Настоящее изобретение относится к дрожжам, содержащим серосодержащее соединение, способу отбора дрожжей и способу культивирования дрожжей. Дрожжи, содержащие высокий процент серосодержащего соединения, в соответствии с настоящим изобретением применимы в области продуктов питания, лекарственных средств, химических продуктов, кормов и т.п.
До сих пор серосодержащие соединения широко используются в области продуктов питания, лекарственных средств, химических продуктов и т.п. Например, известно, что глутатион (GSH), который является трипептидом, состоящим из глутамата, цистеина и глицина, имеет фармацевтический эффект. В настоящее время препарат GSH классифицируют как антидот или офтальмический агент и используют для различных видов зависимостей, хронического заболевания печени, предупреждения нарушений, обусловленных побочными эффектами противораковых агентов и радиотерапии, дерматоза и терапии катаракты или повреждения роговицы (Protein, Nucleic Acid and Enzyme 1988-7 Vol. 33 No. 9 ISSN003909450, дополнительный выпуск “Epoch for research of glutathione” p. 1626).
С другой стороны глутатион известен в качестве вещества, которое придает продуктам питания богатство вкуса (Y.Ueda et al., Biosci. Biotech. Biochem., 61, 1977-1980 (1977)), а дрожжевой экстракт, содержащий высокий процент глутатиона, используется в качестве приправы. Кроме того, сообщается, что γ-глутамилцистеин, который является дипептидом, состоящим из глутамата и цистеина, применим для продукта питания.
Как описано выше, серосодержащие соединения имеют широкое промышленное применение, и по этой причине проводились различные исследования, касающиеся получения микроорганизма, который эффективно продуцирует эти соединения. Например, имеются сообщения, в которых описывается способ, включающий оценку фермента-мишени в пути биосинтеза серосодержащего соединения и повышение содержания серосодержащего соединения в клетках путем модификации функции этого фермента-мишени. Например, сообщается, что содержание глутатиона в клетках повышается при введении в дрожжи гена γ-глутамилцистеинсинтетазы E.coli (Yasuyuki OHTAKE et al., Agric. Biol. Chem., 52(11), 2753-2762, 1988) или гена глутатионсинтетазы E.coli (Yasuyuki OHTAKE et al., Journal of FERMENTATION AND BIOENGINEERING, Vol. 68, No. 6, 390-394, 1989).
Также сообщается способ повышения содержания глутатиона путем введения фермента, вовлеченного в синтез глутатиона в дрожжах (JP 61-52299 А, JP 62-275685 А, JP 63-129985 А и JP 04-179484 А).
Также сообщались способы отбора микроорганизма, имеющего повышенное клеточное содержание серосодержащего соединения, включающие мутирование микроорганизма и распределение мутированных штаммов по питательной среде, содержащей различные виды агентов, и отбор штамма, способного расти на питательной среде (устойчивого к лекарственному средству штамма), или штамма, неспособного расти на питательной среде (чувствительного к лекарственному средству штамма), с помощью метода реплик. Например, сообщается способ, включающий мутирование дрожжей, принадлежащих роду Candida, и отбор штамма, способного расти в питательной среде, содержащей этионин и судьфит, (JP 59-151894 А, JP 03-18872 А и JP 10-191963 А) и способ, включающий мутирование дрожжей, принадлежащих роду Saccharomyces, и отбор штамма, имеющего увеличенную устойчивость к цинку (JP 02-295480 А). Помимо этих способов были исследованы различные агенты для того, чтобы повысить клеточное содержание серосодержащих соединений (JP 06-70752 А и JP 08-70884 А).
Кроме того, недавно сообщалось о получении с помощью случайной мутации штаммов Saccharomyces cerevisiae, содержащих 5 вес.% или более глутатиона в клетках (JP 2004-180509 А). В этом документе содержание глутатиона в одном за другим мутантом определяли для получения штаммов, продуцирующих глутатион в высоком проценте. Хотя в документе говорилось, что способ делает возможной быструю оценку большого числа штаммов, при изучении описанного в документе способа автором настоящего изобретения было обнаружено, что число штаммов, которое один человек может оценить за день, составляет самое большее приблизительно 100 штаммов. Следовательно, очевидно, что операции, описанные в документе, являются очень сложными и являются дорогостоящими и требуют много времени.
Кстати, известно, что у дрожжей, демонстрирующих ауксотрофию по аденину, в частности у дрожжей, имеющих мутацию в гене ADE1 или гене ADE2, проявляется красный цвет. В последние годы механизм красного окрашивания был проанализирован на генетическом уровне. Т.е. обнаружено, что AIR или CAIR в качестве промежуточных продуктов при биосинтезе аденина, которые аккумулируются при мутации гена ADE2, кодирующего фермент, катализирующий шестую ступень биосинтеза пуринов и промежуточного продукта AIR, или при мутации гена ADE1, кодирующего фермент, катализирующий седьмую ступень биосинтеза пуринов и промежуточного продукта CAIR, связываются с глутатионом и переносятся в вакуоль, в результате чего проявляется красный цвет (K.G. Sharma et al., Arch Microbiol. (2003) 180: 108-117). Однако в этом документе сообщается, что при сверхэкспрессии гена GSH1, кодирующего ограничивающий скорость при биосинтезе GSH фермент, изменения интенсивности красного цвета не было. Следовательно, у специалиста со средним уровнем компетентности в данной области техники не могло возникнуть идеи использования ауксотрофии по аденину для отбора дрожжей, содержащих высокий процент GSH.
Задача настоящего изобретения состоит в предоставлении дрожжей, имеющих повышенное клеточное содержание серосодержащего соединения, способа эффективного отбора дрожжей и способа культивирования дрожжей с предоставлением дрожжам возможности аккумулировать в клетках серосодержащее соединение в высоком проценте.
Авторы настоящего изобретения подробно изучили сообщение Sharma и др. (Arch. Microbiol (2003) 180: 108-117) и выдвинули гипотезу, что поскольку биосинтез аденина вовлечен в биосинтез АТФ, являющийся существенным для организмов, в ауксотрофных по аденину штаммах, таких как штаммы с разрушенным геном ADE2 или геном ADE1, в клетках в достаточной степени аккумулируется такой промежуточный продукт, как AIR или CAIR. Авторы настоящего изобретения полагали, что, если гипотеза верна, проявление красного цвета у дрожжей контролирует клеточное содержание глутатиона, и дрожжи, у которых проявляется красный цвет большей интенсивности, могли бы иметь более высокое содержание глутатиона. В результате обширных изучений, в отличие от сообщения Sharma и др., авторы настоящего изобретения обнаружили, что дрожжи, содержащие высокий процент глутатиона, можно отобрать, используя в качестве показателя красный цвет, и обнаружили, что клеточное содержание серосодержащего соединения, в том числе глутатиона, можно повысить сочетанием чувствительности к селеновой кислоте со способностью к проявлению красного цвета, обусловленной приданием ауксотрофии по аденину и культивированием на минимальной питательной среде, дополненной метионином, завершив тем самым настоящее изобретение.
Одним аспектом настоящего изобретения является предоставление дрожжей, имеющих повышенное клеточное содержание серосодержащего соединения, причем указанные дрожжи демонстрируют чувствительность к селеновой кислоте и развивают красный цвет при их культивировании на питательной среде.
Другим аспектом настоящего изобретения является предоставление описанных выше дрожжей, причем указанная питательная среда содержит метионин.
Другим аспектом настоящего изобретения является предоставление описанных выше дрожжей, развивающих красный цвет при придании ауксотрофии по аденину.
Другим аспектом настоящего изобретения является предоставление описанных выше дрожжей, в которых указанная ауксотрофия по аденину придана модификацией гена ADE1 или гена ADE2.
Другим аспектом настоящего изобретения является предоставление описанных выше дрожжей, которые, дополнительно, имеют мутацию в гене ADE4 или гене ADE8, и при этом, развивают белый цвет при их культивировании на питательной среде.
Другим аспектом настоящего изобретения является предоставление описанных выше дрожжей, в которых указанная чувствительность к селеновой кислоте придана модификацией для увеличения экспрессии гена МЕТ25.
Другим аспектом настоящего изобретения является предоставление описанных выше дрожжей, в которых указанным серосодержащим соединением является по меньшей мере одно соединение, выбранное из группы, состоящей из цистеина, γ-глутамилцистеина, глутатиона и цистеинилглицина.
Другим аспектом настоящего изобретения является предоставление способа культивирования описанных выше дрожжей, включающего культивирование указанных дрожжей в условии избытка аденина для увеличения дрожжевых клеток и затем культивирование указанных дрожжей в условии недостаточного содержания аденина для повышения клеточного содержания серосодержащего соединения.
Другим аспектом настоящего изобретения является предоставление способа отбора дрожжей, имеющих повышенное клеточное содержание серосодержащего соединения, включающего подвергание дрожжей, которые демонстрируют ауксотрофию по аденину и чувствительность к селеновой кислоте, обработке для модификации гена, распределение модифицированных дрожжей по питательной среде, на которой дрожжи могут становиться красными в случае недостатка аденина, для образования колоний дрожжей и отбор колонии дрожжей, которая краснее, чем до модификации.
Другим аспектом настоящего изобретения является предоставление способа отбора дрожжей, имеющих повышенное клеточное содержание серосодержащего соединения, включающего подвергание дрожжей, которые демонстрируют чувствительность к селеновой кислоте, обработке для модификации гена, распределение модифицированных дрожжей по минимальной питательной среде, дополненной метионином, для образования колоний дрожжей и отбор колонии дрожжей, которая краснее, чем до модификации.
Другим аспектом настоящего изобретения является предоставление описанного выше способа, в котором указанным серосодержащим соединением является по меньшей мере одно соединение, выбранное из группы, состоящей из цистеина, γ-глутамилцистеина, глутатиона и цистеинилглицина.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
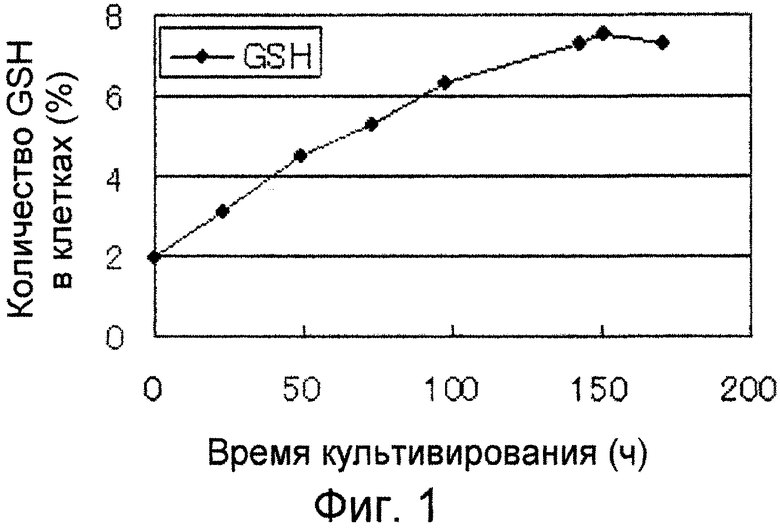
На фиг. 1 демонстрируется изменение с течением времени содержания GSH в штамме Y-3256.
