Изобретение относится к биохимии и биотехнологии и может быть использовано для ферментативного синтеза нуклеозидт- рифосфатов. меченных фосфором-32(33) в «-положении фосфатной группы.
Известен способ получения нуклеазы. базирующийся на традиционных приемах очистки-фракционирование солями, гель- фильтрация, ионообменная хроматогра- фия 1,
Способ не позволяет получать препараты, отвечающие требованиям в отношении примесных активностей, так как не содержат надежного этапа их устранения (особенно АТФ-аз).
Наиболее близким является способ очистки нуклеазы из проростков ячменя 2. Способ включает четыре стадии очистки: сульфатаммонийное осаждение белков в интервале 40-80% насыщения, ионообменную хроматографию на ДЕАЕ-трис-акриле М при рН 8,0, гельфильтрацию на G-75, афинную хроматографию на АМФ-сефаро- зе.
К недостаткам известного способа относятся низкий выход очищенного продукта - не более 1%. нецелесообразность применения афинного сорбента АМФ-сефарозы для крупномасштабной очистки нукле.эзы ввиду малой емкости и нестабильности сорбента, отсутствие характеристики эн- зимологической чистоты препарата нукле- аза.
Целью изобретения является увеличение выхода целевого продукта, свободного от примесных АТ-фазной, 5 -нуклеотидаз- ной и неспецифической фосфатазной активностей.
Согласно способу очистки нуклеазы из проростков ячменя, включающему гомо 4 О СО О СО 00
