффективность профилактики инфекционных заболеваний в конечном итоге оценивают в эпидемиологических исследованиях на популяциях людей, и она часто коррелирует с титрами нейтрализующих антител в сыворотках и индуцированием многофункциональных патоген-специфичных Т-клеточных ответов. Доклиническая оценка может включать устойчивость к инфекции после заражения инфекционным патогеном. Лечение инфекционного заболевания можно оценивать по подавлению роста патогена или устранению патогена (и, таким образом, невыявлению патогена), оно коррелирует со специфическими к патогену антителами и/или иммунными ответами Т-клеток.
[0076] Термин «биологический материал» относится к любому материалу или образцу, полученному из тела субъекта. Это включает, например, образцы цельной крови, сыворотки, плазмы, мочи, мокроты, слюны, вагинальных мазков или спинномозговой жидкости.
[0077] Термин «неживой материал или поверхность» включает растворы, среду, устройства, предметы, пол, поверхность стола.
[0078] Термин «среда» включает воду, воздух или пищу.
[0079] Термины «фармацевтический состав» или «фармацевтическая композиция» относятся к препаратам, находящимся в такой форме, которая обеспечивает возможность несомненно эффективного проявления биологической активности активного(ых) ингредиента(ов), и не содержащим дополнительных компонентов, которые были бы токсичными для субъектов, которым будут вводить указанную композицию.
[0080] Термин «фармацевтически приемлемый» относится к носителю, состоящему из материала, не являющегося нежелательным с биологической или иной точки зрения.
[0081] Термин «носитель» относится к любым компонентам, присутствующим в фармацевтической композиции, кроме активного агента, и, таким образом, включает разбавители, связующие агенты, смазывающие агенты, разрыхлители, наполнители, красители, смачивающие или эмульгирующие агенты, забуферивающие рН агенты, консерванты и тому подобные.
[0082] Термин «вариант» относится к полипептиду, включающему вставки, делеции и/или замены, либо неконсервативные, либо предпочтительно консервативные, относительно нативной аминокислотной последовательности. Например, полипептид может содержать аминокислотную последовательность которая по меньшей мере на 80% идентична нативной аминокислотной последовательности, предпочтительно идентична по меньшей мере на 85%, более предпочтительно идентична по меньшей мере на 90%, еще более предпочтительно идентична по меньшей мере на 95%, еще более предпочтительно идентична по меньшей мере на 96%, еще более предпочтительно идентична по меньшей мере на 97%, еще более предпочтительно идентична по меньшей мере на 98%, еще более предпочтительно идентична по меньшей мере на 99%, еще более предпочтительно идентична по меньшей мере на 99,5% и наиболее предпочтительно идентична по меньшей мере на 99,7% указанной аминокислотой последовательности. Процент идентичности можно определить способами, хорошо известными в данной области техники, используя подходящие компьютерные программы, например MatGAT 2.0 (Myers and Miller, CABIOS (1989). Предпочтительно, % идентичности устанавливают по всей длине сравниваемых последовательностей. Следует понимать, что процент идентичности рассчитывается по отношению к полипептидам, последовательность которых выровнена оптимально. Фрагмент и варианты аминокислотной последовательности могут быть получены с использованием любого из методов белковой инженерии, направленной эволюции и/или сайт-направленно го мутагенеза, хорошо известных в данной области (см., например, Molecular Cloning: a Laboratory Manual, 3rd edition, Sambrook & Russell, 2001, Cold Spring Harbor Laboratory Press). Специалистам будет понятно, что полипептид по изобретению или его фрагмент, вариант или слияние могут содержать производное нативной аминокислотной последовательности, или ее фрагмента, или варианта либо состоять из него. Химические производные одной или более аминокислот могут быть получены посредством реакции с функциональной боковой группой. Такие дериватизированные молекулы включают, например, те молекулы, в которых свободные аминокислотные группы были дериватизированы с образованием аминогидрохлоридов, п-толуолсульфонильных групп, карбоксибензоксигрупп, f-бутилоксикарбонильных групп, хлор ацетильных групп или формильных групп. Свободные карбоксильные группы могут быть дериватизированы с образованием солей, метиловых и этиловых эфиров или других типов сложных эфиров и гидразидов. Свободные гидроксильные группы могут быть дериватизированы с образованием О-ацильных или О-алкильных производных. В качестве химических производных также включены пептиды, которые содержат встречающиеся в природе аминокислотные производные двадцати стандартных аминокислот. Например, 4-гидроксипролин может заменять пролин; 5-гидроксилизин может заменять лизин; 3-метилгистидин может заменять гистидин; гомосерин может заменять серии, а орнитин лизин. Производные также включают пептиды, содержащие одну или более вставок или делеций, при условии, что сохраняется требуемая активность. Другие включаемые модификации представляют собой амидирование, ацилирование по амино-концу (например, ацетилирование или амидирование тиогликолевой кислоты), амидирование концевого карбоксила (например, аммиаком или метиламином) и подобные концевые модификации. Специалистам в области техники также очевидно, что также могкт быть полезными соединения - пептидомиметики. Таким образом, термин «полипептидом» включает соединения - пептидомиметики, которые проявляют активность эндолизина. Термин «пептидомиметик» относится к соединению, которое имитирует конформацию и желательные свойства конкретного полипептида в качестве терапевтического агента.
Эндолизины по изобретению
[0083] Эндолизин по настоящему изобретению обладает антибактериальной активностью в отношении штаммов Gardnerella. Оптимальные значения рН, при которых эндолизин по изобретению проявляет антибактериальную активность, составляют приблизительно от 4 до 6, предпочтительно приблизительно рН 5. Эндолизин по настоящему изобретению содержит или состоит из
(1) N-концевого каталитического домена или его функционального варианта,
(2) С-концевой связывающейся с клеточной стенкой области или ее функционального варианта, где С-концевая связывающаяся с клеточной стенкой область содержит или состоит из по меньшей мере одного связывающегося с клеточной стенкой домена, и
(3) линкерной области между N-концевым каталитическим доменом и С-концевой связывающейся с клеточной стенкой областью,
и обладает уничтожающей активностью в отношении клеток Gardnerella.
В некоторых воплощениях N-концевой каталитический домен имеет происхождение из первого естественного эндолизина, линкерная область и С-концевая связывающаяся с клеточной стенкой область имеют происхождение из второго естественного эндолизина, и первый и второй естественные эндолизины закодированы разными геномами разных профагов. Предполагается, что уничтожающая активность эндолизинов по изобретению в отношении Gardnerella представляет собой видоспецифичную цитолитическщую активность в отношении Gardnerella.
[0084] N-концевой каталитический домен представляет собой функциональный полипептид, функция которого включает способность лизировать клеточную стенку Gardnerella. N-концевой каталитический домен может представлять собой N-ацетилмурамидазу, N-ацетилмурамоил-L-аланинамидазы, L-аланоил-D-глутамат-эндопептидазы, эндопептидазы межпептидного мостика или N-ацетил-бета-D-глюкозаминидазы. Предпочтительно N-концевой каталитический домен представляет собой N-ацетилмурамидазу, наиболее предпочтительно 1,4-бета-N-ацетилмурамидазу. Например, N-концевой каталитический домен может представлять собой полипептид, содержащий или состоящий из аминокислот любой из SEQ ID NO: 1-5, 7 или 10-12, или любой его функциональный вариант, идентичный по меньшей мере на 80% (предпочтительно идентичный по меньшей мере на 85%, более предпочтительно идентичный по меньшей мере на 90%, еще более предпочтительно идентичный по меньшей мере на 95%, еще более предпочтительно идентичный по меньшей мере на 96%, еще более предпочтительно идентичный по меньшей мере на 97%, еще более предпочтительно идентичный по меньшей мере на 98%, еще более предпочтительно идентичный по меньшей мере на 99%, еще более предпочтительно идентичный по меньшей мере на 99,5% и наиболее предпочтительно идентичный по меньшей мере на 99,7%) аминокислотой последовательности любой из SEQ ID NO: 1-5, 7 или 10-12, где указанный полипептид является функциональным, где функция включает способность лизировать клеточную стенку Gardnerella. Предпочтительно, N-концевой каталитический домен представляет собой полипептид, содержащий аминокислоты с SEQ ID NO: 2 или 7, или любой его функциональный вариант, идентичный по меньшей мере на 80% (предпочтительно идентичный по меньшей мере на 85%, более предпочтительно идентичный по меньшей мере на 90%, еще более предпочтительно идентичный по меньшей мере на 95%, еще более предпочтительно идентичный по меньшей мере на 96%, еще более предпочтительно идентичный по меньшей мере на 97%, еще более предпочтительно идентичный по меньшей мере на 98%, еще более предпочтительно идентичный по меньшей мере на 99%, еще более предпочтительно идентичный по меньшей мере на 99,5% и наиболее предпочтительно идентичный по меньшей мере на 99,7%) аминокислотой последовательности SEQ ID NO: 2 или 7, где указанный полипептид является функциональным, где функция включает способность лизировать клеточную стенку Gardnerella.
[0085] Согласно настоящему изобретению, С-концевая связывающаяся с клеточной стенкой область представляет собой функциональный полипептид, функция которого включает способность связываться с клеточной стенкой Gardnerella. С-концевая связывающаяся с клеточной стенкой область может содержать или состоять из одного, двух или трех связывающихся с клеточной стенкой доменов. Например, один, два, три или более связывающихся с клеточной стенкой доменов могут быть независимо выбраны из группы, состоящей из полипептидов, содержащих или состоящих из аминокислотной последовательности из SEQ ID NO: 15-24 и 26-33, соответственно, и любых их вариантов, идентичных по меньшей мере на 80% (предпочтительно идентичных по меньшей мере на 85%, более предпочтительно идентичных по меньшей мере на 90%, еще более предпочтительно идентичных по меньшей мере на 95%, еще более предпочтительно идентичных по меньшей мере на 96%, еще более предпочтительно идентичных по меньшей мере на 97%, еще более предпочтительно идентичных по меньшей мере на 98%, еще более предпочтительно идентичных по меньшей мере на 99%, еще более предпочтительно идентичных по меньшей мере на 99,5% и наиболее предпочтительно идентичных по меньшей мере на 99,7%) аминокислотой последовательности из SEQ ID NO: 15-24 и 26-33, соответственно, где указанные полипептиды являются функциональными, где функция включает способность связываться с клеточной стенкой Gardnerella. Предпочтительно, один, два, три или более связывающихся с клеточной стенкой доменов независимо выбраны из группы, состоящей из полипептида, содержащего аминокислотную последовательность любой из SEQ ID NO: 19, 20 и 28-33, или любого его функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности любой из SEQ ID NO: 19, 20 и 28-33, соответственно, где указанные полипептиды являются функциональными, где функция включает способность связываться с клеточной стенкой Gardnerella. Более предпочтительно, один, два, три или более связывающихся с клеточной стенкой доменов независимо выбраны из группы, состоящей из полипептида, содержащего аминокислотную последовательность любой из SEQ ID NO: 28-33, или любого его функционального варианта, идентичного по меньшей мере на 80% (предпочтительно идентичного по меньшей мере на 85%, более предпочтительно идентичного по меньшей мере на 90%, еще более предпочтительно идентичного по меньшей мере на 95%, еще более предпочтительно идентичного по меньшей мере на 96%, еще более предпочтительно идентичного по меньшей мере на 97%, еще более предпочтительно идентичного по меньшей мере на 98%, еще более предпочтительно идентичного по меньшей мере на 99%, еще более предпочтительно идентичного по меньшей мере на 99,5% и наиболее предпочтительно идентичного по меньшей мере на 99,7%) аминокислотой последовательности любой из SEQ ID NO: 28-33, где указанные полипептиды являются функциональными, где функция включает способность связываться с клеточной стенкой Gardnerella.
Более предпочтительно, С-концевая связывающаяся с клеточной стенкой область содержит первый связывающийся с клеточной стенкой домен и второй связывающийся с клеточной стенкой домен, где указанный первый связывающийся с клеточной стенкой домен выбран из группы, состоящей из SEQ ID NO: 15, 17, 19, 21, 23, 26, 28, 30 и 32, а второй связывающийся с клеточной стенкой домен выбран из группы, состоящей из SEQ ID NO: 16, 18, 20, 22, 24, 27, 29, 31 и 33. В предпочтительных воплощениях указанный первый связывающийся с клеточной стенкой домен расположен в N-концевом направлении от указанного второго связывающегося с клеточной стенкой домена.
[0086] В одном аспекте изобретения линкерная область состоит из полипептида, имеющего длину от 6 до 18 аминокислот, предпочтительно длину от 9 до 15 аминокислот, еще более предпочтительно длину 12 аминокислот. Предпочтительно, линкерная область состоит из полипептида, содержащего или состоящего из аминокислотной последовательности (1) (ХХХ)n, где каждый X может независимо представлять собой G, А или S, предпочтительно где аминокислотная последовательность (ХХХ)n представляет собой (GGS)n, где n соответствует числу повторов последовательности XXX, предпочтительно где n представляет собой 2, 3, 4, 5 или 6, или (2) X1X2GLNGX3X4NGGS, где X1 представляет собой N или K, Х2 представляет собой А или V, Х3 представляет собой Y или С и Х4 представляет собой K или Q. Фрагмент, содержащий линкер, может отсутствовать. Фрагмент, содержащий линкер, может также присутствовать и может усиливать связывание с клеточной стенкой и/или литическую активность полипептида по изобретению.
[0087] В изобретении дополнительно предложен эндолизин, обладающий уничтожающей активностью в отношении Gardnerella, как описано выше, для применения в лечении заболевания или расстройства. Заболевание или расстройство, подлежащее лечению, может представлять собой бактериальную инфекцию, предпочтительно бактериальный вагиноз. Бактериальный вагиноз может быть вызван G. vaginalis sensu stricto, G. leopoldii, G. piotii и/или G. swidsinskii или другими видами рода Gardnerella.
[0088] Эндолизин по изобретению, предпочтительно способен специфически связываться с клетками Gardnerella и/или лизировать их для применения в способе лечения инфекции, вызванной Gardnerella, такой как BV.
[0089] Как отмечено выше, хорошо известно, что многие эндолизины бактериофагов состоят из двух отдельных доменов (например, см. Sheehan et al., 1996, FEMS Microbiology Letters 140: 23-28). Один из них - это каталитический домен, который отвечает за разрушение клеточной стенки, и известно, что они существуют в нескольких формах. Другой домен представляет собой связывающийся с клеточной стенкой домен, который распознает мотив клеточной поверхности и позволяет эндолизинам прикрепляться к этой клетке-мишени. Точное распознавание паттернов, задействованное в последнем, обеспечивает специфичность. Ферментативный домен можно идентифицировать по его аминокислотной гомологии с другими подобными участками литических ферментов, которые обладают таким же типом литической активности. Что касается естественных эндолизинов профага Gardnerella, недавно открытых авторами изобретения, была установлена следующая доменная организация: N-концевой домен из 196 остатков, за которым следует линкерная область из 12 остатков и два повторяющихся домена из 49 остатков, соответственно, за исключением EL6 и EL9, где имеется только один неполный домен из 43 остатков. Нативные аминокислотные последовательности этих недавно открытых эндолизинов обобщены в Таблице 7. Авторы изобретения установили, что N-концевой домен является каталитическим доменом вследствие его гомологии с гликозидгидролазами, семейством 25, и что два повторяющихся домена являются двумя доменами, связывающимися с клеточной стенкой, вследствие их гомологии с С-концевым доменом лизоцима Cpl-7 (см. Пример 2 и Фиг. 3).
[0090] В некоторых воплощениях фрагмент, содержащий ферментативный домен, не модифицирован, то есть соответствует нативной аминокислотной последовательности. В альтернативном воплощении фрагмент, содержащий ферментативный домен, может содержать изменения, такие как замена, делеция, вставка аминокислот или любая комбинация перечисленного. В некоторых воплощениях фрагмент, содержащий ферментативный домен, представляет собой вариант фрагмента, идентичный по меньшей мере на 80% (предпочтительно идентичный по меньшей мере на 85%, более предпочтительно идентичный по меньшей мере на 90%, еще более предпочтительно идентичный по меньшей мере на 95%, еще более предпочтительно идентичный по меньшей мере на 96%, еще более предпочтительно идентичный по меньшей мере на 97%, еще более предпочтительно идентичный по меньшей мере на 98%, еще более предпочтительно идентичный по меньшей мере на 99%, еще более предпочтительно идентичный по меньшей мере на 99,5%, еще более более предпочтительно идентичный по меньшей мере на 99,7% и наиболее предпочтительно идентичный на 100% аминокислотой последовательности любой из SEQ ID NO: 1-5, 7 или 10-12.
[0091] В некоторых воплощениях фрагмент, содержащий связывающийся с клеточной стенкой домен, не модифицирован, то есть соответствует нативной аминокислотной последовательности. В альтернативном воплощении фрагмент, содержащий ферментативный домен, может содержать изменения, такие как замена, делеция, вставка аминокислот или любая комбинация перечисленного. В некоторых воплощениях фрагмент, содержащий связывающийся с клеточной стенкой домен, представляет собой вариант фрагмента, идентичный по меньшей мере на 80%, предпочтительно идентичный по меньшей мере на 85%, более предпочтительно идентичный по меньшей мере на 90%, еще более предпочтительно идентичный по меньшей мере на 95%, еще более предпочтительно идентичный по меньшей мере на 96%, еще более предпочтительно идентичный по меньшей мере на 97%, еще более предпочтительно идентичный по меньшей мере на 98%, еще более предпочтительно идентичный по меньшей мере на 99%, еще более предпочтительно идентичный по меньшей мере на 99,5%, еще более более предпочтительно идентичный по меньшей мере на 99,7% и наиболее предпочтительно идентичный на 100% аминокислотой последовательности любой из SEQ ID NO: 15-24 и 26-33.
[0092] В еще одном аспекте изобретения эндолизин содержит слияние полипептида или его фрагмента, варианта или производного или состоит из него. Под «слиянием» полипептида понимают полипептид, который слит с любым другим полипептидом. Например, полипептид может содержать одну или более дополнительных аминокислот, вставленных внутри и/или на N- и/или С-конце аминокислотной последовательности эндолизина по изобретению или его фрагмента, варианта или производного.
Таким образом, как описано выше, в одном воплощении эндолизин по первому аспекту изобретения содержит фрагмент, состоящий из одного или более связывающихся с клеточной стенкой доменов, содержащих или состоящих из аминокислотной последовательности любой из SEQ ID NO: 15-24 и 26-33 (или варианта такой последовательности домена, который сохраняет свою активность связывания с клеточной стенкой), соответственно, с которыми слит ферментативный домен из другого источника. Примеры других подходящих ферментных доменов включают L-аланоил-D-глутамат-эндопептидазу, D-глутамил-m-DAP эндопептидазу, специфичную в отношении межпептидного мостика эндопептидазу, V-ацетил-β-D-глюкозаминидазу (мурамоилгидролазу), N-ацетил-β-D-мурамидазу (лизоцим) или литическую трансгликозилазу, но не ограничены ими. Также можно использовать N-ацетилмурамоил-L-аланинамидазу из других источников.
[0093] В одном аспекте изобретения эндолизин может быть слит с полипептидом или белком для облегчения очистки указанного эндолизина. Примеры таких слияний хорошо известны специалистам в области техники. Аналогично, эндолизин может быть слит с олигогистидиновой меткой, такой как His6, или с эпитопом, распознаваемым антителом, таким как хорошо известный эпитоп, представляющй собой метку Мус. Слияния с любым вариантом фрагмента или производным эндолизина по настоящему изобретению также включены в объем изобретения. Следует понимать, что предпочтительны слияния (или их варианты или производные), которые сохраняют желаемые свойства, а именно активность эндолизина. Также особенно предпочтительно, чтобы химеры подходили для применения в способах, описанных в данном документе. Например, слияние может включать дополнительную часть, которая придает желаемые свойства эндолизину по изобретению; например, эта часть может быть полезна для выявления или выделения эндолизина, стимуляции поступления эндолизина в клетку или управления секрецией белка из клетки. Эта часть может представлять собой, например, биотиновую группировку, радиоактивную групировку, флуоресцентную группировку, например небольшой флуорофор или флуорофор зеленый флуоресцентный белок (GFP), что хорошо известно специалистам в данной области. Группировка может представлять собой иммуногенную метку, например метку Мус, как известно специалистам в данной области, или может представлять собой липофильную молекулу или полипептидный домен, способный стимулировать поступление эндолизина в клетку, как известно специалистам в данной области.
[0094] Существенной характеристикой эндолизинов по изобретению является способность лизировать клетки рода Gardnerella. Предпочтительно эндолизин способен лизировать клетки нескольких штаммов Gardnerella. Наиболее предпочтительно эндолизин способен лизировать все штаммы рода Gardnerella, включая G. vaginalis sensu stricto, G. leopoldii, G. piotii и G. swidsinskii (Vaneechoutte et al., 2019 Int. J. Syst. Evol. Biol. 898661). В одном воплощении эндолизины по изобретению по существу или полностью не способны лизировать бактерии, которые являются комменсальными представителями микробиоты здорового влагалища (и, как известно, не оказывают неблагоприятного воздействия на хозяина). Например, предпочтительно, чтобы эндолизины не лизировали клетки рода Lactobacilli. Наиболее предпочтительно эндолизины по изобретению по существу или полностью не способны лизировать клетки L. crispatus, L. gasseri и L. jensenii. Возможно эндолизины по изобретению не лизируют клетки L. iners. Предпочтительно эндолизин способен избирательно лизировать клетки патогенных бактерий, то есть в большей степени, чем клетки непатогенных бактерий.
[0095] Уничтожающая активность эндолизина по изобретению в отношении конкретного микроорганизма может быть определена стандартными в данной области процедурами, включая такие, которые основаны на определении минимальных ингибирующих концентраций (MIC) противомикробного агента, определяемых как самая низкая концентрация указанного противомикробного агента, которая подавляет видимый рост микроорганизмов после инкубации в течение ночи, как описано Andrews, 2001, J Antimicrobial Chemotherapy, 48, Suppl. SI, 5-16 или в «Document M7-A7, Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically; Approved standards, 7th Edition, January 2006, vol. 26, No. 2», опубликованном Институтом клинических и лабораторных стандартов. Другой подходящий способ определения уничтожающей активности эндолизина по изобретению описан в разделе Примеров настоящей заявки и состоит в определении уменьшения оптической плотности суспензии бактерий, измеряемой при 610-620 нм, чувствительность которых следует определять в турбидиметрическом анализе in vitro, проводимом в присутствии очищенного эндолизина по изобретению. Согласно другому воплощению, в турбидиметрическом анализе in vitro, согласно приведенному описанию, эндолизин по изобретению снижает OD(610-620 нм) суспензии по меньшей мере одного штамма бактерий Gardnerella более чем на 20%, более чем на 30%, более чем на 40%, более чем на 50%, более чем на 60%, более чем на 70%, более чем на 80%, более чем на 90% или более чем на 95%.
