ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Система комплемента является компонентом врожденной и приобретенной иммунной системы (обзор приведен в Volanakis, J.E., 1998. Chapter 2. In The Human Complement System in Health in Health and Disease. Edited by J. E. Volanakis, and M.M. Frank. Marcel Dekker, Inc., New York pp 9-32). Комплемент играет важную роль в уничтожении микробов и в транспорте и выведении иммунных комплексов. Множество продуктов активации системы коплемента ассоциированы также с провоспалительными или иммунорегуляторными функциями. Система коплемента состоит из ассоциированных с плазмой и мембраной белков, организованных в три каскада активации: классический, лектиновый и альтернативный пути (фигура 1). Все три пути могут приводить к формированию терминального комплекса комплемента (TCC) и ряда биологически активных продуктов.
В некоторых случаях, активация комплемента инициируется или посредством распознавания и связывания специфическими антителами множества патогенов и чужеродных молекул, и/или посредством прямого взаимодействия белков комплемента с чужеродными веществами. При активации, эти пути приводят к формированию комплексов протеаз, C3-конвертаз. C3-конвертаза классического пути, C4b2a, и C3-конвертаза альтернативного пути, C3bBb, обе являются способными расщеплять цепь C3, образуя C3b. C3b обладает способностью ковалентно связываться с биологическими поверхностями. Связывание C3b приводит к опсонизации для фагоцитоза полиморфноядерными клетками и макрофагами. Когда доступен дополнительный C3b, C3-конвертазы могут функционировать как C5-конвертазы, расщепляя C5 и инициируя сборку TCC, или мембраноатакующего комплекса (MAC), опосредующего клеточный лизис посредством вставки порообразующих белковых комплексов в мембраны клеток-мишеней.
В классическом пути, как показано на фигуре 1A, C1q, коллагеновый субкомпонент первого компонента (C1), связывается с иммуноглобулинами внутри иммунных комплексов, и ассоциированные с ним сериновые протеазы, C1r и C1s, становятся активированными. Этот каскад реакций комплемента инициируется последующим расщеплением C4 и C2, с последующей активацией C3. Полученный фрагмент C3b не только действует как опсонин, но также приводит к формированию мембраноатакующего комплекса (MAC) в литическом пути. Во врожденном иммунитете, комплекс, состоящий из узнающей молекулы (лектина) и сериновых протеаз, названный сериновой протеазой, ассоциированной со связывающим маннозу лектином (MBL) (MASP), активирует C4 и C2 при связывании с углеводами на поверхности микроорганизмов через лектиновый путь. Это связывание происходит в отсутствие иммуноглобулинов. Узнающие молекулы из лектинового пути, обнаруженные у челюстноротых позвоночных, представляют собой MBL и фиколины, которые оба характеризуются присутствием коллагеноподобного домена, подобно C1q, и связывающего углевод домена, обладающего общей специфичностью связывания для GlcNAc. MASP и C1r/C1s разделяют одинаковую доменную организацию и формируют подсемейство сериновых протеаз.
Лектиновый путь комплемента во врожденном иммунитете является близко родственным классическому пути активации комплемента в приобретенном иммунитете, например, по отношению к структурам и функциям их компонентов. Оба пути, как правило, инициируют комплексы, состоящие из коллагеновых белков и сериновых протеаз семейства ассоциированной со связывающим маннозу лектином (MBL) сериновой протеазы (MASP)/C1r/C1s. Выдвинуто предположение, что классический путь возник в ходе эволюции после лектинового пути.
Активация альтернативного пути комплемента, показанного на фигуре 1B, как правило, начинается со связывания белка C3b (или C3i) с клеткой и другими компонентами поверхности, например, микроорганизмов. C3b может также связываться с антителами иммуноглобулином G (IgG). Белок фактор B альтернативного пути затем объединяется с белком C3b с формированием C3bB. Затем белок фактор D расщепляет связанный белок фактор B на фрагменты Bb и Ba, образуя C3bBb. Затем пропердин связывается с Bb с образованием C3bBbP, который функционирует как C3 конвертаза, способная к ферментативному расщеплению, как правило, сотен молекул C3 на C3a и C3b. Некоторые из C3b затем связываются с некоторыми из C3bBb с образованием C3bBbC3b, C5 конвертазы, способной расщеплять молекулы C5 на C5a и C5b.
Поскольку C3b является свободным в плазме, он может связываться с поверхностью либо клетки-хозяина, либо патогена. Для предотвращения прохождения активации комплемента в клетке-хозяине, существует несколько различных видов регуляторных белков, нарушающих процесс активации комплемента. Рецептор комплемента 1 (CR1 или CD35) и DAF (известный также как CD55) конкурируют с фактором B за связывание с C3b на поверхности клеток и могут даже удалять Bb из уже сформированного комплекса C3bBb. Образование C3 конвертазы можно также предотвращать, когда протеаза плазмы, называемая фактором I, расщепляет C3b до его неактивной формы, iC3b. Фактор I действует со связывающими C3b белок кофакторами, такими как CR1 и мембранный кофактор протеолиза (MCP или CD46). Хотя регуляторным белком комплемента является фактор H, который либо конкурирует с фактором B, вытесняет Bb из конвертазы, действует как кофактор для фактора I, либо предпочтительно связывается с C3b, связанным с клетками позвоночных.
Точная функция системы комплемента зависит от ее регуляции, поскольку активация каскада реакций комплемента приводит к продукции ряда белков, вносящих вклад в воспаление. Это является преимущественным, когда вносит вклад в защиту хозяина, но может являться вредным, если активируется на собственной ткани. Как правило, активация C3 в крови поддерживается на низком уровне, и накопление C3b ограничено поверхностью патогенов.
Белок фактор комплемента B дикого типа человека представляет собой одноцепочечный гликопротеин из 764 аминокислот (приблизительно 93 кДа), состоящий из пяти доменов белка (Mole et al, 1984 The J. Biol Chem, 259:6, 3407-3412). Белок фактор комплемента B (fB) дикого типа человека, как правило, экспрессируется с N-концевым сигнальным пептидом из 25 аминокислот, например, см. SEQ ID NO:1. Аминоконцевая область (Ba) фактора комплемента B дикого типа человека состоит в первую очередь из трех коротких консенсусных повторов. Средняя область представляет собой домен типа A, сходный с доменами, обнаруженными в факторе фон Виллебранда (Colombatti et al., Blood (1991) 77(11):2305-15). Карбоксиконец представляет собой домен сериновой протеазы (SP) (Perkins and Smith, Biochem J. (1993) 295 (Pt 1): 109-14; Hourcade et al, JBC (1998) 273(40):25996-6000; Hourcade et al. J Immunol. (1999) 162(5):2906-11; Xu et al, J Biol Chem. 2000 275(1):378-85; Milder et al Nat Struct Mol Biol (2007) 14(3):224-8).
Аналоги фактора комплемента B и их использование для ингибирования комплемента и лечения опосредованных комплементом заболеваний описаны в Публикации PCT No. WO08/106644 и Публикации патента США No. US20100120665. Например, аналог белка фактора комплемента B, hfB3 (описанный в Публикации патента США No. 20100120665), представляет собой доминантный негативный вариант белка фактора B человека, который эффективно ингибирует альтернативную активность комплемента (AP). Белок hfB3 (SEQ ID NO:4) имеет пять замен аминокислот по сравнению с белком фактором B дикого типа человека (SEQ ID NO:1). Пять замен аминокислот позволяет белку hfB3 (i) более крепко связывать белок C3b, (ii) являться устойчивым к C3b-зависимому расщеплению белком фактором D, и (iii) более крепко связывать белок фактор D по сравнению с белком фактором B дикого типа. Более крепкое связывание белка hfB3 с белком C3b и белком фактором D разделяет два необходимых компонента альтернативного пути комплемента (ACP) на неактивную C3 конвертазу (hfB3), блокируя активность AP. Поскольку связанный с C3b белок hfB3 не может расщепляться белком фактором D, конформационного изменения белка hfB3 не происходит, и сериновая протеаза на C-конце белка hfB3 не активируется.
Как белок фактор комплемента B дикого типа человека, так и hfB3 содержат 23 аминокислоты цистеина. «Активные» формы обоих имеют все цистеины со сформированными дисульфидными связями с одним из других цистеинов, за исключением цистеина, соответствующего C292 SEQ ID NO:1. C292 «активных форм» hfB3 и фактора B дикого типа представляет собой свободный цистеин (Parkes et al. 1983 Biochem J. 213, 201-209) и является высоко консервативным среди различных видов млекопитающих, например, см. таблицу 1, ниже.
Цитирование или обсуждение ссылок в настоящем документе не следует рассматривать как допущение, что они являются предпосылками настоящего изобретения.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Изобретение относится к полипептидам, содержащим аналог фактора комплемента B. Изобретение относится также к различным аналогам фактора комплемента B. В некоторых вариантах осуществления, аналог фактора комплемента B содержит мутацию свободной аминокислоты цистеина. Изобретение относится также к нуклеиновым кислотам и вирусным векторам, содержащим нуклеотидную последовательность, кодирующую полипептиды и аналоги белка фактора комплемента B по изобретению. Некоторые варианты осуществления изобретения относятся к клеткам, где клетки содержат нуклеиновую кислоту, кодирующую аналог белка фактора комплемента B по изобретению, и где клетки экспрессируют аналог белка фактора комплемента B.
Кроме того, изобретение относится к фармацевтическим препаратам, содержащим полипептид или аналог белка фактора комплемента B по изобретению, нуклеиновую кислоту по изобретению, вирусный вектор по изобретению или любую их комбинацию.
Настоящее изобретение относится также к способам лечения опосредованного комплементом заболевания, включающим в себя введение пациенту фармацевтического препарата по изобретению, полипептида по изобретению, аналога белка фактора комплемента B по изобретению, нуклеиновой кислоты по изобретению, вирусного вектора по изобретению или любой их комбинации.
Изобретение относится также к способам получения полипептида, содержащего аналог белка фактора комплемента B, где способ включает в себя: экспрессию в клетке аналога белка фактора комплемента B по изобретению и очистку указанного аналога белка фактора комплемента B.
Полипептиды по изобретению, аналоги фактора комплемента B по изобретению, и нуклеиновые кислоты и векторы, кодирующие их, можно использовать для модуляции пути комплемента и для исследования и/или лечения различных состояний или заболеваний, связанных с путем комплемента.
Изобретение основано на обнаружениях, что мутация или удаление свободного цистеина (i) улучшает выход активного и/или правильно свернутого аналога белка фактора комплемента B (например, см. примеры 9 и 10); (ii) улучшает термостабильность аналога белка фактора комплемента B (например, см. примеры 13 и 14); и/или (iii) снижает агрегацию аналога белка фактора комплемента B (например, см. пример 6).
Это краткое изложение сущности изобретения не обязательно описывает все признаки или необходимые признаки изобретения. Изобретение может также относиться к субкомбинации описанных признаков.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
С целью иллюстрации изобретения, на фигурах изображены варианты осуществления изобретения. Однако, изобретение не является ограниченным точными организацией и средствами вариантов осуществления, изображенными на фигурах.
На фигуре 1A изображены классический и лектиновый пути комплемента. Классический путь инициируется посредством C1, в то время как лектиновый путь инициируется посредством связывающего маннозу лектина (MBL). C4bC2a представляет собой протеазу, расщепляющую C3 на C3a и C3b, и называемую C3 конвертазой. Подобным образом, C4bC2aC3b расщепляет C5 на C5a и C5b, и его называют C5 конвертазой. C3a, C4a и C5a обладают воспалительными свойствами и привлекают фагоцитирующие клетки. C5b6-9 формирует мембраноатакующий комплекс (MAC), который образует поры в мембране, которые убивают инфекционные агенты, но могут также повреждать клетки-хозяева. MASP представляет собой ассоциированную со связывающим маннан лектином сериновую протеазу.
На фигуре 1B изображен альтернативный путь комплемента. Этот путь является конститутивно активным на низком уровне посредством спонтанного расщепления C3. В присутствии подходящей поверхности, C3b связывается с фактором комплемента B (fB). Этот комплекс затем расщепляется фактором комплемента D (fD) с образованием C3bBb. Спонтанная диссоциация («разложение») этого комплекса в течение минут приводит к его инактивации, в то время как стабилизация пропердином образует комплекс, расщепляющий C3; то есть, C3 конвертазу. Несколько факторов, ослабляющих пути комплемента, осуществляют это посредством усиления разложения C3 и C5 конвертаз. C3b участвует в C3 конвертазе для получения дополнительного C3b, таким образом создавая петлю положительной обратной связи, как показано большой стрелкой. C3bBb представляет собой C3 конвертазу. C3bBbC3b представляет собой C5 конвертазу.
Фигура 2 представляет собой экспрессирующую конструкцию hfB3-292S. CMV - немедленный ранний промотор цитомегаловируса; IRES - внутренний участок связывания рибосомы; Neo - ген неомицин-фосфотрансферазы; SynPolyA - синтетический полиA; Amp - ген устойчивости к ампициллину. SEQ ID NO:8 представляет собой нуклеотидную последовательность экспрессирующей конструкции hfB3-292S, показанной на фигуре 2.
На фигуре 3 показан анализ Вестерн-блоттингом неочищенных супернатантов культур клеток, содержащих белок либо hfB3, либо hfB3-292S после инкубации культуры клеток в течение 72 часов (2×106 клеток/мл). Дорожка 1, один мкл среды культуры клеток от наивных нетрансфицированных клеток 293 FreeStyle, служащей отрицательным контролем; Дорожка 2, сто нанограмм фактора B дикого типа человека (Quidel, Santa Clara, CA), очищенного из плазмы, служащего положительным контролем; Дорожка 3, один мкл среды культуры клеток из продуцирующих hfB3 клеток; Дорожка 4, один мкл среды культуры клеток из продуцирующих hfB3-292S клеток. Маркеры молекулярной массы в кДа указаны справа.
На фигуре 4 показаны результаты гемолитического анализа. Эти результаты показывают ингибирование гемолитической активности альтернативного пути комплемента человека неочищенной культуральной средой продуцирующих белок hfB3 или белок hfB3-292S клеток. Относительную гемолитическую активность оценивают по гемоглобину, высвобожденному после гемолиза rRBC посредством активности альтернативного пути комплемента человека. По X-оси слева направо: истощенная по фактору B сыворотка человека, дополненная 0 мкг очищенного белка фактора B человека (контроль, без лизиса rRBC); истощенная по фактору B сыворотка человека, дополненная в каждой реакции смесью 0,5 мкг очищенного белка фактора B человека и 1,0, 0,5, 0,25 или 0,125 мкг белка hfB3 или белка hfB3-292S, как указано, из культуральной среды продуцирующих белок hfB3 или белок hfB3-292S клеток. Примечание: 100% ингибирование представляет отсутствие лизиса rRBC. Y-ось представляет среднее OD405 и стандартное отклонение (SD).
На фигуре 5A показан белок hfB3 (200 нг), очищенный на трех стадиях процесса хроматографии, подвергнутый анализу SDS-PAGE и окрашиванию серебром. На фигуре 5B показано ингибирование гемолитической активности альтернативного пути комплемента человека очищенным белком hfB3. По X-оси слева направо: истощенная по фактору B сыворотка человека, дополненная 0 мкг очищенного белка фактора B дикого типа человека (wt hfB) (контроль, без лизиса rRBC); истощенная по фактору B сыворотка человека, дополненная в каждой реакции смесью 0,5 мкг очищенного белка фактора B человека и 1,0, 0,5, 0,3, 0,2, 0,1 или 0,05 мкг hfB3, как указано. 100% ингибирование представляет отсутствие лизиса rRBC. Y-ось представляет среднее OD405 и SD.
На фигуре 6 показана биологическая активность двух популяций белка hfB3. Показаны результаты хроматографии гидрофобного взаимодействия (HIC) очищенного белка hfB3 из пика I и пика II для ингибирования гемолитической активности альтернативного пути комплемента человека. По X-оси слева направо: истощенная по фактору B сыворотка человека, дополненная 0 мкг очищенного белка фактора B человека (контроль, без лизиса rRBC); истощенная по фактору B сыворотка человека, дополненная в каждой реакции смесью 0,5 мкг очищенного белка фактора B человека и различными количествами белка hfB3 в диапазоне 1,0-0,05 мкг. 100% ингибирование представляет отсутствие лизиса rRBC. Y-ось представляет среднее OD405 и SD.
На фигуре 7 показана обращеннофазовая высокоэффективная жидкостная хроматография (HPLC) неочищенных супернатантов культур клеток, содержащих либо белок hfB3, либо белок hfB3-292S после инкубации культуры клеток в течение 72 часов (2×106 клеток/мл). A) Супернатанты из продуцирующих hfB3 клеток (………), продуцирующих белок hfB3-292S клеток (______) и наивных клеток 293 (- - - - -), вносили в систему HPLC Agilent HP1 100 с использованием колонки с микронасадкой Jupiter™ C4 (Phenomenex) и элюировали 50-минутным градиентом вода/ацетонитрил (25-70% ацетонитрил), содержащим 0,1% TFA. Элюцию мониторировали при 215 нм с помощью детектора PDA. Показано также положение белка теплового шока 70 (HSP70), присутствующего во всех трех образцах.
B) Увеличенная область хроматограммы, показанной на фигуре 7A, сфокусированной на области (25-29 минут), содержащей пики I и II белка hfB3 и пик, содержащий белок hfB3-292S.
На фигуре 8 показан анализ экспрессии белка hfB3-Fc- посредством подвергания 2 мкл супернатанта культуры клеток, содержащего белок hfB3-Fc-, анализу невосстанавливающим SDS-PAGE и Вестерн-блоттингом. (См. пример 12). Детектировали две полосы белка hfB3-Fc-, маркеры молекулярной массы в кДа указаны слева.
На фигуре 9 показано репрезентативное окрашивание H&E парафиновых срезов суставов правых передних лап мышей из исследования, тестирующего hfB3-292S в модели ревматоидного артрита на мышах - индуцированном антителом против коллагена артрите (CAIA), как описано в примере 16. Группа 1 представляет собой контрольную группу носителя, в которой не вводили коктейля антител против коллагена. Группа 2 представляет собой группу без лечения, в которой вводили коктейль антител против коллагена. Группа 3 представляет собой группу после лечения, в которой вводили коктейль антител против коллагена и которую лечили hfB3-292S.
На фигуре 10 показаны измерения опухания сустава из исследования, тестирующего hfB3-292S в модели ревматоидного артрита на мышах - индуцированном антителом против коллагена артрите (CAIA), как описано в примере 16. Группы являются такими же, как группы на фигуре 8, как описано в разделе. hfB3-292S вызывал статистически значимое (p<0,0003) уменьшение опухания сустава по сравнению с группой без лечения (группа 2).
На фигуре 11 показан анализ Вестерн-блоттингом культуральной среды от клеток, трансфицированных экспрессирующей конструкцией hfB3-292SN480. Этот анализ Вестерн-блоттингом проводили с использованием моноклонального антитела, специфического для hfB3-292S. Левая дорожка содержит hfB3-292S, а правая дорожка представляет собой культуральную среду от клеток, трансфицированных экспрессирующей конструкцией hfB3-292SN480. Анализ детектировал полосу приблизительно 55 кДа из культуральной среды линии клеток hfB3-292SN480 (правая дорожка).
На фигуре 12 показан анализ Вестерн-блоттингом культуральной среды от клеток, трансфицированных экспрессирующей конструкцией hfB3-292S/Fc-mono, как описано в примере 19, ниже. Полосу приблизительно 115 кДа детектировали посредством очищенного антитела козы против фактора B человека после невосстанавливающего SDS-PAGE.
На фигуре 13 показано, что супернатант культуры клеток из клеток, экспрессирующих hfB3-292SN480, ингибировал гемолитическую активность альтернативного пути комплемента человека зависимым от дозы образом.
На фигуре 14 показано, что супернатант культуры клеток из клеток, экспрессирующих hfB3-292S/Fc-mono, ингибировал гемолитическую активность альтернативного пути комплемента человека зависимым от дозы образом.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO:1 - аминокислотная последовательность фактора комплемента B дикого типа человека.
SEQ ID NO:2 - аминокислотная последовательность аналога белка фактора комплемента B человека, hfB3-292S, содержащего следующие мутации: K258A, R259A, K260A, D279G, N285D и C292S.
SEQ ID NO:3 - аминокислотная последовательность аналога фактора комплемента B человека, hfB3-292S-740N, содержащего следующие мутации по сравнению с SEQ ID NO:1: K258A, R259A, K260A, D279G, N285D, D740N и C292S.
SEQ ID NO:4 - аминокислотная последовательность аналога фактора комплемента B человека, hfB3, содержащего следующие мутации по сравнению с SEQ ID NO:1: K258A, R259A, K260A, D279G и N285D.
SEQ ID NO:5 - нуклеотидная последовательность экспрессирующей конструкции hfB3,
конструирование которой описано в примере 1.
SEQ ID NO:6-7 - праймеры для сайт-специфического мутагенеза.
SEQ ID NO:8 - нуклеотидная последовательность экспрессирующей конструкции hfB3-292S, конструирование которой описано в примере 2.
SEQ ID NO:9-14 - частичные аминокислотные последовательности белков факторов комплемента B человека, мыши, крысы, свиньи, обезьяны и овцы, соответственно.
SEQ ID NO:15-16 - праймеры для сайт-специфического мутагенеза
SEQ ID NO:17 - аминокислотная последовательность аналога белка фактора комплемента B человека, hfB4.
SEQ ID NO:18 - нуклеотидная последовательность экспрессирующей конструкции hfB3-Fc-, конструирование которой описано в примере 12.
SEQ ID NO:19-20 - праймеры для сайт-специфического мутагенеза.
SEQ ID NO:21 - аминокислотная последовательность аналога белка фактора комплемента B человека, hfB3-Fc.
SEQ ID NO:22 - аминокислотная последовательность аналога белка фактора комплемента B человека, hfB3-292S-Fc.
SEQ ID NO:23 - аминокислотная последовательность аналога белка фактора комплемента B человека, hfB3-292S-740N-Fc.
SEQ ID NO:24 - нуклеотидная последовательность экспрессирующей конструкции для экспрессии hfB3-292SN480.
SEQ ID NO:25 - нуклеотидная последовательность конструкции для экспрессии гена для hfB3-292S/Fc-mono.
SEQ ID NO:26 - аминокислотная последовательность hfB3-292S/Fc-mono.
SEQ ID NO:27 - аминокислотная последовательность Fc-домена.
ПОДРОБНОЕ ОПИСАНИЕ
В осуществлении настоящего изобретения на практике используют, если не указано иначе, общепринятые способы клеточной биологии, молекулярной биологии, культивирования клеток, вирусологии и т.п., находящиеся в компетенции специалистов в данной области. Эти способы полностью описаны в настоящем документе и/или в современной литературе, например, Sambrook, Fritsch and Maniatis eds., «Molecular Cloning, A Laboratory Manual», 2nd Ed., Cold Spring Harbor Laboratory Press (1989); Celis J. E. «Cell Biology, A Laboratory Handbook» Academic Press, Inc. (1994) и Bahnson et al, J. of Virol. Methods, 54: 131-143 (1995).
Предусматривают, что любой способ, препарат или композицию, описанные в настоящем документе, можно осуществлять по отношению к любому другому способу, препарату или композиции, описанным в настоящем документе. Использование слова «a» или «an» при использовании в сочетании с термином «содержащий» в формуле изобретения и/или в описании может означать «один», но оно согласуется также со значением «один или несколько», «по меньшей мере один» и «один или более одного». Использование термина/фразы «и/или» при использовании со списком означает, что можно использовать один или несколько из перечисленных пунктов, например, он не является ограниченным одним или всеми элементами.
В ходе продукции и очистки аналога белка фактора комплемента B, обозначенного hfB3 (SEQ ID NO:4), детектированы две популяции аналога белка фактора комплемента B. Одна популяция обладала желаемой активностью для аналога белка фактора комплемента B (пик I), в то время как другая популяция обладала по существу меньшей, чем желательная, активностью (пик II), например, см. фигуры 6 и 7. Результаты характеризации двух популяций позволяют предполагать, что две популяции различались по характерам их дисульфидных связей. Когда свободный цистеин (положение 292 SEQ ID NO:4) в результате мутации меняли на серин, пик II не поддавался детекции. На фигуре 6 показано, что фракция пика II обладает некоторой способностью ингибировать активность комплемента/гемолитическую активность, но намного меньшей способностью на мкг белка по сравнению с фракцией пика I. Возможно, что активность ингибирования комплемента/гемолиза пиком II является результатом в основном или единственно того, что пик II содержит некоторое количество hfB3 со свободным цистеином в положении 292, возможно, в результате того, что пик I и пик II не полностью отделены друг от друга.
Цистеин, соответствующий положению 292 SEQ ID NO:1 является высоко консервативным среди белков факторов комплемента B различных видов млекопитающих (например, см. таблицу 1). Высоко консервативные последовательности являются, как правило, важными для функционирования белка. «Нейтральная теория молекулярной эволюции гласит, что мутации аминокислот возникают стохастически постоянным образом, пока мутации не оказывают эффекта на функцию продукта гена [Kimura M: The neutral theory of molecular evolution. Sci Am 1979, 241(5):98-100, 102, 108 passim]. С другой стороны, аминокислоты, являющиеся важными для функции и структуры белка, не могут мутировать без вредного эффекта на активность белка. Таким образом, эти аминокислоты будут меняться очень медленно в данном семействе белков в ходе эволюции». (Liu et al. BMC Bioinformatics 2006, 7:37)

Замена мутацией цистеина в аминокислоте 292 аналога белка фактора комплемента B на серин, например, как показано в SEQ ID NO:2 (MB3-292), существенно уменьшала, если не уничтожала, количество в популяции пика II (по существу менее активной) и аналог белка фактора комплемента B hfB3-292S сохранял свою активность, в этом случае, способность ингибировать или снижать активность комплемента. (Например, см. пример 10, ниже.)
Без желания быть связанными теорией, менее активная популяция (фракция пика II) белка hfB3 может являться результатом неправильного сворачивания белка hfB3. Возможно, образование большей части популяции пика II было обусловлено комбинацией свободного цистеина и мутаций, введенных в белок hfB3, поскольку когда клетки конструировали для экспрессии белка фактора комплемента B дикого типа человека (SEQ ID NO:1) способом, сходным со способом, используемым для белка hfB3, детектировали только одну популяцию белка фактора B дикого типа человека (данные не представлены).
Эта мутация свободного цистеина позволяет более высокий выход активного аналога фактора комплемента B, поскольку большая часть, если не все, из продуцированного аналога белка фактора комплемента B присутствует в активной форме. Кроме того, мутация свободного цистеина приводит к более стабильному белку, поскольку все из оставшихся не подвергшихся мутации цистеинов являются частью дисульфидной связи, и не остается свободного цистеина, который может участвовать в возможно нежелательных и вредных реакциях. Кроме того, мутация свободного цистеина неожиданно, по-видимому, уменьшает или исключает агрегацию аналога белка фактора комплемента B, например, см. пример 6 и фигуру 3.
Аналоги белка фактора комплемента B, описанные в Публикации PCT No. WO08/106644 или Публикации патента США No. US20100120665 (полное содержание обоих из которых приведено в качестве ссылки), могут обладать мутациями свободного цистеина и еще сохранять их желательную функцию, в то же время извлекая пользу из вышеупомянутых преимуществ мутации. Таким образом, настоящее изобретение относится к любым аналогам белка фактора комплемента B, описанного в Публикации PCT No. WO08/106644 или Публикации патента США No. US20100120665, с мутацией свободного цистеина.
Термин «свободный цистеин» относится к цистеину, который является частью белка или пептида, где свободный цистеин не формирует дисульфидную связь с другим цистеином в том же самом белке или пептиде. В некоторых случаях «свободный цистеин» аналога белка не формирует дисульфидную связь с другим цистеином (в том же самом белке или пептиде), когда аналог белка обладает желательной активностью, но может формировать дисульфидную связь с другим цистеином (в том же самом белке или пептиде) в менее активной или неактивной форме аналога белка.
Термин «опосредованный комплементом» относится к процессу или заболеванию, включающему в себя комплемент. Как правило, «опосредованное комплементом» заболевание или состояние представляет собой то, где активность комплемента является одной из первопричин заболевания или состояния, и где ингибирование или блокирование активности комплемента снижает степень заболевания или состояния. Примеры многочисленных опосредованных комплементом заболеваний или состояний описаны в настоящем документе.
Термин «дикий тип» (или дикого типа), который используют взаимозаменяемо с «природный», относится к встречающемуся в природе белку, кодируемому геномом млекопитающих, к встречающейся в природе нуклеиновой кислоте, и т.д. В некоторых случаях, в действительности может существовать более одного белка, соответствующего варианту дикого типа, например, из-за аллельных различий; различных изоформ; и/или генетической изменчивости между различными индивидуумами вида.
Термин «аналог» относится к структурному производному белка (исходного белка). Аналог не обязательно сохраняет все свойства исходного белка и в некоторых случаях обладает по меньшей мере одним измененным свойством по сравнению с соответствующим природным исходным белком. В некоторых вариантах осуществления, исходный белок представляет собой природный (встречающийся в природе) белок. Аналог или вариант белка получают заменой, замещением, делецией и/или добавлением аминокислот по отношению к соответствующей природной аминокислотной последовательности белка. Замены или вставки, как правило, вовлекают встречающиеся в природе аминокислоты, но могут также включать в себя синтетические или редкие аминокислоты. В некоторых вариантах осуществления аналог или вариант получают подверганием белка мутациям, например, подверганием мутациям кодирующей его нуклеиновой кислоты. Аналог может, как правило, сохранять по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,5% аминокислотной последовательности соответствующего природного исходного белка (например, обладать этим процентом идентичности аминокислотной последовательности по сравнению с встречающимся в природе исходным белком, как определено на протяжении длины целого исходного белка или, в конкретных вариантах осуществления, на протяжении конкретного домена или части исходного белка). Аналоги включают в себя также фрагменты полноразмерных аналогов, которые содержат часть аминокислотной последовательности и либо сохраняют один или несколько видов биологической активности исходного белка или полноразмерного аналога, либо ингибируют один или несколько из этих видов биологической активности.
Термин «соответствует» или «соответствующий» при ссылке на аминокислоту в конкретном белке относится к конкретной аминокислоте в этом конкретном белке, а также к аминокислоте в родственном или сходном белке и может обеспечивать сходную функцию белка. Например, может быть обнаружено, что аминокислота в факторе комплемента B человека соответствует аминокислоте в факторе комплемента B мыши, или в аллельном варианте фактора B человека, как обычно определено посредством выравнивания двух аминокислотных последовательностей. Например, специалист в данной области может выравнивать две или более родственных последовательностей, таких как SEQ ID NO:9-14, для определения соответствующих аминокислот, например, с использованием программы BLAST (например, см. таблицу 1, выше). А также, соответствующие аминокислоты можно определять, например, посредством выравнивания мотивов (например, мотива расщепления протеазой) внутри родственных или неродственных белков. Такое выравнивание можно использовать также для выведения консенсусных последовательностей белка-мишени или его доменов.
Как применяют в настоящем документе, термин «ген», как правило, относится к кодирующей области белка. Однако, в некоторых контекстах в настоящем документе, понятно, что термин «ген» относится также к элементам (например, регуляторным элементам), функционально связанным с кодирующей областью, таким как промоторы, энхансеры, участки сплайсинга (акцепторы и/или доноры), сигналы полиаденилирования, интроны, 5' нетранслируемые области, 3'-нетранслируемые области и т.д.
Термин «фармацевтически приемлемый» обозначает одобренный регулирующими органами федерального или государственного правительства или перечисленный в Фармакопее США или другой общепризнанной фармакопее для использования для человека.
