ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается преимущество приоритета в соответствии с предварительной заявкой США №63/044478, поданной 26 июня 2020 г., и предварительной заявкой США №62/991806, поданной 19 марта 2020 г., содержание каждой из которых в полном объеме включено в данный документ посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
В настоящей заявке содержится список последовательностей, который был представлен в электронном виде в формате ASCII и полностью включен в данный документ посредством ссылки. Указанная копия ASCII, созданная 16 марта 2021 г., имеет название P35791-WO_Sequence_Listing и размер 492334 байт.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к селективным в отношении изоформы антителам к TGFβ (например, моноспецифическим антителам к TGFβ2 и антителам к TGFβ3, и антителам с двойной специфичностью к TGFβ2/3) и способам их применения, например, для лечения нарушений, связанных с TGFβ.
УРОВЕНЬ ТЕХНИКИ
TGFβ включает плейотропный набор из трех цитокинов - TGFβ1, TGFβ2 и TGFβ3, которые играют решающие роли в дифференцировке клеток, развитии тканей, заживлении ран, иммунорегуляции, а в случае нарушения регуляции в фиброзе тканей. В случае интерстициальных заболеваний легких (ILD), таких как идиопатический легочный фиброз (IPF), активность TGFβ играет роль во многих аспектах патогенеза заболеваний. Генетический риск IPF обусловлен мутациями в генах, экспрессируемых в эпителиальных клетках легких, которые повышают их восприимчивость к повреждению и/или нарушают их регенеративную способность. Этот эпителиальный стресс или повреждение могут активировать врожденные иммунные клетки, такие как альвеолярные макрофаги, с целью продуцирования цитокинов, которые активируют мезенхимальные клетки с инициированием реакции заживления ран посредством пролиферации, миграции, дифференцировки в миофибробласты и секреции внеклеточного матрикса (ЕСМ). TGFβ может содействовать многим из этих процессов; в частности, было показано, что он способствует апоптозу эпителиальных клеток легких, при этом способствуя активации, дифференцировке и выживанию миофибробластов. Системный склероз (SSc)/склеродермия представляет собой аутоиммунное заболевание, которое начинается с микрососудистого воспаления, прогрессирующего до полиорганной дисфункции соединительной ткани, включающей ткани кожи, легких, сердца, почек и кишечника. TGFβ участвует в нарушении регуляции сосудистой, соединительной ткани и иммунных компонентов при SSc (Lafyatis R. Nat Rev Rheumatol. 2014 Dec; 10(12):706-19).
Передача сигналов TGFβ также играет роль в патогенезе рака, особенно в перитуморальной строме и иммунных компартментах, где он может ингибировать продуктивные противоопухолевые иммунные ответы как посредством стимуляции избыточного продуцирования внеклеточного матрикса, которое предупреждает инфильтрацию Т-клеток в опухолевые ткани, так и посредством стимуляции дифференцировки и активации Т-регуляторных клеток, которые могут подавлять противоопухолевый иммунитет. В совокупности эти данные указывают на то, что TGFβ представляет собой потенциальную терапевтическую мишень при фиброзных заболеваниях и раке. Однако многообразные гомеостатические функции TGFβ и сложность контекстно-зависимых механизмов активации TGFβ способствовали ограничениям как с точки зрения безопасности, так и эффективности для установления благоприятного терапевтического индекса в интервенционных исследованиях ингибиторов TGFβ при фиброзных нарушениях человека. Например, ингибиторы всех видов TGFβ были связаны с нежелательными сигналами безопасности. В частности, низкомолекулярные ингибиторы ALK5 AZ12601011 и AZ12799734 (Anderton et al. (2011) Toxicologic Pathology, 39: 916-924) вызывали микроскопические поражения сердечных клапанов у крыс, а антитело 1D11 ко всем видам TGFβ (Lonning et al. (2011) Current Pharmaceutical Biotechnology, 12, 2176-2189) вызывали у мышей, обработанных 1D11, развитие гистологических поражений, потерю веса, неопухолевую кистозную эпителиальную гиперплазию и воспаление языка, дисплазию зубов и гиперплазию эпителия десен и пищевода. Кроме того, САТ-192 (метелимумаб), антитело, преимущественно селективное в отношении TGFβ1, имело высокую частоту серьезных нежелательных явлений с множественными желудочными кровотечениями, наблюдаемыми в исследовании фазы 1-2 при SSc (Denton A&R 56:323 (2007)).
Таким образом, в данной области техники сохраняется потребность в безопасных и эффективных молекулах, которые нацелены на TGFβ. В настоящем изобретении представлены такие молекулы и соответствующие варианты применения.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В настоящем изобретении представлены селективные в отношении изоформы антитела к TGFβ и способы их применения.
В одном аспекте представлено выделенное антитело к TGFβ3, где указанное антитело содержит: (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9. В некоторых аспектах антитело селективно нейтрализует TGFβ3.
В другом аспекте представлено выделенное антитело к TGFβ3, где антитело селективно нейтрализует TGFβ3, и при этом антитело характеризуется одним или более из следующих свойств: (а) антитело к TGFβ3 специфически связывается со шпилечной областью бета6/бета7 TGFβ3; (б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3; (в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFBR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3; (г) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека; (д) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи; (е) изоформная селективность антитела к TGFβ3 к TGFβ3 по сравнению с TGFβ1 достигается за счет прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека); (ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается за счет прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека); (и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 к TGFβ; (к) антитело к TGFβ3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 к TGFβ; (л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; (м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; (н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом САТ-192 к TGFβl; (п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3; (р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, К331, W332, Н334, Е335, Т387, 1388, L389, Y391, V392, G393, R394, Р396, К397 и V398 TGFβ3 человека; и (т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3.
В другом аспекте представлено выделенное антитело к TGFβ3, где антитело селективно нейтрализует TGFβ3, и при этом антитело характеризуется одним или более из следующих свойств: (а) антитело к TGFβ3 специфически связывается со шпилечной областью бета6/бета7 TGFβ3; (б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3; (в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFBR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3; (г) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека; (д) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи; (е) изоформная селективность антитела к TGFβ3 к TGFβ3 по сравнению с TGFβ1 достигается за счет прямого контакта антигенсвязывающего домена антитела к TGFP3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека); (ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается за счет прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека); (и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 к TGFβ; (к) антитело к TGFβ3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 к TGFβ; (л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; (м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; (н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом САТ-192 к TGFβl; (п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3; (р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 5 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, К331, W332, Н334, Е335, Т387, 1388, L389, Y391, V392, G393, R394, Р396, K397 и V398 TGFβ 33 человека; и (т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность вариабельной области тяжелой цепи (VH), имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37 и 42-50. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 23, 37 и 42-50. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность тяжелой (Н) цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 59, 64 и 65-72. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность тяжелой (Н) цепи, выбранную из группы, состоящей из SEQ ID NO: 29, 59, 64 и 65-72. В дополнительных аспектах антитело к TGFβ3 содержит аминокислотную последовательность вариабельной области легкой цепи (VL), имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41. В еще одних аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность легкой (L) цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63. В некоторых аспектах антитело к TGFβ3 содержит пару VH/VL, где пара VH/VL содержит аминокислотные последовательности (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2A10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9). В некоторых аспектах антитело к TGFβ3 содержит полную пару H/L-цепей, при этом полная пара H/L-цепей содержит аминокислотные последовательности (соответственно), выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.6), SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8) и SEQ ID NO: 72/58 (v2.9). В некоторых аспектах антитело к TGFβ3 содержит VL SEQ ID NO: 22, содержащую одну или несколько модификаций каркасной области, выбранных из группы, состоящей из: 4L или 4М, 38Н или 38Q, 43А или 43Q и 58V. В некоторых аспектах VL антитело к TGFβ3 содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 4L в FR1, 38Н и 43Q в FR2, 581 в FR3 (h2A10.v1 и h2A10.v2); (ii) 4М в FR1 (h2A10.v1.1 и h2A10.v2.1); (iii) 38Q в FR2 (h2A10.v1.2 и h2A10.v2.2); (iv) 43А в FR2 (h2A10.v1.3 и h2A10.v2.3); (v) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); (vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4); (vii) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); и (vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4), где мутации представлены по отношению к VL 2А10 крысы, содержащей SEQ ID NO: 22. В других аспектах антитело к TGFβ3 содержит VH SEQ ID NO: 23, содержащую одну или несколько модификаций каркасной области, выбранных из группы, состоящей из: 47L или 47W; 49А, 49S или 49G; 73D или 73N; и 76N, 78D или 78L, 78А или 78V. В некоторых аспектах VH антитела к TGFβ3 содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 47L, 49А в FR2, 78V в FR3 (h2A10.v1); (ii) 47L, 49А в FR2, 73D, 76S, 78V в FR3 (h2A10.v2); (iii) 47W в FR2 (h2A10.v1.5); (iv): 49G в FR2 (h2A10.v1.6); (v) 78A в FR3 (h2A10.v1.7); (vi) 47W в FR2 (h2A10.v2.5); (vii) 49S в FR2 (h2A10.v2.6); (viii) 73N в FR3 (h2A10.v2.7); (ix) 76N в FR3 (h2A10.v2.8); (x) 78L в FR3 (h2A10.v2.9); и (xi) 49S в FR2, 76N, 78L в FR3 (h2A10.v3 и h2A10.v4), где мутации представлены по отношению к VH 2А10 крысы, содержащей SEQ ID NO: 23.
В определенных вариантах осуществления вышеуказанных антител к TGFJ33 VL антитело к TGFβ3 сохраняет лейцин (L) в положении 4 в каркасной области I и лейцин (L) в положении 47 в каркасной области II (по отношению к 2А10 VL крысы, содержащей SEQ ID №: 22). В некоторых вариантах осуществления VH сохраняет D в положении 73 в каркасной области III VH 2А10 крысы (по отношению к VH 2А10, содержащей SEQ ID NO: 23).
В другом аспекте представлено выделенное антитело к TGFβ3, где антитело селективно нейтрализует TGFP3, и при этом антитело характеризуется одним или более из следующих свойств: (а) антитело к TGFβ3 специфически связывается со шпилечной областью бета6/бета7 TGFβ3; (б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3; (в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFBR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3; (г) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека; (д) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи; (е) изоформная селективность антитела к TGFβ3 к TGFβ3 по сравнению с TGFβ1 достигается за счет прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека); (ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается за счет прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека); (и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 к TGFβ; (к) антитело к TGFβ3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 к TGFβ; (л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; (м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; (н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом САТ-192 к TGFβ1; (п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3; (р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 159 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, К331, W332, Н334, Е335, Т387, 1388, L389, Y391, V392, G393, R394, Р396, К397 и V398 TGFβ3 человека; и (т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3. В некоторых аспектах этого варианта осуществления антитело к TGFβ3 содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 52. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VH SEQ ID NO: 52. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 74. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 22, 36 и 38-41. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63. В дополнительных аспектах антитело к TGFβ3 содержит пару VH/VL, содержащую аминокислотные последовательности (соответственно) SEQ ID NO: 52/36. В некоторых аспектах антитело к TGFβ3 содержит полную пару H/L-цепей, при этом пара H/L-цепей содержит аминокислотные последовательности (соответственно) SEQ ID NO: 74/58.
В другом аспекте представлено выделенное антитело к TGFβ3, где антитело селективно нейтрализует TGFβ3, и при этом антитело характеризуется одним или более из следующих свойств: (а) антитело к TGFβ3 специфически связывается со шпилечной областью бета6/бета7 TGFβ3; (б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3; (в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFBR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3; (г) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека; (д) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи; (е) изоформная селективность антитела к TGFβ3 к TGFβ3 по сравнению с TGFβ1 достигается за счет прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека); (ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается за счет прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека); (и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 к TGFβ; (к) антитело к TGFβ3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 к TGFβ; (л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; (м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; (н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом САТ-192 к TGFβl; (п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3; (р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 34 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, К331, W332, Н334, Е335, Т387, 1388, L389, Y391, V392, G393, R394, Р396, К397 и V398 TGFβ3 человека; и (т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 51 или 55. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VH SEQ ID NO: 51 или 55. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 73 или 77. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 73. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 77. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 22, 36 и 38-41. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63. В некоторых аспектах антитело к TGFβ3 содержит пару VH/VL, где пара VH/VL содержит аминокислотные последовательности (соответственно) SEQ ID NO: 51/36 или SEQ ID NO: 55/54. В некоторых аспектах антитело к TGFβ3 содержит полную пару H/L-цепей, при этом пара H/L-цепей содержит аминокислотные последовательности (соответственно) SEQ ID NO: 73/58 или SEQ ID NO: 77/76.
В другом аспекте представлено выделенное антитело к TGFβ3, где антитело селективно нейтрализует TGFβ3, и при этом антитело характеризуется одним или более из следующих свойств: (а) антитело к TGFβ3 специфически связывается со шпилечной областью бета6/бета7 TGFβ3; (б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3; (в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFBR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3; (г) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека; (д) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи; (е) изоформная селективность антитела к TGFβ3 к TGFβ3 по сравнению с TGFβ1 достигается за счет прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека); (ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается за счет прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека); (и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 к TGFβ; (к) антитело к TGFβ3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 к TGFβ; (л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; (м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; (н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом САТ-192 к TGFβ1; (п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3; (р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, К331, W332, Н334, Е335, Т387, 1388, L389, Y391, V392, G393, R394, Р396, K397 и V398 TGFβ3 человека; и (т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 53 или 57. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VH SEQ ID NO: 53. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VH SEQ ID NO: 57. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 75 или 79. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 75. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 79. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 22, 36 и 38-41. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63. В некоторых аспектах антитело к TGFβ3 содержит пару VH/VL, содержащую аминокислотные последовательности (соответственно) SEQ ID NO: 53/36 или SEQ ID NO: 57/56. В некоторых аспектах антитело к TGFβ3 содержит полную пару H/L-цепей, при этом пара H/L-цепей содержит аминокислотные последовательности (соответственно) SEQ ID NO: 75/58 или SEQ ID NO: 79/78.
В другом аспекте представлено антитело к TGFβ3, при этом антитело содержит: (a) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-Н1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (б) пару VH/VL, VH пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 57, и VL пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 56; или (в) полную пару H/L-цепей, при этом Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79, а L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78.
В другом аспекте представлено выделенное антитело к TGFβ 32/3, где антитело селективно нейтрализует TGFβ2 и TGFβ3, и при этом антитело характеризуется одним или более из следующих свойств: (а) селективность антитела к TGFβ2/3 в отношении TGFβ2 и TGFβ3 по сравнению с TGFβ1 человека в отношении селективной нейтрализации достигается посредством прямого контакта антигенсвязывающего домена антитела с аминокислотным остатком Е373 TGFβ2 или TGFβ3 (нумерация TGFβ2 человека); (б) антитело к TGFβ2/3 нейтрализует TGFβ2 и/или TGFβ3 посредством аллостерического механизма; (в) антитело к TGFβ2/3 индуцирует конформационные изменения в гомодимере TGFβ2 и/или TGFβ3; (г) антитело к TGFβ2/3 индуцирует конформационное изменение в гомодимере TGFβ2 и/или TGFβ3, где конформационное изменение включает совместное сжатие двух мономеров на несколько градусов; (д) антитело к TGFβ2/3 представляет собой двухвалентное антитело или одновалентное антитело; (е) антитело к TGFβ2/3 содержит (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 15; (ж) антитело к TGFβ2/3 специфически связывается с гомодимером TGFβ2, при этом гомодимер TGFβ2 имеет первый и второй мономер TGFβ2, и при этом антитело к TGFβ2/3 содержит антигенсвязывающий домен, который непосредственно контактирует с (i) аминокислотными остатками V313, Q314, D315, R320, L322, Y323, R328, D329, F345 и А347 первого мономера TGFβ2, и (ii) аминокислотными остатками N368, Т369, 1370, N371, Р372, Е373, А374, S375, А376 и S377 второго мономера TGFβ2 (нумерация TGFβ2 человека); (и) антитело к TGFβ2/3, как и в (ж), где антигенсвязывающий домен находится в пределах 5 ангстрем от аминокислотных остатков TGFβ2 и/или TGFβ3; (к) при этом антитело к TGFβ2/3 специфически связывается с тем же эпитопом TGFβ3, что и в (ж); и (л) антитело к TGFβ2/3 не нейтрализует TGFβ2 и/или TGFβ3 в одновалентной форме. В одном аспекте антитело к TGFβ2/3 содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 27, 81, 83, 86-88, 93-100 и 102-105. В одном аспекте антитело к TGFβ2/3 содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 27, 81, 83, 86-88, 93-100 и 102-105. В одном аспекте антитело к TGFβ2/3 содержит полную аминокислотную Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 33, 107, 109, 112-114 и 119-130. В одном аспекте антитело к TGFβ2/3 содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 26, 80, 82, 84, 85, 89-92 и 101. В одном аспекте антитело к TGFβ2/3 содержит аминокислотную последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 26, 80, 82, 84, 85, 89-92 и 101. В одном аспекте антитело к TGFβ2/3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32, 106, 108, ПО, 111, 115-118 и 186. В одном аспекте антитело к TGFβ2/3 содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 32, 106, 108, 110, 111, 115-118 и 186. В одном аспекте антитело к TGFβ2/3 содержит пару VH/VL, при этом пара VH/VL содержит аминокислотные последовательности (соответственно), выбранные из группы, состоящей из SEQ ID NO: 27/26 (4А11 кролика), SEQ ID NO: 81/80 (v1), SEQ ID NO: 81/82 (v2), SEQ ID NO: 83/80 (v3), SEQ ID NO: 83/82 (v4), SEQ ID NO: 81/84 (v5), SEQ ID NO: 81/85 (v6), SEQ ID NO: 83/84 (v7), SEQ ID NO: 86/84 (v7/1), SEQ ID NO: 87/84 (v7.2), SEQ ID NO: 88/84 (v7.3), SEQ ID NO: 83/89 (v7.4), SEQ ID NO: 83/90 (v7.5), SEQ ID NO: 83/91 (v7.6), SEQ ID NO: 83/92 (v7.7), SEQ ID NO: 93/84 (v7.8), SEQ ID NO: 94/84 (v7.9), SEQ ID NO: 95/84 (v7.10), SEQ ID NO: 96/84 (v7.11), SEQ ID NO: 97/84 (v7.12), SEQ ID NO: 98/84 (v7.13), SEQ ID NO: 99/84 (v7.14), SEQ ID NO: 100/84 (v7.15), SEQ ID NO: 102/101 (v7.16), SEQ ID NO: 103/101 (v7.17), SEQ ID NO: 104/101 (v7.18), SEQ ID NO: 105/101 (v7.19) и SEQ ID NO: 83/85 (v8). В одном аспекте антитело к TGFβ2/3 содержит полную пару H/L-цепей, при этом полная пара H/L-цепей содержит аминокислотные последовательности (соответственно), выбранные из группы, состоящей из SEQ ID NO: 32/33 (4А11 кролика), SEQ ID NO: 107/106 (v1), SEQ ID NO: 107/108 (v2), SEQ ID NO: 109/106 (v3), SEQ ID NO: 109/108 (v4), SEQ ID NO: 107/110 (v5), SEQ ID NO: 107/111 (v6), SEQ ID NO: 109/110 (v7), SEQ ID NO: 112/110 (v7.1), SEQ ID NO: 113/110 (v7.2), SEQ ID NO: 114/110 (v7.3), SEQ ID NO: 114/115 (v7.4), SEQ ID NO: 114/116 (v7.5), SEQ ID NO: 114/117 (v7.6), SEQ ID NO: 114/118 (v7.7), SEQ ID NO: 119/110 (v7.8), SEQ ID NO: 120/110 (v7.9), SEQ ID NO: 121/110 (v7.10), SEQ ID NO: 122/110 (v7.11), SEQ ID NO: 123/110 (v7.12), SEQ ID NO: 124/110 (v7.13), SEQ ID NO: 125/110 (v7.14), SEQ ID NO: 126/110 (v7.15), SEQ ID NO: 127/186 (v7.16), SEQ ID NO: 128/186 (v7.17), SEQ ID NO: 129/186 (v7.18), SEQ ID NO: 130/186 (v7.19) и SEQ ID NO: 114/111 (v8). В одном аспекте антитело к TGFβ2/3 содержит VL, содержащую аминокислотную последовательность SEQ ID NO: 26, содержащую одну или несколько модификаций каркасной области, выбранных из группы, состоящей из: 2А или 21, 4L, 36F или 36Y, 43Р или 43А и 58V или 581. В одном аспекте VL антитело к TGFβ2/3 содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 2А и 4L в FR1 и 36F в FR2 (h4A11.v1 и h4A11.v3); (ii) 2А и 4L в FR1 и 36F и 43Р в FR2 (h4A11.v2 и h4A11.v4); (iii) 2А в FR1, 36F и 43Р в FR2 и 58V в FR3 (h4A11.v5 и h4A11.v7); (iv) 2А и 4L в FR1 и 36F в FR2 (h4A11.v6 и h4A11.v8); (v) 21 в FR1 (h4A11.v7.4); (vi) 36Y в FR2 (h4A11.v7.5); (vii) 43A в FR2 (h4A11.v7.6); (viii) 581 в FR3 (h4A11.v7.7); и (ix) 21 в FR1, 43A в FR2, 581 в FR3 (h4A11.v7.16-19), где мутации представлены по отношению к VL, содержащей аминокислотную последовательность SEQ ID NO: 26. В одном аспекте антитело к TGFβ2/3 содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 27, содержащую одну или более модификаций каркасной области, выбранных из группы, состоящей из: делеции IE, 2Q или 2V, 24V, 37V или 371, 481, 49G, 67F или 67V, 71К или 71V, 73S или 73Т, делеции 75К и 76N, 78V или 78F, 91F или 91Y, 105Р или 105Q. В одном аспекте VL антитело к TGFβ2/3 содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 2Q и 24V в FR1, 481 и 49G в FR2, 71К, 73S, 78V и 91F в FR3 и 105Р в FR4 (h4A11.v1, h4A11.v2, h4A11.v5, h4A11.v6); (ii) 2Q в FR1, 37V в FR2, 67F, 71K, 73S, 78V и 91F в FR3 и 105P в FR4 (h4A11.v3, h4A11.v4, h4A11.v7, h4A11.v8); (iii) делеции IE в FR1 (h4A11.v7.1); (iv) делеции 75K и 76N в FR3 (h4A11.v7.2); (v) делеции IE в FR1 и 75K76N в FR3 (h4A11.v7.3); (vi) 2V в FR1 (h4A11.v7.8); (vi) 371 в FR2 (h4A11.v7.9); (vii) 67V в FR3 (h4Al l.v7.10); (viii) 71V в FR3 (h4A11.v7.11); (ix) 73T в FR3 (h4A11.v7.12); (x) 78F в FR3 (h4A11.v7.13); (xi) 91Y в FR3 (h4A11.v7.14); (xii) 105Q в FR4 (h4A11.v7.15); (xiii) 2V в FR1, 371 в FR2, 67V, 73T, 78F в FR3, 105Q в FR4 (h4A11.v7.16); (xiv) 2V в FR1, 371 в FR2, 67V, 73T, 91Y в FR3, 105Q в FR4 (h4A11.v7.17); (xv) 2V в FR1, 371 в FR2, 67V, 73T в FR3, 105Q в FR4 (h4A11.v7.18); и (xvi) 2V в FR1, 371 в FR2, 67V, 73T, делеции 75K и 76N в FR3, 105Q в FR4 (h4A11.v7.19), где мутации представлены по отношению к VH, содержащей аминокислотную последовательность SEQ ID NO: 27.
В другом аспекте представлено выделенное антитело к TGFβ2, где указанное антитело содержит: (a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 18; и (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21. В одном аспекте антитело к TGFβ2 селективно нейтрализует TGFβ2. В некоторых аспектах антитело к TGFβ2: (а) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ; (б) имеет сниженную токсичность у грызунов по сравнению с антителом 1D11 ко всем видам TGFβ; (в) имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; и/или (г) имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 132 и 138-142. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 25, 132 и 138-142. В одном аспекте антитело к TGFβ2 содержит полную аминокислотную Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 31, 146 и 152-156. В одном аспекте антитело к TGFβ2 содержит полную аминокислотную последовательность Н-цепи, выбранную из группы, состоящей из SEQ ID NO: 31, 146 и 152-156. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 24, 131, 133-137, 143 и 144. В другом аспекте антитело к TGFβ2 содержит аминокислотную последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 24, 131, 133-137, 143 и 144. В одном аспекте антитело к TGFβ2 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 30, 145, 147-151, 157 и 158. В одном аспекте антитело к TGFβ2 содержит полную аминокислотную L-цепи, выбранную из группы, состоящей из SEQ ID NO: 30,145, 147-151, 157 и 158. В одном аспекте антитело к TGFβ2 содержит пару VH/VL, при этом пара VH/VL содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 25/24 (6F12 кролика), SEQ ID NO: 132/131 (v1), SEQ ID NO: 132/133 (v1.1), SEQ ID NO: 132/134 (v1.2), SEQ ID NO: 132/135 (v1.3), SEQ ID NO: 132/136 (v1.4), SEQ ID NO: 132/137 (v1.5), SEQ ID NO: 138/131 (v1.6), SEQ ID NO: 139/131 (v1.7), SEQ ID NO: 140/131 (v1.8), SEQ ID NO: 141/131 (v1.9), SEQ ID NO: 142/131 (v2), SEQ ID NO: 132/143 (v3) и SEQ ID NO: 142/144 (v4). В одном аспекте антитело к TGFβ 32 содержит полную пару H/L-цепей, при этом полная пара H/L-цепей содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 31/30 (6F12 кролика), SEQ ID NO: 146/145 (v1), SEQ ID NO: 146/147 (v1.1), SEQ ID NO: 146/148 (v1.2), SEQ ID NO: 146/149 (v1.3), SEQ ID NO: 146/150 (v1.4), SEQ ID NO: 146/151 (v1.5), SEQ ID NO: 152/145 (v1.6), SEQ ID NO: 153/145 (v1.7), SEQ ID NO: 154/145 (v1.8), SEQ ID NO: 155/145 (v1.9), SEQ ID NO: 156/145 (v2), SEQ ID NO: 146/157 (v3) и SEQ ID NO: 156/158 (v4). В одном аспекте антитело к TGFβ2 содержит VL, содержащую аминокислотную последовательность SEQ ID NO: 24, содержащую одну или несколько мутаций каркасной области, выбранных из группы, состоящей 43S или 43А, 66G, 69Т, 71F и 87Y. В одном аспекте VL антитело к TGFβ2 содержит набор мутаций каркасной области, выбранный из группы, состоящей из: (i) 43S в FR2 и 66Е, 69Р, 71Y и 87F в FR3 (h6F12.v1 и h6F12.v2); (ii) 43S в FR2 и 58V, 66Е, 69Р, 71Y и 87F в FR3 (h6F12.v3 и h6F12.v4); (iii) 43А в FR2 (h6F12.v1. 1); (iv) 66G в FR3 (h6F12.v1.2); (v) 69T в FR3 (h6F12.v1.3); (vi) 71F в FR3 (h6F12.v1.4); и (vii) 87Y в FR3 (h6F12.v1.5), где мутации представлены по отношению к VL, содержащей аминокислотную последовательность SEQ ID NO: 24. В одном аспекте антитело к TGFβ2 содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 25, содержащую одну или несколько мутаций каркасной области, выбранных из группы, состоящей из 37V или 371, 48М или 48L, 49G или 49А, 67L, 71К и 78V, и 105Р или 105R. В одном аспекте VH антитело к TGFβ2 содержит набор мутаций каркасной области, выбранный из группы, состоящей из: (i) 37V, 48М и 49G в FR2 и 105Р в FR4 (h6F12.v1 и h6F12.v3); (ii) 37V и 48М в FR2, 67L, 71К и 78V в FR3 и 105Р в FR4 (h6F12.v2 и h6F12.v4); (iii) 371 в FR2 (h6F12.v1.6); (iv) 48L в FR2 (h6F12.v1.7); (v) 49A в FR2 (h6F12.v1.8); (vi) 105R в FR4 (h6F12.v1.9); (vii) 37V, 48M и 49G в FR2 и 105P в FR4 (h6F12.v1 и h6F12.v3); (viii) 37V и 48M в FR2, 67L, 71K и 78V в FR3 и 105Р в FR4 (6F12.v2 и h6F12.v4); (ix) 371 в FR2 (h6F12.v1.6); (х) 48L в FR2 (h6F12.v1.7); (xi) 49А в FR2 (h6F12.v1.8); и (xii) 105R в FR4 (h6F12.v1.9), где мутации представлены по отношению к VH, содержащей аминокислотную последовательность SEQ ID NO: 25.