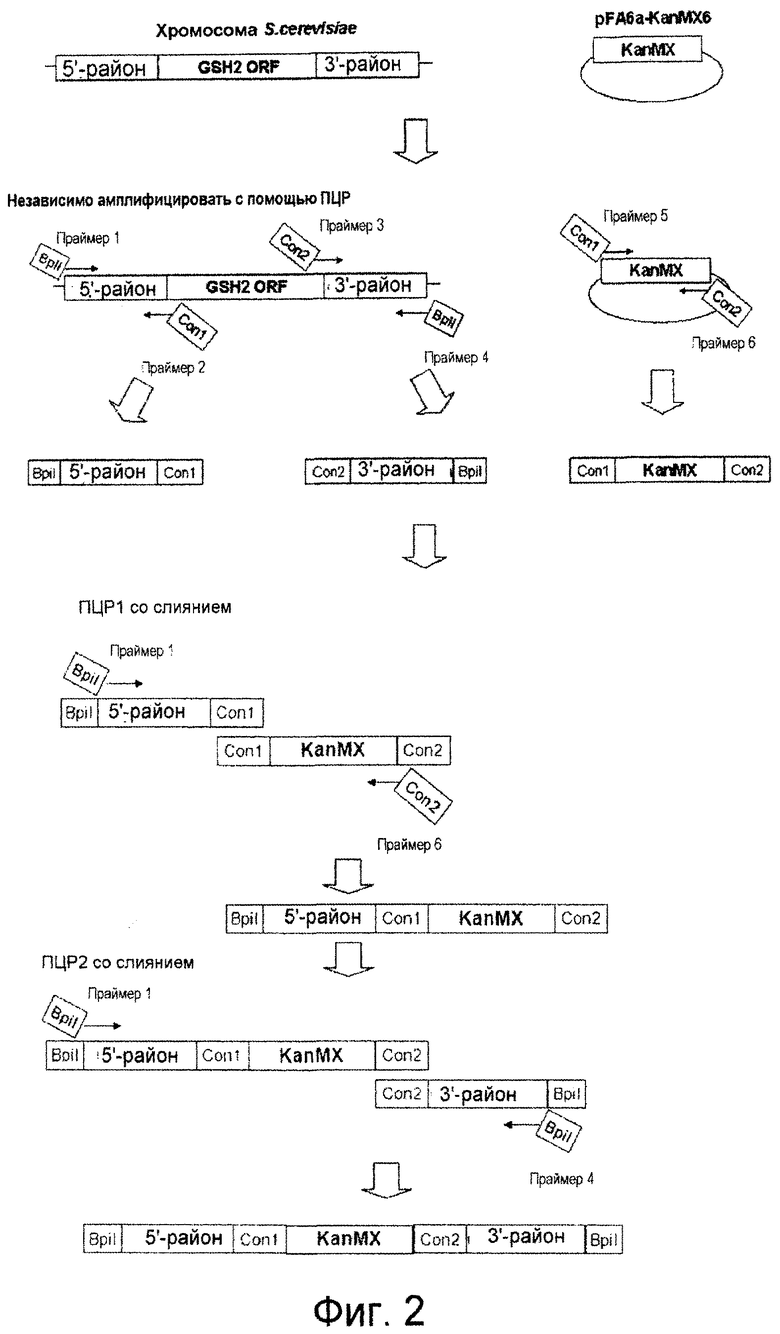
На фиг. 2 демонстрируется схема конструирования кассеты для разрушения гена GSH2.
ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Используемые в настоящем изобретении дрожжи особенно не ограничены при условии, что дрожжи демонстрируют чувствительность к селеновой кислоте и развивают красный цвет при их культивировании на питательной среде, и они, в силу этого, имеют повышенное клеточное содержание серосодержащего соединения. Примеры их включают дрожжи, принадлежащие роду Saccharomyces, например, Saccharomyces cerevisiae, дрожжи, принадлежащие роду Candida, например, Candida utilis, дрожжи, принадлежащие роду Pichia, например, Pichia pastoris, и дрожжи, принадлежащие роду Schizosaccharomyces, например, Schizosaccharomyces pombe. Среди них предпочтительными являются Saccharomyces cerevisiae и Candida utilis, которые часто используются при продукции серосодержащего соединения. Дрожжи настоящего изобретения могут быть гаплоидом или могут быть с двойным или более набором хромосом.
Используемая для оценки проявления красного цвета питательная среда особенно не ограничена при условии, что питательной средой является среда, на которой у дрожжей может проявляться красный цвет при истощении аденина в дрожжевых клетках. В качестве примера приводится питательная среда, имеющая содержание аденина, составляющее 25 мг/л или меньше, и ее конкретные примеры включают питательную среду YPD (1% бактодрожжевого экстракта, 2% бактопептона и 2% глюкозы; METHODS IN YEAST GENETICS 2000 Edition p.171: ISBN 0-87969-588-9) и питательную среду PGC (0,5% казаминовой кислоты (без витаминов), 1% бактопептона и 2% глюкозы). Предпочтительной является питательная среда PGC, на которой у дрожжей может легко проявляться красный цвет большей интенсивности под влиянием очень малого количества соединения. У дрожжей проявляется более ясно красный цвет при добавлении в питательную среду кадмия, и, следовательно, предпочтительно добавлять кадмий. Предпочтительно среду дополняют метионином. Предпочтительно используют агаризованную твердую питательную среда, а не жидкую питательную среду, так что штамм образует колонию красного цвета, и красное окрашивание можно оценить на основе цвета колонии. Способ предоставления красному цвету возможности проявиться особенно не ограничен, и его конкретные примеры включают придание ауксотрофии по аденину и культивирование на минимальной питательной среде, дополненной метионином.
В настоящем изобретении выражение «демонстрирующий ауксотрофию по аденину» означает, что штамм не может образовывать колонию на твердой питательной среде, которая не содержит аденин, или образует колонии небольшого размера, как это делает штамм Y-3219, на питательной среде. Ген, ответственный за ауксотрофию по аденину, может быть определен или может быть не определен, но предпочтительно, когда ген, ответственный за ауксотрофию по аденину, определен. Ген, ответственный за ауксотрофию по аденину можно определить путем анализа полученного ауксотрофного по аденину штамма с помощью анализа комплементации или анализа последовательности. Предпочтительно, когда дрожжи настоящего изобретения демонстрируют ауксотрофию по аденину вследствие модификации для инактивации по меньшей мере одного гена, вовлеченного в биосинтез аденина, такого как ген ADE2 и ген ADE1, которые непосредственно влияют на аккумуляцию промежуточных продуктов, включающих AIR и CAIR, которые связываются с GSH. В настоящем изобретении модификация для инактивации гена включает введение мутации, которая ликвидирует активность продукта гена в отношении гена и модификацию, которая ликвидирует экспрессию гена.
В настоящем изобретении выражение «подвергание обработке для модификации гена» означает, что мутируют нуклеотидную последовательность родительского штамма. Можно использовать обычную технологию мутирования, и можно также использовать технологию рекомбинации генов. Примеры обычных технологий мутирования включают метод мутирования гена облучением УФ-светом или лазером и метод с использованием агента для мутагенеза, такого как EMS, MNNG или DAPA (натрия 4-диметиламинобензолдиазосульфоната). Кроме того, может также быть включена встречающаяся в природе мутация, которая имеет место при культивировании микроорганизма. Ауксотрофные по аденину штаммы можно получить мутированием дрожжей и отбором штамма, который может расти в содержащей аденин питательной среде, но не может расти или плохо растет в питательной среде, не содержащей аденин. Более того, у штамма, имеющего мутацию в гене ADE1 или гене ADE2, демонстрируется ауксотрофия по аденину и проявляется красный цвет при его культивировании на питательной среде.
Модификация с использованием технологии рекомбинации гена включает способ замены гена дикого типа на хромосоме геном мутантного типа (гена инактивированного или разрушенного типа) с помощью гомологичной рекомбинации.
Вышеупомянутую замену гена можно осуществить следующим образом. Т.е. дрожжи трансформируют рекомбинантной ДНК, содержащей мутантный ген ADE1, для вызова рекомбинации между мутантным геном ADE1 и хромосомным геном ADE1. По этой причине маркерный ген, встроенный в рекомбинантную ДНК в зависимости от таких свойств, как аусотрофия хозяина, облегчает манипулирование. Более того, линеаризация вышеупомянутой рекомбинантной ДНК например, с помощью расщепления ферментом рестрикции и, кроме того, удаление контролирующей репликацию области, которая функционирует у дрожжей, из рекомбинантной ДНК может привести к эффективному получению штамма, в котором рекомбинантная ДНК интегрирована в хромосому.
Для трансформации дрожжей можно использовать такие методы, которые обычно используются при трансформации дрожжей, такие как метод протопластов, метод KU, метод KUR, метод электропорации и т.п. Штамм, в котором рекомбинантная ДНК интегрируется в хромосому вышеупомянутым образом, претерпевает рекомбинацию между мутантным геном ADE1 и геном ADE1, врожденно существующим на хромосоме, так что два слитых гена, т.е. ген ADE1 дикого типа и мутантный ген ADE1, становятся встроенными в хромосому так, чтобы между двумя слитыми генами присутствовали другие части рекомбинантной ДНК (сегмент вектора и маркерный ген).
Затем для того, чтобы на хромосомной ДНК остался только мутантный ген ADE1, одна копия гена ADE1 вместе с сегментом вектора (включающим также маркерный ген) удаляется из хромосомной ДНК с помощью рекомбинации двух генов ADE1. По этой причине существуют два случая. В одном случае ген ADE1 дикого типа остается на хромосомной ДНК, а мутантный ген ADE1 исключается из нее. В другом случае, наоборот, мутантный ген ADE1 остается на хромосомной ДНК, а дикий ген ADE1 исключается. В обоих случаях маркерный ген исключается, так что случай второй рекомбинации можно подтвердить по фенотипу, соответствующему маркерному гену. Являющийся целью штамм с заменой гена можно отобрать с помощью амплификации гена ADE1 способом ПЦР и проверки его структуры.
Хотя замена гена объяснена со ссылкой на ген ADE1, таким же образом можно также подвергнуть модификации другие гены, в том числе ген ADE2.
Предпочтительно, когда ген ADE4 или ген ADE8 также мутирован у дрожжей настоящего изобретения, поскольку дрожжи, имеющие мутацию в гене ADE4 или ADE8, демонстрируют нормальный белый, а не красный цвет. Штамм, имеющий мутацию в гене ADE4 или ADE8, помимо мутации в гене ADE1 или гене ADE2, можно получить сначала введением мутации в ген ADE1 или ген ADE2 с отбором штамма, имеющего повышенное содержание серосодержащего соединения, на основе интенсивности красного цвета в качестве показателя и затем введением мутации в ген ADE4 или ADE8, посредством чего можно эффективно получить белые дрожжи, содержащие высокий процент серосодержащего соединения.
Введение мутации в ген ADE4 или ADE8 можно осуществить таким же способом, как и используемый для гена ADE1 и гена ADE2 способ, описанный выше.
