Область техники, к которой относится изобретение
Настоящее изобретение относится к способам отбора мутантных бактерий, вырабатывающих повышенные уровни фолата. В этих способах в качестве агента отбора использовали антифолат метотрексат (МТХ). Также обеспечены пищевые композиции и композиции пищевых добавок, содержащие мутантные бактерии или производные, или экстракты данных бактерий.
Предпосылки к созданию изобретения
Аналоги фолата и, в частности, ингибиторы дигидрофолатредуктазы были предметом изучения в течение многих лет вследствие их потенциальной роли в химиотерапии рака и в качестве агентов против малярийного паразита. Одним из первых аналогов фолата, описанных в литературе, является метотрексат от фирмы Lederle Laboratories. Это соединение применяли уже в 1948 году для лечения острой лейкемии (Hitchings, G. H., Jr., 1989, In Vitro Cell Dev Biol 25:303-10).
Действие метотрексата (МТХ) основано на конкурентном ингибировании фермента-мишени дигидрофолатредуктазы. Этот фермент необходим для продукции и рециклинга тетрагидрофолата (THF) с использованием в качестве субстрата дигидрофолата (DHF). Специфичность метотрексата можно объяснить небольшими структурными различиями дигидрофолатредуктаз у различных видов (Chang et al., 1978, Nature 275:617-24). Метоторексат препятствует выработке клетками достаточного количества тетрагидрофолата для биосинтеза метионина, ДНК и РНК. Тем не менее в присутствии пуринов, тимидина, глицина, метионина и пантотеновой кислоты (все являются метаболитами, которым необходим тетрагидрофолат для их собственного биосинтеза) потребность клеток синтезировать тетрагидрофолат может быть снижена (Harvey, R.J., 1973, J Bacteriol 114:309-22). Несмотря на это клеточный рост всегда каким-либо путем будет затруднен посредством МТХ, поскольку TNF имеет важное значение для формирования метионил-тРНКfmet, соединения, которое необходимо для инициации синтеза белка, вне зависимости от присутствия метаболитов, зависимых от фолата (Baumstark et al., 1977, J Bacteriol 129:457-71).
Молочнокислые бактерии, такие как Lactococcus lactis, Lactobacillus plantarum и Lactobacillus casei, проявляют высокую степень изменчивости их реакции на метотрексат и другой аналог фолата, триметоприм. Известно, что L.lactis является нечувствительным к обоим антифолатам (Leszczynska et al., 1995, Appl Environ Microbiol 61:561-6). Для многочисленных микроорганизмов показано, что продолжительное воздействие антифолатами приводит к отбору устойчивых клеточных типов. Описано несколько основных механизмов, объясняющих механизм устойчивости к метотрексату (МТХ) эукариотических клеточных линий и бактерий. Tamura et al. (1997, Microbiology 143:2639-46) изучали механизмы, приводящие к устойчивости к МТХ у Enterococcus hirae. Они обнаружили некоторые фенотипические эффекты, включая (i) повышенную активность редуктазы фолиевой кислоты (FAR), (ii) повышенную активность дигидрофолатредуктазы (DHFR), (iii) сниженный синтез и внутриклеточное удержание МТХ, содержащего два остатка глутамила, (iv) уменьшенное поглощение МТХ, сопровождаемое сниженным поглощением фолатов; и (v) уменьшение фолат-связывающей способности. Форма фолата, присутствующая в среде в процессе развития устойчивости, влияет на активности DHFR и FAR и транспорт фолатов.
Однако хотя известно, что воздействие с помощью МТХ можно использовать для отбора клеток, имеющих повышенную устойчивость к МТХ, в известном уровне техники не было выявлено связи между устойчивостью к МТХ и уровнями фолатов в данных клетках. В отличие от механизмов устойчивости к МТХ, описанных выше, авторы настоящего изобретения с удивлением обнаружили, что бактериальные клетки обладают другим способом выработки устойчивости к МТХ. Этим механизмом является выработка высоких уровней фолата. Эту находку можно использовать при скрининге и селекции (спонтанных) мутантных бактерий, вырабатывающих высокие уровни фолата, используя в качестве агента отбора метотрексат. Такие бактерии, в особенности пищевые бактерии, можно использовать для обогащения продуктов питания фолатом. Фолат представляет собой растворимый витамин В, необходимый для правильного роста клеток и функционирования. Многие продукты питания, такие как хлеб, зерновые продукты, молочные продукты и тому подобное, обогащены фолиевой кислотой для обеспечения адекватного потребления фолата. Мутантные бактерии по настоящему изобретению облегчают обогащение продуктов питания естественным фолатом посредством, например, ферментации с использованием данных мутантных бактерий.
Определения
«Молочнокислые бактерии», как используется в данном описании, представляют собой бактерии, которые вырабатывают молочную кислоту в качестве конечного продукта ферментации, такие как бактерии родов Lactobacillus, Lactococcus, Streptococcus и Bifidobacterium.
Термины бактериальный «штамм» или «изолят», используемые в данном описании, являются равнозначными и означают бактерию, которая остается генетически неизменной после роста или размножения. Многообразие аналогичных бактерий включено.
Термин «мутантная бактерия» или «мутантный бактериальный штамм или изолят» относится к природной (спонтанной, случающейся в природе) мутантной бактерии или индуцированной мутантной бактерии, содержащей в своем геноме (ДНК) одну или несколько мутаций, которые отсутствуют в ДНК дикого типа. «Индуцированный мутант» представляет собой бактерию, в которой данная мутация была вызвана в результате воздействия, проводимого человеком, например, в результате обработки химическими мутагенами, УФ- или гамма-лучами и тому подобное. В отличие от этого, «природный» или «спонтанный мутант» мутирует без помощи человека. Мутантные бактерии здесь не являются ГМО, а именно не модифицированы с помощью методов рекомбинантной ДНК.
Выражения «штамм дикого типа» или «изолят дикого типа» относятся к немутантной форме бактерии, такой как обнаружена в природе.
«Фолат-зависимые метаболиты» относятся к метаболитам или предшественникам биосинтеза или катаболизма фолата, которые способны маскировать чувствительность клеток к МТХ, когда присутствуют в окружающей среде.
Термин «ингибирующее количество MTX» относится к количеству, которое необходимо для существенного ингибирования клеточного роста. Как правило, для прокариотических клеток, по меньшей мере, количество МТХ около 1,25 мг/л питательной среды является ингибирующим для чувствительных к МТХ клеток.
Термин «общий фолат» относится к внеклеточным (секретируемым) и внутриклеточным уровням фолата, вырабатываемого штаммом.
«Пищевые» микроорганизмы представляют собой, в частности, организмы, которые считаются безвредными, когда проглатываются человеком или животным.
Термин «включающий в себя (содержащий)» должен интерпретироваться как определение наличия заявленных частей, стадий или компонентов, но не исключает наличия одной или нескольких дополнительных частей, стадий или компонентов. Композиция, содержащая бактериальный штамм Х, может, таким образом, содержать дополнительные штаммы, другие компоненты и тому подобное.
