Изобретение относится к ьшкробно- логической промьшшенности, точнее к . тем ее отраслям, где микробиологическим объектом технологического про-: цесса являются дрожжи-сахаромицеты, и может быть использовано для полу- . чения кислых фосфатаз.
Цель изобретения - повышение активности и выхода кислой фосфатазы.
Способ заключается в том, что в качестве продуцента кислой фосфатазы , используют полиплоидный штамм с псевдомицелиальным типом роста Saccharomyces cerevisiae ВКПМ У-618.
Кислые фосфатазы (КФ) относятся к экаоферме.нтам. Количество секретируемой КФ пропорционально площади поверхности клетки. Полиплоидные штам- мы дрожжей с псевдрмицелиальным типом йоста имеют крупные клетки с изменен- ной морфологией, что может существ енно сказаться на площади клеточной поверхности. Кроме того, изменение мор- фологии клеток затрагивает структуру клеточной оболочки, что также может влиять на секреи5ию экзоферментов. Со- ;вокупность перечисленньк характеристик создает реальные предпосылки для получения КФ из полиплоидных штаммов дрожжёй-сахар.омицетов с псевдомицели-. альным роста.
Пример 1. Получение тет.ра- плоида с псевдомицелиальным типом роста.
Для получения полиплоидного штам- ма с псевдомицелиальным типом роста (Rpm) проводят серию скрещивания.
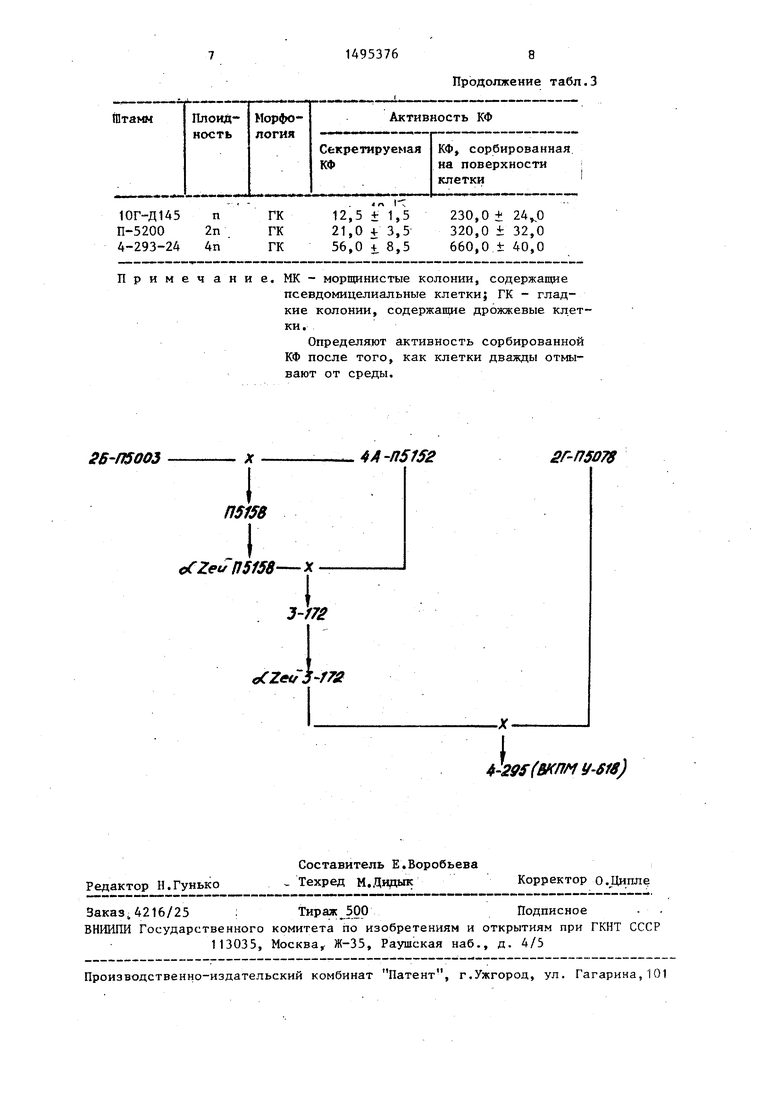
На чертеже представлена схема, реализующая способ.
Гаплоидные штаммы, использованные для получения тетраплоида 4-295, имеют .следующий генотип:
. СО
СП
со --J
05
2Б-П5003 МАТ wl.leu 2-2 ura 3 rpm 3 rpm 4А-П5152-МАТ cithr 4-B15 rpm 3 2Г-П5078-МАТ oCade 1-6 rpm 3.
