иокислоты) и коинтибитору (определенной аминокислоте) 4.
Во всех перечисленных случаях мутантные штаммы, способные продуцировать, аминокислоты, получают за счет однократного или последовательного индуцирования мутаций в генетической структуре (геноме) исходного штамма, не выделяюш,его аминокислоту. До настоящего времени неизвестны штаммы, у которых возрастание продукции аминокислоты достигается за счет увеличения дозы генов, необходимых для ее биосинтеза, или в результате введения в клетку чужеродного генетического материала.
Некоторые практические аспекты применения методов генетической инженерии нашли свое отражение в известном способе получения штаммов Pseudomona§, обеспечивающих деградацию комплекса ортанических веществ (углеводородов нефти) 5. В этой работе гибридные молекулы были получены in vivo путем внутриклеточной рекомбинации.
Однако методы получения штаммов продуцентов аминокислот с использованием приемов генетической инженерии до настоящего времени не разработаны.
В качестве прототипа выбран метод получения штамма Е. соИ ВНИИгенетика 14MG--442, полученного под действием нитрозогуанидина из штамма Е. соИ К12 и отобранного по признаку устойчивости к аналогу треонина р-оксинорвалину и способности продуцировать L-треонин) 6.
Рассматриваемый в качестве прототипа метод предусматривает мутагенную обработку клеток исходного штамма и отбор нужных мутантов, т. е. способных к продукции аминокислоты, на среде со структурньш аналогом аминокислоты, ингибирующим рост немутадтных клеток.
Такой метод позволяет получить продуцирующий аминокислоту штамм, не нуждающийся в дополнительных ростовых факторах (например аминокислотах, витаминах и т. д.). Данный признак был определяющим при выборе прототипа.
Однако основным недостатком такого метода остается сравнительно низкая продуктивность полученных штаммов-продуцентов ряда аминокислот, в-частности продуцентов- L-треокина.
Целью изобретения является увеличение выхода L-треонина.
Для достижения указанной цели получены два штамма-штамм Е. соИ ВНИИгенетика VL334py № 6 и ВНИИгенетика VL334py № 7 - продуценты L-треонина со следующими характеристиками.
Характеристика штамма Е. соИ ВНИИгенетика VL334py № 7.
ШтаммЕ. coliВНИИгенетика
VL334py № 7, продуцирующий L-треонин, хранится в Центральном музее промьгшленных микроорганизмов института «ВНИИгенетика и имеет регистрационный номер ЦМПМ В-1684.
Морфология. Грамотрицательные слабоподвижные тонкие палочки с закругленными концами, размером 1,5-2 мкм в, длину.
Культурально-физиологические признаки.
Мясо-пептонный агар. Через 24 ч роста при 37°С образует колонии диаметром 2- 3 мм, круглые, слегка выпуклые с гладкими краями, структура однородная, прозрачные на свет, консистенция пастообразная, легко- эмульгируются.
Агаризованная минимальная среда (Адамса) с глюкозой (0,2%) и минер альным азотом. Через двое суток роста при 37°С образует колонии диаметром 1,5- 2 мм серовато-белые, круглые с ровными краями, слегка выпуклые, внутренняя структура однородная.
Рост в мя-со-пептоннюм бульоне - после 24 ч роста при 37°С сильное равномерное помутнение, небольшой осадок, запах характерный.
Рост в жидкой минимальной ср-еде Адамса - через одни сутки роста при 37°С с аэрацией сильное равномерное помутнение, запах отсутствует.
Рост по уколу в мясо-пептонном агаре - хороший по всему уколу.
Желатину не разжижает.
На молоке хороший рост с коагуляцией молока.
Индол образует.
Рост на различных углеводах, хорошо растет на глюкозе, лактозе, маннозе, галактозе, ксилозе, фруктозе, глицероле и маннитоле с образоваплсм кислоты и газа.
Потребность в факторах роста. Частичная потребность в изолейцине. Время генерации на глюкозо-минеральной среде (Адамса) с изолейцином 60 мин, на среде без изолейцина 270 мин.
Устойчивость к антибиотикам. Устойчив к пенициллину.
Штамм не пат.огенен.