Кроме того, ссылка на элемент в единственном числе не исключает возможности, что присутствует более одного элемента, если только по контексту определенно не требуется, чтобы присутствовал один и только один из данных элементов. Таким образом, использование определения в единственном числе обычно означает «по меньшей мере, один».
Подробное описание изобретения
При проведении проверки того, имеют ли рекомбинантные бактерии, гиперпродуцирующие фолат, модифицированную восприимчивость к МТХ, было обнаружено, что целый ряд штаммов дикого типа L.plantarum, которые инкубировали в течение более чем приблизительно 40 часов с ингибирующим количеством МТХ (1,25 мг/л), не ингибировались посредством МТХ, а имели скорость роста, схожую с таковой в среде, в которой отсутствует МТХ. При анализе уровней фолата в этих штаммах приблизительно 5% этих колоний проявляли значительно более высокую продукцию фолата по сравнению с (немутантным) штаммом дикого типа. Общий фолат был выше на 4%-63% по сравнению с (средними) уровнями дикого типа. Это наблюдение показывает, что МТХ можно использовать для эффективного отбора природных (не ГМО) бактерий, гиперпродуцирующих фолат. Устойчивость к МТХ поддерживалась, даже когда выращивали более 50 поколений данных штаммов на среде, в которой отсутствует МТХ, что показывает, что данная устойчивость появляется вследствие (спонтанных) стабильных мутаций в бактериальном геноме.
Тот же самый эффект, что повышенная продукция фолата приводит к высокой устойчивости к МТХ, был обнаружен у рекомбинантных штаммов L.plantarum, которые были трансформированы с помощью полного кластера генов биосинтеза фолата под контролем активного промотора. Эти рекомбинантные штаммы продуцировали высокие уровни фолата и были устойчивы к МТХ, когда выращивались в среде, в которой отсутствуют фолат-зависимые метаболиты. Не ограничивая объем данного изобретения, предполагают, что внутриклеточные уровни дигидрофолата конкурируют с аккумулированным МТХ за активный центр фермента дигидрофолатредуктаза (DHFR).
Эту находку применили для развития метода быстрого скрининга, в основе которого лежит рост в присутствии МТХ, для отбора бактериальных штаммов, гиперпродуцирующих фолат. Данные гиперпродуценты фолата, которые были выявлены в данном исследовании, проявляли большое повышение уровней фолата по сравнению с продуцентами фолата дикого типа. Уровни общего фолата колебались в пределах от 20 до 60 раз больше уровней дикого типа. Внутриклеточный фолат, обнаруженный у гиперпродуцентов фолата, был в 10-20 раз выше, по сравнению с уровнями, обнаруженными у штаммов дикого типа. Эти повышенные внутриклеточные уровни фолата приводят к пониженной чувствительности (а именно устойчивости) по отношению к МТХ. Это объясняет, почему МТХ значительно не влиял на рост штаммов, гиперпродуцирующих фолат. Чувствительность дикого типа по отношению к МТХ наблюдали только когда все метаболиты, которые связаны с фолатом, были удалены из данной среды.
Бактерии по данному изобретению
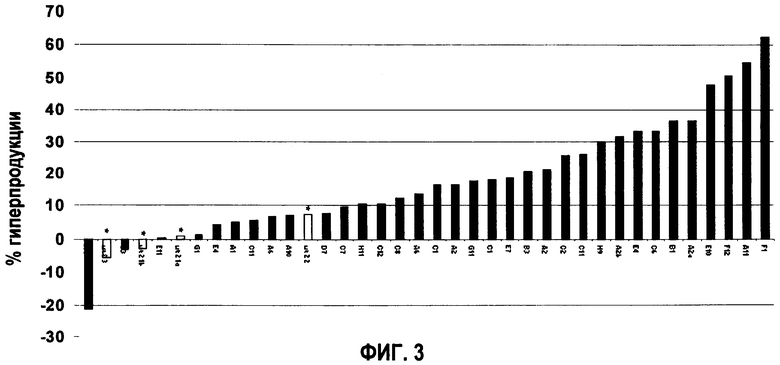
Таким образом, в одном варианте осуществления данное изобретение относится к мутантным бактериям, которые содержат повышенное количество внутриклеточного и/или внеклеточного фолата по сравнению с диким типом, и скорость роста (µ) которых, по меньшей мере, составляет 0,1 ч-1 при выращивании в среде, в которой содержится (по меньшей мере) 1,25 мг/л метотрексата и отсутствуют фолат-зависимые метаболиты (то есть они считаются «устойчивыми» к MTX). Предпочтительно при этих условиях роста скорость роста данной мутантной бактерии значительно не отличается от таковой штамма дикого типа при выращивании штамма дикого типа в такой же среде, в которой отсутствует МТХ (то есть «нормальный рост»). Наиболее важно, что данная мутантная бактерия имеет достоверный рост при концентрации МТХ, которая является ингибирующей для штамма дикого типа. «Ингибирующая концентрация» представляет собой концентрацию МТХ, которая приводит по существу к отсутствию роста штамма дикого типа. На фигуре 3, например, показано, что при концентрациях МТХ, равных 1,25, 1,5, 2,0 и 2,5 мг/л, скорость роста дикого типа близка к нулю (µ<0,05 ч-1), в то время как скорость роста (µ) штаммов, гиперпродуцирующих фолат, выше 0,1 ч-1, главным образом, приблизительно 0,15 ч-1. Данная среда для выращивания не должна быть дополнена фолат-зависимыми метаболитами, поскольку в их присутствии не обнаружено значительного различия устойчивости к МТХ, возможно потому, что штамм дикого типа компенсирует свои низкие урони фолата с помощью метаболитов фолата из среды. Подходящей средой для выращивания является, например, модифицированная среда CDM (среда определенного химического состава), в которой отсутствует глицин, инозин, оротовая кислота, тимидин, гуанин, аденин, урацил и ксантин.
Скорость роста может быть измерена разными способами. Например, в определенные моменты времени после инокуляции среды бактериальным штаммом или изолятом можно провести спектрофотометрическое считывание и вычислить скорость роста в динамике по времени.
Данная мутантная бактерия предпочтительно содержит повышенное количество внутриклеточного и/или внеклеточного фолата, по сравнению со штаммом дикого типа. Предпочтительно, количество общего фолата, по меньшей мере, приблизительно на 4%, 5%, 10%, 15%, 20%, 25%, 30%, 40%, 50%, более предпочтительно, по меньшей мере, приблизительно на 60%, 63% (или более) выше, чем среднее количество внутриклеточного фолата штамма (штаммов) дикого типа. Поскольку у некоторых бактериальных родов штаммы дикого типа показывают значимые вариации уровней внутриклеточного фолата, предпочтительно, чтобы уровни внутриклеточного фолата анализировали в некотором количестве различных штаммов дикого типа (например, 2, 3, 4 или более штаммов) и вычисляли средний уровень фолата.