Bee культуры депонированы во Йсе- союзной коллекции промьгашенных микроорганизмов ВНИИ генетика, где им присвоены следующие номера: 2Б-П5003 У-615, 4А-П5152 - У-616, 2Г-П5078 - У-617, 4-295 - У-618.
При выделении полиплоида с псев- домицелиальньгм типом роста используют метод, основанный на гомозиготи- зации по локусу типа спаривания. ,
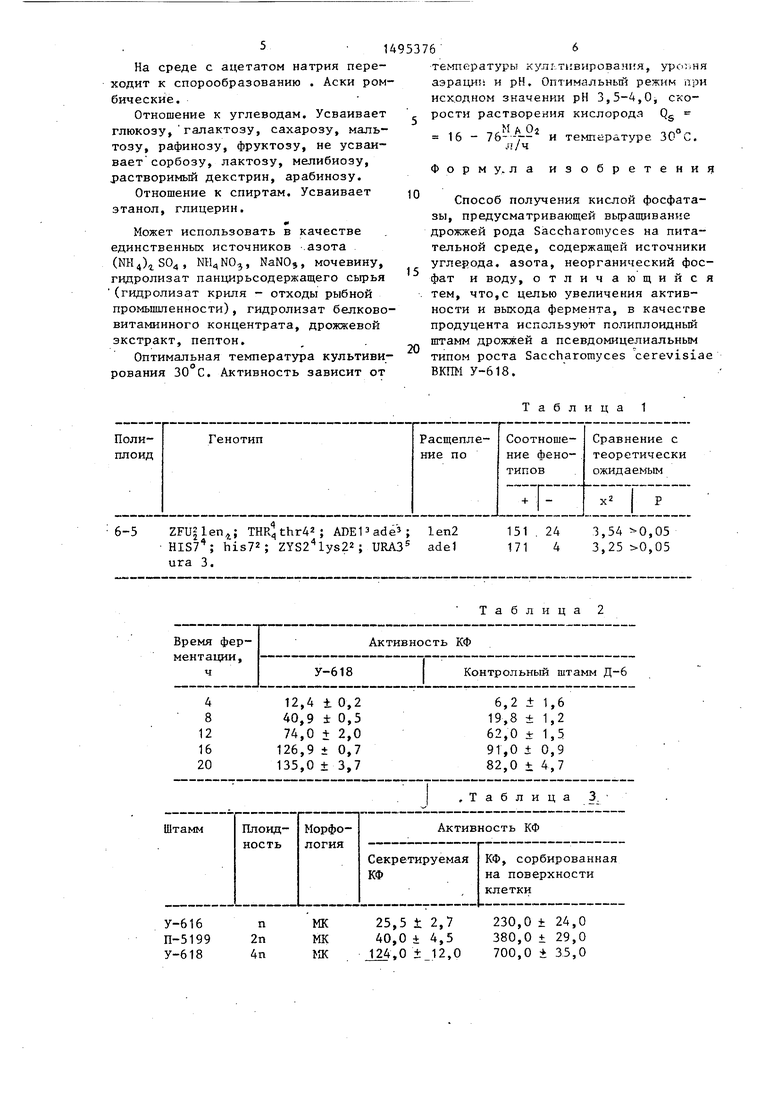
Генетические данные, подтверждаю- щие плоидность штамма ВКПМ-618 представлены в табл.1.
В связи с тем, что вьщеленн й тет- раплоид Rpra не спорулирует, гомозиготу по полу ВКПМ У-618 скрещивают с автодиплоидом 3-П219 генотипа МАТа/ . /МАТа ade1/ade1 his7/his7 Iys2/l yjs2. Анализируют полученный гексаплоид 6-5.. Фертильность аскоспор 6-5 не превышает 50%, поэтому анализирз т случайную выборку аскоспор. Полученные расщепления по ауксотрофным маркерам соответствуют теоретически ожидаемым при предполагаемом уровне Ш1ОИДНОСТИ.
Пример 2. Оценка активности секретируемой кислой фосфатазы (КФ) у штамма БКПМ У-61Й.