[0096] Способы получения эндолизинов или их фрагмента, варианта, слияния или производного для применения в соответствии с изобретением хорошо известны в данной области. Удобно, чтобы эндолизин или его фрагмент, вариант, слияние или производное представляли собой рекомбинантный эндолизин или включали его. Эндолизин по изобретению может быть получен стандартными методами генной инженерии, включающими использование рекомбинантного вектора, содержащего полинуклеотид, кодирующий эндолизин, описанный в данном документе. Могут быть использованы многочисленные системы экспрессии, включая бактериальные плазмиды и производные векторы, транспозоны, дрожжевые эписомы, встраиваемые элементы, элементы дрожжевой хромосомы, вирусы, такие как бакуловирус, вирусы папилломы, такие как SV40, вирусы осповакцины, аденовирусы, вирусы оспы лисицы, вирусы псевдобешенства, ретровирусы, производные космид или фагмид. Нуклеотидная последовательность может быть вставлена в рекомбинантный вектор экспрессии способами, хорошо известными специалистам в данной области, например такими, которые описаны в документе MOLECULAR CLONING: A LABORATORY MANUAL, Sambrook et al., 4th Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 2001. Рекомбинантный вектор может включать нуклеотидные последовательности, которые контролируют регуляцию, экспрессию, транскрипцию и/или трансляцию полинуклеотида, кодирующего эндолизин, эти последовательности выбирают в соответствии с используемыми клетками-хозяевами. Рекомбинантный вектор может дополнительно включать нуклеотидные последовательности, такие как последовательности, кодирующие His-метки, для облегчения стадии очистки. Впоследствии такой рекомбинантный вектор вводят в клетку-хозяина в соответствии со способами, которые хорошо известны специалисту в данной области, например описанными в BASIC METHODS IN MOLECULAR BIOLOGY, Davis et al., 2nd ed., McGraw-Hill Professional Publishing, 1995, and MOLECULAR CLONING: A LABORATORY MANUAL, см. выше, такими как трансфекция с применением фосфата кальция, трансфекция с применением DEAE-декстрана, трансфекция, микроинъекция, трансфекция с применением катионных липидов, электропорация, трансдукция или инфицирование. Клеткой-хозяином могут быть, например, бактериальные клетки, такие как Е.coli, клетки грибов, такие как клетки дрожжей, и клетки Aspergillus, Streptomyces, клетки насекомых, клетки яичника китайского хомячка (СНО), линия клеток мыши С127, линия клеток ВНК сирийского хомячка, эмбриональные клетки почки человека 293 (НЕK 293). Предпочтительно клетка-хозяин представляет собой Е. coli. Указанные клетки-хозяева затем культивируют в соответствующих условиях для получения описанного здесь эндолизина, который затем может быть дополнительно очищен из культуральной среды или из лизата клеток-хозяев любыми стандартными способами очистки, включая аффинную хроматографию с иммобилизованными ионами металлов (IMAC) (Block et al. 2008, Protein Expr. Purif. 27, 244-254).
Композиции по изобретению
[0097] В еще одном аспекте изобретения предложены антибактериальные композиции, содержащие эндолизин согласно первому аспекту изобретения, нуклеиновую кислоту согласно второму аспекту изобретения, вектор/плазмиду согласно третьему аспекту изобретения, клетку-хозяина согласно четвертому аспекту изобретения или бактериофаг, способный экспрессировать эндолизин согласно первому аспекту изобретения, в частности фармацевтические композиции.
[0098] Используемый в данном документе термин «фармацевтическая композиция» означает терапевтически эффективную композицию для применения в способах по изобретению. «Терапевтически эффективное количество», или «эффективное количество», или «терапевтически эффективное» в контексте настоящего описания относится к такому количеству, которое обеспечивает терапевтический эффект для данного состояния и схемы введения. Это заранее определенное количество активного материала, рассчитанное для достижения желаемого терапевтического эффекта, в сочетании с требуемым вспомогательным веществом и разбавителем, то есть носителем или средой для введения. Кроме того, подразумечается, что это означает количество, достаточное для уменьшения и, наиболее предпочтительно, предотвращения клинически значимого дефицита активности, функции и ответа хозяина. В качестве альтернативы, терапевтически эффективного количества достаточно, чтобы вызвать улучшение клинически значимого состояния у хозяина. Специалистам в данной области понятно, что количество соединения может варьировать в зависимости от его специфической активности. Подходящие величины дозы могут содержать заранее определенное количество активной композиции, рассчитанное для получения желаемого терапевтического эффекта, в сочетании с требуемым разбавителем. В способах и применении для изготовления композиций по изобретению берут терапевтически эффективное количество активного компонента. Терапевтически эффективное количество может быть определено обычным квалифицированным медицинским или ветеринарным специалистом на основе характеристик пациента, таких как возраст, вес, пол, состояние, осложнения, другие заболевания и тому подобнок, как хорошо известно в данной области. В одном воплощении изобретения фармацевтическая композиция содержит эндолизин по первому аспекту изобретения. Таким образом, фармацевтическая композиция может содержать количество эндолизина или его фрагмента, варианта, слияния или производного, достаточное для подавления по меньшей мере частично роста клеток рода Gardnerella у пациента, который инфицирован или восприимчив к инфекции такими клетки. Предпочтительно фармацевтический состав содержит количество эндолизина или его фрагмента, варианта, слияния или производного, достаточное для уничтожения клеток рода Gardnerella у пациента. Специалистам в данной области будет понятно, что эндолизины по изобретению обычно вводят в смеси с подходящим фармацевтическим эксципиентом, разбавителем или носителем, выбранным с учетом предполагаемого пути введения и стандартной фармацевтической практики (см., например, Remington: The Science and Practice of Pharmacy, 19th edition, 1995, Ed. Alfonso Gennaro, Mack Publishing Company, Pennsylvania, USA). Например, эндолизины можно вводить местно, то есть местно во влагалище субъекта женского пола и/или субъекту мужского пола, в или на головку полового члена, крайнюю плоть или вход в уретру. В данном документе термин «(введение) в или на головку полового члена» также включает «(введение) в и на головку полового члена». Соответственно, термин «(введение) в или на головку полового члена, крайнюю плоть или вход в уретру субъекта мужского пола» также включает «(введение) в и на головку полового члена и на крайнюю плоть, и на вход в уретру субъекта мужского пола». В другом воплощении эндолизины можно вводить совместно с соединением или композицией, которые доводят рН влагалища. В некоторых воплощениях соединение или композиция доводят рН влагалища до 4,0 -6,0, предпочтительно до рН 5,0.
В альтернативном воплощении изобретения фармацевтические композиции не содержат сам эндолизин, а вместо этого содержат молекулу нуклеиновой кислоты, способную экспрессировать указанный эндолизин. Подходящие молекулы нуклеиновой кислоты, векторы экспрессии и клетки-хозяева подробно описаны выше. Например, можно использовать рекомбинантный пробиотик (штамм LAB, например, Lactococcus lactis или Lactobacillus sp.). В другом воплощении изобретения фармацевтические композиции содержат бактериофаг, способный экспрессировать эндолизин согласно первому аспекту изобретения. Способы проведения такой терапии на основе бактериофагов хорошо известны в данной области (например, см. Watanabe et at., 2007, Antimicrobial Agents & Chemotherapy 51:446-452). Таким образом, для лечения описанных здесь бактериальных инфекций эндолизин по настоящему изобретению можно вводить в виде соответствующего белка, конструкции нуклеиновой кислоты, вектора или клетки-хозяина, экспрессирующих соответствующий белок, в виде части живого организма, экспрессирующей соответствующий белок (включая бактериофаги), или любым другим удобным способом, известным в данной области, чтобы достичь контакта эндолизина с его бактериальной мишенью, будь то патогенная бактерия, такая как G. vaginalis, или другой патоген или потенциальный патоген, как дополнительно описано в данном документе.
[0099] Композиции по изобретению могут содержать один или более полипептидов эндолизина. В этом воплощении полипептиды эндолизина могут присутствовать либо в виде отдельных полипептидов, либо в виде слитых белков, содержащих указанные полипептиды эндолизина или их фрагменты.
[0100] Фармацевтические композиции по настоящему изобретению могут дополнительно содержать один или более дополнительных фармацевтически приемлемых ингредиентов, таких как квасцы, стабилизаторы, антимикробные агенты, буферы, красители, корригенты, адъюванты и тому подобное. Предпочтительно, чтобы фармацевтическая композиция по изобретению не содержала имидазола.
[0101] Эндолизины по изобретению вместе с обычно используемым адъювантом, носителем, разбавителем или эксципиентом могут быть помещены в фармацевтические композиции и их стандартные лекарственные формы, и в таких формах могут использоваться в виде твердых веществ, таких как таблетки или наполненные капсулы, или жидкостей, таких как растворы, суспензии, аэрозоли, эмульсии, эликсиры или капсулы, наполненные ими, все для перорального применения или в форме стерильных растворов для инъекций для парентерального (включая подкожное) применения. Такие фармацевтические композиции и их стандартные лекарственные формы могут содержать ингредиенты в обычных пропорциях, с дополнительными активными соединениями или действующими началами или без них, и такие стандартные лекарственные формы могут содержать любое подходящее эффективное количество активного ингредиента, соизмеримое с предполагаемым диапазоном суточных доз, который следует использовать. Композиции по настоящему изобретению также могут представлять собой жидкие композиции, включая водные или масляные суспензии, растворы, эмульсии, сиропы и эликсиры, но без ограничения ими. Композиции также могут быть приготовлены в виде сухого продукта для востановления водой или другим подходящим носителем перед применением. Такие жидкие препараты могут содержать добавки, включая, без ограничения, суспендирующие агенты, эмульгирующие агенты, неводные носители и консерванты. Суспендирующие агенты включают сорбитовый сироп, метилцеллюлозу, глюкозный/сахарный сироп, желатин, гидроксиэтилцеллюлозу, карбоксиметилцеллюлозу, гель стеарата алюминия и гидрогенизированные пищевые жиры, но не ограничены ими. Эмульгирующие агенты включают лецитин, сорбитана моноолеат и гуммиарабик, но не ограничены ими. Неводные носители включают пищевые масла, миндальное масло, фракционированное кокосовое масло, масляные сложные эфиры, пропиленгликоль и этиловый спирт, но не ограничены ими. Консерванты включают метил- или пропил-п-гидроксибензоат и сорбиновую кислоту, но не ограничены ими. Дополнительные материалы, а также методы обработки и тому подобное изложены в Части 5 Remington's "The Science and Practice of Pharmacy", 22nd Edition, 2012, University of the Sciences in Philadelphia, Lippincott Williams & Wilkins.
[0102] Твердые композиции по настоящему изобретению могут быть в форме таблеток или пастилок, приготовленных стандартным способом. Таблетки могут быть покрыты оболочкой способами, хорошо известными в данной области. Композиции для инъекций обычно имеют в своей основе стерильный физиологический раствор для инъекций или забуференный фосфатом физиологический раствор или другие носители для инъекций, известные в данной области.
[0103] Композиции по настоящему изобретению также могут быть приготовлены в форме суппозиториев, которые могут содержать основы суппозиториев, включая масло какао или глицериды, но без ограничения ими. Композиции по настоящему изобретению также могут быть приготовлены в форме составов для чрескожного введения, содержащих водные или неводные носители, включая кремы, мази, лосьоны, пасты, лечебный пластырь, пластырь или мембрану, но без ограничения ими.
[0104] Композиции по настоящему изобретению также могут быть приготовлены для парентерального введения, включая инъекции или непрерывную инфузию, но без ограничения ими. Инъекционные составы могут быть в форме суспензий, растворов или эмульсий в масляных или водных носителях и могут содержать вспомогательные агенты, включая суспендирующие, стабилизирующие и диспергирующие агенты, но без ограничения ими. Композиции также могут быть представлены в форме порошка для восстановления подходящим носителем, включая, без ограничения, стерильную апирогенную воду.
[0105] Композиции по настоящему изобретению также могут быть приготовлены в виде депо-препаратов, которые можно вводить путем имплантации или внутримышечной инъекции. Композиции могут быть приготовлены с подходящими полимерными или гидрофобными материалами (например, в виде эмульсии в приемлемом масле), ионообменными смолами или в виде труднорастворимых производных (например, в виде труднорастворимой соли).
[0106] Соединения по настоящему изобретению также можно вводить в формах с замедленным высвобождением или при помощи систем доставки с замедленным высвобождением лекарственных средств. Описание типичных материалов с замедленным высвобождением также можно найти в Remington «The Science and Practice of Pharmacy».
Способ введения
[0107] Композиции по настоящему изобретению предпочтительно вводят местно во влагалище субъекта женского пола и/или в или на головку полового члена, крайнюю плоть или вход в уретру субъекта мужского пола. Однако эти композиции также можно вводить любым способом, включая внутривенную инъекцию, внутриартериальную, внутрибрюшинную инъекцию, подкожную инъекцию, внутримышечный, интратекальный, пероральный путь, включая сублингвальное или трансбуккальное введение, местное, кожное нанесение, прямую перфузию тканей во время операции или их комбинации.
[0108] В предпочтительном воплощении эндолизины, полинуклеотиды или фармацевтические композиции по настоящему изобретению, как описано в данном документе, подлежат местному введению. В еще одном предпочтительном воплощении эндолизины, полинуклеотиды или фармацевтические композиции по настоящему изобретению, как описано в данном документе, подлежат введению во влагалище субъекта женского пола и/или в или на головку полового члена, крайнюю плоть или вход в уретру субъекта мужского пола. В еще одном предпочтительном воплощении эндолизины, полинуклеотиды или фармацевтические композиции по настоящему изобретению, как описано в данном документе, вводят местно во влагалище субъекта.
[0109] Дозировка, вводимая индивидууму в виде однократной дозы или многократных доз, будет варьировать в зависимости от множества факторов, включая фармакокинетические свойства, состояние и характеристики пациента (пол, возраст, масса тела, состояние здоровья, размер), выраженность симптомов, сопутствующие лечения, периодичность лечения и желаемый эффект.
[0110] Согласно одному аспекту композиции по изобретению можно вводить пациентам профилактически до полового контакта.
Комбинации
[0111] Согласно изобретению эндолизин можно вводить отдельно или в комбинации с совместно применяемым агентом, полезным для профилактики и/или лечения инфекций или расстройств, вызванных Gardnerella, включая вызванные Gardnerella vaginalis sensu stricto, Gardnerella leopoldii, Gardnerella piotii, Gardnerella swidsinskii и/или другими видами рода Gardnerella.
[0112] Эндолизин по изобретению можно вводить в комбинации с
(а) лечением одним или более чем одним обычным антибиотиком. Такие антибиотики могут включать клиндамицин, метронидазол или любые другие подходящие антибиотики, известные специалисту в данной области;
(б) одним или более чем одним дополнительным эндолизином или молекулой нуклеиновой кислоты, вектором, клеткой-хозяином или бактериофагом, способным их экспрессировать;
(в) соединением или композицией, которые доводят рН влагалища. В некоторых воплощениях соединение или композиция доводят рН влагалища до 4,0 - 6,0, предпочтительно до рН 5,0. Подходящие соединения, которые доводят рН, включают фосфат, молочную кислоту (например, естественное подкисляющее вещество, секретируемое Lactobacilli для создания кислой среды) или другие органические кислоты, например карбоксизамещенные полимеры;
(г) терапией для нейтрализации токсинов, высвобождаемых при лизисе бактериальных клеток Gardnerella во влагалище. Подходящие нейтрализующие терапии могут включать антитела (см. Babcock et al, 2006, Infect. Immun. 74:6339-6347) и вещества, абсорбирующие токсины, такие как толевамер (см. Barker et al., 2006, Aliment. Pharmacol. Ther. 24:1525-1534);
(д) пробиотиком.
Применения и способы по изобретению
[0113] В еще одном аспекте изобретения предложены эндолизин по изобретению, нуклеиновая кислота по изобретению, вектор по изобретению, клетка-хозяин по изобретению, бактериофаг, способный экспрессировать эндолизин по изобретению, или фармакологическая композиция по изобретению для применения в медицине. Так, эндолизины по изобретению можно применять в способе лечения организма человека или животного хирургическим путем или в способах терапии и/или диагностирования, применяемых на организме человека или животного. В частности, в изобретении предложены эндолизин по изобретению, нуклеиновая кислота по изобретению, вектор/плазмида по изобретению, клетка-хозяин по изобретению, бактериофаг, способный экспрессировать эндолизин по изобретению, или фармакологическая композиция по изобретению для применения в лечении заболевания или расстройства.
[0114] В еще одном аспекте изобретения предложены эндолизин по изобретению, нуклеиновая кислота по изобретению, вектор/плазмида по изобретению, клетка-хозяин по изобретению, бактериофаг, способный экспрессировать эндолизин по изобретению, или фармакологическая композиция по изобретению для применения в качестве лекарственного средства.
[0115] В еще одном аспекте изобретения предложено применение эндолизина по изобретению, нуклеиновой кислоты по изобретению, вектора/плазмиды по изобретению, клетки-хозяина по изобретению, бактериофага, способного экспрессировать эндолизин по изобретению, или фармакологической композиции по изобретению в изготовлении лекарственного средства для уничтожения и/или подавления/предотвращения роста микробных клеток у пациента, где микробные клетки выбраны из группы, состоящей из клеток Gardnerella и других бактериальных клеток, чувствительных к лизису указанным эндолизином. В частности, в изобретении предложено применение эндолизина по изобретению, нуклеиновой кислоты по изобретению, вектора/плазмиды по изобретению, клетки-хозяина по изобретению, бактериофага, способного экспрессировать эндолизин по изобретению, или фармакологической композиции по изобретению в изготовлении лекарственного средства для лечения бактериальных инфекций и расстройств.
[0116] В еще одном аспекте изобретения предложено применение эндолизина по изобретению, нуклеиновой кислоты по изобретению, вектора/плазмиды по изобретению, клетки-хозяина по изобретению или фармакологической композиции по изобретению для применения в уничтожении и/или подавлении/предотвращении роста микробных клеток у пациента, где микробные клетки выбраны из группы, состоящей из клеток Gardnerella и других бактериальных клеток, чувствительных к лизису указанным эндолизином.
[0117] В еще одном аспекте изобретения предложен способ уничтожения и/или подавления/предотвращения роста микробных клеток у пациента, включающий введение пациенту эндолизина по изобретению, нуклеиновой кислоты по изобретению, вектора/плазмиды по изобретению, клетки-хозяина по изобретению, бактериофага, способного экспрессировать эндолизин по изобретению, или фармакологической композиции по изобретению, где микробные клетки выбраны из группы, состоящей из клеток Gardnerella и других бактериальных клеток, чувствительных к лизису указанным эндолизином.
[0118] В еще одном аспекте изобретения предложено применение эндолизина по изобретению, нуклеиновой кислоты по изобретению, вектора/плазмиды по изобретению, клетки-хозяина по изобретению, бактериофага, способного экспрессировать эндолизин по изобретению, или фармакологической композиции по изобретению в изготовлении лекарственного средства для лечения или профилактики заболевания или состояния пациента, ассоциированного с микробными клетками, где микробные клетки выбраны из группы, состоящей из клеток Gardnerella и других бактериальных клеток, чувствительных к лизису указанным эндолизином.
[0119] В еще одном аспекте изобретения предложены эндолизин по изобретению, нуклеиновая кислота по изобретению, вектор/плазмида по изобретению, клетка-хозяин по изобретению, бактериофаг, способный экспрессировать эндолизин по изобретению, или фармакологическая композиция по изобретению для применения в лечении или профилактике заболевания или состояния пациента, ассоциированного с микробными клетками, где микробные клетки выбраны из группы, состоящей из клеток Gardnerella и других бактериальных клеток, чувствительных к лизису эндолизинами по изобретению.
[0120] В еще одном аспекте изобретения предложен способ лечения или профилактики заболевания или состояния, ассоциированного с микробными клетками, у нуждающегося в таком лечении пациента, включающий введение пациенту эндолизина по изобретению, нуклеиновой кислоты по изобретению, вектора/плазмиды по изобретению, клетки-хозяина по изобретению, бактериофага, способного экспрессировать эндолизин по изобретению, или фармакологической композиции по изобретению, где микробные клетки выбраны из группы, состоящей из клеток Gardnerella и других бактериальных клеток, чувствительных к лизису эндолизинами по изобретению.
[0121] «Заболевание или состояние пациента, ассоциированное с микробными клетками» включает заболевания и состояния, которые возникают вследствие инфицирования пациента Gardnerella или которые вызваны борьбой с инфекцией Gardnerella. Такие заболевания и состояния включают BV.
[0122] Термин «лечение» включает как терапевтическое, так и профилактическое лечение субъекта (или пациента). В одном воплощении эндолизин по изобретению, нуклеиновая кислота по изобретению, вектор/плазмида по изобретению, клетка-хозяин по изобретению, бактериофаг, способный экспрессировать эндолизин по изобретению, или фармакологическая композиция по изобретению, применения и способы по изобретению предназначены для лечения существующего заболевания или состояния. В качестве альтернативы или дополнения, применения и способы по изобретению могут быть предназначены для профилактики. Термин «профилактический» или «профилактика» используется как включающий применение эндолизина или композиции, описанных в данном документе, которое либо предотвращает, либо снижает вероятность инфицирования пациента или субъекта Gardnerella. Профилактика может представлять собой первичную профилактику (то есть предотвращать развитие заболевания) или вторичную профилактику (если заболевание уже развилось, и пациента защищают от обострения этого процесса). Предпочтительно, чтобы средства и способы, представленные в данном документе, предназначались для лечения существующего заболевания или состояния, в частности для лечения существующего BV.
[0123] Как обсуждалось выше, термин «эффективное количество» используется в данном документе для описания концентраций или количеств эндолизинов по настоящему изобретению, которые можно использовать для достижения благоприятного изменения заболевания или состояния, лечение которого осуществляют, независимо от того, является ли это изменение ремиссией, благоприятным физиологическим результатом, обращением вспять или ослаблением болезненного состояния или состояния, лечение которого осуществляют, предотвращением или снижением вероятности возникновения состояния или болезненного состояния, в зависимости от заболевания или состояния, лечение которого осуществляют. В одном воплощении эндолизин согласно первому аспекту изобретения, нуклеиновую кислоту согласно второму аспекту изобретения, вектор согласно третьему аспекту изобретения, клетку-хозяина согласно четвертому аспекту изобретения, бактериофаг, способный экспрессировать эндолизин согласно первому аспекту изобретения, или фармакологическую композицию согласно шестому аспекту изобретения вводят в виде однократной дозы. В качестве альтернативы эндолизин, нуклеиновую кислоту, вектор/плазмиду, клетку-хозяина, бактериофаг или фармакологическую композицию вводят в виде многократных доз (например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 или более доз). Эндолизин, нуклеиновую кислоту, вектор/плазмиду, клетку-хозяина, бактериофаг или фармакологическую композицию предпочтительно вводят с частотой, достаточной для поддержания постоянного присутствия эндолизина согласно первому аспекту изобретения во влагалище субъекта. Предпочтительно доза и частота введения являются достаточными для предотвращения возникновения или рецидивирования у субъекта заболевания или состояния, ассоциированного с микробными клетками (например, Gardnerella). Предпочтительно доза и частота введения являются достаточными для предотвращения возникновения или рецидивирования у субъекта импеданса роста, ассоциированного с микробными клетками (например, Gardnerella).