Содержание плазмиды. В логарифмической стадии роста клетки содержат около 17 копий плазмидьг рУ № 7 (мол. вес. 5,7 мегадальтон), обеспечивающей устойчивость штамма к пенициллину и несущей гены треонинового оперона.
Характеристика штамма Е. coli ВНИИгенетика VL334py № 6.
ШтаммЕ. соИВНИИгенетика
VL334py № 6, продуцирующий 1-теронин, хранится в Центральном музее промышленных микроорганизмов института «ВНИИгенетика и имеет регистрационный номер В-1649.
Морфология. ГрамотрИцательные слабоподвижные тонкие палочки с закругленными концами, размером 1,5-2,0 мкм в длину.
Культурально-физиологические признаки.
Мясо-пептонный агар. Через 24 ч роста при 37°С образует колонии диаметром 2- 3 мм, круглые, слегка выпуклые с гладкими краями, структура однородная, прозрачные на свет, консистеиция настообразная, легко эмульгируются.
Агаризованная минимальная среда (Адамса) с глюкозой (0,2%) и минеральным азотом. Через двое суток роста нри 37°С абр азует колонии, диаметром 1,5- 2 мм серовато-белые, круглые с ровными краями, слегка выпуклые, внутренняя структура однородная.
Рост в мясо-п-ептонном бульоне - после 24 ч росте при 37°С сильное равномерное помутнение, небольшОй осадок, запах характерный.
Рост Б жидкой минимальной среде Адамса с необходимыми добав.ками - через одни сутки роста при 37°С с аэрацией сильное равномерное помутнение, запах отсутствует.
Рост по уколу В мясо-пептонном агаре - хороший ПО всему уколу.
Желатину не разжижает.
На молоке хороший рост с коагуляцией молока. .
Индол образует.
Рост на различных углеводах: хорошо р астет на глюкозе, лактозе, маннозе, галактозе, фруктозе, глицероле и маннитоле с образованием кислоты и газа.
Потребность в факторах роста. Частичная потребность в изолейциие. Время генерации на глюкозо-минеральной среде (Адамса) с изолейцином 60 мин, на среде без изолейцина 270 мин.
Устойчивость к антибиотикам. Устойчив к пенициллину и тетрациклину.
Штамм не патогенен.
Содержание плазмы. В логарифмической стадии роста клетки содержат около 10 копий плазмиды рУ № 6 (мол. вес 11,2 мегадальтон), обеспечивающей устойчивость штамма к пенициллину и тетрациклину и несущей гены треонинового оперона.
Для достижения поставленной цели в качестве фпагмента ДНК используют ДНК хромосомы донорного микроорганизма, содержащие гены, контролирующие синтез L-треонина и имеющие мутацию, нарушающую негативную регуляцию синтеза данной аминокислоты, а в качестве реципиентного штамма используют шта1мм, имеющий мутацию, частично блокирующую смежный этап метаболизма указанной аминокислоты.
Кооме того, объединение фрагмента ДНК хромосомы донорного штамма Е. соИ осуществляют плазмидой PBR 322, имеющей молекулярный вес 2,8 мегадальтон in vivo.
Кроме того, используют фр агмент ДНК хромосомы донорного щтамма Е. соИ, содержащий гены треонинового оперона, у которого в результате мута-ции ферментпродукт гена thr А устойчив к ингибированию треонином.
Кроме того, в качестве реципиентного щтамма используют штамм Е. соИ ВНИИгенетика VL 334, имеющий двойную ауксотрофность по треонину и изолейцииу.
Кроме того, трансформацию реципиентного щтамма Е. соИ ВНИИгенетика VL 334 проводят гибридной плазмидой рУ № 6, имеющей молекулярный вес 11,2 мегадальтон, состоящей из двух молекул плазмиды рВ R 322 с молекулярным весом 5,6 мегада-льтон и содержащей функционирующие гены треонинового оперона фрагмента ДНК хромосомы донорного штамма Е. соИ.
Кроме того, трансформацию реципиентного щтамма Е. соИ ВНИИгенетика VL 334 проводят гибридной плазмидой рУ № 7, имеющей молекулярный вес 5,7 мегадальтон, состоящей из плазмиды
рВ R 322 с молекулярным весом 2,8 мегадальтон и содержащей функционирующие гены треонинового оперона фрагмента ДНК хромосомы донорного штамма Е. соИ.