Для характеристики секретной активности по КФ дрожжи выращивают в среде, содержащей в 1 л дистиллирот. ванной Hip 20 г глюкозы; 20 г пептона и 30 мг КН2;Р04. Вьфащивание проводят в течение 20-24 ч в колбах Эрленмейе- ра (объем среды - 100 мл), на качалке (235 об/мин-) . Определение биомассы дрожжей проводят нефелометрическим методом на ФЭК-М,
Для количественного определения активности КФ, экскретируемой дрожжа- ми в. культуральную жидкость, используют кашориметрический метод, основанный на способности КФ отщеплять от паранитрофенилфосфата (п-ИФФ) фос- фор, в результате чего образуется паранитрофенол (п-НФ), имеющий максимум поглощения в области 410 нм. Ферментативную активность выражают в мкМ п-НФ, образующегося за ,1 мин при 33 с, принимая Е 125Ш.
Удельную активность КФ выражают в мкМ п-НФ/мин/мг белка клеток дрожжей
Расщепление гексаплоида 6-5 показано в табл.1.
Активность КФ у штамма пока зана в табл.2.
В качестве контрольного использую щтамм Saccharomyces cerevisiae D-6. Статистическая обработка данных по критерию Стьюдента свидетельствует, что после 4 ч выращивания не выявляются достоверные отличия по количеству экскретируемой КФ между контрольным штаммом и У-618 (tdif 3587, -таБл 4,30). Однако по мере роста выявляется достоверное превышение активности КФ штамма над контролем (tdif 12,8; 4,8; 8,8; 8,9 . соответственно для измерений после 8, .12, 16 и 20 ч роста).
Пример 3. Сравнение активности КФ у штаммов различной плоид- ности и морфологии.
Активность КФ оценивают после 16 роста культуры.
Результаты проведенных опытов суммированы в табл.3,
Данные табл.3 указьшают на возрастание активности КФ как секретируемой, так и сорбированной на поверхности клетки в соответствии с пло1ад - ностью клетки. В то же время штаммы, образующие морпщнистые колонии (МК)- с псевдомицелиапьными клетками характеризуются более высоким уровнем секреции: активность секретрфуе мой КФ у морщинистых штаммов превос - ходит вдвое таковую у гладких штаммов той же плоидности.
Физиолого-биохимическое описание штамма Saccharomyces cerevisiae ВКШ У-618.
Штамм является тетраплоидом, получен в результате серии, последовательных скрещиваний.
На средах УАРД, полной с пептоном ПЕПФО и ьшнимальной, штамм образует морщинистые колонии из псевдомицели- альных клеток. В жидких средах того же состава образует быстро осаждающиеся скопления клеток.
Состав сред, УАРД: пептон 20 г, глюкоза 20 г, дрожжевой автолизат 10 МП, 0,9 г, MgS04 0,5 г, (N«4)4204 3,5 г, 0,23 г, агар 30 г; ПЕПФО: пептон 20 г, глюкоза 20 г, 1 г, агар 25 г.
Минимальная: 0,9 г, К НРО. 0,23 г, MgS040,5 г, (NH4)aS04 3,5 г, глюкоза 20.г, агар 30 г, тиамин 200мкг,;5 -аланин 500 мкг, биотин 2 MKF
На среде с ацетатом натрия переходит к спорообразованию . Аски ромбические.
Отношение к углеводам. Усваивает глюкозу, галактозу, сахарозу, мальтозу, рафинозу, фруктозу, не усваивает сорбозу, лактозу, мелибиозу, растворимый декстрин, арабинозу.
Отношение к спиртам. Усваивает этанол, глицерин.
Может использовать в качестве единственных источников .азота (№14)1804, , NaNO,, мочевину, гидролизат панцирьсодержащего сырья (гидролизат криля - отходы рыбной промышленности), гидролизат белково- витаминного концентрата, дрожжевой экстракт, пептон.
Оптимальная температура культивирования 30 С. Активность зависит от
тe mepaтypы культивирования, уро;;ня аэраци : и рН, Оптимальньй режим при исходном значении рН 3,5-4,0, скорости растворения кислорода Qg
f л п„ 16
-
л/ч
и температуре
30°С.
Форму, ла изобретения
Способ получения кислой фосфата- зы, предусматривающей выращивание дрожжей рода Saccharomyces на питательной среде, содержащей источники углерода, азота, неорганический фосфат и воду, отличающийся тем, что,с целью увеличения активности и выхода фермента, в качестве продуцента используют полиплоидный штамм дрожжей а псевдомицелиальным типом роста Saccharomyces cerevisiae ВКПМ У-618.

| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE № 965 (ГКВ) - ПРОДУЦЕНТ ПОВЕРХНОСТНОГО АНТИГЕНА ВИРУСА ГЕПАТИТА B | 2000 |
|
RU2182927C2 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА СЕКРЕТИРУЕМОГО СОМАТОТРОПИНА ЧЕЛОВЕКА И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE - ПРОДУЦЕНТ СЕКРЕТИРУЕМОГО СОМАТОТРОПИНА ЧЕЛОВЕКА | 2011 |
|
RU2460795C1 |
| ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE 1-60-Д578 (MSIL), - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ИНТЕРЛЕЙКИНА-2 ЧЕЛОВЕКА И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2002 |
|
RU2230781C1 |
| Трансформант Komagataella phaffii, содержащий ген HAC1, продуцент рекомбинантного химозина Vicugna pacos в активной форме | 2023 |
|
RU2828277C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА, ШТАММ Saccharomyces cerevisiae - ПРОДУЦЕНТ ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ) | 2010 |
|
RU2427645C1 |
| ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE ВКПМ Y 1678, ПРОДУЦЕНТ ПОВЕРХНОСТНОГО АНТИГЕНА ВИРУСА ГЕПАТИТА B | 1998 |
|
RU2143494C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE И ШТАММ-ПРОДУЦЕНТ ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ) | 2011 |
|
RU2446172C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ФИБРОБЛАСТНОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА КЛЕТКАМИ ДРОЖЖЕЙ, СПОСОБ ЕЕ КОНСТРУИРОВАНИЯ И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE-ПРОДУЦЕНТ ФИБРОБЛАСТНОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА | 1998 |
|
RU2180003C2 |
| КАССЕТА И РЕКОМБИНАНТНАЯ ПЛАЗМИДА ДЛЯ ЗКСПРЕССИИ И СЕКРЕЦИИ МЕХАНОЗАВИСИМОГО ФАКТОРА РОСТА ЧЕЛОВЕКА (MGF), ШТАММ SACCHAROMYCES CEREVISIAE - ПРОДУЦЕНТ MGF И СПОСОБ ПОЛУЧЕНИЯ MGF | 2006 |
|
RU2344173C2 |
| Трансформант Komagataella phaffii - продуцент рекомбинантного химозина в активной форме | 2022 |
|
RU2805486C1 |
Изобретение относится к микробиологической промышленности и может быть использовано для получения кислых фосфатаз. Цель изобретения - повышение активности и выхода фермента. Изобретение заключается в том, что в качестве продуцента кислой фосфатазы используют полиплоидный штамм дрожжей с псевдомицелиальным типом роста +SACCHAROMYCES CEREVISIAE ВКПМ У-618. Активность секретируемой фосфатазы после 16 ч культивирования на среде, содержащей (г/л) глюкозу 20, пептон 20, КН2РО4 0,03, составляет 124,0±12,0 ед, фосфатазы, сорбированной на поверхности клеток, 700±35,0 ед. 1 ил. 3 табл.
Таблица 1
Поли- ГенотипРасщепле- Соотноше- Сравнение с
плоидние по ние фено- теоретически
типов ожидаемым
+ I - х2 Р
6-5 THR thr42 ; ADEPade ; 1еп2151 . 24 3,,05
his72 ; ZYS2 lys22; URA3 adel171 4 3,,05
ura 3.
Таблица 2
Время фер-Активность КФ
ментации,:
чУ-618Контрольный штамм Д-6
412,4 1 0,26,2 ± 1,6
840,9 ± 0,519,8 ± 1,2
1274,0 ± 2,062,0 ± 1,5
16126,9 ± 0,791,0 ± 0,9
20135,0 ±3,782,0 t 4,7
.Таблица 3.
Штамм Плоид- Морфо- Активность КФ ность логия
Секретируемая КФ, сорбированная КФна поверхности
клетки
У-616 п МК 25,5 t 2,7 П-5199 2п МК 40,0 ± 4,5 У-618 4п КК 124,0 ± 12,0
230,0 ± 24,0 380,0 ± 29,0 700,0 ± 35,0
Примечание. МК- морщинистые колонии, содержащие
псевдомицелиальные клетки; ГК - гладкие колонии, содержащие дрожжевые кдет- ки.
Определяют активность сорбированной КФ после того, как клетки дважды отмывают от среды.
2В-П5003
П5т
cCZetfnSfSe-Jf
J-/7
e ZacT;
-Г7
Продолжение табл.3
4А П515г
ЗГ-П5078
4 29f(BKnMy-SlS)