В определенных вариантах осуществления любого из вышеуказанных аспектов антитело к TGFβ3 и/или антитело к TGFβ2/3 специфически связываются с TGFβ3 человека. В некоторых вариантах осуществления любого из вышеуказанных аспектов антитело к TGFβ3 специфически связывается как с незрелой, так и со зрелой формами TGFβ3. В некоторых вариантах осуществления любого из вышеуказанных аспектов антитело к TGFβ2/3 и/или антитело к TGFβ2 специфически связываются с TGFβ2 человека. В некоторых вариантах осуществления любого из вышеуказанных аспектов антитело представляет собой моноклональное антитело. В некоторых вариантах осуществления любого из вышеуказанных аспектов антитело представляет собой человеческое, гуманизированное или химерное антитело. В некоторых вариантах осуществления любого из антител антитело представляет собой фрагмент антитела. В некоторых вариантах осуществления любого из вышеуказанных аспектов антитело содержит Fc-область человека, которая относится к изотипу IgG1 или IgG4. В некоторых вариантах осуществления любого из вышеуказанных аспектов антитело содержит Fc-область человека, которая относится к изотипу IgG1. В некоторых вариантах осуществления любого из вышеуказанных аспектов Fc-область антитела модифицирована с удалением эффекторной функции. В некоторых аспектах Fc-область содержит модификацию, которая приводит к удалению сайта гликозилирования в положении аминокислотного остатка N297 (EU-нумерация согласно Kabat). В некоторых аспектах модификация представляет собой мутацию, выбранную из N297G или N297A. В некоторых аспектах модификация представляет собой мутацию N297G. В некоторых вариантах осуществления любого из вышеуказанных аспектов антитело имеет Cmax, составляющую приблизительно 230-260 мкг/мл, и/или период полужизни (t½), составляющий от приблизительно 15 до 16 дней.
Также предусмотрены выделенные нуклеиновые кислоты, кодирующие антитело в соответствии с любым из вышеуказанных аспектов и вариантов осуществления, и клетки-хозяева, содержащие нуклеиновые кислоты. В некоторых аспектах представлен способ получения антитела. Способ может включать культивирование клетки-хозяина, представленной в данном документе таким образом, чтобы получить антитело. В некоторых аспектах способ дополнительно включает выделение антитела из клетки-хозяина. Также предусмотрено антитело, полученное с помощью способа получения антитела.
В другом аспекте предусмотрен иммуноконъюгат, содержащий антитело, такое как любое из антител, описанных выше, и цитотоксическое средство. В некоторых вариантах осуществления антитело представляет собой антитело к TGFβ3, содержащее: (a1) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 5 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а2) (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 34 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а3) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (а4) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 159 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (б) пару VH/VL, VH пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 57, и VL пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 56; или (в) полную пару H/L-цепей, где Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79, а L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78. В некоторых вариантах осуществления антитело представляет собой антитело к TGFβ2, где антитело содержит: (a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 18; и (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21. В некоторых вариантах осуществления антитело представляет собой антитело к TGFβ2/3, содержащее (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 15.
В другом аспекте представлен фармацевтический состав или иммуноконъюгат, содержащие антитело по любому из вышеуказанных аспектов и вариантов осуществления и фармацевтически приемлемый носитель. В некоторых вариантах осуществления антитело представляет собой антитело к TGFβ3, содержащее: (a1) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 5 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (a2) (i) CDR тяжелой цепи, содержащие CDR-H1, -H2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 34 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а3) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (а4) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 159 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (б) пару VH/VL, VH пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 57, и VL пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 56; или (в) полную пару H/L-цепей, где Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79, а L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78. В некоторых вариантах осуществления антитело представляет собой антитело к TGFβ 32, где антитело содержит: (a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 18; и (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21. В некоторых вариантах осуществления фармацевтического состава или конъюгата антитело представляет собой антитело к TGFβ2/3, содержащее (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-Н3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 15.
В некоторых аспектах фармацевтический состав дополнительно содержит дополнительное терапевтическое средство. В некоторых аспектах дополнительное терапевтическое средство выбрано из группы, состоящей из пирфенидона, нинтеданиба, микофенилата мофетила, ингибитора IL-6 (например, тоцилизумаба), антитела к CTFG (например, FG-3019), ингибитора аутотаксина, ингибитора JAK, ингибитора IL-11 и РТХ2.
Также представлено антитело в соответствии с любым из вышеуказанных аспектов и вариантов осуществления для применения в качестве лекарственного препарата. В некоторых вариантах осуществления антитело для применения в качестве лекарственного препарата представляет собой антитело к TGFβ3, содержащее: (al) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 5 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а2) (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 34 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а3) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (а4) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 159 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 21. В некоторых вариантах осуществления антитело для применения в качестве лекарственного препарата представляет собой антитело к TGFβ2/3, содержащее (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 15.
Также представлено антитело в соответствии с любым из вышеуказанных аспектов и вариантов осуществления для применения в лечении нарушения, связанного с TGFβ. В некоторых вариантах осуществления антитело для применения в лечении нарушения, связанного с TGFβ, представляет собой антитело к TGFβ3, содержащее: (a1) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 5 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а2) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 34 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а3) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (а4) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 159 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (б) пару VH/VL, VH пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 57, и VL пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 56; или (в) полную пару H/L-цепей, где Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79, а L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78. В некоторых вариантах осуществления антитело для применения в лечении нарушения, связанного с TGFβ, представляет собой антитело к TGFβ2, где антитело содержит: (a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 18; и (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21. В некоторых вариантах осуществления антитело для применения в лечении нарушения, связанного с TGFβ, представляет собой антитело к TGFβ2/3, содержащее (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 15.
В одном аспекте представлено антитело к TGFβ3 в соответствии с любым из вышеуказанных аспектов и вариантов осуществления, и представлено антитело к TGFD2 в соответствии с любым из вышеуказанных аспектов и вариантов осуществления для применения в комбинации для лечения нарушения, связанного с TGFβ. В одном варианте осуществления антитело к TGFβ3 для применения в такой комбинации содержит: (a1) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 5 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а2) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 34 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а3) (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (а4) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 159 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (б) пару VH/VL, VH пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 57, и VL пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 56; или (в) полную пару H/L-цепей, где Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79, а L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78. В другом варианте осуществления антитело к TGFβ 32 содержит: (a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 18; и (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21. В одном аспекте антитело к TGFβ2 селективно нейтрализует TGFβ2. В некоторых аспектах антитело к TGFβ2: (а) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ; (б) имеет сниженную токсичность у грызунов по сравнению с антителом 1D11 ко всем видам TGFβ; (в) имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; и/или (г) имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом. В одном аспекте антитело к TGFP2 содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 132 и 138-142. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 25, 132 и 138-142.
Также представлено антитело в соответствии с любым из вышеуказанных аспектов и вариантов осуществления для применения в производстве лекарственного препарата для лечения нарушения, связанного с TGFβ, для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или для ингибирования синтеза нового коллагена. В одном варианте осуществления антитело для такого лекарственного препарата представляет собой антитело к TGFβ3, содержащее: (a1) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 5 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а2) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 34 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а3) (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (а4) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 159 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (б) пару VH/VL, VH пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 57, и VL пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 56; или (в) полную пару H/L-цепей, где Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79, а L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78. В другом варианте осуществления антитело для такого лекарственного препарата представляет собой антитело к TGFβ2, содержащее: (a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 18; и (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21. В одном аспекте антитело к TGFβ2 селективно нейтрализует TGFβ2. В некоторых аспектах антитело к TGFβ2: (а) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ; (б) имеет сниженную токсичность у грызунов по сравнению с антителом 1D11 ко всем видам TGFβ; (в) имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; и/или (г) имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 132 и 138-142. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 25, 132 и 138-142. В другом аспекте антитело представляет собой антитело к TGFβ2/3, содержащее (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 15.
В некоторых вариантах осуществления вышеуказанных вариантов применения и лекарственных препаратов для лечения нарушений, связанных с TGFβ, нарушение, связанное с TGFβ, представляет собой фиброз. В некоторых вариантах осуществления фиброз представляет собой фиброзное состояние легких, печени, сердца, почек, поджелудочной железы, глаз и/или кожи. В некоторых аспектах фиброз представляет собой фиброз легких, выбранный из группы, состоящей из идиопатического фиброза легких (IPF), идиопатического фиброза верхних долей легких (болезнь Амитани), семейного фиброза легких, фиброза легких (например, фиброза легких, вторичного по отношению к системным воспалительным заболеваниям, таким как ревматоидный артрит, склеродермия, волчанка, криптогенный фиброзирующий альвеолит, хроническая обструктивная болезнь легких (COPD) или хроническая астма), муковисцидоз, неспецифическая интерстициальная пневмония (NSIP), криптогенная организующаяся пневмония (СОР), прогрессирующий массивный фиброз, склеродермия/системный склероз (SSc, включая ограниченную кожную (lcSSc) и диффузную кожную (dcSSc) формы и связанное с SSc интерстициальное заболевание легких (SSc-ILD)), облитерирующий бронхиолит с организующейся пневмонией, ILD, связанное с заболеванием соединительной ткани (CT-ILD), гиперчувствительный пневмонит, легочная гипертензия, туберкулез, силикоз, асбестоз легких, острое повреждение легких и острый респираторный дистресс (ARD, включая вызванный бактериальной пневмонией, вызванный повреждением и вызванный вирусной пневмонией, вызванный искусственной вентиляцией легких, вызванный нелегочным сепсисом). В некоторых аспектах фиброз представляет собой фиброзное состояние печени, выбранное из группы, состоящей из цирроза печени, врожденного фиброза печени, ожирения, жировой печени, алкогольного фиброза печени, неалкогольного стеатогепатита (NASH), повреждения желчных протоков, первичного билиарного цирроза (РВС), первичного склерозирующего холангита (PSC), инфекционного или вирусного фиброза печени (например, хронический инфекций, вызванных вирусом гепатита В и С), муковисцидоза, аутоиммунного гепатита, некротизирующего гепатита, первичного склерозирующего холангита, гемохроматоза, нарушений системы желчных протоков и печеночной дисфункции, обусловленной инфекциями. В некоторых аспектах фиброз представляет собой фиброзное состояние сердца и/или перикарда, выбранное из группы, состоящей из эндомиокардиального фиброза, васкулопатии сердечного аллотрансплантата (CAV), инфаркта миокарда, фиброза предсердий, застойной сердечной недостаточности, артериосклероза, атеросклероза, стеноза сосудов, миокардита, застойной кардиомиопатии, коронарных инфарктов, варикозного расширения вен, стеноза коронарных артерий и других постишемических состояний, и идиопатического ретроперитонеального фиброза. В некоторых аспектах фиброз представляет собой фиброзное состояние почек, выбранное из группы, состоящей из гломерулонефрита (включая мембранопролиферативные, диффузно-пролиферативные, быстро прогрессирующие или склерозирующие, постинфекционные и хронические формы), диабетического гломерулосклероза, фокально-сегментарного гломерулосклероза, IgA-нефропатии, диабетической нефропатии, ишемической нефропатии, тубулоинтерстициального фиброза почек, связанной с ВИЧ нефропатии, мембранной нефропатии, гломерулонефрита, вторичного по отношению к системным воспалительным заболеваниям, таким как волчанка, склеродермия и диабетический гломерулонефрит, идиопатического мембранопролиферативного гломерулонефрита, мезангиального пролиферативного гломерулонефрита, серповидного гломерулонефрита, амилоидоза (который среди других тканей поражает почки), аутоиммунного нефрита, почечного тубулоинстерстиального фиброза, почечного атеросклероза, синдрома Альпорта, нефротического синдрома, хронической почечной недостаточности, хронического заболевания почек, перигломерулярного фиброза/атубулярных клубочков, синдрома сочетанной апикальной эмфиземы и базального фиброза (синдрома эмфиземы/фиброза), гломерулярной гипертензии, нефрогенной фиброзирующей дерматопатии, поликистозной болезни почек, болезни Фабри и почечной гипертензии. В некоторых аспектах фиброз представляет собой фиброзное состояние поджелудочной железы, выбранное из группы, состоящей из стромального ремоделирующего панкреатита и стромального фиброза. В некоторых аспектах фиброз представляет собой фиброзное состояние желудочно-кишечного тракта, выбранное из группы, состоящей из болезни Крона, язвенного колита, коллагенозного колита, колоректального фиброза, атрофии ворсинок, гиперплазии крипт, образования полипов, заживления язвы желудка и микроскопического колита. В некоторых аспектах фиброз представляет собой фиброзное состояние глаз, выбранное из группы, состоящей из окулярного фиброза, фиброза глаз, пролиферативной витреоретинопатии, витреоретинопатии любой этиологии, фиброза, связанного с дисфункцией сетчатки, фиброза, связанного с влажной или сухой макулодистрофией, рубцевания роговицы и конъюнктивы, фиброза эндотелия роговицы, передней субкапсулярной катаракты и помутнения задней капсулы хрусталика, фиброзных заболеваний переднего сегмента глаза, фиброза стромы роговицы (например, связанного с помутнением роговицы), фиброза трабекулярной сети (например, связанного с глаукомой), фиброзных заболеваний заднего сегмента глаза, фиброваскулярного рубцевания (например, в сетчатке или хориоидальной сосудистой сети глаз), фиброза сетчатки, эпиретинального фиброза, глиоза сетчатки, субретинального фиброза (например, связанного с возрастной макулодистрофией), тракционной отслойки сетчатки в сочетании с сокращением ткани при диабетической ретинопатии, врожденного фиброза глазницы, фиброза слезных желез, субэпителиального фиброза роговицы и офтальмопатии Грейвса. В некоторых аспектах фиброз выбран из фиброза, возникающего в результате повреждения/фиброза спинного мозга или фиброза центральной нервной системы, таких как фиброз после инсульта, мышечная дистрофия Дюшенна, фиброз, связанный с нейродегенеративным нарушением, таким как болезнь Альцгеймера или рассеянный склероз, рестеноз сосудов, фиброз матки, эндометриоз, фиброиды яичников, болезнь Пейрони, синдром поликистозных яичников, апикальный фиброз легких, связанный с заболеванием анкилозирующим спондилитом, рубцевание и фиброз, возникающие при микробных инфекциях (например, бактериальных, вирусных, паразитарных, грибковых). В конкретном варианте осуществления фиброз представляет собой SSc. В конкретном варианте осуществления фиброз представляет собой IPF. В конкретном варианте осуществления фиброз представляет собой хроническую обструктивную болезнь легких (COPD). В конкретном варианте осуществления фиброз представляет собой прогрессирующее фиброзирующее интерстициальное заболевание легких (PF-ILD). В конкретном варианте осуществления PF-ILD представляет собой заболевание или состояние, выбранное из группы, состоящей из неспецифической интерстициальной пневмонии (NSIP), криптогенной организующейся пневмонии (СОР), прогрессирующего массивного фиброза, осложнения пневмокониоза шахтеров, склеродермии/системной склеродермии, облитерирующего бронхиолита с организующейся пневмонией; ILD, связанное с заболеванием соединительной ткани (CT-ILD), и гиперчувствительный пневмонит.В конкретном варианте осуществления фиброз представляет собой цирроз печени или хронический фиброз печени. В конкретном варианте осуществления фиброз представляет собой фиброз желудочно-кишечного тракта. В конкретном варианте осуществления фиброз представляет собой фиброзное состояние глаз, фиброз, возникающий в результате повреждения спинного мозга, фиброз или фиброз центральной нервной системы, или фиброз, связанный с нейродегенеративным нарушением.
В другом аспекте представлен способ лечения субъекта, у которого имеется нарушение, связанное с TGFβ. В некоторых вариантах осуществления способ включает введение эффективного количества антитела или фармацевтического состава в соответствии с любым из вышеуказанных аспектов и вариантов осуществления субъекту, нуждающемуся в этом. В другом аспекте представлен способ ингибирования TGFBR-зависимой передачи сигналов SMAD, ингибирования сборки сигнальных комплексов TGFβ-TGFBR, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченного эндоглином, и/или ингибирования синтеза нового коллагена у субъекта. В некоторых аспектах способ включает введение субъекту, нуждающемуся в этом, эффективного количества антитела в соответствии с любым из вышеуказанных аспектов и вариантов осуществления для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или для ингибирования синтеза нового коллагена у субъекта. В некоторых аспектах способ включает введение субъекту дополнительного терапевтического средства. В некоторых аспектах дополнительное терапевтическое средство выбрано из группы, состоящей из пирфенидона, нинтеданиба, микофенилата мофетила, ингибитора IL-6 (например, тоцилизумаба, сарилумаба), антитела к CTFG (например, FG-3019), ингибитора аутотаксина и РТХ2. В одном варианте осуществления таких способов антитело представляет собой антитело к TGFβ3, содержащее: (a1) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 5 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а2) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 34 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а3) (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (а4) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 159 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (б) пару VH/VL, VH пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 57, и VL пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 56; или (в) полную пару H/L-цепей, где Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79, а L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78. В другом варианте осуществления таких способов антитело представляет собой антитело к TGFβ2, содержащее: (a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 18; и (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21. В одном аспекте антитело к TGFβ2 селективно нейтрализует TGFβ2. В некоторых аспектах антитело к TGFβ2: (а) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ; (б) имеет сниженную токсичность у грызунов по сравнению с антителом 1D11 ко всем видам TGFβ; (в) имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; и/или (г) имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 132 и 138-142. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 25, 132 и 138-142. В другом варианте осуществления таких способов антитело представляет собой антитело к TGFβ2/3, содержащее (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-Н2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 15.
В дополнительном аспекте вышеуказанных способов способ может включать введение субъекту эффективного количества антитела к TGFβ3 и эффективного количества антитела к TGFβ2. В одном варианте осуществления антитело к TGFβ3 для применения в такой комбинации содержит: (a1) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 5 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (a2) (i) CDR тяжелой цепи, содержащие CDR-H1, -H2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 34 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а3) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (а4) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 159 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (б) пару VH/VL, VH пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 57, и VL пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 56; или (в) полную пару H/L-цепей, где Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79, а L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78. В другом варианте осуществления антитело к TGFβ2 содержит: (a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 18; и (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21. В одном аспекте антитело к TGFβ2 селективно нейтрализует TGFβ2. В некоторых аспектах антитело к TGFβ2: (а) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ; (б) имеет сниженную токсичность у грызунов по сравнению с антителом 1D11 ко всем видам TGFβ; (в) имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; и/или (г) имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 132 и 138-142. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 25, 132 и 138-142.
В дополнительных вариантах осуществления вышеуказанных способов лечения нарушений, связанных с TGFβ, нарушение, связанное с TGFβ, может представлять собой фиброз. В некоторых вариантах осуществления фиброз представляет собой фиброзное состояние легких, печени, сердца, почек, поджелудочной железы, глаз и/или кожи. В некоторых аспектах фиброз представляет собой фиброз легких, выбранный из группы, состоящей из идиопатического фиброза легких (IPF), идиопатического фиброза верхних долей легких (болезнь Амитани), семейного фиброза легких, фиброза легких (например, фиброза легких, вторичного по отношению к системным воспалительным заболеваниям, таким как ревматоидный артрит, склеродермия, волчанка, криптогенный фиброзирующий альвеолит, хроническая обструктивная болезнь легких (COPD) или хроническая астма), муковисцидоз, неспецифическая интерстициальная пневмония (NSIP), криптогенная организующаяся пневмония (СОР), прогрессирующий массивный фиброз, склеродермия/системный склероз (SSc, включая ограниченную кожную (lcSSc) и диффузную кожную (dcSSc) формы и связанное с SSc интерстициальное заболевание легких (SSc-ILD)), облитерирующий бронхиолит с организующейся пневмонией, ILD, связанное с заболеванием соединительной ткани (CT-ILD), гиперчувствительный пневмонит, легочная гипертензия, туберкулез, силикоз, асбестоз легких, острое повреждение легких и острый респираторный дистресс (ARD, включая вызванный бактериальной пневмонией, вызванный повреждением и вызванный вирусной пневмонией, вызванный искусственной вентиляцией легких, вызванный нелегочным сепсисом). В некоторых аспектах фиброз представляет собой фиброзное состояние печени, выбранное из группы, состоящей из цирроза печени, врожденного фиброза печени, ожирения, жировой печени, алкогольного фиброза печени, неалкогольного стеатогепатита (NASH), повреждения желчных протоков, первичного билиарного цирроза (РВС), первичного склерозирующего холангита (PSC), инфекционного или вирусного фиброза печени (например, хронический инфекций, вызванных вирусом гепатита В и С), муковисцидоза, аутоиммунного гепатита, некротизирующего гепатита, первичного склерозирующего холангита, гемохроматоза, нарушений системы желчных протоков и печеночной дисфункции, обусловленной инфекциями. В некоторых аспектах фиброз представляет собой фиброзное состояние сердца и/или перикарда, выбранное из группы, состоящей из эндомиокардиального фиброза, васкулопатии сердечного аллотрансплантата (CAV), инфаркта миокарда, фиброза предсердий, застойной сердечной недостаточности, артериосклероза, атеросклероза, стеноза сосудов, миокардита, застойной кардиомиопатии, коронарных инфарктов, варикозного расширения вен, стеноза коронарных артерий и других постишемических состояний, и идиопатического ретроперитонеального фиброза. В некоторых аспектах фиброз представляет собой фиброзное состояние почек, выбранное из группы, состоящей из гломерулонефрита (включая мембранопролиферативные, диффузно-пролиферативные, быстро прогрессирующие или склерозирующие, постинфекционные и хронические формы), диабетического гломерулосклероза, фокально-сегментарного гломерулосклероза, IgA-нефропатии, диабетической нефропатии, ишемической нефропатии, тубулоинтерстициального фиброза почек, связанной с ВИЧ нефропатии, мембранной нефропатии, гломерулонефрита, вторичного по отношению к системным воспалительным заболеваниям, таким как волчанка, склеродермия и диабетический гломерулонефрит, идиопатического мембранопролиферативного гломерулонефрита, мезангиального пролиферативного гломерулонефрита, серповидного гломерулонефрита, амилоидоза (который среди других тканей поражает почки), аутоиммунного нефрита, почечного тубулоинстерстиального фиброза, почечного атеросклероза, синдрома Альпорта, нефротического синдрома, хронической почечной недостаточности, хронического заболевания почек, перигломерулярного фиброза/атубулярных клубочков, синдрома сочетанной апикальной эмфиземы и базального фиброза (синдрома эмфиземы/фиброза), гломерулярной гипертензии, нефрогенной фиброзирующей дерматопатии, поликистозной болезни почек, болезни Фабри и почечной гипертензии. В некоторых аспектах фиброз представляет собой фиброзное состояние поджелудочной железы, выбранное из группы, состоящей из стромального ремоделирующего панкреатита и стромального фиброза. В некоторых аспектах фиброз представляет собой фиброзное состояние желудочно-кишечного тракта, выбранное из группы, состоящей из болезни Крона, язвенного колита, коллагенозного колита, колоректального фиброза, атрофии ворсинок, гиперплазии крипт, образования полипов, заживления язвы желудка и микроскопического колита. В некоторых аспектах фиброз представляет собой фиброзное состояние глаз, выбранное из группы, состоящей из окулярного фиброза, фиброза глаз, пролиферативной витреоретинопатии, витреоретинопатии любой этиологии, фиброза, связанного с дисфункцией сетчатки, фиброза, связанного с влажной или сухой макулодистрофией, рубцевания роговицы и конъюнктивы, фиброза эндотелия роговицы, передней субкапсулярной катаракты и помутнения задней капсулы хрусталика, фиброзных заболеваний переднего сегмента глаза, фиброза стромы роговицы (например, связанного с помутнением роговицы), фиброза трабекулярной сети (например, связанного с глаукомой), фиброзных заболеваний заднего сегмента глаза, фиброваскулярного рубцевания (например, в сетчатке или хориоидальной сосудистой сети глаз), фиброза сетчатки, эпиретинального фиброза, глиоза сетчатки, субретинального фиброза (например, связанного с возрастной макулодистрофией), тракционной отслойки сетчатки в сочетании с сокращением ткани при диабетической ретинопатии, врожденного фиброза глазницы, фиброза слезных желез, субэпителиального фиброза роговицы и офтальмопатии Грейвса. В некоторых аспектах фиброз выбран из фиброза, возникающего в результате повреждения/фиброза спинного мозга или фиброза центральной нервной системы, таких как фиброз после инсульта, мышечная дистрофия Дюшенна, фиброз, связанный с нейродегенеративным нарушением, таким как болезнь Альцгеймера или рассеянный склероз, рестеноз сосудов, фиброз матки, эндометриоз, фиброиды яичников, болезнь Пейрони, синдром поликистозных яичников, апикальный фиброз легких, связанный с заболеванием анкилозирующим спондилитом, рубцевание и фиброз, возникающие при микробных инфекциях (например, бактериальных, вирусных, паразитарных, грибковых). В конкретном варианте осуществления фиброз представляет собой SSc. В конкретном варианте осуществления фиброз представляет собой IPF. В конкретном варианте осуществления фиброз представляет собой хроническую обструктивную болезнь легких (COPD). В конкретном варианте осуществления фиброз представляет собой прогрессирующее фиброзирующее интерстициальное заболевание легких (PF-ILD). В конкретном варианте осуществления PF-ILD представляет собой заболевание или состояние, выбранное из группы, состоящей из неспецифической интерстициальной пневмонии (NSIP), криптогенной организующейся пневмонии (СОР), прогрессирующего массивного фиброза, осложнения пневмокониоза шахтеров, склеродермии/системной склеродермии, облитерирующего бронхиолита с организующейся пневмонией; ILD, связанное с заболеванием соединительной ткани (CT-ILD), и гиперчувствительный пневмонит.В конкретном варианте осуществления фиброз представляет собой цирроз печени или хронический фиброз печени. В конкретном варианте осуществления фиброз представляет собой фиброз желудочно-кишечного тракта, например, фиброз кишечника, необязательно выбранный из группы, состоящей из фиброза, связанного с болезнью Крона, язвенного колита, коллагенозного колита, колоректального фиброза, атрофии ворсинок, гиперплазии крипт, образования полипов, заживления язвы желудка и микроскопического колита. В конкретном варианте осуществления фиброз представляет собой фиброзное состояние глаз, фиброз, возникающий в результате повреждения спинного мозга, фиброз или фиброз центральной нервной системы, или фиброз, связанный с нейродегенеративным нарушением.
В другом аспекте представлен способ диагностики SSc у субъекта. В некоторых вариантах осуществления способ включает определение уровней экспрессии генов в наборе сигнатур 18 генов, состоящем из PRSS23, PXDN, COL8A1, COL6A3, SERPINE2, TNC, СОМР, THBS1, COL11A1, COL1A1, COL5A2, COL1A2, COL4A1, COL4A2, SFRP4, ALPK2, COL5A1 и TAGLN, и установления диагноза SSc у субъекта, если установлено, что уровни генов повышены по сравнению с уровнями генов в контрольном наборе от здорового субъекта или сигнатуре эталонных генов. В некоторых аспектах уровень гена повышается, если повышение экспрессии по сравнению с контрольным набором от здорового субъекта или сигнатурой эталонных генов является статистически значимым, необязательно по меньшей мере в два раза более высоким, или по меньшей мере в три раза более высоким, или по меньшей мере в четыре раза более высоким по сравнению с контрольным набором от здорового субъекта или сигнатурой эталонных генов. В некоторых аспектах уровень экспрессии гена определяют с применением кПЦР или микроматричного анализа или РНК-секвенирования.
В другом аспекте представлен способ мониторинга ответа на лечение субъекта антителом к TGFβ2 и/или антителом к TGFβ3. В некоторых аспектах способ включает определение уровня экспрессии одного или нескольких индуцируемых TGFβ генов, выбранных из группы, состоящей из serpinel, collal, col1a2 и col3a1 в образце от субъекта, где субъект получал одно или несколько введений антитела к TGFβ2 и/или антитела к TGFβ3. В некоторых аспектах у субъекта определяют наличие ответа на лечение антителом к TGFβ2 и/или антителом к TGFβ3, если уровень экспрессии одного или более индуцируемых TGFβ генов, значительно снижен по сравнению с уровнями одного или более индуцируемых TGFβ генов до лечения, при этом необязательно способ дополнительно включает осуществление одного или более дополнительных введений антитела к TGFβ2 и/или антитела к TGFβ3, если установлено, что уровень экспрессии одного или более индуцируемых TGFβ генов, значимо снижен. В некоторых аспектах субъекту вводят антитело к TGFβ2 в качестве монотерапии. В некоторых аспектах субъекту вводят антитело к TGFβ3 в качестве монотерапии. В некоторых аспектах уровень экспрессии одного или более генов, индуцируемых TGFβ, определяют с помощью кПЦР или микроматричного анализа. В одном варианте осуществления такого способа антитело представляет собой антитело к TGFβ3, содержащее: (a1) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 5 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а2) (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 34 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а3) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (а4) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 159 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (б) пару VH/VL, VH пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 57, и VL пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 56; или (в) полную пару H/L-цепей, где Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79, а L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78. В другом варианте осуществления такого способа антитело представляет собой антитело к TGF(32, содержащее: (a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 18; и (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21. В одном аспекте антитело к TGFβ2 селективно нейтрализует TGFβ2. В некоторых аспектах антитело к TGFβ2: (а) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ; (б) имеет сниженную токсичность у грызунов по сравнению с антителом 1D11 ко всем видам TGFβ; (в) имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; и/или (г) имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 132 и 138-142. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 25, 132 и 138-142.