Пример 1. Конструирование щтамма Е. соИ ВНИИгенетика VL 334 рУ № 6.
В качестве донорного щтамма используют штамм ВНИИгенетика MG-442. Штамм обладает устойчивостью к аналогу треонина (р-оксинорвалину) и несет мутацию в гене thr А, нарушающую аллостерпческос ингибирование треонином активности гомосериндегидрогеназы - ключевого фермента биосинтеза треонина.
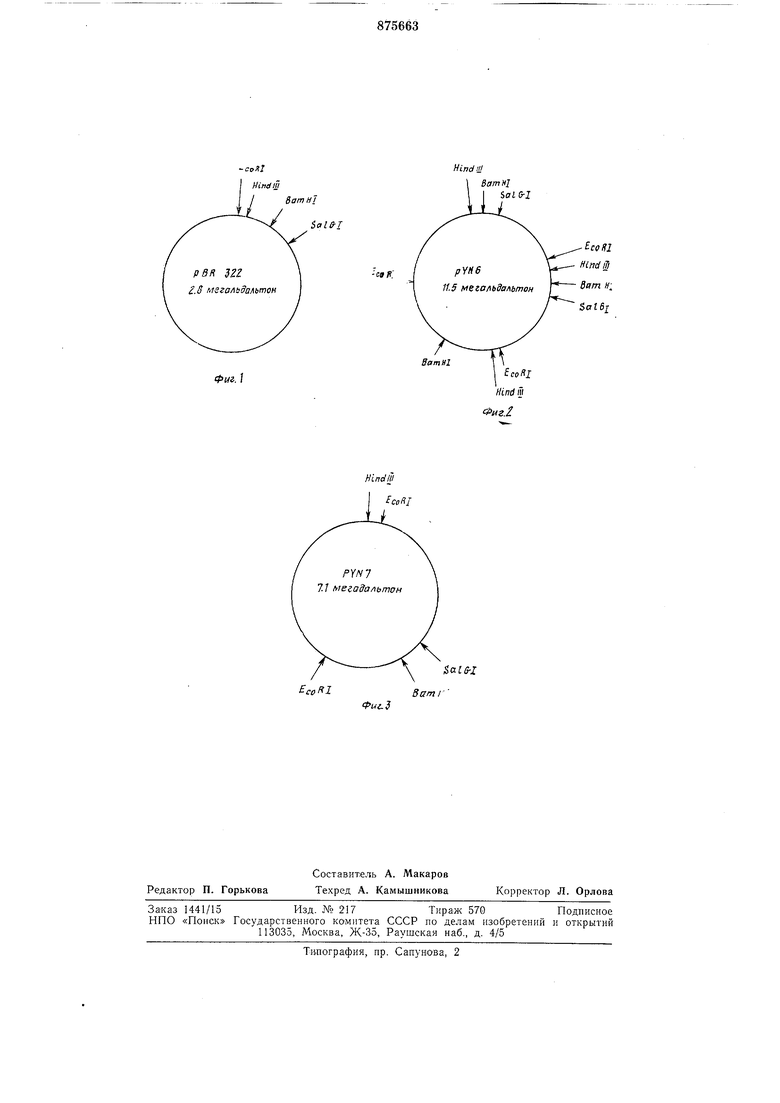
В качестве векторной плазмиды используют плазмиду pBR 322 3. Эта плазмида содержит репликон ColEl, имеет мол. вес. 2,8 мегадальтон, детерминирует устойчивость клеток к ампициллину и тетрациклину. Карта расщепления плазмиды pBR322 специфическими эндонуклеазами приведена на фиг. 1.
