ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] По настоящей заявке испрашивается приоритет по временной заявке США № 62/861901, поданной 14 июня 2019 года, описание которой, таким образом, включено в настоящее описание в качестве ссылки в полном объеме.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка содержит список последовательностей, поданный в электронном виде в формате ASCII и включенный, таким образом, в настоящее описание в качестве ссылки в полном объеме. Указанная копия ASCII, созданная 10 июня 2020 года, названа 36271-808_601_SL.txt и имеет размер 18162 байт.
ЗАЯВЛЕНИЕ ОБ ИССЛЕДОВАНИИ, СПОНСИРУЕМОМ ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА
[0003] Представленное в настоящем описании изобретение создано, по меньшей мере частично, при поддержке правительства США по грантам №№ 5R35 GM118178 и GM128376 и F31 GM128376 National Institutes of Health (NIH) и NSF/DGE-1346837 National Science Foundation (NSF). Таким образом, правительство США обладает некоторыми правами на изобретение.
УРОВЕНЬ ТЕХНИКИ
[0004] Биологическое разнообразие позволяет формам жизни адаптироваться к разной среде и с течением времени эволюционировать в новые формы и функции. Источником этого разнообразия является изменчивость в белковых последовательностях, обеспечиваемая двадцатью природными аминокислотами, изменчивость, закодированная в геноме организма четырьмя природными нуклеотидами ДНК. Хотя функциональное разнообразие, обеспечиваемое природными аминокислотами, может быть высоким, ширина пространства последовательности значительно ограничивает то, что, по сути, можно исследовать, и кроме того, некоторая функциональность просто недоступна. Использование кофакторов в природе для гидридного переноса, окислительно-восстановительной активности и образования электрофильных связей и т.д. является примером этих ограничений. Кроме того, с ростом внимания к разработке белков в качестве терапевтических средств эти ограничения становятся проблемой, т.к. физико-химическое разнообразие природных аминокислот значительно ограничено по сравнению с низкомолекулярными лекарственными средствами, созданными химиками. В принципе, должна существовать возможность обойти эти ограничения, расширив генетический код так, чтобы он включал дополнительные, неканонические аминокислоты (ncAA или "неприродные" аминокислоты) с желаемыми физико-химическими свойствами.
[0005] Почти 20 лет назад был создан способ повышения разнообразия, доступного для живых организмов, посредством расширения генетического кода с использованием янтарного стоп-кодона (UAG) для кодирования ncAA в Escherichia coli. Этого достигали с использованием пары тРНК-аминокислоты и тРНК-синтетазы (aaR) из Methanococcus jannaschii, в которой тРНК перекодирована для супрессии стоп-кодона, и aaRS переделана так, чтобы заряжать тРНК ncAA. С тех пор этот способ супрессии кодонов был расширен на другие стоп-кодоны и даже квадруплетные кодоны, а также до использования нескольких других ортогональных пар тРНК-aaRS (в целом, пара Pyl тРНК-синтетаза из Methanosarcina barkeri/mazei), что расширяет объем ncAA, которые можно включать в белки. Эти способы уже начали революционизировать химическую биологию и белковые терапевтические средства.
[0006] Хотя эти способы позволяют включать до двух разных ncAA в прокариотические и эукариотические клетки, гетерологичные перекодированные тРНК должны конкурировать с эндогенными факторами освобождения (RF) или, в случае квадруплетных кодонов, нормальным декодированием, ограничивающим эффективность и точность встраивания ncAA. Для устранения конкуренции с RF1, распознающим янтарный стоп-кодон и терминирующим трансляцию, усилия были направлены на удаление многих или всех копий янтарного стоп-кодона в геноме организма-хозяина или такую модификацию RF2, чтобы сделать возможной делецию RF1. Однако эукариоты имеют только один фактор освобождения, и, хотя его можно модифицировать, его нельзя подвергать делеции, и в случае прокариот делеция RF1 приводит к более высокой мис-супрессии янтарного стоп-кодона другими тРНК, что снижает точность встраивания ncAA. Попытки дальнейшего исследования избыточности кодонов для выделения природных кодонов для перенаправления на ncAA могут быть затруднены плейотропными эффектами, т.к. кодоны на самом деле не являются избыточными, например, из-за их эффектов в отношении скорости трансляции и фолдинга белка. Кроме того, перераспределение кодонов ограничено проблемами крупномасштабной геномной инженерии, особенно у эукариот.
[0007] Альтернативным подходом для перенаправления природных кодонов является создание совершенно новых кодонов, не имеющих природной функции или затруднения, и распознавание которых в рибосоме, по существу, является более ортогональным. Этого можно достигать посредством создания организмов, несущих пятый и шестой нуклеотид, образующий неприродную пару оснований (UBP). Такие полусинтетические организмы (SSO) должны будут точно реплицировать ДНК, содержащую UBP, эффективно транскрибировать ее в мРНК и тРНК, содержащие неприродные нуклеотиды, а затем эффективно декодировать неприродные кодоны с использованием когнатных неприродных антикодонов. Такие SSO будут иметь практически неограниченное количество новых кодонов для кодирования ncAA.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0008] В некоторых вариантах осуществления настоящее изобретение относится к способам, клеткам, сконструированным микроорганизмам, плазмидам и наборам для повышенной продукции молекулы нуклеиновой кислоты, содержащей неприродный нуклеотид.
[0009] Предусмотрены следующие варианты осуществления.
[0010] Вариант осуществления A1 представляет собой нуклеиновое основание структуры:
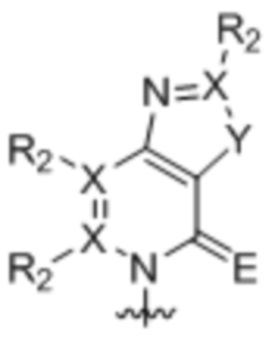 ,
,
где:
каждый X независимо представляет собой углерод или азот;
R2 присутствует, если X представляет собой углерод, и независимо представляет собой водород, алкил, алкенил, алкинил, метоксигруппу, метантиол, метанселеногруппу, галоген, цианогруппу или азидную группу;
Y является серой, кислородом, селеном или вторичным амином; и
E является кислородом, серой или селеном;
где волнистой линией указана точка связывания с остатком рибозила, дезоксирибозила или дидезоксирибозила или его аналогом, где остаток рибозила, дезоксирибозила или дидезоксирибозила или его аналог находится в свободной форме, соединен с монофосфатной, дифосфатной, трифосфатной, α-тиотрифосфатной, β-тиотрифосфатной или γ-тиотрифосфатной группой или включен в РНК, или ДНК, или аналог РНК, или аналог ДНК.
[0011] Вариант осуществления A2 является нуклеиновым основанием по варианту осуществления A1, где X является углеродом.
[0012] Вариант осуществления A3 является нуклеиновым основанием по варианту осуществления A1 или A2, где E является серой.
[0013] Вариант осуществления A4 является нуклеиновым основанием по любому из вариантов осуществления A1-A3, где Y является серой.
[0014] Вариант осуществления A5 является нуклеиновым основанием по варианту осуществления A1, имеющим структуру 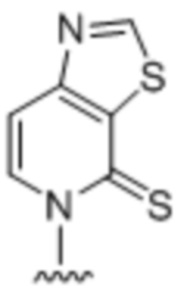 .
.
[0015] Вариант осуществления A6 является нуклеиновым основанием по любому из вариантов осуществления A1-A5, связанным с комплементарным, спаривающимся нуклеиновым основанием для образования неприродной пары оснований (UBP).
[0016] Вариант осуществления A7 является нуклеиновым основанием по варианту осуществления A6, где комплементарное, спаривающееся нуклеиновое основание выбрано из:
[0017] Вариант осуществления A8 представляет собой двухцепочечный олигонуклеотидный дуплекс, где первая олигонуклеотидная цепь содержит нуклеиновое основание по любому из вариантов осуществления A1-A5, и вторая комплементарная олигонуклеотидная цепь содержит комплементарное, спаривающееся нуклеиновое основание в его комплементарном, образующем пару оснований участке.
[0018] Вариант осуществления A9 представляет собой двухцепочечный олигонуклеотидный дуплекс по варианту осуществления A8, где первая олигонуклеотидная цепь содержит ; и вторая цепь содержит комплементарное, спаривающееся нуклеиновое основание, выбранное из:
 в его комплементарном, образующем пару оснований участке.
в его комплементарном, образующем пару оснований участке.
[0019] Вариант осуществления A10 представляет собой двухцепочечный олигонуклеотидный дуплекс по варианту осуществления A9, где вторая цепь содержит комплементарное, спаривающееся нуклеиновое основание 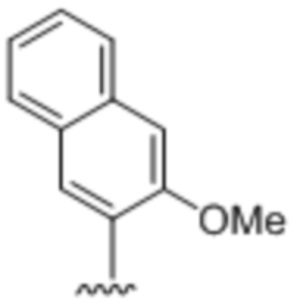 .
.
[0020] Вариант осуществления A11 представляет собой двухцепочечный олигонуклеотидный дуплекс по варианту осуществления A9, где вторая цепь содержит комплементарное, спаривающееся нуклеиновое основание 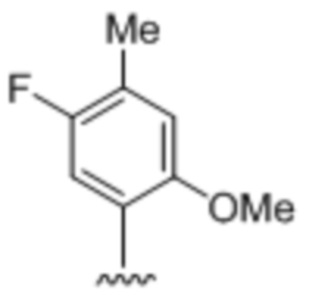 .
.
[0021] Вариант осуществления A12 является плазмидой, содержащей ген, кодирующий транспортную РНК (тРНК), и/или ген, кодирующий интересующий белок, где ген содержит по меньшей мере одно нуклеиновое основание по любому из вариантов осуществления A1-A5 или TPT3 (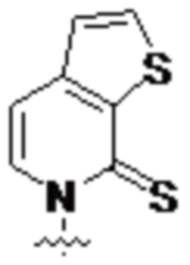 ) и по меньшей мере одно комплементарное, спаривающееся нуклеиновое основание по варианту осуществления A7 или NaM (), где комплементарное, спаривающееся нуклеиновое основание находится в комплементарном, образующем пару оснований участке.
) и по меньшей мере одно комплементарное, спаривающееся нуклеиновое основание по варианту осуществления A7 или NaM (), где комплементарное, спаривающееся нуклеиновое основание находится в комплементарном, образующем пару оснований участке.
[0022] Вариант осуществления A13 является мРНК, кодируемой плазмидой по варианту осуществления A10, кодирующей тРНК.
[0023] Вариант осуществления A14 является мРНК, кодируемой плазмидой по варианту осуществления A10, кодирующей белок.
[0024] Вариант осуществления A15 является транспортной РНК (тРНК), содержащей нуклеиновое основание по любому из вариантов осуществления A1-A5, содержащей:
антикодон, где антикодон содержит нуклеиновое основание, необязательно, где нуклеиновое основание находится в первом, втором или третьем положении антикодона; и
элемент распознавания, где элемент распознавания способствует селективному заряжанию тРНК неприродной аминокислотой посредством аминоацил-тРНК-синтетазы.
[0025] Вариант осуществления A16 является тРНК по варианту осуществления A15, где аминоацил-тРНК-синтетазу получают из Methanosarcina или его варианта или Methanococcus (Methanocaldococcus) или его варианта.
[0026] Вариант осуществления A17 является тРНК по варианту осуществления A15, где неприродная аминокислота содержит ароматический остаток.
[0027] Вариант осуществления A18 является тРНК по варианту осуществления A15, где неприродная аминокислота является лизином или производным фенилаланина.
[0028] Вариант осуществления A19 является структурой, содержащей формулу:
N1 - Zx - N2
где:
каждый Z независимо представляет собой нуклеиновое основание по любому из вариантов осуществления A1-A7, связанное с рибозилом, или дезоксирибозилом, или его аналогом;
N1 представляет собой один или более нуклеотидов или их аналогов или концевую фосфатную группу, присоединенную к 5'-концу рибозила, или дезоксирибозила, или его аналога из Z;
N2 представляет собой один или более нуклеотидов или их аналогов или концевую гидроксильную группу, присоединенную к 3'-концу рибозила, или дезоксирибозила, или его аналога из Z; и
x является целым числом от 1 до 20.
[0029] Вариант осуществления A20 является структурой по варианту осуществления A19, где структура кодирует ген, необязательно, где Zx находится в транслируемой области гена, или где Zx находится в нетранслируемой области гена.
[0030] Вариант осуществления A21 является полинуклеотидной библиотекой, где библиотека содержит по меньшей мере 5000 уникальных полинуклеотидов, и где каждый полинуклеотид содержит по меньшей мере одно нуклеиновое основание по любому из вариантов осуществления A1-A5.
[0031] Вариант осуществления A22 является нуклеозидтрифосфатом, содержащим нуклеиновое основание, где нуклеиновое основание выбрано из:
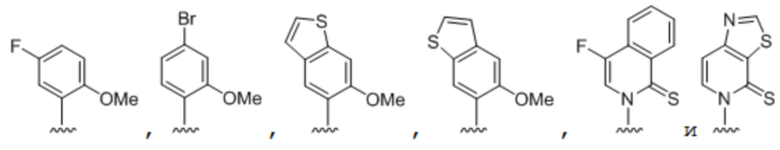 .
.
[0032] Вариант осуществления A23 является нуклеозидтрифосфатом по варианту осуществления A22, где нуклеиновое основание представляет собой .
[0033] Вариант осуществления A24 является нуклеозидтрифосфатом по варианту осуществления A22 или A23, где нуклеозид содержит рибозу или дезоксирибозу.
[0034] Вариант осуществления A25 является ДНК, содержащей нуклеиновое основание, имеющее структуру , и комплементарное, спаривающееся нуклеиновое основание имеющее структуру .
[0035] Вариант осуществления A26 является ДНК, содержащей нуклеиновое основание, имеющее структуру , и комплементарное, спаривающееся нуклеиновое основание, имеющее структуру .
[0036] Вариант осуществления A27 является способом транскрипции ДНК в тРНК или мРНК, кодирующую белок, включающим:
приведение ДНК, содержащей ген, кодирующий тРНК или белок, в контакт с рибонуклеозидтрифосфатами и РНК-полимеразой, где ген, кодирующий тРНК или белок, содержит первое неприродное основание, спаренное и образующее первую неприродную пару оснований со вторым неприродным оснований, и где рибонуклеозидтрифосфаты содержат третье неприродное основание, способное образовывать вторую неприродную пару оснований с первым неприродным основанием, где первая неприродная пара оснований и вторая неприродная пара оснований не являются одинаковыми.
[0037] Вариант осуществления A28 является способом по варианту осуществления A27, где рибонуклеозидтрифосфаты дополнительно содержат четвертое неприродное основание, где четвертое неприродное основание может образовывать вторую неприродную пару оснований с третьим неприродным основанием.
[0038] Вариант осуществления A29 является способом по варианту осуществления A28, где первая неприродная пара оснований и вторая неприродная пара оснований не являются одинаковыми.
[0039] Вариант осуществления A30 является способом по любому из вариантов осуществления A27-29, дополнительно включающим, перед приведением ДНК в контакт с рибонуклеозидтрифосфатами и РНК-полимеразой, репликацию ДНК посредством приведения ДНК в контакт с дезоксирибонуклеозидтрифосфатами и ДНК-полимеразой, где рибонуклеозидтрифосфаты содержат пятое неприродное основание, которое может образовывать пятую неприродную пару оснований с первым неприродным основанием, где первая неприродная пара оснований и пятая неприродная пара оснований не являются одинаковыми.
[0040] Вариант осуществления A30 является способом по любому из вариантов осуществления A27-A30, где первое неприродное основание содержит TPT3, второе неприродное основание содержит CNMO или NaM, третье неприродное основание содержит TAT1, и четвертое неприродное основание содержит NaM или 5FM.
[0041] Вариант осуществления A32 является способом по любому из вариантов осуществления A27-A31, где способ включает использование полусинтетического организма, необязательно, где организм является бактерией, необязательно, где бактерия является Escherichia coli.
[0042] Вариант осуществления A33 является способом по варианту осуществления A32, где организм включает микроорганизм.
[0043] Вариант осуществления A34 является способом по варианту осуществления A32, где организм включает бактерию.
[0044] Вариант осуществления A35 является способом по варианту осуществления A34, где организм включает грамположительную бактерию.
[0045] Вариант осуществления A36 является способом по варианту осуществления A34, где организм включает грамотрицательную бактерию.
[0046] Вариант осуществления A37 является способом по любому из вариантов осуществления A27-A34, где организм включает Escherichia coli.
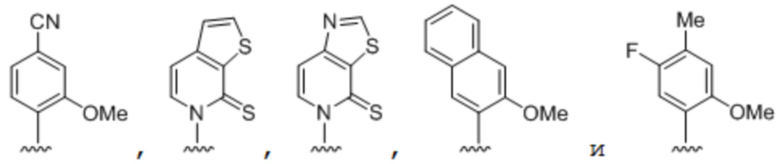
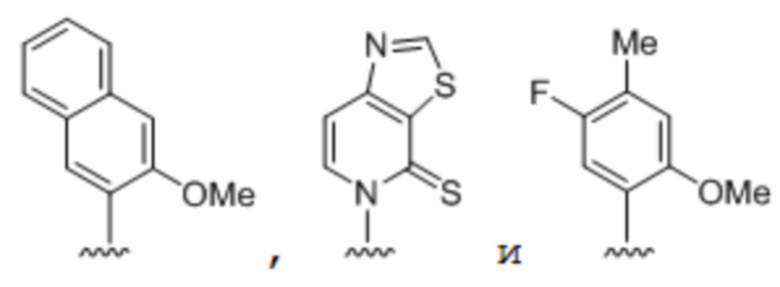
[0047] Вариант осуществления A38 является способом по любому из вариантов осуществления A27-A37, где по меньшей мере одно неприродное основание выбрано из группы, состоящей из
(i) 2-тиоурацила, 2-тиотимина, 2'-дезоксиуридина, 4-тиоурацила, 4-тиотимина, урацил-5-ила, гипоксантин-9-ила (I), 5-галоурацила; 5-пропинил-урацила, 6-азо-тимина, 6-азо-урацила, 5-метиламинометилурацила, 5-метоксиаминометил-2-тиоурацила, псевдоурацила, метилового эфира урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусной кислоты, 5-метил-2-тиоурацила, 3-(3-амино-3-N-2-карбоксипропил)урацила, 5-метил-2-тиоурацила, 4-тиоурацила, 5-метилурацила, 5'-метоксикарбоксиметилурацила, 5-метоксиурацила, урацил-5-оксиуксусной кислоты, 5-(карбоксигидроксилметил)урацила, 5-карбоксиметиламинометил-2-тиоуридина, 5-карбоксиметиламинометилурацила или дигидроурацила;
(ii) 5-гидроксиметилцитозина, 5-трифторметилцитозина, 5-галоцитозина, 5-пропинилцитозина, 5-гидроксицитозина, циклоцитозина, цитозинарабинозида, 5,6-дигидроцитозина, 5-нитроцитозина, 6-азоцитозина, азацитозина, N4-этилцитозина, 3-метилцитозина, 5-метилцитозина, 4-ацетилцитозина, 2-тиоцитозина, феноксазин-цитидин([5,4-b][1,4]бензоксазин-2(3H)-она), фенотиазин-цитидин-(1H-пиримидо[5,4-b][1,4]бензотиазин-2(3H)-она), феноксазин-цитидин-(9-(2-аминоэтокси)-H-пиримидо[5,4-b][1,4]бензоксазин-2(3H)-она), карбазол-цитидин-(2H-пиримидо[4,5-b]индол-2-она) или пиридоиндол-цитидин-(H-пиридо [3',2':4,5]пирроло[2,3-d]пиримидин-2-она);
(iii) 2-аминоаденина, 2-пропиладенина, 2-амино-аденина, 2-F-аденина, 2-амино-пропил-аденина, 2-амино-2'-дезоксиаденозина, 3-деазааденина, 7-метиладенина, 7-деаза-аденина, 8-азааденина, 8-галоген-, 8-амино-, 8-тиол-, 8-тиоалкил- и 8-гидроксил-замещенных аденинов, N6-изопентениладенина, 2-метиладенина, 2,6-диаминопурина, 2-метилтио-N6-изопентениладенина или 6-аза-аденина;
(iv) 2-метилгуанина, 2-пропиловых и алкиловых производных гуанина, 3-деазагуанина, 6-тио-гуанина, 7-метилгуанина, 7-деазагуанина, 7-деазагуанозина, 7-деаза-8-азагуанина, 8-азагуанина, 8-галоген-, 8-амино-, 8-тиол-, 8-тиоалкил- и 8-гидроксил-замещенных гуанинов, 1-метилгуанина, 2,2-диметилгуанина, 7-метилгуанина или 6-аза-гуанина; и
(v) гипоксантина, ксантина, 1-метилинозина, квеуозина, бета-D-галактозилквеуозина, инозина, бета-D-маннозилквеуозина, вибутоксозина, гидроксимочевины, (acp3)w, 2-аминопиридина или 2-пиридона.
[0048] Вариант осуществления A39 является способом по любому из вариантов осуществления A27-A37, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания содержит:
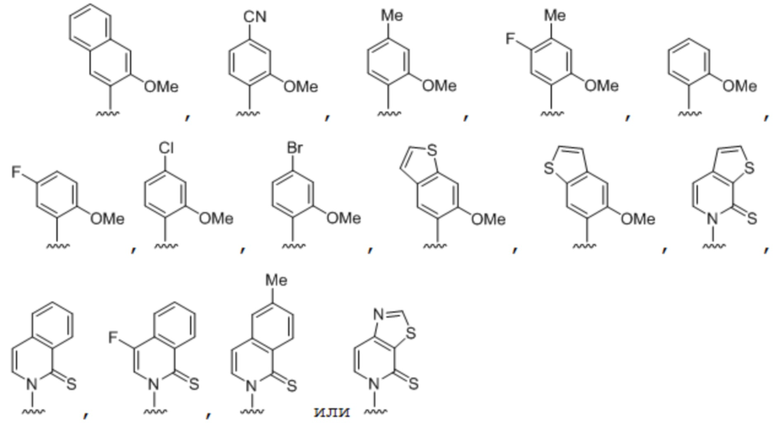
[0049] Вариант осуществления A40 является способом по любому из вариантов осуществления A27-A39, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
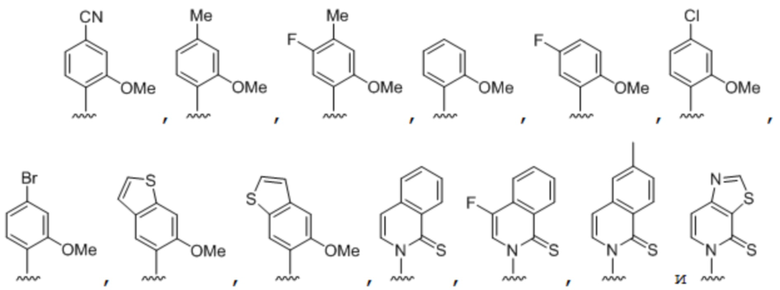 .
.
[0050] Вариант осуществления A41 является способом по варианту осуществления A40, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
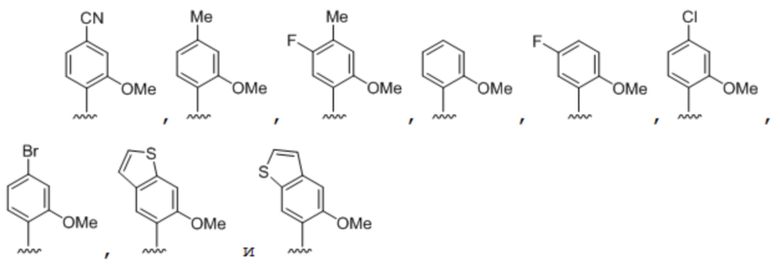
[0051] Вариант осуществления A42 является способом по варианту осуществления A40, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
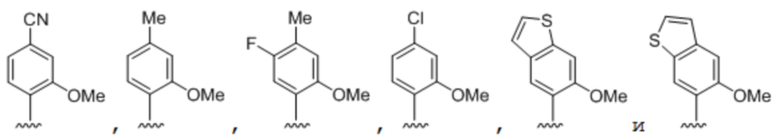 .
.
[0052] Вариант осуществления A43 является способом по варианту осуществления A40, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбран из группы, состоящей из:
 .
.
[0053] Вариант осуществления A44 является способом по варианту осуществления A40, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания содержит:
 .
.
[0054] Вариант осуществления A45 является способом по варианту осуществления A40, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания содержит:
 или .
или .
[0055] Вариант осуществления A46 является способом по варианту осуществления A40, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания содержит:
 .
.
[0056] Вариант осуществления A47 является способом по варианту осуществления A40, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
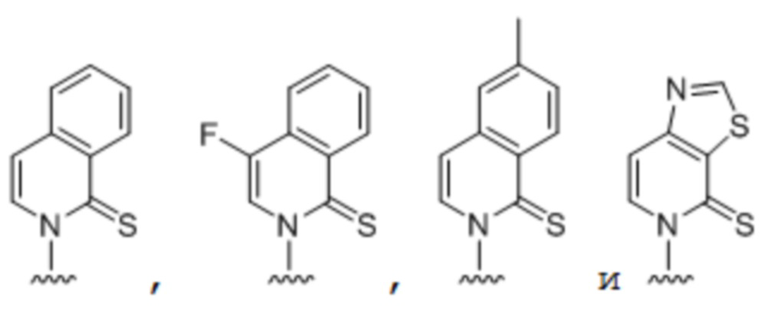 .
.
[0057] Вариант осуществления A48 является способом по варианту осуществления A40, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания представляет собой .
[0058] Вариант осуществления A49 является способом по варианту осуществления A40, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания содержит:
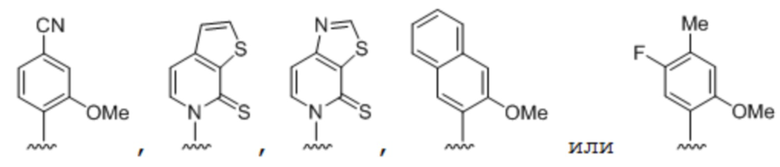 .
.
[0059] Вариант осуществления A50 является способом по варианту осуществления A40, где первое или второе неприродное основание представляет собой .
[0060] Вариант осуществления A51 является способом по варианту осуществления A40, где первое или второе неприродное основание представляет собой 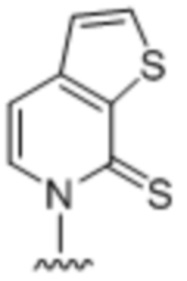 .
.
[0061] Вариант осуществления A52 является способом по любому из вариантов осуществления A27-A40, где первое неприродное основание представляет собой , и второе неприродное основание представляет собой , или первое неприродное основание представляет собой , и второе неприродное основание представляет собой .
[0062] Вариант осуществления A53 является способом по любому из вариантов осуществления A27-A40 и A52, где третье или четвертое неприродное основание представляет собой .
[0063] Вариант осуществления A54 является способом по варианту осуществления A53, где третье неприродное основание представляет собой .
[0064] Вариант осуществления A55 является способом по варианту осуществления A54, где четвертое неприродное основание представляет собой .
[0065] Вариант осуществления A56 является способом по любому из вариантов осуществления A27-A52, где третье или четвертое неприродное основание представляет собой или .
[0066] Вариант осуществления A57 является способом по варианту осуществления A56, где третье неприродное основание представляет собой или .
[0067] Вариант осуществления A58 является способом по варианту осуществления A56, где четвертое неприродное основание представляет собой или .
[0068] Вариант осуществления A59 является способом по любому из вариантов осуществления A27-A40, где первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой , и четвертое неприродное основание представляет собой или .
[0069] Вариант осуществления A60 является способом по любому из вариантов осуществления A27-A40, где первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой , и четвертое неприродное основание представляет собой или .
[0070] Вариант осуществления A61 является способом по любому из вариантов осуществления A27-A40, где первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой или , и четвертое неприродное основание представляет собой .
[0071] Вариант осуществления A62 является способом по любому из вариантов осуществления A27-A40, где третье неприродное основание представляет собой или .
[0072] Вариант осуществления A63 является способом по любому из вариантов осуществления A27-A51, где четвертое неприродное основание представляет собой .
[0073] Вариант осуществления A64 является способом по любому из вариантов осуществления A27-A40, где первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой или , и четвертое неприродное основание представляет собой .
[0074] Вариант осуществления A65 является способом по любому из вариантов осуществления A27-A64, где третье неприродное основание и четвертое неприродное основание содержат рибозу.
[0075] Вариант осуществления A66 является способом по любому из вариантов осуществления A27-A64, где третье неприродное основание и четвертое неприродное основание содержат дезоксирибозу.
[0076] Вариант осуществления A65 является способом по любому из вариантов осуществления A27-A66, где первое и второе неприродные основания содержат дезоксирибозу.
[0077] Вариант осуществления A68 является способом по любому из вариантов осуществления A27-A64, где первое и второе неприродные основания содержат дезоксирибозу, и третье неприродное основание и четвертое неприродное основание содержат рибозу.
[0078] Вариант осуществления A69 является способом по любому из вариантов осуществления A27-A40, где ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из:
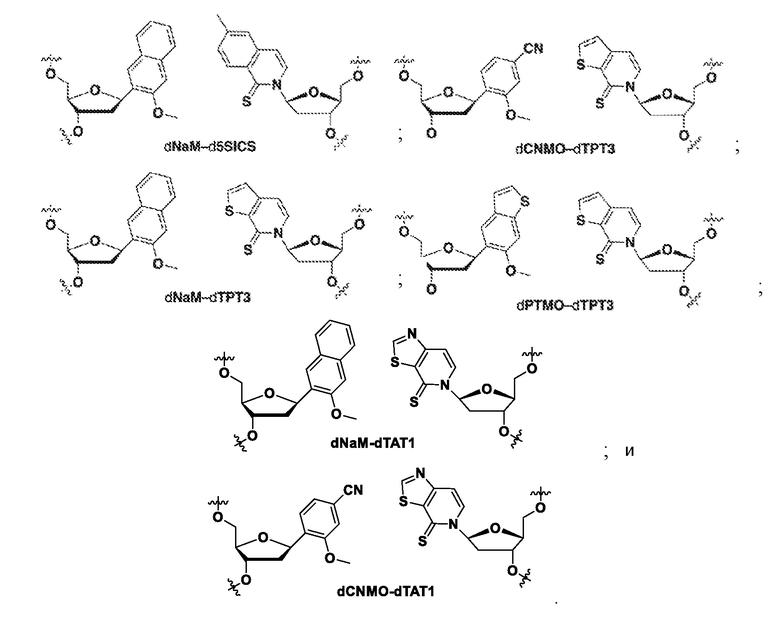
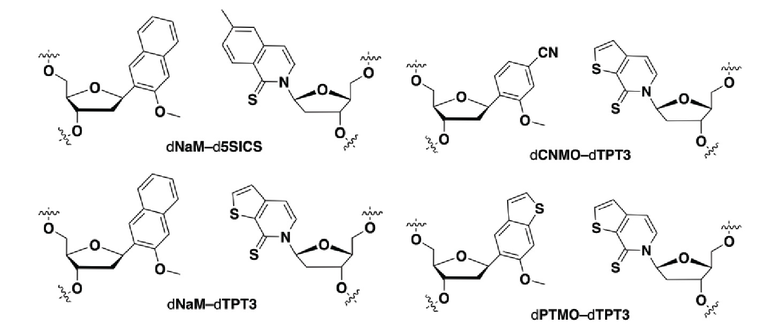
[0079] Вариант осуществления A70 является способом по варианту осуществления A69, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS.
[0080] Вариант осуществления A71 является способом по варианту осуществления A69, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3.
[0081] Вариант осуществления A72 является способом по варианту осуществления A69, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3.
[0082] Вариант осуществления A73 является способом по варианту осуществления A69, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dPTMO-dTPT3.
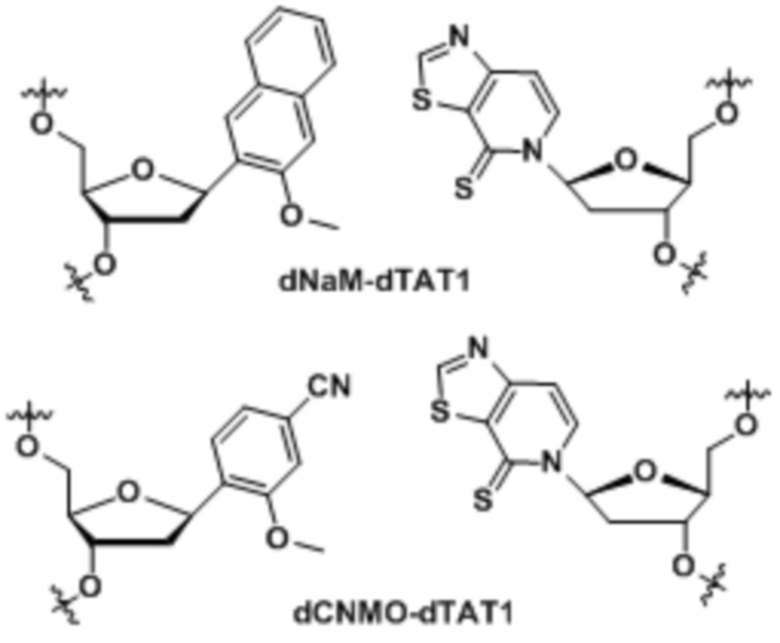
[0083] Вариант осуществления A74 является способом по варианту осуществления A69, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTAT1.
[0084] Вариант осуществления A75 является способом по варианту осуществления A69, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTAT1.
[0085] Вариант осуществления A76 является способом по любому из вариантов осуществления A27-A40, где ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из
 ; и
; и
где мРНК и/или тРНК содержат по меньшей мере одно неприродное основание, выбранное из:
 .
.
[0086] Вариант осуществления A77 является способом по варианту осуществления A76, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS.
[0087] Вариант осуществления A78 является способом по варианту осуществления A76, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3.
[0088] Вариант осуществления A79 является способом по варианту осуществления A76, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3.
[0089] Вариант осуществления A80 является способом по варианту осуществления A76, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dPTMO-dTPT3.
[0090] Вариант осуществления A81 является способом по варианту осуществления A76, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTAT1.
[0091] Вариант осуществления A82 является способом по варианту осуществления A76, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTAT1.
[0092] Вариант осуществления A83 является способом по любому из вариантов осуществления A76-A82, где мРНК и тРНК содержат неприродное основание, выбранное из
 .
.
[0093] Вариант осуществления A84 является способом по варианту осуществления A83, где мРНК и тРНК содержат неприродное основание, выбранное из и .
[0094] Вариант осуществления A85 является способом по варианту осуществления A83, где мРНК содержит неприродное основание, являющееся .
[0095] Вариант осуществления A86 является способом по варианту осуществления A83, где мРНК содержит неприродное основание, являющееся .
[0096] Вариант осуществления A87 является способом по варианту осуществления A83, где мРНК содержит неприродное основание, являющееся .
[0097] Вариант осуществления A88 является способом по любому из вариантов осуществления A76-A87, где тРНК содержит неприродное основание, выбранное из и .
[0098] Вариант осуществления A89 является способом по варианту осуществления A88, где тРНК содержит неприродное основание, являющееся .
[0099] Вариант осуществления A90 является способом по варианту осуществления A88, где тРНК содержит неприродное основание, являющееся .
[00100] Вариант осуществления A91 является способом по любому из вариантов осуществления A76-A87, где тРНК содержит неприродное основание, являющееся .
[00101] Вариант осуществления A92 является способом по любому из вариантов осуществления A27-A40, где первое неприродное основание содержит dCNMO, и второе неприродное основание содержит dTPT3.
[00102] Вариант осуществления A93 является способом по любому из вариантов осуществления A27-40 и A92, где третье неприродное основание содержит NaM, и второе неприродное основание содержит TAT1.
[00103] Вариант осуществления A94 является способом по любому из вариантов осуществления A27-A93, где первое неприродное основание или второе неприродное основание распознается ДНК-полимеразой.
[00104] Вариант осуществления A95 является способом по любому из вариантов осуществления A27-A94, где третье неприродное основание или четвертое неприродное основание распознается РНК-полимеразой.
[00105] Вариант осуществления A96 является способом по любому из вариантов осуществления A27-A95, где мРНК транскрибируют, и способ дополнительно включает трансляцию мРНК в белок, где белок содержит неприродную аминокислоту в положении, соответствующем кодону мРНК, содержащему третье неприродное основание.
[00106] Вариант осуществления A97 является способом по любому из вариантов осуществления A27-A96, где белок содержит по меньшей мере две неприродные аминокислоты.
[00107] Вариант осуществления A98 является способом по любому из вариантов осуществления A27-A96, где белок содержит по меньшей мере три неприродные аминокислоты.
[00108] Вариант осуществления A99 является способом по любому из вариантов осуществления A27-A98, где белок содержит по меньшей мере две разные неприродные аминокислоты.
[00109] Вариант осуществления A100 является способом по любому из вариантов осуществления A27-A98, где белок содержит по меньшей мере три разные неприродные аминокислоты.
[00110] Вариант осуществления A101 является способом по любому из вариантов осуществления A27-A100, где по меньшей мере одна неприродная аминокислота:
является аналогом лизина;
содержит ароматическую боковую цепь;
содержит азидо-группу;
содержит алкиновую группу; или
содержит альдегидную или кетоновую группу.
[00111] Вариант осуществления A102 является способом по любому из вариантов осуществления A27-A101, где по меньшей мере одна неприродная аминокислота не содержит ароматическую боковую цепь.
[00112] Вариант осуществления A103 является способом по любому из вариантов осуществления A27-A102, где по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK), N6-пропаргилэтокси-карбонил-L-лизин (PraK), BCN-L-лизин, норборнен-лизин, TCO-лизин, метилтетразин-лизин, аллилоксикарбониллизин, 2-амино-8-оксононановую кислоту, 2-амино-8-оксооктановую кислоту, p-ацетил-L-фенилаланин, p-азидометил-L-фенилаланин (pAMF), p-йодо-L-фенилаланин, m-ацетилфенилаланин, 2-амино-8-оксононановую кислоту, p-пропаргилоксифенилаланин, p-пропаргил-фенилаланин, 3-метил-фенилаланин, L-ДОФА, фторированный фенилаланин, изопропил-L-фенилаланин, p-азидо-L-фенилаланин, p-ацил-L-фенилаланин, p-бензоил-L-фенилаланин, p-бромфенилаланин, p-амино-L- фенилаланин, изопропил-L-фенилаланин, O-аллилтирозин, O-метил-L-тирозин, O-4-аллил-L-тирозин, 4-пропил-L-тирозин, фосфонотирозин, три-O-ацетил-GlcNAcp-серин, L-фосфосерин, фосфоносерин, L-3-(2-нафтил)аланин, 2-амино-3-((2-((3-(бензилокси)-3-оксопропил)амино)этил)селанил)пропионовую кислоту, 2-амино-3-(фенилселанил)пропионовую кислоту или селеноцистеин.
[00113] Вариант осуществления A104 является способом по варианту осуществления A102 или A103, где по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK) или N6-пропаргилэтокси-карбонил-L-лизин (PraK).
[00114] Вариант осуществления A105 является способом по варианту осуществления A104, где по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK).
[00115] Вариант осуществления A106 является способом по варианту осуществления A104, где по меньшей мере одна неприродная аминокислота содержит N6-пропаргилэтокси-карбонил-L-лизин (PraK).
[00116] Вариант осуществления A107 является мРНК, получаемой способом по любому из вариантов осуществления A27-A106.
[00117] Вариант осуществления A108 является тРНК, получаемой способом по любому из вариантов осуществления A27-A106.
[00118] Вариант осуществления A109 является белком, кодируемым мРНК по варианту осуществления A107, содержащим неприродную аминокислоту в положении, соответствующем кодону мРНК, содержащему третье неприродное основание.
[00119] Вариант осуществления A110 является полусинтетическим организмом, содержащим расширенный генетический алфавит, где генетический алфавит содержит по меньшей мере три разных неприродных основания.
[00120] Вариант осуществления A111 является полусинтетическим организмом по варианту осуществления A110, где организм включает микроорганизм, необязательно, где микроорганизм является Escherichia coli.
[00121] Вариант осуществления A112 является полусинтетическим организмом по варианту осуществления A110 или A111, где организм содержит ДНК, содержащую по меньшей мере одно неприродное нуклеиновое основание, выбранное из:
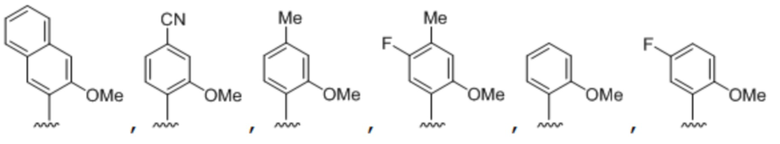
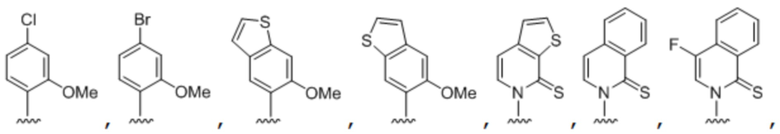
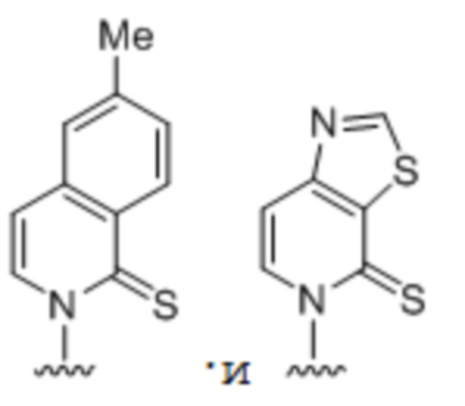 .
.
[00122] Вариант осуществления A113 является полусинтетическим организмом по любому из вариантов осуществления A110-A113, где ДНК содержит по меньшей мере одну неприродную пару оснований (UBP),
[00123] где неприродная пара оснований (UBP) представляет собой dCNMO-dTPT3, dNaM-dTPT3, dCNMO-dTAT1, d5FM-dTAT1 или dNaM-dTAT1.
[00124] Вариант осуществления A114 является полусинтетическим организмом по варианту осуществления A112, где ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, являющееся .
[00125] Вариант осуществления A115 является полусинтетическим организмом по любому из вариантов осуществления A110-A115, где организм экспрессирует гетерологичный транспортер нуклеозидтрифосфата.
[00126] Вариант осуществления A116 является полусинтетическим организмом по варианту осуществления A115, где гетерологичный транспортер нуклеозидтрифосфата является PtNTT2.
[00127] Вариант осуществления A117 является полусинтетическим организмом по любому из вариантов осуществления A110-A116, где организм дополнительно экспрессирует гетерологичную тРНК-синтетазу.
[00128] Вариант осуществления A118 является полусинтетическим организмом по варианту осуществления A117, где гетерологичная тРНК-синтетаза является пирролизил-тРНК-синтетазой M. barkeri (Mb PylRS).
[00129] Вариант осуществления A119 является полусинтетическим организмом по любому из вариантов осуществления A110-A118, где организм дополнительно экспрессирует гетерологичную РНК-полимеразу.
[00130] Вариант осуществления A120 является полусинтетическим организмом по варианту осуществления A119, где гетерологичная РНК-полимераза является RNAP T7.
[00131] Вариант осуществления A121 является полусинтетическим организмом по любому из вариантов осуществления A110-A120, где организм не экспрессирует белок, имеющий функцию рекомбинационной репарации ДНК.
[00132] Вариант осуществления A122 является полусинтетическим организмом по варианту осуществления A121, где организм не экспрессирует RecA.
[00133] Вариант осуществления A123 является полусинтетическим организмом по любому из вариантов осуществления A110-A122, дополнительно содержащим гетерологичную мРНК.
[00134] Вариант осуществления A124 является полусинтетическим организмом по варианту осуществления A123, где гетерологичная мРНК содержит по меньшей мере одно неприродное основание, выбранное из:
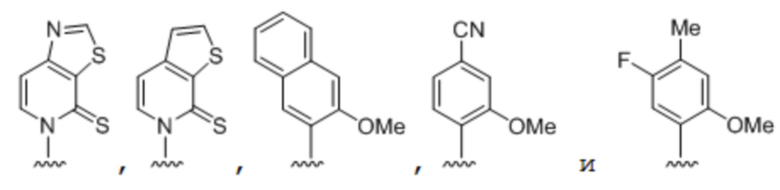 .
.
[00135] Вариант осуществления A125 является полусинтетическим организмом по любому из вариантов осуществления A110-A125, дополнительно содержащим гетерологичную тРНК.
[00136] Вариант осуществления A126 является полусинтетическим организмом по варианту осуществления A125, где гетерологичная тРНК содержит по меньшей мере одно неприродное основание, выбранное из:
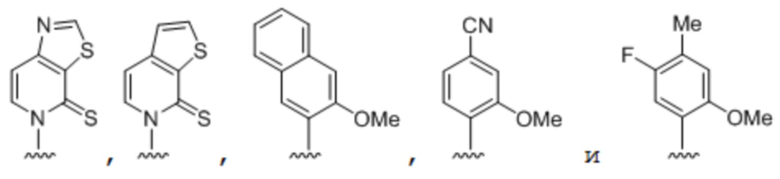 .
.
[00137] Вариант осуществления A127 является способом транскрипции ДНК, включающим:
получение одной или более ДНК, содержащей (1) ген, кодирующий белок, где матричная цепь гена, кодирующего белок, содержит первое неприродное основание, и (2) ген, кодирующий тРНК, где матричная цепь гена, кодирующего тРНК, содержит второе неприродное основание, которое может образовывать пару оснований с первым неприродным основанием;
транскрипцию гена, кодирующего белок, для встраивания третьего неприродного основания в мРНК, третье неприродное основание, которое может образовывать первую неприродную пару оснований с первым неприродным основанием;
транскрипцию гена, кодирующего тРНК, для встраивания четвертого неприродного основания в тРНК, где четвертое неприродное основание может образовывать вторую неприродную пару оснований со вторым неприродным основанием, где первая неприродная пара оснований и вторая неприродная пара оснований не являются одинаковыми.
[00138] Вариант осуществления A128 является способом по варианту осуществления A127, дополнительно включающим трансляцию белка из мРНК с использованием тРНК, где указанный белок содержит неприродную аминокислоту в положении, соответствующем кодону, содержащему третье неприродное основание, в мРНК.
[00139] Вариант осуществления A129 является способом репликации ДНК, включающим:
получение ДНК, содержащей (1) ген, кодирующий белок, где матричная цепь гена, кодирующего белок, содержит первое неприродное основание, и (2) ген, кодирующий тРНК, где матричная цепь гена, кодирующего тРНК, содержит второе неприродное основание, которое может образовывать пару оснований с первым неприродным основанием; и
репликацию ДНК для встраивания первого неприродного основания-заместителя вместо первого неприродного основания и/или для встраивания второго неприродного основания-заместителя вместо второго неприродного основания;
где способ необязательно дополнительно включает:
транскрипцию гена, кодирующего белок, для встраивания третьего неприродного основания в мРНК, при этом третье неприродное основание может образовывать первую неприродную пару оснований с первым неприродным основанием и/или первым неприродным основанием-заместителем; и/или
транскрипцию гена, кодирующего тРНК, для встраивания четвертого неприродного основания в тРНК, где четвертое неприродное основание может образовывать вторую неприродную пару оснований со вторым неприродным основанием и/или вторым неприродным основанием-заместителем, где первая неприродная пара оснований и вторая неприродная пара оснований не являются одинаковыми.
[00140] Вариант осуществления A130 является способом по варианту осуществления A129, дополнительно включающим транскрипцию гена, кодирующего белок, для встраивания третьего неприродного основания в мРНК, при этом третье неприродное основание может образовывать первую неприродную пару оснований с первым неприродным основанием и/или первым неприродным основанием-заместителем.
[00141] Вариант осуществления A131 является способом по варианту осуществления A129 или A130, дополнительно включающим транскрипцию гена, кодирующего тРНК, для встраивания четвертого неприродного основания в тРНК, где четвертое неприродное основание может образовывать вторую неприродную пару оснований со вторым неприродным основанием и/или вторым неприродным основанием-заместителем, где первая неприродная пара оснований и вторая неприродная пара оснований не являются одинаковыми.
[00142] Вариант осуществления A132 является способом по любому из вариантов осуществления A127-A131, где способ включает использование полусинтетического организма.
[00143] Вариант осуществления A133 является способом по варианту осуществления A132, где организм включает микроорганизм.
[00144] Вариант осуществления A134 является способом по варианту осуществления A132 или A133, где способ является способом in vivo, включающим использование полусинтетического организма, являющегося бактерией.
[00145] Вариант осуществления A135 является способом по варианту осуществления A134, где организм включает грамположительную бактерию.
[00146] Вариант осуществления A136 является способом по варианту осуществления A134, где организм включает грамотрицательную бактерию.
[00147] Вариант осуществления A137 является способом по вариантам осуществления A132-A134, где организм включает Escherichia coli.
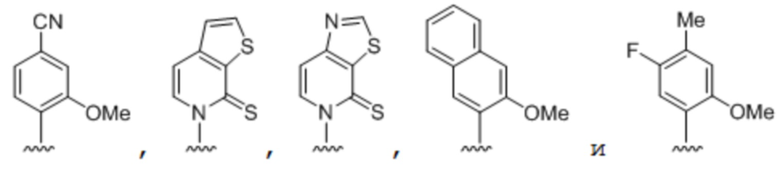
[00148] Вариант осуществления A138 является способом по любому из вариантов осуществления A127-A137, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания содержит:
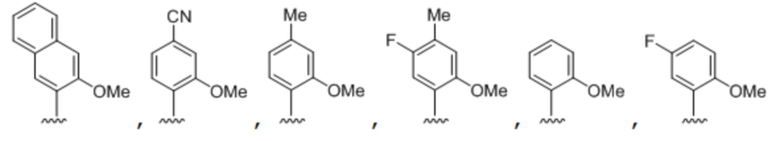
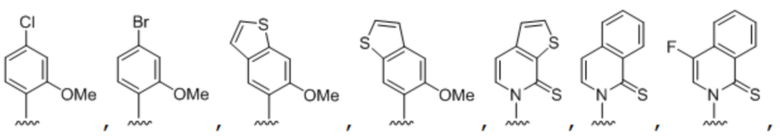
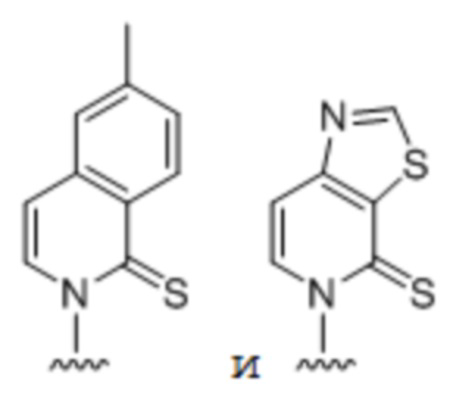 .
.
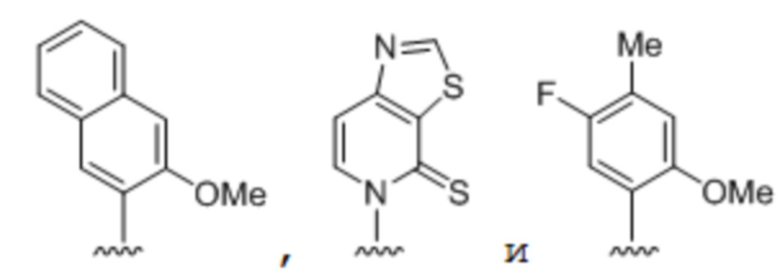
[00149] Вариант осуществления A139 является способом по варианту осуществления A138, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания содержит:
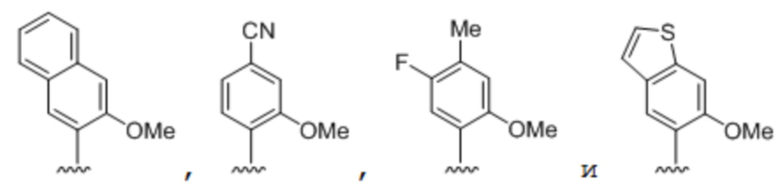 .
.
[00150] Вариант осуществления A140 является способом по варианту осуществления A138, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания содержит:
и .
[00151] Вариант осуществления A141 является способом по варианту осуществления A138, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания содержит
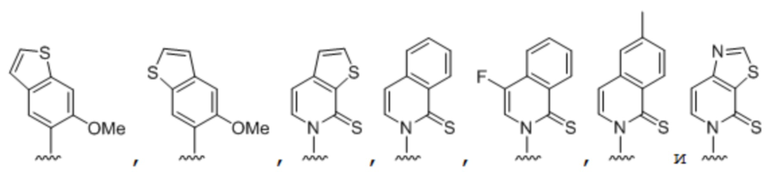 .
.
[00152] Вариант осуществления A142 является способом по варианту осуществления A138, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания представляет собой .
[00153] Вариант осуществления A143 является способом по варианту осуществления A138, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания содержит:
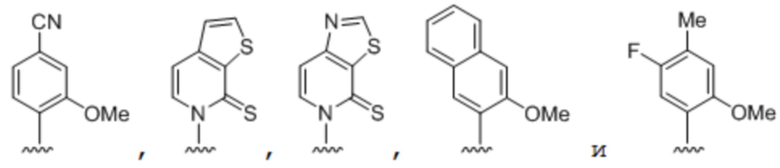 .
.
[00154] Вариант осуществления A144 является способом по варианту осуществления A138, где первое или второе неприродное основание представляет собой .
[00155] Вариант осуществления A145 является способом по варианту осуществления A138, где первое или второе неприродное основание представляет собой .
[00156] Вариант осуществления A146 является способом по варианту осуществления A138, где первое неприродное основание представляет собой , и второе неприродное основание представляет собой .
[00157] Вариант осуществления A147 является способом по варианту осуществления A138, где первое неприродное основание представляет собой , и второе неприродное основание представляет собой .
[00158] Вариант осуществления A148 является способом по любому из вариантов осуществления A138, A146 и A147, где третье или четвертое неприродное основание представляет собой .
[00159] Вариант осуществления A149 является способом по варианту осуществления A148, где третье неприродное основание представляет собой .
[00160] Вариант осуществления A150 является способом по варианту осуществления A148, где четвертое неприродное основание представляет собой .
[00161] Вариант осуществления A151 является способом по любому из вариантов осуществления A138, A146 и A147, где третье или четвертое неприродное основание представляет собой или .
[00162] Вариант осуществления A152 является способом по варианту осуществления A151, где третье неприродное основание представляет собой или .
[00163] Вариант осуществления A153 является способом по варианту осуществления A151, где четвертое неприродное основание представляет собой или .
[00164] Вариант осуществления A154 является способом по любому из вариантов осуществления A138, где первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой , и четвертое неприродное основание представляет собой или .
[00165] Вариант осуществления A155 является способом по варианту осуществления A138, где первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой , и четвертое неприродное основание представляет собой или .
[00166] Вариант осуществления A является способом по варианту осуществления A138, где первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой или и четвертое неприродное основание представляет собой .
[00167] Вариант осуществления A157 является способом по варианту осуществления A138, где третье неприродное основание представляет собой или .
[00168] Вариант осуществления A158 является способом по варианту осуществления A138, где четвертое неприродное основание представляет собой .
[00169] Вариант осуществления A159 является способом по варианту осуществления A138, где первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой или , и четвертое неприродное основание представляет собой .
[00170] Вариант осуществления A160 является способом по любому из вариантов осуществления A127-A159, где третье неприродное основание и четвертое неприродное основание содержат рибозу.
[00171] Вариант осуществления A161 является способом по любому из вариантов осуществления A127-A159, где третье неприродное основание и четвертое неприродное основание содержат дезоксирибозу.
[00172] Вариант осуществления A162 является способом по любому из вариантов осуществления A127-A161, где первое и второе неприродные основания содержат дезоксирибозу.
[00173] Вариант осуществления A163 является способом по любому из вариантов осуществления A127-A159, где первое и второе неприродные основания содержат дезоксирибозу, и третье неприродное основание и четвертое неприродное основание содержат рибозу.
[00174] Вариант осуществления A164 является способом по любому из вариантов осуществления A127-A137, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из группы, состоящей из
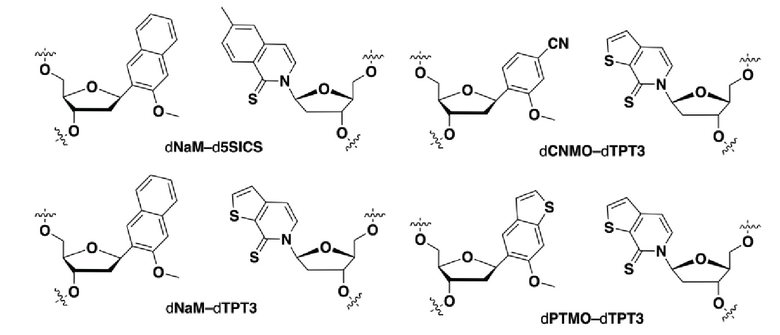
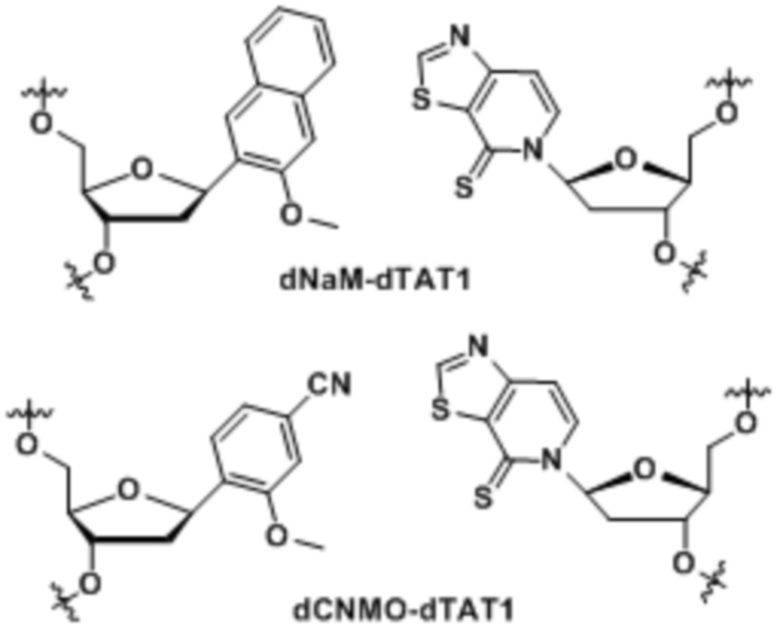 .
.
[00175] Вариант осуществления A165 является способом по варианту осуществления A164, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS.
[00176] Вариант осуществления A166 является способом по варианту осуществления A164, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3.
[00177] Вариант осуществления A167 является способом по варианту осуществления A164, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3.
[00178] Вариант осуществления A168 является способом по варианту осуществления A164, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTAT1.
[00179] Вариант осуществления A169 является способом по варианту осуществления A164, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTAT1.
[00180] Вариант осуществления A170 является способом по любому из вариантов осуществления A127-A137, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из группы, состоящей из
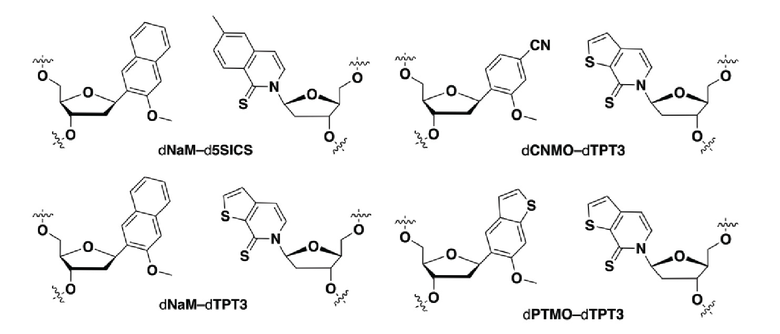
; и
где мРНК и тРНК содержат по меньшей мере одно неприродное основание, выбранное из:
 .
.
[00181] Вариант осуществления A171 является способом по варианту осуществления A170, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS.
[00182] Вариант осуществления A172 является способом по варианту осуществления A170, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3.
[00183] Вариант осуществления A173 является способом по варианту осуществления A170, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3.
[00184] Вариант осуществления A174 является способом по варианту осуществления A170, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTAT1.
[00185] Вариант осуществления A175 является способом по варианту осуществления A170, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTAT1.
[00186] Вариант осуществления A176 является способом по любому из вариантов осуществления A127-A175, где мРНК и тРНК содержат неприродное основание, выбранное из
 .
.
[00187] Вариант осуществления A177 является способом по варианту осуществления A176, где мРНК и тРНК содержат неприродное основание, выбранное из и .
[00188] Вариант осуществления A178 является способом по варианту осуществления A176, где мРНК содержит неприродное основание, являющееся .
[00189] Вариант осуществления A179 является способом по варианту осуществления A176, где мРНК содержит неприродное основание, являющееся .
[00190] Вариант осуществления A180 является способом по варианту осуществления A176, где мРНК содержит неприродное основание, являющееся .
[00191] Вариант осуществления A181 является способом по варианту осуществления A176, где тРНК содержит неприродное основание, выбранное из и .
[00192] Вариант осуществления A182 является способом по варианту осуществления A176, где тРНК содержит неприродное основание, являющееся .
[00193] Вариант осуществления A183 является способом по варианту осуществления A176, где тРНК содержит неприродное основание, являющееся .
[00194] Вариант осуществления A184 является способом по варианту осуществления A176, где тРНК содержит неприродное основание, являющееся .
[00195] Вариант осуществления A185 является способом по любому из вариантов осуществления A127-A137, где первое неприродное основание содержит dCNMO, и второе неприродное основание содержит dTPT3.
[00196] Вариант осуществления A186 является способом по любому из вариантов осуществления A127-A137, где третье неприродное основание содержит NaM, и второе неприродное основание содержит TAT1.
[00197] Вариант осуществления A187 является способом по любому из вариантов осуществления A127-A186, где белок содержит по меньшей мере две неприродных аминокислоты.
[00198] Вариант осуществления A188 является способом по любому из вариантов осуществления A127-A186, где белок содержит по меньшей мере три неприродных аминокислоты.
[00199] Вариант осуществления A189 является способом по любому из вариантов осуществления A127-A186, где белок содержит по меньшей мере две разные неприродные аминокислоты.
[00200] Вариант осуществления A190 является способом по любому из вариантов осуществления A127-A186, где белок содержит по меньшей мере три разные неприродные аминокислоты.
[00201] Вариант осуществления A191 является способом по любому из вариантов осуществления A127-A190, где по меньшей мере одна неприродная аминокислота:
[00202] является аналогом лизина;
[00203] содержит ароматическую боковую цепь;
[00204] содержит азидо-группу;
[00205] содержит алкиновую группу; или
[00206] содержит альдегидную или кетоновую группу.
[00207] Вариант осуществления A192 является способом по любому из вариантов осуществления A127-A191, где по меньшей мере одна неприродная аминокислота не содержит ароматическую боковую цепь.
[00208] Вариант осуществления A193 является способом по вариантам осуществления A191 или A192, где по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK) или N6-пропаргилэтокси-карбонил-L-лизин (PraK).
[00209] Вариант осуществления A194 является способом по варианту осуществления A193, где по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK).
[00210] Вариант осуществления A195 является способом по варианту осуществления A193, где по меньшей мере одна неприродная аминокислота содержит N6-пропаргилэтокси-карбонил-L-лизин (PraK).
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[00211] Различные аспекты изобретения подробно изложены в прилагаемой формуле изобретения. Признаки и преимущества настоящего изобретения будут более понятными со ссылкой на следующее подробное описание, в котором приведены иллюстративные варианты осуществления, в которых использованы принципы настоящего изобретения, и сопутствующие чертежи, на которых:
[00212] На фиг. 1A проиллюстрирована схема использования неприродных пар оснований (UBP) для сайт-специфического встраивания неканонических аминокислот (ncAA) в белок с использованием неприродных пар оснований X-Y. Встраивание трех ncAA в белок показано исключительно в качестве примера; можно встраивать любое количество ncAA.
[00213] На фиг.1 B показаны неприродные пары оснований (UBP).
[00214] На фиг.2 показаны аналоги dXTP. Рибоза и фосфаты опущены для простоты.
[00215] На фиг. 3A показан график удержания UBP (%) в гене sfGFP для оптимизации встраивания AzK в sfGFP с использованием различных dXTP. Каждый столбик соответствует среднему значению, при этом планками погрешностей указана стандартная ошибка (n=3). Незакрашенными кругами указаны данные для каждого независимого исследования. Звездочкой указано, что клетки не могли расти в указанных условиях.
[00216] На фиг. 3B показан график удержания UBP (%) в гене тРНК-Pyl для оптимизации встраивания AzK в sfGFP с использованием различных dXTP. Каждый столбик соответствует среднему значению, при этом планками погрешностей указана стандартная ошибка (n=3). Незакрашенными кругами указаны данные для каждого независимого исследования. Звездочкой указано, что клетки не могли расти в указанных условиях.
[00217] На фиг. 3C показан график наблюдаемой относительной флуоресценции sfGFP, нормализованной по росту клеток (относительные единицы флуоресценции (RFU) на OD600) в присутствии или отсутствие AzK для оптимизации встраивания AzK в sfGFP с использованием различных dXTP. Каждый столбик соответствует среднему значению, при этом планками погрешностей указана стандартная ошибка (n=3). Незакрашенными кругами указаны данные для каждого независимого исследования. Звездочкой указано, что клетки не могли расти в указанных условиях.
[00218] На фиг. 3D показан график относительного белкового сдвига (%), измеренного посредством вестерн-блоттинга, для оптимизации встраивания AzK в sfGFP с использованием различных dXTP. Каждый столбик соответствует среднему значению, при этом планками погрешностей указана стандартная ошибка (n=3). Незакрашенными кругами указаны данные для каждого независимого исследования. Звездочкой указано, что клетки не могли расти в указанных условиях.
[00219] На фиг. 4A показаны аналоги рибонуклеотида XTP. Рибоза и фосфаты опущены для простоты.
[00220] На фиг. 4B показаны аналоги рибонуклеотида YTP. Рибоза и фосфаты опущены для простоты.
[00221] На фиг. 5A показан график анализа трансляции SAR с использованием различных неприродных рибонуклеотидов для встраивания AzK в sfGFP с общей флуоресценцией sfGFP (RFU), наблюдаемой в присутствии AzK для аналогов XTP, на оси y. Каждый столбик соответствует среднему значению, при этом планками погрешностей указана стандартная ошибка (n=4). Незакрашенными кругами указаны данные для каждого независимого исследования.
[00222] На фиг. 5B показан график анализа трансляции SAR с использованием различных неприродных рибонуклеотидов для встраивания AzK в sfGFP с белковым сдвигом (%), измеренным посредством вестерн-блоттинга для аналогов XTP, по оси y. Каждый столбик соответствует среднему значению, при этом планками погрешностей указана стандартная ошибка (n=4). Незакрашенными кругами указаны данные для каждого независимого исследования.
[00223] На фиг. 5C показан график анализа трансляции SAR с использованием различных неприродных рибонуклеотидов для встраивания AzK в sfGFP с общей флуоресценцией sfGFP (RFU), наблюдаемой в присутствии AzK для аналогов YTP, по оси y. Каждый столбик соответствует среднему значению, при этом планками погрешностей указана стандартная ошибка (n=4). Незакрашенными кругами указаны данные для каждого независимого исследования.
[00224] На фиг. 5D показан график анализа трансляции SAR с использованием различных неприродных рибонуклеотидов для встраивания AzK в sfGFP с белковым сдвигом (%), измеряемым посредством вестерн-блоттинга для аналогов YTP, по оси y. Каждый столбик соответствует среднему значению, при этом планками погрешностей указана стандартная ошибка (n=4). Незакрашенными кругами указаны данные для каждого независимого исследования.
[00225] На фиг.6A показан график оптимизации концентраций неприродных рибонуклеотидтрифосфатов по общей флуоресценции sfGFP (RFU) как функции концентраций NaMTP и TAT1TP (мкМ). Планками погрешностей указана стандартная ошибка для каждого значения (n=3).
[00226] На фиг. 6B показан график оптимизации концентраций неприродных рибонуклеотидтрифосфатов по общей флуоресценции sfGFP (RFU) как функции концентраций 5FMTP и TAT1TP (мкМ). Планками погрешностей указана стандартная ошибка для каждого значения (n=3).
[00227] На фиг. 6C показан график оптимизации концентраций неприродных рибонуклеотидтрифосфатов по белковому сдвигу (%) как функции концентраций NaMTP и TAT1TP (мкМ). Планками погрешностей указана стандартная ошибка для каждого значения (n=3).
[00228] На фиг. 6D показан график оптимизации концентраций неприродных рибонуклеотидтрифосфатов по белковому сдвигу (%)как функции концентраций 5FMTP и TAT1TP (мкМ). Планками погрешностей указана стандартная ошибка для каждого значения (n=3).
[00229] На фиг.7A показан график хранения и извлечения неприродной информации более высокой плотности для различных неприродных пар оснований, положений кодонов и общей флуоресценции sfGFP (RFU), наблюдаемой в присутствии AzK. В случае ленточных диаграмм каждый столбик соответствует среднему значению, при этом планками погрешностей указана стандартная ошибка (n=4), и незакрашенными кругами указаны данные для каждого независимого исследования.
[00230] На фиг. 7Ba показан график хранения и извлечения неприродной информации более высокой плотности для различных неприродный пар оснований, положений кодонов и белкового сдвига (%), измеряемого посредством вестерн-блоттинга, наблюдаемого в присутствии AzK. В случае ленточных диаграмм каждый столбик соответствует среднему значению, при этом планками погрешностей указана стандартная ошибка (n=4), и незакрашенными кругами указаны данные для каждого независимого исследования.
[00231] На фиг.7C показан типичный спектр количественного анализа HRMS трижды меченого белка, продуцируемого с использованием dCNMOdTPT3/NaMTP, TAT1TP. Метки пика демонстрируют молекулярную массу интактного белка после деконволюции, при этом показаны аминокислотные остатки в положениях 149, 151 и 153, а ниже показан количественный анализ каждого пика (%, n=3).
[00232] На фиг. 8A показаны примеры неприродных аминокислот. Эта фигура адаптирована с фиг. 2 из Young et al., "Beyond the canonical 20 amino acids: expanding the genetic lexicon," J. of Biological Chemistry 285(15): 11039-11044 (2010).
[00233] На фиг. 8B показаны примеры неприродных производных аминокислоты лизина.
[00234] На фиг. 8C показаны примеры неприродных производных аминокислоты фенилаланина.
[00235] На фиг. 8D-8G показаны примеры неприродных аминокислот. Эти неприродные аминокислоты (UAA) генетически закодированы в белках (фиг. 8D - UAA #1-42; фиг. 8E - UAA # 43-89; фиг. 8F - UAA # 90-128; фиг. 8G - UAA # 129-167). Фиг. 8D-8G адаптированы из таблицы 1 Dumas et al., Chemical Science 2015, 6, 50-69.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Некоторая терминология
[00236] Если не указано иначе, все технические и научные термины, используемые в настоящем описании, обладают тем же значением, которое общепринято понятно специалисту в области, к которой принадлежит изобретение. Следует понимать, что следующее общее описание и подробное описание являются исключительно иллюстративными и пояснительными, и не ограничивают настоящее изобретение каким-либо образом. Если какой-либо материал, включенный в настоящее описание в качестве ссылки, противоречит содержанию настоящего описания, содержание настоящего описания обладает приоритетом. В настоящей заявке использование термина в единственном числе включает множественное число, если конкретно не указано иное. Необходимо отметить, что, как используют в описании и формуле изобретения, формы в единственном числе включают ссылку на множественное число, если контекст четко не указывает на иное. В настоящей заявке использование термина "или" означает "и/или", если не указано иначе. Кроме того, использование термина "включающий", а также других форм, таких как "включают", "включает" и "включенный", не является ограничивающим.
[00237] В рамках изобретения диапазоны и количества можно выражать как "приблизительно" конкретное значение или диапазон. Термин "приблизительно" также включает точное количество. Таким образом "приблизительно 5 мкл" означает "приблизительно 5 мкл", и также включает описание "5 мкл". Как правило, термин "приблизительно" включает количество, которое, как будут ожидать, будет находиться в пределах экспериментальной ошибки.
[00238] Фразы, такие как "в условиях, подходящих для обеспечения" или "в условиях, достаточных для выхода" или т.п., в отношении способов синтеза в рамках изобретения относятся к условиям реакции, таким как время, температура, растворитель, концентрации реагентов и т.п., которые экспериментатор может варьировать в пределах своих навыков, обеспечивающим благоприятное количество или выход продукта реакции. Необязательно, чтобы желаемый продукт реакции являлся единственным продуктом реакции, или чтобы исходные материалы расходовались полностью, при условии, что желаемый продукт реакции можно выделять или иначе использовать впоследствии.
[00239] Термин "химически осуществимый" означает расположение связей или соединение, где общепринято понятные правила органической структуры не нарушены; например, структура по определению пункта формулы, которая в некоторых случаях будет содержать пятивалентный атом углерода, который не будет существовать в природе, будут понимать невходящим в пункт формулы. Структуры, представленные в настоящем описании, во всех вариантах осуществления предназначены для включения только "химически осуществимых" структур, и любые перечисленные структуры, не являющиеся химически осуществимыми, например, в структуре, показанной с переменными атомами или группами, не предназначены для того, чтобы быть представленными или заявленными в настоящем описании.
[00240] В рамках изобретения термин "аналог" химической структуры относится к химической структуре, сохраняющей значительное сходство с родительской структурой, хотя она может не быть полученной синтетически из родительской структуры. В некоторых вариантах осуществления аналог нуклеотида является неприродным нуклеотидом. В некоторых вариантах осуществления аналог нуклеозида является неприродным нуклеозидом. Родственную химическую структуру, легко получаемую синтетически из родительской химической структуры, обозначают как "производное".
[00241] В рамках изобретения термин "основание" или "нуклеиновое основание" относится к по меньшей мере части нуклеинового основания нуклеозида или нуклеотида (нуклеозид и нуклеотид включают рибо- или дезоксирибо-варианты), которые в некоторых случаях могут содержать дополнительные модификации в сахарной части нуклеозида или нуклеотида. В некоторых случаях термин "основание" также используют для обозначения целого нуклеозида или нуклеотида (например, "основание" может встраиваться ДНК-полимеразой в ДНК или РНК-полимеразой в РНК). Однако, термин "основание" не следует интерпретировать как обязательно означающий целый нуклеозид или нуклеотид, если контекст четко не указывает на иное. В химических структурах основания или нуклеинового основания, представленных в настоящем описании, показано только основание нуклеозида или нуклеотида, при этом остаток сахара и, необязательно, любые остатки фосфата опущены для простоты. Как используют в химических структурах основания или нуклеинового основания, представленных в настоящем описании, волнистой линией указано соединение с нуклеозидом или нуклеотидом, в котором сахарная часть нуклеозида или нуклеотид могут быть дополнительно модифицированы. В некоторых вариантах осуществления волнистой линией указано присоединение основания или нуклеинового основания к сахарной части, такой как пентоза, нуклеозида или нуклеотида. В некоторых вариантах осуществления пентоза является рибозой или дезоксирибозой.
[00242] В некоторых вариантах осуществления нуклеиновое основание, как правило, является гетероциклическим основанием нуклеозида. Нуклеиновые основания могут являться природными, могут являться модифицированными, могут не иметь сходства с природными основаниями, и/или их можно синтезировать, например, посредством органического синтеза. В некоторых вариантах осуществления нуклеиновое основание содержит любой атом или группу атомов в нуклеозиде или нуклеотиде, где атом или группа атомов могут взаимодействовать с основанием другой нуклеиновой кислоты с использованием водородных связей или без них. В некоторых вариантах осуществления неприродное нуклеиновое основание не получают из природного нуклеинового основания. Следует отметить, что неприродные нуклеиновые основания могут не обладать основными свойствами, однако, их обозначают как нуклеиновые основания для простоты. В некоторых вариантах осуществления, касающихся нуклеинового основания, "(d)" означает, что нуклеиновое основание можно присоединять к дезоксирибозе или рибозе, в то время как "d" без скобок означает, что нуклеиновое основание присоединено к дезоксирибозе.
[00243] В некоторых вариантах осуществления нуклеозид является соединением, содержащим остаток нуклеинового основания и остаток сахара. Нуклеозиды включают, в качестве неограничивающих примеров, природные нуклеозиды (обнаруживаемые в ДНК и РНК), абазические нуклеозиды, модифицированные нуклеозиды и нуклеозиды, содержащие миметики оснований и/или сахарных групп. Нуклеозиды включают нуклеозиды, содержащие любой спектр заместителей. Нуклеозид может являться гликозидным соединением, образованным с помощью гликозидной связи между основанием нуклеиновой кислоты и восстанавливающей группой сахара.
[00244] В рамках изобретения термин "нуклеотид" относится к соединению, содержащему остаток нуклеозида и фосфатный остаток. Неограничивающие примеры природных нуклеотидов включают аденозинтрифосфат (АТФ), уридинтрифосфат (УТФ), цитидинтрифосфат (ЦТФ), гуанозинтрифосфат (ГТФ), аденозиндифосфат (АДФ), уридиндифосфат (УДФ), цитидиндифосфат (ЦДФ), гуанозиндифосфат (ГДФ), аденозинмонофосфат (АМФ), уридинмонофосфат (УМФ), цитидинмонофосфат (ЦМФ) и гуанозинмонофосфат (ГМФ), дезоксиаденозинтрифосфат (dATP), дезокситимидинтрифосфат (dTTP), дезоксицитидинтрифосфат (dCTP), дезоксигуанозинтрифосфат(dGTP), дезоксиаденозиндифосфат (dADP), тимидиндифосфат (dTDP), дезоксицитидиндифосфат (dCDP), дезоксигуанозиндифосфат (dGDP), дезоксиаденозинмонофосфат (dAMP), дезокситимидинмонофосфат (dTMP), дезоксицитидинмонофосфат (dCMP) и дезоксигуанозинмонофосфат (dGMP). Примеры природных дезоксирибонуклеотидов, содержащие дезоксирибозу в качестве остатка сахара, включают dATP, dTTP, dCTP, dGTP, dADP, dTDP, dCDP, dGDP, dAMP, dTMP, dCMP и dGMP. Примеры природных рибонуклеотидов, содержащие рибозу в качестве остатка сахара, включают АТФ, УТФ, ЦТФ, ГТФ, АДФ, УДФ, ЦДФ, ГДФ, АМФ, УМФ, ЦМФ и ГМФ.
[00245] В рамках изобретения термин "полинуклеотид" относится к ДНК, РНК, ДНК- или РНК-подобным полимерам, таким как пептид нуклеиновые кислоты (ПНК), замкнутые нуклеиновые кислоты (ЗНК), фосфотиоаты и т.п., примеры которых хорошо известны в этой области и могут содержать неприродные основания. Полинуклеотиды можно синтезировать с помощью автоматизированных синтезаторов, например, с использованием фосфорамидатной химии или других химических подходов, адаптированных для использования с помощью синтезатора.
[00246] ДНК включает, в качестве неограничивающих примеров, комплементарную ДНК (кДНК) и геномную ДНК (gDNA). ДНК можно присоединять ковалентными или нековалентными средствами с другой молекулой, включая, в качестве неограничивающих примеров, РНК и пептид. РНК включает кодирующую РНК, например, матричную РНК (мРНК). РНК также включает некодирующую РНК, например, рибосомальную РНК (рРНК). РНК также включает транспортную РНК (тРНК), интерферирующую РНК (РНКи), малую ядрышковую РНК (snoRNA), микроРНК (мкРНК), малую интерферирующую РНК (миРНК), малую ядерную РНК (мяРНК), внеклеточную РНК (exRNA), PIWI-взаимодействующую РНК (piRNA) и длинную некодирующую РНК (длинную ncRNA). В некоторых вариантах осуществления РНК является рРНК, тРНК, РНКи, snoRNA, микроРНК, миРНК, мяРНК, exRNA, piRNA, длинной ncRNA или любой их комбинацией или гибридом. В некоторых случаях РНК является компонентом рибозима. ДНК и РНК могут находиться в любой форме, включая, в качестве неограничивающих примеров, линейную, кольцевую, сверхспиральную, одноцепочечную и двухцепочечную.