«Терапевтическая полезность» не обязательно представляет собой излечение конкретного заболевания или состояния (включая любое заболевание или состояние, описанное в настоящем документе), но вместо этого, охватывает результат, который по большей части, как правило, включает в себя облегчение заболевания или состояния, устранение заболевания или состояния, уменьшение одного или нескольких симптомов, ассоциированных с заболеванием или состоянием, предотвращение или облегчение вторичного заболевания или состояния, возникающего в результате возникновения первичного заболевания или состояния, уменьшение вероятности развития состояния или заболевания, уменьшение тяжести заболевания или состояния, изменение характера заболевания или состояния, сокращение течения заболевания или состояния, замедление или предотвращение прогрессирования или ухудшения заболевания или состояния, и/или предотвращение заболевания или состояния.
Аналоги фактора комплемента B
Настоящее изобретение относится к аналогам белка фактора комплемента B и полипептидам, включающим аналоги фактора комплемента, и их применениям. Некоторые варианты осуществления изобретения относятся к аналогу белка фактора комплемента B, где свободный цистеин подвергали мутации. В некоторых вариантах осуществления эта мутация свободного цистеина может включать в себя делецию свободного цистеина или замену свободного цистеина другой аминокислотой (аминокислотами). Свободный цистеин можно заменять по существу любой аминокислотой, которая еще позволяет аналогу белка фактора комплемента B сохранять по меньшей мере некоторые из желательных характеристик(и), такие как способность к понижающей регуляции, уменьшению или нарушению активности комплемента. Может происходить замена на одну или несколько аминокислот. В некоторых вариантах осуществления, свободный цистеин заменен на серин. В некоторых вариантах осуществления свободный цистеин заменен на одну или несколько аминокислот, выбранных из группы, состоящей из аланина, гистидина, изолейцина, лейцина, метионина, фенилаланина, серина, треонина, тирозина и валина. В некоторых вариантах осуществления свободный цистеин соответствует аминокислоте 292 из SEQ ID NO:1.
В некоторых вариантах осуществления изобретение относится к аналогам белка фактора комплемента B, которые не содержат свободный цистеин. Изобретение относится также к способам получения или продукции аналога белка фактора комплемента B, содержащего мутации свободного цистеина.
Мутацию свободного цистеина можно комбинировать с другими мутациями белка фактора комплемента B, например, другими мутациями, как описано в настоящем документе.
Изобретение относится также к аналогам белка фактора комплемента B, где цистеин, соответствующий аминокислоте 292 из SEQ ID NO:l, подвергают мутации. Эта мутация может представлять собой делецию, вставку или замену, такую как замена на серин или другие мутации, как описано в настоящем документе.
Аналоги могут включать в себя различные мутеины последовательности, отличные от встречающейся в природе аминокислотной последовательности. Например, отдельные или множественные замены аминокислот (например, консервативные или не консервативные замены аминокислот) можно выполнять во встречающейся в природе последовательности. Консервативная замена аминокислоты, как правило, не должна по существу изменять структурные характеристики исходной последовательности (например, замененная аминокислота не должна проявлять тенденцию разрушать спираль, возникающую в исходной последовательности, или разрушать другие типы вторичной структуры, характеризующие исходную последовательность). Примеры известных в данной области вторичных и третичных структур полипептидов описаны в Proteins, Structures and Molecular Principles (Creighton, Ed., W. H. Freeman and Company, New York (1984)); Introduction to Protein Structure (C. Branden и J. Tooze, eds., Garland Publishing, New York, N.Y. (1991)); и Thornton et al. Nature 354: 105 (1991). Консервативные замены включают в себя, но без ограничения, замены из следующих групп: кислые остатки Asp (D) и Glu (E); основные остатки Lys (K), Arg (R) и His (H); гидрофильные незаряженные остатки Ser (S), Thr (T), Asn (N) и Gln (Q); алифатические незаряженные остатки Gly (G), Ala (A), Val (V), Leu (L) и Ile (I); неполярные незаряженные остатки Cys (C), Met (M) и Pro (P); ароматические остатки Phe (F), Tyr (Y) и Trp (W); содержащие спиртовые группы остатки S и T; алифатические остатки I, L, V и M; ассоциированные с циклоалкенилом остатки F, H, W и Y; гидрофобные остатки A, C, F, G, H, I, L, M, R, T, V, W и Y; отрицательно заряженные остатки D и E; полярные остатки C, D, E, H, K, N, Q, R, S и T; положительно заряженные остатки H, K и R; маленькие остатки A, C, D, G, N, P, S, T и V; очень маленькие остатки A, G и S; остатки, вовлеченные в формирование поворота A, C, D, E, G, H, K, N, Q, R, S, P и T; и гибкие остатки Q, T, K, S, G, P, D, E и R.
В некоторых вариантах осуществления используют неконсервативную замену.
В некоторых вариантах осуществления, мутации включают в себя, но без ограничения, замены одной или нескольких аминокислот, делеции одной или нескольких аминокислот или вставки одной или нескольких аминокислот. Мутации включают в себя, но без ограничения, мутации, которые: (1) снижают чувствительность аналога фактора комплемента B к протеолизу, (2) снижают чувствительность аналога фактора комплемента B к окислению, (3) изменяют аффинность связывания аналога фактора комплемента B для формирования белковых комплексов, (4) изменяют (например, увеличивают или уменьшают) аффинности связывания аналога фактора комплемента B, (5) снижают иммуногенность аналога фактора комплемента B; (6) увеличивают стабильность (например, термостабильность) аналога фактора комплемента B; (7) уменьшают агрегацию аналога белка фактора комплемента B; или любые комбинации 1-7.
В некоторых вариантах осуществления, аналог фактора комплемента B человека по изобретению конкурирует со связыванием природного фактора комплемента B. Например, природный фактор комплемента B может связываться с фактором комплемента C3b для формирования C3bB, например, см. фигуру 1B. Фактор B, который является частью комплекса C3bB, может связывать фактор D. Таким образом, в некоторых вариантах осуществления, аналог фактора комплемента B по изобретению может конкурировать со связыванием природного фактора B за (i) связывание с C3b, (ii) связывание с фактором D или (iii) оба.
В некоторых вариантах осуществления аналог белка фактора комплемента B по изобретению представляет собой аналог SEQ ID NO:4, обладающий цистеиновыми аминокислотами, формирующими дисульфидные связи, и свободной цистеиновой аминокислотой, замененной на другую аминокислоту, более, чем одну аминокислоту или делетированной без замены.
В некоторых вариантах осуществления изобретения, аналог белка фактора комплемента B обладает увеличенной аффинностью связывания C3b по сравнению с соответствующим природным белком фактором комплемента B, и аналог белка фактора комплемента B обладает (i) уменьшенной протеазной активностью по сравнению с соответствующим природным белком фактором комплемента B; (ii) уменьшенной способностью к расщеплению белком фактором D по сравнению с соответствующим природным белком фактором комплемента B; или (iii) уменьшенной протеазной активностью по сравнению с соответствующим природным белком фактором комплемента B и уменьшенной способностью к расщеплению белком фактором D по сравнению с соответствующим природным белком фактором комплемента B.
В некоторых вариантах осуществления аналог фактора комплемента B содержит мутацию в связывающем C3b домене, и аналог белка фактора комплемента B обладает увеличенной аффинностью связывания с C3b по сравнению с аффинностью связывания соответствующего природного белка фактора комплемента B с C3b. В некоторых вариантах осуществления мутация в связывающем C3b домене включает (i) замену или делецию аспарагиновой кислоты, соответствующей аминокислоте 279 из SEQ ID NO:1, замену или делецию аспарагина, соответствующего аминокислоте 285 из SEQ ID NO:1 или обе; или (ii) вставку по меньшей мере одной аминокислоты, следующей за указанной аспарагиновой кислотой или указанным аспарагином. В некоторых вариантах осуществления эти аспарагиновая кислота, аспарагин или оба заменены на одну или несколько аминокислот. В некоторых вариантах осуществления аспарагиновая кислота, соответствующая аминокислоте 279 из SEQ ID NO:1 является замененной на глицин, аланин или аспарагин. В некоторых вариантах осуществления, аспарагин, соответствующий аминокислоте 285 из SEQ ID NO:1 является замененной на глицин, аланин или аспарагиновую кислоту. В некоторых вариантах осуществления аспарагиновая кислота, соответствующая аминокислоте 279 из SEQ ID NO:1, является замененной на глицин, и аспарагин, соответствующий аминокислоте 285 из SEQ ID NO:1, является замененной на аспарагиновую кислоту.
В некоторых вариантах осуществления аналог белка фактора комплемента B представляет собой аналог белка фактора комплемента B человека, основанный на белке факторе комплемента человека.
Настоящее изобретение относится к аналогам белка фактора комплемента B, например, которые можно доставлять в форме белков и/или посредством переноса гена для ослабления альтернативного пути активации комплемента. Эти аналоги могут преодолевать препятствия, мешающие развитию некоторых ингибиторов комплемента, включая, например: 1) избегание долгосрочной системной иммунной супрессии; 2) достижение эффективности несмотря на запретительно высокие в ином случае уровни факторов комплемента в крови; 3) достижение достаточных уровней и распределения терапевтического аналога белка фактора комплемента B поблизости от сетчатки и мембраны Бруха для эффективности; 4) достижение активности терапевтического аналога белка фактора комплемента B внутри друз; 5) достижение достаточной продолжительности терапевтической доставки для лечения хронического заболевания; 6) достижение эффективности без вредных помех активности классического пути активации комплемента в задней части глазного яблока; и/или 7) избегание или уменьшение иммунной реакции (например, местной иммунной реакции) на лекарственное средство.
Ослабление петли положительной обратной связи в альтернативном пути является средством понижающей регуляции всего альтернативного пути. Одним из подходящих способов ослабления петли обратной связи является создание помех для функции или уровней белка фактора комплемента B (IB). В некоторых вариантах осуществления изобретения используют аналог фактора комплемента B для ослабления активности комплемента.
Аналог белка фактора комплемента B по изобретению может содержать по меньшей мере одну мутацию, соответствующую мутации SEQ ID NO:1, выбранной из группы, состоящей из K258A, R259A, K260A, D279G, N285D и D740N. В некоторых вариантах осуществления, он содержит мутации, соответствующие K258A, R259A, K260A, D279G и N285D из SEQ ID NO:1. В некоторых вариантах осуществления аналог белка фактора комплемента B содержит мутацию, соответствующую D740N из SEQ ID NO:1.
С целью иллюстрации, конкретные аналоги белка фактора комплемента B описаны в настоящем документе. С белком фактором B можно манипулировать несколькими способами, например, для ингибирования или уменьшения активации альтернативного пути. В некоторых вариантах осуществления конкретные участки в факторе B можно изменять, например, сайт-специфическим мутагенезом, так чтобы молекула больше не функционировала полностью правильно. В некоторых вариантах осуществления часть или домен фермента (например, протеазный домен, представляющий собой сериновую протеазу) молекулы можно изменять, так чтобы молекула больше не обладала ферментативной активностью или обладала сниженной ферментативной активностью (например, сниженной по меньшей мере в 2 раза, 5 раз, 10 раз, 50 раз или 100 раз). В некоторых вариантах осуществления аналог белка фактора комплемента B содержит мутацию в активном участке домена сериновой протеазы, где мутация уменьшает или нарушает способность аналога белка фактора комплемента B расщеплять фактор комплемента C3 по сравнению с соответствующим природным белком фактором комплемента B.
В некоторых вариантах осуществления этого можно достигать изменением остатка, соответствующего аминокислоте 740 из SEQ ID NO:1. В некоторых вариантах осуществления эта мутация включает делецию или замену аспарагиновой кислоты, соответствующей аминокислоте 740 из SEQ ID NO:1. В некоторых вариантах осуществления аспарагиновая кислота (D), соответствующая аминокислоте 740 из SEQ ID NO:1, заменена на другую аминокислоту, такую как аспарагин (N), аланин (A), глутаминовая кислота (E), серин (S), тирозин(Y) или глицин (G). Нумерация конкретных аминокислот фактора B в настоящем документе относится к полному полипептиду, включающему сигнальный пептид и отражен на SEQ ID NO:1. В Hourcade et al. (JBC (1998) 273(40):25996-6000) указано, что аминокислоты 739-746 из SEQ ID NO:l (обозначенные в Hourcade et al. как аминокислоты 714-721, поскольку нумерация в Hourcade et al. не включает сигнальную последовательность/пептид из 25 аминокислот) играет роль в функции сериновой протеазы белка фактора B. Кроме того, N693, T694 и D740 могут составлять участок связывания субстрата или являться частью участка связывания субстрата, и H526, D576 и S699 могут составлять каталитический центр или являться частью каталитического центра, например, см. Xu et al., J Biol Chem. 2000 275(l):378-85. В некоторых вариантах осуществления, аналог белка фактора B содержит мутацию по меньшей мере одной из аминокислот, выбранных из аминокислот 739-746 из SEQ ID NO:1. В некоторых вариантах осуществления аналог белка фактора комплемента B содержит замену аминокислоты, соответствующей аминокислоте 739 из SEQ ID NO:1, на аланин. В некоторых вариантах осуществления аналог белка фактора комплемента B содержит замену аминокислоты, соответствующей аминокислоте 740 из SEQ ID NO:1 на аминокислоту, выбранную из группы, состоящей из аспарагина, глутаминовой кислоты, аланина, серина и тирозина. В некоторых вариантах осуществления аналог белка фактора комплемента B содержит замену аминокислоты, соответствующей аминокислоте 741 из SEQ ID NO:1, на аминокислоту, выбранную из группы, состоящей из триптофана и аланина. В некоторых вариантах осуществления аналог белка фактора комплемента B содержит замену аминокислоты, соответствующей аминокислоте 742 из SEQ ID NO:1, на глутамин. В некоторых вариантах осуществления аналог белка фактора комплемента B содержит замену аминокислоты, соответствующей аминокислоте 743 из SEQ ID NO:1 на фенилаланин. В некоторых вариантах осуществления аналог белка фактора комплемента B содержит замену аминокислоты, соответствующей аминокислоте 745 из SEQ ID NO:1 на фенилаланин. В некоторых вариантах осуществления аналог белка фактора комплемента B содержит замену аминокислоты, соответствующей аминокислоте 746 из SEQ ID NO:1, на триптофан или аланин. В некоторых вариантах осуществления аналог белка фактора B содержит мутацию одной или двух из аминокислот 693 и 694 из SEQ ID NO:1, например, замену или делецию. В некоторых вариантах осуществления аналог белка фактора B содержит мутацию одной или двух из аминокислот 526, 576 и 699 из SEQ ID NO:1, например, замену или делецию.
Другие участки в факторе B, которые можно изменять, включают в себя: 1) участок связывания для пропердина (связывающий пропердин домен), так что связывание происходит с более низкой аффинностью (например, такое как сниженная в 2 раза, 5 раз, 10 раз, 50 раз или 100 раз аффинность по сравнению с белком фактором B дикого типа) или с более высокой аффинностью (такой как увеличенная по меньшей мере в 2 раза, 5 раз, 10 раз, 50 раз или 100 раз аффинность по сравнению с фактором B дикого типа); 2) участок связывания для белка C3b (связывающий C3b домен), так что связывание происходит с более низкой аффинностью (такой как по меньшей мере в 2 раза, 5 раз, 10 раз, 50 раз или 100 раз сниженная аффинность по сравнению с белком фактором B дикого типа) или с более высокой аффинностью (такой как увеличенная по меньшей мере в 2 раза, 5 раз, 10 раз, 50 раз или 100 раз аффинность по сравнению с белком фактором B дикого типа, например, этого можно достигать заменой аминокислоты, соответствующей положению 279 и/или положению 285 из SEQ ID NO:1 на другие аминокислоты, например, где аминокислота в положении, соответствующем положению 279 является замененной на аспарагин (N), аланин (A) или глицин (G), и/или аминокислота в положении, соответствующем положению 285, является замененной на аспарагиновую кислоту (D) или аланин (A)); 3) участок, действующий на фактор D, так что фактор D обладает сниженной способностью к расщеплять или больше не расщепляет фактор B с образованием Bb (например, в участке расщепления фактором D, по меньшей мере одну из аминокислот в положениях, соответствующих положениям 258, 259 или 260 SEQ ID NO:1, например, можно изменять на аланин (A) или; комбинацию из 1, 2 и/или 3 выше).
В некоторых вариантах осуществления аналог белка фактора комплемента B содержит изменение в участке расщепления фактора комплемента D, где изменение уменьшает или нарушает расщепление аналога белка фактора комплемента B белком фактором D. В некоторых вариантах осуществления изменение в участке расщепления фактора D включает (i) замену или делецию аргинина, соответствующего аминокислоте 259 из SEQ ID NO:1, замену или делецию одного или обоих лизинов, соответствующих аминокислоте 258 или 260 из SEQ ID NO:1, или замену или делецию аргинина и обоих лизинов; или (ii) вставку после аргинина, после одного или обоих лизинов, или после аргинина и одного или обоих лизинов. В некоторых вариантах осуществления аналог белка фактора комплемента B обладает аминокислотами, соответствующими аминокислотам 258-260 из SEQ ID NO:1, где каждая заменена на аланин.
Изобретение относится к (i) аналогам белка фактора комплемента B, которые связывают оба фактора C3b и D; (ii) аналогам белка фактора комплемента B с увеличенным связыванием (по сравнению с их природной формой) с обоими факторами C3b и D; (iii) аналогам белка фактора комплемента B с увеличенным связыванием (по сравнению с их природной формой) с белком фактором D; и (iv) аналогам белка фактора комплемента B с увеличенным связыванием (по сравнению с их природной формой) с комплексом C3bB. Изобретение относится также к способам ингибирования пути комплемента с использованием аналогов белка фактора комплемента B по изобретению, таких как i-iv, выше.
В некоторых вариантах осуществления увеличенное связывание является увеличенным в от приблизительно 1,5 до приблизительно 10000, от приблизительно 10 до приблизительно 10000, от приблизительно 100 до приблизительно 10000, от приблизительно 1000 до приблизительно 10000, от приблизительно 1,5 до приблизительно 1000, от приблизительно 1,5 до приблизительно 100, от приблизительно 1,5 до приблизительно 10, от приблизительно 2 до приблизительно 5, от приблизительно 2 до приблизительно 10, от приблизительно 5 до приблизительно 10, от приблизительно 5 до приблизительно 20, от приблизительно 10 до приблизительно 20, от приблизительно 10 до приблизительно 30, от приблизительно 20 до приблизительно 30, от приблизительно 30 до приблизительно 50, от приблизительно 50 до приблизительно 100, от приблизительно 100 до приблизительно 500, от приблизительно 500 до приблизительно 1000, от приблизительно 1000 до приблизительно 5000, или от приблизительно 5000 до приблизительно 10000 раз. В некоторых вариантах осуществления увеличенное связывание является увеличенным более чем в 1,5, 2, 3, 4, 5, 10, 50, 100, 500, 1000, 5000 или 10000 раз. В некоторых вариантах осуществления увеличенное связывание можно измерять посредством иммунопреципитации или с использованием Biacore (GE Healthcare, Piscataway, NJ), например, по сравнению с белком дикого типа. В качестве примера для (i) выше, связывание можно измерять посредством иммунопреципитации белка с помощью связывающей молекулы для белка C3b и затем детекции белка фактора D в иммунопреципитате, например, с использованием иммуноанализа, такого как ELISA или Вестерн, например, где увеличенное связывание показывают в виде полосы увеличенной интенсивности по Вестерну.
Некоторые аналоги белка фактора B по изобретению могут обладать увеличенным связыванием с белком C3b и/или белком фактором D с кратностью в 2 раза, 4 раза, 5 раз, 10 раз, 20 раз, 50 раз, 100 раз, 500 раз, 1000 раз по сравнению со связыванием фактора B дикого типа с C3b и/или фактором D. В некоторых вариантах осуществления, увеличенное связывание можно измерять посредством иммунопреципитации или с использованием Biacore (GE Healthcare, Piscataway, NJ).
Некоторые модифицированные аналоги белка фактора B по изобретению содержат одно или несколько изменений аминокислот, обсуждаемых в настоящем документе, и кроме того, обладают одним или несколькими дополнительными заменами, вставками или изменениями аминокислот (например, по меньшей мере или не более, чем 1, 2, 5, 8, 10, 15 20, 50, 100 или 200 изменений), где аналоги сохраняют увеличенное связывание с C3b и/или фактором D или другую биологическую активность аналогов белка фактора B, обсуждаемую в настоящем документе, опосредующую ингибирование пути комплемента. Такие аналоги могут обладать по меньшей мере 99,9%, 99%, 98%, 95%, 90%, 85%, 80%, 75% или 70% идентичностью аминокислотной последовательности с белком фактором B дикого типа, например, с аминокислотной последовательностью из SEQ ID NO:1, и сохранять увеличенное связывание с C3b и/или фактором D.
В контексте генотерапии и экспрессии аналогов белка фактора комплемента B, изменения/мутации могут быть отражены на уровне кодирующей нуклеиновой кислоты. Таким образом, можно также выполнять модификации нуклеиновой кислоты для усиления экспрессии. Например, конкретная клетка-хозяин может предпочитать конкретные кодоны. Таким образом, можно проводить перекодирование, где конкретные кодоны являются предпочтительными, например, в конкретной системе экспрессии или клетке у млекопитающих.
Некоторые варианты осуществления изобретения относятся к полинуклеотидам и клеткам-хозяевам (или многоклеточным организмам-хозяевам), пригодным для продукции аналога белка фактора комплемента B, и относятся к аналогам, например, hfB3-292S (SEQ ID NO:2), hfB3-292S-740N (SEQ ID NO:3), hfB3-292S-Fc- (SEQ ID NO:22), hfB3-292S-740N-Fc- (SEQ ID NO:23) или hfB3-292S/Fc-mono (SEQ ID NO:26). Представлены также способы выделения и тестирования опосредованной комплементом активности этих аналогов белка фактора комплемента B. Некоторые аспекты изобретения относятся к фармацевтическим композициям/препаратам, где аналог белка фактора комплемента B представляет собой активный ингредиент в терапевтическом и/или профилактическом контекстах. Некоторые варианты осуществления изобретения относятся также к способам лечения опосредованных комплементом нарушений с использованием терапевтически эффективного количества аналога белка фактора комплемента B.
В некоторых вариантах осуществления изобретения аналог фактора комплемента B может обладать характерами гликозилирования, отличающимися от характеров гликозилирования встречающегося в природе фактора комплемента B, или может быть полностью лишен гликозилирования. Углеводы можно добавлять к аналогам фактора B и/или удалять из аналогов фактора B, содержащих последовательности участков гликозилирования для N- и/или O-связанного гликозилирования in vitro, например, с помощью системы микросом поджелудочной железы собаки (например, см. Mueckler and Lodish (1986) Cell 44:629 и Walter, P. (1983) Meth. Enzymol. 96:84) или т.п. Можно получать аналог фактора комплемента B по изобретению, содержащий добавление или делецию/мутацию аминокислотной последовательности, соответствующей участку гликозилирования, например, изменение характера/статуса гликозилирования белка может изменять функциональные характеристики белка. Например, hfB2, hfB3 (SEQ ID NO:4), hfB3-292S (SEQ ID NO:2), hfB3-292S-740N (SEQ ID NO:3), hfB3-292S-Fc (SEQ ID NO:22), hfB3-292S-740N-Fc (SEQ ID NO:23) и hfB3-292S/Fc-mono (SEQ ID NO:26) содержат замену N285D, по сравнению с фактором B дикого типа (SEQ ID NO:1), приводящую к удалению участка N-гликозилирования. (Оба аналога hfB2 и hfB3 белка фактора комплемента B подробно описаны в Публикации патента США No. US20100120665.) Потеря участка N-гликозилирования изменяет характеристики белка. Такого же эффекта можно достигать продукцией белка в клетке, которая обладает измененным характером гликозилирования или не гликозилирует этот N285. Например, белок фактор B или аналог можно продуцировать в клетке E. coli, которая не гликозилирует N285. В некоторых вариантах осуществления аналог белка фактора комплемента B продуцирует клетка (например, E. coli), которая не гликозилирует аминокислоту, соответствующую аминокислоте белка фактора B дикого типа, которая, как правило, является гликозилированной, например, соответствующую аминокислоте N285 из SEQ ID NO:1.
Как показано в Публикации PCT No. WO08/106644 и Публикации патента США No. US20100120665, в некоторых случаях природный белок фактор B из одного вида может обладать активностью в реакции/пути комплемента из других видов. Таким образом, настоящее изобретение относится также к аналогам белка фактора комплемента B из одного вида для ингибирования активности комплемента в других видах.
В некоторых вариантах осуществления аналог белка фактора комплемента B по изобретению является пегилированным, например, см. Roberts et al, Advanced Drug Delivery Reviews 54(4):459-476 (2002); Veronese, Biomaterials 22(5):405-417 (2001); Fee and Alstine, Chemical Engineering Science 61(3):924-939 (2006); Kodera et al, Progress in Polymer Science 23(7): 1233-1271 (1998); Morar, Biopharm International 19(4):34 (2006); и Veronese and Pasut, Drug Discovery Today 10(21): 1451-1458 (2005). Полиэтиленгликоль (PEG) можно присоединять к аналогу белка фактора комплемента B по изобретению. В некоторых вариантах осуществления PEG присоединен в присутствии или в отсутствие многофункционального линкера либо посредством сайт-специфической конъюгации PEG (например, к N-концу или к C концу аналога фактора комплемента B), либо через эпсилон-аминогруппы, присутствующие на остатках лизина. В некоторых вариантах осуществления можно использовать дериватизацию линейным или разветвленным полимером, приводящую к минимальной потере биологической активности. Степень конъюгации можно тщательно контролировать посредством SDS-PAGE и масс-спектрометрии для обеспечения правильной конъюгации молекул PEG с аналогом фактора комплемента B. В некоторых вариантах осуществления непрореагировавший PEG можно отделять от конъюгатов PEG посредством эксклюзионной и/или посредством ионообменной хроматографии.
В конкретных вариантах осуществления карбоксиконец, аминоконец или оба из аналога фактора комплемента B, являются химически модифицированными. Аминоконцевые модификации, такие как ацилирование (например, ацетилирование) или алкилирование (например, метилирование) и карбоксиконцевые модификации, такие как амидирование, так же как другие концевые модификации, включая циклизацию, можно включать в различные варианты осуществления изобретения. Конкретные аминоконцевые и/или карбоксиконцевые модификации и/или удлинения пептида (такие как слияния с гетерологичным полипептидом, таким как альбумин, иммуноглобулин или его часть, такая как Fc-домен иммуноглобулина) от коровой последовательности могут предоставлять преимущественные физические, химические, биохимические и фармакологические свойства, такие как: увеличенная стабильность, увеличенная активность и/или эффективность, устойчивость к протеазам сыворотки, желательные фармакокинетические свойства и другие.
В некоторых вариантах осуществления аналог белка фактора комплемента B по изобретению содержит Fc-домен иммуноглобулина, например, Fc-домен иммуноглобулина человека. В некоторых вариантах осуществления Fc-домен находится на C-конце от аминокислотной последовательности соответствующего аналога фактора комплемента B. В некоторых вариантах осуществления Fc- домен содержит аминокислоты 766-990 из SEQ ID NO:21 или аминокислоты 766-1003 или 786-1003 of SEQ ID NO:26, или состоит из них. В некоторых вариантах осуществления Fc-домен представляет собой аминокислоты 1-239 или 2-239 из SEQ ID NO:27. В некоторых вариантах осуществления, Fc-домен представляет собой Fc-домен человека. В некоторых вариантах осуществления Fc-домен происходит из иммуноглобулина, например, IgG, такого как IgG4. В некоторых вариантах осуществления Fc-домен является способным формировать димер с другим Fc-доменом. В некоторых вариантах осуществления аналог фактора комплемента B по изобретению является способным формировать димеры (например, гомологичные димеры), в качестве неограничивающих примеров, посредством взаимодействия Fc-доменов. В некоторых вариантах осуществления аналог фактора комплемента B не формирует димеры, или основная часть популяции/препарата аналога фактора комплемента B присутствует в форме мономеров. В некоторых вариантах осуществления, аналог фактора комплемента B, содержащий Fc-домен, не формирует димеры или основная часть популяции/препарата этого аналога фактора комплемента B присутствует в форме мономеров. В некоторых вариантах осуществления аналог фактора комплемента B содержит Fc-домен, обладающий одной или несколькими мутациями цистеина(цистеинов) в последовательности Fc-домена, например, цистеина, вовлеченного в димеризацию Fc-домена. В некоторых вариантах осуществления Fc- домен содержит мутацию одного или нескольких цистеинов, соответствующих аминокислотам 17 и/или 20 из SEQ ID NO:27. В некоторых вариантах осуществления аналог фактора комплемента B содержит Fc-домен, обладающий одной или несколькими мутациями свободного цистеина(цистеинов) в последовательности Fc-домена. В некоторых вариантах осуществления цистеин(ы) является замененным на аминокислоту, выбранную из группы, состоящей из гистидина, изолейцина, лейцина, метионина, фенилаланина, серина, треонина, тирозина и валина. В некоторых вариантах осуществления цистеин является делетированным. В некоторых вариантах осуществления аминокислотную последовательность линкера используют между аминокислотными последовательностями для аналога фактора комплемента B и Fc-областью. В некоторых вариантах осуществления этот линкер представляет собой одну аминокислоту, например, аргинин.
В некоторых вариантах осуществления полипептид содержит как усеченный аналог фактора комплемента B, так и Fc-область, например, содержит аминокислоты, соответствующие аминокислотам 26-480 из SEQ ID NO:2, и Fc-область.
Кроме того, изобретение относится к аналогам, представляющим собой фрагменты белка фактора комплемента B или к аналогу, который содержит часть белка фактора комплемента B или аналога по меньшей мере из 20, 30, 50, 70, 100, 150, 200, 300, 400, 480, 500, 600 или 700 аминокислот, и/или содержит 1, 2 или 3 домена белка и имеет или сохраняет один или несколько видов биологической активности белка фактора комплемента B дикого типа или аналога и/или действует как ингибитор одного аспекта системы комплемента (классического пути, альтернативного пути или обоих). Эти фрагменты можно дополнительно модифицировать соединением или слиянием с другим белком или фрагментом, таким как Fc, для увеличения стабильности и/или времени полужизни аналога. В некоторых вариантах осуществления аналог представляет собой фрагмент белка фактора комплемента B или аналога, и фрагмент обладает по меньшей мере 99,5%, 99%, 98%, 95%, 90%, 85% или 62% идентичностью с соответствующей аминокислотной последовательностью фактора комплемента B дикого типа.
Изобретение относится к белкам, содержащим фрагмент или аналог белка фактора комплемента B, где фрагмент обладает усечением по N-концу и/или C-концу. В некоторых вариантах осуществления аналог фактора комплемента B содержит усечение по C-концу, где аналог является усеченным по аминокислоте или после (на C-конце от) аминокислоты, соответствующей аминокислоте 407, 427, 457, 477, 480, 484, 487, 507 или 527 из SEQ ID NO:1, 2 или 4. В некоторых вариантах осуществления аналог фактора комплемента B содержит усечение по C-концу, где аналог является усеченным по аминокислоте между аминокислотами, соответствующими аминокислотам 407-487, 470-495 или 477-487 из SEQ ID NO:1, 2 или 4. В некоторых вариантах осуществления аналог фактора комплемента B по изобретению не содержат аминокислот, соответствующих аминокислотам 408-764, 428-764, 458-764, 478-764, 481-764, 485-764, 488-764, 507-764, 527-764 из SEQ ID NO:1, 2 или 4. В некоторых вариантах осуществления усечение или фрагментация белка фактора B или аналога может создавать свободный цистеин. Например, усечение может приводить к делеции одного цистеина из пары цистеинов, формирующих дисульфидную связь в природном белке факторе комплемента B, таким образом, образуя полипептид, который содержит цистеин, но не содержит его природного цистеинового «партнера». В некоторых из этих вариантов осуществления оставшийся цистеин из пары можно подвергать мутации, например, заменять на другую аминокислоту, такую как аланин, гистидин, изолейцин, лейцин, метионин, фенилаланин, серин, треонин, тирозин и валин, или цистеин можно делетировать, например, для исключения возможных нежелательных дисульфидных связей.
