1
Изобретение относится к области молекулярной биологии, в частности к генной инженерии, и представляет собой способ получения рекомбинатной плазмидной ДНК, содержащей гены рестриктазы и метилазы ,
Известен способ попучения плазмидной рекомбинатной ДНК, включающий обработку донорной и рецийиентной Д«К эндонуклеазой рестрикции, соединение полученных таким образом фрагментов с помоты- полинуклеотидлигазы, трансформацию клеток Esherichid coli рекомбинантными плазмидными ДНК и отбор трансформантов на селективной среде Ij.
Цель изобретения - обеспечение суПерсинтеэа рестриктазы и метилазы.
Для достижения цели в качестве доиорной ДНК используют ДНК плазмиды N 3, в качестве реципентной ДНК плазмиды pIZ 203, а в селективную среду вводят си«1пициллин и сульфаниламид.
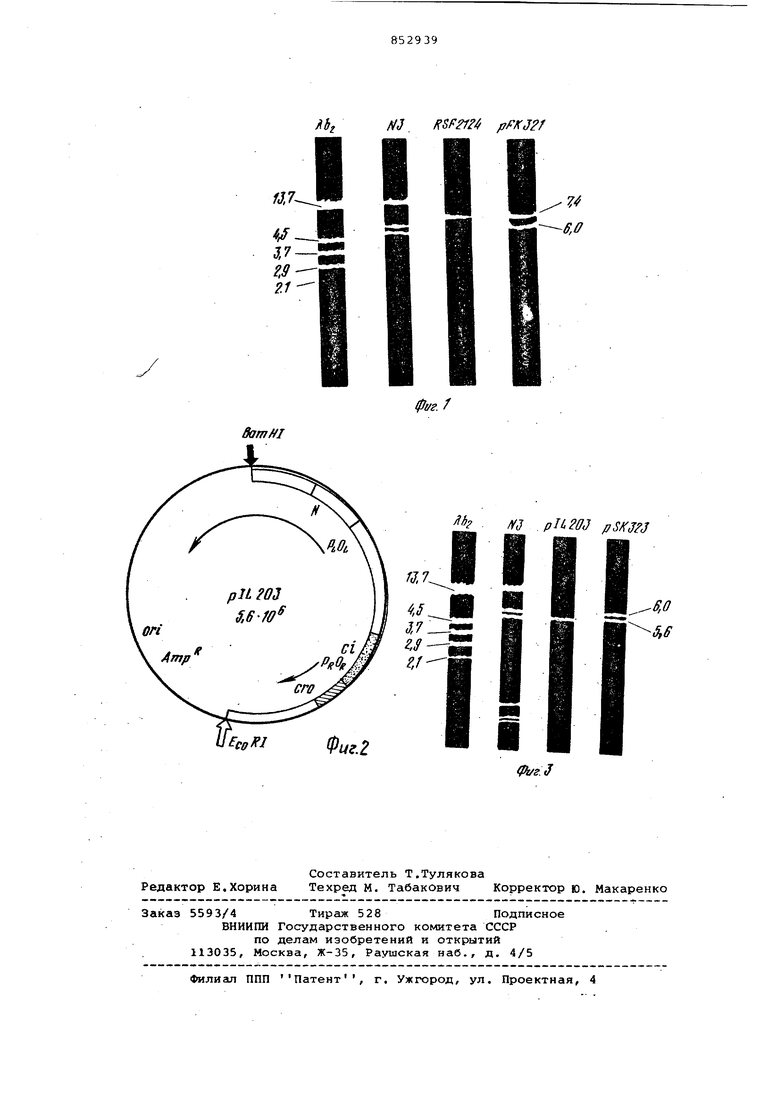
На фиг. 1 приведена диаграмма чадектрофореза в агарозном геле EcoRI фрагментов ДНК фага Л Ь и плазмид: N 3, RSF 2124, pFK 321} цифрами обозначен молекулярный вес фрагментов ДНК в мегадальтонах. На фиг. 2 дана схема плазмиды pIZ 203/Ci 857, сгоб/
.с точкой рестрикции для EcpRI и генетической картой. Стрелками указано направление транскрипции. На фиг, 3 приведена диаграмма электрофореза в агарозном геле EcoRI-фрагментов ДНК ФазаЛ bj и1 плазмид: N 3, pIL 203, pSK 323. Пример.
1.Выделение плазмидных ДНК.