[00247] Пептид-нуклеиновая кислота (ПНК) является синтетическим аналогом ДНК/РНК, где пептид-подобным остовом заменяют сахар-фосфатный остов ДНК или РНК. Олигомеры ПНК демонстрируют более сильное связывание и более высокую специфичность связывания с комплементарной ДНК, при этом ошибочное спаривание оснований ПНК/ДНК является более дестабилизирующим, чем схожее ошибочное спаривание в дуплексе ДНК/ДНК. Эта сила и специфичность связывания также относятся к дуплексам ПНК/РНК. ПНК не распознаются легко нуклеазами или протеазами, что делает их более резистентными к деградации ферментами. ПНК также являются стабильными в широком диапазоне pH. Также см. Nielsen PE, Egholm M, Berg RH, Buchardt O (December 1991). "Sequence-selective recognition of DNA by strand displacement with thymine-substituted polyamide", Science254 (5037): 1497-500. doi:10/1126/science.1962210. PMID 1962210; и, Egholm M, Buchardt O, Christensen L, Behrens C, Freier SM, Driver DA, Berg RH, Kim SK, Nordén B, and Nielsen PE (1993), "PNA Hybridizes to Complementary Oligonucleotides Obeying the Watson-Crick Hydrogen Bonding Rules". Nature365 (6446): 566-8. doi:10.1038/365566a0. PMID 7692304; описание каждой из которой включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00248] Замкнутая нуклеиновая кислота (ЗНК) представляет собой модифицированный нуклеотид РНК, где остаток рибозы нуклеотида ЗНК модифицирован с использованием дополнительного мостика, соединяющего 2'-кислород и 4'-углерод. Мостик "замыкает" рибозу в 3'-эндо (северной) конформации, часто обнаруживаемой в дуплексах A-формы. ЗНК-нуклеотиды, при желании? можно смешивать с остатками ДНК или РНК в олигонуклеотиде. Такие олигомеры можно синтезировать химически, и они являются коммерчески доступными. Замкнутая конформация рибозы повышает стэкинг-взаимодействие и предварительную организацию остова. См., например, Kaur, H; Arora, A; Wengel, J; Maiti, S (2006), "Thermodynamic, Counterion, and Hydration Effects for the Incorporation of Locked Nucleic Acid Nucleotides into DNA Duplexes", Biochemistry45 (23): 7347-55. doi:10.1021/bi060307w. PMID 16752924; Owczarzy R.; You Y., Groth C.L., Tataurov A.V. (2011), "Stability and mismatch discrimination of locked nucleic acid-DNA duplexes.", Biochem.50 (43): 9352-9367. doi:10.1021/bi200904e. PMC3201676. PMID 21928795; Alexei A. Koshkin; Sanjay K. Singh, Poul Nielsen, Vivek K. Rajwanshi, Ravindra Kumar, Michael Meldgaard, Carl Erik Olsen, Jesper Wengel (1998), "LNA (Locked Nucleic Acids): Synthesis of the adenine, cytosine, guanine, 5-methylcytosine, thymine and uracil bicyclonucleoside monomers, oligomerisation, and unprecedented nucleic acid recognition", Tetrahedron54 (14): 3607-30. doi:10,1016/S0040-4020(98)00094-5; и, Satoshi Obika; Daishu Nanbu, Yoshiyuki Hari, Ken-ichiro Morio, Yasuko In, Toshimasa Ishida, Takeshi Imanishi (1997), "Synthesis of 2′-O,4′-C-methyleneuridine and -cytidine. Novel bicyclic nucleosides having a fixed C3'-endo sugar puckering", Tetrahedron Lett.38 (50): 8735-8. doi:10,1016/S0040-4039(97)10322-7; содержание каждой из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00249] В рамках изобретения термин "ген" относится к полинуклеотиду, кодирующему синтез продукта гена, такого как РНК или белка.
[00250] Молекулярный маяк или зонд "молекулярный маяк" является олигонуклеотидным зондом для гибридизации, с помощью которого можно определять наличие специфической последовательности нуклеиновой кислоты в гомогенном растворе. Молекулярные маяки являются шпилечными молекулами с изнутри гасимым флуорофором, флуоресценция которого восстанавливается, когда они связываются с целевой последовательностью нуклеиновой кислоты. См., например, Tyagi S, Kramer FR (1996), "Molecular beacons: probes that fluoresce upon hybridization", Nat Biotechnol.14 (3): 303-8. PMID 9630890; Täpp I, Malmberg L, Rennel E, Wik M, Syvänen AC (2000 Apr), "Homogeneous scoring of single-nucleotide polymorphisms: comparison of the 5'-nuclease TaqMan assay and Molecular Beacon probes", Biotechniques28 (4): 732-8. PMID 10769752; и Akimitsu Okamoto (2011), "ECHO probes: a concept of fluorescence control for practical nucleic acid sensing", Chem. Soc. Rev.40: 5815-5828; описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00251] В рамках изобретения термин "неприродное основание" относится к иному основанию, чем A, C, G, T, U и другие природные основания (например, 5-метилцитозин, псевдоуридин и инозин).
[00252] В рамках изобретения термин "неприродная пара оснований" относится к двум основаниям, связанным друг с другом и лежащим на противоположных цепях двухцепочечного полинуклеотида (который может являться, например, по меньшей мере частично самогибридизующейся молекулой или частично или полностью гибридизованной парой молекул), где по меньшей мере одно из двух оснований является неприродным основанием.
[00253] В рамках изобретения "полусинтетический организм" является организмом, содержащим неприродный компонент, например, расширенный генетический алфавит, включающий одно или более неприродных оснований.
[00254] Заголовки разделов, используемые в настоящем описании, предназначены исключительно для организационных целей, и не следует истолковывать их как ограничения описываемого изобретения.
Способы и композиции, содержащие неприродные пары оснований
[00255] В некоторых вариантах осуществления настоящее изобретение относится к in vitro и in vivo способам и композициям для получения нуклеиновой кислоты с расширенным генетическим алфавитом (фиг. 1). В некоторых случаях нуклеиновая кислота кодирует неприродный белок, где неприродный белок содержит неприродную аминокислоту. В некоторых случаях в способе или композиции in vivo, представленных в настоящем описании, используют полусинтетический организм, или они включают его. В некоторых случаях способ включает встраивание по меньшей мере одной неприродной пары оснований (UBP) в одну или более нуклеиновых кислот. Такая пара оснований образуется посредством спаривания между нуклеиновыми основаниями двух нуклеозидов. На примере схемы ДНК 101, кодирующая белок 102 и тРНК 103, кодирующие области матричной цепи для белка и тРНК, содержащие неприродные нуклеиновые основания (X, Y), являющиеся комплементарными, способными образовывать пару оснований и/или сконфигурированными для образования пары оснований, транскрибируется 104 с образованием тРНК 106 и мРНК 107. После зарядки тРНК неприродной аминокислотой 105, мРНК 107 транслируется 108 с образованием белка 110, содержащего одну или более неприродных аминокислот 109. Способы и композиции, представленные в настоящем описании, в некоторых случаях делают возможным сайт-специфическое встраивание неприродных аминокислот с высокой точностью и выходом. Настоящее изобретение также относится к полусинтетическим организмам, содержащим расширенный генетический алфавит, и способам использования полусинтетических организмов для получения белковых продуктов, включая продукты, содержащие по меньшей мере один неприродный аминокислотный остаток.
[00256] Выбор неприродных нуклеиновых оснований делает возможной оптимизацию одной или более стадий способов, представленных в настоящем описании. Например, нуклеиновые основания выбирают по высокоэффективной репликации, транскрипции и/или трансляции. В некоторых случаях в способах, представленных в настоящем описании, используют более одной пары неприродных нуклеиновых оснований. Например, первый набор нуклеиновых оснований, содержащих дезоксирибо-остаток, используют для репликации ДНК (такой как первое нуклеиновое основание и второе нуклеиновое основание, сконфигурированные для образования первой пары оснований), и второй набор нуклеиновых оснований (такой как третье нуклеиновое основание и четвертое нуклеиновое основание, где третье и четвертое нуклеиновые основания присоединены к рибозе, сконфигурированные для образования второй пары оснований) используют для транскрипции/трансляции. В некоторых вариантах осуществления первый набор нуклеиновых оснований используют для конструирования плазмиды (такой как первое нуклеиновое основание и второе нуклеиновое основание, сконфигурированные для образования первой пары оснований), второй набор нуклеиновых оснований используют для репликации (такой как третье нуклеиновое основание и четвертое нуклеиновое основание, сконфигурированные для образования второй пары оснований), и третий набор оснований используют для транскрипции/трансляции (такой как пятое и шестое нуклеиновое основание, сконфигурированные для образования третьей пары оснований). В некоторых случаях комплементарное спаривание между нуклеиновым основанием из первого набора и нуклеиновым основанием из второго набора делает возможной транскрипцию генов для образования тРНК или белков с матрицы ДНК, содержащей нуклеиновые основания из первого набора. В некоторых случаях комплементарное спаривание между нуклеиновыми основаниями из второго набора (второй пары оснований) делает возможной трансляцию в результате совпадения тРНК, содержащих неприродные нуклеиновые кислоты, и мРНК. В некоторых случаях нуклеиновые основания в первом наборе присоединены к дезоксирибозному остатку. В некоторых случаях нуклеиновые основания в первом наборе присоединены к рибозному остатку. В некоторых случаях нуклеиновые основания из обоих наборов являются уникальными. В некоторых случаях по меньшей мере одно нуклеиновое основание является одним и тем же в обоих наборах. В некоторых случаях первое нуклеиновое основание и третье нуклеиновое основание являются одинаковыми. В некоторых вариантах осуществления первая пара оснований и вторая пара оснований не являются одинаковыми. В некоторых случаях первая пара оснований, вторая пара оснований и третья пара оснований не являются одинаковыми.
[00257] В одном из аспектов настоящее изобретение относится к способу получения in vivo белка, содержащего неприродную аминокислоту, включающему:
транскрипцию матрицы ДНК, содержащей первое неприродное основание и второе неприродное основание, являющееся комплементарным, способное образовывать пару оснований и/или сконфигурированное для образования пары оснований с первым неприродным основанием, для встраивания третьего неприродного основания в мРНК, при этом третье неприродное основание является комплементарным, может образовывать пару оснований и/или сконфигурировано для образования первой неприродной пары оснований с первым неприродным основанием;
транскрипцию матрицы ДНК для встраивания четвертого неприродного основания в тРНК, где четвертое неприродное основание является комплементарным, способным образовывать пару оснований и/или сконфигурировано для образования второй неприродной пары оснований со вторым неприродным основанием, где первая неприродная пара оснований и вторая неприродная пара оснований не являются одинаковыми; и
трансляцию белка с мРНК и тРНК, где указанный белок содержит неприродную аминокислоту.
Молекулы нуклеиновой кислоты
[00258] В некоторых вариантах осуществления нуклеиновая кислота (например, также обозначаемая в настоящем описании как интересующая молекула нуклеиновой кислоты) происходит из любого источника или композиции, таких как ДНК, кДНК, gDNA (геномная ДНК), РНК, миРНК (короткая ингибиторная РНК), РНКи, тРНК, мРНК или рРНК (рибосомальная РНК), например, и находится в любой форме (например, линейной, кольцевой, сверхспиральной, одноцепочечной, двухцепочечной и т.п.). В некоторых вариантах осуществления нуклеиновые кислоты содержат нуклеотиды, нуклеозиды или полинуклеотиды. В некоторых случаях нуклеиновые кислоты включают природные и неприродные нуклеиновые кислоты. В некоторых случаях нуклеиновая кислота также включает неприродные нуклеиновые кислоты, такие как аналоги ДНК или РНК (например, содержащие аналоги оснований, аналоги сахаров и/или ненативный остов и т.п.). Следует понимать, что термин "нуклеиновая кислота" не означает конкретную длину полинуклеотидной цепи, таким образом, полинуклеотиды и олигонуклеотиды также включены в это определение. Неограничивающие примеры природных нуклеотидов включают АТФ, УТФ, ЦТФ, ГТФ, АДФ, УДФ, ЦДФ, ГДФ, АМФ, УМФ, ЦМФ, ГМФ, dATP, dTTP, dCTP, dGTP, dADP, dTDP, dCDP, dGDP, dAMP, dTMP, dCMP и dGMP. Примеры природных дезоксирибонуклеотидов включают dATP, dTTP, dCTP, dGTP, dADP, dTDP, dCDP, dGDP, dAMP, dTMP, dCMP и dGMP. Примеры природных рибонуклеотидов включают АТФ, УТФ, ЦТФ, ГТФ, АДФ, УДФ, ЦДФ, ГДФ, АМФ, УМФ, ЦМФ и ГМФ. В случае природной РНК урацил-содержащим нуклеозидом является уридин. В некоторых случаях нуклеиновая кислота является вектором, плазмидой, фагмидой, автономно реплицирующейся последовательностью (ARS), центромерой, искусственной хромосомой, искусственной дрожжевой хромосомой (например, YAC) или другой нуклеиновой кислотой, способной реплицироваться или быть реплицированной в клетке-хозяине. В некоторых случаях неприродная нуклеиновая кислота является аналогом нуклеиновой кислоты. В дополнительных случаях неприродная нуклеиновая кислота получена из внеклеточного источника. В других случаях неприродная нуклеиновая кислота доступна для внутриклеточного пространства организма, представленного в настоящем описании, например, генетически модифицированного организма. В некоторых вариантах осуществления неприродный нуклеотид не является природным нуклеотидом. В некоторых вариантах осуществления нуклеотид, несодержащий природное основание, содержит неприродное нуклеиновое основание.
Неприродные нуклеиновые кислоты
[00259] Аналог нуклеотида или неприродный нуклеотид включает нуклеотид, содержащий некоторый тип модификации остатков основания, сахара или фосфата. Термин "модификация" (и родственные грамматические формы, такие как "модифицированный") не обязательно означает, что аналог нуклеотида или неприродный нуклеотид получают посредством прямого изменения природного нуклеотида, а скорее то, что аналог нуклеотида или неприродный нуклеотид отличается от природного нуклеотида. В некоторых вариантах осуществления модификация включает химическую модификацию. В некоторых случаях модификации происходят в 3'OΗ- или 5'OΗ-группе, остове, сахарном компоненте или основании нуклеотида. В некоторых случаях модификации, необязательно, включают неприродные линкерные молекулы и/или межцепочечные или внутрицепочечные перекрестные связи. В одном из аспектов модифицированная нуклеиновая кислота содержит модификацию одного или более из 3'OΗ- или 5'OΗ-группы, остова, сахарного компонента или основания нуклеотида и/или добавление неприродных линкерных молекул. В одном из аспектов модифицированный остов включает иной остов, чем фосфодиэфирный остов. В одном из аспектов модифицированный сахар включает иной сахар, чем дезоксирибоза (в модифицированной ДНК) или рибоза (в модифицированной РНК). В одном из аспектов модифицированное основание включает иное основание, чем аденин, гуанин, цитозин или тимин (в модифицированной ДНК) или иное основание, чем аденин, гуанин, цитозин или урацил (в модифицированной РНК). В некоторых вариантах осуществления неприродный нуклеотид включает неприродное основание. В некоторых вариантах осуществления неприродное основание является основанием с кольцом или кольцевой системой, иной, чем пурин или пиримидин (где пурин и пиримидин включает пурины и пиримидины с экзоциклическими заместителями), или содержит кольцо или кольцевую систему, содержащую один или более неазотных гетероатомов и/или несодержащую атомы азота.
[00260] В некоторых вариантах осуществления нуклеиновая кислота содержит по меньшей мере одно модифицированное основание. В некоторых случаях нуклеиновая кислота содержит 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20 или более модифицированных оснований. В некоторых случаях модификации остатка оснований включают природные и синтетические модификации аденина (A), цитозина (C), гуанина (G) и тимина (T)/урацила (U), а также другие пуриновые или пиримидиновые основания. В некоторых вариантах осуществления модификация приводит к модифицированной форме аденина, гуанина, цитозина или тимина (в модифицированной ДНК) или модифицированной форме аденина, гуанина, цитозина или урацила (в модифицированной РНК).
[00261] Модифицированное основание неприродной нуклеиновой кислоты включает, в качестве неограничивающих примеров, урацил-5-ил, гипоксантин-9-ил (I), 2-аминоаденин-9-ил, 5-метилцитозин (5-me-C), 5-гидроксиметилцитозин, ксантин, гипоксантин, 2-аминоаденин, 6-метильные и другие алкильные производные аденина и гуанина, 2-пропильные и другие алкильные производные аденина и гуанина, 2-тиоурацил, 2-тиотимин и 2-тиоцитозин, 5-галоурацил и -цитозин, 5-пропинилурацил и -цитозин, 6-азоурацил, -цитозин и -тимин, 5-урацил (псевдоурацил), 4-тиоурацил, 8-галоген-, 8-амино-, 8-тиол-, 8-тиоалкил-, 8-гидроксил- и другие 8-замещенные аденины и гуанины, 5-галоген-, в частности 5-бром-, 5-трифторметил- и другие 5-замещенные урацилы и цитозины, 7-метилгуанин и 7-метиладенин, 8-азагуанин и 8-азааденин, 7-деазагуанин и 7-деазааденин и 3-деазагуанин и 3-деазааденин. Некоторые неприродные нуклеиновые кислоты, такие как 5-замещенные пиримидины, 6-азапиримидины и N-2-замещенные пурины, N-6-замещенные пурины, O-6-замешенные пурины, 2-аминопропиладенин, 5-пропинилурацил, 5-пропинилцитозин, 5-метилцитозин, соединения, повышающие стабильность образования дуплекса, универсальные нуклеиновые кислоты, гидрофобные нуклеиновые кислоты, смешанные нуклеиновые кислоты, расширенные нуклеиновые кислоты, фторированные нуклеиновые кислоты, 5-замещенные пиримидины, 6-азапиримидины и N-2-, N-6- и O-6-замещенные пурины, включая 2-аминопропиладенин, 5-пропинилурацил и 5-пропинилцитозин. 5-метилцитозин (5-me-C), 5-гидроксиметилцитозин, ксантин, гипоксантин, 2-аминоаденин, 6-метильные и другие алкильные производные аденина и гуанина, 2-пропильные и другие алкильные производные аденина и гуанина, 2-тиоурацил, 2-тиотимин и 2-тиоцитозин, 5-галоурацил, 5-галоцитозин, 5-пропинил (-C≡C-CH3)-урацил, 5-пропинилцитозин, другие алкинильные производные пиримидиновых нуклеиновых кислот, 6-азоурацил, 6-азоцитозин, 6-азотимин, 5-урацил (псевдоурацил), 4-тиоурацил, 8-галоген-, 8-амино-, 8-тиол-, 8-тиоалкил-, 8-гидроксил- и другие 8-замещенные аденины и гуанины, 5-галоген-, в частности, 5-бром-, 5-трифторметил-, другие 5-замещенные урацилы и цитозины, 7-метилгуанин, 7-метиладенин, 2-F-аденин, 2-амино-аденин, 8-азагуанин, 8-азааденин, 7-деазагуанин, 7-деазааденин, 3-деазагуанин, 3-деазааденин, трициклические пиримидины, феноксазина цитидин([5,4-b][1,4]бензоксазин-2(3H)-он), фенотиазина цитидин(1H-пиримидо[5,4-b][1,4]бензотиазин-2(3H)-он), гуанин-фиксирующие основания, феноксазина цитидин (например, 9-(2-аминоэтокси)-H-пиримидо[5,4-b][1,4]бензоксазин-2(3H)-он), карбазолцитидин(2H-пиримидо[4,5-b]индол-2-он), пиридоиндолцитидин(H-пиридо[3',2':4,5]пирроло[2,3-d]пиримидин-2-он), соединения, в которых пуриновое или пиримидиновое основание заменяют другими гетероциклами, 7-деаза-аденин, 7-деазагуанозин, 2-аминопиридин, 2-пиридон, азацитозин, 5-бромцитозин, бромурацил, 5-хлорцитозин, хлорированный цитозин, циклоцитозин, цитозинарабинозид, 5-фторцитозин, фторпиримидин, фторурацил, 5,6-дигидроцитозин, 5-йодцитозин, гидроксимочевина, йодурацил, 5-нитроцитозин, 5-бромурацил, 5-хлорурацил, 5-фторурацил и 5-йодурацил, 2-амино-аденин, 6-тио-гуанин, 2-тио-тимин, 4-тио-тимин, 5-пропинил-урацил, 4-тио-урацил, N4-этилцитозин, 7-деазагуанин, 7-деаза-8-азагуанин, 5-гидроксицитозин, 2'-дезоксиуридин, 2-амино-2'-дезоксиаденозин и соединения, описанные в патентах США №№ 3687808; 4845205; 4910300; 4948882; 5093232; 5130302; 5134066; 5175273; 5367066; 5432272; 5457187; 5459255; 5484908; 5502177; 5525711; 5552540; 5587469; 5594121; 5596091; 5614617; 5645985; 5681941; 5750692; 5763588; 5830653 и 6005096; WO 99/62923; Kandimalla et al., (2001) Bioorg. Med. Chem. 9:807-813; The Concise Encyclopedia of Polymer Science and Engineering, Kroschwitz, J.I., Ed., John Wiley & Sons, 1990, 858-859; Englisch et al., Angewandte Chemie, International Edition, 1991, 30, 613; и Sanghvi, Chapter 15, Antisense Research and Applications, Crookeand Lebleu Eds., CRC Press, 1993, 273-288. Дополнительные модификации оснований можно найти, например, в патенте США № 3687808; Englisch et al., Angewandte Chemie, International Edition, 1991, 30, 613. В некоторых случаях неприродная нуклеиновая кислота содержит нуклеиновое основание с фиг. 2. В некоторых случаях неприродная нуклеиновая кислота содержит нуклеиновое основание с фиг. 4A. В некоторых случаях неприродная нуклеиновая кислота содержит нуклеиновое основание с фиг. 4B.
[00262] В этой области доступны неприродные нуклеиновые кислоты, содержащие различные гетероциклические основания и различные остатки сахаров (и аналоги сахаров), и в некоторых случаях нуклеиновая кислота включает одно или более гетероциклических оснований, иных, чем главные пять оснований-компонентов природных нуклеиновых кислот. Например, в некоторых случаях гетероциклическое основание включает группы урацил-5-ил, цитозин-5-ил, аденин-7-ил, аденин-8-ил, гуанин-7-ил, гуанин-8-ил, 4-аминопирроло[2,3-d] пиримидин-5-ил, 2-амино-4-оксопироло [2,3-d]пиримидин-5-ил, 2-амино-4-оксопирроло[2,3-d]пиримидин-3-ил, где пурины присоединены к сахарному остатку нуклеиновой кислоты через 9-положение, пиримидины - через 1-положение, пирролопиримидины - через 7-положение, и пиразолопиримидины - через 1-положение.
[00263] В некоторых вариантах осуществления модифицированное основание неприродной нуклеиновой кислоты изображено ниже, где волнистой линией указана точка присоединения к сахару нуклеозида или нуклеотида (например, дезоксирибозе или рибозе).
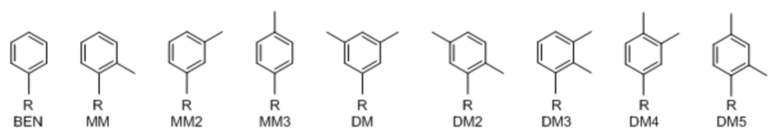
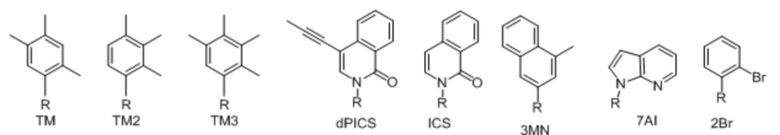
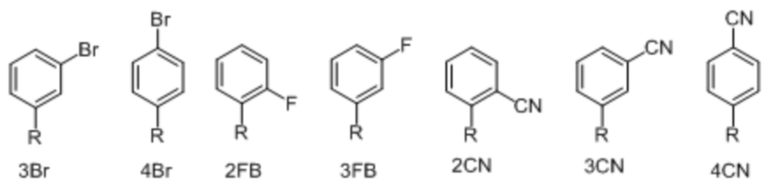
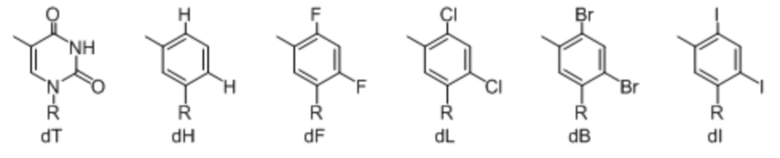
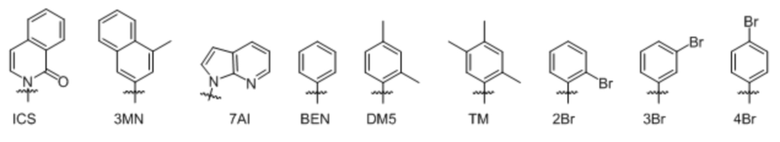
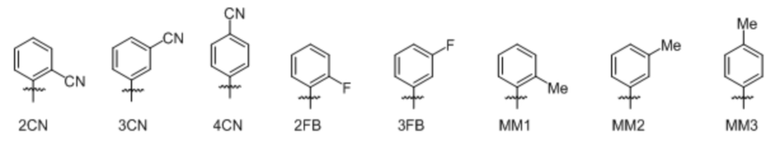
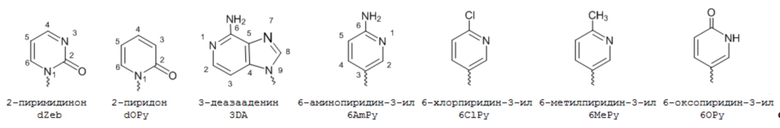
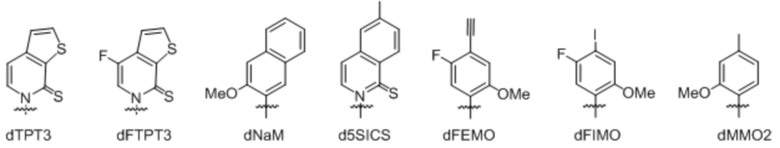
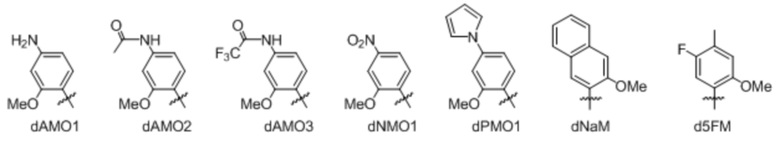
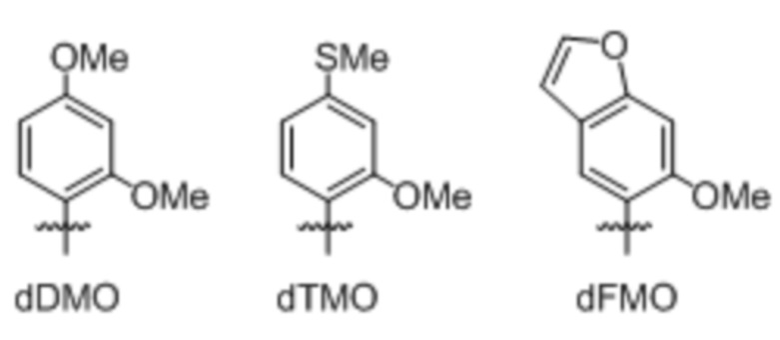
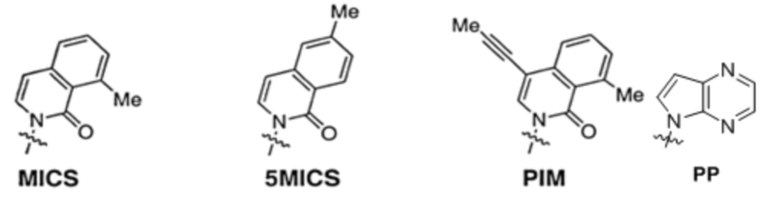
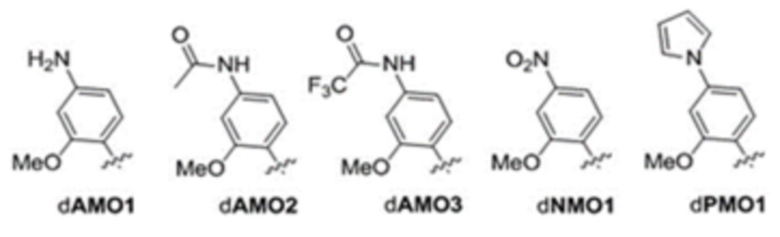
 (NaM),
(NaM),  (CNMO),
(CNMO),  (MMO2),
(MMO2),  (5FM),
(5FM),  (2OMe),
(2OMe),  (5F2OMe),
(5F2OMe),  (ClMO),
(ClMO),  (BrMO),
(BrMO),  (PTMO),
(PTMO),  (MTMO),
(MTMO),  (TPT3),
(TPT3),  (SICS),
(SICS),  (FSICS),
(FSICS),  ,
,  (TAT1), (dNaM), (dCNMO), (dMMO2), (d5FM), (d2OMe), (d5F2OMe), (dClMO), (dBrMO), (dPTMO), (dMTMO), (dTPT3), (dSICS), (dFSICS), и (dTAT1).
(TAT1), (dNaM), (dCNMO), (dMMO2), (d5FM), (d2OMe), (d5F2OMe), (dClMO), (dBrMO), (dPTMO), (dMTMO), (dTPT3), (dSICS), (dFSICS), и (dTAT1).
[00264] В некоторых вариантах осуществления неприродное основание (такое как по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания из способа получения белка, содержащего неприродную аминокислоту, представленного в настоящем описании) выбрано из группы, состоящей из:
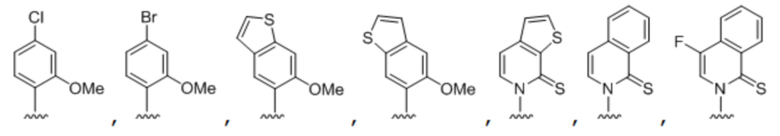
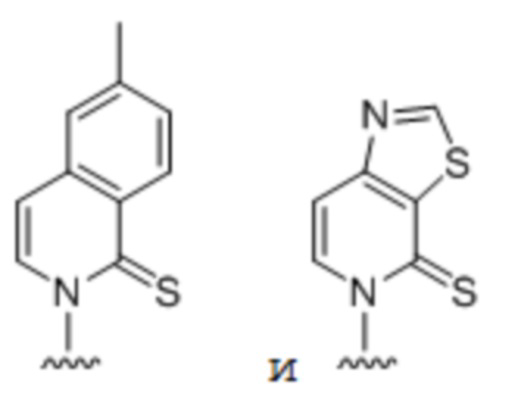 . В некоторых вариантах осуществления неприродное основание выбрано из группы, состоящей из: , , и . В некоторых вариантах осуществления неприродное основание выбрано из группы, состоящей из: и . В некоторых вариантах осуществления неприродное основание выбрано из группы, состоящей из
. В некоторых вариантах осуществления неприродное основание выбрано из группы, состоящей из: , , и . В некоторых вариантах осуществления неприродное основание выбрано из группы, состоящей из: и . В некоторых вариантах осуществления неприродное основание выбрано из группы, состоящей из
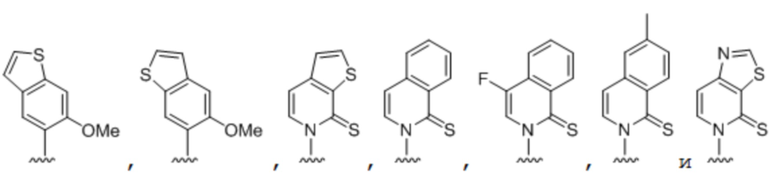 . В некоторых вариантах осуществления неприродное основание представляет собой . В некоторых вариантах осуществления неприродное основание выбрано из:
. В некоторых вариантах осуществления неприродное основание представляет собой . В некоторых вариантах осуществления неприродное основание выбрано из:
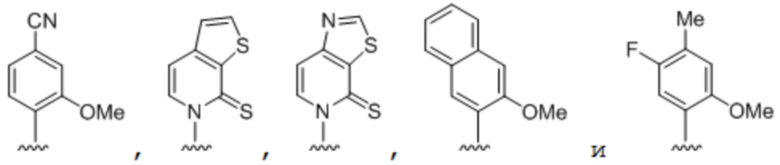 .
.
[00265] В некоторых вариантах осуществления неприродное основание (такое как первое или второе неприродное основание из способа получения белка, содержащего неприродную аминокислоту, представленного в настоящем описании) представляет собой . В некоторых вариантах осуществления неприродное основание (такое как первое или второе неприродное основание из способа получения белка, содержащего неприродную аминокислоту, представленного в настоящем описании) представляет собой . В некоторых вариантах осуществления первое неприродное основание представляет собой , и второе неприродное основание представляет собой .
[00266] В некоторых вариантах осуществления неприродное основание (такое как третье или четвертое неприродное основание из способа получения белка, содержащего неприродную аминокислоту, представленного в настоящем описании) представляет собой . В некоторых вариантах осуществления третье неприродное основание представляет собой . В некоторых вариантах осуществления четвертое неприродное основание представляет собой . В некоторых вариантах осуществления неприродное основание (такое как третье или четвертое неприродное основание из способа получения белка, содержащего неприродную аминокислоту, представленного в настоящем описании) представляет собой или . В некоторых вариантах осуществления третье неприродное основание представляет собой или . В некоторых вариантах осуществления четвертое неприродное основание представляет собой или .
[00267] В некоторых вариантах осуществления первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой , и четвертое неприродное основание представляет собой или . В некоторых вариантах осуществления первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой , и четвертое неприродное основание представляет собой или . В некоторых вариантах осуществления первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой или , и четвертое неприродное основание представляет собой . В некоторых вариантах осуществления третье неприродное основание представляет собой или . В некоторых вариантах осуществления четвертое неприродное основание представляет собой . В некоторых вариантах осуществления первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой или , и четвертое неприродное основание представляет собой .
[00268] В некоторых вариантах осуществления третье неприродное основание и четвертое неприродное основание содержат рибозу. В некоторых вариантах осуществления третье неприродное основание и четвертое неприродное основание содержат дезоксирибозу. В некоторых вариантах осуществления первое и второе неприродные основания содержат дезоксирибозу. В некоторых вариантах осуществления первое и второе неприродные основания содержат дезоксирибозу, и третье неприродное основание и четвертое неприродное основание содержат рибозу.
[00269] В некоторых вариантах осуществления способа получения белка, содержащего неприродную аминокислоту, представленного в настоящем описании, ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из:
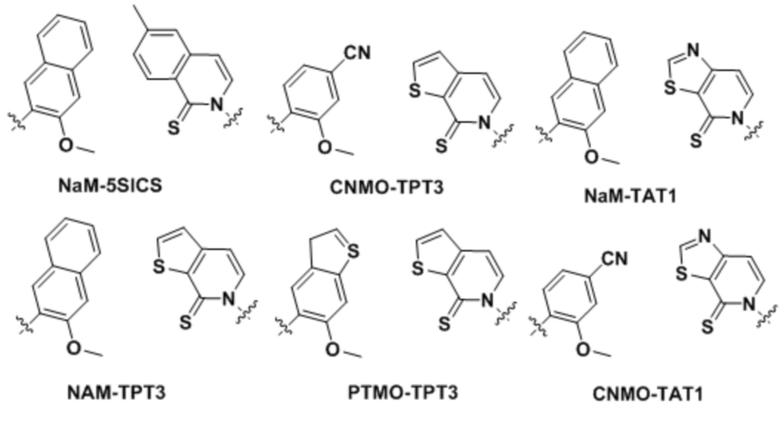 ,
,
где каждый сахарный остаток независимо представляет собой любой вариант осуществления или вариант, представленный в настоящем описании. В некоторых вариантах осуществления сахарный остаток обоих оснований в паре оснований содержит рибозу. В некоторых вариантах осуществления сахарный остаток обоих оснований в паре оснований содержит дезоксирибозу. В некоторых вариантах осуществления сахарный остаток одного основания в паре оснований содержит рибозу, и сахарный остаток другого основания в паре оснований содержит дезоксирибозу. В некоторых вариантах осуществления матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся NaM-5SICS. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся CNMO-TPT3. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся NaM-TPT3. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся NaM-TAT1.В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся CNMO-TAT1.
[00270] В некоторых вариантах осуществления способа получения белка, содержащего неприродную аминокислоту, представленного в настоящем описании, ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из группы, состоящей из
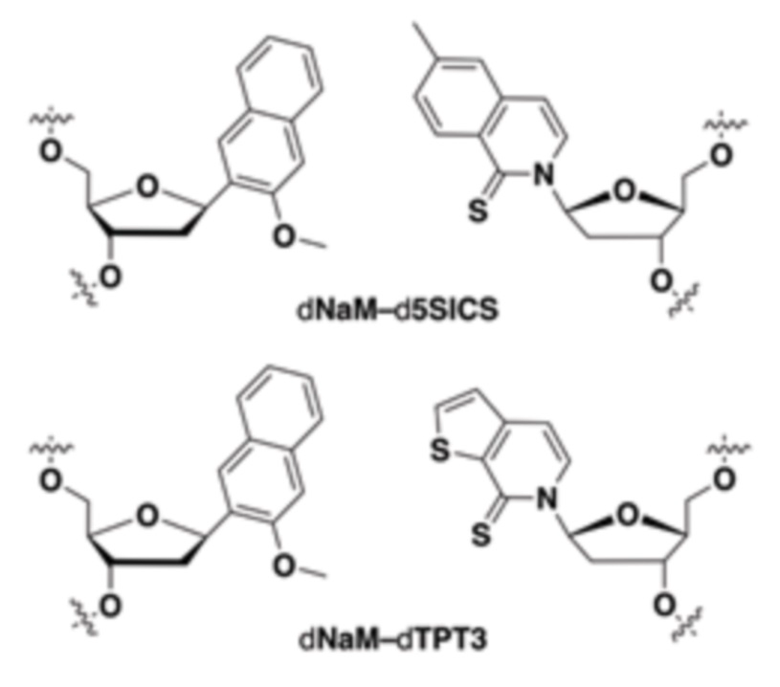
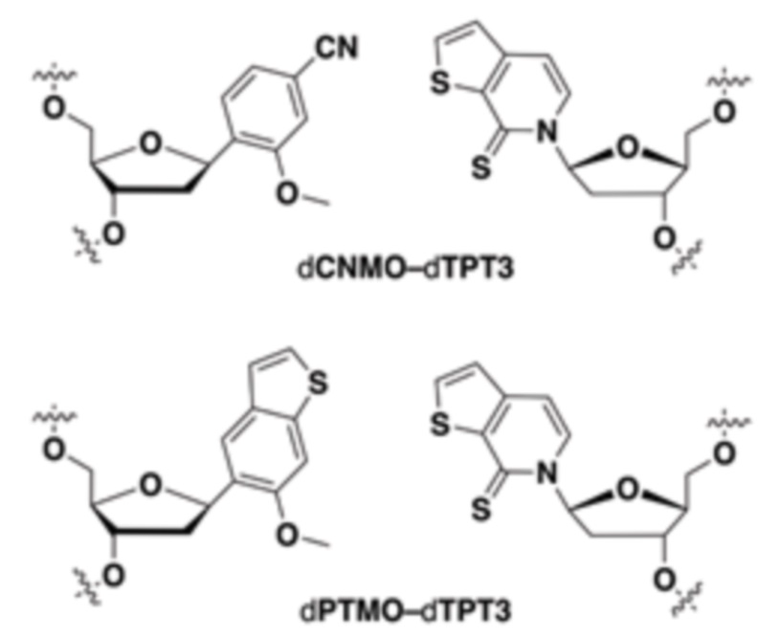
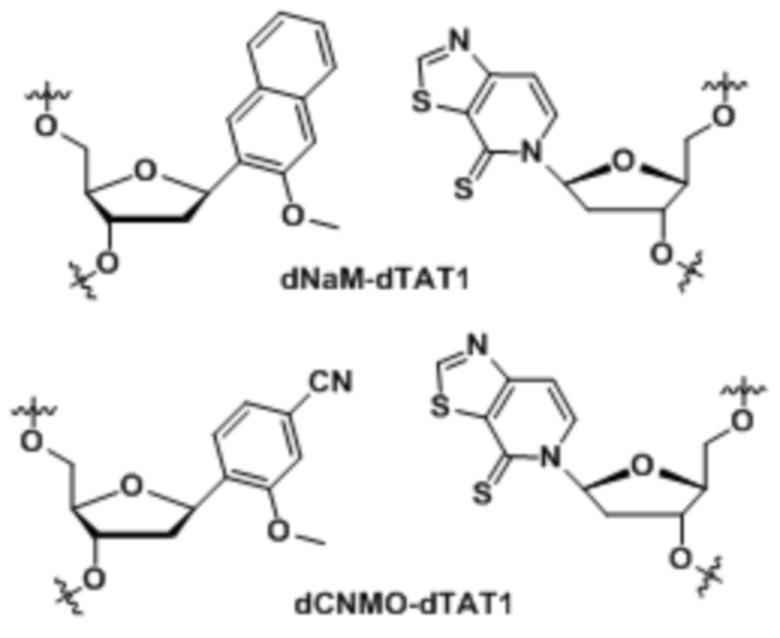 . В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTAT1.В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTAT1.
. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTAT1.В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTAT1.
[00271] В некоторых вариантах осуществления способа получения белка, содержащего неприродную аминокислоту, представленного в настоящем описании, ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из:
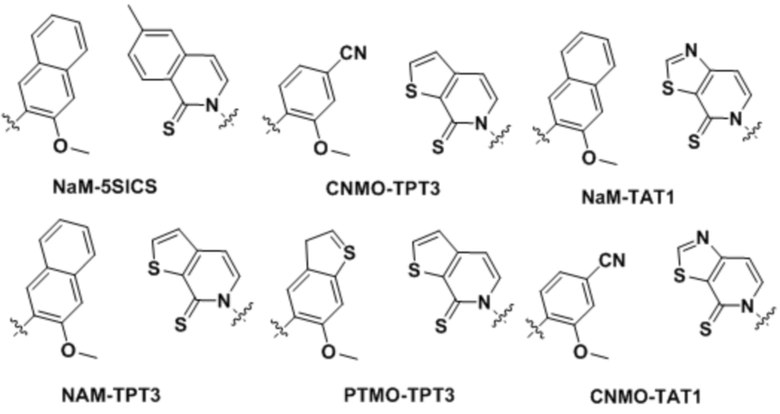 ,
,
где каждый сахарный остаток независимо представляет собой любой вариант осуществления или вариант, представленный в настоящем описании; и где мРНК и тРНК содержат по меньшей мере одно неприродное основание, выбранное из:
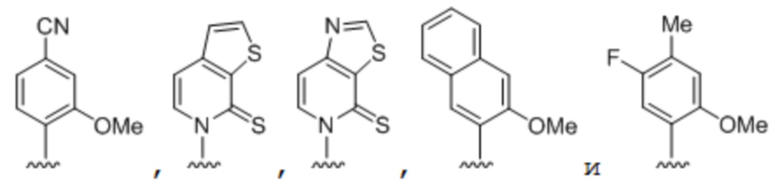 . В некоторых вариантах осуществления сахарный остаток обоих оснований в паре оснований содержит рибозу. В некоторых вариантах осуществления сахарный остаток обоих оснований в паре оснований содержит дезоксирибозу. В некоторых вариантах осуществления сахарный остаток одного основания в паре оснований содержит рибозу, и сахарный остаток другого основания в паре оснований содержит дезоксирибозу. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTAT1. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTAT1.
. В некоторых вариантах осуществления сахарный остаток обоих оснований в паре оснований содержит рибозу. В некоторых вариантах осуществления сахарный остаток обоих оснований в паре оснований содержит дезоксирибозу. В некоторых вариантах осуществления сахарный остаток одного основания в паре оснований содержит рибозу, и сахарный остаток другого основания в паре оснований содержит дезоксирибозу. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTAT1. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTAT1.
[00272] В некоторых вариантах осуществления способа получения белка, содержащего неприродную аминокислоту, представленного в настоящем описании, ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из группы, состоящей из
; и где мРНК и тРНК содержат по меньшей мере одно неприродное основание, выбранное из:
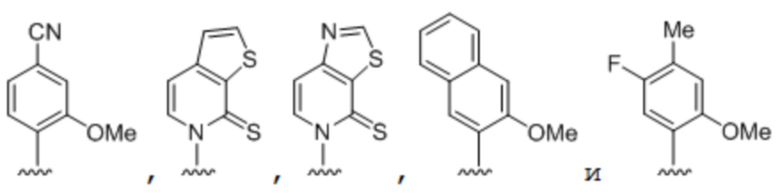 .
.
В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTAT1. В некоторых вариантах осуществления ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTAT1.
[00273] В некоторых вариантах осуществления мРНК и тРНК содержат неприродное основание, выбранное из , и . В некоторых вариантах осуществления мРНК и тРНК содержат неприродное основание, выбранное из и . В некоторых вариантах осуществления мРНК содержит неприродное основание, являющееся . В некоторых вариантах осуществления мРНК содержит неприродное основание, являющееся . В некоторых вариантах осуществления мРНК содержит неприродное основание, являющееся . В некоторых вариантах осуществления тРНК содержит неприродное основание, выбранное из и . В некоторых вариантах осуществления тРНК содержит неприродное основание, являющееся . В некоторых вариантах осуществления тРНК содержит неприродное основание, являющееся . В некоторых вариантах осуществления тРНК содержит неприродное основание, являющееся .
[00274] В некоторых вариантах осуществления способа получения белка, содержащего неприродную аминокислоту, представленного в настоящем описании, первое неприродное основание содержит dCNMO, и второе неприродное основание содержит dTPT3. В некоторых вариантах осуществления третье неприродное основание содержит NaM, и второе неприродное основание содержит TAT1.
[00275] Настоящее изобретение также относится к белку, содержащему по меньшей мере одну неприродную аминокислоту, где белок получают любым из способов, представленных в настоящем описании. В некоторых вариантах осуществления белок содержит по меньшей мере одну неприродную аминокислоту. В некоторых вариантах осуществления белок содержит одну неприродную аминокислоту. В некоторых вариантах осуществления белок содержит две или более неприродных аминокислоты. В некоторых вариантах осуществления белок содержит две неприродные аминокислоты. В некоторых вариантах осуществления белок содержит три или более неприродных аминокислот.
[00276] В некоторых вариантах осуществления аналоги нуклеотидов также являются модифицированными по фосфатному остатку. Модифицированные фосфатные остатки включают, в качестве неограничивающих примеров, фосфатные остатки с модификацией по связи между двумя нуклеотидами и содержат, например, фосфотиоат, хиральный фосфотиоат, фосфодитиоат, фосфотриэфир, аминоалкилфосфотриэфир, метильные и другие алкильные фосфонаты, включая 3'-алкиленфосфонат и хиральные фосфонаты, фосфинаты, фосфорамидаты, включая 3'-аминофосфорамидат и аминоалкилфосфорамидаты, тионофосфорамидаты, тионоалкилфосфонаты, тионоалкилфосфотриэфиры и боранофосфаты. Следует понимать, что эти фосфатные или модифицированные фосфатные связи между двумя нуклеотидами осуществляют через 3'-5'-связь или 2'-5'-связь, и связь включает обратную полярность, например, 3'-5' в 5'-3' или 2'-5' в 5'-2'. Также включены различные соли, смешанные соли и формы свободных кислот. Во многих патентах США описано, как получать и использовать нуклеотиды, содержащие модифицированный фосфаты, и включают, в качестве неограничивающих примеров, патенты США №№ 3687808; 4469863; 4476301; 5023243; 5177196; 5188897; 5264423; 5276019; 5278302; 5286717; 5321131; 5399676; 5405939; 5453496; 5455233; 5466677; 5476925; 5519126; 5536821; 5541306; 5550111; 5563253; 5571799; 5587361 и 5625050; описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00277] В некоторых вариантах осуществления неприродные нуклеиновые кислоты включают 2',3'-дидезокси-2',3'-дидегидро-нуклеозиды (PCT/US2002/006460), 5'-замещенные производные ДНК и РНК (PCT/US2011/033961; Saha et al., J. Org Chem., 1995, 60, 788-789; Wang et al., Bioorganic & Medicinal Chemistry Letters, 1999, 9, 885-890; и Mikhailov et al., Nucleosides & Nucleotides, 1991, 10(1-3), 339-343; Leonid et al., 1995, 14(3-5), 901-905; и Eppacher et al., Helvetica Chimica Acta, 2004, 87, 3004-3020; PCT/JP2000/004720; PCT/JP2003/002342; PCT/JP2004/013216; PCT/JP2005/020435; PCT/JP2006/315479; PCT/JP2006/324484; PCT/JP2009/056718; PCT/JP2010/067560), или 5'-замещенные мономеры, полученные в виде монофосфата с модифицированными основаниями (Wang et al., Nucleosides Nucleotides & Nucleic Acids, 2004, 23 (1 & 2), 317-337); описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00278] В некоторых вариантах осуществления неприродные нуклеиновые кислоты включают модификации в 5'-положении и 2'-положении сахарного кольца (PCT/US94/02993), такие как 5'-CH2-замещенные 2'-O-защищенные нуклеозиды (Wu et al., Helvetica Chimica Acta, 2000, 83, 1127-1143 и Wu et al., Bioconjugate Chem. 1999, 10, 921-924). В некоторых случаях неприродные нуклеиновые кислоты включают амид-связанные димеры нуклеозидов, полученных для встраивания в олигонуклеотиды, где 3'-связанный нуклеозид в димере (5'-3') содержит 2'-OCH3 и 5'-(S)-CH3 (Mesmaeker et al., Synlett, 1997, 1287-1290). Неприродные нуклеиновые кислоты могут включать 2'-замещенные 5'-CH2 (или O-модифицированные нуклеозиды (PCT/US92/01020). Неприродные нуклеиновые кислоты могут включать 5'-метиленфосфонат-мономеры и димеры ДНК и РНК (Bohringer et al., Tet. Lett., 1993, 34, 2723-2726; Collingwood et al., Synlett, 1995, 7, 703-705; и Hutter et al., Helvetica Chimica Acta, 2002, 85, 2777-2806). Неприродные нуклеиновые кислоты могут включать 5'-фосфонатные мономеры, имеющие 2'-замену (US2006/0074035), и другие модифицированные 5'-фосфонатные мономеры (WO1997/35869). Неприродные нуклеиновые кислоты могут включать 5'-модифицированные метиленфосфонатные мономеры (EP614907 и EP629633). Неприродные нуклеиновые кислоты могут включать аналоги 5'- или 6'-фосфонат-рибонуклеозидов, содержащих гидроксильную группу в 5'- и/или 6'-положении (Chen et al., Phosphorus, Sulphur and Silicon, 2002, 777, 1783-1786; Jung et al., Bioorg. Med. Chem., 2000, 8, 2501-2509; Gallier et al., Eur. J. Org. Chem., 2007, 925-933; и Hampton et al., J. Med. Chem., 1976, 19(8), 1029-1033). Неприродные нуклеиновые кислоты могут включать 5'-фосфонат-дезоксирибонуклеозидные мономеры и димеры, имеющие 5'-фосфатную группу (Nawrot et al., Oligonucleotides, 2006, 16(1), 68-82). Неприродные нуклеиновые кислоты могут включать нуклеозиды, имеющие 6'-фосфонатную группу, где 5'- или/и 6'-положение является незамещенным или замещенным тио-трет-бутильной группой (SC(CH3)3) (и ее аналогами); метиленаминогруппой (CH2NH2) (и ее аналогами) или цианогруппой (CN) (и ее аналогами) (Fairhurst et al., Synlett, 2001, 4, 467-472; Kappler et al., J. Med. Chem., 1986, 29, 1030-1038; Kappler et al., J. Med. Chem., 1982, 25, 1179-1184; Vrudhula et al., J. Med. Chem., 1987, 30, 888-894; Hampton et al., J. Med. Chem., 1976, 19, 1371-1377; Geze et al., J. Am. Chem. Soc, 1983, 105(26), 7638-7640; и Hampton et al., J. Am. Chem. Soc, 1973, 95(13), 4404-4414). Описание каждой из ссылок, указанных в этом абзаце, включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00279] В некоторых вариантах осуществления неприродные нуклеиновые кислоты также включают модификации сахарного остатка. В некоторых случаях нуклеиновые кислоты содержат один или более нуклеозидов, где сахарная группа модифицирована. Такие сахар-модифицированные нуклеозиды могут придавать повышенную стабильность против нуклеаз, повышенную аффинность связывания или некоторые другие благоприятные биологические свойства. В некоторых вариантах осуществления нуклеиновые кислоты содержат химически модифицированный остаток рибофуранозного кольца. Неограничивающие примеры химически модифицированных рибофуранозных колец включают добавление групп-заместителей (включая 5'- и/или 2'-группы-заместители; мостиковое соединение двух кольцевых атомов с образованием бициклических нуклеиновых кислот (BNA); замену атома кислорода рибозильного кольца S, N(R) или C(R1)(R2) (R=H, C1-C12-алкил или защитная группа); и их комбинации. Примеры химически модифицированных сахаров можно найти в WO2008/101157, US2005/0130923 и WO2007/134181; описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00280] В некоторых случаях модифицированная нуклеиновая кислота содержит модифицированные сахара или аналоги сахаров. Таким образом, в дополнение к рибозе и дезоксирибозе, сахарный остаток может представлять собой пентозу, дезоксипентозу, гексозу, дезоксигексозу, глюкозу, арабинозу, ксилозу, ликсозу или "аналогичную" сахару циклопентильную группу. Сахар может находиться в пиранозильной или фуранозильной форме. Сахарный остаток может являться фуранозидом рибозы, дезоксирибозы, арабинозы или 2'-O-алкилрибозы, и сахар можно присоединять к соответствующим гетероциклическим основаниям в [альфа]- или [бета]-аномерной конфигурации. Модификации сахаров включают, в качестве неограничивающих примеров, 2'-алкокси-аналоги РНК, 2'-амино-аналоги РНК, 2'-фтор-ДНК и химеры 2'-алкокси- или амино-РНК/ДНК. Например, модификация сахара может включать 2'-O-метил-уридин или 2'-O-метил-цитидин. Модификации сахаров включают 2'-O-алкил-замещенные дезоксирибонуклеозиды и 2'-O-этиленгликоль-подобные рибонуклеозиды. Известно получение этих сахаров или аналогов сахаров и соответствующих "нуклеозидов", где такие сахара или аналоги присоединяют к гетероциклическому основанию (основанию нуклеиновой кислоты). Модификации сахара также можно осуществлять и комбинировать с другими модификациями.
[00281] Модификации сахарного остатка включают природные модификации рибозы и дезоксирибозы, а также неприродные модификации. Модификации сахаров включают, в качестве неограничивающих примеров, следующие модификации в 2'-положении: OH; F; O-, S- или N-алкил; O-, S- или N-алкенил; O-, S- или N-алкинил; или O-алкил-O-алкил, где алкил, алкенил и алкинил может являться замещенным или незамещенным C1-C10-алкилом или C2-C10-алкенилом и -алкинилом. 2'-модификации сахара также включают в качестве неограничивающих примеров, -O[(CH2)nO]m CH3, -O(CH2)nOCH3,-O(CH2)nNH2, -O(CH2)nCH3, -O(CH2)nONH2 и -O(CH2)nON[(CH2)n CH3)]2, где n и m составляют от 1 до приблизительно 10.
[00282] Другие модификации в 2'-положении включают, в качестве неограничивающих примеров: низший C1-C10-алкил, замещенный низший алкил, алкарил, аралкил, O-алкарил, O-аралкил, SH, SCH3, OCN, Cl, Br, CN, CF3, OCF3, SOCH3, SO2 CH3, ONO2, NO2, N3, NH2, гетероциклоалкил, гетероциклоалкарил, аминоалкиламиногруппу, полиалкиламиногруппу, замещенный силил, РНК-отщепляющую группу, репортерную группу, интеркалятор, группу для улучшения фармакокинетических свойств олигонуклеотида или группу для улучшения фармакодинамических свойств олигонуклеотида, и другие заместители, имеющие схожие свойства. Схожие модификации также можно осуществлять в других положениях сахара, в частности, 3'-положении сахара на 3'-концевом нуклеотиде или в 2'-5'-связанных олигонуклеотидах и 5'-положении 5'-концевого нуклеотида. Модифицированные сахара также включают сахара, содержащие модификации в мостиковом кислороде кольца, такие как CH2 и S. Аналоги сахаров нуклеотидов также могут включать миметики сахаров, такие как циклобутильные остатки вместо пентофуранозильного сахара. Существует множество патентов США, в которых описано получение структур таких модифицированных сахаров и диапазон модификаций оснований, такие как патенты США №№ 4981957; 5118800; 5319080; 5359044; 5393878; 5446137; 5466786; 5514785; 5519134; 5567811; 5576427; 5591722; 5597909; 5610300; 5627053; 5639873; 5646265; 5658873; 5670633; 4845205; 5130302; 5134066; 5175273; 5367066; 5432272; 5457187; 5459255; 5484908; 5502177; 5525711; 5552540; 5587469; 5594121, 5596091; 5614617; 5681941 и 5700920, описание каждого из которых включено в настоящее описание в качестве ссылки в полном объеме.
[00283] Неограничивающие примеры нуклеиновых кислот, содержащих остатки модифицированных сахаров, включают нуклеиновые кислоты, содержащие группы-заместители 5'-винил, 5'-метил (R или S), 4'-S, 2'-F, 2'-OCH3 и 2'-O(CH2)2OCH3. Заместитель в 2'-положении также можно выбирать из аллила, аминогруппы, азидогруппы, тиогруппы, O-аллила, O-(C1-C1O-алкила), OCF3, O(CH2)2SCH3, O(CH2)2-O-N(Rm)(Rn) и O-CH2-C(=O)-N(Rm)(Rn), где каждый Rm и Rn независимо представляет собой H или замещенный или незамещенный C1-C10-алкил.
[00284] В некоторых вариантах осуществления нуклеиновые кислоты, представленные в настоящем описании, включают одну или более бициклических нуклеиновых кислот. В некоторых таких вариантах осуществления бициклическая нуклеиновая кислота содержит мостик между 4'- и 2'-атомами рибозильного кольца. В некоторых вариантах осуществления нуклеиновые кислоты, представленные в настоящем описании, включают одну или более бициклических нуклеиновых кислот, где мостик содержит 4'-2'-бициклическую нуклеиновую кислоту. Неограничивающие примеры таких 4'-2'-бициклических нуклеиновых кислот включают одну из формул: 4'-(CH2)-O-2' (ЗНК); 4'-(CH2)-S-2'; 4'-(CH2)2-O-2' (ENA); 4'-CH(CH3)-O-2' и 4'-CH(CH2OCH3)-O-2' и их аналоги (см., патент США № 7399845); 4'-C(CH3)(CH3)-O-2' и его аналоги (см. WO2009/006478, WO2008/150729, US2004/0171570, патент США № 7427672, Chattopadhyaya et al., J. Org. Chem., 209, 74, 118-134, и WO2008/154401). Также см., например: Singh et al., Chem. Commun., 1998, 4, 455-456; Koshkin et al., Tetrahedron, 1998, 54, 3607-3630; Wahlestedt et al., Proc. Natl. Acad. Sci. U. S. A., 2000, 97, 5633-5638; Kumar et al., Bioorg. Med. Chem. Lett., 1998, 8, 2219-2222; Singh et al., J. Org. Chem., 1998, 63, 10035-10039; Srivastava et al., J. Am. Chem. Soc., 2007, 129(26) 8362-8379; Elayadi et al., Curr. Opinion Invens. Drugs, 2001, 2, 558-561; Braasch et al., Chem. Biol, 2001, 8, 1-7; Oram et al., Curr. Opinion Mol. Ther., 2001, 3, 239-243; патенты США №№ 4849513; 5015733; 5118800; 5118802; 7053207; 6268490; 6770748; 6794499; 7034133; 6525191; 6670461 и 7399845; международные патентные публикации №№ WO2004/106356, WO1994/14226, WO2005/021570, WO2007/090071 и WO2007/134181; патентные публикации США №№ US2004/0171570, US2007/0287831 и US2008/0039618; временные патентные заявки США №№ 60/989574, 61/026995, 61/026998, 61/056564, 61/086231, 61/097787 и 61/099844; и международные патентные заявки №№ PCT/US2008/064591, PCT US2008/066154, PCT US2008/068922 и PCT/DK98/00393; описание каждой из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00285] В некоторых вариантах осуществления нуклеиновые кислоты содержат связанные нуклеиновые кислоты. Нуклеиновые кислоты можно связывать друг с другом с использованием любой связи между нуклеиновыми кислотами. Два основных класса групп, связывающих нуклеиновые кислоты, определяют по наличию или отсутствию атома фосфора. Типичные фосфор-содержащие связи между нуклеиновыми кислотами включают, в качестве неограничивающих примеров, фосфодиэфиры, фосфотриэфиры, метилфосфонаты, фосфорамидат и фосфотиоаты (P=S). Типичные не-фосфор-содержащие группы, связывающие нуклеиновые кислоты, включают метиленметилиминогруппу (-CH2-N(CH3)-O-CH2-), тиодиэфир (-O-C(O)-S-), тионокарбамат (-O-C(O)(NH)-S-); силоксан (-O-Si(H)2-O-); и N, N*-диметилгидразин (-CH2-N(CH3)-N(CH3)). В некоторых вариантах осуществления связи между нуклеиновыми кислотами, имеющими хиральный атом, можно получать в виде рацемической смеси или отдельных энантиомеров, например, алкилфосфонатов и фосфотиоатов. Неприродные нуклеиновые кислоты могут содержать отдельную модификацию. Неприродные нуклеиновые кислоты могут содержать множественные модификации в одном из остатков или между разными остатками.
[00286] Фосфатные модификации остова в нуклеиновой кислоте включают, в качестве неограничивающих примеров, метилфосфонат, фосфотиоат, фосфорамидат (мостиковый или немостиковый), фосфотриэфир, фосфодитиоат, фосфодитиоат и боранофосфат, и их можно использовать в любой комбинации. Также можно использовать другие нефосфатные связи.
[00287] В некоторых вариантах осуществления модификации остова (например, метилфосфонатные, фосфотиоатные, фосфорамидатные и фосфодитиоатные межнуклеотидные связи) могут придавать модифицированной нуклеиновой кислоте иммуномодулирующую активность и/или повышать ее стабильность in vivo.
[00288] В некоторых случаях фосфорное производное (или модифицированная фосфатная группа) присоединено к сахарному остатку или аналогу сахара и может представлять собой монофосфат, дифосфат, трифосфат, алкилфосфонат, фосфотиоат, фосфодитиоат, фосфорамидат или т.п. Примеры полинуклеотидов, содержащих модифицированные фосфатные связи или нефосфатные связи, можно найти в Peyrottes et al., 1996, Nucleic Acids Res. 24: 1841-1848; Chaturvedi et al., 1996, Nucleic Acids Res. 24:2318-2323; и Schultz et al., (1996) Nucleic Acids Res. 24:2966-2973; Matteucci, 1997, "Oligonucleotide Analogs: an Overview" в Oligonucleotides as Therapeutic Agent, (Chadwick and Cardew, ed.) John Wiley and Sons, New York, NY; Zon, 1993, "Oligonucleoside Phosphorothioates" в Protocols for Oligonucleotides and Analogs, Synthesis and Properties, Humana Press, pp. 165-190; Miller et al., 1971, JACS 93:6657-6665; Jager et al., 1988, Biochem. 27:7247-7246; Nelson et al., 1997, JOC 62:7278-7287; патент США № 5453496; и Micklefield, 2001, Curr. Med. Chem. 8: 1157-1179; описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00289] В некоторых случаях модификация остова включает замену фосфодиэфирной связи альтернативным остатком, таким как анионная, нейтральная или катионная группа. Примеры таких модификаций включают: анионную межнуклеозидную связь; фосфорамидатную модификацию N3' в P5'; боранофосфат-ДНК; проолигонуклеотиды; нейтральные межнуклеозидные связи, такие как метилфосфонаты; амид-связанную ДНК; метилен(метилимино)-связи; формацеталевые и тиоформацеталевые связи; остовы, содержащие сульфонильные группы; морфолино-олигонуклеотиды; пептид-нуклеиновые кислоты (ПНК); и положительно заряженные дезоксирибонуклеиновые гуанидиновые (DNG) олигонуклеотиды (Micklefield, 2001, Current Medicinal Chemistry 8: 1157-1179), описание которого, таким образом, включено в настоящее описание в качестве ссылки в полном объеме. Модифицированная нуклеиновая кислота может содержать химерный или смешанный остов, содержащий одну или более модификаций, например, комбинацию фосфатных связей, такую как комбинация фосфодиэфирных и фосфотиоатных связей.
[00290] Заместители фосфата включают, например, короткоцепочечные алкильные или циклоалкильные межнуклеозидные связи, смешанные гетероатомные и алкильные или циклоалкильные межнуклеозидные связи, или одну или более короткоцепочечных гетероатомных или гетероциклических межнуклеозидных связей. Они включают остовы, имеющие морфолиновые связи (образованные частично из сахарной части нуклеозида); силоксановые остовы; сульфидные, сульфоксидные и сульфоновые остовы; формацетильные и тиоформацетильные остовы; метиленформацетильные и тиоформацетильные остовы; алкен-содержащие остовы; сульфаматные остовы; метилениминовые и метиленгидразиновые остовы; сульфонатные и сульфонамидные остовы; амидные остовы и другие остовы, имеющие смешанные компоненты N, O, S и CH2. Во многих патентах США описано, как получать и использовать эти типы заместителей фосфатов, и они включают, в качестве неограничивающих примеров, патенты США №№ 5034506; 5166315; 5185444; 5214134; 5216141; 5235033; 5264562; 5264564; 5405938; 5434257; 5466677; 5470967; 5489677; 5541307; 5561225; 5596086; 5602240; 5610289; 5602240; 5608046; 5610289; 5618704; 5623070; 5663312; 5633360; 5677437 и 5677439; описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме. В отношении заместителей нуклеотидов также следует понимать, что можно заменять и сахарные, и фосфатные остатки нуклеотида, например, связью амидного типа (аминоэтилглицин) (ПНК). В патентах США №№ 5539082; 5714331 и 5719262 описано, как получать и использовать молекулы ПНК, каждый из них включен в настоящее описание в качестве ссылки. Также см. Nielsen et al., Science, 1991, 254, 1497-1500. С нуклеотидами или аналогами нуклеотидов также можно связывать другие типы молекул (конъюгаты) для повышения, например, захвата клетками. Конъюгаты можно химически связывать с нуклеотидом или аналогами нуклеотидов. Такие конъюгаты включают, в качестве неограничивающих примеров, липидные остатки, такие как остаток холестерина (Letsinger et al., Proc. Natl. Acad. Sci. USA, 1989, 86, 6553-6556), холевую кислоту (Manoharan et al., Bioorg. Med. Chem. Let., 1994, 4, 1053-1060), тиоэфир, например, гексил-S-тритилтиол (Manoharan et al., Ann. KY. Acad. Sci., 1992, 660, 306-309; Manoharan et al., Bioorg. Med. Chem. Let., 1993, 3, 2765-2770), тиохолестерин (Oberhauser et al., Nucl. Acids Res., 1992, 20, 533-538), алифатическую цепь, например, додекандиоловые или ундециловые остатки (Saison-Behmoaras et al., EM5OJ, 1991, 10, 1111-1118; Kabanov et al., FEBS Lett., 1990, 259, 327-330; Svinarchuk et al., Biochimie, 1993, 75, 49-54), фосфолипид, например, ди-гексадецил-rac-глицерин или 1-ди-O-гексадецил-rac-глицеро-S-H-фосфонат триэтиламмония (Manoharan et al., Tetrahedron Lett., 1995, 36, 3651-3654; Shea et al., Nucl. Acids Res., 1990, 18, 3777-3783), полиаминовую или полиэтиленгликолевую цепь (Manoharan et al., Nucleosides & Nucleotides, 1995, 14, 969-973), или адамантануксусную кислоту (Manoharan et al., Tetrahedron Lett., 1995, 36, 3651-3654), пальмитиловый остаток (Mishra et al., Biochem. Biophys. Acta, 1995, 1264, 229-237) или октадециламиновый или гексиламино-карбонил-оксихолестериновый остаток (Crooke et al., J. Pharmacol. Exp. Ther., 1996, 277, 923-937); описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме. Во многих патентах США описано получение таких конъюгатов, и они включают, в качестве неограничивающих примеров, патенты США №№ 4828979; 4948882; 5218105; 5525465; 5541313; 5545730; 5552538; 5578717, 5580731; 5580731; 5591584; 5109124; 5118802; 5138045; 5414077; 5486603; 5512439; 5578718; 5608046; 4587044; 4605735; 4667025; 4762779; 4789737; 4824941; 4835263; 4876335; 4904582; 4958013; 5082830; 5112963; 5214136; 5082830; 5112963; 5214136; 5245022; 5254469; 5258506; 5262536; 5272250; 5292873; 5317098; 5371241, 5391723; 5416203, 5451463; 5510475; 5512667; 5514785; 5565552; 5567810; 5574142; 5585481; 5587371; 5595726; 5597696; 5599923; 5599928 и 5688941; описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00291] Настоящее изобретение относится к нуклеиновым основаниям, используемым в композициях и способах для репликации, транскрипции, трансляции и встраивания неприродных аминокислот в белки. В некоторых вариантах осуществления нуклеиновое основание, представленное в настоящем описании, содержит структуру:
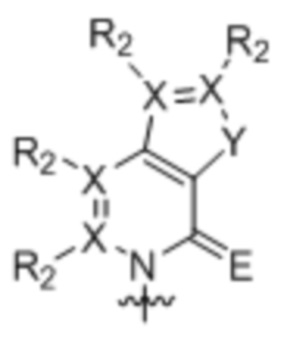 , где
, где
каждый X независимо представляет собой углерод или азот;
R2 присутствует, если X представляет собой углерод, и независимо представляет собой водород, алкил, алкенил, алкинил, метоксигруппу, метантиол, метанселеногруппу, галоген, цианогруппу или азидную группу;
где каждый Y независимо представляет собой серу, кислород, селен или вторичный амин;
где каждый E независимо представляет собой кислород, серу или селен; и
где волнистой линией указана точка связывания с рибозильным, дезоксирибозильным или дидезоксирибозильным остатком или его аналогом, где рибозильный, дезоксирибозильный или дидезоксирибозильный остаток или его аналог находится в свободной форме, соединен с монофосфатной, дифосфатной или трифосфатной группой, необязательно, содержащей α-тиотрифосфатную, β-тиотрифосфатную или γ-тиотрифосфатную группу, или включен в РНК, или ДНК, или аналог РНК, или аналог ДНК.
[00292] В некоторых вариантах осуществления R2 является низшим алкилом (например, C1-C6), водородом или галогеном. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, R2 является фтором. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, X является углеродом. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, E является серой. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, Y является серой.
[00293] В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание имеет структуру: 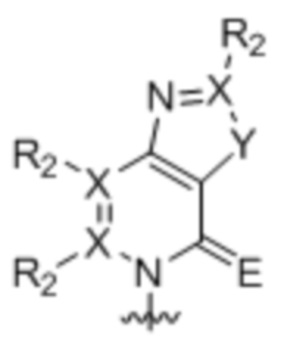 . В некоторых вариантах осуществления нуклеиновое основание, представленное в настоящем описании, имеет структуру: . В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, E является серой, и Y является серой. В некоторых вариантах осуществления волнистой линией указана точка связывания с рибозильным, дезоксирибозильным или дидезоксирибозильным остатком или его аналогом, где рибозильный, дезоксирибозильный или дидезоксирибозильный остаток или его аналог находится в свободной форме, или соединен с монофосфатной, дифосфатной, трифосфатной, α-тиотрифосфатной, β-тиотрифосфатной или γ-тиотрифосфатной группы, или включен в РНК, или ДНК, или аналог РНК, или аналог ДНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, волнистой линией указана точка связывания с рибозильным или дезоксирибозильным остатком. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, волнистой линией указана точка связывания с рибозильным или дезоксирибозильным остатком, соединенным с трифосфатной группой. В некоторых вариантах осуществления нуклеиновое основание, представленное в настоящем описании, является компонентом полимера нуклеиновой кислоты. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание является компонентом тРНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание является компонентом антикодона в тРНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание является компонентом мРНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание является компонентом кодона мРНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание является компонентом РНК или ДНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание является компонентом кодона в ДНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание образует, может образовывать или сконфигурировано для образования пары нуклеиновых оснований с другим (например, комплементарным) нуклеиновым основанием.
. В некоторых вариантах осуществления нуклеиновое основание, представленное в настоящем описании, имеет структуру: . В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, E является серой, и Y является серой. В некоторых вариантах осуществления волнистой линией указана точка связывания с рибозильным, дезоксирибозильным или дидезоксирибозильным остатком или его аналогом, где рибозильный, дезоксирибозильный или дидезоксирибозильный остаток или его аналог находится в свободной форме, или соединен с монофосфатной, дифосфатной, трифосфатной, α-тиотрифосфатной, β-тиотрифосфатной или γ-тиотрифосфатной группы, или включен в РНК, или ДНК, или аналог РНК, или аналог ДНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, волнистой линией указана точка связывания с рибозильным или дезоксирибозильным остатком. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, волнистой линией указана точка связывания с рибозильным или дезоксирибозильным остатком, соединенным с трифосфатной группой. В некоторых вариантах осуществления нуклеиновое основание, представленное в настоящем описании, является компонентом полимера нуклеиновой кислоты. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание является компонентом тРНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание является компонентом антикодона в тРНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание является компонентом мРНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание является компонентом кодона мРНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание является компонентом РНК или ДНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание является компонентом кодона в ДНК. В некоторых вариантах осуществления нуклеинового основания, представленного в настоящем описании, нуклеиновое основание образует, может образовывать или сконфигурировано для образования пары нуклеиновых оснований с другим (например, комплементарным) нуклеиновым основанием.
[00294] В некоторых вариантах осуществления нуклеиновое основание, представленное в настоящем описании, имеет структуру:
где:
каждый X независимо представляет собой углерод или азот;
R2 отсутствует, если X является азотом, и присутствует, если X представляет собой углерод, и независимо представляет собой водород, алкил, алкенил, алкинил, метоксигруппу, метантиол, метанселеногруппу, галоген, цианогруппу или азид;
Y является серой, кислородом, селеном или вторичным амином;
E является кислородом, серой или селеном; и
волнистой линией указана точка связывания с рибозильным, дезоксирибозильным или дидезоксирибозильным остатком или его аналогом, где рибозильный, дезоксирибозильный или дидезоксирибозильный остаток или его аналог находится в свободной форме, соединен с монофосфатной, дифосфатной, трифосфатной, α-тиотрифосфатной, β-тиотрифосфатной или γ-тиотрифосфатной группой или включен в РНК, или ДНК, или аналог РНК, или аналог ДНК.
[00295] В некоторых вариантах осуществления каждый X является углеродом. В некоторых вариантах осуществления по меньшей мере один X является углеродом. В некоторых вариантах осуществления один X является углеродом. В некоторых вариантах осуществления по меньшей мере два X являются углеродом. В некоторых вариантах осуществления два X являются углеродом. В некоторых вариантах осуществления по меньшей мере один X является азотом. В некоторых вариантах осуществления один X является азотом. В некоторых вариантах осуществления по меньшей мере два X являются азотом. В некоторых вариантах осуществления два X являются азотом.
[00296] В некоторых вариантах осуществления Y является серой. В некоторых вариантах осуществления Y является кислородом. В некоторых вариантах осуществления Y является селеном. В некоторых вариантах осуществления Y является вторичным амином.
[00297] В некоторых вариантах осуществления E является серой. В некоторых вариантах осуществления E является кислородом. В некоторых вариантах осуществления E является селеном.
[00298] В некоторых вариантах осуществления R2 присутствует, если X представляет собой углерод. В некоторых вариантах осуществления R2 отсутствует, если X является азотом. В некоторых вариантах осуществления каждый R2, если он присутствует, является водородом. В некоторых вариантах осуществления R2 является алкилом, таким как метил, этил или пропил. В некоторых вариантах осуществления R2 является алкенилом, таким как -CH2=CH2. В некоторых вариантах осуществления R2 является алкинилом, таким как этинил. В некоторых вариантах осуществления R2 является метоксигруппой. В некоторых вариантах осуществления R2 является метантиолом. В некоторых вариантах осуществления R2 является метанселеногруппой. В некоторых вариантах осуществления R2 является галогеном, таким как хлор, бром или фтор. В некоторых вариантах осуществления R2 является цианогруппой. В некоторых вариантах осуществления R2 является азидом.
[00299] В некоторых вариантах осуществления E является серой, Y является серой, и каждый X независимо представляет собой углерод или азот. В некоторых вариантах осуществления E является серой, Y является серой, и каждый X является углеродом.
[00300] В некоторых вариантах осуществления нуклеиновое основание имеет структуру  . В некоторых вариантах осуществления нуклеиновое основание имеет структуру
. В некоторых вариантах осуществления нуклеиновое основание имеет структуру
 .
.
[00301] В некоторых вариантах осуществления нуклеиновое основание имеет структуру .
[00302] В некоторых вариантах осуществления нуклеиновое основание, представленное в настоящем описании, связано (например, нековалентно) с комплементарным, спаривающимся нуклеиновым основанием с образованием неприродной пары оснований (UBP), или может образовывать пары оснований, или сконфигурировано для образования пары оснований с нуклеиновым основанием. В некоторых вариантах осуществления комплементарное, спаривающееся нуклеиновое основание выбрано из:
[00303] В одном из аспектов настоящее изобретение относится к двухцепочечному олигонуклеотидному дуплексу, где первая олигонуклеотидная цепь содержит нуклеиновое основание, представленное в настоящем описании, и вторая комплементарная олигонуклеотидная цепь содержит комплементарное, спаривающееся нуклеиновое основание в его комплементарном, образующем пару оснований участке. В некоторых вариантах осуществления первая олигонуклеотидная цепь содержит ; и вторая цепь содержит комплементарное, спаривающееся нуклеиновое основание, выбранное из группы, состоящей из
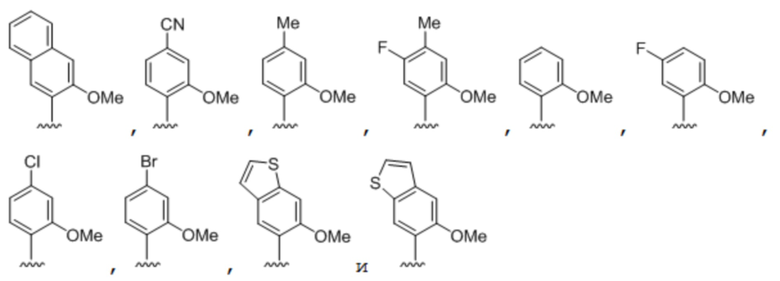
в его комплементарном, образующем пару оснований участке.
[00304] В дополнительном аспекте настоящее изобретение относится к транспортной РНК (тРНК), содержащей нуклеиновое основание, представленное в настоящем описании, содержащее: антикодон, где антикодон содержит нуклеиновое основание; и элемент распознавания, где элемент распознавания способствует селективной зарядке тРНК неприродной аминокислотой посредством аминоацил-тРНК-синтетазы. В некоторых вариантах осуществления нуклеиновое основание находится в области антикодона тРНК. В некоторых вариантах осуществления нуклеиновое основание находится в первом положении антикодона. В некоторых вариантах осуществления нуклеиновое основание находится во втором положении антикодона. В некоторых вариантах осуществления нуклеиновое основание находится в третьем положении антикодона. В некоторых вариантах осуществления аминоацил-тРНК-синтетазу получают из Methanosarcina или его варианта. В некоторых вариантах осуществления аминоацил-тРНК-синтетазу получают из Methanococcus (Methanocaldococcus) или его варианта. В некоторых вариантах осуществления неприродная аминокислота содержит ароматический остаток. В некоторых вариантах осуществления неприродная аминокислота является производным лизина. В некоторых вариантах осуществления неприродная аминокислота является производным фенилаланина.
[00305] Настоящее изобретение также относится к структуре, содержащей формулу:
N1 - Zx - N2
где:
Z является нуклеиновым основание, представленным в настоящем описании, связанным с рибозилом, или дезоксирибозилом, или их аналогом;
N1 является одним или более нуклеотидами или их аналогами или концевой фосфатной группой, присоединенной к 5'-концу рибозила, или дезоксирибозила, или их аналога из Z;
N2 является одним или более нуклеотидами или их аналогами или концевой гидроксильной группой, присоединенной к 3'-концу рибозила, или дезоксирибозила, или их аналога из Z; и
x является целым числом от 1 до 20.
[00306] В некоторых вариантах осуществления N1 является одним или более нуклеотидами или их аналогами, присоединенными к 5'-концу рибозильного или дезоксирибозильного остатка или их аналога из Z. Присоединение к 5'-концу рибозильного или дезоксирибозильного остатка можно осуществлять посредством фосфодиэфира. В некоторых вариантах осуществления N1 является концевой фосфатной группой, присоединенной к 5'-концу рибозильного или дезоксирибозильного остатка или их аналога из Z. В некоторых вариантах осуществления N2 является одним или более нуклеотидами или их аналогами, присоединенными к 3'-концу рибозильного или дезоксирибозильного остатка или их аналога из Z. Присоединение к 3'-концу рибозильного или дезоксирибозильного остатка можно осуществлять посредством фосфодиэфира. В некоторых вариантах осуществления N2 является концевой гидроксильной группой, присоединенной к 3'-концу рибозильного или дезоксирибозильного остатка или их аналога из Z.
[00307] В некоторых вариантах осуществления x является целым числом от 1 до 20. В некоторых вариантах осуществления x является целым числом от 1 до 15. В некоторых вариантах осуществления x является целым числом от 1 до 10. В некоторых вариантах осуществления x является целым числом от 1 до 5. В некоторых вариантах осуществления x является 1. В некоторых вариантах осуществления x является 2. В некоторых вариантах осуществления x является 3. В некоторых вариантах осуществления x является 4. В некоторых вариантах осуществления x является 5. В некоторых вариантах осуществления x является 6. В некоторых вариантах осуществления x является 7. В некоторых вариантах осуществления x является 8. В некоторых вариантах осуществления x является 9. В некоторых вариантах осуществления x является 10. В некоторых вариантах осуществления x является 11. В некоторых вариантах осуществления x является 12. В некоторых вариантах осуществления x является 13. В некоторых вариантах осуществления x является 14. В некоторых вариантах осуществления x является 15. В некоторых вариантах осуществления x является 16. В некоторых вариантах осуществления x является 17. В некоторых вариантах осуществления x является 18. В некоторых вариантах осуществления x является 19. В некоторых вариантах осуществления x является 20.
[00308] В некоторых вариантах осуществления Z имеет структуру , как представлено в настоящем описании. В некоторых вариантах осуществления Z имеет структуру .
[00309] В некоторых вариантах осуществления структура формулы N1 - Zx - N2 кодирует ген. В некоторых вариантах осуществления Zx находится в транслируемой области гена. В некоторых вариантах осуществления Zx находится в нетранслируемой области гена. В некоторых вариантах осуществления структура дополнительно содержит 5'- или 3'-нетранслируемую область (UTR). В некоторых вариантах осуществления структура дополнительно содержит область терминатора. В некоторых вариантах осуществления структура дополнительно содержит промоторную область.