Ген ADE1, ген ADE2, ген ADE4 и ген ADE8 могут быть генами вида дрожжей, одинакового с видом дрожжей, подвергаемых модификации. Например, можно использовать ген ADE1, ген ADE2, ген ADE4 и ген ADE8 из Saccharomyces cerevisiae, имеющие нуклеотидные последовательности SEQ ID NO: 1, 3, 5 и 15, соответственно. Ген ADE1, ген ADE2, ген ADE4 и ген ADE8 не ограничены этими генами при условии, что они имеют гомологию последовательностей, достаточную для вызова гомологичной рекомбинации с геном на хромосоме модифицируемых дрожжей. Следовательно, ген ADE1, ген ADE2, ген ADE4 и ген ADE8 могут быть ДНК, кодирующими аминокислотные последовательности, идентичные на не менее 80%, предпочтительно на не менее 90%, более предпочтительно на не менее 95%, намного более предпочтительно на не менее 98% SEQ ID NO: 2, 4, 6 и 16, соответственно. Ген ADE1, ген ADE2, ген ADE4 и ген ADE8 могут быть ДНК, которые гибридизуются с нуклеотидными последовательностями SEQ ID NO: 1, 3, 5 и 15, соответственно, в жестких условиях. Примерами жестких условий являются условия, при которых промывку проводят один раз, предпочтительно два или три раза при концентрации соли, соответствующей 0,1 × SSC, 0,1% SDS при 60°С.
В настоящем изобретении выражение «имеющие чувствительность к селеновой кислоте» означает, что дрожжи не могут образовывать колонию на питательной среде, содержащей селеновую кислоту и метионин (например, питательной среде, содержащей 5 мМ селеновую кислоту и 1 мМ метионин). Селеновая кислота является структурным аналогом серной кислоты и токсичной для живого организма. Как правило, система поглощения серной кислоты супрессирована сероорганическими соединениями, такими как метионин, в дрожжевых клетках, так что селеновая кислота не поступает в клетки, и дрожжи могут расти в присутствии селеновой кислоты. С другой стороны в чувствительном к селеновой кислоте штамме система поглощения серной кислоты не супрессирована сероорганическими соединениями, и селеновая кислота поступает в клетки. Поскольку система поглощения серной кислоты не супрессирована, продукция серосодержащих соединений увеличена в чувствительном к селеновой кислоте штамме по сравнению со штаммом дикого типа (MOLECULAR AND CELLULAR BIOLOGY Dec, 1995, p. 6526-6534).
Чувствительность к селеновой кислоте можно придать мутационной обработкой или с помощью рекомбинации генов. Например, чувствительный к селеновой кислоте штамм можно получить мутированием дрожжей и отбором штамма, который не может расти в питательной среде, содержащей селеновую кислоту и метионин.
С другой стороны чувствительность к селеновой кислоте можно также придать с помощью рекомбинации генов, например, рекомбинации генов для увеличения экспрессии гена МЕТ25, который кодирует О-ацетилгомосеринсульфгидрилазу, по сравнению со штаммом дикого типа. Экспрессию гена МЕТ25 можно увеличить путем увеличения числа копий гена МЕТ25 с помощью введения плазмиды, несущей ген МЕТ25, или введения гена МЕТ25 в хромосому дрожжей или замены промотора гена МЕТ25 более сильным промотором.
Экспрессию гена МЕТ25 можно также увеличить путем модификации белка, который регулирует транскрипцию гена МЕТ25. Механизмом экспрессии гена МЕТ25 считается следующий механизм. Т.е. продукт гена МЕТ4 функционирует в качестве положительного регулятора экспрессии гена МЕТ25. Обычно продукт гена МЕТ4 образует комплекс SCFMET30 вместе с продуктом гена МЕТ30 и несколькими другими белками, и продукт гена МЕТ4 подвергается убиквитинилированию и разложению вместе с продуктом гена МЕТ30 с помощью протеолитической системы протеасом 26S, посредством чего экспрессия гена МЕТ25 супрессируется. С другой стороны при ухудшении функции комплекса SCFMET30 продукт гена МЕТ4 и продукт гена МЕТ30 не подвергаются разложению, и происходит экспрессия гена МЕТ25 (Patton et al., Genes Dev. 12: 692-705, 1998 и Rouillon et al., EMBO Journal 19: 282-294, 2004). Следовательно, экспрессию гена МЕТ25 можно увеличить введением мутации в ген МЕТ4 или ген МЕТ30.
В качестве мутантного гена МЕТ4, который может увеличить экспрессию гена МЕТ25, сообщается о гене, кодирующем МЕТ4, в котором серин в положении 215 заменен пролином, и гене, кодирующем МЕТ, в котором изолейцин в положении 156 замене серином (Ohmura et al., FEBS Letters 387(1996) 179-183; JP 10-33161А).
В качестве мутантного гена МЕТ30, который может увеличить экспрессию гена МЕТ25, сообщается о генах, кодирующих МЕТ30, в котором мутированы остатки аминокислот в повторе WD40. Кроме того, в JP 2004-201677А сообщается о мутантном гене МЕТ30, кодирующим МЕТ30, в котором серин в положении 569 заменен другими аминокислотами, такими как фенилаланин.
С помощью замены гена дикого типа на хромосоме генами мутантного типа можно увеличить экспрессию гена МЕТ25, посредством чего можно придать чувствительность к селеновой кислоте.
Ген МЕТ25, ген МЕТ30 и ген МЕТ4 могут быть генами вида дрожжей, одинакового с видом дрожжей, подвергаемых модификации. Например, можно использовать ген МЕТ25, ген МЕТ30 и ген МЕТ4 из Saccharomyces cerevisiae, имеющие нуклеотидные последовательности SEQ ID NO: 9, 11 и 7, соответственно. Ген МЕТ25, ген МЕТ30 и ген МЕТ4 не ограничены этими генами при условии, что ген МЕТ25 кодирует белок, имеющий О-ацетилгомосеринсульфгидрилазную активность, а ген МЕТ30 и ген МЕТ4 кодируют белок, имеющий функцию репрессора экспрессии гена МЕТ25. Следовательно, ген МЕТ25, ген МЕТ30 и ген МЕТ4 могут быть ДНК, кодирующими аминокислотные последовательности, идентичные на не менее 80%, предпочтительно на не менее 90%, более предпочтительно на не менее 95%, намного более предпочтительно на не менее 98% SEQ ID NO: 10, 12 и 8, соответственно. Ген МЕТ25, ген МЕТ30 и ген МЕТ4 могут быть ДНК, которые гибридизуются с нуклеотидными последовательностями SEQ ID NO: 9, 11 и 7, соответственно, в жестких условиях. Примеры жестких условий описаны выше.
Благодаря сочетанию ауксотрофии по аденину с чувствительностью к селеновой кислоте клеточное содержание серосодержащего соединения в дрожжах настоящего изобретения выше, чем в аналогичном штамме соответствующих дрожжей, который не демонстрирует ауксотрофию по аденину и демонстрирует чувствительность к селеновой кислоте.
В настоящем изобретении серосодержащее соединение относится к веществу, имеющему группу -SH в химической формуле, и может быть белком, пептидом, аминокислотой или другим веществом. Металлотионеин, в котором 30% составляющих аминокислот состоят из остатков цистеинов, является примером белка, глутатион, γ-глутамилцистеин и цистеинилглицин являются примерами пептида, цистеин является примером аминокислоты, а гомоцистеин является примером другого вещества, но серосодержащее соединение не ограничено ими. В качестве серосодержащих соединений предпочтительными являются глутатион, γ-глутамилцистеин и цистеин, поскольку они широко применяются в промышленности.
При аккумуляции глутатиона в качестве серосодержащего соединения в клетках дрожжей предпочтительно, когда дрожжи дополнительно модифицированы для увеличения внутриклеточной активности глутатионсинтетазы и/или γ-глутамилцистеинсинтетазы. Активности этих ферментов можно увеличить путем увеличения числа копий гена(ов), кодирующих эти ферменты, или замены промотора гена(ов) более сильным геном. В JP 2005-073628 представлены нуклеотидная последовательность гена γ-глутамилцистеинсинтетазы, происходящего из Saccharomyces cerevisiae, и нуклеотидные последовательности гена глутатионсинтетазы и гена γ-глутамилцистеинсинтетазы, происходящие из Candida utilis. Нуклеотидная последовательность гена глутатионсинтетазы, происходящего из Saccharomyces cerevisiae, и кодируемая ею аминокислотная последовательность показаны в SEQ ID NO: 13 и 14, соответственно.
При аккумуляции γ-глутамилцистеина в качестве серосодержащего соединения в клетках дрожжей предпочтительно, когда дрожжи дополнительно модифицированы для уменьшения внутриклеточной активности глутатионсинтетазы, и более предпочтительно, когда дрожжи дополнительно модифицированы для снижения внутриклеточной активности глутатионсинтетазы и для увеличения внутриклеточной активности γ-глутамилцистеинсинтетазы. Активность глутатионсинтетазы можно уменьшить разрушением гена глутатиосинтетазы или введением мутации в ген глутатионсинтетазы, так чтобы уменьшалась активность глутатионсинтетазы.
Мутацией для уменьшения активности глутатионсинтетазы является, например, мутация, которая приводит к замене аргинина в положении 370 в аминокислотной последовательности SEQ ID NO: 14 кодоном терминации.
Другие примеры мутаций для уменьшения активности глутатионсинтетазы включают следующие мутации (WO 03/046155):
(1) мутацию, которая приводит к замене треонина в положении 47 в аминокислотной последовательности SEQ ID NO: 14 изолейцином;
(2) мутацию, которая приводит к замене глицина в положении 387 в аминокислотной последовательности SEQ ID NO: 14 аспарагиновой кислотой;
(3) мутацию, которая приводит к замене пролина в положении 54 в аминокислотной последовательности SEQ ID NO: 14 лейцином.
Мутация может быть единичной мутацией (1) или (2) или любой комбинацией из (1)-(3) и предпочтительно комбинацией (1) с (3) или комбинацией (2) с (3).
Поскольку при разложении γ-глутамилцистеина образуется цистеин, дрожжи, имеющие повышенное клеточное содержание цистеина можно получить тепловой обработкой дрожжей, имеющих повышенное клеточное содержание γ-глутамилцистеина.
Кроме того, при аккумуляции цистеинилглицина в качестве серосодержащего соединения в клетках дрожжей предпочтительно, чтобы дрожжи были дополнительно модифицированы для увеличения внутриклеточной активности ЕСМ38 (Yeast. 2003 Jul. 30; 20(10): 857-63).
Вышеупомянутые дрожжи, которые демонстрируют ауксотрофию по аденину и чувствительность к селеновой кислоте, можно создать так, чтобы они содержали больше серосодержащего соединения, путем подвергания их процедуре модификации гена, предпочтительно мутационной обработке, затем распределения модифицированных дрожжей по питательной среде, на которой дрожжи могут становиться красными в случае недостатка аденина, для образования колоний дрожжей и отбора дрожжевой колонии, которая краснее, чем до модификации.
В настоящем изобретении используемая для отбора питательная среда особо не ограничена при условии, что питательная среда является средой, в которой у дрожжей может проявляться красный цвет при истощении аденина в дрожжевых клетках. В качестве примера приводится питательная среда, имеющая содержание аденина, составляющее 25 мг/л или меньше, и конкретные примеры ее включают питательную среду YPD (1% бактодрожжевого экстракта, 2% бактопептона и 2% глюкозы; METHODS IN YEAST GENETICS 2000 Edition p.171: ISBN 0-87969-588-9) и питательную среду PGC (0,5% казаминовой кислоты (без витаминов), 1% бактопептона и 2% глюкозы). Предпочтительной является питательная среда PGC, на которой у дрожжей может легко проявляться красный цвет большей интенсивности под влиянием очень малого количества соединения. У дрожжей проявляется более ясно красный цвет при добавлении в питательную среду кадмия, и, следовательно, предпочтительно добавлять кадмий. Предпочтительно используют агаризованную твердую питательную среда, а не жидкую питательную среду, так что штамм легко выделить для использования в последующих стадиях. С целью ясного наблюдения за интенсивностью проявления цвета предпочтительным является культивирование дрожжей на питательной среде при 20-30οС в течение приблизительно одной недели.