Общее внутриклеточное или внеклеточное количество фолата, вырабатываемое бактериальным изолятом или штаммом, можно оценить и количественно определить посредством способов, известных в уровне техники, таких как микробиологический анализ L.casei, описанный Sybesma (см. пример 1.3), или его вариантов, посредством анализа методом ВЭЖХ или посредством других известных способов. Например, в микробиологическом анализе можно использовать индикаторный штамм (например, штамм L.casei), который выращен в присутствии фолата, продуцированного данным испытуемым штаммом. Если большие количества фолата вырабатываются испытуемым штаммом, индикаторный штамм растет хорошо, что приводит к высокому показателю OD после установленного периода роста. В отличие от этого, если испытуемым штаммом вырабатываются низкие количества фолата, будут определены более низкие значения OD.
Мутантная бактерия представляет собой, предпочтительно, спонтанный (природный) мутант, отобранный, например, с использованием способа по данному изобретению (см. далее). Данная бактерия относится к любому роду, дикий тип которого проявляет природную чувствительность к МТХ, например, L.plantarum, L.casei, или другие роды из группы кисломолочных бактерий. Быть чувствительным к МТХ означает, что рост бактерии, по существу, ингибируется посредством, по меньшей мере, 1,25 мг/л МТХ, когда данный штамм растет в среде, в которой отсутствуют фолат-зависимые метаболиты. Имеют ли бактериальные роды чувствительность к МТХ, можно без труда определить, выращивая данную бактерию в среде, дополненной МТХ и в которой отсутствуют фолат-зависимые метаболиты. Данная бактерия предпочтительно является пищевой бактерией и в одном варианте осуществления принадлежит к роду, выбранному из группы, состоящей из Lactobacillus, Lactococcus, Streptococcus, Bifidobacterium, Leuconostoc и Streptococcus.
Предпочтительно это пищевая кисломолочная бактерия. Это может быть род, выбранный из группы, состоящей из Lactobacillus reuteri, L.fermentum, L.acidophilus, L.crispatus, L.gasseri, L.johnsonii, L.casei, L.plantarum, L.paracasei, L.murinus, L.jensenii, L.salivarius, L.minutis, L.brevis, L.gallinarum, L.amylovorus, Lactococcus lactis, Streptococcus thermophilus, Leuconostoc mesenteroides, Lc.lactis, Pediococcus damnosus, P.acidilactici, P.parvulus, Bifidobacterium bifidum, B.longum, B.infantis, B.breve, B.adolescente, B.animalis, B.gallinarum, B.magnum и B.thermophilum.
В альтернативном варианте осуществления данная мутантная бактерия представляет собой индуцированный мутант, имеющий повышенный уровень фолата и являющийся устойчивым к МТХ. Такой мутант может быть получен с использованием такого же способа, который используют для отбора природных мутантов (описанный ниже), за исключением того, что перед скринингом включена стадия мутагенеза. Стадия мутагенеза может включать в себя один или несколько известных способов мутагенеза, таких как воздействие химическим и/или физическим мутагеном (например, N-метил-N'-нитро-N-нитрозогуанидин; УФ излучение, гамма-излучение и тому подобное). После мутагенеза данные бактерии выращивают в/на среде, содержащей ингибирующие количества МТХ, и изоляты, которые действительно выросли, отбирают и анализируют на содержание фолата.
В еще одном варианте осуществления мутантные бактерии по данному изобретению могут быть описаны по их характеру экспрессии генов. Это иллюстрируется обнаружением того, что устойчивые к метотрексату штаммы L.plantarum, продуцирующие, по меньшей мере, на 50% больше фолата, по сравнению с диким типом, показывают 2-кратную (или больше) гиперэкспрессию одного или нескольких генов folC в данном геноме. Ген folC кодирует фермент дигидрофолатсинтазу. В одном варианте осуществления бактерии по данному изобретению показывают по сравнению с диким типом гиперэкспрессию одного или нескольких генов, кодирующих ферменты пути биосинтеза фолата. В дополнение к гену, непосредственно вовлеченному в биосинтез фолата, также повышенные уровни транскрипции были обнаружены для следующих генов: FK (кодирует фруктокиназу), POX (пируватоксидаза), LOX (лактатоксидаза) и PDH (пируватдегидрогеназа). По сравнению с диким типом значимо сниженный уровень транскрипции обнаружен для: DAK (дигидроксиацетонфосфотрансфераза или глицеринкиназа). Таким образом, в одном варианте осуществления данного изобретения бактерия/бактерии по данному изобретению содержат значительно повышенные уровни транскриптов мРНК одного или нескольких генов из группы, кодирующих дигидрофолатсинтазу (ЕС 6.3.2.17), фруктокиназу (ЕС 2.7.1.4), пируватоксидазу (ЕС 1.2.3.3), и 6-фосфо-β-глюкозидазу (ЕС 3.2.1.86). Дополнительно или альтернативно, данные бактерии содержат значительно сниженные уровни транскриптов мРНК гена, кодирующего дигидроксиацетонфосфотрансферазу (=глицеринкиназа) (EC. 2.7.1.29).
Термины «активированный» или «супрессированный» означают повышенный уровень транскрипции мРНК или сниженный уровень транскрипции мРНК, по сравнению с геном штамма дикого типа.
Термин «значительно повышенный уровень транскрипта» или «значительно повышенный уровень транскрипции» относится к количеству транскрипта мРНК, по меньшей мере, приблизительно 2-кратному, предпочтительно, по меньшей мере, приблизительно 3-кратному, или даже приблизительно 4-кратному, по сравнению с уровнем транскрипта, обнаруженного в диком типе. Аналогично, выражение «значительно сниженный уровень транскрипта» или «уровень транскрипции» относится к количеству транскрипта мРНК, по меньшей мере, приблизительно в 2 раза, 3 раза и т.д. меньшему, чем обнаруженный в диком типе.
Уровни транскриптов мРНК одного или нескольких генов можно измерить и количественно определить, используя рутинные способы молекулярной биологии, такие как, например, нозерн-анализ и количественная ПЦР с обратной транскриптазой (ОТ-ПЦР). Поскольку информация о последовательности ДНК/кДНК для этих генов доступна, праймеры и зонды, пригодные для гибридизации с целевой мРНК или кДНК, доступны, и необходимо только проведение рутинных экспериментов для анализа уровней транскриптов. Например, последовательности нуклеиновой кислоты L.plantarum (геном которой секвенирован) можно использовать для распознавания (например, посредством анализа in silico), клонирования (например, используя способы на основе ПЦР, способы, основанные на гибридизации, и тому подобное) и/или секвенирования ортологичных генов других родов бактерий. Последовательности нуклеиновых кислот этих генов, или их части, можно затем использовать для создания праймеров или зондов для детекции и количественного определения генов-мишеней.