Хромосомную ДНК Е. соИ ВНИИгенетика MG-442 и плазмидную ДНК выделяют,
как описано ранее 6. Для конструирования гибридных плазмид ДНК из донорного щтамма ВНИИгенетика MG-442 и плазмиды pBR 322 обрабатывают эндонуклеазой. Для этого составляют инкубационную пробу, объемом 100 мкл. которая содержит 60 мМ Na С1; 10 мМ трисНС рН 7,4; 7 мМ MgCb; 10 мМ 2-меркаптоэтанола; 2 мкг pBR 322; 2 мкг Е. соИ 442; 10 единиц эндонуклеазы Hind 1П. Обработку эндонуклеазной проводят в течение 1 ч ПРИ 37°С, затем Пробу прогревают 10 мин. при для инактивации эндонуклеазы. В пробу добавляют 10 мМ дитиотриэтола; 66 мкМ АТФ; 200 г/мл бычьего сывороточного альбумина; 0,01 единиц
полинуклеотидлигазы и инкубируют в течение 24 ч при 6°С.
Полинуклеотидлигазу выделяют по методу Ричардсона. Полученную смесь гибридных молекз л используют для трансформа.ции Е. соИ СбОО (leu thr). Трансформанты высевают на чашки с минимальной средой Адамса, содержащей ампициллин в концентрации 200 мкг/мл. После инкубации в течение 48 ч при 37°С с этих чашек отбирают клоны траисформантов, устойчивые к ампициллину и способные расти без треонина. Из одного произвольно выбранного клона выделяют плазмидную ДНК.
Выделенную гибридную плазмидную ДНК, в- дальнейшем именуемую рУ № 6, исследуют с помощью электронно-микроскопических и электрофоретических методов-. На основании этих результатов строится карта расщепления плазмиды рУ № 6 специфическими эндонуклеазами (см. фиг. 2).
Гибридная плазмида рУ № 6 имеет молекз лярный вес 11,2 мегадальтон и состоит из двух молекул плазмиды pBR 322 (мол. вес 5,6 мегадальтон и фрагмента хромосомы (мол. вес 5,6 мегадальтон) донорного мутантного штамма MG-442, содержащего все три функционирующих гена треонинового оперона,- у которого в результате мутации продукт гена thr А устойчив к ингибированию треонином, и также содерл ащего балластный генетический материал.
Две молекулы плазмяды nBR 322 в составе гибридной плазмиды рУ № 6 осуществляют автономную репликацию плазмиды рУ № 6 в клетке и обеспечивают устойчивость клеток к пенициллину и тетрациклину.
В логарифмической стадии роста клетки мотут содержать 10 копий гибридной плазмиды.
Карта расщепления гибридной плазмиды рУ № 6 специфическими эндонуклеазами приведена на фиг. 2.
Для того, чтобы определить, какие гены треонинового оперона содержатся и функционируют на плазмиде рУ № 6, эту плазмиду ИСПОЛЬЗУЮТ для трансформации в щтаммы Е. соИ с мутациями по различным генам треонинового оперона: GTI4 Cthr А) VL 361 (thr В) и VL334 (thr С). В двух последних щтаммах мутации thr В и thr С из щтаммов G Т25 и G Т28, соответственно, были объединены с мутацией ilv с помощью трандукции.
Трансформантьг высевают на минимальную среду, содержащую ампициллин. Все устойчивые к ампициллину трансформацты щтаммов GT14, VL361 и VL 334.способны расти на среде без треЪнина. Результаты позволяют заключить, что плазмида рУ№6
содержит, все три фуи1кционирующих гена треонинового оперона: thr А, thr В, thr С.
Для получения штамма-продуцента
треонина нлазмида рУ 6 используется
для трансформации в реципиентиый
шта.мм VL 334, несущий М)тации в генах
thr С и thr А.
Характеристика реципиентного штамма ВНИИгенетика VL 334.
Штамм ВНИИгенетика VL334 является производным шта.мма ВНИИгеиетика - MG-442, получен трансдукцией фагом Р1 в этот штамм мутации thr С из щтамма G Т28.
Морфология. Грамотрицательные слабо подвижные тожкие иа,лочки с закругленными концами, размером 1,5-2,0 мм в длину.
Культурально-физиологические призиаМясо-пептонцьгй агар. Через 24 ч роста при 37°С Обравует колонии диаметром 2- 3 мм, круглые, слегка выпуклые с гладкими краями, структура однородная, прозрачные Hai свет, консистенция пастообразная, легко эмульгируется.