Дрожжи, имеющие повышенное клеточное содержание серосодержащего соединения, можно также получить путем подвергания дрожжей, которые демонстрируют чувствительность к селеновой кислоте, процессу модификации гена, предпочтительно мутационной обработке, распределения модифицированных дрожжей по минимальной питательной среде, дополненной метионином, для образования колоний дрожжей и отбора колонии дрожжей, которая краснее, чем до модификации. Используемой для этого способа отбора средой может быть содержащая метионин среда, которая имитирует состояния ауксотрофии по аденину условия, предпочтительно минимальная питательная среда, дополненная метионином в отсутствие биотина, и ее конкретный пример включает описанную ниже чашку со средой min-met(+)-biotin(-).
Дрожжи, имеющие повышенное клеточное содержание серосодержащего соединения, можно продуцировать с помощью культивирования дрожжей настоящего изобретения. Предпочтительно дрожжи культивируют в питательной среде, содержащей достаточное количество аденина, (в условии избытка аденина) для пролиферации дрожжей и затем культивируют в питательной среде, в которой содержание аденина ограничено, (в условии недостаточного содержания аденина) для повышения клеточного содержания серосодержащего соединения. Таким образом можно эффективно продуцировать дрожжи, в которых аккумулируется серосодержащее соединение.
«Достаточное количество» можно определить, например, путем экспериментального измерения количества аденина, требуемого для получения заранее определенного количества клеток, и расчета количества аденина, требуемого для получения желаемого количества клеток. Например, «достаточное количество» может быть меньше 100 мг/л.
Питательную среду для культивирования и условия культивирования, отличные от количества аденина, можно соответствующим образом выбрать, исходя из обычной питательной среды и условий, используемых для культивирования нормальных дрожжей. В зависимости от свойств используемых дрожжей в среду можно необязательно добавить необходимые питательные вещества.
После получения достаточного количества клеток, дрожжи культивируют в питательной среде, в которой количество аденина ограничено. Предпочтительно такая питательная среда имеет содержание аденина, составляющее 25 мг/л или меньше. Клеточное содержание серосодержащего соединения повышается во время культивирования в питательной среде, в которой содержание аденина ограничено.
Предпочтительно культивирование прекращают, когда количество аккумулированного серосодержащего соединения достигает желаемого количества. Обычно время культивирования составляет 10-30 часов, предпочтительно 15-27 часов.
Полученные подвергнутые культивированию клетки или их фракционированный продукт содержат серосодержащие соединения. Подвергнутыми культивированию клетками может быть питательная среда для культивирования, содержащая дрожжевые клетки, или дрожжевые клетки, собранные из среды для культивирования. Фракционированным продуктом, содержащим серосодержащие соединения, могут быть клеточные гомогенаты или дрожжевой экстракт. Приготовление дрожжевого экстракта и т.п. можно осуществить способом, одинаковым с общепринятым способ приготовления дрожжевого экстракта. Дрожжевой экстракт можно получить обработкой дрожжевых клеток горячей водой или обработкой дрожжевых клеток с помощью расщепления ферментом.
Серосодержащие соединения можно выделить из описанных выше дрожжевых клеток. Серосодержащие соединения, а также дрожжевые клетки и их фракционированный продукт, содержащие серосодержащие соединения, можно использовать для получения продуктов питания, лекарственных средств, химических продуктов, кормов для животных и т.п. Примеры продуктов питания включают алкогольные напитки, хлебные продукты и вкусовые вещества ферментированных пищевых продуктов. Продукты питания можно получить смешиванием серосодержащих соединений, подвергнутых культивированию клеток или их фракционированного продукта с исходными материалами продуктов питания и переработкой смеси в продукты питания.
ПРИМЕРЫ
Ниже настоящее изобретение описывается со ссылкой на примеры. Но настоящее изобретение не ограничивается этими примерами.
ПРИМЕР 1
(Выделение ауксотрофных по аденину дрожжей)
Диплоидный штамм Saccharomyces cerevisiae получали скрещиванием гаплоидного штамма AJ14819, который несет мутантный ген МЕТ30 (альфа-тип МАТ), с гаплоидным штаммом AJ14810 (тип МАТ). Штамм AJ14819, который является чувствительным к селеновой кислоте штаммом, был депонирован в Международный депозитарий патентуемых организмов, National Institute of Advanced Industrial Science and Technology (Tsukuba Central 6, 1-1, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken 305-8566, Япония) 1 октября 2003 согласно условиям Будапештского договора и получил входящий номер FERM BP-8502. Штамм AJ14810 был депонирован в Международный депозитарий патентуемых организмов, National Institute of Advanced Industrial Science and Technology 1 ноября 2002 согласно условиям Будапештского договора и получил входящий номер FERM BP-8229. Затем был создан диплоидный штамм для образования спор, и получили с помощью тетрадного анализа следующие гаплоидные штаммы:
А: гаплоид, тип МАТ, с мутированным МЕТ30;
В: гаплоид, альфа-тип МАТ, с мутированным МЕТ30;
С: гаплоид, тип МАТ, с МЕТ30 дикого типа;
D: гаплоид, альфа-тип МАТ, с МЕТ30 дикого типа.
Оценивали уровень экспрессии гена МЕТ25 в полученных 4 штаммах. Основываясь на способах, описанных в примере 1 JP 2004-201677, уровень экспрессии гена МЕТ25 в этих штаммах определяли следующим образом. Каждый штамм инокулировали в питательную среду YPD (50 мл в колбе Сакагучи объемом 500 мл) и культивировали при 30°С с покачиванием. В логарифмической фазе роста клетки каждого штамма собирали, и из клеток экстрагировали РНК, количественный уровень экспрессии гена МЕТ25 определяли, используя ген АСТ1 в качестве внутреннего стандарта. Определение количественного уровня проводили с использованием PCR5700 (Applied Biosystems) и набора для ОТ-ПЦР TaqMan One-Step (Applied Biosystems). АСТ1-986Т и МЕТ25-1077Т (JP 2004-201677) использовали в качестве TaqMan-зондов (Applied Biosystems), и в качестве праймеров использовали АСТ1-963F и АСТ1-1039R (JP 2004-201677) для амплификации АСТ1 и МЕТ25-1056F и МЕТ25-1134R (JP 2004-201677) для амплификации МЕТ25. В результате подтвердили, что уровень экспрессии гена МЕТ25 в штамме А и штамме В, оба из которых обладают мутантным геном МЕТ30 и чувствительностью к селеновой кислоте, выше, чем в штамме С и штамме D, оба из которых не обладают мутантным геном МЕТ30. Штамм В получил неофициальный номер AJ14889, а штамм А получил неофициальный номер AJ14890.
Затем штамм AJ14889 обрабатывали мутагеном MNNG (1-метил-3-нитро-1-нитрозогуанидином), так чтобы степень выживания становилась равной 5-10%, в соответствии с обычным способом, и обработанные клетки распределяли по чашкам YPD и культивировали при 30°С в течение приблизительно одной недели. Среди появившихся колоний отбирали колонии красного цвета. Анализ комплементации проводили в отношении ауксотрофии по аденину полученных штаммов в соответствии с общепринятым способом. В результате получили два штамма, одним из которых был штамм N1, который является ауксотрофным по аденину вследствие мутации в гене ADE1, а другим штаммом был штамм N2, который является штаммом с неполным блокированием аденина вследствие мутации в гене ADE2. Штаммы N1 и N2 были депонированы во Всероссийскую коллекцию промышленных микроорганизмов (ВКПМ) (1 Дорожный проезд, Москва 117545, Россия) 23 декабря 2008 согласно условиям Будапештского договора и получили входящие номера VKPM Y-3218 и VKPM Y-3219, соответственно.
ПРИМЕР 2
(Отбор мутантов с высокой аккумуляцией GSH и интенсивным красным окрашиванием)
Штамм Y-3218, штамм, имеющий мутацию в гене ADE1, обрабатывали мутагеном MNNG, так чтобы степень выживания становилась равной 5-10%, в соответствии с обычным способом, и обработанные клетки распределяли по чашке PGC или чашке YPD и культивировали при 30°С в течение приблизительно одной недели. Отбирали колонии, демонстрирующие более интенсивный красный цвет, чем цвет Y-3218, и содержание в них GSH сравнивали с содержанием GSH в штамме AJ14889 (исходном штамме) и штамме Y-3218. Т.е. отобранные штаммы, а также штамм AJ14889 и штамм Y-3218 инокулировали соответственно в 5 мл жидкой питательной среды YPD и культивировали при 30°С с покачиванием при 250 оборотах/минуту в течение 24 часов. Затем полученную культуру каждого штамма инокулировали в 50 мл жидкой питательной среды YPD и культивировали при 30°С с покачиванием при 250 оборотах/минуту в течение 24 часов. Содержание GSH в каждом штамме определяли в соответствии с общепринятой процедурой, и в результате было обнаружено, что содержание GSH в штамме Y-3218 выше, чем содержание GSH в штамме AJ14889 (таблица 1). Также было обнаружено, что среди отобранных штаммов (всего 180 штаммов) семь штаммов имели более высокое содержание GSH, чем штамм Y-3218 (таблица 1).
Эксперименты такого же рода были выполнены с использованием штамма Y-3219, штамма, имеющего мутацию в гене ADE2, вместо штамма Y-3218. Штамм Y-3219 обрабатывали мутагеном MNNG, так чтобы степень выживания становилась равной 5-10%, в соответствии с обычным способом, и обработанные клетки распределяли по чашке PGC или чашке YPD и культивировали при 30οС в течение приблизительно одной недели. Отбирали колонии, демонстрирующие более интенсивный красный цвет, чем цвет Y-3219, и содержание в них GSH сравнивали с содержанием GSH в штамме AJ14889 (исходном штамме) и штамме Y-3219. Т.е. отобранные штаммы, а также штамм AJ14889 и штамм Y-3219 инокулировали соответственно в 5 мл жидкой питательной среды YPD и культивировали при 30οС с покачиванием при 250 оборотах/минуту в течение 24 часов. Затем полученную культуру каждого штамма инокулировали в 50 мл жидкой питательной среды YPD и культивировали при 30οС с покачиванием при 250 оборотах/минуту в течение 24 часов. Содержание GSH в каждом штамме определяли в соответствии с общепринятой процедурой, и в результате было обнаружено, что содержание GSH в штамме Y-3219 выше, чем содержание GSH в штамме AJ14889 (таблица 2). Также было обнаружено, что десять из отобранных штаммов (всего 118 штаммов) имели более высокое содержание GSH, чем штамм Y-3219 (таблица 2).
Таблица 1 и таблица 2 показали, что было возможным выделение мутантов с более высоким содержанием GSH с использованием в качестве показателя интенсивного красного окрашивания аусотрофии по аденину.
