Изобретение относится к микробиологии, биотехнологии и микробиологической промышленности и представляет собой способ защиты от фаголизиса культур Escherichia coli, ряд объектов для его осуществления.
Целью предлагаемого изобретения является повышение устойчивости культур E. coli к различным бактериофагам, в том числе к бактериофагам, обладающим антирестрикционным механизмом.
Клетки трансформируют рекомбинантными плазмидами, содержащими Bgl II-фрагмент ДНК области иммунитета фага лямбда, обеспечивающий повышенный уровень экспрессии генов rex AB и вследствие этого устойчивость клеток к колифагам.
Для осуществления способа сконструирована рекомбинантная плазмида pIL 323 с областью иммунитета фага лямбда, содержащая репликон плазмиды р15А.
В способе применяется рекомбинантная плазмида pIL RV8, содержащая область иммунитета фага лямбда с интактными генами rex AB и генами, кодирующими систему рестрикции-модификации EcoRV, обеспечивающую дополнительную устойчивость к бактериофагам.
В рекомбинантных плазмидах, применяемых в способе, используется Bgl II-фрагмент ДНК области иммунитета фага лямбда, который в составе рекомбинантной ДНК определяет повышенную резистентность культур к фаголизису.
П р и м е р 1. Конструирование плазмиды pIL 323 из ДНК плазмиды pACУC 184 и ДНК фага лямбда с 1857.
Клетки бактерий E.coli К 802, содержащие плазмидную ДНК рАСУС 184, выращивают в 10 мл бульона LB при 37оС до титра 1х108 кл/мл и выделяют из них плазмидную ДНК по следующему методу.
Клетки осаждают центрифугированием (5000 g, 5 мин 4оС) и ресуспендируют в 0,1 мл лизирующего раствора (лизоцим 2 мкг/мл, трис-НСl25 мМ, рН 8,0, ЭДТА 10 мМ, глюкоза 50 мМ, выдерживают 5 мин при 0оС. Далее прибавляют 0,2 мл раствора (0,2 М додецилсульфат натрия, 1%) и перемешивают до осветления лизатов. 150 мкл раствора 3 М ацетата калия (рН 4,8) вносят в осветленные лизаты, осторожно перемешивают до заметного снижения вязкости раствора, выдерживают 1 ч при 4оС и образовавшиеся осадки удаляют центрифугированием (15000 g, 5 мин 4оС). К полученным супернатантам добавляют 2,5 объема охлажденного при -20оС этилового спирта и выдерживают 2 ч при -20оС. Образовавшиеся осадки собирают центрифугированием (1500 g, 5 мин, 120оС) и суспендируют в 0,2 мл буфера (10 мМ трис-HCl, рН 8,0, 1 мМ ЭДТА). Полученные препараты ДНК используют для конструирования плазмид.
ДНК фага лямбда выделяют по методике, описанной ранее.
Конструирование плазмид проводят следующим образом: ДНК плазмиды раСУС 184 и ДНК фага лямбда смешивают в соотношении 1:20 (1 мкг и 20 мкг) в 100 мкл раствора и смесь ДНК (21 мкг) гидролизуют одновременно эндонуклеазами Bgl II и BamHI (по 10 ед соответственно) в растворе, содержащем 50 мМ трис-HCl, рН 7,6; 10 мМ MgCl2 мМ 2-меркаптоэтанола; 50 мМ NaCl, при 37оС 1 ч. После прогревания реакционной смеси при 65оС в течение 10 мин проводят соединения фрагментов ДНК ДНК-лидазой фага Т4 в присутствии АТФ (500 мкм) при 10оС в течение 8 ч.