генизацию сырья, сульфатаммонийное осаждение белков, ионообменную хрома- тографию при рН 8,0, в качестве сырья используют 4-6-дневные проростки ячменя, гомогенат из сырья подвергают очистке пу- тем сульфатаммонийного осаждения белков в интервале 40-80% насыщения с последующим нагревом суспензии до 60±1°С в течение 15 мин, после чего проводят хроматографию на ДЭАЭ-целлюлозе с элю- цией ферментов раствором 0,1 М NaCI и псевдоафинную хроматографию на силох- роме 50/80 красно-коричневом 2К при рН 8,0 с предварительной элюцией примесных белков раствором 0,1 М NaCI и элюцией нуклеазы раствором 0,5 М NaCI.
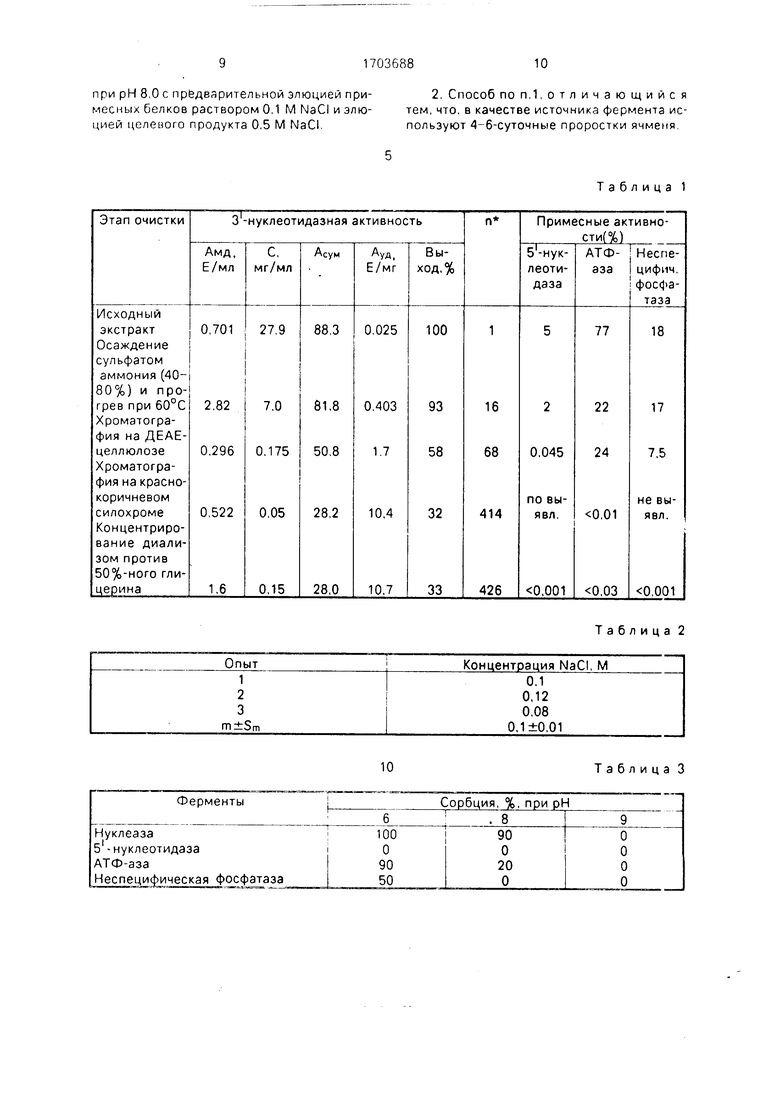
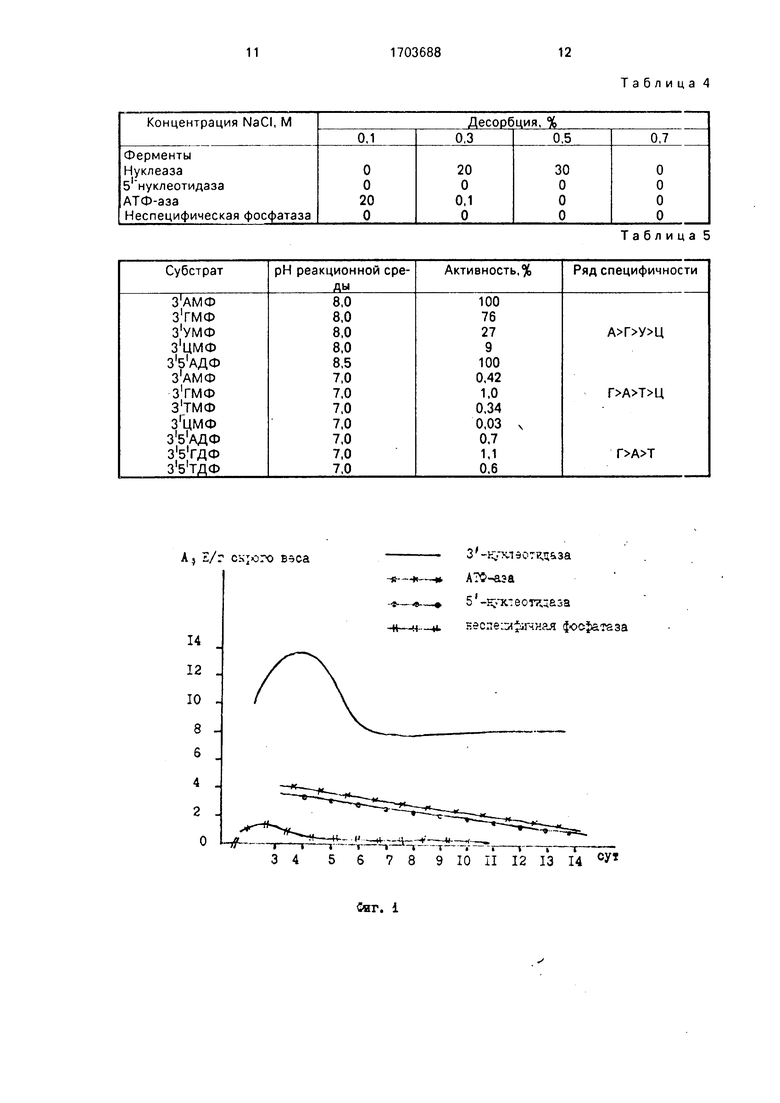
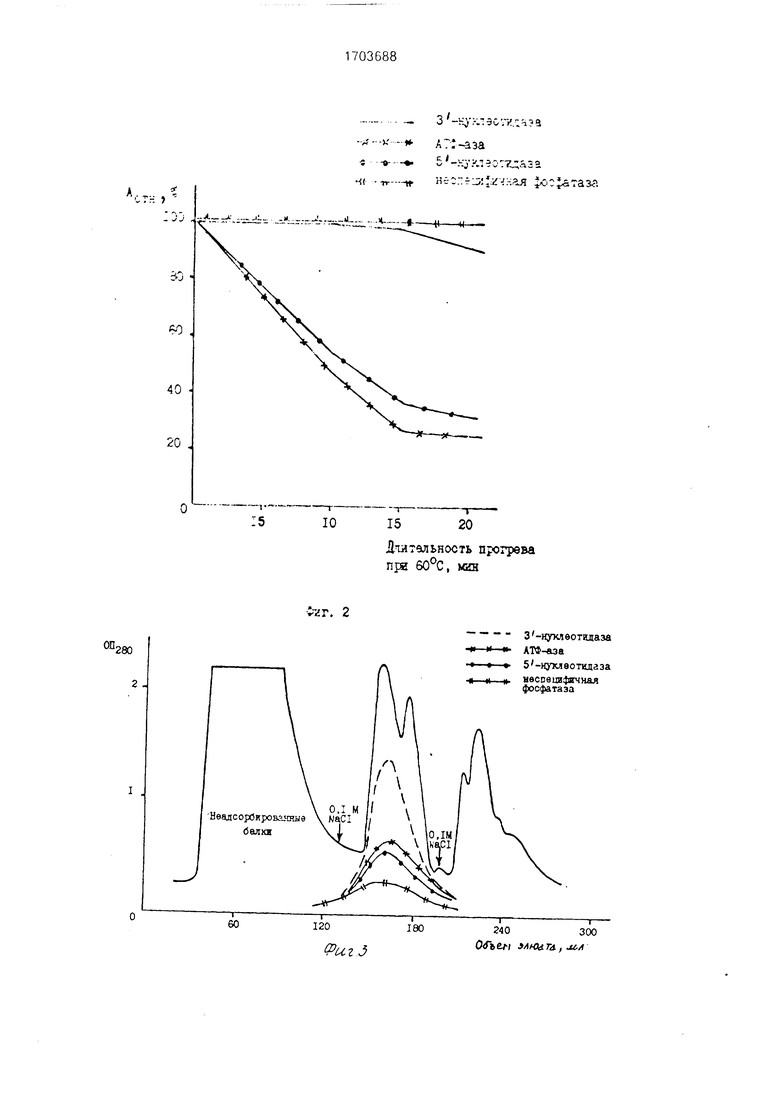
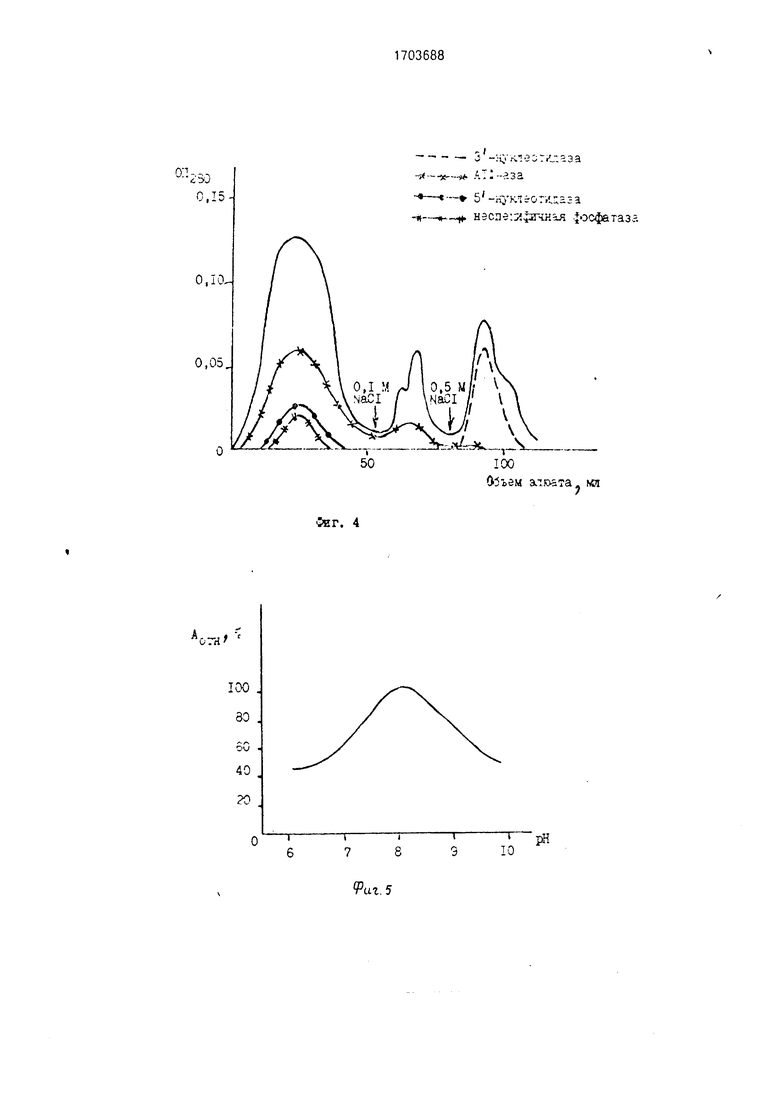
На фиг.1 показана динамика изменения активности целевого и примесных ферментов в зависимости от возраста (максимальная 3-нуклеотидазная активность обнаружена в 4-6-дневных проростках, а не в 6-7-дневных, как в известном способе); на фиг.2 - устойчивость нуклеазы к нагреванию до 60°С в течение 15 мин (при тепловой обработке происходит частичная инактивация примесных дефосфорилирующих ферментов 5 -нуклеотидазы и АТФ-азы (до 60-70%), отсюда целесообразность введения этого этапа в процедуру очистки); на фиг.