В любом из вышеперечисленных аспектов и вариантов осуществления вариантов применения, лекарственных препаратов и способов лечения, диагностики и мониторинга ответа на лечение субъектом может быть человек, например, пациент-человек.
В другом аспекте представлен набор, содержащий антитело по любому из вышеуказанных аспектов и вариантов осуществления. В одном варианте осуществления такого набора антитело представляет собой антитело к TGFβ3, содержащее: (a1) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 5 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (a2) (i) CDR тяжелой цепи, содержащие CDR-H1, -H2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 34 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (а3) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (а4) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 159 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; или (б) пару VH/VL, VH пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 57, и VL пары VH/VL, содержащую аминокислотную последовательность SEQ ID NO: 56; или (в) полную пару H/L-цепей, где Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79, а L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78. В другом варианте осуществления такого набора антитело представляет собой антитело к TGFβ2, содержащее: (a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 18; и (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21. В одном аспекте антитело к TGFβ2 селективно нейтрализует TGFβ2. В некоторых аспектах антитело к TGFβ2: (а) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ; (б) имеет сниженную токсичность у грызунов по сравнению с антителом 1D11 ко всем видам TGFβ; (в) имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; и/или (г) имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 132 и 138-142. В одном аспекте антитело к TGFβ2 содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 25, 132 и 138-142. В другом варианте осуществления такого набора антитело представляет собой антитело к TGFβ2/3, содержащее (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 15.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 изображено выравнивание первичной аминокислотной последовательности TGFβ1, 2 и 3 человека, имеющей следующие номера доступа: huTGFβ1: ХР 011525544.1, huTGFβ2: NP_003229.1, huTGFβ3: ABQ59024.1, при этом домены TGFβ1 (α-листы и β-цепи, лассо латентности, застежка, интегрин-связывающий, расщепление фурином и цитокин), указанные в соответствии с Shi et al., Nature 474:343 (2011). Остатки пронумерованы от предполагаемого сайта начала транскрипции ATG (метионина); нумерация в соответствии с Shi et al. начинается с L30 в начале спирали α1. Стрелка черного цвета под последовательностями указывает на начало рецептор-связывающего домена. Мутации сдвига рамки считывания, преждевременной остановки и сайта сплайсинга, о которых сообщалось для TGFβ2 и TGFβ3, опущены для лучшего понимания. Последовательности, изображенные на фигуре, соответствуют SEQ ID NO: 1-3 соответственно.
На Фиг. 2А изображен фармакокинетический профиль антитела 6F12 muIgG2a, вводимого в виде однократной дозы, составляющей 1 или 10 мг/кг I.V., и 10 мг/кг I.P., у мышей C57BL6 (n=3 на временную точку).
На Фиг. 2Б изображен фармакокинетический профиль 16С10 muIgG2a, 15А7 muIgG2a, 18 В5 muIgG2a, 2А10 muIgG2a и несвязывающего контрольного антитела к gD huIgG1, вводимых в виде однократной дозы, составляющей 10 мг/кг I.V., у мышей C57BL6 (n=3 на временную точку).
На Фиг. 2В изображен фармакокинетический профиль 4А11 muIgG2a, вводимого в виде однократной дозы, составляющей 1 или 10 мг/кг I.V., и 10 мг/кг I.P., и антитела 6F12 muIgG2a, вводимого в виде однократной дозы, составляющей 10 мг/кг I.V., у мышей C57BL6 (n=3 на временную точку).
На Фиг. 3А представлен график, на котором изображены уровни экспрессии, определенные с помощью микроматричного анализа изоформ TGFβ в объемной ткани биоптата легких из контрольных легких (n=8) и легких, пораженных IPF (n=40). Данные показывают повышенную экспрессию TGFβ2 и TGFβ3, но не TGFβ1 при IPF. **Р<0,01; **** Р<0,0001 (непарные двусторонние t-критерии Стьюдента).
На Фиг. 3Б представлено изображение иммуногистохимического анализа (IHC) pSMAD2/3 ткани контрольных легких и ткани легких, пораженных IPF, на котором изображено повышенное окрашивание ядер pSMAD в очагах фибробластов ткани легких, пораженных IPF.
На Фиг. 4А представлен график t-SNE, на котором представлено РНК-секвенирование отдельных клеток легких, пораженных IPF (n=3, комбинированный), на котором изображено представление нескольких линий эпителиальных, мезенхимальных и гемопоэтических клеток, как указано. Экспрессия отдельных изоформ TGFβ показала широкую экспрессию TGFβ1 преимущественно в гемопоэтических и эндотелиальных клетках; экспрессию TGFβ2 преимущественно в эпителиальных клетках; и TGFβ3 преимущественно в мезенхимальных клетках. Экспрессия генов-мишеней TGFβ показала сильное перекрытие SERPINE1, COL1A1, POSTN и COMP с экспрессией TGFβ3 в фибробластах и миофибробластах. SERPINE1 также перекрывается с экспрессией TGFβ1 в эндотелиальных клетках и макрофагах и POSTN в эндотелиальных клетках.
На Фиг. 4Б представлено изображение двойного IHC для pSMAD3 и гибридизации in situ (ISH) для TGFβ3, на котором показана колокализация мРНК TGFβ3 и ядерного pSMAD3 в нескольких клетках в фибробластическом фокусе в ткани легких, пораженных IPF.
Фиг. 5А включает графики, на которых изображена экспрессия изоформ TGFβ, определенная с помощью микроматричного анализа биоптатов кожи, взятых у здоровых контролей (НС) и пациентов с системным склерозом (SSc) при исходном уровне, включенных в исследование FaSScinate.
На Фиг. 5Б изображена сигнатура зависимой от сигнала TGFβR экспрессии генов кожи, полученная посредством сравнения генов, индуцированных в фибробластах, обработанных in vitro рекомбинантным рецептор-связывающим доменом TGFβ1, и генов, экспрессия которых значительно активирована в биоптате кожи, пораженной SSc, по сравнению с контрольным биоптатом кожи. Гены, которые были значительно повышены при обоих состояниях, выбирали в качестве кандидата на сигнатуру TGFβ в коже, пораженной SSc.
На Фиг. 6А представлен график, на котором изображен главный компонент 1 сигнатуры, определенной на Фиг. 5Б, полученный в виде непрерывной переменной для величины экспрессии TGFβR-зависимого гена по всем генам сигнатуры в биоптатах кожи, пораженной SSc, и по сравнению с уровнями экспрессии изоформы TGFβ в этих биоптатах. У пациентов с SSc как «с высоким уровнем TGFβ», так и «с низким уровнем TGFβ» имели место более высокие уровни сигнатуры этого гена, чем у здоровых контролей, а сигнатура гена TGFβ биоптата кожи высоко коррелировала с экспрессией TGFβ3, но не с экспрессией TGFβ1 или TGFβ2.
На Фиг. 6Б представлена таблица, в которой изображены интеркорреляции между показателем сигнатуры TGFβ-индуцируемого гена кожи, как определено на Фиг. 4В, и уровнями экспрессии генов кожи изоформ TGFβ, POSTN и COMP; и интеркорреляции с уровнями белков POSTN и СОМР в сыворотке крови в исследовании FaSScinate.
На Фиг. 6В представлена обобщенная таблица корреляции уровней периостина и СОМР в сыворотке крови с модифицированной шкалой оценки кожи Роднана (MRSS), клиническим показателем системного фиброза кожи, используемым в исследовании FaSScinate.
Фиг. 7 включает графики, на которых показано распределение изменения MRSS от исходного уровня до 48 недель у пациентов в исследовании FaSScinate, стратифицированных по кластеру исходного уровня сигнатуры генов кожи TGFβ. РВО: плацебо; TCZ: лечение тоцилизумабом; кумулят.вероятн.: кумулятивная вероятность изменения MRSS.
На Фиг. 8А-Е изображена экспрессия, полученная на основе микроматрицы в указанные временные точки после i.t. инсталляции блеомицина, TGFβ1 (Фиг. 8А), TGFβ2 (Фиг. 8Б), TGFβ3 (Фиг. 8В), COL1A1 (Фиг. 8Г), Serpinel (Фиг. 8Д) и Fn1 (Фиг. 8Е). Экспрессия TGFβ2, TGFβ3, COL1A1, Serpine1 и Fn1 следовала сходной кинетике, при этом достижение пика имело место между днем 7 и 14 после введения блеомицина. *, Р<0,05; **, Р<0,01; ***, P<10+-3; ****, P<10+-4 на основе непарного двустороннего t-критерия Стьюдента. Данные представляют собой средние значения ± SEM.
На Фиг. 9А представлена схема in vivo экспериментов с I.T. введением блеомицина для оценки эффективности антител к TGFβ. Отдельные когорты животных умерщвляли в день 14 или день 24 для оценки экспрессии генов легких во время пиковой экспрессии TGFβ2/3 или коллагена легких ближе к концу «фиброзной» стадии соответственно. Дейтерированную питьевую воду давали животным с дня 9 по день 23 для оценки доли выработки нового коллагена в течение этого интервала (дейтерированный гидроксипролин).
На Фиг. 9Б представлен график, на котором изображены сниженные уровни доли дейтерированного к общему гидроксипролина легких в день 24 при профилактическом введении антител к TGFβ2 и/или TGFβ3 (10 мг/кг, TIW). Каждая точка представляет отдельное животное.
На Фиг. 9В представлен график, на котором изображены сниженные уровни экспрессии гена FN1 в легких в день 14 после i.t. инсталляции блеомицина с превентивным введением антител к TGFβ2 и/или TGFβ3 (10 мг/кг, TIW). **Р<0,01; *** Р<0,001; ****P<0,0001 (однофакторный дисперсионный анализ с критерием Даннета).
На Фиг. 9Г и Фиг. 9Д представлены графики, на которых изображено влияние совместного введения 6F12 и 2А10 (10 мг/кг, TIW) или 4А11 в дозе 10 мг/кг TIW, 10 мг/кг QW или 2,5 мг/кг QW на содержание общего (Фиг. 9Г) и дейтерированного (Фиг. 9Д) гидроксипролина в день 24 после i.t. введения блеомицина. Аналогичное снижение наблюдали для дозы 10 мг/кг 4А11 QW по сравнению с TIW, но сниженное влияние в дозе 2,5 мг/кг QW. **Р<0,01; *** Р<0,001; ****Р<0,0001 (однофакторный дисперсионный анализ с критерием Даннета).
На Фиг. 10 представлена схема, на которой изображены попытки представления различных гуманизированных вариантов mAb 2А10.
На Фиг. 11 представлена таблица, в которой изображена потеря экспрессии и стабильности для большинства вариантов h2A10v1 во время завершающей очистки каркасных областей и CDR.
Фиг. 12 включает выравнивания аминокислотных последовательностей последовательностей вариабельных областей легкой цепи (верхняя панель) и тяжелой цепи (нижняя панель) антитела 2А10 крысы и его гуманизированных вариантов v1-v4; аминокислотные последовательности, изображенные на фигуре, представляют собой SEQ ID NO: 22, 36, 36, 54 и 56 (верхняя панель, от верхней последовательности к нижней последовательности при выравнивании) и SEQ ID NO: 23, 37, 45, 55 и 57 (нижняя панель, от верхней последовательности к нижней последовательности при выравнивании). Нумерация остатков представлена в соответствии с Kabat; «*» обозначает зону Верньера. Области CDR в соответствии с Chothia или Kabat (как указано) представлены в прямоугольниках; мутированные остатки выделены.
Фиг. 13 включает выравнивание аминокислотной последовательности вариабельной области легкой цепи 2А10 крысы и гуманизированных вариантов h2A10 v2, v2-v2.9; изображенные последовательности соответствуют SEQ ID NO: 22, 36, 38, 39, 40, 41, 36, 36, 36, 36 и 36 (от верхней последовательности к нижней последовательности при выравнивании). Нумерация остатков представлена в соответствии с Kabat; «*» обозначает зону Верньера. Области CDR в соответствии с Chothia или Kabat (как указано) представлены в прямоугольниках; мутированные остатки выделены.
Фиг. 14 включает выравнивание аминокислотной последовательности вариабельной области тяжелой цепи 2А10 крысы и гуманизированных вариантов h2A10 v2, v2-v2.9; изображенные последовательности соответствуют SEQ ID NO: 23, 45, 45, 45, 45, 45 и 46-50 (от верхней последовательности к нижней последовательности при выравнивании). Нумерация остатков представлена в соответствии с Kabat; «*» обозначает зону Верньера. Области CDR в соответствии с Chothia или Kabat (как указано) представлены в прямоугольниках; мутированные остатки выделены.
Фиг. 15 включает выравнивания аминокислотных последовательностей последовательностей вариабельных областей легкой цепи (верхняя панель) и тяжелой цепи (нижняя панель) антитела 2А10 крысы и гуманизированных вариантов v2; аминокислотные последовательности, изображенные на фигуре, представляют собой SEQ ID NO: 22, 36, 36, 36 и 36 (верхняя панель, от верхней последовательности к нижней последовательности при выравнивании) и SEQ ID NO: 23, 45 и 51-53 (нижняя панель, от верхней последовательности к нижней последовательности при выравнивании). Нумерация остатков представлена в соответствии с Kabat; «*» обозначает зону Верньера. Области CDR в соответствии с Chothia или Kabat (как указано) представлены в прямоугольниках; мутированные остатки выделены.
Фиг. 16 включает выравнивания аминокислотных последовательностей последовательностей вариабельных областей легкой цепи (верхняя панель) и тяжелой цепи (нижняя панель) антитела 2А10 крысы и его гуманизированного варианта v4; аминокислотные последовательности, изображенные на фигуре, представляют собой SEQ ID NO: 22 и 56 (верхняя панель, от верхней последовательности к нижней последовательности при выравнивании) и SEQ ID NO: 23 и 57 (нижняя панель, от верхней последовательности к нижней последовательности при выравнивании). Нумерация остатков представлена в соответствии с Kabat; «*» обозначает зону Верньера. Области CDR в соответствии с Chothia или Kabat (как указано) представлены в прямоугольниках; мутированные остатки выделены.
На Фиг. 17 представлен линейный график, на котором изображена средняя (±SD) концентрация h2A10v3 и h2A10v4 в сыворотке крови после однократного I.V. введения дозы 10 мг/кг в ФК исследовании у яванских макаков (n=4 на группу).
Фиг. 18 включает выравнивание аминокислотной последовательности вариабельной области легкой цепи mAb кролика 4А11 и его гуманизированных вариантов, v1-v8; изображенные последовательности соответствуют SEQ ID NO: 26, 80, 82, 80, 82, 84, 85, 84 и 84, от верхней последовательности к нижней последовательности при выравнивании. Нумерация остатков представлена в соответствии с Kabat; «*» обозначает зону Верньера. Области CDR в соответствии с Chothia или Kabat (как указано) представлены в прямоугольниках; мутированные остатки выделены.
Фиг. 19 включает выравнивание аминокислотной последовательности вариабельной области легкой цепи 4А11 кролика и гуманизированных вариантов, v1-v8; изображенные последовательности соответствуют SEQ ID NO: 27, 81, 81, 83, 83, 81, 81, 83 и 83 (от верхней последовательности к нижней последовательности при выравнивании). Нумерация остатков представлена в соответствии с Kabat; «*» обозначает зону Верньера. Области CDR в соответствии с Chothia или Kabat (как указано) представлены в прямоугольниках; мутированные остатки выделены.
Фиг. 20 включает выравнивание аминокислотной последовательности вариабельных областей легкой цепи гуманизированных вариантов 4А11 v7-v7.19; изображенные последовательности соответствуют SEQ ID NO: 84, 84, 84, 84, 89-92, 101, 101, 101 и 101 (от верхней последовательности к нижней последовательности при выравнивании). Нумерация остатков представлена в соответствии с Kabat; «*» обозначает зону Верньера. Области CDR в соответствии с Chothia или Kabat (как указано) представлены в прямоугольниках; мутированные остатки выделены.
Фиг. 21 включает выравнивание аминокислотной последовательности вариабельной области тяжелой цепи гуманизированных вариантов 4А11 v7-v7.19; изображенные последовательности соответствуют SEQ ID NO: 83, 86, 87, 88, 83, 83, 83, 83, 93-100 и 102-105 (от верхней последовательности к нижней последовательности при выравнивании). Нумерация остатков представлена в соответствии с Kabat; «*» обозначает зону Верньера. Области CDR в соответствии с Chothia или Kabat (как указано) представлены в прямоугольниках; мутированные остатки выделены.
Фиг. 22 включает выравнивания аминокислотных последовательностей последовательностей вариабельных областей легкой цепи (верхняя панель) и тяжелой цепи (нижняя панель) антитела 6F12 кролика и гуманизированных вариантов v1-v4; аминокислотные последовательности, изображенные на фигуре, представляют собой SEQ ID NO: 24, 131, 131, 143 и 144 (верхняя панель, от верхней последовательности к нижней последовательности при выравнивании) и SEQ ID NO: 25, 132, 142, 132 и 142 (нижняя панель, от верхней последовательности к нижней последовательности при выравнивании). Нумерация остатков представлена в соответствии с Kabat; «*» обозначает зону Верньера. Области CDR в соответствии с Chothia или Kabat (как указано) представлены в прямоугольниках; мутированные остатки выделены.
Фиг. 23 включает выравнивание аминокислотной последовательности вариабельных областей легкой цепи гуманизированных вариантов 6F12 v1 v1-v1.9; изображенные последовательности соответствуют SEQ ID NO: 131, 133-137, 131, 131, 131 и 131 (от верхней последовательности к нижней последовательности при выравнивании). Нумерация остатков представлена в соответствии с Kabat; «*» обозначает зону Верньера. Области CDR в соответствии с Chothia или Kabat (как указано) представлены в прямоугольниках; мутированные остатки выделены.
Фиг. 24 включает выравнивание аминокислотной последовательности вариабельных областей тяжелой цепи гуманизированных вариантов 6F12 v1 v1-v1.9; изображенные последовательности соответствуют SEQ ID NO: 132, 132, 132, 132, 132, 132 и 138-141 (от верхней последовательности к нижней последовательности при выравнивании). Нумерация остатков представлена в соответствии с Kabat; «*» обозначает зону Верньера. Области CDR в соответствии с Chothia или Kabat (как указано) представлены в прямоугольниках; мутированные остатки выделены.
На Фиг. 25 представлено изображение кристаллической структуры 2А10 в комплексе с TGFβ3 человека.
На Фиг. 26 представлено изображение TGFβ3, связанного с 2А10, и сравнение с TGFβ1 с комплексом TGFBR1/TGFBR2. Это указывает на то, что 2А10 стерически блокирует рекрутинг TGFBR2, но, вероятно, не TGFBR1.
На Фиг. 27 представлено изображение фрезолимумаба («фрезо»), антитела ко всем видам TGFβ, по сравнению с 2А10. Фрезолимумаб блокирует связывание TGFβ3 как с TGFBR1, так и с TGFBR2, но 2А10 блокирует связывание только с TGFBR2 вследствие другого угла связывания.
На Фиг. 28 изображен эпитоп на TGFβ3, связанном 2А10. 2А10 связывает бета-шпильку на кончике бета-6-бета7 «пальца» TGFβ3 посредством R394. Полярные контакты также образованы между R325 и К331.
На Фиг. 29 изображен эпитоп на TGFβ3, связанном 2А10; остатки, участвующие в связывании TGFβ3 посредством 2А10 в пределах 5,0 Å, помечены.
На Фиг. 30 изображен паратоп антитела 2А10 (нумерация остатков согласно Kabat). R394 на TGFβ3 находится в контакте с D50 2А10.
На Фиг. 31 изображен эпитоп TGFβ3/2A10 и сравнение с TGFβ1. TGβF1 имеет три изменения остатков эпитопа по сравнению с TGFβ3. Замены Р387Т, L389V и T395K могут приводить к незначительному изменению конформации бета-шпильки и пальцев β6-β7 TGFβ2 и удалению некоторых полезных контактов с 2А10.
На Фиг. 32 изображен эпитоп TGFβ3/2A10 и сравнение с TGFβ2. TGFβ2 имеет четыре изменения остатков эпитопа по сравнению с TGFβ3. Основные причины специфичности TGFβ3 по сравнению с TGFβ2, вероятно, основаны на изменении R394K, которое приводит к удалению оптимизированного солевого мостика в эпитопе R394-D50, и V398I, добавляя потенциальное стерическое столкновение.
На Фиг. 33 изображено сравнение связывания TGFβ2 с 4A11v2 (левое изображение) и 4A11v7 (правое изображение). Комплекс антиген-антитело является очень похожим. Тяжелые цепи сближаются при связывании, но угол колена связывания второго Fab отличается.
На Фиг. 34 изображено сравнение 4A11v2/TGFβ2 с комплексом TGFβ3/рецептор. 4A11v2 не конкурирует напрямую с сайтами связывания TGFBR1/2; скорее, 4A11v2 слегка изменяет структуру TGFβ2, приводя к совместному «сжатию» пальцев. Это может быть результатом стерического столкновения между Fab 4A11v2.
На Фиг. 35 изображен эпитоп TGFβ2, связываемый 4А11. Глутаминовая кислота в положении Е373 аминокислотной последовательности TGFβ2 (нумерация TGFβ2) находится в центре поверхности контакта. TGFβ3 также имеет глутаминовую кислоту в этом положении, подобно TGFβ2, в то время как TGFβ1 имеет глицин.
Фиг. 36 включает изображения, на которых изображены конформационные изменения в TGFβ2, индуцированные 4A11v2 (правое изображение) и TGFβ2 из комплекса фрезолимумаба («фрезо») (левое изображение). Конформационные изменения в структуре TGFβ2 могут приводить к изменению способности сигнальных рецепторов связывать TGFβ2. 4А11 может также блокировать доступ мембраносвязанных рецепторов посредством связывания проксимальной стороны мембраны с TGFβ2 или TGFβ3.
На Фиг. 37 представлено изображение, на котором изображен TGFβ2 из комплекса 4A11v2 (левое изображение) и TGFβ3 из комплекса TGFβR1/TGFβR2. Конформационные изменения в структуре TGFβ2 могут приводить к изменению способности TGFBR1 связываться с одним мономером.
На Фиг. 38 изображено взаимодействие остатков 4A11v2 (левая структура) и 4A11v7 (правая структура) тяжелой цепи; взаимодействия каркасных областей VH являются комплементарными в обеих структурах.
На Фиг. 39 изображены сравнения каркасных областей VH 4A11v2 и 4A11v7. В 4A11v2 Q81 упакован напротив R19 в высокоэнергетическом ротамере и неудобном положении. В 4A11v7 K81 образует слабую Н-связь с S79, поэтому остатки каркасной области в v2, вероятно, являются отталкивающими, в то время как остатки v7 являются более пермиссивными.
Фиг. 40 включает изображения, на которых иллюстрируется одновалентное (левое изображение) и двухвалентное (правое изображение) связывание 4А11 с TGFβ2.
На Фиг. 41 представлен график, на котором изображена относительная активность люциферазы репортерных клеток MLEC после кокультивирования с клетками 293Т, трансфицированными плазмидой, кодирующей FL-TGFβ1 или FL-TGFβ3 (дикого типа или мутанты по RGE) с плазмидами, кодирующими интегрины αv и β6, или без них. **, Р<0,01; Р<10-3; ****, Р<10-4; н. зн., Р>0,05 на основе непарного двустороннего t-критерия Стьюдента.
На Фиг. 42 представлен график, на котором изображена относительная активность люциферазы в супернатантах (SN) репортерных клеток MLEC (измерение активности TGFβ). SN собирали из клеток, трансфицированных плазмидой, кодирующей FL-TGFβ1, FL-TGFβ2, FL-TGFβ3, или пустым вектором (контроль). Супернатанты подкисляли, как указано (обработка HCl). **, Р<0,01; ***, P<10-3; ****, Р<10-4; н. зн., Р>0,05 на основе непарного двустороннего t-критерия Стьюдента.
На Фиг. 43А и Фиг. 43Б представлены графики, на которых изображена активность TGFβ, оцененная с помощью щелочной фосфатазы, высвобождаемой репортерными клетками HEK-Blue TGFβ (Фиг. 43А), или с помощью анализа репортерных клеток MLEC (относительная активность люциферазы) (Фиг. 43Б). На Фиг. 43А супернатанты (SN) собирали из клеток 293Т, трансфицированных плазмидой, кодирующей FL-TGFβ1, FL-TGFβ2, FL-TGFβ3, или пустым вектором (контроль). На Фиг. 43Б SN собирали из клеток COS-7, трансфицированных плазмидой, кодирующей FL-TGFβ1, FL-TGFβ3, или пустым вектором (контроль). **, Р<0,01; ***, Р<0,001; ****, Р<10-4; н. зн., Р>0,05 на основе непарного двустороннего t-критерия Стьюдента. Данные представляют собой средние значения ± SEM на основе экспериментов, проведенных в трех повторностях. Планки погрешностей не отображаются, если они короче размеров соответствующих им символов.
На Фиг. 44 представлен график, на котором изображена активность TGFβ, оцененная с помощью анализа репортерных клеток MLEC (относительная активность люциферазы). Клетки 293Т трансфицировали плазмидами, кодирующими различные формы FL-TGFβ и интегрины αv и β6. Активности TGFβ измеряли с помощью репортерных клеток MLEC после кокультивирования. Цитохалазин D (30 мкМ) добавляли в начале кокультивирования, если было указано («Цитохал. D»). «MLE» представляет собой контроль. **, Р<0,01; ***, Р<0,001; ****, Р<10-4; н. зн., Р>0,05 на основе непарного двустороннего t-критерия Стьюдента.
На Фиг. 45 представлен график, на котором изображена активность TGFβ, оцененная с помощью анализа репортерных клеток MLEC (относительная активность люциферазы) в супернатантах, собранных из клеток 293Т, трансфицированных плазмидами, кодирующими белки FL-TGFβ дикого типа или с мутацией по сайту фурина, показывающий потерю активностей TGFβ при использовании мутантных белков. **, Р<0,01; Р<0,001; ****., P<10-4; н. зн., Р>0,05 на основе непарного двустороннего t-критерия Стьюдента. Данные представляют собой средние значения ± SEM на основе экспериментов, проведенных в трех повторностях. Планки погрешностей не отображаются, если они короче размеров соответствующих им символов.
На Фиг. 46 представлено изображение вестерн-блоттинга, показывающее кинетику фосфорилирования smad2 в клетках NHLF, обработанных рекомбинантными белками или супернатантами. В качестве контролей клетки NHLF также обрабатывали супернатантом из нетрансфицированных клеток (SN.ctr1) или рекомбинантным ВМР2 в течение 1 часа. Для контроля загрузок между различными образцами концентрации белка в супернатантах измеряли с помощью ВСА, и на каждую дорожку загружали 20 мкг белка.
На Фиг. 47 представлен график, на котором изображена активность TGFβ, измеряемая в репортерных клетках MLEC (относительная активность люциферазы) после инкубации с серией концентраций слитых белков изоформ Fc-FL-TGFβ человека. «Фон» означает, что слитый белок не добавляли. Данные представляют собой средние значения ± SEM на основе экспериментов, проведенных в трех повторностях, и планки погрешностей не отображаются, если они короче, чем размеры соответствующих им символов.
На Фиг. 48 представлен график, изображающий активность TGFβ, измеряемую в репортерных клетках MLEC (относительная активность люциферазы) после инкубации со слитыми белками Fc-FL-TGFβ2 или Fc-FL-TGFβ3 человека (30 нг/мл) и специфическими в отношении изоформы антителами: 19D8 (антитело к TGFβ1); 6F12 (антитело к TGFβ2); 2А10 (антитело к TGFβ3); и 1D11 (антитело ко всем видам TGFβ) в концентрации 3 мкг/мл. «Фон» означает, что слитый белок не добавляли. «Отсутствует» означает, что антитело не добавляли. Данные представляют собой средние значения ± SEM на основе экспериментов, проведенных в трех повторностях, и планки погрешностей не отображаются, если они короче, чем размеры соответствующих им символов.
На Фиг. 49 представлена таблица, в которой изображено сходство первичных аминокислотных последовательностей (%) среди изоформ TGFβ человека пептидов, ассоциированных с латентностью (LAP), и зрелых доменов (активных).