[00310] В дополнительном аспекте настоящее изобретение относится к библиотеке полинуклеотидов, где библиотека содержит по меньшей мере 5000 уникальных полинуклеотидов, и где каждый полинуклеотид содержит по меньшей мере одно нуклеиновое основание, представленное в настоящем описании. В некоторых вариантах осуществления библиотека полинуклеотидов кодирует по меньшей мере один ген.
[00311] В еще одном аспекте настоящее изобретение относится к нуклеозидтрифосфату, где нуклеиновое основание выбрано из
, , и . В некоторых вариантах осуществления нуклеиновое основание является . В некоторых вариантах осуществления нуклеиновое основание является . В некоторых вариантах осуществления нуклеиновое основание является . В некоторых вариантах осуществления нуклеиновое основание является . В некоторых вариантах осуществления нуклеозид содержит рибозу. В некоторых вариантах осуществления нуклеозид содержит дезоксирибозу.
Свойства спаривания оснований нуклеиновой кислоты: примеры пар оснований
[00312] В некоторых вариантах осуществления неприродный нуклеотид образует пару оснований (неприродную пару оснований; UBP) с другим неприродным нуклеотидом во время или после встраивания в ДНК или РНК. В некоторых вариантах осуществления стабильно интегрированная неприродная нуклеиновая кислота является неприродной нуклеиновой кислотой, которая может образовывать пару оснований с другой нуклеиновой кислотой, например, природной или неприродной нуклеиновой кислотой. В некоторых вариантах осуществления стабильно интегрированная неприродная нуклеиновая кислота является неприродной нуклеиновой кислотой, которая может образовывать пару оснований с другой неприродной нуклеиновой кислотой (неприродную пару оснований нуклеиновой кислоты (UBP)). Например, первая неприродная нуклеиновая кислота может образовывать пару оснований со второй неприродной нуклеиновой кислотой. Например, одна пара неприродных нуклеозидтрифосфатов, которые могут спариваться во время и после встраивания в нуклеиновые кислоты, включает трифосфат (d)5SICS ((d)5SICSTP) и трифосфат (d)NaM((d)NaMTP). Другие неограничивающие примеры включают: трифосфат (d)CNMO ((d)CNMOTP) и трифосфат (d)TPT3((d)TPT3TP). Такие неприродные нуклеотиды могут содержать сахарный остаток рибозы или дезоксирибозы (указанный с помощью "(d)"). Например, одна пара неприродных нуклеозидтрифосфатов, которые могут спариваться при встраивании в нуклеиновые кислоты, включает трифосфат TAT1 ((d)TAT1TP) и трифосфат NaM((d)NaMTP). В некоторых вариантах осуществления одна пара неприродных нуклеозидтрифосфатов, которые могут спариваться при встраивании в нуклеиновые кислоты, включает трифосфат dCNMO (dCNMOTP) и трифосфат TAT1(TAT1TP). В некоторых вариантах осуществления одна пара неприродных нуклеозидтрифосфатов, которые могут спариваться при встраивании в нуклеиновые кислоты, включает трифосфат dTPT3 (dTPT3TP) и трифосфат NaM(NaMTP). В некоторых вариантах осуществления неприродная нуклеиновая кислота, по существу, не образует пару оснований с природной нуклеиновой кислотой (A, T, G, C). В некоторых вариантах осуществления стабильно интегрированная неприродная нуклеиновая кислота может образовывать пару оснований с природной нуклеиновой кислотой.
[00313] В некоторых вариантах осуществления стабильно интегрированный неприродный (дезокси)рибонуклеотид является неприродным (дезокси)рибонуклеотидом, который может образовывать UBP, но, по существу, не образует пару оснований с каждым из природных (дезокси)рибонуклеотидов. В некоторых вариантах осуществления стабильно интегрированный неприродный (дезокси)рибонуклеотид является неприродным (дезокси)рибонуклеотидом, который может образовывать UBP, но, по существу, не образует пару оснований с одной или более природными нуклеиновыми кислотами. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с A, T и C, но может образовывать пару оснований с G. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с A, T и G, но может образовывать пару оснований с C. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с C, G и A, но может образовывать пару оснований с T. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с C, G и T, но может образовывать пару оснований с A. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с A и T, но может образовывать пару оснований с C и G. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с A и C, но может образовывать пару оснований с T и G. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с A и G, но может образовывать пару оснований с C и T. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с C и T, но может образовывать пару оснований с A и G. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с C и G, но может образовывать пару оснований с T и G. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с T и G, но может образовывать пару оснований с A и G. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с G, но может образовывать пару оснований с A, T и C. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с A, но может образовывать пару оснований с G, T и C. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с T, но может образовывать пару оснований с G, A и C. Например, стабильно интегрированная неприродная нуклеиновая кислота может, по существу, не образовывать пару оснований с C, но может образовывать пару оснований с G, T и A.
[00314] Неограничивающие примеры неприродных нуклеотидов, способных образовывать неприродную пару оснований (UBP) ДНК или РНК в условиях in vivo, включают 5SICS, d5SICS, NaM, dNaM, dTPT3, dMTMO, dCNMO, TAT1 и их комбинации. В некоторых вариантах осуществления неприродные нуклеотиды, способные образовывать неприродную пару оснований (UBP) ДНК или РНК в условиях in vivo, включают, в качестве неограничивающих примеров, 5SICS, NaM, TPT3, MTMO, CNMO, TAT1 и их комбинации, где сахарный остаток нуклеотида является сахаром дезоксирибозой. В некоторых вариантах осуществления неприродные нуклеотиды, способные образовывать неприродную пару оснований (UBP) ДНК или РНК в условиях in vivo, включают, в качестве неограничивающих примеров, 5SICS, NaM, TPT3, MTMO, CNMO, TAT1 и их комбинации, где сахарный остаток нуклеотида является сахаром рибозой. В некоторых вариантах осуществления неприродные нуклеотиды, способные образовывать неприродную пару оснований (UBP) ДНК или РНК в условиях in vivo, включают, в качестве неограничивающих примеров, (d)5SICS, (d)NaM, (d)TPT3, (d)MTMO, (d)CNMO, (d)TAT1 и их комбинации. В некоторых вариантах осуществления неприродные пары оснований нуклеотидов включают, в качестве неограничивающих примеров:
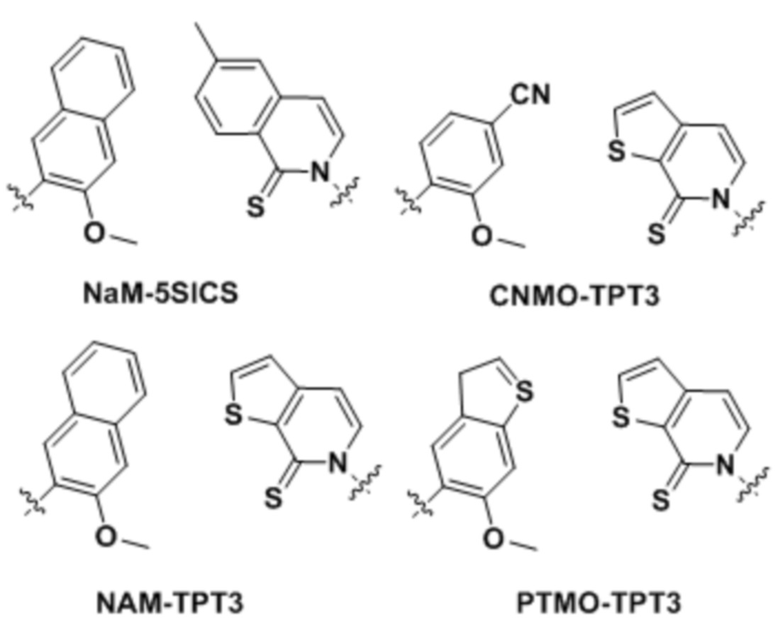 , где сахарный остаток представляет собой любой вариант осуществления или вариант, представленный в настоящем описании. В некоторых вариантах осуществления неприродные пары оснований нуклеотидов включают, в качестве неограничивающих примеров:
, где сахарный остаток представляет собой любой вариант осуществления или вариант, представленный в настоящем описании. В некоторых вариантах осуществления неприродные пары оснований нуклеотидов включают, в качестве неограничивающих примеров:
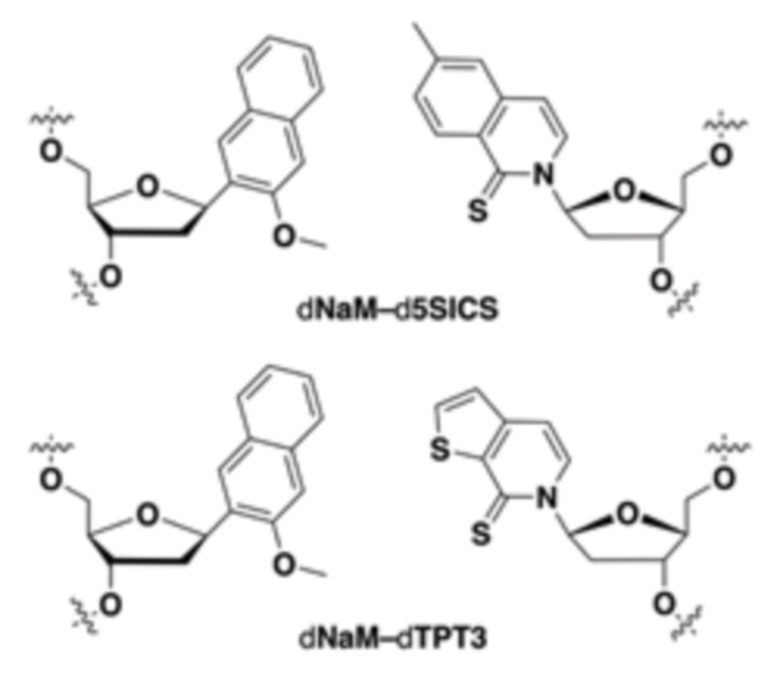
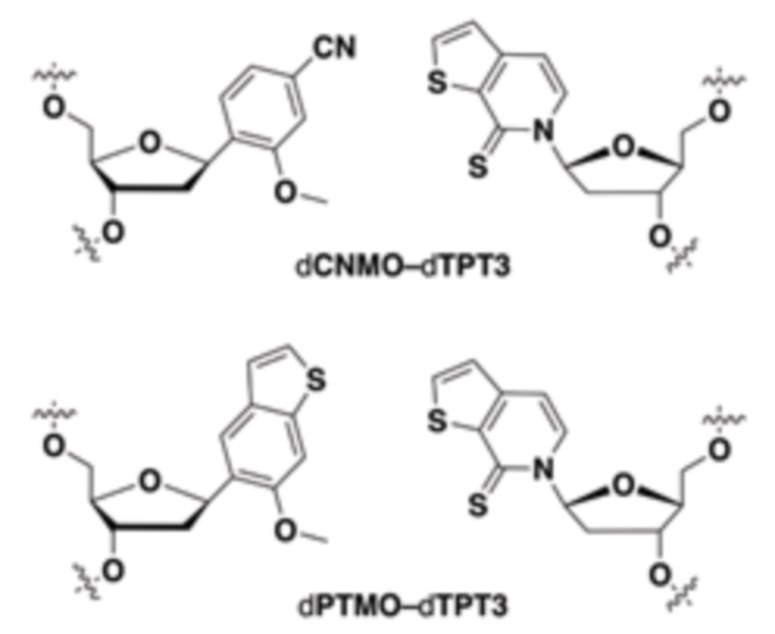 . В любом таком варианте осуществления одна или обе дезоксирибозы, присоединенные к неприродным основаниям, могут быть замещены рибозой.
. В любом таком варианте осуществления одна или обе дезоксирибозы, присоединенные к неприродным основаниям, могут быть замещены рибозой.
Сконструированные организмы
[00315] В некоторых вариантах осуществления способы и плазмиды, представленные в настоящем описании, дополнительно используют для получения сконструированного организма, например организма, включающего и реплицирующего неприродный нуклеотид или неприродную пару оснований нуклеиновой кислоты (UBP), и в них также можно использовать нуклеиновую кислоту, содержащую неприродный нуклеотид, для транскрипции мРНК и тРНК, которую используют для трансляции белков, содержащих неприродный аминокислотный остаток. В некоторых случаях организм является неявляющимся человеком полусинтетическим организмом (SSO). В некоторых случаях организм является полусинтетическим организмом (SSO). В некоторых случаях SSO является клеткой. В некоторых случаях способ in vivo включает полусинтетический организм (SSO). В некоторых случаях полусинтетический организм включает микроорганизм. В некоторых случаях организм включает бактерию. В некоторых случаях организм включает грамотрицательную бактерию. В некоторых случаях организм включает грамположительную бактерию. В некоторых случаях организм включает Escherichia coli (E. coli). Такие модифицированные организмы содержат различные дополнительные компоненты, такие как аппарат репарации ДНК, модифицированные полимеразы, транспортеры нуклеотидов или другие компоненты. В некоторых случаях SSO содержит штамм E. coli YZ3. В некоторых случаях SSO содержит штамм E. coli ML1 или ML2, такой как штаммы, показанные на фигуре 1 (B-D) из Ledbetter, et al. J. Am Chem. Soc. 2018, 140(2), 758, описание которой включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00316] В некоторых случаях используемую клетку генетически трансформируют с использованием экспрессирующей кассеты, кодирующей гетерологичный белок, например, транспортера нуклеозидтрифосфата, способного транспортировать неприродные нуклеозидтрифосфаты в клетку, и, необязательно, систему CRISPR/Cas9 для удаления ДНК, утратившей неприродный нуклеотид (например, штамм E. coli YZ3, ML1 или ML2). В некоторых случаях клетки дополнительно имеют повышенную активность захвата неприродной нуклеиновой кислоты. В некоторых случаях клетки дополнительно имеют повышенную активность импорта неприродной нуклеиновой кислоты.
[00317] В некоторых вариантах осуществления Cas9 и соответствующая гидовая РНК (sgRNA) кодируются разными плазмидами. В некоторых случаях Cas9 и sgRNA кодируются одной плазмидой. В некоторых случаях молекула нуклеиновой кислоты, кодирующая Cas9, sgRNA, или молекула нуклеиновой кислоты, содержащая неприродный нуклеотид, находятся на одной или более плазмидах. В некоторых случаях Cas9 кодируется первой плазмидой, и sgRNA и молекула нуклеиновой кислоты, содержащая неприродный нуклеотид, кодируются второй плазмидой. В некоторых случаях Cas9, sgRNA и молекула нуклеиновой кислоты, содержащая неприродный нуклеотид, кодируются одной плазмидой. В некоторых случаях молекула нуклеиновой кислоты содержит два или более неприродных нуклеотида. В некоторых случаях Cas9 встраивают в геном организма-хозяина, и sgRNA кодируются на плазмиде или в геноме организма.
[00318] В некоторых случаях первую плазмиду, кодирующую Cas9 и sgRNA, и вторую плазмиду, кодирующую молекулу нуклеиновой кислоты, содержащую неприродный нуклеотид, встраивают в сконструированный микроорганизм. В некоторых случаях первую плазмиду, кодирующую Cas9, и вторую плазмиду, кодирующую sgRNA и молекулу нуклеиновой кислоты, содержащую неприродный нуклеотид, встраивают в сконструированный микроорганизм. В некоторых случаях плазмиду, кодирующую Cas9, sgRNA и молекулу нуклеиновой кислоты, содержащую неприродный нуклеотид, встраивают в сконструированный микроорганизм. В некоторых случаях молекула нуклеиновой кислоты содержит два или более неприродных нуклеотида.
[00319] В некоторых вариантах осуществления получают живую клетку, включающую в своей ДНК (плазмиде или геноме) по меньшей мере один неприродный нуклеотид и/или по меньшей мере одну неприродную пару оснований (UBP). В некоторых случаях неприродная пара оснований включает пару неприродных, взаимно спаривающихся нуклеотидов, способных образовывать неприродную пару оснований в условиях in vivo, когда неприродные, взаимно образующие пару нуклеотиды, как их соответствующие трифосфаты, захватываются клеткой благодаря действию транспортера нуклеотидтрифосфатов. В некоторых случаях неприродная пара оснований включает пару неприродных, взаимно спаривающихся нуклеотидов, сконфигурированных для образования неприродной пары оснований в условиях in vivo, когда неприродные, взаимно образующие пару нуклеотиды, как их соответствующие трифосфаты, захватываются клеткой благодаря действию транспортера нуклеотидтрифосфатов. Клетку можно генетически трансформировать посредством экспрессирующей кассеты, кодирующей транспортер нуклеотидтрифосфатов таким образом, что транспортер нуклеотидтрифосфатов экспрессируется и доступен для транспорта неприродных нуклеотидов в клетку. Клетка может являться прокариотической или эукариотической клеткой, и пара неприродных, взаимно спаривающихся нуклеотидов, как их соответствующие трифосфаты, могут являться трифосфатом dTPT3(dTP3TP) и трифосфатом dNaM(dNaMTP) или dCNMO (dCNMOTP).
[00320] В некоторых вариантах осуществления клетки являются клетками, генетически трансформированными с использованием нуклеиновой кислоты, например, экспрессирующей кассеты, кодирующей транспортер нуклеотидтрифосфатов, способный транспортировать такие неприродные нуклеотиды в клетку. Клетка может содержать гетерологичный транспортер нуклеозидтрифосфата, где гетерологичный транспортер нуклеозидтрифосфата может транспортировать природные и неприродные нуклеозидтрифосфаты в клетку.
[00321] В некоторых случаях способ, представленный в настоящем описании, также включает приведение генетически трансформированной клетки в контакт с соответствующими трифосфатами в присутствии фосфата калия и/или ингибитора фосфатаз или нуклеотидаз. Во время или после такого контакта клетку можно помещать в поддерживающую среду, подходящую для роста и деления клетки. Клетку можно поддерживать в поддерживающей среде таким образом, что соответствующие трифосфатные формы неприродных нуклеотидов встраиваются в нуклеиновые кислоты в клетках, и в ходе по меньшей мере одного цикла деления клетки. Пара неприродных, взаимно спаривающихся нуклеотидов, как их соответствующие трифосфаты, может содержать трифосфат dTPT3 или (dTPT3TP) и трифосфат dCNMO или dNaM(dCNOM или dNaMTP), клетка может являться E.coli, и dTPT3TP и dNaMTP могут импортироваться в E. coli с помощью транспортера PtNTT2, где полимераза E. coli, такая как Pol III или Pol II, может использовать неприродные трифосфаты для репликации ДНК, содержащей UBP, таким образом, встраивая неприродные нуклеотиды и/или неприродные пары оснований в клеточные нуклеиновые кислоты в клеточном микроокружении. Кроме того, в некоторых случаях рибонуклеотиды, такие как NaMTP и TAT1TP, 5FMTP и TPT3TP, импортируются в E. coli с помощью транспортера PtNTT2.
[00322] Настоящее изобретение относится к композициям и способам, включающим использование трех или более неприродных спаривающихся нуклеотидов. В некоторых случаях такие спаривающиеся нуклеотиды проникают в клетку с помощью транспортеров нуклеотидов, или используют стандартные способы трансформации нуклеиновых кислот, известные в этой области (например, электропорацию, химическую трансформацию или другой способ). В некоторых случаях спаривающийся неприродный нуклеотид проникает в клетку как часть полинуклеотида, такого как плазмида. Один или более спаривающихся неприродных нуклеотидов, проникающих в клетку как часть полинуклеотида (РНК или ДНК), могут не реплицироваться сами по себе in vivo. Например, двухцепочечную плазмиду ДНК или другую нуклеиновую кислоту, содержащую первый неприродный дезоксирибонуклеотид и второй неприродный дезоксирибонуклеотид с основаниями, сконфигурированными для образования первой неприродной пары оснований, подвергают электропорации в клетку. Клеточные среды обрабатывают третьим неприродным дезоксирибонуклеотидом, четвертым неприродным дезоксирибонуклеотидом с основаниями, сконфигурированными для образования второй неприродной пары оснований друг с другом, где основание первого неприродного дезоксирибонуклеотида и основание третьего неприродного дезоксирибонуклеотида образуют вторую неприродную пару оснований, и где основание второго неприродного дезоксирибонуклеотида и основание четвертого неприродного дезоксирибонуклеотида образуют третью неприродную пару оснований. В некоторых случаях репликация in vivo исходно трансформированной двухцепочечной ДНК плазмиды приводит к получению реплицирующихся впоследствии плазмид, содержащих третий неприродный дезоксирибонуклеотид и четвертый неприродный дезоксирибонуклеотид. Альтернативно или в комбинации, варианты рибонуклеотидов третьего неприродного дезоксирибонуклеотида и четвертого неприродного дезоксирибонуклеотида добавляют в клеточные среды. В некоторых случаях эти рибонуклеотиды встраиваются в РНК, такую как мРНК или тРНК. В некоторых случаях первый, второй, третий и четвертый дезоксинуклеотиды содержат разные основания. В некоторых случаях первый, третий и четвертый дезоксинуклеотиды содержат разные основания. В некоторых случаях первый и третий дезоксинуклеотиды содержат одно и то же основание.
[00323] Осуществляя способ по изобретению на практике, специалист может получать популяцию живых и делящихся клеток, содержащих по меньшей мере один неприродный нуклеотид и/или по меньшей мере одну неприродную пару оснований (UBP) по меньшей мере в одной нуклеиновой кислоте, поддерживаемой по меньшей мере в некоторых из отдельных клеток, где по меньшей мере одна нуклеиновая кислота стабильно реплицируется в клетке, и где клетка экспрессирует транспортер нуклеотидтрифосфатов, подходящий для обеспечения захвата клеткой трифосфатных форм одного или более неприродных нуклеотидов, при приведении в контакт (например, выращивании в присутствии) с неприродными нуклеотидами в поддерживающей среде, подходящей для роста и репликации организма.
[00324] После транспорта в клетку с помощью транспортера нуклеотидтрифосфатов неприродные спаривающиеся нуклеотиды встраиваются в нуклеиновые кислоты в клетке с помощью клеточного аппарата, например, собственных ДНК- и/или РНК-полимераз клетки, гетерологичной полимеразы или полимеразы, полученной с использованием направленной эволюции (Chen T, Romesberg FE, FEBS Lett. 2014 Jan 21;588(2):219-29; Betz K et al., J Am Chem Soc. 2013 Dec 11;135(49):18637-43; описание каждой из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме). Неприродные нуклеотиды могут встраиваться в клеточные нуклеиновые кислоты, такие как геномная ДНК, геномная РНК, мРНК, тРНК, структурная РНК, микроРНК и автономно реплицирующиеся нуклеиновые кислоты (например, плазмиды, вирусы или векторы).
[00325] В некоторых случаях генетически сконструированные клетки получают посредством встраивания в них нуклеиновых кислот, например, гетерологичных нуклеиновых кислот. Любая клетка, представленная в настоящем описании, может являться клеткой-хозяином и может содержать экспрессирующий вектор. В одном из вариантов осуществления клетка-хозяин является прокариотической клеткой. В другом варианте осуществления клетка-хозяин является E. coli. В некоторых вариантах осуществления клетка содержит один или более гетерологичных полинуклеотидов. Реагенты-нуклеиновые кислоты можно встраивать в микроорганизмы различными способами. Неограничивающие примеры способов, используемых для встраивания гетерологичных нуклеиновых кислот в различные организмы, включают: трансформацию, трансфекцию, трансдукцию, электропорацию, опосредованную ультразвуком трансформацию, конъюгацию, бомбардировку микрочастицами и т.п. В некоторых случаях добавление молекул-носителей (например, бис-бензоимидазолиловых соединений, например, см. патент США № 5595899) может повышать захват ДНК клетками, которые, как правило, считают трудными для трансформации общепринятыми способами. Общепринятые способы трансформации доступны специалисту в этой области, и их можно найти в Maniatis, T., E. F. Fritsch and J. Sambrook (1982) Molecular Cloning: a Laboratory Manual; Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., описание которого, таким образом, включено в настоящее описание в качестве ссылки в полном объеме.
[00326] В некоторых случаях генетическую трансформацию осуществляют с использованием прямого переноса экспрессирующей кассеты, в качестве неограничивающих примеров, в плазмидах, вирусных векторах, вирусных нуклеиновых кислотах, фаговых нуклеиновых кислотах, фагах, космидах и искусственных хромосомах или посредством переноса генетического материала в клетках или носителях, таких как катионные липосомы. Такие способы доступны в этой области, и их легко можно адаптировать для использования в способах, представленных в настоящем описании. Векторы для переноса могут являться любой нуклеотидной конструкцией, используемой для доставки генов в клетки (например, плазмидой) или в качестве части общей стратегии доставки генов, например, как часть рекомбинантного ретровируса или аденовируса (Ram et al. Cancer Res. 53:83-88, (1993)). Соответствующие средства для трансфекции, включая вирусные векторы, химические трансфектанты или физико-механические способы, такие как электропорация и направленная диффузия ДНК, описаны, например, в Wolff, J. A., et al., Science, 247, 1465-1468, (1990); и Wolff, J. A. Nature, 352, 815-818, (1991), описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00327] Например, ДНК, кодирующую экспрессирующую кассету и/или вектор транспортера нуклеозидтрифосфата или полимеразы, можно встраивать в клетку любым способом, включая, в качестве неограничивающих примеров, кальций-опосредованную трансформацию, электропорацию, микроинъекцию, липофекцию, бомбардировку микрочастицами и т.п.
[00328] В некоторых случаях клетка содержит неприродные нуклеозидтрифосфаты, встроенные в одну или более нуклеиновых кислот в клетке. Например, клетка может являться живой клеткой, способной к встраиванию по меньшей мере одного неприродного нуклеотида в ДНК или РНК, поддерживаемой в клетке. В клетке также можно встраивать по меньшей мере одну неприродную пару оснований (UBP), включающую пару неприродных, взаимно спаривающихся нуклеотидов, в нуклеиновые кислоты в клетке в условиях in vivo, где неприродные, взаимно образующие пару нуклеотиды, например, их соответствующие трифосфаты, захватываются клеткой под действием транспортера нуклеозидтрифосфата, ген которого присутствует (например, был встроен) в клетке в результате генетической трансформации. Например, после встраивания в нуклеиновую кислоту, поддерживаемую в клетке, dTPT3 и dCNMO могут образовывать стабильную неприродную пару оснований, которые могут стабильно реплицироваться с помощью аппарата репликации ДНК организма, например, при выращивании в поддерживающей среде, содержащей dTPT3TP и dCNMOTP.
[00329] В некоторых случаях клетки могут реплицировать нуклеиновую кислоту, содержащую неприродный нуклеотид. Такие способы могут включать генетическую трансформацию клетки с использованием экспрессирующей кассеты, кодирующей транспортер нуклеозидтрифосфата, способный транспортировать в клетку один или более неприродных нуклеотидов в виде соответствующего трифосфата в условиях in vivo. Альтернативно, можно использовать клетку, ранее генетически трансформированную с использованием экспрессирующей кассеты, которая может экспрессировать кодируемый транспортер нуклеозидтрифосфата. Способ также может включать приведение генетически трансформированной клетки в контакт или подвергание воздействию фосфата калия и соответствующих трифосфатных форм по меньшей мере одного неприродного нуклеотида (например, двух взаимно спаривающихся нуклеотидов, способных образовывать неприродную пару оснований (UBP)) в поддерживающей среде, подходящей для роста и репликации клетки, и поддержание трансформированной клетки в поддерживающей среде в присутствии соответствующих трифосфатных форм по меньшей мере одного неприродного нуклеотида (например, двух взаимно спаривающихся нуклеотидов, способных образовывать неприродную пару оснований (UBP)) в условиях in vivo в результате по меньшей мере одного цикла деления клетки. Способ также может включать приведение генетически трансформированной клетки в контакт или подвергание воздействию фосфата калия и соответствующих трифосфатных форм по меньшей мере одного неприродного нуклеотида (например, двух взаимно спаривающихся нуклеотидов, сконфигурированных для образования неприродной пары оснований (UBP)) в поддерживающей среде, подходящей для роста и деления клетки, и поддержание трансформированной клетки в поддерживающей среде в присутствии соответствующих трифосфатных форм по меньшей мере одного неприродного нуклеотида (например, двух взаимно спаривающихся нуклеотидов, сконфигурированных для образования неприродной пары оснований (UBP)) в условиях in vivo в результате по меньшей мере одного цикла деления клетки.
[00330] В некоторых вариантах осуществления клетка содержит стабильно встроенную неприродную нуклеиновую кислоту. Некоторые варианты осуществления включают клетку (например, E. coli), имеющую стабильно встроенные нуклеотиды, иные, чем A, G, T и C, в нуклеиновых кислотах, поддерживаемых в клетке. Например, нуклеотиды, иные, чем A, G, T и C, могут являться d5SICS, dCNMO, dNaM и/или dTPT3, которые после встраивания в нуклеиновые кислоты клетки, могут образовывать стабильную неприродную пару оснований в нуклеиновых кислотах. В одном из аспектов неприродные нуклеотиды и неприродные пары оснований могут стабильно реплицироваться с помощью аппарата репликации организма, если организм, трансформированный с использованием гена транспортера трифосфата, выращивают в поддерживающей среде, включающей фосфат калия и трифосфатные формы d5SICS, dNaM, dCNMO и/или dTPT3.
[00331] В некоторых случаях клетка имеет расширенный генетический алфавит. Клетка может содержать стабильно встроенную неприродную нуклеиновую кислоту. В некоторых вариантах осуществления клетка с расширенным генетическим алфавитом содержит неприродную нуклеиновую кислоту, содержащую неприродный нуклеотид, который может образовывать пару с другим неприродным нуклеотидом. В некоторых вариантах осуществления клетка с расширенным генетическим алфавитом содержит неприродную нуклеиновую кислоту, связанную водородной связью с другой нуклеиновой кислотой. В некоторых вариантах осуществления клетка с расширенным генетическим алфавитом содержит неприродную нуклеиновую кислоту, не связанную водородной связью с другой нуклеиновой кислотой, с которой она спарена. В некоторых вариантах осуществления клетка с расширенным генетическим алфавитом содержит неприродную нуклеиновую кислоту, содержащую неприродный нуклеотид с нуклеиновым основанием, образующим пару с нуклеиновым основанием другого неприродного нуклеотида посредством гидрофобных и/или упаковывающих взаимодействий. В некоторых вариантах осуществления клетка с расширенным генетическим алфавитом содержит неприродную нуклеиновую кислоту, образующую пару оснований с другой нуклеиновой кислотой посредством неводородных взаимодействий. Клетка с расширенным генетическим алфавитом может являться клеткой, которая может копировать гомологичную нуклеиновую кислоту с образованием нуклеиновой кислоты, содержащей неприродную нуклеиновую кислоту. Клетка с расширенным генетическим алфавитом может являться клеткой, содержащей неприродную нуклеиновую кислоту, спаренную с другой неприродной нуклеиновой кислотой (неприродной парой оснований нуклеиновой кислоты (UBP)).
[00332] В некоторых вариантах осуществления клетки образуют неприродные пары оснований ДНК (UBP) из импортированных неприродных нуклеотидов в условиях in vivo. В некоторых вариантах осуществления действие фосфата калия и/или ингибиторов фосфатазы и/или нуклеотидазы может облегчать транспорт неприродных нуклеотидов. Способы включают использование клетки, экспрессирующей гетерологичный транспортер нуклеозидтрифосфата. Если такую клетку приводят в контакт с одним или более нуклеозидтрифосфатами, нуклеозидтрифосфаты транспортируются в клетку. Клетка может находиться совместно с фосфатом калия и/или ингибиторами фосфатаз и нуклеотидаз. Неприродные нуклеозидтрифосфаты могут встраиваться в нуклеиновые кислоты в клетке под действием природного аппарата клетки (т.е. полимераз) и, например, взаимно образовывать неприродные пары оснований в нуклеиновых кислотах клетки. В некоторых вариантах осуществления UBP образуются между нуклеотидами ДНК и РНК, несущими неприродные основания.
[00333] В некоторых вариантах осуществления UBP можно встраивать в клетку или популяцию клеток под воздействием неприродных трифосфатов. В некоторых вариантах осуществления UBP можно встраивать в клетку или популяцию клеток при, по существу, непрерывном воздействии неприродных трифосфатов.
[00334] В некоторых вариантах осуществления индукция экспрессии гетерологичного гена, например, транспортера нуклеозидтрифосфата (NTT), в клетке может приводить к более медленному росту клеток и повышенному захвату неприродных трифосфатов по сравнению с ростом и захватом одного или более неприродных трифосфатов в клетке без индукции экспрессии гетерологичного гена. Захват в разной степени включает транспорт нуклеотидов в клетку, например, посредством диффузии, осмоса или под действием транспортеров. В некоторых вариантах осуществления индукция экспрессии гетерологичного гена, например, NTT, в клетке может приводить к повышенному росту клеток и повышенному захвату неприродной нуклеиновой кислоты по сравнению с ростом и захватом клеткой без индукции экспрессии гетерологичного гена
[00335] В некоторых вариантах осуществления UBP встраивается во время логарифмической фазы роста. В некоторых вариантах осуществления UBP встраивается во время нелогарифмической фазы роста. В некоторых вариантах осуществления UBP встраивается во время, по существу, линейной фазы роста. В некоторых вариантах осуществления UBP стабильно встраивается в клетку или популяцию клеток после выращивания в течение периода времени. Например, UBP может стабильно встраиваться в клетку или популяцию клеток после выращивания в течение по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45 или 50 или более удвоений. Например, UBP может стабильно встраиваться в клетку или популяцию клеток после выращивания в течение по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23 или 24 часов. Например, UBP может стабильно встраиваться в клетку или популяцию клеток после выращивания в течение по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 или 31 дня. Например, UBP может стабильно встраиваться в клетку или популяцию клеток после выращивания в течение по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 месяцев. Например, UBP может стабильно встраиваться в клетку или популяцию клеток после выращивания в течение по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45 или 50 лет.
[00336] В некоторых вариантах осуществления полусинтетический организм, представленный в настоящем описании, содержит ДНК, содержащую по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
 и
и  . В некоторых вариантах осуществления ДНК полусинтетического организма, содержащая по меньшей мере одно из неприродных оснований, образует неприродную пару оснований (UBP). В некоторых вариантах осуществления неприродная пара оснований (UBP) представляет собой dCNMO-dTPT3, dNaM-dTPT3, dCNMO-dTAT1 или dNaM-dTAT1. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
. В некоторых вариантах осуществления ДНК полусинтетического организма, содержащая по меньшей мере одно из неприродных оснований, образует неприродную пару оснований (UBP). В некоторых вариантах осуществления неприродная пара оснований (UBP) представляет собой dCNMO-dTPT3, dNaM-dTPT3, dCNMO-dTAT1 или dNaM-dTAT1. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
и . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
, , и . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
 . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:  . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из и . В некоторых вариантах осуществления ДНК содержит по меньшей мере два неприродных нуклеиновых основания, выбранных из и . В некоторых вариантах осуществления ДНК содержит две цепи, первая цепь содержит по меньшей мере одно нуклеиновое основание, являющееся , и вторая цепь содержит по меньшей мере одно нуклеиновое основание, являющееся . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, являющееся .
. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из и . В некоторых вариантах осуществления ДНК содержит по меньшей мере два неприродных нуклеиновых основания, выбранных из и . В некоторых вариантах осуществления ДНК содержит две цепи, первая цепь содержит по меньшей мере одно нуклеиновое основание, являющееся , и вторая цепь содержит по меньшей мере одно нуклеиновое основание, являющееся . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, являющееся .
[00337] В некоторых вариантах осуществления клетка дополнительно использует РНК-полимеразу для образования мРНК, содержащей один или более неприродных нуклеотидов. В некоторых вариантах осуществления РНК-полимераза является гетерологичной РНК-полимеразой. В некоторых случаях клетка дополнительно использует полимеразу для образования тРНК, содержащей антикодон, содержащий один или более неприродных нуклеотидов. В некоторых вариантах осуществления тРНК является гетерологичной тРНК. В некоторых случаях тРНК заряжена неприродной аминокислотой. В некоторых случаях неприродный антикодон тРНК спаривается с неприродным кодоном мРНК во время трансляции для синтеза белка, содержащего неприродную аминокислоту.
[00338] В некоторых вариантах осуществления полусинтетический организм, представленный в настоящем описании, экспрессирует гетерологичный транспортер нуклеозидтрифосфата. В некоторых вариантах осуществления гетерологичный транспортер нуклеозидтрифосфата является PtNTT2. В некоторых вариантах осуществления полусинтетический организм дополнительно экспрессирует гетерологичную тРНК-синтетазу. В некоторых вариантах осуществления гетерологичная тРНК-синтетаза является пирролизил-тРНК-синтетазой M. barkeri (Mb PylRS). В некоторых вариантах осуществления полусинтетический организм экспрессирует транспортер нуклеозидтрифосфата PtNTT2 и дополнительно экспрессирует тРНК-синтетазу пирролизил-тРНК-синтетазу M. barkeri (Mb PylRS). В некоторых вариантах осуществления полусинтетический организм дополнительно экспрессирует гетерологичную РНК-полимеразу. В некоторых вариантах осуществления гетерологичная РНК-полимераза является RNAP T7. В некоторых вариантах осуществления полусинтетический организм не экспрессирует белок, имеющий функцию рекомбинационной репарации ДНК. В некоторых вариантах осуществления полусинтетический организм является E. coli, и организм не экспрессирует RecA.
[00339] В некоторых вариантах осуществления полусинтетический организм дополнительно содержит гетерологичную мРНК. В некоторых вариантах осуществления гетерологичная мРНК содержит по меньшей мере одно неприродное основание, выбранное из , и . В некоторых вариантах осуществления гетерологичная мРНК содержит по меньшей мере одно неприродное основание, являющееся. В некоторых вариантах осуществления гетерологичная мРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления гетерологичная мРНК содержит по меньшей мере одно неприродное основание, являющееся .
[00340] В некоторых вариантах осуществления полусинтетический организм дополнительно содержит гетерологичную тРНК. В некоторых вариантах осуществления гетерологичная тРНК содержит по меньшей мере одно неприродное основание, выбранное из , и . В некоторых вариантах осуществления гетерологичная тРНК содержит по меньшей мере одно неприродное основание, являющееся. В некоторых вариантах осуществления гетерологичная тРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления гетерологичная тРНК содержит по меньшей мере одно неприродное основание, являющееся .
[00341] В некоторых вариантах осуществления полусинтетический организм, представленный в настоящем описании, дополнительно содержит гетерологичную мРНК и гетерологичную тРНК. В некоторых вариантах осуществления полусинтетический организм дополнительно содержит (a) гетерологичный транспортер нуклеозидтрифосфата, (b) гетерологичную мРНК, (c) гетерологичную тРНК, (d) гетерологичную тРНК-синтетазу и (e) гетерологичную РНК-полимеразу, и где организм не экспрессирует белок, имеющий функцию рекомбинационной репарации ДНК. В некоторых вариантах осуществления транспортер нуклеозидтрифосфата является PtNTT2, тРНК-синтетаза является пирролизил-тРНК-синтетазой M. barkeri (Mb PylRS), и РНК-полимераза является RNAP T7. В некоторых вариантах осуществления полусинтетический организм является E. coli, и организм не экспрессирует RecA. В некоторых вариантах осуществления полусинтетический организм гиперэкспрессирует одну или более ДНК-полимераз. В некоторых вариантах осуществления организм гиперэкспрессирует ДНК-полимеразу II.
Природные и неприродные аминокислоты
[00342] В рамках изобретения термин "аминокислотный остаток" может относиться к молекуле, содержащей аминогруппу и карбоксильную группу. Подходящие аминокислоты включают, в качестве неограничивающих примеров, D- и L-изомеры природных аминокислот, а также неприродные аминокислоты, полученные посредством органического синтеза или любым другим способом. В рамках изобретения термин "аминокислота" включает, в качестве неограничивающих примеров, α-аминокислоты, β-аминокислоты, природные аминокислоты, неканонические аминокислоты, неприродные аминокислоты и аналоги аминокислот.
[00343] Термин "α-аминокислота" может относиться к молекуле, содержащей аминогруппу и карбоксильную группу, связанную с углеродом, обозначаемым как α-углерод. Например:
 .
.
[00344] Термин "β-аминокислота" может относиться к молекуле, содержащей аминогруппу и карбоксильную группу в β-конфигурации.
[00345] Термин "природная аминокислота" может относиться к любой из двадцати аминокислот, обычно обнаруживаемых в пептидах, синтезируемых в природе, и известных по однобуквенным сокращениям A, R, N, C, D, Q, E, G, H, I, L, K, M, F, P, S, T, W, Y и V.
[00346] В следующей таблице показаны свойства природных аминокислот:
Нейтральный (90%)
[00347] "Гидрофобные аминокислоты" включают небольшие гидрофобные аминокислоты и крупные гидрофобные аминокислоты. "Небольшая гидрофобная аминокислота" может представлять собой глицин, аланин, пролин и их аналоги. "Крупные гидрофобные аминокислоты" могут представлять собой валин, лейцин, изолейцин, фенилаланин, метионин, триптофан и их аналоги. "Полярные аминокислоты" может представлять собой серин, треонин, аспарагин, глутамин, цистеин, тирозин и их аналоги. "Заряженные аминокислоты" может представлять собой лизин, аргинин, гистидин, аспартат, глутамат и их аналоги.
[00348] "Аналог аминокислоты" может являться молекулой, структурно схожей с аминокислотой, и которая может замещать аминокислоту при образовании пептидомиметического макроцикла. Аналоги аминокислот включают, в качестве неограничивающих примеров, β-аминокислоты и аминокислоты, в которых аминогруппа или карбоксильная группа замещена схожей реакционноспособной группой (например, замена первичного амина вторичным или третичным амином или замена карбоксильной группы сложным эфиром).
[00349] "Неканоническая аминокислота (ncAA)" или "неприродная аминокислота" может являться аминокислотой, не являющейся одной из двадцати аминокислот, обычно обнаруживаемых в пептидах, синтезируемых в природе, и известных по однобуквенным сокращениям A, R, N, C, D, Q, E, G, H, I, L, K, M, F, P, S, T, W, Y и V. В некоторых случаях неприродные аминокислоты представляют собой подгруппу неканонических аминокислот.
[00350] Аналоги аминокислот могут включать аналоги β-аминокислот. Неограничивающие примеры аналогов β-аминокислот включают следующие: циклические аналоги β-аминокислот; β-аланин; (R)-β-фенилаланин; (R)-1,2,3,4-тетрагидро-изохинолин-3-уксусную кислоту; (R)-3-амино-4-(1-нафтил)-масляную кислоту; (R)-3-амино-4-(2,4-дихлорфенил)масляную кислоту; (R)-3-амино-4-(2-хлорфенил)-масляную кислоту; (R)-3-амино-4-(2-цианофенил)-масляную кислоту; (R)-3-амино-4-(2-фторфенил)-масляную кислоту; (R)-3-амино-4-(2-фурил)-масляную кислоту; (R)-3-амино-4-(2-метилфенил)-масляную кислоту; (R)-3-амино-4-(2-нафтил)-масляную кислоту; (R)-3-амино-4-(2-тиенил)-масляную кислоту; (R)-3-амино-4-(2-трифторметилфенил)-масляную кислоту; (R)-3-амино-4-(3,4-дихлорфенил)масляную кислоту; (R)-3-амино-4-(3,4-дифторфенил)масляную кислоту; (R)-3-амино-4-(3-бензотиенил)-масляную кислоту; (R)-3-амино-4-(3-хлорфенил)-масляную кислоту; (R)-3-амино-4-(3-цианофенил)-масляную кислоту; (R)-3-амино-4-(3-фторфенил)-масляную кислоту; (R)-3-амино-4-(3-метилфенил)-масляную кислоту; (R)-3-амино-4-(3-пиридил)-масляную кислоту; (R)-3-амино-4-(3-тиенил)-масляную кислоту; (R)-3-амино-4-(3-трифторметилфенил)-масляную кислоту; (R)-3-амино-4-(4-бромфенил)-масляную кислоту; (R)-3-амино-4-(4-хлорфенил)-масляную кислоту; (R)-3-амино-4-(4-цианофенил)-масляную кислоту; (R)-3-амино-4-(4-фторфенил)-масляную кислоту; (R)-3-амино-4-(4-йодфенил)-масляную кислоту; (R)-3-амино-4-(4-метилфенил)-масляную кислоту; (R)-3-амино-4-(4-нитрофенил)-масляную кислоту; (R)-3-амино-4-(4-пиридил)-масляную кислоту; (R)-3-амино-4-(4-трифторметилфенил)-масляную кислоту; (R)-3-амино-4-пентафтор-фенилмасляную кислоту; (R)-3-амино-5-гексеновую кислоту; (R)-3-амино-5-гексиновую кислоту; (R)-3-амино-5-фенилпентановую кислоту; (R)-3-амино-6-фенил-5-гексеновую кислоту; (S)-1,2,3,4-тетрагидро-изохинолин-3-уксусную кислоту; (S)-3-амино-4-(1-нафтил)-масляную кислоту; (S)-3-амино-4-(2,4-дихлорфенил)масляную кислоту; (S)-3-амино-4-(2-хлорфенил)-масляную кислоту; (S)-3-амино-4-(2-цианофенил)-масляную кислоту; (S)-3-амино-4-(2-фторфенил)-масляную кислоту; (S)-3-амино-4-(2-фурил)-масляную кислоту; (S)-3-амино-4-(2-метилфенил)-масляную кислоту; (S)-3-амино-4-(2-нафтил)-масляную кислоту; (S)-3-амино-4-(2-тиенил)-масляную кислоту; (S)-3-амино-4-(2-трифторметилфенил)-масляную кислоту; (S)-3-амино-4-(3,4-дихлорфенил)масляную кислоту; (S)-3-амино-4-(3,4-дифторфенил)масляную кислоту; (S)-3-амино-4-(3-бензотиенил)-масляную кислоту; (S)-3-амино-4-(3-хлорфенил)-масляную кислоту; (S)-3-амино-4-(3-цианофенил)-масляную кислоту; (S)-3-амино-4-(3-фторфенил)-масляную кислоту; (S)-3-амино-4-(3-метилфенил)-масляную кислоту; (S)-3-амино-4-(3-пиридил)-масляную кислоту; (S)-3-амино-4-(3-тиенил)-масляную кислоту; (S)-3-амино-4-(3-трифторметилфенил)-масляную кислоту; (S)-3-амино-4-(4-бромфенил)-масляную кислоту; (S)-3-амино-4-(4-хлорфенил) масляную кислоту; (S)-3-амино-4-(4-цианофенил)-масляную кислоту; (S)-3-амино-4-(4-фторфенил)масляную кислоту; (S)-3-амино-4-(4-йодфенил)-масляную кислоту; (S)-3-амино-4-(4-метилфенил)-масляную кислоту; (S)-3-амино-4-(4-нитрофенил)-масляную кислоту; (S)-3-амино-4-(4-пиридил)-масляную кислоту; (S)-3-амино-4-(4-трифторметилфенил)-масляную кислоту; (S)-3-амино-4-пентафтор-фенилмасляную кислоту; (S)-3-амино-5-гексеновую кислоту; (S)-3-амино-5-гексиновую кислоту; (S)-3-амино-5-фенилпентановую кислоту; (S)-3-амино-6-фенил-5-гексеновую кислоту; 1,2,5,6-тетрагидропиридин-3-карбоновую кислоту; 1,2,5,6-тетрагидропиридин-4-карбоновую кислоту; 3-амино-3-(2-хлорфенил)-пропионовую кислоту; 3-амино-3-(2-тиенил)-пропионовую кислоту; 3-амино-3-(3-бромфенил)-пропионовую кислоту; 3-амино-3-(4-хлорфенил)-пропионовую кислоту; 3-амино-3-(4-метоксифенил)-пропионовую кислоту; 3-амино-4,4,4-трифтор-масляную кислоту; 3-аминоадипиновую кислоту; D-β-фенилаланин; β-лейцин; L-β-гомоаланин; γ-бензиловый сложный эфир L-β-гомоаспарагиновой кислоты; δ-бензиловый сложный эфир L-β-гомоглутаминовой кислоты; L-β-гомоизолейцин; L-β-гомолейцин; L-β-гомометионин; L-β-гомофенилаланин; L-β-гомопролин; L-β-гомотриптофан; L-β-гомовалин; L-Nω-бензилоксикарбонил-β-гомолизин; Nω-L-β-гомоаргинин; O-бензил-L-β-гомогидроксипролин; O-бензил-L-β-гомосерин; O-бензил-L-β-гомотреонин; O-бензил-L-β-гомотирозин; γ-тритил-L-β-гомоаспарагин; (R)-β-фенилаланин; γ-трет-бутиловый сложный эфир L-β-гомоаспарагиновой кислоты; δ-трет-бутиловый сложный эфир L-β-гомоглутаминовой кислоты; L-Nω-β-гомолизин; Nδ-тритил-L-β-гомоглутамин; Nω-2,2,4,6,7-пентаметил-дигидробензофуран-5-сульфонил-L-β-гомоаргинин; O-трет-бутил-L-β-гомогидрокси-пролин; O-трет-бутил-L-β-гомосерин; O-трет-бутил-L-β-гомотреонин; O-трет-бутил-L-β-гомотирозин; 2-аминоциклопентан-карбоновую кислоту и 2-аминоциклогексан-карбоновую кислоту.
[00351] Аналоги аминокислот могут включать аналоги аланина, валина, глицина или лейцина. Неограничивающие примеры аналогов аминокислот аланина, валина, глицина и лейцина включают следующие: α-метоксиглицин; α-аллил-L-аланин; α-аминоизомасляную кислоту; α-метил-лейцин; β-(1-нафтил)-D-аланин; β-(1-нафтил)-L-аланин; β-(2-нафтил)-D-аланин; β-(2-нафтил)-L-аланин; β-(2-пиридил)-D-аланин; β-(2-пиридил)-L-аланин; β-(2-тиенил)-D-аланин; β-(2-тиенил)-L-аланин; β-(3-бензотиенил)-D-аланин; β-(3-бензотиенил)-L-аланин; β-(3-пиридил)-D-аланин; β-(3-пиридил)-L-аланин; β-(4-пиридил)-D-аланин; β-(4-пиридил)-L-аланин; β-хлор-L-аланин; β-циано-L-аланин; β-циклогексил-D-аланин; β-циклогексил-L-аланин; β-циклопентен-1-ил-аланин; β-циклопентил-аланин; соль β-циклопропил-L-Ala-OH.дициклогексиламмония; β-трет-бутил-D-аланин; β-трет-бутил-L-аланин; γ-аминомасляную кислоту; L-α,β-диаминопропионовую кислоту; 2,4-динитро-фенилглицин; 2,5-дигидро-D-фенилглицин; 2-амино-4,4,4-трифтормасляную кислоту; 2-фтор-фенилглицин; 3-амино-4,4,4-трифтор-масляную кислоту; 3-фтор-валин; 4,4,4-трифтор-валин; соль 4,5-дегидро-L-leu-OH.дициклогексиламмония; 4-фтор-D-фенилглицин; 4-фтор-L-фенилглицин; 4-гидрокси-D-фенилглицин; 5,5,5-трифтор-лейцин; 6-аминогексановую кислоту; соль циклопентил-D-Gly-OH.дициклогексиламмония; соль циклопентил-Gly-OH.дициклогексиламмония; D-α,β-диаминопропионовую кислоту; D-α-аминомасляную кислоту; D-α-трет-бутилглицин; D-(2-тиенил)глицин; D-(3-тиенил)глицин; D-2-аминокапроновую кислоту; D-2-инданилглицин; соль D-аллилглицин-дициклогексиламмония; D-циклогексилглицин; D-норвалин; D-фенилглицин; β-аминомасляную кислоту; β-аминоизомасляную кислоту; (2-бромфенил)глицин; (2-метоксифенил)глицин; (2-метилфенил)глицин; (2-тиазолил)глицин; (2-тиенил)глицин; 2-амино-3-(диметиламино)-пропионовую кислоту; L-α,β-диаминопропионовую кислоту; L-α-аминомасляную кислоту; L-α-трет-бутилглицин; L-(3-тиенил)глицин; L-2-амино-3-(диметиламино)-пропионовую кислоту; соль L-2-аминокапроновой кислоты и дициклогексиламмония; L-2-инданилглицин; соль L-аллилглициндициклогексиламмония; L-циклогексилглицин; L-фенилглицин; L-пропаргилглицин; L-норвалин; N-α-аминометил-L-аланин; D-α,γ-диаминомасляную кислоту; L-α,γ-диаминомасляную кислоту; β-циклопропил-L-аланин; (N-β-(2,4-динитрофенил))-L-α,β-диаминопропионовую кислоту; (N-β-1-(4,4-диметил-2,6-диоксоциклогекс-1-илиден)этил)-D-α,β-диаминопропионовую кислоту; (N-β-1-(4,4-диметил-2,6-диоксоциклогекс-1-илиден)этил)-L-α,β-диаминопропионовую кислоту; (N-β-4-метилтритил)-L-α,β-диаминопропионовую кислоту; (N-β-аллилоксикарбонил)-L-α,β-диаминопропионовую кислоту; (N-γ-1-(4,4-диметил-2,6-диоксоциклогекс-1-илиден)этил)-D-α,γ-диаминомасляную кислоту; (N-γ-1-(4,4-диметил-2,6-диоксоциклогекс-1-илиден)этил)-L-α,γ-диаминомасляную кислоту; (N-γ-4-метилтритил)-D-α,γ-диаминомасляную кислоту; (N-γ-4-метилтритил)-L-α,γ-диаминомасляную кислоту; (N-γ-аллилоксикарбонил)-L-α,γ-диаминомасляную кислоту; D-α,γ-диаминомасляную кислоту; 4,5-дегидро-L-лейцин; циклопентил-D-Gly-OH; циклопентил-Gly-OH; D-аллилглицин; D-гомоциклогексилаланин; L-1-пиренилаланин; L-2-аминокапроновую кислоту; L-аллилглицин; L-гомоциклогексилаланин и N-(2-гидрокси-4-метокси-Bzl)-Gly-OH.
[00352] Аналоги аминокислот могут включать аналоги аргинина или лизина. Неограничивающие примеры аналогов аминокислот аргинина и лизина включают следующие: цитруллин; L-2-амино-3-гуанидинопропионовую кислоту; L-2-амино-3-уреидопропионовую кислоту; L-цитруллин; Lys(Me)2-OH; Lys(N3)-OH; Nδ-бензилоксикарбонил-L-орнитин; Nω-нитро-D-аргинин; Nω-нитро-L-аргинин; α-метил-орнитин; 2,6-диаминопимелиновую кислоту; L-орнитин; (Nδ-1-(4,4-диметил-2,6-диоксо-циклогекс-1-илиден)этил)-D-орнитин; (Nδ-1-(4,4-диметил-2,6-диоксо-циклогекс-1-илиден)этил)-L-орнитин; (Nδ-4-метилтритил)-D-орнитин; (Nδ-4-метилтритил)-L-орнитин; D-орнитин; L-орнитин; Arg(Me)(Pbf)-OH; Arg(Me)2-OH (асимметричный); Arg(Me)2-OH (симметричный); Lys(ivDde)-OH; Lys(Me)2-OH.HCl; хлорид Lys(Me3)-OH; Nω-нитро-D-аргинин и Nω-нитро-L-аргинин.
[00353] Аналоги аминокислот могут включать аналоги аспарагиновой или глутаминовой кислот. Неограничивающие примеры аналогов аминокислот аспарагиновой или глутаминовой кислот включают следующие: α-метил-D-аспарагиновую кислоту; α-метил-глутаминовую кислоту; α-метил-L-аспарагиновую кислоту; γ-метилен-глутаминовую кислоту; (N-γ-этил)-L-глутамин; [N-α-(4-аминобензоил)]-L-глутаминовую кислоту; 2,6-диаминопимелиновую кислоту; L-α-аминосубериновую кислоту; D-2-аминоадипиновую кислоту; D-α-аминосубериновую кислоту; α-аминопимелиновую кислоту; иминодиуксусную кислоту; L-2-аминоадипиновую кислоту; трео-β-метил-аспарагиновую кислоту; γ,γ-ди-трет-бутиловый сложный эфир γ-карбокси-D-глутаминовой кислоты; γ,γ-ди-трет-бутиловый сложный эфир γ-карбокси-L-глутаминовой кислоты; Glu(OAll)-OH; L-Asu(OtBu)-OH и пироглутаминовую кислоту.
[00354] Аналоги аминокислот могут включать аналоги цистеина и метионина. Неограничивающие примеры аналогов аминокислот цистеина и метионина включают следующие: Cys(фарнезил)-OH, Cys(фарнезил)-OMe, α-метил-метионин, Cys(2-гидроксиэтил)-OH, Cys(3-аминопропил)-OH, 2-амино-4-(этилтио)масляную кислоту, бутионин, бутионинсульфоxимин, этионин, хлорид метионин-метилсульфония, селенометионин, цистеиновую кислоту, [2-(4-пиридил)этил]-DL-пеницилламин, [2-(4-пиридил)этил]-L-цистеин, 4-метоксибензил-D-пеницилламин, 4-метоксибензил-L-пеницилламин, 4-метилбензил-D-пеницилламин, 4-метилбензил-L-пеницилламин, бензил-D-цистеин, бензил-L-цистеин, бензил-DL-гомоцистеин, карбамоил-L-цистеин, карбоксиэтил-L-цистеин, карбоксиметил-L-цистеин, дифенилметил-L-цистеин, этил-L-цистеин, метил-L-цистеин, трет-бутил-D-цистеин, тритил-L-гомоцистеин, тритил-D-пеницилламин, цистатионин, гомоцистин, L-гомоцистин, (2-аминоэтил)-L-цистеин, селено-L-цистин, цистатионин, Cys(StBu)-OH и ацетамидометил-D-пеницилламин.
[00355] Аналоги аминокислот могут включать аналоги фенилаланина и тирозина. Неограничивающие примеры аналогов аминокислот фенилаланина и тирозина включают следующие: β-метил-фенилаланин, β-гидроксифенилаланин, α-метил-3-метокси-DL-фенилаланин, α-метил-D-фенилаланин, α-метил-L-фенилаланин, 1,2,3,4-тетрагидроизохинолин-3-карбоновую кислоту, 2,4-дихлор-фенилаланин, 2-(трифторметил)-D-фенилаланин, 2-(трифторметил)-L-фенилаланин, 2-бром-D-фенилаланин, 2-бром-L-фенилаланин, 2-хлор-D-фенилаланин, 2-хлор-L-фенилаланин, 2-циано-D-фенилаланин, 2-циано-L-фенилаланин, 2-фтор-D-фенилаланин, 2-фтор-L-фенилаланин, 2-метил-D-фенилаланин, 2-метил-L-фенилаланин, 2-нитро-D-фенилаланин, 2-нитро-L-фенилаланин, 2,4,5-тригидрокси-фенилаланин, 3,4,5-трифтор-D-фенилаланин, 3,4,5-трифтор-L-фенилаланин, 3,4-дихлор-D-фенилаланин, 3,4-дихлор-L-фенилаланин, 3,4-дифтор-D-фенилаланин, 3,4-дифтор-L-фенилаланин, 3,4-дигидрокси-L-фенилаланин, 3,4-диметокси-L-фенилаланин, 3,5,3′-трийод-L-тиронин, 3,5-дийод-D-тирозин, 3,5-дийод-L-тирозин, 3,5-дийод-L-тиронин, 3-(трифторметил)-D-фенилаланин, 3-(трифторметил)-L-фенилаланин, 3-амино-L-тирозин, 3-бром-D-фенилаланин, 3-бром-L-фенилаланин, 3-хлор-D-фенилаланин, 3-хлор-L-фенилаланин, 3-хлор-L-тирозин, 3-циано-D-фенилаланин, 3-циано-L-фенилаланин, 3-фтор-D-фенилаланин, 3-фтор-L-фенилаланин, 3-фтор-тирозин, 3-йодо-D-фенилаланин, 3-йодо-L-фенилаланин, 3-йодо-L-тирозин, 3-метокси-L-тирозин, 3-метил-D-фенилаланин, 3-метил-L-фенилаланин, 3-нитро-D-фенилаланин, 3-нитро-L-фенилаланин, 3-нитро-L-тирозин, 4-(трифторметил)-D-фенилаланин, 4-(трифторметил)-L-фенилаланин, 4-амино-D-фенилаланин, 4-амино-L-фенилаланин, 4-бензоил-D-фенилаланин, 4-бензоил-L-фенилаланин, 4-бис(2-хлорэтил)амино-L-фенилаланин, 4-бром-D-фенилаланин, 4-бром-L-фенилаланин, 4-хлор-D-фенилаланин, 4-хлор-L-фенилаланин, 4-циано-D-фенилаланин, 4-циано-L-фенилаланин, 4-фтор-D-фенилаланин, 4-фтор-L-фенилаланин, 4-йодо-D-фенилаланин, 4-йодо-L-фенилаланин, гомофенилаланин, тироксин, 3,3-дифенилаланин, тиронин, этилтирозин и метилтирозин.
[00356] Аналоги аминокислот могут включать аналоги пролина. Неограничивающие примеры аналогов аминокислоты пролина включают следующие: 3,4-дегидро-пролин, 4-фтор-пролин, цис-4-гидрокси-пролин, тиазолидин-2-карбоновую кислоту и транс-4-фтор-пролин.
[00357] Аналоги аминокислот могут включать аналоги серина и треонина. Неограничивающие примеры аналогов аминокислот серина и треонина включают следующие: 3-амино-2-гидрокси-5-метилгексановую кислоту, 2-амино-3-гидрокси-4-метилпентановую кислоту, 2-амино-3-этоксибутановую кислоту, 2-амино-3-метоксибутановую кислоту, 4-амино-3-гидрокси-6-метилгептановую кислоту, 2-амино-3-бензилоксипропионовую кислоту, 2-амино-3-бензилоксипропионовую кислоту, 2-амино-3-этоксипропионовую кислоту, 4-амино-3-гидроксибутановую кислоту и α-метилсерин.
[00358] Аналоги аминокислот могут включать аналоги триптофана. Неограничивающие примеры аналогов аминокислоты триптофана включают следующие: α-метил-триптофан; β-(3-бензотиенил)-D-аланин; β-(3-бензотиенил)-L-аланин; 1-метил-триптофан; 4-метил-триптофан; 5-бензилокси-триптофан; 5-бром-триптофан; 5-хлор-триптофан; 5-фтор-триптофан; 5-гидрокси-триптофан; 5-гидрокси-L-триптофан; 5-метокси-триптофан; 5-метокси-L-триптофан; 5-метил-триптофан; 6-бром-триптофан; 6-хлор-D-триптофан; 6-хлор-триптофан; 6-фтор-триптофан; 6-метил-триптофан; 7-бензилокси-триптофан; 7-бром-триптофан; 7-метил-триптофан; D-1,2,3,4-тетрагидро-норгарман-3-карбоновую кислоту; 6-метокси-1,2,3,4-тетрагидроноргарман-1-карбоновую кислоту; 7-азатриптофан; L-1,2,3,4-тетрагидро-норгарман-3-карбоновую кислоту; 5-метокси-2-метил-триптофан и 6-хлор-L-триптофан.
[00359] Аналоги аминокислот могут являться рацемическими. В некоторых случаях используют D-изомер аналога аминокислоты. В некоторых случаях используют L-изомер аналога аминокислоты. В некоторых случаях аналог аминокислоты содержит хиральные центры, находящиеся в R- или S-конфигурации. В некоторых случаях аминогруппы аналогов β-аминокислот замещены защитной группой, например, трет-бутилоксикарбонилом (группой BOC), 9-флуоренилметилоксикарбонилом (FMOC), тозилом и т.п. В некоторых случаях функциональная группа карбоновой кислоты аналога β-аминокислоты защищена, например, в виде сложноэфирного производного. В некоторых случаях используют соль аналога аминокислоты.
[00360] В некоторых вариантах осуществления неприродная аминокислота является неприродной аминокислотой, описанной в Liu C.C., Schultz, P.G. Annu. Rev. Biochem. 2010, 79, 413, описание которой, таким образом, включено в настоящее изобретение в качестве ссылки в полном объеме. В некоторых вариантах осуществления неприродная аминокислота включает N6(2-азидоэтокси)-карбонил-L-лизин.
[00361] В некоторых вариантах осуществления аминокислотный остаток, представленный в настоящем описании, (например, в белке) подвергают мутагенезу в неприродную аминокислоту перед связыванием с конъюгирующим остатком. В некоторых случаях мутация в неприродную аминокислоту предотвращает или минимизирует ответ иммунной системы на аутоантигены. В рамках изобретения термин "неприродная аминокислота" относится к аминокислоте, иной, чем 20 аминокислот, встречающихся в природе в белке. Неограничивающие примеры неприродных аминокислот включают: p-ацетил-L-фенилаланин, p-йодо-L-фенилаланин, p-метоксифенилаланин, O-метил-L-тирозин, p-пропаргилоксифенилаланин, p-пропаргил-фенилаланин, L-3-(2-нафтил)аланин, 3-метил-фенилаланин, O-4-аллил-L-тирозин, 4-пропил-L-тирозин, три-O-ацетил-GlcNAcp-серин, L-ДОФА, фторированный фенилаланин, изопропил-L-фенилаланин, p-азидо-L-фенилаланин, p-ацил-L-фенилаланин, p-бензоил-L-фенилаланин, p-боронофенилаланин, O-пропаргилтирозин, L-фосфосерин, фосфоносерин, фосфонотирозин, p-бромфенилаланин, селеноцистеин, p-амино-L-фенилаланин, изопропил-L-фенилаланин, азидо-лизин (N6-азидоэтокси-карбонил-L-лизин, AzK), неприродный аналог аминокислоты тирозина; неприродный аналог аминокислоты глутамина; неприродный аналог аминокислоты фенилаланина; неприродный аналог аминокислоты серина; неприродный аналог аминокислоты треонина; алкил-, арил-, ацил-, азидо-, циано-, галоген-, гидразин-, гидразид-, гидроксил-, алкенил-, алкинил-, простой эфир, тиол-, сульфонил-, селено-, сложный эфир, тиокислота-, борат-, боронат-, фосфо-, фосфоно-, фосфин-, гетероцикл-, енон-, имин-, альдегид-, гидроксиламин-, кето- или амино-замещенную аминокислоту или их комбинацию; аминокислоту с фотоактивируемым кросслинкером; аминокислоту со спиновой меткой; флуоресцентную аминокислоту; металл-связывающую аминокислоту; металл-содержащую аминокислоту; радиоактивную аминокислоту; фотолабильную и/или фотоизомеризуемую аминокислоту; аминокислоту, содержащую биотин или аналог биотина; кето-содержащую аминокислоту; аминокислоту, содержащую полиэтиленгликоль или полиэфир; аминокислоту, замещенную тяжелым атомом; химически расщепляемую или фоторасщепляемую аминокислоту; аминокислоту с удлиненной боковой цепью; аминокислоту, содержащую токсичную группу; сахар-замещенную аминокислоту; углерод-связанную сахар-содержащую аминокислоту; редокс-активную аминокислоту; α-гидрокси-содержащую кислоту; аминотиокислоту; α,α-дизамещенную аминокислоту; β-аминокислоту; циклическую аминокислоту, иную, чем пролин или гистидин, и ароматическую аминокислоту, иную, чем фенилаланин, тирозин или триптофан.
[00362] В некоторых вариантах осуществления неприродная аминокислота содержит селективную реакционноспособную группу или реакционноспособную группу для сайт-специфического мечения целевого белка или полипептида. В некоторых случаях химическая реакция представляет собой биортогональную реакцию (например, биосовместимые и селективные реакции). В некоторых случаях химическая реакция представляет собой Cu(I)-катализируемую или "безмедную" алкин-азидную реакцию образования триазола, лигирование по Штаудингеру, реакцию Дильса-Альдера с обращенными электронными требованиями (IEDDA), "фото-клик-реакцию" или металл-опосредованный способ, такой как метатезис олефинов и реакция Сузуки-Мияуры или реакция перекрестного сочетания Соногаширы. В некоторых вариантах осуществления неприродная аминокислота содержит фотореактивную группу, подвергающуюся перекрестной сшивке, после облучения, например, УФ. В некоторых вариантах осуществления неприродная аминокислота содержит фотолабильную аминокислоту. В некоторых случаях неприродная аминокислота является пара-замещенным, мета-замещенным или орто-замещенным производным аминокислоты.
[00363] В некоторых случаях неприродная аминокислота включает p-ацетил-L-фенилаланин, p-азидометил-L-фенилаланин (pAMF), p-йодо-L-фенилаланин, O-метил-L-тирозин, p-метоксифенилаланин, p-пропаргилоксифенилаланин, p-пропаргил-фенилаланин, L-3-(2-нафтил)аланин, 3-метил-фенилаланин, O-4-аллил-L-тирозин, 4-пропил-L-тирозин, три-O-ацетил-GlcNAcp-серин, L-ДОФА, фторированный фенилаланин, изопропил-L-фенилаланин, p-азидо-L-фенилаланин, p-ацил-L-фенилаланин, p-бензоил-L-фенилаланин, L-фосфосерин, фосфоносерин, фосфонотирозин, p-бромфенилаланин, p-амино-L-фенилаланин или изопропил-L-фенилаланин.
[00364] В некоторых случаях неприродная аминокислота является 3-аминотирозином, 3-нитротирозином, 3,4-дигидрокси-фенилаланином или 3-йодтирозином. В некоторых случаях неприродная аминокислота является фенилселеноцистеином. В некоторых случаях неприродная аминокислота является бензофенон-, кетон-, йодид-, метокси-, ацетил-, бензоил- или азид-содержащим производным фенилаланина. В некоторых случаях неприродная аминокислота является бензофенон-, кетон-, йодид-, метокси-, ацетил-, бензоил- или азид-содержащим производным лизина. В некоторых случаях неприродная аминокислота содержит ароматическую боковую цепь. В некоторых случаях неприродная аминокислота не содержит ароматическую боковую цепь. В некоторых случаях неприродная аминокислота содержит азидо-группу. В некоторых случаях неприродная аминокислота содержит группу акцептора Михаэля. В некоторых случаях группы акцепторов Михаэля содержат ненасыщенный остаток, способный образовывать ковалентную связь посредством реакции 1,2-присоединения. В некоторых случаях группы акцепторов Михаэля содержат электрондефицитные алкены или алкины. В некоторых случаях группы акцепторов Михаэля включают, в качестве неограничивающих примеров, альфа, бета-ненасыщенные: кетоны, альдегиды, сульфоксиды, сульфоны, нитрилы, имины или ароматические соединения. В некоторых случаях неприродная аминокислота является дегидроаланином. В некоторых случаях неприродная аминокислота содержит альдегидную или кетоновую группу. В некоторых случаях неприродная аминокислота является производным лизина, содержащим альдегидную или кетоновую группу. В некоторых случаях неприродная аминокислота является производным лизина, содержащим один или более атомов O, N, Se или S в бета-, гамма- или дельта-положении. В некоторых случаях неприродная аминокислота является производным лизина, содержащим атомы O, N, Se или S в гамма-положении. В некоторых случаях неприродная аминокислота является производным лизина, где эпсилон-атом N заменен атомом кислорода. В некоторых случаях неприродная аминокислота является производным лизина, не являющимся природным посттрансляционно модифицированным лизином.
[00365] В некоторых случаях неприродная аминокислота является аминокислотой, содержащей боковую цепь, где шестой атом от альфа-положения содержит карбонильную группу. В некоторых случаях неприродная аминокислота является аминокислотой, содержащей боковую цепь, где шестой атом от альфа-положения содержит карбонильную группу, и пятый атом от альфа-положения является азотом. В некоторых случаях неприродная аминокислота является аминокислотой, содержащей боковую цепь, где седьмой атом от альфа-положения является атомом кислорода.
[00366] В некоторых случаях неприродная аминокислота является производным серина, содержащим селен. В некоторых случаях неприродная аминокислота является селеносерином (2-амино-3-гидроселенопропионовой кислотой). В некоторых случаях неприродная аминокислота является 2-амино-3-((2-((3-(бензилокси)-3-оксопропил)амино)этил)селанил)пропионовой кислотой. В некоторых случаях неприродная аминокислота является 2-амино-3-(фенилселанил)пропионовой кислотой. В некоторых случаях неприродная аминокислота содержит селен, где окисление селена приводит к образованию неприродной аминокислоты, содержащей алкен.
[00367] В некоторых случаях неприродная аминокислота содержит циклооктинильную группу. В некоторых случаях неприродная аминокислота содержит трансциклоктенильную группу. В некоторых случаях неприродная аминокислота содержит норборненильную группу. В некоторых случаях неприродная аминокислота содержит циклопропенильную группу. В некоторых случаях неприродная аминокислота содержит диазириновую группу. В некоторых случаях неприродная аминокислота содержит тетразиновую группу.
[00368] В некоторых случаях неприродная аминокислота является производным лизина, где азот боковой цепи является карбамилированным. В некоторых случаях неприродная аминокислота является производным лизина, где азот боковой цепи является ацилированным. В некоторых случаях неприродная аминокислота является 2-амино-6-{[(трет-бутокси)карбонил]амино}гексановой кислотой. В некоторых случаях неприродная аминокислота является 2-амино-6-{[(трет-бутокси)карбонил]амино}гексановой кислотой. В некоторых случаях неприродная аминокислота является N6-Boc-N6-метиллизином. В некоторых случаях неприродная аминокислота является N6-ацетиллизином. В некоторых случаях неприродная аминокислота является пирролизином. В некоторых случаях неприродная аминокислота является N6-трифторацетиллизином. В некоторых случаях неприродная аминокислота является 2-амино-6-{[(бензилокси)карбонил]амино}гексановой кислотой. В некоторых случаях неприродная аминокислота является 2-амино-6-{[(p-йодбензилокси)карбонил]амино}гексановой кислотой. В некоторых случаях неприродная аминокислота является 2-амино-6-{[(p-нитробензилокси)карбонил]амино}гексановой кислотой. В некоторых случаях неприродная аминокислота является N6-пролиллизином. В некоторых случаях неприродная аминокислота является 2-амино-6-{[(циклопентилокси)карбонил]амино}гексановой кислотой. В некоторых случаях неприродная аминокислота является N6-(циклопентанкарбонил)лизином. В некоторых случаях неприродная аминокислота является N6-(тетрагидрофуран-2-карбонил)лизином. В некоторых случаях неприродная аминокислота является N6-(3-этинилтетрагидрофуран-2-карбонил)лизином. В некоторых случаях неприродная аминокислота является N6-((проп-2-ин-1-илокси)карбонил)лизином. В некоторых случаях неприродная аминокислота является 2-амино-6-{[(2-азидоциклопентилокси)карбонил]амино}гексановой кислотой. В некоторых случаях неприродная аминокислота является N6-((2-азидоэтокси)карбонил)лизином. В некоторых случаях неприродная аминокислота является 2-амино-6-{[(2-нитробензилокси)карбонил]амино}гексановой кислотой. В некоторых случаях неприродная аминокислота является 2-амино-6-{[(2-циклооктинилокси)карбонил]амино}гексановой кислотой. В некоторых случаях неприродная аминокислота является N6-(2-аминобут-3-иноил)лизином. В некоторых случаях неприродная аминокислота является 2-амино-6-((2-аминобут-3-иноил)окси)гексановой кислотой. В некоторых случаях неприродная аминокислота является N6-(аллилоксикарбонил)лизином. В некоторых случаях неприродная аминокислота является N6-(бутенил-4-оксикарбонил)лизином. В некоторых случаях неприродная аминокислота является N6-(пентенил-5-оксикарбонил)лизином. В некоторых случаях неприродная аминокислота является N6-((бут-3-ин-1-илокси)карбонил)-лизином. В некоторых случаях неприродная аминокислота является N6-((пент-4-ин-1-илокси)карбонил)-лизином. В некоторых случаях неприродная аминокислота является N6-(тиазолидин-4-карбонил)лизином. В некоторых случаях неприродная аминокислота является 2-амино-8-оксононановой кислотой. В некоторых случаях неприродная аминокислота является 2-амино-8-оксооктановой кислотой. В некоторых случаях неприродная аминокислота является N6-(2-оксоацетил)лизином.
[00369] В некоторых случаях неприродная аминокислота является N6-пропиониллизином. В некоторых случаях неприродная аминокислота является N6-бутириллизином. В некоторых случаях неприродная аминокислота является N6-(бут-2-еноил)лизином. В некоторых случаях неприродная аминокислота является N6-((бицикло[2.2.1]гепт-5-ен-2-илокси)карбонил)лизином. В некоторых случаях неприродная аминокислота является N6-((спиро[2.3]гекс-1-ен-5-илметокси)карбонил)лизином. В некоторых случаях неприродная аминокислота является N6-(((4-(1-(трифторметил)циклопроп-2-ен-1-ил)бензил)окси)карбонил)лизином. В некоторых случаях неприродная аминокислота является N6-((бицикло[2,2.1]hept-5-ен-2-илметокси)карбонил)лизином. В некоторых случаях неприродная аминокислота является цистеиниллизином. В некоторых случаях неприродная аминокислота является N6-((1-(6-нитробензо[d][1.3]диоксол-5-ил)этокси)карбонил)лизином. В некоторых случаях неприродная аминокислота является N6-((2-(3-метил-3H-диазирин-3-ил)этокси)карбонил)лизином. В некоторых случаях неприродная аминокислота является N6-((3-(3-метил-3H-диазирин-3-ил)пропокси)карбонил)лизином. В некоторых случаях неприродная аминокислота является N6-((мета-нитробензилокси)N6-метилкарбонил)лизином. В некоторых случаях неприродная аминокислота является N6-((бицикло[6.1.0]нон-4-ин-9-илметокси)карбонил)-лизином. В некоторых случаях неприродная аминокислота является N6-((циклогепт-3-ен-1-илокси)карбонил)-L-лизином.
[00370] В некоторых случаях неприродная аминокислота является 2-амино-3-(((((бензилокси)карбонил)амино)метил)селанил)пропионовой кислотой. В некоторых вариантах осуществления неприродную аминокислоту встраивают в белок с помощью видоизмененного янтарного, опалового или охряного стоп-кодона. В некоторых вариантах осуществления неприродную аминокислоту встраивают в белок с помощью кодона из 4 оснований. В некоторых вариантах осуществления неприродную аминокислоту встраивают в белок с помощью видоизмененного редкого смыслового кодона.
[00371] В некоторых вариантах осуществления неприродную аминокислоту встраивают в белок с помощью неприродного кодона, содержащего неприродный нуклеотид.
[00372] В некоторых вариантах осуществления белок содержит по меньшей мере две неприродных аминокислоты. В некоторых вариантах осуществления белок содержит по меньшей мере три неприродных аминокислоты. В некоторых вариантах осуществления белок содержит по меньшей мере две разные неприродные аминокислоты. В некоторых вариантах осуществления белок содержит по меньшей мере три разные неприродные аминокислоты. По меньшей мере одна неприродная аминокислота: является аналогом лизина; содержит ароматическую боковую цепь; содержит азидо-группу; содержит алкиновую группу; или содержит альдегидную или кетоновую группу. В некоторых вариантах осуществления по меньшей мере одна неприродная аминокислота не содержит ароматическую боковую цепь. В некоторых вариантах осуществления по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK) или N6-пропаргилэтокси-карбонил-L-лизин (PraK). В некоторых вариантах осуществления по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK). В некоторых вариантах осуществления по меньшей мере одна неприродная аминокислота содержит N6-пропаргилэтокси-карбонил-L-лизин (PraK).
[00373] В некоторых случаях встраивание неприродной аминокислоты в белок опосредовано ортогональной, модифицированной парой синтетаза/тРНК. Такие ортогональные пары содержат природную или мутантную синтетазу, способную заряжать неприродную тРНК специфической неприродной аминокислотой, зачастую минимизируя зарядку a) другими эндогенными аминокислотами или альтернативными неприродными аминокислотами неприродной тРНК и b) любых других (включая эндогенные) тРНК. Такие ортогональные пары содержат тРНК, способные заряжаться синтетазой, избегая зарядки другими эндогенными аминокислотами под действием эндогенных синтетаз. В некоторых вариантах осуществления такие пары идентифицированы в различных организмах, таких как бактерии, дрожжи, археи или человек. В некоторых вариантах осуществления ортогональная пара синтетаза/тРНК содержит компоненты из одного организма. В некоторых вариантах осуществления ортогональная пара синтетаза/тРНК содержит компоненты из двух разных организмов. В некоторых вариантах осуществления ортогональная пара синтетаза/тРНК содержит компоненты, которые перед модификацией способствуют трансляции разных аминокислот. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной аланин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной аргинин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной аспарагин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной синтетазой аспарагиновой кислоты. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной цистеин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной глутамин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной синтетазой глутаминовой кислоты. В некоторых вариантах осуществления ортогональная синтетаза является модифицированным аланин-глицином. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной гистидин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной лейцин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной изолейцин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной лизин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной метионин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной фенилаланин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной пролин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной серин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной треонин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной триптофан-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной тирозин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной валин-синтетазой. В некоторых вариантах осуществления ортогональная синтетаза является модифицированной фосфосерин-синтетазой. В некоторых вариантах осуществления ортогональная тРНК является модифицированной аланин тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной аргинин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной аспарагин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной тРНК аспарагиновой кислоты. В некоторых вариантах осуществления ортогональная тРНК является модифицированной цистеин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной глутамин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной тРНК глутаминовой кислоты. В некоторых вариантах осуществления ортогональная тРНК является модифицированным аланин-глицином. В некоторых вариантах осуществления ортогональная тРНК является модифицированной гистидин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной лейцин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной изолейцин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной лизин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной метионин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной фенилаланин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной пролин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной серин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной треонин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной триптофан-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной тирозин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной валин-тРНК. В некоторых вариантах осуществления ортогональная тРНК является модифицированной фосфосерин-тРНК. В любом из этих вариантов осуществления тРНК может являться гетерологичной тРНК.