З - профиль элюции белков с ДЕАЕ- целлюлозы (рН 8,0) при осуществлении ступенчатой элюции раствором 0,1 М NaCI (на этом этапе происходит дополнительная очистки фермента в 4.1 раза, однако примесные дефосфорилирующие ферменты не отделяются от целевого фермента); на фиг.4 - профиль элюции белков при рН 8,0 с псевдоаффинного сорбента силохром 50/80 красно-коричневый 2К при осуществлении ступенчатой элюции примесных белков раствором 0,1 М NaCI и элюции нуклеазы с З -нуклеотидазной активностью раствором 0,5 М NaCI (в этих условиях проведения хроматографии неспецифичная фосфатаза и б -нуклеотидаза не адсорбируется, АТФ-аза связывается слабо и полностью элюируется с 0,1 М NaCI, тогда как нуклеаза связывается прочно и элюируется с 0,5 М NaCI); на фиг.5 - оптимум рН З -нуклеотидазной активности (рН 8,2); на фиг.6 - диаграмма, поясняющая предлагаемый способ.
Пороговые концентрации растворов NaCf примененных при хроматографии на ДЕАЕ-целлюлозе и силохроме красно-коричневом 2К, а также рН буфера при хроматографии на силохроме красно-коричневом 2К, подобраны экспериментально.
Схема процедуры очистки нуклеазы
Гомогенизация растительного материала
Экстракция белков буфером
I
Осаждение белков сульфатом аммония (40-80% насыщения)
t Тепловая обработка (60°С, 15 мин)
роматография на ДЕАЕ-целлюлозе, рН «,0
0,1 М NaCI Хроматография на силохроме красно-коричневом, рН 8,0
0,5МЫаС1 оТм NaCI
неадсорбированные белки
NT