ПРИМЕР 3
(Выделение белой колонии, имеющей высокое содержание GSH)
Цвет штамма дрожжей дикого типа не красный, а белый или бело-желтоватый. Поэтому авторы настоящего изобретения пытались выделить штамм, который образует колонию нормального цвета, из полученных мутантов интенсивного красного цвета. Сначала штамм Y-3219 обрабатывали мутагеном MNNG таким же образом, как описано в примере 2, и обработанные клетки распределяли по чашке PGC или чашке YPD для получения мутанта более интенсивного красного цвета. Штамм Y-3219-20, представленный в таблице 3, получали таким же образом, как описано в примере 2. Затем штамм Y-3219-20 обрабатывали мутагеном MNNG, так чтобы степень выживания становилась равной 5-10%, в соответствии с обычным способом, и обработанные клетки распределяли по чашке PGC и культивировали при 30°С в течение приблизительно одной недели. Среди появившихся колоний отобрали три белых колонии (Y-3219-20-52, Y-3219-20-53, Y-3219-20-56) и одну красную колонию (Y-3219-20-1), и содержание в них GSH сравнивали с содержанием GSH в штамме Y-3219 и штамме Y-3219-20. Т.е. отобранные штаммы, а также штаммы Y-3219 и Y-3219-20 инокулировали соответственно в 5 мл жидкой питательной среды YPD и культивировали при 30οС с покачиванием при 250 оборотах/минуту в течение 24 часов. Затем полученную культуру каждого штамма инокулировали в 50 мл жидкой питательной среды YPD и культивировали при 30°С с покачиванием при 250 оборотах/минуту в течение 24 часов. Содержание GSH в каждом штамме определяли в соответствии с общепринятой процедурой. Результаты представлены в таблице 3.
Белые колонии генетически анализировали с использованием штаммов-тестеров. В результате было обнаружено, что штамм Y-3219-20-52, который имел уменьшенное содержание GSH, потерял ауксотрофию по аденину. С другой стороне штамм Y-3219-20-56, в котором сохранялось высокое содержание GSH, имел мутацию в гене ADE4, помимо мутации в гене ADE2, штамм Y-3219-20-53 имел мутацию в гене ADE8, помимо мутации в гене ADE2. На основании этих результатов обнаружено, что ауксотрофный по аденину штамм, который образует красную колонию, может быть превращен в штамм, который образует колонию нормального цвета, путем мутирования гена ADE4 или гена ADE8. Штамм Y-3219-20-56 был депонирован во Всероссийскую коллекцию промышленных микроорганизмов (ВКПМ) (1 Дорожный проезд, Москва 117545, Россия) 23 декабря 2008 согласно условиям Будапештского договора и получил входящий номер VKPM Y-3256.
ПРИМЕР 4
(Высокая аккумуляция GSH, вызванная недостатком аденина)
Исследовали связь между содержанием GSH и недостатком аденина. Штамм Y-3256 культивировали в 50 мл питательной среды YPD (20 г/л D-глюкозы, 20 г/л бактопептона, 10 г/л дрожжевого экстракта), содержащейся в колбе Сакагучи объемом 500 мл, при 30°С с покачиванием при 120 оборотах/мин в течение 24 часов, и полученную культуру инокулировали в 50 мл питательной среды YPD, содержащейся в колбе Сакагучи объемом 500 мл, дополненной различными концентрациями аденина (конечная концентрация аденина 0 мг/л, 10 г/л или 20 г/л), чтобы сделать исходную оптическую плотность при 660 нм равной 0,1, и культивировали при 30°С с покачиванием при 120 оборотах/мин. Таблица 4 демонстрирует временной курс содержания GSH.
Эти результаты указывают на то, что содержание GSH в ауксотрофном по аденину штамме Y-3256 обратно пропорционально концентрации аденина в питательной среде. Следует отметить, что штамм Y-3256 может расти в питательной среде YPD без дополнения аденином (аденин 0 мг/л), поскольку питательная среда YPD содержит очень малое количество аденина (в соответствии с анализом содержание аденина в среде YPD составляет приблизительно 10 мг/л).
ПРИМЕР 5
(Оценка в широкогорлом биореакторе)
Штамм Y-3256 культивировали в широкогорлом биореакторе, и оценивали временной курс содержания GSH в штамме Y-3256. Клетки штамма брали с агаризованной твердой питательной среды YPD и инокулировали в три конические колбы объемом 750 мл, содержащие 50 мл жидкой питательной среды YPD, и культивировали при 30°С с покачиванием при 250 оборотах/мин в течение 20 часов. Полученную посевную культуру (120 мл) инокулировали в 1,2 л питательной среды для основного культивирования (среды YPD), содержащейся в широкогорлом биореакторе объемом 3 л, и культивирование проводили при 30°С с покачиванием при 1100 оборотах/мин. Питательную среду аэрировали со сменой 1/1 (объем/объем)/мин, и рН регулировали водным аммиаком так, чтобы он составлял 6. Подаваемую питательную среду подавали со скоростью 1,5 мл/час в пределах от 0 до 24 часов, и затем со скоростью 1,8 мл/час. Состав подаваемой среды включал 600 г глюкозы, 10 г бактодрожжевого экстракта, 10 г кукурузного экстракта, 10 г бактопептона, 0,274 г (NH4)2SO4, 0,11 г KH2PO4, 0,732 г KCl, 0,466 г MgSO4, 0,0012 г CuSO4, 0,014 г ZnSO4, 0,00334 г MnSO4, 0,00012 г NaMoO4, 0,002 г KCl, 0,00004 г H3B03, 0,0001 г CoSO4, 0,28 г CaCl2, 0,2 г FeSO4, 0,05 мг биотина, 0,2 мг рибофлавина и 0,5 мг тиамина на 1 л. На фиг. 1 демонстрируется временной курс содержания GHS в штамме Y-3256.
ПРИМЕР 6
(Получение диплоидного штамма и исследование состава питательной среды)
В соответствии с общепринятым способом получали гомодиплоидный штамм Y-3219-20-53. Экспериментальными условиями были следующие условия. Штамм Y-3219-20-53 обрабатывали мутагеном MNNG, так чтобы степень выживания становилась равной 5-10%, в соответствии с обычным способом, и обработанные клетки распределяли по агаризованной твердой питательной среде YPD так, чтобы появилось 100-200 колоний. После культивирования при 30οС в течение пяти дней колонии перепечатывали на агаризованную твердую питательную среду SD, дополненную аденином, и на агаризованную твердую питательную среду YPD, и отобрали штамм, который мог расти на питательной среде YPD, но не мог расти на питательной среде SD, дополненной аденином. Определяли ауксотрофию, отличную от ауксотрофии по аденину, и получили штамм Y-3219-20-53-aux1 и штамм Y-3219-20-53-aux2, которые обладали отличными друг от друга аусотрофиями. Делали посев вертикальными штрихами штамма Y-3219-20-53-aux1 на агаризованную твердую питательную среду SD, дополненную аденином, и делали посев горизонтальными штрихами штамма Y-3219-20-53-aux2 на ту же агаризованную твердую питательную среду SD, дополненную аденином, так чтобы вертикальный штрих и горизонтальный штрих пересекались в одной точке. Эту агаризованную твердую питательную среду SD, дополненную аденином, инкубировали при 30οС в течение 20 дней, и отобрали колонию, которая появилась в точке пересечения. Таким образом получили аусотрофный по аденину диплоидный штамм D1-3. Штамм D1-3 был депонирован во Всероссийскую коллекцию промышленных микроорганизмов (ВКПМ) (1 Дорожный проезд, Москва 117545, Россия) 23 декабря 2008 согласно условиям Будапештского договора и получил входящий номер VKPM Y-3309. При посеве штрихами штамма D1-3 на агаризованную твердую питательную среду SD, дополненную аденином, и культивировании при 30°С в течение 5 дней образовывались колонии, так что было подтверждено, что этому штамму не была придана аусотрофия, отличная от аусотрофии по аденину.
Диплоидный штамм Dip был получен в качестве контрольного не являющегося аусотрофным по аденину штаммом скрещиванием штамма AJ14889 со штаммом AJ14890.
Штамм D1-3 и штамм Dip инокулировали в 5 мл жидкой питательной среды YPD, содержащейся в пробирке, и культивировали при 30°С в течение 24 часов с покачиванием при 250 оборотах/мин. Полученную культуру инокулировали в 50 мл каждой из питательных сред, представленных в таблице 5, и культивирование проводили при 30°С в течение 24 часов с покачиванием при 250 оборотах/мин. Определяли содержание GSH в каждом из штаммов, подвергнутых культивированию в каждой из питательных сред. Результаты представлены в таблице 6.
Эти данные показывают, что аусотрофия по аденину может повысить содержание GSH при использовании каждой из питательных сред.