На Фиг. 50А, Фиг. 50Б и Фиг. 50В представлены графики, на которых показаны кривые титрования на основе анализа репортерных клеток MLEC (активность TGFβ) после инкубации зрелого пептида (1 нг/мл) TGFβ1, TGFβ2 или TGFβ3 (графики слева направо) с серией концентраций слитых белков Fc-LAP человека, как указано. Данные представляют собой средние значения ± SEM на основе экспериментов, проведенных в трех повторностях, и планки погрешностей не отображаются, если они короче, чем размеры соответствующих им символов.
На Фиг. 51 представлена таблица, в которой изображены IC50 на основе кривых титрования, изображенных на Фиг. 50.
На Фиг. 52А и Фиг. 52Б представлены графики, на которых изображены процентные содержания (%) нового гидроксипролина (среднее ± SEM), измеряемые в качестве индикатора вновь синтезированного коллагена у мышей WT или изоформы TGFβ CKO. На Фиг. 52А n=5 (каждая группа с применением солевого раствора), n=20 (WT, обработанные блеомицином (BLM»)), n=24 мыши (р2.cKO, BLM); на Фиг. 52Б n=7 (WT, солевой раствор), n=14 (WT, BLM), n=13 (β3.cKO, BLM) и n=13 (β2/3.cDKO, BLM); * Р<0,05, *** Р<0,001 согласно однофакторному ANOVA с применением критерия Даннетта.
На Фиг. 53А и Фиг. 53Б представлены столбиковые диаграммы, на которых изображена экспрессия генов Serpine1, Fn1 и Col1a1 в целом легком, как определено с помощью количественной ОТ-ПЦР через 14 дней после I.T. инсталляции солевого раствора или блеомицина и обработки антителом изотипического контроля, антителом к TGFβ2 (6F12) (Фиг. 53А) или антителом к TGFβ3 (2А10) (Фиг. 53Б). На Фиг. 53А n=5 (солевой раствор), n=15 (контроль, блеомцин (βLM»)) и n=14 (6F12, BLM); на Фиг. 53Б n=5 (солевой раствор), n=19 (контроль, BLM) и n=20 мышей (2А10, BLM). *Р<0,05, **Р<0,01, *** Р<0,001, **** Р<0,0001, н. зн., Р>0,05 согласно ANOVA с применением критерия Даннета.
На Фиг. 53В представлена столбиковая диаграмма, на которой изображены уровни вновь синтезированного коллагена, определенные в день 24 после инсталляции солевого раствора (n=10) или блеомицина (BLM»). Животных обрабатывали изотипическим контролем (n=21), комбинацией (n=23) 6F12 и 2А10 или антителом к TGFβ2/3 (4А11). Уровни доз 4А11 были высокими (10 мг/кг; 3 раза/неделя, n=23); средними (10 мг/кг; 1 раз/неделя, n=24) или низкими (2,5 мг/кг; 1 раз/неделя, n=23) соответственно. *Р<0,05, **Р<0,01, *** Р<0,001, **** Р<0,0001, н. зн., Р>0,05 согласно ANOVA с применением критерия Даннета.
На Фиг. 53Г представлена столбиковая диаграмма, на который изображены индексы заболевания, определенные с помощью анализов патологии. Легкие мышей собирали через 24 дня после инсталляции либо солевого раствора (n=5), либо блеомицина (BLM») с помощью обработки изотипическим контролем (n=7) или 4А11 (высокая доза, n=12). *Р<0,05, **Р<0,01, *** Р<0,001, **** Р<0,0001, н. зн., Р>0,05 согласно ANOVA с применением критерия Даннета.
Фиг. 53Д включает репрезентативные изображения легких мыши, полученные при иммуногистохимическом окрашивании коллагена III.
Фиг. 54А включает столбиковые диаграммы, на которых изображены относительные уровни экспрессии генов изоформ TGFp в печени пациентов с NASH с легким фиброзом (F0 и F1) (N=40) по сравнению с тяжелым фиброзом (F3 и F4) (N=32).
Фиг. 54Б включает точечные диаграммы, на которых изображены относительные уровни экспрессии генов изоформ TGFβ в печени мышей, определенные с помощью количественной ОТ-ПЦР через 6 недель после начала обработки несущей средой или CCl4; n=8 для обеих групп.
Фиг. 54В включает столбиковые диаграммы, на которых изображена экспрессия генов Col1al и Col3a1 в печени, определенная с помощью количественной ОТ-ПЦР через 6 недель после начала обработки несущей средой или CCl4; животных профилактически обрабатывали либо изотипическим контролем, либо 4А11; n=8 для всех групп.*, Р<0,05; **, Р<0,01; P<10-4; н. зн., Р>0,05 согласно непарного двустороннего t-критерия Стьюдента (А и Б); **Р<0,01 согласно однофакторного ANOVA с применением критерия Даннетта.
На Фиг. 54Г представлена столбиковая диаграмма, на которой изображены индексы патологии печени, определенные с помощью гистопатологического анализа; печень мышей собирали через 6 недель после обработки несущей средой или CCl4; n=8 для всех групп.*, Р<0,05; **, Р<0,01; ****, Р<10-4; н. зн., Р>0,05 согласно непарного двустороннего t-критерия Стьюдента (А и Б); **Р<0,01 согласно однофакторного ANOVA с применением критерия Даннетта.
На Фиг. 55А представлена таблица, в которой изображена аффинность связывания (KD) антител 6F12 и 4А11 с TGFβ2 человека и мыши, определенная с помощью Biacore SPR.
Фиг. 55Б включает линейные графики, на которых изображены активности TGFβ, измеренные с помощью репортерных клеток MLEC. Зрелый пептид (1 нг/мл) TGFβ2 человека или мыши инкубировали с серией концентраций антител 6F12 или 4А11. Кривые более всего соответствуют модели ингибирования доза-ответ. Фон (фоновый уровень), зрелый пептид не добавляли.
На Фиг. 56А представлена схема модели колита для оценки повышенных воспалительных ответов, связанных с антителами к TGFβ.
На Фиг. 56Б представлен вес толстой кишки (в граммах), измеренный в день 24 в модели колита из Фиг. 56А. N=10 для всех групп, кроме группы без обработки (n=6).
На Фиг. 56В представлена относительная экспрессия генов воспаления (проанализированных на основе РНК толстой кишки) в модели колита из Фиг. 56А. Анализировали от трех до пяти мышей из каждой группы, как указано.
На Фиг. 56Г представлено количество иммунных клеток в собственной пластинке у остальных мышей в каждой группе (3-5/группа), определенное с помощью проточной цитометрии с использованием их поверхностных маркеров, в модели колита из Фиг. 56А.
ПОДРОБНОЕ ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
I. ОПРЕДЕЛЕНИЯ
Используемые в данном документе термины «фактор некроза опухоли β» и «TGFp» используются взаимозаменяемо и относятся к любой нативной изоформе TGFp из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), если не указано другое. Этот термин охватывает «полноразмерный» непроцессированный TGFβ, а также любую форму TGFβ, полученную в результате процессинга в клетке. Данный термин также включает встречающиеся в природе варианты TGFβ, например, сплайс-варианты или аллельные варианты. Изоформы TGFβ имеют формы-предшественники (незрелые) и зрелые формы. Пептид, ассоциированный с латентностью, как изображено на Фиг. 1, расщепляется фуриновыми протеазами внутриклеточно и образует нековалентное взаимодействие с рецептор-связывающим доменом. Этот комплекс секретируется из клетки отдельно или ковалентно связывается посредством пептида, ассоциированного с латентностью, с молекулами «окружающей среды», такими как GARP, LRRC33 или латентные TGFβ-связывающие белки (LTBP)1-4. Комплекс LAP и рецептор-связывающего домена называется малым латентным комплексом (SLC), а комплекс LAP, рецептор-связывающего домена и молекулы окружающей среды называется большим латентным комплексом (LLC).
Аминокислотные последовательности для TGFβ1, TGFβ2 и TGFβ3 представлены далее:
TGFβ1 человека
MPPSGLRLLPLLLPLLWLLVLTPGRPAAGLSTCKTIDMELVKRKRIEAIRGQILSKLRLASPPSQGEVPPGPLPEAVLALYNSTRDRVAGESAEPEPEPEADYYAKEVTRVLMVETHNEIYDKFKQSTHSIYMFFNTSELREAVPEPVLLSRAELRLLRLKLKVEQHVELYQKYSNNSWRYLSNRLLAPSDSPEWLSFDVTGVVRQWLSRGGEIEGFRLSAHCSCDSRDNTLQVDINGFTTGRRGDLATIHGMNRPFLLLMATPLERAQHLQSSRHRRALDTNYCFSSTEKNCCVRQLYIDFRKDLGWKWIHEPKGYHANFCLGPCPYIWSLDTQYSKVLALYNQHNPGASAAPCCVPQALEPLPIVYYVGRKPKVEQLSNMIVRSCKCS (SEQ ID NO: 1).
TGFβ2 человека
MHYCVLSAFLILHLVTVALSLSTCSTLDMDQFMRKRIEAIRGQILSKLKLTSPPEDYPEPEEVPPEVISIYNSTRDLLQEKASRRAAACERERSDEEYYAKEVYKIDMPPFFPSENAIPPTFYRPYFRIVRFDVSAMEKNASNLVKAEFRVFRLQNPKARVPEQRIELYQILKSKDLTSPTQRYIDSKVVKTRAEGEWLSFDVTDAVHEWLHHKDRNLGFKISLHCPCCTFVPSNNYIIPNKSEELEARFAGIDGTSTYTSGDQKTIKSTRKKNSGKTPHLLLMLLPSYRLESQQTNRRKKRALDAAYCFRNVQDNCCLRPLYIDFKRDLGWKWIHEPKGYNANFCAGACPYLWSSDTQHSRVLSLYNTINPEASASPCCVSQDLEPLTILYYIGKTPKIEQLSNMIVKSCKCS (SEQ ID NO: 2).
TGFβ3 человека
MKMHLQRALVVLALLNFATVSLSLSTCTTLDFGHIKKKRVEAIRGQILSKLRLTSPPEPTVMTHVPYQVLALYNSTRELLEEMHGEREEGCTQENTESEYYAKEIHKFDMIQGLAEHNELAVCPKGITSKVFRFNVSSVEKNRTNLFRAEFRVLRVPNPSSKRNEQRIELFQILRPDEHIAKQRYIGGKNLPTRGTAEWLSFDVTDTVREWLLRRESNLGLEISIHCPCHTFQPNGDILENIHEVMEIKFKGVDNEDDHGRGDLGRLKKQKDHHNPHLILMMIPPHRLDNPGQGGQRKKRALDTNYCFRNLEENCCVRPLYIDFRQDLGWKWVHEPKGYYANFCSGPCPYLRSADTTHSTVLGLYNTLNPEASASPCCVPQDLEPLTILYYVGRTPKVEQLSNMVVKSCKCS (SEQ ID NO: 3).
Термин «TGFβ1», используемый в данном документе, относится к любому нативному TGFβ1 из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), если не указано иное. Данный термин охватывает «полноразмерный» непроцессированный TGFβ1, а также любую форму TGFβ1, полученную в результате процессинга в клетке. Данный термин также включает встречающиеся в природе варианты TGFβ1, например, сплайс-варианты или аллельные варианты. Аминокислотная последовательность иллюстративного TGFβ1 человека изображена на Фиг. 1 (SEQ ID NO: 1).
Используемый в данном документе термин «TGFβ2» относится к любому нативному TGFβ2 из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), если не указано иное. Этот термин охватывает «полноразмерный» непроцессированный TGFβ2, а также любую форму TGFβ2, полученную в результате процессинга в клетке. Данный термин также включает встречающиеся в природе варианты TGFβ2, например, сплайс-варианты или аллельные варианты. Аминокислотная последовательность иллюстративного TGFβ2 человека изображена на Фиг. 1 (SEQ ID NO: 2).
Термин «TGFβ3», используемый в данном документе, относится к любому нативному TGFβ3 из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), если не указано иное. Данный термин охватывает «полноразмерный» непроцессированный TGFβ3, а также любую форму TGFβ3, полученную в результате процессинга в клетке. Данный термин также включает встречающиеся в природе варианты TGFβ3, например, сплайс-варианты или аллельные варианты. Аминокислотная последовательность иллюстративного TGFβ3 человека изображена на Фиг. 1 (SEQ ID NO: 3).
Используемый в данном документе термин «TGFβ» означает любую одну, две или все три изоформы TGFβ TGFβ1, TGFβ2 и TGFβ3, описанные выше.
Используемые в данном документе термины «специфически связывается» и «связывается специфически с» относятся к антителу, селективуно или предпочтительно связывающемуся с его антигеном-мишенью. Предпочтительно аффинность связывания с антигеном характеризуется KD, составляющей 10-9 моль/л или ниже (например, КD-10 моль/л), предпочтительно KD, составляющей 10-10 моль/л или ниже (например, 10-12 моль/л). Аффинность связывания определяют с помощью стандартного анализа связывания, такого как методика поверхностного плазмонного резонанса (BIACORE®).
Используемый в данном документе термин «антитело к TGFβ» означает антитело, которое специфически связывается с одной или более изоформами TGFβ. Таким образом, используемый в данном документе термин «антитело к TGFβ3» относится к моноспецифическому антителу, которое специфически связывается с TGFβ3; термин «антитело к TGFβ2» относится к моноспецифическому антителу, которое специфически связывается с TGFβ2 (например, TGFβ2 человека); термин «антитело к TGFβ2/3» относится к антителу с двойной специфичностью, которое специфически связывается с TGFβ2 (например, TGFβ2 человека) и TGFβ3 (например, TGFβ3 человека); термин «антитело к TGFβ1» относится к моноспецифическому антителу, которое специфически связывается с TGFβ1 (например, TGFβ1 человека); и термин «специфическое антитело ко всем видам TGFβ» относится к антителу, которое связывается со всеми тремя изоформами TGFβ (TGFβ1, TGFβ2 и TGFβ3, например, TGFβ1, TGFβ2 и TGFβ3 человека)). Антитело к TGFβ, описанное в данном документе, которое является моноспецифическим или характеризуется двойной специфичностью в отношении определенной(ых) изоформы(форм) TGFβ, также упоминается в данном документе как «селективное в отношении изоформы антитело к TGFβ». В одном варианте осуществления степень связывания селективного в отношении изоформы антитела к TGFβ (например, антитела к TGFβ1, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) с изоформой(ами) TGFβ, для которой(ых) антитело не является специфическим, составляет менее приблизительно 10% связывания антитела со своей(ими) изоформой(ами)-мишенью(ями) TGFβ, как измерено, например, с помощью радиоиммуноанализа (RIA) или поверхностного плазмонного резонанса (SPR). В другом варианте осуществления степень связывания селективного в отношении изоформы антитела к TGFβ с изоформой(ами) TGFβ, в отношении которой(ых) антитело не является специфичным, составляет менее приблизительно 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или предпочтительно менее приблизительно 1% связывания антитела с его изоформой(ами)-мишенью(ями) TGFβ, как измерено, например, с помощью RIA или SPR. В одном варианте осуществления селективное в отношении изоформы антитело к TGFβ относится к антителу, которое способно связывать изоформу(ы) TGFβ, к которой(ым) антитело является специфическим, с достаточной аффинностью, так что антитело можно применять в качестве диагностического и/или терапевтического средства при нацеливании на изоформу(ы) TGFβ. В одном варианте осуществления степень связывания селективного а отношении изоформы антитела к TGFβ с неродственным белком составляет менее приблизительно 10% или менее приблизительно 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или предпочтительно менее приблизительно 1% связывания селективного в отношении изоформы антитела к TGFβ, измеренного, например, с помощью радиоиммунологического анализа (RIA). В определенных вариантах осуществления антитело к TGFβ имеет константу диссоциации (K-D) ≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например, 10-8 М или меньше, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М) в отношении своей изомормы(форм)-мишени(ей) TGFβ. В предпочтительном варианте осуществления селективное в отношении изоформы антитело к TGFβ имеет K-D ≤250 мкМ.
Используемый в данном документе термин «селективно нейтрализовать» и его грамматические вариации в отношении селективного в отношении изоформы антитела к TGFβ, описанного в данном документе, означает, что антитело специфически связывается и нейтрализует изоформу(ы), в отношении которой(ых) антитело является селективным, но не нейтрализует другую(ие) изоформу(ы). Так, например, антитело к TGFβ1, которое селективно нейтрализует TGFβ1, не нейтрализует TGFβ2 или TGFβ3; антитело к TGFβ2, которое селективно нейтрализует TGFβ2, не нейтрализует TGFβ1 или TGFβ3; антитело к TGFβ3, которое селективно нейтрализует TGFβ3, не нейтрализует TGFβ1 или TGFβ2; антитело к TGFβ2/3, которое селективно нейтрализует TGFβ2 и TGFβ3, не нейтрализует TGFβ1. В определенных вариантах осуществления селективное в отношении изоформы антитело к TGFβ, описанное в данном документе, специфически связывается с эпитопом изоформы TGFβ, который является консервативным у разных видов. В некоторых вариантах осуществления способность антитела (например, селективного в отношении изоформы антитела к TGFβ, описанного в данном документе) селективно нейтрализовать одну или более изоформ TGFβ можно определить в анализе ингибирования in vitro, например, клеточном анализе ингибирования TGFβ, описанном ниже.
Используемый в данном документе термин «TGFBR» означает рецептор TGFβ. Димерные рецептор-связывающие домены всех трех изоформ TGFβ связываются с парами гетеродимерных рецепторных комплексов TGFBR1 и TGFBR2; затем тетрамерный рецепторный комплекс активирует внутриклеточную передачу сигналов посредством активности рецепторной тирозинкиназы (RTK) TGFBR1, также известной как ALK5 (Weiss et al., Wiley Interdiscip.Rev. Dev. Biol. 2:47-63 (2013)). В канонической передаче сигналов TGFBR ALK5 фосфорилирует SMAD2 и SMAD3, которые затем ассоциируются с SMAD4, транслоцируются в ядро и направляют транскрипцию генов. Комплекс Smad активирует транскрипцию генов миофибробластов, включая αSMA, кальпонин и коллаген (Usuki et al., J. Nippon Med. Sch. 79:46-59 (2012); Carthy et al., PloS one 6:el9809 (2011); и Gu et al., Acta Pharmacol. Sin. 28:382-391 (2007)). Существуют дополнительные сигнальные пути, не зависящие от SMAD, которые могут быть активированы TGFβ в определенных контекстах, включая МАР-киназы, AKT, JAK-STAT и NFβκB. Биохимические и структурные исследования показали, что сборка сигнальных комплексов TGFβ-TGFβR может иметь тонкие специфические в отношении изоформы различия: TGFβ1 и TGFβ3 более сильно связываются с TGFβR2 и образуют сильные взаимодействия только с TGFβR1 в комплексе с TGFβR2, в то время как TGFβ2 слабо связывается как с TGFBR1, так и с TGFBR2 в отдельности, и авидность может стимулировать образование полного комплекса (Radaev JBC 2009; 285, 14806-14814). TGFβ1 и TGFβ2 кристаллизуются в «закрытые» структуры, которые облегчают связывание с TGFBR1 и TGFBR2, в то время как TGFβ3 может принимать аналогичную «закрытую» или менее упорядоченную «открытую» структуру в кристаллической форме, что может приводить к различиям в авидности сборки комплекса лиганд-рецептор (Hinck, А.P. et al. 2016; Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a02210). Кроме того, TGFβR3 (бетагликан), несигнальный рецептор, может облегчать связывание TGFβ2 с комплексами TGFβR1/2, но, видимо, не играет аналогичной роли в связывании рецепторов TGFβ1 или TGFβ3 (del Re, JBC 2004; 279, 22765-22772). В то время как в большинстве клеток (включая эндотелиальные клетки) TGFβ передает сигналы посредством комплексов TGFBR1/2, в эндотелиальных клетках TGFβ1 и TGFβ3 также могут передавать сигналы посредством комплекса TGFBR2/ALK1, облегченные эндоглином, что способствует SMAD1/5-зависимой пролиферации эндотелия сосудов и ангиогенезу (EMBO J. 2004 Oct 13; 23(20): 4018-28). В конечном итоге, несмотря на эти различия в сборке сигнальных комплексов, все рекомбинантные рецептор-связывающие домены TGFβ1, 2 и 3 способны индуцировать TGFBR-зависимую передачу сигналов SMAD в одинаковой степени в клеточных анализах in vitro. Следовательно, любые биологические различия в активности эндогенных изоформ TGFββ скорее связаны с различиями в их паттернах экспрессии и механизмах высвобождения из SLC или LLC, чем с различиями в их рецептор-связывающих доменах.
Используемый в данном документе термин «нейтрализовать» и его грамматические вариации в отношении селективного в отношении изоформ антитела к TGFβ, описанного в данном документе, означает, что антитело в измеримой степени ингибирует свою(и) изоформу(мы)-мишень(ни) TGFβ от индукции передачи сигналов посредством комплекса TGFBR.
Используемый в данном документе термин «непосредственно контактирует» и его грамматические варианты в отношении антигенсвязывающего домена антитела к TGFβ означает, что антигенсвязывающий домен находится в пределах 15-8, 8, 8-5 или предпочтительно в пределах 5 ангстрем от остатка аминокислоты в своем соответствующем эпитопе.
«Нарушения TGFβ или «нарушения, связанные с TGFβ относятся к любому нарушению, заболеванию или состоянию, при котором лечение селективным в отношении изоформы антитела к TGFβ, представленным в данном документе, может оказывать благоприятное действие. Они включает хронические и острые нарушения или заболевания, включая патологические состояния, которые предрасполагают млекопитающего к развитию рассматриваемого нарушения. Заболевания, подлежащие лечению в данном документе, включают заболевания, характеризующиеся накоплением внеклеточного матрикса, заболевания, вызванные циркулирующим TGFβ или TGFβ, активированным в локальном участке, включая одну или более изоформ TGFβ, состояния, вызванные подавлением иммунной системы вследствие эндогенного продуцирования TGFβ, формы острой иммунной недостаточности, возникающие в результате тяжелых травм, ожогов и заболеваний, таких как вирусные или бактериальные инфекции, полиорганные системные заболевания вследствие продуцирования или сверхпродуцирования TGFβ, и опухоли, продуцирующие TGFβ.
Используемые в данном документе термины «фиброз», «фиброзные состояния» и «фибротические состояния» имеют одинаковое значение. В определенных вариантах осуществления фиброзные состояния представляют собой состояния, опосредованные стимулятором фиброза. Иллюстративные стимуляторы фиброза включают без ограничения TGFβ, эндотелии, молочную кислоту (посредством лактатдегидрогеназы), IL-1, Thy-1 (CD90), фактор роста соединительной ткани («CTGF»), а также их комбинации. В определенных вариантах осуществления фиброзное состояние представляет собой состояние, которое опосредовано TGFβ. В определенных вариантах осуществления фиброзное состояние представляет собой состояние, которое опосредовано одним или более из TGFβ1, TGFβ2 и TGFββ3. В определенных вариантах осуществления фиброзное состояние представляет собой состояние, которое опосредовано одним или обоими из TGFβ2 и TGFβ3. В определенных вариантах осуществления фиброзное состояние представляет собой состояние, которое опосредовано TGFβ2. В определенных вариантах осуществления фиброзное состояние представляет собой состояние, которое опосредовано TGFβ3. Иллюстративные фиброзные состояния более подробно описаны в данном документе ниже.
«Системный склероз» (SSc) или «склеродермия» представляет собой сложное и гетерогенное заболевание, основными признаками которого являются фиброз кожи и тканей, сосудистые изменения и аутоантитела к различным клеточным антигенам. Клинические проявления системного склероза могут варьировать от ограниченного поражения кожи до тяжелой дисфункции внутренних органов. Патология внутренних органов является основным фактором заболеваемости при данном заболевании, при этом наиболее часто поражаются почки, пищевод, сердце и легкие. В общепринятой классификации SSc выделяют две основные подгруппы: ограниченный кожный SSc (1cSSc) и диффузный кожный SSc (dcSSc). Gabrielli et al. Mechanisms of disease. Scleroderma. N Engl J Med 360:1989-2003 (2009). В одном варианте осуществления пациент с системным склерозом был классифицирован в соответствии с критериями Американского колледжа ревматологов (ранее Американская ассоциация ревматизма) для классификации системной склеродермии на основании: основного критерия: диффузный (туловищный) склероз (уплотнение, утолщение кожи и - индурация без ямок при вдавливании); и второстепенных критериев: (1) склеродактилия (только пальцы рук и/или пальцев ног), (2) пальцевые рубцы с ямками при вдавливании или потеря вещества подушечек пальцев (потеря пульпы) и (3) двусторонний базилярный легочный фиброз, при этом пациент с системным склерозом должен соответствовать основному критерию или двум из трех второстепенных критериев. См. Subcommittee for Scleroderma Criteria of the American Rheumatism Association, Diagnostic and Therapeutic Criteria Committee. Preliminary criteria for the classification of systemic sclerosis (scleroderma). Arthritis Rheum 23:581-90 (1980).
Используемый в данном документе термин «хроническая обструктивная болезнь легких» («COPD») представляет собой общий термин, используемый для описания группы заболеваний дыхательных путей, обычно характеризующихся обструкцией или ограничением воздушного потока. Это состояние также может быть известно под терминами хроническое обструктивное заболевание дыхательной системы (CORD), хроническое обструктивное заболевание дыхательных путей (COAD), хроническое обструктивное заболевание легких (COLD) или хроническое ограничение дыхательных путей (CAL). Используемый в данном документе термин COPD предназначен для включения всех таких ссылок. Клиническое течение COPD характеризуется хронической инвалидизацией с периодическими обострениями, чаще возникающими в зимние месяцы. Острое обострение COPD может быть определено как устойчивое ухудшение симптомов пациента по сравнению с его или ее обычным стабильным состоянием, которое выходит за рамки нормальных ежедневных вариаций и имеет острое начало. Когда возникают острые обострения, они обычно проявляются увеличением образования мокроты, более гнойной мокротой, изменением цвета мокроты, усилением кашля, симптомами со стороны верхних дыхательных путей (например, простудой и болью в горле), усилением свистящего дыхания, стеснением в груди, снижением толерантности к физической нагрузке, повышенной утомляемостью, задержкой жидкости, острой спутанностью сознания и усилением одышки. Хотя инфекционная этиология является причиной большинства обострений, воздействие аллергенов, загрязняющих веществ или вдыхаемых раздражителей также может играть роль. Инфекционные средства, которые, как известно, вызывают обострения COPD, включают: риновирусы, вирус гриппа, вирус парагриппа, коронавирус, аденовирус, респираторно-синцитиальный вирус, Chlamydia pneumoniae, Haemophilus influenzae, Streptococcus pneumoniae, Moraxella catarrhalis, Staphylococcus aureus, Mycloplasma pneumoniae и Pseudomonas aeruginosa. Известно, что загрязняющие вещества, вызывающие обострения, включают двуокись азота, твердые частицы, двуокись серы и озон. Несмотря на эти известные причины, точная причина обострений может быть неустановленной в случае до 30% диагностированных случаев обострения COPD. Глобальная инициатива по хронической обструктивной болезни легких (GOLD) определяет COPD как болезненное состояние, характеризующееся не полностью обратимым ограничением скорости воздушного потока. Ограничение воздушного потока обычно прогрессирует и связано с аномальным воспалительным ответом легких на вредные частицы или газы. Американское торакальное общество (ATS) определяет COPD как болезненный процесс, включающий прогрессирующую хроническую обструкцию дыхательных путей вследствие хронического бронхита, эмфиземы или того и другого. Хронический бронхит клинически определяется как чрезмерный кашель и выделение мокроты в течение большей части дней в течение по меньшей мере трех месяцев в течение по меньшей мере двух лет подряд. Эмфизема характеризуется хронической одышкой (нехваткой воздуха), возникающей в результате разрушения легочной ткани и расширения воздушных пространств. Еще одним состоянием, которое обычно включает термином COPD, является бронхоэктатическая болезнь, которая представляет собой аномальное растяжение и расширение дыхательных путей, вызванное скоплением и закупоркой слизи. В таких условиях ослабленные проходы могут покрыться рубцами и деформироваться, способствуя накоплению большего количества слизи и бактерий, что приведет к циклу инфекции и блокировке дыхательных путей.
Используемый в данном документе термин «ILD» относится к интерстициальному заболеванию легких. Интерстициальные заболевания легких включают большую и разнообразную группу из более чем 200 заболеваний легких и состояний дыхательной системы, характеризующихся воспалением и фиброзом интерстиция, ткани и пространства между воздушными мешками легких (см., например, du Bois, Nat. Rev. Drug Discov. 2010, 9, 129-140). При «прогрессирующих фиброзирующих интерстициальных заболеваниях легких (PF-ILD)» ответ на повреждение легких при фиброзирующих ILD включает развитие фиброза, который становится прогрессирующим, самоподдерживающимся и независимым от исходной клинической связи или триггера.