[00374] В некоторых вариантах осуществления неприродную аминокислоту встраивают в белок с помощью пары аминоацил (aaRS или RS)-тРНК синтетаза-тРНК. Неограничивающие примеры пар aaRS-тРНК включают пары aaRS/тРНК Methanococcus jannaschii (Mj-Tyr), пары TyrRS E. coli (Ec-Tyr)/тРНКCUA B. stearothermophilus, пары LeuRS E. coli (Ec-Leu)/тРНКCUA B. stearothermophilus и пары пирролизил-тРНК. В некоторых случаях неприродную аминокислоту встраивают в белок с помощью пары Mj-TyrRS/тРНК. Неограничивающие примеры неприродных аминокислот (UAA), которые можно встраивать с помощью пары Mj-TyrRS/тРНК, включают пара-замещенные производные фенилаланина, такие как p-аминофенилаланин и p-метилфенилаланин; мета-замещенные производные тирозина, такие как 3-аминотирозин, 3-нитротирозин, 3,4-дигидроксифенилаланин и 3-йодтирозин; фенилселеноцистеин; p-боронофенилаланин; и o-нитробензилтирозин.
[00375] В некоторых случаях неприродную аминокислоту встраивают в белок с помощью пары Ec-Tyr/тРНКCUA или Ec-Leu/тРНКCUA. Неограничивающие примеры UAA, которые можно встраивать с помощью пары Ec-Tyr/тРНКCUA или Ec-Leu/тРНКCUA, включают производные фенилаланина, содержащие бензофеноновые, кетоновые, йодидные или азидные заместители; O-пропаргилтирозин; α-аминокапроновую кислоту, O-метилтирозин, O-нитробензилцистеин и 3-(нафталин-2-иламино)-2-амино-пропионовую кислоту.
[00376] В некоторых случаях неприродную аминокислоту встраивают в белок с помощью пары пирролизил-тРНК. В некоторых случаях PylRS получают из видов архебактерий, например, метаногенных архебактерий. В некоторых случаях PylRS получают из Methanosarcina barkeri, Methanosarcina mazei или Methanosarcina acetivorans. Неограничивающие примеры UAA, которые можно встраивать с помощью пары пирролизил-тРНК включают амид- и карбамат-замещенные лизины, такие как 2-амино-6-((R)-тетрагидрофуран-2-карбоксамид)гексановая кислота, N-ε-D-пролил-L-лизин и N-ε-циклопентилоксикарбонил-L-лизин; N-ε-акрилоил-L-лизин; N-ε-[(1-(6-нитробензо[d][1.3]диоксол-5-ил)этокси)карбонил]-L-лизин и N-ε-(1-метилциклопро-2-енкарбоксамидо)лизин.
[00377] В некоторых случаях неприродную аминокислоту встраивают в белок, представленный в настоящем описании, с помощью синтетазы, описанной в патентах США №№ 9988619 и 9938516, описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме. Примеры UAA, которые можно встраивать с помощью таких синтетаз, включают пара-метилазидо-L-фенилаланин, аралкил-, гетероциклил-, гетероаралкил- неприродные аминокислоты и др. В некоторых вариантах осуществления такие UAA содержат пиридил, пиразинил, пиразолил, триазолил, оксазолил, тиазолил, тиофенил или другой гетероцикл. В некоторых вариантах осуществления такие аминокислоты содержат азиды, тетразины или другую химическую группу, способные к конъюгации с партнером по сочетанию, таким как водорастворимый остаток. В некоторых вариантах осуществления такие синтетазы экспрессируют и используют для встраивания UAA в белки in vivo. В некоторых вариантах осуществления такие синтетазы используют для встраивания UAA в белки с использованием бесклеточной системы трансляции.
[00378] В некоторых случаях неприродную аминокислоту встраивают в белок, представленный в настоящем описании, с помощью природной синтетазы. В некоторых вариантах осуществления неприродную аминокислоту встраивают в белок с помощью организма, являющегося ауксотрофным по одной или более аминокислотам. В некоторых вариантах осуществления синтетазы, соответствующие ауксотрофной аминокислоте, могут заряжать соответствующую тРНК неприродной аминокислотой. В некоторых вариантах осуществления неприродная аминокислота является селеноцистеином или его производным. В некоторых вариантах осуществления неприродная аминокислота является селенометионином или его производным. В некоторых вариантах осуществления неприродная аминокислота является ароматической аминокислотой, где ароматическая аминокислота содержит арилгалид, такой как йодид. В вариантах осуществления неприродная аминокислота является структурно схожей с ауксотрофной аминокислотой.
[00379] В некоторых случаях неприродная аминокислота включает неприродную аминокислоту, показанную на фиг. 8A.
[00380] В некоторых случаях неприродная аминокислота содержит производное или аналог лизина или фенилаланина. В некоторых случаях неприродная аминокислота содержит производное лизина или аналог лизина. В некоторых случаях неприродная аминокислота содержит пирролизин (Pyl). В некоторых случаях неприродная аминокислота содержит производное фенилаланина или фенилаланин аналог. В некоторых случаях неприродная аминокислота является неприродной аминокислотой, описанной в Wan, et al., "Pyrrolysyl-tRNA synthetase: an ordinary enzyme but an outstanding genetic code expansion tool," Biocheim Biophys Aceta 1844(6): 1059-4070 (2014). В некоторых случаях неприродная аминокислота включает неприродную аминокислоту, показанную на фиг. 8B и фиг. 8C.
[00381] В некоторых вариантах осуществления неприродная аминокислота включает неприродную аминокислоту, показанную на фиг. 8D-фиг. 8G (адаптированную из таблицы 1 из Dumas et al., Chemical Science 2015, 6, 50-69).
[00382] В некоторых вариантах осуществления неприродная аминокислота, встроенная в белок, представленный в настоящем описании, описана в US 9840493; US 9682934; US 2017/0260137; US 9938516 или US 2018/0086734; описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме. Примеры UAA, которые можно встраивать с помощью таких синтетаз, включают пара-метилазидо-L-фенилаланин, аралкильное, гетероциклильное и гетероаралкильное и лизиновое производное неприродных аминокислот. В некоторых вариантах осуществления такие UAA содержат пиридил, пиразинил, пиразолил, триазолил, оксазолил, тиазолил, тиофенил или другой гетероцикл. В некоторых вариантах осуществления такие аминокислоты содержат азиды, тетразины или другую химическую группу, способную к конъюгации с партнером по сочетанию, таким как водорастворимый остаток. В некоторых вариантах осуществления UAA содержит азид, присоединенный к ароматическому остатку с помощью алкильного линкера. В некоторых вариантах осуществления алкильный линкер является C1-C10-линкером. В некоторых вариантах осуществления UAA содержит тетразин, присоединенный к ароматическому остатку с помощью алкильного линкера. В некоторых вариантах осуществления UAA содержит тетразин, присоединенный к ароматическому остатку с помощью аминогруппы. В некоторых вариантах осуществления UAA содержит тетразин, присоединенный к ароматическому остатку с помощью алкиламиногруппы. В некоторых вариантах осуществления UAA содержит азид, присоединенный к концевому азоту (например, N6 производного лизина или N5, N4 или N3 производного, содержащего более короткую алкильную боковую цепь) боковой цепи аминокислоты с помощью алкильной цепи. В некоторых вариантах осуществления UAA содержит тетразин, присоединенный к концевому азоту боковой цепи аминокислоты с помощью алкильной цепи. В некоторых вариантах осуществления UAA содержит азид или тетразин, присоединенный к амиду с помощью алкильного линкера. В некоторых вариантах осуществления UAA является азид- или тетразин-содержащим карбаматом или амидом 3-аминоаланина, серина, лизина или их производного. В некоторых вариантах осуществления такие UAA встраивают в белки in vivo. В некоторых вариантах осуществления такие UAA встраивают в белки в бесклеточной системе.
Типы клеток
[00383] В некоторых вариантах осуществления используют множество типов клеток/микроорганизмов, например, для трансформации или генетической инженерии. В некоторых вариантах осуществления клетка является прокариотической или эукариотической клеткой. В некоторых вариантах осуществления прокариотическая клетка является бактериальной клеткой. В некоторых вариантах осуществления эукариотическая клетка является клеткой гриба или одноклеточным простейшим. В некоторых вариантах осуществления клетка гриба является дрожжевой клеткой. В других случаях эукариотическая клетка является культивируемой клеткой животного, растения или человека. В дополнительных случаях эукариотическая клетка находится в организме, таком как растение, многоклеточный гриб или животное.
[00384] В некоторых вариантах осуществления сконструированный микроорганизм является одноклеточным организмом, зачастую способным делиться и пролиферировать. В рамках изобретения "сконструированный микроорганизм" является микроорганизмом, генетический материал которого изменен способами генетической инженерии (т.е. технологии рекомбинантных ДНК). Микроорганизм может включать один или более из следующих признаков: аэроб, анаэроб, нитчатый, ненитчатый, моноплоидный, диплоидный, ауксотрофный и/или неауксотрофный. В некоторых вариантах осуществления сконструированный микроорганизм является прокариотическим микроорганизмом (например, бактерией), и в некоторых вариантах осуществления сконструированный микроорганизм является непрокариотическим микроорганизмом, таким как эукариотический микроорганизм. В некоторых вариантах осуществления сконструированный микроорганизм является эукариотическим микроорганизмом (например, дрожжами, другим грибом, амебой). В некоторых вариантах осуществления сконструированный микроорганизм является грибом. В некоторых вариантах осуществления сконструированный организм является дрожжами.
[00385] В качестве микроорганизма-хозяина, сконструированного микроорганизма, генетически модифицированного организма или источника гетерологичного или модифицированного полинуклеотида можно выбирать любые подходящие дрожжи. Дрожжи включают, в качестве неограничивающих примеров, дрожжи Yarrowia (например, Y. lipolytica (ранее классифицируемые как Candida lipolytica)), дрожжи Candida (например, C. revkaufi, C. viswanathii, C. pulcherrima, C. tropicalis, C. utilis), дрожжи Rhodotorula (например, R. glutinus, R. graminis), дрожжи Rhodosporidium (например, R. toruloides), дрожжи Saccharomyces (например, S. cerevisiae, S. bayanus, S. pastorianus, S. carlsbergensis), дрожжи Cryptococcus, дрожжи Trichosporon (например, T. pullans, T. cutaneum), дрожжи Pichia (например, P. pastoris, K. phaffii) и дрожжи Lipomyces (например, L. starkeyii, L. lipoferus). В некоторых вариантах осуществления подходящие дрожжи принадлежат роду Arachniotus, Aspergillus, Aureobasidium, Auxarthron, Blastomyces, Candida, Chrysosporuim, Chrysosporium, Debaryomyces, Coccidiodes, Cryptococcus, Gymnoascus, Hansenula, Histoplasma, Issatchenkia, Kluyveromyces, Lipomyces, Issatchenkia, Microsporum, Myxotrichum, Myxozyma, Oidiodendron, Pachysolen, Penicillium, Pichia, Rhodosporidium, Rhodotorula, Rhodotorula, Saccharomyces, Schizosaccharomyces, Scopulariopsis, Sepedonium, Trichosporon или Yarrowia. В некоторых вариантах осуществления подходящие дрожжи принадлежат роду Arachniotus flavoluteus, Aspergillus flavus, Aspergillus fumigatus, Aspergillus niger, Aureobasidium pullulans, Auxarthron thaxteri, Blastomyces dermatitidis, Candida albicans, Candida dubliniensis, Candida famata, Candida glabrata, Candida guilliermondii, Candida kefyr, Candida krusei, Candida lambica, Candida lipolytica, Candida lustitaniae, Candida parapsilosis, Candida pulcherrima, Candida revkaufi, Candida rugosa, Candida tropicalis, Candida utilis, Candida viswanathii, Candida xestobii, Chrysosporuim keratinophilum, Coccidiodes immitis, Cryptococcus albidus var. diffluens, Cryptococcus laurentii, Cryptococcus neofomans, Debaryomyces hansenii, Gymnoascus dugwayensis, Hansenula anomala, Histoplasma capsulatum, Issatchenkia occidentalis, Issatchenkia orientalis, Kluyveromyces lactis, Kluyveromyces marxianus, Kluyveromyces thermotolerans, Kluyveromyces waltii, Lipomyces lipoferus, Lipomyces starkeyii, Microsporum gypseum, Myxotrichum deflexum, Oidiodendron echinulatum, Pachysolen tannophilis, Penicillium notatum, Pichia anomala, Pichia pastoris, Pichia stipitis, Rhodosporidium toruloides, Rhodotorula glutinus, Rhodotorula graminis, Saccharomyces cerevisiae, Saccharomyces kluyveri, Schizosaccharomyces pombe, Scopulariopsis acremonium, Sepedonium chrysospermum, Trichosporon cutaneum, Trichosporon pullans, Yarrowia lipolytica или Yarrowia lipolytica (ранее классифицируемые как Candida lipolytica). В некоторых вариантах осуществления дрожжи представляют собой штамм Y. lipolytica, включая, в качестве неограничивающих примеров, штаммы ATCC20362, ATCC8862, ATCC18944, ATCC20228, ATCC76982 и LGAM S(7)1 (Papanikolaou S., and Aggelis G., Bioresour. Technol. 82(1):43-9 (2002)). В некоторых вариантах осуществления дрожжи представляют собой вид дрожжей Candida (т.е. Candida spp.). Любой подходящий вид Candida можно использовать и/или генетически модифицировать для получения жирной дикарбоновой кислоты (например, октандиовой кислоты, декандиовой кислоты, додекандиовой кислоты, тетрадекандиовой кислоты, гексадекандиовой кислоты, октадекандиовой кислоты, эйкозандиовой кислоты). В некоторых вариантах осуществления подходящий вид Candida включают, в качестве неограничивающих примеров, Candida albicans, Candida dubliniensis, Candida famata, Candida glabrata, Candida guilliermondii, Candida kefyr, Candida krusei, Candida lambica, Candida lipolytica, Candida lustitaniae, Candida parapsilosis, Candida pulcherrima, Candida revkaufi, Candida rugosa, Candida tropicalis, Candida utilis, Candida viswanathii, Candida xestobii и любыми дрожжами Candida spp., представленными в настоящем описании. Неограничивающие примеры штаммов Candida spp. включают штаммы sAA001 (ATCC20336), sAA002 (ATCC20913), sAA003 (ATCC20962), sAA496 (US2012/0077252), sAA106 (US2012/0077252), SU-2 (ura3-/ura3-), H5343 (с заблокированным бета-окислением; патент США № 5648247). В качестве родительских штаммов для генетической модификации можно использовать любые подходящие штаммы дрожжей Candida spp.
[00386] Рода, виды и штаммы дрожжей зачастую настолько близкородственны по генетическому содержимому, что их может быть трудно различать, классифицировать и/или называть. В некоторых случаях штаммы C. lipolytica и Y. lipolytica может быть трудно различать, классифицировать и/или называть, и в некоторых случаях их можно считать одним и тем же организмом. В некоторых случаях различные штаммы C. tropicalis и C. viswanathii может быть трудно различать, классифицировать и/или называть (см., например, Arie et.al., J. Gen. Appl. Microbiol., 46, 257-262 (2000)). Некоторые штаммы C. tropicalis и C. viswanathii, полученные из ATCC, а также других коммерческих или научных источников можно считать эквивалентными и в равной степени подходящими для вариантов осуществления, представленных в настоящем описании. В некоторых вариантах осуществления некоторые родительские штаммы C. tropicalis и C. viswanathii считают отличающимися только названиями.
[00387] В качестве микроорганизма-хозяина, сконструированного микроорганизма или источника гетерологичного полинуклеотида можно выбирать любой подходящий гриб. Неограничивающие примеры грибов включают грибы Aspergillus (например, A. parasiticus, A. nidulans), грибы Thraustochytrium, грибы Schizochytrium и грибы Rhizopus (например, R. arrhizus, R. oryzae, R. nigricans). В некоторых вариантах осуществления гриб представляет собой штамм A. parasiticus, включающий, в качестве неограничивающих примеров, штамм ATCC24690, и в некоторых вариантах осуществления гриб представляет собой штамм A. nidulans, включающий, в качестве неограничивающих примеров, штамм ATCC38163.
[00388] В качестве микроорганизма-хозяина, сконструированного микроорганизма или источника гетерологичного полинуклеотида можно выбирать любых подходящих прокариот. Можно выбирать грамотрицательные или грамположительные бактерии. Неограничивающие примеры бактерий включают бактерии Bacillus (например, B. subtilis, B. megaterium), бактерии Acinetobacter, бактерии Norcardia, бактерии Xanthobacter, бактерии Escherichia (например, E. coli (например, штаммы DH10B, Stbl2, DH5-альфа, DB3, DB3,1), DB4, DB5, JDP682 и ccdA-over (например, заявка США № 09/518188))), бактерии Streptomyces, бактерии Erwinia, бактерии Klebsiella, бактерии Serratia (например, S. marcessans), бактерии Pseudomonas (например, P. aeruginosa), бактерии Salmonella (например, S. typhimurium, S. typhi), бактерии Megasphaera (например, Megasphaera elsdenii). Бактерии также включают, в качестве неограничивающих примеров, фотосинтезирующие бактерии (например, зеленые несерные бактерии (например, бактерии Choroflexus (например, C. aurantiacus), бактерии Chloronema (например, C. gigateum)), зеленые серные бактерии (например, бактерии Chlorobium (например, C. limicola), бактерии Pelodictyon (например, P. luteolum), пурпурные серные бактерии (например, бактерии Chromatium (например, C. okenii)), и пурпурные несерные бактерии (например, бактерии Rhodospirillum (например, R. rubrum), бактерии Rhodobacter (например, R. sphaeroides, R. capsulatus) и бактерии Rhodomicrobium (например, R. vanellii)).
[00389] В качестве микроорганизма-хозяина, сконструированного микроорганизма или источника гетерологичного полинуклеотида можно использовать клетки из немикробных организмов. Неограничивающие примеры таких клеток включают клетки насекомых (например, Drosophila (например, D. melanogaster), Spodoptera (например, клетки Sf9 или Sf21 S. frugiperda) и Trichoplusa (например, клетки High-Five); клетки нематод (например, клетки C. elegans); клетки птиц; клетки амфибий (например, клетки Xenopus laevis); клетки рептилий; клетки млекопитающих (например, клетки NIH3T3, 293, CHO, COS, VERO, C127, BHK, Per-C6, клетки меланомы Боуэса и HeLa); и растительные клетки (например, Arabidopsis thaliana, Nicotania tabacum, Cuphea acinifolia, Cuphea aequipetala, Cuphea angustifolia, Cuphea appendiculata, Cuphea avigera, Cuphea avigera var. pulcherrima, Cuphea axilliflora, Cuphea bahiensis, Cuphea baillonis, Cuphea brachypoda, Cuphea bustamanta, Cuphea calcarata, Cuphea calophylla, Cuphea calophylla subsp. mesostemon, Cuphea carthagenensis, Cuphea circaeoides, Cuphea confertiflora, Cuphea cordata, Cuphea crassiflora, Cuphea cyanea, Cuphea decandra, Cuphea denticulata, Cuphea disperma, Cuphea epilobiifolia, Cuphea ericoides, Cuphea flava, Cuphea flavisetula, Cuphea fuchsiifolia, Cuphea gaumeri, Cuphea glutinosa, Cuphea heterophylla, Cuphea hookeriana, Cuphea hyssopifolia (мексиканский вереск), Cuphea hyssopoides, Cuphea ignea, Cuphea ingrata, Cuphea jorullensis, Cuphea lanceolata, Cuphea linarioides, Cuphea llavea, Cuphea lophostoma, Cuphea lutea, Cuphea lutescens, Cuphea melanium, Cuphea melvilla, Cuphea micrantha, Cuphea micropetala, Cuphea mimuloides, Cuphea nitidula, Cuphea palustris, Cuphea parsonsia, Cuphea pascuorum, Cuphea paucipetala, Cuphea procumbens, Cuphea pseudosilene, Cuphea pseudovaccinium, Cuphea pulchra, Cuphea racemosa, Cuphea repens, Cuphea salicifolia, Cuphea salvadorensis, Cuphea schumannii, Cuphea sessiliflora, Cuphea sessilifolia, Cuphea setosa, Cuphea spectabilis, Cuphea spermacoce, Cuphea splendida, Cuphea splendida var. viridiflava, Cuphea strigulosa, Cuphea subuligera, Cuphea teleandra, Cuphea thymoides, Cuphea tolucana, Cuphea urens, Cuphea utriculosa, Cuphea viscosissima, Cuphea watsoniana, Cuphea wrightii, Cuphea lanceolata).
[00390] Микроорганизмы или клетки, используемые в качестве организмов-хозяев или источника гетерологичного полинуклеотида, являются коммерчески доступными. Микроорганизмы и клетки, представленные в настоящем описании, и другие подходящие микроорганизмы и клетки доступны, например, в Invitrogen Corporation, (Carlsbad, CA), American Type Culture Collection (Manassas, Virginia) и Agricultural Research Culture Collection (NRRL; Peoria, Illinois). Микроорганизмы-хозяева и сконструированные микроорганизмы можно получать в любой подходящей форме. Например, такие микроорганизмы можно получать в жидкой культуре или культуре на твердом субстрате (например, среде на основе агара), которая может являться первичной культурой или подвергнута пассажам (например, разведена и культивирована) один или более раз. Микроорганизмы также можно получать в замороженной форме или сухой форме (например, лиофилизированной). Микроорганизмы можно получать в любой подходящей концентрации.
Полимеразы
[00391] Особенно полезной функцией полимераз является катализ полимеризации цепи нуклеиновой кислоты с использованием существующей нуклеиновой кислоты в качестве матрицы. Другие полезные функции описаны в других местах в настоящем описании. Примеры полимераз, которые можно использовать, включают ДНК-полимеразы и РНК-полимеразы.
[00392] Способность улучшать специфичность, процессивность или другие признаки полимераз неприродных нуклеиновых кислот будут крайне желательными в различных контекстах, например, если желательным является встраивание неприродной нуклеиновой кислоты, включая амплификацию, секвенирование, мечение, детекцию, клонирование и др.
[00393] В некоторых случаях настоящее изобретение включает полимеразы, встраивающие неприродные нуклеиновые кислоты в растущую копию матрицы, например, во время амплификации ДНК. В некоторых вариантах осуществления полимеразы можно модифицировать таким образом, что активный центр полимеразы модифицируют для снижения стерического ингибирования поступления неприродной нуклеиновой кислоты в активный центр. В некоторых вариантах осуществления полимеразы можно модифицировать для обеспечения комплементарности с одним или более неприродными признаками неприродных нуклеиновых кислот. Такие полимеразы можно экспрессировать или конструировать в клетках для стабильного встраивания UBP в клетки. Таким образом, настоящее изобретение включает композиции, включающие гетерологичную или рекомбинантную полимеразу, и способы их применения.
[00394] Полимеразы можно модифицировать способами, относящимися к белковой инженерии. Например, можно осуществлять молекулярное моделирование с учетом кристаллических структур для идентификации локализации полимераз, при этом можно осуществлять мутации для модификации целевой активности. Остаток, идентифицированный в качестве мишени для замены, можно заменять остатком, выбранным с использованием моделирования минимизации энергии, моделирования гомологии и/или консервативных аминокислотных замен, как описано в Bordo, et al. J Mol Biol 217: 721-729 (1991) и Hayes, et al. Proc Natl Acad Sci, USA 99: 15926-15931 (2002), описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00395] В способе или композиции, приведенных в настоящем описании, можно использовать любую из множества полимераз, включая, например, белковые ферменты, выделенные из биологических систем и их функциональных вариантов. Ссылку на конкретную полимеразу, такую как примеры ниже, будут понимать как включающую ее функциональные варианты, если не указано иначе. В некоторых вариантах осуществления полимераза является полимеразой дикого типа. В некоторых вариантах осуществления полимераза является модифицированной или мутантной полимеразой. В некоторых вариантах осуществления полимераза может являться гетерологичной полимеразой.
[00396] Также можно использовать полимеразы с признаками улучшения поступления неприродных нуклеиновых кислот в области активного центра и координирования с неприродными нуклеотидами в области активного центра. В некоторых вариантах осуществления модифицированная полимераза имеет модифицированный участок связывания нуклеотида.
[00397] В некоторых вариантах осуществления модифицированная полимераза имеет специфичность к неприродной нуклеиновой кислоте, составляющую по меньшей мере приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,99% специфичности полимеразы дикого типа в отношении неприродной нуклеиновой кислоты. В некоторых вариантах осуществления модифицированная полимераза или полимераза дикого типа имеет специфичность к неприродной нуклеиновой кислоте, содержащей модифицированный сахар, составляющую по меньшей мере приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,99% специфичности полимеразы дикого типа в отношении природной нуклеиновой кислоты и/или неприродной нуклеиновой кислоты без модифицированного сахара. В некоторых вариантах осуществления модифицированная полимераза или полимераза дикого типа имеет специфичность к неприродной нуклеиновой кислоте, содержащей модифицированное основание, составляющую по меньшей мере приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,99% специфичности полимеразы дикого типа в отношении природной нуклеиновой кислоты и/или неприродной нуклеиновой кислоты без модифицированного основания. В некоторых вариантах осуществления модифицированная полимераза или полимераза дикого типа имеет специфичность к неприродной нуклеиновой кислоте, содержащей трифосфат, составляющую по меньшей мере приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,99% специфичности полимеразы дикого типа в отношении нуклеиновой кислоты, содержащей трифосфат, и/или неприродной нуклеиновой кислоты без трифосфата. Например, модифицированная полимераза или полимераза дикого типа может иметь специфичность к неприродной нуклеиновой кислоте, содержащей трифосфат, составляющую по меньшей мере приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,99% специфичности полимеразы дикого типа в отношении неприродной нуклеиновой кислоты с дифосфатом или монофосфатом, или без фосфата, или их комбинации.
[00398] В некоторых вариантах осуществления модифицированная полимераза или полимераза дикого типа имеет сниженную специфичность к неприродной нуклеиновой кислоте. В некоторых вариантах осуществления модифицированная полимераза или полимераза дикого типа имеет специфичность к неприродной нуклеиновой кислоте и специфичность к природной нуклеиновой кислоте, составляющей по меньшей мере приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,99% специфичности полимеразы дикого типа в отношении природной нуклеиновой кислоты. В некоторых вариантах осуществления модифицированная полимераза или полимераза дикого типа имеет специфичность к неприродной нуклеиновой кислоте, содержащей модифицированный сахар, и специфичность к природной нуклеиновой кислоте, составляющей по меньшей мере приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,99% специфичности полимеразы дикого типа в отношении природной нуклеиновой кислоты. В некоторых вариантах осуществления модифицированная полимераза или полимераза дикого типа имеет специфичность к неприродной нуклеиновой кислоте, содержащей модифицированное основание, и специфичность к природной нуклеиновой кислоте, составляющей по меньшей мере приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,99% специфичности полимеразы дикого типа в отношении природной нуклеиновой кислоты.
[00399] Отсутствие экзонуклеазной активности может являться характеристикой дикого типа или характеристикой, придаваемой вариантом или сконструированной полимеразой. Например, экзонуклеаза без фрагмента Кленова является мутантной версией фрагмента Кленова, в котором отсутствует 3'-5'-корректирующая экзонуклеазная активность.
[00400] Способ по изобретению можно использовать для расширения субстратного диапазона любой ДНК-полимеразы, у которой отсутствует внутренняя 3-5'-экзонуклеазная корректирующая активность, или где 3-5'-экзонуклеазная корректирующая активность заблокирована, например, в результате мутации. Примеры ДНК-полимераз включают polA, polB (см., например, Parrel & Loeb, Nature Struc Biol 2001), polC, polD, polY, polX и обратные транскриптазы (RT), но, предпочтительно, являются процессивными, высокоточными полимеразами (PCT/GB2004/004643). В некоторых вариантах осуществления модифицированная полимераза или полимераза дикого типа, по существу, не имеет 3'-5'-корректирующей экзонуклеазной активности. В некоторых вариантах осуществления модифицированная полимераза или полимераза дикого типа, по существу, не имеет, 3'-5'-корректирующей экзонуклеазной активности в отношении неприродной нуклеиновой кислоты. В некоторых вариантах осуществления модифицированная полимераза или полимераза дикого типа имеет 3'-5'-корректирующую экзонуклеазную активность. В некоторых вариантах осуществления модифицированная полимераза или полимераза дикого типа имеет 3'-5'-корректирующую экзонуклеазную активность в отношении природной нуклеиновой кислоты и, по существу, не имеет 3'-5'-корректирующей экзонуклеазной активности в отношении неприродной нуклеиновой кислоты.
[00401] В некоторых вариантах осуществления модифицированная полимераза имеет 3'-5'-корректирующую экзонуклеазную активность, составляющую по меньшей мере приблизительно 60%, 70%, 80%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,99% корректирующей экзонуклеазной активности полимеразы дикого типа. В некоторых вариантах осуществления модифицированная полимераза имеет 3'-5' корректирующую экзонуклеазную активность в отношении неприродной нуклеиновой кислоты, составляющей по меньшей мере приблизительно 60%, 70%, 80%, 90%, 95%, 97%, 98%, 99%, 99,5%, 99,99% корректирующей экзонуклеазной активности полимеразы дикого типа в отношении природной нуклеиновой кислоты. В некоторых вариантах осуществления модифицированная полимераза имеет 3'-5'-корректирующую экзонуклеазную активность в отношении неприродной нуклеиновой кислоты и 3'-5'-корректирующую экзонуклеазную активность в отношении природной нуклеиновой кислоты, составляющую по меньшей мере приблизительно 60%, 70%, 80%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,99% корректирующей экзонуклеазной активности полимеразы дикого типа в отношении природной нуклеиновой кислоты. В некоторых вариантах осуществления модифицированная полимераза имеет 3'-5'-корректирующую экзонуклеазную активность в отношении природной нуклеиновой кислоты, составляющую по меньшей мере, приблизительно 60%, 70%, 80%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,99% корректирующей экзонуклеазной активности полимеразы дикого типа в отношении природной нуклеиновой кислоты.
[00402] В некоторых вариантах осуществления полимеразы отличаются по скорости диссоциации нуклеиновых кислот. В некоторых вариантах осуществления полимераза имеет относительно низкую скорость диссоциации в отношении одной или более природных и неприродных нуклеиновых кислот. В некоторых вариантах осуществления полимераза имеет относительно высокую скорость диссоциации в отношении одной или более природных и неприродных нуклеиновых кислот. Скорость диссоциации представляет собой активность полимеразы, которую можно корректировать для настраивания скоростей реакции в способах, представленных в настоящем описании.
[00403] В некоторых вариантах осуществления полимеразы отличаются своей точностью при использовании с конкретной природной и/или неприродной нуклеиновой кислотой или группой природных и/или неприродных нуклеиновых кислот. Термин "точность", как правило, относится к точности, с которой полимераза встраивает правильные нуклеиновые кислоты в растущую цепь нуклеиновой кислоты при создании копии матрицы нуклеиновой кислоты. Точность ДНК-полимеразы можно измерять как соотношение правильных и неправильных встраиваний природных и неприродных нуклеиновых кислот, если присутствуют природные и неприродные нуклеиновые кислоты, например, в равных концентрациях, для конкуренции за синтез цепи в одном и том же участке в бинарном комплексе полимераза-цепь-матричная нуклеиновая кислота. Точность ДНК-полимеразы можно вычислять как соотношение (kcat/Km) для природной и неприродной нуклеиновой кислоты и (kcat/Km) для неправильной природной и неприродной нуклеиновой кислоты; где kcat и Km представляют собой параметры Михаэлиса-Ментен для равновесной кинетики фермента (Fersht, A. R. (1985) Enzyme Structure and Mechanism, 2nd ed., p 350, W. H. Freeman & Co., New York., включенный в настоящее описание в качестве ссылки). В некоторых вариантах осуществления полимераза имеет значение точности по меньшей мере приблизительно 100, 1000, 10000, 100000 или 1×106 с корректирующей активностью или без нее.
[00404] В некоторых вариантах осуществления полимеразы из нативных источников или их варианты подвергают скринингу с использованием анализа, в котором определяют встраивание неприродной нуклеиновой кислоты, имеющей конкретную структуру. В одном из примером полимеразы можно подвергать скринингу на способность встраивать неприродную нуклеиновую кислоту или UBP; например, d5SICSTP, dCNMOTP, dTPT3TP, dNaMTP, UBP dCNMOTP-dTPT3TP, или d5SICSTP-dNaMTP. Можно использовать полимеразу, например, гетерологичную полимеразу, демонстрирующую модифицированное свойство в отношении неприродной нуклеиновой кислоты по сравнению с полимеразой дикого типа. Например, модифицированное свойство может представлять собой, например, Km, kcat, Vmax, процессивность полимеразы в присутствии неприродной нуклеиновой кислоты (или природного нуклеотида), средняя длина рида матрицы полимеразой в присутствии неприродной нуклеиновой кислоты, специфичность полимеразы в отношении неприродной нуклеиновой кислоты, скорость связывания неприродной нуклеиновой кислоты, скорость высвобождения продукта (пирофосфата, трифосфата и т.д.), скорость ветвления или любую их комбинацию. В одном из вариантов осуществления модифицированное свойство представляет собой сниженную Km для неприродной нуклеиновой кислоты и/или повышенное kcat/Km или Vmax/Km для неприродной нуклеиновой кислоты. Аналогично, полимераза, необязательно, имеет повышенную скорость связывания неприродной нуклеиновой кислоты, повышенную скорость высвобождения продукта и/или сниженную скорость ветвления по сравнению с полимеразой дикого типа.
[00405] В то же время, полимераза может встраивать природные нуклеиновые кислоты, например, A, C, G и T, в растущую копию нуклеиновой кислоты. Например, полимераза, необязательно, демонстрирует специфическую активность в отношении природной нуклеиновой кислоты, составляющую по меньшей мере до приблизительно 5% (например, 5%, 10%, 25%, 50%, 75%, 100% или более) активности соответствующей полимеразы дикого типа, и процессивность в отношении природных нуклеиновых кислот в присутствии матрицы, составляющую по меньшей мере до 5% (например, 5%, 10%, 25%, 50%, 75%, 100% или более) процессивности полимеразы дикого типа в присутствии природной нуклеиновой кислоты. Необязательно, полимераза демонстрирует kcat/Km или Vmax/Km для природного нуклеотида, составляющее по меньшей мере до приблизительно 5% (например, приблизительно 5%, 10%, 25%, 50%, 75%, 100% или более) значений для полимеразы дикого типа.
[00406] Полимеразы, используемые в настоящем описании, которые могут иметь способность встраивать неприродную нуклеиновую кислоту с конкретной структурой, также можно получать с использованием подхода направленной эволюции. Анализ синтеза нуклеиновой кислоты можно использовать для скрининга на варианты полимеразы, имеющие специфичность в отношении любой из множества неприродных нуклеиновых кислот. Например, варианты полимераз можно подвергать скринингу на способность встраивать неприродный нуклеозидтрифосфат напротив неприродного нуклеотида в матрице ДНК; например, dTPT3TP напротив dCNMO, dCNMOTP напротив dTPT3, NaMTP напротив dTPT3 или TAT1TP напротив dCNMO или dNaM. В некоторых вариантах осуществления такой анализ является анализом in vitro, например, с использованием рекомбинантного варианта полимеразы. В некоторых вариантах осуществления такой анализ является анализом in vivo, например, экспрессии варианта полимеразы в клетке. Такие способы направленной эволюции можно использовать для скрининга вариантов любой подходящей полимеразы на активность в отношении любой из неприродных нуклеиновых кислот, приведенных в настоящем описании. В некоторых случаях полимеразы, используемые в настоящем описании, обладают способностью встраивать неприродные рибонуклеотиды в нуклеиновую кислоту, такую как РНК. Например, рибонуклеотиды NaM или TAT1 встраиваются в нуклеиновые кислоты с использованием полимераз, представленных в настоящем описании.
[00407] Модифицированные полимеразы из описываемых композиций, необязательно, могут являться модифицированной и/или рекомбинантной ДНК-полимеразой Φ29-типа. Необязательно, полимераза может являться модифицированной и/или рекомбинантной полимеразой Φ29, B103, GA-1, PZA, Φ15, BS32, M2Y, Nf, G1, Cp-1, PRD1, PZE, SF5, Cp-5, Cp-7, PR4, PR5, PR722 или L17.
[00408] Модифицированные полимеразы из описываемых композиций, необязательно, могут являться модифицированной и/или рекомбинантной прокариотической ДНК-полимеразой, например, ДНК-полимеразой II (Pol II), ДНК-полимеразой III (Pol III), ДНК-полимеразой IV (Pol IV), ДНК-полимеразой V (Pol V). В некоторых вариантах осуществления модифицированные полимеразы включают полимеразы, опосредующие синтез ДНК среди неинструктивных поврежденных нуклеотидов. В некоторых вариантах осуществления гены, кодирующие Pol I, Pol II (polB), Poll IV (dinB) и/или Pol V (umuCD), конститутивно экспрессируются или гиперэкспрессируются в сконструированной клетке или SSO. В некоторых вариантах осуществления повышение экспрессии или гиперэкспрессия Pol II вносит вклад в повышенное удержание неприродных пар оснований (UBP) в сконструированной клетке или SSO.
[00409] Полимеразы нуклеиновых кислот, которые, как правило, можно использовать в изобретении, включают ДНК-полимеразы, РНК-полимеразы, обратные транскриптазы и их мутантные или измененные формы. ДНК-полимеразы и их свойства подробно описаны, помимо прочего, в DNA Replication 2nd edition, Kornberg and Baker, W. H. Freeman, New York, N. Y. (1991). Известные общепринятые ДНК-полимеразы, которые можно использовать в изобретении, включают, в качестве неограничивающих примеров, ДНК-полимеразу Pyrococcus furiosus (Pfu) (Lundberg et al., 1991, Gene, 108: 1, Stratagene), ДНК-полимеразу Pyrococcus woesei (Pwo) (Hinnisdaels et al., 1996, Biotechniques, 20:186-8, Boehringer Mannheim), ДНК-полимеразу Thermus thermophilus (Tth) (Myers and Gelfand 1991, Biochemistry 30:7661), ДНК-полимеразу Bacillus stearothermophilus (Stenesh and McGowan, 1977, Biochim Biophys Acta 475:32), ДНК-полимеразу Thermococcus litoralis (TIi) (также обозначаемую как ДНК-полимераза Vent™, Cariello et al, 1991, Polynucleotides Res, 19: 4193, New England Biolabs), ДНК-полимеразу 9 Nm™ (New England Biolabs), фрагмент Штоффеля, Thermo Sequenase® (Amersham Pharmacia Biotech UK), Therminator™ (New England Biolabs), ДНК-полимеразу Thermotoga maritima (Tma) (Diaz and Sabino, 1998 Braz J Med. Res, 31:1239), ДНК-полимеразу Thermus aquaticus (Taq) (Chien et al, 1976, J. Bacteoriol, 127: 1550), ДНК-полимеразу, ДНК-полимеразу Pyrococcus kodakaraensis KOD (Takagi et al., 1997, Appl. Environ. Microbiol. 63:4504), ДНК-полимеразу JDF-3 (из Thermococcus sp. JDF-3, патентная заявка № WO 0132887), ДНК-полимеразу Pyrococcus GB-D (PGB-D) (также обозначаемую как ДНК-полимераза Deep Vent™, Juncosa-Ginesta et al., 1994, Biotechniques, 16:820, New England Biolabs), ДНК-полимеразу UlTma (из термофильных Thermotoga maritima; Diaz and Sabino, 1998 Braz J. Med. Res, 31:1239; PE Applied Biosystems), ДНК-полимеразу Tgo (из Thermococcus gorgonarius, Roche Molecular Biochemicals), ДНК-полимеразу I E. coli (Lecomte and Doubleday, 1983, Polynucleotides Res. 11:7505), ДНК-полимеразу T7 (Nordstrom et al, 1981, J Biol. Chem. 256:3112) и ДНК-полимеразу II архей DP1I/DP2 (Cann et al, 1998, Proc. Natl. Acad. Sci. USA 95:14250). Предусмотрены и мезофильные полимеразы и термофильные полимеразы. Термофильные ДНК-полимеразы включают, в качестве неограничивающих примеров, ThermoSequenase®, 9 Nm™, Therminator™, Taq, Tne, Tma, Pfu, TfI, Tth, TIi, фрагмент Штоффеля, ДНК-полимеразу Vent™ и Deep Vent™, ДНК-полимеразу KOD, Tgo, JDF-3 и их мутантов, варианты и производные. Также предусмотрена полимераза, являющаяся 3'-экзонуклеаза-дефектным мутантом. Обратные транскриптазы, которые можно использовать в изобретении, включают, в качестве неограничивающих примеров, обратные транскриптазы из ВИЧ, HTLV-I, HTLV-II, FeLV, FIV, SIV, AMV, MMTV, MoMuLV и других ретровирусов (см. Levin, Cell 88:5-8 (1997); Verma, Biochim Biophys Acta. 473:1-38 (1977); Wu et al, CRC Crit Rev Biochem. 3:289-347(1975)). Дополнительные неограничивающие примеры полимераз включают ДНК-полимеразу 9 N, ДНК-полимеразу Taq, ДНК-полимеразу Phusion®, ДНК-полимеразу Pfu, ДНК-полимеразу RB69, ДНК-полимеразу KOD и ДНК-полимеразу VentR® (Gardner et al. (2004) "Comparative Kinetics of Nucleotide Analog Incorporation by Vent DNA Polymerase" J. Biol. Chem., 279(12), 11834-11842; Gardner and Jack "Determinants of nucleotide sugar recognition in an archaeon DNA polymerase" Nucleic Acids Research, 27(12) 2545-2553.). Полимеразы, выделенные из нетермофильных организмов, могут являться термически инактивируемыми. Примерами являются ДНК-полимеразы из фага. Следует понимать, что полимеразы из любого из множества источников можно модифицировать для повышения или снижения их толерантности к условиям высоких температур. В некоторых вариантах осуществления полимераза может являться термофильной. В некоторых вариантах осуществления термофильная полимераза может являться термически инактивируемой. Термофильные полимеразы, как правило, можно использовать для условий высоких температур или условий термоциклирования, таких как используемые для способов полимеразной цепной реакции (ПЦР).
[00410] В некоторых вариантах осуществления полимераза включает Φ29, B103, GA-1, PZA, Φ15, BS32, M2Y, Nf, G1, Cp-1, PRD1, PZE, SF5, Cp-5, Cp-7, PR4, PR5, PR722, L17, ThermoSequenase®, ДНК-полимеразу 9 Nm™, Therminator™, Tne, Tma, TfI, Tth, TIi, фрагмент Штоффеля, ДНК-полимеразу Vent™ и Deep Vent™, ДНК-полимеразу KOD, Tgo, JDF-3, Pfu, Taq, ДНК-полимеразу T7, РНК-полимеразу T7, PGB-D, ДНК-полимеразу UlTma, ДНК-полимеразу I E. coli, ДНК-полимеразу III E. coli, ДНК-полимеразу II DP1I/DP2 архей, ДНК-полимеразу 9 N, ДНК-полимеразу Taq, ДНК-полимеразу Phusion®, ДНК-полимеразу Pfu, РНК-полимеразу SP6, ДНК-полимеразу RB69, обратную транскриптазу вируса миелобластоза птиц (AMV), обратную транскриптазу вируса мышиного лейкоза Молони (MMLV), обратную транскриптазу SuperScript® II или обратную транскриптазу SuperScript® III.
[00411] В некоторых вариантах осуществления полимераза является ДНК-полимеразой I (или фрагментом Кленова), полимеразой Vent, ДНК-полимеразой Phusion®, ДНК-полимеразой KOD, полимеразой Taq, ДНК-полимеразой T7, РНК-полимеразой T7, ДНК-полимеразой Therminator™, полимеразой POLB, РНК-полимеразой SP6, ДНК-полимеразой I E. coli, ДНК-полимеразой III E. coli, обратной транскриптазой вируса миелобластоза птиц (AMV), обратной транскриптазой вируса мышиного лейкоза Молони (MMLV), обратной транскриптазой SuperScript® II или обратной транскриптазой SuperScript® III.
Транспортер нуклеотидов
[00412] Транспортеры нуклеотидов (NT) представляют собой группу мембранных транспортных белков, облегчающих перенос субстратов-нуклеотидов через мембраны клетки и везикулы. В некоторых вариантах осуществления существует два типа NT, консервативные транспортеры нуклеозидов и равновесные транспортеры нуклеозидов. В некоторых случаях NT также включают транспортеры органических анионов (OαΤ) и транспортеры органических катионов (OCT). В некоторых случаях нуклеотид транспортер является транспортером нуклеозидтрифосфата (NTT).
[00413] В некоторых вариантах осуществления транспортер нуклеозидтрифосфата (NTT) происходит из бактерий, растений или водорослей. В некоторых вариантах осуществления транспортер нуклеозидтрифосфата нуклеотида является TpNTT1, TpNTT2, TpNTT3, TpNTT4, TpNTT5, TpNTT6, TpNTT7, TpNTT8 (T. pseudonana), PtNTT1, PtNTT2, PtNTT3, PtNTT4, PtNTT5, PtNTT6 (P. tricornutum), GsNTT (Galdieria sulphuraria), AtNTT1, AtNTT2 (Arabidopsis thaliana), CtNTT1, CtNTT2 (Chlamydia trachomatis), PamNTT1, PamNTT2 (Protochlamydia amoebophila), CcNTT (Caedibacter caryophilus) или RpNTT1 (Rickettsia prowazekii).
[00414] В некоторых вариантах осуществления NTT является CNT1, CNT2, CNT3, ENT1, ENT2, OAT1, OAT3 или OCT1.
[00415] В некоторых вариантах осуществления NTT импортирует неприродные нуклеиновые кислоты в организм, например, клетку. В некоторых вариантах осуществления NTT можно модифицировать таким образом, что участок связывания нуклеотида NTT модифицируют для снижения стерического ингибирования поступления неприродной нуклеиновой кислоты в участок связывания нуклеотида. В некоторых вариантах осуществления NTT можно модифицировать для обеспечения усиленного взаимодействия с одним или более природными или неприродными признаками неприродных нуклеиновых кислот. Такие NTT можно экспрессировать или конструировать в клетках для стабильного импорта UBP в клетки. Таким образом, настоящее изобретение включает композиции, включающие гетерологичные или рекомбинантные NTT, и способы их применения.
[00416] NTT можно модифицировать способами, относящимися к белковой инженерии. Например, можно осуществлять молекулярное моделирование с учетом кристаллических структур для идентификации локализации NTT, где можно осуществлять мутации для модификации целевой активности или участка связывания. Остаток, идентифицированный как мишень для замены, можно заменять остатком, выбранным с использованием моделирования минимизации энергии, моделирования гомологии и/или консервативных аминокислотных замен, как описано в Bordo, et al. J Mol Biol 217: 721-729 (1991) и Hayes, et al. Proc Natl Acad Sci, USA 99: 15926- 15931 (2002), описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
[00417] Любые из множества NTT можно использовать в способе или композиции, приведенных в настоящем описании, включая, например, белковые ферменты, выделенные из биологических систем, и их функциональные варианты. Ссылку на конкретный NTT, такой как примеры ниже, следует понимать как включающую его функциональные варианты, если не указано иначе. В некоторых вариантах осуществления NTT является NTT дикого типа. В некоторых вариантах осуществления NTT является модифицированным или мутантным NTT.
[00418] Также можно использовать NTT с признаками для улучшения проникновения неприродных нуклеиновых кислот в клетки и координации с неприродными нуклеотидами в участке связывания нуклеотида. В некоторых вариантах осуществления модифицированный NTT имеет модифицированный участок связывания нуклеотида. В некоторых вариантах осуществления модифицированный NTT или NTT дикого типа имеет сниженную специфичность в отношении неприродной нуклеиновой кислоты. Например, NTT, необязательно, демонстрирует специфическую активность импорта в отношении неприродного нуклеотида, составляющую до по меньшей мере приблизительно 0,1% (например, приблизительно 0,1%, 0,2%, 0,5%, 0,8%, 1%, 1,1%, 1,2%, 1,5%, 1,8%, 2%, 3%, 4%, 5%, 10%, 25%, 50%, 75%, 100% или более) активности соответствующего NTT дикого типа. Необязательно, NTT демонстрирует kcat/Km или Vmax/Km в отношении неприродного нуклеотида, составляющее по меньшей мере до приблизительно 0,1% (например, приблизительно 0,1%, 0,2%, 0,5%, 0,8%, 1%, 1,1%, 1,2%, 1,5%, 1,8%, 2%, 3%, 4%, 5%, 10%, 25%, 50%, 75%, 100% или более) значения для NTT дикого типа.
[00419] NTT можно охарактеризовывать по их аффинности к трифосфату (т.е. Km) и/или скорости импорта (т.е. Vmax). В некоторых вариантах осуществления NTT имеет относительно Km или Vmax для одного или более природных и неприродных трифосфатов. В некоторых вариантах осуществления NTT имеет относительно высокую Km или Vmax для одного или более природных и неприродных трифосфатов.
[00420] NTT из нативных источников или их варианты можно подвергать скринингу с использованием анализа, с помощью которого определяют количество трифосфата (с использованием массового спектра или радиоактивности, если трифосфат мечен соответствующим образом). В одном из примеров NTT можно подвергать скринингу на способность импортировать неприродный трифосфат; например, dTPT3TP, dCNMOTP, d5SICSTP, dNaMTP, NaMTP и/или TPT1TP. Можно использовать NTT, например, гетерологичный NTT, демонстрирующий модифицированное свойство в отношении неприродной нуклеиновой кислоты по сравнению с NTT дикого типа. Например, модифицированное свойство может представлять собой, например, Km, kcat, Vmax для импорта трифосфата. В одном из вариантов осуществления модифицированное свойство представляет собой сниженную Km для неприродного трифосфата и/или повышенное kcat/Km или Vmax/Km для неприродного трифосфата. Аналогично, NTT, необязательно, имеет повышенную скорость связывания неприродного трифосфата, повышенную скорость внутриклеточного высвобождения и/или повышенную скорость импорта в клетку по сравнению с NTT дикого типа.
[00421] В то же время, NTT может импортировать в клетку природные трифосфаты, например, dATP, dCTP, dGTP, dTTP, АТФ, ЦТФ, ГТФ и/или ТТФ. В некоторых случаях NTT, необязательно, демонстрирует специфическую активность импорта природной нуклеиновой кислоты, способной поддерживать репликацию и транскрипцию. В некоторых вариантах осуществления NTT, необязательно, демонстрирует kcat/Km или Vmax/Km для природной нуклеиновой кислоты, способной поддерживать репликацию и транскрипцию.
[00422] NTT, используемые в настоящем описании, которые могут обладать способностью импортировать неприродный трифосфат с конкретной структурой, также можно получать с использованием подхода направленной эволюции. Анализ синтеза нуклеиновой кислоты можно использовать для скрининга на варианты NTT, имеющие специфичность в отношении любого из множества неприродных трифосфатов. Например, варианты NTT можно подвергать скринингу на способность импортировать неприродный трифосфат; например, d5SICSTP, dNaMTP, dCNMOTP, dTPT3TP, NaMTP и/или TPT1TP. В некоторых вариантах осуществления такой анализ является анализом in vitro, например, с использованием рекомбинантного варианта NTT. В некоторых вариантах осуществления такой анализ является анализом in vivo, например, экспрессией варианта NTT в клетке. Такие способы можно использовать для скрининга вариантов любого подходящего NTT на активность в отношении любых из неприродных трифосфатов, приведенных в настоящем описании.
Реагенты и инструменты-нуклеиновые кислоты
[00423] Реагент-нуклеотид и/или нуклеиновая кислота (или полинуклеотид) для использования со способом, клеткой или сконструированным микроорганизмом, представленным в настоящем описании, содержит одну или более ORF с неприродным нуклеотидом или без него. ORF может быть из любого подходящего источника, в некоторых случаях, из геномной ДНК, мРНК, обратно-транскрибированной РНК или комплементарный ДНК (кДНК), или библиотека нуклеиновых кислот, содержащей одно или более из указанного выше, и из любого вида организмов, имеющего интересующую последовательность нуклеиновой кислоты, интересующий белок или интересующую активность. Неограничивающие примеры организмов, из которых можно получать ORF, включают, например, бактерии, дрожжи, грибы, человека, насекомых, нематод, крупный рогатый скот, лошадей, собак, кошек, крыс или мышей. В некоторых вариантах осуществления реагент-нуклеотид и/или нуклеиновую кислоту или другой реагент, представленный в настоящем описании, выделяют или очищают. Можно получать ORF, включающие неприродные нуклеотиды с помощью описанных способов in vitro. В некоторых случаях реагент-нуклеотид или нуклеиновая кислота содержит неприродное нуклеиновое основание.
[00424] В некоторых случаях реагент-нуклеиновая кислота содержит нуклеотидную последовательность, смежную с ORF, транслируемую в комбинации с ORF, и кодирует аминокислотную метку. Нуклеотидная последовательность, кодирующая метку, находится в 3'- и/или 5'-направлении от ORF в реагенте-нуклеиновой кислоте, таким образом, кодирующей метку на C-конце или N-конце белка или пептида, кодируемого ORF. Можно использовать любую метку, устраняющую транскрипцию и/или трансляцию in vitro, и ее соответствующим образом может выбирать специалист в этой области. Метки могут облегчать выделение и/или очистку желаемого продукта ORF из культуры или ферментационных сред. В некоторых случаях библиотеки с реагентами-нуклеиновыми кислотами используют вместе со способами и композициями, представленными в настоящем описании. Например, библиотека из по меньшей мере 100, 1000, 2000, 5000, 10000 или более 50000 уникальных полинуклеотидов находится в библиотеке, где каждый полинуклеотид содержит по меньшей мере одно неприродное нуклеиновое основание.
[00425] Нуклеиновая кислота или реагент-нуклеиновая кислота с неприродным нуклеотидом или без него может содержать некоторые элементы, например, регуляторные элементы, зачастую выбранные в зависимости от предполагаемого использования нуклеиновой кислоты. Любой из следующих элементов можно включать или исключать из реагента-нуклеиновой кислоты. Реагент-нуклеиновая кислота, например, может включать один, или более, или все из следующих нуклеотидных элементов: один или более промоторных элементов, одну или более 5'-нетранслируемых областей (5'UTR), одну или более областей, в которые можно встраивать целевую нуклеотидную последовательность ("инсерционный элемент"), одну или более целевые нуклеотидные последовательности, одну или более 3'-нетранслируемых областей (3'UTR) и один или более селективных элементов. Реагент-нуклеиновую кислоту можно получать с одним или более из таких элементов, и в нуклеиновую кислоту можно встраивать другие элементы до встраивания нуклеиновой кислоты в желаемый организм. В некоторых вариантах осуществления реагент-нуклеиновая кислота по изобретению содержит промотор, 5'UTR, необязательную 3'UTR и инсерционные элементы, с помощью которых встраивают (т.е. клонируют) целевую нуклеотидную последовательность в реагент-нклеиновую кислоту. В некоторых вариантах осуществления реагент-нуклеиновая кислота по изобретению содержит промотор, инсерционные элементы и необязательную 3'UTR, и 5'UTR/целевую нуклеотидную последовательность встраивают с необязательной 3'UTR. Элементы можно располагать в любом порядке, подходящем для экспрессии в выбранной экспрессионной системе (например, экспрессии в выбранном организме или экспрессии в бесклеточной системе), и в некоторых вариантах осуществления реагент-нуклеиновая кислота содержит следующие элементы в 5'-3'-направлении: (1) промоторный элемент, 5'UTR и инсерционные элементы; (2) промоторный элемент, 5'UTR и целевую нуклеотидную последовательность; (3) промоторный элемент, 5'UTR, инсерционные элементы и 3'UTR; и (4) промоторный элемент, 5'UTR, целевую нуклеотидную последовательность и 3'UTR. В некоторых вариантах осуществления UTR можно оптимизировать для изменения или повышения транскрипции или трансляции ORF, являющейся полностью природной или содержащей неприродные нуклеотиды.
[00426] Реагенты-нуклеиновые кислоты, например, экспрессирующие кассеты и/или экспрессирующие векторы, могут включать различные регуляторные элементы, включая промоторы, энхансеры, последовательности инициации трансляции, последовательности терминации транскрипции и другие элементы. "Промотор", как правило, является последовательностью или последовательностями ДНК, функционирующими в относительно фиксированной локации относительно участка инициации транскрипции. Например, промотор может находиться выше сегмента нуклеиновой кислоты транспортера нуклеотидтрифосфатов. "Промотор" содержит основные элементы, необходимые для основного взаимодействия РНК-полимеразы и факторов транскрипции, и может содержать вышележащие элементы и чувствительные элементы. Термин "энхансер", как правило, относится к последовательности ДНК, функционирующей без фиксированного расстояния от участка инициации транскрипции, и он может находиться в 5'- или 3''-направлении от единицы транскрипции. Кроме того, энхансеры могут находиться в интроне, а также самой кодирующей последовательности. Как правило, они составляют от 10 до 300 п.н. в длину и функционируют в цис-положении. Энхансеры функционируют, усиливая транскрипцию с близлежащих промоторов. Энхансеры, подобно промоторам, зачастую также содержат чувствительные элементы, опосредующие регуляцию транскрипции. Энхансеры зачастую определяют регуляцию экспрессии, и их можно использовать для изменения или оптимизации экспрессии ORF, включая ORF, являющиеся полностью природными или содержащими неприродные нуклеотиды.
[00427] Как указано выше, реагенты-нуклеиновые кислоты также могут содержать одну или более 5'UTR и одну или более 3'UTR. Например, экспрессирующие векторы, используемые в эукариотических клетках-хозяевах (например, клетках дрожжей, грибов, насекомых, растений, животных, человека или ядросодержащих клетках) и прокариотических клетках-хозяевах (например, вирусе, бактериях), могут содержать последовательности, являющиеся сигналом терминации транскрипции, который может влиять на экспрессию мРНК. Эти области могут транскрибироваться в виде полиаденилированных сегментов в нетранслируемой части мРНК, кодирующей белок тканевого фактора. 3''-нетранслируемые области также включают участки терминации транскрипции. В некоторых предпочтительных вариантах осуществления единица транскрипции содержит область полиаденилирования. Одним из преимуществ этой области является то, что она повышает вероятность того, что транскрибируемая единица будет процессироваться и транспортироваться подобно мРНК. Идентификация и использование сигналов полиаденилирования в экспрессирующих конструкциях хорошо известны. В некоторых предпочтительных вариантах осуществления гомологичные сигналы полиаденилирования можно использовать в трансгенных конструкциях.
[00428] 5'UTR может содержать один или более элементов, эндогенных для нуклеотидной последовательности, из которой она происходит, и в некоторых случаях включает один или более экзогенных элементов. 5'UTR может происходить из любой подходящей нуклеиновой кислоты, такой как геномная ДНК, плазмида ДНК, РНК или мРНК, например, из любого подходящего организма (например, вируса, бактерии, дрожжей, грибов, растений, насекомых или млекопитающих). Специалист в этой области может выбирать подходящие элементы для 5'UTR в зависимости от выбранной системы экспрессии (например, экспрессии в выбранном организме или экспрессии в бесклеточной системе). В некоторых случаях 5'UTR содержит один или более из следующих элементов, известных специалисту в этой области: энхансерные последовательности (например, транскрипционные или трансляционные), участок инициации транскрипции, участок связывания фактора транскрипции, участок регуляции трансляции, участок инициации трансляции, участок связывания фактора трансляции, участок связывания акцессорного белка, участки связывания средств регуляции с обратной связью, Прибнов-бокс, TATA-бокс, элемент -35, E-бокс (связывающий элемент спираль-петля-спираль), участок связывания рибосомы, репликон, внутренний участок связывания рибосомы (IRES), сайленсерный элемент и т.п. В некоторых вариантах осуществления промоторный элемент можно выделять таким образом, что все элементы 5'UTR, необходимые для правильной зависящей от условий регуляции, содержатся во фрагменте промоторного элемента или в функциональной подпоследовательности фрагмента промоторного элемента.
[00429] 5'UTR в реагенте-нуклеиновой кислоте может содержать нуклеотидную последовательность трансляционного энхансера. Нуклеотидная последовательность трансляционного энхансера зачастую находится между промотором и целевой нуклеотидной последовательностью в реагенте-нуклеиновой кислоте. Последовательность трансляционного энхансера зачастую связывается с рибосомой, в некоторых случаях является 18S рРНК-связывающей рибонуклеотидной последовательностью (т.е. последовательностью, связывающейся с рибосомой 40S) и в некоторых случаях является внутренним участком посадки рибосомы (IRES). IRES, как правило, образует каркас РНК с точно расположенными третичными структурами РНК, контактирующими с субъединицей рибосомы 40S с помощью ряда специфических взаимодействий. Известны примеры рибосомальных энхансерных последовательностей, и их может определять специалист в этой области (например, Mignone et al., Nucleic Acids Research 33: D141-D146 (2005); Paulous et al., Nucleic Acids Research 31: 722-733 (2003); Akbergenov et al., Nucleic Acids Research 32: 239-247 (2004); Mignone et al., Genome Biology 3(3): reviews0004.1-0001,10 (2002); Gallie, Nucleic Acids Research 30: 3401-3411 (2002); Shaloiko et al., DOI: 10.1002/bit.20267; и Gallie et al., Nucleic Acids Research 15: 3257-3273 (1987); описание каждой из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме).
[00430] В некоторых случаях трансляционная энхансерная последовательность является эукариотической последовательностью, такой как консенсусная последовательность Козак или другая последовательность (например, последовательность гидроидного полипа, регистрационный номер GenBank U07128). В некоторых случаях трансляционная энхансерная последовательность является прокариотической последовательностью, такой как консенсусная последовательность Шайна-Дальгарно. В некоторых вариантах осуществления трансляционная энхансерная последовательность является вирусной нуклеотидной последовательностью. В некоторых случаях трансляционная энхансерная последовательность получена из 5'UTR вируса растений, такого как вирус табачной мозаики (TMV), вирус мозаики люцерны (AMV); вирус гравировки табака (ETV); вирус картофеля Y (PVY); вирус мозаики турнепса (poty) и вирус мозаики семян гороха. В некоторых вариантах осуществления последовательность омега длиной приблизительно 67 оснований из TMV включена в реагент-нуклеиновую кислоту в качестве трансляционной энхансерной последовательности (например, не содержит гуанозиновые нуклеотиды и включает центральную поли(CAA)-область длиной 25 нуклеотидов).
[00431] 3'UTR может содержать один или более элементов, эндогенных для нуклеотидной последовательности, из которой она происходит, и в некоторых случаях включает один или более экзогенных элементов. 3'UTR может происходить из любой подходящей нуклеиновой кислоты, такой как геномная ДНК, плазмида ДНК, РНК или мРНК, например, из любого подходящего организма (например, вируса, бактерии, дрожжей, грибов, растений, насекомых или млекопитающего). Специалист в этой области может выбирать подходящие элементы для 3'UTR в зависимости от выбранной системы экспрессии (например, экспрессии в выбранном организме). В некоторых случаях 3'UTR содержит один или более из следующих элементов, известных специалисту в этой области: участок регуляции транскрипции, участок инициации транскрипции, участок терминации транскрипции, участок связывания фактора транскрипции, участок регуляции трансляции, участок терминации трансляции, участок инициации трансляции, участок связывания фактора трансляции, участок связывания рибосомы, репликон, энхансерный элемент, сайленсерный элемент и полиаденозиновый хвост. 3'UTR зачастую включает полиаденозиновый хвост, а в некоторых случаях не включает, и если полиаденозиновый хвост присутствует, один или более остатков аденозина могут быть добавлены или делетированы из него (например, приблизительно 5, приблизительно 10, приблизительно 15, приблизительно 20, приблизительно 25, приблизительно 30, приблизительно 35, приблизительно 40, приблизительно 45 или приблизительно 50 остатков аденозина могут быть добавлены или удалены).
[00432] В некоторых вариантах осуществления модификацию 5'UTR и/или 3'UTR используют для изменения (например, повышения, добавления, снижения или, по существу, устранения) активности промотора. Изменение активности промотора, в свою очередь, может приводить к изменению активности пептида, полипептида или белка (например, активности фермента) посредством изменения транскрипции интересующих нуклеотидных последовательностей с функционально связанного промоторного элемента, содержащего модифицированную 5'- или 3'UTR. Например, в некоторых вариантах осуществления микроорганизм можно конструировать посредством генетической модификации для экспрессии реагента-нуклеиновой кислоты, содержащей модифицированную 5'- или 3'UTR, что может добавлять новую активность (например, активность, в норме не обнаруживаемую в организме-хозяине) или повышать экспрессию существующей активности посредством повышения транскрипции с гомологичного или гетерологичного промотора, функционально связанного с интересующей нуклеотидной последовательностью (например, гомологичной или гетерологичной интересующей нуклеотидной последовательностью). В некоторых вариантах осуществления микроорганизм можно конструировать посредством генетической модификации для экспрессии реагента-нуклеиновой кислоты, содержащей модифицированную 5'- или 3'UTR, что в некоторых вариантах осуществления может снижать экспрессию активности посредством снижения или, по существу, устранения транскрипции с гомологичного или гетерологичного промотора, функционально связанного с интересующей нуклеотидной последовательностью.
[00433] Экспрессию транспортера нуклеотидтрифосфатов из экспрессирующей кассеты или экспрессирующего вектора можно контролировать с помощью любого промотора, способного к экспрессии в прокариотических клетках или эукариотических клетках. Промоторный элемент, как правило, необходим для синтеза ДНК и/или синтеза РНК. Промоторный элемент часто содержит область ДНК, которая может облегчать транскрипцию конкретного гена, обеспечивая инициаторный участок для синтеза РНК, соответствующей гену. Промоторы, как правило, находятся вблизи генов, которые они регулируют, находятся выше гена (например, в 5'-направления от гена), и в некоторых вариантах осуществления находятся на той же цепи ДНК, которая является смысловой цепью гена. В некоторых вариантах осуществления промоторный элемент можно выделять из гена или организма и встраивать в функциональной связи с полинуклеотидной последовательностью, чтобы сделать возможной измененную и/или регулируемую экспрессию. Ненативный промотор (например, промотор, в норме неассоциированный с указанной последовательностью нуклеиновой кислоты), используемый для экспрессии нуклеиновой кислоты, часто обозначают как гетерологичный промотор. В некоторых вариантах осуществления гетерологичный промотор и/или 5'UTR можно встраивать в функциональной связи с полинуклеотидом, кодирующим полипептид, имеющий желаемую активность, представленную в настоящем описании. В рамках изобретения термины "функционально связанный" и "в функциональной связи с" в отношении промоторов относятся к взаимосвязи между кодирующей последовательностью и промоторным элементом. Промотор функционально связан или находится в функциональной связи с кодирующей последовательностью, когда экспрессия кодирующей последовательности посредством транскрипции регулируется или контролируется промоторным элементом. Термины "функционально связанный" и "в функциональной связи с" используют в настоящем описании взаимозаменяемо в отношении промоторных элементов.
[00434] Промотор часто взаимодействует с РНК-полимеразой. Полимераза является ферментом, катализирующим синтез нуклеиновых кислот с использованием уже существующего реагента-нуклеиновой кислоты. Если матрица является матрицей ДНК, молекула РНК транскрибируется перед синтезом белка. Ферменты, имеющие полимеразную активность, подходящую для использования в способах по изобретению, включают любую полимеразу, являющуюся активной в выбранной системе с выбранной матрицей для синтеза белка. В некоторых вариантах осуществления промотор (например, гетерологичный промотор), также обозначаемый в настоящем описании как промоторный элемент, может быть функционально связан с нуклеотидной последовательностью или открытой рамкой считывания (ORF). Транскрипция с промоторного элемента может катализовать синтез РНК, соответствующей нуклеотидной последовательности или последовательности ORF, функционально связанной с промотором, что, в свою очередь, приводит к синтезу желаемого пептида, полипептида или белка.
[00435] В некоторых случаях промоторные элементы демонстрируют чувствительность к регуляторному контролю. В некоторых случаях промоторные элементы также могут регулироваться селективным средством. Т.е. в некоторых случаях транскрипция с промоторных элементов может включаться, выключаться, подвергаться положительной регуляции или отрицательной регуляции в ответ на изменение обусловленных окружением, питательных или внутренних условий или сигналов (например, термически индуцибельные промоторы, светорегулируемые промоторы, регулируемые обратной связью промоторы, гормон-регулируемые промоторы, тканеспецифические промоторы, кислород- и pH-регулируемые промоторы, промоторы, чувствительные к селективным средствам (например, канамицину) и т.п.). На промоторы, на которые влияют обусловленные окружением, питательные или внутренние сигналы, часто влияет сигнал (прямой или непрямой), связывающийся в промоторе или вблизи него и в некоторых условиях повышающий или снижающий экспрессию целевой последовательности. Как и во всех способах, представленных в настоящем описании, включение природных или модифицированных промоторов можно использовать для изменения или оптимизации экспрессии полностью природной ORF (например, NTT или aaRS) или ORF, содержащей неприродный нуклеотид (например, мРНК или тРНК).
[00436] Неограничивающие примеры селективных или регуляторных средств, влияющих на транскрипцию с промоторного элемента, используемого в вариантах осуществления, представленных в настоящем описании, включают: (1) сегменты нуклеиновой кислоты, кодирующие продукты, обеспечивающие резистентность против в ином случае токсичных соединений (например, антибиотиков); (2) сегменты нуклеиновой кислоты, кодирующие продукты, в ином случае отсутствующие в клетке-реципиенте (например, основные продукты, гены тРНК, ауксотрофные маркеры); (3) сегменты нуклеиновой кислоты, кодирующие продукты, супрессирующие активность продукта гена; (4) сегменты нуклеиновой кислоты, кодирующие продукты, которые легко можно идентифицировать (например, фенотипические маркеры, такие как антибиотики (например, β-лактамаза), β-галактозидаза, зеленый флуоресцентный белок (GFP), желтый флуоресцентный белок (YFP), красный флуоресцентный белок (RFP), голубой флуоресцентный белок (CFP) и белки поверхности клетки); (5) сегменты нуклеиновой кислоты, связывающиеся с продуктами, в ином случае неблагоприятными для выживания и/или функционирования клеток; (6) сегменты нуклеиновой кислоты, в ином случае ингибирующие активность любых из сегментов нуклеиновой кислоты, описанных в №№ 1-5 выше (например, антисмысловые олигонуклеотиды); (7) сегменты нуклеиновой кислоты, связывающиеся с продуктами, модифицирующими субстрат (например, эндонуклеазы рестрикции); (8) сегменты нуклеиновой кислоты, которые можно использовать для выделения или идентификации желаемой молекулы (например, участки связывания специфического белка); (9) сегменты нуклеиновой кислоты, кодирующие специфическую нуклеотидную последовательность, которая в ином случае может являться нефункциональной (например, для ПЦР-амплификации субпопуляций молекул); (10) сегменты нуклеиновой кислоты, которые, если отсутствуют, прямо или косвенно придают резистентность или чувствительность к конкретным соединениям; (11) сегменты нуклеиновой кислоты, кодирующие продукты, являющиеся токсичными или превращающими относительно нетоксичное соединение в токсичное соединение (например, тимидинкиназа вируса простого герпеса, цитозиндезаминаза) в клетках-реципиентах; (12) сегменты нуклеиновой кислоты, ингибирующие репликацию, разделение или наследование молекулы нуклеиновой кислоты, содержащей их; (13) сегменты нуклеиновой кислоты, кодирующие функции зависящей от условий репликации, например, репликацию в некоторых организмах-хозяевах или штаммах клеток-хозяев или в некоторых условиях окружающей среды (например, температуре, питательных условиях и т.п.); и/или (14) нуклеиновые кислоты, кодирующие одну или более мРНК или тРНК, содержащих неприродные нуклеотиды. В некоторых вариантах осуществления регуляторное или селективное средство можно добавлять для изменения существующих условий роста, которым подвергают организм (например, рост в жидкой культуре, рост в ферментере, рост на твердых подложках с питательными веществами и т.п.).
[00437] В некоторых вариантах осуществления регуляцию промоторного элемента можно использовать для изменения (например, повышения, добавления, снижения или, по существу, устранения) активности пептида, полипептида или белка (например, ферментативной активности). Например, в некоторых вариантах осуществления микроорганизм можно конструировать посредством генетической модификации для экспрессии реагента-нуклеиновой кислоты, который может придавать новую активность (например, активность, в норме не обнаруживаемую в организме-хозяине) или повышать экспрессию существующей активности посредством повышения транскрипции с гомологичного или гетерологичного промотора, функционально связанного с интересующей нуклеотидной последовательностью (например, интересующей гомологичной или гетерологичной нуклеотидной последовательностью). В некоторых вариантах осуществления микроорганизм можно конструировать посредством генетической модификации для экспрессии реагента-нуклеиновой кислоты, который может снижать экспрессию активности посредством снижения или, по существу, устранения транскрипции с гомологичного или гетерологичного промотора, функционально связанного с интересующей нуклеотидной последовательностью.
[00438] Нуклеиновые кислоты, кодирующие гетерологичные белки, например, транспортеры нуклеотидтрифосфатов, можно встраивать в любую подходящую систему экспрессии или использовать с ними. В некоторых вариантах осуществления реагент-нуклеиновая кислота стабильно интегрирована в хромосому организма-хозяина, или в некоторых вариантах осуществления реагент-нуклеиновая кислота может представлять собой делецию части хромосомы хозяина (например, генетически модифицированных организмов, где изменение генома хозяина придает способность селективно или преференциально поддерживать желаемый организм, несущий генетическую модификацию). Такие реагенты-нуклеиновые кислоты (например, нуклеиновые кислоты или генетически модифицированные организмы, измененный геном которых придает организму подвергаемый селекции признак) можно выбирать по их способности направлять продукцию желаемого белка или молекулы нуклеиновой кислоты. При желании, реагент-нуклеиновую кислоту можно изменять таким образом, что кодоны кодируют (i) ту же аминокислоту с использованием иной тРНК, чем определенная в нативной последовательности, или (ii) иную аминокислоту, чем в норме, включая необщепринятые или неприродные аминокислоты (включая детектируемо меченые аминокислоты).
[00439] Рекомбинантную экспрессию эффективно осуществляют с использованием экспрессирующей кассеты, которая может являться частью вектора, такого как плазмида. Вектор может включать промотор, функционально связанный с нуклеиновой кислотой, кодирующей транспортер нуклеотидтрифосфатов. Вектор также может включать другие элементы, необходимые для транскрипции и трансляции, представленных в настоящем описании. Экспрессирующая кассета, экспрессирующий вектор и последовательности в кассете или векторе могут являться гетерологичными для клетки, с которой неприродные нуклеотиды приводят в контакт. Например, последовательность транспортер нуклеотидтрифосфатов может являться гетерологичной в отношении клетки.
[00440] Можно получать множество прокариотических и эукариотических экспрессирующих векторов, подходящих для переноса, кодирования и/или экспрессии транспортеров нуклеотидтрифосфатов. Такие экспрессирующие векторы включают, например, pET, pET3d, pCR2.1, pBAD, pUC и дрожжевые векторы. Векторы можно использовать, например, в различных условиях in vivo и in vitro. Неограничивающие примеры прокариотических промоторов, которые можно использовать, включают промоторы SP6, T7, T5, tac, bla, trp, gal, lac или промоторы мальтозы. Неограничивающие примеры эукариотических промоторов, которые можно использовать, включают конститутивные промоторы, например, вирусные промоторы, такие как промоторы CMV, SV40 и RSV, а также регулируемые промоторы, например, индуцибельный или репрессируемый промотор, такой как промотор tet, промотор hsp70, и синтетический промотор, регулируемый CRE. Векторы для бактериальной экспрессии включают pGEX-5X-3, и векторы для эукариотической экспрессии включают pCIneo-CMV. Вирусные векторы, которые можно использовать, включают векторы, относящиеся к лентивирусу, аденовирусу, аденоассоциированному вирусу, вирусу герпеса, вирусу осповакцины, вирусу полиомиелита, вирусу СПИД, нейротропному вирусу, вирусу Синдбис и другим вирусам. Также можно использовать любые семейства вирусов, обладающие общими свойствами с этими вирусами, что делает их подходящими для использования в качестве векторов. Ретровирусные векторы, которые можно использовать, включают вирусы, описанные в Verma, American Society for Microbiology, pp. 229-232, Washington, (1985). Например, такие ретровирусные векторы могут включать вирус мышиного лейкоза Молони, MMLV и другие ретровирусы, демонстрирующие желаемые свойства. Как правило, вирусные векторы содержат неструктурные ранние гены, структурные поздние гены, транскрипт РНК-полимеразы III, инвертированные концевые повторы, необходимые для репликации и капсидирования, и промоторы для контроля транскрипции и репликации вирусного генома. При конструировании в качестве векторов, у вирусов, как правило, удаляют один или более из ранних генов и встраивают ген или кассету ген/промотор в вирусный геном вместо удаленной вирусной нуклеиновой кислоты.
Клонирование
[00441] Для встраивания элемента, такого как ORF, в реагент-нуклеиновую кислоту можно использовать любую общепринятую стратегию клонирования, известную в этой области. Для встраивания элемента в матрицу, независимо от инсерционного элемента можно использовать известные способы, такие как (1) расщепление матрицы по одному или более существующим участкам распознавания рестрикционных ферментов и лигирование интересующего элемента и (2) добавление участков распознавания рестрикционных ферментов в матрицу посредством гибридизации олигонуклеотидных праймеров, включающих один или более подходящих участков распознавания рестрикционных ферментов, и амплификацию посредством полимеразной цепной реакции (подробно описанной в настоящем описании). В других стратегиях клонирования используют один или более участков инсерции, присутствующих или встроенных в реагент-нуклеиновую кислоту, таких как участок гибридизации олигонуклеотидного праймера для ПЦР, например, и другие, представленные в настоящем описании. В некоторых вариантах осуществления стратегию клонирования можно комбинировать с генетической манипуляцией, такой как рекомбинация (например, рекомбинация реагента-нуклеиновой кислоты с интересующей последовательностью нуклеиновой кислоты в геноме организма, подлежащего модификации, как представлено в настоящем описании). В некоторых вариантах осуществления клонированные ORF могут приводить к продукции (прямо или косвенно) модифицированных транспортеров нуклеотидтрифосфатов, и/или полимераз или транспортеров нуклеотидтрифосфатов, и/или полимераз дикого типа) посредством конструирования микроорганизма с одной или более интересующих ORF, где микроорганизм имеет измененные активности транспортера нуклеотидтрифосфатов или полимеразы.
[00442] Нуклеиновую кислоту можно специфически расщеплять посредством приведения нуклеиновой кислоты в контакт с одним или более специфическими расщепляющими средствами. Специфические расщепляющие средства зачастую будут специфически расщеплять конкретную нуклеотидную последовательность по конкретному участку. Неограничивающие примеры ферментативных специфических расщепляющих средств включают эндонуклеазы (например, ДНКазу (например, ДНКазу I, II); РНКазу (например, РНКазу E, F, H, P); фермент Cleavase™; ДНК-полимеразу Taq; ДНК-полимеразу E. coli I и эукариотические структура-специфические эндонуклеазы; эндонуклеазы FEN-1 мыши; эндонуклеазы рестрикции типа I, II или III, такие как Acc I, Afl III, Alu I, Alw44 I, Apa I, Asn I, Ava I, Ava II, BamH I, Ban II, Bcl I, Bgl I. Bgl II, Bln I, BsaI, Bsm I, BsmBI, BssH II, BstE II, Cfo I, CIa I, Dde I, Dpn I, Dra I, EcIX I, EcoR I, EcoR I, EcoR II, EcoR V, Hae II, Hae II, Hind II, Hind III, Hpa I, Hpa II, Kpn I, Ksp I, Mlu I, MIuN I, Msp I, Nci I, Nco I, Nde I, Nde II, Nhe I, Not I, Nru I, Nsi I, Pst I, Pvu I, Pvu II, Rsa I, Sac I, Sal I, Sau3A I, Sca I, ScrF I, Sfi I, Sma I, Spe I, Sph I, Ssp I, Stu I, Sty I, Swa I, Taq I, Xba I, Xho I); гликозилазы (например, урацил-ДНК-гликозилазу (UDG), 3-метиладенин-ДНК-гликозилазу, 3-метиладенин-ДНК-гликозилазу II, пиримидин гидрат-ДНК-гликозилазу, FaPy-ДНК-гликозилазу, мисматч-специфическую тимин-ДНК-гликозилазу, гипоксантин-ДНК-гликозилазу, 5-гидроксиметилурацил-ДНК-гликозилазу (HmUDG), 5-гидроксиметилцитозин-ДНК-гликозилазу или 1,N6-этено-аденин-ДНК-гликозилазу); экзонуклеазы (например, экзонуклеазу III); рибозимы и дезоксирибозимы. Образец нуклеиновой кислоты можно обрабатывать химическим средством или синтезировать с использованием модифицированных нуклеотидов, и можно расщеплять модифицированную нуклеиновую кислоту. В неограничивающих примерах образец нуклеиновой кислоты можно обрабатывать (i) алкилирующими средствами, такими как метилнитрозомочевина, с помощью которых получают несколько алкилированных оснований, включая N3-метиладенин и N3-метилгуанин, распознаваемых и расщепляемых алкилпурин-ДНК-гликозилазой; (ii) бисульфит натрия, вызывающий дезаминирование остатков цитозина в ДНК с образованием остатков урацила, которые могут расщепляться урацил-N-гликозилазой; и (iii) химическим средством, превращающим гуанин в его окисленную форму, 8-гидроксигуанин, который может расщепляться формамидопиримидин-ДНК-N-гликозилазой. Неограничивающие примеры способов химического расщепления включают алкилирование (например, алкилирование фосфотиоат-модифицированной нуклеиновой кислоты); расщепление кислотолабильной P3'-N5'-фосфорамидат-содержащей нуклеиновой кислоты; и обработка нуклеиновой кислоты тетраоксидом осмия и пиперидином.