ПРИМЕР 7
(Получение аусотрофных по аденину дрожжей с высоким содержанием γ-глутамилцистеина и оценка повышенного содержания γ-глутамилцистеина)
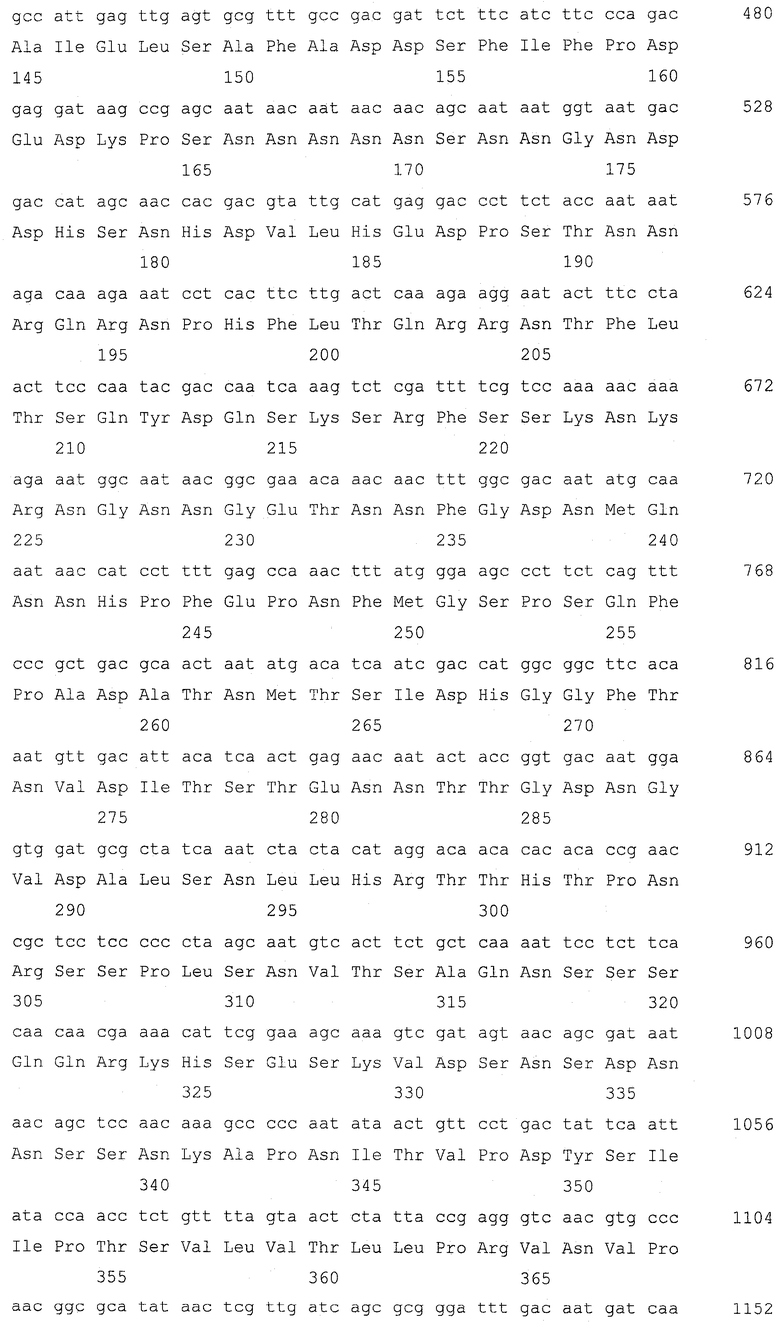
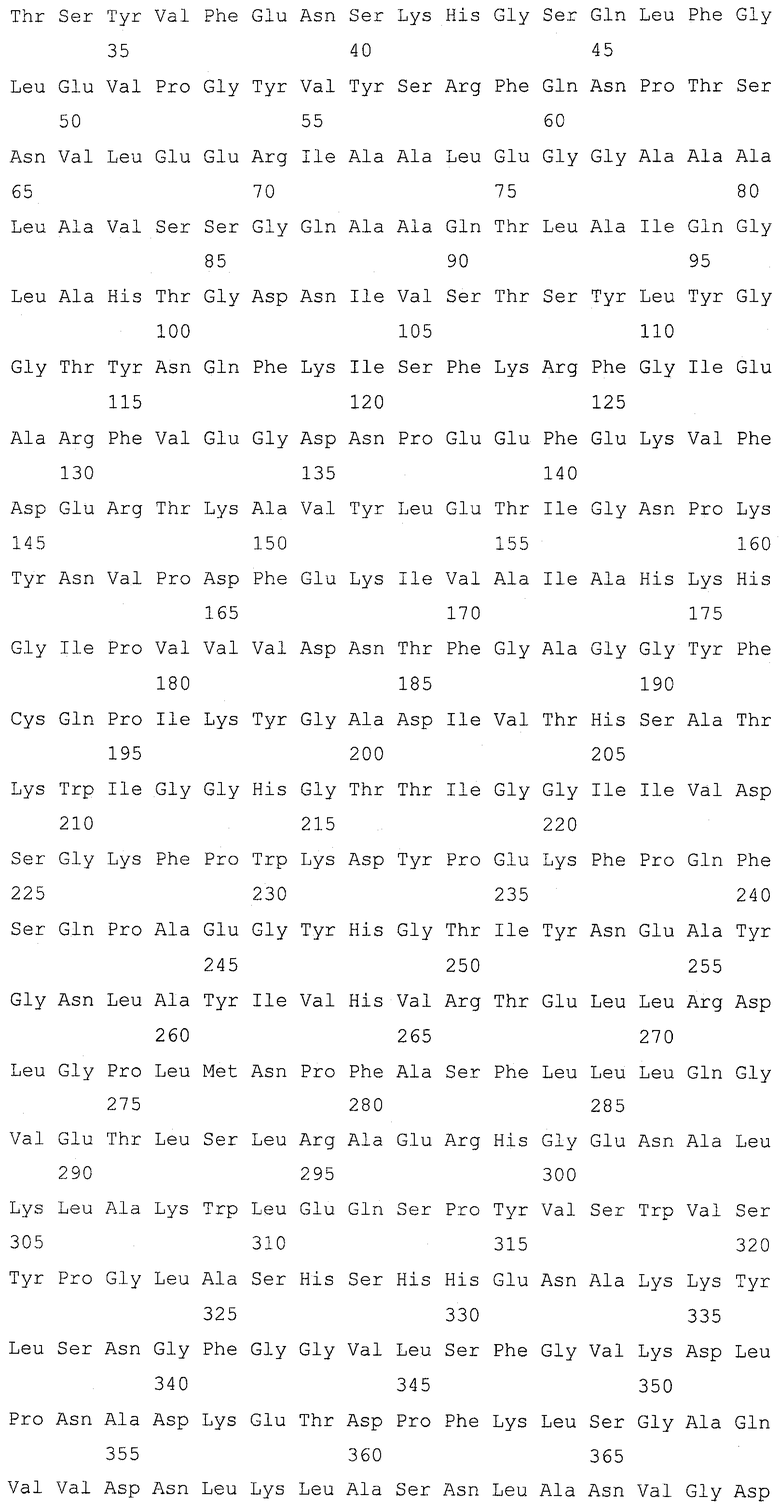
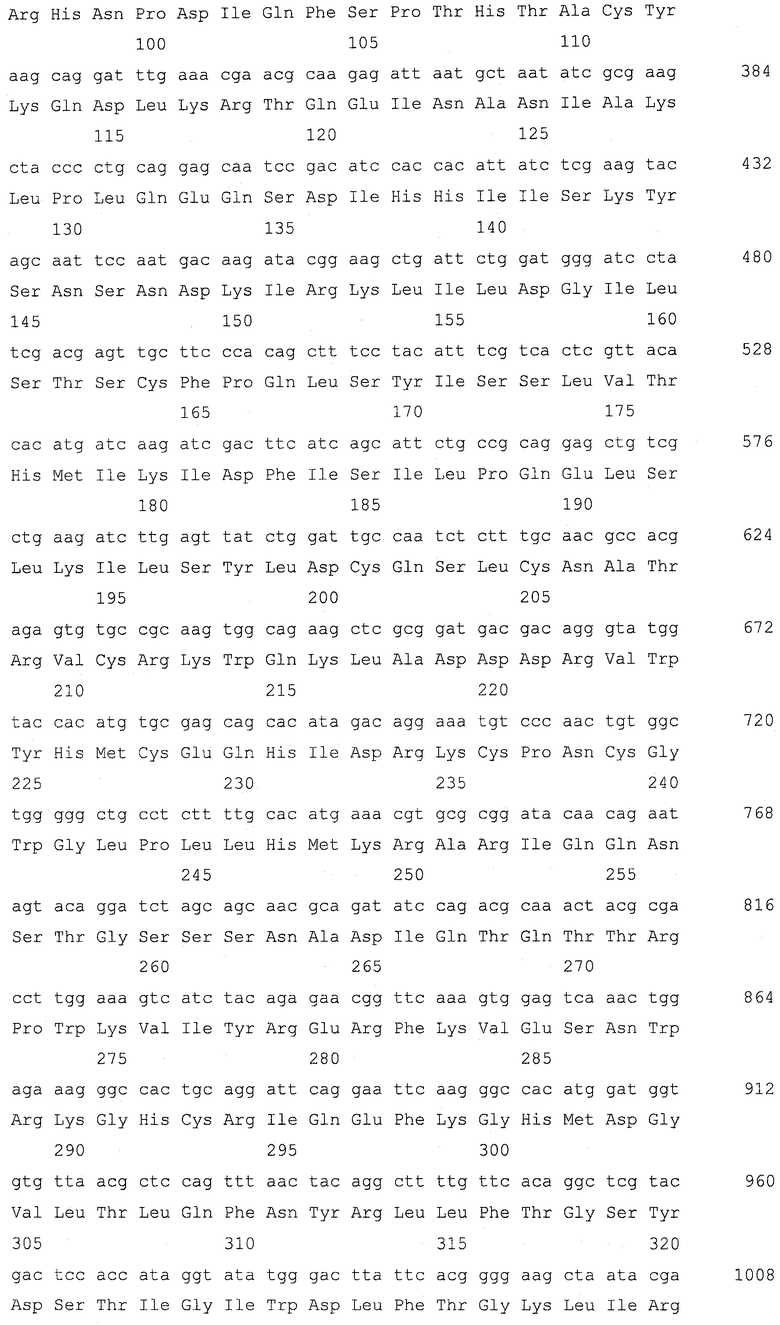
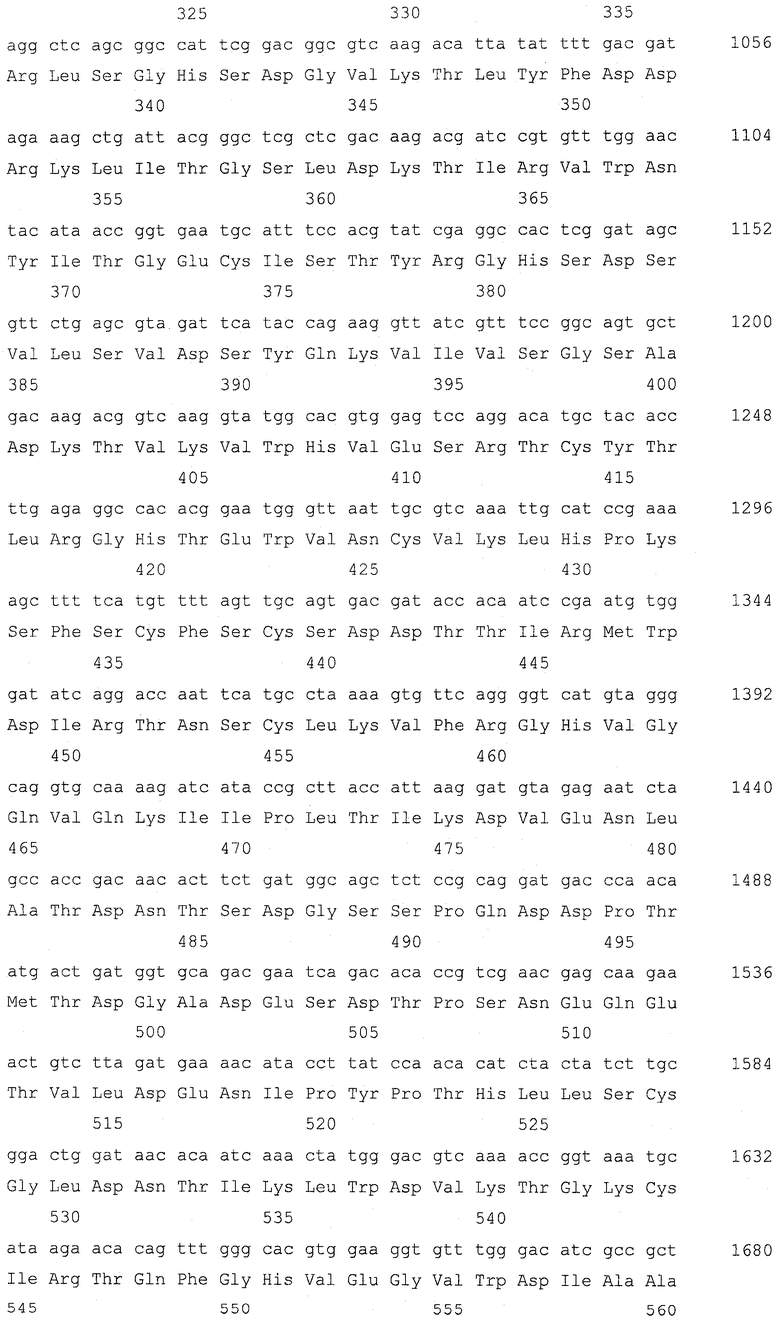
Ген GSH2, кодирующий глутатионсинтетазу, подвергали разрушению в штамме Y-3256, полученном в примере 3, для получения гаплоидного штамма, который аккумулирует γ-глутамилцистеин. Для конструирования кассеты для разрушения гена GSH2 проводили ПЦР, используя праймеры SEQ ID NO: 17-22 и геномную ДНК S.cerevisiae (штамма дикого типа) и плазмиду pFA6a-KanMX6 (Chiara et al., Yeast 2000; 16: 1089-1097) в качестве матриц (фиг. 2). Детальными условиями были следующие условия.