Смесью образовавшихся молекул ДНК трансформируют клетки E.coli DH 1 следующим образом. 0,1 мл суспензии клеток E.coli вносят в 10 мл питательного бульона и выращивают до титра 3х108 кл/мл. После 10 мин охлаждения (0оС) клетки собирают центрифугированием (15000 g, 10 мин, 0оС), суспендируют в 10 мл 0,1 М раствора CaCl2 и выдерживают при 0оС 1 ч. Клетки повторно центрифугируют (5000 g, 10 мин, 0оС), затем суспендируют в 0,5 мл 0,1 М CaCl2 (0oC) и используют для трансформации. Смесь соединенных фрагментов ДНК инкубируют с компонентными (обработанными CaCl2) клетками 1 ч при 0оС и 10 мин при комнатной температуре. После десятикратного разбавления бульоном LB клетки подращивают 2 ч и высевают на агаризованную среду LB, содержащую 30 мкг/мл хлорамфеникола, устойчивость к которому определяют векторная плазмида рАСУС 184. Выросшие трансформанты проверяют в spot-тесте на наличие иммунитета к фагу λ 126 и на чувствительность к тетрациклину, так как вставка чужеродного фрагмента в Bam HI-сайт рАСУС 184 приводит к инактивации гена, определяющего устойчивость к тетрациклину.
Из клеток клонов, резистентных к хлорамфениколу, иммунных к фагу лямбда с126 и чувствительных к тетрациклину, выделяют ДНК по описанной выше методике и анализируют с помощью эндонуклеазы рестрикции Hind II.
Этот анализ позволяет определить, что фрагмент Bg1 II (2,392 пар оснований) встроился в обеих ориентациях. Вариант плазмиды, в которой направления тетрациклинового промотора и промотора фага лямбда Pr противоположны, был назван pIL 323.
П р и м е р 2. Способ защиты клеток E.coli В 834 от фаговой инфекции.
Клетки E.coli В 834 выращивают в 10 мл мясопептонного бульона (МПБ) до плотности 4х108 кл/мл, осаждают центрифугированием 10 мин при 0оС (К-23, 6000 об/мин), промывают дважды 20 мл холодного раствора, содержащего трис-HCl 0,01 М, рН 7,7; CaCl2 0,1М, ресуспендируют в 10 мл этого раствора и выдерживают 20 мин в ледяной бане, ресуспендируют в 1 мл раствора. К 0,3 мл суспензии клеток добавляют 20 мкл раствора ДНК плазмиды pIL 323 с концентрацией 50 мкг/мл, инкубируют 50 мин при 0оС, 5 мин при 42оС, добавляют два объема среды LB, выдерживают 30 мин при 37оС и высевают на агар LB, содержащий 30 мкг/мл хлорамфеникола. Выросшие клоны отбирают и определяют им устойчивость к бактериофагам.
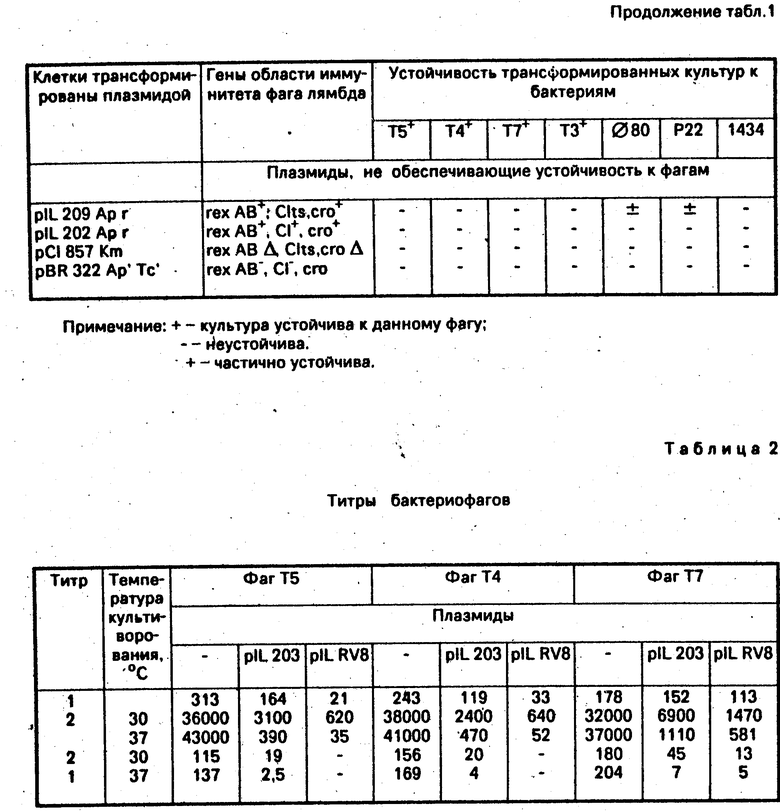