неспецифичная фосфатаза, 5/-нуклеотидаза. часть АТФ-азной активности
нуклеаза с АТФ-аза
З -нуклеотидазнойактивностью
I
Концентрированно (диализ против
50%-ного глицерина) П р и м е р 1. Все этапы очистки, кроме тепловой обработки и хроматографии на си- лохроме с иммобилизованным красителем, проводят при .
Результаты выделения и очистки по этапам сведены в табл. 1, где показаны данные очистки нуклеазы с З -нуклеотидазной активностью из 75 г проростков ячменя.
Этап 1. Получение исходного экстракта. Зерновки ячменя (сорт Пиркка) проращивают в кюветах 20 х 30 см на деионизо- ванной воде в темноте при 20°С. 4-6-суточные проростки отделяют от семени и корней, растирают в ступке или гомогенизируют в ножевом гомогенизаторе с 0,1 М трис-HCI буфером, рН 6,5. Гомогенат ставят на качалку на 1 ч для полноты экст- ракции фермента, после чего центрифугируют 1 ч при 8000 д, осадок отделяют, а супернатант фильтруют через Miracloch. Концентрация белка в исходном экстракте 20-30 мг/мл.
Этап 2. Осаждение белков сульфатом аммония.
К исходному энзимологическому :жс- тракту медленно при непрерывном помешивании добавляют кристаллический сульфат аммония до 40% насыщения (243 г/л). Осадок, сформировавшийся в течение 1 ч, отделяют центрифугированием при 10000 g 30 мин. Осадок отбрасывают, супернзтант насыщают сульфатом аммония до 80% (561 г/л). Оставляют на 2 ч для формирования осадка.