Используемые в данном документе термины «идиопатический легочный фиброз» и «IPF» относятся к ограниченному заболеванию легких, характеризующемуся прогрессирующим интерстициальным фиброзом легочной паренхимы, от которого страдают примерно 100000 пациентов в Соединенных Штатах Америки (Raghu et al., Am J Respir Crit Care Med 174:810-816 (2006)). Данный интерстициальный фиброз, связанный с IPF, приводит к прогрессирующей потере функции легких, что приводит к смерти вследствие дыхательной недостаточности у большинства пациентов. Медиана выживаемости с момента постановки диагноза составляет 2-3 года (Raghu et al., Am J Respir Crit Care Med 183:788-824 (2011)). Этиология и ключевые молекулярные и патофизиологические факторы IPF неизвестны. В некоторых вариантах осуществления диагноз IPF подтверждается обнаружением обычной интерстициальной пневмонии (UIP) при гистопатологической оценке легочной ткани, полученной при биопсии вследствие хирургического вмешательства. Критерии диагностики IPF известны. Ryu et al. (1998) Mayo Clin. Proc. 73:1085-1101.
Используемый в данном документе термин «фиброз желудочно-кишечного тракта» относится к фиброзу желудочно-кишечного тракта, включая, например, полость рта, пищевод, желудок, тонкую кишку, толстую кишку и задний проход. Таким образом, «фиброз желудочно-кишечного тракта» включает фиброз кишечника. «Фиброз кишечника» является частым осложнением воспалительного заболевания кишечника (IBD) и обычно определяется как чрезмерное накопление рубцовой ткани в стенке кишечника. Фиброз кишечника может возникать при обеих формах IBD: язвенном колите и болезни Крона. «Фиброз желудочно-кишечного тракта» включает без ограничения фиброз, связанный с болезнью Крона, язвенного колита, коллагенозного колита, колоректального фиброза, атрофии ворсинок, гиперплазии крипт, образования полипов, заживления язвы желудка и микроскопического колита.
Используемый в данном документе термин «мониторинг прогрессирования заболевания» относится к оценке субъекта (например, субъекта, страдающего нарушением, связанным с TGFβ, например, субъекта, проходящего лечение антителом к TGFβ, как описано в других разделах данного документа) через последовательные интервалы времени для определения того, ухудшились, стабилизировались или нормализовались (т.е. стали менее выраженными) симптомы заболевания. Например, мониторинг прогрессирования фиброза (например, SSc или IPF или другого фиброза) у субъекта может, в некоторых случаях, включать мониторинг изменений в наборе сигнатур 18 генов TGFβ, описанном в таблице 2 ниже, общей частоте ответов, продолжительности ответа, качестве жизни, экспрессии и/или активности маркеров заболевания (например, экспрессии некоторых других генов и/или белков) или других критериев, известных в данной области техники. Можно использовать дополнительные подходы к мониторингу прогрессирования заболевания у пациента с нарушением, связанным с TGFβ, включая, например, измерение ответа на лечение с помощью методов визуализации, которые более подробно описаны в других разделах данного документа.
Используемые в данном документе термины «мониторинг хода лечения» или «мониторинг ответа на лечение» используются взаимозаменяемо и относятся к оценке субъекта (например, субъекта, страдающего нарушением, связанным с TGFβ, например, субъекта, проходящего лечение антителом к TGFβ, как описано в других разделах данного документа) через последовательные интервалы времени во время или после лечения, чтобы определить, ухудшились, стабилизировались или нормализовались (т.е. стали менее тяжелыми) симптомы заболевания в результате лечения. Например, ход лечения у субъекта (например, у субъекта, который проходит или получает лечение иммунотерапевтическим средством, таким как без ограничения антитело к TGFβ, описанное в данном документе), можно отслеживать с использованием тех же критериев, которые используются для мониторинга прогрессирования заболевания.
Используемый в данном документе термин «обнаружение» включает любые средства обнаружения, включая прямое и косвенное обнаружение.
Используемый в данном документе термин «диагностика» используется в данном документе для обозначения идентификации или классификации молекулярного или патологического состояния, заболевания или состояния. Например, «диагноз» может относиться к выявлению определенного типа фиброза (например, SSc, IPF и т.д.) или другого опосредованного TGFβ нарушения. «Диагноз» может также относиться к классификации конкретного подтипа фиброзного состояния, например, по гистопатологическим или рентгенологическим критериям или по молекулярным признакам (например, подтип, характеризующийся экспрессией одного или комбинации определенных генов или белков, кодируемых указанными генами).
Термин «прогноз» используется в данном документе для обозначения прогнозирования вероятности выживания с течением времени, а также ухудшения одного или нескольких симптомов заболевания, связанного с TGFβ, с течением времени.
Используемый в данном документе термин «контрольный субъект» относится к здоровому субъекту, у которого не было диагностировано заболевание или состояние, представляющие интерес, например, фиброз, например, IPF, SSc и т.д., и который не страдает каким-либо признаком или симптомом, связанным заболеванием или состоянием.
Термин «образец», используемый в данном документе, относится к композиции, которая получена от субъекта, представляющего интерес, который содержит клеточный и/или другой молекулярный материал, который необходимо охарактеризовать и/или идентифицировать, например, на основании физических, биохимических, химических и/или физиологических характеристик. Например, выражение «патологический образец» и его вариации относится к любому образцу, полученному от представляющего интерес субъекта, который, как ожидается или как известно, содержит клеточный и/или молекулярный материал, который необходимо охарактеризовать.
Под «тканевым образцом» или «клеточным образцом» подразумевается сбор аналогичных клеток, полученных из ткани субъекта или пациента. Источником тканевого или клеточного образца может быть солидная ткань, например, из свежего, замороженного и/или законсервированного органа, тканевого образца, биоптата и/или аспирата; кровь или любые компоненты крови, такие как плазма крови; жидкости организма, такие как спинномозговая жидкость, околоплодная жидкость, перитонеальная жидкость или интерстициальная жидкость; клетки, полученные на любой стадии созревания плода или развития субъекта. Тканевый образец также может представлять собой первичные или культивируемые клетки или линии клеток. Необязательно тканевый или клеточный образец получен из пораженных заболеванием тканей/органов. Тканевый образец может содержать соединения, которые естественным образом не присутствуют в ткани в природе, такие как консерванты, антикоагулянты, буферы, фиксаторы, питательные вещества, антибиотики и т.п. Термины «эталонный образец», «эталонная клетка», «эталонная ткань», «контрольный образец», «контрольная клетка» или «контрольная ткань», используемые в данном документе, относятся к образцу, клетке или ткани, полученным из известного источника, или считается, что он не поражен заболеванием или состоянием, для идентификации которого используется способ или композиция по настоящему изобретению. В одном варианте осуществления эталонный образец, эталонную клетку, эталонную ткань, контрольный образец, контрольную клетку или контрольную ткань получают из здоровой части тела того же субъекта или пациента, у которого заболевание или состояние обнаруживают с помощью композиции или способа по настоящему изобретению. В одном варианте осуществления эталонный образец, эталонную клетку, эталонную ткань, контрольный образец, контрольную клетку или контрольную ткань получают из здоровой части тела индивидуума, который не представляет собой субъекта или пациента, у которого заболевание или состояние обнаруживают с помощью композиции или способа по настоящему изобретению.
Используемый в данном документе термин «сигнатура гена» используется взаимозаменяемо с «сигнатурой экспрессии гена» и относится к одному или комбинации генов, экспрессия которых указывает на субъекта или ткань или другой образец, выделенный от субъекта, имеющие высокую активность TGFβ, и/или указывает на то, что, вероятно, лечение ингибитором изоформы TGFβ, характеризующееся определенными молекулярными, патологическими, гистологическими, рентгенологическими и/или клиническими признаками, будет оказывать положительное действие на субъекта. В некоторых вариантах осуществления экспрессия одного или более генов, содержащих сигнатуру гена, повышена по сравнению с экспрессией у контрольных субъектов.
Используемый в данном документе термин «повышенный уровень экспрессии» или «повышенные уровни» относится к повышенной экспрессии мРНК или белка у субъекта (например, у субъекта, например, пациента, у которого имеется подозрение на наличие заболевания, связанного с TGFβ, или установлен его диагноз, например, фиброза, например, IPF, COPD, PF-ILD (например, SSc), фиброза печени (например, цирроза печени или хронического фиброза печени)) по сравнению с контролем, таким как индивидуум или индивидуумы, которые не страдает связанным с TGFβ нарушением.
В целях данного документа «акцепторная каркасная область человека» представляет собой каркасную область, содержащую аминокислотную последовательность каркасной области вариабельного домена легкой цепи (VL) или каркасной области вариабельного домена тяжелой цепи (VH), полученную из каркасной области человеческого иммуноглобулина или человеческой консенсусной каркасной области, как определено ниже. Человеческая акцепторная каркасная область, «полученная из» каркасной области человеческого иммуноглобулина или человеческой консенсусной каркасной области, может содержать такую же самую аминокислотную последовательность или может содержать изменения в аминокислотной последовательности. В некоторых вариантах осуществления число аминокислотных изменений составляет 10 или меньше, 9 или меньше, 8 или меньше, 7 или меньше, 6 или меньше, 5 или меньше, 4 или меньше, 3 или меньше, или 2 или меньше. В некоторых вариантах осуществления человеческая акцепторная каркасная область VL идентична по последовательности с последовательностью каркасной области VL человеческого иммуноглобулина или последовательностью человеческой консенсусной каркасной области.
Термин «аффинность» относится к силе суммарных общих нековалентных взаимодействий между одиночным сайтом связывания молекулы (например, антитела) и ее партнера по связыванию (например, антигена). Если не указано иное, в данном документе термин «аффинность связывания» относится к присущей молекуле аффинности связывания, отражающему взаимодействие между членами пары связывающихся компонентов (например, антителом и антигеном) при их соотношении 1:1. Аффинность молекулы X в отношении ее партнера Y обычно может быть представлена константой диссоциации (KD). Аффинность можно измерять с помощью общепринятых методов, известных в данной области техники, включая описанные в данном документе. Конкретные иллюстративные и типовые варианты осуществления для определения аффинности связывания описаны ниже.
Антитело с «созревшей аффинностью» относится к антителу с одним или более изменениями в одной или более гипервариабельных областях (HVR) по сравнению с исходным антителом, которое не содержит такие изменения, при этом такие изменения приводят к улучшению аффинности антитела в отношении антигена.
Термин «антитело» в данном документе используется в самом широком смысле и включает различные структуры антител, включая без ограничения моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они проявляют необходимую антигенсвязывающую активность.
«Фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, которая связывает антиген, с которым связывается интактное антитело. Примеры фрагментов антител включают без ограничения Fv, Fab, Fab', Fab'-SH, F(ab')-2; диатела; линейные антитела; молекулы одноцепочечных антител (например, scFv); и мультиспецифические антитела, образованные из фрагментов антител.
«Антитело, которое связывается с тем же эпитопом», как и эталонное антитело, относится к антителу, которое блокирует связывание эталонного антитела с его антигеном на 50% или больше в конкурентном анализе связывания, и наоборот, эталонное антитело блокирует связывание антитела с его антигеном на 50% и больше в конкурентном анализе связывания. Иллюстративный конкурентный анализ связывания представлен в данном документе.
Термин «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи получена от определенного источника или вида, тогда как остальная часть тяжелой и/или легкой цепи получена от другого источника или вида.
«Класс» антитела относится к типу константного домена или константной области, которые содержит его тяжелая цепь. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, а некоторые из них можно дополнительно разделить на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, соответствующие разным классам иммуноглобулинов, называются α, β, ε, γ и μ соответственно.
Термин «цитотоксическое средство», используемый в данном документе, относится к веществу, которое ингибирует или предупреждает функционирование клеток и/или приводит к гибели или разрушению клеток. Цитотоксические средства включают без ограничения радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu); химиотерапевтические средства или лекарственные средства (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие средства); средства, ингибирующие рост; ферменты и их фрагменты, такие как нуклеолитические ферменты; антибиотики; токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или варианты; и различные противоопухолевые или противораковые средства, раскрытые ниже.
«Эффекторные функции» относятся к видам биологической активности, характерным для Fc-области антитела, которые варьируются в зависимости от изотипа антитела. Примеры эффекторных функций антител включают: связывание C1q и комплементзависимую цитотоксичность (CDC); связывание с рецептором Fc; антителозависимую клеточно-опосредованную цитотоксичность (ADCC); фагоцитоз; понижающую регуляцию клеточных поверхностных рецепторов (например, В-клеточного рецептора) и активацию В-клетки.
«Эффективное количество» средства, например, фармацевтического состава, относится к количеству, эффективному в дозах и в течение периодов времени, необходимых для достижения требуемого терапевтического или профилактического результата.
Термин «терапевтически эффективное количество» относится к количеству, например, иммунотерапевтического средства (такого как иммунотерапевтическое средство, описанное в других разделах в данном документе), эффективному для «лечения» заболевания или нарушения у субъекта (например, млекопитающего, такого как человек).
Используемый в данном документе термин «тоцилизумаб» представляет собой рекомбинантное гуманизированное моноклональное антитело, которое связывается с человеческим рецептором интерлейкина-6 (IL-6R). Он представляет собой антитело на основе IgG1κ (гамма 1, каппа) с двумя тяжелыми цепями и двумя легкими цепями, образующими два антигенсвязывающих сайта. В предпочтительном варианте осуществления аминокислотные последовательности легкой и тяжелой цепей тоцилизумаба содержат SEQ ID NO. 187 и 188 соответственно.
В данном документе термин «Fc-область» используется для определения С-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере часть константной области. Этот термин включает Fc-области с нативной последовательностью и вариантные Fc-области. В одном варианте осуществления Fc-область тяжелой цепи человеческого IgG простирается от Cys226 или от Pro230 до карбокси-конца тяжелой цепи. При этом С-концевой лизин (Lys447) Fc-области может присутствовать или может не присутствовать. Если в данном документе не указано иное, нумерация аминокислотных остатков в Fc-области или константной области соответствует системе нумерации EU, также называемой EU-индексом, как это описано в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
«Каркасная область» или «FR» относится к остаткам вариабельного домена, отличным от остатков гипервариабельной области (HVR). FR вариабельного домена обычно состоит из четырех доменов FR: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR обычно находятся в VH (или VL) в следующем порядке: FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Термины «полноразмерное антитело», «интактное антитело» и «полное антитело» используются взаимозаменяемо в данном документе для обозначения антитела, имеющего структуру, по сути, сходную со структурой нативного антитела, или имеющего тяжелые цепи, которые содержат Fc-область, как определено в данном документе.
Термины «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используются взаимозаменяемо и относятся к клеткам, в которые была внесена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают «трансформантов» и «трансформированные клетки», которые включают первично трансформированные клетки и полученное от них потомство вне зависимости от числа пассажей. Потомство может не быть полностью идентичным исходной клетке по содержанию нуклеиновой кислоты и может содержать мутации. В данный документ включено мутантное потомство, которое имеет такую же функцию или биологическую активность, в отношении которых проводится скрининг или отбор изначально трансформированных клеток.
«Человеческое антитело» представляет собой антитело, которое имеет аминокислотную последовательность, соответствующую последовательности антитела, вырабатываемого человеком или клеткой человека, или полученного из источника, отличного от человека, в котором используются репертуары человеческих антител или другие последовательности, кодирующие человеческие антитела. Это определение человеческого антитела явным образом исключает гуманизированное антитело, содержащее отличные от человеческих антигенсвязывающие остатки.
«Человеческая консенсусная каркасная область» представляет собой каркасную область, которая представляет наиболее часто встречающиеся аминокислотные остатки в наборе каркасных последовательностей VL или VH человеческого иммуноглобулина. Обычно набор последовательностей VL или VH человеческого иммуноглобулина получают из подгруппы последовательностей вариабельных доменов. В общем случае подгруппа последовательностей представляет собой подгруппу согласно Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), тома. 1-3. В одном варианте осуществления в случае VL подгруппа представляет собой подгруппу каппа I согласно Kabat et al., см. выше. В одном варианте осуществления в случае VH подгруппа представляет собой подгруппу III согласно Kabat et al., см. выше.
«Гуманизированное» антитело относится к химерному антителу, содержащему аминокислотные остатки из отличных от человеческих HVR и аминокислотные остатки человеческих FR. В определенных вариантах осуществления гуманизированное антитело содержит, по сути, все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или, по сути, все HVR (например, CDR) соответствуют таковым из нечеловеческого антитела, а все или практически все FR соответствуют таковым из человеческого антитела. Гуманизированное антитело необязательно может содержать по меньшей мере часть константной области антитела, полученную из человеческого антитела. «Гуманизированная форма» антитела, например, нечеловеческого антитела, относится к антителу, подвергшемуся гуманизации.
Термин «гипервариабельная область» или «HVR», используемый в данном документе, относится к каждой из областей вариабельного домена антитела, которые являются гипервариабельными по последовательности («определяющие комплементарность области» или «CDR») и/или образуют структурно определенные петли («гипервариабельные петли»), и/или содержат контактирующие с антигенами остатки («антигенные контакты»). В общем случае антитела содержат шесть HVR (например, CDR): три в VH (H1, Н2, Н3) и три в VL (L1, L2, L3). Иллюстративные HVR в данном документе включают:
(а) гипервариабельные петли, находящиеся в аминокислотных остатках 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) (Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987));
(б) CDR, находящиеся в аминокислотных остатках 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2) и 95-102 (Н3) (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991));
(в) антигенные контакты, находящиеся в аминокислотных остатках 27 с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (Н2) и 93-101 (Н3) (MacCallum et al. J. Mol. Biol. 262: 732-745 (1996)); и
(г) комбинации из (а), (б) и/или (в), включающие аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (H1), 26-35b (H1), 49-65 (Н2), 93-102 (Н3) и 94-102 (Н3).
В одном варианте осуществления остатки HVR включают идентифицированные на Фиг. 12-16 и 18-24 или в другом месте описания.
Если не указано иное, остатки HVR и другие остатки в вариабельном домене (например, остатки FR) нумеруются в данном документе в соответствии с Kabat et al., см. выше.
«Иммуноконъюгат» представляет собой антитело, конъюгированное с одной или более гетерологичной(ыми) молекулами(ой), включая без ограничения цитотоксическое средство.
«Индивидуум» или «субъект» представляет собой млекопитающее. Млекопитающие включают без ограничения одомашненных животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и отличных от человека приматов, таких как обезьяны), кроликов и грызунов (например, мышей и крыс). В определенных вариантах осуществления индивидуум или субъект представляет собой человека.
«Выделенное» антитело представляет собой антитело, которое было отделено от компонента его естественного окружения. В некоторых вариантах осуществления антитело является очищенным до более чем 95% или 99% чистоты по определению, например, электрофоретическими (например, SDS-PAGE, изоэлектрическое фокусирование (IEF), капиллярный электрофорез) или хроматографическими (например, ионообменная или обращенно-фазовая ВЭЖХ) методами. Обзор методов оценки чистоты антител см., например, в Flatman et al., J. Chromatogr. В 848:79-87 (2007).
«Выделенная» нуклеиновая кислота относится к молекуле нуклеиновой кислоты, которая была отделена от компонента ее естественного окружения. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно содержат эту молекулу нуклеиновой кислоты, но при этом молекула нуклеиновой кислоты присутствует внехромосомно или в хромосомном положении, которое отличается от ее природного хромосомного положения.
«Выделенная нуклеиновая кислота, кодирующая селективное в отношении изоформы антитело к TGFβ» относится к одной или более молекулам нуклеиновых кислот, кодирующим тяжелые и легкие цепи антитела (или их фрагменты), включая такую(ие) молекулу(ы) нуклеиновых кислот в одном векторе или отдельных векторах и такую(ие) молекулу(ы) нуклеиновых кислот, присутствующую(ие) в одной или более местах в клетке-хозяине.
Термин «моноклональное антитело», используемый в данном документе, относится к антителу, полученному из популяции по сути гомогенных антител, т.е. отдельные антитела, составляющие популяцию, идентичны и/или связываются с одним и тем же эпитопом, за исключением возможных вариантных антител, например, содержащих мутации, имеющие природное происхождение или возникающие во время получения препарата моноклональных антител, при этом такие варианты в целом присутствуют в незначительных количествах. В отличие от препаратов поликлональных антител, которые обычно содержат разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело препарата моноклональных антител направлено против одной детерминанты на антигене. Таким образом, определение «моноклональный» указывает на характеристику антитела, как полученного из, по сути, гомогенной популяции антител, и его не следует интерпретировать как требование получения антитела каким-либо конкретным способом. Например, моноклональные антитела для применения в соответствии с настоящим изобретением могут быть получены с помощью различных методик, включая без ограничения метод гибридомы, методы рекомбинантной ДНК, методы фагового дисплея и методы с использованием трансгенных животных, содержащих все или часть локусов человеческого иммуноглобулина, причем такие методы и другие типичные методы получения моноклональных антител описаны в данном документе.
«Голое антитело» означает антитело, которое не конъюгировано с гетерологическим компонентом (например, цитотоксическим компонентом) или радиометкой. Голое антитело может присутствовать в фармацевтическом составе.
«Нативные антитела» относятся к молекулам иммуноглобулинов природного происхождения с различной структурой. Например, нативные антитела на основе IgG представляют собой гетеротетрамерные гликопротеины массой приблизительно 150000 дальтон, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, которые связаны дисульфидной связью. В направлении от N-конца к С-концу каждая тяжелая цепь имеет вариабельную область (VH), также называемую вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, за которой следуют три константных домена (CH1, СН2 и СН3). Аналогично, в направлении от N-конца к С-концу каждая легкая цепь имеет вариабельную область (VL), также называемую вариабельным легким доменом или вариабельным доменом легкой цепи, за которой следует константный домен легкой цепи (CL). Легкая цепь антитела может быть отнесена к одному из двух типов, называемых каппа (κ) и лямбда (λ), на основании аминокислотной последовательности ее константного домена.
Термин «вкладыш в упаковку» используется для обозначения инструкций, обычно включаемых в коммерческие упаковки терапевтических продуктов, которые содержат информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предостережениях относительно применения таких терапевтических продуктов.
«Процент (%) идентичности аминокислотной последовательности» относительно эталонной полипептидной последовательности определяется как процентная доля аминокислотных остатков в кандидатной последовательности, которые являются идентичными с аминокислотными остатками в эталонной полипептидной последовательности, после выравнивания последовательностей и внесения, в случае необходимости, гэпов для достижения максимального процента идентичности последовательностей, и без учета каких-либо консервативных замен в качестве части идентичности последовательностей. Выравнивание в целях определения процента идентичности аминокислотной последовательности можно осуществлять различными способами, которые известны в данной области техники, например, используя общедоступное компьютерное программное обеспечение, такое как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области техники могут определить подходящие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. При этом в целях данного документа значения % идентичности аминокислотных последовательностей получают, используя компьютерную программу для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 была разработана компанией Genentech, Inc., а исходный код был подан вместе с пользовательской документацией в Бюро регистрации авторских прав США, Вашингтон, округ Колумбия, 20559, где он зарегистрирован под номером регистрации авторского права США TXU510087. Программа ALIGN-2 находится в свободном доступе от Genentech, Inc., Южный Сан-Франциско, Калифорния, или может быть скомпилирована из исходного кода. Программу ALIGN-2 следует компилировать для использования в операционной системе UNIX, включая Digital UNIX V4.0D. Все параметры сравнения последовательностей установлены программой ALIGN-2 и не варьируются.
В тех случаях, когда для сравнения аминокислотных последовательностей используют ALIGN-2, % идентичности аминокислотной последовательности заданной аминокислотной последовательности А с или по сравнению с заданной аминокислотной последовательностью В (что в альтернативном варианте можно сформулировать как заданная аминокислотная последовательность А, которая имеет или содержит определенный % идентичности аминокислотной последовательности с или по сравнению с заданной аминокислотной последовательностью В) рассчитывают следующим образом:
100 умножить на соотношение X/Y,
где X представляет собой число аминокислотных остатков, оцененных программой выравнивания последовательностей ALIGN-2 как идентичные совпадения при программном выравнивании А и В, и где Y представляет общее количество аминокислотных остатков в В. Следует понимать, что там, где длина аминокислотной последовательности А не равняется длине аминокислотной последовательности В, % идентичности аминокислотной последовательности А к В не будет равен % идентичности аминокислотной последовательности В к А. Если специально не указано иное, все значения % идентичности аминокислотной последовательности, которые используется в данном документе, получают, как описано в предыдущем абзаце с использованием компьютерной программы ALIGN-2.
Термин «фармацевтический состав» относится к препарату, который находится в форме, обеспечивающей биологическую активность содержащегося в нем активного ингредиента, и который при этом не содержит дополнительные компоненты, являющиеся неприемлемо токсичными для субъекта, которому будет введен состав.
«Фармацевтически приемлемый носитель» относится к ингредиенту в фармацевтическом составе, отличному от активного ингредиента, который нетоксичен для субъекта. Фармацевтически приемлемый носитель включает без ограничения буфер, вспомогательное вещество, стабилизатор или консервант.
Используемый в данном документе термин «лечение» (и его грамматические варианты, такие как «лечить» или «лечащий») относится к клиническому вмешательству с целью изменения естественного течения заболевания индивидуума, подлежащего лечению, и может проводиться как для профилактики, так и в течении клинической патологии. Необходимые эффекты лечения включают без ограничения предупреждение появления или повторного появления заболевания, смягчение симптомов, уменьшение каких-либо прямых или косвенных патологических последствий заболевания, предупреждение метастазирования, снижение скорости прогрессирования заболевания, уменьшение интенсивности или временное облегчение болезненного состояния и ремиссию или улучшение прогноза. В некоторых вариантах осуществления антитела по настоящему изобретению используют для задержки развития заболевания или замедления прогрессирования заболевания.
Термин «вариабельная область» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL соответственно) нативного антитела обычно имеют сходные структуры, причем каждый домен содержит четыре консервативные каркасные области (FR) и три гипервариабельные области (HVR). (См., например, Kindt et al. Kuby Immunology, 6th ed., W.H. Freeman and Co., page 91 (2007). Одного домена VH или VL может быть достаточно для придания антигенсвязывающей специфичности. Кроме того, антитела, которые связывают конкретный антиген, можно выделять, используя домен VH или VL из антитела, которое связывает антиген, для скрининга библиотеки комплементарных доменов VL или VH, соответственно. См., например, Portolano et al., J. Immunol. 150:880-887 (1993); Clarkson et al., Nature 352:624-628 (1991).
Термин «вектор», используемый в данном документе, относится к молекуле нуклеиновой кислоты, способной увеличивать число копий другой нуклеиновой кислоты, с которой она связана. Этот термин включает вектор в виде самореплицирующейся структуры нуклеиновой кислоты, а также вектор, включенный в геном клетки-хозяина, в которую его внесли. Некоторые векторы способны управлять экспрессией нуклеиновых кислот, с которыми они функционально связаны. Такие векторы называются в данном документе «векторами экспрессии».
При использовании в данном документе «и/или» следует понимать как конкретное раскрытие каждой из двух указанных функций или компонентов с другими или без них. Например, «А и/или Б» следует понимать как конкретное раскрытие каждого из (i) A, (ii) Б и (iii) А и Б, точно так же, как если бы каждый из них был изложен в данном документе отдельно.
II. КОМПОЗИЦИИ И СПОСОБЫ
В одном аспекте настоящее изобретение частично основано на получении селективных в отношении изоформы антител к TGFβ (например, моноспецифических антител к TGFβ2 и TGFβ3 и антител с двойной специфичностью к TGFβ2/3) и способов их применения. TGFβ участвует в контроле нескольких ключевых клеточных функций, включая клеточную пролиферацию, дифференцировку, миграцию, апоптоз и продуцирование внеклеточного матрикса. Как следствие, фактор роста влияет на многие биологические процессы, включая эмбриональное развитие, заживление ран, иммунную функцию, злокачественную трансформацию и старение. Таким образом, селективные в отношении изоформы антитела к TGFβ по настоящему изобретению можно применять, например, для диагностики или лечения нарушений, связанных с TGFβ, таких как без ограничения фиброзное заболевание и рак.