10
Плазмидные ДНК получают центрифугированием осветленного клеточного экстракта 2 с зтидиумбромидом 31.
2.Конструиррдание рекомбинатных плазмидных ДНК.
15
ДНК плазмид рестриктируют Е RI в реакдионной среде,содержащей 0,1 М Трис-НС рН 7,4, 0,01 М MgCI , 0,05 М NaCI, 1 ч при 37 С. К гид;роли3ату ДНК затем добавляют 1/10
20 объема . 5 М NaCI и осаждают 2,5 объемами этанола Осадок ресуспендируют в лигазном буфере (0,666М рис-НС1 рН 7,5, 0,01М MgCI., 1(Г М ЭДТА, 0,01 М дитиоэритритол, 100 мкг
25 мл бычий сывороточный альбумин,10 М АТФ), добавляют лйгазу и инкубируют 10-24 ч при 10°С,
3, конструирование RSF2124 и N3 рекомбинантной ДНК, Высокоочищеииые
3CI ДНК RSF2124 и N3 рестриктируют эндоиуклеазой EcoRI. Продукты гидролиза анализируют электрофорезом в 1%-ном агарозном геле. Для получения рекомбинантных ДНК проводят сшивку EcoRI-j фрагментов ДНК ппаэмкд RSF2124 и N3 В реакцию берут ,6 мкг ДНК RSF2124 и 4 мкг N3, после гидролиза EcoRI и анализа продуктов гидролиза электрофорезом смесь фрагг нтов ДНК переосаждают спиртом, после этого осадок ресуспендируют в 100 мкл лигазного буфера и добавляют 2 мкл лигазы. 4.Трансформация и отбор клонов, содержащих рекомбинантные ДНК. Для трансформации получгиот клетки штамма Е.соИСбОО, обработанные раствором CaCto С 4.Для проверки ком петентности клеток, про.водят трансфекци о ДИК фага лямбда. Эффективность инфекционности составляет 2-510 фаговых колоний мкг/ДНК. К 100 мкл ДНК, полученной после лигазной обработки, добавляют 200 мк клеток, обработанных CaCIa, Смесь инкубируют 30 мин при , затем 2 м при и 10 мин при 24С. Добавляют 2,7 мл питательного бульона, инкубируют 30 мин при с аэрацией. Суспензию высевают на чашки с ан тибиотиками (ампициллин 25 мгк/мп и колицин в соответствующем разведении Выросшие колонии проверяют на способ ность синтезировать колиции, для этого их перекапывают на чашки, подращивают 6 ч при , облучают УФсветом 30 с и засевают чашки индика торной культурой Е.colic 600. Отобранные клоны, не синтезирующие колицин, после перечисленных операций затем проверяют на способность рест риктировать и модифицировать фаг лямбда, 5.Анализ клонов, содержащих RSF 2124 и N3 рекомбинантную ДНК. Отобранные клоны проверяют на рестрикцию и модификацию системой. EcoRII. Степень рестрикции фага лямбда, выращенного на штамме СбОО (г mJ ), определяют сравнением титра фага при посеве его на штаммы E.coIiC 600 (г т+ ) и E.coli НВ110 (г т ) N3 и клоны, содержащие рекомбинантные плазмиды. При высеве ф га на клоны, содержащие рекомбинатные плазмиды, несущие гены Egi-yRII, они ограничивают рост по сравнению со штаммом С600 на два порядка, как и штамм НВ 1100/N3. Фаги, выросшие на этих клонах, содержащих рекомбинантные плазмиды, высевали на штамм С600 и штамм HB1100/N3. Фата, вырос шие на клонах, содержащих ракомбинантные плазмиды, нерущие гены EcoRII, дали одинаковый титр как на штамме СбОО, так и на штамме НВ 1100/N3. В табл. 1 показаны данные рестрикции и модификации одного кло на, содержащего рекомбинантную ДНК с генами EcoRII, обозначенную как РК 321, При проверке этих клонов на стойчивость к антибиотикам, которые, есла плазмида N3, они показывают СТОЙЧИВОСТЬ к сульфаниламиду. Таблица 1 ффективность рестрикции и модификации фага Л клетками Е.соП НВ1100 { nit )/N3 «С600- () FK321 6.