Этап 3. Тепловая обработка.
Суспензию белков насыщения сульфатом аммония подвергают нагреву до 60°С в течение 15 мин, после чего охлаждают в ледяной бане 20-30 мин. Суспензию центрифугируют в том же режиме. Осадок растворяют в 50 мМ трис-HCI буфере. рН 8,0 и подвергают диализу против 1 л этого же буфера с трехкратной заменой буфера.
Этап 4. Хроматография на ДЕАЕ-целлю- лозе.
Обессоленный раствор белка наносят на колонку с ДЕАЕ-целлюлозой (2,5 х 18 см), уравновешенной 50 мМ трис-HCI буфером, при скорости 25 мл/ч. После нанесения белка и промывки колонки буфером (100 мл) подают раствор 0.1 М NaCI для элюции фермента с 3 -нуклеотидазной активностью (фиг.З). Фракции с 3 -нуклеотидазной активностью объединяют и диализуют против 1 л 50 мМ трис-HCI буфера, рН 8.0.
Этап 5. Хроматография на силохроме 50/80 красно-коричневом 2К.
Отдиализованный раствор белка наносят на небольшую колонку (2x6 см) с силохромом красно-коричневым, уравновешенным 50 мМ трис-HCI буфером, рН 8.0, при 20°С, после чего колонку промывают 20 мл того же буфера для удаления неспецифичной фосфатазы и 5 -нуклеотидазы. Примесные белки с АТФ-азной активностью устраняют раствором 0.1 М NaCI (20 мл), нуклеазу с З -нуклеотидазной активностью элюируют 20 мл 05 М раствора NaCI. Скорость элюции 40-50 мл/ч.
Этап 6. Концентрирование.
Объединение фракции с 3 -нуклеотидазной активностью диализуют против 50%-но- го раствора глицерина, приготовленного на 50 мМ Na-ацетатном буфере рН 6.0 и хранят при-10-20°С.
Определение активности ферментов.
Определение 3 -нуклеотидазной активности нуклеазы.
З -Нуклеотидазную активность определяют по колориметрическому методу. За единицу активности принимают количество Рн в микромолях, освобожденного за одну минуту гидролиза 3 АМФ при 37°С.
Метод определения следующий: в пробирку для микропроб объемом 1,5 см3 помещают 50 мкл исследуемого образца белка, 100 мкл раствора З АМФ концентрации 7.5 мМ, 300 мкл буферного раствора 50 мМ TEA-HCI - Гм эОН. рН 8.0: проб/ июу- бируют 30 мин при 37 С. Реакцию с ,г;м;н;- ливают добавлением 50 мг.г. х;;ор ion кислоты (9 н.). Из аналитических гробирск
отбирают по 100 мкл раствора. ;;ССЗЕ- : 0,4 мл 1 М ацетата натрия, 1 м.; С .: ..4t татного буфера, рН 4, 0.4 мл а:.,;- О 2 м;: 1%-ного раствора аскорбиновой кислоты. приготовленного на 1 мМ раствора сернокислой меди. Оставляют смесь на 10 мин после чего добавляют 0,2 мл 1%-ного раствора молибдата аммония, приготовленного на 0,05 н. H2S04. Через 10 мин замеряют поглощенные пробы на спектрофотометре
при длине волны 660 нм. Расчет активности производят по формуле
20
ДЕббО 14.6 100 Амд ---Гз1----где Д Еббо - разность поглощения при /. 660 нм между опытным образцом и контролем (проба без белка);
14.6 - коэффициент пересчета единиц поглощения, мкг Рн (определяемый по калибровке);
г - время инкубации (30 мин); 31 - атомная масса Р. Определение активностей примесны дефосфорилирующих ферментов.
Определение активностей неспециф: ч- ной фосфатазы, 5 -нуклеотидазы и АТФ-азы проводят аналогичным образом, используя в качестве субстратов соответственно /3-глицерофосфат, 5 АМФ, АТФ. При выявлении фосфатазной активности с у5-глицеро- фосфатом применяют 50 мМ имидазольный буфер, рН 6,5.
П р и м е р 2. Выбор условий тепловой обработки.