В некоторых аспектах представлены эпитопы, связываемые селективными в отношении изоформы антителами к TGFβ. Антигенсвязывающие домены селективных в отношении изоформы антител к TGFβ определяли на основании их кристаллической структуры. В качестве примера, связывающий эпитоп антител 2А10 и 4А11 картировали путем определения кристаллических структур их комплекса антитело/TGFβ. См. пример 10 ниже. Как будет понятно специалисту в данной области техники, результаты из примера 10 демонстрируют, где антитело 4А11 к TGFβ2/3 взаимодействует с TGFβ2 (и, следовательно, где оно также связывается в той же высококонсервативной области в TGFβ3), и где антитело 2А10 к TGFβ3 взаимодействует с TGFβ3. Таким образом, антитела, которые взаимодействуют с любым из этих остатков в TGFJ32 или TGFβ3 или блокируют их, могут быть использованы в качестве антител, нейтрализующих TGFβ2 или TGFβ3 соответственно. В некоторых вариантах осуществления предполагается, что антитела, которые при связывании с изоформой(ами)-мишенью(ями) TGFβ взаимодействуют с остатками изоформы TGFβ или блокируют их, или находятся в пределах 15-8, 8, 8-7, 8-6, 8-5 или 5 ангстрем от этих остатков, обеспечивают полезную нейтрализацию изоформ(ы) TGFβ. В качестве неограничивающего примера было определено, что антитело 2А10 к TGFβ3 связывается с эпитопом на TGFβ3, содержащим аминокислотные остатки R325, K331, W332, Н334, Е335, Т387, 1388, L389, Y391, V392, G393, R394, Р396, K397 и V398 на TGFβ3 человека (т.е. антигенсвязывающий домен непосредственно связывался с этими остатками на TGFβ3), и это связывание приводило к нейтрализации TGFβ3. Таким образом, в некоторых вариантах осуществления предполагается, что антитела, которые при связывании с TGFβ3 взаимодействуют с этими остатками на TGFβ3 или блокируют их, находятся в пределах 15-8, 8, 8-5 или предпочтительно в пределах 5 ангстрем от этих остатков, обеспечивают полезную нейтрализацию TGFβ3. В качестве дополнительного неограничивающего примера было определено, что антитело 4А11 к TGFβ2/3 связывается с гомодимером TGFβ2 и непосредственно контактирует с аминокислотными остатками V313, Q314, D315, R320, L322, Y323, R328, D329, F345 и А347 в первом мономере TGFβ2 гомодимера, и аминокислотными остатками N368, Т369, 1370, N371, Р372, Е373, А374, S375, А376 и S377 во втором мономере TGFβ2. Таким образом, в некоторых вариантах осуществления предполагается, что антитела, которые при связывании с TGFβ2 взаимодействуют с этими остатками на TGFβ2 или блокируют их, находятся в пределах 15-8, 8, 8-5 или предпочтительно в пределах 5 ангстрем от этих остатков, обеспечивают полезную нейтрализацию TGFβ2. В некоторых вариантах осуществления антигенсвязывающий домен связывается в пределах 30, 30-25, 25-20, 20-15, 15-8, 8, 8-5, 5, 5-4, 4 или менее ангстрем от одного или нескольких из вышеуказанных остатков. В некоторых вариантах осуществления антигенсвязывающий домен при связывании с изоформой TGFβ находится в пределах по меньшей мере одного из указанных выше расстояний для более чем одного из вышеуказанных остатков. Например, в некоторых вариантах осуществления антигенсвязывающий домен находится в пределах одного из указанных расстояний (например, 30, 30-25, 25-20, 20-15, 15-8, 8, 8-5, 5, 5-4, 4 или меньше) для по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 20-25, 25-30, 30-35, 35-40, 40-45, 45-50, 50-55, 55-60, 60-65, 65-70, 70-75 или более вышеуказанных остатков. В некоторых вариантах осуществления антигенсвязывающий домен находится в пределах одного из указанных расстояний для по меньшей мере 1-10, 10-20, 20-30, 30-40, 40-50, 50-60, 60-70, 70-80, 80-90, 90-95, 95-99, 99-100% остатков, идентифицированных в каждой группе или их подгруппе (например, только поверхностных остатков в группе). Если специально не указано иное, расстояние между антигенсвязывающим доменом и изоформой TGFβ представляет собой кратчайшее расстояние между ковалентно связанным атомом изоформы TGFβ и ковалентно связанным атомом антигенсвязывающего домена, которые являются ближайшими атомами изоформы TGFβ и антигенсвязывающего домена. Аналогичным образом, если специально не указано иное, расстояние между остатком в антигенсвязывающем домене и изоформой TGFβ, для которой он является специфическим, представляет собой расстояние от ближайшей точки на идентифицированном остатке до ближайшей ковалентно связанной части изоформы TGFβ, или наоборот. В некоторых вариантах осуществления расстояние можно измерять от остова аминокислотных цепей. В некоторых вариантах осуществления можно измерить расстояние между краем паратопа и краем (ближайшим друг к другу) эпитопа. В некоторых вариантах осуществления расстояние можно измерить между центром поверхности паратопа и центром поверхности эпитопа. Как будет понятно специалисту в данной области техники, настоящее описание применимо для каждого из индивидуальных наборов остатков, перечисленных в данном документе. Например, вышеуказанные диапазоны рассматриваются в целом и конкретно для остатков эпитопа и паратопа, перечисленных в примере 10.
А. Иллюстративные селективные в отношении изоформы антитела к TGFβ
Антитела к TGFβ2
В одном аспекте в настоящем изобретении представлены выделенные антитела, которые связываются с TGFβ2. В определенных вариантах осуществления антитело к TGFβ2 селективно нейтрализует TGFβ2. В определенных вариантах осуществления антитело к TGFβ2 имеет одно или более из следующих свойств: (а) селективно нейтрализует TGFβ2; (б) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ; (в) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ; (г) имеет сниженную токсичность у грызунов по сравнению с антителом 1D11 ко всем видам TGFβ; (д) имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; и/или (е) имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом.
В определенных аспектах антитело к TGFβ2, представленное в данном документе, связывается с TGFβ2 с KD, составляющей менее 10 пМ, и/или клеточной IC50, составляющей менее 250 пМ. В одном аспекте антитело к TGFβ2, представленное в данном документе, связывается с TGFβ2 с KD, составляющей менее или равную приблизительно 5 пМ, приблизительно 4 пМ, приблизительно 3 пМ, приблизительно 2 пМ или приблизительно 1 пМ. В одном аспекте антитело к TGFβ2, представленное в данном документе, связывается с TGFβ2 с KD, составляющей менее 1 пМ. В одном аспекте антитело к TGFβ2, представленное в данном документе, имеет клеточную IC50, составляющую менее или равную приблизительно 250 пМ, приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ или приблизительно 50 пМ. В одном аспекте антитело к TGFβ2, представленное в данном документе, имеет клеточную IC50 для ингибирования (нейтрализации) TGFβ2, составляющую 40 пМ.
В одном аспекте в настоящем изобретении представлено антитело к TGFβ2, содержащее по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 17; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 19; (д) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 20; и (е) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21.
В одном аспекте в настоящем изобретении представлено антитело к TGFβ2, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 17; и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 18. В другом аспекте в настоящем изобретении представлено антитело, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17; и (в) HVR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 18.
В другом аспекте в настоящем изобретении представлено антитело к TGFJ32, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 19; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 20; и (в) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21. В одном варианте осуществления антитело содержит (a) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 19; (б) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 20; и (в) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21.
В другом варианте осуществления антитело к TGFβ2 содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18, и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21. В дополнительном варианте осуществления антитело к TGFβ2 содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18, HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17.
В другом аспекте антитело к TGFβ2 по настоящему изобретению содержит (а) домен VH, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 16, (ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 17, и (iii) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 18; и (б) домен VL, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VL, выбранные из (i) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 19, (ii) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 20, и (iii) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21.
В другом аспекте в настоящем изобретении представлено антитело к TGFβ2, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 19; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 20; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21.
В любом из вышеуказанных вариантов осуществления антитело к TGFβ2 является гуманизированным. В одном варианте осуществления антитело к TGFβ2 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область. В любом из вышеуказанных вариантов осуществления гуманизированное антитело к TGFβ2 содержит одну или более мутаций в каркасной области VH, выбранных из группы, состоящей из 37V или 37I, 48М или 48L, 49G или 49А, 67L, 71K и 78V, и 105Р или 105R. В некоторых вариантах осуществления антитело к TGFβ2 содержит VH, где VH содержит аминокислотную последовательность SEQ ID NO: 25, при этом VH содержит набор мутаций каркасной области, выбранный из группы, состоящей из: (i) 37V, 48М и 49G в FR2 и 105Р в FR4 (h6F12.v1 и h6F12.v3); (ii) 37V и 48М в FR2, 67L, 71K и 78V в FR3 и 105Р в FR4 (h6F12.v2 и h6F12.v4); (iii) 37I в FR2 (h6F12.v1.6); (iv) 48L в FR2 (h6F12.v1.7); (v) 49A в FR2 (h6F12.v1.8); (vi) 105R в FR4 (h6F12.v1.9); (vii) 37V, 48M и 49G в FR2 и 105P в FR4 (h6F12.v1 и h6F12.v3); (viii) 37V и 48M в FR2, 67L, 71K и 78V в FR3 и 105Р в FR4 (6F12.V2 и h6F12.v4); (ix) 37I в FR2 (h6F12.v1.6); (х) 48L в FR2 (h6F12.v1.7); (xi) 49А в FR2 (h6F12.v1.8); и (xii) 105R в FR4 (h6F12.v1.9). В любом из вышеуказанных вариантов осуществления гуманизированное антитело к TGFβ2 содержит одну или более мутаций в каркасной области VL, выбранных из группы, состоящей из 43 S или 43A, 66G, 69Т, 71F и 87Y. В некоторых вариантах осуществления антитело к TGFβ2 содержит VL, где VL содержит аминокислотную последовательность SEQ ID NO: 24, при этом VL содержит набор мутаций каркасной области, выбранный из группы, состоящей из: (i) 43S в FR2 и 66Е, 69Р, 71Y и 87F в FR3 (h6F12.v1 и h6F12.v2); (ii) 43S в FR2 и 58V, 66Е, 69Р, 71Y и 87F в FR3 (h6F12.v3 и h6F12.v4); (iii) 43А в FR2 (h6F12.v1. 1); (iv) 66G в FR3 (h6F12.v1.2); (v) 69T в FR3 (h6F12.v1.3); (vi) 71F в FR3 (h6F12.v1.4); и (vii) 87Y в FR3 (h6F12.v1.5).
В другом аспекте антитело к TGFβ2 содержит последовательность вариабельного домена тяжелой цепи (VH), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 132 и 138-142. В определенных вариантах осуществления последовательность VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности, содержит замены (например, консервативные замены), вставки или делеции по отношению к эталонной последовательности, но селективное в отношении изоформы антитело к TGFβ, содержащее эту последовательность, сохраняет способность связываться с TGFβ2 и нейтрализовать его. В определенных вариантах осуществления в общей сложности 1-10 аминокислот были замещены, вставлены и/или делетированы в SEQ ID NO: 25. В определенных вариантах осуществления замены, вставки или делеции находятся в областях за пределами HVR (т.е. в FR). Необязательно, антитело к TGFβ2 содержит последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 25, 132 и 138-142, включая посттрансляционные модификации этой последовательности. В конкретном варианте осуществления VH содержит одну, две или три HVR, выбранные из: (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 16, (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 17, (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 18.
В другом аспекте представлено антитело к TGFβ2, где антитело содержит последовательность вариабельного домена легкой цепи (VL), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 24, 131, 133-137, 143 и 144. В определенных вариантах осуществления последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности, содержит замены (например, консервативные замены), вставки или делеции по отношению к эталонной последовательности, но антитело к TGFβ2, содержащее эту последовательность, сохраняет способность связываться с PRO. В определенных вариантах осуществления в общей сложности 1-10 аминокислот были замещены, вставлены и/или делетированы в SEQ ID NO: 24. В определенных вариантах осуществления замены, вставки или делеции встречаются в областях за пределами HVR (т.е. в FR). Необязательно, антитело к TGFβ2 содержит последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 24, 131, 133-137, 143 и 144, включая посттрансляционные модификации этой последовательности. В конкретном варианте осуществления VL содержит одну, две или три HVR, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 19; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 20; и (в) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21.
В другом аспекте представлено антитело к TGFβ2, где антитело содержит VH, как в любом из вышеуказанных вариантов осуществления, представленных выше, и VL, как в любом из вышеуказанных вариантов осуществления, представленных выше. В одном варианте осуществления антитело содержит последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 25/24 (6F12 кролика), SEQ ID NO: 132/131 (v1), SEQ ID NO: 132/133 (v1.1), SEQ ID NO: 132/134 (v1.2), SEQ ID NO: 132/135 (v1.3), SEQ ID NO: 132/136 (v1.4), SEQ ID NO: 132/137 (v1.5), SEQ ID NO: 138/131 (v1.6), SEQ ID NO: 139/131 (v1.7), SEQ ID NO: 140/131 (v1.8), SEQ ID NO: 141/131 (v1.9), SEQ ID NO: 142/131 (v2), SEQ ID NO: 132/143 (v3) и SEQ ID NO: 142/144 (v4), включая посттрансляционные модификации этих последовательностей.
В другом аспекте представлено антитело к TGFβ2, где антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 31, 146 и 152-156, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 30, 145, 147-151, 157 и 158. В некоторых вариантах осуществления антитело к TGFβ2 содержит полную пару H/L-цепей, при этом полная пара H/L-цепей (соответственно) содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 31/30 (6F12 кролика), SEQ ID NO: 146/145 (v1), SEQ ID NO: 146/147 (v1.1), SEQ ID NO: 146/148 (v1.2), SEQ ID NO: 146/149 (v1.3), SEQ ID NO: 146/150 (v1.4), SEQ ID NO: 146/151 (v1.5), SEQ ID NO: 152/145 (v1.6), SEQ ID NO: 153/145 (v1.7), SEQ ID NO: 154/145 (v1.8), SEQ ID NO: 155/145 (v1.9), SEQ ID NO: 156/145 (v2), SEQ ID NO: 146/157 (v3) и SEQ ID NO: 156/158 (v4).
В дополнительном аспекте в настоящем изобретении представлено антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ2, представленное в данном документе. Например, в определенных вариантах осуществления представлено антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ2, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 19; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 20; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21.
Антитела к TGFβ2/3
В другом аспекте в настоящем изобретении представлены выделенные антитела, которые связываются как с TGFβ2, так и с TGFβ3 (антитела к TGFβ2/3). В определенных аспектах антитело селективно нейтрализует TGFβ2 и TGFβ3 и характеризуется одним или более из следующих свойств:
(а) селективность антитела к TGFβ2/3 в отношении TGFβ2 и TGFβ3 по сравнению с TGFβ1 человека в отношении селективной нейтрализации достигается посредством прямого контакта антигенсвязывающего домена антитела с аминокислотным остатком Е373 TGFβ2 или TGFβ3 (нумерация TGFβ2 человека);
(б) антитело к TGFβ2/3 нейтрализует TGFβ2 и/или TGFβ3 посредством аллостерического механизма;
(в) антитело к TGFβ2/3 индуцирует конформационные изменения в гомодимере TGFβ2;
(г) антитело к TGFβ2/3 индуцирует конформационное изменение в гомодимере TGFβ2, при этом конформационное изменение включает совместное сжатие двух мономеров на несколько градусов;
(д) антитело к TGFβ2/3 представляет собой двухвалентное антитело или одновалентное антитело;
(е) антитело к TGFβ2/3 содержит (i) CDR тяжелой цепи, содержащую CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащую CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 15;
(ж) антитело к TGFβ2/3 связывается с гомодимером TGFβ2, при этом гомодимер TGFβ2 имеет первый и второй мономер TGFβ2, и при этом антитело к TGFβ2/3 содержит антигенсвязывающий домен, который непосредственно контактирует с (i) аминокислотными остатками V313, Q314, D315, R320, L322, Y323, R328, D329, F345 и А347 первого мономера TGFβ2, и (ii) аминокислотными остатками N368, Т369, 1370, N371, Р372, Е373, А374, S375, А376 и S377 второго мономера TGFβ2 (нумерация TGFβ2 человека);
(и) антитело к TGFβ2/3, как в (ж), где антигенсвязывающий домен находится в пределах 5 ангстрем от аминокислотных остатков TGFβ2 и/или TGFβ3;
(к) при этом антитело к TGFβ2/3 связывается с эпитопом, по сути сходным с эпитопом, как в (ж) в TGFβ3; и
(л) антитело к TGFβ2/3 не нейтрализует TGFβ2 и/или TGFβ3 в форме с одним плечом.
В определенных вариантах осуществления антитело к TGFβ2/3 селективно нейтрализует TGFβ2 и TGFβ3. В определенных аспектах антитело к TGFβ2/3, представленное в данном документе, связывается с TGFβ2/3 с KD, составляющей менее 10 пМ, и/или клеточной IC50, составляющей менее 250 пМ. В одном аспекте антитело к TGFβ2/3, представленное в данном документе, связывается с TGFβ2 и/или TGFβ3 с KD, составляющей менее приблизительно 10 пМ, приблизительно 9 пМ, приблизительно 8 пМ, приблизительно 7 пМ, приблизительно 6 пМ, приблизительно 5 пМ, приблизительно 4 пМ, приблизительно 3 пМ, приблизительно 2 пМ или приблизительно 1 пМ. В одном аспекте антитело к TGFβ2/3, представленное в данном документе, связывается с TGFβ2 с KD, составляющей приблизительно 5 пМ. В одном аспекте антитело к TGFβ2/3, представленное в данном документе, имеет клеточную 1С50 для ингибирования TGFβ2, составляющую приблизительно 250 пМ, приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ или приблизительно 50 пМ. В одном аспекте антитело к TGFβ2/3, представленное в данном документе, имеет клеточную IC50 для ингибирования TGFβ3, составляющую приблизительно 250 пМ, приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ, приблизительно 50 пМ, приблизительно 40 пМ, приблизительно 30 пМ или менее чем приблизительно 30 пМ. В одном аспекте антитело к TGFβ2/3, представленное в данном документе, имеет клеточную IC50 для ингибирования TGFβ2, составляющую приблизительно 250 пМ, и/или клеточную IC50 для ингибирования (нейтрализации) TGFβ3, составляющую приблизительно 30 пМ.
В одном аспекте в настоящем изобретении представлено антитело к TGFβ2/3, содержащее по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 11; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 12; (г) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 13; (д) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 14; и (е) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 15.
В одном аспекте в настоящем изобретении представлено антитело к TGFβ2/3, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащей аминокислотную последовательность антитела к TGFβ2/3 SEQ ID NO: 11; и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 12. В одном варианте осуществления антитело содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 12. В другом варианте осуществления антитело содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 12, и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 15. В дополнительном варианте осуществления антитело содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 12, HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 15, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 11. В дополнительном варианте осуществления антитело содержит (а) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 11; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 12.
В другом аспекте в настоящем изобретении представлено антитело к TGFP2/3, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 13; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 14; и (в) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 15. В одном варианте осуществления антитело к TGFβ2/3 содержит (a) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 13; (б) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 14; и (в) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 15.
В другом аспекте антитело к TGFβ2/3 по настоящему изобретению содержит (а) домен VH, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 10, (ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 11, и (iii) HVR-Н3, содержащей аминокислотную последовательность SEQ ID NO: 12; и (б) домен VL, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VL, выбранные из (i) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 13, (ii) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 14, и (iii) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 15.
В другом аспекте в настоящем изобретении представлено антитело к TGFβ2/3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 11; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 12; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 13; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 14; и (e) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 15.
В любом из вышеуказанных вариантов осуществления антитело к TGFβ2/3 является гуманизированным. В одном варианте осуществления антитело к TGFβ2/3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область. В другом варианте осуществления антитело к TGFP2/3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VH, содержащую модификации FR, выбранные из группы, состоящей из 1E, 2Q или 2V, 24V, 37V или 37I, 48I, 49G, 67F или 67V, 71K или 71V, 73S или 73Т, делеции 75K и 76N, 78V или 78F, 91F или 91Y, 105Р или 105Q. В некоторых вариантах осуществления антитело к TGFβ2/3 содержит VH, где VH содержит аминокислотную последовательность SEQ ID NO: 27, при этом VH содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 2Q и 24V в FR1, 48I и 49G в FR2, 71K, 73S, 78V и 91F в FR3 и 105Р в FR4 (h4A11.v1, h4A11.v2, h4A11.v5, h4A11.v6); (ii) 2Q в FR1, 37V в FR2, 67F, 71K, 73S, 78V и 91F в FR3 и 105P в FR4 (h4A11.v3, h4A11.v4, 4A11.v7, h4A11.v8); (iii) делеции 1E в FR1 (h4A11.v7.1); (iv) делеции 75K и 76N в FR3 (h4A11.v7.2); (v) делеции 1E в FR1 и 75K76N в FR3 (h4A11.v7.3); (vi) 2V в FR1 (h4A11.v7.8); (vi) 37I в FR2 (h4A11.v7.9); (vii) 67V в FR3 (h4A11.v7.10); (viii) 71V в FR3 (h4A11.v7.11); (ix) 73T в FR3 (h4A11.v7.12); (x) 78F в FR3 (h4A11.v7.13); (xi) 91Y в FR3 (h4A11.v7.14); (xii) 105Q в FR4 (h4A11.v7.15); (xiii) 2V в FR1, 37I в FR2, 67V, 73T, 78F в FR3, 105Q в FR4 (h4A11.v7.16); (xiv) 2V в FR1, 37I в FR2, 67V, 73T, 91Y в FR3, 105Q в FR4 (h4Al l.v7.17); (xv) 2V в FR1, 37I в FR2, 67V, 73T в FR3, 105Q в FR4 (h4A11.v7.18); и (xvi) 2V в FR1, 371 в FR2, 67V, 73T, делеции 75K и 76N в FR3, 105Q в FR4 (h4A11.v7.19). В другом варианте осуществления антитело к TGFβ2/3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VL, содержащую модификации FR, выбранные из группы, состоящей из: 2А или 2I, 4L, 36F или 36Y, 43Р или 43А и 58V или 58I. В некоторых вариантах осуществления антитело к TGFβ2/3 содержит VL, где VL содержит аминокислотную последовательность SEQ ID NO: 26, при этом VL содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 2А и 4L в FR1 и 36F в FR2 (h4A11.v1 и h4A11.v3); (ii) 2А и 4L в FR1 и 36F и 43Р в FR2 (h4A11.v2 и h4A11.v4); (iii) 2А в FR1, 36F и 43Р в FR2 и 58V в FR3 (h4A11.v5 и h4A11.v7); (iv) 2А и 4L в FR1 и 36F в FR2 (h4A11.v6 и h4A11.v8); (v) 21 в FR1 (h4A11.v7.4); (vi) 36Y в FR2 (h4A11.v7.5); (vii) 43A в FR2 (h4A11.v7.6); (viii) 581 в FR3 (h4A11.v7.7); и (ix) 2I в FR1, 43A в FR2, 58I в FR3 (h4A11.v7.16-19).
В другом аспекте антитело к TGFβ2/3 содержит последовательность вариабельного домена тяжелой цепи (VH), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 27, 81, 83, 86-88, 93-100 и 102-105. В определенных вариантах осуществления последовательность VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности, содержит замены (например, консервативные замены), вставки или делеции по отношению к эталонной последовательности, но антитело к TGFβ2/3, содержащее эту последовательность, сохраняет способность связываться с TGFβ2 и с TGFβ3. В определенных вариантах осуществления в общей сложности 1-10 аминокислот были замещены, вставлены и/или делетированы в SEQ ID NO: 27. В определенных вариантах осуществления замены, вставки или делеции находятся в областях за пределами HVR (т.е. в FR). Необязательно, антитело к TGFβ2/3 содержит последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 27, 81, 83, 86-88, 93-100 и 102-105, включая посттрансляционные модификации этой последовательности. В конкретном варианте осуществления VH содержит одну, две или три HVR, выбранные из: (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 10, (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 11, и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 12.
В другом аспекте представлено антитело к TGFβ2/3, где антитело к TGFβ2/3 содержит последовательность вариабельного домена легкой цепи (VL), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 26, 80, 82, 84, 85, 89-92 и 101. В определенных вариантах осуществления последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности, содержит замены (например, консервативные замены), вставки или делеции по отношению к эталонной последовательности, но антитело к TGFβ2/3, содержащее эту последовательность, сохраняет способность связываться с TGFβ2 и с TGFβ3. В определенных вариантах осуществления в общей сложности 1-10 аминокислот были замещены, вставлены и/или делетированы в SEQ ID NO: 26. В определенных вариантах осуществления замены, вставки или делеции встречаются в областях за пределами HVR (т.е. в FR). Необязательно, антитело к TGFβ2/3 содержит последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 26, 80, 82, 84, 85, 89-92 и 101, включая посттрансляционные модификации этой последовательности. В конкретном варианте осуществления VL содержит одну, две или три HVR, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 13; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 14; и (в) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 15.
В другом аспекте предложено антитело к TGFβ2/3, содержащее VH, как в любом из вышеуказанных вариантов осуществления, и VL, как в любом из вышеуказанных вариантов осуществления. В одном варианте осуществления антитело к TGFβ2/3 содержит последовательности VH и VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 27/26 (4А11 кролика), SEQ ID NO: 81/80 (v1), SEQ ID NO: 81/82 (v2), SEQ ID NO: 83/80 (v3), SEQ ID NO: 83/82 (v4), SEQ ID NO: 81/84 (v5), SEQ ID NO: 81/85 (v6), SEQ ID NO: 83/84 (v7), SEQ ID NO: 86/84 (v7/l), SEQ ID NO: 87/84 (v7.2), SEQ ID NO: 88/84 (v7.3), SEQ ID NO: 83/89 (v7.4), SEQ ID NO: 83/90 (v7.5), SEQ ID NO: 83/91 (v7.6), SEQ ID NO: 83/92 (v7.7), SEQ ID NO: 93/84 (v7.8), SEQ ID NO: 94/84 (v7.9), SEQ ID NO: 95/84 (v7.10), SEQ ID NO: 96/84 (v7.11), SEQ ID NO: 97/84 (v7.12), SEQ ID NO: 98/84 (v7.13), SEQ ID NO: 99/84 (v7.14), SEQ ID NO: 100/84 (v7.15), SEQ ID NO: 102/101 (v7.16), SEQ ID NO: 103/101 (v7.17), SEQ ID NO: 104/101 (v7.18), SEQ ID NO: 105/101 (v7.19), SEQ ID NO: 83/85 (v8), включая посттрансляционные модификации этих последовательностей.
В другом аспекте представлено антитело к TGFβ2/3, где антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 33, 107, 109, 112-114 и 119-130, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 32, 106, 108, 110, 111, 115-118 и 186. В некоторых вариантах осуществления антитело к TGFβ2/3 содержит полную пару H/L-цепей, при этом полная пара H/L-цепей (соответственно) содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 32/33 (4А11 кролика), SEQ ID NO: 107/106 (v1), SEQ ID NO: 107/108 (v2), SEQ ID NO: 109/106 (v3), SEQ ID NO: 109/108 (v4), SEQ ID NO: 107/110 (v5), SEQ ID NO: 107/111 (v6), SEQ ID NO: 109/110 (v7), SEQ ID NO: 112/110 (v7.1), SEQ ID NO: 113/110 (v7.2), SEQ ID NO: 114/110 (v7.3), SEQ ID NO: 114/115 (v7.4), SEQ ID NO: 114/116 (v7.5), SEQ ID NO: 114/117 (v7.6), SEQ ID NO: 114/118 (v7.7), SEQ ID NO: 119/110 (v7.8), SEQ ID NO: 120/110 (v7.9), SEQ ID NO: 121/110 (v7.10), SEQ ID NO: 122/110 (v7.11), SEQ ID NO: 123/110 (v7.12), SEQ ID NO: 124/110 (v7.13), SEQ ID NO: 125/110 (v7.14), SEQ ID NO: 126/110 (v7.15), SEQ ID NO: 127/186 (v7.16), SEQ ID NO: 128/186 (v7.17), SEQ ID NO: 129/186 (v7.18), SEQ ID NO: 130/186 (v7.19) и SEQ ID NO: 114/111 (v8).
В дополнительном аспекте в настоящем изобретении представлено антитело к TGFβ2/3, которое связывается с тем же эпитопом, что и антитело к TGFβ2/3, представленное в данном документе. Например, в определенных вариантах осуществления представлено антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ2/3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 11; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 12; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 13; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 14; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 15. В определенных вариантах осуществления представлено антитело к TGFβ2/3, которое связывается с эпитопом, охватывающим гомодимер TGFβ2, при этом гомодимер TGFβ2 имеет первый и второй мономер TGFβ2, при этом антитело к TGFβ2/3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками V313, Q314, D315, R320, L322, Y323, R328, D329, F345 и А347 первого мономера TGFβ2, и аминокислотными остатками N368, Т369, 1370, N371, Р372, Е373, А374, S375, А376 и S377 второго мономера TGFβ2, и при этом в некоторых вариантах осуществления антитело к TGFβ2/3 связывается с тем же эпитопом в TGFβ3. TGFβ2 и TGFβ3 являются высококонсервативными в области, содержащей положения аминокислот 368-377 (нумерация TGFβ2), таким образом, антитело к TGFβ2/3 связывается с той же областью как в TGFβ2, так и в TGFβ3.