Агар.изованная и минимальная среда (Адамса) с глюкозой (0,2%) и минер-альным азотом. Через двое суток ро,ста при
37°С образует колонии диаметром 1,5-
2.0 мм серовато-белые, круглые с ровными
краями, слегка выпуклые, внутренняя
структура ол-нородная.
Рост в мясо-пептонном бульоне. После
24 ч роста при 37°С сильное равномерное иом5 тнение, небольщой осадок, запах характерный.
Рост в жидкой минимальной среде Аламса с нео бходимыми добавками. Через
одни сутки роста при 37°С с аэрацией сильное равномерное помутнение., запах отсутствует.
Рост по уколу в мясо-пептонном агаре ХОРОШИЙ по всему Зколу.
Желатину не разжижает.
На молоке хорошда рост с коаг)ляцией мОЛОка..
Индол образует. Рост на различных углеводах: хорощо
растет на глюкозе, лактозе, маннозе, галактоэе, ксилозе, фруктозе, глицироле н маинитоле с образованнем кислоты и газа. Потребность в факторах роста. Нуждается для роста в треонине и изолейцине
(20 .мкг/мл). Потребность в треонине связана с мутацией thrClOlO, иа-рушающей синтез треонина:, а потребность в изолейцине - с мутацией ilv А 442, блокирующей цервую реакцию на пути преврашения
треонина; в изолейцин. Фермент треониндезаминаза, поврежденный мутацией ilv А 442, сохраняет остаточную активность (менее 1% от исходного уровня). Поэтому при повыщении внутриклеточной конщентр ацкк треониря происходит частичная
компенсация мутации ilv А 442, т. е. клетки приобретатот способность к росту в отсутствие изолейцина в среде.
Мутация thr С необходима для отбора трансформаторов, получивших гибридную плазмиду. Эти трансфор.маторы должны быть устойчивы к1 ампициллину и расти на среде без треонина. Мутация активирует выражение треонинового оперона, так как блокирует синтез изолейцина, участвующего в репрессии треонинового ошероHai. Кроме того, она частично блокирует превращений треонина в п редшественник изолейцина-а-кетомас-дяную кислоту. Рецилиект является ауксотрофом по треоницу W изолейцину, однако расчет иа соеле без изолейцина при высокой концентрации треонина.
Трансформанты, получившие плазмиду пУ № 6, отбирают на среде, содержащей 200 MKir/мл: ампи,цилдина, но без твеонина. Клетки одного произвольно -выбранного клоиа трансфо|рмантов вьшащи;вают на бульоие Хоттингера с ампициллином и из них выделяют илазмидную ДНК и сравнивают ее физико-химические параметры (мол. вес, ра.спределение участков расшепления специфическими эндонуклеазам.и) с параметрами плазмиды рУ № 6, установленными ранее (см. фиг. 2). Совпадение указанных свойств свидетельствует о том, что клетки выбранного клона содержат гглазмилу рУ № 6, и плазмида стабильно существует в них в указанных условиях культирлрозания. Полученный штамм в ля.лънейшел1 именуют ВНИИген.етика VL334py № 6.
Полученный штамм УСТОЙЧИВ к ампициллину и тетрациклину и способен расти на среде без треонина и изолейцина. СпосОбность расти без изолейцина сВЯза1на с высоким уровнем синтеза треонина в этих клетках. Генотип реципиентного штамма (мутация ily А) обусловливает, тем самым, селективное преимущество клеткам, содержащим плазадиду рУ № 6.
Пример- 2. Конструирование штамма Е. соИ ВНИИгенетика-VL 334 рУ № 7.
Фрагмент ДНК Е. соИ в соста ве плазм.иды РУ № 6 больше размера треонинового оперона. Удавление балластного генетического материала может увеличить жиэнеспособность ба.ктерий и оказать ноложительное действие на стабильность плазмиды. В связи с этим плазмиду рУ № 6 обрабатывают специфическими эндонуклеазами Hind П1 и Ват Hi. Инкубационная Проба, объемом 100 мкл, имеет тот же состав, ЧТО и В примере 1, но кроме того, содержит б единиц эндонуклеазы Ват Hi. После ина ктивации ферментов прогреванием при 65°С в течение 10 мин смесь обрабатывают полинуклеотидлигазой (оример 1),
Полученная смесь гибридных молекул используется для трансформации штамма Е. соИ СбОО. Отбор клонов и выделение плазмидной ДНК производят как в примере I. Выделенная гибридная плазмида, в лальне 1шеМ именуемая рУ № 7, имеет МОЛ. 55ес ,4 мег-адальтон. Карта расщепления этой плазмиды специфическими эндонуклеазами приведена- на рис. 3. Трансформация рУ Л9 7 в мутантные щтаммы Е. coll показывает, что эта плазм-ида имеет все три гена треониноваго оперона: thr А, thr В и thr С.