Аналоги по изобретению можно получать различными способами, включая, но без ограничения, химический синтез или экспрессию рекомбинантного аналога.
Нуклеотидные последовательности генов и кодирующих областей, кодирующих фактор B человека, так же как аминокислотные последовательности, известны в данной области. Например, ген, кодирующий фактор B человека, обнаружен в базе данных NCBI с номером доступа No. NG_000013. Кодирующая последовательность фактора B человека обнаружена в базе данных NCBI с номером доступа No. NM_001710 и аминокислотная последовательность препробелка фактора комплемента B человека обнаружена в базе данных NCBI с номером доступа No. NP_001701 или P00751. Последовательности из других видов животных также известны в данной области. Для сравнения, в белковой последовательности фактора B мыши (например, см. базу данных NCBI, номер доступа No. P04186), третий домен SCR локализован в положениях 160-217 этого пребелка из 761 аминокислоты, и зрелый белок фактор B мыши перекрывает положения 23-761. Первые 22 аминокислоты белка фактора B мыши содержат сигнальную последовательность.
Как правило, пребелок фактор B человека представляет собой белок из 764 аминокислот (например, см. SEQ ID NO:1) с сигнальным пептидом, перекрывающим положения аминокислот 1-25. Зрелая цепь фактора B соответствует положениям 26-764 (например, см. SEQ ID NO:l). Три области SCR фактора B человека представляют собой SCR1, известную также как Sushi 1, перекрывающую от приблизительно положения 35 до приблизительно положения 100, SCR2, известную также как Sushi 2, перекрывающую от приблизительно положения 101 до приблизительно положения 160, и SCR3, известную также как Sushi 3, перекрывающую от приблизительно положения 163 до приблизительно положения 220.
В Публикации PCT No. WO08/106644 и Публикации патента США No. US20100120665 описаны, среди прочих, три специфических доминантно негативных аналога белка фактора B человека, обозначенные как hfB1, hfB2 и hfB3. Первый из этих трех аналогов, названный fB1, содержит мутированную аминокислоту в участке протеазы фактора B (fB). Эта группа fB связывает C3b с нормальной аффинностью и кинетикой, но после действия на него фактора D (fD) и стабилизации посредством пропердина, не функционирует как протеаза и не формирует C3 конвертазу. fB1 содержит замену на N аминокислоты, соответствующей аминокислоте 740 из SEQ ID NO:l (например, D740N). Во втором из этих аналогов фактора комплемента B, обозначенном fB2, изменена та же самая аминокислота, как в fB1, но кроме того, изменены две дополнительные аминокислоты в связывающем C3b домене (замены аминокислот, соответствующих аминокислотам 279 и 285 из SEQ ID NO:1), что увеличивает аффинность связывания fB2 с C3b, например, замены D279G, N285D и D740N. Замена N285D удаляет предположительный участок N-гликозилирования. В третьем из этих аналогов белка фактора комплемента B, названном hfB3, объединены мутации, увеличивающие связывание C3b, из fB2 с мутацией, выключающей участок расщепления фактором D, в частности, с заменами аминокислот, соответствующих остаткам 258, 259 и 260 из SEQ ID NO:1, так же как с заменами аминокислот, соответствующих остаткам 279 и 285, например, заменами K258A, R259A, K260A, D279G и N285D. Расщепление fB дикого типа фактором D активирует fB протеазу. Таким образом, hfB3, с его пятью заменами аминокислот, эффективно связывает C3b, но обладает минимальной протеазной активностью.
hfB1, hfB2 и hfB3 являются примерами аналогов фактора B человека, которые можно далее модифицировать до аналогов фактора комплемента B по настоящему изобретению посредством мутации свободного цистеина, соответствующего аминокислоте 292 из SEQ ID NO:1, например, посредством замены цистеина на серин, но изобретение не является ограниченным этими конкретными аналогами. Некоторые варианты осуществления изобретения относятся к любому аналогу фактора комплемента B, который модулирует путь комплемента и не содержит свободный цистеин. В некоторых вариантах осуществления аналог фактора комплемента B содержит, в дополнение к мутации свободного цистеина, одну или несколько мутаций аминокислот, соответствующих одной или нескольким из следующих аминокислот в SEQ ID NO:1: аминокислота 258, 259, 260, 279, 285, 739, 740, 741, 742, 743, 744, 745 и 746. Эти одна или несколько мутаций могут представлять собой замену или делецию аминокислоты или добавление по меньшей мере одной аминокислоты сразу после или в пределах 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот из соответствующих аминокислот. В некоторых вариантах осуществления, это добавление нарушает, изменяет, усиливает или ингибирует функцию перечисленных аминокислот, например, нарушает их роль (i) в расщеплении другого белка (например, 740), (ii) в качестве участка расщепления другим белком (например, аминокислоты, соответствующие остаткам 258, 259 и/или 260 из SEQ ID NO:1), или (iii) их роль в связывании другого белка (например, аминокислоты, соответствующие остаткам 279 или 285 из SEQ ID NO:1).
Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей одной или нескольким из аминокислот, соответствующих 258, 259 и/или 260 из SEQ ID NO:l, например, на аминокислоту, выбранную из группы, состоящей из аланина, глицина, валина, лейцина и изолейцина. Некоторые варианты осуществления изобретения содержат делецию аминокислоты, соответствующей одной, двум или трем из аминокислот, соответствующих аминокислотам 258, 259 и/или 260 из SEQ ID NO:l. Некоторые варианты осуществления изобретения содержат по меньшей мере одно добавление 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот непосредственно после аминокислоты, соответствующей аминокислотам 258, 259 и/или 260 из SEQ ID NO:1.
Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 739 из SEQ ID NO:1. Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 739 из SEQ ID NO:l, например, на аминокислоту, выбранную из группы, состоящей из аланина, глицина, валина, лейцина и изолейцина. Некоторые варианты осуществления изобретения содержат делецию аминокислоты, соответствующей аминокислоте 739 из SEQ ID NO:1.
Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 740 из SEQ ID NO:l, например, на аминокислоту, выбранную из группы, состоящей из глутаминовой кислоты, аспарагина, аланина, серина, глицина и тирозина. Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 740 из SEQ ID NO:l, на аминокислоту, выбранную из группы, состоящей из валина, лейцина, изолейцина, треонина, цистеина, метионина, глутамина, фенилаланина, тирозина, триптофана, глутаминовой кислоты, аспарагина, аланина, серина, глицина и тирозина. Некоторые варианты осуществления изобретения содержат делецию аминокислоты, соответствующей аминокислоте 740 из SEQ ID NO:1.
Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 741 из SEQ ID NO:l, например, на аминокислоту, выбранную из группы, состоящей из триптофана и аланина. Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 741 из SEQ ID ΝΟ: 1, на аминокислоту, выбранную из группы, состоящей из аланина, глицина, валина, лейцина и изолейцина. Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 741 из SEQ ID NO:l, на аминокислоту, выбранную из группы, состоящей из триптофана, тирозина и фенилаланина. Некоторые варианты осуществления изобретения содержат делецию аминокислоты, соответствующей аминокислоте 741 из SEQ ID NO:1.
Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 742 из SEQ ID NO:l, например, на глутамин. Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 742 из SEQ ID NO:1, на аминокислоту, выбранную из группы, состоящей из глутамина, глутаминовой кислоты, аспарагина и аспарагиновой кислоты. Некоторые варианты осуществления изобретения содержат делецию аминокислоты, соответствующей аминокислоте 742 из SEQ ID NO:1.
Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 743 и/или 745 из SEQ ID NO:1, например, на фенилаланин. Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 743 и/или 745 из SEQ ID NO:1, на аминокислоту, выбранную из группы, состоящей из фенилаланина, тирозина и триптофана. Некоторые варианты осуществления изобретения содержат делецию одной или нескольких из аминокислот, соответствующих аминокислотам 743, 744 и/или 745 из SEQ ID NO:1.
Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 746 из SEQ ID NO:1, например, на аминокислоту, выбранную из группы, состоящей из триптофана и аланина. Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 746 из SEQ ID NO:1, на аминокислоту, выбранную из группы, состоящей из аланина, глицина, валина, лейцина и изолейцина. Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 746 из SEQ ID NO:1, на аминокислоту, выбранную из группы, состоящей из триптофана, тирозина и фенилаланина. Некоторые варианты осуществления изобретения содержат делецию аминокислоты, соответствующей аминокислоте 746 из SEQ ID NO:1.
Некоторые варианты осуществления изобретения содержат вставку или замену 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот, непосредственно после или вместо любой одной или нескольких из аминокислот, соответствующих аминокислотам 739, 740, 741, 742, 743, 744, 745 и/или 746 из SEQ ID NO:1.
Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 279 из SEQ ID NO:l, например, на аминокислоту, выбранную из группы, состоящей из глицина, аланина и аспарагина. Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 279 из SEQ ID NO:1, на аминокислоту, выбранную из группы, состоящей из глицина, аланина, валина, лейцина и изолейцина. Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 279 из SEQ ID NO:1, на аминокислоту, выбранную из группы, состоящей из аспарагина, глутаминовой кислоты и глутамина. Некоторые варианты осуществления изобретения содержат делецию аминокислоты, соответствующей аминокислоте 279 из SEQ ID NO:1. Некоторые варианты осуществления изобретения содержат вставку или замену 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот непосредственно после или вместо аминокислоты, соответствующей аминокислоте 279 из SEQ ID NO:1.
Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 285 из SEQ ID NO:l, например, на аминокислоту, выбранную из группы, состоящей из аланина и аспарагиновой кислоты. Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 285 из SEQ ID NO:1, на аминокислоту, выбранную из группы, состоящей из глицина, аланина, валина, лейцина и изолейцина. Некоторые варианты осуществления изобретения содержат замену аминокислоты, соответствующей аминокислоте 285 из SEQ ID NO:1, на аминокислоту, выбранную из группы, состоящей из аспарагиновой кислоты, глутаминовой кислоты и глутамина. Некоторые варианты осуществления изобретения содержат делецию аминокислоты, соответствующей аминокислоте 285 из SEQ ID NO:1. Некоторые варианты осуществления изобретения содержат вставку или замену 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот непосредственно после или вместо аминокислоты, соответствующей аминокислоте 285 из SEQ ID NO:1.
Некоторые варианты осуществления изобретения содержат замену одной или нескольких из аминокислот, соответствующих аминокислотам 279, 282, 283, 284 и 285 из SEQ ID NO:1. В некоторых вариантах осуществления эти аминокислоты заменены на глицин, изолейцин, пролин, гистидин и аспарагиновую кислоту, соответственно.
Некоторые варианты осуществления изобретения содержат мутации аминокислот, соответствующих 258, 259, 260, 279 и 285 из SEQ ID NO:1, как описано в настоящем документе.
В некоторых конкретных вариантах осуществления изобретение относится к аналогу белка фактора B, содержащему аминокислотную последовательность из SEQ ID NO:2, 3, 21, 22 или 23 (необязательно, без какой-либо сигнальной последовательности, например, аминокислот 1-25, содержащихся в ней). Эти аналоги белка фактора B можно использовать в способах по изобретению.
Изобретение относится к аналогам белка фактора комплемента B, содержащим комбинацию мутаций (замен, делеций и добавлений) обсуждаемых в настоящем документе. В некоторых вариантах осуществления эти аналоги фактора комплемента B сохраняют один или несколько атрибутов аналогов hfB1, hfB2, hfB3, hfB3-292S или hfB3-292S-740N или любого другого аналога фактора комплемента B, обсуждаемого в настоящем документе. Изобретение, кроме того, относится к аналогам, которые представляют собой фрагменты (например, содержащие один или несколько доменов белка фактора комплемента B, обладающих одной или несколькими мутациями аминокислот, указанных в настоящем документе) этих аналогов, обладающие одним или несколькими из атрибутов аналогов, обсуждаемых выше, например, hfB3-292SN480. Кроме того, аналоги могут содержать дополнительные замены, делеции или вставки аминокислот (например, консервативные замены аминокислот, усечения с N-конца или C-конца и т.д.), так что аналог обладает по меньшей мере 99,5%, 99%, 98%, 95%, 90%, 85%, 80%, 75% или 75% идентичностью с соответствующим фактором комплемента B дикого типа или, в случае аналога, содержащего фрагмент фактора комплемента B, аналог обладает по меньшей мере 99,5%, 99%, 98%, 95%, 90%, 85%, 80%, 75% или 70% идентичностью между соответствующими частями аналога и фактора комплемента B дикого типа.
Белки hfB3-292S и hfB3-292SN480 обладают следующими характерными признаками от N-конца к C-концу: 1) природная сигнальная последовательность белка фактора B для эффективной секреции из экспрессирующих клеток млекопитающих, таких как клетки 293 человека, клетки CHO, клетки BHK; 2) мутированный участок C3b-зависимого расщепления белком фактором D (изменение аминокислот с K258R259K260 на A258A259A260); 3) мутированный участок связывания белка C3b (изменение аминокислоты с D279 на G279; и аминокислоты с N285 на D285), чтобы позволять его тесное связывание и захват белка C3b; и 4) мутантный свободный цистеин (изменение аминокислоты с C292 на S292) для увеличения количества «активного» или правильного свернутого белка.
В некоторых вариантах осуществления изобретения, аналог белка фактора комплемента B человека является по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 98% или по меньшей мере на 99% идентичным (i) SEQ ID NO:1, 2, 3, 22 или 23; (ii) аминокислотам 26-764 из SEQ ID NO:1, 2 или 3; (iii) аминокислотам 1-990 или 26-990 из любого из SEQ ID NO:22 или 23; аминокислотам 26-480 из SEQ ID NO:2; и (iv) аминокислотам 1-1003 или 26-1003 из SEQ ID NO:26. В некоторых вариантах осуществления аналог белка фактора комплемента B человека по изобретению содержит (i) SEQ ID NO:2, 3, 22, 23 или 26; (ii) аминокислоты 1-480 из SEQ ID NO:2; (iii) аминокислоты 26-480 из SEQ ID NO:2; (iv) аминокислоты 26-764 из SEQ ID NO:2 или 3; (v) аминокислоты 26-990 из SEQ ID NO:22 или 23; или (vi) аминокислоты 26-1003 из SEQ ID NO:26. В некоторых вариантах осуществления аналог белка фактора комплемента B состоит в основном из (i) аминокислот 26-764 из SEQ ID NO:2 или 3; (ii) аминокислот 26-480 из SEQ ID NO:2 или 3; (iii) аминокислот 26-990 из SEQ ID NO:22 или 23; или (iv) аминокислот 26-1003 из SEQ ID NO:26.
На фигуре 1 изображены классический и лектиновый пути комплемента (фигура 1a), и альтернативный путь комплемента (фигура 1b). Оба пути используют C3b. Комплекс C3bBb представляет собой C3 конвертазу, превращающую C3 в C3b. Спонтанная диссоциация («разложение») C3bBb в течение минут приводит к его инактивации, в то время как пропердин стабилизирует комплекс C3bBb. C3b участвует в C3 конвертазе для образования дополнительного C3b, таким образом создавая петлю положительной обратной связи, как показано большой стрелкой. Некоторые из факторов, ослабляющих пути комплемента, такие как усиливающие разложение фактор (DAF), осуществляют это посредством усиления разложения C3 конвертазы, C3bBb. Без намерения быть связанным теорией, некоторые из аналогов фактора комплемента B, описанных в настоящем документе, связывают C3b вместо природного фактора комплемента B, таким образом конкурируя с природным фактором комплемента B за связывание с C3b. Некоторые аналоги фактора комплемента B по изобретению связывают C3b для образования неактивного комплекса или комплекса со значительно сниженной активностью C3 конвертазы по сравнению с природным комплексом C3bBb.
Изобретение относится также к аналогам фактора комплемента B, содержащим мутации аминокислот, соответствующих тем аминокислотам в факторе комплемента B, которые взаимодействуют с факторами/молекулами, усиливающими разложение комплекса C3bBb. В некоторых вариантах осуществления аналог фактора комплемента B содержит мутации аминокислот, взаимодействующих с DAF. Эти мутации включают в себя, но без ограничения, одну или нескольких мутаций аминокислот, соответствующих аминокислотам 290, 291, 323, 363, 364 или 407 из SEQ ID NO:1. В некоторых вариантах осуществления эти мутации представляют собой замену или делецию одной или нескольких из аминокислот, соответствующих аминокислотам 290, 291, 323, 363, 364 или 407 из SEQ ID NO:1. В некоторых вариантах осуществления аналог фактора комплемента B содержит одну или несколько из следующих мутаций, соответствующих K323E, K290A, K291A, Y363A, S364A или D407N из SEQ ID NO:1. В некоторых вариантах осуществления аналог фактора комплемента B содержит одну из следующих комбинаций мутаций, соответствующих K290A/K291A, Y363A/S364A или K290A/K291A/Y363A/S364A из SEQ ID NO:1. Без желания быть связанными теорией, мутации аминокислот в аналоге фактора комплемента B, взаимодействующих с факторами/молекулами, которые усиливают разложение комплекса C3bBb, могут ингибировать разложение комплексов C3b и аналога фактора комплемента B по изобретению, таким образом, позволяя аналогу фактора комплемента B лучше ингибировать активность комплемента.
Иллюстративные способы получения кДНК (например, fB дикого типа человека и трех аналогов фактора комплемента B, так же как четырех аналогичных мышиных последовательностей) и их включения в векторы подробно описаны в Примере 8 Публикации PCT No. WO08/106644 и в Публикации патента США No. US20100120665.
Нуклеиновые кислоты
Изобретение относится к нуклеиновым кислотам, содержащим нуклеотидную последовательность, кодирующую аналог белка фактора комплемента B по изобретению, и относится к векторам, содержащим эти нуклеиновые кислоты.
Для обеспечения местной и долгосрочной экспрессии интересующей нуклеиновой кислоты, некоторые варианты осуществления изобретения включают в себя трансдукцию клетки нуклеиновой кислоты или вектором, кодирующими аналог фактора комплемента B по изобретению. Настоящее изобретение не следует рассматривать как ограниченное каким-либо конкретным способом доставки нуклеиновых кислот, и любой доступный носитель для способа доставки нуклеиновых кислот либо in vivo, либо in vitro, или применение модифицированных клеток (например, по способу Neurotech, Lincoln, RI, например, см. Патенты США No. 6231879; 6262034; 6264941 ; 6303136; 6322804; 6436427; 6878544), так же как нуклеиновые кислоты по изобретению, кодирующие аналог фактора комплемента B, сами по себе (например, «голую ДНК»), можно использовать в практическом осуществлении настоящего изобретения. Различные носители для доставки, такие как векторы, можно использовать по настоящему изобретению. Например, вирусные векторы, амфитрофные липиды, катионные полимеры, такие как полиэтиленимин (PEI) и полилизин, дендримеры, такие как гребенчатые молекулы и звездообразные молекулы, неионные липиды, анионные липиды, носители, липосомы и другие средства доставки генов из синтетических нуклеиновых кислот (например, см. Патенты США No. 6958325 и 7098030; Langer, Science 249: 1527-1533 (1990); Treat et al, in «Liposomes» in «The Therapy of Infectious Disease and Cancer»; и Lopez-Berestein & Fidler (eds.), Liss, New York, pp. 317-327 и 353-365 (1989); «голые» нуклеиновые кислоты и т.д. можно использовать в осуществлении настоящего изобретения на практике.
Вектор представляет собой средство, посредством которого интересующую нуклеиновую кислоту (например, терапевтическую нуклеиновую кислоту, например, нуклеиновую кислоту, которая может кодировать терапевтический белок) вводят в интересующую клетку-мишень. Вектор, как правило, конструируют или получают из исходного материала, такого как нуклеиновая кислота, которая способна нести чужеродный ген или трансген, и которую можно вводить в клетку-мишень и экспрессировать в ней. Пригодные исходные материалы, из которых можно получить вектор, включают в себя транспозоны, плазмиды, вирусы, продукты ПЦР, кДНК, мРНК и т.д., как известно в данной области. Способы получения или конструирования интересующего вектора включают в себя, но без ограничения, общепринятые способы манипуляций с генами, реакции секвенирования, расщепления рестрикционными ферментами, полимеразные реакции, ПЦР, ПЦР SOE, лигирования, рекомбиназные реакции (например, способ GATEWAY® от Invitrogen), другие ферменты, активные по отношению к нуклеиновым кислотам, материалы и методы для размножения бактерий и вирусов, химические вещества и реагенты, способы сайт-специфического мутагенеза и т.д., как известно в данной области, см., например, the Maniatis et al. text, «Molecular Cloning.»
Нуклеиновые кислоты по изобретению могут, как правило, содержать промотор, функционально связанный с кодирующей последовательностью аналога белка фактора комплемента B. Промотор может представлять собой, например, тканеспецифический промотор, специфический для клетки промотор, индуцируемый промотор, репрессируемый промотор, конститутивный промотор, синтетический промотор или гибридный промотор. Промоторы, пригодные для конструкций по изобретению, включают в себя, но без ограничения, промотор фага лямбда (PL); ранний промотор SV40; промотор вируса простого герпеса (HSV); промотор цитомегаловируса (CMV), такой как немедленный ранний промотор CMV человека; контролируемая тетрациклином отвечающая на транс-активаторы промоторная система (tet); промотор длинного концевого повтора (LTR), такой как MoMLV LTR, BIV LTR или HIV LTR; промотор области U3 вируса саркомы мышей Молони; промотор гранзима A; регуляторная последовательность(последовательности) гена металлотионеина; промотор CD34; промотор CD8; промотор тимидинкиназы (TK); промотор парвовируса B19; промотор PGK; индуцируемый глюкокортикоидами промотор; промотор белка теплового шока (HSP), такой как промоторы HSP65 и HSP70; промотор иммуноглобулина; промотор MMTV; промотор вируса саркомы Рауса (RSV); lac промотор; промотор CaMV 35S; и промотор нопалин-синтетазы. В некоторых вариантах осуществления промотор представляет собой промотор MND (Robbins et al., 1997, J. Virol. 71:9466-9474), или промотор MNC, который представляет собой производное промотора MND, в котором энхансеры LTR скомбинированы с минимальным промотором CMV (Haberman et al., J. Virol. 74(18):8732-8739, 2000).
В некоторых вариантах осуществления вектор по изобретению содержит интрон, например, как часть гена, кодирующего аналог белка фактора комплемента B. Известны гетерологичные интроны, и неограничивающие примеры включают в себя интрон гена β-глобина человека. Интрон может происходить из гена фактора комплемента B или представлять собой гетерологичный интрон.
Сигнальные последовательности или лидерные последовательности известны и могут быть использованы в экспрессирующих конструкциях аналога фактора комплемента B. Сигнальные последовательности транслируют в рамке считывания в форме пептида, присоединенного к амино-концу выбранного полипептида, секреторная сигнальная последовательность может вызывать секрецию полипептида посредством взаимодействия с аппаратом клетки-хозяина. В качестве части процесса секреции, эта секреторная сигнальная последовательность отщепляется. Секреторная сигнальная последовательность щелочной фосфатазы плаценты человека представляет собой пример пригодной сигнальной последовательности. Настоящее изобретение не является ограниченным конкретными секреторными сигнальными последовательностями, и другие известны специалистам в данной области. Термин «сигнальная последовательность» относится также к последовательности нуклеиновой кислоты, кодирующей секреторный пептид. Если включена сигнальная последовательность, она может представлять собой последовательность фактора комплемента B дикого типа, гомологичную последовательность или гетерологичную последовательность.
Вирусные векторы
Настоящее изобретение относится к вирусным векторам, кодирующим аналог(и) фактора комплемента B по изобретению. Примеры вирусных векторов, пригодных по настоящему изобретению, описаны в Публикации PCT No. WO08/106644 и Публикации патента США No. US20100120665. Настоящее изобретение не является ограниченным конкретным вирусным вектором. Вирусные векторы включают в себя, но без ограничения, ретровирусные векторы, лентивирусные векторы, аденовирусные векторы (см., например, Патент США No. 7045344), AAV векторы (например, см. Патент США No. 7105345), векторы из вируса герпеса (например, см. Патенты США No. 5830727 и 6040172), векторы из вируса гепатита (например, гепатита D) (например, см. Патент США No. 5225347), векторы SV40, векторы EBV (например, см. Патент США No. 6521449) и векторы из вируса болезни Ньюкасла (например, см. Патенты США No. 6146642, 7442379, 7332169 и 6719979). В некоторых вариантах осуществления лентивирусный вектор представляет собой вектор HIV, EIAV, SIV, FIV или BIV. Изобретение относится также к клетке, продуцирующей вирусный вектор по изобретению.
Примеры систем BIV описаны, например, в Matukonis et al., 2002 Hum. Gene Ther. 13, 1293-1303; Molina et al, 2002 Virology. 304, 10-23; Molina et al, 2004 Hum. Gene Ther., 15, 65-877; Патентах США No. 6864085, 7125712, 7153512; Публикации PCT No. WO08/106644 и Публикации патента США No. US20100120665.
Вирионы вектора по изобретению можно вводить in vivo или in vitro в клетки (например, клетки млекопитающих). Векторы (вирусные или невирусные) можно использовать для трансдукции или трансформации клеток, включая, но без ограничения, недифференцированные клетки, дифференцированные клетки, соматические клетки, сперматогонии и/или стволовые клетки. В некоторых вариантах осуществления стволовые клетки предназначены для введения человеку, а не для имплантации подходящей псевдобеременной женщине для дифференцировки и развития грудного ребенка.
В некоторых вариантах осуществления, вирусный вектор по изобретению содержит фактор ускорения распада (DAF). Например, оболочечный вирусный вектор содержит DAF на вирусной мембране. В некоторых вариантах осуществления DAF представляет собой DAF дикого типа. В некоторых вариантах осуществления DAF является частью белка, слитого с белком оболочки, например, см. Guibinga et al. Mol Ther. 2005 1 1(4):645-51. В некоторых вариантах осуществления клетка-продуцент BIV экспрессирует DAF.
Получение аналогов фактора комплемента B по изобретению
Аналоги белка фактора комплемента B по изобретению можно продуцировать в клетке. В некоторых вариантах осуществления аналог белка фактора комплемента B затем очищают из клетки и/или из культуральной среды.
Изобретение относится к (i) клеткам, содержащим нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую аналог фактора комплемента B по изобретению, и/или к (ii) клеткам, экспрессирующим аналог белка фактора комплемента B по изобретению. В некоторых вариантах осуществления используют клетки млекопитающих. В некоторых вариантах осуществления используют прокариотическую клетку.
Клетки-хозяева, как правило, трансфицируют или трансдуцируют экспрессирующим или клонирующим вектором для продукции белка и культивируют в общепринятой питательной среде, модифицированной подходящим образом для индукции промоторов, отбора трансформантов, амплификации генов, кодирующих желательные последовательности, или для нижестоящих способов очистки и/или концентрации.
Пригодные клетки-хозяева для клонирования или экспрессии кодирующей области в векторе представляют собой клетки прокариот, дрожжей или высших эукариот. Пригодные прокариоты для этой цели включают в себя, но без ограничения, эубактерии, такие как грамотрицательные или грамположительные организмы, например, энтеробактерии, такие как Escherichia, например, E. coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например, Salmonella typhimurium, Serratia, например, Serratia marcescans, и Shigella, так же как бациллы, такие как B. subtilis и B. licheniformis (например, B. licheniformis 41P), Pseudomonas, такие как P. aeruginosa, и Streptomyces. В некоторых вариантах осуществления клонирующий хозяин E. coli представляет собой E. coli 294 (например, ATCC 31446), хотя другие штаммы, такие как E. coli B, E. coli X1776 (например, ATCC 31537) и E. coli W3110 (например, ATCC 27325), могут являться пригодными.
В дополнение к прокариотам, эукариотические микроорганизмы, такие как мицелиальные грибы или дрожжи, являются пригодными хозяевами для клонирования или экспрессии. Saccharomyces cerevisiae, или пекарские дрожжи, являются общепринятыми среди микроорганизмов-хозяев - низших эукариот. Однако, ряд других родов, видов и штаммов являются общепринятыми и пригодными по настоящему изобретению, такие как Schizosaccharomyces pombe; хозяева Kluyveromyces, например, такие как, K. lactis, K. fragilis (например, ATCC 12424), K. bulgaricus (например, ATCC 16045), K. wickeramii (например, ATCC 24178), K. waltii (например, ATCC 56500), K. drosophilarum (например, ATCC 36906), K. thermotolerans и K. marxiamis; yarrowia (например, EP402226); Pichia pastoris (например, EP 183070); Candida; Trichoderma reesia (например, EP 244234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и мицелиальные грибы, например, такие как Neurospora, Penicillium, Tolypocladium, и хозяева Aspergillus, такие как A. nidulans и A. niger.
Пригодные клетки-хозяева, например, для экспрессии гликозилированных белков, можно получать из многоклеточных организмов. Примеры клеток беспозвоночных включают в себя клетки растений и клетки насекомых. Многочисленные бакуловирусные штаммы и варианты и соответствующие пермиссивные клетки-хозяева из таких хозяев, как Spodoptera frugiperda (листовертка), Aedes aegypti (комар), Aedes albopictus (комар), Drosophila melanogaster (плодовая мушка) и Bombyx mori, идентифицированы и могут быть использованы для экспрессии белков. Множество вирусных штаммов для трансфекции можно использовать для экспрессии белка, и они являются общедоступными, например, вариант L-1 NPV Autographa californica и штамм Bm-5 NPV Bombyx mori, и такие вирусы можно использовать в соответствии с настоящим изобретением, например, для трансфекции клеток Spodoptera frugiperda. Культуры клеток растений хлопчатника, кукурузы, картофеля, сои, петунии, томата и табака также можно использовать в качестве хозяев.
В некоторых вариантах осуществления изобретения используют клетки позвоночных или млекопитающих, и размножение клеток позвоночных в культуре (культуре ткани) может представлять собой общепринятый способ.
Примеры пригодных линий клеток-хозяев млекопитающих представляют собой линию клеток почки обезьяны CVI, трансформированную SV40 (например, COS-7, ATCC CRL 1651); линию эмбриональной почки человека (например, клетки 293 или 293T, включая любую линию клеток, субклонированных для выращивания в суспензионной культуре, Graham et al, J. Gen Virol. 36:59 (1977) such as 293 Freestyle (Invitrogen, Carlsbad, CA)) или 293FT; клетки почки детеныша хомяка (например, BHK, ATCC CCL 10); клетки яичника китайского хомяка; клетки яичника китайского хомяка/-DHFR (например, CHO, Urlaub et al, Proc. Natl. Acad. Sci. USA 77:4216 (1980)); клетки Сертоли мыши (например, TM4, Mather, Biol. Reprod. 23:243-251 (1980)); клетки почки обезьяны (например, CVI ATCC CCL 70); клетки почки африканской зеленой мартышки (например, VERO-76, ATCC CRL-1587); клетки цервикальной карциномы человека (например, HELA, ATCC CCL 2); клетки почки собаки (например, MOCK, ATCC CCL 34); клетки CF2TH; клетки печени крысы буффало (например, BRL 3A, ATCC CRL 1442); клетки легкого человека (например, W138, ATCC CCL 75); клетки печени человека (например, Hep G2, HB 8065); клетки опухоли молочной железы мыши (например, MMT 060562, ATCC CCL51); клетки TRI (Mather et al, Annals N.Y. Acad. Sci. 383:44-68 (1983)); клетки MRC 5; и клетки FS4.