[0124] В одном воплощении применения и способы по изобретению, клетку-хозяина или фармакологическую композицию, содержащую клетку-хозяина, применяют для доставки эндолизина по первому аспекту изобретения (предпочтительно, клетку-хозяина).
[0125] Следует понимать, что описанные в данном документе лекарственные средства можно вводить субъекту в комбинации с одним или несколькими дополнительными терапевтическими агентами. Например, описанные в данном документе лекарственные средства можно вводить субъекту в комбинации с:
(а) лечением одним или более чем одним обычным антибиотиком. Такие антибиотики могут включать клиндамицин, метронидазол или любые другие подходящие антибиотики, известные специалисту в данной области;
(б) одним или более чем одним дополнительным эндолизином или молекулой нуклеиновой кислоты, вектором, клеткой-хозяином или бактериофагом, способным их экспрессировать;
(в) соединением или композицией, которые доводят рН влагалища, предпочтительно до 4,0-6,0, более предпочтительно приблизительно до рН 5,0. Такие соединения, которые доводят рН, могут включать фосфат, молочную кислоту (например, естественное подкисляющее вещество, секретируемое Lactobacilli для создания кислой среды) или другие органические кислоты, например карбоксизамещенные полимеры;
(г) терапией для нейтрализации токсинов, высвобождаемых при лизисе бактериальных клеток G.vaginalis во влагалище. Подходящие нейтрализующие терапии могут включать антитела (см. Babcock et al, 2006, Infect. Immun. 74:6339-6347) и вещества, абсорбирующие токсины, такие как толевамер (см. Barker et al., 2006, Aliment. Pharmacol. Ther. 24:1525-1534);
(д) пробиотиком.
[0126] В еще одном аспекте изобретения предложено применение эндолизина, обладающего лизирующей активностью в отношении клеток Gardnerella, или молекулы нуклеиновой кислоты, вектора/плазмиды, клетки-хозяина или бактериофага, способного его экспрессировать, для уничтожения микробных клеток и/или подавления/предотвращения их роста in vitro и/или ex vivo, где микробные клетки выбраны из группы, состоящей из клеток Gardnerella и других бактериальных клеток, чувствительных к лизису эндолизинами по изобретению. Например, эндолизины, обладающие указанной активностью, можно применять для очистки поверхностей, таких как поверхности в больницах, на кухнях и тому подобное, которые могут быть подвержены загрязнению такими бактериальными клетками. Предпочтительно микробные клетки содержат или представляют собой клетки G. vaginalis sensu stricto, G. leopoldii, G. piotii, G. swidsinskii или другие виды рода Gardnerella.
[0127] В еще одном аспекте настоящего изобретения предложен набор. Указанный набор содержит эндолизин, описанный в данном документе, и инструкции по применению, в частности, для лечения заболевания или расстройства, предпочтительно BV. Указанный набор можно применять в терапевтических или профилактических целях и может также содержать соединение или композицию, которые доводят рН влагалища до 4,0-6,0, предпочтительно до 4,5-5,5, более предпочтительно до приблизительно 5. Однако набор по настоящему изобретению также можно применять для выявления присутствия микробных клеток в образце, причем набор содержит полипептид, обладающий лизирующей клетки активностью и/или специфичностью связывания с клетками эндолизина по изобретению, или молекулу нуклеиновой кислоты, вектор/плазмиду, клетку-хозяина или бактериофаг, способный его экспрессировать, причем микробные клетки выбраны из группы, состоящей из клеток Gardnerella и других бактериальных клеток, чувствительных к лизису указанным эндолизином.
[0128] В связанных аспектах изобретения предложены:
(а) применение полипептида, обладающего связывающей клеточную стенку активностью и/или лизирующей клетки активностью эндолизина по изобретению, или молекулы нуклеиновой кислоты, вектора/плазмиды, клетки-хозяина или бактериофага, способного его экспрессировать, в получении диагностического агента заболевания или состояния, ассоциированного с микробными клетками, выбранными из группы, состоящей из клеток Gardnerella и других бактериальных клеток, чувствительных к лизису указанным эндолизином;
(б) применение полипептида, обладающего связывающей клеточную стенку активностью и/или лизирующей клетки активностью эндолизина по изобретению, или молекулы нуклеиновой кислоты, вектора/плазмиды, клетки-хозяина или бактериофага, способного его экспрессировать, в диагностике заболевания или состояния, ассоциированного с микробными клетками, выбранными из группы, состоящей из клеток Gardnerella и других бактериальных клеток, чувствительных к лизису указанным эндолизином;
(в) применение полипептида, обладающего связывающей клеточную стенку активностью и/или лизирующей клетки активностью эндолизина по изобретению, или молекулы нуклеиновой кислоты, вектора/плазмиды, клетки-хозяина или бактериофага, способного его экспрессировать, для выявления присутствия в образце микробных клеток in vitro и/или ex vivo, где микробные клетки выбраны из группы, состоящей из клеток Gardnerella и других бактериальных клеток, чувствительных к лизису указанным эндолизином; и
(г) способ in vitro диагностирования заболевания или состояния, которое можно лечить эндолизином согласно настоящему изобретению, включающий стадии: (1) приведения полученного от субъекта образца в контакт с полипептидом, содержащим или состоящим из С-концевой связывающейся с клеточной стенкой области эндолизина по настоящему изобретению и возможно N-концевого каталитического домена эндолизина по настоящему изобретению, где образец содержит микробные клетки и где С-концевая связывающаяся с клеточной стенкой область указанного эндолизина возможно является меченой; (2) тестирования, связывается ли полипептид с микробными клетками образца и/или лизирует ли их; и (3) установления того, что заболевание или состояние можно лечить эндолизином по настоящему изобретению, если полипептид связывается с микробными клетками и/или лизирует их. Микробные клетки могут представлять собой клетки Gardnerella, предпочтительно клетками G. vaginalis sensu stricto, G. leopoldii, G. piotii, G. swidsinskii или другими видами рода Gardnerella.
Таким образом, в изобретении предложен способ in vitro диагностирования заболевания или состояния, которое можно лечить эндолизином по изобретению, где способ включает приведение полученного от субъекта образца клеток в контакт с полипептидом, обладающим связывающей клеточную стенку активностью и/или лизирующей клетки активностью эндолизина по изобретению, или молекулой нуклеиновой кислоты, вектором/плазмидой, клеткой-хозяином или профагом, способным его экспрессировать, и установление того, были ли таким образом лизированы клетки в образце, где микробные клетки выбраны из группы, состоящей из клеток Gardnerella и других бактериальных клеток, чувствительных к лизису указанным эндолизином. Предпочтительно микробные клетки содержат или представляют собой клетки G. vaginalis sensu stricto, G. leopoldii, G. piotii, G. swidsinskii или другие виды рода Gardnerella. В таких диагностических применениях и способах лизис клеток может быть выявлен с использованием методов, хорошо известных в данной области. Например, можно измерять уровни АТР (аденозинтрифосфат) как индикатор лизиса клеток.
[0129] В альтернативном воплощении определенных выше применений и способов по изобретению полипептид содержит или состоит из связывающегося с клеточной стенкой домена эндолизина по изобретению. Чтобы сделать возможным обнаружение, такой полипептид может быть слит с магнитными бусинами или использован в виде слитого белка, содержащего подходящий репортер или метку (например, зеленый флуоресцентный белок или фермент, дающий окраску, такой как HRP (пероксидаза хрена)). Такие диагностические подходы хорошо разработаны для эндолизинов из других систем, таких как эндолизины Listeria (например, см. Loessner et al., 2002, Mol Microbiol 44, 335-49; Kretzer et al., 2007, Applied Environ. Microbiol. 73:1992-2000).
[0130] Воплощения изобретения, служащие в качестве иллюстрации, описаны в приведенных ниже Примерах, не являющихся ограничивающими, со ссылкой на приведенные ниже графические материалы.
ПРИМЕРЫ
[0131] Пример 1 - Обнаружение естественных эндолизинов в геномах Gardnerella
[0132] Эндолизины представляют собой гидролитические ферменты, продуцируемые бактериофагами для расщепления клеточной стенки хозяина во время заключительной стадии литического цикла. Они обладают способностью нацеливаться на одну из пяти связей в пептидогликане (муреине), основном компоненте бактериальных клеточных стенок, что позволяет высвобождать потомство вирионов из лизированной клетки. На сегодняшний день не выделено бактериофагов, обладающих литической активностью против Gardnerella. Поэтому также было неизвестно, могут ли быть успешно идентифицированы эндолизины бактериофагового происхождения, обладающие литической активностью против Gardnerella. Авторы изобретения исследовали, можно ли обнаружить в различных геномах Gardnerella эндолизины, кодируемые последовательностями профагов. Профаги представляют собой геномы бактериофагов, встроенные и интегрированные в кольцевую бактериальную ДНК-хромосому или существующие в виде внехромосомной плазмиды. Это латентная форма фага, в которой вирусные гены присутствуют в бактерии, не вызывая разрушения бактериальной клетки. Последовательности профагов в бактериальных геномах и плазмидах могут быть идентифицированы с помощью веб-приложений, известных специалистам в данной области. Например, такие приложения включают, без ограничения, PHASTER (Arndt et al., 2016 Nucleic Acids Res. 44, W16-W21.), PROPHINDER (Lima-Mendez et al., 2008 Bioinformatics 24, 863 865) или подобные. Авторам настоящего изобретения удалось идентифицировать последовательности, которые, согласно прогнозам, представляют собой интактные или частичные профаги, в 14 геномах Gardnerella. Последовательности были найдены в результате идентификации участков ДНК, в которых сгруппированы гены, которые, согласно прогнозам, имеют вирусное происхождение. Также были включены кластеры вирусных генов, которые, согласно прогнозам, являются частичными профагами, в отличие от полных профагов.
[0133] Затем, чтобы идентифицировать предполагаемые эндолизины, предполагаемые последовательности профагов аннотировали путем анализа в BLAST предсказанных кодирующих последовательностей. В частности, проводили поиск белковых последовательностей, гомологичных ферментам, способным расщеплять любую из ключевых химических связей, составляющих пептидогликан. В частности, проводили поиск белковых последовательностей, гомологичных N-актилмурамидазам, N-актилмурамоил-L-аланинамидазам, L-аланоил-D-глутамат эндопептидазам, эндопептидазам межпептидных мостиков или N-ацетил-бета-D-глюкозаминидазам. В каждом проанализированном отдельном профаге или частичном профаге авторы изобретения обнаружили кодирующие последовательности белков, гомологичных 1,4-бета-N-ацетилмурамидазам, и обозначили их от EL1 до EL14. Присвоенные названия и геномы, служившие источниками, показаны в Таблице 1.
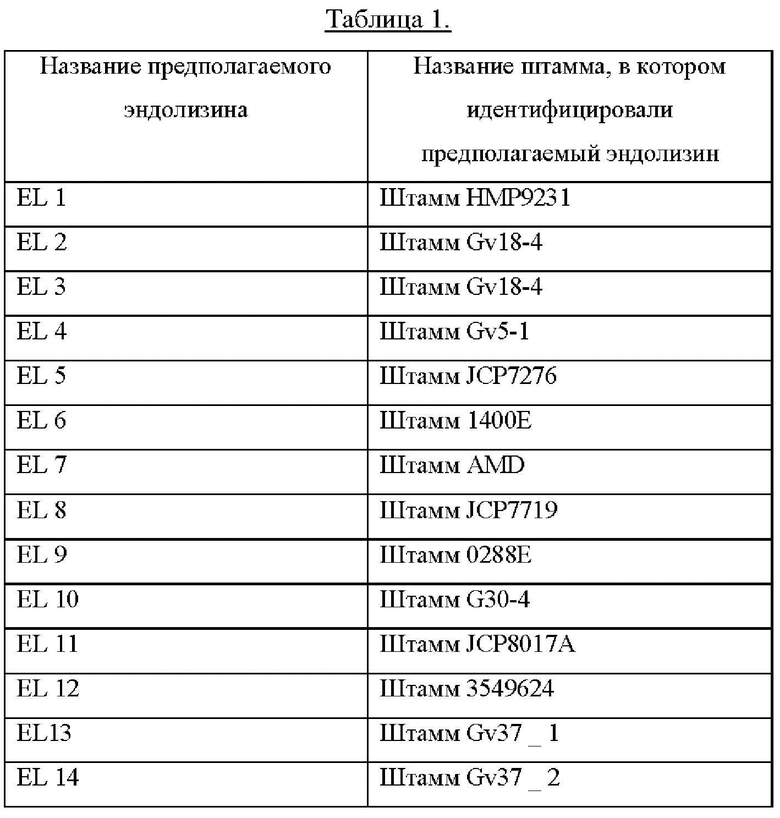
В каждом случае была обнаружена только одна копия на профаг и не обнаружены никакие другие кодирующие последовательности, которые, согласно прогнозам, являются ферментами, способными лизировать клеточную стенку бактерий. Для понимания гомологии и доменной структуры осуществляли выравнивание предполагаемых 1,4-бета-N-ацетилмурамидаз (см. Фиг. 1). Как видно на Фиг. 1, большинство эндолизинов, даже из разных профагов разных геномов, имеет ровно 306 остатков. Двумя исключениями являются EL6 и EL9. EL6 усечен на С-конце сдвигом рамки. EL9 заканчивается в том же положении, что и EL6, однако в этом случае заканчивается весь контиг. Среди эндолизинов нет идентичных пар, несмотря на их высокую гомологию, как можно видеть на Фиг. 2.
[0134] Пример 2 - Определение доменной структуры естественных эндолизинов профага Gardnerella
[0135] Доменную структуру недавно открытых эндолизинов устанавливали при помощи InterPro (Mitchell et al., 2019 Nucleic Acids Res. 47, D351-D360). Вкратце, InterPro - это база данных семейств белков, доменов и функциональных сайтов, в которой идентифицируемые признаки, обнаруженные у известных белков, можно применить к последовательностям новых белков, чтобы охарактеризовать их с функциональной точки зрения. Содержимое InterPro состоит из диагностических сигнатур и белков, которым они по существу соответствуют. Сигнатуры представляют собой модели, например, простого типа, такие как регулярные выражения, или более сложные, такие как скрытые Марковские модели, описывающие семейства белков, домены или сайты. Как видно на Фиг. 3, все эндолизины с 306 остатками имеют одинаковое расположение доменов. N-концевой домен из 196 остатков идентифицирован как каталитический домен вследствие его гомологии с гидролазами гликозидов, семейством 25. За указанным каталитическим доменом следует линкерная область и два домена, связывающих клетки, гомологичных С-концевому домену лизоцима Cpl-7, также обозначаемые как домены CW 7 (Garcia et al, 1990 Gene 86, 81-88; Lopez and Garcia, 2004 FEMS Microbiol. Rev. 28, 553-580; Bustamante et al., 2010 J. Biol. Chem. 285, 33184-33196, 2012 PLoS One 7, e46654). В следующих примерах указанный выше каталитический домен представляет собой «N-концевой каталитический домен» или «Н-домен», где, например, «Н2» относится к Н-домену естественного EL2. В следующих примерах «линкерная область» и «С-концевая связывающаяся с клеточной стенкой область», где последняя содержит или состоит из одного или более связывающихся с клеточной стенкой доменов, или «В-доменов», вместе представляют собой «В-область», где, например, В10 относится к В-области естественного EL10. Аналогично, В11_N относится к N-концевому связывающемуся с клеточной стенкой домену естественного EL11, В12_С относится к С-концевому связывающемуся с клеточной стенкой домену естественного EL12 и так далее.
[0136] Пример 3 - Определение ферментативной активности естественных эндолизинов профага Gardnerella на клетках Gardnerella
[0137] Авторы изобретения исследовали ферментативную активность недавно открытых эндолизинов в отношении Gardnerella. Были ли активными обнаруженные последовательности, гомологичные 1,4-бета-N-ацетилмурамидазам, невозможно предсказать in silico. Как хорошо известно в данной области техники, у бактерий могут мутировать их последовательности профагов, и соответствующие профаги, следовательно, могут потерять свою способность к размножению. Более того, даже если недавно открытые кодируемые фагами гидролазы пептидогликанов действительно были активными белками, все еще не было продемонстрировано, что указанные белки способны ферментативно разрушать специфический пептид о гликановый слой Gardnerella. Действительно, особенностью Gardnerella является то, что это грамвариабельный вид: он не образует внешнюю мембрану, характерную для истинных грамотрицательных видов. Ее клеточная стенка обычно очень тонкая, а содержание в ней пептидогликанов составляет лишь 10% или менее. Таким образом, специалист в данной области мог бы подумать, что фермент, разрушающий пептид о гликаны, такой как белки эндолизины, не может эффективно лизировать бактериальные клеточные стенки Gardnerella.
[0138] 14 обнаруженных эндолизинов EL1 до EL14 клонировали с His-меткой, экспрессировали в Е. coli и очищали в одну стадию с помощью колонки Ni-NTA с применением способа, описанного в Справочном Примере 1. Обозначения, присвоенные эндолизинам, и геномы, служившие их источникам, показаны в Таблице 1. Используемые штаммы Gardnerella показаны в Таблице 2.


Чтобы проверить активность очищенных эндолизинов, измеряли изменение мутности суспензий Gardnerella (см. Таблицу 2) при 610-620 нм, используя метод, по существу описанный в Справочном Примере 2, где 95 мкл бактериальной суспензии в бульоне Харди при указанном рН смешивали с 5 мкл раствора эндолизина в фотометрической кювете в аэробных условиях при комнатной температуре. В турбидиметрических анализах уменьшение светорассеяния (то есть уменьшение мутности) суспензии живых клеток можно использовать для анализа активности гидролаз пептидогликанов в спектрофотометре. Уменьшение оптической плотности с течением времени (в минутах) можно использовать для расчета скорости гидролиза. Результаты сравнивают с контрольным препаратом «без добавления фермента, только буфер», обработанным идентично в течение того же периода времени. Таким образом, специфическая активность ферментного препарата может быть представлена как ΔOD/время/мкл белка лизина. Как можно видеть на фигурах 4А-4С, падение мутности было гораздо более выраженным для групп, обработанных эндолизином, а не буфером, что указывает на ферментативную активность. Неожиданно авторы изобретения обнаружили, что недавно открытые эндолизины EL1, EL2, EL3, EL4, EL5, EL7, EL10, EL11 и EL12 были активными белками, обладающими способностью лизировать клеточные стенки Gardnerella. Как объяснено выше, из-за низкого содержания пептид о гликана в клеточной стенке, тот факт, что фермент, разрушающий пептид о гликаны, такой как обнаруженные эндолизины, может эффективно лизировать стенки бактериальных клеток, был неожиданным и удивительным открытием.
[0139] Пример 4 - Обнаружение наиболее активных доменов у искусственных эндолизинов с замененными доменами
[0140] Затем авторы изобретения оценили, могут ли различные N-концевые обладающие ферментативной активностью домены иметь разную литическую активность и может ли В-область (включающая линкер и связывающиеся с клеточной стенкой домены) опосредовать специфичность к разным штаммам. Для этого искусственно создавали эндолизины с замененными доменами.
[0141] 4.1 - Конструкции эндолизинов
[0142] Чтобы искусственным путем создать эндолизины с замененными доменами, N-концевые 196 остатков первого естественного эндолизина, включая каталитический домен, переносили блоком на полную С-концевую область из 110 остатков второго естественного эндолизина, содержащего линкерную область и два связывающихся с клеточной стенкой домена. Белки с замененными доменами получали с применением следующих методов. Исходные конструкции EL1-14 заказывали в GeneWiz как синтетические гены с оптимизацией кодонов для Е. coli. В GeneWiz эти конструкции клонировали в вектор pETM14_ccdB при помощи рестриктазно-лигазного метода с использованием сайтов, распознаваемых ферментами NcoI и NotI
В Таблице 3 ниже приведены использованные праймеры. Для замены доменов у выбранных 10 конструкций каждый Н-домен вместе с промотором Т7 амплифицировали с использованием одинаковых прямых праймеров (№2) и специфичных для конструкции обратных праймеров (№3-12), используя полимеразу PhusionFlash (Thermo, F -548L). Аналогично, каждую В-область амплифицировали с использованием специфичных для конструкции внутренних праймеров (№13-21) и одинаковых обратных праймеров (№1), включавших терминатор Т7. Все праймеры содержали удлинения, несущие сайт распознавания BsaI, что делало внешние концы совместимыми с каркасом вектора pETMdest, производным от рЕТМ14. Между доменами предусматривали липкие концы, имеющие последовательность «GGCT», располагая их между двух аминокислот GL линкерной последовательности. Таким образом, эти 2 аминокислоты представляли точную границу между доменами для задач этого эксперимента.
Таким образом, амплифицированные и очищенные путем электрофореза в геле домены (набор для очистки GeneJet Gel, Thermo, K0692) затем объединяли в 90 новых экспрессионных конструкций с использованием стратегии клонирования GoldenGate посредством рестрикции BsaI (BsaI-HFv2, NEB, Рч.3733S)/лигазной циклической реакции (ДНК-лигаза Т4, Thermo, EL0011).
Для трансформации использовали штамм E.coli NEB10beta (NEB, С3019) и очищали плазмиды с использованием набора GeneGet Plasmid Miniprep (Thermo, K0502).

Для краткости комбинацию доменов искусственных эндолизинов обозначали с помощью Н-кода для N-концевого каталитического домена (в дальнейшем обозначаемого как Н-домен) и В-кода для части, содержащей С-концевую связывающуюся с клеточной стенкой область и линкерную область (далее обозначаемой как В-область). Например, Н2В10 относится к эндолизину с замененными доменами, имеющему N-концевой домен из естественного эндолизина EL2 и линкерную область и С-концевую связывающуюся с клеточной стенкой область из естественного эндолизина EL10. Другими словами, Н2В10 относится к эндолизину с замененными доменами, состоящему из 196 N-концевых остатков естественного эндолизина EL2 (SEQ ID NO: 2) и 110 С-концевых остатков естественного эндолизина EL10. В этом примере В-область В10, соответствующая 110 С-концевым остаткам естественного эндолизина EL10, включает в направлении от С-конца к N-концу: С-концевой связывающийся с клеточной стенкой домен «В10_С» (SEQ ID NO: 29), N-концевой связывающийся с клеточной стенкой домен «B10_N» (SEQ ID NO: 28) и линкерную область «L10» (NAGLNGYKNGGS). Номенклатура и соответствующие аминокислотные последовательности подробно представлены в Таблице 7.
Следовательно, согласно приведенной выше номенклатуре, естественный эндолизин, например EL3, может быть взаимозаменяемо описан как Н3В3 или H3-L3-(В3_N)(B3_С). Аналогичным образом, рекомбинантный эндолизин, например Н2В10, может также взаимозаменяемо быть обозначен как H2-L10- (B10_N)(B10_С). Специалисту в данной области техники понятно, что в других воплощениях элементы «Lx», «(Bx_N)» и «(Bx_C)» также могут быть независимо заменены. Номенклатура дополнительно представлена в Таблице 7 ниже.
[0143] 4.2 - Оптимизация параметров анализа
[0144] Проанализировали зависимость активности от трех потенциально важных параметров, то есть рН, анэробных/микроаэрофильных/аэробных условий, отсутствия/присутствия имидиазола. Три критерия для оценки выбирали по следующим причинам.