Сначала амплифицировали 5'-район размером приблизительно 400 п.о. от открытой рамки считывания (ОРС) GSH2 с геномной ДНК Х2180-1В (штамма дикого типа S.cerevisiae, доступного из АТСС, с входящим номером АТСС204505) для получения 5'-фрагмента GSH2, используя праймер GSH2-up-F (SEQ ID NO: 17) и праймер GSH2-up-R (SEQ ID NO: 18). 5'-фрагмент GSH2 имел последовательность сайта для фермента рестрикции BpiI на одном конце и последовательность con1 для ПЦР со слиянием, которая описывается позже, на другом конце согласно конструкции праймера GSH2-up-F и праймера GSH2-up-R. Также амплифицировали 3'-район размером приблизительно 300 п.о. от ОРС GSH2 с геномной ДНК Х2180-1В S.cerevisiae для получения 3'-фрагмента GSH2, используя праймер GSH2-down-F (SEQ ID NO: 19) и праймер GSH2-down-R (SEQ ID NO: 20). 3'-фрагмент GSH2 имел последовательность con2 для ПЦР со слиянием, которая описывается позже, на одном конце и последовательность сайта для фермента рестрикции BpiI на другом конце согласно конструкции праймера GSH2-down-F и праймера GSH2-down-R. Затем с помощью ПЦР амплифицировали ген KanMX с плазмиды pFA6a-KanMX6, используя праймер Marker-F (SEQ ID NO: 21) и праймер Marker-R (SEQ ID NO: 22). Этот амплифицированный с помощью ПЦР фрагмент гена KanMX имел последовательность con1 на одном конце и последовательность con2 на другом конце для ПЦР со слиянием согласно конструкции праймера Marker-F и праймера Marker-R. Эти три амплификации с помощью ПЦР проводили при следующих условиях. Для каждой реакции использовали смесь ДНК-полимераз (Pfu:Taq = 1:10, обе из которых доступны от Fermentas, Литва). ПЦР проводили повтором 30 раз цикла, состоящего из 94°С в течение 30 секунд, 50°С в течение 30 секунд и 68°С в течение 3 минут.
Затем эти три полученных фрагмента подвергали лигированию для получения кассеты для разрушения гена GSH2 с помощью ПЦР со слиянием. ПЦР со слиянием проводили, используя праймер GSH2-up-F и праймер Marker-R в качестве праймеров для ПЦР и 5'-фрагмента GSH2 и фрагмента гена KanMX в качестве матриц. Поскольку 5'-фрагмент GSH2 и фрагмент гена KanMX имеют одинаковую последовательность con1 на своих концах, 5'-фрагмент GSH2 и фрагмент гена KanMX подвергались лигированию, и с помощью этой ПЦР со слиянием был получен фрагмент (5'-фрагмент GSH2)-(фрагмент гена KanMX). Эту ПЦР со слиянием проводили с использованием той же смеси ДНК-полимераз. ПЦР проводили повтором 5 раз цикла, состоящего из 94°С в течение 30 секунд, 61°С в течение 30 секунд и 68°С в течение 4,5 минут, и затем повтором 25 раз цикла, состоящего из 94°С в течение 30 секунд, 50οС в течение 30 секунд и 68οС в течение 4,5 минут. Затем провели другую ПЦР со слиянием, используя праймер GSH2-up-F и праймер GSH2-down-R в качестве праймеров для ПЦР и фрагмент (5'-фрагмент GSH2)-(фрагмент гена KanMX) и 3'-фрагмент GSH2 в качестве матриц. Поскольку фрагмент (5'-фрагмент GSH2)-(фрагмент гена KanMX) и 3'-фрагмент GSH2 имеют одинаковую последовательность con2 на своих концах, фрагмент (5'-фрагмент GSH2)-(фрагмент гена KanMX) и 3'-фрагмент GSH2 подвергались лигированию, и была получена кассета для разрушения гена GSH2. Эту ПЦР со слиянием проводили с использованием той же смеси ДНК-полимераз. ПЦР проводили повтором 5 раз цикла, состоящего из 94°С в течение 30 секунд, 61°С в течение 30 секунд и 68°С в течение 5,3 минут, и затем повтором 25 раз цикла, состоящего из 94°С в течение 30 секунд, 50°С в течение 30 секунд и 68°С в течение 5,3 минут.
Последовательности праймеров для ПЦР были следующими последовательностями.
(1) GSH2-up-F, CCGAAGACCTTCGTTTGGTGTTATGGT (SEQ ID NO: 17)
(2) GSH2-up-R, GAGAGGGGGGGGGTGGGGGGAAGGTGGATAGTGTGCC (SEQ ID NO: 18)
(3) GSH2-down-F, CCTCCTCCCCCCGCCCACGGCAGGATTCGGATGTTTG (SEQ ID NO: 19)
(4) GSH2-down-R, CGAAGACTCAGTACGAGCATTACGCAA (SEQ ID NO: 20)
(5) Marker-F, 5`-CCCCACCCCCCCCCTCTCTACCGTTCGTATAATGTATGCTATACGAAGTTATACTGGATGGCGGCGTTAG (SEQ ID NO: 21)
(6) Marker-R, GTGGGCGGGGGGAGGAGGTACCGTTCGTATAGCATACATTATACGAAGTTATGTTTAGCTTGCCTCGTCC (SEQ ID NO: 22)
Штамм Y-3256 трансформировали кассетой для разрушения гена GSH2 для вызова гомологичной рекомбинации, и трансформанты распределяли по агаризованной твердой питательной среды YPD, содержащей G418 (50 мкг/мл). Среди появившихся колоний был получен штамм N8ΔGSH2, в котором разрушен ген GSH2.
Штамм N8ΔGSH2 культивировали в 50 мл питательной среды YPD (20 г/л D-глюкозы, 20 г/л бактопептона, 10 г/л дрожжевого экстракта), содержащейся в колбе Сакагучи объемом 500 мл, при 30°С с покачиванием при 120 оборотах/мин в течение 24 часов, и полученную культуру инокулировали в 50 мл питательной среды YPD, содержащейся в колбе Сакагучи объемом 500 мл, дополненной различными концентрациями аденина (конечная концентрация аденина 0 мг/л, 10 г/л или 20 г/л), чтобы сделать исходную оптическую плотность при 660 нм равной 0,1, и культивировали при 30°С с покачиванием при 120 оборотах/мин. Концентрацию γ-глутамилцистеина определяли в соответствии с общепринятым способом. Таблица 7 демонстрирует временной курс содержания γ-глутамилцистеина. Было обнаружено, что содержание γ-глутамилцистеина можно повысить приданием ауксотрофии по аденину.
ПРИМЕР 8
(Оценка на питательной среде, которая имитирует состояния ауксотрофии по аденину)
В соответствии с общепринятым способом штамм AJ14889 подвергали обработке мутагеном MNNG, вследствие чего коэффициент выживаемости становился равным приблизительно 10%, и обработанные клетки распределяли по чашке с агаризованной средой min-met(+)-biotin(-), которая содержит представленные в таблице 8 компоненты и метионин в концентрации 150 мг/л, вследствие чего должно появляться не более 350 колоний. После культивирования при 30°С в течение восьми-десяти дней среди колоний, появившихся на 30 планшетах, было отобрано 106 колоний от светло-розового до красного цвета.
Затем определяли внутриклеточное содержание GSH в отобранных штаммах и штамме AJ14889 (исходном штамме) при их культивирования на питательной среде YPD. Т.е. отобранные штаммы, а также штамм AJ14889, подвергнутые культивированию на чашке с питательной средой YPD, инокулировали соответственно в 5 мл жидкой питательной среды YPD, содержащейся в пробирке, и культивировали при 30°С с покачиванием при 250 оборотах/минуту в течение 24 часов. Затем полученную культуру каждого штамма инокулировали в 50 мл жидкой питательной среды YPD, содержащейся в конической колбе объемом 750 мл, так чтобы оптическая плотность при 600 нм стала равной 0,1, и культивировали при 30°С с покачиванием при 250 оборотах/минуту в течение 24 и 48 часов, соответственно. В результате было установлено, что среди 106 отобранных штаммов восемь штаммов демонстрируют содержание GSH, превышающее содержание GSH в штамме AJ14889 (таблица 9) через 24 и/или 48 часов. Установлено, что этот способ отбора является очень эффективным для получения штамма, имеющего высокое содержание GSH, поскольку восемь штаммов среди 108 штаммов продемонстрировали повышенное содержание GSH.
Затем из группы из восьми штаммов были отобраны штамм 89-6, штамм 89-28 и штамм 89-31, и их рост и содержание в них GSH сравнивали с ростом и содержанием GSH в штамме AJ14889. Эти штаммы культивировали, как описано выше, за исключение того, что клетки инокулировали так, чтобы оптическая плотность при 600 нм стала равной 0,3. Рост определяли на основе сухого клеточного веса (DCW). Как продемонстрировано в таблице 10, три штамма продемонстрировали почти такой же рост, как и штамм AJ14889, и содержание GSH, превышающее содержание GSH в штамме AJ14889 через 24 и 48 часов.
Для исследования вероятности того, что этим трем штаммам была придана неожиданная способность - ауксотрофия по питательному веществу, эти три штамма и штамм AJ14889 инокулировали в питательную среду SD, и определяли их рост. Конкретно, штаммы, подвергнутые культивированию на чашке с питательной средой YPD, инокулировали соответственно в 5 мл питательной среды SD, содержащейся в пробирке, и культивировали при 30°С с покачиванием при 250 оборотах/минуту в течение 24 и 48 часов, соответственно. В результате было установлено, что рост трех штаммов является почти таким же, как и рост штамма AJ14889 в среде SD (таблица 11), что означает, что этим трем штаммам не была придана неожиданная способность - ауксотрофия по питательному веществу.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
В соответствии с настоящим изобретением предоставляются ауксотрофные по аденину дрожжи, имеющие повышенное содержание серосодержащего соединения в клетках, способ отбора дрожжей и способ культивирования дрожжей. Настоящее изобретения может широко использоваться в области продуктов питания, лекарственных средств, химических продуктов, кормов для животных и т.п.