[00443] В некоторых вариантах осуществления реагент-нуклеиновая кислота включает один или более участков инсерции рекомбиназы. Участок инсерции рекомбиназы является последовательностью распознавания на молекуле нуклеиновой кислоты, участвующей в реакции интеграции/рекомбинации с помощью рекомбинационных белков. Например, участком рекомбинации для рекомбиназы Cre является loxP, являющийся последовательностью из 34 пар оснований, состоящей из двух инвертированных повторов по 13 пар оснований (служащих в качестве участков связывания рекомбиназы), фланкирующих основную последовательность из 8 пар оснований (например, Sauer, Curr. Opin. Biotech. 5:521-527 (1994)). Другие примеры участков рекомбинации включают последовательности attB, attP, attL и attR и их мутанты, фрагменты, варианты и производные, распознаваемые белком рекомбинации λ Int и вспомогательными белками клеточным фактором интеграции (IHF), FIS и эксцизионазой (Xis) (например, патенты США №№ 5888732; 6143557; 6171861; 6270969; 6277608 и 6720140; патентные заявки США 3№ 09/517466 и 09/732914; патентную публикацию США № US2002/0007051; и Landy, Curr. Opin. Biotech. 3:699-707 (1993); описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме).
[00444] Примерами рекомбиназ, клонирующих нуклеиновые кислоты, являются системы Gateway® (Invitrogen, California), включающие по меньшей мере один участок рекомбинации для клонирования желаемых молекул нуклеиновой кислоты in vivo или in vitro. В некоторых вариантах осуществления в системе используют векторы, содержащие по меньшей мере два разных участка сайт-специфической рекомбинации, зачастую на основе системы бактериофага лямбда (например, att1 и att2) и мутантные относительно участков дикого типа (att0). Каждый мутантный участок имеет уникальную специфичность для его когнатного партнера-участка att (т.е. участка рекомбинации-его партнера по связыванию) одного типа (например, attB1 с attP1, или attL1 с attR1) и не будет перекрестно реагировать с участками рекомбинации другого мутантного типа или с участком att0 дикого типа. Разные специфичности участков делают возможным направленное клонирование или связывание желаемых молекул, таким образом, приводящее к желаемой ориентации клонированных молекул. Фрагменты нуклеиновых кислот, фланкированные участками рекомбинации, клонируют и субклонируют с использованием системы Gateway® посредством замены селективного маркера (например, ccdB), фланкированного участками att, на молекуле плазмиды-реципиента, в некоторых случаях обозначая ее как принимающий вектор. Затем желаемые клоны подвергают селекции посредством трансформации ccdB-чувствительного штамма организма-хозяина и положительной селекции по маркеру на молекуле-реципиенте. Схожие стратегии для отрицательной селекции (например, использование токсических генов) можно использовать в других организмах, например, тимидинкиназу (TK) у млекопитающих и насекомых.
[00445] В некоторых случаях реагент-нуклеиновая кислота содержит один или более элементов участков начала репликации (ORI). В некоторых вариантах осуществления матрица содержит два или более ORI, где один из них функционирует эффективно в одном организме (например, бактерии), а другой функционирует эффективно в другом организме (например, эукариоте, такой как дрожжи). В некоторых вариантах осуществления ORI может эффективно функционировать в одном биологическом виде (например, S. cerevisiae), а другой ORI может эффективно функционировать в другом биологическом виде (например, S. pombe). В некоторых случаях реагент-нуклеиновая кислота также включает один или более участков регуляции транскрипции.
[00446] Реагент-нуклеиновая кислота, например, экспрессирующая кассета или вектор, может включать последовательность нуклеиновой кислоты, кодирующую маркерный продукт. Маркерный продукт используют для определения того, доставлен ли ген в клетку и экспрессируется ли онпосле доставки. Примеры маркерных генов включают ген lacZ E. coli, кодирующий β-галактозидазу и зеленый флуоресцентный белок. В некоторых вариантах осуществления маркер может являться селективным маркером. Когда такие селективные маркеры успешно переносят в клетку-хозяина, трансформированная клетка-хозяин может выживать при помещении в условия селекции. Существует две широко используемые, отличающиеся категории селективных режимов. Первая категория основана на метаболизме клетки и использовании мутантной линии клеток, у которой отсутствует способность расти, независимо от дополненных сред. Вторая категория представляет собой доминантную селекцию, что означает схему селекции, используемую в любом типе клеток, и не требует использования мутантной линии клеток. В этих схемах, как правило, используют лекарственное средство для прекращения роста клетки-хозяина. Эти клетки, имеющие новый ген, будут экспрессировать белок, придающий резистентность к лекарственным средствам, и переживут селекцию. В примерах такой доминантной селекции используют лекарственные средства неомицин (Southern etal., J. Molec. Appl. Genet. 1: 327 (1982)), микофеноловую кислоту, (Mulligan et al., Science 209: 1422 (1980)) или гигромицин (Sugden, et al., Mol. Cell. Biol. 5: 410-413 (1985); описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме).
[00447] Реагент-нуклеиновая кислота может включать один или более селекционных элементов (например, элементов для селекции наличия реагента-нуклеиновой кислоты, а не для активации промоторного элемента, который может являться селективно регулируемым). Селекционные элементы зачастую используют с известными способами определения того, включена ли реагент-нуклеиновая кислота в клетку. В некоторых вариантах осуществления реагент-нуклеиновая кислота включает два или более селекционных элемента, где один функционирует эффективно в одном организма, а другой функционирует эффективно в другом организма. Неограничивающие примеры селекционных элементов включают: (1) сегменты нуклеиновой кислоты, кодирующие продукты, обеспечивающие резистентность к, в ином случае, токсичным соединениям (например, антибиотикам); (2) сегменты нуклеиновой кислоты, кодирующие продукты, которые, в ином случае, отсутствуют в клетке-реципиенте (например, незаменимые продукты, гены тРНК, ауксотрофные маркеры); (3) сегменты нуклеиновой кислоты, кодирующие продукты, супрессирующие активность продукта гена; (4) сегменты нуклеиновой кислоты, кодирующие продукты, которые легко можно идентифицировать (например, фенотипические маркеры, такие как антибиотики (например, β-лактамаза), β-галактозидаза, зеленый флуоресцентный белок (GFP), желтый флуоресцентный белок (YFP), красный флуоресцентный белок (RFP), голубой флуоресцентный белок (CFP) и белки поверхности клетки); (5) сегменты нуклеиновой кислоты, связывающие продукты, в ином случае неблагоприятные для выживания и/или функции клетки; (6) сегменты нуклеиновой кислоты, в ином случае ингибирующие активность любых из сегментов нуклеиновой кислоты, описанных под №№ 1-5 выше (например, антисмысловые олигонуклеотиды); (7) сегменты нуклеиновой кислоты, связывающие продукты, модифицирующие субстрат (например, рестрикционные эндонуклеазы); (8) сегменты нуклеиновой кислоты, которые можно использовать для выделения или идентификации желаемой молекулы (например, специфические участки связывания белков); (9) сегменты нуклеиновой кислоты, кодирующие специфическую нуклеотидную последовательность, которая, в ином случае, может являться нефункциональной (например, для ПЦР-амплификации субпопуляций молекул); (10) сегменты нуклеиновой кислоты, которые, если отсутствуют, прямо или косвенно придают резистентность или чувствительность к конкретным соединениям; (11) сегменты нуклеиновой кислоты, кодирующие продукты, являющиеся токсичными или превращающими относительно нетоксичное соединение в токсичное соединение (например, тимидинкиназа вируса простого герпеса, цитозиндезаминаза) в клетках-реципиентах; (12) сегменты нуклеиновой кислоты, ингибирующие репликацию, разделение или наследование молекул нуклеиновой кислоты, содержащих их; и/или (13) сегменты нуклеиновой кислоты, кодирующие функции зависящей от условий репликации, например, репликации в некоторых организмах-хозяевах или штаммах клеток-хозяев или в некоторых условиях окружающей среды (например, температура, пищевые условия и т.п.).
[00448] Реагент-нуклеиновая кислота может иметь любую форму, которую можно использовать для транскрипции и/или трансляции in vivo. В некоторых случаях нуклеиновая кислота является плазмидой, такой как сверхспиральная плазмида, в некоторых случаях является искусственной дрожжевой хромосомой (например, YAC), в некоторых случаях является линейной нуклеиновой кислотой (например, линейной нуклеиновой кислотой, получаемой посредством ПЦР или расщепления ферментами рестрикции), в некоторых случаях является одноцепочечной и в некоторых случаях является двухцепочечной. В некоторых случаях реагент-нуклеиновая кислота получают способом амплификации, таким как полимеразная цепная реакция (ПЦР) или опосредованной транскрипцией амплификации (TMA). При TMA используют два фермента в изотермической реакции для получения продуктов амплификации, определяемых по испусканию света (например, Biochemistry 1996 Jun 25;35(25):8429-38). Известны стандартные способы ПЦР (например, патенты США №№ 4683202; 4683195; 4965188 и 5656493), и, как правило, их осуществляют циклами. Каждый цикл включает термическую денатурацию, при которой гибридные нуклеиновые кислоты диссоциируют; охлаждение, при котором олигонуклеотидные праймеры гибридизуются; и элонгацию олигонуклеотидов под действием полимеразы (т.е. Taq-полимеразы). Пример циклического способа ПЦР представляет собой обработку образца при 95°C в течение 5 минут; повторяющиеся сорок пять циклов 95°C в течение 1 минуты, 59°C в течение 1 минуты 10 секунд, и 72°C в течение 1 минуты 30 секунд; а затем обработку образца при 72°C в течение 5 минут. Часто осуществляют множество циклов с использованием коммерчески доступного термоциклера. В некоторых случаях продукты ПЦР-амплификации хранят в течение периода времени при более низкой температуре (например, при 4°C) и в некоторых случаях замораживают (например, при -20°C) перед анализом.
[00449] Для получения ДНК, содержащей неприродные нуклеотиды, можно использовать стратегии клонирования, аналогичные описанным выше. Например, олигонуклеотиды, содержащие неприродные нуклеотиды в желаемых положениях, синтезируют с использованием стандартного твердофазного синтеза и очищать посредством ВЭЖХ. Затем олигонуклеотиды встраивают в плазмиду, содержащую необходимый контекст последовательности (т.е. UTR и кодирующую последовательность), способом клонирования (таким как Golden Gate Assembly) с использованием участков клонирования, таких как участки BsaI (хотя можно использовать и другие, описанные выше).
Наборы/промышленное изделие
[00450] В некоторых вариантах осуществления настоящее изобретение относится к наборам и промышленным изделиям для применения с одним или более способами, представленными в настоящем описании. Такие наборы включают носитель, упаковку или контейнер, компартментализованный для включения одного или более контейнеров, таких как флаконы, пробирки и т.п., где каждый из контейнеров содержит один из отдельных элементов для использования в способе, представленном в настоящем описании. Подходящие контейнеры включают, например, бутыли, флаконы, шприцы и пробирки. В одном из вариантов осуществления контейнеры получены из различных материалов, таких как стекло или пластик.
[00451] В некоторых вариантах осуществления набор включает подходящий упаковочный материал для вмещения содержимого набора. В некоторых случаях упаковочный материал получают хорошо известными способами, предпочтительно, для обеспечения стерильной среды без загрязнений. Упаковочные материалы, используемые в настоящем описании, могут включать, например, общепринято используемые в коммерческих наборах, продаваемых для использования с системами секвенирования нуклеиновых кислот. Неограничивающие примеры упаковочных материалов включают стекло, пластик, бумагу, фольгу и т.п., способные удерживать в фиксированных пределах компонент, представленный в настоящем описании.
[00452] Упаковочный материал может включать ярлык, на котором указано конкретное использование компонентов. Использование набора, указанное на ярлыке, может представлять собой один или более из способов, представленных в настоящем описании, при необходимости, для конкретной комбинации компонентов, присутствующих в наборе. Например, на ярлыке можно указывать, что набор можно использовать для способа синтеза полинуклеотида или способа определения последовательности нуклеиновой кислоты.
[00453] Инструкции по использованию упакованных реагентов или компонентов также можно включать в набор. Инструкции, как правило, будут включать ясное выражение, в котором описывают параметры реакции, такие как относительные количества компонентов набора и образца, подлежащие смешиванию, время поддержания смесей реагентов/образцов, температура, буферные условия и т.п.
[00454] Следует понимать, что не все компоненты, необходимые для конкретной реакции, должны обязательно находиться в конкретном наборе. Вместо этого, можно предоставлять один или более дополнительных компонентов из других источников. В инструкциях, предоставляемых вместе с набором, можно указывать дополнительные компоненты, которые необходимо предоставить, и где их можно получать.
[00455] В некоторых вариантах осуществления настоящее изобретение относится к набору, который можно использовать для стабильного встраивания неприродной нуклеиновой кислоты в клеточную нуклеиновую кислоту, например, способами, представленными в настоящем описании, для получения генетически сконструированных клеток. В одном из вариантов осуществления набор, представленный в настоящем описании, включает генетически сконструированную клетку и одну или более неприродных нуклеиновых кислот. В другом варианте осуществления набор, представленный в настоящем описании, включает выделенную и очищенную плазмиду, содержащую последовательность, выбранную из SEQ ID NO: 1-2. В дополнительном варианте осуществления набор, представленный в настоящем описании, включает праймер, содержащий последовательность, выбранную из SEQ ID NO: 3-20.
[00456] В дополнительных вариантах осуществления набор, представленный в настоящем описании, обеспечивает получение клетки и молекулы нуклеиновой кислоты, содержащей гетерологичный ген для встраивания в клетку, чтобы таким образом обеспечивать получение генетически сконструированной клетки, такой как экспрессирующие векторы, содержащие нуклеиновую кислоту по любому из вариантов осуществления, представленных выше в этом абзаце.
Примеры вариантов осуществления
[00457] Настоящее изобретение дополнительно описано с помощью следующих вариантов осуществления. Признаки каждого из вариантов осуществления можно комбинировать с любым из других вариантов осуществления, если это уместно и целесообразно.
[00458] Вариант осуществления 1. Способ in vivo получения белка, содержащего неприродную аминокислоту, включающий:
транскрипцию матрицы ДНК, содержащей первое неприродное основание и комплементарное второе неприродное основание, для встраивания третьего неприродного основания в мРНК, где третье неприродное основание сконфигурировано для образования первой неприродной пары оснований с первым неприродным основанием;
транскрипцию матрицы ДНК для встраивания четвертого неприродного основания в тРНК, где четвертое неприродное основание сконфигурировано для образования второй неприродной пары оснований со вторым неприродным основанием, где первая неприродная пара оснований и вторая неприродная пара оснований не являются одинаковыми; и
трансляцию белка из мРНК и тРНК, где указанный белок содержит неприродную аминокислоту.
[00459] Вариант осуществления 2. Способ по варианту осуществления 1, где способ in vivo включает использование полусинтетического организма.
[00460] Вариант осуществления 3. Способ по варианту осуществления 2, где организм включает микроорганизм.
[00461] Вариант осуществления 4. Способ по варианту осуществления 3, где организм включает бактерию.
[00462] Вариант осуществления 5. Способ по варианту осуществления 4, где организм включает грамположительную бактерию.
[00463] Вариант осуществления 6. Способ по варианту осуществления 4, где организм включает грамотрицательную бактерию.
[00464] Вариант осуществления 7. Способ по любому из вариантов осуществления 2-4, где организм включает Escherichia coli.
[00465] Вариант осуществления 8. Способ по любому из вариантов осуществления 1-7, где по меньшей мере одно неприродное основание выбрано из группы, состоящей из
(i) 2-тиоурацила, 2-тио-тимина, 2'-дезоксиуридина, 4-тио-урацила, 4-тио-тимина, урацил-5-ила, гипоксантин-9-ила (I), 5-галоурацила; 5-пропинил-урацила, 6-азо-тимина, 6-азо-урацила, 5-метиламинометилурацила, 5-метоксиаминометил-2-тиоурацила, псевдоурацила, метилового эфира урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусной кислоты, 5-метил-2-тиоурацила, 3-(3-амино-3-N-2-карбоксипропил)урацила, 5-метил-2-тиоурацила, 4-тиоурацила, 5-метилурацила, 5'-метоксикарбоксиметилурацила, 5-метоксиурацила, урацил-5-оксиуксусной кислоты, 5-(карбоксигидроксилметил)урацила, 5-карбоксиметиламинометил-2-тиоуридина, 5-карбоксиметиламинометилурацила или дигидроурацила;
(ii) 5-гидроксиметилцитозина, 5-трифторметилцитозина, 5-галоцитозина, 5-пропинилцитозина, 5-гидроксицитозина, циклоцитозина, цитозинарабинозида, 5,6-дигидроцитозина, 5-нитроцитозина, 6-азоцитозина, азацитозина, N4-этилцитозина, 3-метилцитозина, 5-метилцитозина, 4-ацетилцитозина, 2-тиоцитозина, феноксазинцитидин([5,4-b][1,4]бензоксазин-2(3H)-она), фенотиазина цитидин(1H-пиримидо[5,4-b][1,4]бензотиазин-2(3H)-она), феноксазина цитидин(9-(2-аминоэтокси)-H-пиримидо[5,4-b][1,4]бензоксазин-2(3H)-она), карбазолцитидин(2H-пиримидо[4,5-b]индол-2-она) или пиридоиндолцитидин(H-пиридо [3',2':4,5]пирроло [2,3-d]пиримидин-2-она);
(iii) 2-аминоаденина, 2-пропиладенина, 2-амино-аденина, 2-F-аденина, 2-амино-пропил-аденина, 2-амино-2'-дезоксиаденозина, 3-деазааденина, 7-метиладенина, 7-деаза-аденина, 8-азааденина, 8-галоген-, 8-амино-, 8-тиол-, 8-тиоалкил- и 8-гидроксил-замещенных аденинов, N6-изопентениладенина, 2-метиладенина, 2,6-диаминопурина, 2-метилтио-N6-изопентениладенина или 6-аза-аденина;
(iv) 2-метилгуанина, 2-пропиловых и алкиловых производных гуанина, 3-деазагуанина, 6-тио-гуанина, 7-метилгуанина, 7-деазагуанина, 7-деазагуанозина, 7-деаза-8-азагуанина, 8-азагуанина, 8-галоген-, 8-амино-, 8-тиол-, 8-тиоалкил- и 8-гидроксил-замещенных гуанинов, 1-метилгуанина, 2,2-диметилгуанина, 7-метилгуанина или 6-аза-гуанина; и
(v) гипоксантина, ксантина, 1-метилинозина, квеуозина, бета-D-галактозилквеуозина, инозина, бета-D-маннозилквеуозина, вибутоксозина, гидроксимочевины, (acp3)w, 2-аминопиридина или 2-пиридона.
[00466] Вариант осуществления 9. Способ по любому из вариантов осуществления 1-7, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
 .
.
[00467] Вариант осуществления 10. Способ по любому из вариантов осуществления 1-7, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
 .
.
[00468] Вариант осуществления 11. Способ по варианту осуществления 9, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
 .
.
[00469] Вариант осуществления 12. Способ по варианту осуществления 9, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
 .
.
[00470] Вариант осуществления 13. Способ по варианту осуществления 9, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
 .
.
[00471] Вариант осуществления 14. Способ по варианту осуществления 9, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
 .
.
[00472] Вариант осуществления 15. Способ по варианту осуществления 9, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
и .
[00473] Вариант осуществления 16. Способ по варианту осуществления 9, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из
 .
.
[00474] Вариант осуществления 17. Способ по варианту осуществления 9, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
 .
.
[00475] Вариант осуществления 18. Способ по варианту осуществления 9, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания представляет собой .
[00476] Вариант осуществления 19. Способ по варианту осуществления 9, где по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из:
 .
.
[00477] Вариант осуществления 20. Способ по варианту осуществления 9, где первое или второе неприродное основание представляет собой .
[00478] Вариант осуществления 21. Способ по варианту осуществления 9, где первое или второе неприродное основание представляет собой .
[00479] Вариант осуществления 22. способ по варианту осуществления 9, где первое неприродное основание представляет собой , и второе неприродное основание представляет собой .
[00480] Вариант осуществления 23. Способ по варианту осуществления 9, где первое неприродное основание представляет собой , и второе неприродное основание представляет собой .
[00481] Вариант осуществления 24. способ по варианту осуществления 9, где третье или четвертое неприродное основание представляет собой .
[00482] Вариант осуществления 25. Способ по варианту осуществления 9, где третье неприродное основание представляет собой .
[00483] Вариант осуществления 26. Способ по варианту осуществления 9, где четвертое неприродное основание представляет собой .
[00484] Вариант осуществления 27. Способ по варианту осуществления 9, где третье или четвертое неприродное основание представляет собой или .
[00485] Вариант осуществления 28. Способ по варианту осуществления 9, где третье неприродное основание представляет собой или .
[00486] Вариант осуществления 29. Способ по варианту осуществления 9, где четвертое неприродное основание представляет собой или .
[00487] Вариант осуществления 30. Способ по варианту осуществления 9, где первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой , и четвертое неприродное основание представляет собой или .
[00488] Вариант осуществления 31. Способ по варианту осуществления 9, где первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой , и четвертое неприродное основание представляет собой или .
[00489] Вариант осуществления 32. Способ по варианту осуществления 9, где первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой или , и четвертое неприродное основание представляет собой .
[00490] Вариант осуществления 33. Способ по варианту осуществления 9, где третье неприродное основание представляет собой или .
[00491] Вариант осуществления 34. Способ по варианту осуществления 9, где четвертое неприродное основание представляет собой .
[00492] Вариант осуществления 35. Способ по варианту осуществления 9, где первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой или , и четвертое неприродное основание представляет собой .
[00493] Вариант осуществления 36. Способ по любому из вариантов осуществления 9-35, где третье неприродное основание и четвертое неприродное основание содержат рибозу.
[00494] Вариант осуществления 37. Способ по любому из вариантов осуществления 9-35, где третье неприродное основание и четвертое неприродное основание содержат дезоксирибозу.
[00495] Вариант осуществления 38. Способ по любому из вариантов осуществления 9-35, где первое и второе неприродные основания содержат дезоксирибозу.
[00496] Вариант осуществления 39. Способ по любому из вариантов осуществления 9-35, где первое и второе неприродные основания содержат дезоксирибозу, и третье неприродное основание и четвертое неприродное основание содержат рибозу.
[00497] Вариант осуществления 40. Способ по варианту осуществления 9, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из группы, состоящей из
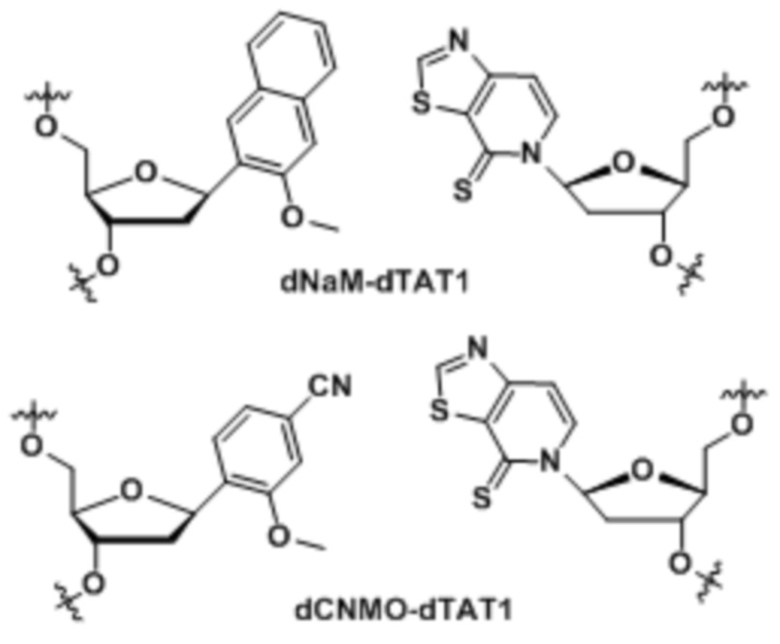 .
.
[00498] Вариант осуществления 41. Способ по варианту осуществления 40, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS.
[00499] Вариант осуществления 42. Способ по варианту осуществления 40, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3.
[00500] Вариант осуществления 43. способ по варианту осуществления 40, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3.
[00501] Вариант осуществления 44. Способ по варианту осуществления 40, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dPTMO-dTPT3.
[00502] Вариант осуществления 45. Способ по варианту осуществления 40, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTAT1.
[00503] Вариант осуществления 46. Способ по варианту осуществления 40, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTAT1.
[00504] Вариант осуществления 47. Способ по варианту осуществления 1, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из группы, состоящей из
; и
где мРНК и тРНК содержат по меньшей мере одно неприродное основание, выбранное из:
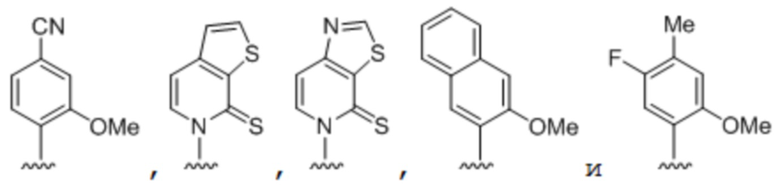 .
.
[00505] Вариант осуществления 48. Способ по варианту осуществления 47, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS.
[00506] Вариант осуществления 49. Способ по варианту осуществления 47, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3.
[00507] Вариант осуществления 50. Способ по варианту осуществления 47, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3.
[00508] Вариант осуществления 51. Способ по варианту осуществления 47, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dPTMO-dTPT3.
[00509] Вариант осуществления 52. Способ по варианту осуществления 47, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTAT1.
[00510] Вариант осуществления 53. Способ по варианту осуществления 47, где матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTAT1.
[00511] Вариант осуществления 54. Способ по любому из вариантов осуществления 47-53, где мРНК и тРНК содержат неприродное основание, выбранное из, и .
[00512] Вариант осуществления 55. Способ по варианту осуществления 54, где мРНК и тРНК содержат неприродное основание, выбранное из и .
[00513] Вариант осуществления 56. Способ по варианту осуществления 54, где мРНК содержит неприродное основание, являющееся .
[00514] Вариант осуществления 57. Способ по варианту осуществления 54, где мРНК содержит неприродное основание, являющееся .
[00515] Вариант осуществления 58. Способ по варианту осуществления 54, где мРНК содержит неприродное основание, являющееся .
[00516] Вариант осуществления 59. Способ по варианту осуществления 54, где тРНК содержит неприродное основание, выбранное из и .
[00517] Вариант осуществления 60. Способ по варианту осуществления 54, где тРНК содержит неприродное основание, являющееся .
[00518] Вариант осуществления 61. Способ по варианту осуществления 54, где тРНК содержит неприродное основание, являющееся .
[00519] Вариант осуществления 62. Способ по варианту осуществления 54, где тРНК содержит неприродное основание, являющееся .
[00520] Вариант осуществления 63. Способ по любому из вариантов осуществления 1-7, где первое неприродное основание содержит dCNMO, и второе неприродное основание содержит dTPT3.
[00521] Вариант осуществления 64. Способ по любому из вариантов осуществления 1-7 или 41, где третье неприродное основание содержит NaM, и второе неприродное основание содержит TAT1.
[00522] Вариант осуществления 65. Способ по любому из вариантов осуществления 1-64, где первое неприродное основание или второе неприродное основание распознается ДНК-полимеразой.
[00523] Вариант осуществления 66. Способ по любому из вариантов осуществления 1-65, где третье неприродное основание или четвертое неприродное основание распознается РНК-полимеразой.
[00524] Вариант осуществления 67. Способ по любому из вариантов осуществления 1-66, где белок содержит по меньшей мере две неприродных аминокислоты.
[00525] Вариант осуществления 68. Способ по любому из вариантов осуществления 1-66, где белок содержит по меньшей мере три неприродных аминокислоты.
[00526] Вариант осуществления 69. Способ по любому из вариантов осуществления 1-66, где белок содержит по меньшей мере две разные неприродные аминокислоты.
[00527] Вариант осуществления 70. Способ по любому из вариантов осуществления 1-66, где белок содержит по меньшей мере три разные неприродные аминокислоты.
[00528] Вариант осуществления 71. Способ по любому из вариантов осуществления 1-70, где по меньшей мере одна неприродная аминокислота:
является аналогом лизина;
содержит ароматическую боковую цепь;
содержит азидо-группу;
содержит алкиновую группу; или
содержит альдегидную или кетоновую группу.
[00529] Вариант осуществления 72. Способ по любому из вариантов осуществления 1-70, где по меньшей мере одна неприродная аминокислота не содержит ароматическую боковую цепь.
[00530] Вариант осуществления 73. Способ по любому из вариантов осуществления 1-70, где по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK), N6-пропаргилэтокси-карбонил-L-лизин (PraK), BCN-L-лизин, норборнен-лизин, TCO-лизин, метилтетразин-лизин, аллилоксикарбониллизин, 2-амино-8-оксононановую кислоту, 2-амино-8-оксооктановую кислоту, p-ацетил-L-фенилаланин, p-азидометил-L-фенилаланин (pAMF), p-йодо-L-фенилаланин, m-ацетилфенилаланин, 2-амино-8-оксононановую кислоту, p-пропаргилоксифенилаланин, p-пропаргил-фенилаланин, 3-метил-фенилаланин, L-ДОФА, фторированный фенилаланин, изопропил-L-фенилаланин, p-азидо-L-фенилаланин, p-ацил-L-фенилаланин, p-бензоил-L-фенилаланин, p-бромфенилаланин, p-амино-L-фенилаланин, изопропил-L-фенилаланин, O-аллилтирозин, O-метил-L-тирозин, O-4-аллил-L-тирозин, 4-пропил-L-тирозин, фосфонотирозин, три-O-ацетил-GlcNAcp-серин, L-фосфосерин, фосфоносерин, L-3-(2-нафтил)аланин, 2-амино-3-((2-((3-(бензилокси)-3-оксопропил)амино)этил)селанил)пропионовую кислоту, 2-амино-3-(фенилселанил)пропионовую кислоту или селеноцистеин.
[00531] Вариант осуществления 74. Способ по варианту осуществления 73, где по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK) и N6-пропаргилэтокси-карбонил-L-лизин (PraK).
[00532] Вариант осуществления 75. Способ по варианту осуществления 72, где по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK).
[00533] Вариант осуществления 76. Способ по варианту осуществления 72, где по меньшей мере одна неприродная аминокислота содержит N6-пропаргилэтокси-карбонил-L-лизин (PraK).
[00534] Вариант осуществления 77. Полусинтетический организм, содержащий расширенный генетический алфавит, где генетический алфавит содержит по меньшей мере два уникальных неприродных основания.
[00535] Вариант осуществления 78. Полусинтетический организм по варианту осуществления 77, где организм включает микроорганизм.
[00536] Вариант осуществления 79. Полусинтетический организм по любому из вариантов осуществления 77-78, где организм включает бактерию.
[00537] Вариант осуществления 80. Полусинтетический организм по варианту осуществления 79, где организм включает грамположительную бактерию.
[00538] Вариант осуществления 81. Полусинтетический организм по варианту осуществления 79, где организм включает грамположительную бактерию.
[00539] Вариант осуществления 82. Полусинтетический организм по любому из вариантов осуществления 77-79, где организм включает Escherichia coli.
[00540] Вариант осуществления 83. Полусинтетический организм по любому из вариантов осуществления 77-82, где по меньшей мере одно из неприродных оснований выбрано из группы, состоящей из 2-аминоаденин-9-ила, 2-аминоаденина, 2-F-аденина, 2-тиоурацила, 2-тио-тимина, 2-тиоцитозина, 2-пропиловых и алкиловых производных аденина и гуанина, 2-амино-аденина, 2-амино-пропил-аденина, 2-аминопиридина, 2-пиридона, 2'-дезоксиуридина, 2-амино-2'-дезоксиаденозина, 3-деазагуанина, 3-деазааденина, 4-тио-урацила, 4-тио-тимина, урацил-5-ила, гипоксантин-9-ила (I), 5-метил-цитозина, 5-гидроксиметилцитозина, ксантина, гипоксантина, 5-бром- и 5-трифторметил-урацилов и -цитозинов; 5-галоурацила, 5-галоцитозина, 5-пропинил-урацила, 5-пропинилцитозина, 5-урацила, 5-замещенных, 5-галоген, 5-замещенных пиримидинов, 5-гидроксицитозина, 5-бромцитозина, 5-бромурацила, 5-хлорцитозина, хлорированного цитозина, циклоцитозина, цитозинарабинозида, 5-фторцитозина, фторпиримидина, фторурацила, 5,6-дигидроцитозина, 5-йодцитозина, гидроксимочевины, йодурацила, 5-нитроцитозина, 5-бромурацила, 5-хлорурацила, 5-фторурацила и 5-йодурацила, 6-алкиловых производных аденина и гуанина, 6-азапиримидинов, 6-азо-урацила, 6-азоцитозина, азацитозина, 6-азо-тимина, 6-тио-гуанина, 7-метилгуанина, 7-метиладенина, 7-деазагуанина, 7-деазагуанозина, 7-деаза-аденина, 7-деаза-8-азагуанина, 8-азагуанина, 8-азааденина, 8-галоген-, 8-амино-, 8-тиол, 8-тиоалкил- и 8-гидроксил-замещенных аденинов и гуанинов; N4-этилцитозина, N-2-замещенных пуринов, N-6 замещенных пуринов, O-6-замещенных пуринов, соединений, повышающих стабильность образования дуплекса, универсальных нуклеиновых кислот, гидрофобных нуклеиновых кислот, смешанных нуклеиновых кислот, расширенных нуклеиновых кислот, фторированных нуклеиновых кислот, трициклических пиримидинов, феноксазина цитидин([5,4-b][1,4]бензоксазин-2(3H)-она), фенотиазина цитидин(1H- пиримидо[5,4-b][1,4]бензотиазин-2(3H)-она), гуанин-фиксирующих оснований, феноксазина цитидин(9-(2-аминоэтокси)-H-пиримидо[5,4-b][1,4]бензоксазин-2(3H)-она), карбазолцитидин (2H-пиримидо[4,5-b]индол-2-она), пиридоиндолцитидин(H-пиридо [3',2':4,5]пирроло [2,3-d]пиримидин-2-она), 5-фторурацила, 5-бромурацила, 5-хлорурацила, 5-йодурацила, гипоксантина, ксантина, 4-ацетилцитозина, 5-(карбоксигидроксилметил)урацила, 5-карбоксиметиламинометил-2-тиоуридина, 5-карбоксиметиламинометилурацила, дигидроурацила, бета-D-галактозилквеуозина, инозина, N6-изопентениладенина, 1-метилгуанина, 1-метилинозина, 2,2-диметилгуанина, 2-метиладенина, 2-метилгуанина, 3-метилцитозина, 5-метилцитозина, N6-аденина, 7-метилгуанина, 5-метиламинометилурацила, 5-метоксиаминометил-2-тиоурацила, бета-D-маннозилквеуозина, 5'-метоксикарбоксиметилурацила, 5-метоксиурацила, 2-метилтио-N6-изопентениладенина, урацил-5оксиуксусной кислоты, вибутоксозина, псевдоурацила, квеуозина, 2-тиоцитозина, 5-метил-2-тиоурацила, 2-тиоурацила, 4-тиоурацила, 5-метилурацила, метилового эфира урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусной кислоты, 5-метил-2-тиоурацила, 3-(3-амино-3-N-2-карбоксипропил)урацила, (acp3)w, 2,6-диаминопурина и соединений, в которых пуриновое или пиримидиновое основание заменяют гетероциклом.
[00541] Вариант осуществления 84. Полусинтетический организм по любому из вариантов осуществления 77-82, где организм содержит ДНК, содержащую по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
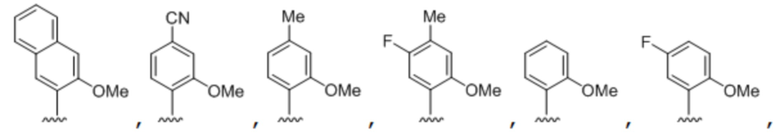
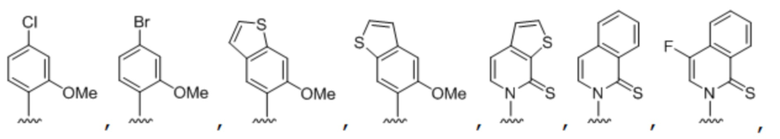
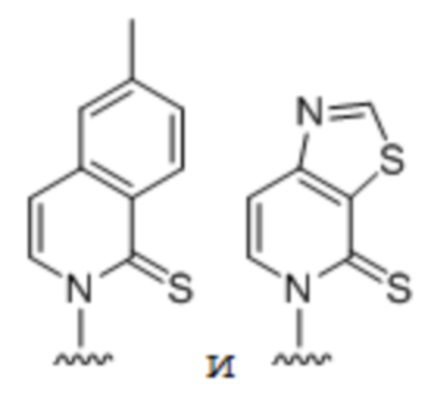 .
.
[00542] Вариант осуществления 85. Полусинтетический организм по любому из вариантов осуществления 77-84, где ДНК, содержащая по меньшей мере одно из неприродных оснований, образует неприродную пару оснований (UBP).
[00543] Вариант осуществления 86. Полусинтетический организм по варианту осуществления 85, где неприродная пара оснований (UBP) представляет собой dCNMO-dTPT3, dNaM-dTPT3, dCNMO-dTAT1 или dNaM-dTAT1.
[00544] Вариант осуществления 87. Полусинтетический организм по варианту осуществления 84, где ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
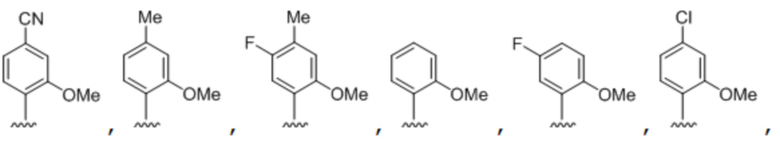
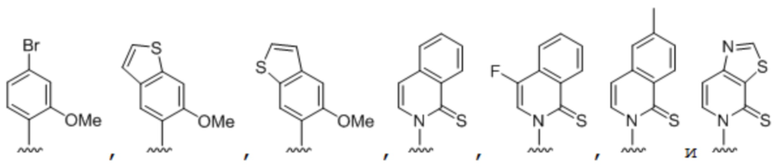 .
.
[00545] Вариант осуществления 88. Полусинтетический организм по варианту осуществления 87, где ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
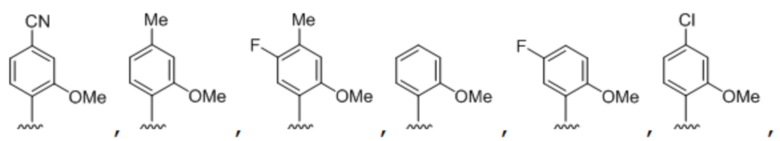
 .
.
[00546] Вариант осуществления 89. Полусинтетический организм по варианту осуществления 88, где ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
 .
.
[00547] Вариант осуществления 90. Полусинтетический организм по варианту осуществления 88, где ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
 .
.
[00548] Вариант осуществления 91. Полусинтетический организм по варианту осуществления 88, где ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
, и .
[00549] Вариант осуществления 92. Полусинтетический организм по варианту осуществления 88, где ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
и .
[00550] Вариант осуществления 93. Полусинтетический организм по варианту осуществления 88, где ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
 .
.
[00551] Вариант осуществления 94. Полусинтетический организм по варианту осуществления 88, где ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
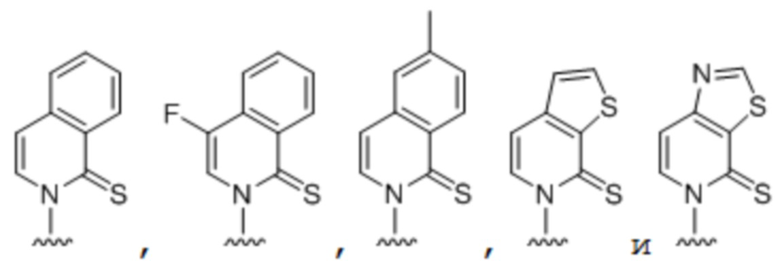 .
.
[00552] Вариант осуществления 95. Полусинтетический организм по варианту осуществления 88, где ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из и .
[00553] Вариант осуществления 96. Полусинтетический организм по варианту осуществления 88, где ДНК содержит по меньшей мере два неприродных нуклеиновых основания, выбранных из и .
[00554] Вариант осуществления 97. Полусинтетический организм по варианту осуществления 88, где ДНК содержит две цепи, где первая цепь содержит по меньшей мере одно нуклеиновое основание, являющееся , и вторая цепь содержит по меньшей мере одно нуклеиновое основание, являющееся .
[00555] Вариант осуществления 98. Полусинтетический организм по варианту осуществления 88, где ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, являющееся .
[00556] Вариант осуществления 99. Полусинтетический организм по любому из вариантов осуществления 77-98, где организм экспрессирует транспортер нуклеозидтрифосфата.
[00557] Вариант осуществления 100. Полусинтетический организм по варианту осуществления 99, где организм экспрессирует транспортер нуклеозидтрифосфата, являющийся PtNTT2.
[00558] Вариант осуществления 101. Полусинтетический организм по любому из вариантов осуществления 77-100, где организм дополнительно экспрессирует тРНК-синтетазу.
[00559] Вариант осуществления 102. Полусинтетический организм по варианту осуществления 101, где тРНК-синтетаза является пирролизил-тРНК-синтетазой M. barkeri (Mb PylRS).
[00560] Вариант осуществления 103. Полусинтетический организм по варианту осуществления 99, где организм экспрессирует транспортер нуклеозидтрифосфата PtNTT2 и дополнительно экспрессирует тРНК-синтетазу пирролизил-тРНК-синтетазу M. barkeri (Mb PylRS).
[00561] Вариант осуществления 104. Полусинтетический организм по любому из вариантов осуществления 77-103, где организм дополнительно экспрессирует РНК-полимеразу.
[00562] Вариант осуществления 105. Полусинтетический организм по варианту осуществления 104, где РНК-полимераза является RNAP T7.
[00563] Вариант осуществления 106. Полусинтетический организм по любому из вариантов осуществления 77-105, где организм не экспрессирует белок, имеющий функцию рекомбинационной репарации ДНК.
[00564] Вариант осуществления 107. Полусинтетический организм по любому из вариантов осуществления 77-105, где организм является E coli, и организм не экспрессирует RecA.
[00565] Вариант осуществления 108. Полусинтетический организм по любому из вариантов осуществления 77-107, дополнительно содержащий мРНК.
[00566] Вариант осуществления 109. Полусинтетический организм по варианту осуществления 108, где мРНК содержит по меньшей мере одно неприродное основание, выбранное из , и .
[00567] Вариант осуществления 110. Полусинтетический организм по варианту осуществления 108, где мРНК содержит по меньшей мере одно неприродное основание, являющееся .
[00568] Вариант осуществления 111. Полусинтетический организм по варианту осуществления 108, где мРНК содержит по меньшей мере одно неприродное основание, являющееся .
[00569] Вариант осуществления 112. Полусинтетический организм по варианту осуществления 108, где мРНК содержит по меньшей мере одно неприродное основание, являющееся .
[00570] Вариант осуществления 113. Полусинтетический организм по любому из вариантов осуществления 77-112, дополнительно содержащий тРНК.
[00571] Вариант осуществления 114. Полусинтетический организм по варианту осуществления 113, где тРНК содержит по меньшей мере одно неприродное основание, выбранное из , и .
[00572] Вариант осуществления 115. Полусинтетический организм по варианту осуществления 113, где тРНК содержит по меньшей мере одно неприродное основание, являющееся .
[00573] Вариант осуществления 116. Полусинтетический организм по варианту осуществления 113, где тРНК содержит по меньшей мере одно неприродное основание, являющееся .
[00574] Вариант осуществления 117. Полусинтетический организм по варианту осуществления 113, где тРНК содержит по меньшей мере одно неприродное основание, являющееся .
[00575] Вариант осуществления 118. Полусинтетический организм по любому из вариантов осуществления 77-107, дополнительно содержащий мРНК и тРНК.
[00576] Вариант осуществления 119. Полусинтетический организм по любому из вариантов осуществления 77-98, где организм дополнительно содержит (a) транспортер нуклеозидтрифосфата, (b) мРНК, (c) тРНК, (d) тРНК-синтетазу и (e) РНК-полимеразу, и где организм не экспрессирует белок, имеющий функцию рекомбинационной репарации ДНК.
[00577] Вариант осуществления 120. Полусинтетический организм по варианту осуществления 119, где транспортер нуклеозидтрифосфата является PtNTT2, тРНК-синтетаза является пирролизил-тРНК-синтетазой M. barkeri (Mb PylRS), и РНК-полимераза является RNAP T7.
[00578] Вариант осуществления 121. Полусинтетический организм по варианту осуществления 119, где организм является E. coli, и организм не экспрессирует RecA.
[00579] Вариант осуществления 122. Полусинтетический организм по варианту осуществления 118 или 119, где организм гиперэкспрессирует одну или более ДНК-полимераз.
[00580] Вариант осуществления 123. Полусинтетический организм по варианту осуществления 122, где организм гиперэкспрессирует ДНК-полимеразу II.
[00581] Вариант осуществления 124. Полусинтетический организм по любому из вариантов осуществления 77-123, где по меньшей мере одно неприродное основание дополнительно содержит неприродный сахарный остаток.
[00582] Вариант осуществления 125. Полусинтетический организм по варианту осуществления 124, где неприродный сахарный остаток выбран из группы, состоящей из:
модификации в 2'-положении:
OH, замещенного низшего алкила, алкарила, аралкила, O-алкарила или O-аралкила, SH, SCH3, OCN, Cl, Br, CN, CF3, OCF3, SOCH3, SO2CH3, ONO2, NO2, N3, NH2F;
O-алкила, S-алкила, N-алкила;
O-алкенила, S-алкенила, N-алкенила;
O-алкинила, S-алкинила, N-алкинила;
O-алкил-O-алкила, 2'-F, 2'-OCH3, 2'-O(CH2)2OCH3, где алкил, алкенил и алкинил могут являться замещенными или незамещенными C1-C10-алкилом, C2-C10-алкенилом, C2-C10-алкинилом, -O[(CH2)nO]mCH3, -O(CH2)nOCH3, -O(CH2)nNH2, -O(CH2)nCH3, -O(CH2)n-NH2, и -O(CH2)nON[(CH2)nCH3)]2, где n и m составляют от 1 до приблизительно 10;
и/или модификации в 5'-положении:
5'-винила, 5'-метила (R или S);
модификации в 4'-положении:
4'-S, гетероциклоалкила, гетероциклоалкарила, аминоалкиламиногруппы, полиалкиламиногруппы, замещенного силила, РНК-расщепляющей группы, репортерной группы, интеркалятора, группы для улучшения фармакокинетических свойств олигонуклеотида или группы для улучшения фармакодинамических свойств олигонуклеотида и любой их комбинации.
[00583] Вариант осуществления 126. Полусинтетический организм по любому из вариантов осуществления 77-125, где по меньшей мере одно неприродное основание распознается ДНК-полимеразой.
[00584] Вариант осуществления 127. Полусинтетический организм по любому из вариантов осуществления 73-126, где по меньшей мере одно неприродное основание распознается РНК-полимеразой.
[00585] Вариант осуществления 128. Композиция, содержащая аналог нуклеинового основания структуры:
,
где
каждый X независимо представляет собой углерод или азот;
R2 является необязательным и, если присутствует, независимо представляет собой водород, алкил, алкенил, алкинил, метоксигруппу, метантиол, метанселеногруппу, галоген, цианогруппу или азидную группу;
где каждый Y независимо представляет собой серу, кислород, селен или вторичный амин;
где каждый E независимо представляет собой кислород, серу или селен; и
где волнистой линией указана точка связывания с рибозильным, дезоксирибозильным или дидезоксирибозильным остатком или его аналогом, где рибозильный, дезоксирибозильный или дидезоксирибозильный остаток или его аналог находится в свободной форме, соединенной с монофосфатной, дифосфатной или трифосфатной группой, необязательно, содержащей α-тиотрифосфатную, β-тиотрифосфатную или γ-тиотрифосфатную группу, или включен в РНК, или ДНК, или аналог РНК, или аналог ДНК.
[00586] Вариант осуществления 129. Соединение по варианту осуществления 128, где рибозильный или дезоксирибозильный остаток несет трифосфатную или α-тиотрифосфатную группу, связанную с его 5'-гидроксилом.
[00587] Вариант осуществления 130. Соединение по любому из вариантов осуществления 128 или 129, где рибозильный или дезоксирибозильный остаток встроен в олигонуклеотидную цепь РНК или ДНК, соответственно, или рибозильный или дезоксирибозильный остаток или его аналог встроен в полинуклеотид.
[00588] Вариант осуществления 131. Соединение по любому из вариантов осуществления 128-130, где X является углеродом.
[00589] Вариант осуществления 132. Соединение по любому из вариантов осуществления 128-131, где E является серой.
[00590] Вариант осуществления 133. Соединение по любому из вариантов осуществления 128-132, где Y является серой.
[00591] Вариант осуществления 134. Соединение по варианту осуществления 128, где нуклеиновое основание содержит структуру .
[00592] Вариант осуществления 135. Соединение по любому из вариантов осуществления 128-134, где нуклеиновое связано с комплементарным, спаривающимся нуклеиновым основанием для образования неприродной пары оснований (UBP).
[00593] Вариант осуществления 136. Соединение по варианту осуществления 135, где комплементарное, спаривающееся нуклеиновое основание выбрано из:
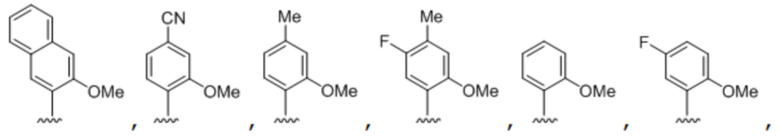
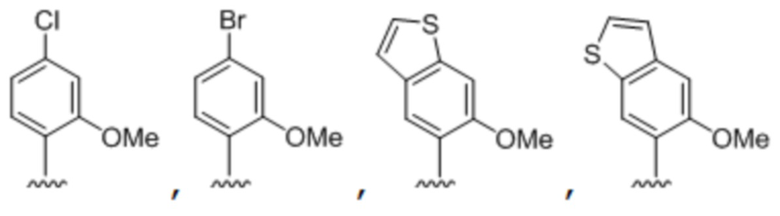 .
.
[00594] Вариант осуществления 137. Двухцепочечный олигонуклеотидный дуплекс, где первая олигонуклеотидная цепь содержит соединение по любому из вариантов осуществления 128-134, и вторая комплементарная олигонуклеотидная цепь содержит комплементарное, спаривающееся нуклеиновое основание в его комплементарном, образующем пару оснований участке.
[00595] Вариант осуществления 138. Двухцепочечный олигонуклеотидный дуплекс по варианту осуществления 137, где первая олигонуклеотидная цепь содержит ; и вторая цепь содержит комплементарное, спаривающееся нуклеиновое основание, выбранное из группы, состоящей из
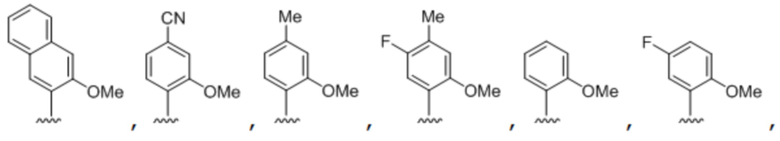
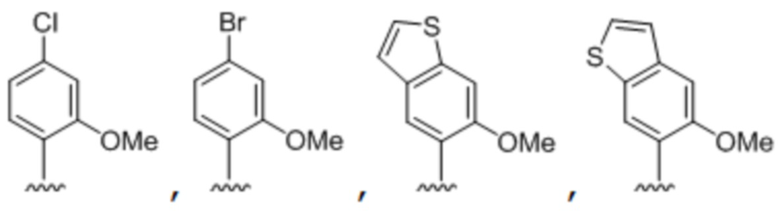
в его комплементарном, образующем пару оснований участке.
[00596] Вариант осуществления 139. Транспортная РНК (тРНК), содержащая аналог нуклеинового основания по любому из вариантов осуществления 128-136, содержащий:
антикодон, где антикодон содержит аналог нуклеинового основания; и
элемент распознавания, где элемент распознавания способствует селективной зарядке тРНК неприродной аминокислотой посредством аминоацил-тРНК-синтетазы.
[00597] Вариант осуществления 140. тРНК по варианту осуществления 139, где аналог нуклеинового основания находится в области антикодона тРНК.
[00598] Вариант осуществления 141. тРНК по варианту осуществления 140, где аналог нуклеинового основания находится в первом положении антикодона.
[00599] Вариант осуществления 142. тРНК по варианту осуществления 140, где аналог нуклеинового основания находится во втором положении антикодона.
[00600] Вариант осуществления 143. тРНК по варианту осуществления 140, где аналог нуклеинового основания находится в третьем положении антикодона.
[00601] Вариант осуществления 144. тРНК по варианту осуществления 139, где аминоацил-тРНК-синтетаза получена из термофильного организма.
[00602] Вариант осуществления 145. тРНК по варианту осуществления 139, где аминоацил-тРНК-синтетаза получена из Methanosarcina или его варианта.
[00603] Вариант осуществления 146. тРНК по варианту осуществления 139, где аминоацил-тРНК-синтетаза получена из Methanococcus (Methanocaldococcus) или его варианта.
[00604] Вариант осуществления 147. тРНК по варианту осуществления 139, где неприродная аминокислота содержит ароматический остаток.
[00605] Вариант осуществления 148. тРНК по варианту осуществления 139, где неприродная аминокислота является производным лизина.
[00606] Вариант осуществления 149. тРНК по варианту осуществления 139, где неприродная аминокислота является производным фенилаланина.
[00607] Вариант осуществления 150. тРНК по варианту осуществления 139, где неприродная аминокислота выбрана из группы, состоящей из: N6-азидоэтокси-карбонил-L-лизина (AzK), N6-пропаргилэтокси-карбонил-L-лизина (PraK), BCN-L-лизина, норборнен-лизина, TCO-лизина, метилтетразин-лизина, аллилоксикарбониллизина, 2-амино-8-оксононановой кислоты, 2-амино-8-оксооктановой кислоты, p-ацетил-L-фенилаланина, p-азидометил-L-фенилаланина (pAMF), p-йодо-L-фенилаланина, m-ацетилфенилаланина, 2-амино-8-оксононановой кислоты, p-пропаргилоксифенилаланина, p-пропаргил-фенилаланина, 3-метил-фенилаланина, L-ДОФА, фторированного фенилаланина, изопропил-L-фенилаланиан, p-азидо-L-фенилаланина, p-ацил-L-фенилаланина, p-бензоил-L-фенилаланина, p-бромфенилаланина, p-амино-L-фенилаланина, изопропил-L-фенилаланина, O-аллилтирозина, O-метил-L-тирозина, O-4-аллил-L-тирозина, 4-пропил-L-тирозина, фосфонотирозина, три-O-ацетил-GlcNAcp-серина, L-фосфосерина, фосфоносерина, L-3-(2-нафтил)аланина, 2-амино-3-((2-((3-(бензилокси)-3-оксопропил)амино)этил)селанил)пропионовой кислоты, 2-амино-3-(фенилселанил)пропионовой кислоты и селеноцистеина.
[00608] Вариант осуществления 151. Структура, содержащая формулу:
N1 - Zx - N2
где N1 является нуклеотидом, или его аналогом, или концевой фосфатной группой; где N2 является нуклеотидом, или его аналогом, или концевой гидроксильной группой; где Z является соединением по любому из вариантов осуществления 128-136, и где x является целым числом от 1 до 20.
[00609] Вариант осуществления 152. Структура по варианту осуществления 151, где структура кодирует ген.
[00610] Вариант осуществления 153. Структура по варианту осуществления 151, где Zx находится в транслируемой области гена.
[00611] Вариант осуществления 154. Структура по варианту осуществления 151, где Zx находится в нетранслируемой области гена.
[00612] Вариант осуществления 155. Структура по варианту осуществления 151, где структура дополнительно содержит 5'- или 3'-нетранслируемую область (UTR).
[00613] Вариант осуществления 156. Структура по варианту осуществления 151, где структура дополнительно содержит область терминатора.
[00614] Вариант осуществления 157. Структура по варианту осуществления 151, где структура дополнительно содержит промоторную область.
[00615] Вариант осуществления 158. Библиотека полинуклеотидов, где библиотека содержит по меньшей мере 5000 уникальных полинуклеотидов, и где каждый полинуклеотид содержит, по меньшей мере одно соединение по любому из вариантов осуществления 128-136.
[00616] Вариант осуществления 159. Библиотека по варианту осуществления 158, где библиотека полинуклеотидов кодирует по меньшей мере один ген.
[00617] Вариант осуществления 160. Нуклеозидтрифосфат, где нуклеиновое основание выбрано из
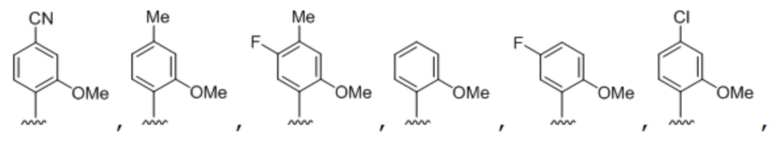
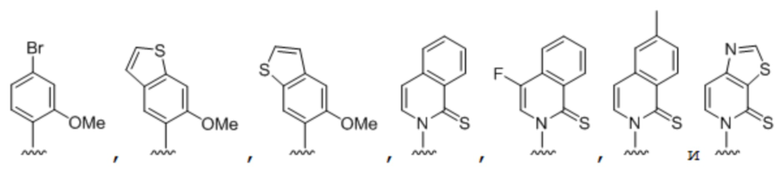 .
.
[00618] Вариант осуществления 161. Нуклеозидтрифосфат по варианту осуществления 160, где нуклеозид содержит рибозу.
[00619] Вариант осуществления 162. Нуклеозидтрифосфат по варианту осуществления 160, где нуклеозид содержит дезоксирибозу.
[00620] Вариант осуществления 163. Нуклеозидтрифосфат по любому из вариантов осуществления 160-162, где нуклеиновое основание выбрано из
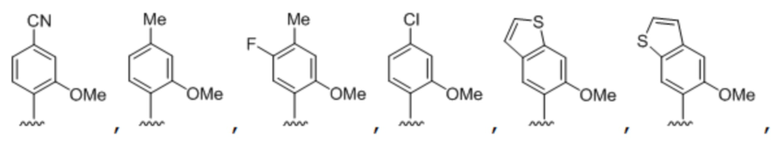
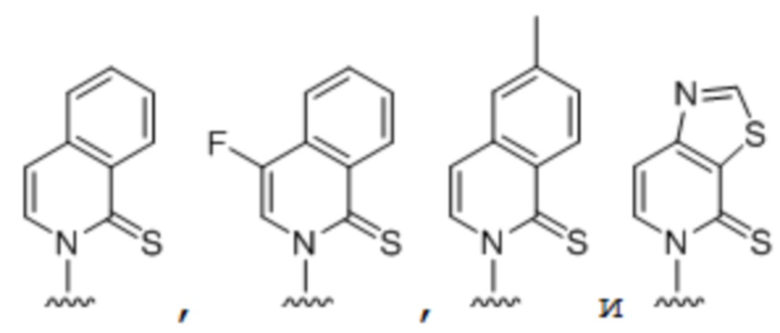 .
.
[00621] Вариант осуществления 164. Нуклеозидтрифосфат по варианту осуществления 163, где нуклеиновое основание выбрано из
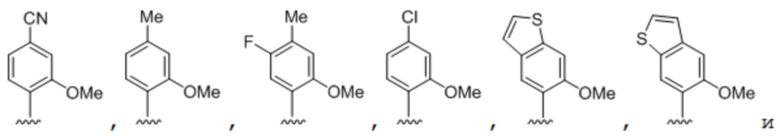 .
.
[00622] Вариант осуществления 165. Нуклеозидтрифосфат по варианту осуществления 163, где нуклеиновое основание выбрано из
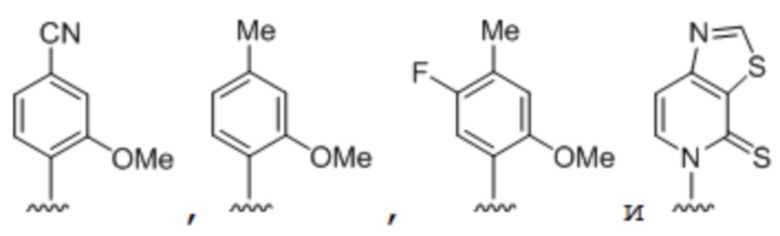 .
.
[00623] Вариант осуществления 166. Нуклеозидтрифосфат по варианту осуществления 163, где нуклеиновое основание выбрано из
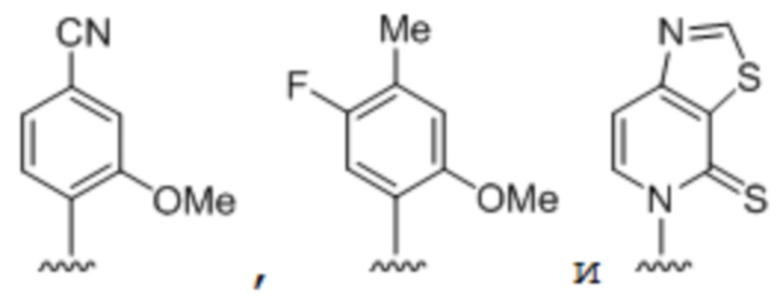 .
.
[00624] Вариант осуществления 167. Нуклеозидтрифосфат по варианту осуществления 165, где нуклеиновое основание представляет собой .
[00625] Вариант осуществления 168. Нуклеозидтрифосфат по варианту осуществления 167, где нуклеозид содержит рибозу.
[00626] Вариант осуществления 169. Нуклеозидтрифосфат по варианту осуществления 167, где нуклеозид содержит дезоксирибозу.
[00627] Вариант осуществления 170. Нуклеозидтрифосфат по варианту осуществления 165, где нуклеиновое основание представляет собой .
[00628] Вариант осуществления 171. Нуклеозидтрифосфат по варианту осуществления 170, где нуклеозид содержит рибозу.
[00629] Вариант осуществления 172. Нуклеозидтрифосфат по варианту осуществления 170, где нуклеозид содержит дезоксирибозу.
[00630] Вариант осуществления 173. Нуклеозидтрифосфат по варианту осуществления 165, где нуклеиновое основание представляет собой .
[00631] Вариант осуществления 174. Нуклеозидтрифосфат по варианту осуществления 173, где нуклеозид содержит рибозу.
[00632] Вариант осуществления 175. Нуклеозидтрифосфат по варианту осуществления 173, где нуклеозид содержит дезоксирибозу.
[00633] Настоящее изобретение относится к способам in vivo получения белка, содержащего неприродную аминокислоту, включающий: транскрипцию матрицы ДНК, содержащей первое неприродное основание и комплементарное второе неприродное основание для встраивания третьего неприродного основания в мРНК, при этом третье неприродное основание сконфигурировано для образования первой неприродной пары оснований с первым неприродным основанием; транскрипцию матрицы ДНК для встраивания четвертого неприродного основания в тРНК, где четвертое неприродное основание сконфигурировано для образования второй неприродной пары оснований со вторым неприродным основанием, где первая неприродная пара оснований и вторая неприродная пара оснований не являются одинаковыми; и трансляцию белка из мРНК и тРНК, где указанный белок содержит неприродную аминокислоту. В некоторых вариантах осуществления способ in vivo включает использование полусинтетического организма. В некоторых вариантах осуществления организм включает микроорганизм. В некоторых вариантах осуществления организм включает бактерию. В некоторых вариантах осуществления организм включает грамположительную бактерию. В некоторых вариантах осуществления организм включает грамотрицательную бактерию. В некоторых вариантах осуществления организм включает Escherichia coli. В некоторых вариантах осуществления по меньшей мере одно неприродное основание выбрано из группы, состоящей из (i) 2-тиоурацила, 2-тио-тимина, 2'-дезоксиуридина, 4-тио-урацила, 4-тио-тимина, урацил-5-ила, гипоксантин-9-ила (I), 5-галоурацила; 5-пропинил-урацила, 6-азо-тимина, 6-азо-урацила, 5-метиламинометилурацила,5-метоксиаминометил-2-тиоурацила, псевдоурацила, метилового эфира урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусной кислоты, 5-метил-2-тиоурацила, 3-(3-амино-3-N-2-карбоксипропил)урацила, 5-метил-2-тиоурацила, 4-тиоурацила, 5-метилурацила, 5'-метоксикарбоксиметилурацила, 5-метоксиурацила, урацил-5-оксиуксусной кислоты, 5-(карбоксигидроксилметил)урацила, 5-карбоксиметиламинометил-2-тиоуридина, 5-карбоксиметиламинометилурацила или дигидроурацила; (ii) 5-гидроксиметилцитозин, 5-трифторметилцитозина, 5-галоцитозина, 5-пропинилцитозина, 5-гидроксицитозина, циклоцитозина, цитозинарабинозида, 5,6-дигидроцитозина, 5-нитроцитозина, 6-азоцитозина, азацитозина, N4-этилцитозина, 3-метилцитозина, 5-метилцитозина, 4-ацетилцитозина, 2-тиоцитозина, феноксазина цитидин([5,4-b][1,4]бензоксазин-2(3H)-она), фенотиазина цитидин (1H-пиримидо[5,4-b][1,4]бензотиазин-2(3H)-она), феноксазина цитидин(9-(2-аминоэтокси)-H-пиримидо[5,4-b][1,4]бензоксазин-2(3H)-она), карбазолцитидин(2H-пиримидо[4,5- b]индол-2-она) или пиридоиндолцитидин(H-пиридо [3',2':4,5]пирроло [2,3-d]пиримидин-2-она); (iii) 2-аминоаденина, 2-пропиладенина, 2-амино-аденина, 2-F-аденина, 2-амино-пропил-аденина, 2-амино-2'-дезоксиаденозина, 3-деазааденина, 7-метиладенина, 7-деаза-аденина, 8-азааденина, 8-галоген-, 8-амино-, 8-тиол-, 8-тиоалкил- и 8-гидроксил-замещенных аденинов, N6-изопентениладенина, 2-метиладенина, 2,6-диаминопурина, 2-метилтио-N6-изопентениладенина или 6-аза-аденина; (iv) 2-метилгуанина, 2-пропиловых и алкиловых производных гуанина, 3-деазагуанина, 6-тио-гуанина, 7-метилгуанина, 7-деазагуанина, 7-деазагуанозина, 7-деаза-8-азагуанина, 8-азагуанина, 8-галоген-, 8-амино-, 8-тиол-, 8-тиоалкил- и 8-гидроксил-замещенных гуанинов, 1-метилгуанина, 2,2-диметилгуанина, 7-метилгуанина или 6-аза-гуанина; и (v) гипоксантина, ксантина, 1-метилинозина, квеуозина, бета-D-галактозилквеуозина, инозина, бета-D-маннозилквеуозина, вибутоксозина, гидроксимочевины, (acp3)w, 2-аминопиридина или 2-пиридона. В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбран из группы, состоящей из:
 . В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
. В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из: 
 . В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
. В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
 . В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
. В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
 . В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
. В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
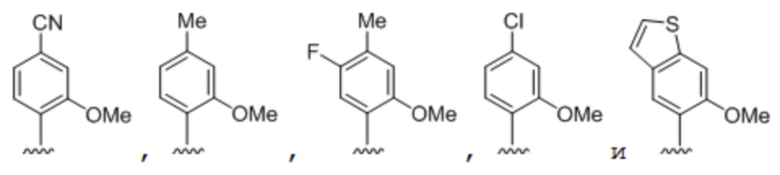 . Показано только основание нуклеозида или нуклеотида, при этом сахарный остаток и, необязательно, любые фосфатные остатки опущены для простоты. Здесь и на всем протяжении описания, волнистой линией указано соединение с дезокси- или рибонуклеозидом или нуклеотидом, в котором сахарная часть нуклеотида может быть дополнительно модифицирована. В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из: , и . В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из: и . В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
. Показано только основание нуклеозида или нуклеотида, при этом сахарный остаток и, необязательно, любые фосфатные остатки опущены для простоты. Здесь и на всем протяжении описания, волнистой линией указано соединение с дезокси- или рибонуклеозидом или нуклеотидом, в котором сахарная часть нуклеотида может быть дополнительно модифицирована. В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из: , и . В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из: и . В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
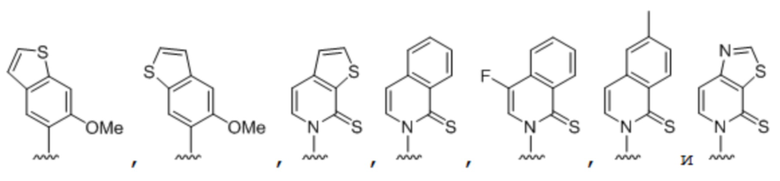 . В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
. В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из группы, состоящей из:
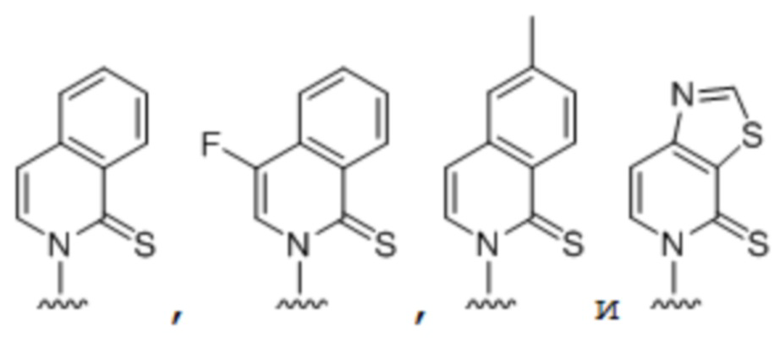 . В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания представляет собой . В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из:
. В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания представляет собой . В некоторых вариантах осуществления по меньшей мере одно из первого неприродного основания, второго неприродного основания, третьего неприродного основания или четвертого неприродного основания выбрано из:
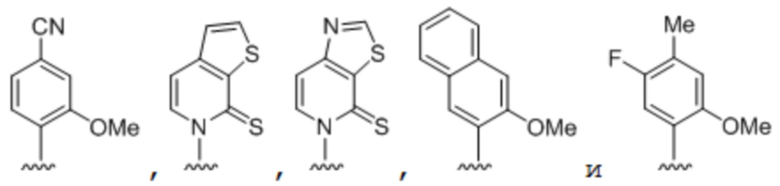 . В некоторых вариантах осуществления первое или второе неприродное основание представляет собой . В некоторых вариантах осуществления первое или второе неприродное основание представляет собой . В некоторых вариантах осуществления первое неприродное основание представляет собой , и второе неприродное основание представляет собой . В некоторых вариантах осуществления первое неприродное основание представляет собой , и второе неприродное основание представляет собой . В некоторых вариантах осуществления третье или четвертое неприродное основание представляет собой . В некоторых вариантах осуществления третье неприродное основание представляет собой . В некоторых вариантах осуществления четвертое неприродное основание представляет собой . В некоторых вариантах осуществления третье или четвертое неприродное основание представляет собой или . В некоторых вариантах осуществления третье неприродное основание представляет собой или . В некоторых вариантах осуществления четвертое неприродное основание представляет собой или . В некоторых вариантах осуществления первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой , и четвертое неприродное основание представляет собой или . В некоторых вариантах осуществления первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой , и четвертое неприродное основание представляет собой или . В некоторых вариантах осуществления первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой или , и четвертое неприродное основание представляет собой . В некоторых вариантах осуществления третье неприродное основание представляет собой или . В некоторых вариантах осуществления четвертое неприродное основание представляет собой . В некоторых вариантах осуществления первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой или , и четвертое неприродное основание представляет собой . В некоторых вариантах осуществления третье неприродное основание и четвертое неприродное основание содержат рибозу. В некоторых вариантах осуществления третье неприродное основание и четвертое неприродное основание содержат дезоксирибозу. В некоторых вариантах осуществления первое и второе неприродные основания содержат дезоксирибозу. В некоторых вариантах осуществления первое и второе неприродные основания содержат дезоксирибозу, и третье неприродное основание и четвертое неприродное основание содержат рибозу. В некоторых вариантах осуществления матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из группы, состоящей из:
. В некоторых вариантах осуществления первое или второе неприродное основание представляет собой . В некоторых вариантах осуществления первое или второе неприродное основание представляет собой . В некоторых вариантах осуществления первое неприродное основание представляет собой , и второе неприродное основание представляет собой . В некоторых вариантах осуществления первое неприродное основание представляет собой , и второе неприродное основание представляет собой . В некоторых вариантах осуществления третье или четвертое неприродное основание представляет собой . В некоторых вариантах осуществления третье неприродное основание представляет собой . В некоторых вариантах осуществления четвертое неприродное основание представляет собой . В некоторых вариантах осуществления третье или четвертое неприродное основание представляет собой или . В некоторых вариантах осуществления третье неприродное основание представляет собой или . В некоторых вариантах осуществления четвертое неприродное основание представляет собой или . В некоторых вариантах осуществления первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой , и четвертое неприродное основание представляет собой или . В некоторых вариантах осуществления первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой , и четвертое неприродное основание представляет собой или . В некоторых вариантах осуществления первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой или , и четвертое неприродное основание представляет собой . В некоторых вариантах осуществления третье неприродное основание представляет собой или . В некоторых вариантах осуществления четвертое неприродное основание представляет собой . В некоторых вариантах осуществления первое неприродное основание представляет собой , второе неприродное основание представляет собой , третье неприродное основание представляет собой или , и четвертое неприродное основание представляет собой . В некоторых вариантах осуществления третье неприродное основание и четвертое неприродное основание содержат рибозу. В некоторых вариантах осуществления третье неприродное основание и четвертое неприродное основание содержат дезоксирибозу. В некоторых вариантах осуществления первое и второе неприродные основания содержат дезоксирибозу. В некоторых вариантах осуществления первое и второе неприродные основания содержат дезоксирибозу, и третье неприродное основание и четвертое неприродное основание содержат рибозу. В некоторых вариантах осуществления матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из группы, состоящей из:
. В некоторых вариантах осуществления матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS. В некоторых вариантах осуществления матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3. В некоторых вариантах осуществления матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3. В некоторых вариантах осуществления матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dPTMO-dTPT3. В некоторых вариантах осуществления матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), выбранную из группы, состоящей из:
; и где мРНК и тРНК содержат по меньшей мере один неприродный рибонуклеотид с неприродным основанием, выбранным из: , , , и . В некоторых вариантах осуществления матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-d5SICS. В некоторых вариантах осуществления матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dCNMO-dTPT3. В некоторых вариантах осуществления матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dNaM-dTPT3. В некоторых вариантах осуществления матрица ДНК содержит по меньшей мере одну неприродную пару оснований (UBP), являющуюся dPTMO-dTPT3. В некоторых вариантах осуществления мРНК и тРНК содержат неприродное основание, выбранное из, и . В некоторых вариантах осуществления мРНК и тРНК содержат неприродное основание, выбранное из и . В некоторых вариантах осуществления мРНК содержит неприродное основание, являющееся . В некоторых вариантах осуществления мРНК содержит неприродное основание, являющееся . В некоторых вариантах осуществления мРНК содержит неприродное основание, являющееся . В некоторых вариантах осуществления тРНК содержит неприродное основание, выбранное из и . В некоторых вариантах осуществления тРНК содержит неприродное основание, являющееся . В некоторых вариантах осуществления тРНК содержит неприродное основание, являющееся . В некоторых вариантах осуществления тРНК содержит неприродное основание, являющееся . В некоторых вариантах осуществления первое неприродное основание содержит dCNMO, и второе неприродное основание содержит dTPT3. В некоторых вариантах осуществления третье неприродное основание содержит NaM, и второе неприродное основание содержит TAT1. В некоторых вариантах осуществления первое неприродное основание или второе неприродное основание распознается ДНК-полимеразой. В некоторых вариантах осуществления третье неприродное основание или четвертое неприродное основание распознается РНК-полимеразой. В некоторых вариантах осуществления белок содержит по меньшей мере две неприродных аминокислоты. В некоторых вариантах осуществления белок содержит по меньшей мере три неприродные аминокислоты. В некоторых вариантах осуществления по меньшей мере одна неприродная аминокислота: является аналогом лизина; содержит ароматическую боковую цепь; содержит азидо-группу; содержит алкиновую группу; или содержит альдегидную или кетоновую группу. В некоторых вариантах осуществления по меньшей мере одна неприродная аминокислота не содержит ароматическую боковую цепь. В некоторых вариантах осуществления по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK), N6-пропаргилэтокси-карбонил-L-лизин (PraK), BCN-L-лизин, норборнен-лизин, TCO-лизин, метилтетразин-лизин, аллилоксикарбониллизин, 2-амино-8-оксононановую кислоту, 2-амино-8-оксооктановую кислоту, p-ацетил-L-фенилаланин, p-азидометил-L-фенилаланин (pAMF), p-йодо-L-фенилаланин, m-ацетилфенилаланин, 2-амино-8-оксононановую кислоту, p-пропаргилоксифенилаланин, p-пропаргил-фенилаланин, 3-метил-фенилаланин, L-ДОФА, фторированный фенилаланин, изопропил-L-фенилаланин, p-азидо-L-фенилаланин, p-ацил-L-фенилаланин, p-бензоил-L-фенилаланин, p-бромфенилаланин, p-амино-L- фенилаланин, изопропил-L-фенилаланин, O-аллилтирозин, O-метил-L-тирозин, O-4-аллил-L-тирозин, 4-пропил-L-тирозин, фосфонотирозин, три-O-ацетил-GlcNAcp-серин, L-фосфосерин, фосфоносерин, L-3-(2-нафтил)аланин, 2-амино-3-((2-((3-(бензилокси)-3-оксопропил)амино)этил)селанил)пропионовую кислоту, 2-амино-3-(фенилселанил)пропионовую кислоту или селеноцистеин. В некоторых вариантах осуществления по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK) и N6-пропаргилэтокси-карбонил-L-лизин (PraK). В некоторых вариантах осуществления по меньшей мере одна неприродная аминокислота содержит N6-азидоэтокси-карбонил-L-лизин (AzK). В некоторых вариантах осуществления по меньшей мере одна неприродная аминокислота содержит N6-пропаргилэтокси-карбонил-L-лизин (PraK).