Известно, что нуклеаза ячменя термостабильна при t 60°C. Экспериментально показано (фиг.2), что при этой температуре
происходит инактивация примесных деФос- форилирующих ферментов: б -нуклеотидззы 60%. АТФ-азы 70%. Поэтому целесообразно проведение при очистке целевого Фермента тепловой обработки при 60±1°С
(погрешность термометра). Длительность прогрева подбирают экспериментально. Наибольшее падение активности 5 -нукле- отидазы и АТФ-азы происходит за первые 15 мин нагрева при 60°С (фиг.2). При более
длительном воздействии примесные активности остаются на прежнем уровн.з, в то время как З -нуклеотидэзная активность снижается, Исходя из полученных данных, длительность прогрева выбрана 15 мин.
П р и м е р 3. Подбор концентрации NaCI для ступенчатой десорбции нуклеазы с ДЕАЕ-целлюлозы.
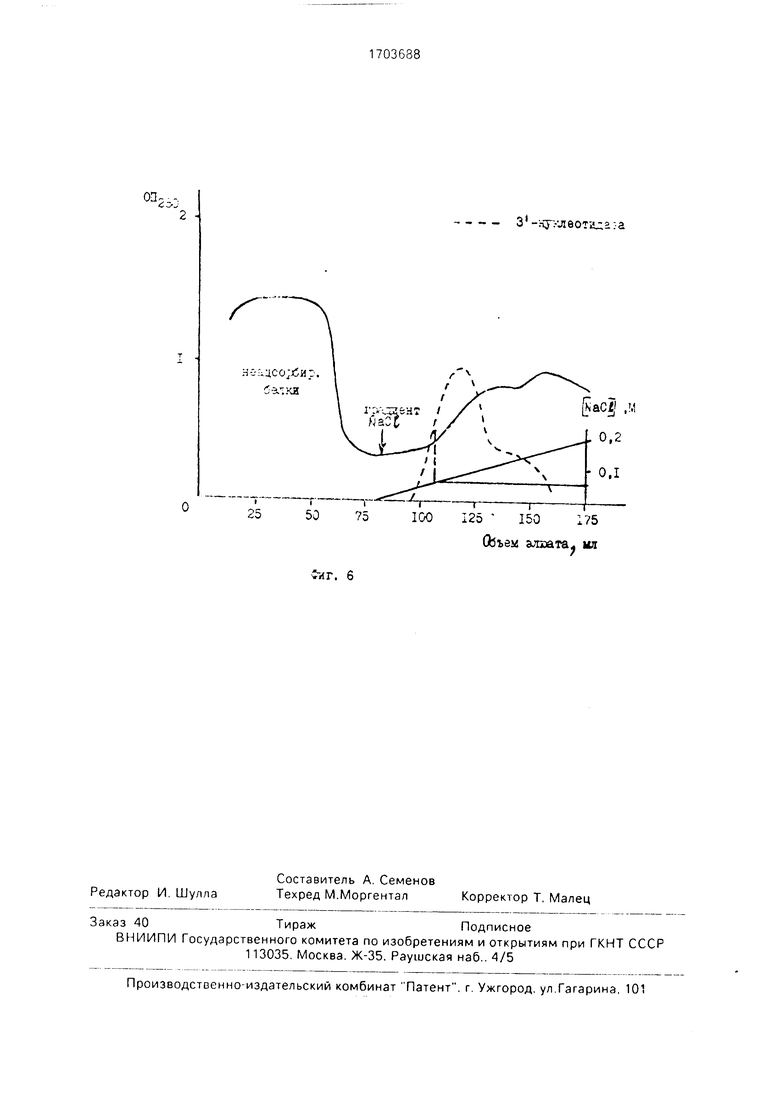
Процедура выполнения до этапа 4 аналогична примеру 1. после чего осуществляют в аналитическом варианте десорбцию целевого фермента в градиенте NaCI 0-0,2 М (150 + 150 мл) и определяют среднеарифметическое со среднеквадратичной ошибкой концентрации NaCI (фиг.6, срединная точка переднего фронта элюции).
В табл. 2 показаны срединные точки переднего фронта элюции нуклеазы в М NaCI при хроматографии на ДЕАЕ-целлюло- зе в градиенте NaCI (0-0,2) М.
Как видно из табл. 2, концентрация NaCI, необходимая для десорбции нуклеазы, 0,1 tO.01 М.
П р и м е р 4. Подбор рН буферного раствора и концентрации NaCI для проведения хроматографии на силохроме красно- коричневом.