- рН: Уничтожающая активность эндолизинов по изобретению была успешно продемонстрирована в экспериментах, проведенных при значениях рН приблизительно 7. Однако в здоровом влагалище рН составляет приблизительно 3,5, в то время как при BV рН во влагалище достигает приблизительно 5,5 и даже выше. Таким образом, исследовали зависимость активности эндолизинов от рН.
- Кислород: В литературе Gardnerella описывается как анаэроб или микроаэрофил. Поэтому исследовали, выживало ли в отсутствие воздействия большее количество клеток в анаэробных, микроаэрофильных или аэробных условиях в течение инкубационного периода в ходе эксперимента (обычно 5 часов).
- Имидазол: В соответствии со способом, описанным в Справочном Примере 1, эндолизины по изобретению очищают в ходе одной стадии с помощью колонки с Ni-NTA, где буфер, используемый для элюирования эндолизинов из матрикса Ni-NTA, содержал имидазол. Следовательно, в отсутствие последующего этапа диализа образца полученные растворы элюата содержат 250 мМ имидазола. В этой связи исследовали влияние имидазола на Gardnerella.
Сначала оценивали чувствительность выживания G. vaginalis Gv_9 к инкубации в среде с имидазолом или без него при различных значениях рН (см. Фиг. 6). 5×107 CFU/мл клеток инкубировали в условиях, указанных под графиком, в течение 5 часов при 37°С в анаэробных условиях. Затем определяли количество выживших CFU/мл путем количественного посева. Как показано на Фиг. 6, при рН 6,0 клетки в медианном количестве 1×107 и 1×106 выживали при инкубации без имидазола и с имидазолом, соответственно. При рН 7,0 медианное количество выживших клеток составляло только 2×106 и 3×104 без имидазола и с имидазолом, соответственно. В необработанном контроле при рН 5,0 после процедуры выживало 1е7 клеток, в то время как медианная выживаемость в присутствии имидазола при рН 7, составляла 3е4, то есть на 3 log ниже. Следовательно, выживание G. vaginalis Gv_9 сильно зависит от отсутствия имидазола, особенно при рН более 6,0 и низких значениях рН.
Во-вторых, оценивали чувствительность G. vaginalis Gv_9 к воздействию рекомбинантнго эндолизина Н10В1 по сравнению с контролем, содержащим имидазол, при различных значениях рН (см. Фиг. 7). 5×107 CFU/мл клеток инкубировали в условиях, указанных под графиком, в течение 5 часов при 37°С в анаэробных условиях. Затем определяли количество выживших CFU/мл путем количественного посева. Столбцы, обозначенные контролем с имидазолом, отображают те же данные, что и на Фиг. 5. Как показано на Фиг. 7, эндолизин в высокой степени активен при рН до 5,0, и при таком низком рН относительное снижение по сравнению с контролем даже намного более выражено, происходит снижение жизнеспособных CFU на 2,5 log. В то время как при рН 7,0 разница в выживаемости между обработанными Н10В1 и необработанными клетками составляла менее 1 log10, при рН 5,0 разница составляла 2 log10. Выживаемость клеток, не обработанных Н10В1, не увеличивалась при значениях рН ниже 6,0 в присутствии имидазола. Следовательно, активность эндолизина Н10В1 в высокой степени зависит от рН. Когда аналогичные эксперименты проводили в аэробных условиях, выживаемость контрольных клеток снижалась на несколько log10 по сравнению с анаэробными условиями (данные не показаны).
Таким образом, сделали вывод, что оптимальные параметры для проведения экспериментов с эндолизинами по изобретению представляли собой анаэробные условия при рН 5,0 и со стадией удаления имидазола из элюатов эндолизина.
[0145] 4.3 - Уровни экспрессии
[0146] Концентрации всех конструкций эндолизина сведены в Таблицу 4. У каждой конструкции, концентрация которой превышала 0,2 мг/мл после удаления имидазола, доводили концентрацию до 0,2 мг/мл путем разбавления. Конструкции с более низкой концентрацией оставляли как есть и тестировали их активность. Естественный эндолизин EL6 (не показан в Таблице 4) имел концентрацию ниже 0,2 мг/мл. Н4, Н11 и H12, по-видимому, приводят к низкой растворимости и низким уровням экспрессии, поскольку у большинства конструкций не достигалось пороговое значение 0,2 мг/мл. Также низкую концентрацию имели несколько конструкций с H1.

[0147] 4.4 - Количественная оценка лизиса четырех основных видов Gardnerella под действием эндолизина в суспензии в оптимизированных условиях
[0148] Литическую активность 91 конструкции (естественные эндолизины и эндолизины с замененными доменами) в отношении четырех основных видов Gardnerella оценивали количественно с использованием метода, описанного в Справочном Примере 2. Вкратце, 90 мкл 5е7 CFU/мл указанного штамма инкубировали в течение 5 часов при рН 5,0 в анаэробных условиях с 10 мкл эндолизина (концентрацию доводили до 0,2 мг/мл, где это возможно, см. Таблицу 4).
Результаты показаны на Фиг. 8А - 8D и в Таблицах 5А - 5С. На Фиг. 8А - 8D по логарифмической оси Y отложено количество выживших клеток. Пунктирная линия указывает предел обнаружения (LOD), полученный при посеве 2 мкл реакционной смеси (500 CFU/мл). Оценивали каждую комбинацию естественных 10 Н-доменов и естественных 9 В-областей, включая естественные эндолизины (Н1В1, Н2В2, Н3В3 и так далее), а также Н6В6. Никакие другие конструкции с В-доменом В6 не тестировали, поскольку В6 делал все Н-домены, слитые с ним, неактивными, как показывало измерение OD (данные не приведены). Активность каждой конструкции измеряли на каждом из 4 основных видов Gardnerella: G. vaginalis sensu stricto, G. leopoldii, G. piotii и G. swidsinkii. В Таблице 5 обобщены результаты снижения log10 CFU, сгруппированные по Н-домену и В-области. Все измерения проводили в трипликатах. Выживаемость после 5 часов инкубации при рН 5,0 в анаэробных условиях с эндолизинами или с буфером измеряли путем количественного посева. Значения показывают log10 отношения выживших CFU для обработанных и необработанных клеток. Использовали среднее значение из трех измерений. В Таблицах 5А и 5В высокие отрицательные значения log10, например -6,7, -5,5, -4,8 и так далее, коррелируют с высокой ферментативной активностью, тогда как значения log10, близкие к нулю или даже положительные значения log10, коррелируют с низкой ферментативной активностью или ее отсутствием. В качестве примера, если бы в контроле среднее из 3 измерений значение CFU для Gv_9 составляло 1,0×107 CFU/мл, а среднее значение для 3 образцов, обработанных Н2В10, составляло 2,5×103 CFU/мл, тогда значение log10 снижения CFU под влиянием Н2В10 для Gv_9 составляло бы log10(2×103/107)=-3,7. И наоборот, величина снижения -3,7 означает 103,7-кратное (=5012-кратное) снижение жизнеспособных CFU в обработанном образце по сравнению с необработанным контролем. - В случае воздействия на Gv_9 неактивного эндолизина, например, Н4В3, CFU Gv_9 после воздействия будет равно CFU, измеренному в контрольном образце, и соотношение двух значений CFU будет равно единице. Следовательно, величина снижения Gv_9 под воздействием Н4В3 составляет log10 (1) = 0,0.
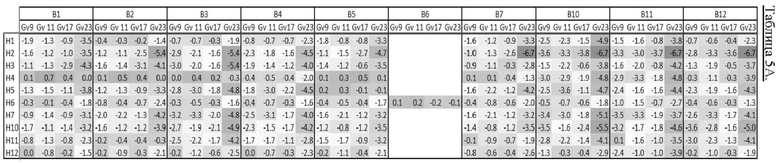
В Таблице 5В ниже представлены те же данные, что и в таблице 5А, но отображенные как средние значения log 10 активности каждой конструкции в отношении четырех штаммов Gardnerella. Справа и внизу показаны средние значения каждого естественного Н-домена в сочетании со всеми естественными В-областями (кроме В6) и каждой естественной В-области (кроме В6) в сочетании со всеми естественными Н-доменами, соответственно, наряду с рангом активности соответствующей естественной Н-области и естественной В-области.
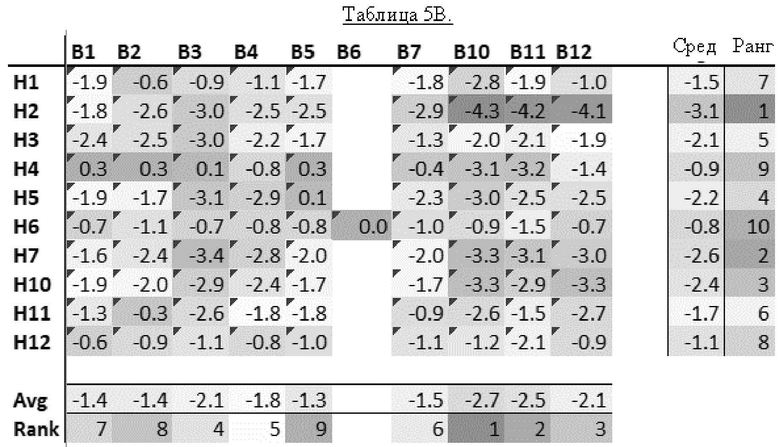
В таблице 5С приведены рагни активности всех эндолизинов на основе данных Таблицы 5 В.
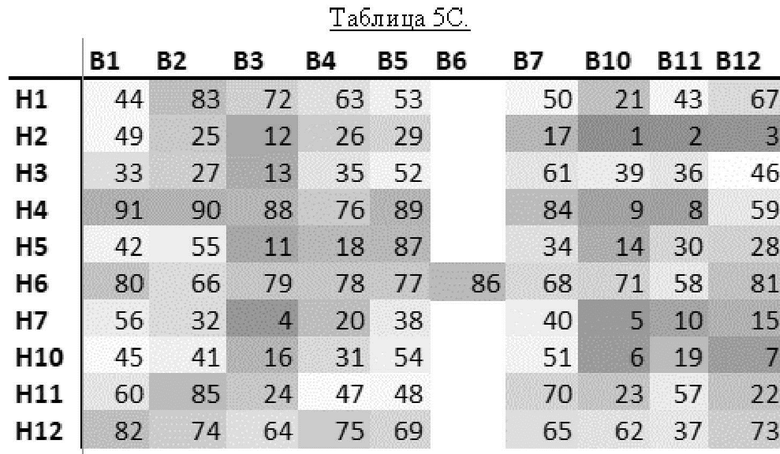
В таблице 5D показано среднее значение log10 лизиса каждого использованного штамма Gardnerella. Средние значения log10 активности рассчитали для всех протестированных конструкций.

В Таблицах 5А-5С показано, что активность эндолизинов была высокоспецифичной в зависимости от комбинации Н-домена/В-области и бактериального штамма, на котором ее тестировали. Каждую конструкцию тестировали на четырех штаммах Gardnerella (см. Таблицу 5А). В среднем, наиболее активным Н-доменом являлся Н2, со средним снижением CFU на 3,1 log10 единиц в сочетании со всеми В-областями (кроме В6), за ним следовали Н7, Н10 и Н5 (см. Таблицу 5В). Среди В-областей наиболее активной являлась В10 со средним снижением CFU на 2,7 log10 единиц, за которой следовали B11, В12 и В3.
Удивительным и неожиданным оказалось то, что некоторые рекомбинантные эндолизины обладали более высокой активностью, чем любой естественный эндолизин (от H1B1 до Н12В12), особенно при рассмотрении всех 4 протестированных штаммов Gardnerella (см. Фиг. 8A-8D). В частности, Н2В10, Н2В11 и Н2В12 занимали 1, 2 и 3 место по активности, соответственно, и каждый из них был более активен, чем любой естественный эндолизин (см. Таблицу 5С). Н7В3 занимает в целом 4 место (см. Таблицу 5С) и также более активен, чем любой другой естественный эндолизин, включенный в эксперимент. Фактически, единственный естественный эндолизин, входящий в десятку наиболее активных, это Н10В10 (6 место), следующим по активности естественным эндолизином является Н3В3 (13 место). Таким образом, рекомбинантные эндолизины по настоящему изобретению могут проявлять значительно более высокую активность, чем естественные эндолизины.
Не ограничиваясь какой-либо конкретной теорией, неожиданное увеличение активности, вызывающей гибель Gardnerella, наблюдаемое при замене доменов эндолизинов по изобретению, можно объяснить следующим образом. Естественные эндолизины профагов подвергаются процессу дарвиновской эволюции в ходе которого оптимизируется распространение всего профага. Однако вероятность мутаций, приводящих к лучшему размножению соответствующего профага за счет одновременного улучшения каталитической активности эндолизина и в то же время расширения круга его хозяев между видами только в пределах Gardnerella, очень мала. Напротив, у некоторых профагов Gardnerella должен был развиться до наивысшей активности N-концевой каталитический домен эндолизина, а у некоторых других профагов должна была произойти оптимизация С-концевой области эндолизина до наиболее широкой активности в отношении видов Gardnerella. Таким образом, в результате комбинации высокоразвитого N-концевого каталитического домена одного из эндолизинов по изобретению с высокоразвитой С-концевой областью другого эндолизина по изобретению, кодируемого другим геномом другого профага, могут быть получены рекомбинантные эндолизины с более оптимизированной уничтожающей активностью в отношении видов Gardnerella, по сравнению с естественными эндолизинами по изобретению. Это достигается, например, у рекомбинантных эндолизинов по изобретению Н2В10, Н2В11, Н2В12 и Н7В3, все из которых обладают более высокой уничтожающей активностью, чем естественный эндолизин EL2 или любые другие естественные эндолизины, и это справедливо для всех видов Gardnerella (см. Фиг. 8А - 8D и Таблицу 5А).
Более того, большинство конструкций гораздо более активны в отношении Gv_23 (G. swidsinskii), чем в отношении трех других протестированных штаммов (см. Таблицу 5А). Средняя активность всех конструкций в отношении каждого штамма Gardnerella (см. Таблицу 5D) подтверждает, что Gv_23 является наиболее чувствительным к эндолизинам, за ним следуют Gv_9 и Gv_11, a Gv_17 (G. piotii) является наименее чувствительным. Такое ранжирование чувствительности в большей мере справедливо для эндолизиновых конструкций. Хотя Gv_23 является наиболее чувствительным штаммом, ко многим конструкциям его чувствительность выше на несколько log10 единиц, иногда Gv_17 более чувствителен, чем Gv_9 или Gv_11 (например, к Н2В10, наиболее активному эндолизину в целом). Не ограничиваясь какой-либо конкретной теорией, разницу в чувствительности можно объяснить либо структурным недостатком, таким как более слабая/более тонкая/более доступная клеточная стенка Gv_23, либо более сильной ферментативной активностью протестированных эндолизинов в отношении Gv_23.
Кроме того, можно сделать вывод, что концентрация эндолизинов в растворе имеет решающее значение для их активности в анализе. При тестировании активности конструкции с низкой концентрацией согласно Таблице 4 обычно также имеют низкую активность, особенно конструкции с Н-доменами Н4, Н11 и Н12 (которые придают низкую растворимость с любыми В-областями). Есть отдельные сюрпризы, такие как Н12В11, который имел очень низкий уровень экспрессии, но сравнительно высокую активность.
[0149] 4.5 - Анализ паттернов активности
[0150] Последовательности естественных Н-доменов выравнивали и сравнивали, чтобы соотнести с паттернами активности, как показано на дендрограмме на Фиг. 9. Ожидалось, что наиболее активные Н-домены будут также наиболее близкородственными. Однако неожиданно оказалось, что наиболее активный N-концевой домен, Н2, является наиболее близким гомологом Н6, который является наименее активным. Следующие по активности Н-домены, Н7 и Н10, также довольно отдалены друг от друга и от Н2. Четвертый по активности Н-домен, Н5, наиболее близок к Н7. Также комбинации с В-областями, которые делают рекомбинантные эндолизины наиболее активными, не позволяют получить предсказуемый паттерн. Н2 наиболее активен в сочетании с B10, В11 и В12. Однако второй по активности Н-домен, Н7, наиболее активен в комбинации с В3, как и его ближайший гомолог Н5.
Также осуществляли выравнивание В-областей, чтобы установить гомологию по паттерну активности, как показано на дендрограмме на Фиг. 10. Наиболее активными В-областями по результатам анализа данных из Таблиц 5А - 5С являются В10, В11, В12, за которыми следует В3, все из которых имеют средние значения снижения CFU выше 2 log10 единиц. В отличие от паттерна, наблюдаемого для Н-доменов, эти 4 наиболее активных В-области являются 4 ближайшими гомологами в группе протестированных В-областей. Интересно, что области В5 и В7 идентичны (см. Фиг. 10). В целом, наилучшие результаты были получены для Н2В10, Н2В11 и Н2В12, как следует из Фиг. 8А - 8D.
Как объяснено в Примере 2, каждая естественная В-область содержит два В-домена, а именно N-концевой связывающийся с клеточной стенкой домен и С-концевой связывающийся с клеточной стенкой домен. Последовательности каждого естественного В-домена в составе В-области также выравнивали и сравнивали, как показано на Фиг. 11 и 12. Границы В-доменов можно установить как путем анализа последовательности при помощи InterPro (Mitchell et al., 2019 Nucleic Acids Res. 47, D351-D360), так и путем выравнивания двух повторяющихся мотивов в каждой В-области. С-конец всех В-доменов представляет собой консервативную последовательность (VNELL или VNKLL), гомолог которой можно найти также на С-конце мотивов CW_7 (VNELL или VNEIL) белка Cpl-7, таким образом определяя границы двух В-доменов в каждой В-области. Будучи исключением, В6 имеет только один усеченный В-домен, что, вероятно, является причиной полного отсутствия активности у EL6.
В заключение, было показано, что определенная комбинация Н-домена и В-области имеет решающее значение, и каждая из комбинаций Н/В, результатом которой стали эндолизины с более высокой уничтожающей активностью по сравнению с естественными эндолизинами, явилась неожиданным и непредсказуемым открытием.
[0151] Пример 5 - Анализ активности в отношении полезных Lactobacilli
[0152] Здоровое влагалище заселяют в основном 3 вида Lactobacilli: L. crispatus, L. gasseri и L. jensenii. Они поддерживают кислые значения рН 3,5-4,5, производя молочную кислоту, и защитную окислительную среду, производя Н2О2. Выздоровление после BV ассоциировано с заселением влагалища заново этими лактобациллами, и лекарство против BV предпочтительно не должно мешать этому процессу. Очевидно, что антибиотики мешают, поэтому по-прежнему существует острая медицинская потребность в улучшенных способах и композициях для лечения инфекций и BV, вызванных Gardnerella. Успешно продемонстрировав высокую активность эндолизинов по изобретению в отношении Gardnerella, авторы изобретения исследовали, могут ли эти эндолизины лизировать штаммы 3 видов Lactobacilli, наиболее часто встречающихся в здоровом влагалище. Эксперимент проводили, используя способ, описанный в Справочном Примере 2, при рН 5,0 в анаэробных условиях. Как показано на Фиг. 13, протестированные рекомбинантные эндолизины не проявляли какой-либо уничтожающей активности в отношении трех видов использованных полезных Lactobacilli, а именно L. crispatus, L. gasseri и L. jensenii. Эндолизины по изобретению, хотя и проявляют выраженную уничтожающую активность в отношении Gardnerella, неэффективны против наиболее часто встречающихся полезных Lactobacilli. Таким образом, эти результаты подтверждают селективную в отношении рода активность эндолизинов по изобретению и их статус лекарственного средства-кандидата в качестве инновационного лекарственного средства против BV. В этой связи, лечение BV эндолизинами по изобретению имеет большие преимущества по сравнению с доступными в настоящее время способами лечения, такими как лечение антибиотиками метронидазолом и клиндамицином.
[0153] Пример 6 - Анализ активности антибиотиков метронидазола и клиндамицина, являющихся стандартом лечения, в отношении роста штаммов Gardnerella в суспензии
[0154] Одним из основных недостатков лечения BV является высокая частота рецидивов у многих женщин, что приводит к многократному введению антибиотиков и сопутствующей дестабилизации микробиома и другим побочным эффектам. Далее, используя метод, описанный в Справочных Примерах 6 и 7, измеряли минимальную ингибирующую концентрацию (MIC) и минимальную бактерицидную концентрацию (МВС) в отношении клеток штаммов Gardnerella, которые также использовали в анализе активности эндолизинов. Вкратце, протокол измерений пришлось значительно адаптировать по сравнению с международным стандартом, который не подходит для измерения MIC и МВС на Gardnerella. Основными параметрами, которые требовалось изменить, были питательная среда (Gardnerella не растут в бульоне Мюллера-Хинтона, который обычно используют для измерения MIC), анаэробные условия, время инкубации, а также начальная концентрация бактерий в первом раунде экспериментов. Начальную концентрацию изменяли со стандартной 5×105 CFU/мл на 2,5×107 CFU/мл, главным образом потому, что во влагалище пациента с BV клетки также очень сконцентрированы, и действие антибиотика следует измерять при плотности клеток, более сопоставимой с той, которую использовали для анализа активности эндолизинов. Оценивали влияние метронидазола (полученного от компании Gatt-Koller) и клиндамицина (полученного от компании Ratiopharm) на рост штаммов Gardnerella в суспензии. По существу, суспензии Gardnerella в концентрации 2,5×107 CFU/мл инкубировали с антибиотиками в указанных концентрациях и инкубировали при 37°С в течение 48 ч в анаэробных условиях. MIC определяли как минимальную концентрацию антибиотика, при которой рост через 48 часов не детектировался путем измерения OD. OD(610) измеряли в начале и в конце эксперимента. В конце эксперимента 2 мкл каждой реакционной смеси наносили на агар для определения МВС. Результаты эксперимента, показанные на Фиг. 14, обобщены в Таблице 6А. Устойчивость (R) определяли как >= 32 мкг/мл для метронидазола и >8 мг/мл для клиндамицина. В соответствии с международными стандартами, чувствительность (S) определяли как <= 8 мкг/мл для метронидазола и <= 2 мкг/мл для клиндамицина.