Необязательно, характер транскрипции (то есть повышенные или сниженные уровни транскриптов одного или нескольких генов, упомянутых выше) можно использовать в качестве критерия отбора. Например, можно сначала провести скрининг бактерий на предмет измененных уровней экспрессии при первоначальном скрининге, а затем анализировать устойчивость к метотрексату и/или уровни фолата (как описано ниже). Аналогично это можно использовать в качестве дополнительной стадии скрининга/отбора в способе, описанном ниже (например, между стадиями (b) и (c) или после стадии (d), или даже можно заменить стадии (a) и (b).
Характер транскрипции одного из устойчивых к метотрексату штаммов, описанных в данном документе (Lactobacillus plantarum NIZO B2550), сравнивали с таковым исходного штамма (Lactobacillus plantarum WCFS1 (полная последовательность L.plantarum WCFS1 была депонирована в базе данных EMBL под регистрационным № AL935263)). Это осуществили посредством микрочипов с цельной геномной ДНК Lactobacillus plantarum WCFS1. Образцы общей мРНК исходного штамма и мутанта, устойчивого к метотрексату, выделяли и затем обрабатывали обратной транскриптазой для синтеза флуоресцентно меченной кДНК. Образцы исходного штамма и мутанта метили дифференцированно. Затем смесь образцов меченой кДНК гибридизовали на ДНК-микрочипе для определения относительного содержания мРНК всех генов у мутанта по отношению к исходному штамму L.plantarum.
В одном варианте осуществления данные мутантные бактерии представляют собой бактериальные штаммы (или любые их производные), депонированные в банке данных NIZO Food Research, P.O. Box 20, 6710 BA Ede, the Netherlands at the CBS (Centraalbureau voor Schimmelcultures, Uppsalalaan 8, 3584 CT Utrecht, The Netherlands), согласно Будапештскому Соглашению под регистрационным номером CBS 117120.
Композиции по данному изобретению
В еще одном аспекте данное изобретение относится к композициям, содержащим мутантные бактерии, как описано в данном документе выше, или их производные или экстракты, а также к способам получения таких композиций. Бактерии по данному изобретению культивировали в соответствующих условиях, при желании восстанавливали из среды для культивирования и при желании готовили в виде композиции, пригодной для предполагаемого использования. Способы получения таких композиций, по существу, известны.
Предпочтительно, данные мутантные бактерии, их производные или экстракты используют для получения пищевых композиций и композиций пищевых добавок, содержащих природный фолат.
Предпочтительная композиция по данному изобретению пригодна для потребления субъектом, предпочтительно человеком или животным. Такие композиции могут быть представлены в форме пищевой добавки или цельного продукта, или пищевой композиции, в которой, помимо бактерии по данному изобретению, также содержится подходящая пищевая основа. Продукт питания или пищевая композиция, как понимается в данном описании, включают в себя не только твердые или полутвердые композиции, но также жидкости для потребления человеком или животным, то есть напитки или питье. Данный продукт питания или пищевая композиция может быть твердым, полутвердым и/или жидким продуктом питания или пищевой композицией и, в частности, может быть молочным продуктом, таким как ферментированный молочный продукт, включая, но не ограничиваясь ими, йогурт, напиток на основе йогурта или напиток из пахты. Такие продукты питания или пищевые композиции могут быть приготовлены по существу известным способом, например путем добавления одной или нескольких мутантных бактерий по данному изобретению в подходящий продукт питания или пищевую основу в подходящем количестве.
Также могут быть получены пищевые добавки, содержащие соответствующие количества одного или нескольких мутантных бактериальных штаммов, или их экстрактов (например, частично или по существу очищенный природный фолат). Пищевые добавки включают в себя, например, витаминные таблетки, пилюли, капсулы или порошки, содержащие рекомендованное дневное количество различных витаминов, включая фолат. Синтетическую фолиевую кислоту, обычно используемую в таких композициях, можно заменить на природный фолат по данному изобретению.
В еще одном предпочтительном варианте осуществления живые бактерии используют при или для получения продукта питания или пищевой композиции, например путем ферментации. При этом бактерии по данному изобретению можно использовать в известном по существу способе получения таких ферментированных продуктов питания или пищевых композиций, например посредством по существу известного способа получения ферментированных молочных продуктов с использованием кисломолочных бактерий. В таких способах бактериальные клетки по данному изобретению можно использовать в дополнение к обычно используемому микроорганизму, и/или можно заменить один или несколько, или часть обычно используемых микроорганизмов. Например, при приготовлении ферментированного молочного продукта, такого как йогурт или напитки на основе йогурта, пищевые мутантные бактерии по данному изобретению могут быть добавлены к, или использованы как часть заквасочной культуры, или могут быть соответственно добавлены во время такой ферментации. В этом случае «среда для выращивания» данных бактерий представляет собой пищевую среду для выращивания, например, на молочной основе.
В еще одном варианте осуществления используемые бактерии могут быть мертвыми (например, лизированными) или нежизнеспособными, или лиофилизированными.
Композиции, изготовленные с использованием одного или нескольких мутантных бактериальных штаммов по данному изобретению, или дополненные соответствующим количеством одного или нескольких данных штаммов, содержат увеличенное количество природного фолата, вырабатываемого данными бактериями. Один или несколько штаммов, которые являются высокими гиперпродуцентами фолата, могут быть смешаны или могут быть последовательно добавлены в данную композицию, или во время ее получения. Общее количество фолата, представленное в конечном продукте, можно проанализировать, используя, например, анализ методом ВЭЖХ.
Способы и применения согласно данному изобретению
Метотрексат используют для отбора бактерий, имеющих повышенные внутриклеточные уровни фолата и повышенные уровни общего фолата.
В одном варианте осуществления обеспечен способ отбора бактерий, имеющих повышенные внутриклеточные и/или уровни общего фолата. Этот способ включает следующие стадии:
(a) выращивание бактерий на (или в) среде, содержащей метотрексат, где данная среда не дополнена фолат-зависимыми метаболитами,
(b) отбор бактерий, имеющих скорость роста (µ), по меньшей мере, 0,1 в час,
(c) определение уровня фолата в бактериях, отобранных на стадии b,
(d) отбор бактерий из имеющих повышенные уровни фолата по сравнению с диким типом или другим подходящим контролем,
(e) необязательное повторение стадии (b), (c) и/или (d).