| название | год | авторы | номер документа |
|---|---|---|---|
| Штамм дрожжей Saccharomyces cerevisiae для тестирования мутагенов окружающей среды | 2020 |
|
RU2757018C1 |
| СПОСОБ ПОЛУЧЕНИЯ ВОДНОГО РАСТВОРА МЕДА И СПОСОБ ПРОВЕРКИ ЕГО ПОДЛИННОСТИ | 2012 |
|
RU2506813C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ПРОЯВЛЯЮЩАЯ АНТИМУТАГЕННОЕ ДЕЙСТВИЕ (ВАРИАНТЫ) | 2012 |
|
RU2477142C1 |
| Штамм Saccharomyces cerevisiae 1-TAE-2 для тестирования мутагенов окружающей среды | 2023 |
|
RU2825780C1 |
| ПРИМЕНЕНИЕ ШТАММА ДРОЖЖЕЙ Komagataella pastoris В КАЧЕСТВЕ РЕЦИПИЕНТА ДЛЯ КОНСТРУИРОВАНИЯ ПРОДУЦЕНТОВ ЦЕЛЕВОГО БЕЛКА | 2013 |
|
RU2522479C1 |
| ВАРИАНТ ГЛУТАМАТ-ЦИСТЕИНЛИГАЗЫ И СПОСОБ ПОЛУЧЕНИЯ ГЛУТАТИОНА С ЕГО ПРИМЕНЕНИЕМ | 2021 |
|
RU2809326C1 |
| Новый промотор и способ получения глутатиона с его использованием | 2021 |
|
RU2806289C1 |
| Трансформант дрожжей Pichia pastoris, продуцирующий ксиланазу | 2019 |
|
RU2714113C1 |
| Способ определения мутагенной активности химических веществ | 2022 |
|
RU2810923C1 |
| Штамм дрожжей Yarrowia lipolytica - продуцент янтарной кислоты (варианты) | 2016 |
|
RU2631922C1 |

















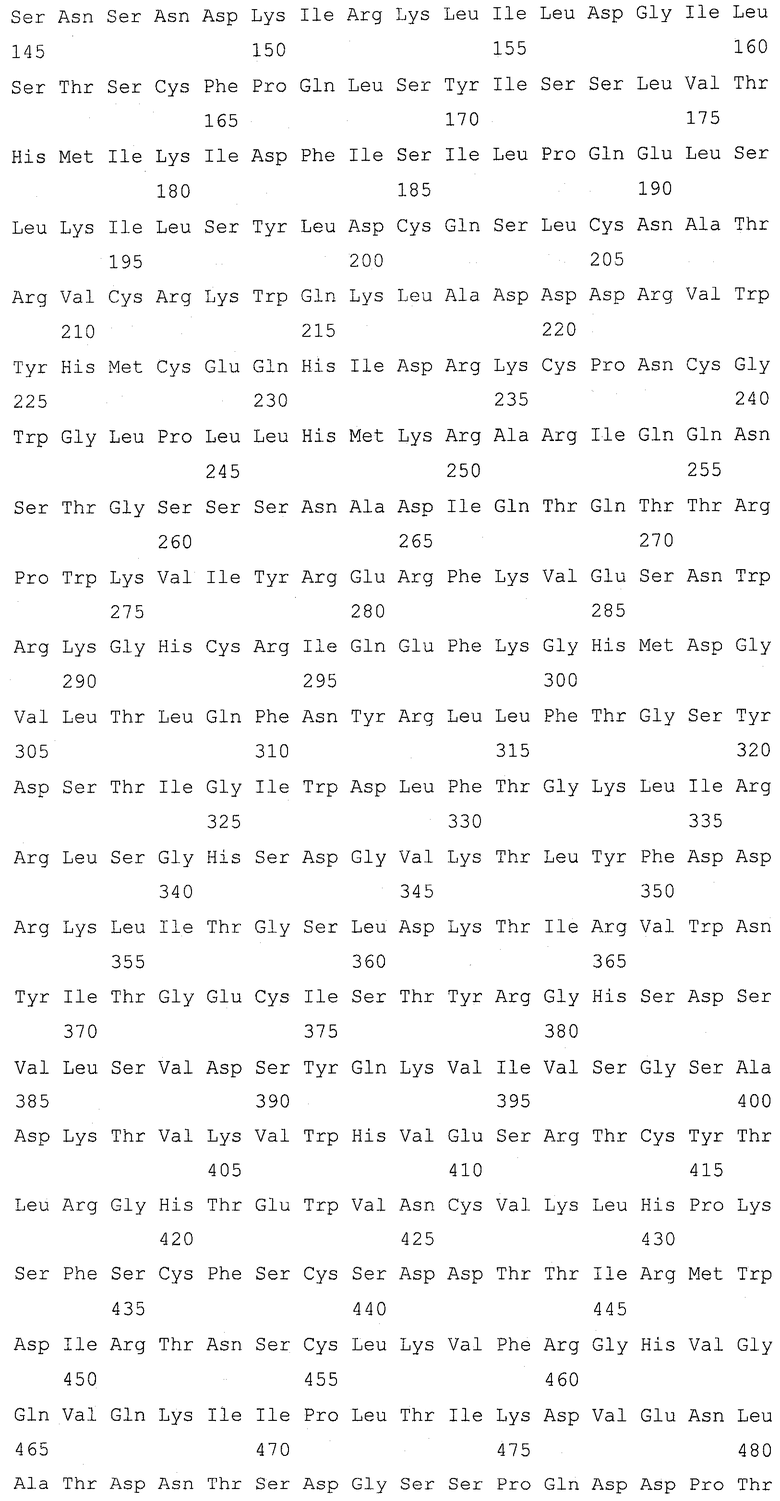





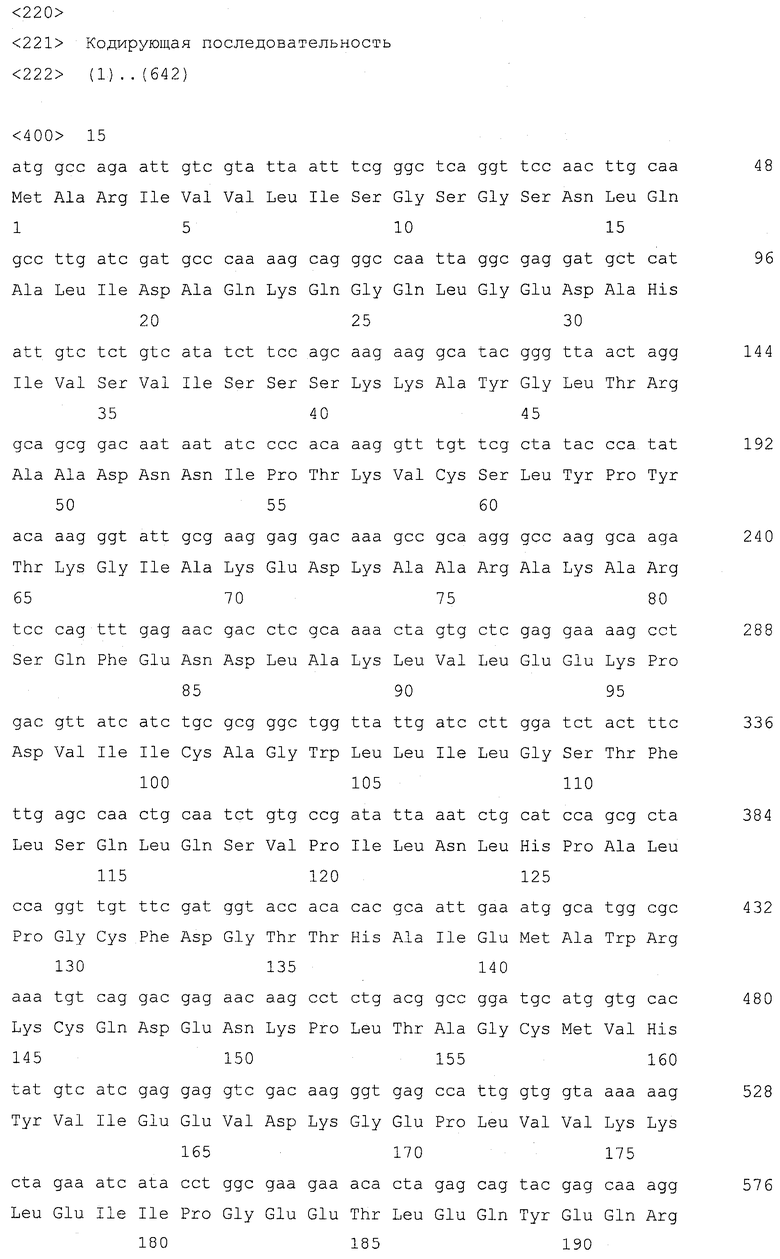



Изобретение относится к биотехнологии и представляет собой способ получения дрожжей, имеющих повышенное клеточное содержание серосодержащего соединения, выбранного из группы, состоящей из цистеина, γ-глутамилцистеина, глутатиона и цистеинилглицина, по сравнению с родительским штаммом дрожжей который демонстрирует ауксотрофию по аденину и имеет модификацию для усиления экспрессии гена МЕТ25, включающий подвергание родительского штамма обработке агентом для мутагенеза, распределение модифицированных дрожжей по питательной среде, имеющей содержание аденина 25 мг/л или менее, для образования колоний дрожжей, отбор колонии дрожжей, которая краснее по сравнению с родительским штаммом до модификации, и отбор дрожжей, имеющих повышенное клеточное содержание указанного серосодержащего соединения по сравнению с родительским штаммом. Изобретение позволяет эффективно получить дрожжи с высоким содержанием серосодержащего соединения. 3 н. и 3 з.п. ф-лы, 2 ил., 11 табл., 8 пр.
1. Способ получения дрожжей, имеющих повышенное клеточное содержание серосодержащего соединения, выбранного из группы, состоящего из цистеина, γ-глутамилцистеина, глутатиона и цистеинилглицина, по сравнению с родительским штаммом дрожжей который демонстрирует ауксотрофию по аденину и имеет модификацию для усиления экспрессии гена МЕТ25, включающий:
подвергание родительского штамма обработке агентом для мутагенеза,
распределение модифицированных дрожжей по питательной среде, имеющей содержание аденина 25 мг/л или менее, для образования колоний дрожжей,
отбор колонии дрожжей, которая краснее по сравнению с родительским штаммом до модификации, и
отбор дрожжей, имеющих повышенное клеточное содержание указанного серосодержащего соединения по сравнению с родительским штаммом.
2. Способ по п. 1, в котором указанные дрожжи дополнительно модифицированы для снижения межклеточной активности глутатион-синтетазы.
3. Способ получения дрожжей, имеющих повышенное клеточное содержание серосодержащего соединения, выбранного из группы, состоящего из цистеина, γ-глутамилцистеина, глутатиона и цистеинилглицина, по сравнению с родительским штаммом дрожжей который демонстрирует ауксотрофию по аденину и имеет модификацию для усиления экспрессии гена МЕТ25, включающий:
подвергание родительского штамма обработке агентом для мутагенеза,
распределение модифицированных дрожжей по минимальной питательной среде, дополненной метионином, для образования колоний дрожжей,
отбор колонии дрожжей, которая краснее по сравнению с родительским штаммом до модификации, и
отбор дрожжей, имеющих повышенное клеточное содержание указанного серосодержащего соединения по сравнению с родительским штаммом.
4. Способ по п. 3, в котором указанные дрожжи дополнительно модифицированы для снижения межклеточной активности глутатион-синтетазы.
5. Способ получения серосодержащего соединения, выбранного из группы, состоящего из цистеина, γ-глутамилцистеина, глутатиона и цистеинилглицина, включающий культивирование дрожжей, полученных способом по любому из пп. 1-4.
6. Способ по п. 5, в котором указанные дрожжи культивируют в богатых аденином условиях при содержании аденина 100 мг/л или более для увеличения количества дрожжевых клеток, а затем культивируют в бедных аденином условиях при содержании аденина 25 мг/л или менее для увеличения клеточного содержания серосодержащего соединения, выбранного из группы, состоящего из цистеина, γ-глутамилцистеина, глутатиона и цистеинилглицина.
| SHARMA KG The glutathione-mediated detoxification pathway in yeast: an analysis using the red pigment that accumulates in certain adenine biosynthetic mutants of yeasts reveals the involvement of novel genes, Arch Microbiol | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| JP 2004180509 A, 02.07.2004 | |||
| Устройство для гашения вибрации | 1986 |
|
SU1428873A2 |
| ОПРЕДЕЛЕНИЕ ВНУТРИКЛЕТОЧНЫХ КОНЦЕНТРАЦИЙ ЦИСТЕИНА И ГЛУТАТИОНА | 1997 |
|
RU2216020C2 |