[00634] Настоящее изобретение относится к полусинтетическим организмам, содержащим расширенный генетический алфавит, где генетический алфавит содержит по меньшей мере три уникальных неприродных основания. В некоторых вариантах осуществления организм включает микроорганизм. В некоторых вариантах осуществления организм включает бактерию. В некоторых вариантах осуществления организм включает грамположительную бактерию. В некоторых вариантах осуществления организм включает грамположительную бактерию. В некоторых вариантах осуществления организм включает Escherichia coli. В некоторых вариантах осуществления по меньшей мере одно из неприродных оснований выбрано из группы, состоящей из 2-аминоаденин-9-ила, 2-аминоаденина, 2-F-аденина, 2-тиоурацила, 2-тио-тимина, 2-тиоцитозина, 2-пропиловых и алкиловых производных аденина и гуанина, 2-амино-аденина, 2-амино-пропил-аденина, 2-аминопиридина, 2-пиридона, 2'-дезоксиуридина, 2-амино-2'-дезоксиаденозина, 3-деазагуанина, 3-деазааденина, 4-тио-урацила, 4-тио-тимина, урацил-5-ила, гипоксантин-9-ила (I), 5-метил-цитозина, 5-гидроксиметилцитозина, ксантина, гипоксантина, 5-бром- и 5-трифторметил-урацилов и -цитозинов; 5-галоурацила, 5-галоцитозина, 5-пропинил-урацила, 5-пропинилцитозина, 5-урацила, 5-замещенных, 5-галоген, 5-замещенных пиримидинов, 5-гидроксицитозина, 5-бромцитозина, 5-бромурацила, 5-хлорцитозина, хлорированного цитозина, циклоцитозина, цитозинарабинозида, 5-фторцитозина, фторпиримидина, фторурацила, 5,6-дигидроцитозина, 5-йодцитозина, гидроксимочевины, йодурацила, 5-нитроцитозина, 5-бромурацила, 5-хлорурацила, 5-фторурацила и 5-йодурацила, 6-алкиловых производных аденина и гуанина, 6-азапиримидинов, 6-азо-урацила, 6-азоцитозина, азацитозинов, 6-азо-тимина, 6-тио-гуанина, 7-метилгуанина, 7-метиладенина, 7-деазагуанина, 7-деазагуанозина, 7-деаза-аденина, 7-деаза-8-азагуанина, 8-азагуанина, 8-азааденина, 8-галоген-, 8-амино-, 8-тиол-, 8-тиоалкил- и 8-гидроксил-замещенных аденинов и гуанинов; N4-этилцитозина, N-2-замещенных пуринов, N-6-замещенных пуринов, O-6-замещенных пуринов, соединений, повышающих стабильность образования дуплекса, универсальных нуклеиновых кислот, гидрофобных нуклеиновых кислот, смешанных нуклеиновых кислот, расширенных нуклеиновых кислот, фторированных нуклеиновых кислот, трициклических пиримидинов, феноксазина цитидин([5,4-b][й,4]бензоксазин-2(3H)-она), фенотиазина цитидин(1H- пиримидо[5,4-b][1,4]бензотиазин-2(3H)-она), гуанин-фиксирующих оснований, феноксазина цитидин(9-(2-аминоэтокси)-H-пиримидо[5,4-b][1,4]бензоксазин-2(3H)-она), карбазолцитидин(2H-пиримидо[4,5- b]индол-2-она), пиридоиндолцитидин(H-пиридо[3',2':4,5]пирроло[2,3-d]пиримидин-2-она), 5-фторурацила, 5-бромурацила, 5-хлорурацила, 5-йодурацила, гипоксантина, ксантина, 4-ацетилцитозина, 5-(карбоксигидроксилметил)урацила, 5-карбоксиметиламинометил-2-тиоуридина, 5-карбоксиметиламинометилурацила, дигидроурацила, бета-D-галактозилквеуозина, инозина, N6-изопентениладенина, 1-метилгуанина, 1-метилинозина, 2,2-диметилгуанина, 2-метиладенина, 2-метилгуанина, 3-метилцитозина, 5-метилцитозина, N6-аденина, 7-метилгуанина, 5-метиламинометилурацила, 5-метоксиаминометил-2-тиоурацила, бета-D-маннозилквеуозина, 5'-метоксикарбоксиметилурацила, 5-метоксиурацила, 2-метилтио-N6-изопентениладенина, урацил-5-оксиуксусной кислоты, вибутоксозина, псевдоурацила, квеуозина, 2-тиоцитозина, 5-метил-2-тиоурацила, 2-тиоурацила, 4-тиоурацила, 5-метилурацила, метилового эфира урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусной кислоты, 5-метил-2-тиоурацила, 3-(3-амино-3-N-2-карбоксипропил)урацила, (acp3)w, 2,6-диаминопурина и соединений, в которых пуриновое или пиримидиновое основание заменяют гетероциклом. В некоторых вариантах осуществления организм содержит ДНК, содержащую по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
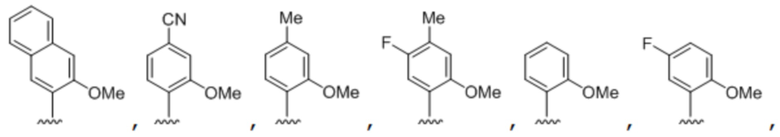
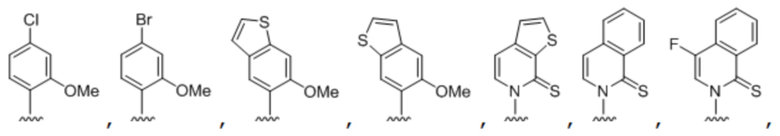
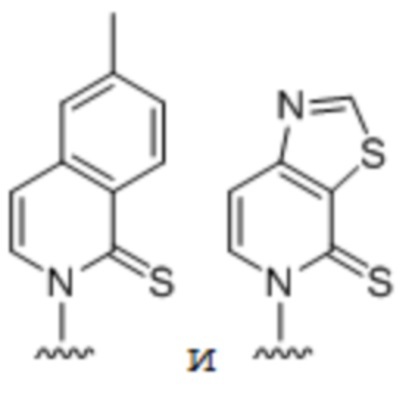 . В некоторых вариантах осуществления ДНК, содержащая по меньшей мере одно из неприродных оснований, образует неприродную пару оснований (UBP). В некоторых вариантах осуществления неприродная пара оснований (UBP) является dCNMO-dTPT3, dNaM-dTPT3, dCNMO-dTAT1 или dNaM-dTAT1. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
. В некоторых вариантах осуществления ДНК, содержащая по меньшей мере одно из неприродных оснований, образует неприродную пару оснований (UBP). В некоторых вариантах осуществления неприродная пара оснований (UBP) является dCNMO-dTPT3, dNaM-dTPT3, dCNMO-dTAT1 или dNaM-dTAT1. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
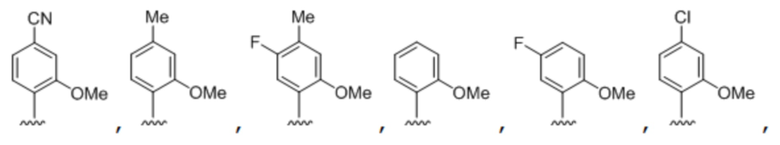
 . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из: 
 . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:  . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:  . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:  . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из: и . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из: и . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:  . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:
. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из группы, состоящей из:  . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из и . В некоторых вариантах осуществления ДНК содержит по меньшей мере два неприродных нуклеиновых основания, выбранных из и . В некоторых вариантах осуществления ДНК содержит две цепи, при этом первая цепь содержит по меньшей мере одно нуклеиновое основание, являющееся , и вторая цепь содержащий по меньшей мере одно нуклеиновое основание, являющееся . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, являющееся . В некоторых вариантах осуществления организм экспрессирует транспортер нуклеозидтрифосфата. В некоторых вариантах осуществления организм экспрессирует транспортер нуклеозидтрифосфата является PtNTT2. В некоторых вариантах осуществления организм дополнительно экспрессирует тРНК-синтетазу. В некоторых вариантах осуществления тРНК-синтетаза является пирролизил-тРНК-синтетазой M. barkeri (Mb PylRS). В некоторых вариантах осуществления организм экспрессирует транспортер нуклеозидтрифосфата PtNTT2 и дополнительно экспрессирует тРНК-синтетазу пирролизил-тРНК-синтетазу M. barkeri (Mb PylRS). В некоторых вариантах осуществления организм дополнительно экспрессирует РНК-полимеразу. В некоторых вариантах осуществления РНК-полимераза является RNAP T7. В некоторых вариантах осуществления организм не экспрессирует белок, имеющий функцию рекомбинационной репарации ДНК. В некоторых вариантах осуществления организм является E coli и организм не экспрессирует RecA. В некоторых вариантах осуществления организм гиперэкспрессирует ДНК-полимеразу. В некоторых вариантах осуществления организм гиперэкспрессирует ДНК-полимеразу II. В некоторых вариантах осуществления полусинтетический организм дополнительно содержит мРНК. В некоторых вариантах осуществления мРНК содержит по меньшей мере одно неприродное основание, выбранное из , и . В некоторых вариантах осуществления мРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления мРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления мРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления полусинтетический организм дополнительно содержит тРНК. В некоторых вариантах осуществления тРНК содержит по меньшей мере одно неприродное основание, выбранное из , и . В некоторых вариантах осуществления тРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления тРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления тРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления полусинтетический организм дополнительно содержит мРНК и/или тРНК. В некоторых вариантах осуществления организм дополнительно содержит (a) транспортер нуклеозидтрифосфата, (b) мРНК, (c) тРНК, (d) тРНК-синтетазу и (e) РНК-полимеразу, и где организм не экспрессирует белок, имеющий функцию рекомбинационной репарации ДНК. В некоторых вариантах осуществления транспортер нуклеозидтрифосфата является PtNTT2, тРНК-синтетаза является пирролизил-тРНК-синтетазой M. barkeri (Mb PylRS), и РНК-полимераза является RNAP T7. В некоторых вариантах осуществления организм является E coli, и организм не экспрессирует RecA. В некоторых вариантах осуществления организм гиперэкспрессирует одну или более ДНК-полимераз. В некоторых вариантах осуществления организм гиперэкспрессирует одну ДНК-полимеразу II. В некоторых вариантах осуществления по меньшей мере одно неприродное основание дополнительно содержит неприродный сахарный остаток. В некоторых вариантах осуществления неприродный сахарный остаток выбран из группы, состоящей из: модификации в 2'-положении: OH, замещенного низшего алкила, алкарила, аралкила, O-алкарила или O-аралкила, SH, SCH3, OCN, Cl, Br, CN, CF3, OCF3, SOCH3, SO2CH3, ONO2, NO2, N3, NH2F; O-алкила, S-алкила, N-алкила; O-алкенила, S-алкенила, N-алкенила; O-алкинила, S-алкинила, N-алкинила; O-алкил-O-алкила, 2'-F, 2'-OCH3, 2'-O(CH2)2OCH3, где алкил, алкенил и алкинил могут являться замещенным или незамещенным C1-C10-алкилом, C2-C10-алкенилом, C2-C10-алкинилом, -O[(CH2)nO]mCH3, -O(CH2)nOCH3, -O(CH2)nNH2, -O(CH2)nCH3, -O(CH2)n-NH2, и -O(CH2)nON[(CH2)nCH3)]2, где n и m составляют от 1 до приблизительно 10; и/или модификации в 5'-положении: 5'-винила, 5'-метила (R или S); модификации в 4'-положении: 4'-S, гетероциклоалкила, гетероциклоалкарила, аминоалкиламиногруппы, полиалкиламиногруппы, замещенного силила, РНК-отщепляющей группы, репортерной группы, интеркалятора, группы для улучшения фармакокинетических свойств олигонуклеотида или группы для улучшения фармакодинамических свойств олигонуклеотида, и любой их комбинации. В некоторых вариантах осуществления по меньшей мере одно неприродное основание распознается ДНК-полимеразой. В некоторых вариантах осуществления по меньшей мере одно неприродное основание распознается РНК-полимеразой.
. В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, выбранное из и . В некоторых вариантах осуществления ДНК содержит по меньшей мере два неприродных нуклеиновых основания, выбранных из и . В некоторых вариантах осуществления ДНК содержит две цепи, при этом первая цепь содержит по меньшей мере одно нуклеиновое основание, являющееся , и вторая цепь содержащий по меньшей мере одно нуклеиновое основание, являющееся . В некоторых вариантах осуществления ДНК содержит по меньшей мере одно неприродное нуклеиновое основание, являющееся . В некоторых вариантах осуществления организм экспрессирует транспортер нуклеозидтрифосфата. В некоторых вариантах осуществления организм экспрессирует транспортер нуклеозидтрифосфата является PtNTT2. В некоторых вариантах осуществления организм дополнительно экспрессирует тРНК-синтетазу. В некоторых вариантах осуществления тРНК-синтетаза является пирролизил-тРНК-синтетазой M. barkeri (Mb PylRS). В некоторых вариантах осуществления организм экспрессирует транспортер нуклеозидтрифосфата PtNTT2 и дополнительно экспрессирует тРНК-синтетазу пирролизил-тРНК-синтетазу M. barkeri (Mb PylRS). В некоторых вариантах осуществления организм дополнительно экспрессирует РНК-полимеразу. В некоторых вариантах осуществления РНК-полимераза является RNAP T7. В некоторых вариантах осуществления организм не экспрессирует белок, имеющий функцию рекомбинационной репарации ДНК. В некоторых вариантах осуществления организм является E coli и организм не экспрессирует RecA. В некоторых вариантах осуществления организм гиперэкспрессирует ДНК-полимеразу. В некоторых вариантах осуществления организм гиперэкспрессирует ДНК-полимеразу II. В некоторых вариантах осуществления полусинтетический организм дополнительно содержит мРНК. В некоторых вариантах осуществления мРНК содержит по меньшей мере одно неприродное основание, выбранное из , и . В некоторых вариантах осуществления мРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления мРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления мРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления полусинтетический организм дополнительно содержит тРНК. В некоторых вариантах осуществления тРНК содержит по меньшей мере одно неприродное основание, выбранное из , и . В некоторых вариантах осуществления тРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления тРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления тРНК содержит по меньшей мере одно неприродное основание, являющееся . В некоторых вариантах осуществления полусинтетический организм дополнительно содержит мРНК и/или тРНК. В некоторых вариантах осуществления организм дополнительно содержит (a) транспортер нуклеозидтрифосфата, (b) мРНК, (c) тРНК, (d) тРНК-синтетазу и (e) РНК-полимеразу, и где организм не экспрессирует белок, имеющий функцию рекомбинационной репарации ДНК. В некоторых вариантах осуществления транспортер нуклеозидтрифосфата является PtNTT2, тРНК-синтетаза является пирролизил-тРНК-синтетазой M. barkeri (Mb PylRS), и РНК-полимераза является RNAP T7. В некоторых вариантах осуществления организм является E coli, и организм не экспрессирует RecA. В некоторых вариантах осуществления организм гиперэкспрессирует одну или более ДНК-полимераз. В некоторых вариантах осуществления организм гиперэкспрессирует одну ДНК-полимеразу II. В некоторых вариантах осуществления по меньшей мере одно неприродное основание дополнительно содержит неприродный сахарный остаток. В некоторых вариантах осуществления неприродный сахарный остаток выбран из группы, состоящей из: модификации в 2'-положении: OH, замещенного низшего алкила, алкарила, аралкила, O-алкарила или O-аралкила, SH, SCH3, OCN, Cl, Br, CN, CF3, OCF3, SOCH3, SO2CH3, ONO2, NO2, N3, NH2F; O-алкила, S-алкила, N-алкила; O-алкенила, S-алкенила, N-алкенила; O-алкинила, S-алкинила, N-алкинила; O-алкил-O-алкила, 2'-F, 2'-OCH3, 2'-O(CH2)2OCH3, где алкил, алкенил и алкинил могут являться замещенным или незамещенным C1-C10-алкилом, C2-C10-алкенилом, C2-C10-алкинилом, -O[(CH2)nO]mCH3, -O(CH2)nOCH3, -O(CH2)nNH2, -O(CH2)nCH3, -O(CH2)n-NH2, и -O(CH2)nON[(CH2)nCH3)]2, где n и m составляют от 1 до приблизительно 10; и/или модификации в 5'-положении: 5'-винила, 5'-метила (R или S); модификации в 4'-положении: 4'-S, гетероциклоалкила, гетероциклоалкарила, аминоалкиламиногруппы, полиалкиламиногруппы, замещенного силила, РНК-отщепляющей группы, репортерной группы, интеркалятора, группы для улучшения фармакокинетических свойств олигонуклеотида или группы для улучшения фармакодинамических свойств олигонуклеотида, и любой их комбинации. В некоторых вариантах осуществления по меньшей мере одно неприродное основание распознается ДНК-полимеразой. В некоторых вариантах осуществления по меньшей мере одно неприродное основание распознается РНК-полимеразой.
[00635] Настоящее изобретение относится к композициям, содержащим аналог нуклеинового основания структуры: 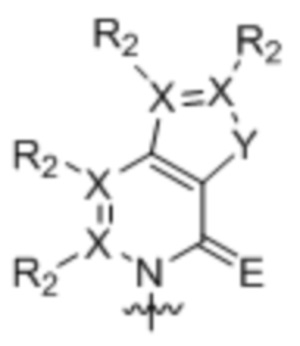 ,где каждый X независимо представляет собой углерод или азот; R2 является необязательным и, если присутствует, независимо представляет собой водород, алкил, алкенил, алкинил, метоксигруппу, метантиол, метанселеногруппу, галоген, цианогруппу или азидную группу; где каждый Y независимо представляет собой серу, кислород, селен или вторичный амин; где каждый E независимо представляет собой кислород, серу или селен; и где волнистой линией указана точка связывания с рибозильным, дезоксирибозильным или дидезоксирибозильным остатком или его аналогом, где рибозильный, дезоксирибозильный или дидезоксирибозильный остаток или его аналог находится в свободной форме, соединенной с монофосфатной, дифосфатной или трифосфатной группой, необязательно, содержащей α-тиотрифосфатную, β-тиотрифосфатную или γ-тиотрифосфатную группа, или включен в РНК, или ДНК, или аналог РНК, или аналог ДНК. В некоторых вариантах осуществления рибозильный или дезоксирибозильный остаток несет трифосфатную или α-тиотрифосфатную группу, связанную с его 5'-гидроксилом. В некоторых вариантах осуществления рибозильный или дезоксирибозильный остаток встроен в олигонуклеотидную цепь РНК или ДНК, соответственно, или рибозильный или дезоксирибозильный остаток или его аналог встроен в аналог РНК или ДНК. В некоторых вариантах осуществления X является углеродом. В некоторых вариантах осуществления E является серой. В некоторых вариантах осуществления Y является серой. В некоторых вариантах осуществления нуклеиновое основание содержит структуру
,где каждый X независимо представляет собой углерод или азот; R2 является необязательным и, если присутствует, независимо представляет собой водород, алкил, алкенил, алкинил, метоксигруппу, метантиол, метанселеногруппу, галоген, цианогруппу или азидную группу; где каждый Y независимо представляет собой серу, кислород, селен или вторичный амин; где каждый E независимо представляет собой кислород, серу или селен; и где волнистой линией указана точка связывания с рибозильным, дезоксирибозильным или дидезоксирибозильным остатком или его аналогом, где рибозильный, дезоксирибозильный или дидезоксирибозильный остаток или его аналог находится в свободной форме, соединенной с монофосфатной, дифосфатной или трифосфатной группой, необязательно, содержащей α-тиотрифосфатную, β-тиотрифосфатную или γ-тиотрифосфатную группа, или включен в РНК, или ДНК, или аналог РНК, или аналог ДНК. В некоторых вариантах осуществления рибозильный или дезоксирибозильный остаток несет трифосфатную или α-тиотрифосфатную группу, связанную с его 5'-гидроксилом. В некоторых вариантах осуществления рибозильный или дезоксирибозильный остаток встроен в олигонуклеотидную цепь РНК или ДНК, соответственно, или рибозильный или дезоксирибозильный остаток или его аналог встроен в аналог РНК или ДНК. В некоторых вариантах осуществления X является углеродом. В некоторых вариантах осуществления E является серой. В некоторых вариантах осуществления Y является серой. В некоторых вариантах осуществления нуклеиновое основание содержит структуру 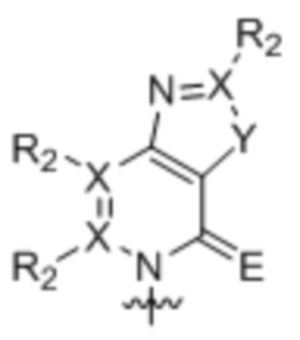 . В некоторых вариантах осуществления нуклеиновое основание содержит структуру . В некоторых вариантах осуществления пары нуклеиновых оснований с комплементарным нуклеиновым основанием образуют неприродную пару оснований (UBP). В некоторых вариантах осуществления комплементарное, спаривающееся нуклеиновое основание выбрано из:
. В некоторых вариантах осуществления нуклеиновое основание содержит структуру . В некоторых вариантах осуществления пары нуклеиновых оснований с комплементарным нуклеиновым основанием образуют неприродную пару оснований (UBP). В некоторых вариантах осуществления комплементарное, спаривающееся нуклеиновое основание выбрано из:
 .
.
[00636] Настоящее изобретение относится к двухцепочечным олигонуклеотидным дуплексам, где первая олигонуклеотидная цепь содержит соединение, представленное в настоящем описании, и вторая, комплементарная олигонуклеотидная цепь содержит комплементарное, спаривающееся нуклеиновое основание в его комплементарном, образующем пару оснований участке. В некоторых вариантах осуществления первая олигонуклеотидная цепь содержит ; и вторая цепь содержит комплементарное, спаривающееся нуклеиновое основание, выбранное из группы, состоящей из:
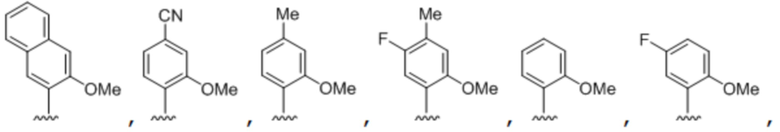
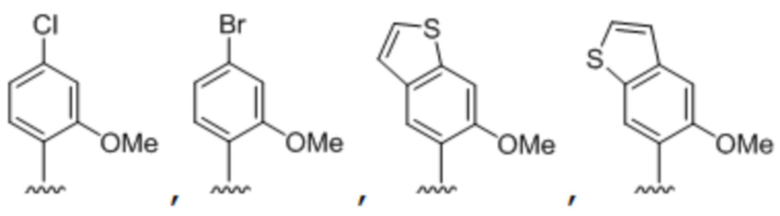 в его комплементарном, образующем пару оснований участке.
в его комплементарном, образующем пару оснований участке.
[00637] Настоящее изобретение относится к транспортной РНК (тРНК), содержащей аналог рибонуклеинового основания, представленного в настоящем описании, содержащий: антикодон, где антикодон содержит аналог рибонуклеинового основания; и элемент распознавания, где элемент распознавания способствует селективной зарядке тРНК неприродной аминокислотой посредством аминоацил-тРНК-синтетазы. В некоторых вариантах осуществления тРНК аналог нуклеинового основания находится в области антикодона тРНК. В некоторых вариантах осуществления тРНК аналог нуклеинового основания находится в первом положении антикодона. В некоторых вариантах осуществления тРНК аналог нуклеинового основания находится во втором положении антикодона. В некоторых вариантах осуществления тРНК аналог нуклеинового основания находится в третьем положении антикодона. В некоторых вариантах осуществления тРНК аминоацил-тРНК-синтетазу получают из термофильного организма. В некоторых вариантах осуществления тРНК аминоацил-тРНК-синтетазу получают из Methanosarcina или его варианта. В некоторых вариантах осуществления тРНК аминоацил-тРНК-синтетазу получают из Methanococcus (Methanocaldococcus) или его варианта. В некоторых вариантах осуществления тРНК неприродная аминокислота содержит ароматический остаток. В некоторых вариантах осуществления тРНК неприродная аминокислота является производным лизина. В некоторых вариантах осуществления тРНК неприродная аминокислота является производным фенилаланина. В некоторых вариантах осуществления тРНК неприродная аминокислота выбрана из группы, состоящей из: N6-азидоэтокси-карбонил-L-лизина (AzK), N6-пропаргилэтокси-карбонил-L-лизина (PraK), BCN-L-лизина, норборнен-лизина, TCO-лизина, метилтетразин-лизина, аллилоксикарбониллизина, 2-амино-8-оксононановой кислоты, 2-амино-8-оксооктановой кислоты, p-ацетил-L-фенилаланина, p-азидометил-L-фенилаланина (pAMF), p-йодо-L-фенилаланина, m-ацетилфенилаланина, 2-амино-8-оксононановой кислоты, p-пропаргилоксифенилаланина, p-пропаргил-фенилаланина, 3-метил-фенилаланина, L-ДОФА, фторированного фенилаланина, изопропил-L-фенилаланина, p-азидо-L-фенилаланина, p-ацил-L-фенилаланина, p-бензоил-L-фенилаланина, p-бромфенилаланина, p-амино-L-фенилаланина, изопропил-L-фенилаланина, O-аллилтирозина, O-метил-L-тирозина, O-4-аллил-L-тирозина, 4-пропил-L-тирозина, фосфонотирозина, три-O-ацетил-GlcNAcp-серина, L-фосфосерина, фосфоносерина, L-3-(2-нафтил)аланина, 2-амино-3-((2-((3-(бензилокси)-3-оксопропил)амино)этил)селанил)пропионовой кислоты, 2-амино-3-(фенилселанил)пропионовой кислоты и селеноцистеина.
[00638] Настоящее изобретение относится к структурам, содержащим формулу: N1 - Zx - N2, где N1 является нуклеотидом, или его аналогом, или концевой фосфатной группой; где N2 является нуклеотидом, или его аналогом, или концевой гидроксильной группой; где Z является соединением, представленным в настоящем описании, и где x является целым числом от 1 до 20. В некоторых вариантах осуществления структура кодирует ген. В некоторых вариантах осуществления Zx находится в кодирующей области гена. В некоторых вариантах осуществления Zx находится в кодоне. В некоторых вариантах осуществления структура дополнительно содержит 5'- или 3'-нетранслируемую область (UTR). В некоторых вариантах осуществления структура дополнительно содержит область терминатора. В некоторых вариантах осуществления структура дополнительно содержит промоторную область.
[00639] Настоящее изобретение относится к библиотекам полинуклеотидов, где библиотеки содержат по меньшей мере 5000 уникальных полинуклеотидов, и где каждый полинуклеотид содержит по меньшей мере одно неприродное нуклеиновое основание, представленное в настоящем описании. В некоторых вариантах осуществления библиотека полинуклеотидов кодирует по меньшей мере один ген.
[00640] Настоящее изобретение относится к нуклеозидтрифосфатам, где нуклеиновое основание выбрано из 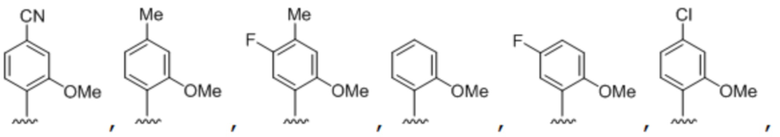
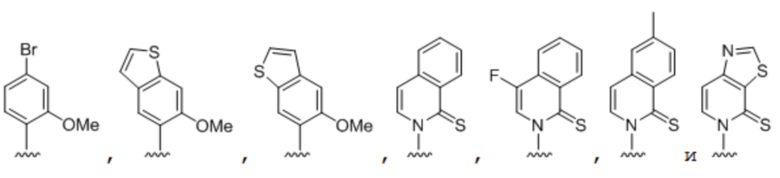 . В некоторых вариантах осуществления нуклеозид содержит рибозу. В некоторых вариантах осуществления нуклеозид содержит дезоксирибозу. В некоторых вариантах осуществления нуклеиновое основание выбрано из
. В некоторых вариантах осуществления нуклеозид содержит рибозу. В некоторых вариантах осуществления нуклеозид содержит дезоксирибозу. В некоторых вариантах осуществления нуклеиновое основание выбрано из 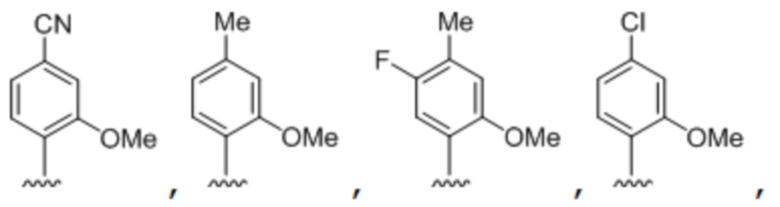
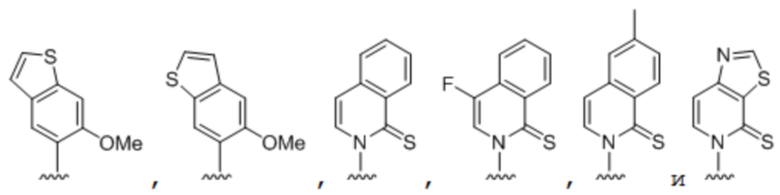 . В некоторых вариантах осуществления нуклеиновое основание выбрано из
. В некоторых вариантах осуществления нуклеиновое основание выбрано из 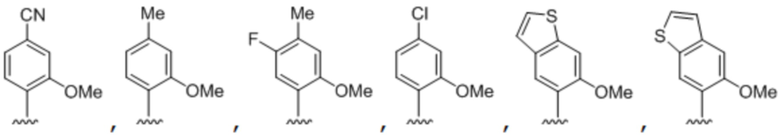 и . В некоторых вариантах осуществления нуклеиновое основание выбрано из
и . В некоторых вариантах осуществления нуклеиновое основание выбрано из 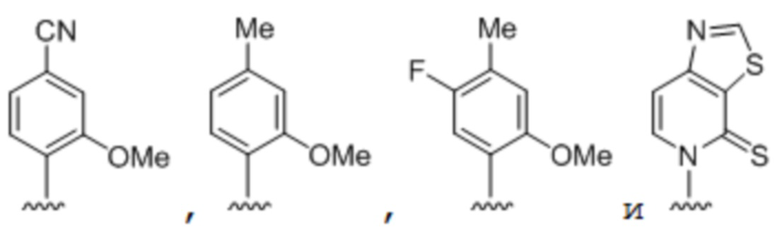 . В некоторых вариантах осуществления нуклеиновое основание выбрано из, и . В некоторых вариантах осуществления нуклеиновое основание представляет собой . В некоторых вариантах осуществления нуклеозид содержит рибозу. В некоторых вариантах осуществления нуклеозид содержит дезоксирибозу. В некоторых вариантах осуществления нуклеиновое основание представляет собой . В некоторых вариантах осуществления нуклеозид содержит рибозу. В некоторых вариантах осуществления нуклеозид содержит дезоксирибозу. В некоторых вариантах осуществления нуклеиновое основание представляет собой . В некоторых вариантах осуществления нуклеозид содержит рибозу. В некоторых вариантах осуществления нуклеозид содержит дезоксирибозу.
. В некоторых вариантах осуществления нуклеиновое основание выбрано из, и . В некоторых вариантах осуществления нуклеиновое основание представляет собой . В некоторых вариантах осуществления нуклеозид содержит рибозу. В некоторых вариантах осуществления нуклеозид содержит дезоксирибозу. В некоторых вариантах осуществления нуклеиновое основание представляет собой . В некоторых вариантах осуществления нуклеозид содержит рибозу. В некоторых вариантах осуществления нуклеозид содержит дезоксирибозу. В некоторых вариантах осуществления нуклеиновое основание представляет собой . В некоторых вариантах осуществления нуклеозид содержит рибозу. В некоторых вариантах осуществления нуклеозид содержит дезоксирибозу.
ПРИМЕРЫ
[00641] Эти примеры приведены исключительно в иллюстративных целях, и не ограничивают объем формулы изобретения, представленной в настоящем описании.
[00642] Пример 1A: Обзор репликации и сборки на матрице при транскрипции.
[00643] Плазмиды конструировали с двумя UBP dNaM-dTPT3, таким образом, что последовательность AXC (здесь и на всем протяжении описании "X" относится к (d)NaM или аналогу (d)NaM) была помещена так, чтобы служить матрицей кодона 151 мРНК sfGFP (sfGFP151(AXC)), и последовательность GYT (здесь и на всем протяжении описании "Y" относится к (d)TPT3 или аналогу (d)TPT3) была помещена так, чтобы служить матрицей антикодона Pyl-тРНК M. mazei (тРНК-Pyl(GYT)), селективно заряжающего ncAAN6-(2-азидоэтокси)-карбонил-L-лизином (AzK) с помощью пирролизил-тРНК-синтетазы M. barkeri (Mb PylRS). Эти плазмиды использовали для трансформации E. coli, экспрессирующих транспортер нуклеозидтрифосфата PtNTT2 (штамм YZ38) и несущих плазмиду, кодирующую Mb PylRS. После трансформации колонии подвергали селекции и выращивали до OD600 ~1,0 в жидких средах, дополненных dTPT3TP (10 мкМ) и одним из семи разных dXTP (фиг.2), добавляемых в разных концентрациях (150 мкМ, 10 мкМ или 5 мкМ). Затем клетки разводили свежей экспрессионной средой, содержащей те же неприродные дезоксирибонуклеотиды, а также NaMTP (250 мкМ), TPT3TP (30 мкМ) и AzK (10 мМ). После кратковременной инкубации экспрессию RNAP T7 и тРНК-Pyl(GYT) индуцировали добавлением изопропил-β-d-тиогалактозида (IPTG, 1 мМ). После еще 1 ч. инкубации экспрессию sfGFP151(AXC) индуцировали добавления ангидротетрациклина (aTc, 100 нг/мл).
[00644] Анализ продуктов ПЦР. Через 2,5 ч. выделяли плазмиды, а затем интересующие гены независимо подвергали ПЦР-амплификации с использованием d5SICSTP и биотинилированного аналога dNaMTP7. Полученный продукт ПЦР анализировали посредством анализа сдвига электрофоретической подвижности с использованием стрептавидина для количественного анализа удерживаемого UBP как сдвига процента общего амплифицированного продукта (далее обозначаемого как анализ сдвига электрофоретической подвижности стрептавидина) (фиг. 3a и фиг. 3b). При наибольшей исследуемой концентрации dXTP (150 мкМ), удержание в гене sfGFP151(AXC) было схожим для каждого аналога dXTP, варьируясь от 99% в случае dNaMTP до 92% в случае dMTMOTP. Удержание в гене тРНК-Pyl(GYT) являлось немного меньшим, в диапазоне от 82% в случае dMTMOTP до 74% в случае dMMO2TP. При добавлении 10 мкМ каждого dXTP удержание в гене sfGFP151(AXC) находилось в диапазоне от 96% в случае d5FMTP до 73% в случае dNaMTP. В случае гена тРНК-Pyl(GYT) удержание снова, в целом, было немного ниже, в диапазоне от 82% в случае d5FMTP и dPTMOTP до 71% в случае dNaMTP. При наименьшей концентрации (5 мкМ) удержание находилось в диапазоне от 94% в случае d5FMTP до 64% в случае dMTMOTP в гене sfGFP151(AXC) и от 83% в случае d5FMTP до 71% в случае dMTMOTP в гене тРНК-Pyl(GYT). В основном, при наибольшей концентрации все dXTP функционировали с практически количественным удержанием UBP в гене sfGFP. Однако, при наименьших концентрациях dCNMO-dTPT3, d5FM-dTPT3 и dPTMO-dTPT3 реплицировались со значимо более высоким удержанием. Удержание в гене тРНК значимо меньше зависело от концентрации dXTP.
[00645] Анализ продукции репортерного белка. Для характеризации количества продуцируемого белка совокупную флуоресценцию культуры, нормализованную по росту клеток, измеряли через 2,5 ч. после индукции экспрессии белка (фиг. 3C). В отсутствие AzK флуоресценция, как правило, была низкой, за исключением dPTMOTP и dMTMOTP в более низких концентрациях, где она казалась немного более высокой. При добавлении AzK в среды клетки, выращенные с каждым dXTP при 150 или 10 мкМ, как правило, демонстрировали значимые и схожие уровни флуоресценции, за исключением dNaMTP, когда наблюдали значимо меньшую флуоресценцию при 10 мкМ, чем 150 мкМ. При наименьшей концентрации (5 мкМ) добавление AzK приводило к меньшей флуоресценции в случае dMTMOTP, в то время как она оставалась той же в случае dCNMOTP, d5FMTP, dClMOTP, dMMO2TP или dPTMOTP. Роста клеток не наблюдали при использовании лишь 5 мкМ dNaMTP. Хотя результаты схожи при высоких концентрациях, при более низких концентрациях использование каждого аналога dXTP приводит к более высокому AzK-зависимому повышению флуоресценции, чем при использовании dNaMTP. В частности, dCNMOTP, d5FMTP и dClMOTP демонстрировали наибольшее AzK-зависимое повышение флуоресценции.
[00646] Анализ точности репортерного белка. Для прямой оценки точности продукции неприродного белка клетки собирали через 2,5 ч. после индукции экспрессии белка, и продуцируемый sfGFP очищали и подвергали промотируемой напряжением реакции циклоприсоединения азид-алкина с дибензоциклооктином (DBCO), связанным с красителем TAMRA с помощью четырех единиц PEG. В дополнение к мечению белков, содержащих ncAA с детектируемым флуорофором, конъюгация приводит к сдвигу электрофоретической миграции, что делает возможным количественный анализ белка, содержащего AzK, в виде процента от общего продуцируемого белка (т.е. точности встраивания ncAA; фиг. 3D). Как и в случае dNaM, использование каждого dXTP при наибольшей концентрации (150 мкМ) приводило к практически полным сдвигам очищенного белка, что отражает высокоточное встраивание ncAA. При выращивании в присутствии 10 мкМ dXTP точность встраивания ncAA остается высокой в случае dCNMOTP, d5FMTP, dClMOTP, dMMO2TP, dPTMOTP и dMTMOTP, но резко снижается в случае dNaMTP. И наконец, при концентрации 5 мкМ dXTP точность встраивания ncAA остается схожей с наблюдаемой при 10 мкМ в случае dCNMOTP, d5FMTP, dClMOTP, dMMO2TP и dPTMOTP, но резко снижается в случае dMTMOTP (и снова из-за эффективности, точность в случае dNaMTP нельзя измерить при этой концентрации).
[00647] Результаты. Неприродными основаниями, приводящими к наибольшему количеству чистого неприродного белка, в частности, при более низких концентрациях, являются dCNMOTP и d5FMTP. Однако, относительно d5FMTP ранее показано, что использование dCNMOTP приводит к более высокому удержанию UBP в более трудных для репликации последовательностях. UBP dCNMO-dTPT3 подходит для хранения и извлечения увеличенной информации. Ранее описанные SSO хранили информацию с помощью UBP dNaM-dTPT3, dPTMO-dTPT3 или dMTMO-dTPT3 и извлекали эту информацию с использованием NaMTP и TPT3TP. Для исследования оптимизации SSO удержание UBP, транскрипцию в мРНК sfGFP и тРНК-Pyl и декодирование на рибосоме изучали с использованием коллекции ранее и недавно описанных аналогов дезокси- и рибонуклеотидтрифосфатов. Исследовали способность хранить информацию с использованием семи разных UBP dX-dTPT3. В каждом случае контекст цепи UBP был одним и тем же, при этом dTPT3 и dX находились в соответствующих антисмысловых (матричных) цепях генов sfGFP и тРНК-Pyl, соответственно. При использовании высоких концентраций каждого dXTP каждая UBP dX-dTPT3 удерживается на высоком уровне в гене мРНК с вариацией от 92% в случае dMTMOTP до 96-99% в случае dCNMOTP, dPTMOTP и dNaMTP. Удержание в гене тРНК являлось, в некоторой степени, сниженным, варьируясь от 74% до 82%. Со снижением концентраций dXTP удержание остается приблизительно постоянным в гене тРНК, но снижалось в зависимости от dXTP в гене мРНК, снижаясь до 64% в случае dMTMOTP, но оставаясь относительно высоким при ~94% в случае dCNMOTP и d5FMTP. Не желая быть связанными какой-либо теорией, полагают, что разные зависимости от концентрации в генах тРНК и мРНК, вероятно, являются результатом эффектов контекста последовательностей, делая встраивание нуклеотида скорость-лимитирующим для мРНК, а продолжающуюся элонгацию - скорость-лимитирующей для гена тРНК. Исключения наблюдали в случае dNaMTP, где при 10 мкМ удержание снижалось до 73%, и клетки не выживали, когда dNaMTP присутствовал в концентрации 5 мкМ. Кроме того, в случае dCNMOTP и d5FMTP удержание оставалось высоким в мРНК (~93%) даже при наименьших исследуемых концентрациях. Не желая быть связанными какой-либо теорией, полагают, что потеря удержания в гене тРНК может приводить к меньшей продукции неприродного белка и, возможно, что более неблагоприятно, сниженной точности встраивания ncAA из-за повышенной конкуренции за декодирование неприродного кодона "практически когнатными" природными тРНК. Однако, точность встраивания ncAA коррелировала с удержанием в гене мРНК (данные не представлены). Таким образом, данные свидетельствуют о том, что каждый dX служит матрицей для транскрипции тРНК с эффективностью и точностью, достаточной, чтобы не ограничивать точность продукции неприродного белка. Учитывая эти результаты, d5FM-dTPT3 и dCNMO-dTPT3 являются UBP, способными к увеличенному хранению информации, при этом их полезность относительно dNaM-dTPT3 является результатом, главным образом, их более высокого удержания и продукции белка при более низких концентрациях неприродного трифосфата.
[00648] Пример 1B: Подробные способы репликации и сборки на матрице при транскрипции.
[00649] Сборка плазмид Golden Gate. Вставки, содержащие UBP (и, при необходимости, гены, кодирующие sfGFP или тРНК-Pyl), получали посредством ПЦР олигонуклеотидов, содержащих неприродные основание, O3 и O6-12 (см. последовательности олигонуклеотидов в таблице S4), синтезированных вручную (см. Синтез олигонуклеотидов) или полученных из Synthorx, с использованием dTPT3TP и dNaMTP и праймеров P1-2 (при амплификации O3 или O7-12) и P3-4 (при амплификации O6) (см. последовательности праймеров в таблице S1) для создания концевых участков распознавания BsaI, в которых после расщепления образуются липкие концы, совместимые с подходящей принимающей плазмидой p[sfGFP(gg)151 тРНК-Pyl(gg)] GG для трансляции sfGFP, последовательность которой показана ниже (SEQ ID NO: 2). ПЦР-амплификацию осуществляли с помощью термоциклера PTC-200 Peltier (MJ Research). Олигонуклеотиды-матрицы (0,025 нг на 50 мкл реакционной смеси) подвергали ПЦР-амплификации с использованием следующих концентраций реагентов: стандартный реакционный буфер OneTaq (1-кратный, New England BioLabs), dNTP (0,2 мМ), dTPT3TP (0,1 мМ), dNaMTP (0,1 мМ), MgSO4 (1,2 мМ), праймеры (по 1 мкМ каждый) и полимераза OneTaq (1,25 ед.). Природные олигонуклеотиды амплифицировали в отсутствие неприродных трифосфатов. Их амплифицировали в следующих условия термоциклирования (время обозначено как мин:сек): [94°C 0:30| 20 раз (94°C 0:30 | 54°C 0:30 | 68°C 4:00)]. Оставшийся раствор очищали с помощью центрифужной колонки (DNA Clean and Concentrator-5; Zymo Research), затем количественно анализировали по поглощению при 280 нм с использованием спектрофотометра для чтения микропланшетов Infinite M200 Pro Multimode (Tecan).
[00650] Синтез олигонуклеотидов. Модифицированные олигонуклеотиды синтезировали в масштабе 0,2 мкмоль с использованием генетического синтезатора Expedite 8909, колонок с сукцинил-связанным LCAA-CPG (длинноцепочечный алкиламин-стекло с контролируемым размером пор) с размером пор 1000 Å и стандартных способов встраивания фосфорамидитов ДНК dABz, dCBz, dGiBu и dT. Для встраивания мономера dNaM использовали следующие условия сочетания (15 мин; тетразол в CH3CN; >95%). Модифицированные фосфорамидиты использовали в 50-кратном молярном избытке и концентрации 0,05 M в безводном CH3CN. Отщепление от твердой подложки и удаление защитных групп осуществляли с использованием ~30% водного раствора аммиака (55°C, 16 ч.). Очистку неочищенных олигонуклеотидов DMT-on осуществляли с использованием системы UHPLC Vanquish, оборудованной колонкой Hypersil Gold C18 (5 мкм, 4,6×150 мм) с использованием 0-50% B за 25 мин (буфер A=0,05 M TEAA, pH 7; буфер B=25% H2O в CH3CN; скорость потока=1,0 мл/мин). Объединяли соответствующие фракции, выпаривали до сухости, а затем осуществляли детритилирование (80% водн. AcOH) и осаждение (NaOAc/NaClO4/ацетон, -80°C в течение 3 ч.). Чистоту проверяли с помощью ион-парной обращенно-фазовой ВЭЖХ, осуществляемой в аналитическом режиме (>90%).
[00651] Плазмиды собирали в объеме реакционной смеси 80 мкл в следующих условиях: принимающая плазмида (1 мкг), вставки для ПЦР (молярное соотношение 4:1 вставка:плазмида), ДНК-лигаза T4 (532 ед.), BsaI-HF (53,4 ед.) и АТФ (1 мМ) комбинировали в буфере CutSmart (1-кратном, New England BioLabs) и подвергали термоциклированию в следующих условиях (время обозначено как мин:сек): [37°C 20:00 | 40 раз (37°C 5:00 | 16°C 10:00 | 22°C 5:00) | 37°C 20:00 | 50°C 15:00 | 70°C 30:00].
После реакции Golden Gate добавляли экзонуклеазу T5 (13,3 ед.) и BsaI-HF (26,6 ед.) и реакционную смесь инкубировали при 37°C в течение 1 ч. для расщепления оставшихся фрагментов ДНК и невстроившейся принимающей плазмиды. Собранные плазмиды очищали с помощью колонок Zymo-Spin I и количественно анализировали с использованием аналитического набора Qubit dsDNA HS (Thermo Fisher Scientific).
[00652] Трансформация YZ3/ML2 с помощью pGEX-MbPylRS TetR. Ночную культуру SSO штамма YZ3 или ML2 выращивали в 2×YT, дополненной 50 мМ фосфатом калия и 5 мкг/мл хлорамфеникола (в настоящем описании обозначаемой в этом разделе как "среда"), затем разводили обратно до OD600 0,03 той же средой и выращивали до OD600 0,4-0,6. Затем культуру охлаждали при встряхивании на ледяной бане в течение 5 мин, затем осаждали посредством центрифугирования при 3200×g в течение 10 мин. Затем клетки ресуспендировали и промывали ледяным ddH2O дважды, затем ресуспендировали в ледяном ddH2O до OD600 50-60. Электрокомпетентные клетки (50 мкл) и 2 нг плазмиды pGEX-MbPylRS TetR (SEQ ID NO: 1) переносили в предварительно охлажденную кювету для электропорации (щель 0,2 см), а затем осуществляли электропорацию (Gene Pulser II; Bio-Rad) по инструкциям производителя (напряжение 25 кВ, конденсатор 2,5 мкФ, резистор 200 Ом), затем незамедлительно разбавляли 950 мкл среды. Затем аликвоту этой реакционной смеси для электропорации (40 мкл) разводили в пять раз средой до конечного объема 200 мкл, а затем позволяли восстанавливаться при 37°C в течение 1 ч. Двукратные разведения восстановленных клеток высевали на твердую среду 2×YT, дополненную 50 мМ фосфата калия, 5 мкг/мл хлорамфеникола, 100 мкг/мл карбенициллина и 2% масс./об. агара, а затем позволяли им расти при 37°C в течение ночи. Отбирали отдельные колонии для инокуляции культур в жидких средах, дополненных 100 мкг/мл карбенициллина, а затем хранили в глицерине (25% об./об.) при - 80°C.
[00653] Скрининг трансляции in vivo dXTP. Ночную культуру SSO штамма YZ3, несущего плазмиду pGEX-MbPylRS TetR, выращивали в 2×YT, дополненной 50 мМ фосфата калия, 5 мкг/мл хлорамфеникола и 100 мкг/мл карбенициллина (в настоящем описании обозначаемой в этом разделе как "среда"), затем разводили обратно до OD600 0,03 в той же среде и выращивали до OD600 0,4-0,6. Затем культуру охлаждали при встряхивании на ледяной бане в течение 15 минут, затем осаждали посредством центрифугирования при 3200×g в течение 10 мин. Затем клетки ресуспендировали и промывали ледяной ddH2O дважды, затем ресуспендировали в ледяной ddH2O до OD600 55-65. Электрокомпетентные клетки (50 мкл) и 1 нг собранной плазмиды Golden Gate с UBP в генах sfGFP и тРНК-Pyl (см. Сборка плазмид Golden Gate) переносили в предварительно охлажденную кювету для электропорации (щель 0,2 см), затем осуществляли электропорацию (Gene Pulser II; Bio-Rad) по инструкциям производителя (напряжение 25 кВ, конденсатор 2,5 мкФ, резистор 200 Ом), и затем незамедлительно разводили 950 мкл среды. Затем аликвоту (40 мкл) смеси для трансформации разводили в пять раз в среде, дополненной dNaMTP (150 мкМ) и dTPT3TP (10 мкМ), до конечного объема 200 мкл, а затем позволяли ей восстанавливаться в течение 1 ч. при встряхивании при 37°C. Двукратные разведения восстановленной смеси высевали на твердые среды, дополненные зеоцином (50 мкг/мл), dNaMTP (150 мкМ), dTPT3TP (10 мкМ) и агаром (2% масс./об.), затем выращивали при 37°C в течение ночи. Отбирали отдельные колонии и выращивали в среде (300 мкл), дополненной 50 мкг/мл зеоцина (в настоящем описании далее обозначаемой в этом разделе как "среда для выращивания") и добавляли dNaMTP (150 мкМ) и dTPT3TP (10 мкМ). Культуру подвергали мониторингу роста клеток (многоканальный спектрофотометр для чтения планшетов Envision 2103 с фильтром 590/20 нм), а затем собирали при OD600 ~1,0. Аликвоту (50 мкл) подвергали выделению плазмиды с использованием набора ZR Plasmide Prep Kit (Zymo Research). Затем выделенные плазмиды подвергали анализу сдвига электрофоретической подвижности с использованием стрептавидина, описанному ниже (с использованием праймеров P3-4 и P5-6, таблица S1), для определения удержания UBP. Затем колонии с исключительным удержанием UBP хранили в глицерине (25% об./об.) при -80°C.
[00654] Десятикратные разведения глицеринового стока снова высевали на твердые среды для выращивания, дополненные dNaMTP (150 мкМ), dTPT3TP (10 мкМ) и 2% масс./об. агара, затем выращивали при 37°C в течение ночи. Отбирали отдельные колонии и выращивали в средах для выращивания (300 мкл), дополненных соответствующим dXTP (различные концентрации) и dTPT3TP (10 мкМ), затем осуществляли мониторинг роста клеток и собирали их при OD600 ~1,0. После сбора всех образцов, их оставляли на льду на ночь. Затем образцы повторно разводили до OD600 ~0,1 до конечного объема 300 мкл и добавляли соответствующий dXTP (различные концентрации) и dTPT3TP (10 мкМ). Оставшуюся неиспользуемую культуру осаждали и хранили при -80°C для последующего выделения плазмиды с использованием набора ZR Plasmide Prep Kit (Zymo Research) для определения удержания UBP. Когда культуры достигали OD600 ~0,4-0,6, в них добавляли NaMTP, (250 мкМ) TPT3TP (30 мкМ) и AzK (10 мМ) (или ddH2O в случае культур, выращенных в отсутствие AzK). После добавления AzK образцы закрывали от света для предотвращения фотодеградации. Затем эти образцы выращивали при 37°C в течение 20 мин перед добавлением 1 мМ IPTG и выращивали при 37°C еще в течение 1 ч. для индуцирования RNAP T7 и транскрипции тРНК-Pyl и PylRS. С этого момента клетки подвергали мониторингу роста и флуоресценции. Затем экспрессию sfGFP индуцировали ангидротетрациклином (100 нг/мл). После дополнительных 2,5 ч. выращивания при 37°C клетки собирали (50 мкл для выделения плазмиды для определения удержания UBP, 230 мкл для аффинной очистки sfGFP), охлаждали, а затем осаждали и хранили при -80°C перед оценкой удержания UBP и очистки белка.
[00655] Скрининг трансляции XTP/YTP in vivo. Ночную культуру SSO штамма ML2, несущего плазмиду pGEX-MbPylRS TetR, выращивали в 2×YT, дополненной 50 мМ фосфата калия, 5 мкг/мл хлорамфеникола и 100 мкг/мл карбенициллина (в настоящем описании обозначаемой в этом разделе как "среда"), затем разводили обратно до OD600 0,03 в той же среде и выращивали до OD600 0,4-0,6. Затем культуру охлаждали при встряхивании на ледяной бане в течение 15 минут, затем осаждали посредством центрифугирования при 3200×g в течение 10 мин. Затем клетки ресуспендировали и промывали ледяной ddH2O дважды, затем ресуспендировали ледяной ddH2O до OD600 55-65. Электрокомпетентные клетки (50 мкл) и 1 нг собранной плазмиды Golden Gate с UBP в генах sfGFP и тРНК-Pyl (см. Сборка плазмид Golden Gate) переносили в предварительно охлажденную кювету для электропорации (щель 0,2 см), затем осуществляли электропорацию (Gene Pulser II; Bio-Rad) по инструкциям производителя (напряжение 25 кВ, конденсатор 2,5 мкФ, резистор 200 Ом), а затем незамедлительно разводили 950 мкл среды. Затем аликвоту (40 мкл) смеси для трансформации разводили в пять раз в среде, дополненной dCNMOTP (25 мкМ) и dTPT3TP (10 мкМ) до конечного объема 200 мкл, затем ей позволяли восстановиться в течение 1 ч. при встряхивании и 37°C. Двукратные разведения восстановленной смеси высевали на твердые среды, дополненные зеоцином (50 мкг/мл), dCNMOTP (10 мкМ), dTPT3TP (10 мкМ) и агаром (2% масс./об.), затем выращивали при 37°C в течение 18 ч. Отбирали отдельные колонии и выращивали в среде (300 мкл), дополненной 50 мкг/мл зеоцина, и добавляли dCNMOTP (25 мкМ) и dTPT3TP (10 мкМ) (в настоящем описании далее обозначаемой в этом разделе как "среды для выращивания"). Культуру подвергали мониторингу роста клеток (многоканальный спектрофотометр для чтения планшетов Envision 2103 с фильтром 590/20 нм), а затем собирали при OD600 ~1,0. Аликвоту (50 мкл) подвергали выделению плазмиды с использованием набора ZR Plasmide Prep Kit (Zymo Research). Затем выделенные плазмиды подвергали анализу сдвига электрофоретической подвижности с использованием стрептавидина, описанному ниже (с использованием праймеров P3-4 и P5-6, таблица S1) для определения удержания UBP.
[00656] Затем колонию с исключительным удержанием UBP разводили до OD600 ~0,01 в средах для выращивания и выращивали до OD600 ~0,4-0,6. Затем эту культуру разделяли на множество культур по 300 мкл и для скрининга аналогов XTP, каждый из которых добавляли с соответствующим XTP (в количестве 25 мкМ или 250 мкМ), или ddH2O для образцов "-XTP", TPT3TP (30 мкМ) и AzK (10 мМ). Для скрининга аналогов YTP в каждые 300 мкл культуры добавляли соответствующий YTP (в количестве 25 мкМ или 250 мкМ) или ddH2O для образцов "-YTP", NaMTP (250 мкМ) и AzK (10 мМ). После добавления AzK образцы закрывали от света для предотвращения фотодеградации. Затем эти образцы выращивали при 37°C в течение 20 мин перед добавлением 1 мМ IPTG и выращивали при 37°C еще в течение 1 ч. для индуцирования RNAP T7 и транскрипции тРНК-Pyl и PylRS. С этого момента клетки подвергали мониторингу роста и флуоресценции. Затем экспрессию sfGFP индуцировали ангидротетрациклином (100 нг/мл). Еще через 3 ч. выращивания при 37°C клетки собирали (230 мкл для аффинной очистки sfGFP), охлаждали, а затем осаждали и хранили при -80°C перед очисткой белка.
[00657] Анализ сдвига электрофоретической подвижности с использованием стрептавидина. ПЦР-амплификацию осуществляли с помощью системы CFX Connect Real-Time PCR Detection System (Bio-Rad). Минипрепараты плазмид или собранные плазмиды Golden Gate (от 0,5 мкл до 2 мкл, от 0,5 нг/мкл до 5 нг/мкл) или dNaM-содержащие олигонуклеотиды (0,025 нг) подвергали ПЦР-амплификации в общих объемах реакционной смеси 15 мкл в следующих условиях: стандартный реакционный буфер OneTaq (1-кратный, New England BioLabs), dNTP (400 мкМ), SYBR Green I (1-кратный, Life Technologies), MgSO4 (2,2 мМ), праймеры P3-4 или P5-6 (по 1 мкМ каждый, см. последовательности праймеров в таблице S4), d5SICSTP (65 мкМ), dMMO2BIOTP (65 мкМ; структура показана ниже, при этом сахар и фосфат опущены для простоты), ДНК-полимераза OneTaq (0,27 ед., New England BioLabs), ДНК-полимераза DeepVent (0,105 ед., New England BioLabs).
 dMMO2BIOTP
dMMO2BIOTP
Плазмиды, выделенные во время экспериментов по трансляции, амплифицировали с использованием следующих условий термоциклирования (время обозначено как мин:сек): [96°C 5:00 | 20 раз (95°C 00:15 | 54°C 00:15 | 68°C 4:00]. Затем стрептавидин (2,5 мкл, 2 мкг/мкл; Promega) смешивали с каждой реакционной смесью (1 мкл) и инкубировали в течение 5 мин при комнатной температуре. Каждый из образцов со стрептавидином (3,5 мкл) и без него (1 мкл) смешивали с нагрузочным буфером и разделяли на 6% (масс./об.) полиакриламидном геле (29:1 акриламид:бис-акриламид)Трис/борат/ЭДТА (TBE) при 120 В в течение ~30 мин. Затем гели окрашивали красителем 1-кратным SYBR Gold (Thermo Fisher) и визуализировали с использованием системы Molecular Imager Gel Doc XR+ (Bio-Rad), оборудованной фильтром 520DF30 (Bio-Rad). Затем удержание UBP можно определять посредством сравнения соотношения полос со сдвигом и без сдвига в каждой дорожке относительно результатов, полученных для входной плазмиды (плазмиды, используемой в начальной трансформации). Этот анализ ранее определен как количественный.
[00658] Анализ сдвига электрофоретической подвижности с использованием стрептавидина чувствителен к различным факторам, таким как чистота минипрепаратов, и его можно повторять множество раз, в некоторых случаях с большей концентрацией плазмид, для достижения приемлемой амплификации UBP-содержащей плазмиды для детекции удержания UBP. В случаях, когда концентрация плазмиды является слишком низкой, удержание UBP кажется более низким в соответствии с анализом сдвига электрофоретической подвижности с использованием стрептавидина, его повторяли с использованием большего количества плазмиды минипрепарата.
n.d.a Значение нельзя определить
[00659] Аффинная очистка sfGFP. Клеточные осадки, собранные во время экспериментов по трансляции, ресуспендировали в BugBuster (100 мкл 1-кратного, EMD Millipore) и встряхивали при комнатной температуре в течение 15 мин. Затем лизат клеток разводили буфером W (380 мкл; 50 мМ HEPES, pH 8, 150 мМ NaCl, 1 мМ ЭДТА) и добавляли магнитные бусы Strep-Tactin (20 мкл, суспензию 5% (об./об.) бус MagStrep "тип 3" XT, IBA Lifesciences), уравновешенных в буфере W. Смесь осторожно переворачивали при 4°C в течение 30 мин. Затем бусы притягивали с помощью магнитной подставки и промывали буфером W (2 × 500 мкл), а затем добавляли 25 мкл буфера BXT (100 мМ Трис-HCl, pH 8, 50 мМ D-биотина), и суспензии позволяли инкубироваться при комнатной температуре в течение 10 мин с периодическим перемешиванием. Бусы снова притягивали для сбора элюентов. Очищенные белки количественно анализировали с использованием набора Qubit Protein Assay Kit (Thermo Fisher Scientific).
[00660] Безмедная клик-конъюгация TAMRA. Очищенный белок (300 нг) смешивали с TAMRA-DBCO (0,1 мМ, Click Chemistry Tools; продукт №A131), добавляли воду до конечного объема реакционной смеси 6 мкл, инкубировали в течение ночи при комнатной температуре и закрывали от света. После конъюгации предпринимали меры для минимизации воздействия света на образцы, гели и блоты для минимизации фотообесцвечивания TAMRA.
[00661] Вестерн-блоттинг sfGFP. Очищенный белок (300 нг), конъюгированный с TAMRA-DBCO, смешивали (2:1 об.:об.) с нагрузочным буфером/красителем (250 мМ Трис-HCl, 30% (об./об.) глицерин, 2% (масс./об.) SDS, pH 6,8), затем нагревали при 95°C в течение 5 мин. Белковый маркер молекулярной массы (Color Prestained Protein Standard, Broad Range, New England BioLabs) и часть каждой реакционной смеси для клик-реакции (~ 60 нг белка) наносили на гель ПААГ с SDS (концентрирующий гель: 5% (масс./об.) акриламид:бис-акриламид 29:1 (Fisher), 0,125 M Трис-HCl и 0,1% SDS, pH 6,8 (ProtoGel Stacking Buffer, National Diagnostics); разделяющий гель: 15% (масс./об.) акриламид:бис-акриламид 29:1 (Fisher), 0,375 M Трис-HCl и 0,1% SDS, pH 8,8 (ProtoGel Resolving Buffer, National Diagnostics)), а затем прогоняли при 50 В в течение 15 мин, затем при 120 В еще в течение ~4,5 ч. в буфере для ПААГ с SDS (25 мМ Трис-основания, 200 мМ глицина, 0,1% (масс./об.) SDS). Затем образцы переносили на PVDF с низкой флуоресценцией (0,2 мкМ, Bio-Rad) посредством полусухого переноса в соответствующем буфере (20% (об./об.) MeOH, 50 мМ Трис-основания, 400 мМ глицина, 0,0373% (масс./об.) SDS), прогоняли при 15 В в течение ~30-45 мин. Затем мембрану встряхивали в течение ночи при 4°C в блокирующем растворе (5% (масс./об.) обезжиренного молока (Carnation) в PBS-T (PBS, pH 7,4, 0,01% (об./об.) Tween-20), солюбилизировали при 37°C в течение 1 ч). Затем мембрану промывали 2 раза PBS-T, затем встряхивали с антителом кролика против GFP (1:3000 в PBS-T, продукт №G1544, лот № 046M4871V, SigmaAldrich) в течение 1 ч. при комнатной температуре. Затем мембрану промывали в течение 5 мин PBS-T, а затем встряхивали с AlexaFluor 647-конъюгированным антителом козы против кролика (1:20000 в PBS-T, продукт №A32733, лот №SD250298, Thermo Fisher Scientific) при комнатной температуре в течение 45 мин. Затем мембрану промывали (3 раза по 5 мин) PBS-T и визуализировали с помощью лазерного сканера плоскостного типа (Typhoon 9410, GE Amersham Biosciences) со следующими настройками: разрешение 50 мкМ, возбуждение лазера 532 нм и эмиссионный фильтр 580/30 нм с 400 В PMT для TAMRA, и возбуждение лазера 633 нм и эмиссионный фильтр 670/30 нм с 500 В PMT для AlexaFluor 647.
[00662] Для оценки встраивания AzK очищенный белок конъюгировали с DBCO-TAMRA для получения электрофоретического сдвига с помощью ПААГ с SDS. Разные размеры полос соответствуют нативному sfGFP и sfGFP, конъюгированному с TAMRA. Затем эти полосы количественно анализировали с использованием программного обеспечения Image Studio Lite 5.2.5 (LI-COR Biosciences) посредством денситометрического количественного анализа полос для получения соотношения сдвинутой (конъюгированный) полосы и общего сигнала, образованного сдвинутыми (конъюгированный) и несдвинутыми (нативный) полосами sfGFP. Это можно интерпретировать как процент белкового сдвига или процент sfGFP в образце, в который встроен ncAA AzK.
[00663] Количественная масс-спектрометрия высокого разрешения интактных белков. Получение образцов и получение данных. Очищенный белок (4 мкг) разводили 450 мкл воды для масс-спектрометрии и наносили на устройство с центрифужным фильтром (Amicon Ultra-0,5 - Millipore) для обессоливания. Устройство центрифугировали при 14000×g в течение 10 мин и полученный концентрированный раствор разводили водой для масс-спектрометрии обратно до общего объема 500 мкл. Это действие повторяли еще для четырех центрифугирований, при этом последнее центрифугирование осуществляли в течение 18 минут. После последнего центрифугирования образцы выделяли, переворачивая фильтровальное устройство в новую микроцентрифужную пробирку и центрифугируя при 1000×g в течение 2 мин. Затем обессоленный белок инъецировали (6 мкл, ~250 нг) в Waters I-Class LC, соединенный с Waters G2-XS TOF. Условия потока представляли собой 0,4 мл/мин 50:50 вода:ацетонитрил с 0,1% муравьиной кислотой. Ионизацию осуществляли посредством ESI+, при этом данные собирали при m/z 500-m/z 2000. Комбинирование спектров осуществляли в основной части пика и объединенный спектр подвергали деконволюции с использованием Waters MaxEnt1.
[00664] Валидационный количественный анализ. Аутентичные образцы белков, в которых положение 151 sfGFP содержало остаток тирозина или остаток AzK, получали с использованием идентичных условий получения плазмиды и экспрессии белка, как описано выше, но с использованием sfGFP151(TAC) и sfGFP151(TAG), соответственно, и без дополнения неприродными трифосфатами. Используя эти аутентичные белка, получали смеси Y151 и AzK151 sfGFP в определенных соотношениях, находящихся в диапазоне от 100:0 до 0:100 AzK и Y. Затем эти смеси получали и количественно анализировали посредством HRMS, как описано выше. Наблюдаемые данные являлись линейными на всем протяжении диапазона образцов и точно совпадали с ожидаемыми соотношениями, что позволяет валидировать способ как высоко количественный.
[00665] Исследование токсичности рибонуклеозидтрифосфатов. Ночную культуру ML2 выращивали в среде 2×YT, дополненной 50 мМ фосфата калия и хлорамфениколом (5 мкг/мл), обозначаемой на всем протяжении этого раздела как "среда". Эту культуру разводили обратно до OD600 0,01, разделяли на множество культур по 300 мкл, каждая из которых дополнена различными концентрациями TPT3TP, TAT1TP, NaMTP или 5FMTP, вместе с контролем без трифосфата. Затем культуры инкубировали при 37°C с встряхиванием при 230 об./мин. Затем клетки подвергали мониторингу роста клеток (многоканальный спектрофотометр для чтения планшетов Envision 2103 с фильтром 590/20 нм) каждый час всего в течение восьми часов. Затем строили график OD600 как функции времени для каждого нуклеотида при каждой концентрации для визуализации различий скорости роста.
[00666] Пример 2: Синтез и первый анализ SAR рибонуклеотидов-кандидатов.
[00667] Извлечение информации, доступное с помощью UBP, ранее исследовали только с использованием NaMTP и TPT3TP. Для начала выявления SAR, регулирующих эффективную транскрипцию и трансляцию в SSO, создавали и синтезировали девять новых аналогов NaMTP (фиг. 4A) и четыре новых аналога TPT3TP (фиг. 4B). Эти аналоги создавали для исследования роли формы нуклеинового основания, площади поверхности ароматических групп и дериватизации гетероатома. Как правило, синтез аналогов XTP осуществляли посредством литирования соответствующего арилгалида с последующим сочетанием литированных молекул с бензил- или TBS-защищенным риболактоном. Восстановление полученного полуацеталевого производного в присутствии диэтилэфирата трифторида бора и триэтилсилана, получая желаемый защищенный нуклеозид в каждом случае. После снятия защитной группы полученные аналоги нуклеозида X превращали в трифосфаты с использованием стандартных условий фосфорилирования Людвига. NaMTP и MMO2TP синтезировали, как описано ранее. Синтез аналогов YTP, как правило, осуществляли посредством внутримолекулярной перегруппировки Курциуса соответствующего ацилазида с последующим опосредованным кислотами Льюиса сочетанием с 1-O-ацетил-2,3,5-три-O-бензоил-b-D-рибофуранозой, получая чистый β-аномер желаемого защищенного нуклеозида. После превращения пиридина в соответствующий тио-пиридон и последующего снятия бензоильной защитной группы, соответствующие свободные нуклеозиды превращали в трифосфаты с использованием стандартных условий фосфорилирования Людвига. 5SICSTP синтезировали, как описано ранее.
[00668] Анализ SAR начинали с NaMTP и девяти аналогов XTP. Учитывая их параметры при скрининге dXTP, описанном выше, и для устранения переменных потерь на уровне ДНК в качестве осложняющего фактора, неприродную информацию кодировали с помощью UBP dCNMO-dTPT3. Кроме того, использовали E. coli штамма ML2, экспрессирующие транспортер нуклеозидтрифосфата PtNTT2, но также генетически сконструированные для репликации UBP с более высокой точностью посредством делеции гена, кодирующего RecA (что распространено в клонирующих штаммах), и гиперэкспрессии ДНК-полимеразы II. Использовали ту же плазмиду, описанную выше в примерах 1A и 1B, несущую sfGFP151(AXC) и тРНК-Pyl(GYT), и для фокусирования скрининга на отдельном неприродном рибонуклеотиде, тРНК Pyl M. mazei транскрибировали в присутствии 30 мкМ TPT3TP. Клетки выращивали и индуцировали для продукции белка, как описано выше, за исключением того, что различные XTP добавляли в высокой (250 мкМ) или низкой (25 мкМ) концентрации в экспрессионной среде (фиг. 5A-5D). Экспрессируемый sfGFP очищали через 3 ч. после индуцирования и анализировали содержание ncAA с использованием DBCO-опосредованного анализа электрофоретического сдвига, описанного выше (фиг. 5B). Использование каждого XTP в концентрации 250 мкМ приводило к электрофоретическому сдвигу sfGFP по меньшей мере 63%. Вместе с отсутствием наблюдаемого сдвига в отсутствие XTP, XTP импортируется с помощью PtNTT2 и участвует в транскрипции и трансляции на рибосоме с по меньшей мере приемлемой эффективностью. Хотя каждый из CNMOTP и 5F2OMeTP хорошо функционировал, приводя к электрофоретическим сдвигам 92% и 94%, соответственно, NaMTP, MMO2TP и 5FMTP действовали лучше остальных, при этом электрофоретические сдвиги белков составляли 98%, 97% и 98%, соответственно. При аналогично высоких уровнях чистоты белка, относительную флуоресценцию, получаемую с использованием этих трех XTP, сравнивали в отсутствие любых затруднений, являющихся результатом значительных уровней загрязнения природным sfGFP. Клетки, выращенные с 250 мкМ MMO2TP или 5FMTP, демонстрировали 63% и 90% совокупной флуоресценции, соответственно, по сравнению с клетками, выращенными с NaMTP в той же концентрации (фиг. 5A).
[00669] При более низкой тестируемой концентрации (25 мкМ) использование NaMTP приводило к более низкой точности встраивания ncAA с белковым сдвигом, снижающимся до 86%. Хотя 7 из 9 аналогов NaMTP также демонстрировали значительное снижение точности, использование каждого из MMO2TP и 5FMTP приводило к белковым сдвигам 94%. При сравнении относительной флуоресценции клеток, выращенных с этими рибонуклеотидами (что также возможно из-за схожей и высокой точности продуцируемого неприродного белка), 5FMTP приводил к на 34% большей флуоресценции, чем MMO2TP при этой более низкой концентрации. Проводили схожие эксперименты, где обеспечивали постоянное количество NaMTP (250 мкМ) и высокую (250 мкМ) или низкую (25 мкМ) концентрацию аналога YTP. В соответствии с полученными ранее данными, добавление TPT3TP в более высокой концентрации приводило к значимо сниженному росту клеток и низкой флуоресценции относительно контрольного образца, в который не добавляли YTP (фиг. 5C). В отличие от этого, каждый из других аналогов YTP демонстрировал флуоресценцию выше фоновой и белковые сдвиги по меньшей мере 51% (фиг. 5D). Использование TAT1TP приводило к по меньшей мере в 2,6 раз большей флуоресценции, чем любой другой YTP, при сохранении белкового сдвига 96%. Добавление TAT1TP в этой концентрации приводило к умеренному уровню сниженного роста клеток (данные не представлены). TPT3TP является, в некоторой степени, менее токсичным при использовании в меньших концентрациях, и соответственно, при добавлении в концентрации 25 мкМ клетки продуцируют значительные количества чистого белка. В этих условиях TPT3TP приводил к в 2, 5 и 6 раз более высокой флуоресценции, чем SICSTP, FSICSTP и 5SICSTP, соответственно. Токсичность, наблюдаемая в случае TAT1TP при более высоких концентрациях, была почти полностью устранена при более низких концентрациях (данные не представлены), и его использование приводило к большей флуоресценции, чем при более высокой концентрации. При добавлении в концентрации 25 мкМ, TAT1TP приводит к на 41% большей флуоресценции, чем в концентрации 250 мкМ, и примечательно, он приводит к на 57% большей флуоресценции, чем TPT3TP в концентрации 25 мкМ. Использование TAT1TP в этих концентрациях приводило к продукции белка с встраиванием ncAA 98%.
[00670] Способы синтеза.
[00671] Общие материалы и способы. Для способов синтеза все реакции проводили в высушенной в печи стеклянной посуде в инертной атмосфере. Растворители дистиллировали и/или сушили над молекулярными ситами 4 Å. Если не указано иначе, все другие химические реагенты использовали без дополнительной очистки. Спектры 1H, 13C и 31P получали с помощью спектрометра Bruker ЯМР (AV-600, DRX-500, или DPX-400). Данные масс-спектрометрии получали из центральных лабораторий The Scripps Research Institute. Неприродный дезоксирибонуклеозидтрифосфаты синтезировали, как описано ранее (см. Dien, V. T. et al.J. Am. Chem. Soc. 2018, 140, 16115-16123; Lavergne, T. et al. J. Am. Chem. Soc. 2013, 135, 5408-5419; Matsuda, S. et al. J. Am. Chem. Soc. 2007, 129, 5551-5557; Seo, Y. J. et al. J. Am. Chem. Soc. 2009, 131, 3246-3252, описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме). Нуклеозидтрифосфаты dNaMTP, dTPT3TP, dCNMOTP и TPT3TP любезно предоставлены Synthorx Inc.
[00672] Синтез нуклеинового основания 13a. Нуклеиновое основание TAT1 (соединение 13a) синтезировали способами на основе описанных в литературе (см. New, J. S. et al.J. Med. Chem. 1989, 32, 1147-1156; Asagarasu, A. et al. Chem. Pharm. Bull. 2009, 57, 34-42, описание каждого из которых включено, таким образом, в настоящее описание в качестве ссылки в полном объеме). В кратком изложении, соответствующее промежуточное соединение акриловой кислоты (соединение 11) получали посредством конденсации тиазол-4-карбоксальдегида (1 экв.) с малоновой кислотой при 100°C с использованием пиридина в качестве растворителя и пиперидина в качестве катализатора (2:1 пиридин:пиперидин, конечная концентрация в реакционной смеси 2,75 M) в течение 13 ч. с последующим кипячением с обратным холодильником в течение 1 ч. Хлорирование этой кислоты (1 экв.) тионилхлоридом (1,1 экв.) в хлороформе (концентрация в реакционной среде 1,3 M) в присутствии каталитического DMF (0,2 экв.) приводило к получению ацилхлорида. Без дальнейшей очистки ацилхлорид превращали в соответствующий азид (соединение 12) в двухфазной смеси 1:1 1,4-диоксана и воды при 5°C с азидом натрия (2 экв.). Неочищенный ацилазид растворяли в хлороформе и по каплям добавляли в дифениловый простой эфир до 0,2 M. Затем полученную смесь нагревали до 230°C в течение 2 ч. После охлаждения до комнатной температуры добавляли гексаны и фильтровали реакционную смесь с использованием дополнительных гексанов для промывки. Полученный неочищенный продукт очищали посредством флэш-хроматографии с использованием силикагеля в качестве стационарной фазы для получения желаемого нуклеинового основания (соединение 13a).
Схема S1.(g) Пиперидин, пиридин, 100°C, 12 ч., затем кипячение с обратным холодильником в течение 1 ч., (h) SOCl2, DMF, CHCl3, кипячение с обратным холодильником, 3 ч., (i) NaN3, 1,4-диоксан, H2O, 5°C, 0,5 ч., (j) дифениловый простой эфир, 230°C, 1 ч.
[00673] Реакция сочетания нуклеинового основания и снятие защитной группы с TBS-защищенного риболактона 2. Арилгалид (1,0 экв.) растворяли в безводном THF до 0,1 M и охлаждали до -78°C. После охлаждения раствора n-BuLi (1,6 экв., 1,6 M в гексанах) по каплям добавляли в реакционную колбу. Через 30 мин перемешивания этой смеси при -78°C по каплям добавляли раствор TBS-защищенного риболактона (соединения 2, 1,6 экв.) в безводном THF в количестве ~0,8 M. Затем реакционной смеси позволяли перемешиваться еще в течение 1 ч. при -78°C перед тушением насыщенным водным раствором хлорида аммония. После нагревания до комнатной температуры летучие вещества удаляли под вакуумом и полученный остаток экстрагировали этилацетатом и водой. Органические вещества сушили над сульфатом натрия, фильтровали и снова удаляли летучие вещества под вакуумом.
[00674] Остаточный растворитель удаляли под высоким вакуумом в течение 1 ч. при комнатной температуре перед повторным растворением в безводном DCM до 0,1 M. Раствор охлаждали до -78°C перед добавлением по каплям триэтилсилана (3 экв.) с последующим добавлением по каплям 48% раствора диэтилэфирата трифторида бора (3 экв.). Затем реакционной смеси позволяли перемешиваться при -78°C в течение 15-20 мин перед тушением раствором 1:1 метанола и триэтиламина (добавляли объем 30% к объему реакционной смеси). Этой смеси позволяли перемешиваться в течение 3 мин при -78°C перед добавлением насыщенного раствора бикарбоната натрия (50% исходного объема реакционной смеси). Затем смесь разводили DCM в 2 раза и отделяли органический слой. Водный слой экстрагировали еще два раза с помощью DCM и объединенные органические вещества сушили над сульфатом натрия, фильтровали и концентрировали под вакуумом.
[00675] Какие-либо остаточные растворители удаляли из остатка под сильным вакуумом в течение 1 ч. при комнатной температуре. В это время остаток растворяли в THF до 0,1 M и добавляли раствор фторида тетрабутиламмония (3,3 экв., 1,0 M в THF). Реакционной смеси позволяли перемешиваться в течение 1 ч. перед разведением этилацетатом и экстракцией водой. Объединенные органические вещества сушили сульфатом натрия, фильтровали и концентрировали под вакуумом. Затем полученный остаток очищали посредством флэш-хроматографии на силикагеле с использованием 5-10% этанола в DCM в качестве подвижной фазы.
[00676] Реакция сочетания нуклеинового основания и снятие защитных групп с бензил-защищенного риболактона 10. Арилгалид (1,0 экв.) растворяли в безводном THF до 0,1 M и охлаждали до -78°C. После охлаждения раствор n-BuLi (1,6 экв., 1,6 M в гексанах) по каплям добавляли в реакционную колбу. Через 30 мин перемешивания этой смеси при -78°C по каплям добавляли раствор бензил-защищенного риболактона (соединение 10, 1,6 экв.) в безводном THF в количестве ~0,8 M. Затем реакционной смеси позволяли перемешиваться в течение еще 1 ч. при -78°C перед тушением насыщенным водным раствором хлорида аммония. После нагревания до комнатной температуры летучие вещества удаляли под вакуумом и полученный остаток экстрагировали этилацетатом и водой. Органические вещества сушили над сульфатом натрия, фильтровали и снова удаляли летучие вещества под вакуумом.
[00677] Остаточный растворитель удаляли под сильным вакуумом в течение 1 ч. при комнатной температуре перед повторным растворением в безводном DCM до 0,1 M. Раствор охлаждали до -78°C перед добавлением по каплям триэтилсилана (3 экв.) с последующим добавлением по каплям 48% раствора диэтилэфирата трифторида бора (3 экв.). Затем реакционной смеси позволяли перемешиваться при -78°C в течение 15-20 мин перед тушением раствором 1:1 метанола и триэтиламина (добавляют объем 30% объема реакционной смеси). Этой смеси позволяли перемешиваться в течение 3 мин при -78°C перед добавлением насыщенного раствора бикарбоната натрия (50% исходного объема реакционной смеси). Затем смесь разводили DCM в 2 раза и отделяли органический слой. Водный слой экстрагировали еще два раза с помощью DCM и объединенные органические вещества сушили над сульфатом натрия, фильтровали и концентрировали под вакуумом. Затем бензил-защищенное промежуточное соединение нуклеозида очищали посредством флэш-хроматографии на силикагеле с использованием градиента от 0 до 8-10% этилацетата в гексанах в качестве подвижной фазы.
[00678] Очищенный бензил-защищенный нуклеозид разводили в метаноле до 0,1 M вместе с 10% палладия на угле (0,1 экв.). Раствор замораживали жидким азотом и, пока он был заморожен, реакционную колбу три раза барботировали водородом, оставляя колбу под баллоном с водородом после последнего барботирования. Затем реакционной смеси позволяли размораживаться и интенсивно перемешивали в атмосфере водорода в течение 3 ч. при комнатной температуре. Затем реакционную смесь фильтровали через прокладку силикагеля, промывая этилацетатом. Затем летучие вещества удаляли под вакуумом и полученный неочищенный остаток очищали посредством флэш-хроматографии на силикагеле с использованием 5-10% этанола в DCM в качестве подвижной фазы.
[00679] Цианирование арилбромидов. В сухой сосуд для микроволновой печи помещали CuCN (8 экв.), Pd2(dba)3 (0,1 экв.) и DPPF (0,1 экв.) и три раза барботировали аргоном. Арилбромид (1 экв.) растворяли в дегазированном безводном DMF до 0,05-0,1 M и переносили в реакционную колбу. Затем сосуд запаивали в атмосфере аргона и интенсивно перемешивали при 155°C в течение ночи (~16 ч.). Затем реакционную смесь охлаждали до комнатной температуры и фильтровали через тонкую прокладку силикагеля, промывая этилацетатом. Летучие вещества удаляли под вакуумом и полученный неочищенный остаток очищали посредством флэш-хроматографии на силикагеле с использованием градиента 70-95% этилацетата в гексанах в качестве подвижной фазы.
[00680] Общий способ трифосфорилирования нуклеозидов. В атмосфере инертного газа в сухой сосуд для микроволновой печи помещали протонную губку (1,3 экв.) и свободный нуклеозид (1 экв.). В колбу добавляли безводный триметилфосфат (40 экв.) и полученную смесь охлаждали до -15°C на льдосоляной бане. Свежедистилированный оксихлорид фосфора (1,3 экв.) добавляли по каплям в охлажденную колбу и реакционной смеси позволяли перемешиваться при -15°C в течение 3 ч. В это время пирофосфат трибутиламмония (5 экв.) растворяли в безводном DMF в концентрации 0,5 M в отдельной сухой колбе и добавляли по каплям в реакционную колбу с последующим добавлением по каплям безводного трибутиламина (6 экв.). Полученную смесь медленно нагревали до 0°C и позволяли ей перемешиваться при этой температуре еще в течение 30 мин. Затем реакционную смесь тушили добавлением водного раствора бикарбоната триэтиламмония (0,5 M, pH 7,5, 1:1 с общим объемом реакционной смеси) и перемешивали при комнатной температуре в течение 15 мин. Затем эту подвергнутую тушению реакционную смесь очищали посредством анионообменной хроматографии с использованием DEAE Sephadex A-25 в качестве стационарной фазы и медленного градиента 0-1,2 M TEAB при pH 7,5 в качестве подвижной фазы. Фракции, содержащие нуклеозидтрифосфат, объединяли и концентрировали посредством SpeedVac для получения сухого желаемого продукта.
[00681] Реакция сочетания нуклеинового основания и снятие защитной группы с бензоил-защищенного риболактона 14. Смесь нуклеинового основания (1,0 экв.) и N, O-бис(триметилсилил)ацетамида (1,2 экв.) в ацетонитриле до 0,1 M перемешивали в атмосфере аргона при комнатной температуре в течение 30 мин. В это время добавляли 1-O-ацетил-2,3,5-три- O-бензоил-β-D-рибофуранозу (соединение 14, 1,2 экв.) и полученную реакционную смесь охлаждали до 0°C. Затем в реакционную смесь добавляли SnCl4 (1 экв.) и раствор перемешивали в течение ночи при комнатной температуре. Реакционную смесь экстрагировали этилацетатом и насыщенным водным раствором NaHCO3 и объединенные органические слои сушили над безводным Na2SO4. После фильтрации и выпаривания остаток очищали посредством флэш-хроматографии на силикагеле.
[00682] Превращение пиридонов в тио-пиридоны. Пиридон (1 экв.) сушили посредством повторного соиспарения с сухим толуолом. Добавляли реагент Лавессона (3 экв.) в атмосфере аргона и смесь нагревали в течение ночи при 120°C. После фильтрации на хлопке фильтрат концентрировали и неочищенный продукт очищали посредством флэш-хроматографии на силикагеле.