Процедура выполнения до этапа 5 аналогична примеру 1, после чего устанавливают интервал рН буфера для эффективной сорбции нуклеазы на силохроме при слабой сорбции дефосфорилирующих ферментов.
В табл. 3 представлены данные сорбции нуклеазы с З -нуклеотидазной активностью и примесных ферментов на силохроме красно-коричневом при разных рН.
Как видно из табл. 3, оптимальным значением рН является 8,0, так как несмотря на незначительную потерю активности при сорбции (порядка 10%) при этом рН резко ослаблено связывание примесных ферментов по сравнению с рН 6.0. При рН 9,0 рассматриваемые белки не адсорбируются.
Выбор концентрации NaCI для ступенчатой адсорбции примесного и целевого ферментов определяется эффективностью десорбции при рН8,0.
В табл. 4 показана десорбция нуклеазы и примесных ферментов с силохрома красно-коричневого при различных концентрациях NaCI (рН 8,0).
Как видно из табл. 4. при концентрации 0,1 М NaCI происходит десорбция оставшейся АТФ-азы и других белков, в то время как нуклеаза остается связанной с сорбентом. Десорбция целевого фермента начинается с концентрации 0,3 М NaCI и достигает максимальной эффективности с 0,5 М NaCI.
Методика испытаний препарата.
Нуклеазу из проростков ячменя применяют в синтезе (5 - Р) dAMO и (5 33Р) АМФ на стадии перехода метки из (у- ЗЗР) АТФ в
(а -33) (d) АМФ: (у -33Р) АТФ + 3(d) АМФ полинуклеотид/киназа 3х - (5 - 33Р)(Ь)АДФ; (3, 5 -33PXd) АДФ нуклеаза/ячменя ( 33РХй)АМФ + фн.
Гидролиз З -фосфатной связи осуществляют как с очищенным после первой реакции меченым продуктом (дифосфатом), т.ж и в сквозном синтезе без предварительной очистки дифосфата. Существенно, что в обоих вариантах синтеза (a -33PXd) АМФ рН реакционной среды 7,0-7,5.
Реакцию проводят с 0,2 Е нуклеазы в течение 30 мин при 37°С; загрузка по радиоактивности составляет 45 мКи (у- Р) АТФ.
Переход метки из (3,5 -33Р)-дифосфата в (5 -33Р)-монофосфат составляет более 90%, что рассчитывают по соотношению площадей пиков на хроматограмме HPLC., Специфичность нуклеазы из ячменя к 3
-субстратам,
Исследована субстратная специфичность нуклеазы по отношению к З -нуклео- зид-моно- и дифосфатам рибо- и дезокси- риборяда.
В табл. 5 приведены относительные активности (с З АМФ-100%), проявляемые в области рН 7,0-8,5 (субстратная специфичность нуклеазы из проростков ячменя). Из табл. 5 видно, что нуклеаза из ячменя
проявляет З -нуклеотидазную активность и по отношению к З -субстратам дезокси-ря- да. в том числе к З ТМФ и 3, 5 ТДФ, однако скорость гидролиза приблизительно в 100 раз меньше, чем с З -рибонуклеотидами. Оптимум рН для рибонуклеотидов находится в области рН 8,0-8,5, для дезоксирибонукле- отидов - около 7,0.
Формула изобретения
1. Способ очистки нуклеазы из проростков ячменя, включающий гомогенизацию сырья, экстракцию гомогената, фракционирование экстракта сульфатом аммония в интервале концентраций 40-80% нэсыщения, ионообменную хроматографию на носителе, содержащем диэтиламиноэтиль- ные группы, и хроматографическую очистку, отличающийся т.ем, что, с целью увеличения выхода целевого продукта, свобедного от примесных АТ-фазной, 5 -нукле- отидазной и неспецифической фосфатазной активностей, суспензию белков после фракционирования сульфатом аммония подчер- гают термообработке при 60±1°С в течение
15 мин, ионообменную хроматографию проводят на ДЭАЭ-целлюлозе при рН 8,0 с элю- цией фермента 0,1 М NaCI, после чего проводят псевдоаффинную хроматографию на силохроме 50/80 красно-коричневом 2К
при рН 8,0 с предварительной элюцией примесных белков раствором 0.1 М NaCI и элюцией целевого продукта 0.5 М NaCI.