В некоторых случаях клетку-хозяина можно модифицировать для уменьшения или прекращения экспрессии эндогенного белка. Например, если аналог белка фактора комплемента B предназначен для продукции в конкретной клетке-хозяине (например, в клетке CHO), тогда клетку-хозяина можно модифицировать так, чтобы экспрессия природного белка фактора B клетки-хозяина (например, фактора B хомяка) снижена или прекращена. Это может являться преимущественным для нижестоящей очистки аналога белка фактора комплемента B. Таким образом, изобретение относится к способу получения аналога белка комплемента по изобретению, включающему в себя снижение или прекращение экспрессии соответствующего природного белка комплемента в клетке-хозяине. Способы снижения, прекращения или нокаута экспрессии белка клетки-хозяина известны в данной области. Например, уровень экспрессии белка можно снижать или прекращать посредством модификации клетки-хозяина для экспрессии ингибирующей РНК (например, РНКи), специфической для РНК, кодирующей белок. Например, Clontech (Mountain View, CA) продает различные векторы и наборы, такие как векторы и наборы, обозначаемые как часть систем KNOCKOUT™ RNAi, для нокдауна экспрессии белков в клетке-хозяине. Другие способы включают в себя нацеливание на гены посредством гомологичной рекомбинации, позволяющей введение конкретных мутаций в любой клонированный ген, например, см. Current Protocols in Molecular Biology, John Wiley & Sons, Inc., 1994-1998, Sections 9,16 и 9,17. Это можно использовать для нокаута гена для экспрессии белка клетки-хозяина.
Другой способ, который можно использовать для снижения экспрессии эндогенного белка, включает в себя использование направленного фактора транскрипции, репрессирующего экспрессию эндогенного белка. Например, домен репрессора из фактора транскрипции можно соединять или сливать с ДНК-связывающим доменом, таким как полипептид с цинковыми пальцами. Специалист в данной области может разрабатывать полипептиды с цинковыми пальцами, которые связывают специфические последовательности ДНК, например, см. Патенты США No. 6140081; и 7067617; и опубликованные Патентные заявки США 20060078880; 20040224385; и 20070213269. Специалист в данной области может связывать разработанные полипептиды с цинковыми пальцами с доменом репрессора транскрипции (например, домен KRAB (Kriippel-ассоциированный бокс)). Примеры таких молекул и способов описаны в Beerli et al. (Proc Natl Acad Sci U S A. 2000 97(4): 1495-1500) и опубликованной Патентной заявке США 20070020627. В некоторых вариантах осуществления изобретения, клетку-хозяина можно трансдуцировать вектором, экспрессирующим репрессор транскрипции. Этот способ обладает преимуществом над нокаутом интересующего гена с использованием гомологичной рекомбинации, поскольку, в большинстве случаев, клетка-хозяин является диплоидной и может быть желательно провести нокаут обеих копий гена. В то же время, экспрессия репрессора транскрипции должна репрессировать экспрессию обеих копий гена.
Экспрессию конкретного эндогенного белка можно также снижать с использованием соединений, которые могут напрямую или опосредованно снижать экспрессию конкретного эндогенного белка. С использованием fB в качестве примера, различные соединения можно использовать для снижения экспрессии эндогенного fB. Например, показано, что экспрессию белка fB ингибирует гистамин (Falus & Meretey, Immunology 1987 60:547-551 и Falus & Meretey, Mol Immunol 1988 25(11): 1093-97), бутират натрия (Andoh et al. Clin Exp Immuno 1999 118:23-29), глюкокортикоид, такой как дексаметазон (Dauchel et al Eur J Immunol 1990 20(8): 1669-75), тромбоцитарный фактор роста (Circolo et al 1990 The Journal of Biol Chem 265(9):5066-5071), эпидермальный фактор роста (Circolo et al. 1990), и фактор роста фибробластов (Circolo et al. 1990). Клетку-хозяина по изобретению можно культивировать в присутствии любой или комбинации из этих молекул для снижения эндогенной экспрессии белка фактора комплемента B. Таким образом, в некоторых вариантах осуществления изобретения, клетку-хозяина, экспрессирующую аналог белка фактора комплемента B, культивируют в присутствии любого одного или нескольких соединений, выбранных из группы, состоящей из гистамина, бутирата натрия, глюкокортикоида (например, дексаметазона), тромбоцитарного фактора роста, эпидермального фактора роста или фактора роста фибробластов.
Показано, что различные соединения и белки осуществляют повышающую регуляцию или поддерживают экспрессию белка фактора комплемента B. Например, показано, что осуществляют повышающую регуляцию или поддерживают экспрессию белка фактора комплемента B фактор некроза опухоли (TNF) (Andoh et al. Clin Exp Immuno 1999 118:23-29), эстроген (Sheng-Hsiang et al. Biology of Reproduction 2002 66:322-332), интерлейкин-1 (Dauchel et al. Eur J Immunol 1990 20(8): 1669-75), дексаметазон (Lappin & Whaley, Biochem J 1991 280: 117-123), преднизолон (Lappin & Whaley 1991), cortical (Lappin & Whaley 1991) и интерферон-гамма (Huang et al. 2001 Eur J Immunol 31:3676-3686). Клетку-хозяина по изобретению можно культивировать в отсутствие любой или комбинации из этих молекул для снижения эндогенной экспрессии белка фактора комплемента B. Кроме того, клетку-хозяина можно культивировать в присутствии ингибитора любого одного или нескольких из этих соединений. Таким образом, в некоторых вариантах осуществления изобретения, клетку-хозяина, экспрессирующую аналог белка фактора комплемента B, культивируют в присутствии любого одного или нескольких соединений, ингибирующих соединение, выбранное из группы, состоящей из TNF, эстрогена, интерлейкина-1, дексаметазона, преднизолона, cortical и интерферона-гамма. В некоторых вариантах осуществления экспрессию клеткой-хозяином одного или нескольких из этих соединений снижают, например, с использованием способов, как описано в настоящем документе. Ингибиторы эстрогена включают в себя, в качестве неограничивающих примеров, тамоксифен. Ингибиторы включают в себя также антитела, которые связывают соединение и снижают его активность. Например, различные антитела, которые связывают и инактивируют TNF, известны в данной области.
Композицию, содержащую аналог белка фактора комплемента B, полученный из клеток, можно очищать с использованием, например, хроматографии на гидроксилапатите, электрофореза в геле, диализа, эксклюзионной хроматографии, аффинной хроматографии, иммуноаффинной хроматографии, очистки в тангенциальном потоке, диафильтрации, ионообменной хроматографии, хроматографии гидрофобного взаимодействия (HIC), обращеннофазовой хроматографии, аффинной хроматографии на гепаринсефарозе и других известных форм разделения и концентрации. После любой стадии(стадий) предварительной очистки, смесь, содержащую аналог белка фактора комплемента B и загрязнения, если присутствуют, можно подвергать хроматографии гидрофобного взаимодействия при низком pH, например, с использованием буфера для элюции при pH приблизительно 2,5-4,5, в некоторых случаях проводимой при низких концентрациях соли (например, от приблизительно 0-0,25 М соли), или другим способам для дополнительной очистки.
В некоторых вариантах осуществления аналог белка фактора комплемента B по изобретению является по меньшей мере на 90%, по меньшей мере на 93%, по меньшей мере на 95%, по меньшей мере на 98%, по меньшей мере на 99,5% или по меньшей мере на 99,9% чистым по отношению к общему белку.
Изобретение относится также к способам получения аналога белка фактора комплемента B, включающим в себя экспрессию в клетке аналога белка фактора комплемента B по изобретению и очистки аналога белка фактора комплемента B.
Опосредованные комплементом состояния/заболевания
Существуют три пути активации комплемента, классический путь, альтернативный путь и лектиновый путь (фигура 1). В настоящем документе описаны примеры аналогов белка фактора комплемента B по изобретению. В некоторых вариантах осуществления эти аналоги белка фактора комплемента B могут ослаблять альтернативный путь активации комплемента. Однако, основываясь на области, где все три пути комплемента пересекаются, эти аналоги могут уменьшать воспаление, вызванное любым из трех путей комплемента, и таким образом обеспечивать лечение для любого заболевания, в этиологию которого вовлечена, по меньшей мере частично, активация комплемента. Они включают в себя, но без ограничения, раннюю AMD, влажную AMD, и географическую атрофию. На фигурах 1A и 1B изображены пути комплемента. Следует отметить, что они пересекаются на C3b.
Изобретение относится к способам лечения опосредованного комплементом заболевания, включающим в себя введение пациенту фармацевтического препарата по изобретению, аналога фактора комплемента B по изобретению или нуклеиновой кислоты или вектора, кодирующего аналог фактора комплемента B по изобретению. Изобретение относится также к способам ингибирования активности комплемента, где способ включает в себя введение субъекту-человеку мутированного аналога фактора комплемента B человека в количестве, достаточном для ингибирования пути комплемента посредством конкуренции со связыванием природного фактора комплемента B у субъекта, где мутированный аналог фактора комплемента B человека представляет собой активный аналог фактора комплемента B из SEQ ID NO:4, обладающий цистеиновыми аминокислотами, формирующими дисульфидные связи, и свободной цистеиновой аминокислотой, замененной на аминокислоту, выбранную из группы, состоящей из гистидина, изолейцина, лейцина, метионина, фенилаланина, серина, треонина, тирозина и валина. В некоторых вариантах осуществления мутированный аналог фактора комплемента B содержит SEQ ID NO:2, 3, 22 или 23.
Изобретение относится также к способам ингибирования активности комплемента, где способы включают в себя введение в участок активности комплемента аналога фактора комплемента B по изобретению нуклеиновой кислоты по изобретению, вирусного вектора по изобретению, фармацевтической композиции/препарата по изобретению или любой их комбинации в количестве, достаточном для ингибирования активности комплемента. В некоторых вариантах осуществления в способе по изобретению используют фактор комплемента B, представляющий собой аналог SEQ ID NO:4, обладающий цистеиновыми аминокислотами, формирующими дисульфидные связи, и свободной цистеиновой аминокислотой, замененной на другую аминокислоту.
Изобретение относится к способам ингибирования активности комплемента, где способы включают в себя введение в участок активности комплемента мутантного аналога фактора комплемента B человека в количестве, достаточном для ингибирования активности комплемента мутантным аналогом фактора комплемента B человека, конкурирующим со связыванием природного фактора комплемента B, где мутантный аналог фактора комплемента B человека представляет собой активный аналог фактора комплемента B из SEQ ID NO:4, обладающий цистеиновыми аминокислотами, формирующими дисульфидные связи, и свободной цистеиновой аминокислотой, замененной на какую-либо аминокислоту. В некоторых вариантах осуществления в этом способе используют аналог фактора комплемента B, содержащий аминокислоты 26-764 из SEQ ID NO:2, аминокислоты 26-764 из SEQ ID NO:3, аминокислоты 26-990 из SEQ ID NO:22 или аминокислоты 26-990 из SEQ ID NO:23.
Пути комплемента являются частью иммунной системы, известной как врожденная иммунная система, обеспечивающая немедленную защиту от инфекции до активации гуморальной и опосредованной клетками ветвей иммунной системы. Они активируются и инактивируются посредством каскадных реакций, которые обладают кинетикой высокого порядка, но поразительно хорошей регуляцией. Альтернативный путь комплемента, в частности, развивается циклично с высокой скоростью посредством петли положительной обратной связи.
Аналоги белка фактора комплемента B по изобретению и/или экспрессирующие их векторы можно преимущественно использовать для местного и/или системного введения млекопитающему и/или для лечения хронических заболеваний. В некоторых вариантах осуществления изобретения аналог фактора комплемента B ингибирует путь комплемента посредством конкуренции со связыванием природного фактора комплемента B, например, с белком C3b и/или белком фактором D. Это может позволять ослабление активности комплемента в отличие от полного блокирования пути. Таким образом, возможна понижающая регуляция активности комплемента до уровня, который является терапевтическим (например, облегчает некоторые симптомы или их тяжесть) без полного блокирования активности комплемента. Таким образом уничтожают или уменьшают риски, ассоциированные с блокированием активности комплемента, такие как увеличенный риск инфекции. Таким образом, настоящее изобретение относится к способам лечения опосредованного комплементом заболевания (например, хронического заболевания) посредством местного или системного введения (например, i.v., внутрибрюшинного или перорального) аналога белка фактора комплемента B по изобретению.
В некоторых вариантах осуществления настоящее изобретение относится к композициям и способам для модуляции, регуляции, ингибирования и/или усиления активности комплемента. Относящиеся к комплементу пути включают в себя, но без ограничения, классический, лектиновый и альтернативный пути комплемента. В некоторых случаях, относящийся к комплементу путь может играть роль в конкретном состоянии, заболевании или заболеваниях. Таким образом, некоторые варианты осуществления изобретения относятся к способам регуляции, модификации, излечения, ингибирования, предотвращения, облегчения, замедления прогрессирования и/или лечение стадии заболевания, опосредованного одним или несколькими относящимися к комплементу путями, посредством введения аналога фактора комплемента B по изобретению или нуклеиновой кислоты или вектора, кодирующего аналог фактора комплемента B по изобретению. Такие стадии или состояния заболевания включают в себя, но без ограничения, образование друз, дегенерацию желтого пятна, AMD, сухость глаз, язвы роговицы, атеросклероз, диабетическую ретинопатию, витреоретинопатию (Grisanti ei al Invest. Ophthalmol. Vis. Sci. 32:271 1-2717), воспаление роговицы, гиперчувствительность дыхательных путей, связанные с иммунитетом заболевания, связанные с аутоиммунитетом заболевания, волчаночный нефрит, системную красную волчанку (SLE), артрит (например, ревматоидный артрит), ревматологические заболевания, антифосфолипидный синдром, I/R повреждение кишечника и почек, астму, атипичный гемолитико-уремический синдром, мембранопролиферативный гломерулонефрит типа II, непролиферативный гломерулонефрит, выкидыш (например, спонтанный выкидыш), глаукому, увеит, глазную гипертензию, повреждение головного мозга (например, травматическое повреждение головного мозга), инсульт (например, см. Arumugam et al. PNAS 93(12):5872-6 (1996)), посттравматическое повреждение органов, температурную травму (например, ожог), постинфарктное повреждение органов (например, кардиологическое, неврологическое), васкулит, болезнь Кавасаки, наследственный ангионевротический отек (HAE), пароксизмальную ночную гемоглобинурию (PNH, иногда обозначаемая как синдром Маркиафавы-Микели), колиты, воспалительное заболевание кишечника, метастазирование опухолей, ишемически-реперфузионное повреждение, острое нарушение мозгового кровообращения, болезнь Альцгеймера, отторжение трансплантата (например, ксено и алло), инфекции, сепсис, септический шок, синдром Шегрена, миастению, опосредованные антителами заболевания кожи, все опосредованные антителами органоспецифические заболевания (включая сахарный диабет типа I и типа II, тиреоидит, идиопатическую тромбоцитопеническую пурпуру и гемолитическую анемию и невропатии), синдром устойчивости к инсулину (например, см. Weyer et al. (2000) Diabetes Care, 23(6):779-785), гестационный диабет, рассеянный склероз, псориаз, повреждение при искусственном кровообращении, узелковый полиартериит, пурпуру Шенлейна-Геноха, сывороточную реакцию, заболевание Гудпасчера, системный некротизирующий васкулит, постстрептококковый гломерулонефрит, идиопатический пульмонарный фиброз (обычный интерстициальный пневмонит), мембранный гломерулонефрит, миокардит (например, аутоиммунный миокардит) (Kaya et al. Nat Immunol. 2001 ;2(8):739-45), инфаркт миокарда, мышечную дистрофию (например, ассоциированную с недостаточностью дистрофина), синдром острого инфаркта легкого, синдром острой дыхательной недостаточности у взрослых, реперфузию и/или опосредованное комплементом заболевание.
В некоторых вариантах осуществления опосредованное комплементом заболевание представляет собой заболевание глаза. В некоторых вариантах осуществления аналог фактора комплемента B или фармацевтическую композицию вводят в глаз, например, посредством интравитреальной инъекции, субретинальной инъекции, инъекции в переднюю камеру глаза, инъекции или местного введения в роговицу, субконъюнктивальной инъекции, субтеноновой инъекции или посредством глазных капель. В некоторых вариантах осуществления фармацевтическую композицию вводят в глаз, где фармацевтическая композиция содержит по меньшей мере один аналог фактора комплемента B, например, выбранный из группы hfB3-292S (SEQ ID NO:2), hfB3-292S-740N (SEQ ID NO:3), hfB3-292S-Fc (SEQ ID NO:22) и hfB3-292S-740N-Fc (SEQ ID NO:23).
Связанная с возрастом дегенерация желтого пятна (AMD) является наиболее распространенной причинной снижения зрения у индивидуумов в возрасте старше 65 в развитых странах. Сухая AMD характеризуется прогрессирующей дегенерацией желтого пятна, вызывая потерю центрального поля зрения. Более острая инвалидизирующая AMD включает в себя флоридную неоваскуляризацию и просачивание в сетчатку, известные как влажная AMD. В настоящее время не существует эффективного лечения AMD.
Характерным для AMD является накопление друз, локализованных между базальной мембраной пигментного эпителия сетчатки (RPE) и внутренним слоем мембраны Бруха (Pauleikhoff et al, 1990 Am. J. Ophthalmol. 109, 38-43; Bressler et al, 1990 Arch. Ophthalmol. 108, 1442-1447). Считают, что друзы, так же как другие связанные с возрастом изменения, происходящие поблизости от мембраны Бруха, вносят вклад в дисфункцию и дегенерацию RPE и сетчатки, посредством индукции ишемии, так же как ограничения обмена питательных веществ и продуктов жизнедеятельности между сетчаткой и хориоидом (обзор приведен в Bird, 1992 Pathophysiology of AMD. In Age-Related Macular Degeneration: Principles and Practice (Hampton, G., and Nelsen, P.T., eds.) Chap. 3, Raven Press, New York). В нескольких исследованиях показаны опосредованные иммунитетом процессы в развитии AMD. Важно, что аутоантитела детектировали в сыворотке пациентов с AMD (Penfold et al, 1990 Graefes Arch. Clin. Exp. Ophthalmol. 228, 270-274), как предсказано гипотезой, что опосредованные иммунитетом и воспалением процессы вовлечены в развитие и/или удаление друз.
Формирование друз в глазу может быть ассоциировано с различными заболеваниями, такими как дегенерация желтого пятна. В некоторых случаях подразумевают, что формирование друз и/или его ассоциация с заболеванием связана с активностью комплемента. Некоторые варианты осуществления по изобретению относятся к композициям и способам модуляции, регуляции, ингибирования, снижения, задержки и/или обращения формирования или роста друз у животного, такого как человек. Например, композиции или молекулы по изобретению можно доставлять к друзам (например, прямой инъекцией в друзы (интрадрузальной инъекцией), поблизости к друзам или интравитреальной инъекцией). Некоторые варианты осуществления изобретения можно использовать для замедления прогрессирования дегенерации желтого пятна, возможно, посредством ингибирования формирования друз. Витронектин, наиболее распространенный компонент друз, является также компонентом внеклеточных отложений, ассоциированных с атеросклерозом (Niculescu et al., 1989 Atherosclerosis, 78, 197-203), амилоидозом (Dahlback et al., 1993 J. Invest. Dermatol. 100, 166- 170), эластозом (Dahlback et al., 1988 Acta Derm. Venereol. 68, 107- 1 15), и MPGN типа II (Jansen et al, 1993 Am. J. Pathol. 143, 1356-1365). Витронектин представляет собой многофункциональный белок, функционирующий в адгезии клеток, поддержании гемостаза и ингибирования индуцированного комплементом лизиса клеток (Preissner, 1991 Ann. Rev. Cell Biol. 7, 275-310). Более того, атеросклеротические бляшки разделяют с друзами ряд других составляющих, таких как компоненты комплемента и аполипопротеин E. Ассоциация между AMD на поздних стадиях и атеросклерозом сонных артерий опубликована в эпидемиологическом исследовании (Vingerling et al., 1995 Am. J. Epidemiol. 142, 404-409), и в другом исследовании идентифицирована значимая корреляция между эластотической дегенерацией не подверженной воздействию солнечных лучей дермы и хориоидальной неоваскуляризацией у пациентов AMD (Blumenkranz et al., 1986 Ophthalmology, 93, 552-558). Наконец, пептид β-амилоида, главное составляющее нейритических бляшек при болезни Альцгеймера, также обнаружен в друзах (Johnson et al, 2002 Proc. Natl. Acad. Sci. USA, 99, 11830-11835). Пептид β-амилоида вовлечен в качестве первичного активатора комплемента (Bradt et al, 1998 J. Exp. Med. 188, 431-438).
Сравнительный анализ молекулярного состава друз человека, так же как клеток RPE, фланкирующих или покрывающих друзы, показал иммунореактивность по отношению к иммуноглобулинам и компонентам системы комплемента, ассоциированным с накоплением иммунных комплексов (Johnson et al, 2000 Exp. Eye Res. 70, 441-449). Друзы содержат также многофункциональные белки, такие как витронектин (Hageman et al, 1999 FASEB J. 13, 477-484) и аполипопротеин E (Anderson et al, 2001 Am. J. Ophthalmol. 131, 767-781), играющие роль в модуляции иммунной системы. Кроме того, молекулы, вовлеченные в ответ острой фазы на воспаление, такие как компонент амилоида P и α1-антитрипсин, также идентифицированы в друзах (Mullins et al, 2000 The FASEB Journal, 14, 835-846), так же как белки, вовлеченные в свертывание и фибринолиз (фактор X, тромбин и фибриноген) (Mullins et al, 2000 The FASEB Journal, 14, 835-846). Предполагают, что формирование друз и ассоциированная патология RPE вносят вклад в хронический воспалительный ответ, активирующий каскад реакций комплемента (Hageman et al, 2001 Prog. Retin, Eye Res. 20, 705-732; Johnson et al, 2001 Exp. Eye Res. 73, 887-896).
Одной из других форм нарушения зрения, возникающего из-за AMD и приводящего к нарушениям сетчатки, является географическая атрофия, приводящая к гибели путей палочковых и колбочковых клеток, так же как клеток RPE.
Показано, что атеросклероз, как правило, вовлекает связанные с комплементом пути, например, см. Niculescu et al Immunologic Research, 3Q(l):73-80(8) (2004) и Niculescu и Horea, Immunologic Research 30(l):73-80 (2004). Активация комплемента и накопление C5b-9, как правило, происходит как при атеросклерозе человека, так и при экспериментальном атеросклерозе. C5b-9 могут отвечать за лизис клеток, и сублитическая сборка C5b-9 индуцирует активацию и пролиферацию гладкомышечных клеток (SMC) и эндотелиальных клеток (EC). Недостаточность C6 комплемента оказывает защитный эффект на индуцированный диетой атеросклероз, позволяя предполагать, что сборка C5b-9 необходима, или по меньшей мере играет значительную роль, в прогрессировании атеросклеротических бляшек, например, см. Niculescu and Horea, Immunologic Research 30(l):73-80 (2004). Некоторые варианты осуществления изобретения можно использовать для ингибирования формирования C5b-9 и/или ингибирования атеросклероза. В некоторых вариантах осуществления аналог белка фактора комплемента B по изобретению вводят в участок или потенциальный участок атеросклероза. Этот аналог белка фактора комплемента B ингибирует путь (например, классический и/или альтернативный путь комплемента), который, в свою очередь, ингибирует формирование или активацию C5b-9 или другого относящегося к пути комплемента соединения, вовлеченного в атеросклероз. Могут существовать другие относящиеся к комплементу белки, вовлеченные в атеросклероз, образование и/или активацию которых можно ингибировать или блокировать сходным образом.
Гиперчувствительность дыхательных путей (AHR) является характерной для различных заболеваний, включая, но без ограничения, астму (например, аллергическая астма). Показано, что AHR, как правило, включает в себя относящиеся к комплементу пути, например, см. Taube et al, 2006 PNAS 103(21):8084-8089; Park et al, American Journal of Respiratory и Critical Care Medicine 169:726-732, (2004); Thurman and Holers, J Immunology 176: 1305-1310 (2006) и Публикация патента США No. 20050260198. Park et al. показали, что Crry-Ig, вводимый внутрибрюшинной инъекцией, оказывает эффект на AHR. Некоторые варианты осуществления изобретения относятся к композициям и способам модуляции, регуляции, ингибирования и/или уменьшения AHR у животного, такого как человек. Конкретные родственные AHR заболевания, которые можно лечить, ингибировать и/или облегчать, включают в себя, но без ограничения, астму, хроническое обструктивное заболевание легких (COPD), аллергический бронхолегочный аспергиллез, аллергическую пневмонию, эозинофильную пневмонию, эмфизему, бронхит, аллергический бронхит, бронхоэктаз, кистозный фиброз, туберкулез, пневмонит гиперчувствительности, профессиональную астму, саркоид, реактивный синдром дисфункции дыхательных путей, интерстициальную легочную болезнь, гиперэозинофильный синдром, ринит, синусит, индуцированную физическим усилием астму, индуцированную загрязнениями астму, кашлевую астму, вызванное паразитами заболевание легких, инфекцию респираторно-синцитиальным вирусом (RSV), инфекцию вирусом парагриппа (PIV), инфекцию риновирусом (RV), инфекцию вирусом Хантаана (например, штамм four-comers) и аденовирусную инфекцию. Показано, что связанные с иммунитетом заболевания, такие как связанные с аутоиммунитетом заболевания, ассоциированные с HLA-B27 воспалительные заболевания, волчаночный нефрит и системная красная волчанка (SLE), как правило, вовлекают относящиеся к комплементу пути, например, см. Thurman and Holers, J Immunology 176:1305-1310 (2006). Волчаночный нефрит является одним из осложнений SLE. Он относится к аутоиммунному процессу при волчанке, когда иммунная система продуцирует антитела (антиядерное антитело и другие) против компонентов организма. Комплексы этих антител и компонентов комплемента, как правило, накапливаются в почках и приводят к воспалительному ответу. Некоторые варианты осуществления изобретения относятся к способам и композициям для регуляции, модификации, излечения, ингибирования, предотвращения, облегчения и/или лечения связанного с иммунитетом заболевания, например, вовлекающего путь комплемента или относящегося к пути комплемента, такого как SLE.
Показано, что артрит, как правило, вовлекает связанные с комплементом пути, например, см. Thurman and Holers, J Immunology 176: 1305-1310 (2006) и Banda et al. J Immunol. 177(3): 1904-12 (2006). Альтернативный путь комплемента играет значительную роль в индукции артрита, и альтернативный путь комплемента может даже являться необходимым. Некоторые варианты осуществления изобретения относятся к способам и композициям для регуляции, модификации, излечения, ингибирования, предотвращения, облегчения и/или лечения артрита, например, ревматоидного артрита или воспалительного артрита.
Пароксизмальная ночная гемоглобинурия (PNH) представляет собой потенциально опасное для жизни заболевание крови, характеризующееся индуцированными комплементом внутрисосудистой гемолитической анемии и тромбоза из-за внутрисосудистого разрушения эритроцитов (RBC) комплементом, возникающего в результате неконтролируемого усиления системы комплемента, приводящего к разрушению мембраны RBC. Лица с этим заболеванием, как правило, имеют клетки крови, в которых отсутствует ген, называемый PIG-A. Этот ген позволяет веществу, называемому гликозилфосфатидилинозитол (GPI), помогать конкретным белкам прилипать к клеткам. Без PIG-A регулирующие комплемент белки не могут присоединяться к поверхности клетки и защищать клетки от комплемента. Некоторые варианты осуществления изобретения относятся к способам и композициям для регуляции, модификации, излечения, ингибирования, предотвращения, облегчения и/или лечения пароксизмальной ночной гемоглобинурии.
Наследственный ангионевротический отек (HAE) представляет собой потенциально опасное для жизни генетическое состояние, как правило, вызванное недостаточностью ингибитора C1, белка системы комплемента. Симптомы включают в себя приступы отека (опухания) различных частей организма, включая руки, ноги, лицо и дыхательные пути. Наследственный ангионевротический отек (HAE) существует в трех формах, все из которых вызваны генетической мутацией, наследуемой в аутосомно доминантной форме. Типы I и II вызваны мутациями в гене SERPING1 gene, приводящими либо к сниженным уровням, либо к дисфункциональным формам белка C1-ингибитора (HAE типа I). HAE типа III связана с мутациями в гене F12, кодирующем белок свертывания фактор XII. Все формы HAE приводят к аномальной активации системы комплемента. Некоторые современные лекарственные средства включают в себя экаллантид, пептидный ингибитор калликреина (например, см. Публикацию патента США No. US20070213275), Icatibant (Firazyr, Jerini), представляющий собой избирательный антагонист рецептора брадикинина, и цинриз (Viropharma, Inc.), ингибитор C1-эстеразы. Некоторые варианты осуществления изобретения относятся к способам и композициям для регуляции, модификации, излечения, ингибирования, предотвращения, облегчения и/или лечения пароксизмальной ночной гемоглобинурии.
Глаукома представляет собой группу заболеваний зрительного нерва, вовлекающих потерю клеток ганглия сетчатки с характерной картиной зрительной невропатии. Приблизительно 25% пациентов с глаукомой с потерей клеток ганглия сетчатки имеют нормальное внутриглазное давление. Внутриглазная гипертензия (OHT) является важным фактором риска для развития глаукомы, и ее снижение посредством лекарственных средств или хирургии в настоящее время является основным направлением лечения глаукомы. Показано, что внутриглазная гипертензия и глаукома, как правило, вовлекают связанные с комплементом пути, например, см. Khalyfa et al, Molecular Vision, 13:293-308 (2007); Stasi et al. IOVS 47(3): 1024-1029 (2007); и Kuehn et al, Experimental Eye Research 83:620-628 (2006). Показано, что экспрессия и/или присутствие C1q и C3 являются более высокими в сетчатке, подвергавшейся OHT. Некоторые варианты осуществления изобретения относятся к способам и композициям для регуляции, модификации, излечения, ингибирования, предотвращения, облегчения и/или лечения глаукомы.
Показано, что увеит, как правило, является ассоциированным с путем комплемента, например, см. Mondino and Rao, Investigative Ophthalmology & Visual Science 24:380-384 (1983) и Jha et al. Molecular Immunology 44:3901-3908 (2007). Mondino и Rao обнаружили, что средние значения для всех измерений тестированных компонентов комплемента во внутриглазной жидкости по отношению к измерениям в сыворотке были увеличены у пациентов с предшествующими операциями на глазах в анамнезе и являлись наиболее высокими у пациентов с передним увеитом. Некоторые варианты осуществления изобретения относятся к способам и композициям для регуляции, модификации, излечения, ингибирования, предотвращения, облегчения и/или лечения увеита.
Диабетическая ретинопатия является одной из основных причин потери зрения у индивидуумов среднего возраста. Считают, что активация системы комплемента играет важную роль в патогенезе диабетической ретинопатии (например, см. Jha et al. Molecular Immunology 44:3901-3908 (2007)). Некоторые варианты осуществления изобретения относятся к способам и композициям для регуляции, модификации, излечения, ингибирования, предотвращения, облегчения и/или лечения диабетической ретинопатии.
Пролиферативная витреоретинопатия (PV) является одним из самых распространенных осложнений отслоения сетчатки. PV связана с активностью комплемента, например, см. Grisante et al. Invest Ophthalmol Vis Sci. 1991;32(10):271 1-7 и Grisante et al. Ophthalmologe. 1992; 89 (1): 50-4. Некоторые варианты осуществления изобретения относятся к способам и композициям для регуляции, модификации, излечения, ингибирования, предотвращения, облегчения и/или лечения PV.
Показано, что антифосфолипидный синдром, повреждение кишечника и почек при ишемии/реперфузии I/R, атипичный гемолитико-уремический синдром, мембранопролиферативный гломерулонефрит типа II и выкидыш (например, спонтанный выкидыш), как правило, вовлекают относящиеся к комплементу пути, например, см. Ihurman and Holers, J Immunology 176: 1305- 1310 (2006).
Показано, что повреждение головного мозга (например, травматическое повреждение головного мозга), как правило, вовлекают относящиеся к комплементу пути, например, см. Leinhase et al., J Neuroinflammation 4: 13 (2007) и BMC Neurosci.7:55 (2006). В Leinhase 2006, показано, что после экспериментального травматического повреждения головного мозга у мышей дикого типа (fB+/+), присутствовала зависимая от времени системная активация комплемента. В отличие от этого, степень системной активации комплемента была значительно снижена у мышей fB-/-. Некоторые варианты осуществления изобретения относятся к способам и композициям для регуляции, модификации, излечения, ингибирования, предотвращения, облегчения и/или лечения гибели нейрональных клеток, травматического повреждения нервной системы (например, головного мозга), опосредованного комплементом нейровоспаления и/или нейропатологии.