[00683] Фторирование защищенных нуклеозидов. Защищенный нуклеозид (1 экв.) растворяли в MeOH-CH3CN (1:1 об./об.) до 0,1 M и добавляли Selectfluor (1,1 экв.). Смесь нагревали с обратным холодильником в течение 3 ч. Затем летучие вещества удаляли под вакуумом и полученный остаток экстрагировали этилацетатом и вода. Объединенные органические слои выпаривали и полученный неочищенный остаток сушили посредством соиспарения с безводным толуолом три раза. Остаток растворяли в TfOH-DCM (1:1 об./об.) до 0,1 M и смесь перемешивали при комнатной температуре в течение 1 ч. В это время реакционную смесь концентрировали под вакуумом и полученный неочищенный продукт очищали посредством флэш-хроматографии на силикагеле.
[00684] Характеризация соединений.
[00685] Соединение 1b. (0,387 ммоль, выход 38%, 3 стадии). 1H ЯМР (600 МГц, хлороформ-d) 7,46 (дд, J=8,2, 0,9 Гц, 1H), 7,01 (дд, J=8,2, 1,9 Гц, 1H), 6,87 (д, J=1,9 Гц, 1H), 5,32 (д, J=3,3 Гц, 1H), 4,43 (дд, J=4,6, 3,3 Гц, 1H), 4,37 (дд, J=8,1, 4,6 Гц, 1H), 4,05-4,01 (м, 1H), 3,97 (дд, J=12,0, 3,2 Гц, 1H), 3,83 (с, 3H), 3,77 (дд, J=11,9, 4,1 Гц, 1H). 13C ЯМР (151 МГц, CDCl3) 156,40, 134,51, 128,81, 124,00, 121,01, 111,03, 82,18, 78,55,73,31, 72,25, 62,57, 55,85.HRMS (ESI-TOF+) вычислено для C12H15ClO5 [M+Na]+ 297,0500; найдено 297,0501.
[00686] Соединение 3b. (0,333 ммоль, выход 52%, 3 стадии). 1H ЯМР (600 МГц, метанол-d4) 7,53 (д, J=7,4, 1,5 Гц, 1H), 7,24 (тд, J=7,8, 1,8 Гц, 1H), 6,98-6,92 (м, 2H), 5,33 (д, J=2,9 Гц, 1H), 4,35-4,28 (м, 2H), 4,03 (ддд, J=8,2, 4,7, 2,6 Гц, 1H), 3,89 (дд, J=13,1, 3,8 Гц, 1H), 3,84 (с, 3H), 3,70 (дд, J=12,0, 4,7 Гц, 1H). 13C ЯМР (151 МГц, MeOD) 157,46, 129,07, 129,03, 127,68, 121,09, 110,69, 83,00, 79,66, 74,04, 73,61, 63,32, 55,72. HRMS (ESI-TOF+) вычислено для C12H16O5 [M+Na]+ 263,0890; 263,0888.
[00687] Соединение 4b. (0,765 ммоль, выход 62%, 3 стадии). 1H ЯМР (600 МГц, хлороформ-d) 7,23 (дд, J=9,2, 3,1 Гц, 1H), 6,94 (тд, J=8,4, 3,2 Гц, 1H), 6,77 (дд, J=8,9, 4,2 Гц, 1H), 5,28 (д, J=3,3 Гц, 1H), 4,43-4,38 (м, 1H), 4,30 (дд, J=8,2, 4,6Гц, 1H), 3,99 (дт, J=7,8, 3,6 Гц, 1H), 3,90 (дд, J=12,0, 3,2 Гц, 1H), 3,79 (с, 3H), 3,72 (дд, J=12,0, 4,0 Гц, 1H). 13C ЯМР (151 МГц, CDCl3) 158,28, 156,69, 151,87, 151,86, 127,64, 127,59, 115,09, 114,92, 114,62, 114,46, 110,95, 110,90, 81,92, 78,51, 73,10, 72,30, 62,37, 56,02. HRMS (ESI-TOF+) вычислено для C12H15FO5 [M+Na}+ 281,0796; 281,0799.
[00688] Соединение 5b. (0,366 ммоль, выход 50%, 3 стадии). 1H ЯМР (600 МГц, метанол-d4) δ 7,95 (с, 1H), 7,43 (с, 1H), 7,32 (д, J=5,4 Гц, 1H), 7,26 (д, J=5,4 Гц, 1H), 5,38 (д, J=3,0 Гц, 1H), 4,40-4,35 (м, 1H), 4,32 (дд, J=8,5, 4,4 Гц, 1H), 4,06 (ддд, J=8,2, 4,6, 2,6 Гц, 1H), 3,92 (дд, J=12,0, 2,5 Гц, 1H), 3,90 (с, 3H), 3,71 (дд, J=12,0, 4,7 Гц, 1H). 13C ЯМР (151 МГц, метанол-d4) δ 154,39, 139,73, 133,33, 124,79, 123,31, 122,99, 122,24, 102,01, 81,60, 78,62, 72,69, 72,23, 61,98, 54,69. HRMS (ESI-TOF+) вычислено для C14H16O5S [M+Na]+ 319,0611; 319,0612.
[00689] Соединение 6b. (0,160 ммоль, выход 51%, 3 стадии). 1H ЯМР (600 МГц, метанол-d4) δ 7,98 (с, 1H), 7,48 (д, J=5,4 Гц, 1H), 7,34 (с, 1H), 7,29 (д, J=5,4 Гц, 1H), 5,39 (д, J=3,7 Гц, 1H), 4,40-4,35 (м, 1H), 4,32 (дд, J=8,5, 4,4 Гц, 1H), 4,05 (ддд, J=8,4, 4,7, 2,6 Гц, 1H), 3,91 (дд, J=11,9, 2,6 Гц, 1H), 3,89 (с, 3H), 3,71 (дд, J=12,0, 4,7 Гц, 1H). 13C ЯМР (151 МГц, метанол-d4) δ154,41, 139,63, 131,85, 126,26, 125,11, 123,24, 120,94, 102,92, 81,60, 78,66, 72,68, 72,29, 61,95. HRMS (ESI-TOF+) вычислено для C14H16O5S [M+Na]+ 319,0611; 319,0609.
[00690] Соединение 7b. (1,248 ммоль, выход 61%, 3 стадии). 1H ЯМР (600 МГц, хлороформ-d) δ 7,41 (дд, J=8,1, 0,9 Гц, 1H), 7,17 (дд, J=8,1, 1,8 Гц, 1H),7,02 (д, J=1,8 Гц, 1H), 5,32 (д, J=3,3 Гц, 1H), 4,44 (дд, J=4,6, 3,4 Гц, 1H), 4,38 (дд, J=8,1, 4,6 Гц, 1H), 4,04 (дт, J=7,7, 3,6 Гц, 1H), 3,98 (дд, J=11,9, 3,2 Гц, 1H), 3,83 (с, 3H), 3,79 (дд, J=11,9, 4,1 Гц, 1H). 13C ЯМР (151 МГц, CDCl3) δ 155,82, 128,57, 123,94, 123,44, 121,77, 113,26, 81,62, 78,00, 72,72, 71,60, 61,99, 55,28. HRMS (ESI-TOF-) вычислено для C12H15BrO5 [M+Cl]- 352,9797; найдено 352,9799.
[00691] Соединение 8a. (0,066 ммоль, выход 42% из соединения 6b). 1H ЯМР (600 МГц, метанол-d4) δ 7,68 (дд, J=7,8, 0,9 Гц, 1H), 7,34 (дд, J=7,8, 1,5 Гц, 1H), 7,28 (д, J=1,5 Гц, 1H), 5,32 (д, J=3,1 Гц, 1H), 4,39 (дд, J=4,5, 3,2 Гц, 1H), 4,30 (дд, J=8,6, 4,5 Гц, 1H), 4,03 (ддд, J=8,6, 4,6, 2,5 Гц, 1H), 3,93-3,89 (м, 4H), 3,70 (дд, J=12,0, 4,6 Гц,1H). 13C ЯМР (151 МГц, MeOD) δ 157,65, 134,76, 129,90, 125,39, 120,07, 113,77, 112,30, 83,14, 79,64, 73,80, 73,50, 63,17, 56,31, 50,87. HRMS (ESI-TOF-) вычислено для C13H15NO5 [M-H]- 264,0877; найдено 264,0869.
[00692] Соединение 9b. (0,288 ммоль, выход 29%, 3 стадии). 1H ЯМР (600 МГц, хлороформ-d) δ 7,12 (д, J=9,9 Гц, 1H), 6,73 (д, J=6,1 Гц, 1H), 5,02 (д, J=6,1 Гц, 1H), 4,17 (дд, J=5,9, 4,8 Гц, 1H), 4,13-4,09 (м, 1H), 4,08 (т, J=6,0 Гц, 1H), 3,98 (дд, J=11,9, 3,3 Гц, 1H), 3,87 (с, 3H), 3,85, 3,81 (м, 1H), 2,29 (д, J=2,0 Гц, 3H). 13C ЯМР (151 МГц, CDCl3) δ 156,19, 154,61, 151,45, 126,24, 124,08, 123,95, 113,10, 83,98, 80,69, 71,56, 62,65, 55,76, 14,22. HRMS (ESI-TOF+) вычислено для C13H17FO5 [M+Na]+ 295,0952; 295,0952.
[00693] Соединение 13a (3,184 ммоль, 11% за 4 стадии). 1H ЯМР (500 МГц, DMSO-d6) δ 11,87 (ш.с., 1 H, N-H), 9,59 (с, 1H, Ar-H), 7,50 (д, J=5,0 Гц, 1 H, Ar-H), 6,98 (д, J=5,0 Гц, 1 H, Ar-H). 13C ЯМР (125 МГц, CDCl3) δ 161,9, 150,3, 159,2, 133,1, 124,3, 102,5. HRMS (ESI-TOF+) вычислено для C6H5N2OS+ [M+H]+ 153,0117; найдено 153,0115.
[00694] Соединение 13b. (0,530 ммоль, выход 53%). 1H ЯМР (500 МГц, CDCl3) δ 9,13 (с, 1 H, Ar-H), 7,36-8,16 (м, 16H, Ar-H), 6,87 (д, J=5,0 Гц, 1H, H-1'), 6,71 (д, J=5,0 Гц, 1 H, Ar-H), 5,99-6,01 (м, 1 H, H-3'), 5,90-5,92 (м, 1 H, H-2'), 4,92 (дд, J1=15 Гц, J2=5 Гц, 1 H, H-5'a), 4,80-4,82 (м, 1 H, H-4'), 4,72 (дд, J1=15 Гц, J2=5 Гц, 1 H, H-5'b). 13C ЯМР (125 МГц, CDCl3) δ166,5. 165,7, 165,6, 160,7, 156,7, 134,1, 133,9, 130,9, 130,3, 130,2, 130,1, 130,1, 130,1, 129,8, 129,1, 129,0, 128,9, 128,8, 104,5, 89,2, 80,8, 75,3, 71,4, 64,0. HRMS (ESI-TOF+) вычислено для C32H25N2O8S [M+H]+ 597,1326; найдено 597,1330.
[00695] Соединение 13c. (0,056 ммоль, выход 56%). 1H ЯМР (500 МГц, CDCl3) δ 9,16 (с, 1 H, Ar-H), 7,31-8,24 (м, 17 H, Ar-H), 7,12 (д, J=5 Гц, 1H, H-1'), 5,95-5,96 (м, 1 H, H-2'), 5,87-5,90 (м, 1 H, H-3'), 4,99 (дд, J1=15 Гц, J2=5 Гц, 1 H, H-5'a), 4,89-4,91 (м, 1H, H-4'), 4,73 (дд, J1=15 Гц, J2=5 Гц, 1 H, H-5'b). 13C ЯМР (125 МГц, CDCl3) δ175,7, 166,4, 165,6, 165,3, 163,7, 151,8, 142,1, 134,2, 134,1, 134,0, 132,5, 130,5, 130,3, 130,1, 129,2, 129,2, 128,9, 128,9, 109,7, 92,2, 80,8, 75,9, 69,9, 63,0. HRMS (ESI-TOF+) вычислено для C32H25N2O7S2 [M+H]+ 613,1098; найдено 613,1095.
[00696] Соединение 13d. (0,043 ммоль, выход 89%). 1H ЯМР (500 МГц, DMSO-d6) δ 9,58 (с, 1 H, Ar-H), 8,68 (д, J=10 Гц, 1 H, Ar-H), 7,45 (д, J=10 Гц, 1 H, Ar-H), 6,81 (д, J=2,5 Гц, 1 H, H-1'), 5,48 (д, J=5 Гц, 1 H, -OH), 5,39-5,41 (м, 1 H,-OH), 5,18 (д, J=5 Гц, 1 H, -OH), 4,03-4,05 (м, 2 H, H-2', H-3'), 3,98-4,00 (м, 1 H, H-4'), 3,78 (дд, J1=10Гц, J2=2,5 Гц, 1 H, H-5'a), 3,64 (дд, J1=10Гц, J2=2,5 Гц, 1 H, H-5'b). 13C ЯМР (125 МГц, DMSO-d6) δ 174,2, 166,6, 152,2, 135,5, 109,4, 99,9, 95,1, 85,5, 76,6, 69,0, 60,2. HRMS (ESI-TOF+) вычислено для C11H13N2O4S2 [M+H]+ 301,0311; найдено 301,0309.
[00697] Соединение 15b. (0,931 ммоль, выход 93%). 1H ЯМР (500 МГц, CDCl3) δ 9,05 (д, J=10 Гц, 1 H, H-Ar), 7,30-8,16 (м, 20 H, Ar-H), 6,72 (д, J= 5 Гц, 1 H, H-1'), 5,97-5,99 (м, 1 H, H-2'), 5,85-5,88 (м, 1 H, H-3'), 4,98 (дд, J1=10 Гц, J2=5Гц, 1 H, H-5'a), 4,89-4,90 (м, 1 H, H-4'), 4,72 (дд, J1=10 Гц, J2=5 Гц, 1 H, H-5'b). 13C ЯМР (125 МГц, CDCl3) δ182,1, 179,6, 166,6, 165,5, 165,5, 165,2, 151,1, 143,9, 134,0, 133,9, 133,6, 130,4, 130,3, 130,2, 130,1, 129,6, 129,4, 129,1, 128,9, 128,9, 128,8, 92,8, 80,4,75,7, 69,7, 62,8. HRMS (ESI-TOF+) вычислено для C35H28NO8 [M+H]+ 590,1809; найдено 590,1811.
[00698] Соединение 15c. (0,088 ммоль, выход 87%). 1H ЯМР (500 МГц, CDCl3) δ 9,07 (д, J=10 Гц, 1 H, Ar-H), 7,30-8,17 (м, 20 H, Ar-H, H-1'), 5,98-5,99 (м, 1 H, H-2'), 5,82-5,84 (м, 1 H, H-3'), 4,89-4,94 (м, 2 H, H-5'a, H-4'), 4,80 (дд, J1=15 Гц,J2=5 Гц, 1 H, H-5'b). 13C ЯМР (125 МГц, CDCl3) δ184,9, 166,5, 165,6, 165,3, 134,0, 133,9, 133,3, 132,7, 130,5,130,3, 130,2, 129,8, 129,5, 129,1, 128,9, 128,9, 128,8, 127,3, 127,1, 112,8, 92,6, 80,3, 75,9, 70,1, 63,2. HRMS (ESI-TOF+) вычислено для C35H28NO7S [M+H]+ 606,1581; найдено 606,1579.
[00699] Соединение 15d. (0,044 ммоль, выход 92%). 1H ЯМР (500 МГц, MeOH-d4) δ 8,33 (д, J=10 Гц, 1 H, Ar-H), 7,52-7,80 (м, 4 H, Ar-H), 6,71 (д, J=5 Гц, 1 H, Ar-H), 6,36 (д, J=2,5 Гц, 1 H, H-1'), 4,22-4,26 (м, 2 H, H-2', H-3'), 4,10-4,11 (м,1 H, H-4'), 3,94 (дд, J1=15 Гц, J2=5 Гц, 1 H, H-5'a), 3,82 (дд, J1=15 Гц, J2=5 Гц, 1 H, H-5'b). 13C ЯМР (125 МГц, MeOH-d4) δ 163,0, 137,6, 133,0, 127,5, 127,4, 127,1, 126,3, 125,6, 106,9, 90,1, 85,1, 75,5, 70,3, 61,4. HRMS (ESI-TOF+) вычислено для C14H16NO4S [M+H]+ 294,0795; найдено 294,0799.
[00700] Соединение 16a. (0,077 ммоль, выход 71%). 1H ЯМР (500 МГц, CDCl3) δ 7,33-8,42 (м, 20 H, Ar-H), 6,82 (д, J=5 Гц, 1 H, H-1'), 5,94-5,97 (м, 1 H, H-2'), 5,83-5,85 (м, 1 H, H-3'), 4,87 (д, J=10 Гц, 1 H, H-5'a), 4,74-4,79 (м, 2 H, H-4', H-5'b). 13C ЯМР (125 МГц, CDCl3) δ166,6, 165,7, 165,6, 162,9, 160,8, 134,0, 134,0, 133,9, 133,5, 130,3, 130,2, 130,1, 129,7, 129,1, 129,1, 129,1, 128,9, 128,8, 128,8, 120,3, 111,1, 88,9, 87,8,85,3, 80,6, 80,4, 74,8, 71,4, 64,1. 19F ЯМР (376 МГц, CDCl3) δ -145,6. HRMS (ESI-TOF+) вычислено для C35H27FNO8 [M+H]+ 608,1715; найдено 608,1717.
[00701] Соединение 16b. (0,066 ммоль, выход 65%). 1H ЯМР (500 МГц, CDCl3) δ 9,07 (д, J=10 Гц,1 H, Ar-H), 7,32-8,18 (м, 20 H, Ar-H, H-1'), 5,96-5,98 (м, 1 H, H-2'), 5,87-5,89 (м, 1 H, H-3'), 4,97 (дд, J1=15 Гц, J2=5 Гц, 1 H, H-5'a), 4,89-4,91 (м, 1H, H-4'), 4,76 (дд, J1=15 Гц, J2=5 Гц, 1 H, H-5'b). 13C ЯМР (125 МГц, CDCl3) δ184,3, 166,6, 165,6, 165,3, 134,7, 134,1, 134,0, 133,9, 133,9, 133,3, 133,2, 130,5, 130,3, 130,3, 129,8, 129,7, 129,3, 129,2, 128,9, 128,9, 128,9, 128,8, 128,3, 126,8, 108,4, 100,0, 92,3, 80,7, 75,8, 70,3, 63,2, 56,0. HRMS (ESI-TOF+) вычислено для C35H27FNO7S [M+H]+ 624,1487; найдено 624,1490.
[00702] Соединение 16c. (0,042 ммоль, выход 90%). 1H ЯМР (500 МГц, MeOH-d4) δ 9,04 (д, J=10 Гц, 1 H, H-Ar), 8,73 (д, J=10 Гц, 1 H, H-Ar),7,83-7,87 (м, 2 H, Ar-H), 7,65-7,68 (м, 1 H, H-Ar), 6,96 (с, 1 H, H-1'), 4,23-4,29 (м, 2 H, H-2', H-3'), 4,18-4,21 (м, 1 H, H-4'), 4,11 (дд, J1=10 Гц, J2=5 Гц, H-5'a), 3,91 (дд, J1=10 Гц, J2=5 Гц, H-5'b). 13C ЯМР (125 МГц, MeOH-d4) δ 180,5, 150,5, 148,6, 134,2, 134,1, 133,2, 132,0, 132,0, 129,2, 126,1, 125,9, 119,6, 119,6, 115,5, 115,1, 96,2, 84,4, 76,1, 68,1, 59,4. HRMS (ESI-TOF)+ вычислено для C14H15FNO4S [M+H]+ 312,0700; найдено 312,0702.
[00703] Соединение 1c. (0,048, выход 24%). 31P ЯМР (162 МГц, H2O) δ -8,79 - -10,99 (м), -21,21 - -21,58 (м), -21,77 - -23,23 (м). MS (MALDI-TOF-, матрица: 9-аминоакридин) (m/z) [M-H]- вычислено для C12H17ClO14P3-, 512,95; найдено, 512,7.
[00704] Соединение 3c. (0,031 ммоль, выход 31%). 31P ЯМР (162 МГц, H2O) δ -11,19 - -11,91 (м), -22,06 - -22,73 (м), -23,87 - -24,46 (м). MS (MALDI-TOF-, матрица: 9-аминоакридин) (m/z) [M-H]- вычислено для C12H18O14P3-,479,00; найдено,497,4.
[00705] Соединение 4c. (0,014 ммоль, выход 14%). 31P ЯМР (162 МГц, H2O) δ -7,31 (д), -11,47 (д), -22,79 - -23,21 (м). MS (MALDI-TOF-, матрица: 9-аминоакридин) (m/z): [M-H]- вычислено для C12H17FO14P3-, 496,97; найдено, 497,3.
[00706] Соединение 5c. (0,042 ммоль, выход 23%). 31P ЯМР (162 МГц, D2O) δ -8,48 (д, J=17,8 Гц), -11,21 (д, J=20,7 Гц), -22,96 (т, J=21,6 Гц). MS (MALDI-TOF-, матрица: 9-аминоакридин) (m/z): [M-H]- вычислено для C14H18O14P3S-, 534,97; найдено,535,4.
[00707] Соединение 6c. (0,017 ммоль, выход 21%). 31P ЯМР (162 МГц, D2O) δ -9,25 (д, J=21,0 Гц), -10,93 (д, J=19,5 Гц), -22,94 (т, J=20,5 Гц). MS (MALDI-TOF-, матрица: 9-аминоакридин) (m/z): [M-H]- вычислено для C14H18O14P3S-, 534,97; найдено, 535,4.
[00708] Соединение 7c. (0,009 ммоль, выход 12%). 31P ЯМР (162 МГц, H2O) δ -10,07 - -11,45 (м), -21,8 - -22,21 (м), -24,06 - -24,60 (м). MS (MALDI-TOF-, матрица: 9-аминоакридин) (m/z) [M-H]- вычислено для C12H17BrO14P3-, 556,91; найдено, 556,6.
[00709] Соединение 8b. (0,023 ммоль, выход 22%). 31P ЯМР (162 МГц, H2O) δ -9,96 - -12,34 (м), -21,91 - -22,45 (м), -22,88 - -24,4 (м). MS (MALDI-TOF-, матрица: 9-аминоакридин) (m/z) [M-H]- вычислено для C13H17NO14P3-, 503,99; найдено,504,3.
[00710] Соединение 9c. (0,021 ммоль, выход 21%). 31P ЯМР (162 МГц, H2O) δ -9,77 - -10,37 (м), -10,8 - -11,17 (м), -22,0 - -22,2 (м). MS (MALDI-TOF-, матрица: 9-аминоакридин) (m/z) [M-H]- вычислено для C13H19FO14P3-, 511,0; найдено, 511,0.
[00711] Соединение 13e. (0,007 ммоль, выход 22%). 31P ЯМР (400 МГц, D2O): δ -10,82-10,94 (м), -11,68-11,74 (м), -23,21-23,45 (м). MS (MALDI-TOF, матрица: 9-аминоакридин) (m/z): [M-H]- вычислено для C11H14N2O13P3S2-, 538,9, найдено, 538,3.
[00712] Соединение 15e. (0,012 ммоль, выход 35%). 31P ЯМР (400 МГц, D2O): δ -10,74-10,84 (м), -11,55-11,67 (м), -23,17-23,42 (м). MS (MALDI-TOF, матрица: 9-аминоакридин) (m/z): [M-H]- вычислено для C14H17NO13P3S-, 532,0, найдено, 532,5.
[00713] Соединение 16d. (0,009 ммоль, выход 29%). 31P ЯМР (400 МГц, D2O): δ -10,65-10,79 (м), -11,45-11,54 (м), -22,16-23,40 (м). MS (MALDI-TOF, матрица: 9-аминоакридин) (m/z): [M-H]- вычислено для C14H16FNO13P3S-, 550,0, найдено, 550,6.
Схема S2. (a) 1,6 M n-BuLi, THF, -78°C, 1 ч, (b) триэтилсилан, DCM, BF3·OEt2, -78°C, 15-20 мин, (c) 1,0 M TBAF, THF, 1 ч., (d) протоновая губка, POCl3, Bu3N, Bu3NPPi, (MeO)3P, DMF, -15°C, 3 ч.
Схема S3. (a) 1,6 M n-BuLi, THF, -78°C, 1 ч., (b) триэтилсилан, DCM, BF3·OEt2, -78°C, 15-20 мин, (c) 1,0 M TBAF, THF, 1 ч., (d) протоновая губка, POCl3, Bu3N, Bu3NPPi, (MeO)3P, DMF, -15°C, 3 ч.
Схема S4.(a) 1,6 M n-BuLi, THF, -78°C, 1 ч., (b) триэтилсилан, DCM, BF3·OEt2, -78°C, 15-20 мин, (c) 1,0 M TBAF, THF, 1 ч., (d) протоновая губка, POCl3, Bu3N, Bu3NPPi, (MeO)3P, DMF, -15°C, 3 ч.
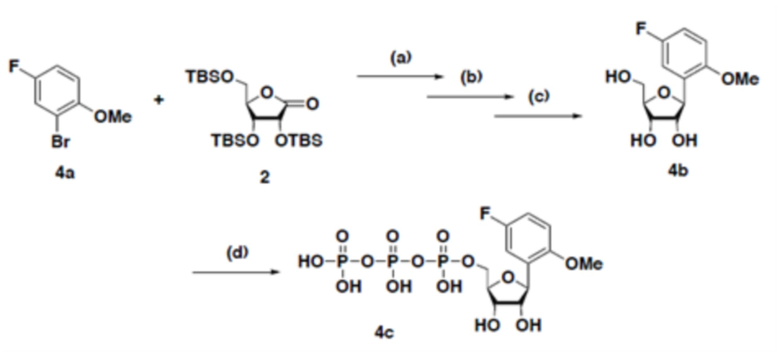
Схема S5.(a) 1,6 M n-BuLi, THF, -78°C, 1 ч., (b) триэтилсилан, DCM, BF3·OEt2, -78°C, 15-20 мин, (c) 1,0 M TBAF, THF, 1 ч., (d) протоновая губка, POCl3, Bu3N, Bu3NPPi, (MeO)3P, DMF, -15°C, 3 ч.
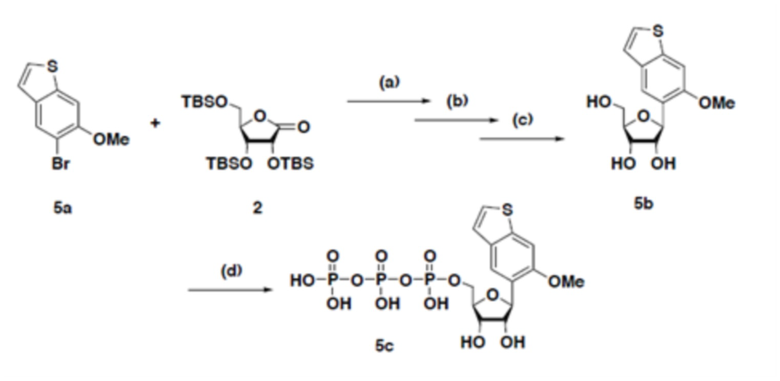
Схема S6. (a) 1,6 M n-BuLi, THF, -78°C, 1 ч., (b) триэтилсилан, DCM, BF3·OEt2, -78°C, 15-20 мин, (c) 1,0 M TBAF, THF, 1 ч., (d) протоновая губка, POCl3, Bu3N, Bu3NPPi, (MeO)3P, DMF, -15°C, 3 ч.
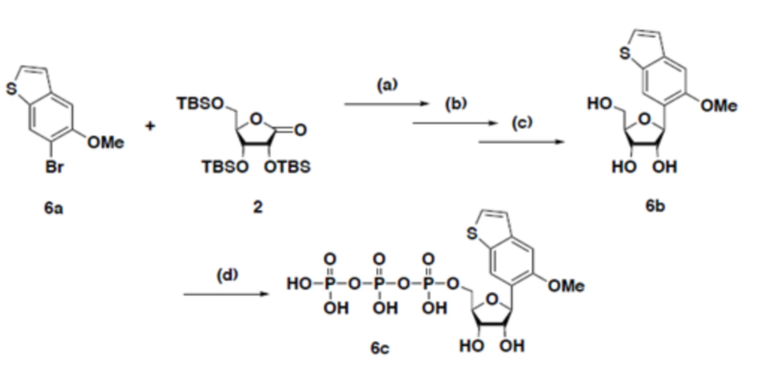
Схема S7. (a) 1,6 M n-BuLi, THF, -78°C, 1 ч., (b) триэтилсилан, DCM, BF3·OEt2, -78°C, 15-20 мин, (c) 1,0 M TBAF, THF, 1 ч., (d) протоновая губка, POCl3, Bu3N, Bu3NPPi, (MeO)3P, DMF, -15°C, 3 ч.
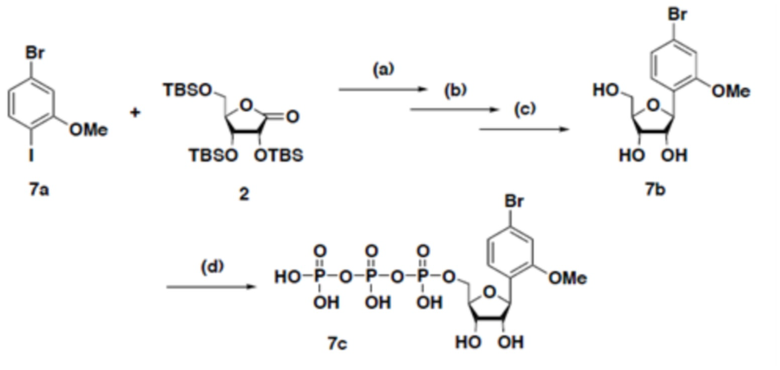
Схема S8. (d) протоновая губка, POCl3, Bu3N, Bu3NPPi, (MeO)3P, DMF, -15°C, 3 ч., (e) CuCN, Pd2(dba)3, DPPF, DMF, 155°C, 16 ч.
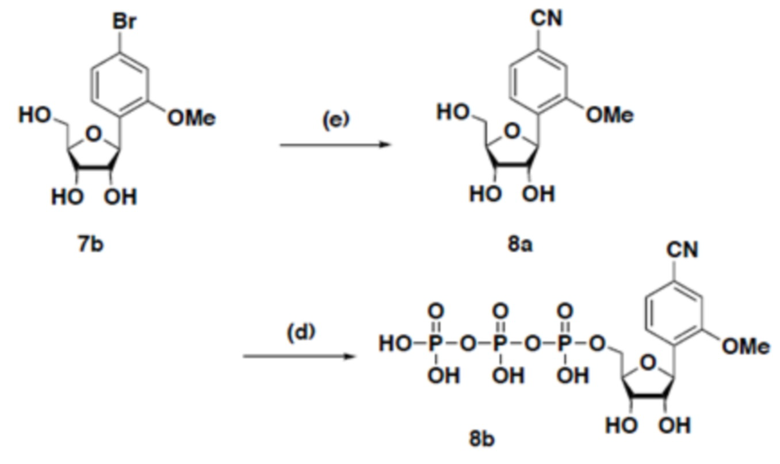
Схема S9. (a) 1,6 M n-BuLi, THF, -78°C, 1 ч., (b) триэтилсилан, DCM, BF3·OEt2, -78°C, 15-20 мин, (d) протоновая губка, POCl3, Bu3N, Bu3NPPi, (MeO)3P, DMF, -15°C, 3 ч., (f) H2(газ), Pd/C (10%), MeOH, RT, 3 ч.
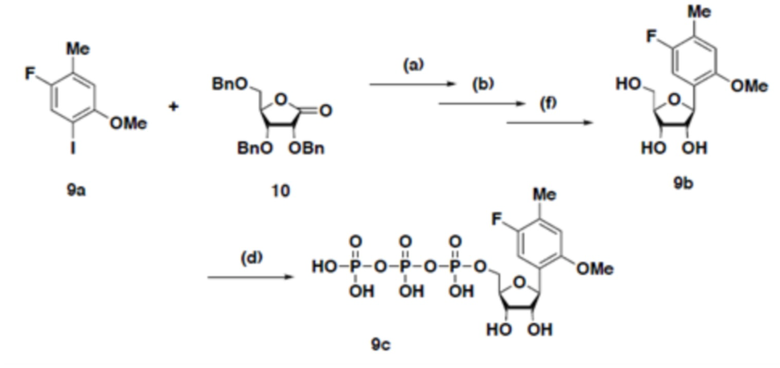
Схема S10. (k) N, O-бис(триметилсилил)ацетамид, SnCl4, CH3CN, RT, в течение ночи, (l) реагент Лавессона, 120°C, в течение ночи, (m) 30% NaOMe, DCM, 0°C, 2 ч., (d) протоновая губка, POCl3, Bu3N, Bu3NPPi, (MeO)3P, DMF, -15°C, 3 ч.
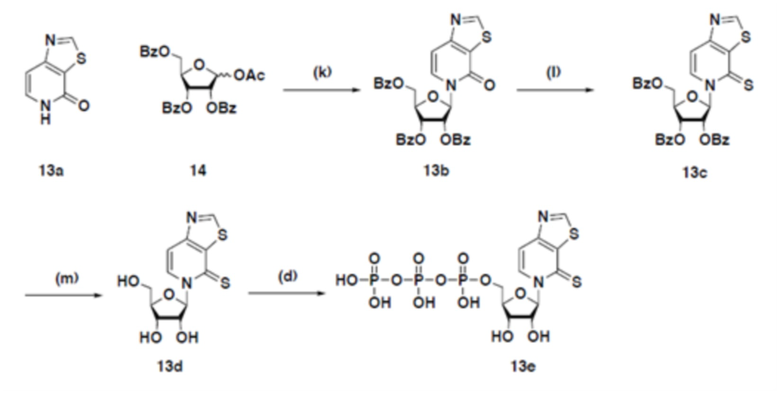
Схема S11. (k) N, O-бис(триметилсилил)ацетамид, SnCl4, CH3CN, RT, в течение ночи, (l) реагент Лавессона, 120°C, в течение ночи, (m) 30% NaOMe, DCM, 0°C, 2 ч., (d) протоновая губка, POCl3, Bu3N, Bu3NPPi, (MeO)3P, DMF, -15°C, 3 ч.
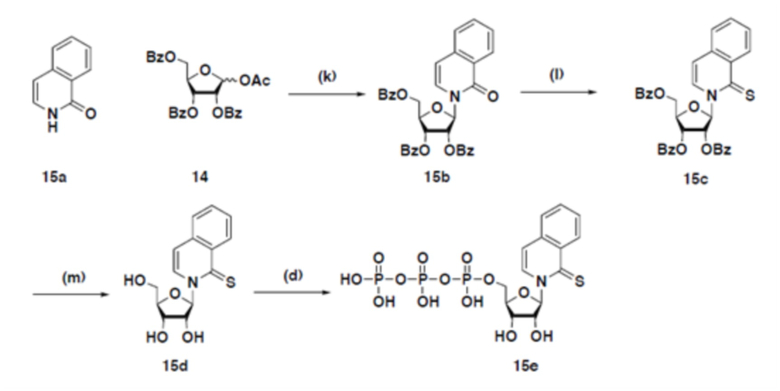
Схема S12. (n) i. Selectfluor, MeOH/CH3CN (1:1 об./об.), кипячение с обратным холодильником, 3 ч.; ii. TfOH/DCM (1:1 об./об.), 1 ч., (l) реагент Лавессона, 120°C, в течение ночи, (m) 30% NaOMe, DCM, 0°C, 2 ч., (d) протоновая губка, POCl3, Bu3N, Bu3NPPi, (MeO)3P, DMF, -15°C, 3 ч.
[00714] Пример 3:Оптимизация продукции неприродного белка.
[00715] В примерах 1A, 1B и 2 идентифицировали dCNMO-dTPT3 в качестве подходящего UBP для хранения информации, и TAT1TP и NaMTP или 5FMTP в качестве подходящих рибонуклеозидтрифосфатов для ее извлечения. После трансформации с использованием тех же плазмид, которые использовали выше, 10 мкМ dTPT3TP и 25 мкМ dCNMOTP добавляли в среды для выращивания, которые затем также дополняли TAT1TP в концентрациях в диапазоне от 100 мкМ до 12,5 мкМ и NaMTP или 5FMTP в концентрациях в диапазоне от 200 мкМ до 12,5 мкМ, все в сериях 2-кратных разведений, и после добавления 1 мМ AzK клетки индуцировали для экспрессии sfGFP. Общая флуоресценция sfGFP, наблюдаемая в клетках, дополненных TAT1TP и NaMTP, как правило, была выше, чем наблюдаемая в клетках, дополненных TAT1TP и 5FMTP (фиг. 6A и фиг. 6B). В обоих случаях флуоресценция являлась более высокой при более низких концентрациях NaMTP или 5FMTP (из-за повышенной продукции примесного природного sfGFP, см. ниже). Кроме того, клетки, как правило, демонстрировали более высокую флуоресценцию при более высоких концентрациях TAT1TP. Однако, из-за небольшого снижения роста, клетки, дополненные 100 мкМ TAT1TP, демонстрировали меньшую флуоресценцию, чем дополненные 50 мкМ TAT1TP. Продукцию белка снова количественно анализировали посредством анализа электрофоретического сдвига (фиг. 6C и фиг. 6D). Как правило, со снижением концентрации NaMTP ниже 200 мкМ, наблюдали постепенное снижение точности встраивания AzK в sfGFP, в то время как при использовании 5FMTP это снижение точности наблюдали только при концентрации ниже 50 мкМ. Очевидно более низкие концентрации 5FMTP можно использовать без нарушения точности. Клетки, дополненные высокой концентрации 5FMTP (≥50 мкМ), демонстрировали высокие белковые сдвиги при всех исследуемых концентрациях TAT1TP (100 мкМ, 50 мкМ, 25 мкМ или 12,5 мкМ). Однако если концентрация 5FMTP составляла 25 мкМ или менее, снижение концентрации TAT1TP приводило к снижению белкового сдвига. Если NaMTP добавляли в концентрации 200 мкМ, все исследуемые концентрации TAT1TP приводили к продукции белка с высокой точностью встраивания ncAA, но при более низких концентрациях NaMTP снижение концентрации TAT1TP снова приводило к снижению белкового сдвига. В целом, эти исследования показали, что комбинированной оптимизации чистоты и выхода белка достигали с использованием NaMTP и TAT1TP, добавляемых в концентрациях 200 мкМ и 50 мкМ, соответственно, или с использованием 5FMTP и TAT1TP, добавляемых в концентрации 50 мкМ. В терминах исключительно продукции белка использованием NaMTP и TAT1TP является оптимальным, в то время как использование 5FMTP и TAT1TP приводит к немного более низким выходам чистого ncAA-меченого белка, но требует значимо более низких концентрациях XTP.
[00716] Оптимизация условий трансляции in vivo. Ночную культуру SSO штамма ML2, несущего плазмиду pGEX-MbPylRS TetR, выращивали в 2×YT, дополненной 50 мМ фосфата калия, 5 мкг/мл хлорамфеникола и 100 мкг/мл карбенициллина (в настоящем описании обозначаемой в этом разделе как "среда"), затем разводили обратно до OD600 0,03 в той же среде и выращивали до OD600 0,4-0,6. Затем культуру охлаждали при встряхивании на ледяной бане в течение 15 мин, затем осаждали посредством центрифугирования при 3200×g в течение 10 мин. Затем клетки ресуспендировали и промывали ледяной ddH2O дважды, затем ресуспендировали в ледяной ddH2O до OD600 55-65. Электрокомпетентные клетки (50 мкл) и ~1 нг собранной плазмиды Golden Gate с UBP в генах sfGFP и тРНК-Pyl (см. Сборка плазмид Golden Gate) переносили в предварительно охлажденную кювету для электропорации (щель 0,2 см), затем осуществляли электропорацию (Gene Pulser II; Bio-Rad) по инструкциям производителя (напряжение 25 кВ, конденсатор 2,5 мкФ, резистор 200 Ом), а затем незамедлительно разводили 950 мкл среды. Затем аликвоту (40 мкл) смеси для трансформации разводили в пять раз в среде, дополненной dCNMOTP (25 мкМ) и dTPT3TP (10 мкМ), до конечного объема 200 мкл, затем ей позволяли восстанавливаться в течение 1 ч. с встряхиванием при 37°C. Двукратные разведения восстановленных клеток высевали на твердые среды, дополненные зеоцином (50 мкг/мл), dCNMOTP (10 мкМ), dTPT3TP (10 мкМ) и агаром (2% масс./об.), затем выращивали при 37°C в течение 18 ч. Отбирали отдельные колонии и выращивали в среде (300 мкл), дополненной 50 мкг/мл зеоцина и dCNMOTP (25 мкМ) и dTPT3TP (10 мкМ) (в настоящем описании обозначаемой в этом разделе как "среды для выращивания"). Культуру подвергали мониторингу роста клеток (многоканальный спектрофотометр для чтения планшетов Envision 2103 с фильтром 590/20 нм), а затем собирали при OD600 ~1,0. Аликвоту (50 мкл) подвергали выделению плазмиды с использованием набора ZR Plasmide Prep Kit (Zymo Research). Затем выделенные плазмиды подвергали анализу сдвига электрофоретической подвижности с использованием стрептавидина, описанному ниже (с использованием праймеров P3-4 и P5-6, таблица S1), для определения удержания UBP.
[00717] Затем колонию с исключительным удержанием UBP разводили до OD600 ~0,01 в средах для выращивания и выращивали до OD600 ~0,4-0,6. Затем эту культуру разделяли на множество культур по 300 мкл и в каждую добавляли 5FMTP, TAT1TP или NaMTP, TAT1TP в различных концентрациях каждого XTP (12,5 мкМ, 25 мкМ, 50 мкМ, 100 мкМ или 200 мкМ) и TAT1TP (12,5 мкМ, 25 мкМ, 50 мкМ, или 100 мкМ). В это время в клетки также добавляли AzK (10 мМ). Образцы закрывали от света после добавления AzK для предотвращения фотодеградации. Затем эти образцы выращивали при 37°C в течение 20 мин перед добавлением 1 мМ IPTG и выращивали при 37°C еще в течение 1 ч. для индуцирования RNAP T7 и транскрипции тРНК-Pyl и PylRS. С этого момента клетки подвергали мониторингу роста и флуоресценции. Затем экспрессию sfGFP индуцировали ангидротетрациклином (100 нг/мл). Еще через 3 ч. выращивания при 37°C клетки собирали (230 мкл для аффинной очистки sfGFP), охлаждали, а затем осаждали и хранили при -80°C перед очисткой белка.
[00718] Трансляция in vivo неприродных кодонов высокой плотности. Ночную культуру SSO штамма ML2, несущего плазмиду pGEX-MbPylRS TetR, выращивали в 2×YT, дополненной 50 мМ фосфата калия, 5 мкг/мл хлорамфеникола и 100 мкг/мл карбенициллина (в настоящем описании обозначаемой в этом разделе как "среда"), затем разводили обратно до OD600 0,03 в той же среде и выращивали до OD600 0,4-0,6. Затем культуру охлаждали со встряхиванием на ледяной бане в течение 15 минут, затем осаждали посредством центрифугирования при 3200×g в течение 10 мин. Затем клетки ресуспендировали и промывали ледяной ddH2O дважды, затем ресуспендировали в ледяной ddH2O до OD600 55-65. Электрокомпетентные клетки (50 мкл) и ~1 нг собранных плазмид Golden Gate с UBP в положении кодона 149 или 153 sfGFP, двух из трех валидированных положений кодона, или всех трех положениях (sfGFP149(AXC), sfGFP151(AXC), sfGFP149,151(AXC, AXC), sfGFP151,153(AXC, AXC), sfGFP149,153(AXC, AXC) и (sfGFP149151153(AXC, AXC,AXC)) (см. Сборка плазмид Golden Gate), переносили в предварительно охлажденную кювету для электропорации (щель 0,2 см). Затем клетки подвергали электропорации (Gene Pulser II; Bio-Rad) по инструкциям производителя (напряжение 25 кВ, конденсатор 2,5 мкФ, резистор 200 Ом), а затем незамедлительно разводили 950 мкл среды. Затем аликвоту (40 мкл) смеси для трансформации разводили в пять раз в среде, дополненной dNaMTP (150 мкМ) и dTPT3TP (10 мкМ) (далее в настоящем описании обозначаемые в этом разделе как "прежние условия") или dCNMOTP (150 мкМ) и dTPT3TP (10 мкМ) (далее в настоящем описании обозначаемые в этом разделе как "новые условия"), до конечного объема 200 мкл, затем ей позволяли восстанавливаться в течение 1 ч. с встряхиванием при 37°C. Двукратные разведения восстановленных клеток высевали на твердые среды, дополненные зеоцином (50 мкг/мл), соответствующими неприродными трифосфатами и агаром (2% масс./об.), затем выращивали при 37°C в течение ~18 ч. Отбирали отдельные колонии и выращивали в среде (300 мкл), дополненной 50 мкг/мл зеоцина (далее в настоящем описании обозначаемой в этом разделе как "среды для выращивания"), и добавляли соответствующие неприродные трифосфаты для прежних условий или новых условий. Культуру подвергали мониторингу роста клеток (многоканальный спектрофотометр для чтения планшетов Envision 2103 с фильтром 590/20 нм), а затем собирали при OD600 ~1,0. Собранные клетки охлаждали на льду в течение ночи.
[00719] Затем клетки повторно разводили до OD600 ~0,1 в средах для выращивания и добавляли соответствующие неприродные трифосфаты для прежних условий или новых условий, затем выращивали до OD600 ~0,4-0,6. Затем к клеткам, соответствующим прежним условиям, добавляли NaMTP (250 мкМ) и TPT3TP (30 мкМ), в то время как в клетки, соответствующие новым условиям, добавляли NaMTP (200 мкМ) и TAT1TP (50 мкМ). В это время в клетки также добавляли AzK (10 мМ) (или ddH2O для культур, выращенных в отсутствие AzK). Образцы закрывали от света после добавления AzK для предотвращения фотодеградации. Затем эти образцы выращивали при 37°C в течение 20 мин перед добавлением 1 мМ IPTG и выращивали при 37°C еще в течение 1 ч. для индуцирования RNAP T7 и транскрипции тРНК-Pyl и PylRS. С этого момента клетки подвергали мониторингу роста и флуоресценции. Затем экспрессию sfGFP индуцировали ангидротетрациклином (100 нг/мл). Еще через 3 ч. выращивания при 37°C клетки охлаждали, собирали (50 мкл для выделения плазмиды для определения удержания UBP, 230 мкл для аффинной очистки sfGFP), а затем осаждали и хранили при -80°C перед очисткой белка и оценкой удержания UBP.
[00720] Результаты. Все десять исследуемых XTP в высокой концентрации могут опосредовать продукцию белков с по меньшей мере умеренной точностью встраивания ncAA (от 98% до 63%). Как правило, со снижением концентраций XTP, точность встраивания ncAA снижалась, что свидетельствует о снижении точности, с которой транскрибируется неприродная мРНК. Это согласовывается со значительной флуоресценцией, наблюдаемой при удержании XTP.
[00721] В этом примере представлены SAR для транскрипции и трансляции преимущественно гидрофобных рибонуклеотидов. В случае XTP, если рассматривать NaMTP, PTMOTP и MTMOTP, сужение кольца и/или дериватизация гетероатома в некоторых случаях является неблагоприятной. Однако, в случае моноциклического нуклеинового основания XTP, за исключением ClMOTP, наблюдают более высокую точность продукции белка относительно MTMOTP и PTMOTP. В некоторых случаях специфические взаимодействия между неприродными нуклеиновыми основаниями или с полимеразой являются благоприятными для спаривания оснований. Например, замена в обоих 4- и 5-положениях моноциклических нуклеиновых оснований в некоторых случаях приводит к спариванию оснований. По сравнению с 2OMeTP, заместитель Cl или Br в 4-положении (ClMOTP и BrMOTP) в некоторых случаях является неблагоприятным, в то время как метильная группа в 4-положении (MMO2TP) снижает общую флуоресценцию, но с повышением точности белка. Нитрильный заместитель в 4-положении (CNMOTP) приводит к продукции наибольшего количества неприродного белка, а также умеренно повышает точность, с которой встраивается ncAA. Добавление фторного заместителя в 5-положении (5F2OMeTP) также повышает продукцию белка и точность относительно 2OMeTP. Эффекты замены в 4- и 5-положениях в некоторых случаях кажутся по меньшей мере приблизительно аддитивными, т.к. комбинирование заместителей 5-фтора и 4-метила (5FMTP) делает возможным высокий выход продукции чистого неприродного белка при более низких концентрациях относительно других неприродных трифосфатов. Однако, хоть и требуя более высоких концентраций, NaMTP обеспечивает сбалансированную комбинацию выхода и чистоты среди исследуемых аналогов XTP. В некоторых случаях SAR, полученные для аналогов XTP, значительно отличаются от полученных при репликации аналогов dXTP. Например, хотя dPTMOTP, dClMOTP и dCNMO в некоторых случаях являются более оптимальными, чем dNaMTP, ClMOTP и PTMOTP являются умеренно и значимо менее оптимальными в некоторых случаях, чем NaMTP. Кроме того, хотя dCNMO является наиболее оптимальным партнером для dTPT3, обнаруженным к настоящему времени, использование CNMOTP приводит к немного сниженной точности встраивания ncAA относительно NaMTP, хотя его использование не приводит к наибольшей продукции неприродного белка. В наиболее высокой концентрации все пять исследуемых YTP эффективно встраивались в антикодон тРНК-Pyl и могли опосредовать продукцию белков с по меньшей мере умеренной точностью встраивания ncAA (от 98% до 51%). Не желая быть связанными какой-либо теорией, данные об удержании UBP позволяют полагать, что точность продукции белка не является чувствительной к умеренной утрате неприродной тРНК, что позволяет предполагать, что в случае YTP, приводящих к меньшим сдвигам электрофоретической подвижности белков, транскрипция тРНК являлась весьма неэффективной или имела низкую точность. Все из TPT3TP, SICSTP, FSICSTP, и TAT1TP являлись по меньшей мере немного токсичными в наибольшей концентрации (данные не представлены), и совокупная флуоресценция клеток повышалась со снижением концентраций. В отличие от XTP и транскрипции мРНК, точность неприродного белка не снижалась, что позволяет предполагать, что повышенная продукция белка просто являлась результатом повышенного роста клеток. В отличие от этого, 5SICSTP демонстрировал минимальную токсичность (данные не представлены), и продукция неприродного белка и точность снижались со снижением концентраций снова, по-видимому, из-за значительно нарушенной продукции неприродной тРНК. При рассмотрении этого SAR YTP и использовании SICSTP в качестве референса, 7-фторный заместитель (FSICSTP) в некоторых случаях является неблагоприятным, и продуцируется лишь небольшое количество белка и с низкой точностью. Добавление метильной группы в 5-положении (5SICSTP) снижает токсичность, но также в некоторых случаях снижает продукцию неприродной тРНК. В некоторых случаях сужение кольца и дериватизация гетероатома (TPT3TP) улучшает продукцию белка и точность, но в высокой концентрации он зачастую токсичен по сравнению с другими аналогами YTP. Дальнейшая дериватизация гетероатома тиофенового кольца TPT3TP для получения тиазола TAT1TP приводит к продукции большего неприродного белка, чем в случае TPT3TP, со снижением токсичности. Учитывая количество продуцируемого белка и точность встраивания ncAA, в некоторых случаях TAT1TP используют в качестве YTP.
[00722] Пример 4: Хранение и извлечение неприродной информации более высокой плотности.
[00723] Оценивали хранение и извлечение информации из гена, имеющего более высокую плотность UBP. С этой целью валидировали способность SSO реплицировать ДНК, содержащую ген sfGFP с UBP, расположенными так, чтобы кодировать кодоны 149 или 153, каждый из которых отделен от кодона, описанного выше (кодона 151), одним природным кодоном. Таким образом, конструировали экспрессирующие плазмиды, как описано выше, но в которых последовательность AXC расположена так, чтобы кодировать кодон 149 или 153 (sfGFP149(AXC) или sfGFP153(AXC), соответственно). После трансформации ML2, клетки выращивали в присутствии неприродных нуклеозидтрифосфатов, соответствующих системе, ранее описанной авторами настоящего изобретения (дезоксирибонуклеотиды dNaMTP и dTPT3TP и рибонуклеотиды NaMTP и TPT3TP, обозначенные как dNaM-dTPT3/NaMTP, TPT3TP), или оптимизированной системе, обнаруженной в настоящем исследовании (dCNMOdTPT3/NaMTP, TAT1TP). Затем удержание UBP охарактеризовывали с помощью анализа электрофоретического сдвига с использованием стрептавидина, как описано выше. Высокое удержание соответствующей UBP в генах sfGFP149(AXC) и sfGFP153(AXC) (≥95%), а также в гене тРНК (≥91%) наблюдали в обоих условиях (данные не представлены).
[00724] Общая флуоресценция sfGFP, наблюдаемая через 3 ч. после индуцирования, позволяла выявить значимую продукцию белка с обеих конструкций в присутствии AzK в обоих условиях (фиг. 7A).
[00725] Однако, флуоресценция клеток, экспрессирующих конструкцию sfGFP149(AXC), в которые добавляли dCNMOdTPT3/NaM, TAT1, была на 58% большей, чем клетки, в которые добавляли dNaM-dTPT3/NaM, TPT3. В случае гена sfGFP153(AXC) наблюдали на 43% больше флуоресценции в случае dCNMOdTPT3/NaMTP, TAT1TP, чем dNaMdTPT3/NaMTP, TPT3TP. В обоих условиях наблюдали в приблизительно 2 раза более высокую флуоресценцию в случае sfGFP153(AXC), чем sfGFP149(AXC). Очищали белок и анализировали встраивание AzK, как описано выше (фиг. 7B). В случае dNaM-dTPT3/NaMTP, TPT3TP наблюдали белковые сдвиги 86% и 94% в случае sfGFP149(AXC) и sfGFP153(AXC), соответственно. Однако, в случае dCNMOdTPT3/NaMTP, TAT1TP наблюдали 96% сдвиг для белка, продуцируемого с любой из конструкций. Эти результаты четко демонстрируют, что два дополнительных положения кодона транскрибируются и транслируются эффективно, но опять же они транскрибируются и транслируются лучше в случае недавно идентифицированной системы dCNMO-dTPT3/NaMTP, TAT1TP. Конструировали экспрессирующие плазмиды с неприродным кодоном, одновременно кодируемым в двух или всех трех из исследуемых положений (sfGFP149,151(AXC, AXC), sfGFP151,153(AXC, AXC), sfGFP149,153(AXC, AXC) и sfGFP149151153(AXC, AXC,AXC), соответственно). Клетки ML2 подвергали трансформации, выращивали в присутствии dNaMdTPT3/NaMTP, TPT3TP или dCNMOdTPT3/NaMTP, TAT1TP и индуцировали экспрессию белка, как описано выше. Хотя удержание UBP в гене тРНК-Pyl(GYT) оставалось высоким (≥88%) во всех случаях (данные не представлены), анализ электрофоретического сдвига биотина с использованием генов мРНК показал комплексные и неинтерпретируемые паттерны полос, вероятно, по меньшей мере частично, из-за образования смеси комплексов с отдельными продуктами ПЦР, связанными с множеством стрептавидинов. Исследовали белок, получаемый посредством конъюгации с DBCO-TAMRA, как описано выше (фиг. 7B). Примечательно, что относительно сдвига, наблюдаемого в случае отдельной ncAA, наблюдали значимо дополнительно сдвинутую полосу в случае белков, экспрессирующихся с конструкций sfGFP149,151(AXC, AXC), sfGFP151,153(AXC, AXC) и sfGFP149,153(AXC, AXC), что свидетельствует о конъюгации двух молекул DBCO-TAMRA с sfGFP, несущим два остатка AzK. При анализе очищенных белков, экспрессируемых с dNaM-dTPT3/NaMTP, TPT3TP, количественный анализ этих двойных сдвинутых полос относительно общего sfGFP показал, что 80%, 87% и 83% белка, соответственно, имели два остатка AzK, и 20%, 13% или 9%, соответственно, имели один AzK при использовании конструкций sfGFP149,151(AXC, AXC), sfGFP151,153(AXC, AXC) или sfGFP149,153(AXC, AXC), соответственно. В случае dCNMOdTPT3/NaMTP, TAT1TP 81%, 89% и 93% белка имели два ncAA с sfGFP149,151(AXC, AXC), sfGFP151,153(AXC, AXC) и sfGFP149,153(AXC, AXC), соответственно, в то время как 19%, 11% и 6% имели один ncAA. Клетки, трансформированные с использованием конструкции sfGFP149,151,153(AXC, AXC,AXC), экспрессировали белок, демонстрирующий даже более сдвинутую полосу, что четко свидетельствует о встраивании трех остатков AzK. Количественный анализ каждой полосы относительно общего sfGFP показал, что использование dNaM-dTPT3/NaMTP, TPT3TP приводило к 39%, 24% и 33% белка, имеющего три, два и один ncAA, соответственно, и при этом флуоресценция и белковые сдвиги сильно варьировались (фигуры 7A и 7B). В отличие от этого, использование системы dCNMO-dTPT3/NaMTP, TAT1TP приводило к тому, что 90% продуцируемого белка имели все три ncAA, а остальная часть содержала два ncAA.
[00726] Для дальнейшей проверки успешного встраивания всех трех ncAA с использованием системы dCNMO-dTPT3/NaMTP, TAT1TP выделенные sfGFP анализировали посредством количественной масс-спектрометрии интактного белка. В кратком изложении, очищенные белки обессоливали с использованием устройств с центрифужным фильтром (AmiconR Ultra-0,5 - Millipore) и анализировали посредством HRMS (ESI-TOF). Затем полученные массовые спектры подвергали деконволюции с использованием программного обеспечения Waters MaxEnt 1, подтверждая, что они являлись количественными, после интегрирования пиков (данные не представлены). В соответствии с анализом сдвига электрофоретической подвижности, этот анализ показал, что 88% выделенного белка содержала ожидаемые три основания AzK, в то время как остальные 12% содержали два остатка AzK и один остаток Ile или Leu (фиг. 7C).
Результаты. В целом, с помощью SAR идентифицировали систему dCNMOdTPT3/NaMTP, TAT1TP как продуцирующую большие количества белка с высокой точностью встраивания ncAA. Использование dCNMOdTPT3/5FMTP, TAT1TP приводит к продукции белка с той же высокой точностью, и хотя она приводит к продукции немного меньшего количества белка, она требует использования значимо меньшего количества неприродных рибонуклеотидов. В некоторых случаях систему dCNMO-dTPT3/NaMTP, TAT1TP можно использовать для кодирования и извлечения неприродной информации более высокой плотности. С помощью обеих систем продуцировали белок с двумя ncAA с высокой точностью, но система dCNMOdTPT3/NaMTP, TAT1TP, как правило, продуцировала желаемый белок в более высоких количествах. Кроме того, при кодировании трех ncAA система dNaM-dTPT3/NaMTP, TPT3TP приводила к продукции белка с тройной меткой со значимо сниженной и более вариабельной точностью и выходом, в то время как точность и выход в случае системы dCNMO-dTPT3/NaMTP, TAT1TP оставались воспроизводимо высокими. Не желая быть связанными какой-либо теорией, полагают, что маловероятно, что примесь, в которой Ile или Leu заменяли одну ncAA, является результатом продукции неприродной тРНК, т.к. удержание UBP в гене тРНК являлось высоким и схожим для обеих систем, и даже при небольших различиях, данные о встраивании отдельного ncAA позволяют предполагать, что они не должны вызывать значительное снижение точности продукции неприродного белка. Также маловероятно, что это является результатом транскрипции мРНК, которая должна быть идентичной для двух систем (в обоих случаях dTPT3 направляет встраивание NaM в мРНК). Не желая быть связанными какой-либо теорией, полагают, что причиной примеси Leu/Ile, вероятно, является утрата UBP в гене мРНК во время репликации; это также соответствует наиболее распространенной ожидаемой мутации (dX в dT), которая будет приводить к образованию кодона Ile.
[00727] Описание всех публикаций, патентов, патентных заявок и опубликованных патентных заявок, на которые ссылаются в настоящем описании, указывая цитирование, таким образом, включено в настоящее описание в качестве ссылки в полном объеме.
Последовательности
SEQ ID NO:1. Последовательность экспрессирующей плазмиды pGEX-MbPylRS TetR PylRS (5923 п.н.)
GTAAATCACTGCATAATTCGTGTCGCTCAAGGCGCACTCCCGTTCTGGATAATGTTTTTTGCGCCGACAT
CATAACGGTTCTGGCAAATATTCTGAAATGAGCTGTTGACAATTAATCATCGGCTCGTATAATGTGTGGA
ATTGTGAGCGGATAACAATTTCACACAGGAAACAGTATTCATGGATAAAAAACCGCTGGACGTTCTGATC
TCCGCTACGGGTCTGTGGATGAGCCGCACGGGTACGCTGCATAAAATTAAACACCACGAAGTGTCACGTT
CGAAAATCTATATCGAAATGGCGTGCGGTGATCATCTGGTGGTTAACAATAGCCGTTCTTGTCGCACCGC
GCGTGCCTTTCGCCATCACAAATACCGCAAAACGTGCAAACGTTGTCGCGTGTCAGATGAAGACATTAAC
AATTTCCTGACCCGTAGTACGGAATCCAAAAACTCAGTGAAAGTTCGCGTCGTGAGTGCTCCGAAAGTTA
AAAAAGCGATGCCGAAAAGTGTCTCCCGTGCCCCGAAACCGCTGGAAAACTCAGTGTCGGCAAAAGCTTC
CACCAATACGAGCCGCTCTGTTCCGTCGCCGGCAAAAAGCACCCCGAACAGCTCTGTCCCGGCAAGCGCA
CCGGCACCGTCTCTGACGCGTAGTCAGCTGGATCGCGTGGAAGCCCTGCTGTCCCCGGAAGACAAAATCT
CACTGAATATGGCAAAACCGTTTCGTGAACTGGAACCGGAACTGGTTACCCGTCGCAAAAACGATTTCCA
ACGTCTGTATACGAATGATCGCGAAGACTACCTGGGTAAACTGGAACGTGATATCACCAAATTTTTCGTG
GACCGCGGCTTTCTGGAAATCAAATCTCCGATTCTGATCCCGGCTGAATATGTTGAACGCATGGGTATTA
ACAATGATACCGAACTGAGTAAACAGATTTTTCGTGTGGATAAAAACCTGTGCCTGCGGCCGATGCTGGC
ACCGACGCTGTATAATTACCTGCGTAAACTGGATCGCATTCTGCCGGGTCCGATTAAAATCTTTGAAGTG
GGCCCGTGTTATCGTAAAGAATCGGATGGCAAAGAACACCTGGAAGAATTTACCATGGTTAACTTCTGCC
AAATGGGCAGCGGTTGTACGCGCGAAAATCTGGAAGCGCTGATCAAAGAATTCCTGGATTACCTGGAAAT
CGACTTCGAAATCGTCGGTGATTCTTGCATGGTGTATGGCGATACCCTGGACATCATGCATGGTGACCTG
GAACTGAGTTCCGCTGTTGTCGGTCCGGTCAGCCTGGATCGTGAATGGGGCATTGACAAACCGTGGATCG
GCGCGGGTTTTGGCCTGGAACGCCTGCTGAAAGTTATGCACGGCTTCAAAAACATCAAACGTGCGTCTCG
CTCGGAATCGTATTACAACGGCATCTCAACCAATCTGTAATAATGACTGACGATCTGCCTCGCGCGTTTC
GGTGATGACGGTGAAAACCTCTGACACATGCAGCTCCCGGAGACGGTCACAGCTTGTCTGTAAGCGGATG
CCGGGAGCAGACAAGCCCGTCAGGGCGCGTCAGCGGGTGTTGGCGGGTGTCGGGGCGCAGCCATGACCCA
GTCACGTAGCGATAGCGGAGTGTATAATTCTTGAAGACGAAAGGGCCTCGTGATACGCCTATTTTTATAG
GTTAATGTCATGATAATAATGGTTTCTTAGACGTCAGGTGGCACTTTTCGGGGAAATGTGCGCGGAACCC
CTATTTGTTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAATAACCCTGATAAATGCT
TCAATAATATTGAAAAAGGAAGAGTATGAGTATTCAACATTTCCGTGTCGCCCTTATTCCCTTTTTTGCG
GCATTTTGCCTTCCTGTTTTTGCTCACCCAGAAACGCTGGTGAAAGTAAAAGATGCTGAAGATCAGTTGG
GTGCACGAGTGGGTTACATCGAACTGGATCTCAACAGCGGTAAGATCCTTGAGAGTTTTCGCCCCGAAGA
ACGTTTTCCAATGATGAGCACTTTTAAAGTTCTGCTATGTGGCGCGGTATTATCCCGTGTTGACGCCGGG
CAAGAGCAACTCGGTCGCCGCATACACTATTCTCAGAATGACTTGGTTGAGTACTCACCAGTCACAGAAA
AGCATCTTACGGATGGCATGACAGTAAGAGAATTATGCAGTGCTGCCATAACCATGAGTGATAACACTGC
GGCCAACTTACTTCTGACAACGATCGGAGGACCGAAGGAGCTAACCGCTTTTTTGCACAACATGGGGGAT
CATGTAACTCGCCTTGATCGTTGGGAACCGGAGCTGAATGAAGCCATACCAAACGACGAGCGTGACACCA
CGATGCCTGCAGCAATGGCAACAACGTTGCGCAAACTATTAACTGGCGAACTACTTACTCTAGCTTCCCG
GCAACAATTAATAGACTGGATGGAGGCGGATAAAGTTGCAGGACCACTTCTGCGCTCGGCCCTTCCGGCT
GGCTGGTTTATTGCTGATAAATCTGGAGCCGGTGAGCGTGGGTCTCGCGGTATCATTGCAGCACTGGGGC
CAGATGGTAAGCCCTCCCGTATCGTAGTTATCTACACGACGGGGAGTCAGGCAACTATGGATGAACGAAA
TAGACAGATCGCTGAGATAGGTGCCTCACTGATTAAGCATTGGTAACTGTCAGACCAAGTTTACTCATAT
ATACTTTAGATTGATTTAAAACTTCATTTTTAATTTAAAAGGATCTAGGTGAAGATCCTTTTTGATAATC
TCATGACCAAAATCCCTTAACGTGAGTTTTCGTTCCACTGAGCGTCAGACCCCGTAGAAAAGATCAAAGG
ATCTTCTTGAGATCCTTTTTTTCTGCGCGTAATCTGCTGCTTGCAAACAAAAAAACCACCGCTACCAGCG
GTGGTTTGTTTGCCGGATCAAGAGCTACCAACTCTTTTTCCGAAGGTAACTGGCTTCAGCAGAGCGCAGA
TACCAAATACTGTCCTTCTAGTGTAGCCGTAGTTAGGCCACCACTTCAAGAACTCTGTAGCACCGCCTAC
ATACCTCGCTCTGCTAATCCTGTTACCAGTGGCTGCTGCCAGTGGCGATAAGTCGTGTCTTACCGGGTTG
GACTCAAGACGATAGTTACCGGATAAGGCGCAGCGGTCGGGCTGAACGGGGGGTTCGTGCACACAGCCCA
GCTTGGAGCGAACGACCTACACCGAACTGAGATACCTACAGCGTGAGCTATGAGAAAGCGCCACGCTTCC
CGAAGGGAGAAAGGCGGACAGGTATCCGGTAAGCGGCAGGGTCGGAACAGGAGAGCGCACGAGGGAGCTT
CCAGGGGGAAACGCCTGGTATCTTTATAGTCCTGTCGGGTTTCGCCACCTCTGACTTGAGCGTCGATTTT
TGTGATGCTCGTCAGGGGGGCGGAGCCTATGGAAAAACGCCAGCAACGCGGCCTTTTTACGGTTCCTGGC
CTTTTGCTGGCCTTTTGCTCACATGTTCTTTCCTGCGTTATCCCCGAGCTCTTAGCGCGAATTGTCGAGG
GAAATTTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAATAACCCTGATAAATGCTTCAATA
ATATTAAATATGGCTGGTTCTCGCAGAAAGAAACATATCCATGAAATCCCGCCCCGAATTGATATGTCCA
GATTAGATAAAAGTAAAGTGATTAACAGCGCATTAGAGCTGCTTAATGAGGTCGGAATCGAAGGTTTAAC
AACCCGTAAACTCGCCCAGAAGCTAGGTGTAGAGCAGCCTACATTGTATTGGCATGTAAAAAATAAGCGG
GCTTTGCTCGACGCCTTAGCCATTGAGATGTTAGATAGGCACCATACTCACTTTTGCCCTTTAGAAGGGG
AAAGCTGGCAAGATTTTTTACGTAATAACGCTAAAAGTTTTAGATGTGCTTTACTAAGTCATCGCGATGG
AGCAAAAGTACATTTAGGTACACGGCCTACAGAAAAACAGTATGAAACTCTCGAAAATCAATTAGCCTTT
TTATGCCAACAAGGTTTTTCACTAGAGAATGCATTATATGCACTCAGCGCTGTGGGGCATTTTACTTTAG
GTTGCGTATTGGAAGATCAAGAGCATCAAGTCGCTAAAGAAGAAAGGGAAACACCTACTACTGATAGTAT
GCCGCCATTATTACGACAAGCTATCGAATTATTTGATCACCAAGGTGCAGAGCCAGCCTTCTTATTCGGC
CTTGAATTGATCATTTGCGGATTAGAAAAACAACTTAAATGTGAAAGTGGGTCTTAAGCACTAGGTCTAG
GGCGGCGGATTTGTCCTACTCAGGAGAGCGTTCACCGACAAACAACAGATAAAACGAAAGGCCCAGTCTT
TCGACTGAGCCTTTCGTTTTATTTGATGCCTCTAGCACGCGTAGAGCTAGAGCCTTCAACCCAGTCAGCT
CCTTCCGGTGGGCGCGGGGCATGACTAACATGAGAATTACAACTTATATCGTATGGGGCTGACTTCAGGT
GCTACATTTGAAGAGATAAATTGCACTGAAATCTAGATGATTCTGTGGATAACCGTATTACCGCCTTTGA
GTGAGCTGATACCGCTCGCCGCAGCCGAACGACCGAGCGCAGCGAGTCAGTGAGCGAGGAAGCGGAAGAG
CGCCTGATGCGGTATTTTCTCCTTACGCATCTGTGCGGTATTTCACACCGCATAAATTCCGACACCATCG
AATGGTGCAAAACCTTTCGCGGTATGGCATGATAGCGCCCGGAAGAGAGTCAATTCAGGGTGGTGAATGT
GAAACCAGTAACGTTATACGATGTCGCAGAGTATGCCGGTGTCTCTTATCAGACCGTTTCCCGCGTGGTG
AACCAGGCCAGCCACGTTTCTGCGAAAACGCGGGAAAAAGTGGAAGCGGCGATGGCGGAGCTGAATTACA
TTCCCAACCGCGTGGCACAACAACTGGCGGGCAAACAGTCGTTGCTGATTGGCGTTGCCACCTCCAGTCT
GGCCCTGCACGCGCCGTCGCAAATTGTCGCGGCGATTAAATCTCGCGCCGATCAACTGGGTGCCAGCGTG
GTGGTGTCGATGGTAGAACGAAGCGGCGTCGAAGCCTGTAAAGCGGCGGTGCACAATCTTCTCGCGCAAC
GCGTCAGTGGGCTGATCATTAACTATCCGCTGGATGACCAGGATGCCATTGCTGTGGAAGCTGCCTGCAC
TAATGTTCCGGCGTTATTTCTTGATGTCTCTGACCAGACACCCATCAACAGTATTATTTTCTCCCATGAA
GACGGTACGCGACTGGGCGTGGAGCATCTGGTCGCATTGGGTCACCAGCAAATCGCGCTGTTAGCGGGCC
CATTAAGTTCTGTCTCGGCGCGTCTGCGTCTGGCTGGCTGGCATAAATATCTCACTCGCAATCAAATTCA
GCCGATAGCGGAACGGGAAGGCGACTGGAGTGCCATGTCCGGTTTTCAACAAACCATGCAAATGCTGAAT
GAGGGCATCGTTCCCACTGCGATGCTGGTTGCCAACGATCAGATGGCGCTGGGCGCAATGCGCGCCATTA
CCGAGTCCGGGCTGCGCGTTGGTGCGGATATCTCGGTAGTGGGATACGACGATACCGAAGACAGCTCATG
TTATATCCCGCCGTTAACCACCATCAAACAGGATTTTCGCCTGCTGGGGCAAACCAGCGTGGACCGCTTG
CTGCAACTCTCTCAGGGCCAGGCGGTGAAGGGCAATCAGCTGTTGCCCGTCTCACTGGTGAAAAGAAAAA
CCACCCTGGCGCCCAATACGCAAACCGCCTCTCCCCGCGCGTTGGCCGATTCATTAATGCAGCTGGCACG
ACAGGTTTCCCGACTGGAAAGCGGGCAGTGAGCGCAACGCAAT
SEQ ID NO: 2. Последовательность принимающей плазмиды p[sfGFP(gg)151; тРНК-Pyl(gg)] GG для экспрессии sfGFP и тРНК-Pyl (3101 п.н.)