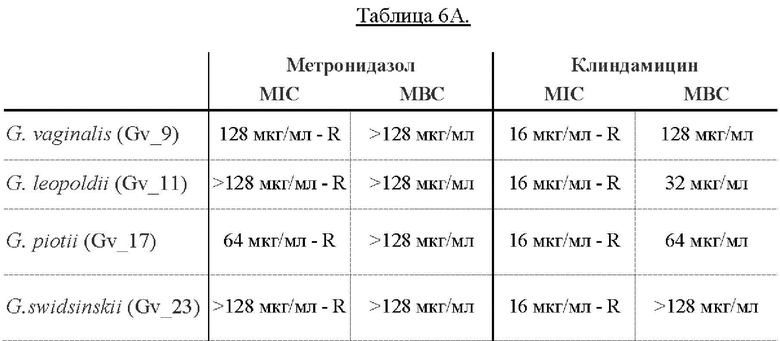
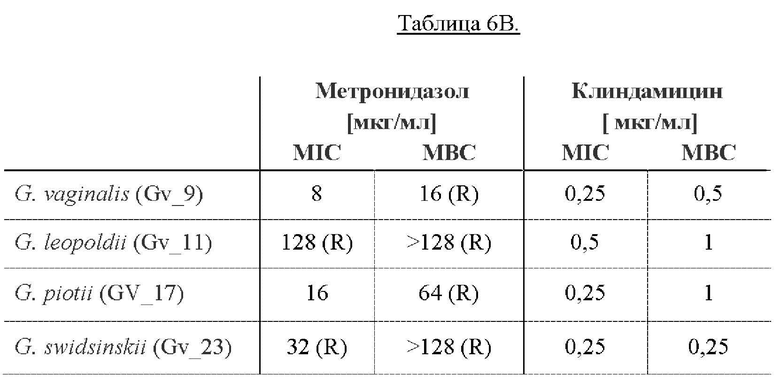
Согласно результатам, представленным на Фиг. 14, все штаммы Gardnerella имеют низкую чувствительность как к метронидазолу, так и к клиндамицину. Условия, при которых измеряли MIC и МВС, более строгие, чем стандартные. Обычно измеряют МВС90, то есть концентрацию антибиотика, вызывающую гибель 90% клеток в течение определенного времени, в то время как в настоящей заявке МВС определяли как минимальную концентрацию, вызывающую полную эрадикацию суспензии 2,5×107 CFU/мл. Тем не менее, эти условия более сопоставимы с тем, что обнаруживается во влагалище пациента с BV. Высокие значения MIC и МВС, измеренные в этих условиях, могут объяснить высокую частоту рецидивирования BV. Исследуемые эндолизины, напротив, являются бактерицидными по определению, поскольку они вызывают полное разрушение бактериальной клетки. Таким образом, эти результаты подтверждают, что эндолизины по изобретению превосходят антибиотики при лечении BV.
Проводили второй раунд экспериментов с определением MIC и МВС, где изменяли некоторые параметры эксперимента: (1) начальное количество клеток 1×105-1×106 теперь находилось в соответствии со стандартами CLSI (Институт клинических и лабораторных стандартов), (2) использовали гидрохлорид клиндамицина в порошке (Sigma Aldrich, кат. номер С5269), (3) использовали бульон NYC-III вместо бульона Харди и (iv) использовали 96-луночный планшет вместо 384-луночных планшетов. Как показано на Фиг. 15 и обобщено в Таблице 6В, MIC метронидазола по-прежнему была очень низкой для всех штаммов Gardnerella (8-128 мкг/мл), тогда как гидрохлорид клиндамицина, в отличие от клиндамицина, представленного на Фиг. 14 и в Таблице 6А, теперь оказывал ингибирующее и бактерицидное действие при низкой концентрации (MIC ≤ 1 мкг/мл).
Пример 7 - Анализ активности репрезентативного эндолизина с замененным доменом (Н2В10) в отношении роста различных штаммов Gardnerella в суспензии. Для анализа MIC и МВС с эндолизином Н2 В10 использовали 1×105-1×106 клеточных суспензий. Н2В10 демонстрировал MIC в диапазоне низких мкг/мл (0,5-4 мкг/мл), что указывает на высокую чувствительность клеток Gardnerella к эндолизинам (Фиг. 16, Таблица 7). Условия, в которых измеряли МВС, являются более строгими по сравнению со стандартными. Например, обычно измеряют МВС90, то есть концентрацию противомикробного средства, вызывающую гибель 90% клеток в течение определенного времени, в то время как в настоящей заявке МВС определяли как дозу, уменьшающую количество клеток по меньшей мере на 99,5% по сравнению с исходным. Н2В10, как репрезентативный представитель заявленных эндолизинов, демонстрировал значительно более высокие значения MIC и МВС по сравнению со стандартным антибиотиком метронидазолом, который не эффективен в отношении многих штаммов Gardnerella из-за формирования устойчивости. Однако, с клиндамицином получали противоречивые результаты. В соответствии с международными стандартами, все четыре штамма Gardnerella оказались устойчивыми (MIC > 8 мкг/мл) к клиндамицину, полученному от компании Ratiopharm (Фиг. 14). тогда как гидрохлорид клиндамицина (Sigma Aldrich) был гораздо более эффективен, оказывая бактерицидное действие уже в концентрациях ≤1 мкг/мл (Фиг. 16). В целом известно, что антибиотики обеспечивают лишь недостаточную эрадикацию биопленки Gardnerella, характерного признака BV, что является одной из предполагаемых причин очень высокой частоты рецидивирования BV. Оставляя остаточную жизнеспособную биопленку, антибиотики лишают микробном влагалища части благотворных организмов, что впоследствии открывает экологическую нишу для других патогенов, например, грибов. Таким образом, лечение на основе эндолизина, которое обеспечивает избирательную эрадикацию бактериальных клеток рода Gardnerella и предположительно обеспечивает эрадикацию биопленки, не нанося вреда полезным Lactobacilli, должно превосходить стандартную антибиотикотерапию BV.

[0155] Справочный пример 1 - Клонирование, экспрессия и очистка эндолизинов
Материалы
• 96-луночный полистироловый фильтрующий планшет Multiscreen HTS Durapore Filterplatten (Labshop, каталожный номер 44.MSGVS22)
• Обессоливающие колонки PD MiniTrap с Sephadex G-25 (GE Lifescience, каталожный номер 28918007)
• Устройство для диализа Slide-A-Lyzer™ MINI, НОММ 10K, 2 мл (Thermo Scientific, каталожный номер 88404)
• Реагент Fastbreak (Promega, каталожный номер V8571)
• Лизирующий буфер: 50 мМ фосфат рН 6, 150 мМ NaCl, 20 мМ имидазол, 1 мМ ТСЕР, 1x FastBreak, бензоназа.
• Отмывочный буфер I: 50 мМ фосфат рН 6, 150 мМ NaCl, 20 мМ имидазол, 1 мМ ТС ЕР (1,5 мл)
• Отмывочный буфер II: 50 мМ фосфат рН 6, 150 мМ NaCl, 40 мМ имидазол, 1 мМ ТС ЕР (1,5 мл)
• Элюирующий буфер: 50 мМ фосфат рН 6, 150 мМ NaCl, 250 мМ имидазол, 1 мМ ТС ЕР (1,5 мл)
Способ:
Штамм Е. coli Bl 21(DE3) трансформировали экспрессируемыми конструкциями и проводили отбор с использованием соответствующих антибиотиков. Клетки из 2 мл культуры (ТВ + лактоза, 25°С, в течение ночи) ресуспендировали в 1,5 мл лизирующего буфера и лизировали реагентом FastBreak (Promega). Внутриклеточную растворимую фракцию выделяли центрифугированием при 15000 g в течение 30 мин при 4°С. Фракцию растворимого белка загружали в 100 мкл аффинного сорбента с ионами никеля, промывали 15 объемами колонки (CV) каждого из отмывочных буферов I и II и элюировали 10 CV элюирующего буфера. Затем можно было провести замену буфера элюата на 20 мМ фосфат, рН 6,0, 150 мМ NaCl, чтобы удалить имидазол, используя обессоливающие колонки. После элюирования (или замены буфера, при необходимости) концентрацию очищенного белка доводили до 0,2 мг/мл, затем растворы стерилизовали фильтрованием с использованием 96-луночного фильтрующего планшета.
[0156] Справочный Пример 2 - Анализ активности в бактериальных суспензиях
Материалы
1. Бульон Харди, автоклавированный в течение 20 минут при 121°С:
• 12,0 г панкреатического гидролизата казеина (SigmaAldrich, каталожный номер 70172-100G)
• 10,0 г протеозопептона (SigmaAldrich, каталожный номер 82450-100G)
• 5,0 г продуктов переваривания животных тканей пепсином (SigmaAldrich, каталожный номер 70174-100G)
• 5,0 г хлорида натрия (CarlRoth, каталожный номер 3957.1)
• 3,0 г мясного экстракта (SigmaAldrich, каталожный номер B4888-50G)
• 3,0 г дрожжевого экстракта (SigmaAldrich, каталожный номер Y1625-250G)
• 1,0 г растворимого крахмала (Sigma Aldrich, каталожный номер S9765-250G)
• деионизированная H2O до 1 литра (полученная в лаборатории PhagoMed с применением Millipore RiOs Essential 16)
2. Агар Харди, автоклавированный в течение 20 минут при 121°С: Аналогично бульону Харди, но с добавлением 15 г бактериологического агара (OXOID, каталожный номер LP0011)
3. Агар Харди для верхнего слоя, автоклавированный в течение 20 минут при 121°С: Аналогично бульону Харди, но с добавлением 7 г бактериологического агара (OXOID, каталожный номер LP0011)
4. Среда NYC-III, рН 5,0, автоклавированная в течение 20 минут при 121°С с последующим добавлением лошадиной сыворотки (NYC-III-HS-5.0)
• 12 г HEPES (Sigma Life Science, каталожный номер H4034-100G)
• 7,5 г протеозопептона No. 3 (BD, каталожный номер 211693)
• 1,9 г дрожжевого экстракта (SigmaAldrich, каталожный номер Y1625-250G)
• 2,5 г хлорида натрия (SigmaAldrich, каталожный номер S9888-lkg-M)
• 2,5 г глюкозы (молекулярная масса 180,16 г/моль) (Sigma Aldrich, каталожный номер G6152-1KG)
• деионизированная вода до общего объема 450 мл
• 50 мл лошадиной сыворотки (HS), инактивированной нагреванием, 100 мл (Thermo Fisher Scientific, кат.26050070), добавляемой после автоклавирования
5. Общие материалы:
• Чашки с шоколадным агаром BD для Gardnerella (BD, каталожный номер 254060)
• Пластины BD Schaedler / 5% овечьей крови для Lactobacilli (BD, каталожный номер 254042)
• Isovitalex (BD, каталожный номер 211876)
• Бульон Харди + Isovitalex (см. выше), рН доводили до указанного значения
• Агар Харди + Isovitalex (см. выше),
• Агар Харди для верхнего слоя + Isovitalex (см. выше),
• 96-луночный круглодонный планшет (Sigma Aldrich, каталожный номер М2311-100ЕА)
• 96-луночный плоскодонный планшет с крышкой (Labshop, каталожный номер 44.781662)
• 384-луночные планшеты Greiner CELLSTAR® (Sigma-Aldrich, каталожный номер М1937-32ЕА)
• Пакеты для создания анаэробной атмосферы (Sigma-Aldrich, каталожный номер 68061-10SACHETS-F)
• Индикаторный тест для анаэробной атмосферы (Sigma-Aldrich, каталожный номер 59886-1PAK-F)
• Анаэростат (Sigma-Aldrich, каталожный номер 28029-1EA-F) или пищевой пластиковый контейнер с резиновым уплотнителем для герметичного закрывания, купленный в местном магазине бытовых принадлежностей
6. Штаммы бактерий: Штаммы Gardnerella:
• Gv_1: UGent 09.07
• Gv_8: UGent 25.49
• Gv_9: ATCC 14018
• Gv_10: UGent 06.41
• Gv_11: UGent 09.48
• Gv_17: UGent 18.01
• Gv_23: GS 10234 (FC2)
Штаммы Lactobacilli:
• L. jensenii PB2003-013-T2-2
• L. gasseri 020566
• L. crispatus LAB117
Способ:
Клетки гарднереллы извлекали из криогенного хранилища, высевали на чашки с шоколадным агаром (Beckton Dickinson) и инкубировали в течение 48 часов при 37°С в анаэробных условиях. Для лактобацилл вместо этого использовали чашки BD Schaedler с 5% овечьей крови. Колонии соскребали с чашки, ресуспендировали в бульоне Харди или NYC-III-HS-5,0 с указанным рН, и суспензию доводили до OD (при длине волны 610 или 620 нм, как указано) 0,1. Следует отметить, что в экспериментах взаимозаменяемо использовали два считывающих устройства для микропланшетов Tecan, имеющие соответственно фильтр 610 нм или 620 нм. Хотя это не имеет значения для экспериментов, точная длина волны указана в каждом примере. Если не указано иное, в 384-луночных планшетах смешивали 90 мкл клеточной суспензии и 10 мкл раствора эндолизина в различных комбинациях вид/эндолизин. OD(610-620 нм, как указано) измеряли либо в двух точках в начале и в конце реакции, либо проводили непрерывное измерение кинетическим методом при помощи считывающего устройства для микропланшетов Tecan F200. Реакции проводили в течение 5 часов (или другого указанного времени) при 37°С в анаэробных, микроаэрофильных или аэробных условиях, как указано. Анаэробные условия предполагают, что в контейнере, в котором инкубируовали бактерии (анаэростат Sigma-Aldrich или герметичный пищевой контейнер), кислород был полностью истощен при помощи анаэробного пакета, и отсутствие кислорода подтверждалось анаэробным индикатором внутри контейнера. Там, где указаны микроаэрофильные условия, использовали метод свечи в банке (свечу зажигали в соответствующем герметичном контейнере, что снижало уровень кислорода до тех пор, пока пламя не погаснет). Затем для обнаружения и количественного определения жизнеспособных CFU содержимое каждой лунки разводили в 5 шагов (от 10-1 до 10-5) с использованием 96-луночных круглодонных планшетов и 2 мкл каждого разведения каждой реакционной смеси высевали на чашки с шоколадным агаром BD либо на чашки с агаром BD Schaedler с 5% овечьей крови в случае Gardnerella и Lactobacilli, соответственно. Планшеты, в которых проводили детекцию, инкубировали при 37°С в течение 48 часов в анаэробных условиях.
[0157] Справочные Примеры 6 и 7 - измерения MIC и МВС
Материалы
Общие материалы:
• Метронидазол (Gatt-Koller, Metronidazolum mikronisiert, 10 г, 606293914)
• Клиндамицин (Ratiopharm, ампулы по 300 мг/2 мл, 5 шт)
• Гидрохлорид клиндамицина (Sigma Aldrich, 10 мг, каталожный номер С5269)
• Эндолизин Н2В10 [530 мкг/мл] в буфере MES (50 мМ MES, 100 мМ NaCl, 8 мМ MgSO4, рН=5,5)
• Чашки с шоколадным агаром BD для Gardnerella (BD, каталожный номер 254060)
• Пластины BD Schaedler / 5% овечьей крови для Lactobacilli (BD, каталожный номер 254042)
• Isovitalex (BD, каталожный номер 211876)
• Бульон Харди + Isovitalex (см. выше), рН доводили до указанного значения
• среда NYC-III+HS, рН 5,5 Чашки с агаром NYC-III-HS: Среда NYC-III+HS, как описано выше, но с добавлением 1,5% агара перед автоклавированием
• Агар Харди + Isovitalex (см. выше),
• Агар Харди для верхнего слоя + Isovitalex (см. выше),
• 96-луночный круглодонный планшет (Sigma Aldrich, каталожный номер М2311-100ЕА)
• 96-луночный плоскодонный планшет с крышкой (Labshop, каталожный номер 44.781662)
• 384-луночные планшеты Greiner CELLSTAR® (Sigma-Aldrich, каталожный номер М1937-32ЕА)
• Пакеты для создания анаэробной атмосферы (Sigma-Aldrich, каталожный номер 68061-10SACHETS-F)
• Индикаторный тест для анаэробной атмосферы (Sigma-Aldrich, каталожный номер 59886-1PAK-F)
• Анаэростат (Sigma-Aldrich, каталожный номер 28029-1EA-F) или пищевой пластиковый контейнер с резиновым уплотнителем для герметичного закрывания, купленный в местном магазине бытовых принадлежностей
Способ:
Бактерии из криогенного хранилища высевали на чашки с шоколадным агаром BD (Gardnerella) и инкубировали в течение 48 часов при 37°С в анаэробных условиях. Колонии соскребали с чашки, ресуспендировали в бульоне Харди или NYC-III-HS-pH 5,0, и суспензию доводили до OD (при длине волны 610 или 620 нм, как указано) 0,05. Следует отметить, что в экспериментах взаимозаменяемо использовали два считывающих устройства для микропланшетов Tecan, имеющие фильтр 610 нм или 620 нм, соответственно. Хотя это не имеет значения для экспериментов, точная длина волны указана в каждом примере. Готовили антибиотики в виде 20-кратного концентрированного раствора для каждой из требуемых конечных концентраций. В 384-луночном планшете смешивали 95 мкл клеточной суспензии и 5 мкл раствора антибиотика. Измеряли OD (610-620, как указано) в начале реакции, затем планшет инкубировали при 37°С в течение 48 часов в анаэробных условиях. После этого снова измеряли OD (610-620, как указано) для определения MIC, при этом MIC определяли как самую низкую концентрацию антибиотика, при которой OD (610-620, как указано) не превышала уровня, измеренного в начале эксперимента. После измерения OD 2 мкл из каждой лунки наносили на чашку с агаром NYC-III+HS. Затем планшеты инкубировали еще 48 часов при 37°С в анаэробных условиях. После инкубации оценивали рост клеток в каждой колонии, и МВС определяли как самую низкую концентрацию антибиотиков, при которой бактерии на чашке не росли. Эксперименты проводили в трипликатах для каждого условия.
Во втором раунде экспериментов для определения MIC и МВС антибиотиков суспензию клеток Gardnerella доводили до стандарта Макфарланда 0,5 (OD(610) приблизительно 0,07), а затем разводили 1:75 в соответствии со стандартами CLSI (Института клинических и лабораторных стандартов). Антибиотики готовили в соответствии со стандартами CLSI и смешивали 50 мкл клеточной суспензии в 96-луночном планшете с 50 мкл антибиотиков. В противном случае MIC и МВС определяли, как описано выше.
Для определения MIC и МВС эндолизина Н2В10 с замененным доменом 50 мкл суспензии клеток Gardnerella смешивали в 96-луночном планшете с 50 мкл раствора, содержащего Н2В10, последовательно разведенного 1:1. Измеряли OD610 в начале реакции, затем планшет инкубировали при 37°С в течение 48 часов в анаэробных условиях. После этого снова измеряли OD(610) для определения MIC, при этом MIC определяли как самую низкую концентрацию Н2В10, при которой OD не превышала или незначительно превышала уровень, измеренный в начале эксперимента.