Повреждение ишемии-реперфузии может вызывать увеличения продукции или окисления различных потенциально опасных соединений, продуцируемых клетками и тканями, которые могут приводить к окислительному повреждению или смерти клеток и тканей. Например, повреждение почек при ишемии-реперфузии может приводить к гистологическому повреждению почек, включая тубулярное повреждение почек, и изменяет характеристики острого тубулярного некроза. Результирующая дисфункция почек позволяет накопление азотсодержащих отходов, обычно выделяемых почками, таких как азот мочевины сыворотки (SUN). Ишемия-реперфузия может также вызывать повреждение удаленных органов, таких как легкое. В некоторых вариантах осуществления изобретения используют модуляторы, такие как ингибиторы, пути комплемента (например, ингибиторы активности фактора B), например, при введении животному, имеющему ишемию-реперфузию, или подверженного риску испытания или развития ишемии-реперфузии. В некоторых вариантах осуществления эти модуляторы предотвращают, снижают или ингибируют по меньшей мере один симптом повреждения из-за ишемии-реперфузии. Другие типы повреждения при ишемии-реперфузии, которые можно предотвращать или снижать с использованием способов и композиций по изобретению, включают в себя, но без ограничения, кардиологическое повреждение при ишемии-реперфузии, такое как инфаркт миокарда, или операцию коронарного шунтирования, повреждение центральной нервной системы при ишемии-реперфузии, повреждение конечностей или пальцев при ишемии-реперфузии, ишемию-реперфузию внутренних органов, таких как, печень или кишечник, или повреждение при ишемии-реперфузии какого-либо трансплантированного органа или ткани. См., например, Публикацию PCT No. WO03/061765, в которой обсуждают инфаркт миокарда и пути комплемента.
Воспаление является главным этиологическим фактором инфаркта миокарда (Ridker, 2007 Nutr. Rev. 65(12 Pt 2):S253-9). Показано также, что доставка (например, интракоронарная) (стволовых) клеток костного мозга приводит к улучшению систолической функции после острого инфаркта миокарда (Wollert, 2008, Curr. Opin. Pharmacol. Jan 31 [Epub]). А также, стволовые клетки костного мозга могут регенерировать миокард после инфаркта (Orlic et al. 2003 Pediatr. Transplant. 7 Suppl 3:86-88). Показано, что мезенхимальные стволовые клетки оказывают кардиологический защитный эффект при ишемической болезни сердца (Guo et al. 2007 Inflammation 30(3-4):97-104). По настоящему изобретению, доставку стволовых клеток можно осуществлять любыми способами, такими как интракоронарная инъекция, инъекция непосредственно в миокард (например, в пораженный заболеванием и/или здоровый миокард (например, рядом с пораженной областью)). В некоторых вариантах осуществления млекопитающее лечат цитокинами для мобилизации его стволовых клеток костного мозга в кровотоке, позволяющей стволовым клеткам мигрировать в участок инфаркта миокарда.
Различные стволовые клетки используют in vivo для различных применений. Одной из проблем с использованием стволовых клеток in vivo являются меньшие, чем желательно, выживаемость и/или оседание стволовых клеток, например, в интересующей области. Одной из важных причин низких оседания и выживаемости стволовых клеток может являться воспаление в участке. Таким образом, настоящее изобретение относится к способу лечения и/или к способу улучшения выживаемости и/или оседания стволовых клеток. В некоторых вариантах осуществления эти способы включают в себя введение композиции по изобретению до, во время и/или после введения или мобилизации стволовых клеток. В некоторых вариантах осуществления ингибиторы комплемента по изобретению действуют как противовоспалительные средства, которые могут создавать благоприятное окружение для стволовых клеток для хоминга и выживания в области желательного оседания (например, поврежденном сердце или костном мозге) и таким образом, репарации или замены поврежденной ткани. Стволовые клетки можно вводить в растворе, содержащем также аналог белка фактора комплемента B по изобретению. Стволовые клетки могут представлять собой, но без ограничения, гематопоэтические стволовые клетки, эмбриональные стволовые клетки, мезенхимальные стволовые клетки, нейрональные стволовые клетки, стволовые клетки молочной железы, обонятельные стволовые клетки, стволовые клетки островков Лангерганса, тотипотентные стволовые клетки, мультипотентые стволовые клетки или плюрипотентные стволовые клетки. Стволовые клетки могут являться аутологичными, аллогенными или сингенными.
Активность комплемента, по-видимому, вовлечена в мышечную дистрофию (например, ассоциированную с недостаточностью дистрофина). Например, см. Публикацию PCT No. WO2007130031, Spuler & Engel 1998 Neurology 50:41-46, и Selcen el al 2001 Neurology 56: 1472-1481.
Таким образом, некоторые варианты осуществления изобретения относятся к способам и композициям для регуляции, модификации, излечения, ингибирования, предотвращения, облегчения и/или лечения мышечной дистрофии.
Активность комплемента может вносить вклад в воспаление роговицы. Таким образом, некоторые варианты осуществления изобретения относятся к способам и композициям для регуляции, модификации, излечения, ингибирования, предотвращения, облегчения и/или лечения воспаления роговицы, например, после хирургической операции. В некоторых вариантах осуществления аналог фактора комплемента B по изобретению вводят посредством глазных капель или иным способом, описанным в настоящем документе.
В некоторых вариантах осуществления аналоги фактора комплемента B по изобретению используют для регуляции, модификации, излечения, ингибирования, предотвращения, облегчения и/или лечения неоваскуляризации роговицы.
Некоторые варианты осуществления изобретения относятся к способам усиления эффективности после трансплантации шунта или ангиопластики коронарной или периферической артерии. В некоторых вариантах осуществления вектор по изобретению, кодирующий аналог белка фактора комплемента B по изобретению, (например, hfB3-292S или hfB3-292S-740N) используют для трансдукции клеток кровеносного сосуда (например, эндотелиальных клеток). В некоторых вариантах осуществления, клетки кровеносного сосуда трансдуцируют до имплантации животному. В некоторых вариантах осуществления клетки кровеносного сосуда трансдуцируют in vivo.
Облегчение боли и страданий, и воспаления у пациентов после операции является областью особого внимания в клинической медицине, особенно с ростом количества амбулаторных операций, проводимых каждый год. Аналоги фактора комплемента B по настоящему изобретению можно использовать для ингибирования воспаления, например, посредством ингибирования активности комплемента. Таким образом, аналоги фактора комплемента B можно использовать для уменьшения воспаления, например, у пациентов после операции. В некоторых вариантах осуществления аналог фактора комплемента B вводят местно (например, периоперационная доставка) в участок хирургической операции для ингибирования воспаления, что в некоторых случаях уменьшает боль и страдания. В некоторых вариантах осуществления аналог фактора комплемента B вводят в растворе, например, в физиологически электролитном жидком носителе. В некоторых вариантах осуществления аналог фактора комплемента B доставляют посредством непосредственно в участок хирургической операции раствора для орошения, содержащего композицию. В некоторых вариантах осуществления, благодаря способу местной периоперационной доставки по настоящему изобретению, желательного терапевтического эффекта можно достигать с помощью более низких доз средств, чем необходимо при использовании других способов доставки, таких как внутривенный, внутримышечный, подкожный и пероральный. В некоторых вариантах осуществления, при периоперационном использовании, раствор может приводить к клинически значимому снижению боли и/или воспаления в участке операции, таким образом, позволяя снижать необходимость пациента в послеоперационном анальгетическом средстве (например, опиате) и, когда это целесообразно, позволяя более раннее использование пациентом участка после операции. В некоторых вариантах осуществления не требуется дополнительных усилий со стороны хирурга и персонала операционной для использования настоящего раствора по сравнению с общепринятыми жидкостями для орошения. В некоторых вариантах осуществления композицию по изобретению (например, жидкость для орошения) используют для артроскопии, сердечно-сосудистой и общей сосудистой терапии и диагностики, урологических процедур, общих послеоперационных ран и ран в целом. Композиции по изобретению можно доставлять посредством, но без ограничения, инъекции (например, посредством шприца), посредством орошающей жидкости, в качестве части повязки на рану или посредством местного введения, например, раствора, крема, геля или т.п.
В некоторых вариантах осуществления изобретения аналог фактора комплемента B и/или вектор по изобретению вводят в комбинации с ингибирующим комплемент фактором, до, во время или после введения аналога фактора комплемента B и/или вектора. Ингибирующий комплемент фактор включает в себя, но без ограничения, фактор H, подобный фактору H фактор 1, MCP, DAF или растворимую форму MCP.
В некоторых вариантах осуществления изобретения аналог фактора комплемента B или вектор по изобретению вводят в комбинации с антиангиогенным фактором. Антиангиогенные факторы включают в себя, но без ограничения, эндостатин, связывающую VEGF молекулу, PEDF, T2-TrpRS (например, см. Патент США No. 7273844), sFLT (например, см. Kong et al. Hum Gene Ther (1998) 9:823-833), афлиберцепт (ловушка VEGF), ловушка VEGF глазная, кининостатин, ранибизумаб и бевацизумаб.
В некоторых вариантах осуществления аналог фактора комплемента B и/или вектор по изобретению вводят в комбинации с ЛУЦЕНТИСОМ® (ранибизумабом), АВАСТИНОМ® (бевацизумабом), ловушкой VEGF глазной, афлиберцептом или молекулой(молекулами), которые связывают VEGF и/или ингибируют ангиогенез. ЛУЦЕНТИС® используют для лечения влажной AMD. Некоторые варианты осуществления изобретения также можно использовать для лечения влажной AMD. Таким образом, настоящее изобретение относится к способам и композициям для лечения влажной AMD, включающим в себя введение, вместе или по отдельности, композиции по изобретению в комбинации с ЛУЦЕНТИСОМ® (ранибизумаб), АВАСТИНОМ® (бевацизумабом), ловушкой VEGF глазной, афлиберцептом и/или молекулой(молекулами), которые связывают VEGF и/или ингибируют ангиогенез. Кроме того, внутриглазное воспаление является одной из наиболее распространенных неблагоприятных реакций, опубликованных после введения ЛУЦЕНТИСА®, например, см. «Полную инструкцию по применению лекарственного препарата» для ЛУЦЕНТИСА®. Настоящее изобретение относится к способу ингибирования или уменьшения внутриглазного воспаления (например, вызванного введением ЛУЦЕНТИСА®), включающему в себя введение молекулы или композиции по изобретению до, во время и/или после введения ЛУЦЕНТИСА®, ловушки VEGF глазной или афлиберцепта.
В некоторых вариантах осуществления изобретения аналог фактора комплемента B или вектор по изобретению вводят в комбинации с другим соединением(соединениями), таким как соединение, ингибирующее активацию T-клеток, B-клеток, TNF, интерлейкин-1 (например, интерлейкин-1b), интерлейкин-6 и/или интерферон-гамма. Аналог фактора комплемента B или вектор по изобретению можно вводить также в комбинации с соединением(соединениями), ингибирующими активность комплемента, например, активность альтернативного пути комплемента. Соединения, которые можно использовать, и которые ингибируют TNF, включают в себя, но без ограничения, соединения, связывающие TNF, такие как антитела (например, инфликсимаб (РЕМИКЕЙД®), голимумаб (СИМПОНИ®) и адалимумаб (ХУМИРА®)), или растворимые рецепторы, связывающие TNF, такие как этанерцепт (ЭНБРЕЛ®). Другие соединения, которые можно использовать, и которые ингибируют активацию T-клеток, включают в себя, но без ограничения, соединения, связывающие B7, такие как абатацепт (ОРЕНЦИЯ®). Можно использовать также соединения, осуществляющие понижающую регуляцию B-клеток, включая, но без ограничения, соединения, связывающие CD20, такие как ритуксимаб (РИТУКСАН® и МАБТЕРА®).
Пути комплемента, вносящие вклад в заболевание и/или вызывающие заболевание, можно модулировать, регулировать, ингибировать и/или активировать с использованием различных способов и/или аналогов белка фактора комплемента B, являющихся частью настоящего изобретения.
Композиции, составы и препараты
Некоторые варианты осуществления изобретения относятся к композициям, например, фармацевтическим композициям, содержащим аналог фактора комплемента B по изобретению, например, для терапевтических применений. В некоторых вариантах осуществления фармацевтическая композиция содержит аналог фактора комплемента B, содержащий аминокислоты 26-764 из SEQ ID NO:2, аминокислоты 26-764 из SEQ ID NO:3, аминокислоты 26-990 из SEQ ID NO:22 или аминокислоты 26-990 из SEQ ID NO:23, например, hfB3-292S (SEQ ID NO:2), hfB3-292S-740N (SEQ ID NO:3), hfB3-292S-Fc (SEQ ID NO:22) или hfB3-292S-740N-Fc (SEQ ID NO:23). В некоторых вариантах осуществления фармацевтическая композиция содержит аналог фактора комплемента B, состоящий из аминокислот 26-764 из SEQ ID NO:2, аминокислот 26-764 из SEQ ID NO:3, аминокислот 26-990 из SEQ ID NO:22 или аминокислот 26-990 из SEQ ID NO:23. Примеры фармацевтических композиций и составов, которые можно использовать с аналогами фактора комплемента B по изобретению, описаны в Публикации PCT No. WO08/106644 и Публикации патента США No. US20100120665.
Некоторые варианты осуществления по изобретению включают в себя фармацевтические препараты, содержащие аналог белка фактора комплемента B по изобретению, нуклеиновую кислоту по изобретению, вирусный вектор по изобретению или любую их комбинацию.
Составы (например, для инъекции) представляют собой в основном, но не обязательно, биосовместимые растворы активного ингредиента, например, содержащие раствор Хенкса или раствор Рингера.
Составы для чрескожного или чресслизистого введения, как правило, включают в себя, но без ограничения, пенетранты, такие как фусидовая кислота или соли желчных кислот в комбинации с детергентами или поверхностно-активными средствами. В некоторых вариантах осуществления составы можно изготавливать в форме аэрозолей, суппозиториев или пластырей. В некоторых вариантах осуществления пероральное введение может не являться благоприятным для белковых или пептидных активных ингредиентов; однако, этот тип композиций можно пригодным образом составлять, например, в форме с кишечнорастворимым покрытием, в депо, в капсуле и т.д., для того, чтобы они были защищены от пищеварительных ферментов, так что пероральное введение также можно использовать. Некоторые составы по изобретению содержат сбалансированный солевой раствор (Alcon Laboratories, Inc., Fort Worth, Texas) или сбалансированный солевой раствор плюс (Alcon Laboratories, Inc.). В некоторых вариантах осуществления состав содержит одно или несколько из следующего: цитрат, NaCl (например, 0,64%), хлорид калия (KC1) (например, 0,075%), дигидрат хлорида кальция (CaCl2·2H2O) (например, 0,048%), гексагидрат хлорида магния (MgCl·6H2O) (например, 0,03%), тригидрат ацетата натрия (CH3CO2Na·3H2O) (например, 0,39%), дигидрат цитрата натрия (C6H5O7Na3·2H2O) (например, 0,17%), сахарозу и гидроксид натрия и/или соляную кислоту (для доведения pH) и воду. Предшествующий список включает в себя некоторые молекулы, которые перечислены как конкретные гидраты, например, дигидрат, тригидрат, гексагидрат и т.д. Понятно, что различные гидраты этих соединений можно использовать по настоящему изобретению, и изобретение не является ограниченным этими конкретными гидратными формами перечисленных молекул. В некоторых вариантах осуществления, состав содержит одно или несколько из следующего: NaCl, моногидрат одноосновного фосфата, гептагидрат двухосновного фосфата натрия и соляную кислоту и/или гидроксид натрия для доведения pH, и воду. В некоторых вариантах осуществления, фармацевтическая композиция содержит по меньшей мере один ингредиент, выбранный из группы, состоящей из гистидина, MgCl2, трегалозы, полисорбата, полисорбата 20, NaCl, сахарозы, аргинина и пролина. В некоторых вариантах осуществления состав содержит одно или несколько из следующего: гистидин (например, приблизительно 10 мМ); дегидрат α,α-трегалозы (например, приблизительно 10% или приблизительно 50 мМ); MgCl2 (например, приблизительно 10 мМ); полисорбат, такой как полисорбат 20 (например, приблизительно 0,01%); и NaCl (например, приблизительно 0,1%). В некоторых вариантах осуществления, a состав может содержать одно или несколько из следующего: сахароза, аргинин или пролин. В некоторых вариантах осуществления состав содержит молекулу (молекулы) по настоящему изобретению, 10 мМ гистидин, 10 мМ MgCl2, 50 мМ трегалозу и 0,01% полисорбат 20 или состоит из этого. В некоторых вариантах осуществления состав содержит молекулу (молекулы) по настоящему изобретению, 1,0% NaCl и 10 мМ MgCl2 или состоит из этого. В некоторых вариантах осуществления состав содержит молекулу (молекулы) по настоящему изобретению и сбалансированный солевой раствор, обогащенный бикарбонатом, декстрозой и глутатионом, такой как BSS PLUS®, или состоит из этого. В некоторых вариантах осуществления, состав не содержит трегалозу. В некоторых вариантах осуществления состав или композиция находится при pH приблизительно 5,5. В некоторых вариантах осуществления состав или композиция находится при pH между приблизительно 5,0 и 9,0, приблизительно 5,0 и 5,5, приблизительно 5,3 и 5,7, приблизительно 5,5 и 6,0, приблизительно 5,8 и 6,2, приблизительно 6,0 и 6,5, приблизительно 6,3 и 6,7, приблизительно 6,5 и 7,0, приблизительно 6,8 и 7,2, приблизительно 7,0 и 7,5, приблизительно 7,3 и 7,7, приблизительно 7,5 и 8,0, приблизительно 7,8 и 8,2, приблизительно 8,0 и 8,5, приблизительно 8,3 и 8,7 и приблизительно 8,5 и 9,0, что бы ни было пригодно для сохранения биологической активности и стабильности активного ингредиента (ингредиентов).
Некоторые составы по изобретению можно изготавливать в форме аэрозолей, суппозиториев, глазных капель или пластырей.
Примеры пригодных составов и способов составления для желательного способа введения можно обнаружить в Remington's Pharmaceutical Sciences, latest edition, Mack Publishing Co., Easton, PA и в Патенте США No. 7208577.
В некоторых вариантах осуществления композиция для использования in vivo содержит «носитель» или «фармацевтически приемлемый носитель». Термин «носитель» относится к разбавителю, адъюванту, наполнителю или носителю, с которым вводят интересующий вектор. Термин «носитель» включает в себя, но без ограничения, либо твердый, либо жидкий материал, который может являться неорганическим или органическим и быть синтетического или природного происхождения, с которым активный компонент(ы) композиции смешивают или составляют для облегчения введения субъекту.
В целом, пригодное масло (масла), солевой раствор, водные растворы декстрозы (глюкозы) и родственных сахаров, и гликоли, такие как пропиленгликоль или полиэтиленгликоли, являются, как правило, пригодными носителями для парентеральных растворов. В некоторых вариантах осуществления растворы для парентерального введение содержат водорастворимую соль активного ингредиента, пригодные стабилизаторы, и, желательно или обязательно, буферные вещества. Средства против окисления, такие как бисульфит натрия, сульфит натрия или аскорбиновая кислота, либо по отдельности, либо в комбинации, можно использовать в качестве стабилизаторов. Используют также лимонную кислоту и ее соли и натрий ЭДТА. Кроме того, растворы для парентерального введения могут содержать консерванты, такие как хлорид бензалкония, метил- или пропилпарабен и хлорбутанол.
Носители могут включать в себя углеводы, такие как трегалоза, маннит, глутатион, ксилит, сахароза, лактоза и сорбит. Другие ингредиенты для использования в составах могут включать в себя, например, DPPC (1,2-дидеканоил-sn-глицеро-3-фосфохолин), DOPE (1,2-диолеоил-sn-глицеро-3-фосфоэтаноламин), DSPC (1,2-дистеароил-sn-глицеро-3-фосфохолин 1,2-Дистеароил-sn-глицеро-3-фосфохолин) и DOPC (1,2-диолеоил-sn-глицеро-3-фосфохолин). Можно использовать природные или синтетические поверхностно-активные вещества. Можно использовать полиэтиленгликоль (даже наряду с его использованием при дериватизации белка). Можно использовать декстраны, такие как циклодекстран. В некоторых вариантах осуществления можно использовать циклодекстрин, третичные амины и/или бета-циклодекстрин. Можно использовать соли желчных кислот и другие родственные усилители. Можно использовать целлюлозу и производные целлюлозы. Можно использовать аминокислоты, например, использовать при составлении буфера. Предусмотрено также использование липосом, микрокапсул или микросфер, комплексов включения, или других типов носителей.
Пригодные фармацевтические наполнители включают в себя, но без ограничения, крахмал, глюкозу, лактозу, сахарозу, желатин, антибиотики, консерванты, солод, рис, муку, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое снятое молоко, глицерин, пропилен, гликоль, воду, этанол и т.п. Композиция, если желательно, может также содержать увлажняющие и/или эмульгирующие, и/или забуферивающие pH средства. При необходимости, композиция может включать в себя также солюбилизирующее средство и/или местный анестетик, такой как лигнокаин, для облегчения боли в участке инъекции.
В настоящем документе предусмотрена также доставка в легкие средства или белка (или его производного) по настоящему изобретению. В некоторых вариантах осуществления аналог(и) фактора комплемента B доставляют в легкие млекопитающих в ходе ингаляции, и он может в основном оставаться в легких или в некоторых вариантах осуществления проходит через эпителиальную выстилку легких в кровоток, (например, см. Adjei et al., Pharmaceutical Research 7:565-569 (1990); Adjei et al., International Journal of Pharmaceutics 63: 135-144 (1990); Braquet et al., Journal of Cardiovascular Pharmacology 13(suppl. 5):s. l43-146 (1989); Hubbard et al., Annals of Internal Medicine 3:206-212 (1989); Smith et al., J. Clin. Invest. 84: 1145-1 146 (1989); Oswein et al., Proceedings of Symposium on Respiratory Drug Delivery II, Keystone, Colo., March, 1990; Debs et al., The Journal of Immunology 140:3482-3488 (1988) и Platz et al., Патент США No. 5284656). Для использования в осуществлении этого изобретения на практике предусмотрен широкий диапазон механических устройств, разработанных для доставки в легкие терапевтических продуктов, включая, но без ограничения, распылители, дозирующие ингаляторы и порошковые ингаляторы. Некоторыми конкретными примерами коммерчески доступных устройств, пригодных для осуществления на практике некоторых вариантов осуществления изобретения, являются распылитель ULTRA VENT™, изготовленный Mallinckrodt, Inc., St. Louis, Mo.; распылитель ACORN II®, изготовленный Marquest Medical Products, Englewood, Colo.; дозирующий ингалятор VENTOLIN, изготовленный Glaxo Inc., Research Triangle Park, N.C.; и порошковый ингалятор SPINHALER, изготовленный Fisons Corp., Bedford, Mass.
В некоторых вариантах осуществления белок получают в форме частиц. В некоторых вариантах осуществления эта форма частиц обладает средним размером частиц менее 10 мкм (или микрон), наиболее предпочтительно, 0,5-5 мкм, для доставки на периферию легкого.
Составы, пригодные для использования с распылителем (например, струйным или ультразвуковым), как правило, содержат аналог фактора комплемента B, растворенный в воде, в некоторых вариантах осуществления, в концентрации от приблизительно 0,1 до приблизительно 25 мг биологически активного белка на мл раствора. Состав может включать также буфер и/или простой сахар (например, для стабилизации белка и регуляции осмотического давления). Состав для распылителя может также содержать поверхностно-активное вещество для снижения или предотвращения индуцированной поверхностью агрегацию белка (белков), вызванную атомизацией раствора при формировании аэрозоля.
Составы для использования с дозирующим ингалятором, как правило, содержат тонко измельченный порошок, содержащий аналог фактора комплемента B по изобретению, суспендированный в пропелленте, например, с помощью поверхностно-активного вещества. Пропеллент может представлять собой любой общепринятый материал, применяемый для этой цели, такой как хлорфторуглерод, гидрохлорфторуглерод, гидрофторуглерод или углеводород, включая трихлорфторметан, дихлордифторметан, дихлортетрафторэтанол, и 1,1,1,2-тетрафторэтан или их комбинации. Пригодные поверхностно-активные вещества включают в себя триолеат сорбитана и соевый лецитин. Олеиновая кислота также может являться пригодной в качестве поверхностно-активного вещества. В некоторых вариантах осуществления, составы для распределения из порошкового ингалятора содержат тонко измельченный сухой порошок, содержащий аналог фактора комплемента B по изобретению, и могут содержать также наполнитель, такой как лактоза, сорбит, сахароза, маннит, трегалоза или ксилит, в количествах, облегчающих распределение порошка из устройства, например, 50-90% по массе состава.
Введение и доставка
Понятно, что при обсуждении введения или доставки нуклеиновой кислоты, кодирующей аналог белка фактора комплемента B, это изобретение относится также к введению или доставке собственно аналога белка фактора комплемента B. Понятно, что при обсуждении введения аналога фактора комплемента B, это изобретение относится также к введению нуклеиновой кислоты, кодирующей аналог белка фактора комплемента B.
В некоторых вариантах осуществления, аналоги фактора комплемента B или композиции по изобретению можно вводить местно или системно. Пригодные способы введения описаны в настоящем документе и известны в данной области. Способы введения или доставки включают в себя, но без ограничения, внутрикожный, внутримышечный, внутрибрюшинный, внутривенный, подкожный, интраназальный, интратрахеальный, местный, ингаляцию, чрескожный, ректальный, парентеральные способы, эпидуральный, интракраниальный, в головной мозг, интравентрикулярный, субдуральный, внутрисуставной, интратекальный, интракардиальный, интракоронарный, интравитреальный, субретинальный, в переднюю камеру глаза, в форме частиц, местно в роговицу, субконъюнктивальный, субтеноновую инъекцию, использование глазных капель, пероральные способы, посредством баллонного катетера, посредством стента или любую их комбинацию. В некоторых вариантах осуществления композицию или аналог фактора комплемента B по изобретению вводят в друзы, например, посредством инъекции непосредственно в друзы. Системное введение может представлять собой, но без ограничения, внутривенную или внутриартериальную инъекцию или чрезслизистое, подкожное и/или чрескожное введение. В некоторых вариантах осуществления композицию по изобретению можно первоначально направлять в участок, отличный от участка заболевания. Например, что касается AHR, возникающей в легких животного, внутрибрюшинная инъекция белка, вектора или нуклеиновой кислоты по изобретению может приводить к изменению AHR в легких, например, см. Park et al, American Journal of Respiratory и Critical Care Medicine 169:726-732, (2004). В некоторых вариантах осуществления уровень дозирования и/или способ введения композиции может зависеть от характера композиции, характера состояния (состояний), подлежащего лечению, и/или анамнеза индивидуального пациента. В некоторых вариантах осуществления вводят клетки, экспрессирующие аналог фактора комплемента B по изобретению. Эти клетки могут представлять собой линию клеток, ксеногенную, аллогенную или аутологичную.
В некоторых вариантах осуществления, например, включающих в себя введение в глаз, аналог белка фактора комплемента B или вектор по изобретению вводят приблизительно один раз в каждую неделю, месяц, 2 месяца, 3 месяца, 6 месяцев, 9 месяцев, год, 18 месяцев, 2 года, 30 месяцев, 3 года, 5 лет, 10 лет или по необходимости. В некоторых вариантах осуществления, например, включающих в себя введение в глаз, молекулу или вектор по изобретению вводят приблизительно каждые 1-4 недели, приблизительно каждые 4-8 недель, приблизительно каждые 1-4 месяца, приблизительно каждые 3-6 месяцев, приблизительно каждые 4-8 месяцев, приблизительно каждые 6-12 месяцев, приблизительно каждые 9-15 месяцев, приблизительно каждые 12-18 месяцев, приблизительно каждые 15-21 месяц, приблизительно каждые 18-24 месяца, приблизительно каждые 1-2 года, приблизительно каждые 1,5-3 года, приблизительно каждые 2-4 года, приблизительно каждые 3-5 лет, приблизительно каждые 5-7 лет, приблизительно каждые 7-10 лет или приблизительно каждые 10-20 лет. Ожидают, что введение вектора, кодирующего аналог белка фактора комплемента B, будет менее частым, чем введение аналога белка фактора комплемента B. В некоторых вариантах осуществления изобретения фармацевтический препарат содержит вектор, кодирующий аналог фактора комплемента B по изобретению, и фармацевтический препарат вводят пациенту только один раз.
В некоторых вариантах осуществления, например, включающих в себя введение в глаз, вектор, кодирующий аналог фактора комплемента B, вводят 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более раз пациенту на протяжении его жизни. В некоторых вариантах осуществления, например, включающих в себя введение в глаз, лентивирусный вектор по изобретению вводят 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более раз пациенту на протяжении его жизни.
В некоторых вариантах осуществления аналог белка фактора комплемента B по изобретению вводят интравитреальной инъекцией в глаз человека. В некоторых вариантах осуществления, от приблизительно 15 мкг до приблизительно 5 мг; от приблизительно 15 мкг до приблизительно 500 мкг; от приблизительно 100 мкг до приблизительно 900 мкг; от приблизительно 300 мкг до приблизительно 700 мкг; от приблизительно 500 мкг до приблизительно 1 мг; от приблизительно 1 мг до приблизительно 5 мг; приблизительно 1 мг; или приблизительно 500 мкг аналога белка фактора комплемента B вводят интравитреальной инъекцией в глаз человека.
В некоторых вариантах осуществления, a аналог белка фактора комплемента B по изобретению вводят субретинальной инъекцией или интравитреальной инъекцией лентивирусного или аденоассоциированного вирусного (AAV) вектора. В некоторых вариантах осуществления, от приблизительно 5×106 до приблизительно 5×108; от приблизительно 5×106 до приблизительно 5×107; от приблизительно 5×107 до приблизительно 5×108; от приблизительно 1×107 до приблизительно 1×108; от приблизительно 3×107 до приблизительно 5×107; от приблизительно 2,5×107; от приблизительно 5×107; от приблизительно 7,5×107; или от приблизительно 1×108 трансдуцирующих единиц лентивирусного вектора вводят субретинальной инъекцией. В некоторых вариантах осуществления, от приблизительно 5×108 до приблизительно 1×109; от приблизительно 5×108 до приблизительно 7,5×108; от приблизительно 7,5×108 до приблизительно 1×109; от приблизительно 6×108 до приблизительно 9×108; от приблизительно 7×108 до приблизительно 8×108; от приблизительно 5×108; приблизительно 6×108; приблизительно 7×108; приблизительно 8×108; приблизительно 9×108; или приблизительно 1×109 трансдуцирующих единиц AAV вектора вводят субретинальной инъекцией.
В некоторых вариантах осуществления, от приблизительно 5×108 до приблизительно 1×1010; от приблизительно 5×108 до приблизительно 5×109; от приблизительно 5×108 до приблизительно 2×109; от приблизительно 2×109 до приблизительно 5×109; от приблизительно 5×109 до приблизительно 1×1010; от приблизительно 5×108 до приблизительно 1×109; от приблизительно 1×109 до приблизительно 3×109; от приблизительно 3×109 до приблизительно 6×109; от приблизительно 6×109 до приблизительно 1×1010; или от приблизительно 1×109 до приблизительно 1×1010 трансдуцирующих единиц AAV вектора вводят интравитреальной инъекцией.