А. Бактериофаг Т5+, выращенный на культуре E.coli В 834, высевают на газон культуры E.coli В 834, трансформированной плазмидой pIL 323. К 2 мл мягкого агара. приготовленного на среде LB с добавкой 10 мМ CaCl2, добавляют при 37оС 1 х 108 свежих клеток в 0,2 мл среды и по 102 частиц фага Т5+ на первые две чашки, по 104- на вторые две чашки и по 107- на третьи две чашки. Для контроля фаг Т5 высевают аналогично на чашки с газоном культуры E. coli В 834 без плазмиды. Чашки инкубируют при 37оС 16 ч и определяют эффективность высева фага в опытных чашках и контрольных. На газоне в опытных чашках не видно негативных колоний фага Т5. Трансформированные клетки устойчивы к фагу Т5. По аналогичной схеме проверяют культуру E.coli В 834, трансформированную плазмидой рIL 323 на устойчивость к бактериофагам Т4, Т7, Т3, ⊘ 80 , Р2 и λimm 434. В результате проведенного анализа показано, что плазмида pIL 323 обеспечивает полную устойчивость к фагам ⊘ 80 , Р22, Т4, Т5, частичную к фагам Т3 и Т7 (размер негативных колоний на чашках уменьшается с 10 мм (контроль) до 1 мм (опыт) и не обеспечивает устойчивости к фагу лямбда i 434 (см. табл. 1).
Б. Зависимость устойчивости клеток E.coli В 834/pIL 323 к бактериофагам от температуры культивирования.
Способ осуществляют аналогично примеру 2, используя клетки E.coli В 834, трансформированные плазмидой pIL 323, чашки с высеянным фагом Т5 инкубируют при температуре 26, 32 и 38оС.
При 26оС культура E.coli частично устойчива к фагу Т5. На газоне опытных чашек - мелкие негативные колонии фага в количестве около 10% от числа колоний в контрольных чашках с культурой E.coli В 834, не содержащей плазмиды pIL 323.
При 32оС культура E.coli частично устойчива к фагу Т5. На газоне опытных чашек видны очень мелкие колонии фага (около 0,1% от числа колоний на контрольных чашках).
При 38оС культура E.coli устойчива к фагу т/5. На газонах опытных чашек не видно колоний фага т5.
При анализе зависимости устойчивости клеток E.coli В 834 pIL 323 к бактериофагу Т4 от температуры культивирования показано, что по сравнению с контрольными чашками при 26оС на газоне опытных чашек наблюдают 10% мелких фаговых колоний, при 32оС около 3% очень мелких, при 38оС колонии не видны.
В. Определение устойчивости клеток E.coli В 834/pIL 323 к бактериофагам при культивировании в жидкой среде.
К культуре клеток E.coli В 834, трансформированных плазмидой pIL 323, с плотностью 5х107 кл/мл в МПБ добавляют фаг Т5+ с множественностью 0,3 и инкубируют при 37оС в условиях интенсивной аэрации 5 ч. За это время культура выходит в стационарную фазу, лизиса культуры не наблюдают так как клетки устойчивы к бактериофагу Т5.