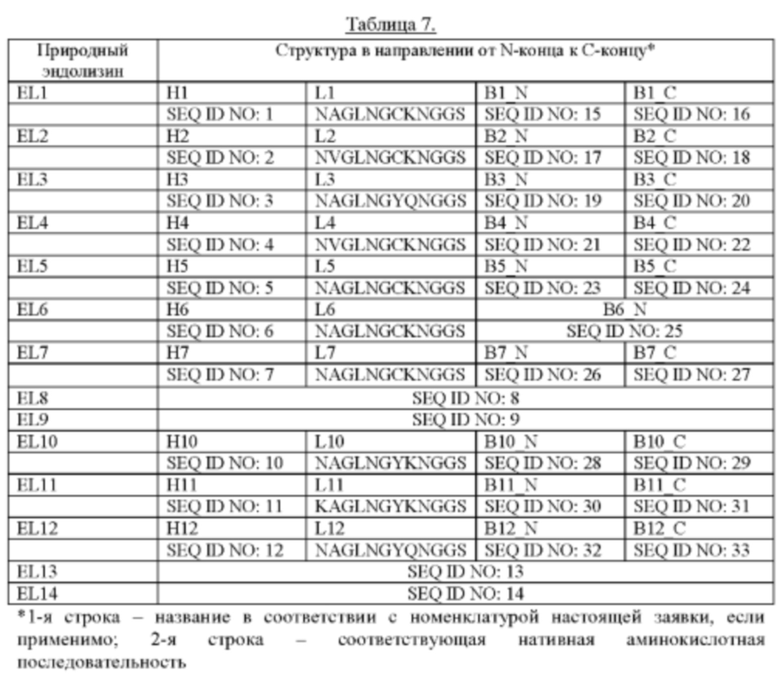
--->
Перечень последовательностей
SEQUENCE LISTING
<110> PhagoMed Biopharma GmbH
<120> Novel Gardnerella endolysins and uses thereof
<130> AC1230 PCT S3
<150> EP19 17 3389.8
<151> 2019-05-08
<160> 97
<170> BiSSAP 1.3.6
<210> 1
<211> 196
<212> PRT
<213> Artificial Sequence
<220>
<223> H1
<400> 1
Met Ser Lys Lys Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Leu Met Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ala His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Asn Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Asn Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys
195
<210> 2
<211> 196
<212> PRT
<213> Artificial Sequence
<220>
<223> H2
<400> 2
Met Ser Lys Arg Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Cys Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Gly Phe Cys Asn Lys Leu Glu Ser Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ala His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys
195
<210> 3
<211> 196
<212> PRT
<213> Artificial Sequence
<220>
<223> H3
<400> 3
Met Ser Lys Lys Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ser Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly His Lys Asp Lys Trp Phe Glu Glu Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ser Tyr Trp Tyr Ser Tyr
50 55 60
Ala Ser Ser Ala Gly Glu Ala Ser Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Ile Leu Ser Gly Lys Ser Phe Glu Tyr Pro Ile Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Asp Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Val Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asp Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Asn
165 170 175
Gly Ile Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Val Ile Lys
195
<210> 4
<211> 196
<212> PRT
<213> Artificial Sequence
<220>
<223> H4
<400> 4
Met Ser Lys Lys Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Cys Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Ser Lys Leu Glu Thr Tyr Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Val Val Asn Asn Leu Val Ser Ala His Val Arg
130 135 140
Asp Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys
195
<210> 5
<211> 196
<212> PRT
<213> Artificial Sequence
<220>
<223> H5
<400> 5
Met Ser Lys Lys Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Ser Ser Glu Ala Ala Glu Glu Ala Gln Ser Cys Ala Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Gly Phe Cys Ser Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ala His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Asn Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Lys Asp Tyr Pro
180 185 190
Ser Ile Ile Lys
195
<210> 6
<211> 196
<212> PRT
<213> Artificial Sequence
<220>
<223> H6
<400> 6
Met Ser Lys Lys Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Cys Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Ser Lys Leu Glu Ser Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ala His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Asp Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Lys Asn Tyr Pro
180 185 190
Ser Ile Ile Lys
195
<210> 7
<211> 196
<212> PRT
<213> Artificial Sequence
<220>
<223> H7
<400> 7
Met Ser Lys Lys Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Ala Ser Glu Ala Ala Glu Glu Ala Gln Ser Cys Ala Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Ser Lys Leu Glu Thr Tyr Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Lys Asp Tyr Pro
180 185 190
Ser Ile Ile Lys
195
<210> 8
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL8
<400> 8
Met Ser Lys Lys Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Ser Ser Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Ser Lys Leu Glu Thr Tyr Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Asn Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Ala Gly Leu Asn Gly Tyr Lys Asn Gly Gly Ser
195 200 205
Tyr Thr Ala Pro Gln Thr Ser Ser Ile Asp Asp Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
225 230 235 240
Gln Ala Gly Tyr Asp Tyr Thr Ser Val Gln Asn Lys Val Asn Lys Leu
245 250 255
Leu Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Thr Trp Gly Asn Gly Asn Glu Arg Lys Asn Arg Leu
275 280 285
Thr Gln Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 9
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> EL9
<400> 9
Met Ser Lys Lys Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Cys Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Val Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Asp Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Ala Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
195 200 205
Asp Gln Ala Ala Arg Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu Thr
225 230 235 240
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Ala Lys
245 250
<210> 10
<211> 196
<212> PRT
<213> Artificial Sequence
<220>
<223> H10
<400> 10
Met Ser Lys Arg Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly His Lys Asp Lys Trp Phe Glu Glu Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Val Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Ser Ser Ala Gly Glu Ala Ser Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Ile Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Asp Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Val Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asp Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Asn
165 170 175
Gly Ile Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Val Ile Lys
195
<210> 11
<211> 196
<212> PRT
<213> Artificial Sequence
<220>
<223> H11
<400> 11
Met Ser Lys Arg Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly His Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Thr Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Ser Ser Ala Gly Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Ile Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Asp Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Thr Tyr Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Val Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asp Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Asp Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Asp
165 170 175
Gly Ile Ala Gly Arg Val Asp Met Asp Tyr Thr Tyr Val Asp Tyr Pro
180 185 190
Ser Val Ile Lys
195
<210> 12
<211> 196
<212> PRT
<213> Artificial Sequence
<220>
<223> H12
<400> 12
Met Ser Lys Lys Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly His Lys Asp Lys Trp Phe Glu Glu Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ser Tyr Trp Tyr Ser Tyr
50 55 60
Ala Ser Ser Ala Gly Glu Val Ala Leu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Ile Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Asp Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Val Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asp Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Asn
165 170 175
Gly Ile Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Val Ile Lys
195
<210> 13
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL13
<400> 13
Met Ser Lys Lys Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Glu Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ser Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Pro Ala His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Asp Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Ala Gly Leu Asn Gly Tyr Lys Asn Gly Glu Ser
195 200 205
His Gln Ala Thr Arg Thr Thr Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
225 230 235 240
Gln Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Glu Leu
245 250 255
Leu Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Thr Trp Gly Asn Gly Asn Glu Arg Lys Asn Arg Leu
275 280 285
Thr Ser Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 14
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL14
<400> 14
Met Ser Lys Lys Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Cys Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ala His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Ala Gly Leu Asn Gly Tyr Lys Asn Gly Glu Ser
195 200 205
His Gln Ala Thr Arg Thr Thr Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
225 230 235 240
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Glu Leu
245 250 255
Leu Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Ala Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu
275 280 285
Thr Ser Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 15
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B1_N
<400> 15
Asp Gln Ala Ala Arg Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
1 5 10 15
Ile Asn Gly Ala Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu Thr
20 25 30
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Glu Leu
35 40 45
Leu
<210> 16
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B1_C
<400> 16
Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Leu Ala Arg Glu Val
1 5 10 15
Ile Arg Gly Ala Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu Ala
20 25 30
Gln Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu Leu
35 40 45
Leu
<210> 17
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B2_N
<400> 17
Asp Gln Ala Ala Arg Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
1 5 10 15
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
20 25 30
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Glu Leu
35 40 45
Leu
<210> 18
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B2_C
<400> 18
Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Ile Ala Arg Glu Val
1 5 10 15
Ile Arg Gly Thr Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu Thr
20 25 30
Gln Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu Leu
35 40 45
Leu
<210> 19
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B3_N
<400> 19
Tyr Thr Ala Pro Gln Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
1 5 10 15
Ile Asn Gly Asp Trp Gly Asn Gly Asn Asp Arg Lys Asn Arg Leu Ile
20 25 30
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Glu Leu
35 40 45
Leu
<210> 20
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B3_C
<400> 20
Gly Val Lys Ala Tyr Arg Lys Ser Val Asp Glu Leu Ala Arg Glu Val
1 5 10 15
Ile Arg Gly Thr Trp Gly Asn Gly Ser Met Arg Lys His Arg Leu Thr
20 25 30
Gln Ala Gly Tyr Asp Tyr Asp Ala Val Gln Lys Arg Val Asn Glu Leu
35 40 45
Leu
<210> 21
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B4_N
<400> 21
Asp Gln Ala Thr Arg Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
1 5 10 15
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
20 25 30
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Lys Leu
35 40 45
Leu
<210> 22
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B4_C
<400> 22
Gly Val Lys Ala Tyr Arg Lys Ser Val Asp Glu Leu Ala Arg Glu Val
1 5 10 15
Ile Arg Gly Thr Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Ala
20 25 30
Gln Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu Leu
35 40 45
Leu
<210> 23
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B5_N
<400> 23
Asp Gln Ala Ala Arg Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
1 5 10 15
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
20 25 30
Gln Ala Gly Tyr Asp Tyr Thr Ser Val Gln Asn Lys Val Asn Lys Leu
35 40 45
Leu
<210> 24
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B5_C
<400> 24
Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Leu Ala Arg Glu Val
1 5 10 15
Ile Arg Gly Thr Trp Gly Asn Gly Asn Glu Arg Lys Asn Arg Leu Thr
20 25 30
Gln Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu Leu
35 40 45
Leu
<210> 25
<211> 43
<212> PRT
<213> Artificial Sequence
<220>
<223> B6_N
<400> 25
Asn Gln Ala Ala Arg Thr Ser Ser Ile Asp Asp Val Ala Arg Glu Val
1 5 10 15
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
20 25 30
Gln Ala Gly Tyr Asp Tyr Ala Ser Val Ala Lys
35 40
<210> 26
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B7_N
<400> 26
Asp Gln Ala Ala Arg Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
1 5 10 15
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
20 25 30
Gln Ala Gly Tyr Asp Tyr Thr Ser Val Gln Asn Lys Val Asn Lys Leu
35 40 45
Leu
<210> 27
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B7_C
<400> 27
Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Leu Ala Arg Glu Val
1 5 10 15
Ile Arg Gly Thr Trp Gly Asn Gly Asn Glu Arg Lys Asn Arg Leu Thr
20 25 30
Gln Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu Leu
35 40 45
Leu
<210> 28
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B10_N
<400> 28
Tyr Thr Ala Pro Gln Ile Ser Ser Ile Asp Glu Val Ala Arg Glu Val
1 5 10 15
Ile Asn Gly Asp Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
20 25 30
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Glu Leu
35 40 45
Leu
<210> 29
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B10_C
<400> 29
Gly Val Lys Ala Tyr Arg Lys Ser Val Asp Glu Leu Ala Arg Glu Val
1 5 10 15
Ile Arg Gly Thr Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu Thr
20 25 30
Gln Ala Gly Tyr Asp Tyr Asn Ala Val Gln Lys Arg Val Asn Glu Leu
35 40 45
Leu
<210> 30
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B11_N
<400> 30
Tyr Thr Ala Pro Gln Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
1 5 10 15
Ile Asn Gly Asp Trp Gly Asn Gly Asn Glu Arg Lys Asn Arg Leu Thr
20 25 30
Ser Ala Gly Tyr Asp Tyr Thr Ser Val Gln Asn Lys Val Asn Glu Leu
35 40 45
Leu
<210> 31
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B11_C
<400> 31
Gly Val Lys Ala Tyr Arg Lys Ser Val Asp Glu Leu Ala Arg Glu Val
1 5 10 15
Ile Arg Gly Thr Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu Thr
20 25 30
Gln Ala Gly Tyr Asp Tyr Asp Ala Val Gln Lys Arg Val Asn Glu Leu
35 40 45
Leu
<210> 32
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B12_N
<400> 32
Tyr Thr Ala Pro Gln Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
1 5 10 15
Ile Asn Gly Asp Trp Gly Asn Gly Ile Glu Arg Lys Asn Arg Leu Thr
20 25 30
Ser Ala Gly Tyr Asp Tyr Thr Ser Val Gln Asn Lys Val Asn Glu Leu
35 40 45
Leu
<210> 33
<211> 49
<212> PRT
<213> Artificial Sequence
<220>
<223> B12_C
<400> 33
Gly Val Lys Ala Tyr Arg Lys Ser Val Asp Glu Leu Ala Arg Glu Val
1 5 10 15
Ile Arg Gly Thr Trp Gly Asn Gly Lys Thr Arg Lys Gln Arg Leu Thr
20 25 30
Gln Ala Gly Tyr Asp Tyr Asn Ala Val Gln Lys Arg Val Asn Glu Leu
35 40 45
Leu
<210> 34
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> linker polypeptide
<220>
<221> VARIANT
<222> 1
<223> Asn = Lys
<220>
<221> VARIANT
<222> 8
<223> Xaa = Lys or Gln
<400> 34
Asn Ala Gly Leu Asn Gly Tyr Xaa Asn Gly Gly Ser
1 5 10
<210> 35
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<220>
<221> VARIANT
<222> 1..6
<223> Xaa = Gly, Ala or Ser
<400> 35
Xaa Xaa Xaa Xaa Xaa Xaa
1 5
<210> 36
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<220>
<221> VARIANT
<222> 1..9
<223> Xaa = Gly, Ala or Ser
<400> 36
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
1 5
<210> 37
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<220>
<221> VARIANT
<222> 1..12
<223> Xaa = Gly, Ala or Ser
<400> 37
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
1 5 10
<210> 38
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<220>
<221> VARIANT
<222> 1..15
<223> Xaa = Gly, Ala or Ser
<400> 38
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
1 5 10 15
<210> 39
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<220>
<221> VARIANT
<222> 1..18
<223> Xaa = Gly, Ala or Ser
<400> 39
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
1 5 10 15
Xaa Xaa
<210> 40
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 40
Gly Gly Ser Gly Gly Ser
1 5
<210> 41
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 41
Gly Gly Ser Gly Gly Ser Gly Gly Ser
1 5
<210> 42
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 42
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
1 5 10
<210> 43
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 43
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
1 5 10 15
<210> 44
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 44
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
1 5 10 15
Gly Ser
<210> 45
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 45
Asn Ala Gly Leu Asn Gly Tyr Lys Asn Gly Gly Ser
1 5 10
<210> 46
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<220>
<221> VARIANT
<222> 1
<223> Xaa = Asn or Lys
<220>
<221> VARIANT
<222> 2
<223> Xaa = Ala or Val
<220>