В некоторых вариантах осуществления, от приблизительно 50 мкл до приблизительно 100 мкл, от приблизительно 50 мкл до приблизительно 75 мкл, от приблизительно 75 мкл до приблизительно 100 мкл, от приблизительно 60 мкл до приблизительно 90 мкл, от приблизительно 70 мкл до приблизительно 80 мкл, приблизительно 50 мкл; приблизительно 60 мкл; приблизительно 70 мкл; приблизительно 80 мкл; приблизительно 90 мкл; или приблизительно 100 мкл аналога белка фактора комплемента B или вектора, кодирующего аналог белка фактора комплемента B, инъецируют субретинально. В некоторых вариантах осуществления от приблизительно 50 мкл до приблизительно 1 мл, от приблизительно 50 мкл до приблизительно 500 мкл, от приблизительно 500 мкл до приблизительно 1 мл, от приблизительно 250 мкл до приблизительно 750 мкл, от приблизительно 250 мкл до приблизительно 500 мкл, от приблизительно 500 мкл до приблизительно 750 мкл, от приблизительно 400 мкл до приблизительно 600 мкл, или от приблизительно 750 мкл до приблизительно 1 мл аналога белка фактора комплемента B или вектора, кодирующего аналог белка фактора комплемента B, инъецируют интравитреально.
В некоторых вариантах осуществления противовоспалительное средство можно доставлять в комбинации с аналогом белка фактора комплемента B (например, hfB3-292S или hfB3-292S-740N), вектором или нуклеиновой кислотой по изобретению. Противовоспалительное средство можно доставлять до, во время и/или после введения молекулы или вектора по изобретению. В некоторых вариантах осуществления противовоспалительное средство вводят в том же самом растворе и/или в том же самом шприце, что и аналог белка фактора комплемента B, нуклеиновую кислоту или вектор по изобретению. В некоторых вариантах осуществления аналог белка фактора комплемента B или вектор по изобретению и противовоспалительное средство совместно вводят в глаз, например, как описано в настоящем документе.
Многие противовоспалительные лекарственные средства известны в данной области и включают в себя, но без ограничения, дексаметазон, метасульфобензоат дексаметазона натрия, фосфат дексаметазона натрия, фторметолон, бромфенак, пранопрофен, РЕСТАСИС™, офтальмологическую эмульсию циклоспорина, напроксен, глюкокортикоиды, кеторолак, ибупрофен, толметин, нестероидные противовоспалительные лекарственные средства, стероидные противовоспалительный лекарственные средства, диклофенак, флурбипрофен, индометацин и супрофен.
Некоторые варианты осуществления по изобретению включают в себя введение как аналога белка фактора комплемента B, так и кодирующего его вектора. Аналог белка фактора комплемента B по изобретению можно доставлять до, во время и/или после введения вектора по изобретению. В некоторых вариантах осуществления аналог белка фактора комплемента B по изобретению вводят в том же самом растворе и/или в том же самом шприце, что и вектор по изобретению. В некоторых вариантах осуществления аналог белка фактора комплемента B по изобретению и вектор по изобретению совместно вводят в глаз, например, как описано в настоящем документе.
Кроме того, аналог фактора комплемента B или кодирующую его нуклеиновую кислоту можно доставлять или вводить животному посредством клетки, например, в качестве клеточной терапии. Например, это можно осуществлять введением или доставкой клетки (клеток), экспрессирующих аналог(и) фактора комплемента B. В некоторых вариантах осуществления, аналог(и) фактора комплемента B экспрессируют из клеток посредством регулируемого, индуцируемого и/или репрессируемого промотора. В некоторых вариантах осуществления, инкапсулированные клетки, экспрессирующие аналог(и) фактора комплемента B, доставляют животному, например, см. Публикацию PCT No. WO07078922 относительно инкапсулированных клеток. В некоторых вариантах осуществления клетки вводят местно (например, в сустав, интравитреально, интраретинально, интракраниально и т.д.) или системно (например, i.v.).
Клетки, подлежащие введению животному, могут являться аутологичными, аллогенными или ксеногенными. В некоторых вариантах осуществления аутологичные клетки подвергают манипуляциям ex vivo, чтобы заставить их продуцировать аналог белка фактора комплемента B по изобретению, и, в некоторых вариантах осуществления, клетки вводят обратно животному. Перенос нуклеиновой кислоты, содержащей кодирующую область, в клетки ex vivo может представлять собой любой способ, такой как электропорация, микроинъекция, слияние клеток, опосредованный хромосомами перенос генов, опосредованный микроклетками перенос генов, слияние сферопластов, липофекция, бомбардировка микрочастицами, опосредованная фосфатом кальция трансфекция, вирусная инфекция и т.д. Необязательно, в клетки можно вводить также селективный маркер. Если используют селективный маркер, клетки потом подвергают отбору, например, для усиления экспрессии и/или для выделения клеток, экспрессирующих перенесенную кодирующую область (см., например, Loeffler & Behr, Meth. Enzymol. 217:599-618 (1993); Cohen et al., Meth. Enzymol. 217:618-644 ( 1993); и Cline, Pharmac. Ther. 29:69-92 (1985)).
Рекомбинантные клетки (например, аутологичные или аллогенные клетки, трансдуцированные in vitro) можно доставлять пациенту различными способами, известными в данной области. Например, клетки можно инкапсулировать перед введением, как известно в данной области. В некоторых вариантах осуществления при инкапсуляции, клетки не являются аутологичными. В некоторых вариантах осуществления, рекомбинантные клетки крови (например, гематопоэтические стволовые клетки и/или клетки-предшественники) вводят внутривенно. В некоторых вариантах осуществления, клетки глаза и/или плюрипотентные клетки можно инъецировать непосредственно в глаз. Необходимое количество клеток зависит от желательного эффекта, состояния животного и т.д.
В некоторых вариантах осуществления изобретения система доставки генов может приводить к трансдукции и/или стабильной интеграции гена или кодирующей области для аналога фактора комплемента B в клетку-мишень. В некоторых вариантах осуществления клетки-мишени представляют собой клетки млекопитающих, такие как клетки приматов и клетки человека. В некоторых вариантах осуществления клетки-мишени представляют собой клетки глаза, такие как клетки пигментного эпителия сетчатки, клетки сетчатки или плюрипотентные клетки. Клетки-мишени могут находиться in vitro, ex vivo или in vivo. В некоторых вариантах осуществления клетка-мишень представляет собой стволовую клетку. Стволовые клетки включают в себя, но без ограничения, плюрипотентные стволовые клетки, тотипотентные стволовые клетки, гематопоэтические стволовые клетки, стволовые клетки злокачественных опухолей и эмбриональные стволовые клетки. В некоторых вариантах осуществления плюрипотентные клетки, предусмотренные в настоящем документе, не являются клетками для размножения живого существа из зиготы или бластомера. По настоящему изобретению предусматривают также использование частично недифференцированных клеток для имплантации в глаз пациента, нуждающегося в лечении, например, для регенерации клеток глаза.
Трансгенные животные
Некоторые варианты осуществления изобретения относятся к трансгенному животному (например, не относящемуся к человеку), экспрессирующему аналог фактора комплемента B по изобретению. Способы получения трансгенных животных известны в данной области. В некоторых вариантах осуществления, трансгенное животное (такое как мышь) также может содержать мутацию, делецию или нарушение гена Fas, например, см. Macmicking et al. Cell. 81:641-650 (1995).
ПРИМЕРЫ
Изобретение в настоящее время описано по отношению к следующим примерам. Эти примеры представлены только с целью иллюстрации, и изобретение никаким образом не следует рассматривать как ограниченное этими примерами, но следует рассматривать как охватывающее все без исключения варианты, которые становятся очевидными в результате объяснений, представленных в настоящем документе.
В то время как конкретные варианты осуществления изобретения описаны в настоящем документе с целью описания, специалистам в данной области очевидно, что многочисленные варианты частностей можно осуществлять без отклонения от изобретения, как описано в прилагаемой формуле изобретения.
Пример 1. Получение экспрессирующей hfB3 конструкции
Плазмида разработана для включения кодирующей последовательности hfB3 человека с селективным маркером IRES-Neo (hfB3-IRES-Neo). Плазмиду синтезировали в GENEART AG (Regensburg, Germany, plasmid), договор о возмездном оказании услуг. Участок рестрикции Nhe I включали на 5' и 3' концах кодирующей последовательности. Кодирующую hfB3 последовательность нуклеиновой кислоты подвергали оптимизации кодонов для оптимальной экспрессии в клетках млекопитающих.
Плазмиду для экспрессии гена, pCI (Promega, Madison, WI), модифицировали. Во-первых, полиA BGH (бычьего гормона роста) удаляли из pCI и заменяли на синтетический полиA. Затем, последовательность, кодирующую hfB3, с селективным маркером (hfB3-IRES-Neo) вырезали посредством Nhe I из плазмиды и клонировали в участок Sal I модифицированной pCI посредством лигирования по тупым концам для получения экспрессирующей hfB3 конструкции. Плазмиду полностью секвенировали для подтверждения целостности конструкции (SEQ ID NO:5).
Пример 2. Получение экспрессирующей hfB3-292S конструкции
Дополнительную модификацию вводили в белок hfB3. Белок фактор B дикого типа человека и белок hfB3 обладают 23 цистеиновыми аминокислотами (C), что позволяет предполагать, что по меньшей мере один неспаренный свободный цистеин присутствует в белке. Картирование дисульфидных связей позволяет предполагать, что свободный C в биологически активном hfB3 локализован в аминокислоте 292. C в 292 является высоко консервативным в белке факторе B среди различных видов (таблица 1, выше). В этом примере, этот C в 292 заменили на серин (S), получив hfB3-292S.
Для получения экспрессирующей hfB3-292S конструкции, сайт-специфическую мутацию вводили в экспрессирующую hfB3 конструкцию (SEQ ID NO:5).
Экспрессирующую hfB3 конструкцию использовали в качестве матрицы для получения участка мутации, изменяющей C в положении 292 на S с использованием набора для сайт-специфического мутагенеза от Stratagene согласно рекомендациям производителя, получая экспрессирующую hfB3-292S конструкцию. Использовали два праймера: прямой праймер 5'-CACCGGCGCCAAGAAGAGCCTGGTCAACCTGATC-3' (SEQ ID NO:6) и обратный праймер 5'-GATCAGGTTGACCAGGCTCTTCTTGGCGCCGGTG-3' (SEQ ID NO:7). Подчеркнутые нуклеотиды обозначают аминокислоту с мутацией с C до S. Экспрессирующая hfB3-292S кассета включает, от 5' до 3', промотор CMV, химерный интрон, кодирующую последовательность для hfB3-292S с оптимизированными кодонами, селективный маркер IRES-Neo и синтетический полиA (фигура 2). Всю конструкцию секвенировали для подтверждения мутации и целостности конструкции (SEQ ID NO:8). Ожидаемая аминокислотная последовательность для hfB3-292S показана на SEQ ID NO:2.
Пример 3. Получение стабильных экспрессирующих hfB3 и hfB3-292S линий клеток
Стабильные линии клеток, экспрессирующие белок hfB3 или hfB3-292S, получали трансфекцией клеток 293 FreeStyle (Invitrogen, кат. No. R79007) экспрессирующей hfB3 или hfB3-292S конструкцией. Трансфекция клеток 293 FreeStyle плазмидной ДНК являлась опосредованной PEI (полиэтиленимин, Sigma, кат. No. 23966) трансфекцией. Раствор PEI получали в стерильной воде с конечной концентрацией 1 мг/мл. pH доводили до 7,0 5Н HC1. Раствор стерилизовали с использованием фильтра 0,22 мкм. Аликвоты PEI хранили замороженным при -80°C до использования.
Способ трансфекции является следующим:
за сутки до трансфекции клетки рассевали при 1×106 клеток/мл в бессывороточной экспрессионной среде 293F (Invitrogen, кат. No. 12338-018).
На следующие сутки клетки промывали базовой средой RPMI1640 (Invitrogen, кат. No. 22400-089), дополненной только HT Supplement (кат. No. 11067-030, Invitrogen), ресуспендировали в такой же среде при 2×106 клеток/мл и распределяли в новый 6-луночный планшет по 1 мл в каждую лунку.
Исходные растворы ДНК и PEI получали в стерильном 150 мМ NaCl следующим образом: 2 мкг ДНК экспрессирующей hfB3 или hfB3-292S конструкции (в 2,5 мкл) разводили в 47,5 мкл 150 мМ NaCl и перемешивали пипетированием (раствор ДНК). Десять микролитров (10 мкл) раствора PEI разводили в 40 мкл 150 мМ NaCl, с последующим осторожным встряхиванием (раствор PEI). Растворы ДНК и PEI инкубировали при комнатной температуре в течение 5 минут. Затем раствор PEI добавляли к раствору ДНК и смеси позволяли инкубироваться при комнатной температуре в течение дополнительных 10 минут и затем смесь ДНК/PEI добавляли к клеткам в 6-луночном планшете, и клетки инкубировали с покачиванием (200 об./мин) в течение 5 часов при 37°C в инкубаторе с 8% CO2 и 85% влажностью. Через 5 часов, 1,1 мл полной экспрессионной среды 293F (без добавок) добавляли в каждую лунку и инкубацию продолжали в течение 72 часов.
Затем клетки собирали, один раз промывали экспрессионной средой 293F, и помещали в свежую экспрессионную среду 293F, содержащую 300 мкг/мл G418 (Teknova). В качестве отрицательного контроля, равное количество нетрансфицированных наивных клеток 293F культивировали в такой же содержащей G418 среде. Клетки подвергали отбору на G418 в течение приблизительно 3 недель. К этому времени нетрансфицированные клетки в содержащей G418 среде погибали. Трансфицированные клетки пропускали через градиент FICOL (Sigma) для удаления мертвых или умирающих клеток от устойчивой к G418 живой популяции. Устойчивую к G418 живую популяцию далее размножали в течение периода приблизительно 2 недель, в течение которого клетки каждые 2-3 суток осаждали центрифугированием и ресуспендировали в свежей экспрессионной среде 293F, содержащей 300 мкг/мл G418.
Пример 4. Продукция hfB3 и hfB3-292S
Устойчивые к G418 продуцирующие hfB3 или hfB3-292S клетки рассевали при плотности 2×106 клеток/мл в экспрессирующей среде 293F в 6-луночных планшетах с 2 мл культуры в каждой лунке, в 500 мл роллерных колбах с 100 мл культуры в каждой колбе, или в 3000 мл роллерных колбах с 1000 мл культуры в каждой колбе. Клетки инкубировали в течение 72 часов с встряхиванием при 100 об./мин на орбитальном встряхивателе в инкубаторе при 37°C с 8% CO2 и 80% влажностью.
Затем супернатант культуральной среды, содержащий белок hfB3 или белок hfB3-292S, собирали и центрифугировали при 2000 об./мин в течение 10 минут для осветления клеточного дебриса, после чего культуральную среду фильтровали через 0,22 мкм фильтр.
Пример 5. Количественное определение белков hfB3 и hfB3-292S
Разработан электрохемилюминесцентный анализ (ECL) для количественного определения hfB3 и hfB3-292S с фактором B дикого типа человека в качестве стандарта. Анализ представлял собой сэндвич-иммуноанализ на основе способа ECL BioVeris. Кратко, анализ ECL разработан в формате сэндвич-анализа в 96-луночном планшете, с одной стадией, и без промывки. Образцы для контроля качества (очищенный фактор B из плазмы человека, Quidel, кат. No. A408) и тестируемые образцы инкубировали с готовой смесью реагентов, содержащей биотинилированное моноклональное антитело против hfB (моноклональное антитело против фактора B человека, R&D Systems, кат. No. MAB2739), меченное BV-TAG плюс поликлональное антитело против hfB (поликлональное антитело против фактора B человека, R&D Systems, кат. No. AF2739), и покрытые стрептавидином парамагнитные бусины. Смесь инкубировали в течение 150 минут. После инкубации добавляли останавливающий раствор (боратный буфер, 250 мМ, pH 9,2, содержащий 500 мМ хлорид натрия и 1,6 мг/мл BSA), и затем планшет считывали на анализаторе M1MR. Оцениваемый динамический диапазон для анализа составлял 9,0-950 нг/мл.
Пример 6. Анализ Вестерн-блоттингом hfB3 и hfB3-292S
Электрофорез в полиакриламидном геле белка hfB3 и белка hfB3-292S в денатурирующих, но невосстанавливающих условиях (SDS-PAGE) проводили посредством смешивания образцов белка hfB3 или белка hfB3-292S с невосстанавливающим буфером для образцов белка (Pierce). Белок фактор B человека, очищенный из плазмы (100 нг на образец, Quidel) использовали в качестве положительного контроля. Каждый гель также содержал лунку с предварительно окрашенными маркерами молекулярной массы белков (15 мкл/дорожку (Invitrogen). Образцы и маркеры нагревали при 95°C в течение 5 минут в невосстанавливающем буфере для образцов белка. Образцы наносили в предварительно залитый 7,5% Tris-HCL мини-гель (Bio-Rad). Разделение в геле проводили (10 Х SDS/Трис/глициновый буфер для разделения, Bio-Rad) при 75 В в течение 15 минут или пока фронт красителя не проходил через концентрирующий гель в разделяющий гель. Когда фронт красителя входил в разделяющий гель, вольтаж увеличивали до 100 В, и электрофорез продолжали, пока фронт красителя не выходил из геля.
Анализ Вестерн-блоттингом проводили посредством промывки и уравновешивания геля в буфере для переноса (10 Х Трис/глициновый буфер для переноса, Bio-Rad) в течение 20 минут с осторожным покачиванием. Нитроцеллюлозную мембрану (Bio-Rad) и бумагу для блоттинга также уравновешивали в буфере для переноса. Белки, разделенные в SDS-PAGE, переносили электрофоретически на нитроцеллюлозу с использованием ячейки для полусухого переноса (Bio-Rad) (20 В в течение 45 минут). После окончания переноса мембрану блокировали в 1 Х растворе казеина (Vector Laboratories) в течение по меньшей мере часа при комнатной температуре с осторожным покачиванием на качалке. Мембрану обрабатывали в качестве зонда первичным антителом (моноклональное антитело против фактора B человека, R&D Systems, кат. No. MAB2739), разведенным до 1:10000 в 1 Х растворе казеина при комнатной температуре, в течение 1 часа с осторожным покачиванием и промывали в 10 мл 1 Х раствора казеина 3 раза в течение 5 минут каждый при комнатной температуре с осторожным покачиванием на качалке. Мембрану инкубировали с биотинилированным антителом козы против IgG мыши (вторичное антитело, R&D Systems, кат. No. BAF007), разведенным до 1:20000 в 1 Х растворе казеина, в течение 1 часа при комнатной температуре с осторожным покачиванием на качалке и промывали в 10 мл 1 Х раствора казеина 3 раза в течение 5 минут каждый при комнатной температуре с осторожным покачиванием. Мембрану инкубировали в реагенте Vectastain ABC-AmP (Vector Laboratories) в течение 45 минут в 20 мл 1 Х раствора казеина, содержащем 40 мкл реагента A и 40 мкл реагента B. Мембрану промывали в 10 мл 1 Х раствора казеина 3 раза в течение 5 минут каждый при комнатной температуре с осторожным покачиванием.
Для получения хемилюминесцентного сигнала от Вестерн-блотов, мембраны уравновешивали в 20 мл 0,1 M Трис pH 9,5 в течение 5 минут без покачивания. Избыток буфера удаляли с мембраны посредством удерживания мембраны вертикально и касания углом мембраны Kimwipe. Мембрану помещали в новый контейнер интересующей стороной вверх. Субстрат Duolox (7 мл, Vector Laboratories) помещали непосредственно на интересующую сторону мембраны, которую инкубировали в течение 5 минут в темноте. Избыток Duolox удаляли с мембраны посредством удерживания мембраны вертикально и касания углом мембраны Kimwipe. Мембрану промывали посредством ее погружения в 20 мл 0,1 M Трис pH 9,5 на 5 минут с покачиванием в темноте. Избыток буфера удаляли с мембраны посредством удерживания мембраны вертикально и касания углом мембраны Kimwipe. Мембрану помещали в сложенный лист пищевой пленки и экспонировали с рентгеновской пленкой Kodak BioMax MS в кассете для пленки в течение 1-5 минут. Пленку помещали в раствор проявителя Kodak (разводили 26 мл раствора проявителя в 92 мл ddH2O) на 1 минуту. Пленку удаляли из раствора проявителя и помещали в раствор фиксажа Kodak (разводили 26 мл раствора фиксажа в 92 мл ddH2O) на 1 минуту. Наконец, пленку споласкивали водопроводной водой и позволяли высохнуть при комнатной температуре.
Как показано на фигуре 3, устойчивые к G418 продуцирующие hfB3 и hfB3-292S клетки продуцировали, в культуральной среде, белок hfB3 (дорожка 3) и белок hfB3-292S (дорожка 4) соответствующего размера (приблизительно 100 кДа). Эти белки мигрировали приблизительно с такой же скоростью, как фактор B дикого типа человека, очищенный из плазмы человека (дорожка 2). Не детектировали видимой полосы в культуральной среде нетрансфицированных клеток (отрицательный контроль), что указывает на специфичность моноклонального антитела против фактора B человека (дорожка 1). Интересно, что предполагаемые агрегаты приблизительно 200-260 кДа легко детектировали в образцах hfB3 (дорожка 3), в то время как не детектировали агрегатов в образцах hfB3-292S (дорожка 4), полученных в тех же условиях эксперимента. Агрегаты могли быть образованы неправильно свернутыми популяциями hfB3 в культуральной среде. Когда белки неправильно сворачиваются, они экспонируют гидрофобные области, подверженные формированию агрегатов посредством гидрофобно-гидрофобных взаимодействий. Эти данные позволяют предполагать, что в препарате белка hfB3-292S неправильное сворачивание являлось либо исключенным, либо значительно сниженным по сравнению с белком hfB3.
Пример 7. Анализ гемолитической активности альтернативного пути комплемента
Активность альтернативного пути комплемента человека можно измерять с использованием анализа гемолиза, как описано в этом примере.
Один миллилитр (1 мл) суспензии эритроцитов кролика (rRBC) (Lampire Biological Laboratory, кат. No. 7246408) промывали свежеприготовленным холодным буфером Mg2+-EGTA. Эритроциты переносили в 50 мл коническую центрифужную пробирку, добавляли 30 мл буфера Mg2+-EGTA и клетки осторожно перемешивали. rRBC осаждали в центрифуге Beckman Allegra 6KR при 1200 об./мин при 4°C без торможения в течение 5 минут и ресуспендировали в буфере Mg2+-EGTA. Эту стадию промывки повторяли дважды. rRBC ресуспендировали в 2 мл ледяном буфере Mg2+-EGTA, и количество клеток получали с использованием гемоцитометра.
Реакционную смесь для гемолитической активности составляли в 96-луночных планшетах с V-образным дном, помещенных в лед. Чтобы определить, может ли белок hfB3 или белок hfB3-292S конкурировать с белком фактором B человека дикого типа и ингибировать его гемолитическую активность, конкурентный анализ проводили в общем объеме 40 мкл, содержащем 500 нг фактора B дикого типа человека, увеличивающиеся количества белка hfB3 или белка hfB3-292S, и буфер GVB++ (Sigma, кат. No. G6415). Пятьдесят микролитров (50 мкл) истощенной по фактору B сыворотки человека, разведенной в 25 раз буфером Mg2+-EGTA добавляли в каждую лунку, затем 10 мкл промытых Mg2+-EGTA 5×107 rRBC. После добавления rRBCs, каждый образец осторожно смешивали в 96-луночном планшете с использованием многоканальной пипетки.
96-луночный планшет помещали в стеклянный лоток со слоем воды при 37°C, погружая дно планшета. Затем лоток помещали в водяную баню при 37°C с орбитальным встряхиванием при 110 об./мин в течение 40 минут. После инкубации планшет помещали на лед, 150 мкл ледяного 0,9% солевого раствора добавляли в каждую лунку, и каждую реакционную смесь осторожно перемешивали для остановки реакции. 96-луночный планшет центрифугировали при 2000 об./мин в центрифуге Eppendorf 5810R в течение 5 минут (мин) при 4°C без торможения для осаждения rRBC на дно планшета. Супернатант (180 мкл) удаляли из каждой лунки, не затрагивая осадок, и переносили в соответствующую лунку нового 96-луночного планшета. Поглощение каждого образца измеряли при 405 нм в считывателе для микропланшетов.
Пример 8. Биологическая активность hfB3 и hfB3-292S
Биологическую активность белка hfB3 и белка hfB3-292S исследовали посредством измерения опосредованного альтернативным путем комплемента гемолиза rRBC. Анализ, описанный в примере 7, использовали для тестирования активности белка hfB3 или белка hfB3-292S в ингибировании альтернативного пути комплемента человека. Каждую реакцию проводили в трех повторах. Как показано на фигуре 4, необработанная культуральная среда от продуцирующих белок hfB3-292S и белок hfB3 клеток эффективно ингибировала/снижала активность альтернативного пути комплемента зависимым от дозы образом. Несмотря на нежелание быть связанными теорией, белки hfB3 и hfB3-292S могут ингибировать активность альтернативного пути комплемента посредством конкуренции с белком фактором B дикого типа и/или посредством секвестрирования C3b и/или фактора комплемент D.
Пример 9. Очистка белков hfB3 и hfB3-292S
Культуральную среду линии клеток человека 293 FreeStyle, трансфицированных и стабильно экспрессирующих и секретирующих белок hfB3 или белок hfB3-292S, использовали в качестве исходного материала для очистки белка hfB3 или белка hfB3-292S. Растворимый секретированный белок hfB3 или hfB3-292S очищали из культурального супернатанта посредством комбинации анионообменной хроматографии (AEX), хроматографии гидрофобного взаимодействия (HIC) и эксклюзионной хроматографии (SEC) для стадий связывания, промежуточной очистки и финальной очистки, соответственно, с использованием установки для очистки GE AKTA. Две схемы очистки описаны ниже. Эти две схемы отличаются тем, что в одной используют одну стадию хроматографии HIC, и в другой используют две стадии хроматографии HIC.
Культуральный супернатант, содержащий белок hfB3, разводили дистиллированной водой в соотношении 4:1 по объему супернатант/вода для снижения проводимости до ~8 миллиСименс на сантиметр (мСм/см) и доводили до pH 7,5 50 мМ фосфатным буфером. Этот материал наносили непосредственно на предварительно набитую ионообменную колонку (POROS HQ 50, ABI) на установке для очистки AKTA с использованием насоса P-960 со скоростью потока 30 мл в минуту. Колонку предварительно уравновешивали буфером A (50 мМ фосфатный буфер (PB), pH7,5, проводимость ~8 мСм/см), с линейной скоростью потока 600 мл/час (~1 объем колонки/мин, CV/мин). Элюат мониторировали посредством детекции в УФ при 280 нм. После отмывки всего несвязавшегося материала, оставшийся материал элюировали нелинейным градиентом буфера A и буфера B (50 мМ PB и 1M NaCl, pH 7,5) для последовательного повышения проводимости подвижной фазы ступенчато от 8 мСм/см (0% буфера B для 2 CV), до 33 мСм/см (25% буфера B для 8 CV) и наконец, до 105 мСм/см (100% буфера B для 5 CV).
Чтобы дополнительно способствовать удалению загрязнений белками хозяина, вводили промежуточную стадию хроматографии гидрофобного взаимодействия (HIC), очищающую и разделяющую белки на основании различий в их поверхностной гидрофобности. Основные фракции (со стадии AEX хроматографии), содержащие белок hfB3, объединяли и доводили до содержания приблизительно 1,4 M сульфата аммония и 47 мМ фосфатного буфера, pH 7,5 (проводимость 216 мСм/см) (1,0 M сульфат аммония и 33,7 мМ фосфатный буфер, pH 7,5, проводимость 169 мСм/см для hfB3-292S) добавлением буфера C (1,5 M сульфат аммония и 50 мМ фосфатный буфер, pH 7,5) к образцу при соотношении по объему приблизительно 30:1 сульфат аммония/фосфатный буфер по отношению к образцу. Объединенный образец фильтровали через фильтр 0,2 мкм и наносили на колонку гидрофобного взаимодействия (HiTrap Phenyl HP, GE Healthcare), предварительно уравновешенную 1,5 M сульфатом аммония и 50 мМ фосфатным буфером, pH 7,5 (буфер C) (1,0 M сульфат аммония и 33,7 мМ фосфатный буфер для hfB3-292S) со скоростью потока 300 мл/час (1 CV/мин). Оставшийся материал элюировали посредством уменьшения концентрации сульфата аммония нелинейным образом.
Альтернативно, промежуточную стадию HIC хроматографии можно заменять двухступенчатой HIC хроматографией, например, для облегчения масштабирования процесса очистки. В этом способе двухступенчатой очистки HIC, большую часть белков клетки-хозяина отделяли от fB3 или fB3-292S посредством первой стадии HIC хроматографии, HIC с отрицательным отбором, посредством связывания с колонкой HIC (HiTrap Phenyl HP, GE Healthcare) в условиях низкой соли (0,75 M сульфат аммония и 25 мМ фосфатный буфер, pH 7,5 для fB3 и 0,6 M сульфат аммония и 20 мМ фосфатный буфер, pH 7,5, проводимость 100 мСм/см для hfB3-292S). Фракцию проскока со стадии HIC с отрицательным отбором, содержащую белок hfB3 или fB3-292S, затем повторно доводили до содержания приблизительно 1,5 M сульфата аммония и 50 мМ фосфатного буфера, pH 7,5 (проводимость 216 мСм/см) для fB3 или 1,0 M сульфата аммония и 33,7 мМ фосфатного буфера, pH 7,5, (проводимость 169 мСм/см) для hfB3-292S. Для второй стадии HIC хроматографии, образец фильтровали через фильтр 0,2 мкм и наносили на колонку гидрофобного взаимодействия (HiTrap Phenyl HP, GE Healthcare), предварительно уравновешенную 1,5 M сульфатом аммония и 50 мМ фосфатным буфером, pH 7,5 для fB3 и 1,0 M сульфатом аммония и 33,7 мМ фосфатным буфером для hfB3-292S со скоростью потока 300 мл/час (1 CV/мин). Оставшийся материал элюировали посредством уменьшения концентрации сульфата аммония нелинейным образом для дополнительного отделения белка fB3 или fB3-292S от оставшихся белков клетки-хозяина.
Фракции, содержащие биологически активный белок hfB3 со стадии HIC хроматографии объединяли и концентрировали с помощью центрифугирующего фильтрующего устройства (Millipore, Amicon Ultra, кат. No. 901024, отсекание 10000 MW). Концентрированный образец подвергали эксклюзионной хроматографии на колонке Sephacryl S300 26/60 HR (максимальный объем нанесения: 3% CV, ~10 мл), уравновешенной в буфере PBS (4 мМ фосфат, 150 мМ NaCl, pH 7,4, (GIBCO)). Элюцию белка hfB3 проводили при постоянной линейной скорости потока 60 см/час с использованием PBS, с мониторированием элюата детекцией УФ при 280 нм. Очищенный белок hfB3 сохраняли при -80°C в аликвотах.
Этот способ позволял почти полное отделение белка hfB3 от других примесей. Чистота белка hfB3 после трех стадий процесса хроматографии являлась достаточно высокой, на что указывает тот факт, что при анализе SDS-PAGE с окрашиванием серебром для 0,2 мкг очищенного белка hfB3 получали только отдельную четкую полосу (фигура 5, левая панель). Когда белок hfB3, очищенный вышеописанным способом трехстадийной хроматографии, подвергали конкурентному анализу против фактора B дикого типа человека в анализе гемолиза, он супрессировал гемолитическую активность альтернативного пути комплемента человека, показывая, что очищенный белок hfB3 являлся биологически активным (фигура 5, правая панель). Неожиданно, две популяции hfB3 детектировали посредством HIC, одна являлась биологически активной (обозначена как пик I или активная популяция), и другая обладала намного сниженной биологической активностью (обозначена как пик II или менее активная популяция) (фигура 6). Эти две формы легко детектировали обращенно-фазовой HPLC (RP-HPLC) в культуральной среде необработанных клеток для стабильно продуцирующих белок hfB3 клеток (Фигура 7).