Антитела к TGFβ3
В одном аспекте в настоящем изобретении представлены выделенные антитела, которые связываются с TGFβ3. В определенных вариантах осуществления антитело к TGFβ3 селективно нейтрализует TGFβ3. Кроме того, в настоящее время обнаружено, что антитела к TGFβ3, описанные в данном документе, имеют улучшенные профили безопасности (например, сниженную токсичность) по сравнению с ингибиторами всех видов TGFβ, а также по сравнению с селективными в отношении изоформы антителами, специфическими в отношении TGFβ2 (пример 2) и селективными антителами к TGFβ1. Несколько ингибиторов передачи сигналов TGFβ были исследованы в доклинических токсикологических исследованиях, включая как низкомолекулярные ингибиторы киназной активности TGFBR1 (ALK5), например, галунисертиб, так и основанные на антителах ингибиторы взаимодействий TGFβ-TGFBR, например, фрезолимумаб, антитело ко всем видам TGFβ. Токсикологические исследования низкомолекулярных ингибиторов ALK5 из разных химических серий на крысах последовательно показали геморрагические, воспалительные и дегенеративные поражения сердечного клапана и метафизарную дисплазию (Frazier Toxicol Pathol 35, 284-95. 2007; Anderton Toxicol Pathol 39: 916, 2011). В ранних клинических исследованиях галунисертиб вводили в дозе при уровнях, которые не полностью ингибируют активность ALK5, в течение относительно коротких периодов времени, а в клинической практике еще не было выявлено данных в отношении сердечной деятельности. У мышей, обработанных антителом 1D11 ко всем видам TGFβ (Lonning et al. (2011) Current Pharmaceutical Biotechnology, 12, 2176-2189), развились гистологические поражения, потеря веса, неопухолевая кистозная гиперплазия эпителия и воспаление языка, дисплазия зубов и гиперплазия эпителия десен и пищевода. Фрезолимумаб, гуманизированная форма антитела GC1008, которое связывается со всеми тремя изоформами TGFβ и ингибирует их активность с сопоставимыми аффинностями в отношении изоформ TGFβ, что и антитело 1D11, исследовали на яванских макаках, и он приводил к дозозависимым кровотечениям, анемии и гиперплазии мочевыводящих путей, эпителия носа и мочевого пузыря. У людей лечение фрезолимумабом приводило к анемии и кровотечениям (десневым, носовым и субконъюнктивальным), а также к увеличению частоты кератоакантом (предраковых плоскоклеточных поражений кожи), которые исчезали после прекращения лечения (Rice, JCI 125:2795 (2015); Lacouture, Cancer Immunol Immunother 64:437 (2015)). CAT-192, антитело, преимущественно селективное в отношении TGFβ1, имело высокую частоту серьезных нежелательных явлений с множественными желудочными кровотечениями, наблюдаемыми в исследовании фазы 1-2 при SSc (Denton A&R 56:323 (2007); см. также World Wide Web по адресу tripod.nih.gov/ginas/app/substance/4AR6718OL0). В совокупности эти наблюдения позволяют предположить, что кровотечение, поражение сердца и гиперплазия эпителия являются серьезной проблемой при длительном хроническом фармакологическом ингибировании TGFβ.
В отличие от проблем безопасности, которые наблюдались при предыдущих попытках нейтрализовать TGFβ in vivo, в определенных аспектах в настоящем изобретении представлены антитела к TGFβ3, имеющие улучшенные профили безопасности (например, сниженную токсичность). Например, в приведенном ниже примере 2 у мышей, обработанных антителом к TGFβ3 в дозах до 50 мг/кг три раза в неделю в общей сложности в течение 4 недель, побочные эффекты отсутствовали или были очень незначительными (метафизарная дисплазия при наиболее высокой дозе), и ни один из серьезных побочных эффектов, вызванных низкомолекулярными ингибиторами, антителами к TGFβ1 или ко всем видам TGFβ, как обсуждалось выше, не наблюдался. Также было обнаружено, что антитела к TGFβ3, описанные в данном документе, имеют улучшенные профили безопасности по сравнению с селективными в отношении изоформы антителами к TGFβ2/3 и к TGFβ2 (пример 2, см. таблицу 8).
В определенных аспектах представлено выделенное антитело к TGFβ3, где антитело селективно нейтрализует TGFβ3, и при этом антитело характеризуется одним или более из следующих свойств:
(а) антитело к TGFβ3 связывается со шпилечной областью бета6/бета7 TGFβ3;
(б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3;
(в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFBR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3;
(г) антитело к TGFβ3 непосредственно связывает аминокислотный остаток R394 в шпилечной области бета6/бета7 TGFP3 человека;
(д) антитело к TGFβ3 непосредственно связывает аминокислотный остаток R394 в шпилечной области бета6/бета7 TGFβ3 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи;
(е) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ1 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека);
(ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека);
(и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ;
(к) антитело к TGFβ3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 ко всем видам TGFβ;
(л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом CAT-192 к TGFβ1;
(п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3;
(р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9;
(с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, I388, L389, Y391, V392, G393, R394, Р396, K397 и V398 TGFβ3 человека; и
(т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3.
В определенных аспектах антитело к TGFβ3, представленное в данном документе, связывается с TGFβ3 с KD, составляющей менее 10 пМ, и/или имеет клеточную IC50, составляющую менее 250 пМ. В одном аспекте антитело к TGFβ3, представленное в данном документе, связывается с TGFβ3 с KD, составляющей менее приблизительно 5 пМ, приблизительно 4 пМ или приблизительно 3 пМ. В одном аспекте антитело к TGFβ3, представленное в данном документе, связывается с TGFβ3 с KD, составляющей менее приблизительно 2 пМ. В одном аспекте антитело к TGFβ3, представленное в данном документе, имеет клеточную IC50, составляющую менее приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ или приблизительно 50 пМ. В одном аспекте антитело к TGFβ3, представленное в данном документе, имеет клеточную IC50 для ингибирования (нейтрализации) TGFβ3, составляющую менее приблизительно 20 пМ.
В другом аспекте антитело к TGFβ3, представленное в данном документе, селективно нейтрализует TGFβ3 и имеет сниженную токсичность у мышей по сравнению с антителом 1D11 к TGFβ (см. Lonning et al. (2011)). В другом аспекте антитело к TGFJ33, представленное в данном документе, селективно нейтрализует TGFJ33 и имеет улучшенный профиль безопасности, например, по сравнению с ингибиторами всех видов TGFβ, такими как ингибиторы ALK5, описанные Anderton et al., и/или антителом 1D11, описанным Lonning et al. (2011). В другом аспекте антитело к TGFβ3, представленное в данном документе, селективно нейтрализует TGFβ3 и имеет сниженную токсичность у мышей по сравнению с антителом САТ-192/метелимумаб к TGFβ1.
В определенных аспектах антитело CAT-192 имеет следующие вариабельные аминокислотные последовательности тяжелой и легкой цепи:


В любом из вышеуказанных аспектов представлено антитело к TGFβ3, содержащее по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащей аминокислотную последовательность, выбранную из SEQ ID NO: 5, 34, 35 или 159; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 9.
В одном аспекте в настоящем изобретении представлено антитело к TGFβ3, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 5; и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6. В другом аспекте в настоящем изобретении представлено антитело к TGFβ3, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 34; и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6. В другом аспекте в настоящем изобретении представлено антитело к TGFβ3, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 35; и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6. В другом аспекте в настоящем изобретении представлено антитело к TGFβ3, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 159; и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6.
В одном варианте осуществления антитело содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6. В другом варианте осуществления антитело содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6, и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9. В дополнительном варианте осуществления антитело содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6, HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5. В дополнительном варианте осуществления антитело к TGFβ3 содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6, HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 34. В дополнительном варианте осуществления антитело к TGFβ3 содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6, HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 35. В дополнительном варианте осуществления антитело к TGFβ3 содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6, HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 159. В дополнительном варианте осуществления антитело к TGFβ3 содержит (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6. В дополнительном варианте осуществления антитело к TGFβ3 содержит (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 34; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6. В дополнительном варианте осуществления антитело к TGFβ3 содержит (a) HVR-Н1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-Н2, содержащую аминокислотную последовательность SEQ ID NO: 35; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6. В дополнительном варианте осуществления антитело к TGFβ3 содержит (a) HVR-Н1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-Н2, содержащую аминокислотную последовательность SEQ ID NO: 159; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6.
В другом аспекте в настоящем изобретении представлено антитело к TGFβ3, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 7; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 8; и (в) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 9. В одном варианте осуществления антитело содержит (a) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (в) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9.
В другом аспекте антитело к TGFβ3 по настоящему изобретению содержит (а) домен VH, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4, (ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 5, и (iii) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6; и (б) домен VL, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VL, выбранные из (i) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 7, (ii) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 8, и (iii) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 9. В другом аспекте антитело к TGFβ3 по настоящему изобретению содержит (а) домен VH, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4, (ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 34, и (iii) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6; и (б) домен VL, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VL, выбранные из (i) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 7, (ii) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 8, и (iii) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 9. В другом аспекте антитело к TGFβ3 по настоящему изобретению содержит (а) домен VH, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4, (ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 35, и (iii) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6; и (б) домен VL, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VL, выбранные из (i) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 7, (ii) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 8, и (iii) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 9. В другом аспекте антитело к TGFβ3 по настоящему изобретению содержит (а) домен VH, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4, (ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 159, и (iii) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6; и (б) домен VL, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VL, выбранные из (i) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 7, (ii) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 8, и (iii) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 9.
В другом аспекте в настоящем изобретении представлено антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9. В другом аспекте в настоящем изобретении представлено антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 34; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9. В другом аспекте в настоящем изобретении представлено антитело к TGFβ3, содержащее (a) HVR-Н1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-Н2, содержащую аминокислотную последовательность SEQ ID NO: 35; (в) HVR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (e) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9. В другом аспекте в настоящем изобретении представлено антитело к TGFβ3, содержащее
(а) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4;
(б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 159;
(в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
(г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7;
(д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и
(е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9.
В определенных вариантах осуществления любая одна или более аминокислот антитела к TGFβ3, как указано выше, замещены в следующих положениях HVR:
- в HVR-H2 (SEQ ID NO: 5): в положении N54 (например, N54S, N54Q) или Т56 (например, Т56А).
В некоторых вариантах осуществления замены представляют собой консервативные замены, как представлено в данном документе.
В любом из вышеуказанных вариантов осуществления антитело к TGFβ3 является гуманизированным. В некоторых аспектах антитело к TGFβ3, представленное в данном документе, которое подверглось одной или более этапам гуманизации, имеет способность блокировать TGFβ, аналогичную исходному антителу, и/или связывание TGFβ человека, аналогичное исходному антителу, и/или сохраняет растворимость, и/или способно экспрессироваться, в то время как такие возможности могут быть неизвестны или не наблюдаться в вариантах, потенциально или фактически полученных из других антител к TGFβ3 (пример 6, таблицы 9 и 10; Фиг. 10, 11). В одном варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область. В другом варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VH, содержащую модификации FR, выбранные из группы, состоящей из 47L или 47W; 49А, 49S или 49G; 73D или 73N; и 76N, 78D или 78L, 78А или 78V. В некоторых вариантах осуществления антитело к TGFβ3 содержит VH, где VH содержит аминокислотную последовательность SEQ ID NO: 23, при этом VH содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 47L, 49А в FR2, 78V в FR3 (h2A10.v1); (ii) 47L, 49А в FR2, 73D, 76S, 78V в FR3 (h2A10.v2); (iii) 47W в FR2 (h2A10.v1.5); (iv): 49G в FR2 (h2A10.v1.6); (v) 78A в FR3 (h2A10.v1.7); (vi) 47W в FR2 (h2A10.v2.5); (vii) 49S в FR2 (h2A10.v2.6); (viii) 73N в FR3 (h2A10.v2.7); (ix) 76N в FR3 (h2A10.v2.8); (x) 78L в FR3 (h2A10.v2.9); и (xi) 49S в FR2, 76N, 78L в FR3 (h2A10.v3 и h2A10.v4). В другом варианте осуществления антитело к TGF(33 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VL, содержащую модификации FR, выбранные из группы, состоящей из: 4L или 4М, 38Н или 38Q, 43А или 43Q и 58V (по отношению к аминокислотной последовательности VL SEQ ID NO: 22). В другом варианте осуществления антитело содержит VL SEQ ID NO: 22, где VL содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 4L в FR1, 38Н и 43Q в FR2, 58I в FR3 (h2A10.v1 и h2A10.v2); (ii) 4М в FR1 (h2A10.v1.1 и h2A10.v2.1); (iii) 38Q в FR2 (h2A10.v1.2 и h2A10.v2.2); (iv) 43А в FR2 (h2A10.v1.3 и h2A10.v2.3); (v) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); (vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4); (vii) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); и (vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4).
В другом аспекте антитело к TGFβ3 содержит последовательность вариабельного домена тяжелой цепи (VH), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37, 42-50, 55 и 57. В другом аспекте антитело к TGFβ3 содержит последовательность вариабельного домена тяжелой цепи (VH), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37 и 42-50. В некоторых аспектах антитело к TGFβ3 содержит последовательность вариабельного домена тяжелой цепи (VH), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 57. В определенных вариантах осуществления последовательность VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности, содержит замены (например, консервативные замены), вставки или делеции по отношению к эталонной последовательности, но антитело к TGFβ3, содержащее эту последовательность, сохраняет способность связываться с TGFβ3. В определенных вариантах осуществления в общей сложности 1-10 аминокислот были замещены, вставлены и/или делетированы в SEQ ID NO: 23. В определенных вариантах осуществления замены, вставки или делеции находятся в областях за пределами HVR (т.е. в FR). Необязательно, антитело к TGFβ3 содержит последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 23, 37, 42-50, 55 и 57, включая посттрансляционные модификации этой последовательности. Необязательно, антитело к TGFβ3 содержит последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 23, 37 и 42-50, включая посттрансляционные модификации этой последовательности. В конкретном варианте осуществления VH содержит одну, две или три HVR, выбранные из: (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4, (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 5, и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6. В конкретном варианте осуществления VH содержит одну, две или три HVR, выбранные из: (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4, (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 34, и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6. В конкретном варианте осуществления VH содержит одну, две или три HVR, выбранные из: (а) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4, (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 35, и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6. В конкретном варианте осуществления VH содержит одну, две или три HVR, выбранные из: (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4, (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 159, и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6.
В другом аспекте представлено антитело к TGFβ3, где антитело содержит последовательность вариабельного домена легкой цепи (VL), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41, 54 и 56. В другом аспекте представлено антитело к TGFβ3, где антитело содержит последовательность вариабельного домена легкой цепи (VL), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41. В некоторых аспектах представлено антитело к TGFβ3, где антитело содержит последовательность вариабельного домена легкой цепи (VL), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 56. В определенных вариантах осуществления последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности последовательности, содержит замены (например, консервативные замены), вставки или делеции по отношению к эталонной последовательности, но антитело к TGFβ3, содержащее эту последовательность, сохраняет способность связываться с TGFβ3. В определенных вариантах осуществления в общей сложности 1-10 аминокислот были замещены, вставлены и/или делетированы в SEQ ID NO: 22. В определенных вариантах осуществления замены, вставки или делеции встречаются в областях за пределами HVR (т.е. в FR). Необязательно, антитело к TGFβ3 содержит последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 22, 36, 38-41, 54 и 56, включая посттрансляционные модификации этой последовательности. Необязательно, антитело к TGFβ3 содержит последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 22, 36 и 38-41, включая посттрансляционные модификации этой последовательности. В конкретном варианте осуществления VL содержит одну, две или три HVR, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 7; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 8; и (в) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 9.
В другом аспекте представлено антитело к TGFβ3, где антитело содержит VH, как в любом из вышеуказанных вариантов осуществления, и VL, как в любом из вышеуказанных вариантов осуществления. В одном варианте осуществления антитело к TGFβ3 содержит последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8), SEQ ID NO: 50/36 (v2.9), SEQ ID NO: 55/54 (v3) и SEQ ID NO: 57/56 (v4), включая посттрансляционные модификации этих последовательностей. В одном варианте осуществления антитело к TGFβ3 содержит последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9), включая посттрансляционные модификации этих последовательностей. В некоторых вариантах осуществления антитело к TGFβ3 содержит последовательности VH/VL (соответственно) SEQ ID NO: 57/56.
В другом аспекте представлено антитело к TGFβ3, где антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 59, 64, 65-72, 77 и 79, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 28, 58, 60-63, 76 и 78. В другом аспекте представлено антитело к TGFβ3, где антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 59, 64 и 65-72, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63. В некоторых вариантах осуществления антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности последовательности SEQ ID NO: 79, и/или полную аминокислотную последовательность L-цепи SEQ ID NO: 78. В некоторых вариантах осуществления антитело к TGFβ3 содержит полную пару H/L-цепей, при этом полная пара H/L-цепей (соответственно) содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.6), SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8), SEQ ID NO: 72/58 (v2.9), SEQ ID NO: 77/76 (v3) и SEQ ID NO: 79/78 (v4). В некоторых вариантах осуществления антитело к TGFβ3 содержит полную пару H/L-цепей, при этом полная пара H/L-цепей содержит (соответственно) аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.l), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.6), SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8) и SEQ ID NO: 72/58 (v2.9).
В другом варианте осуществления представлено антитело к TGFβ3, где антитело содержит VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 52. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41. В некоторых аспектах антитело содержит последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 52/36 (h2A10.v2.N54Q). В некоторых аспектах антитело содержит полную Н-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74. В некоторых аспектах антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63. В некоторых аспектах антитело к TGFβ3 содержит полную Н-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74, и полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63. В некоторых аспектах антитело содержит полные последовательности H/L-цепи (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 74/58.
В другом варианте осуществления представлено антитело к TGFβ3, где антитело содержит VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 51 или 55. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41 и 54. В некоторых аспектах антитело к TGFβ3 содержит последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 51/36 или SEQ ID NO: 55/54 (h2A10.v3). В некоторых аспектах антитело содержит полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 73 или 77, и/или антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58, 60-63 и 76. В некоторых аспектах антитело содержит полные последовательности H/L-цепи (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 73/58 или SEQ ID NO: 77/76.
В другом варианте осуществления представлено антитело к TGFβ3, где антитело содержит VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 53 или 57. В некоторых аспектах антитело к TGFβ3 содержит аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41 и 56. В некоторых аспектах антитело к TGFP3 содержит VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 57. В других аспектах антитело к TGFβ3 содержит VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 56. В некоторых аспектах антитело к TGFβ3 содержит VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности, аминокислотной последовательности SEQ ID NO: 57, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 56. В некоторых аспектах антитело к TGFβ3 содержит последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 53/36 или SEQ ID NO: 57/56 (h2A10.v4). В некоторых аспектах антитело к TGFβ3 содержит последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 57/56 (h2A10.v4). В некоторых аспектах антитело содержит полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74 или 79, и/или полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58, 60-63, 76 и 78. В некоторых аспектах антитело к TGFβ3 содержит полную Н-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 79. В других аспектах антитело к TGFβ3 содержит полную L-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 78. В еще одних дополнительных аспектах антитело к TGFβ3 содержит полную Н-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 79; и содержит полную L-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 78. В некоторых аспектах антитело содержит полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 79, и/или полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 78. В некоторых аспектах антитело к TGFβ3 содержит полные последовательности H/L-цепи (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 75/58 или SEQ ID NO: 79/78. В определенных аспектах антитело к TGFβ3 содержит полные последовательности H/L-цепи (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 79/78.
В дополнительном аспекте в настоящем изобретении представлено антитело к TGFβ3, которое связывается с тем же эпитопом, что и антитело к TGFβ3, представленное в данном документе. Например, в некоторых вариантах осуществления представлено антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6. В одном варианте осуществления представлено антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 34; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6. В другом варианте осуществления представлено антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 35; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6. В другом варианте осуществления представлено антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 159; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6.
В другом аспекте представлено антитело к TGFβ3, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 53/36 (h2A10.v2.t56A), SEQ ID NO: 57/56 (h2A10.v4), SEQ ID NO: 51/36 (h2A10.v2.N54S), SEQ ID NO: 55/54 (h2A10.v3), SEQ ID NO: 52/36 (h2A10.v2.N54Q), SEQ ID NO: 23/22 (2A10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.l), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8), SEQ ID NO: 50/36 (v2.9), SEQ ID NO: 55/54 (v3) и SEQ ID NO: 57/56 (v4). В другом аспекте представлено антитело к TGFβ3, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 53/36 (h2A10.v2.t56A), SEQ ID NO: 57/56 (h2A10.v4), SEQ ID NO: 51/36 (h2A10.v2.N54S), SEQ ID NO: 55/54 (h2A10.v3), SEQ ID NO: 52/36 (h2A10.v2.N54Q), SEQ ID NO: 23/22 (2A10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9).
В одном варианте осуществления представлено антитело к TGFβ3, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно) SEQ ID NO: 57/56 (h2A10.v4).
В некоторых вариантах осуществления представлено выделенное антитело к TGFβ3, содержащее антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, 1388, L389, Y391, V392, G393, R394, Р396, K397 и V398 на TGFβ3 человека, где антитело к TGFβ3 селективно нейтрализует TGFβ3.
В некоторых аспектах в данном документе представлено выделенное антитело к TGFβ3, которое содержит: CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9. В некоторых вариантах осуществления CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35. В определенных вариантах осуществления выделенное антитело к TGFβ3 дополнительно содержит VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 57. В некоторых вариантах осуществления антитело к TGFβ3 дополнительно содержит VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 56. В определенных вариантах осуществления антитело к TGFβ3 дополнительно содержит полную Н-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 79. В других вариантах осуществления антитело к TGFβ3 дополнительно содержит полную L-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 78.
В другом варианте осуществления представлено выделенное антитело к TGFβ3, которое селективно нейтрализует TGFβ3 и имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ. В некоторых вариантах осуществления антитело к TGFβ3, представленное в данном документе, имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ в дозе 50 мг/кг. Антитело 1D11 описано в Lonning et al. (Current Pharmaceutical Biotechnology, 2011, 12, 2176-2189) и имеет следующие аминокислотные последовательности:

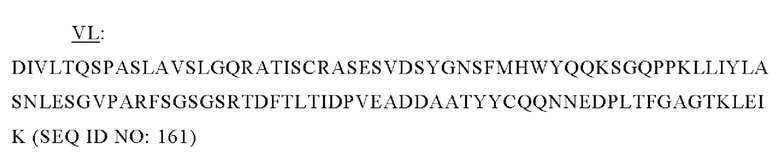
В дополнительном аспекте настоящего изобретения селективное в отношении изоформы антитело к TGFβ в соответствии с любым из вышеуказанных вариантов осуществления (например, моноспецифические антитела к TGFβ2 и к TGFβ3 и антитела с двойной специфичностью к TGFβ2/3, описанные в данном документе) представляет собой моноклональное антитело, включая химерное, гуманизированное или человеческое антитело. В одном варианте осуществления селективное в отношении изоформы антитело к TGFβ представляет собой фрагмент антитела, например, фрагмент Fv, Fab, Fab', scFv, диатело или фрагмент F(ab')2. В другом варианте осуществления антитело представляет собой полноразмерное антитело, например, интактное антитело на основе IgG1 или IgG4, предпочтительно IgG1 человека и еще более предпочтительно IgG1 человека, содержащие мутацию N297 (EU-нумерация согласно Kabat), например, N297A или N297G, предпочтительно N297G, или другого класса или изотипа антитела, как определено в данном документе.
В дополнительном аспекте антитело к TGFβ в соответствии с любым из вышеуказанных вариантов осуществления может включать любые из признаков, по отдельности или в комбинации, как описано ниже в разделах 1-7:
1. Аффинность антител
В определенных вариантах осуществления антитело, представленное в данном документе, имеет константу диссоциации (KD) ≤ 1 мкМ, ≤ 100 нМ, ≤ 10 нМ, ≤ 1 нМ, ≤ 0,1 нМ, ≤ 0,01 нМ или ≤ 0,001 нМ (например, 10-8 М или меньше, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М).
В одном варианте осуществления KD измеряется с помощью анализа связывания антигена, меченного радиоактивным изотопом (RIA). В одном варианте осуществления RIA проводят с Fab-версией антитела, представляющего интерес, и его антигеном. Например, аффинность связывания Fab с антигеном в растворе измеряют путем уравновешивания Fab с минимальной концентрацией (125I)-меченого антигена в присутствии ряда титрующих концентраций немеченого антигена с последующим захватом связанного антигена на планшете, покрытом антителом к Fab (см., например, Chen et al., J. Mol. Biol. 293:865-881(1999)). Для определения условий анализа многолуночные планшеты MICROTITER® (Thermo Scientific) покрывают на ночь 5 мкг/мл захватывающего антитела к Fab (Cappel Labs) в 50 мМ растворе карбоната натрия (рН 9,6), а затем блокируют 2% (масс./об.) бычьим сывороточным альбумином в PBS в течение двух-пяти часов при комнатной температуре (примерно 23°С). В неадсорбентном планшете (Nunc №269620) смешивают 100 пМ или 26 пМ [125I]-антигена с серийными разведениями Fab, представляющего интерес (например, в соответствии с оценкой антитела к VEGF, Fab-12, в Presta et al., Cancer Res. 57:4593-4599 (1997)). Затем Fab, представляющий интерес, инкубировали в течение ночи; однако инкубацию можно продолжать в течение более длительного периода (например, приблизительно 65 часов), чтобы обеспечить достижение равновесия. Затем смесь переносят на планшет для захвата и инкубируют при комнатной температуре (например, в течение часа). Затем раствор удаляют и восемь раз промывают планшет 0,1% полисорбатом 20 (TWEEN-20®) в PBS. После высушивания планшетов добавляют 150 мкл/лунка сцинтиллятора (MICROSCINT-20™; Packard) и производят подсчет планшетов на гамма-счетчике TOPCOUNT™ (Packard) в течение десяти минут. Концентрации каждого Fab, который дает 20% или меньше от максимального связывания, отбирают для использования в конкурентном анализе связывания.
В соответствии с другим вариантом осуществления KD определяют с применением анализа на основе поверхностного плазмонного резонанса BIACORE®. Например, анализ с применением BIACORE®-2000 или BIACORE®-3000 (BIAcore, Inc., Пискатауэй, Нью-Джерси) проводят при 25°С с чипами СМ5 с иммобилизованным антигеном при ~10 единицах ответа (RU). В одном варианте осуществления биосенсорные чипы на основе карбоксиметилированного декстрана (СМ5, BIACORE, Inc.) активируют N-этил-N'-(3-диметиламинопропил)-карбодиимида гидрохлоридом (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разводят в 10 мМ растворе ацетате натрия, рН 4,8, до 5 мкг/мл (~ 0,2 мкМ) перед вводом при скорости потока 5 мкл/минута для достижения примерно 10 единиц ответа (RU) связанного белка. После ввода антигена вводят 1 М этаноламина, чтобы блокировать не вступившие в реакцию группы. Для измерений кинетики двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) вводят в PBS с содержащим 0,05% полисорбата 20 (TWEEN-20™) поверхностно-активным веществом (PBST) при 25°С при скорости потока примерно 25 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) рассчитывают, используя простую один-к-одному модель связывания Лэнгмюра (оценочное программное обеспечение BIACORE®, версия 3.2), путем одновременной аппроксимации сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (KD) рассчитывали, как соотношение koff/kon. См., например, Chen et al., J. Mot Biol. 293:865-881 (1999). Если скорость ассоциации превышает 106 М-1 с-1 по данным вышеописанного анализа на основе поверхностного плазмонного резонанса, то скорость ассоциации можно определить, используя метод гашения флуоресценции, в котором измеряют повышение или снижение интенсивности флуоресценции (возбуждение = 295 нм; испускание = 340 нм, 16 нм полоса пропускания) при 25°С для 20 нМ антитела к антигену (Fab-формы) в PBS, рН 7,2, в присутствии возрастающих концентраций антигена, с измерением на спектрометре, таком как спектрофотометр с остановкой потока (Aviv Instruments) или спектрофотометр серии 8000 SLM-AMINCO™ (ThermoSpectronic) с кюветой с перемешиванием.
2. Фрагменты антител
В определенных вариантах осуществления антитело, представленное в данном документе, представляет собой фрагмент антитела. Фрагменты антител включают без ограничения фрагменты Fab, Fab', Fab'-SH, F(ab')2, Fv и scFv и другие фрагменты, описанные ниже. Для обзора некоторых фрагментов антител см. Hudson et al. Nat. Med. 9:129-134 (2003). Для обзора фрагментов scFv см., например, Pluckthün, в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., (Springer-Verlag, New York), pp. 269-315 (1994); см. также WO 93/16185; и патенты США №№5571894 и 5587458. Обсуждение фрагментов Fab и F(ab')2, содержащих остатки эпитопа связывания рецептора реутилизации и имеющих повышенное время полужизни in vivo, см. в патенте США №5869046.