Рестрикция рекомбинатных плазИз полученных клонов выделяют плазмидную ДНК и подвергают рестрикции эндонуклеазой EcoRI 1 ч при З7с. затем смесь подогревают при 10 мин, добавляют глицерин до конечной концентрации 20% с бромфеноловым голубым (0,001%). Образцы после гидролиза наносят на 1%-ный агарозный гель и проводят электрофорез при напряжении 10 В/см в течение 2 ч. Используют электрофоретический буфер 0,04 М трис-ацетат, рН 8,00,002 С ЭДТА, 0,002 М ацетат натрия. Гель окрашивают этидиумбромидом и фотографируют в проходящем УФ-свете. Картина электрофоретического разделения фрагментов ДНК показывает, что рекомбинатная плазмида состоит из вектора RSF2124 и включенного в него фрагмента плазмида N3, молекулярный вес которого составляет 6,0-1(1 Дальтон (фиг. 1). 7.Клонирование генов Е.11 в векторе pIZ203. Обнаружение после клонирования ЕСО RI фрагмента плазмиды N3 с генами RII свидетельствует о том, что этот фрагмент наряду с генами несет устойчивость к антибиотику сульфаниламиду, это дало возможность клонировать этот фрагмент в векторе i- pIL203, который не имеет селекции при встройке чужеродных Е RI фрагментов, если эти фрагменты не несут селектируемых маркеров. Вектор p3L 203 получен на основе плазмиды PBR и BamHI-Ej RI фрагмента фага лямбда, в котором находятся левый и правый промоторы, кроме того, в этом фрагменте содержится температурочувствительная мутация в гене репрессора CJ и супрессируемая аббер-мутация гене его. Таким образом, используя вектор PIL203, можно увеличить Доз и количество транскриптов клонируе мых генов. Плазмидные ДНК N3 и pIL 203 пол чают центрифугированием в CsCI, ре стриктируют эндонуклеазой Е RI и анализируют рестриктированную ДНК электрофорезом в агароэном геле (фиг. 3). Рестриктированные ДНК N3 (4 мкг) и pIL 203 (4 мкг) смешивают переосазкдают спиртом и ресуспендируют в лигазном буфере. После доба ки ли газы (2 мкл) смесь инкубируют 16 ч при 10С. Лигированную ДНК трансформируют в Elcoli (802/г т Трансформанты отбираиот по устойчиво ти к с1мпициллину и сульфаниламиду. Выросшие колонии проверяют на способность рестриктировать и модифици ровать фаг лямбда. Таким образом, отбирают клоны, которые проявляют устойчивость к ампициллину и сульфаниламиду и могут рестриктировать модифицировать фаг лямбда со специфичностью системы . Данные по рестрикции и модификации клона, нес щего такую рекомбинантную ДНК, обоз наченную как pSK 323, представлены в табл. 2. Таблица 2 Эффективность рестрикции и модифик ции фага клетками E.coii -о .4 Л -802()1 то 1-10 Л- 802{г;т)ЫЗ1 Л. .802(г-тМ PSK3231 Выделения плазмида ДНК из этого клона после рестрикции эндонуклеазо Ejjp RI образуют два фрагмента при электрофорезе, соответствующие по молекулярному весу вектору pIL 203 фрагменту плазмифл N3, молекулярный вес которого 6,0 10 дальтон. 8, Экспрессия растриктазы и мети лазы в штаммах, содержащих рекомбинантную плазмиду pSK323. Плазмида pIL 203, используемая качестве донора при получении реко бинатной ДНК pIL 203-3, несет терм чувствит ьную мутацию в гене репрессора Ci и супрессируемую аббермутацию в гене CRO, которые приводя к понижению репрессии транскрипции с фаговых промоторов. В зависимости от бактериального штамма наличие продукта гена CRO колеблется от 1 (полная супрессия) до уровня . Таким образом, можно увеличить транскрипцию как за счет температурных условий культивирования, так и за. счет использования штаммов Е coli, содержащих различные супрессоры, для трансформации рекомбинатной плазмидой. Трансформировали рекомбинатную плазмиду pSK 323 в штаммы Elcoli, несущие различные супрессоры. После выращивания этих штаммов при (при этой температуре резко снижен синтез репрессора Ci) была проанализирована активность рескриктазы и метилазы Е RII в бесклеточных экстрактах зтих штампов. 9. Определение активности рестриктазы и метилазы . Клетки Е.соП выращивают до поздней логарифмиче.