Аналогично проводят определение устойчивости клеток E.coli к фагам Т4, Т3, Т7, ⊘ 80 . Р22, λimm 434. В случае фагов Т4. ⊘ 80 и Р22 лизиса не наблюдают, так как клетки устойчивы к перечисленным бактериофагам. В случае фагов Т3 и Т7 лизис культуры E.coli В 834, трансформированной плазмидой pIL 323, наступает значительно позднее, чем лизис незащищенной плазмидой культуры E.coli В 834, из-за частичной устойчивости клеток. В случае фага λimm 434 наблюдают обычный лизис культуры.
П р и м е р 3. Трансформацию клеток E.coli С 600 плазмидой pIL 323 проводят аналогично трансформации клеток E.coli В 834, как описано в примере 2.
А. Определение устойчивости клеток E.coli С 600/pIL 323 к бактериофагам на агаризированной среде проводят, как описано в примере 2. пункт А. В результате анализа показано, что клетки E.coli C 600/pIL 323 устойчивы к фагам Т4 и Р22, частично устойчивы к фагам Т3 и Т7,к фагу λimm 434 клетки не устойчивы. Чувствительность клеток С 600/pIL 323 к бактериофагам Т5 и ⊘ 80 не анализировали, так как клетки E.coli С 600 устойчивы к этим фагам за счет ton A мутации.
Б. зависимости устойчивости клеток E.coli С 600/pIL 323 к бактериофагам Т4, Т7 и Р22 от температуры культивирования, проводимой, как описано в примере 2, пункт Б показано, что 37оС является оптимальной температурой культивирования, при которой клетки E.coli C 600/pIL 323 наиболее устойчивы к бактериофагам Т4, Т3, Т7 и Р22.
В. При определении устойчивости клеток E.coli С 600/pIL 323 к фаговой инфекции в жидкой среде, которая определялась, как описано в примере 2, пункт В, показано, что культура устойчива к бактериофагам Р22, Т4, частично устойчива к фагам Т3 и Т7 и не устойчива к фагу λimm434.
П р и м е р 4. Способ осуществляют аналогично способу, описанному в примере 2, используя вместо клеток E.coli В 834 клетки E.coli C.
Проведенный анализ позволяет заключить, что культура E.coli C/pIL 323 устойчива к бактериофагам Т4, Т22, Т5, ⊘ 80 , частично устойчива к фагам Т7 и Т3 и чувствительна к бактериофагу λimm 434 при культивировании на агаризованной среде, а также и в жидкой среде.
П р и м е р 5. Способ осуществляют аналогично способу, описанному в примере 2, используя вместо плазмиды pIL 323 плазмиду pIL RV8.
А. Проведенный анализ позволяет заключить, что культура E.coli В 834/pIL RV8 устойчива ко всем используемым в данной работе бактериофагам, в том числе и к λimm 434 и к Т3, и частично устойчива к Т7 (менее 50% мелких колоний с диаметром около 1 мм). Ограничение фагов Т3 и λimm 434 определяется наличием в плазмиде генов системы рестрикции-модификации EcoRV и присутствием сайтов, узнаваемых этой эндонуклеазой, на ДНК этих бактериофагов. Фаг Т3 ограничивается активностью генов rex AB фага лямбда и системой рестрикции EcoRV, фаг Т7, который не имеет на своей ДНК сайтов EcoRV, только активностью генов rex AB, а фаг λimm 434 ограничивается только активностью гена, кодирующего эндонуклеазу рестрикции EcoRV.
Б. Зависимость устойчивости культур E.coli В 834/pIL RV8 к бактериофагам от температуры определяли, как описано в примере 2, пункт Б.
Показано, что при 26оС на агаризованной среде падение титра фага Т5 оставляет около 90%, при 32оС падение титра достигает 99,9%, при 38оС культура полностью устойчива к фагу Т5.
В. Определение устойчивости клеток E.coli D 834/pIL RV8 к бактериофагам в жидкой среде проводят, как описано в примере 2 пункт В. Показано, что клетки E.coli B 834/pIL RV8 устойчивы к фагам Т4, Т5, Т3, ⊘ 80 , λimm 434, Р22.