«Выращивание» на стадии (a) может также означать контактирование данных бактерий с такой средой для культивирования. Среда для культивирования может содержать различные концентрации MTX. Например, первую стадию отбора можно проводить при относительно низкой концентрации, например 1,25, 1,5, 2,0, 2,5 мг/л культуральной среды. Изоляты, выращиваемые при одной или нескольких из этих концентраций, затем могут быть скринированы непосредственно на предмет содержания фолата, или могут быть подвергнуты одному или нескольким дополнительным стадиям отбора с помощью МТХ, с использованием, например, более высоких концентраций МТХ, например 4, 5, 10 или более мг/литр. Строгость процесса отбора может измениться на более строгую, например, только изоляты с высокой скоростью роста при высокой концентрации, или на менее строгую, например, изоляты, проявляющие умеренную скорость роста при относительно низких концентрациях МТХ. Предпочтительно, на каждой стадии отбора с помощью МТХ, данные изоляты растут на, или в данной среде для культивирования, по меньшей мере, в течение 40 часов, более предпочтительно, в течение, по меньшей мере, 50 часов или более. Подходящие контроли, например изоляты дикого типа, всегда должны быть включены, поскольку мутанты отбираются на основании различий в росте между мутантом и контролем (например, диким типом). Контроль необязательно должен быть диким типом, но также может быть ранее отобранным штаммом. Оптимальная температура инкубирования, значение рН среды для культивирования и другие параметры могут быть без труда определены квалифицированным специалистом. Точные условия зависят от используемых бактериальных родов.
Предпочтительно, данный способ отработан таким образом, чтобы проводить одновременно скрининг большого количества изолятов, например, в 96-луночных микротитрационных планшетах или тому подобное.
Данный способ может быть применен ко всем бактериям, которые проявляют природную чувствительность к метотрексату. Используя аналог фолата метотрексат, специалист может отобрать и выделить бактерии, эффективно гиперпродуцирующие природный фолат, которые можно использовать для получения ферментированных пищевых продуктов с повышенной концентрацией фолата (натуральное обогащение или ферментационное обогащение), как описано выше.
Любая бактерия, получаемая данным способом и имеющая значительно повышенные уровни фолата, включена в объем притязаний.
Если не указано иначе, практическое применение данного изобретения будет задействовать стандартные способы молекулярной биологии, вирусологии, микробиологии и биохимии. Такие способы описаны в Sambrook et al. (1989) Molecular Cloning, A Laboratory Manual (2nd edition), Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press; in Sambrook and Russell (2001) Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Laboratory Press, NY; in Volumes 1 and 2 of Ausubel et al. (1994) Current Protocols in Molecular Biology, Current Protocols, USA; и в Volumes I and II of Brown (1998) Molecular Biology LabFax, Second Edition, Academic Press (UK); Oligonucleotide Synthesis (N. Gait editor); Nucleic Acid Hybridization (Hames and Higgins, eds.), все включены в данное описание в качестве ссылки.
Описание чертежей
Фиг.1:
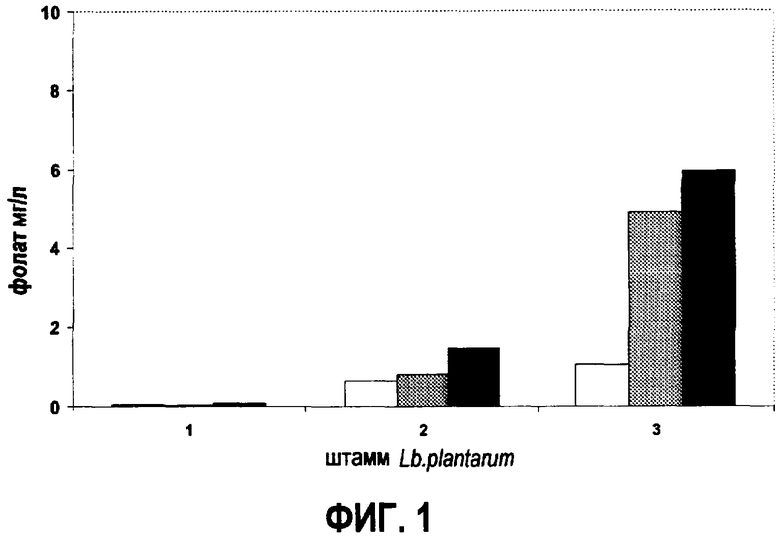
Продукция фолата диким штаммом L.plantarum (1) и штаммами L.plantarum, несущими в себе pNZ7019 (2) и pNZ7021 (3), культивированных на CDM. Расшифровка обозначений: внеклеточные уровни фолата; (белый столбец), внутриклеточные уровни фолата; (серый столбец), уровни общего фолата; (черный столбец).
Фиг.2:
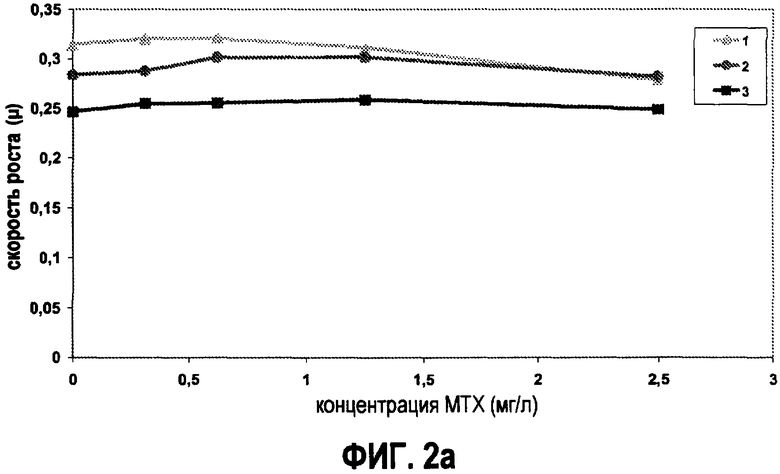
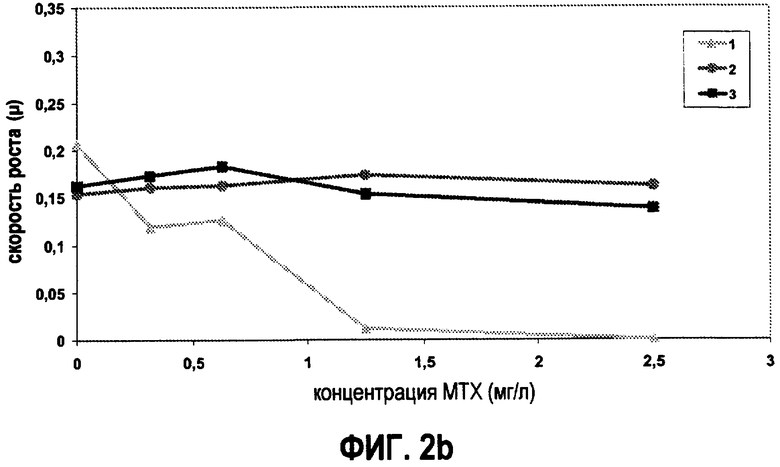
Скорость роста штамма дикого типа L.plantarum (1) и двух штаммов L.plantarum, несущих в себе вектор pNZ7019 (2) и pNZ7021 (3). Динамику роста трех штаммов L.plantarum тестировали на CDM (a), и на модифицированнной CDM (b), содержащей возрастающие концентрации метотрексата 0, 0,3125, 0,625, 1,25 и 2,5 мг/л (MTX).