Пример 10. Продуцирующая белок hfB3-292S линия клеток не продуцирует популяцию пика II
Необработанную культуральную среду клеток с белком hfB3-292S, содержащую аналог фактора комплемента B со свободным цистеином, замененным на серин в положении 292, подвергали анализу RP-HPLC с необработанной культуральной средой наивных клеток 293 FreeStyle в качестве отрицательного контроля и с необработанной культуральной средой клеток с белком hfB3 в качестве положительного контроля (фигура 7). Культуральная среда клеток с белком hfB3-292S не продуцировала какой-либо поддающейся детекции популяции пика II, в то время как анализ культуральной среды клеток с белком hfB3 показал, что 35% из hfB3 находилось в менее активной популяции, пика II. (фигуры 7A и B.)
Пример 11. Получение и характеризация экспрессирующей hfB4 конструкции и белка hfB4
Белок hfB4 разработан для замены аспарагиновой кислоты в аминокислоте 740 в hfB3, на аспарагин. Считают, что это изменение ослабляет или ингибирует функцию сериновой протеазы этого аналога белка фактора комплемента B.
Для получения экспрессирующей hfB4 конструкции сайт-специфическую мутацию вводили в экспрессирующую hfB3 конструкцию.
Экспрессирующую hfB3 конструкцию (SEQ ID NO:5), описанную в примере 1, использовали в качестве матрицы для получения мутации, изменяющую аспарагиновую кислоту (D) в положении 740 в SEQ ID NO:4 на аспарагин (N) с использованием набора для сайт-специфического мутагенеза от Stratagene (Stratagene, Santa Clara, CA) согласно инструкциям производителя, получая экспрессирующую hfB4 конструкцию. Использовали два праймера: прямой праймер 5'-GTCCCCGCCCACGCCCGGAACTTCCACATCAACCTGTTCC-3' (SEQ ID NO:15) и обратный праймер 5'-GGAACAGGTTGATGTGGAAGTTCCGGGCGTGGGCGGGGAC-3' (SEQ ID NO:16). Подчеркнутые нуклеотиды обозначают замену нуклеотида, приводящую к замене аминокислоты с D на N. Эта экспрессирующая hfB4 конструкция содержит, от 5' до 3', промотор CMV, химерный интрон, кодирующую последовательность для hfB4 с оптимизированными кодонами, селективный маркер IRES-Neo и синтетический полиA. Всю конструкцию секвенировали для подтверждения мутации и целостности конструкции. Ожидаемая аминокислотная последовательность для hfB4 показана на SEQ ID NO:17. Единственным различием между аминокислотной последовательностью hfB3 и hfB4 является замена D740N.
Стабильную линию клеток, экспрессирующую белок hfB4, получали опосредованной PEI трансфекцией и отбором с помощью лекарственного средства клеток 293, как описано в примере 3. Концентрацию белка hfB4 в культуральной среде популяции клеток после отбора измеряли посредством ECL, как описано в примере 5.
Биологическую активность белка hfB4, очищенного, как описано в примере 9 с использованием способа с одной стадией HIC хроматографии, исследовали анализом гемолитической активности, как описано в примере 7. В таблице 2 показано ингибирование гемолитической активности альтернативного пути комплемента человека культуральной средой, содержащей белок hfB4. Относительную гемолитическую активность оценивали по гемоглобину, высвобождаемому после гемолиза rRBC посредством гемолитической активности альтернативного пути комплемента человека. Как показано в таблице 2, hfB4 белок эффективно ингибировал активность альтернативного пути комплемента.
Пример 12. Получение и характеризация экспрессирующей hfB3-Fc конструкции и белка hfB3-Fc
hfB3-Fc представляет собой белок, слитый из hfB3 и Fc IgG. Конкретно, полноразмерный белок hfB3 слит с Fc IgG4 человека.
Для получения экспрессирующей hfB3-Fc конструкции продукт ПЦР амплифицировали с экспрессирующей hfB3 конструкции (SEQ ID NO:5), описанной в примере 1, с использованием двух праймеров: прямого праймера 5'-GCGCACCGGTGCTAGCGAATTCGGCGACAAGAAGGGCAGCTGCGA-3' (SEQ ID NO:19); и обратного праймера 5'-GCGCAGATCTCAGGAAGCCCAGGTCCTCAT-3' (SEQ ID NO:20). Продукт ПЦР 377 п.о., содержащий кодирующую область для C-конца hfB3-Fc, расщепляли затем Age I и Bgl II и лигировали в pFUSE-hIgG4Fc (Invivogen, кат. код: pfuse-hg4fcl), предварительно расщепленный Age I и Bgl II, получая плазмиду phfB3Cterm-Fc. Экспрессирующую hfB3 конструкцию (SEQ ID NO:5, описанную в примере 1) расщепляли EcoR I и EcoR V. Фрагмент EcoR I/EcoR V, содержащий N-конец hfB3, лигировали в phfB3Cterm-Fc, предварительно расщепленный EcoR I и EcoR V, получая phfB3-Fc. Плазмиду phfB3-Fc расщепляли Nhe I, и фрагмент, содержащий кодирующие hfB3 и Fc последовательности, лигировали в модифицированную конструкцию pCI с IRES-Neo, описанную в примере 1, получая экспрессирующую hfB3-Fc конструкцию (SEQ ID NO:18). Эта экспрессирующая hfB3-Fc конструкция включает, от 5' до 3', промотор CMV, химерный интрон, кодирующую последовательность для hfB3-Fc, селективный маркер IRES-Neo и синтетический полиA. Всю конструкцию секвенировали для подтверждения целостности конструкции (SEQ ID NO:18). SEQ ID NO:21 представляет собой аминокислотную последовательность белка hfB3-Fc.
Стабильную линию клеток, экспрессирующих белок hfB3-Fc, получали опосредованной PEI трансфекцией и отбором клеток 293 с лекарственным средством, как описано в примере 3. Отобранные с лекарственным средством клетки культивировали при 2×106 клеток/мл в течение 72 часов. Затем экспрессию белка hfB3-Fc исследовали подверганием 2 мкл супернатанта культуры клеток невосстанавливающему SDS-PAGE и анализу Вестерн-блоттингом. Как показано на фигуре 8, две полосы белка hfB3-Fc детектировали посредством специфического антитела козы против фактора B (R&D Systems, кат. No. AF2739). Маркеры молекулярной массы в кДа указаны слева. Без желания быть связанными теорией, эти две полосы белка hfB3-Fc могут представлять мономеры и димеры белка. Биологическую активность белка hfB3-Fc (в супернатанте культуры клеток) исследовали анализом гемолитической активности, как описано в примере 7. Как показано в таблице 3, белок hfB3-Fc ингибировал активность альтернативного пути комплемента.
Пример 13. Эффект повторяющегося замораживания/размораживания на ингибирование комплемента hfB3-292S
Белок hfB3-292S очищали, как описано в примере 9, с использованием способа с одной стадией HIC хроматографии. Очищенный белок hfB3-292S в PBS, удаленный из морозильника с -80°C и размороженный при комнатной температуре, учитывали как первый цикл замораживания и размораживания.
После этого размораживания концентрацию белка hfB3-292S доводили до 2 мг/мл с помощью PBS и одну аликвоту hfB3-292S отбирали в качестве образца и сохраняли во льду как образец первого замораживания и размораживания.
Затем пробирку с белком hfB3-292S замораживали помещением пробирки в баню с метанолом/сухим льдом на 20 минут и затем размораживали при комнатной температуре до полного размораживания (второй цикл замораживания и размораживания). Одну аликвоту образца отбирали в качестве образца и оставляли перед повторением следующего цикла замораживания и размораживания. Биологическую активность образцов с каждого цикла замораживания и размораживания (всего 7 раз) анализировали измерением их способности конкурировать с фактором B дикого типа человека в анализе гемолиза, опосредованного альтернативным путем комплемента, как описано в примере 7. Каждая реакционная смесь содержала фиксированное количество фактора B дикого типа человека (0,5 мкг) с увеличивающимися количествами hfB3-292S. Результаты в таблице 4 представляют процент ингибирования в гемолитическом анализе.
Эти результаты показывают, что белок hfB3-292S еще может эффективно ингибировать опосредованный альтернативным путем гемолиз даже после семи циклов замораживания и размораживания (таблица 4). Сходный уровень ингибирования гемолиза наблюдали для всех образцов, показав, что повторное замораживание и размораживание (вплоть до 7 раз) не влияет на способность белка hfB3-292S ингибировать опосредованный комплементом гемолиз.
Пример 14. Большая термостабильность белка hfB3-292S, чем белка hfB3
Белки hfB3 и hfB3-292S очищали, как описано в примере 9, с использованием способа с одной стадией HIC хроматографии. Очищенные белки hfB3 и hfB3-292S, оба в PBS, удаляли с -80°C. Концентрацию белка повторно доводили до 2 мг/мл в PBS (pH 7,4). Белки hfB3 и hfB3-292S разделяли на равные аликвоты в 0,6 мл пробирки эппендорф (40 мкл на пробирку) и затем сохраняли в условиях 4°C, -80°C и 37°C в течение 7 суток. Биологическую активность (способность ингибировать опосредованный комплементом гемолиз) каждого из сохраняемых образцов анализировали измерением их способности ингибировать опосредованный альтернативным путем комплемента гемолиз, как описано в примере 7. Результаты этого гемолитического анализа показали, что хранение при 4°C или -80°C в течение 7 суток не влияет на способность hfB3 или hfB3-292S ингибировать опосредованный альтернативным путем комплемента гемолиз. Однако, белок hfB3, хранившийся при 37°C в течение 7 суток, потерял по существу всю свою биологическую активность во всех четырех тестированных образцах. Примечательно, что hfB3-292S, сохраняемый при 37°C в течение 7 суток, хорошо сохранял свою биологическую активность и еще мог эффективно конкурировать фактором B дикого типа человека (таблица 5). Результаты в таблице 5 представляют процент ингибирования активности альтернативного пути комплемента человека (со стандартным отклонением) посредством hfB3 или hfB3-292S. Эти результаты показывают, что белок hfB3-292S обладает большей термостабильностью, чем белок hfB3, при 37°C.
Пример 15. Определение температуры плавления белка для белков фактора B человека, fB3 и fB3-292S
Температура плавления белка (Tm) является показателем термостабильности белка, и изменения аминокислотной последовательности белка могут влиять, наряду с другим, на термостабильность белка. Белок фактор B человека (hfB) содержит двадцать три остатка цистеина, двадцать два из которых встречаются в форме пар с дисульфидной связью (цистина) и один из которых, C292, присутствует в неспаренной сульфгидрильной форме.
Профили температуры плавления белка hfB3 (K258A, R259A, K260A, D279G, N285D) и белка hfB3-292S (единственный неспаренный остаток цистеина белка hfB3 модифицирован до серина) сравнивали с профилями температуры плавления белка hfB посредством инкубации каждого белка в присутствии 1-анилинонафтален-8-сульфоновой кислоты (ANS) и измерения увеличения флуоресценции ANS при 460 нм. ANS связывается с гидрофобными областями белка (Stryer, J. Molecular Biology (1965) 13:482-495,), и ее использовали для исследования эффекта температуры на гидрофобность поверхности белка hfB (Takada, et. al., Complement (1985) 2: 193-203). Образцы, содержащие белок hfB (35 мкг), белок hfB3 (50 мкг) и белок hfB3-292S (39 мкг) получали в 100 мкл буфере PBS (137 мМ NaCl, 2,7 мМ KC1, 10 мМ фосфат, pH 7,4), содержащем 10 мМ ANS (Invitrogen, каталожный # A-47). Образцы каждого белка (в трех повторах) инкубировали в закрытых полипропиленовых пробирках в течение тридцати минут при 21°C, 30°C, 37°C, 44°C, 47°C, 50°C, 55°C, 60°C и 65°C. Образцы переносили в прозрачный 96-луночный микропланшет (Costar, каталожный # 3635), и флуоресценцию измеряли в спектрофлуориметре для микропланшетов Perceptive Biosystems Cytofluor 4000 (возбуждение = 60/40 нм, излучение = 460/40 нм). Результаты анализировали с использованием соответствия нелинейной кривой 4PL. Значения Tm (среднее из результатов из двух экспериментов) для белка hfB дикого типа, белка hfB3 и белка hfB3-292S определили как 46,4°C, 45,1°C и 47,0°C, соответственно. Изменения пяти аминокислот, выполненные в белке hfB3 по отношению к белку hfB дикого типа (K258A, R259A, K260A, D279G, N285D) приводили к ΔTm=-1,3°C, указывая на то, что белок hfB3 являлся менее термостабильным по сравнению с соответствующим белком hfB дикого типа. Однако, Tm белка hfB3-292S (47,0°C) приводила к ΔTm=+1,9°C по отношению к белку hfB3 и указывает на то, что белок hfB3-292S являлся по меньшей мере настолько же термостабильным, как белок hfB дикого типа (46,4°C).
Таким образом, в отличие от результатов Culajay, et. al. (Biochemistry (2000) 39:7153-7158), которые показали, что замена остатка цистеина на серин в белке факторе роста фибробластов человека (FGF-1) (C83S или C117S) уменьшала Tm на 13°C и 2°C, соответственно, замена на серин в hfB3 в аминокислоте C292 приводила к hfB3-292S, являющемуся более термостабильным, чем hfB3.
Пример 16. Белок hfB3-292S предотвращает воспаление и повреждение суставов в модели ревматоидного артрита на мышах
Индуцированный антителами против коллагена артрит (CAIA) представляет собой модель агрессивного ревматоидного артрита на мышах (Terato K et al, J. Immunol. (1992) 148(7):2103-8; Terato K et al, Autoimmunity (1995) 22(3): 137-47). В этой модели коктейль антител против коллагена, содержащий 4 моноклональных антитела против коллагена (Chondrex, Inc., каталожный номер: 10010) с бустер-инъекцией LPS использовали для индукции артрита у самцов мышей DBA/1J дикого типа в возрасте шести недель (Jackson Laboratory).
Сорок мышей разделяли на три группы. Группа 1 содержала 10 мышей, служивших группой контроля - носителя, где 100 мкл PBS (фосфатно-солевой буфер, pH 7,4) инъецировали в хвостовую вену на сутки 0, бустер-инъекцию 25 мкг LPS (List Biological Lab, Campbell, California, каталожный номер: 421) на мышь вводили внутрибрюшинно (LPS находился в PBS в концентрации 500 мкг/мл) на сутки 3, и 100 мкл PBS через хвостовую вену снова на сутки 3, 5, 7 и 9. Группа 2 содержала 15 мышей, которым инъецировали коктейль антител против коллагена (0,25 мг в 100 мкл PBS на мышь) через хвостовую вену на сутки 0, вводили бустер-инъекцию 25 мкг LPS на сутки 3, и вводили 100 мкл PBS через хвостовую вену на сутки 3, 5, 7 и 9. Группа 3 содержала 15 мышей, которым инъецировали 0,25 мг коктейля антител против коллагена и 1 мг белка hfB3-292S, оба вместе в 100 мкл PBS на мышь через хвостовую вену на сутки 0, вводили бустер-инъекцию 25 мкг LPS на сутки 3, и вводили 1 мг белка hfB3-292S в 100 PBS через хвостовую вену на сутки 3, 5, 7 и 9.
Мышей осматривали каждые сутки. Массу каждой мыши регистрировали при измерении суставов. Для суставов передних и задних конечностей измеряли с использованием штангенциркулей ширину и толщину на сутки -1, 4, 6, 9 и 11. Последовательность измерения представляла собой левую переднюю конечность, левую заднюю конечность, правую заднюю конечность и правую переднюю конечность. На сутки 11 всех животных умерщвляли, и все конечности (левую переднюю конечность, правую переднюю конечность, левую заднюю конечность и правую заднюю конечность) собирали и сохраняли в пластиковых кассетах с индивидуальными ярлыками. Каждую кассету помещали в бокс для гистологических контейнеров, содержащий 10% раствор нейтрального забуференного формалина.
Для этих конечностей мышей получали парафиновые срезы и окрашивали H&E для исследования патогенеза суставов. Конкретно, после фиксации, каждую конечность от каждой мыши переносили в пластиковую кассету по отдельности. Конечности споласкивали проточной водой в мерном стакане в течение 30 минут (мин) при комнатной температуре (RT) для удаления фиксирующего раствора. Затем каждую кассету переносили в мерный стакан, содержащий декальцифицирующий раствор (Thermo Scientific, каталожный номер 8340) посредством погружения кассеты в раствор. Передние конечности декальцифицировали в течение 8 часов и задние конечности декальцифицировали в течение 9 часов. Через 8 часов или 9 часов, конечности снова споласкивали проточной водой в течение 30 мин при RT для удаления декальцифицирующего раствора. После декальцификации, конечности сохраняли в 70% этаноле в течение ночи (O/N) для следующей стадии дегидратации. Конечности дегидратировали последовательным погружением в: 75% этанол (полученный из 100% этанола) два раза; 85% этанол два раза, 95% этанол два раза и 100% этанол два раза, каждый раз на 15 мин при RT с покачиванием. Затем конечности погружали в смесь 1:1 100% этанола и кедрового масла (Fisher, каталожный номер: 040-1) в течение 15 мин при RT с встряхиванием, и повторяли еще два раза, всего три раза. Затем конечности погружали в 100% кедровое масло и инкубировали при 40°C в течение 5 час. После 5 часов инкубации конечности погружали в смесь 1:1 кедрового масла и метилсалицилата (ACROS, каталожный номер 119-36-8) на 60 мин при RT. Затем конечности погружали в другую смесь 1:1 кедрового масла и метилсалицилата O/N при RT. После инкубации O/N, конечности погружали в 100% метилсалицилат на 40 мин при RT, повторяли еще один раз (всего два раза). Наконец, конечности погружали в парафин, который подготавливали инкубацией при 60°C в течение 7 час.
Парафиновые срезы конечностей мыши получали с использованием микротома и нарезали до толщины 7 мкм. Срезы инкубировали в водяной бане при 40°C и переносили на предметное стекло Superfrost Plus. Срезы высушивали O/N при RT, и дополнительно высушивали инкубацией O/N на столике с подогревом. Стекла сохраняли при RT до окрашивания.
Иммобилизованные парафиновые срезы конечностей мышей подвергали окрашиванию H&E. Срезы депарафинизировали и регидратировали погружением в ксилол на 3 мин, повторяя 2 раза, всего 3 раза. Затем избыток ксилола промокали, и срезы погружали в 100% этанол на 3 мин, всего 3 раза, затем в 95% этанол на 3 мин, один раз, в 80% этанол на 3 мин, один раз, и в деионизированную воду на 5 мин, один раз. Все инкубации проводили при RT. Для окрашивания гематоксилином стекла погружали в гематоксилин на 4 мин, один раз, и споласкивали деионизированной водой. Стекла погружали в водопроводную воду на 5 мин один раз, чтобы позволить окраске проявиться. Стекла быстро погружали, 8-12 раз, в кислый этанол (200 мл 70% этанола плюс 150 мкл концентрированной HCL) для удаления окраски срезов. Затем стекла споласкивали дважды в течение 1 мин в водопроводной воде, и затем один раз в течение 2 мин в деионизированной воде. Избыток воды промокали со стекол перед окрашиванием эозином.
Для окрашивания эозином, стекла погружали в эозин один раз на 20 секунд. Затем стекла дегидратировали погружением в 95% этанол 3 раза на 5 мин. Стекла инкубировали в 100% этаноле 3 раза по 5 мин. Избыток этанола промокали, и стекла инкубировали в ксилоле три раза по 15 мин. Покровные стекла приклеивали к стеклам с использованием основанного на ксилоле Permount (EMS, каталожный номер 17986-01) посредством помещения капли Permount на стекло с использованием стеклянной палочки, следя за тем, чтобы не образовались пузыри. Затем покровное стекло располагали под углом на стекле и осторожно опускали на стекло. Permount позволяли распределяться под покровным стеклом, покрывая весь срез. Стекла высушивали O/N при RT под тягой.
Как показано на фигуре 10, в группе, подвергнутой инъекции коктейля антител против коллагена в отсутствие hfB3-292S (группа 2) индуцировано тяжелое опухание передней лапы. Для мышей, подвергнутых инъекции коктейля антител, но подвергнутой лечению белком hfB3-292S (группа 3), показано 65% уменьшение размера лапы (p<0,0003) (фигура 10). Эти результаты показывают значимый ингибирующий эффект hfB3-292S на артрит суставов в этой модели. При дозе 250 мкг коктейля антител против коллагена на мышь для индукции CAIA, для задних конечностей мышей не показано очевидного опухания. Не наблюдали значительного неблагоприятного эффекта на массу мышей для лечения белком hfB3-292S. Средняя масса у всех мышей стабильно сохранялась. Средняя масса для всех мышей во всех трех группах составляла 20,4±1,1 грамм на конец исследования.
Как показано на фигуре 9, мыши в группе 1, по-видимому, имели нормальные суставы, не имели поддающейся детекции инфильтрации воспалительных клеток в сустав, и хрящ и кости выглядели нормально (фигура 9, верхняя панель). Мыши в группе 2 страдали тяжелым воспалением суставов, инфильтрацией воспалительных клеток, формирование паннуса, разрушение хряща и эрозию кости (фигура 9, средняя панель). Мыши в группе 3 после лечения hfB3-292S имели нормальную структуру суставов, отсутствие инфильтрации воспалительных клеток, отсутствие эрозии или повреждения хряща или кости (фигура 9, нижняя панель).
Эти данные показывают, что hfB3-292S являлся значительно эффективным для предотвращения воспаления и повреждения суставов в этой модели на мышах CAIA, демонстрируя терапевтическую полезность белка hfB3-292S для ревматоидного артрита.
Пример 17. Получение и характеризация экспрессирующей hfB3-292S-Fc конструкции и белка hfB3-292S-Fc
Стабильную линию клеток, экспрессирующую белок hfB3-292S-Fc (SEQ ID NO:22) получали опосредованной PEI трансфекцией и отбором с лекарственным средством клеток 293, как описано в примере 3. Отобранные с лекарственным средством клетки культивировали при 2×106 клеток/мл в течение 72 часов. Затем исследовали экспрессию белка hfB3-292S-Fc подверганием 2 мкл супернатанта культуры клеток невосстанавливающему SDS-PAGE и анализу Вестерн-блоттингом. Две полосы белка hfB3-292S-Fc детектировали посредством специфического антитела козы против фактора B. (Данные не представлены.) Без желания быть связанными теорией, эти две полосы белка hfB3-292S-Fc могут представлять мономеры и димеры белка.
hfB3-292S-Fc очищали на колонке с белком A. Биологическую активность очищенного белка hfB3-292S-Fc исследовали анализом гемолитической активности, как описано в примере 7. Как показано в таблице 6, белок hfB3-292S-Fc ингибировал активность альтернативного пути комплемента зависимым от дозы образом.
-0,0
Пример 18. Усеченный по C-концу hfB3-292S
Поучали конструкцию для экспрессии гена, экспрессирующую усеченную форму hfB3-292S с делецией 284 аминокислот на C-конце (домен сериновой протеазы). Молекула обозначена как hfB3-292SN480, составляющая 480 N-концевых аминокислот из hfB3-292S (аминокислоты 1-480 из SEQ ID NO:2 или аминокислоты 26-480 из SEQ ID NO:2 после отщепления секретирующего пептида). Последовательность ДНК экспрессирующей hfB3-292SN480 конструкции показана на SEQ ID NO:24, где нуклеотиды 1064-2509 представляют собой кодирующую последовательность для hfB3-292SN480. Клетки 293 FreeStyle трансфицировали экспрессирующей конструкцией и отбирали с G418, как описано ранее в примере 3. Культуральную среду устойчивых к G418 не клонированных клеток подвергали анализу Вестерн-блоттингом по экспрессии hfB3-292SN480 с использованием полноразмерного hfB3-292S в качестве контроля (левая дорожка). Как показано на фигуре 11, анализ Вестерн-блоттингом с моноклональным антителом, специфическим для hfB3-292S детектировал полосу приблизительно 55 кДа в культуральной среде линии клеток hfB3-292SN480 (правая дорожка), позволяя предполагать, что даже при делеции 280 аминокислот с C-конца hfB3-292S, 480 N-концевых аминокислот можно экспрессировать с соответствующим размером.
Анализ активности альтернативного пути комплемента проводили, как описано ранее, чтобы определить, может ли hfB3-292SN480 ингибировать активность альтернативного пути комплемента. Как показано на фигуре 13, культуральный супернатант от клеток, экспрессирующих hfB3-292SN480, ингибировал активность альтернативного пути комплемента зависимым от дозы образом. Это демонстрирует, что фрагменты hfB3-292S еще могут сохранять способность ингибировать активность комплемента и таким образом, их можно использовать так же, как описано в настоящем документе для hfB3-292S (SEQ ID NO:2)
Пример 19. Мономерный hfB3-292S/Fc слитый белок hfB3-292S/Fc-mono
Разработана конструкция для экспрессия гена, кодирующего полноразмерный hfB3-292S, «слитый» с Fc IgG4 человека. hfB3-292S представляет собой мономер при продукции в клетках млекопитающих, таких как клетки человека, как описано ранее, например, см. фигуру 3, описанную в примере 6, где показано, что hfB3-292S детектировали как одну полосу с MW приблизительно 100 кДа в невосстанавливающих условиях, позволяя предполагать, что hfB3-292S представляет собой мономер. Два цистеина в шарнирной области Fc IgG4 человека подвергали мутации для обеспечения того, чтобы слитый белок был мономером и сохранял биологическое свойство hfB3-292S ингибировать активность комплемента. Два цистеина подвергали мутации посредством замены каждого из них на серин. Этот слитый белок из hfB3-292S и мутантного Fc IgG4 человека обозначили hfB3-292S/Fc-mono. Последовательность ДНК экспрессирующей этот слитый белок конструкции показана на SEQ ID NO:26. Соответствующая аминокислотная последовательность hfB3 292S/Fc-mono показана на SEQ ID NO:25, где аминокислоты 1-764 представляют собой область hfB3-292S, и аминокислоты 765-1003 представляют собой область Fc IgG4 человека, где аминокислоты 782 и 785 представляют собой сериновые аминокислоты, которыми заменены цистеиновые остатки, обнаруженные в природном Fc IgG4 человека.
Клетки 293 FreeStyle человека трансфицировали конструкцией для экспрессии гена hfB3-292S/Fc-mono (SEQ ID NO:26). Затем клетки подвергали отбору с G418. Культуральную среду устойчивых к лекарственному средству клеток подвергали анализу слитого белка Вестерн-блоттингом. Как показано на фигуре 12, полосу приблизительно 115 кДа детектировали посредством очищенного антитела козы против фактора B человека в этом анализе невосстанавливающим SDS-PAGE и Вестерн-блоттингом. Две более высоких полосы, наиболее вероятно, представляли собой агрегаты мономерного слитого белка. Данные позволяют предполагать, что мономерный слитый белок из hfB3-292S и Fc IgG4 человека (hfB3-292S/Fc-mono) успешно экспрессировался в клетках млекопитающих, и что большинство слитого белка, по-видимому, являлось мономерным.
Для исследования, сохранил ли hfB3-292S/Fc-mono свойство hfB3-292S блокировать активность альтернативного пути комплемента, анализ активности альтернативного пути комплемента проводили, как описано ранее. Как показано на фигуре 14, культуральный супернатант от клеток, продуцирующих hfB3-292S/Fc-mono, ингибировал активность альтернативного пути комплемента зависимым от дозы образом, позволяя предполагать, что активность ингибирования комплемента hfB3-292S не была утрачена этим мономерным слитым белком hfB3-292S/Fc-mono.
Полное содержание всех публикаций, патентов и патентных заявок, упомянутых в этом описании, приведено в этом описании в той же степени, как если бы было конкретно и отдельно указано, что содержание каждой отдельной публикации, патента или патентной заявки приведено в настоящем документе в качестве ссылки.

























