Г. Определение урожая фага Т5.
Клетки E. coli B 834/pIL RV8 выращивают в 10 мл МПБ до плотности 3х108 кл/мл, осаждают центрифугированием (6000g, 2oC 10 мин) и ресуспендируют в 1 мл МПБ. В суспензию клеток добавляют фаг Т5 с множ. 0,5-0,8 и проводят адсорбцию фага 10 мин при 10оС. В этих условиях на клетках адсорбируется свыше 90% всего фага (в среднем 95%). Затем быстро делают разведения клеток в 106 раз в пробирках с холодным МПБ (10оС) так, чтобы в последней пробирке было 3х103 кл/мл. Из этой пробирки отбирают аликвоты по 0,1 мл для титрования фага (титр 1) на газоне E.coli В 834 при 37оС, затем содержимое пробирки делят пополам, нагревают на водяной бане одну пробу до 30оС, вторую - до 37оС и инкубируют пробы при указанной температуре и усиленной аэрации 1,5 ч. Затем отбирают аликвоты по 0,1 мл из проб для второго титрования (титр 2).
Титр 1 равен сумме инфицированных клеток, дающих фаговое потомство, и неадсорбированных инфекционных фаговых частиц.
Титр 2 соответствует урожаю фага из инфицированных клеток. Отношение титр 2/титр 1 приближенно характеризует выход фага на 1 клетку. Результаты одного из трех независимых опытов представлены в табл. 2, в которой приведены также данные, полученные при анализе урожая фагов Т4 и Т7, а также при анализе урожая фагов Т5, Т4 и Т7, выросших на культуре E.coli В 834 без плазмиды и с плазмидой pIL 203, которая содержит область иммунитета фага лямбда с интактными генами rex AB и мутантными генами репрессоров с 1857 и croam6.
Из данных табл. 2 следуют выводы.
1. В клетках E.coli B 834, трансформированных плазмидами с повышенной активностью генов rex AВ, выход фагового потомства значительно ниже, чем в бесплазмидных клетках.
2. При повышении температуры культивирования с 30 до 37оС выход фага из клеток E.coli В 834, трансформированных плазмидами pIL 203, pIL RV8, значительно снижается.
3. При инфицировании клеток E.coli B 834, трансформированных плазмидой pIL RV8, фагами Т4 или Т5 при 37оС инфицированные клетки не дают фагового потомства, т.е. инфекция в этих условиях протекает абортивно.
4. Плазмида pIL RV8 сильнее подавляет рост фагов по сравнению с pIL 203, что возможно объясняется слабой супрессией мутации am 6 гена cro в штамме E.coli B 834.
П р и м е р 6. Способ осуществляют аналогично способу, описанному в примере 5, используя вместо клеток E.coli B 834 клетки E.coli с 600. Проведенный анализ позволяет заключить, что клетки E.coli C600/pIL RV8 устойчивы к бактериофагам Т4, Т5, ⊘ 80 , Т3, Р22, λimm 434, к фагу Т7 наблюдают частичную устойчивость при выращивании культур на агаризованной и жидких средах. Урожай бактериофагов Т5, Т4 и Т7 падает соответственно в 100, 1000 и 10 раз.
П р и м е р 7. Способ осуществляют аналогично способу, описанному в примере 5, используя вместо клеток E.coli В 834 клетки E.coli C. Приведенный анализ позволяет заключить, что культура E.coli C/pIL RV8 устойчива ко всем используемым бактериофагам, кроме фага Т7 при культивировании как в жидкой среде, так и на агаризованной сpеде. К фагу Т7 наблюдают частичную устойчивость. Эффективность защиты от фаголизиса зависит от температуры, оптимальная температура для осуществления способа 37-38оС. Урожай бактериофагов при росте на культуре E.coli C/pIL RV8 резко понижен по сpавнению с урожаем при росте на бесплазмидном штамме.