ской фазы. Клетки собирают центрифугированием и ресуспендируют в РЕМд буфере (0,01 М KjPOji, , рН 7,0 мМ ЭДТА, 7 мМ 2-меркаптоэтанол, 5% глицерин). Клетки разрушают ультразвуком. После разрушения центрифугируют при 100000 g 1ч при . Бесклеточный экстрат используют для определения ферментативной активности. Активность метилазы определяют в реакционной смеси общим объемом 150 мкл, содержащей 40 мМ КНРОд рН 8,0, 1 мМ ЭДТА, 7 мМ 2-меркаптоэтанола, 20 мкг ДНК спермы лосося. 4 мкМ (метил-н) S-аденозилметионина и 50 мкл фермента при различных разведениях . Реакцию проводят при в течение 1 ч, добавляют равный объем 0,75 М натриевой щело. чи, инкубируют 30 мин при . Затем к пробам добавляют 2 мл и 2мл 10% ТХУ. Осажденную ДНК наносят на фильтры, промывают 0,01 М HCI и спиртом. После просушки фильтров считают радиоактивность. Для опрег деления специфичности включения метки в азотистые основания ДНК последнюю элюируют с фильтров, гидролизуют до азотистых оснований 57%-ной хлорной кислотой и азотистые основания разделяют хроматографией на бумаге. Радиоактивность азотистых оснований определяют в вырезанных образцах бумаги б. Активность рестриктазы Ecohl I -определяют в буфере{, содержащем 1 мкг ДНК плазмиды pBR322, 50 мМ трис-НС рН 7,5, 10 мМ MgCIj , 10 мМ 2-меркаптоэтанола 1 мМ ЭДТА и фермент в различных разведениях. Общий объем смеси составляет 40 мкл, реакцию проводят при 37°С 15 мин. Полученные фрагменты ДНК анализируют электрофоре зом в 1%-ном агарозном пластинчатом геле и трис-ацетатном буфере f6.
Результаты этих опытов представлены в таОл. 3 и 4.
Таблица 3
Активность ме илаэы в Сесклеточных экстрактах штаммов, содержащих плаэмиду N3 и .pSK 323.
W3350 (Su)
С R 63 (SuI)l,
802 (SuIII) 1,1.10
Таблица 4
Активность рестриктазы RII в в бесклеточных экстрактах штаммов, содержащих плаэмиду N3 и pSK 323 W 3350 3200 W 3350 (SU-) С R 63 2000 С R 63 (Sul) (Sul) 8021500 802(SuIIl) 200 (SuIII) За единицу активности фермен .принимали количество фермента которое необходимо для полного гид лиза 1 мкг ДНК плаэмнды рВН 322 в течение 15 мин при ,
Как видно из этих данных, наибольший синтез рескриктазы и метилазы наблюдается, когда рекомбинантная плазмида находится в штамме W 3350 (S|J ) . Активность рескриктазы и мее тилаэы в этом штамме в 15 раз увеличена по сравнению с контрольным штаммом W 3350/N3/.
Таким образом, полученная ракомбинантная плазмидная ДНК, несущая . « гены рестриктазы и метилазы , содержащаяся в различных штаммах Е . со)i .Обеспечивает суперсинтез ферментов Ef fjRII бактериальной клеткой,
Формула изобретения
способ получения плазмидной рекомбинатной ДНК, содержащей гены рескриктазы и метилазы , включающий обработку донорной и реципиентной ДНК, эндонуклеазой рестрикции, соединение полученных таким образом фрагментов с помощью полинуклеотидлигазы, трансформацию клеток Escheri5 chia coli рекомбинатными плазмидными ДНК и отбор трансформантов на селективной среде, отличаюиийс я тем, что, с целью обеспечения суперсинтеэа рестриктазы и метилазы, в качестве донорной ДНК используют ДНК плазмиды N3, в качестве реципиентной ДНК плазмиды pZ203, а в селективную среду вводят Ампициллин и сульфаниламид. Источники информации, принятые во внимание при экспертизе 1.Заявка Франции 2281426, кл. С 12 К 3/00, 1976. 2.Guerry Р. et а1тj. Bacter., 1973, v.H6, 1064-1066 3.Tanaka Т. etal- 5.Bacter- , 1975, 121, 354-362. 4.Mandel M. etal.-j.Mol. Biol 1970, V.5, 159-162. 5.Солонин и др. ДАН СССР, 1979, Т. 245. 722-725. 6.Бурьянов И.И. и др. ДАН СССР 1977, т. 237, 465-468.
AJ. RSF 



Bamf/I /KJ pIl.20J