TAACTAGCATAACCCCTTGGGGCCTCTAAACGGGTCTTGAGGGGTTTTTTGCTGAAAGGAGGAACTATAT
CCGGATTGGTTAATACGACTCACTATAGGGGAATTGTGAGCGGATAACAATTCCCCTCTAGAAAAGCATT
GGAAACCGAGACCGGTACCGGTCTCTTAGATTCCCGGGGTTTCCGCCAAATTCGAAAAGCCTGCTCAACG
AGCAGGCTTTTTTGCATCTAGCATAACCCCTTGGGGCCTCTAAACGGGTCTTGAGGGGTTTTTTGCCTGA
ACGAGCAGGCTTTTTTGCATAAGCTTCCTAGTGGCAGCGGCTAACTAAGCGGCCTGCTGACTTTCTCGCC
GATCAAAAGGCATTTTGCTATTAAGGGATTGACGAGGGCGTATCTGCGCAGTAAGATGCGCCCCGCATTG
GAGACGCCATGGCGTCTCGGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACT
TGAAAAAGTGGCACCGAGTCGGTGCTTTTTTTAATTCGAAAAGCCTGCTCAACGAGCAGGCTTTTTTGGT
CGACAGTAGTGGCAGCGGCTAACTAAGCGGCCTGCTGACTTTCTCGCCGATCAAAAGGCATTTTGCTATT
AAGGGATTGACGAGGGCGTATCTGCGCAGTAAGATGCGCCCCGCATGAGACGGCATGCCGTCTCTAGAGC
TAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTTT
TTTAATTCGAAAAGCCTGCTCAACGAGCAGGCTTTTTTGGTCGACAGTTCATAGGTGATTGCGGATCCCG
TCGTTGACAATTAATCATCGGCATAGTATATCGGCATAGTATAATACGACAAGGTGAGGAACTAAACCAT
GGCCAAGTTGACCAGTGCCGTTCCGGTGCTCACCGCGCGCGACGTCGCCGGAGCGGTCGAGTTCTGGACC
GACCGGCTCGGGTTCTCCCGCGACTTCGTGGAGGACGACTTCGCCGGTGTGGTCCGGGACGACGTGACCC
TGTTCATCAGCGCGGTCCAGGACCAGGTGGTGCCGGACAACACCCTGGCCTGGGTGTGGGTGCGCGGCCT
GGACGAGCTGTACGCCGAGTGGTCGGAGGTCGTGTCCACGAACTTCCGGGACGCCTCCGGGCCGGCCATG
ACCGAGATCGGCGAGCAGCCGTGGGGGCGGGAGTTCGCCCTGCGCGACCCGGCCGGCAACTGCGTGCACT
TCGTGGCCGAGGAGCAGGACTGAGAGCTCGCTTGGACTCCTGTTGATAGATCCAGTAATGACCTCAGAAC
TCCATCTGGATTTGTTCAGAACGCTCGGTTGCCGCCGGGCGTTTTTTATTGGTGAGAATCCAAGCACTAG
CTAGTAACAACTTATATCGTATGGGGCTGACTTCAGGTGCTACATTTGAAGAGATAAATTGCACTGAAAT
CTAGTAATATTTTATCTGATTAATAAGATGATCTTCTTGAGATCGTTTTGGTCTGCGCGTAATCTCTTGC
TCTGAAAACGAAAAAACCGCCTTGCAGGGCGGTTTTTCGAAGGTTCTCTGAGCTACCAACTCTTTGAACC
GAGGTAACTGGCTTGGAGGAGCGCAGTCACCAAAACTTGTCCTTTCAGTTTAGCCTTAACCGGCGCATGA
CTTCAAGACTAACTCCTCTAAATCAATTACCAGTGGCTGCTGCCAGTGGTGCTTTTGCATGTCTTTCCGG
GTTGGACTCAAGACGATAGTTACCGGATAAGGCGCAGCGGTCGGACTGAACGGGGGGTTCGTGCATACAG
TCCAGCTTGGAGCGAACTGCCTACCCGGAACTGAGTGTCAGGCGTGGAATGAGACAAACGCGGCCATAAC
AGCGGAATGACACCGGTAAACCGAAAGGCAGGAACAGGAGAGCGCACGAGGGAGCCGCCAGGGGGAAACG
CCTGGTATCTTTATAGTCCTGTCGGGTTTCGCCACCACTGATTTGAGCGTCAGATTTCGTGATGCTTGTC
AGGGGGGCGGAGCCTATGGAAAAACGGCTTTGCCGCGGCCCTCTCACTTCCCTGTTAAGTATCTTCCTGG
CATCTTCCAGGAAATCTCCGCCCCGTTCGTAAGCCATTTCCGCTCGCCGCAGTCGAACGACCGAGCGTAG
CGAGTCAGTGAGCGAGGAAGCGGAATATATCCCTTAATACGACTCACTATAGGGTCCCTATCAGTGATAG
AGAGGTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACATATGTCGAAAGGCGAAGAACTGTT
TACGGGAGTGGTGCCTATCCTGGTAGAGCTCGACGGAGATGTAAACGGTCACAAATTTTCAGTCCGCGGG
GAAGGCGAAGGCGATGCGACCAACGGTAAATTAACTTTGAAGTTTATTTGCACCACCGGCAAATTACCGG
TGCCTTGGCCGACGCTTGTGACGACCCTGACTTACGGGGTGCAGTGTTTCAGTCGCTACCCAGATCACAT
GAAACGCCATGACTTCTTCAAATCTGCGATGCCGGAAGGCTATGTGCAGGAACGTACAATTAGCTTTAAA
GACGACGGCACGTATAAAACGCGGGCAGAGGTTAAATTTGAGGGAGATACCCTGGTAAACCGTATTGAAC
TGAAAGGCATCGATTTTAAAGAAGATGGGAACATCTTGGGCCACAAGAGACCGGTACCGGTCTCGGAATC
AAAGCAAATTTCAAGATCCGTCATAACGTGGAGGACGGTTCCGTGCAGCTTGCAGATCACTATCAGCAGA
ATACGCCGATTGGCGATGGCCCGGTGCTGCTGCCCGATAATCACTACCTCTCTACTCAGAGTGTTTTATC
GAAAGACCCGAACGAGAAGCGTGATCACATGGTGCTGCTTGAATTTGTTACCGCGGCAGGTATTACACAC
GGCATGGATGAGTTGTATAAGGGATCCGCTTGGAGCCACCCGCAGTTCGAGAAAGGTGGAGGTTCCGGAG
GTGGATCGGGAGGTTCGGCGTGGAGCCACCCGCAGTTCGAAAAATAAAAGCTTAATTAGCTGAGCTTGGA
CTCCCTGCCACCGCTGAGCAA
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> THE SCRIPPS RESEARCH INSTITUTE
<120> РЕАГЕНТЫ И СПОСОБЫ ДЛЯ РЕПЛИКАЦИИ, ТРАНСКРИПЦИИ И ТРАНСЛЯЦИИ
В ПОЛУСИНТЕТИЧЕСКИХ ОРГАНИЗМАХ
<130> 36271-808.601
<140>
<141>
<150> 62/861,901
<151> 2019-06-14
<160> 20
<170> PatentIn version 3.5
<210> 1
<211> 5923
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1
gtaaatcact gcataattcg tgtcgctcaa ggcgcactcc cgttctggat aatgtttttt 60
gcgccgacat cataacggtt ctggcaaata ttctgaaatg agctgttgac aattaatcat 120
cggctcgtat aatgtgtgga attgtgagcg gataacaatt tcacacagga aacagtattc 180
atggataaaa aaccgctgga cgttctgatc tccgctacgg gtctgtggat gagccgcacg 240
ggtacgctgc ataaaattaa acaccacgaa gtgtcacgtt cgaaaatcta tatcgaaatg 300
gcgtgcggtg atcatctggt ggttaacaat agccgttctt gtcgcaccgc gcgtgccttt 360
cgccatcaca aataccgcaa aacgtgcaaa cgttgtcgcg tgtcagatga agacattaac 420
aatttcctga cccgtagtac ggaatccaaa aactcagtga aagttcgcgt cgtgagtgct 480
ccgaaagtta aaaaagcgat gccgaaaagt gtctcccgtg ccccgaaacc gctggaaaac 540
tcagtgtcgg caaaagcttc caccaatacg agccgctctg ttccgtcgcc ggcaaaaagc 600
accccgaaca gctctgtccc ggcaagcgca ccggcaccgt ctctgacgcg tagtcagctg 660
gatcgcgtgg aagccctgct gtccccggaa gacaaaatct cactgaatat ggcaaaaccg 720
tttcgtgaac tggaaccgga actggttacc cgtcgcaaaa acgatttcca acgtctgtat 780
acgaatgatc gcgaagacta cctgggtaaa ctggaacgtg atatcaccaa atttttcgtg 840
gaccgcggct ttctggaaat caaatctccg attctgatcc cggctgaata tgttgaacgc 900
atgggtatta acaatgatac cgaactgagt aaacagattt ttcgtgtgga taaaaacctg 960
tgcctgcggc cgatgctggc accgacgctg tataattacc tgcgtaaact ggatcgcatt 1020
ctgccgggtc cgattaaaat ctttgaagtg ggcccgtgtt atcgtaaaga atcggatggc 1080
aaagaacacc tggaagaatt taccatggtt aacttctgcc aaatgggcag cggttgtacg 1140
cgcgaaaatc tggaagcgct gatcaaagaa ttcctggatt acctggaaat cgacttcgaa 1200
atcgtcggtg attcttgcat ggtgtatggc gataccctgg acatcatgca tggtgacctg 1260
gaactgagtt ccgctgttgt cggtccggtc agcctggatc gtgaatgggg cattgacaaa 1320
ccgtggatcg gcgcgggttt tggcctggaa cgcctgctga aagttatgca cggcttcaaa 1380
aacatcaaac gtgcgtctcg ctcggaatcg tattacaacg gcatctcaac caatctgtaa 1440
taatgactga cgatctgcct cgcgcgtttc ggtgatgacg gtgaaaacct ctgacacatg 1500
cagctcccgg agacggtcac agcttgtctg taagcggatg ccgggagcag acaagcccgt 1560
cagggcgcgt cagcgggtgt tggcgggtgt cggggcgcag ccatgaccca gtcacgtagc 1620
gatagcggag tgtataattc ttgaagacga aagggcctcg tgatacgcct atttttatag 1680
gttaatgtca tgataataat ggtttcttag acgtcaggtg gcacttttcg gggaaatgtg 1740
cgcggaaccc ctatttgttt atttttctaa atacattcaa atatgtatcc gctcatgaga 1800
caataaccct gataaatgct tcaataatat tgaaaaagga agagtatgag tattcaacat 1860
ttccgtgtcg cccttattcc cttttttgcg gcattttgcc ttcctgtttt tgctcaccca 1920
gaaacgctgg tgaaagtaaa agatgctgaa gatcagttgg gtgcacgagt gggttacatc 1980
gaactggatc tcaacagcgg taagatcctt gagagttttc gccccgaaga acgttttcca 2040
atgatgagca cttttaaagt tctgctatgt ggcgcggtat tatcccgtgt tgacgccggg 2100
caagagcaac tcggtcgccg catacactat tctcagaatg acttggttga gtactcacca 2160
gtcacagaaa agcatcttac ggatggcatg acagtaagag aattatgcag tgctgccata 2220
accatgagtg ataacactgc ggccaactta cttctgacaa cgatcggagg accgaaggag 2280
ctaaccgctt ttttgcacaa catgggggat catgtaactc gccttgatcg ttgggaaccg 2340
gagctgaatg aagccatacc aaacgacgag cgtgacacca cgatgcctgc agcaatggca 2400
acaacgttgc gcaaactatt aactggcgaa ctacttactc tagcttcccg gcaacaatta 2460
atagactgga tggaggcgga taaagttgca ggaccacttc tgcgctcggc ccttccggct 2520
ggctggttta ttgctgataa atctggagcc ggtgagcgtg ggtctcgcgg tatcattgca 2580
gcactggggc cagatggtaa gccctcccgt atcgtagtta tctacacgac ggggagtcag 2640
gcaactatgg atgaacgaaa tagacagatc gctgagatag gtgcctcact gattaagcat 2700
tggtaactgt cagaccaagt ttactcatat atactttaga ttgatttaaa acttcatttt 2760
taatttaaaa ggatctaggt gaagatcctt tttgataatc tcatgaccaa aatcccttaa 2820
cgtgagtttt cgttccactg agcgtcagac cccgtagaaa agatcaaagg atcttcttga 2880
gatccttttt ttctgcgcgt aatctgctgc ttgcaaacaa aaaaaccacc gctaccagcg 2940
gtggtttgtt tgccggatca agagctacca actctttttc cgaaggtaac tggcttcagc 3000
agagcgcaga taccaaatac tgtccttcta gtgtagccgt agttaggcca ccacttcaag 3060
aactctgtag caccgcctac atacctcgct ctgctaatcc tgttaccagt ggctgctgcc 3120
agtggcgata agtcgtgtct taccgggttg gactcaagac gatagttacc ggataaggcg 3180
cagcggtcgg gctgaacggg gggttcgtgc acacagccca gcttggagcg aacgacctac 3240
accgaactga gatacctaca gcgtgagcta tgagaaagcg ccacgcttcc cgaagggaga 3300
aaggcggaca ggtatccggt aagcggcagg gtcggaacag gagagcgcac gagggagctt 3360
ccagggggaa acgcctggta tctttatagt cctgtcgggt ttcgccacct ctgacttgag 3420
cgtcgatttt tgtgatgctc gtcagggggg cggagcctat ggaaaaacgc cagcaacgcg 3480
gcctttttac ggttcctggc cttttgctgg ccttttgctc acatgttctt tcctgcgtta 3540
tccccgagct cttagcgcga attgtcgagg gaaatttttt ctaaatacat tcaaatatgt 3600
atccgctcat gagacaataa ccctgataaa tgcttcaata atattaaata tggctggttc 3660
tcgcagaaag aaacatatcc atgaaatccc gccccgaatt gatatgtcca gattagataa 3720
aagtaaagtg attaacagcg cattagagct gcttaatgag gtcggaatcg aaggtttaac 3780
aacccgtaaa ctcgcccaga agctaggtgt agagcagcct acattgtatt ggcatgtaaa 3840
aaataagcgg gctttgctcg acgccttagc cattgagatg ttagataggc accatactca 3900
cttttgccct ttagaagggg aaagctggca agatttttta cgtaataacg ctaaaagttt 3960
tagatgtgct ttactaagtc atcgcgatgg agcaaaagta catttaggta cacggcctac 4020
agaaaaacag tatgaaactc tcgaaaatca attagccttt ttatgccaac aaggtttttc 4080
actagagaat gcattatatg cactcagcgc tgtggggcat tttactttag gttgcgtatt 4140
ggaagatcaa gagcatcaag tcgctaaaga agaaagggaa acacctacta ctgatagtat 4200
gccgccatta ttacgacaag ctatcgaatt atttgatcac caaggtgcag agccagcctt 4260
cttattcggc cttgaattga tcatttgcgg attagaaaaa caacttaaat gtgaaagtgg 4320
gtcttaagca ctaggtctag ggcggcggat ttgtcctact caggagagcg ttcaccgaca 4380
aacaacagat aaaacgaaag gcccagtctt tcgactgagc ctttcgtttt atttgatgcc 4440
tctagcacgc gtagagctag agccttcaac ccagtcagct ccttccggtg ggcgcggggc 4500
atgactaaca tgagaattac aacttatatc gtatggggct gacttcaggt gctacatttg 4560
aagagataaa ttgcactgaa atctagatga ttctgtggat aaccgtatta ccgcctttga 4620
gtgagctgat accgctcgcc gcagccgaac gaccgagcgc agcgagtcag tgagcgagga 4680
agcggaagag cgcctgatgc ggtattttct ccttacgcat ctgtgcggta tttcacaccg 4740
cataaattcc gacaccatcg aatggtgcaa aacctttcgc ggtatggcat gatagcgccc 4800
ggaagagagt caattcaggg tggtgaatgt gaaaccagta acgttatacg atgtcgcaga 4860
gtatgccggt gtctcttatc agaccgtttc ccgcgtggtg aaccaggcca gccacgtttc 4920
tgcgaaaacg cgggaaaaag tggaagcggc gatggcggag ctgaattaca ttcccaaccg 4980
cgtggcacaa caactggcgg gcaaacagtc gttgctgatt ggcgttgcca cctccagtct 5040
ggccctgcac gcgccgtcgc aaattgtcgc ggcgattaaa tctcgcgccg atcaactggg 5100
tgccagcgtg gtggtgtcga tggtagaacg aagcggcgtc gaagcctgta aagcggcggt 5160
gcacaatctt ctcgcgcaac gcgtcagtgg gctgatcatt aactatccgc tggatgacca 5220
ggatgccatt gctgtggaag ctgcctgcac taatgttccg gcgttatttc ttgatgtctc 5280
tgaccagaca cccatcaaca gtattatttt ctcccatgaa gacggtacgc gactgggcgt 5340
ggagcatctg gtcgcattgg gtcaccagca aatcgcgctg ttagcgggcc cattaagttc 5400
tgtctcggcg cgtctgcgtc tggctggctg gcataaatat ctcactcgca atcaaattca 5460
gccgatagcg gaacgggaag gcgactggag tgccatgtcc ggttttcaac aaaccatgca 5520
aatgctgaat gagggcatcg ttcccactgc gatgctggtt gccaacgatc agatggcgct 5580
gggcgcaatg cgcgccatta ccgagtccgg gctgcgcgtt ggtgcggata tctcggtagt 5640
gggatacgac gataccgaag acagctcatg ttatatcccg ccgttaacca ccatcaaaca 5700
ggattttcgc ctgctggggc aaaccagcgt ggaccgcttg ctgcaactct ctcagggcca 5760
ggcggtgaag ggcaatcagc tgttgcccgt ctcactggtg aaaagaaaaa ccaccctggc 5820
gcccaatacg caaaccgcct ctccccgcgc gttggccgat tcattaatgc agctggcacg 5880
acaggtttcc cgactggaaa gcgggcagtg agcgcaacgc aat 5923
<210> 2
<211> 3101
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 2
taactagcat aaccccttgg ggcctctaaa cgggtcttga ggggtttttt gctgaaagga 60
ggaactatat ccggattggt taatacgact cactataggg gaattgtgag cggataacaa 120
ttcccctcta gaaaagcatt ggaaaccgag accggtaccg gtctcttaga ttcccggggt 180
ttccgccaaa ttcgaaaagc ctgctcaacg agcaggcttt tttgcatcta gcataacccc 240
ttggggcctc taaacgggtc ttgaggggtt ttttgcctga acgagcaggc ttttttgcat 300
aagcttccta gtggcagcgg ctaactaagc ggcctgctga ctttctcgcc gatcaaaagg 360
cattttgcta ttaagggatt gacgagggcg tatctgcgca gtaagatgcg ccccgcattg 420
gagacgccat ggcgtctcgg ttttagagct agaaatagca agttaaaata aggctagtcc 480
gttatcaact tgaaaaagtg gcaccgagtc ggtgcttttt ttaattcgaa aagcctgctc 540
aacgagcagg cttttttggt cgacagtagt ggcagcggct aactaagcgg cctgctgact 600
ttctcgccga tcaaaaggca ttttgctatt aagggattga cgagggcgta tctgcgcagt 660
aagatgcgcc ccgcatgaga cggcatgccg tctctagagc tagaaatagc aagttaaaat 720
aaggctagtc cgttatcaac ttgaaaaagt ggcaccgagt cggtgctttt tttaattcga 780
aaagcctgct caacgagcag gcttttttgg tcgacagttc ataggtgatt gcggatcccg 840
tcgttgacaa ttaatcatcg gcatagtata tcggcatagt ataatacgac aaggtgagga 900
actaaaccat ggccaagttg accagtgccg ttccggtgct caccgcgcgc gacgtcgccg 960
gagcggtcga gttctggacc gaccggctcg ggttctcccg cgacttcgtg gaggacgact 1020
tcgccggtgt ggtccgggac gacgtgaccc tgttcatcag cgcggtccag gaccaggtgg 1080
tgccggacaa caccctggcc tgggtgtggg tgcgcggcct ggacgagctg tacgccgagt 1140
ggtcggaggt cgtgtccacg aacttccggg acgcctccgg gccggccatg accgagatcg 1200
gcgagcagcc gtgggggcgg gagttcgccc tgcgcgaccc ggccggcaac tgcgtgcact 1260
tcgtggccga ggagcaggac tgagagctcg cttggactcc tgttgataga tccagtaatg 1320
acctcagaac tccatctgga tttgttcaga acgctcggtt gccgccgggc gttttttatt 1380
ggtgagaatc caagcactag ctagtaacaa cttatatcgt atggggctga cttcaggtgc 1440
tacatttgaa gagataaatt gcactgaaat ctagtaatat tttatctgat taataagatg 1500
atcttcttga gatcgttttg gtctgcgcgt aatctcttgc tctgaaaacg aaaaaaccgc 1560
cttgcagggc ggtttttcga aggttctctg agctaccaac tctttgaacc gaggtaactg 1620
gcttggagga gcgcagtcac caaaacttgt cctttcagtt tagccttaac cggcgcatga 1680
cttcaagact aactcctcta aatcaattac cagtggctgc tgccagtggt gcttttgcat 1740
gtctttccgg gttggactca agacgatagt taccggataa ggcgcagcgg tcggactgaa 1800
cggggggttc gtgcatacag tccagcttgg agcgaactgc ctacccggaa ctgagtgtca 1860
ggcgtggaat gagacaaacg cggccataac agcggaatga caccggtaaa ccgaaaggca 1920
ggaacaggag agcgcacgag ggagccgcca gggggaaacg cctggtatct ttatagtcct 1980
gtcgggtttc gccaccactg atttgagcgt cagatttcgt gatgcttgtc aggggggcgg 2040
agcctatgga aaaacggctt tgccgcggcc ctctcacttc cctgttaagt atcttcctgg 2100
catcttccag gaaatctccg ccccgttcgt aagccatttc cgctcgccgc agtcgaacga 2160
ccgagcgtag cgagtcagtg agcgaggaag cggaatatat cccttaatac gactcactat 2220
agggtcccta tcagtgatag agaggtctag aaataatttt gtttaacttt aagaaggaga 2280
tatacatatg tcgaaaggcg aagaactgtt tacgggagtg gtgcctatcc tggtagagct 2340
cgacggagat gtaaacggtc acaaattttc agtccgcggg gaaggcgaag gcgatgcgac 2400
caacggtaaa ttaactttga agtttatttg caccaccggc aaattaccgg tgccttggcc 2460
gacgcttgtg acgaccctga cttacggggt gcagtgtttc agtcgctacc cagatcacat 2520
gaaacgccat gacttcttca aatctgcgat gccggaaggc tatgtgcagg aacgtacaat 2580
tagctttaaa gacgacggca cgtataaaac gcgggcagag gttaaatttg agggagatac 2640
cctggtaaac cgtattgaac tgaaaggcat cgattttaaa gaagatggga acatcttggg 2700
ccacaagaga ccggtaccgg tctcggaatc aaagcaaatt tcaagatccg tcataacgtg 2760
gaggacggtt ccgtgcagct tgcagatcac tatcagcaga atacgccgat tggcgatggc 2820
ccggtgctgc tgcccgataa tcactacctc tctactcaga gtgttttatc gaaagacccg 2880
aacgagaagc gtgatcacat ggtgctgctt gaatttgtta ccgcggcagg tattacacac 2940
ggcatggatg agttgtataa gggatccgct tggagccacc cgcagttcga gaaaggtgga 3000
ggttccggag gtggatcggg aggttcggcg tggagccacc cgcagttcga aaaataaaag 3060
cttaattagc tgagcttgga ctccctgcca ccgctgagca a 3101
<210> 3
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический олигонуклеотид
<400> 3
ctcgagtaca actttaactc acacaatgta tagatcacgg cagacaaaca aaagaatgga 60
atc 63
<210> 4
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический олигонуклеотид
<400> 4
ctcgagtaca actttaactc acacaatgta tacatcacgg cagacaaaca aaagaatgga 60
atc 63
<210> 5
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический олигонуклеотид
<220>
<221> модифицированное_основание
<222> (32)..(32)
<223> Неприродный нуклеотид
<400> 5
ctcgagtaca actttaactc acacaatgta ancatcacgg cagacaaaca aaagaatgga 60
atc 63
<210> 6
<211> 52
<212> ДНК
<213> Methanosarcina mazei
<400> 6
gaatctaacc cggctgaacg gatttagagt ccgttcgatc tacatgatca gg 52
<210> 7
<211> 52
<212> ДНК
<213> Methanosarcina mazei
<400> 7
gaatctaacc cggctgaacg gatttacagt ccgttcgatc tacatgatca gg 52
<210> 8
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический олигонуклеотид
<220>
<221> модифицированное_основание
<222> (26)..(26)
<223> Неприродный нуклеотид
<400> 8
gaatctaacc cggctgaacg gattancagt ccgttcgatc tacatgatca gg 52
<210> 9
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический олигонуклеотид
<220>
<221> модифицированное_основание
<222> (26)..(26)
<223> Неприродный нуклеотид
<400> 9
ctcgagtaca actttaactc acacancgta tacatcacgg cagacaaaca aaagaatgga 60
atc 63
<210> 10
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический олигонуклеотид
<220>
<221> модифицированное_основание
<222> (38)..(38)
<223> Неприродный нуклеотид
<400> 10
ctcgagtaca actttaactc acacaatgta tacatcancg cagacaaaca aaagaatgga 60
atc 63
<210> 11
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический олигонуклеотид
<220>
<221> модифицированное_основание
<222> (26)..(26)
<223> Неприродный нуклеотид
<220>
<221> модифицированное_основание
<222> (32)..(32)
<223> Неприродный нуклеотид
<400> 11
ctcgagtaca actttaactc acacancgta ancatcacgg cagacaaaca aaagaatgga 60
atc 63
<210> 12
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический олигонуклеотид
<220>
<221> модифицированное_основание
<222> (32)..(32)
<223> Неприродный нуклеотид
<220>
<221> модифицированное_основание
<222> (38)..(38)
<223> Неприродный нуклеотид
<400> 12
ctcgagtaca actttaactc acacaatgta ancatcancg cagacaaaca aaagaatgga 60
atc 63
<210> 13
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический олигонуклеотид
<220>
<221> модифицированное_основание
<222> (26)..(26)
<223> Неприродный нуклеотид
<220>
<221> модифицированное_основание
<222> (38)..(38)
<223> Неприродный нуклеотид
<400> 13
ctcgagtaca actttaactc acacancgta tacatcancg cagacaaaca aaagaatgga 60
atc 63
<210> 14
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический олигонуклеотид
<220>
<221> модифицированное_основание
<222> (26)..(26)
<223> Неприродный нуклеотид
<220>
<221> модифицированное_основание
<222> (32)..(32)
<223> Неприродный нуклеотид
<220>
<221> модифицированное_основание
<222> (38)..(38)
<223> Неприродный нуклеотид
<400> 14
ctcgagtaca actttaactc acacancgta ancatcancg cagacaaaca aaagaatgga 60
atc 63
<210> 15
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический праймер
<400> 15
atgggtctca cacaaactcg agtacaactt taactcacac 40
<210> 16
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический праймер
<400> 16
atgggtctcg attccattct tttgtttgtc tgc 33
<210> 17
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический праймер
<400> 17
atgggtctcg aaacctgatc atgtagatcg aacgg 35
<210> 18
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический праймер
<400> 18
atgggtctca tctaacccgg ctgaacgg 28
<210> 19
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический праймер
<400> 19
ctcgagtaca actttaactc acac 24
<210> 20
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический праймер
<400> 20
gattccattc ttttgtttgt ctgc 24
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ НА ОСНОВЕ НЕПРИРОДНЫХ ПАР ОСНОВАНИЙ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2799441C2 |
| КОНЪЮГАТЫ ЦИТОКИНОВ ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ | 2018 |
|
RU2787201C2 |
| ГЕНЕТИЧЕСКИ ЗАПРОГРАММИРОВАННАЯ ЭКСПРЕССИЯ БЕЛКОВ, СОДЕРЖАЩИХ НЕ ВСТРЕЧАЮЩУЮСЯ В ПРИРОДЕ АМИНОКИСЛОТУ ФЕНИЛСЕЛЕНОЦИСТЕИН | 2007 |
|
RU2464315C2 |
| КОНЪЮГАТЫ ЦИТОКИНОВ ДЛЯ ЛЕЧЕНИЯ ПРОЛИФЕРАТИВНЫХ И ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ | 2018 |
|
RU2786116C2 |
| Конъюгаты IL-15 и пути их применения | 2019 |
|
RU2793754C2 |
| АМИНОАЦИЛ-тРНК-СИНТЕТАЗА ДЛЯ ЭФФЕКТИВНОГО ВВЕДЕНИЯ ПРОИЗВОДНОГО ЛИЗИНА В БЕЛОК | 2020 |
|
RU2799794C2 |
| КОНЪЮГАТЫ ИНТЕРЛЕЙКИНА 10 И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2815891C2 |
| ГЕНЕТИЧЕСКИ КОДИРУЕМЫЕ ФЛЮОРЕСЦЕНТНЫЕ КУМАРИНОВЫЕ АМИНОКИСЛОТЫ | 2007 |
|
RU2473690C2 |
| СЕЛЕКТИВНАЯ ПОСТТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ ЭКСПОНИРУЕМЫХ ФАГАМИ ПОЛИПЕПТИДОВ | 2006 |
|
RU2459866C2 |
| ГИСТИДИЛ-тРНК-СИНТЕТАЗЫ ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ И ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2659423C2 |
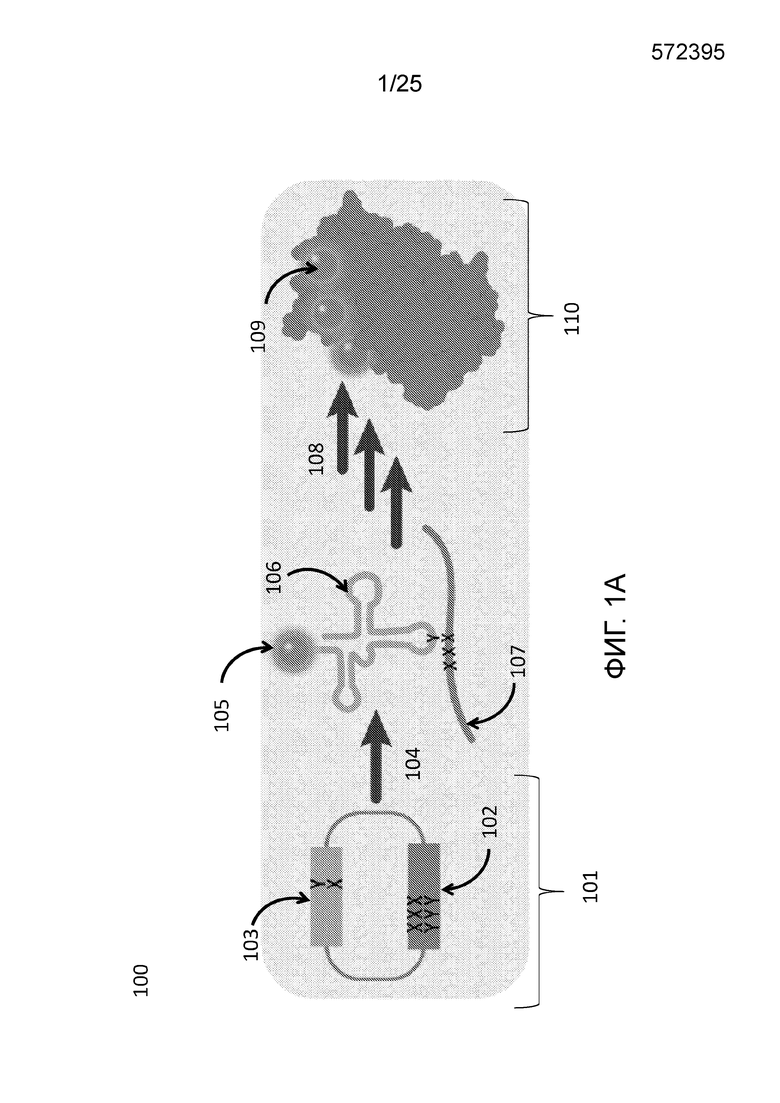
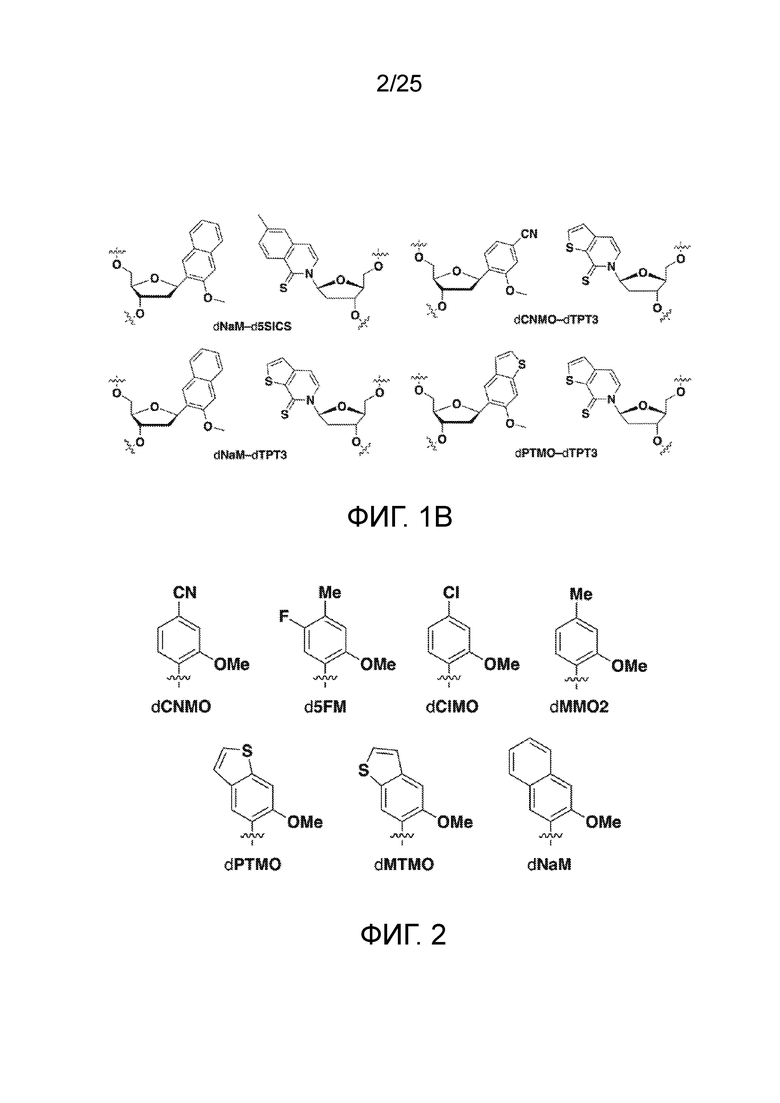
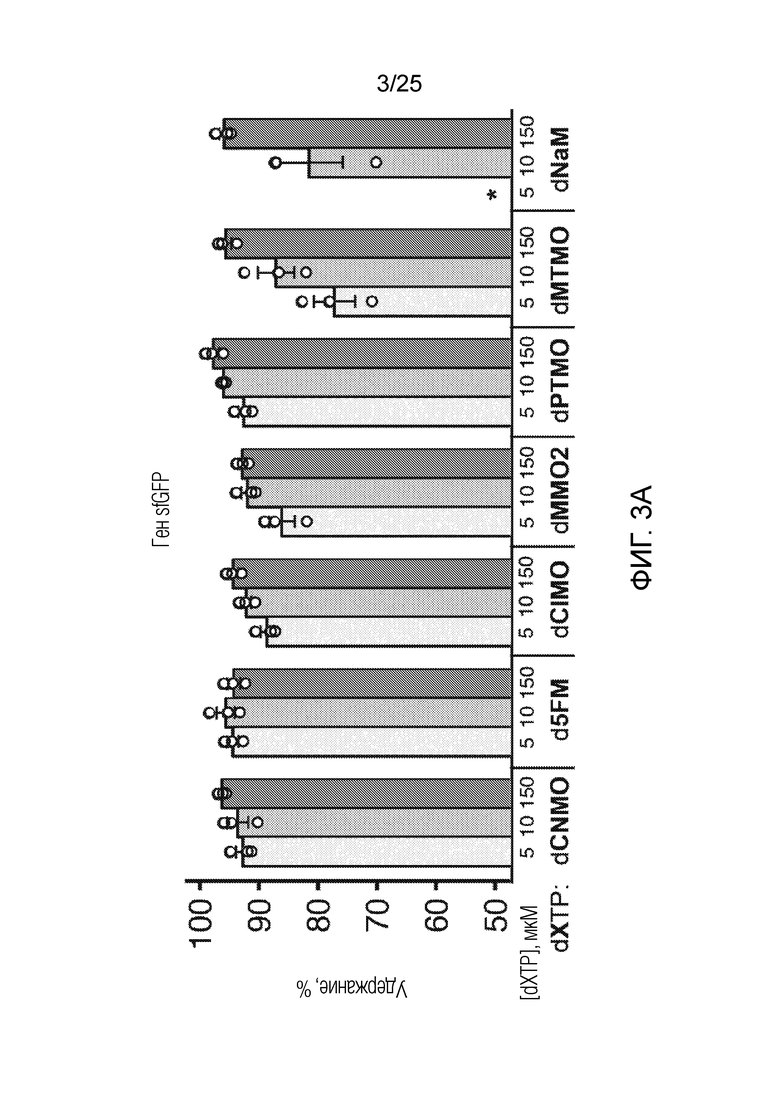
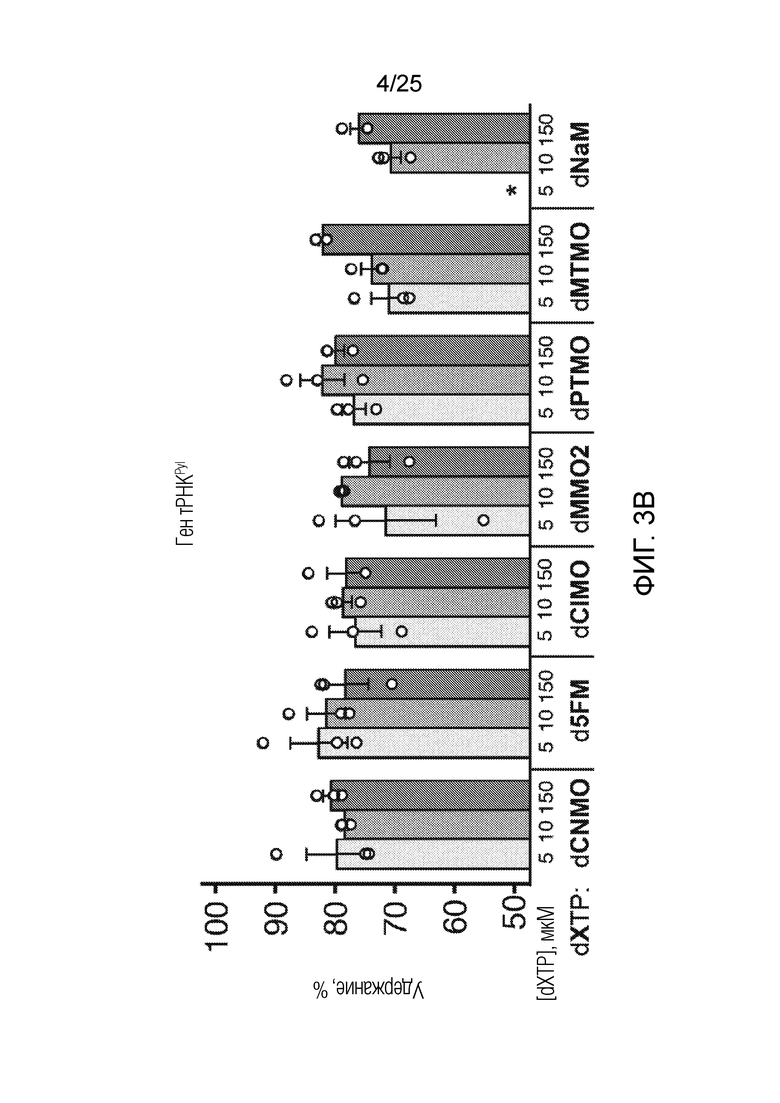

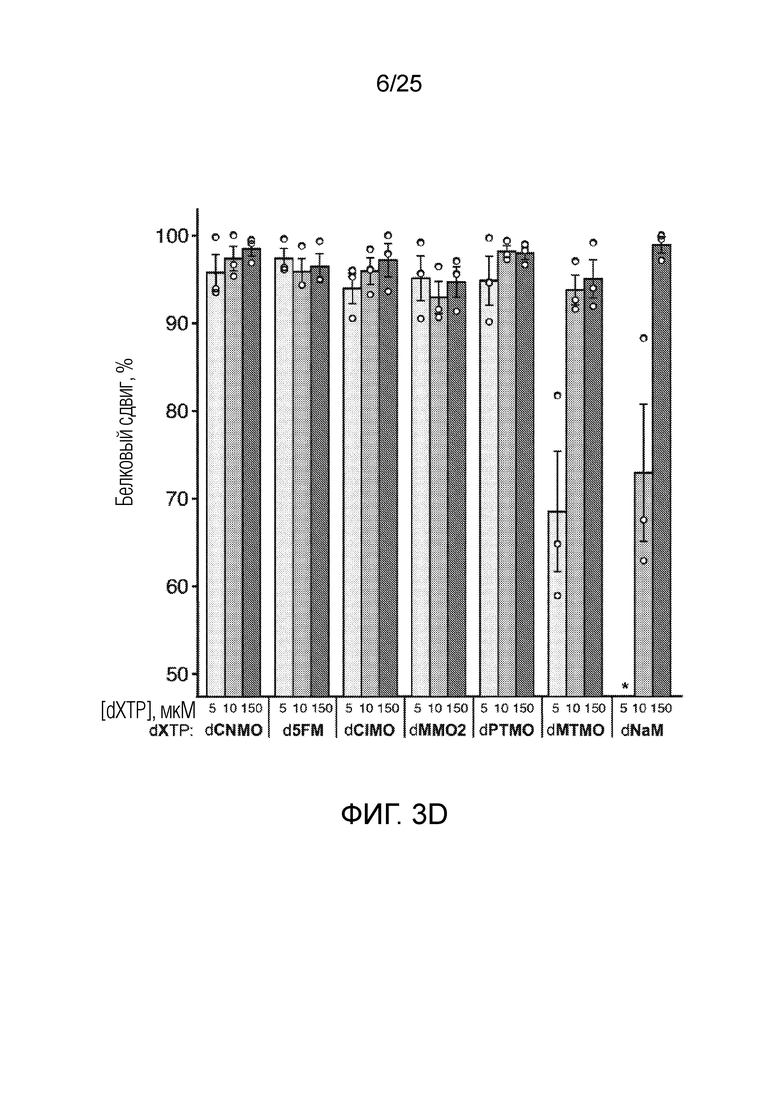
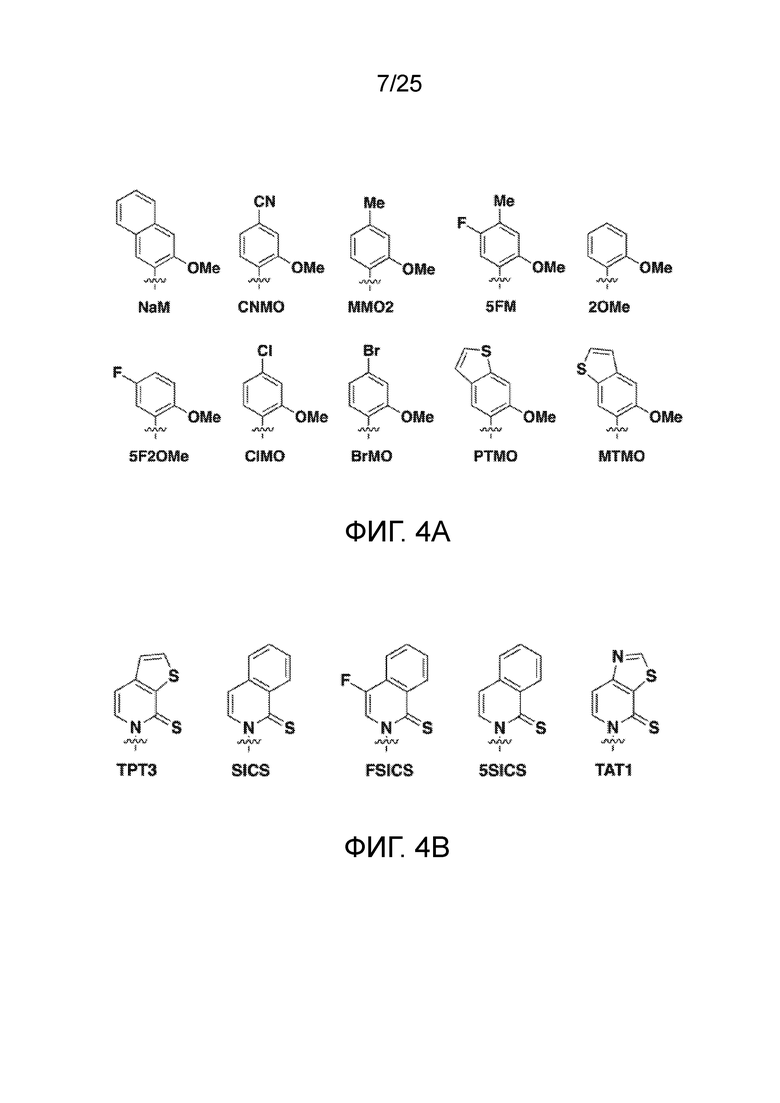
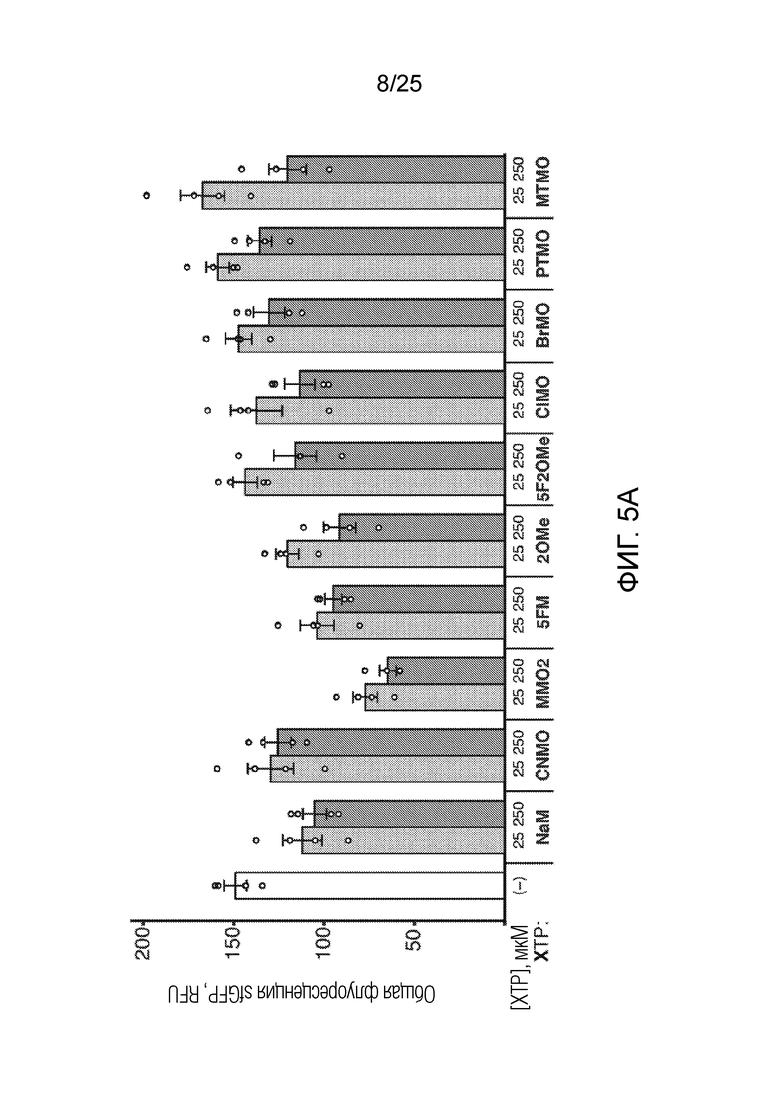
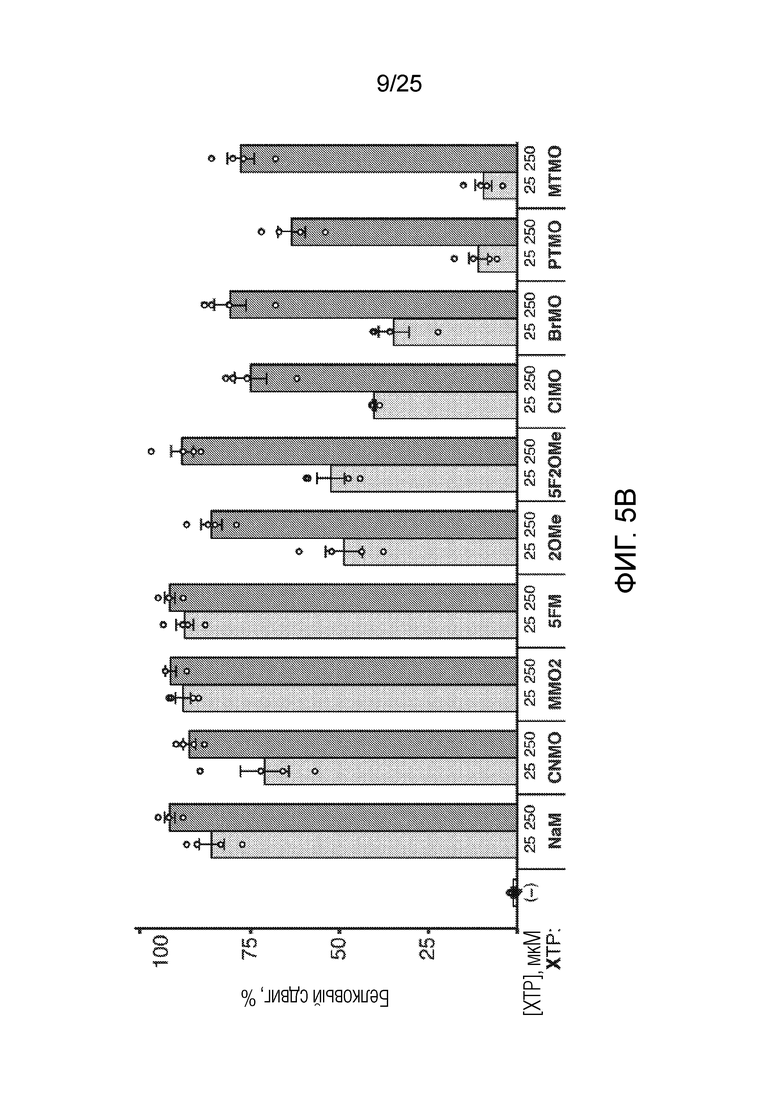
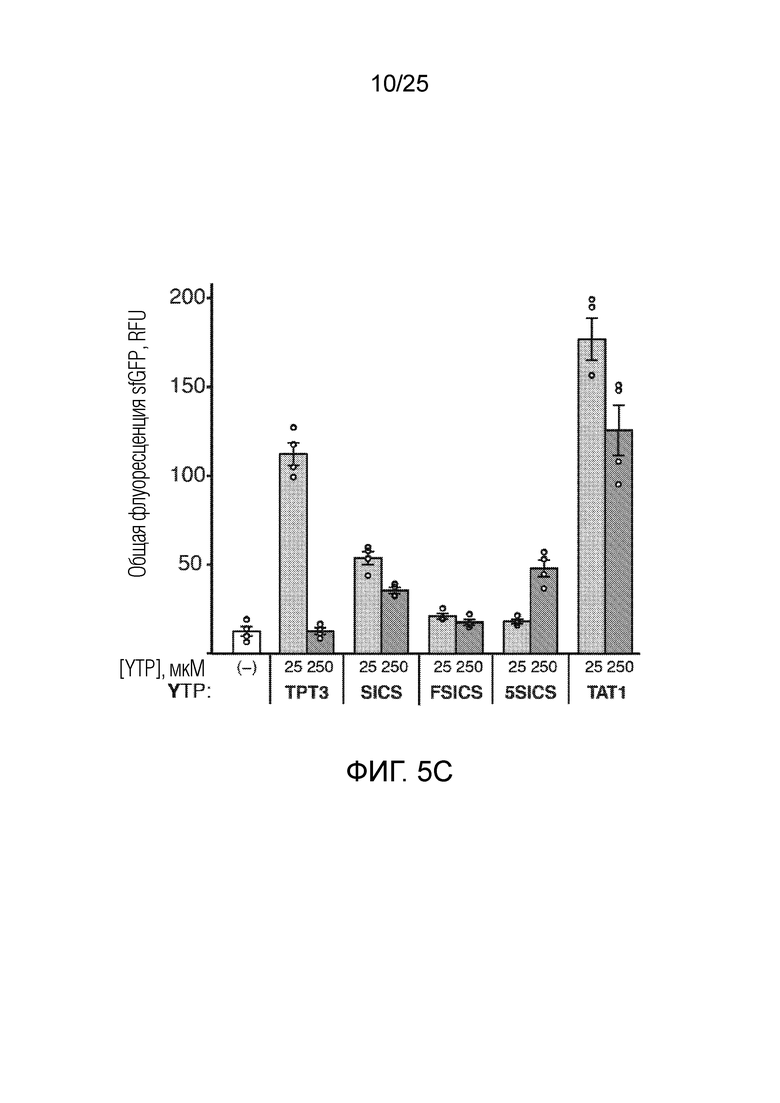
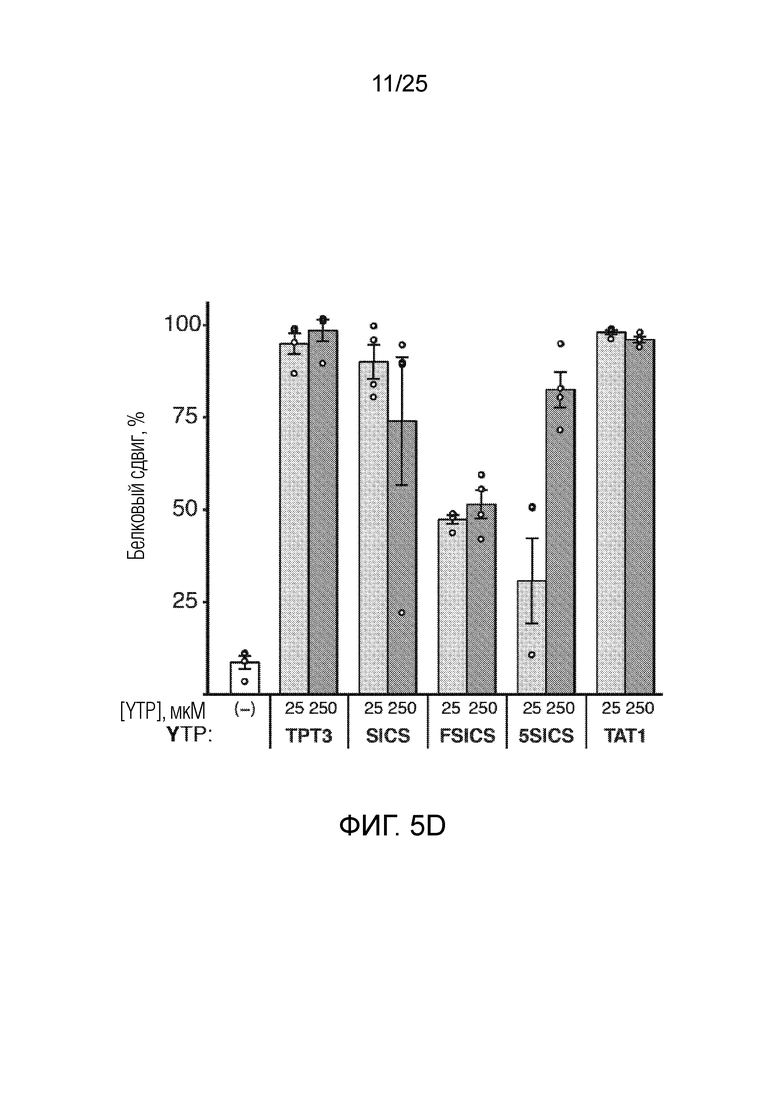
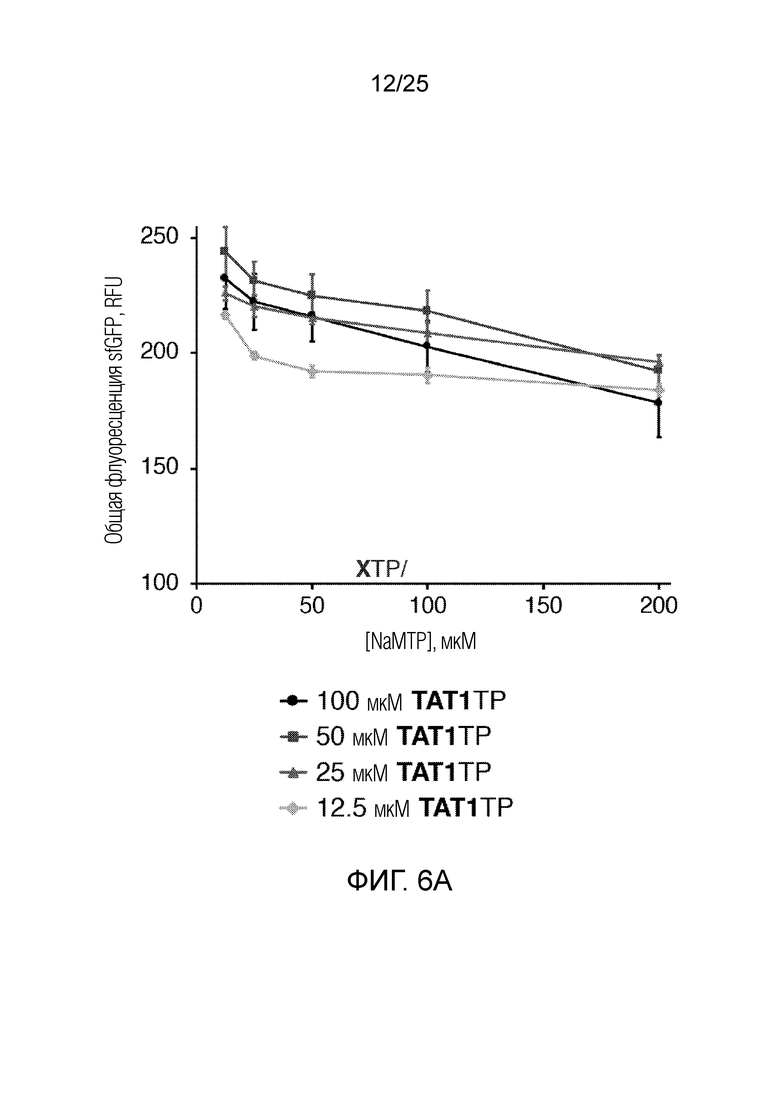
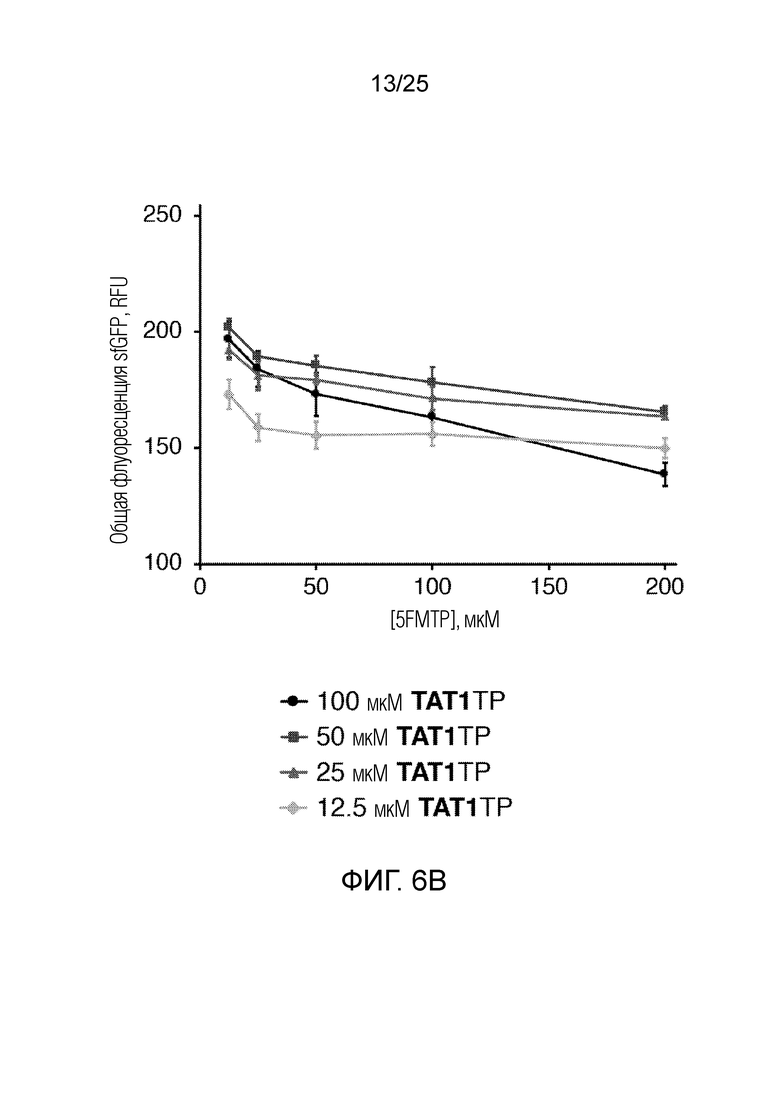
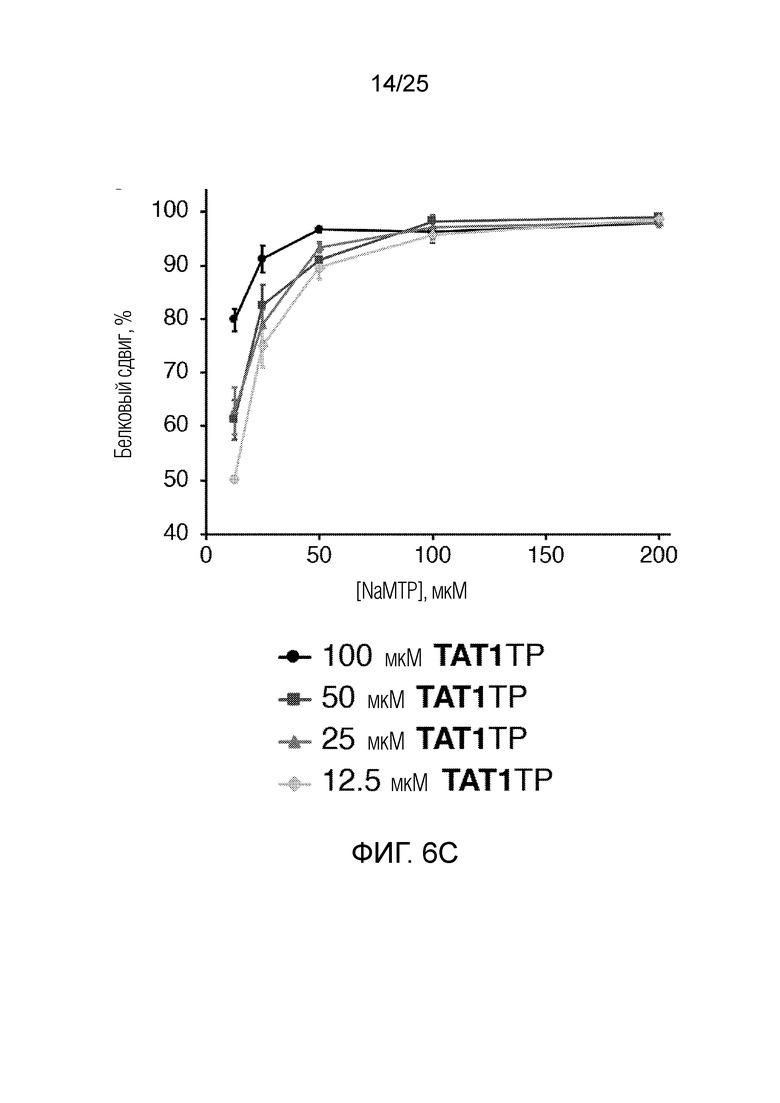
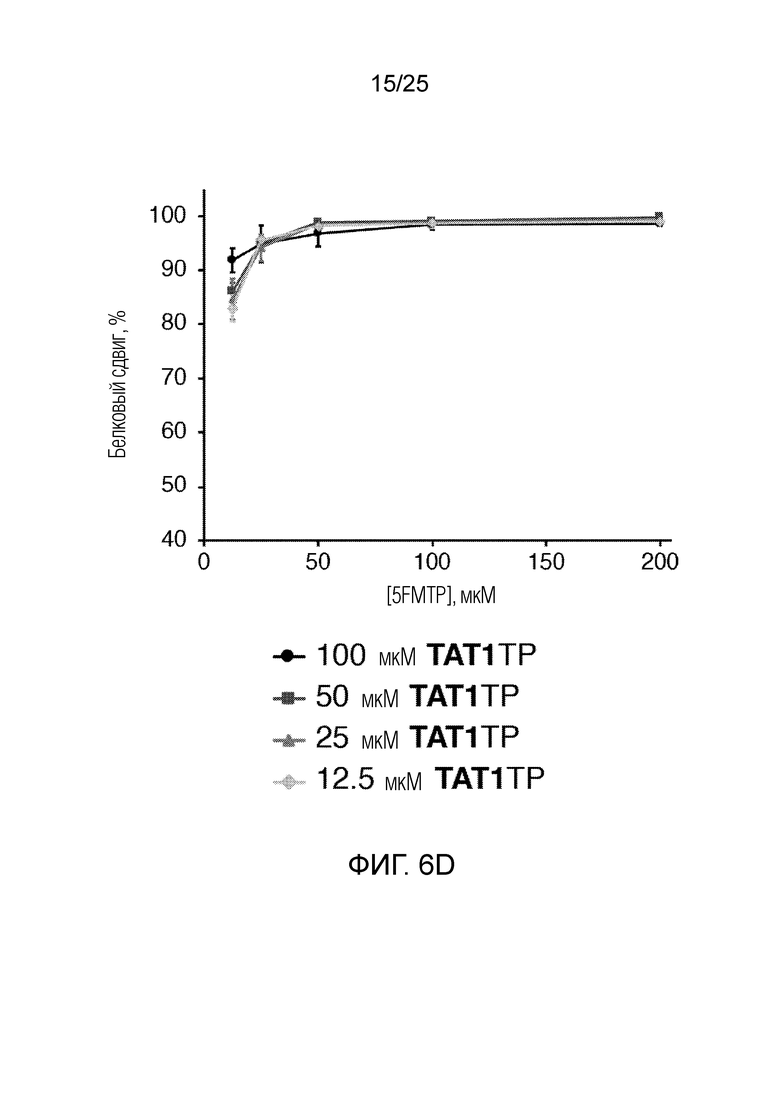
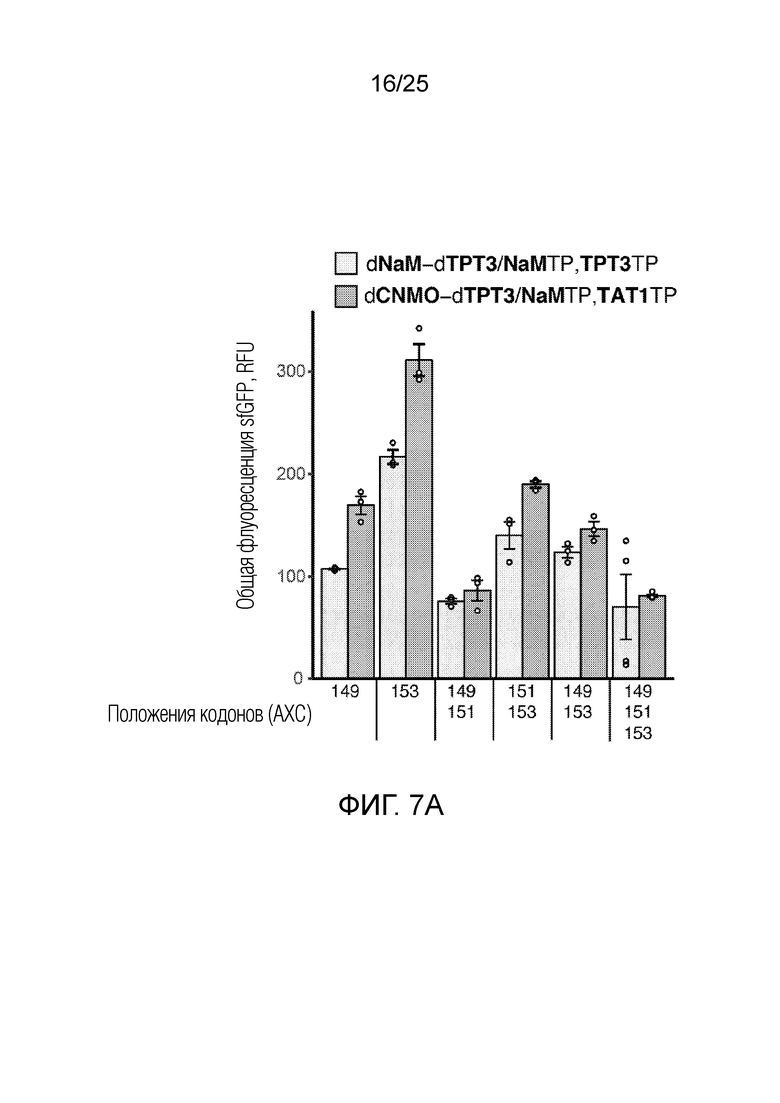
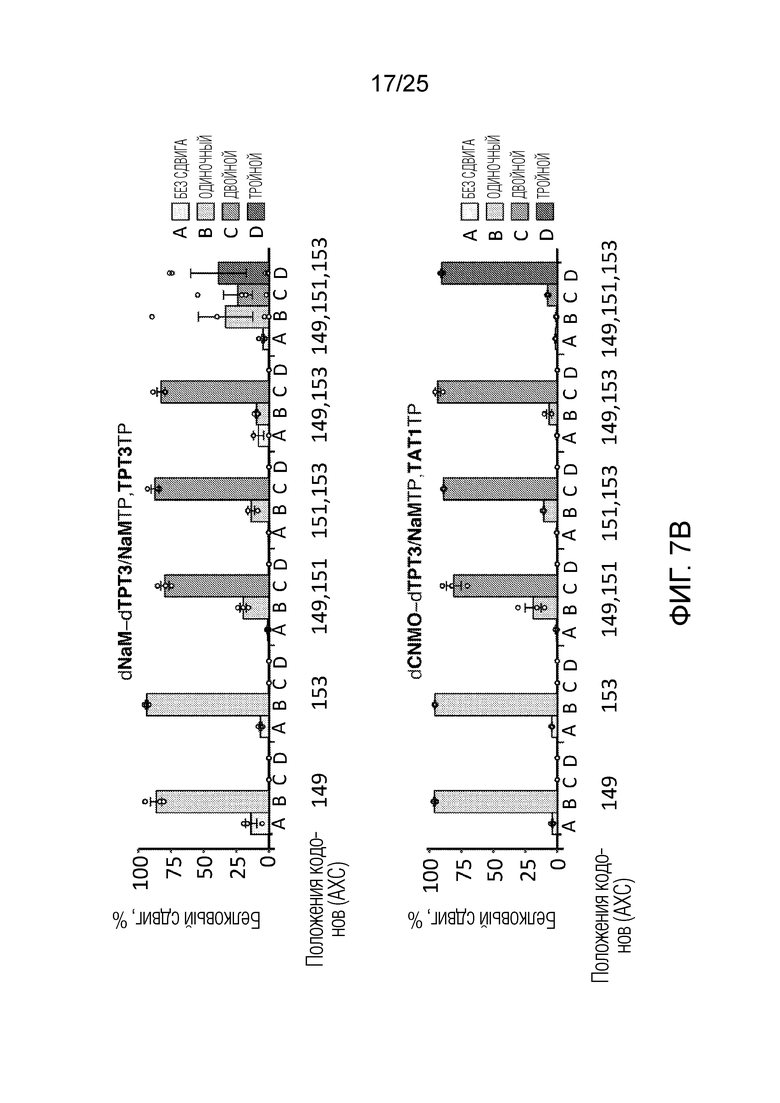
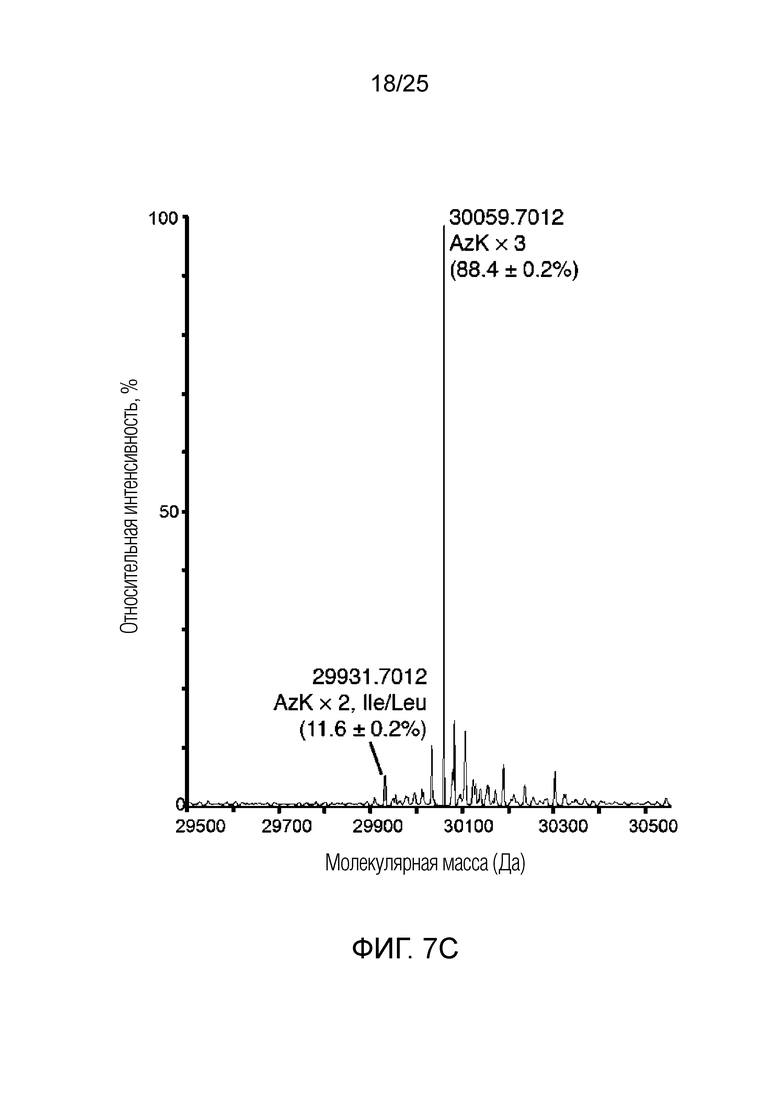
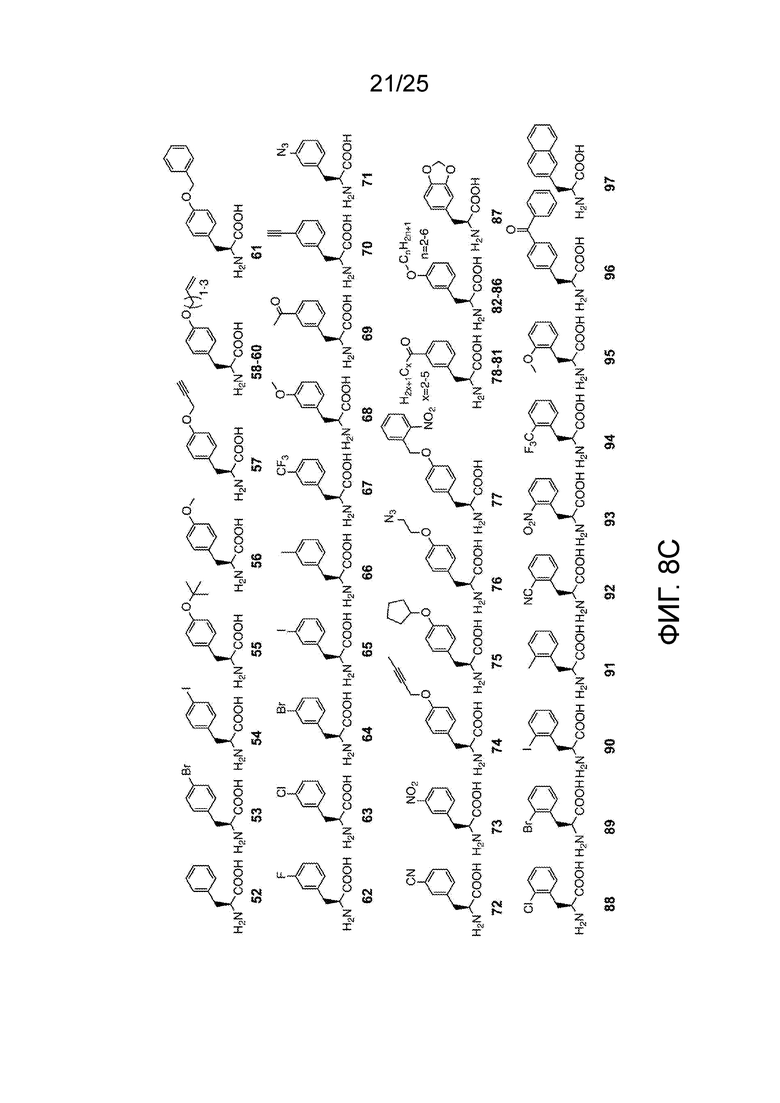

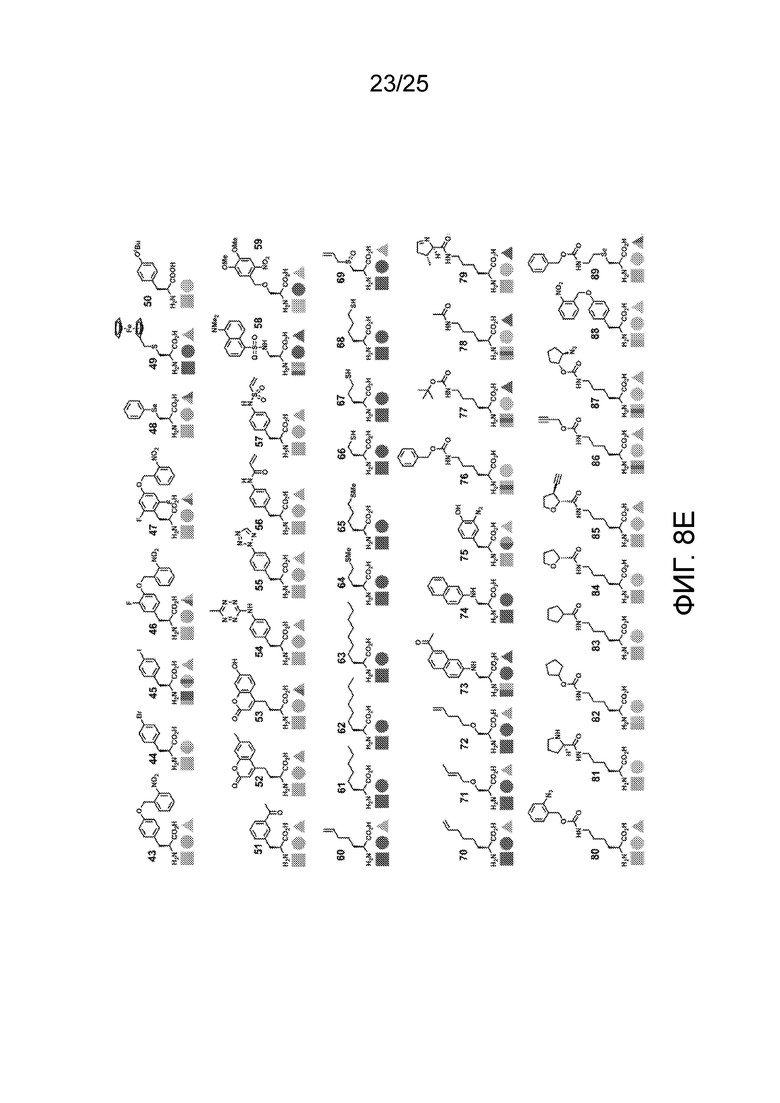
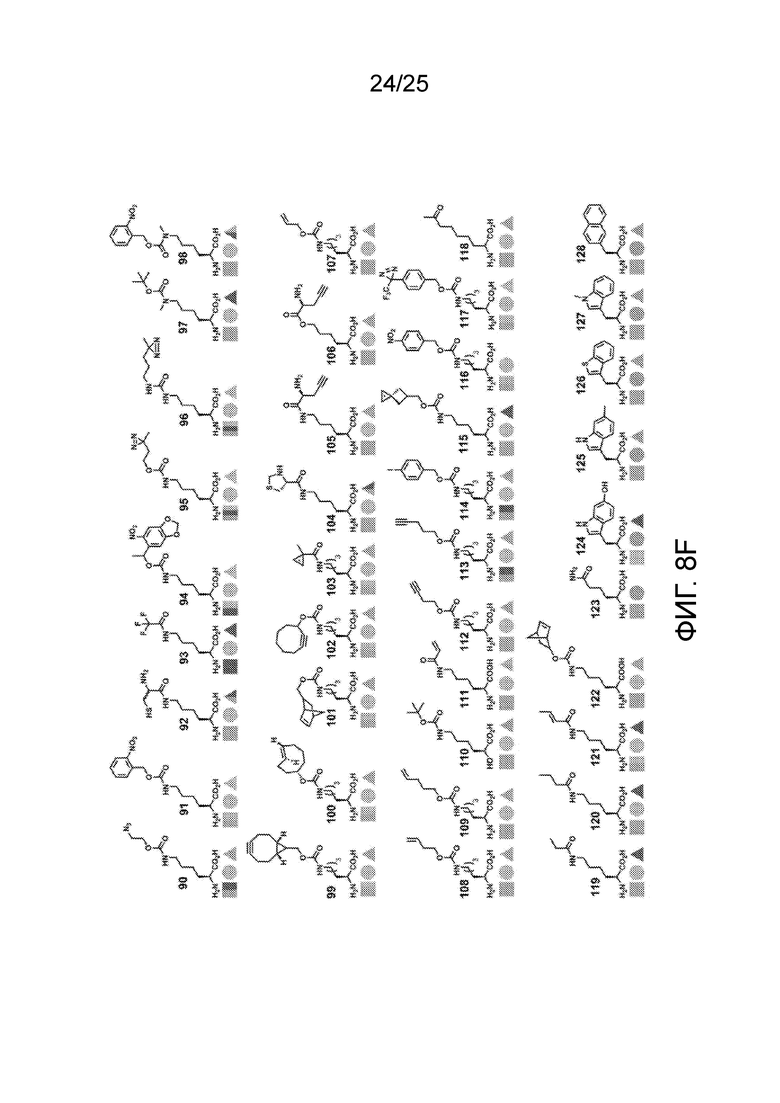

Настоящее изобретение относится к области биотехнологии. Раскрывается синтетический аналог природных нуклеиновых оснований. Изобретение может быть применимо для расширения генетического кода, в частности при получении новых кодонов, не имеющих природной функции и затруднений, однако распознавание которых в рибосомах, по существу, является более ортогональным. 6 н. и 16 з.п. ф-лы, 8 ил., 3 табл., 4 пр.
1. Нуклеиновое основание структуры:
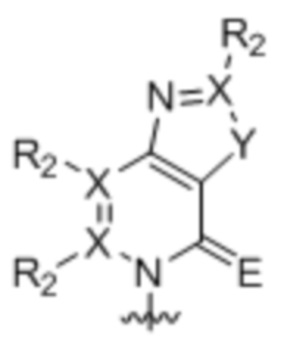 ,
,
где:
каждый X представляет собой углерод;
каждый R2 независимо представляет собой водород, алкил, алкенил, алкинил, метоксигруппу, метантиол, метанселеногруппу, галоген, цианогруппу или азидную группу;
Y является серой, кислородом или селеном; и
E является кислородом, серой или селеном;
где волнистая линия представляет присоединение нуклеинового основания к рибозильному, дезоксирибозильному или дидезоксирибозильному остатку, где рибозильный, дезоксирибозильный или дидезоксирибозильный остаток находится в свободной форме, соединен с монофосфатной, дифосфатной, трифосфатной, α-тиотрифосфатной, β-тиотрифосфатной или γ-тиотрифосфатной группой, или включен в полинуклеотид,
необязательно где полинуклеотид представляет собой РНК, ДНК, бициклическую нуклеиновую кислоту, связанную нуклеиновую кислоту, пептид нуклеиновую кислоту (ПНК), замкнутую нуклеиновую кислоту (ЗНК), нуклеиновую кислоту, содержащую фосфоротиоат.
2. Нуклеиновое основание по п.1, где E является серой.
3. Нуклеиновое основание по любому из пп.1 или 2, где Y является серой.
4. Нуклеиновое основание по п.1, имеющее структуру 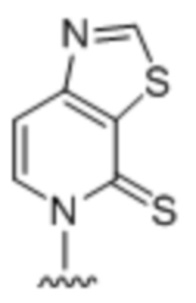 .
.
5. Нуклеиновое основание по любому из пп.1-4, связанное с комплементарным, спаривающимся нуклеиновым основанием для образования неприродной пары оснований (UBP).
6. Нуклеиновое основание по п.5, где комплементарное, спаривающееся нуклеиновое основание выбрано из:
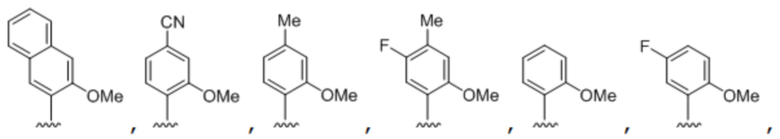
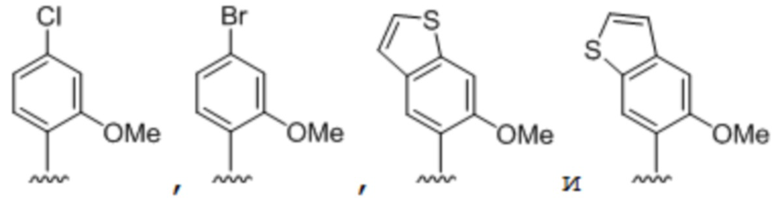 .
.
7. Двухцепочечный олигонуклеотид для репликации или транскрипции, содержащий первую олигонуклеотидную цепь, которая содержит нуклеиновое основание по любому из пп.1-4, и вторую олигонуклеотидную цепь, которая является комплементарной к первой олигонуклеотидной цепи, где вторая олигонуклеотидная цепь содержит комплементарное, спаривающееся нуклеиновое основание в его комплементарном, образующем пару оснований участке.
8. Двухцепочечный олигонуклеотид по п.7, где первая олигонуклеотидная цепь содержит ; и вторая олигонуклеотидная цепь содержит комплементарное, образующее пару нуклеиновое основание, выбранное из:
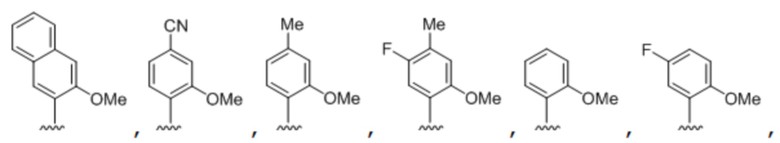
 в его комплементарном, образующем пару оснований участке.
в его комплементарном, образующем пару оснований участке.
9. Двухцепочечный олигонуклеотид по п.8, где вторая олигонуклеотидная цепь содержит комплементарное, спаривающееся нуклеиновое основание  .
.
10. Двухцепочечный олигонуклеотид по п.8, где вторая олигонуклеотидная цепь содержит комплементарное, спаривающееся нуклеиновое основание  .
.
11. Транспортная РНК (тРНК), содержащая по меньшей мере одно нуклеиновое основание по любому из пп.1-4.
12. Транспортная РНК (тРНК), содержащая:
антикодон, содержащий нуклеиновое основание по любому из пп.1-4, где антикодон спаривается с неприродным кодоном, который кодирует неприродную аминокислоту; и
элемент распознавания, где элемент распознавания способствует селективной зарядке тРНК с неприродной аминокислотой посредством аминоацил-тРНК-синтетазы.
13. тРНК по п.12, где аминоацил-тРНК-синтетазу получают из Methanosarcina, или его варианта, или Methanococcus (Methanocaldococcus), или его варианта.
14. тРНК по п.12, где неприродная аминокислота содержит ароматический остаток.
15. тРНК по п.12, где неприродная аминокислота является производным лизина или производным фенилаланина.
16. Структура, которая кодирует ген, содержащая формулу:
N1 - Zx - N2
где:
каждый Z независимо представляет собой нуклеиновое основание по любому из пп.1-4, связанное с рибозилом, или дезоксирибозилом;
N1 является одним или более нуклеотидами, или концевой фосфатной группой, присоединенной к 5'-концу рибозила, или дезоксирибозила из Z;
N2 является одним или более нуклеотидами, или концевой гидроксильной группой, присоединенной к 3'-концу рибозила, или дезоксирибозила из Z; и
x является 1.
17. Нуклеозидтрифосфат, содержащий нуклеиновое основание по любому из пп.1-3.
18. Нуклеозидтрифосфат по п.17, где нуклеиновое основание представляет собой .
19. Нуклеозидтрифосфат по п.17 или 18, где нуклеозид содержит рибозу или дезоксирибозу.
20. Нуклеиновое основание по любому из пп.1-6, где полинуклеотид представляет собой бициклическую нуклеиновую кислоту, связанную нуклеиновую кислоту, пептидную нуклеиновую кислоту (ПНК), замкнутую нуклеиновую кислоту (ЗНК) или нуклеиновую кислоту, содержащую фосфоротиоат.
21. Нуклеиновое основание по любому из пп.1-6, где полинуклеотид представляет собой РНК.
22. Нуклеиновое основание по любому из пп. 1-6, где полинуклеотид представляет собой ДНК.
| EP 3041854 A4, 03.05.2017 | |||
| Malyshev D | |||
| A | |||
| et al | |||
| A semi-synthetic organism with an expanded genetic alphabet //Nature, 2014, 509(7500), p | |||
| Саморазгружающаяся платформа | 1922 |
|
SU385A1 |
| RU 2008122948 A, 20.12.2009 | |||
| US 8846883 B2, 30.09.2014 | |||
| Способ торможения электровозов | 1928 |
|
SU28928A1 |