<221> VARIANT
<222> 7
<223> Xaa = Tyr or Cys
<220>
<221> VARIANT
<222> 8
<223> Xaa = Lys or Gln
<400> 46
Xaa Xaa Gly Leu Asn Gly Xaa Xaa Asn Gly Gly Ser
1 5 10
<210> 47
<211> 39
<212> PRT
<213> Artificial Sequence
<220>
<223> CW_7 motif
<220>
<221> VARIANT
<222> 38
<223> Xaa is Ile for motif No. 1 and No. 2
Xaa is Leu for motif No. 3
<400> 47
Thr Val Ala Asn Glu Val Ile Gln Gly Leu Trp Gly Asn Gly Gln Glu
1 5 10 15
Arg Tyr Asp Ser Leu Ala Asn Ala Gly Tyr Asp Pro Gln Ala Val Gln
20 25 30
Asp Lys Val Asn Glu Xaa Leu
35
<210> 48
<211> 42
<212> DNA
<213> Artificial Sequence
<220>
<223> primer T7casette-16-rev
<400> 48
aacaggtctc aatacaatcc ggatatagtt cctcctttca gc 42
<210> 49
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> primer T7casette-01-for
<400> 49
aacaggtctc aacctccgcg aaattaatac gactcactat agg 43
<210> 50
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_H1-rev
<400> 50
aacaggtctc aagccggcat ttttgatgat gctcggg 37
<210> 51
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_H2-rev
<400> 51
aacaggtctc aagccaacat ttttaataat gctcggataa tcc 43
<210> 52
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_H3-rev
<400> 52
aacaggtctc aagccggcgt ttttgataac gctcggg 37
<210> 53
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_H4-rev
<400> 53
aacaggtctc aagcccacgt tcttgatgat gctcgg 36
<210> 54
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_H5-rev
<400> 54
aacaggtctc aagccggcgt ttttgatgat gctcgg 36
<210> 55
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_H6-rev
<400> 55
aacaggtctc aagccggcat tcttaataat gctcggata 39
<210> 56
<211> 42
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_H7-rev
<400> 56
aacaggtctc aagccggcgt ttttaatgat gctcggataa tc 42
<210> 57
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_H10-rev
<400> 57
aacaggtctc aagccggcgt ttttgatcac gctcgg 36
<210> 58
<211> 42
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_H11-rev
<400> 58
aacaggtctc aagccggcct tcttgataac gctcggataa tc 42
<210> 59
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_H12-rev
<400> 59
aacaggtctc aagccggcat tcttgatcac gctcgg 36
<210> 60
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_B6-for
<400> 60
aacaggtctc aggcttaaac ggctgcaaaa atggcgg 37
<210> 61
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_B7-for
<400> 61
aacaggtctc aggcttaaac ggctgcaaaa acggtgg 37
<210> 62
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_B10-for
<400> 62
aacaggtctc aggcttaaac ggctataaaa acggcggc 38
<210> 63
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_B11-for
<400> 63
aacaggtctc aggcttaaat ggttacaaga atggcggcag 40
<210> 64
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_B12-for
<400> 64
aacaggtctc aggcttaaat ggctaccaga acggcgg 37
<210> 65
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_B1-for
<400> 65
aacaggtctc aggcttaaac ggctgcaaga atggtgg 37
<210> 66
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_B2-for
<400> 66
aacaggtctc aggcttaaat ggttgcaaga acggcgg 37
<210> 67
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_B3-for
<400> 67
aacaggtctc aggcttaaat ggctaccaga atggcggc 38
<210> 68
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> primer EL_B4-for
<400> 68
aacaggtctc aggcttaaat ggctgcaaaa acggtggc 38
<210> 69
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> primer T7term-STOP-for
<400> 69
aacaggtctc atgacgccat taacctgatg ttctggg 37
<210> 70
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> linker region L10
<400> 70
Asn Ala Gly Leu Asn Gly Tyr Lys Asn Gly Gly Ser
1 5 10
<210> 71
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> C-terminus of B-domains
<400> 71
Val Asn Glu Leu Leu
1 5
<210> 72
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> C-terminus of B-domains
<400> 72
Val Asn Lys Leu Leu
1 5
<210> 73
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> C-terminus of CW_7 motifs
<400> 73
Val Asn Glu Ile Leu
1 5
<210> 74
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> L1
<400> 74
Asn Ala Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
1 5 10
<210> 75
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> L2
<400> 75
Asn Val Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
1 5 10
<210> 76
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> L3
<400> 76
Asn Ala Gly Leu Asn Gly Tyr Gln Asn Gly Gly Ser
1 5 10
<210> 77
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> L4
<400> 77
Asn Val Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
1 5 10
<210> 78
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> L5
<400> 78
Asn Ala Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
1 5 10
<210> 79
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> L6
<400> 79
Asn Ala Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
1 5 10
<210> 80
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> L7
<400> 80
Asn Ala Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
1 5 10
<210> 81
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> L10
<400> 81
Asn Ala Gly Leu Asn Gly Tyr Lys Asn Gly Gly Ser
1 5 10
<210> 82
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> L11
<400> 82
Lys Ala Gly Leu Asn Gly Tyr Lys Asn Gly Gly Ser
1 5 10
<210> 83
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> L12
<400> 83
Asn Ala Gly Leu Asn Gly Tyr Gln Asn Gly Gly Ser
1 5 10
<210> 84
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL11_strain_JCP8017A
<400> 84
Met Ser Lys Arg Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly His Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Thr Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Ser Ser Ala Gly Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Ile Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Asp Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Thr Tyr Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Val Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asp Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Asp Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Asp
165 170 175
Gly Ile Ala Gly Arg Val Asp Met Asp Tyr Thr Tyr Val Asp Tyr Pro
180 185 190
Ser Val Ile Lys Lys Ala Gly Leu Asn Gly Tyr Lys Asn Gly Gly Ser
195 200 205
Tyr Thr Ala Pro Gln Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Asp Trp Gly Asn Gly Asn Glu Arg Lys Asn Arg Leu Thr
225 230 235 240
Ser Ala Gly Tyr Asp Tyr Thr Ser Val Gln Asn Lys Val Asn Glu Leu
245 250 255
Leu Gly Val Lys Ala Tyr Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Thr Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu
275 280 285
Thr Gln Ala Gly Tyr Asp Tyr Asp Ala Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 85
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL3_strain_Gv18-4
<400> 85
Met Ser Lys Lys Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ser Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly His Lys Asp Lys Trp Phe Glu Glu Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ser Tyr Trp Tyr Ser Tyr
50 55 60
Ala Ser Ser Ala Gly Glu Ala Ser Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Ile Leu Ser Gly Lys Ser Phe Glu Tyr Pro Ile Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Asp Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Val Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asp Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Asn
165 170 175
Gly Ile Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Val Ile Lys Asn Ala Gly Leu Asn Gly Tyr Gln Asn Gly Gly Ser
195 200 205
Tyr Thr Ala Pro Gln Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Asp Trp Gly Asn Gly Asn Asp Arg Lys Asn Arg Leu Ile
225 230 235 240
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Glu Leu
245 250 255
Leu Gly Val Lys Ala Tyr Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Thr Trp Gly Asn Gly Ser Met Arg Lys His Arg Leu
275 280 285
Thr Gln Ala Gly Tyr Asp Tyr Asp Ala Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 86
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL10_strain_G30-4
<400> 86
Met Ser Lys Arg Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly His Lys Asp Lys Trp Phe Glu Glu Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Val Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Ser Ser Ala Gly Glu Ala Ser Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Ile Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Asp Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Val Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asp Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Asn
165 170 175
Gly Ile Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Val Ile Lys Asn Ala Gly Leu Asn Gly Tyr Lys Asn Gly Gly Ser
195 200 205
Tyr Thr Ala Pro Gln Ile Ser Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Asp Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
225 230 235 240
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Glu Leu
245 250 255
Leu Gly Val Lys Ala Tyr Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Thr Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu
275 280 285
Thr Gln Ala Gly Tyr Asp Tyr Asn Ala Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 87
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL12_strain_3549624_non-prophage
<400> 87
Met Ser Lys Lys Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly His Lys Asp Lys Trp Phe Glu Glu Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ser Tyr Trp Tyr Ser Tyr
50 55 60
Ala Ser Ser Ala Gly Glu Val Ala Leu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Ile Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Asp Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Val Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asp Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Asn
165 170 175
Gly Ile Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Val Ile Lys Asn Ala Gly Leu Asn Gly Tyr Gln Asn Gly Gly Ser
195 200 205
Tyr Thr Ala Pro Gln Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Asp Trp Gly Asn Gly Ile Glu Arg Lys Asn Arg Leu Thr
225 230 235 240
Ser Ala Gly Tyr Asp Tyr Thr Ser Val Gln Asn Lys Val Asn Glu Leu
245 250 255
Leu Gly Val Lys Ala Tyr Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Thr Trp Gly Asn Gly Lys Thr Arg Lys Gln Arg Leu
275 280 285
Thr Gln Ala Gly Tyr Asp Tyr Asn Ala Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 88
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL8_strain_JCP7719
<400> 88
Met Ser Lys Lys Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Ser Ser Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Ser Lys Leu Glu Thr Tyr Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Asn Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Ala Gly Leu Asn Gly Tyr Lys Asn Gly Gly Ser
195 200 205
Tyr Thr Ala Pro Gln Thr Ser Ser Ile Asp Asp Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
225 230 235 240
Gln Ala Gly Tyr Asp Tyr Thr Ser Val Gln Asn Lys Val Asn Lys Leu
245 250 255
Leu Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Thr Trp Gly Asn Gly Asn Glu Arg Lys Asn Arg Leu
275 280 285
Thr Gln Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 89
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL5_strain_JCP7276
<400> 89
Met Ser Lys Lys Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Ser Ser Glu Ala Ala Glu Glu Ala Gln Ser Cys Ala Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Gly Phe Cys Ser Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ala His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Asn Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Lys Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Ala Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
195 200 205
Asp Gln Ala Ala Arg Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
225 230 235 240
Gln Ala Gly Tyr Asp Tyr Thr Ser Val Gln Asn Lys Val Asn Lys Leu
245 250 255
Leu Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Thr Trp Gly Asn Gly Asn Glu Arg Lys Asn Arg Leu
275 280 285
Thr Gln Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 90
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL7_strain_AMD
<400> 90
Met Ser Lys Lys Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Ala Ser Glu Ala Ala Glu Glu Ala Gln Ser Cys Ala Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Ser Lys Leu Glu Thr Tyr Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Lys Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Ala Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
195 200 205
Asp Gln Ala Ala Arg Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
225 230 235 240
Gln Ala Gly Tyr Asp Tyr Thr Ser Val Gln Asn Lys Val Asn Lys Leu
245 250 255
Leu Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Thr Trp Gly Asn Gly Asn Glu Arg Lys Asn Arg Leu
275 280 285
Thr Gln Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 91
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL4_strain_Gv5-1
<400> 91
Met Ser Lys Lys Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Cys Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Ser Lys Leu Glu Thr Tyr Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Val Val Asn Asn Leu Val Ser Ala His Val Arg
130 135 140
Asp Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Val Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
195 200 205
Asp Gln Ala Thr Arg Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
225 230 235 240
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Lys Leu
245 250 255
Leu Gly Val Lys Ala Tyr Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Thr Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu
275 280 285
Ala Gln Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 92
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> EL6_strain_1400E_frameshift
<400> 92
Met Ser Lys Lys Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Cys Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Ser Lys Leu Glu Ser Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ala His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Asp Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Lys Asn Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Ala Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
195 200 205
Asn Gln Ala Ala Arg Thr Ser Ser Ile Asp Asp Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
225 230 235 240
Gln Ala Gly Tyr Asp Tyr Ala Ser Val Ala Lys
245 250
<210> 93
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL1_strain_HMP9231
<400> 93
Met Ser Lys Lys Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Leu Met Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ala His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Asn Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Asn Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Ala Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
195 200 205
Asp Gln Ala Ala Arg Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu Thr
225 230 235 240
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Glu Leu
245 250 255
Leu Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Ala Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu
275 280 285
Ala Gln Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 94
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> EL9_strain_0288E_incomplete_contig
<400> 94
Met Ser Lys Lys Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Cys Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Val Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ser His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Asp Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Ala Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
195 200 205
Asp Gln Ala Ala Arg Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu Thr
225 230 235 240
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Ala Lys
245 250
<210> 95
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL2_strain_Gv18-4
<400> 95
Met Ser Lys Arg Gly Ile Asp Val Ser Glu Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Cys Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Gly Phe Cys Asn Lys Leu Glu Ser Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ala His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Val Gly Leu Asn Gly Cys Lys Asn Gly Gly Ser
195 200 205
Asp Gln Ala Ala Arg Thr Ser Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