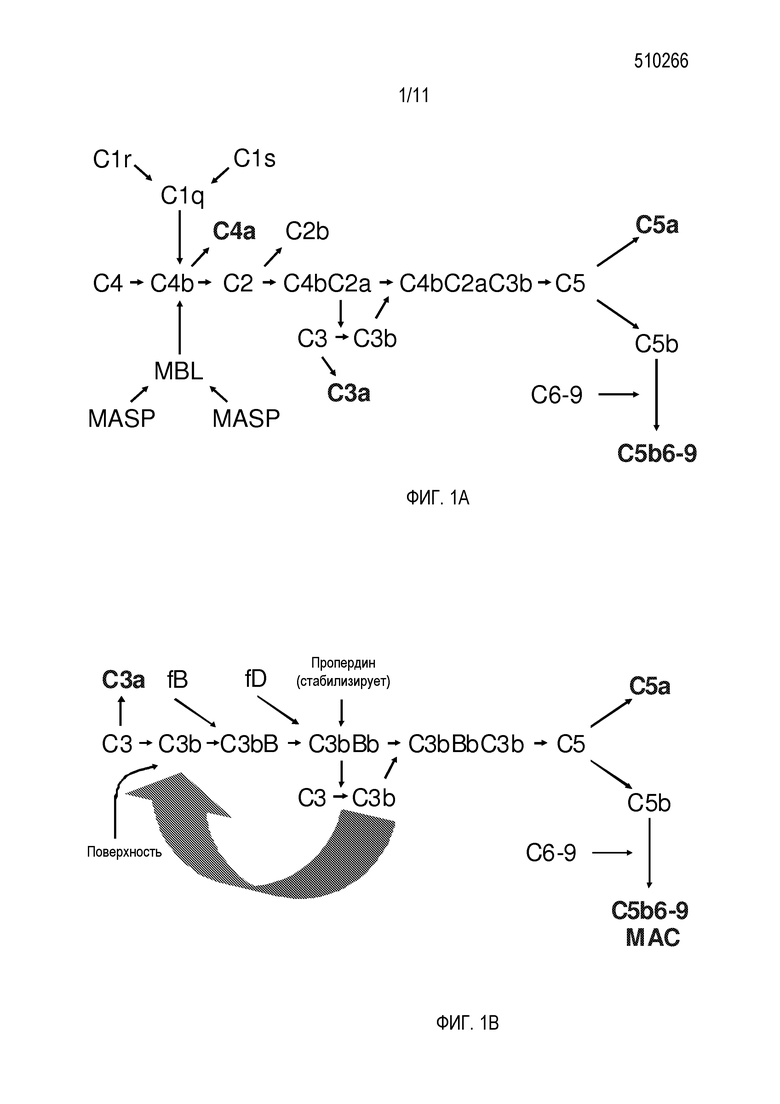
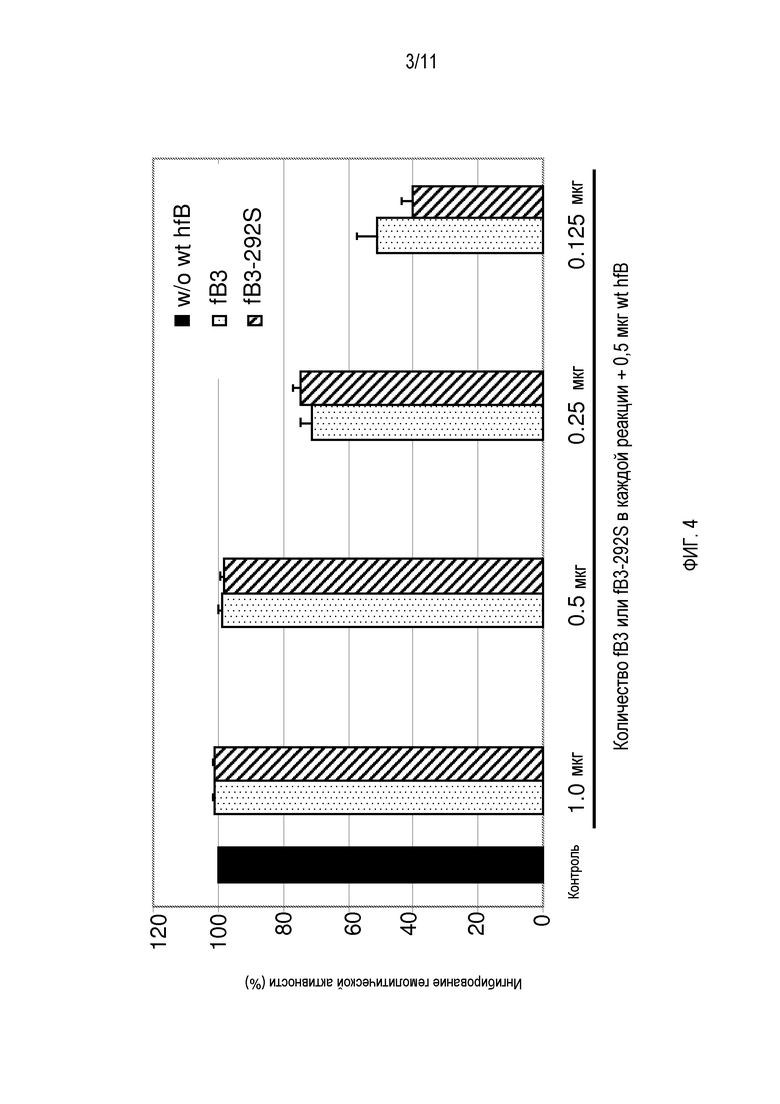
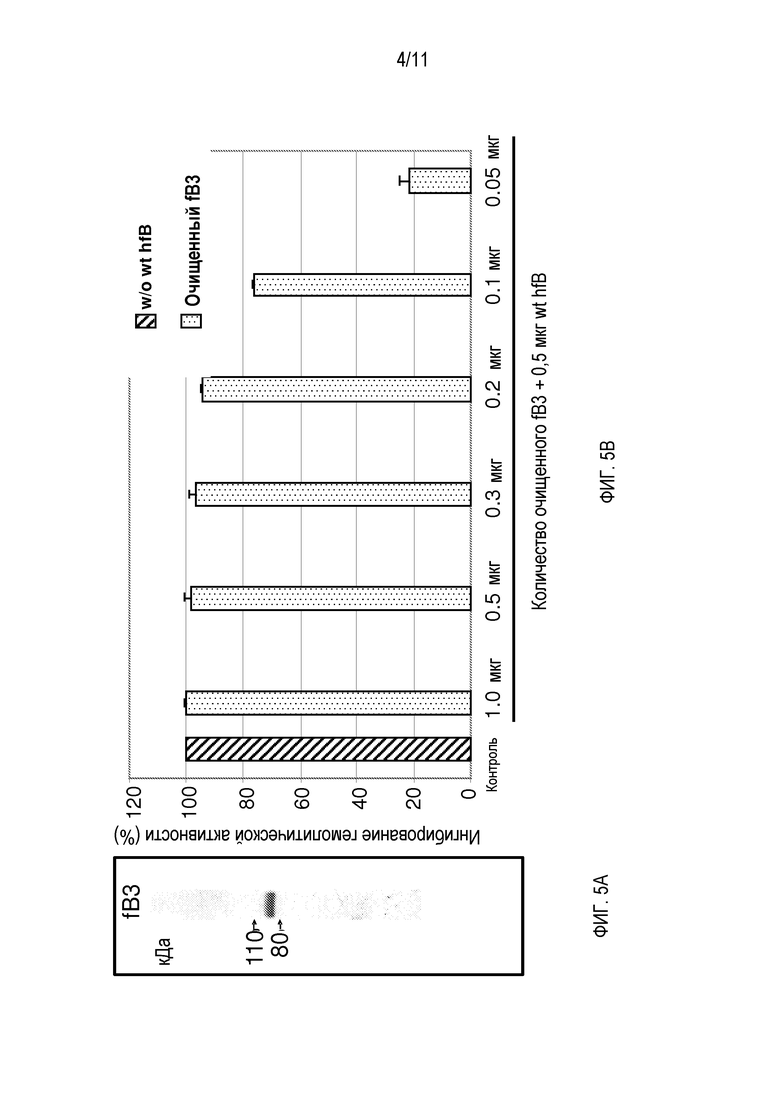
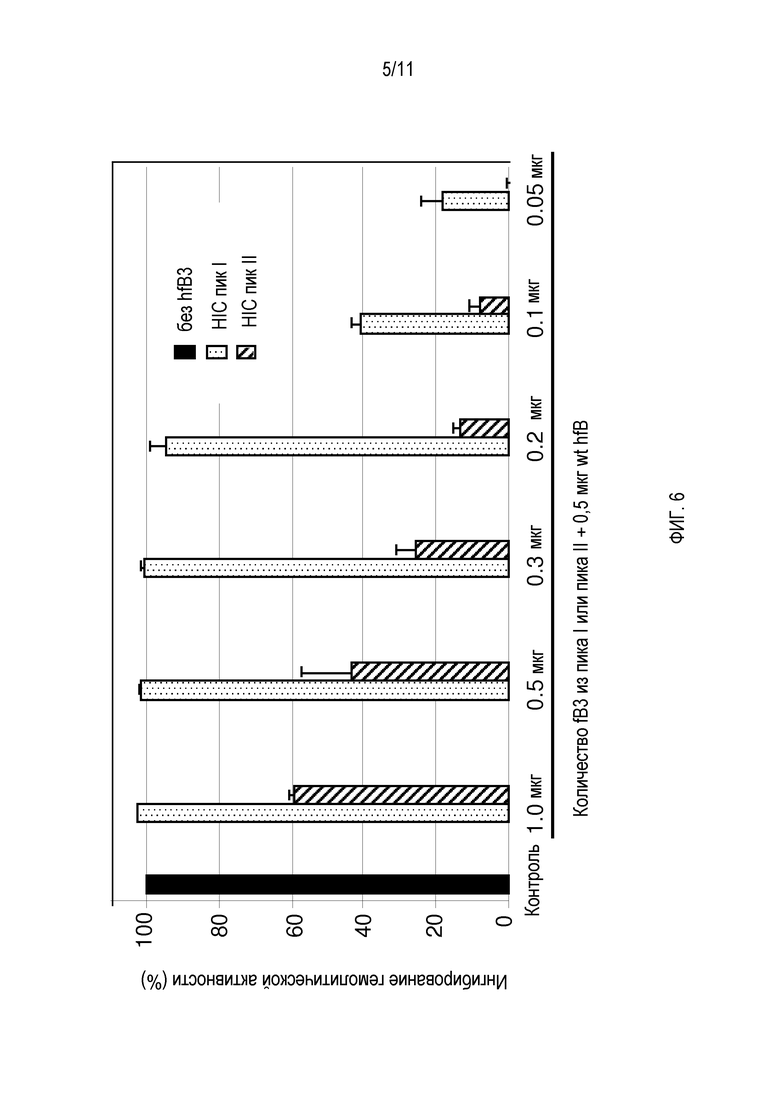
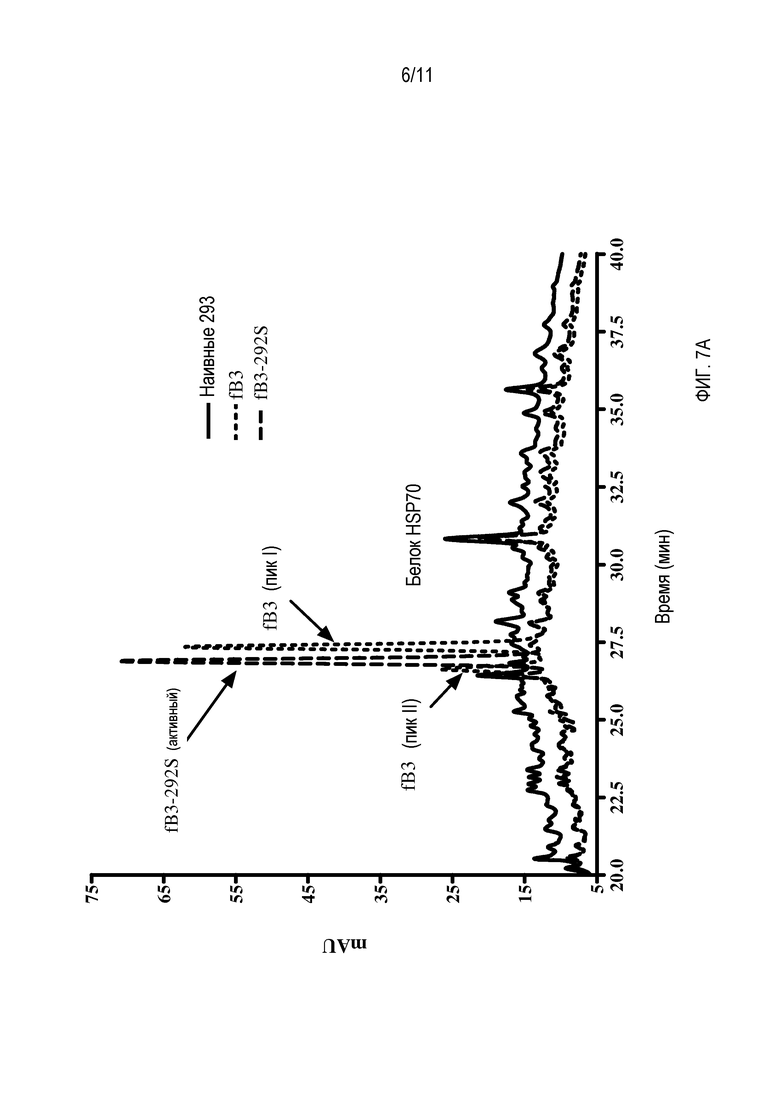
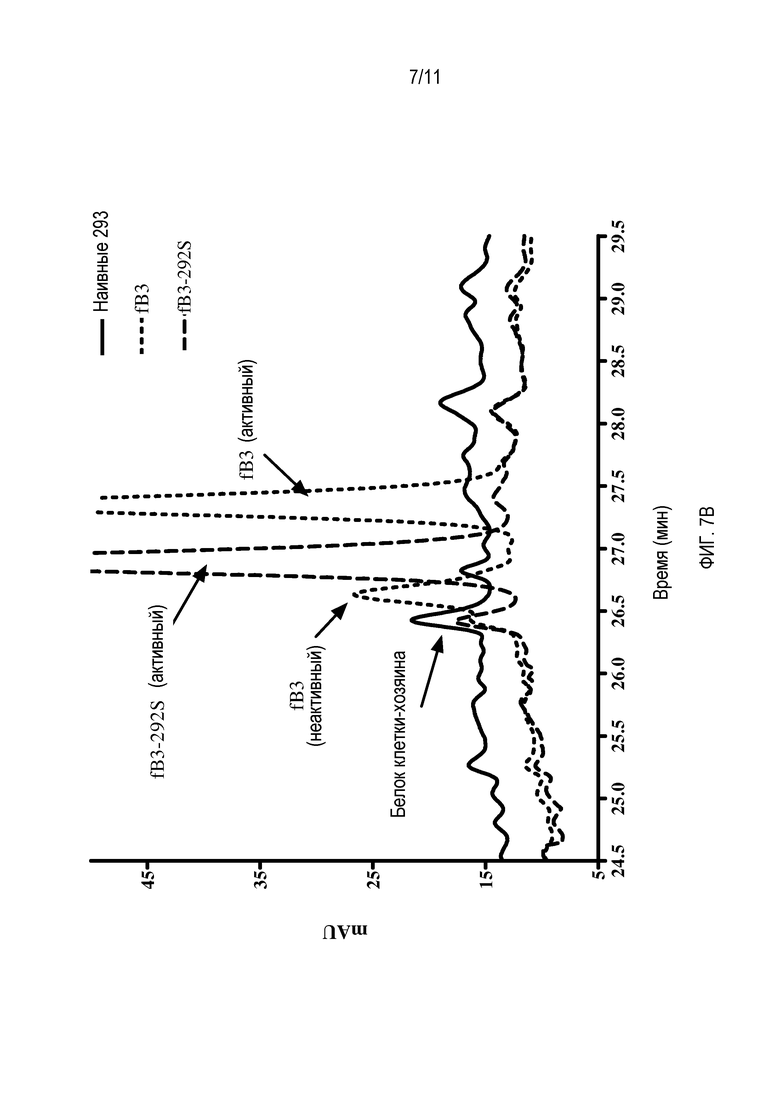
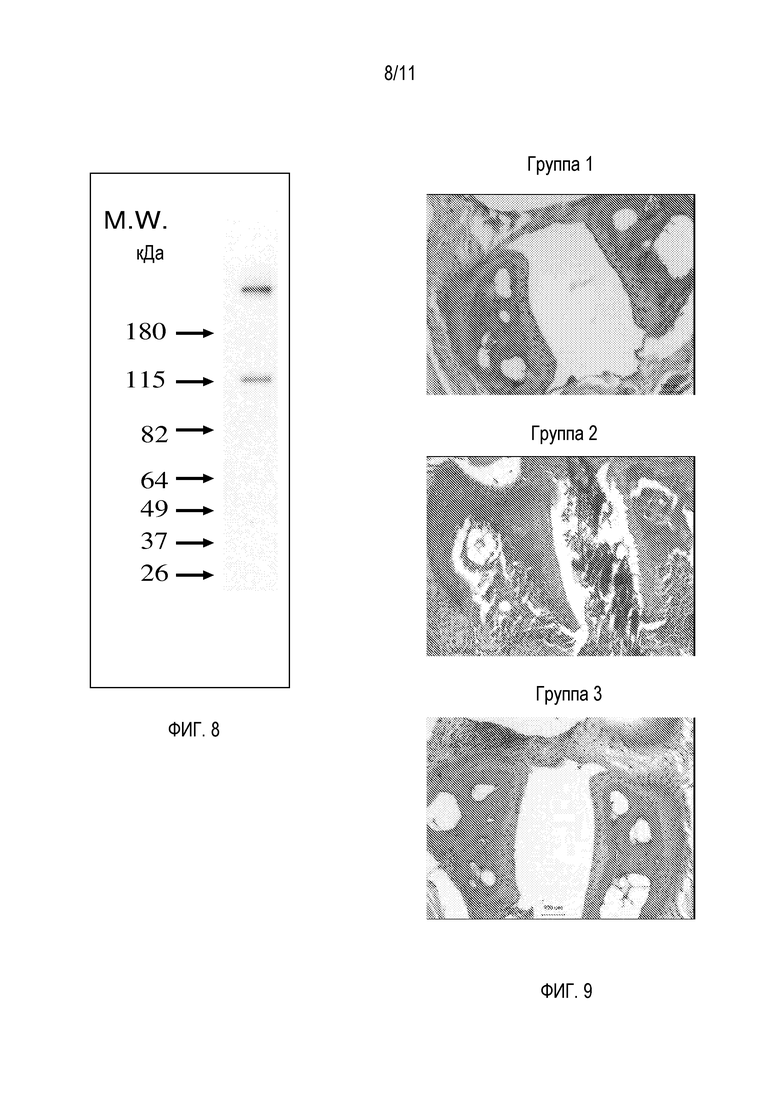
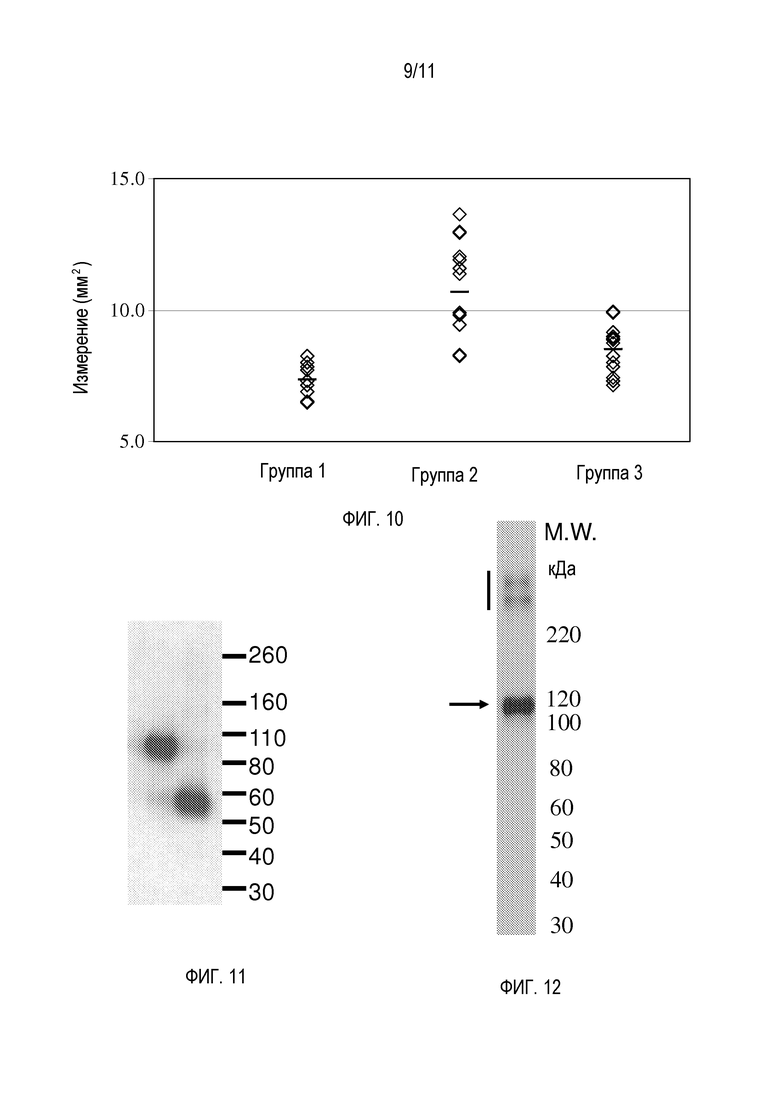
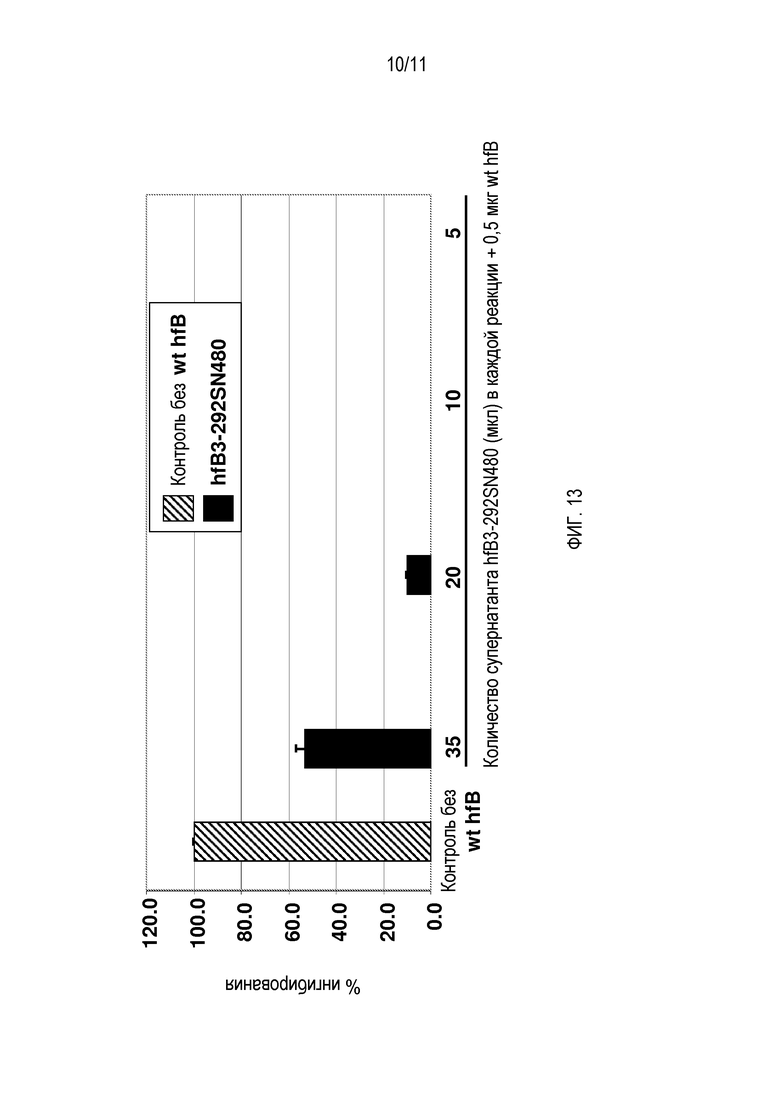
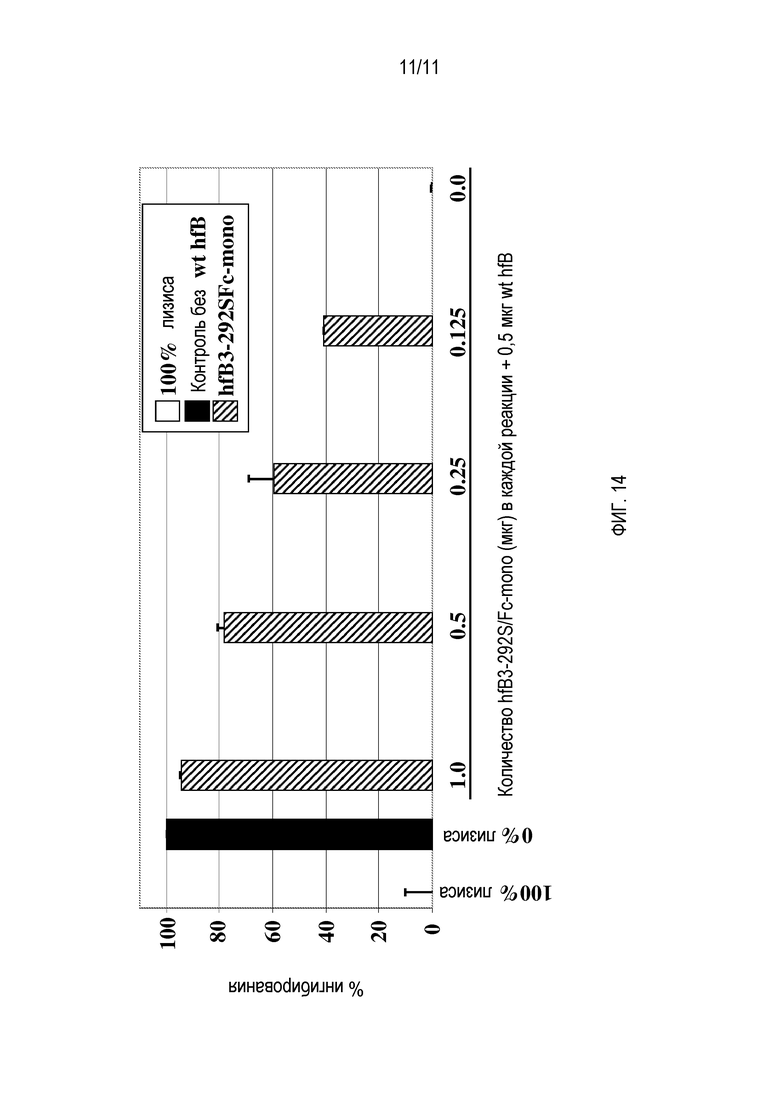
Изобретение относится к области биотехнологии, конкретно к аналогам фактора комплемента B, и может быть использовано в медицине для лечения заболевания, опосредованного активацией альтернативного пути комплемента. Получают полипептид, состоящий из hfB3-292S (аминокислоты 1-764 или 26-764 в SEQ ID NO: 2), hfB3-292SN480 (аминокислоты 1-480 или 26-480 в SEQ ID NO: 2) или hfB3-292S-Fc (аминокислоты 1-990 или 26-990 в SEQ ID NO: 22). Изобретение позволяет эффективно ингибировать активность альтернативного пути комплемента при использовании полученного полипептида, кодирующей его нуклеиновой кислоты или вектора экспрессии. 5 н. и 7 з.п. ф-лы, 14 ил., 6 табл., 19 пр.
1. Полипептид для ингибирования активности альтернативного пути комплемента, где полипептид выбран из группы, состоящей из hfB3-292S (аминокислоты 1-764 или 26-764 в SEQ ID NO: 2), hfB3-292SN480 (аминокислоты 1-480 или 26-480 в SEQ ID NO: 2) и hfB3-292S-Fc (аминокислоты 1-990 или 26-990 в SEQ ID NO: 22).
2. Нуклеиновая кислота для ингибирования активности альтернативного пути комплемента, кодирующая полипептид по п. 1.
3. Вирусный вектор экспрессии, содержащий нуклеиновую кислоту по п. 2.
4. Вирусный вектор экспрессии по п. 3, где указанный вирусный вектор выбран из группы, состоящей из ретровирусного вектора, лентивирусного вектора, аденовирусного вектора, вектора из вируса герпеса, вектора из вируса гепатита, вектора из вируса SV40, вектора AAV, вектора EBV и вектора NDV.
5. Вирусный вектор по п. 4, где указанный лентивирусный вектор представляет собой вектор BIV, HIV, EIAV, SIV или FIV.
6. Фармацевтическая композиция для лечения заболевания, опосредованного активацией альтернативного пути комплемента, содержащая (i) полипептид по п. 1, нуклеиновую кислоту по п. 2, вирусный вектор по любому из пп. 3-5 или любую их комбинацию в количестве, достаточном для ингибирования активности альтернативного пути комплемента; и (ii) фармацевтически приемлемый носитель.
7. Способ лечения заболевания, опосредованного активацией альтернативного пути комплемента у пациента, включающий введение пациенту полипептида по п. 1, нуклеиновой кислоты по п. 2, вирусного вектора по любому из пп. 3-5 или любой их комбинации в количестве, достаточном для ингибирования активности комплемента, где полипептид ингибирует активность комплемента.
8. Способ по п. 7, где заболевание, опосредованное активацией альтернативного пути комплемента, представляет собой заболевание глаза.
9. Способ по п. 8, где фармацевтическую композицию вводят в глаз.
10. Способ по п. 9, где фармацевтическую композицию вводят интравитреальной инъекцией, субретинальной инъекцией, инъекцией в переднюю камеру глаза, инъекцией или местным введением в роговицу, субконъюнктивальной инъекцией, субтеноновой инъекцией или посредством глазных капель.
11. Способ по п. 7, где заболевание, опосредованное активацией альтернативного пути комплемента, представляет собой дегенерацию желтого пятна, связанную с возрастом дегенерацию желтого пятна (AMD), географическую атрофию, влажную AMD, инфаркт миокарда, сухую AMD, образование друз, артрит, инсульт, ишемически-реперфузионное повреждение, диабетическую ретинопатию, витреоретинопатию, травматическое повреждение органов, воспаление роговицы, неоваскуляризацию роговицы, увеит, глазную гипертензию или глаукому.
12. Способ по п. 7, где заболевание, опосредованное активацией альтернативного пути комплемента, выбрано из группы, состоящей из атеросклероза, гиперчувствительности дыхательных путей, связанных с иммунитетом заболеваний, связанных с аутоиммунитетом заболеваний, волчаночного нефрита, системной красной волчанки (SLE), артритом, ревматологических заболеваний, антифосфолипидного синдрома, I/R повреждения кишечника и почек, астмы, атипичного гемолитико-уремического синдрома, мембранопролиферативного гломерулонефрита типа II, непролиферативного гломерулонефрита, выкидыша, повреждения головного мозга, посттравматического повреждения органов, постинфарктного повреждения органов, васкулита, наследственного ангионевротического отека, пароксизмальной ночной гемоглобинурии, острого нарушения мозгового кровообращения, болезни Альцгеймера, отторжения трансплантата, инфекций, сепсиса, септического шока, синдрома Шегрена, миастении, опосредованных антителами заболеваний кожи, сахарного диабета типа I и типа II, синдрома устойчивости к инсулину, гестационного диабета, тиреоидита, идиопатической тромбоцитопенической пурпуры и гемолитической анемии, невропатий, рассеянного склероза, повреждения при искусственном кровообращении, узелкового полиартериита, пурпуры Шенлейна - Геноха, сывороточной реакции, заболевания Гудпасчера, системного некротизирующего васкулита, постстрептококкового гломерулонефрита, идиопатического пульмонарного фиброза, мембранного гломерулонефрита, синдрома острого инфаркта легкого, синдрома острой дыхательной недостаточности у взрослых и реперфузии.
| US 20100120665 A1, 13.05.2010 | |||
| LEPOCK J | |||
| R | |||
| et al | |||
| Contribution of conformational stability and reversibility of unfolding to the increased thermostability of human and bovine superoxide dismutase mutated at free cysteines, 1990, J | |||
| Biol | |||
| Chem., v | |||
| САННЫЙ ВЕЛОСИПЕД С ВЕДУЩИМ КОЛЕСОМ, СНАБЖЕННЫМ ШИПАМИ | 1921 |
|
SU265A1 |
| Скоропечатный станок для печатания со стеклянных пластинок | 1922 |
|
SU35A1 |
| ГИДРАВЛИЧЕСКАЯ ПЕРЕДАЧА | 1929 |
|
SU21612A1 |
| HOURCADE D | |||
| et al | |||
| Analysis of the short consensus repeats of human complement factor B by site-directed mutagenesis, 1995, J | |||
| Biol | |||
| Chem., v | |||
| Приспособление для уменьшения дымовой тяги паровоза | 1920 |
|
SU270A1 |
| Нивелир для отсчетов без перемещения наблюдателя при нивелировании из средины | 1921 |
|
SU34A1 |
| Паровая форсунка | 1930 |
|
SU19716A1 |
| GOLD B | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Способ сужения чугунных изделий | 1922 |
|
SU38A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Приспособление для отопления печей нефтью | 1922 |
|
SU458A1 |
| PETERSEN M.T | |||
| et al., Amino acid neighbours and detailed conformational analysis of cysteines in proteins, Protein Eng., 1999, v | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| ЯКОРЬ ДЛЯ МАШИНЫ ВЫСОКОЙ ЧАСТОТЫ | 1922 |
|
SU535A1 |
| TATARA Y | |||
| et al., A Single Free Cysteine Residue and Disulfide Bond Contribute to the Thermostability of Aspergillus saitoi 1,2-α-Mannosidase, BIOSCIENCE, BIOTECHNOLOGY, AND BIOCHEMISTRY, 2005, v | |||
| Способ приготовления пищевого продукта сливкообразной консистенции | 1917 |
|
SU69A1 |
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Прибор для измерения профилей самолетных частей | 1925 |
|
SU2101A1 |
| LEE J | |||
| et al., Structural Basis of Conserved Cysteine in the Fibroblast Growth Factor Family: Evidence for a Vestigial Half-Cystine, J | |||
| Mol | |||
| Biol., 2009, v | |||
| Транспортир | 1922 |
|
SU393A1 |
| MALIK T.H | |||
| et al, The alternative pathway is critical for pathogenic complement activation in endotoxin- and diet-induced atherosclerosis in low-density lipoprotein receptor-deficient mice, Circulation, 2010, v | |||
| Схема обмотки ротора для пуска в ход индукционного двигателя без помощи реостата, с применением принципа противосоединения обмоток при трогании двигателя с места | 1922 |
|
SU122A1 |
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| HOLERS V | |||
| M., The spectrum of complement alternative pathway-mediated diseases, Immunological Reviews, 2008, v | |||
| Способ исправления пайкой сломанных алюминиевых предметов | 1921 |
|
SU223A1 |
| ТКАЦКИЙ СТАНОК | 1920 |
|
SU300A1 |
| RICKLIN D | |||
| et al, Complement: a key system for immune surveillance and homeostasis, Nature Immunology, 2010, v | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Гильза для удерживания скрепляющих подошвенные части винтов | 1916 |
|
SU785A1 |
| ПРИМЕНЕНИЕ ИНГИБИТОРОВ ПУТИ КОМПЛЕМЕНТА ДЛЯ ЛЕЧЕНИЯ ГЛАЗНЫХ БОЛЕЗНЕЙ | 2006 |
|
RU2417099C2 |