Хара чтеристика плазмиды рУ N° 7. 15 Гибридная плазм.ила рУ ЛЬ 7 имеет МОлекуляпный вес 5,7 мегадальтон и состоит из пл:а.змиды рВ R 322 (молекулярный вес 2,8 метадальтон) и фрагмента хромосомы (молекулярный вес 2,9 мегадальтон) му0 тантного донорного штамма MG 442, содержащего все три функционирующих гена треонинового оперона, у которого в результате мутащии продукт гена thr А устойчив к иигибирова1НИЮ треонино-м. 5
Плазмида пВ R 322 в составе гибридной плазмилы рУ .NO 7 представлена фрагментом ДНК, способным осуществлять автономную репликацию гибридной плазмиды 0 и обеспечивать устойчивость клеток к пенициллину.
В логарифмической стадии роста клетки могут содержать 17 копий гибридной плазмиды.
5 Карта ра.сщепления гибридной плазмиды рУ № 7 специфическими эндонуклеазами приведена на фиг. 3.
После трансформации рУ № 7 в реципиентный штамм VL 334 (пример 1) отбирают клоН, в л а-льнейшем именуемый ВНИИгенетика VL 334 рУ 7, устойчивый к а.Мпициллину и способный расти на среде без изолейцина и треонина.
Пример 3. Получение L-треонина с но5 мощью Е. СОИ, несущих гибридные плазмиды.
Штаммы, полученные по способу, описанному в примерах 1,2 и перечисленные в таблице, засевают петлей с косяка ага0 ризованной среды Адамса, содержащей 0,5 мг/мл калиевой соли бензилпенициллина, в конические колбы, емкОСтью 250 мл, содержащие по 30 мл лодкой среды Адамса (глюкоза 1 %, тиамин 100 мкг/л). 5 После посева колбы устанавливают на круговую качалку (200 об/мин) и инкубируют в течение 18 ч цри 37°С. Выращенный та.ким способом посевной материал используют в количестве 1 мл д.ля засева 0 предварительно простерилизованной ниже описанным методом ферментационной среды, разлитой ио 15 мл в конические колбы, емкостью 250 мл.
Ферментационные среды имеют следующи и состав, г/Л: Тиамин 0,0001 0,0001 0,0001 Фепментационные спеды стерилизуют в ячтоклаве .прИ избыточном давлении 0,5 а.мт в течение 15 мин. Мел стерилизуют отдельно- и вносят в среду после степилизации. После добавления мела среда имеет пН 6,8-7,2. Колбы, содепжащие указанкне соеды, после посева кх-лътупами указанных в та блкпе штаммов устанавливают wa. КПУГОВУЮ качалку С9ПП обмин) и инкубипуют 48 ч ппи 37°С. 95% клеток иосле сЬепментации сохраняют способность оастч без треонина и изолейцина и устойчивы к ампициллину, что свидетельствует о наличии плазмид в клетках данных штаммов. Количество треонина, образованного штяммами. показанов таблице. ПрИ получении L-тпеонина в фепментер е посевной материал штаммаУЬ334 пУ № 6, по.лученный, как Описано в примере 3, В ко.пичестве 25 мл вносят е лабора торный ферментер марки Biflo С230, куда предва.рИтельно помеща.ют 250 мл ферментационной среды. 1. Режим ферментации следующий: температура 37°С, количество поступающего в аПпарат воздуха 1.1 (по ротаметру), скорость мешалки 900 Об/мик. Через- 28 ч. после начала ферментации приступают к полаче подпитки, представляющей собой десятикратный концентрат ферментационной среды 1, из состава которой исключен мея. Подпитку производят с помощью перистальтического насоса со скоростью 1.5 мл/ч. Через 51 ч после начала ферментации в среде на-каплива.ется треонин в количестве 20 г/л 95% клеток после ферментации содержат плавмиду рУ № 6. L-треонин выделяют из кулътуральной жидкости известными методами. Как видно из данных таблицы наиболее высокий уровень образования аминокислоты имеет место у штаммов, содержащих т-ибпитную плазмиду. Использование этого Me orrai позволяет по.