Фиг.3:
Уровни продукции фолата 15 штаммами, гиперпродуцирующими фолат, которые отобраны по устойчивости к МТХ. Продукция фолата этими 15 штаммами сравнивали с продукцией фолата штаммом дикого типа.
ПРИМЕРЫ
1. Материалы и методы
1.1 Штаммы и условия культивирования
Штаммы L.plantarum WCFS1 дикого типа и ее производные выращивали при 37°C на среде для культивирования определенного химического состава (CDM), содержащей 19 г/л β-фосфоглицерата (6), или на модифицированной CDM. В этой модифицированной CDM глицин, инозин, оротовая кислота, тимидин, гуанин, аденин, урацил и ксантин были удалены из нормальной CDM. Дополнительно во все используемые среды было добавлено 8 нг/мл хлороамфеникола. Метотрексат (Sigma, аминоптерин) добавляли в оба типа CDM, в концентрациях в пределах от 0 до 10 мг/л.
1.2 ДНК-технологии, конструирование плазмид и трансформации
Плазмиды, использованные в данном исследовании, перечислены в таблице 1.
Вектор pNZ8148 представляет собой пустой вектор, содержащий индуцибельный промотор низина, многочисленные сайты клонирования и маркер устойчивости к хлорамфениколу. Производные pNZ8148 и геномную ДНК L.plantarum выделяли, используя общепринятые способы. ПЦР осуществляли с использованием Pfx ДНК полимеразы (Invitrogen, Paisley, UK) на приборе Mastercycler PCR (Eppendorf, Hamburg, Germany), используя следующие циклы: денатурация при 94°C в течение 30 с (3 минуты, первый цикл), отжиг при 45°C в течение 25 с и удлинение при 68°C в течение 1 минуты на 1000 п.о., в итоге в течение 30 циклов.
Вектор pNZ8148 расщепляли, используя SphI и BglII, таким образом, удаляя промотор низина. В параллельном эксперименте конститутивный промотор pepN L.lactis (Van Alen-Boerrigter et al., 1991. Appl. Environ. Microbiol. 57:2555-61) вырезали из вектора pNZ7017, используя SphI и BglII. Затем оба фрагмента лигировали, используя T4 ДНК лигазу (Invitrogen), полученную плазмиду обозначали как вектор pNZ7020. Кластер генов биосинтеза фолата штамма L.plantarum амплифицировали из хромосомной ДНК, используя следующие праймеры: (i) folBKpn-F (5'-GAAAGAGGCTGGGTACCATTATGGGCATGATTC-3'), который удлинен на 5'-конце сайтом рестрикции для KpnI, таким образом, перекрывая инициирующий кодон гена folB, и (ii) обратный праймер LpfPxba-R (5'-CTTAACCCCATCTAGACGTAATATCG-3'), который удлинен на 5'-конце для создания сайта рестрикции для XbaI, что позволяет слияние между стоп-кодоном гена folC и данным вектором. Амплифицированный фрагмент ДНК расщепляли с помощью XbaI и KpnI, что приводит к укороченной версии кластера генов folB-folP, (folP содержит сайт рестрикции для XbaI). Вектор pNZ7020 также расщепляли с помощью XbaI и KpnI, затем оба фрагмента лигировали с помощью T4 ДНК лигазы, полученный вектор обозначали как pNZ7020a. Недостающую часть гена folP амплифицировали посредством ПЦР, с использованием следующих праймеров lpfP-F (5'-CATGGCATCGATATTGAACGAATTG-3') и праймер lpfPxba-R (5'-CTTAACCCCATCTAGACGTAATATCG-3'). Амплифицированную ДНК расщепляли с помощью XbaI. Затем вектор pNZ7020a был расщеплен с помощью XbaI и затем фосфорилирован с использованием щелочной фосфатазы (Pharmacia Biotech) для предотвращения самолигирования. Оба расщепленных с помощью XbaI участка ДНК лигировали с помощью T4 ДНК лигазы, местоположение встроенного фрагмента проверяли с помощью ПЦР. Полученный вектор обозначали как pNZ7021. Три вектора, указанные в таблице 1, затем переносили в компетентные клетки L.lactis NZ9000 с фенотипом ylgG. Далее, мидипрепы этих трех векторов трансформировали в L.plantarum, используя общепринятые технологии.
1.3 Количественное определение фолата с использованием микробиологического анализа
Уровни фолата в трех штаммах L.plantarum, созданных методом генетической инженерии, определяли, используя микробиологический анализ Lactobacillus casei, как описано ранее (Sybesma et al, 2003. Appl. Environ. Microbiol. 69:3069-76.)
1.4 Мониторинг роста данных штаммов
Для определения скорости роста трех штаммов L.plantarum, несущих различные векторы, использовали микротитрационные планшеты. Рост этих трех штаммов контролировали в 96-луночном микротитрационном планшете в течение 35 часов, используя спектрофотометр (SPECTRAmax®, Molecular Devices, Sunnyvale, CA, USA). Рост контролировали на двух типах среды для культивирования и каждую конструкцию проверяли в трех повторах на обеих средах при следующих концентрациях МТХ: 0, 0,3125, 0,625, 1,25 и 2,5 мг/л.
1.5 Отбор гиперпродуцентов природного фолата
Дикий тип L.plantarum WCFS1, несущий в себе pNZ8148, повторно культивировали в модифицированной CDM, содержащей 2,5 мг/л MTX, и эту среду для выращивания разливали в 96 лунок микротитрационного планшета. Этот микротитрационный планшет инкубировали в течение 3 дней, а затем аликвоты из каждой из 96 лунок переносили в два свежих 96-луночных микротитрационных планшета, содержащие модифицированную CDМ, в этот раз с 10 мг/л МТХ. На протяжении каждой стадии исходный микротитрационный планшет заливали 60% глицерином и хранили в морозильной камере при -80°C. Микротитрационные планшеты, хранившиеся в глицерине, использовали для инокуляции мутированными или адаптированными клетками на свежей модифицированной среде CDM. Уровни фолата в этих 288 культурах, устойчивых к МТХ, проверяли, используя метод быстрого скрининга. В основе этого метода лежит стандартный микробиологический анализ, описанный ранее, в который были внесены некоторые изменения. В этом методе быстрого скрининга фолат, выработанный диким типом сравнивали с фолатом, выработанным мутантными штаммами. Культивирование индикаторного штамма на фолате, выработанном диким типом, приводит к относительно низкой оптической плотности. Оптическая плотность индикаторного штамма на фолате, выработанном гиперпродуцентами, будет выше, чем на фолате, выработанном штаммом дикого типа. Штаммы, образующие повышенную оптическую плотность индикаторного штамма, повторно анализировали, используя развернутый микробиологический анализ Lb.casei.