225 230 235 240
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Glu Leu
245 250 255
Leu Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Ile Ala Arg Glu
260 265 270
Val Ile Arg Gly Thr Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu
275 280 285
Thr Gln Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 96
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL13_strain_Gv37_1
<400> 96
Met Ser Lys Lys Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Glu Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Ala Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ser Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Pro Ala His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Asp Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Ala Gly Leu Asn Gly Tyr Lys Asn Gly Glu Ser
195 200 205
His Gln Ala Thr Arg Thr Thr Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
225 230 235 240
Gln Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Glu Leu
245 250 255
Leu Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Thr Trp Gly Asn Gly Asn Glu Arg Lys Asn Arg Leu
275 280 285
Thr Ser Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<210> 97
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<223> EL14_strain_Gv37_2
<400> 97
Met Ser Lys Lys Gly Ile Asp Val Ser Val Trp Gln Gly Asp Ile Asp
1 5 10 15
Phe Asn Ala Val Lys Ala Ser Gly Val Glu Phe Val Ile Ile Arg Ala
20 25 30
Gly Tyr Gly Ile Gly Cys Lys Asp Lys Trp Phe Glu Gln Asn Tyr Arg
35 40 45
Lys Ala Lys Thr Cys Gly Leu Asp Val Gly Ala Tyr Trp Tyr Ser Tyr
50 55 60
Ala Asn Ser Gly Phe Glu Ala Ala Glu Glu Ala Gln Ser Cys Val Asn
65 70 75 80
Met Leu Ser Gly Lys Ser Phe Glu Tyr Pro Val Tyr Phe Asp Leu Glu
85 90 95
Glu Lys Ser Gln Leu Asn Arg Gly Arg Ala Phe Cys Asp Ser Leu Ile
100 105 110
Thr Ser Phe Cys Asn Lys Leu Glu Ala Cys Gly Tyr Tyr Ala Gly Phe
115 120 125
Tyr Thr Ser Leu Ser Thr Ala Asn Asn Leu Val Ser Ala His Val Arg
130 135 140
Asn Arg Tyr Ala Leu Trp Ile Ala Gln Trp Asn Thr His Cys Ser Tyr
145 150 155 160
Gln Gly Ser Tyr Gly Leu Trp Gln Tyr Ser Ser Ser Gly Ser Val Pro
165 170 175
Gly Val Ala Gly Arg Val Asp Met Asp Tyr Ala Tyr Val Asp Tyr Pro
180 185 190
Ser Ile Ile Lys Asn Ala Gly Leu Asn Gly Tyr Lys Asn Gly Glu Ser
195 200 205
His Gln Ala Thr Arg Thr Thr Ser Ile Asp Glu Val Ala Arg Glu Val
210 215 220
Ile Asn Gly Ala Trp Gly Asn Gly Asn Glu Arg Lys Gln Arg Leu Thr
225 230 235 240
Ser Ala Gly Tyr Asp Tyr Ala Ser Val Gln Asn Lys Val Asn Glu Leu
245 250 255
Leu Gly Val Lys Ala Cys Arg Lys Ser Val Asp Glu Leu Ala Arg Glu
260 265 270
Val Ile Arg Gly Ala Trp Gly Asn Gly Ser Thr Arg Lys Gln Arg Leu
275 280 285
Thr Ser Ala Gly Tyr Asp Tyr Asp Thr Val Gln Lys Arg Val Asn Glu
290 295 300
Leu Leu
305
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-CLL1-СПЕЦИФИЧЕСКИЕ ОДНОЦЕПОЧЕЧНЫЕ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ (scCAR) ДЛЯ ИММУНОТЕРАПИИ РАКА | 2016 |
|
RU2731543C2 |
| СВЯЗЫВАЮЩИЕСЯ С CD38 И PD-L1 МОЛЕКУЛЫ | 2017 |
|
RU2764201C2 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ ДЛЯ ЛЕЧЕНИЯ ВИРУСНОЙ ИНФЕКЦИИ ГЕПАТИТА В И СВЯЗАННЫХ СОСТОЯНИЙ | 2016 |
|
RU2733496C2 |
| Т-КЛЕТКИ С ДВУМЯ ХИМЕРНЫМИ АНТИГЕННЫМИ РЕЦЕПТОРАМИ CAR | 2021 |
|
RU2839662C1 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С TFR | 2019 |
|
RU2810756C2 |
| УЛУЧШЕННЫЕ ВАРИАБЕЛЬНЫЕ ДОМЕНЫ ИММУНОГЛОБУЛИНА | 2015 |
|
RU2746738C2 |
| БИСПЕЦИФИЧЕСКИЕ ТРЕХВАЛЕНТНЫЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С КЛАУДИНОМ 6 ИЛИ КЛАУДИНОМ 18.2, И CD3, ДЛЯ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ С ЭКСПРЕССИЕЙ КЛАУДИНА | 2017 |
|
RU2798988C2 |
| АНТИ-PVRIG АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2732042C2 |
| СВЯЗЫВАЮЩИЙ TNF-α ПОЛИПЕПТИД И СПОСОБ ЛЕЧЕНИЯ СВЯЗАННЫХ С TNF-α ЗАБОЛЕВАНИЙ | 2017 |
|
RU2794974C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) С АНТИГЕНСВЯЗЫВАЮЩИМИ ДОМЕНАМИ К КОНСТАНТНОЙ ОБЛАСТИ β Т-КЛЕТОЧНОГО РЕЦЕПТОРА | 2015 |
|
RU2830064C2 |





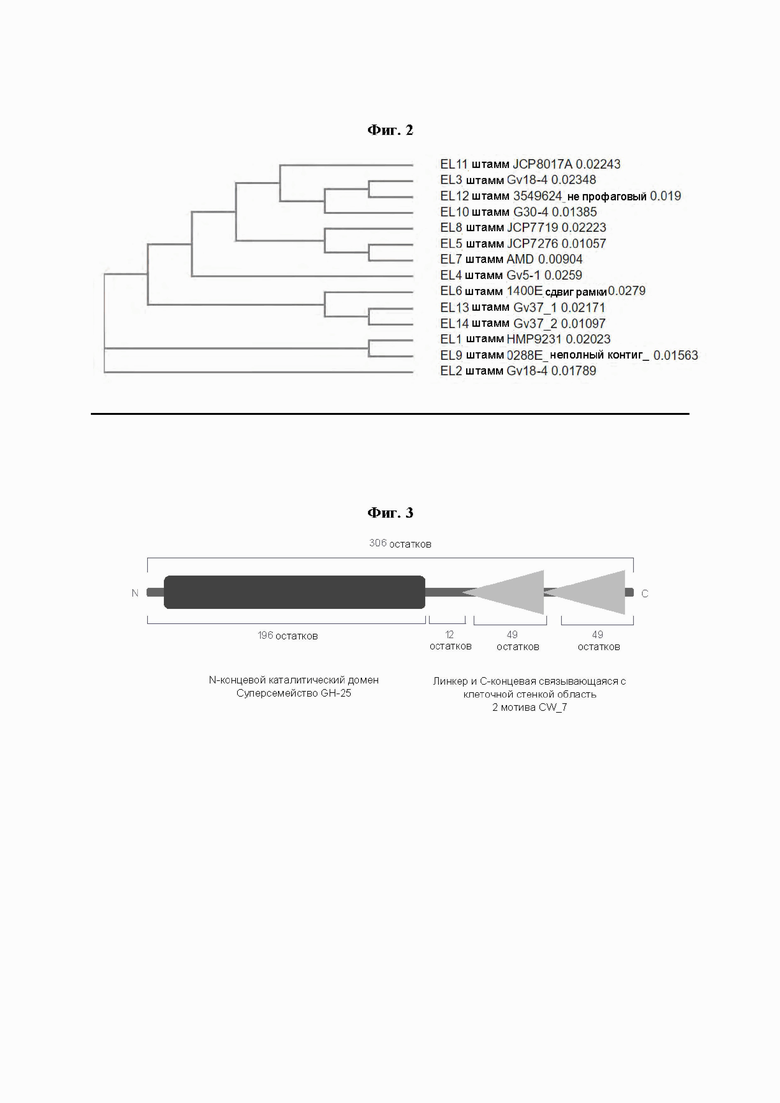
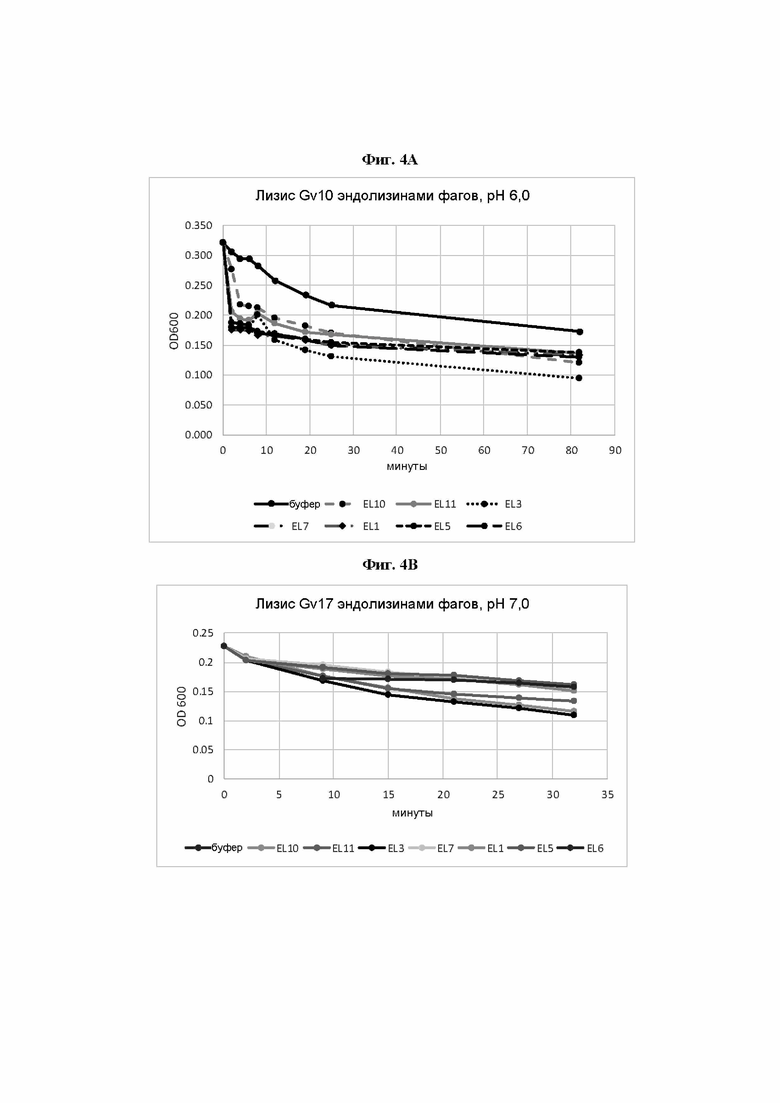
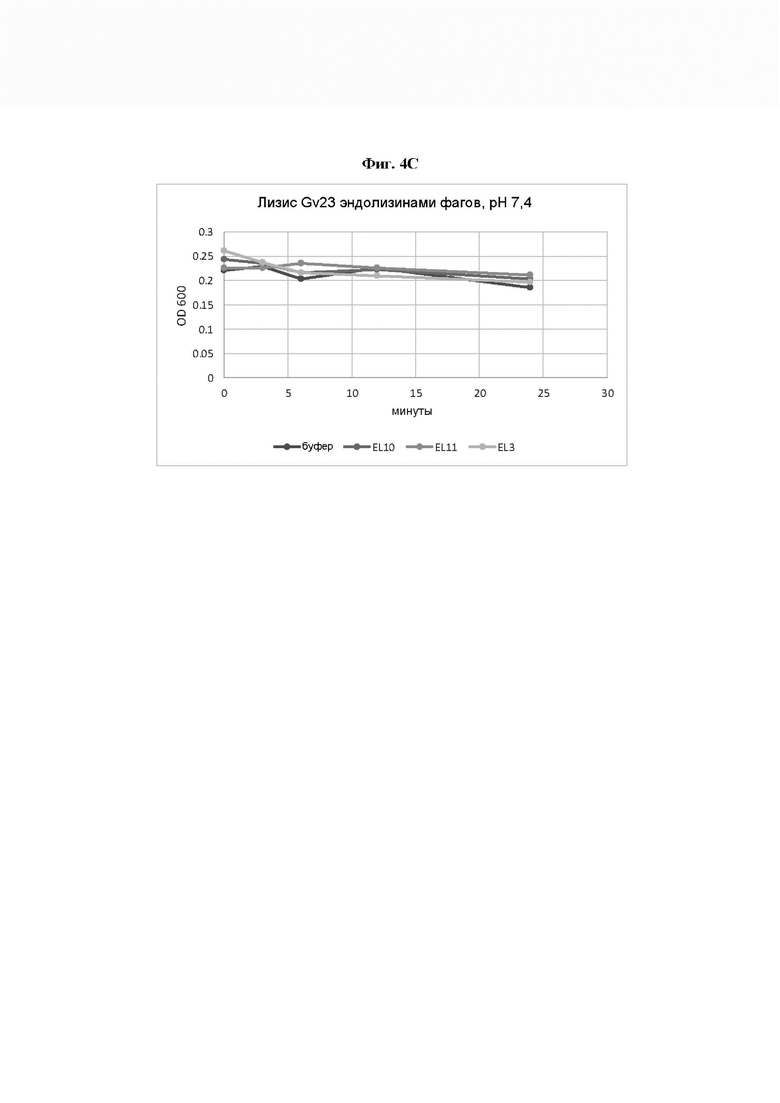

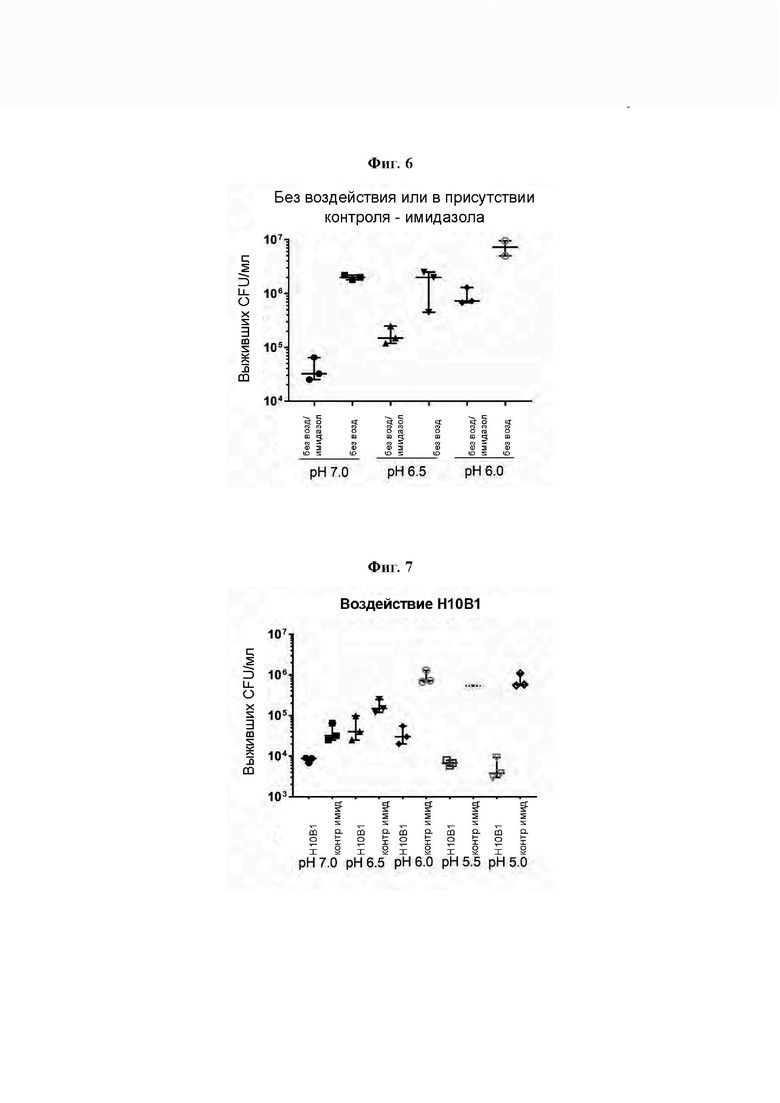
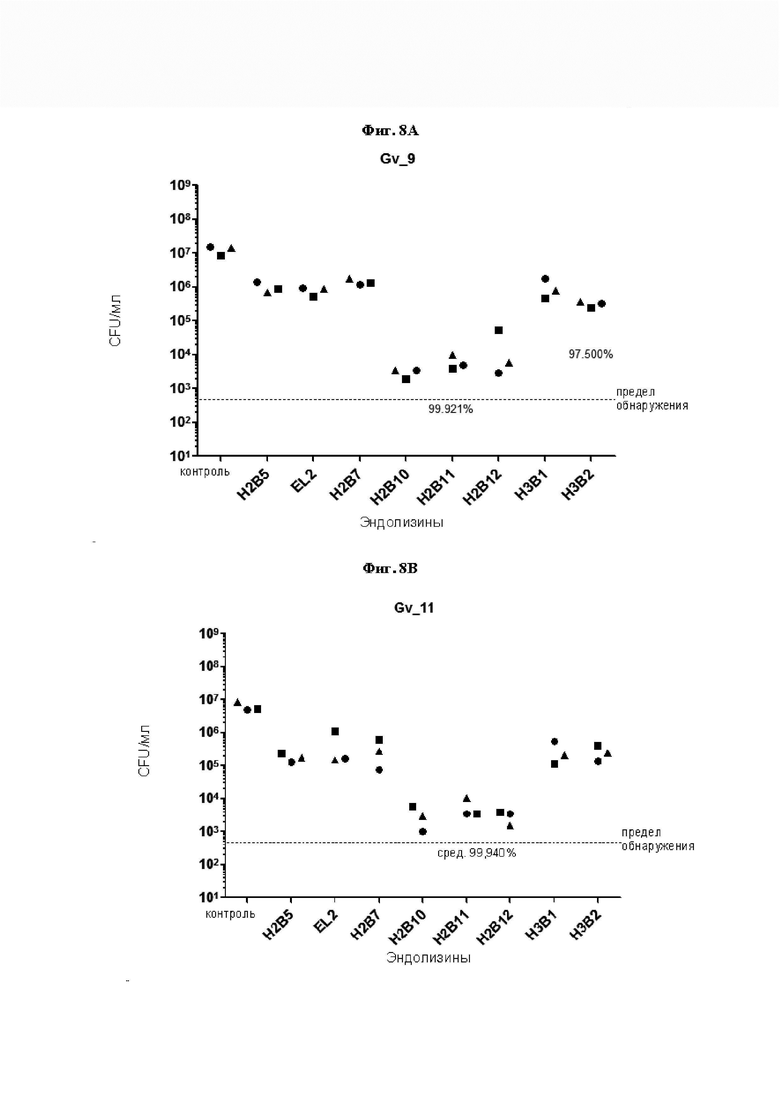
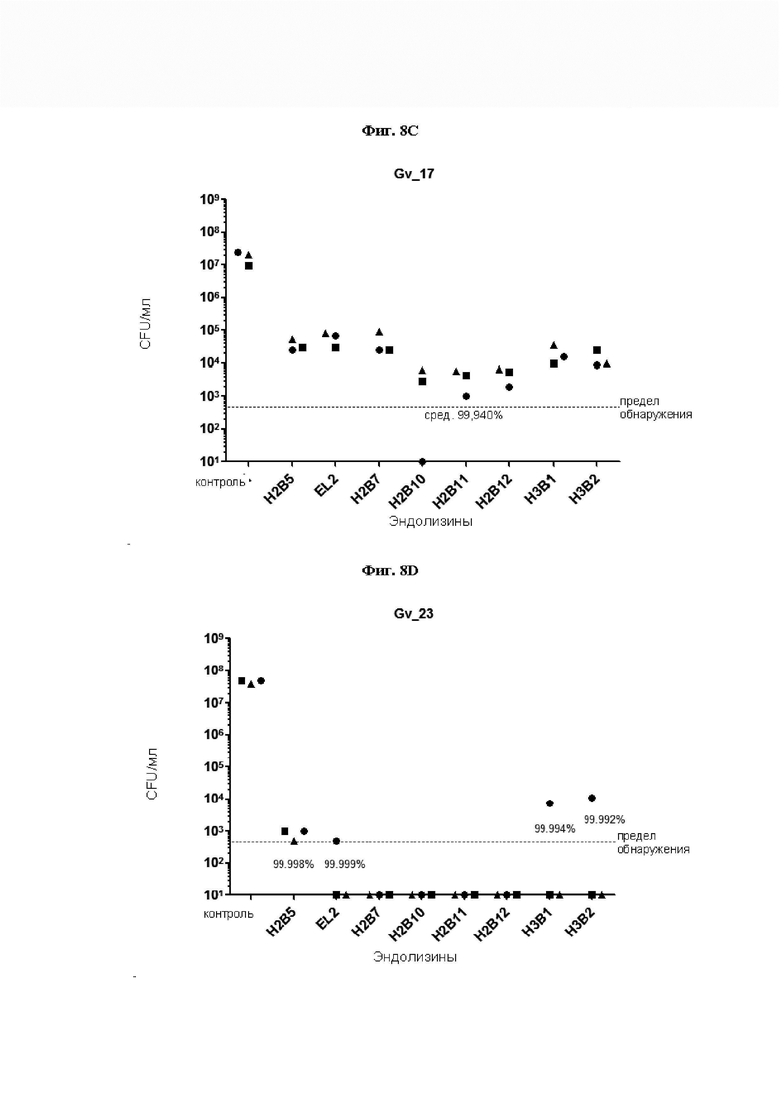
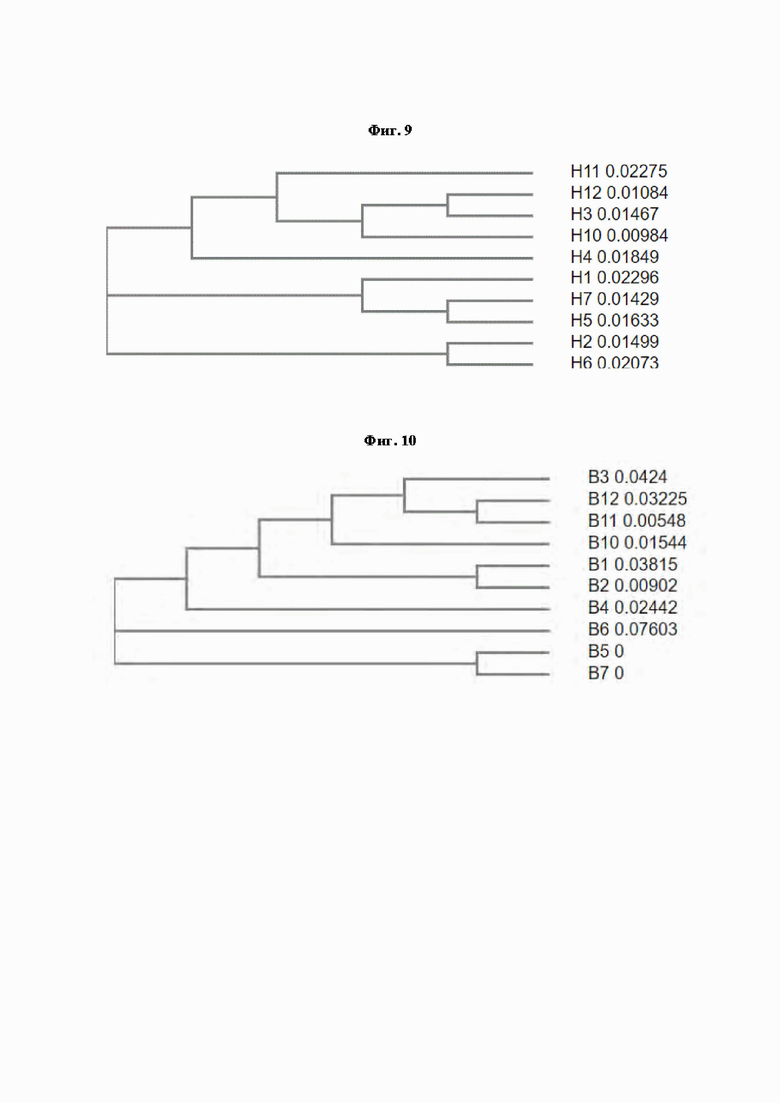

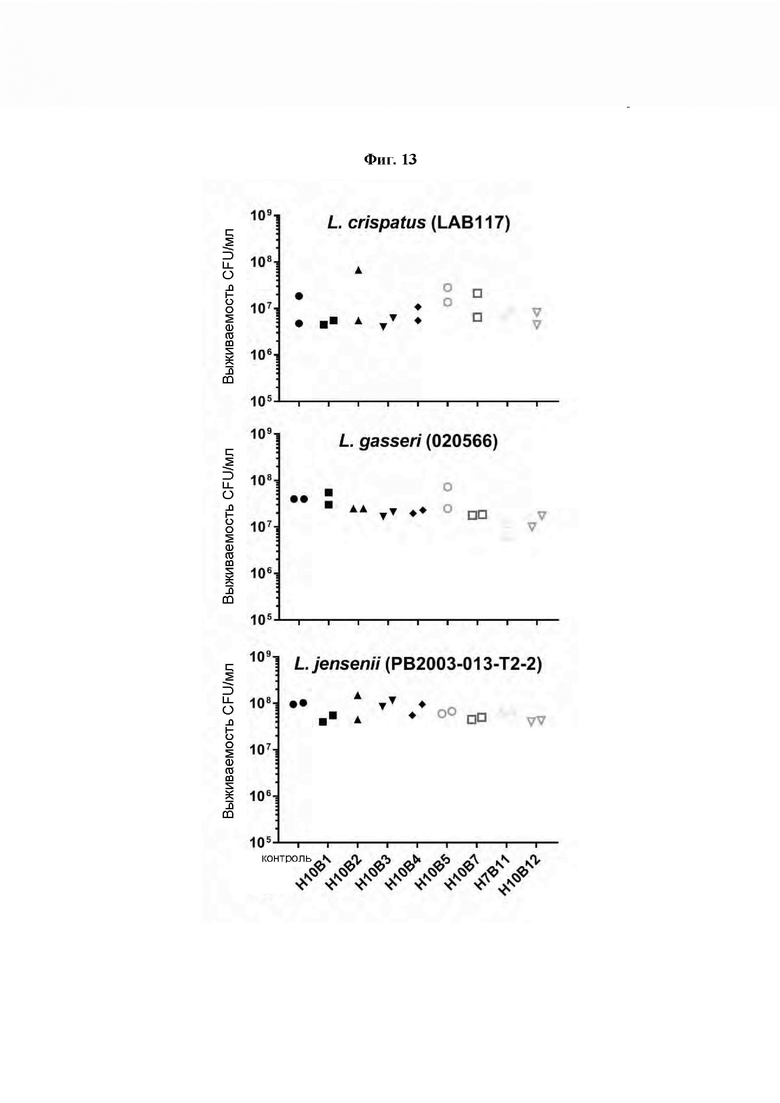
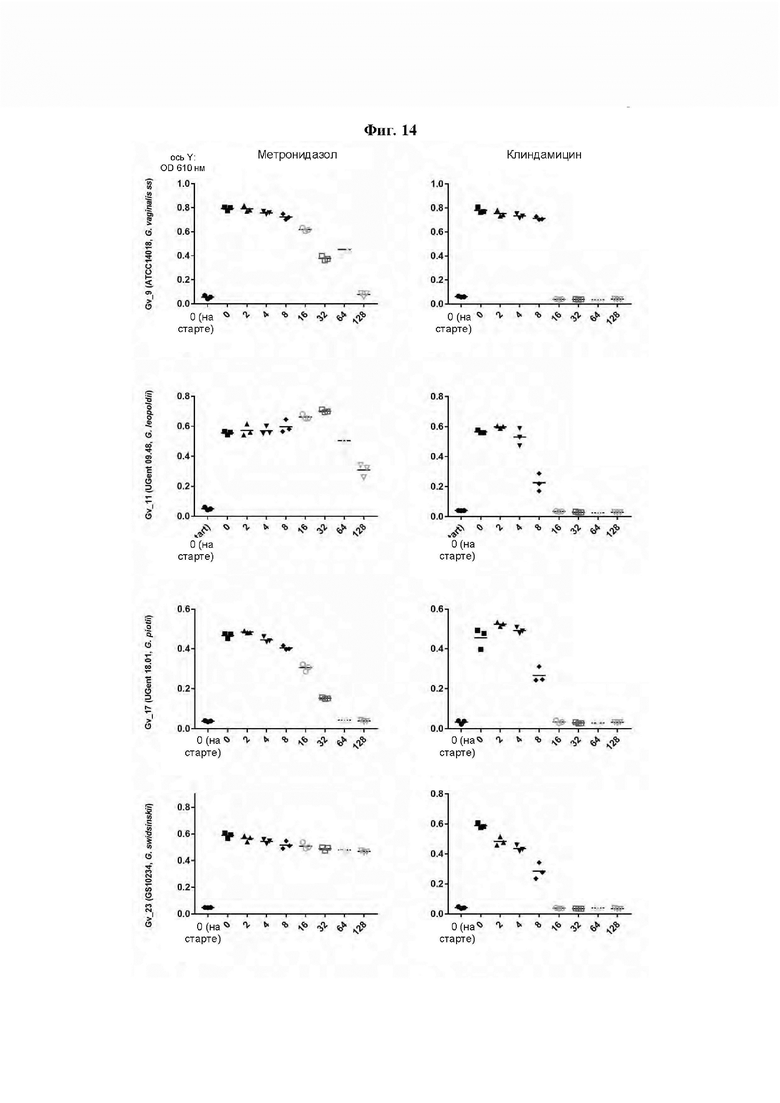
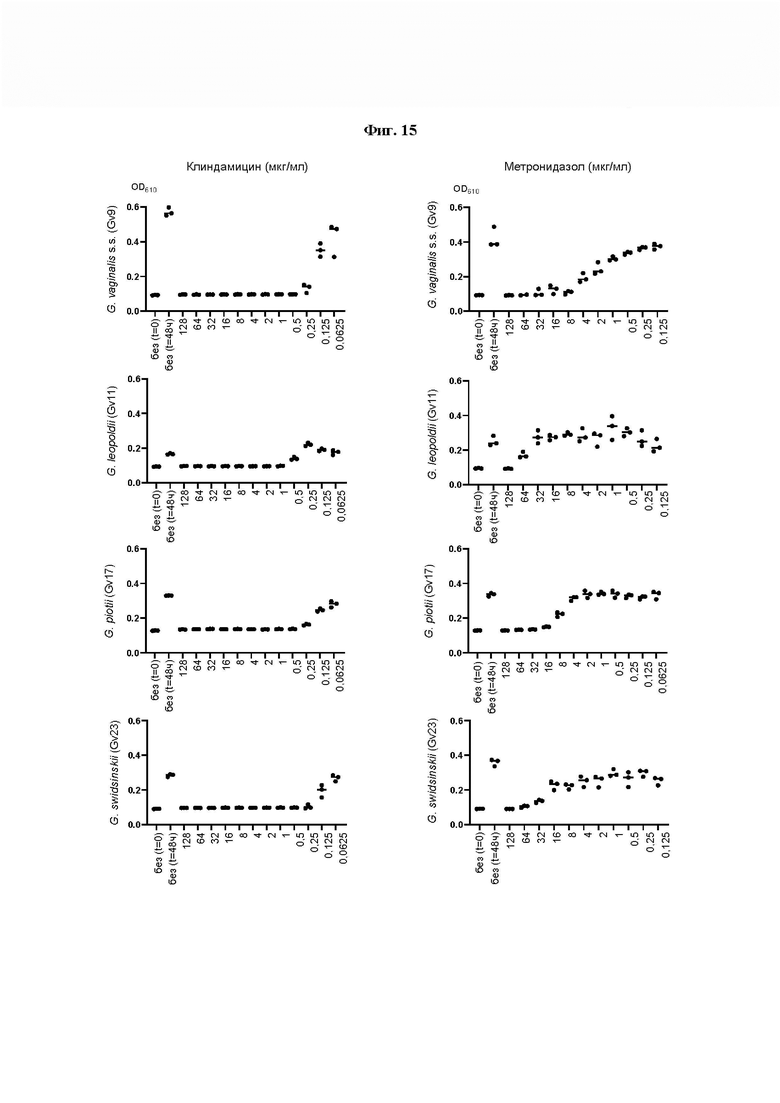
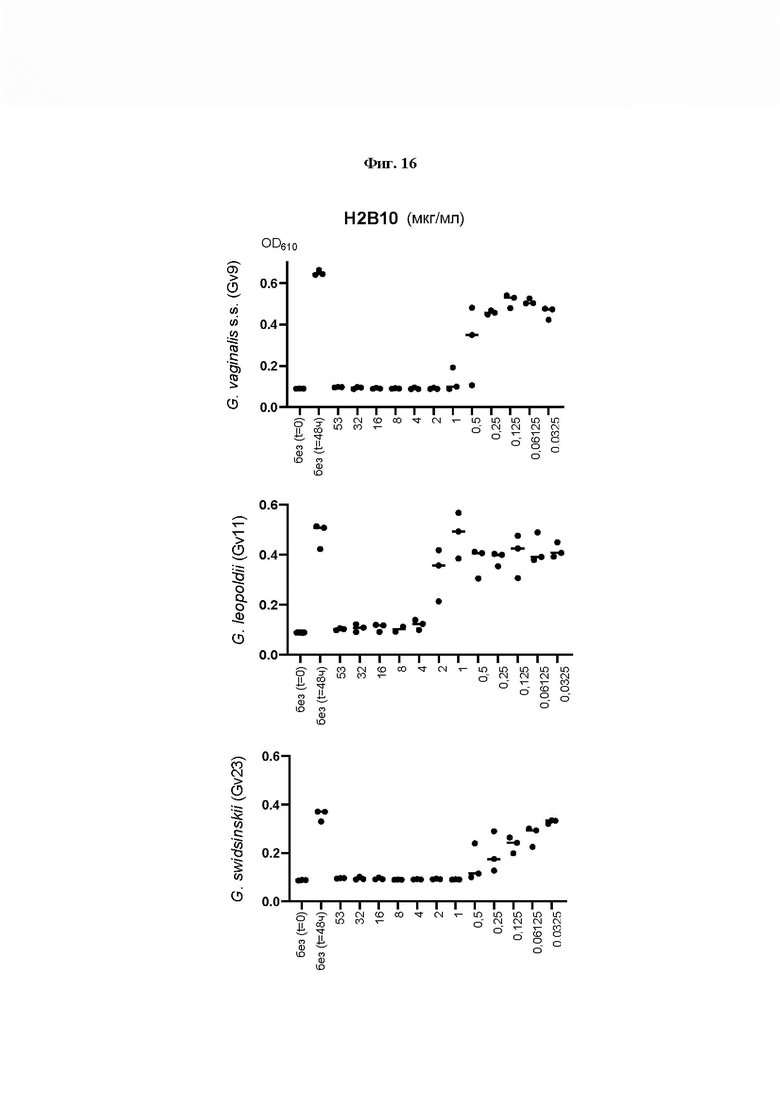
Настоящее изобретение относится к области биотехнологии, в частности к новым селективным в отношении вида эндолизинам фагов и их применению для лечения бактериального вагиноза (BV). В настоящем изобретении предложены рекомбинантные эндолизины, то есть эндолизины с замененными доменами. Изобретение также относится к указанным эндолизинам для применения в лечении заболеваний или расстройств, таких как бактериальные инфекции, в частности BV. Изобретение также относится к полинуклеотидам, кодирующим указанные эндолизины. Указанные полинуклеотиды также можно применять для лечения таких заболеваний или расстройств. В настоящем изобретении также предложена фармацевтическая композиция, содержащая эндолизин по настоящему изобретению для применения в лечении таких заболеваний или расстройств. Указанные эндолизины, полинуклеотиды и фармацевтическую композицию можно вводить местно, в частности местно во влагалище. Изобретение обеспечивает способы лечения, деконтаминации или обнаружения бактериальных инфекций и расстройств, связанных с Gardnerella. 7 н. и 20 з.п. ф-лы, 16 ил., 7 табл., 7 пр.
1. Рекомбинантный эндолизин, содержащий или состоящий из
(1) N-концевого каталитического домена,
(2) С-концевой связывающейся с клеточной стенкой области, где С-концевая связывающаяся с клеточной стенкой область содержит или состоит из одного, двух или трех связывающихся с клеточной стенкой доменов, и
(3) линкерной области между N-концевым каталитическим доменом и С-концевой связывающейся с клеточной стенкой областью;
где N-концевой каталитический домен имеет происхождение из первого естественного эндолизина С-концевая связывающаяся с клеточной стенкой область и возможно линкерная область имеют происхождение из второго естественного эндолизина, и где первый и второй естественные эндолизины закодированы разными геномами разных профагов,
где N-концевой каталитический домен представляет собой полипептид, содержащий или состоящий из аминокислотной последовательности любой из SEQ ID NO: 1-5, 7 или 10-12 или любого ее варианта, идентичного по меньшей мере на 80% аминокислотной последовательности любой из SEQ ID NO: 1-5, 7 или 10-12, где указанный полипептид или указанный его вариант является функциональным, где функция включает способность лизировать клеточную стенку Gardnerella.
где один, два или три связывающихся с клеточной стенкой домена независимо выбраны из группы, состоящей из полипептидов, содержащих или состоящих из аминокислотной последовательности SEQ ID NO: 15-24 и 26-33, соответственно, и любых их вариантов, идентичных по меньшей мере на 80% аминокислотной последовательности SEQ ID NO: 15-24 и 26-33, соответственно, где указанные полипептиды или указанные их варианты являются функциональными, где функция включает способность связываться с клеточной стенкой Gardnerella; и где указанный рекомбинантный эндолизин обладает уничтожающей активностью в отношении Gardnerella.
2. Рекомбинантный эндолизин по п. 1, где N-концевой каталитический домен представляет собой полипептид, содержащий или состоящий из аминокислотной последовательности SEQ ID NO: 2.
3. Рекомбинантный эндолизин по п. 1 или 2, где один, два или три связывающихся с клеточной стенкой домена независимо выбраны из группы, состоящей из полипептидов, содержащих или состоящих из аминокислотной последовательности SEQ ID NO: 28-33.
4. Рекомбинантный эндолизин по п. 1, где N-концевой каталитический домен представляет собой полипептид, содержащий аминокислотную последовательность SEQ ID NO: 7 или состоящий из нее, и где один, два или три связывающихся с клеточной стенкой домена независимо выбраны из группы, состоящей из полипептидов, содержащих или состоящих из аминокислотной последовательности SEQ ID NO: 19 и 20.
5. Рекомбинантный эндолизин по любому из пп. 1, 2, где С-концевая связывающаяся с клеточной стенкой область содержит или состоит из первого связывающегося с клеточной стенкой домена и второго связывающегося с клеточной стенкой домена, где указанный первый связывающийся с клеточной стенкой домен выбран из группы, состоящей из SEQ ID NO: 15, 17, 19, 21, 23, 26, 28, 30 и 32, а второй связывающийся с клеточной стенкой домен выбран из группы, состоящей из SEQ ID NO: 16, 18, 20, 22, 24, 27, 29, 31 и 33.
6. Рекомбинантный эндолизин по п. 1, где N-концевой каталитический домен представляет собой полипептид, содержащий или состоящий из аминокислотной последовательности SEQ ID NO: 2, и С-концевая связывающаяся с клеточной стенкой область содержит или состоит из первого связывающегося с клеточной стенкой домена и второго связывающегося с клеточной стенкой домена, причем указанный первый связывающийся с клеточной стенкой домен выбран из группы, состоящей из полипептидов, содержащих или состоящих из аминокислотной последовательности, имеющей по меньшей мере 90% идентичности с любой из аминокислотных последовательностей SEQ ID NO: 28, 30 и 32, и указанный второй связывающийся с клеточной стенкой домен выбран из группы, состоящей из полипептидов, содержащих или состоящих из аминокислотной последовательности, имеющей по меньшей мере 95% идентичность с любой аминокислотной последовательностью из SEQ ID NO: 29, 31 и 33, причем указанные первый и второй связывающиеся с клеточной стенкой домены являются функциональными, и эта функция включает способность связываться с клеточной стенкой Gardnerella.
7. Рекомбинантный эндолизин по п. 1, где N-концевой каталитический домен представляет собой полипептид, содержащий или состоящий из аминокислотной последовательности SEQ ID NO: 2, и С-концевая связывающаяся с клеточной стенкой область содержит или состоит из первого связывающегося с клеточной стенкой домена и второго связывающегося с клеточной стенкой домена, причем указанный первый связывающийся с клеточной стенкой домен выбран из группы, состоящей из SEQ ID NO: 28, 30 и 32, и указанный второй связывающийся с клеточной стенкой домен выбран из группы, состоящей из SEQ ID NO: 29, 31 и 33.
8. Рекомбинантный эндолизин по п. 1, где N-концевой каталитический домен представляет собой полипептид, содержащий или состоящий из аминокислотной последовательности SEQ ID NO: 2, и С-концевая связывающаяся с клеточной стенкой область содержит или состоит из первого связывающегося с клеточной стенкой домена и второго связывающегося с клеточной стенкой домена, причем указанный первый связывающийся с клеточной стенкой домен определен в SEQ ID NO: 28, и указанный второй связывающийся с клеточной стенкой домен определен в SEQ ID NO: 29.
9. Рекомбинантный эндолизин по п. 1, где N-концевой каталитический домен представляет собой полипептид, содержащий или состоящий из аминокислотной последовательности SEQ ID NO: 2, и С-концевая связывающаяся с клеточной стенкой область содержит или состоит из первого связывающегося с клеточной стенкой домена и второго связывающегося с клеточной стенкой домена, причем указанный первый связывающийся с клеточной стенкой домен определен в SEQ ID NO: 28, и указанный второй связывающийся с клеточной стенкой домен определен в SEQ ID NO: 31.
10. Рекомбинантный эндолизин по любому из пп. 5-9, где указанный первый связывающийся с клеточной стенкой домен расположен в N-концевом направлении от указанного второго связывающегося с клеточной стенкой домена.
11. Рекомбинантный эндолизин по любому из пп. 1-10, где линкерная область представляет собой полипептид, содержащий или состоящий из аминокислотной последовательности:
(1) (ХХХ)n, где каждый X может независимо представлять собой G, А или S, предпочтительно где аминокислотная последовательность (ХХХ)n представляет собой (GGS)n, где n соответствует числу повторов последовательности XXX, предпочтительно где n представляет собой 2, 3, 4, 5 или 6; или
(2) X1X2GLNGX3X4NGGS, где X1 представляет собой N или K, Х2 представляет собой А или V, Х3 представляет собой Y или С и Х4 представляет собой K или Q.
12. Рекомбинантный эндолизин по любому из пп. 1-11, где указанный эндолизин обладает избирательной уничтожающей активностью в отношении рода Gardnerella.
13. Рекомбинантный эндолизин по любому из пп. 1-12, где указанный эндолизин обладает уничтожающей активностью в отношении Gardnerella vaginalis sensu stricto, Gardnerella leopoldii, Gardnerella piotii и/или Gardnerella swidsinskii.
14. Рекомбинантный эндолизин по любому из пп. 1-13, где указанный эндолизин не обладает уничтожающей активностью в отношении Lactobacilli.
15. Рекомбинантный эндолизин по любому из пп. 1-14, где указанный эндолизин не обладает уничтожающей активностью в отношении Lactobacilli crispatus, Lactobacilli gasseri и/или Lactobacilli jensenii.
16. Полинуклеотид, кодирующий рекомбинантный эндолизин по любому из пп. 1-15.
17. Фармацевтическая композиция, обладающая уничтожающей активностью в отношении Gardnerella, содержащая рекомбинантный эндолизин по любому из пп. 1-15 или полинуклеотид по п. 16 и дополнительно содержащая фармацевтически приемлемый носитель и/или разбавитель.
18. Применение рекомбинантного эндолизина по любому из пп. 1-15, полинуклеотида по п. 16 или фармацевтической композиции по п. 17 в лечении бактериальной инфекции, вызванной по меньшей мере одним штаммом рода Gardnerella.
19. Применение по п. 18, где указанная бактериальная инфекция представляет собой бактериальный вагиноз.
20. Применение по п. 19, где указанный бактериальный вагиноз вызван Gardnerella vaginalis sensu stricto, Gardnerella leopoldii, Gardnerella piotii и/или Gardnerella swidsinskii.
21. Применение по любому из пп. 18-20, где указанный рекомбинантный эндолизин, полинуклеотид или фармацевтическую композицию следует вводить местно, предпочтительно где указанный рекомбинантный эндолизин, полинуклеотид или фармацевтическую композицию следует вводить местно во влагалище субъекта женского пола и/или в или на головку полового члена, крайнюю плоть или вход в уретру субъекта мужского пола.
22. Применение по любому из пп. 18-20, где указанный рекомбинантный эндолизин, полинуклеотид или фармацевтическую композицию следует вводить во влагалище субъекта женского пола и/или в или на головку полового члена, крайнюю плоть или вход в уретру субъекта мужского пола.
23. Применение по любому из пп. 18-22, где указанный рекомбинантный эндолизин, полинуклеотид или фармацевтическую композицию следует вводить совместно с соединением или композицией, доводящими рН во влагалище до 4,0-6,0.
24. Плазмида для получения эндолизина, обладающего уничтожающей активностью в отношении Gardnerella, содержащая полинуклеотид по п. 16.
25. Бактериальная клетка-хозяин для получения эндолизина, обладающего уничтожающей активностью в отношении Gardnerella, содержащая плазмиду по п. 24.
26. Бактериальная клетка-хозяин по п. 25, представляющая собой клетку Е. coli.
27. Способ in vitro диагностирования заболевания или состояния, которое можно лечить эндолизином по любому из пп. 1-15, включающий стадии:
(1) приведения полученного от субъекта образца в контакт с полипептидом, содержащим или состоящим из С-концевой связывающейся с клеточной стенкой области эндолизина по любому из пп. 1-15 и возможно N-концевого каталитического домена эндолизина по любому из пп. 1-15, где образец содержит микробные клетки, и где С-концевая связывающаяся с клеточной стенкой область указанного эндолизина возможно является меченой;
(2) тестирования, связывается ли полипептид с микробными клетками образца и/или лизирует ли их; и
(3) установления того, что указанное заболевание или состояние можно лечить эндолизином по любому из пп. 1-15, если полипептид связывается с микробными клетками и/или лизирует их.
| SCHMELCHER M | |||
| et al., Bacteriophage endolysins as novel antimicrobials, Future Microbiol., 2012, Volume 7 (10), pp | |||
| ШАХТНАЯ ТОПКА ДЛЯ ДРОВ С НАКЛОННЫМИ И ГОРИЗОНТАЛЬНЫМИ КОЛОСНИКАМИ | 1919 |
|
SU1147A1 |
| АНТОНОВА Н.П | |||
| и др | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |
| WO 2018185634 | |||