2. Способ по п.1,отличающийся тем, что, в качестве источника фермента используют 4-6-суточные проростки ячменя.



| название | год | авторы | номер документа |
|---|---|---|---|
| Способ выделения полинуклеотидфосфорилазы | 1981 |
|
SU1076445A1 |
| Способ получения рестриктазы, способной узнавать и расщеплять последовательность нуклеотидов GTCGAC | 1989 |
|
SU1752769A1 |
| Способ очистки ферментных препаратов | 1975 |
|
SU551339A1 |
| Способ получения фосфолипазы @ | 1983 |
|
SU1125246A1 |
| Способ получения сорбента для очистки белков | 1982 |
|
SU1060217A1 |
| Способ выделения ингибитора ангиотензин-1-превращающего фермента из яда животного происхождения | 1990 |
|
SU1730088A1 |
| Способ получения ДНК-полимеразы 1 ЕSснеRIснIа coLI | 1988 |
|
SU1622393A1 |
| СПОСОБ ПОЛУЧЕНИЯ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ SST 12 I | 2001 |
|
RU2233877C2 |
| СПОСОБ ПОЛУЧЕНИЯ МОДИФИЦИРОВАННОЙ ФОРМЫ ТРИПТОФАНИЛ-тРНК-СИНТЕТАЗЫ | 2012 |
|
RU2495123C2 |
| Способ получения лизил-тРНК-синтетазы | 1984 |
|
SU1195649A1 |
Изобретение относится к биохимии и биотехнологии и может быть использовано для ферментативного синтеза нуклеозпдт- рифосфатов. меченных фосфором -32(33) в «-положении фосфатной группы. Целью изобретения является увеличение выхода целевого продукта, свободного от примесных АТ-фазной, 5 -нуклеотидазной и неспецифической фосфатазной активностей. Способ заключается в том, что проростки ячменя 4- 6 дней гомогенизируют, экстрагируют, фракционируют экстракт сульфатом аммония 40- 80%, суспензию прогревают 15 мин при 60°С. хроматографируют на ДЭАЭ-целлю- лозе и силохроме 50/80 красно-коричневом 2К. Выход по активности 33%, степень очистки - 426 раз. 1 з.п. ф-лы., 6 ил., 5 табл. ё
Таблица 1
Таблица 2
10
Таблица 3
А, Е/г веса
14
12
10
8
6
4
2
О
34 5 6 7 8 9 10 II 12 13 14 СУ
Таблица 4
Таблица 5
З -ктхлзотядьза АТФ-аэа
5 -цух-еоти,ч&за кеспе:;иЈгчная фос| атаза
Сет. i
Я--.
Ю
ео ,
40
.-,.- .5
10
v-иг. 2
on,
280
I .
Неадсор(3ярова.чныа балкж
J
т-
60
-I-
120
.........3 -ку хл э о ТУЛ ч а
, за
« - -ну .чл о :7j;a з а
ч
нес.г : гч:-:ая ос атазя
15
20
Детальность прогрева при 60°С, vcffl
З -нуклвотидаза AW-аза
5 -нухл80тидаэа
неспвиифтчнал фосфатаэа
-т-
240
-i-
300
J
Ofbtfl ЗЛЮ&ТА , М.Л
оо
СО UD О
О
L. О
Ь- СТ
- 03
U t
(-, S
U U)
s
о о
D R
Pi
C-rJ
2 Г
ногасо;йи
25
Сиг. 6
.,„.. З -нуулеотидаг-а
ОЬъем элвата« кя
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Sung S.C. | |||
| Laskowski M.S | |||
| A nuclease from bean sprouts.- Journal of Biological Chemistry | |||
| Водоотводчик | 1925 |
|
SU1962A1 |
| v | |||
| Прибор для корчевания пней | 1921 |
|
SU237A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| p | |||
| Способ получения бумажной массы из стеблей хлопчатника | 1912 |
|
SU506A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Prentice N. | |||
| Heisel S | |||
| Characterization of nuclease from Barley Shoots.- Phytochemistry | |||
| Пневматический водоподъемный аппарат-двигатель | 1917 |
|
SU1986A1 |
| v | |||
| Видоизменение пишущей машины для тюркско-арабского шрифта | 1923 |
|
SU25A1 |
| p | |||
| Контактный детектор | 1925 |
|
SU2057A1 |