2. Результаты
2.1 Рекомбинантные штаммы
Уровни продукции фолата диким типом L.plantarum, несущим pNZ8148 и две генетические конструкции pNZ7019 и pNZ7021, исследовали на нормальной среде CDM. Продукция фолата штаммом L.plantarum WCFS1, несущим в себе пустой вектор pNZ8148, составляла 100 нг фолата/мл среды (фиг.1). Штамм L.plantarum, несущий pNZ7019 (кластер генов биосинтеза фолата штамма L.lactis), вырабатывает вплоть до 2000 нг/мл фолата в культуре (фиг.1). Это соответствует приблизительно 20-кратной гиперпродукции по сравнению с диким типом. Трансформация L.plantarum WCFS1 вектором pNZ7021, несущим кластер генов биосинтаза фолата штамма L.plantarum, приводит к штаммовой выработке уровней фолата вплоть до 6000 нг/мл культуры (фиг.1).
Затем специфические скорости роста двух штаммов, гиперпродуцирующих фолат, и штамма дикого типа сравнивали на CDM с и без фолат-зависимых метаболитов в присутствии различных концентраций метотрексата (МТХ) (фиг.2). В присутствии фолат-зависимых метаболитов ни один из тестируемых штаммов не был ингибирован посредством MTX (фиг.2a). Скорость роста L.plantarum, несущего в себе плазмиду pNZ7021, ниже по сравнению с двумя другими штаммами при всех протестированных концентрациях MTX. На среде, в которой отсутствуют фолат-зависимые метаболиты, наблюдается совершенно другая картина. На этой среде гиперпродуценты фолата явно более устойчивы к МТХ, по сравнению со штаммом дикого типа (фиг.2b). При концентрации МТХ, равной 2,5 мг/л, в модифицированной CDM наблюдали полное ингибирование роста штамма дикого типа, тогда как оба штамма, гиперпродуцирующе фолат, хорошо росли на этой среде. При концентрациях МТХ между 0 и 2,5 мг/л, наблюдали дозозависимое ингибирование для штамма L.plantarum дикого типа. Эти результаты показывают, что, если анализировать на подходящей среде для выращивания, гиперпродукция фолата приводит к устойчивости к МТХ. Таким образом, МТХ можно использовать для различения штаммов, гиперпродуцирующих фолат, от штаммов дикого типа.
2.2 Штаммы дикого типа
Длительное инкубирование (>40 часов) штамма L.plantarum WCFS1 дикого типа на модифицированной среде CDM, дополненной 2,5 мг/л MTX, приводит к значительному росту данной культуры. Если эту культуру разбавляли свежей модифицированной средой CDM, дополненной 2,5 мг/л МТХ, наблюдали немедленный рост данной культуры. Одиночные колонии выделяли из этой культуры для определения того, были ли отобраны природные мутанты с повышенной устойчивостью к МТХ, или имеет место какая-либо временная адаптация. Модифицированную среду CDM, содержащую 2,5 мг/л МТХ, инокулировали одиночной колонией, полученной из культуры, устойчивой к МТХ. Затем данную среду разделяли на две культуры; выращивали 50 поколений одной культуры в модифицированной среде CDM с 2,5 мг/л МТХ, выращивали 50 поколений другой культуры в такой же среде в отсутствие МТХ. После 50 поколений обе культуры снова перемещали в модифицированную CDM, содержащую 2,5 мг/л МТХ. Оба штамма, прошедшие через различные процессы, проявляли сопоставимую скорость роста на этой среде, что свидетельствует о том, что стабильная мутация приводит к фенотипу устойчивости к МТХ. Авторы данного изобретения изолировали 288 колоний из культур, содержащих 2,5 мг/л или 10 мг/л МТХ.
Эти колонии анализировали на предмет гиперпродукции фолата, используя метод быстрого предварительного скрининга. На основании этого быстрого скрининга 5 колоний из 288 дали рост культурам с более высокими уровнями фолата по сравнению с диким типом. Затем эти 5 культур помещали в чашку с модифицированной CDM, дополненной 2,5 мг/л МТХ. Из каждой чашки выделяли 3 одиночные колонии, сохраняли и снова анализировали продукцию фолата следующим образом. Контролировали уровни фолата в культурах, полученных от 15 положительных колоний (фиг.3). Средние уровни фолата были на 20%-60% выше, чем таковые, наблюдаемые у дикого типа. Для подтверждения того, что все колонии все еще были L.plantarum, авторы данного изобретения провели RAPD-анализ. Все проверенные колонии проявляли высокое сходство с исходным штаммом.
2.3. Анализ характера транскрипции.
Используя метод олигонуклеотидных микрочипов для определения характера транскрипции полного генома, авторы данного изобретения выявили у устойчивого к метотрексату мутанта L.plantarum NIZO B2550 (происходящий от L.plantarum WCFS1) гены, которые специфически активированы или супрессированы по сравнению с диким типом. Культуры L.plantarum WCFS1 и NIZO B2550 выращивали на модифицированной CDM и при оптической мутности (OD600), равной 1, клетки собирали и выделяли мРНК. Образцы мРНК подвергали анализу на ДНК-микрочипах. Относительное содержание специфических мРНК выражали как кратность изменения экспрессии генов в штамме, устойчивом к метотрексату, по сравнению с соответствующими генами в исходном штамме. Анализ характера транскриптона выявил, что по сравнению с исходным штаммом, folC1, ген, кодирующий дигидрофолатсинтазу, в частности, активируется в 4 раза (см. таблицу 2) у мутанта, устойчивого к метотрексату, по сравнению с диким типом. Этот фермент соединяет L-глутамат с дигидроптероатом с образованием дигидрофолата, который является структурным аналогом метотрексата и выполняет функцию субстрата для фермента дигидрофолатредуктазы. Ожидается, что большее количество дигидрофолатсинтазы в результате гиперэкспрессии folC1 приводит к большему содержанию внутриклеточного дигидрофолата, который, в свою очередь, может конкурировать с метотрексатом за трансформирование посредством дигидрофолатредуктазы. Это наблюдение объясняет механизм устойчивости к метотрексату у выделенных мутантов. Это также объясняет, почему эти мутанты вырабатывают более высокие уровни фолата в отсутствие метотрексата.
L.plantarum WCFS1 имеет два гена folC (folC1 и folC2). Интересно, что только folC1 активирован у устойчивого к метотрексату мутанта. Этот ген расположен не в функциональном кластере, в отличие от folC2, который расположен в кластере биосинтеза фолата.
Кроме folC1, было обнаружено некоторое количество других метаболических генов (кодирующих фруктокиназу, пируватоксидазу и 6-фосфо-бета-глюкозидазу), активированных в штамме, устойчивом к метотрексату (см. таблицу 2). Обнаружено, что три гена (dak1A, dak1B и dak2), кодирующие глицеринкиназу (дигидроксиацетонфосфотрансферазу), супрессированы у мутанта, по сравнению с исходным штаммом.