ЯУЧИть штаммы Е. coli ВНИИ-енетикя VL 334 ру № 6 и ВНИИгенетика VL 334 ру № 7, способные накапливать до 20 г/л треонина пои культивировании на минеральной среде с глюкозой, тиаминам. Полученные штаммы обладают высокими технологическими показателями - они превосходят по продуктивности все известные в мире штаммы и не требуют сложных или ДОРОГИХ питательных сред. Формула изобретения - I. Штаммы Е. coli ВНИИгенетика VL 334 рУ № 6 и ВНИИгенетика VL 334р У № 7 продуценты L-треонина хранятся в центральном музее ПРОМЫТО денных микроорганизмов института «ВНИИгенетика и имеют пегистрациоиные номера В-1649 и В-1в84. МОРФОЛОГИЯ. Грамотрицательные слабополвижные тонкие лалочки с закругленными концами, размером 1,5-2,0 мм в длину. Культурально-физиологические призна.ки. Мясо-пептонный a raip. Через 24 ч роста при 37°С образует колонии диаметром 2- 3 мм, круглые, слегка выпуклые с гладкимн краями, структура однородная, прозрачные на свет, ковсистен1ция пастообразная, легко эмульгируются. Агапизованная и минимальная среда; (Адамса) с глюкозой (0,2%) и минеральным азотом. Через двое суток роста при 37°С образует колонии диаметром 1,5- 2 мм, серовато-белые, круглые с ровными кра.ями, слегка выпуклые, внутренняя структура однородная. Рост в мясо-пептонном бульоне - после 24 ч роста при 37°С сильное равномерное помутнение, небольшой осадок, запах характерный. Рост в жидкой минимальной среде Адамса с необходимыми добавками - через одни сутки роста при 37°С -с аэрацией сильное равномерное помутнение, запах отсутствует. Рост по уколу в мясо-пептонном агаре- хороший по всему уколу. Желатину не разжижает. На молоке хороший рост с коагуляцией молока;. Индол образует. Рост на различных углеводах: хорошо растет па глюкозе, лактазе, маннозе, галактозе, ксилозе, фруктозе, глицероле и маннитоле с образованием кислоты и газа. Потребность в факторах роста. Частичпая потребность в изолейцине. Время генерации на глюкозо-минеральной среде Адамса с изолейцином 60 мин, на среде без изслейцина 270 мин. Устойчивость к антибиотикам. Устойчив к пенициллину и тетрациклину. Штамм не патогенен. Содержание нлазмиды. Для штамма Е. соИ ВНИИгенетика VL 334 фУ №.б в логарифмической стадии роста клетки содержат около 10 копий плазмиды рУ № 6 (мол. вес. 11,2 мегадальтон), обеспечивающей устойчивость штамма к пенициллину и тетрациклину и несущей гены треонииового оперона. Для щтамма Е. соИ ВНИИгенетика VL 334 рУ № 7 в логарифмической стадии роста клетки содержат около 17 копий плавмиды рУ № 7 (мол. вес 5,7 мегадальтон), обеспечивающей устойчивость штамма к пенициллину и несущей гены треонинового оперона. 2.Способ получения штаммов Е. соИ продуцентов L-треонина по п. 1, включающий объединение фрагмента ДНК хромосомы донорного микроорганизма с векторной молекулой ДНК, предпочтительно с плазмидой, и трансформацию полученной гибридной молекулой ДНК реципиентного микроорганизма, отличающийся тем, что, с целью увеличения выхода L-треонина, в качестве фрагмента ДНК используют ДНК хромосомы донориого микроорганизма, содержащие гены, контролирующие синтез L-треонина и имеющие мутацию, нарушающую негативную регуляцию синтеза данной аминокислоты, а в качест1ве реципиентного щтамма используют штамм, имеюший мутацию, частично блокирующую смежный этап метаболизма указанной аминокислоты. 