П р и м е p 8. Способ осуществляют аналогично способу, описанному в примере 2, используя вместо плазмиды pIL 323 плазмиду pFRD 35.
Трансформацию клеток E.coli В 834 плазмидой pFR 35 проводят, как описано в примере 2. Селективная среда содержит 20 мкг/кг ампициллина. Плазмида pFR 35 размером 7,85 т.п.о. содержит BamHI-фрагмент ДНК рВR 322, BgL II-фрагмент ДНК области иммунитета фага лямбда с 1857 размером 2,39 т.п.о., Hind III-фрагмент ДНК фага Т4 размером 1,1 т.п.о с геном frd, кодирующим дигидрофолатредуктазу, которая определяют устойчивость клеток к триметаприму, ген беталактамазы, определяющий устойчивость клеток к ампициллину, репликон pMBI. Направление транскрипции с промторов Pr и Pfrd конвергентно.
Проведенный анализ показал, что клетки E.coli B 834 достаточно устойчивы при температуре 37оС к бактериофагам Т5, Т4, ⊘ 80 и Р22, частично устойчивы к фагам Т7 и Т3 при росте как в жидкой, так и на твердой среде при наличии в клетках плазмиды PFRL 35.
П р и м е р 9. Способ осуществляют аналогично способу, описанному в примере 2, используя вместо плазмиды pIL 323 плазмиду pIL 203. Проведенный анализ показал, что эта плазмида повышает устойчивость клеток к фаговой инфекции. Урожай фагов при росте на E.coli В 834/pIL 203 при 37оС в среднем приблизительно в 100 раз ниже урожая при росте на бесплазмидном штамме в случае фагов Т4 и Т5 и в 10-20 раз ниже в случае фага Т7 (см. табл. 2).

Изобретение относится к биотехнологии, в частности к технологии рекомбинантных ДНК, и позволяет защитить культуры E.coli - продуценты белков, гормонов, незаменимых аминокислот и т.п. при крупномасштабных ферментациях от случайных фаговых инфекций, приводящих к лизису клеток и потере целевого продукта. С целью повышения устойчивости культур E.coli к различным бактериофагам проводят трансформацию клеток E.coli рекомбинантными плазмидами, содержащими фрагмент ДНК области иммунитета фага лямбда с интактными генами rex AB, ts 857, мутацией в гене cz и мутацией в структурной части гена cro, обеспечивающий в составе плазмиды повышенную экспрессию генов rex AB фага лямбда по сравнению с лизогенной культурой. Повышенная экспрессия генов rex AB подавляет развитие различных бактериофагов в клетках вплоть до абортивной инфекции. Сконструирована защитная плазмида pIL 323 с репликоном p15A группы несовместимости incY, содержащая Bgl II-фрагмент ДНК области иммунитета фага лямбда. В качестве средств, обеспечивающих повышение устойчивости к колифагам , используются рекомбинантная плазмида pIL R 8 и Bgl II-фрагмент ДНК области иммунитета фага лямбда. 1 з.п.ф-лы, 2 с.п.ф-лы, 2 табл.
- Bgl II-фрагмент ДНК области иммунитета фага лямбда, ограниченной сайтами Bgl II с координатами на ДНК фага лямбда 35711 и 38103 п.н.,
- Bam HI-фрагмент плазмиды pACYC 184,
- уникальные сайты рестрикации EcoRI, EcoRV, SalI, ClaI,
- четыре сайта для Hind III, разделяющие плазмиду на фрагменты 125, 564, 990, 4700 п.н.,
- гены rex AB фага лямбда,
- ген Cl с термочувствительной мутацией 857,
- генетический маркер - ген хромфениколацетилтрансферазы, определяющий резистентность к хлорамфениколу,
- репликон p15A со строгим контролем группы несовместимости.
| Патент США N 453904, кл.435-172, 3, 1985. |