Гены, которые специфически активированы или супрессированы в L.plantarum NIZO B2550 по сравнению с L.plantarum WCFS1
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ УЛУЧШЕНИЯ ПРИЗНАКОВ РАСТЕНИЙ | 2016 |
|
RU2775712C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ УЛУЧШЕНИЯ ПРИЗНАКОВ РАСТЕНИЙ | 2016 |
|
RU2769873C2 |
| ПРИМЕНЕНИЕ БАКТЕРИЙ, ОТНОСЯЩИХСЯ К ГРУППЕ НОКАРДИОФОРМНЫХ АКТИНОМИЦЕТОВ, ДЛЯ ПОЛУЧЕНИЯ ФАРМАЦЕВТИЧЕСКОЙ КОМПОЗИЦИИ И СПОСОБ ИСПОЛЬЗОВАНИЯ ТАКОЙ ФАРМАЦЕВТИЧЕСКОЙ КОМПОЗИЦИИ | 2010 |
|
RU2543663C2 |
| ПРОТИВОМИКРОБНАЯ КОМПОЗИЦИЯ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2000 |
|
RU2241457C2 |
| ГЕНЫ-МИШЕНИ ДЛЯ НАПРАВЛЕННОГО ВОЗДЕЙСТВИЯ НА АЗОТФИКСАЦИЮ ДЛЯ УЛУЧШЕНИЯ КАЧЕСТВ РАСТЕНИЙ | 2018 |
|
RU2805085C2 |
| ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ АСПАРТОКИНАЗУ III, СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 1993 |
|
RU2113484C1 |
| СПОСОБ ПОЛУЧЕНИЯ АТТЕНУИРОВАННОГО ШТАММА БАКТЕРИЙ SALMONELLA | 1988 |
|
RU2114172C1 |
| СПОСОБ ПРОДУЦИРОВАНИЯ АМИНОКИСЛОТ СЕМЕЙСТВА АСПАРТАТА С ИСПОЛЬЗОВАНИЕМ МИКРООРГАНИЗМОВ | 2010 |
|
RU2535973C2 |
| МУТАНТНАЯ АЛЬФА-ИЗОПРОПИЛМАЛАТ СИНТАЗА (IPMS), ДНК, КОДИРУЮЩАЯ МУТАНТНУЮ IPMS, СПОСОБ ПОЛУЧЕНИЯ ШТАММА ESCHERICHIA COLI, СПОСОБ ПОЛУЧЕНИЯ L-ЛЕЙЦИНА | 1999 |
|
RU2201454C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 2006 |
|
RU2402610C2 |
Изобретение относится к биотехнологии, в частности к мутантной бактерии, продуцирующей повышенные уровни внутриклеточного и/или внеклеточного фолата по сравнению с диким типом, который является чувствительным к метотрексату, и имеет скорость роста, по меньшей мере, 0,1 µ в час, при росте на среде, в которой содержится 1,25 мг/л метотрексата и отсутствуют фолат-зависимые метаболиты. Также изобретение относится к композиции для обогащения пищевого продукта или пищевой добавки, содержащей вышеуказанную бактерию, и к способу отбора вышеуказанных бактерий. Изобретение позволяет получить бактерии с повышенным уровнем фолата. 4 н. и 9 з.п. ф-лы, 3 ил., 2 табл.
1. Мутантная бактерия - продуцент фолата, где мутантная бактерия представляет собой спонтанную, встречающуюся в природе мутантную бактерию или бактерию с индуцированной мутацией, в которой мутация индуцирована воздействием человека, отличающаяся тем, что бактерия содержит повышенные уровни внутриклеточного и/или внеклеточного фолата по сравнению с диким типом, который является чувствительным к метотрексату, и имеет скорость роста, по меньшей мере, 0,1 мкм/ч, при росте на среде, в которой содержится 1,25 мг/л метотрексата и отсутствуют фолатзависимые метаболиты.
2. Бактерия по п.1, где указанная бактерия представляет собой природный мутант или индуцированный мутант.
3. Бактерия по п.2, где указанная бактерия проявляет по сравнению с диким типом гиперэкспрессию одного или нескольких генов, кодирующих ферменты метаболического пути биосинтеза фолата.
4. Бактерия по п.2, где экспрессия одного или нескольких генов, выбранных из генов, кодирующих дигидрофолатсинтазу, фруктокиназу, пируватоксидазу и 6-фосфо-β-глюкозидазу, активирована, по меньшей мере, в 2 раза по сравнению с диким типом.
5. Бактерия по п.2, где экспрессия гена, кодирующего дигидроксиацетонфосфотрансферазу, супрессирована, по меньшей мере, в 2 раза по сравнению с диким типом.
6. Бактерия по п.1, где количество общего фолата в мутантной бактерии, по меньшей мере, на 10% больше такового в штамме дикого типа.
7. Бактерия по любому из предшествующих пунктов, где бактерия представляет собой род Lactobacillus plantarum, L.casei, L.reuteri, L.fermentum, L.acidophilus, L.crispatus, L.gasseri, L.johnsonii, L.paracasei, L.murinus, L.jensenii, L.salivarius, L.mmutis, L.brevis, L.gallinarum, L.amylovorus, Lactococcus lactis, Streptococcus thermophilus, Leuconostoc mesenteroides, Lc.lactis, Pediococcus damnosus, P.acidilactici, P.parvulus, Bifidobacterium bifidum, B.longum, B.infantis, B.breve, B.adolescentis, B.animalis, B.gallinarum, B.magnum и B.thermophilum.
8. Бактерия по п.1, где бактерия относится к штамму Lactobacillus plantarum CBS 117120.
9. Композиция для обогащения пищевого продукта или пищевой добавки фолатом, содержащая эффективное количество бактерий по любому из предшествующих пунктов.
10. Композиция по п.9, где композиция представляет собой пищевую композицию или композицию пищевой добавки.
11. Композиция по п.10, где пищевая композиция представляет собой ферментированную пищевую композицию.
12. Применение метотрексата для отбора бактерий по п.1.
13. Способ отбора бактерий по п.1, где указанный способ включает следующие стадии:
(a) выращивание бактерий на или в среде, содержащей, по меньшей мере 1,25 мг/л метотрексата, в течение по меньшей мере, 40 ч, где указанная среда не дополнена фолатзависимыми метаболитами,
(b) отбор бактерий, имеющих скорость роста, по меньшей мере, 0,1 мкм/ч,
(c) определение уровня фолата в бактериях, отобранных на стадии (b), и отбор бактерий из имеющих повышенные уровни фолата по сравнению с диким типом,
(d) необязательное повторение стадий (а), (b) и/или (с).
| TAMURA T | |||
| at al | |||
| The from of folate affects the mechanisms of methotrexate resistance in Enterococcus faecium, Microbiology, 1997, Aug, 143 (Pt 8), p.2639-2646 | |||
| SYBESMA W | |||
| at al | |||
| Effects of cultivation conditions on folate production by lactic acid bacteria | |||
| Appi Environ Microbiol | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| SYBESMA W | |||
| at al | |||
| Increased production | |||