3.Способ па п. 2, отличающийся тем, что объединение фрагмента ДНКхромосомьБ донорного щтамма Е. соИ осуществляют с плазмидой pBR 322, имеющей молекулярный вес 2,8 мегадальтон. 4.Способ ПО п. 2, отличающийся тем, что используют фрагмент ДНК хромосомы донорного щтамма Е. соИ, содержащий гены треовинового оперона, у которого в результате мутации фермент8755 10 15 20 25 2Q 35 40 45 3 продукт гена thr А, устойчив к ингибированию треонином. 5.Способ по пш 1-4, отличающийся тем, что в качестве реципиентного щтамма используют щтамм Е. соИ ВНИИгенетика VL 334, имеющий двойную ауксотрофность по треонину и изолейцину. 6.Способ по П1П. 1-5, отличающийс я тем, что трансформацию реципиентного штамма Е. соИ ВНИИгенетика VL 334 проводят гибридной плазмидой рУ № 6, имеющей молекулярный вес 11,2 мегадальтон, состоящей из двух молекул плазмиды pBR 322 с молекулярным весом 5,6 мегаДальтон и содержащей функционирующие гены треониновОГо Оперона фрагмента ДНК хромосомы донорного щтамма Е. соИ. 7.Способ по пп. 1-5, отличающийс я тем, что трансформацию реципиентного штамма Е. со-И ВНИИгенетика VL 334 проводят гибридной плазмидой рУ № 7, имеющей молекулярный вес 5,7 мегадальтон, состоящей из плазмиды p-BR 322 с молекулярным весом 2,8 мегадальтон и содержащий функционирующие гены треонинового Оперона фрагмента ДНК хромосомы донорного штамма Е. соИ. Источники информации, принятые во внимание лри экспертизе 1.Патент США 3002889, кл. 195-28.30, 1969. 2.Патент Великобритании 1258380, кл. С 6F, 1969. 3.Патент Франции 1579835, С 12D, 3/06, 1972. 4.Патент США 3756916, кл. 195-78.96, 1974. 5. Патент США 3923603, кл. 195-103,5, 1975. 6. Гусятинер М. М., Жданова Н. И. и Лившиц В. А. Исследование фракции гена ге1 А в выражении аминокислотных оперонов. Сообщение П. Влияние аминного состояния гена ге1 А на сверхсинтез треонина мутантов Е. coli К-12, устойчивых к р-оксинорвалину. ж. Генетика, 14, № 6, 1978, с. 957.
Фиг. /
Hind a/
Bam HI Sal 6-1
Sam HI






| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения L-треонина | 1979 |
|
SU943282A1 |
| ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ АМИНОКИСЛОТЫ (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ АМИНОКИСЛОТ (ВАРИАНТЫ) | 2000 |
|
RU2212447C2 |
| ФРАГМЕНТ ДНК rhtB, КОДИРУЮЩИЙ СИНТЕЗ БЕЛКА RhtB, ПРИДАЮЩЕГО УСТОЙЧИВОСТЬ К L-ГОМОСЕРИНУ БАКТЕРИЯМ ESCHERICHIA COLI, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 1998 |
|
RU2144564C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ, ШТАММ Escherichia coli - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2001 |
|
RU2215782C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-треонина С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2006 |
|
RU2351646C2 |
| МИКРООРГАНИЗМ С ВЫКЛЮЧЕННЫМ ГЕНОМ fadR НА ХРОМОСОМЕ И СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА ПОСРЕДСТВОМ ФЕРМЕНТАЦИИ С ДАННЫМ МУТАНТОМ | 2003 |
|
RU2305132C2 |
| Способ получения L-треонина | 1980 |
|
SU904325A1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ, ШТАММ Escherichia coli - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2001 |
|
RU2215784C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ ESCHERICHIA COLI-ПРОДУЦЕНТ L-ТРЕОНИНА | 2013 |
|
RU2546237C1 |
| Штамм Escherichia coli - продуцент L-треонина | 2023 |
|
RU2817252C1 |