Диатела представляют собой фрагменты антител с двумя антигенсвязывающими сайтами, которые могут быть двухвалентными или биспецифическими. См., например, ЕР 404097; WO 1993/01161; Hudson et al., Nat. Med. 9:129-134 (2003); и Hollinger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993). Тритела и тетратела также описаны в Hudson et al., Nat. Med. 9:129-134 (2003).
Однодоменные антитела представляют собой фрагменты антител, содержащие весь или часть вариабельного домена тяжелой цепи или весь или часть вариабельного домена легкой цепи антитела. В определенных вариантах осуществления однодоменное антитело представляет собой человеческое однодоменное антитело (Domantis, Inc., Уолтем, Массачусетс; см., например, патент США №6248516 В1).
Фрагменты антител можно получать различными способами, включая без ограничения протеолитическое расщепление интактного антитела, а также выработку рекомбинантными клетками-хозяевами (например, Е. coli или фагом), как описано в данном документе.
3. Химерные и гуманизированные антитела
В определенных вариантах осуществления антитело, представленное в данном документе, представляет собой химерное антитело. Некоторые химерные антитела описаны, например, в патенте США №4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). В одном примере химерное антитело содержит отличную от человеческой вариабельную область (например, вариабельную область, полученную из антитела мыши, крысы, хомяка, кролика или отличного от человека примата, например, обезьяны) и константную область человека. В дополнительном примере химерное антитело представляет собой антитело с «переключенным классом», класс или подкласс которого был изменен по сравнению с исходным антителом. Химерные антитела включают их антигенсвязывающие фрагменты.
В определенных вариантах осуществления химерное антитело представляет собой гуманизированное антитело. Как правило, антитело отличного от человеческого происхождения гуманизируют с целью снижения иммуногенности для человека, сохраняя при этом специфичность и аффинность исходного антитела отличного от человеческого происхождения. Как правило, гуманизированное антитело содержит один или более вариабельных доменов, в которых HVR, например, CDR (или их фрагменты) происходят от антитела отличного от человеческого происхождения, a FR (или их фрагменты) происходят из последовательностей человеческого антитела. Гуманизированное антитело необязательно также содержит по меньшей мере часть человеческой константной области. В некоторых вариантах осуществления некоторые FR остатки гуманизированного антитела заменены соответствующими остатками из антитела отличного от человеческого происхождения (например, антитела, из которого получены остатки HVR), например, для восстановления или улучшения специфичности или аффинности антитела.
Гуманизированные антитела и способы их получения рассмотрены, например, в Almagro and Fransson, Front. Biosci. 13:1619-1633 (2008), и дополнительно описаны, например, в Riechmann et al., Nature 332:323-329 (1988); Queen et al., Proc. Nat'l Acad. Sci. USA 86:10029-10033 (1989); патентах США №№5821337, 7527791, 6982321 и 7087409; Kashmiri et al., Methods 36:25-34 (2005) (где описано прививание определяющей специфичность области (SDR)); Padlan, Mol. Immunol. 28:489-498 (1991) (где описано «изменение поверхности»); Dall'Acqua et al., Methods 36:43-60 (2005) (где описана «шаффлинг FR»); и Osbourn et al., Methods 36:61-68 (2005) и Klimka et al., Br. J. Cancer, 83:252-260 (2000) (где описан подход «направленного отбора» при шаффлинге FR).
Человеческие каркасные области, которые можно применять для гуманизации, включают без ограничения: каркасные области, выбранные методом «наилучшего соответствия» (см., например, Sims et al. J. Immunol. 151:2296 (1993)); каркасные области, полученные из консенсусной последовательности человеческих антител конкретной подгруппы вариабельных областей легких или тяжелых цепей (см., например, Carter et al. Proc. Natl. Acad. Sci. USA, 89:4285 (1992); и Presta et al. J. Immunol., 151:2623 (1993)); человеческие зрелые (соматически мутированные) каркасные области или каркасные области зародышевой линии человека (см., например, Almagro and Fransson, Front. Biosci. 13:1619-1633 (2008)); и каркасные области, полученные при скрининге библиотек FR (см., например, Baca et al., J. Biol. Chem. 272:10678-10684 (1997) и Rosok et al., J. Biol. Chem. 271:22611-22618 (1996)).
4. Человеческие антитела
В некоторых вариантах осуществления антитело, представленное в данном документе, представляет собой человеческое антитело. Человеческие антитела можно получать, используя различные методики, известные в данной области техники. Человеческие антитела, описаны в целом в van Dijk and van de Winkel, Curr. Opin. Pharmacol. 5: 368-74 (2001) and Lonberg, Curr. Opin. Immunol. 20:450-459 (2008).
Человеческие антитела можно получать путем введения иммуногена трансгенному животному, модифицированному для выработки интактных человеческих антител или интактных антител с человеческими вариабельными областями в ответ на антигенную стимуляцию. Такие животные, как правило, содержат все или часть локусов иммуноглобулина человека, которые замещают эндогенные локусы иммуноглобулина или которые находятся вне хромосом или случайным образом интегрированы в хромосомы животного. Как правило, у таких трансгенных мышей инактивированы эндогенные локусы иммуноглобулина. Обзор способов получения человеческих антител из трансгенных животных см. в Lonberg, Nat. Biotech. 23:1117-1125 (2005). Также см., например, патенты США №№6075181 и 6150584, в которых описан способ XENOMOUSE™; патент США №5770429, в котором описан способ HUMAB®; патент №7041870, в котором описан способ K-М MOUSE®, и публикацию заявки на патент США № US 2007/0061900, в которой описан способ VELOCIMOUSE®). Человеческие вариабельные области из интактных антител, образуемых такими животными, можно дополнительно модифицировать, например, путем объединения с разными человеческими консервативными областями.
Человеческие антитела можно получать с помощью способов на основе гибридом. Описаны линии клеток миеломы человека и гетеромиеломы мыши и человека для продуцирования человеческих моноклональных антител. (См., например, Kozbor J. Immunol., 133: 3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al., J. Immunol., 147: 86 (1991).) Человеческие антитела, полученные посредством способа человеческой В-клеточной гибридомы, также описаны в Li et al., Proc. Natl. Acad. Sci. USA, 103:3557-3562 (2006). Дополнительные способы включают способы, описанные, например, в патенте США №7189826 (где описано получение моноклональных человеческих антител на основе IgM из линий клеток гибридомы) и Ni, Xiandai Mianyixue, 26(4):265-268 (2006) (где описаны гибридомы типа человек-человек). Способ получения человеческой гибридомы (способ триомы) также описан в Vollmers and Brandlein, Histology and Histopathology, 20(3):927-937 (2005), и Vollmers and Brandlein, Methods and Findings in Experimental and Clinical Pharmacology, 27(3): 185-91 (2005).
Человеческие антитела также можно получать путем выделения последовательностей вариабельного домена Fv-клона из библиотек фагового дисплея человеческого происхождения. Такие последовательности вариабельного домена затем можно объединять с необходимым человеческим константным доменом. Методики отбора человеческих антител из библиотек антител описаны ниже.
5. Антитела, полученные из библиотек
Антитела по настоящему изобретению могут быть выделены путем скрининга комбинаторных библиотек антител с необходимой активностью или свойствами. Например, в данной области техники известны различные способы получения библиотек фагового дисплея и скрининг таких библиотек в отношении антител, обладающих необходимыми характеристиками связывания. Такие способы рассматриваются, например, в Hoogenboom et al. в Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, 2001) и далее описаны, например, в McCafferty et al., Nature 348:552-554; Clackson et al., Nature 352: 624-628 (1991); Marks et al., J. Mol. Biol. 222: 581-597 (1992); Marks and Bradbury, в Methods in Molecular Biology 248: 161-175 (Lo, ed., Human Press, Totowa, NJ, 2003); Sidhu et al., J. Mol. Biol. 338 (2): 299-310 (2004); Lee et al., J. Mol. Biol. 340 (5): 1073-1093 (2004); Fellouse, Proc. Natl. Acad. Sci. USA 101 (34): 12467-12472 (2004); и Lee et al., J. Immunol. Methods 284 (1-2): 119-132 (2004).
В некоторых способах фагового дисплея репертуары VH- и VL-генов по отдельности клонируют с помощью полимеразной цепной реакции (ПЦР) и случайным образом рекомбинируют в фаговых библиотеках, которые впоследствии можно подвергать скринингу в отношении антигенсвязывающего фага, как описано в Winter et al., Ann. Rev. Immunol., 12: 433-455 (1994). Как правило, фаг представляет фрагменты антител в виде одноцепочечных Fv-фрагментов (scFv) или Fab-фрагментов. Библиотеки из иммунизированных источников позволяют получать антитела с высокой аффинностью к иммуногену без необходимости конструирования гибридом. В качестве альтернативы можно клонировать наивный репертуар (например, от человека) с получением единого источника антител к широкому спектру не собственных, а также собственных антигенов без какой-либо иммунизации, как описано в Griffiths et al., EMBO J, 12: 725-734 (1993). В конечном итоге наивные библиотеки также можно получать синтетически, клонируя неаранжированные сегменты V-генов из стволовых клеток и используя ПЦР-праймеры, содержащие случайную последовательность, для кодирования высоковариабельных областей CDR3 и осуществления реаранжировки in vitro, как описано в Hoogenboom and Winter, J. Mol. Biol., 227: 381-388 (1992). Патентные публикации, в которых описаны фаговые библиотеки человеческих антител, включают, например: патент США №5750373 и патентные публикации США №№2005/0079574, 2005/0119455, 2005/0266000, 2007/0117126, 2007/0160598, 2007/0237764, 2007/0292936 и 2009/0002360.
Антитела или фрагменты антител, выделенные из библиотек человеческих антител, считаются в данном документе человеческими антителами или фрагментами человеческих антител.
6. Мультиспецифические антитела
В определенных вариантах осуществления антитело, представленное в данном документе, представляет собой мультиспецифическое антитело, например, биспецифическое антитело. Мультиспецифические антитела представляют собой моноклональные антитела, которые имеют специфичность связывания в отношении по меньшей мере двух разных сайтов. В определенных вариантах осуществления одна из специфичностей связывания направлена на изоформу TGFβ (например, TGFβ1, TGFβ2, TGFβ3 или TGFβ2/3), а другая направлена на любой другой антиген. В определенных вариантах осуществления одна из специфичностей связывания направлена на одну изоформу TGFβ (например, TGFβ1, TGFβ2 или TGFβ3), а другая направлена на другую изоформу TGFβ.В определенных вариантах осуществления биспецифические антитела могут связываться с двумя разными эпитопами в одной изоформе TGFβ. Биспецифические антитела также можно использовать для локализации цитотоксических средств в клетках, которые экспрессируют одну или более изоформ TGFβ. Биспецифические антитела можно получать в виде полноразмерных антител или фрагментов антител.
Методики получения мультиспецифических антител включают без ограничения рекомбинантную коэкспрессию двух пар тяжелая цепь легкая цепь иммуноглобулина, имеющих разные специфичности (см. Milstein et al. Nature 305: 537, (1983)), WO 93/08829 и Traunecker et al. EMBO J. 10: 3655, (1991)), и конструирование по типу «выступ-во-впадину» (см., например, патент США №5731168). Мультиспецифические антитела также можно получать путем конструирования с использованием эффекта электростатического взаимодействия для получения Fc-гетеродимерных молекул антител (WO 2009/089004 А1); перекрестного сшивания двух или более антител или фрагментов (см., например, патент США №4676980, и Brennan et al., Science, 229: 81 (1985)); использования лейциновых молний для получения биспецифических антител (см., например, Kostelny et al., J. Immunol., 148 (5): 1547-1553 (1992)); использования способа «диател» для получения фрагментов биспецифических антител (см., например, Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993)); и использования димеров одноцепочечных Fv (sFv) (см., например, Gruber et al., J. Immunol, 152: 5368 (1994)); и получения триспецифических антител, как описано, например, в Tint et al. J. Immunol. 147: 60 (1991).
Сконструированные антитела с тремя или более функциональными антигенсвязывающими сайтами, включая «антитела-осьминоги», также включены в данный документ (см., например, US 2006/0025576 А1).
Антитело или фрагмент в данном документе также включает в себя «Fab двойного действия» или «DAF», содержащий антигенсвязывающий сайт, который связывается с одной или более изоформами TGFβ (например, TGFβ1, TGFβ2 и/или TGFβ3), а также с другим, отличным от него антигеном (см., например, US 2008/0069820).
7. Варианты антител
В определенных вариантах осуществления в данном документе предусмотрены варианты аминокислотной последовательности антител. Например, может существовать потребность в улучшении аффинности связывания и/или других биологических свойств антитела. Варианты аминокислотной последовательности антитела можно получать путем внесения соответствующих модификаций в нуклеотидную последовательность, кодирующую антитело, или с помощью пептидного синтеза. Такие модификации включают, например, делеции, и/или вставки, и/или замены остатков в аминокислотных последовательностях антитела. Для получения конечной конструкции можно осуществлять любую комбинацию делеций, вставок и замен при условии, что конечная конструкция обладает требуемыми характеристиками, например, связывания антигена.
а) Варианты с заменами, вставками и делениями
В определенных вариантах осуществления представлены варианты антитела, содержащие одну или более аминокислотных замен. Сайты, представляющие интерес для заместительного мутагенеза, включают HVR и FR. Консервативные замены показаны в таблице 1 под заголовком «предпочтительные замены». Более существенные изменения представлены в таблице 1 под заголовком «иллюстративные замены» и дополнительно описаны ниже по отношению к классам боковой цепи аминокислот. Аминокислотные замены можно вносить в антитело, представляющее интерес, и проводить скрининг полученных продуктов в отношении необходимой активности, например, сохранения/улучшения связывания антигена, снижения иммуногенности или улучшения ADCC или CDC.
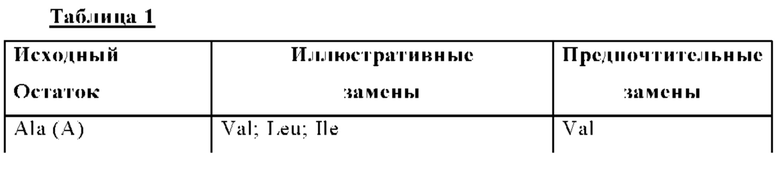
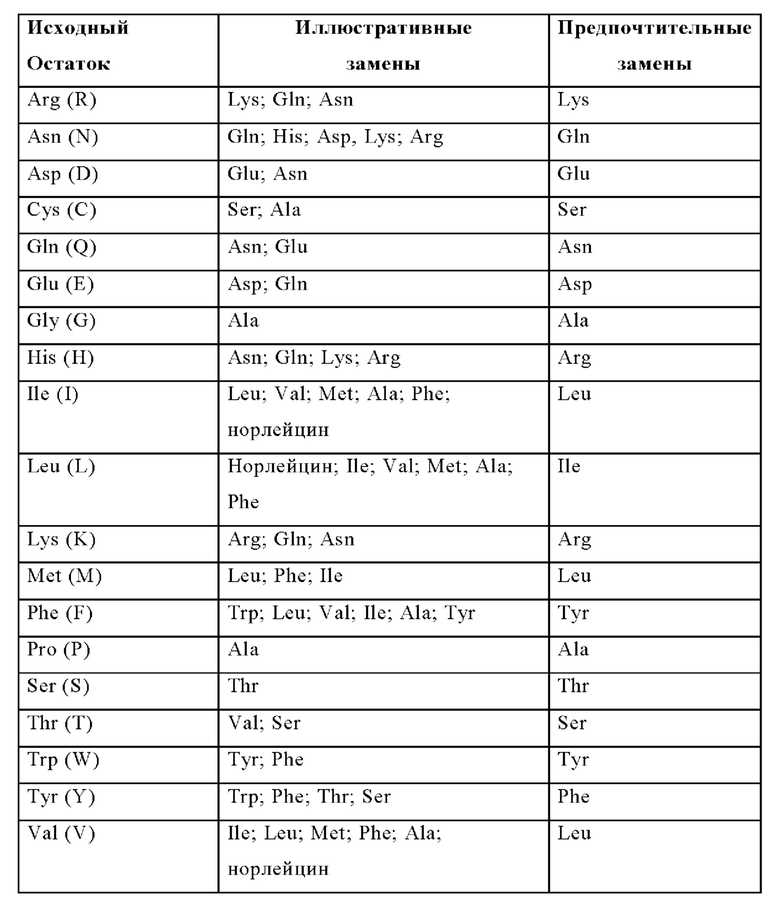
Аминокислоты можно разделить на группы в соответствии с общими свойствами боковых цепей:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gin;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, влияющие на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены повлекут за собой замену члена одного из этих классов другим классом.
Один тип заместительного варианта включает замену одного или более остатков гипервариабельной области исходного антитела (например, гуманизированного или человеческого антитела). Как правило, полученный (ые) вариант (ы), выбранный (ые) для дальнейшего изучения, будет иметь модификации (например, улучшения) определенных биологических свойств (например, повышенную аффинность, сниженную иммуногенность) по сравнению с исходным антителом и/или будет иметь, по сути, сохраненные некоторые биологические свойства исходного антитела. Иллюстративный заместительный вариант представляет собой антитело с созревшей аффинностью, которое удобно получать, например, используя методики созревания аффинности на основе фагового дисплея, такие как описанные в данном документе. Вкратце один или более аминокислотных остатков HVR подвергают мутации и вариантные антитела представляют на поверхности фага и проводят скрининг в отношении конкретного вида биологической активности (например, аффинности связывания).
Модификации (например, замены) в HVR можно осуществить, например, с целью улучшения аффинности антитела. Такие изменения могут быть сделаны в «горячих точках» HVR, т.е. в остатках, кодируемых кодонами, которые с высокой частотой подвергаются мутации во время процесса соматического созревания (см., например, Chowdhury, Methods Mol. Biol. 207: 179-196 (2008)), и/или в остатках, которые контактируют с антигеном, при этом испытывают аффинность связывания полученного варианта VH или VL. Созревание аффинности путем конструирования и повторного отбора из вторичных библиотек было описано, например, в Hoogenboom et al. в Methods in Molecular Biology 178: 1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, (2001)). В некоторых вариантах осуществления созревания аффинности многообразие вводится в вариабельные гены, выбранные для созревания любым из множества способов (например, ПЦР с внесением ошибок, шаффлинг цепей или олигонуклеотид-направленный мутагенез). Затем создают вторичную библиотеку. После этого библиотеку подвергают скринингу для идентификации любых вариантов антител с требуемой аффинностью. Другой способ внесения разнообразия включает подходы, направленные на HVR, в которых несколько остатков HVR (например, 4-6 остатков подряд) являются рандомизированными. Остатки HVR, вовлеченные в связывание антигена, могут быть конкретно идентифицированы, например, с использованием сканирующего аланином мутагенеза или моделирования. В частности, мишенями часто являются CDR-H3 и CDR-L3.
В определенных вариантах осуществления замены, вставки или делеции могут находиться в одной или более HVR до той меры, пока такие изменения существенно не снижают способность антитела связывать антиген. Например, в HVR можно проводить консервативные изменения (например, консервативные замены, представленные в данном документе), которые существенно не снижают аффинность связывания. Такие изменения могут, например, находиться за пределами контактирующих с антигеном остатков в HVR. В определенных вариантах осуществления вариантных последовательностей VH и VL, представленных выше, каждая HVR остается неизменной или содержит не более одной, двух или трех аминокислотных замен.
Удобный способ идентификации остатков или областей антитела, которые могут быть мишенями для мутагенеза, называется «аланин-сканирующим мутагенезом», описанным Cunningham and Wells (1989) Science, 244: 1081-1085. В этом способе идентифицируют аминокислотный остаток или группу целевых аминокислотных остатков (например, заряженные аминокислотные остатки, такие как Arg, Asp, His, Lys и Glu) и заменяют нейтральной или отрицательно заряженной аминокислотой (например, аланином или полиаланином) для определения, повлияло ли это на взаимодействие антитела с антигеном. Дополнительные замены можно вносить в аминокислотные положения, демонстрирующие функциональную чувствительность к исходным заменам. В качестве альтернативы или в качестве дополнения используют кристаллическую структуру комплекса антиген-антитело, чтобы идентифицировать точки контакта между антителом и антигеном. Такие контактные остатки и соседние остатки можно использовать для нацеливания или удалять из кандидатов для замены. Можно проводить скрининг вариантов, чтобы определить, имеют ли они необходимые свойства.
Вставки аминокислотных последовательностей включают амино- и/или карбокси-концевые слияния с диапазоном длины от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки внутри последовательности одного или некоторого количества аминокислотных остатков. Примеры концевых вставок включают антитело с N-концевым остатком метионила. Другие инсерционные варианты молекулы антитела включают слияние N- или С-конца антитела с ферментом (например, для терапии ADEPT) или полипептидом, который увеличивает период полужизни антитела в сыворотке крови.
б) Варианты по гликозилинованию
В определенных вариантах осуществления антитело, представленное в данном документе, изменяют с повышением или снижением степени гликозилирования антитела. Добавление или удаление сайтов гликозилирования к антителу удобно осуществлять путем изменения аминокислотной последовательности так, чтобы создать или удалить один или более сайтов гликозилирования.
Если антитело содержит Fc-область, можно изменять присоединенный к ней углевод. Нативные антитела, продуцируемые клетками млекопитающих, обычно содержат разветвленный биантеннарный олигосахарид, который обычно присоединен посредством N-связи к Asn297 СН2-домена Fc-области. См., например, Wright et al. TIBTECH 15: 26-32 (1997). Олигосахарид может содержать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в «стебле» биантеннарной олигосахаридной структуры. В некоторых вариантах осуществления модификации олигосахарида в антителе по настоящему изобретению можно осуществлять с целью создания вариантов антитела с определенными улучшенными свойствами.
В одном варианте осуществления представлены варианты антител, содержащие углеводную структуру с недостаточным количеством фукозы, присоединенной (непосредственно или опосредованно) к Fc-области. Например, количество фукозы в таком антителе может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Количество фукозы определяют путем расчета среднего количества фукозы в сахарной цепи в Asn297 по отношению к сумме всех гликоструктур, прикрепленных к Asn 297 (например, сложных, гибридных и высокоманнозных структур), согласно результатам измерения методом масс-спектрометрии MALDI-TOF, например, как описано в WO 2008/077546. Asn297 относится к остатку аспарагина, расположенному приблизительно в положении 297 в Fc-области (согласно EU-нумерации остатков Fc-области); при этом Asn297 также может быть расположен на приблизительно ±3 аминокислоты выше или ниже положения 297, т.е. между положениями 294 и 300, вследствие незначительных вариаций последовательностей в антителах. Такие варианты по фукозилированию могут иметь улучшенную функцию ADCC. См., например, публикации патентов США №№. US 2003/0157108 (Presta, L.); US 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Примеры публикаций, относящихся к «дефукозилированным» или «фукозодефицитным» вариантам антител, включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; WO 2002/031140; Okazaki et al. J. Mol. Biol. 336: 1239-1249 (2004); Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004). Примеры линий клеток, способных продуцировать дефукозилированные антитела, включают клетки СНО Lecl3, дефицитные по фукозилированию белка (Ripka et al. Arch. Biochem. Biophys. 249: 533-545 (1986); заявка на патент США №US 2003/0157108 Al, Presta, L; и WO 2004/056312 Al, Adams et al, в особенности пример 11), и нокаутные линии клеток, такие как клетки СНО с нокаутом гена альфа-1,6-фукозилтрансферазы, FUT8 (смотрите, например, Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004); Kanda, Y. et al., Biotechnol. Bioeng., 94 (4): 680-688 (2006); и WO2003/085107).
В определенных вариантах осуществления сайт N-гликозилирования в домене СН2 селективного в отношении изоформы антитела к TGFβ по настоящему изобретению мутирован для предупреждения гликозилирования. Например, селективное в отношении изоформы антитело к TGFβ с агликозилированной Fc-областью может быть получено путем мутагенизации аминокислотного остатка в положении 297, как в EU-индексе в домене СН2 Fc-области (например, N297). В определенных вариантах осуществления, гликозилирование в домене СН2 Fc-области может быть устранено путем изменения консенсусного сайта гликозилирования, т.е. Asn в положении 297, за которым следует любой аминокислотный остаток (в случае IgG, Ser человека) и Thr. Сайт гликозилирования может быть изменен аминокислотными вставками, делециями и/или заменами. Например, один или более аминокислотных остатков могут быть вставлены между Asn и Ser, или между Ser, и Thr, чтобы изменить исходный сайт гликозилирования, где вставки не регенерируют сайт N-гликозилирования. В некоторых конкретных вариантах осуществления аминокислотный остаток в положении 297, как в EU-индексе (например, N-гликозилированный сайт в Fc) в домене СН2 IgG Fc человека, мутирован с устранением сайта гликозилирования. В некоторых конкретных вариантах осуществления аминокислотный остаток в положении 297, как в EU-индексе (например, N297) заменен на Gly, Ala, Gln, Asp или Glu. В некоторых конкретных вариантах осуществления аминокислотный остаток в положении 297, как в EU-индексе (например, N297) заменен на Gly или Ala. В других конкретных вариантах осуществления аминокислотный остаток в положении 297, как в EU-индексе (например, N297) заменен на Gly. В некоторых других вариантах осуществления аминокислотный остаток в положении 299, как в EU-индексе, может быть замещен другой аминокислотой, например, Ala, Val или Gly. В определенных конкретных вариантах осуществления мутации, которые приводят к агликозилированному Fc, не влияют на структуру и/или стабильность селективного в отношении изоформы антитела к TGFβ.
В определенных вариантах осуществления селективное в отношении изоформы антитело к TGFβ содержит Fc-область, в которой аминокислотный остаток в положении 297, как в EU-индексе в домене СН2, мутирован. В определенных вариантах осуществления аминокислотный остаток в положении 297, как в EU-индексе, изменен на Gly или Ala, предпочтительно на Gly. В определенных других вариантах осуществления аминокислотный остаток в положении 297, как в EU-индексе, делетирован. В других вариантах осуществления N-гликан, присоединенный к аминокислотному остатку дикого типа в положении 297, как в EU-индексе (например, N297), может быть удален ферментативно, например, путем гликозилирования. Подходящие гликолитические ферменты включают без ограничения пептид-N-гликозидазу (PNGasy).
Дополнительно представлены варианты антител с раздвоенными олигосахаридами, например, в которых диантенарный олигосахарид, прикрепленный к Fc-области антитела, раздвоен GlcNAc. Такие варианты антител могут иметь сниженный уровень фукозилирования и/или улучшенную функцию ADCC. Примеры таких вариантов антител описаны, например, в WO 2003/011878 (Jean-Mairet et al.); патенте США №6602684 (Umana et al.,); и патенте США №2005/0123546 (Umana et al.). Также представлены варианты антител с по меньшей мере одним остатком галактозы в олигосахариде, прикрепленном к Fc-области. Такие варианты антител могут иметь улучшенную функцию CDC. Такие модификации антител описаны, например, в WO 1997/30087 (Patel et al.); WO 1998/58964 (Raju, S.); и WO 1999/22764 (Raju, S.).
в) Варианты по Fc-области
В определенных вариантах осуществления одна или более модификаций аминокислот могут быть введены в Fc-область антитела, представленного в данном документе, за счет чего получают вариант Fc-области. Вариант Fc-области может содержать последовательность человеческой Fc-области (например, Fc-область IgG1, IgG2, IgG3 или IgG4 человека), содержащую аминокислотную модификацию (например, замену) в одном или более положениях аминокислот.
В определенных вариантах осуществления в настоящем изобретении предусмотрен вариант антитела, который обладает некоторыми, но не всеми эффекторными функциями, что делает его желательным кандидатом для вариантов применения, в которых важен период полужизни антитела in vivo, но при этом определенные эффекторные функции (такие как комплемент и ADCC) не нужны или вредны. Цитотоксические анализы могут быть проведены in vitro и/или in vivo для подтверждения восстановления/ослабления активности CDC и/или ADCC. Например, исследования связывания рецептора Fc (FcR) могут быть проведены, чтобы убедиться в том, что антитело не способно к связыванию FcγR (следовательно, вероятно, не обладает ADCC-активностью), но сохраняет способность связывать FcRn. Первичные клетки для опосредования ADCC, NK-клетки, экспрессируют только Fc (RIII, тогда как моноциты экспрессируют Fc (RI, Fc (RII и Fc (RIII. Экспрессия FcR на гемопоэтических клетках обобщенно описана в таблице 3 на странице 464 Ravetch and Kinet, Annu. Rev. Immunol. 9: 457-492 (1991). Неограничивающие примеры способов in vitro анализа для оценки ADCC-активности молекулы, представляющей интерес, описаны в патенте США №5500362 (см., например, Hellstrom, I. et al. Proc. Nat'l Acad. Sci. USA 83: 7059-7063 (1986)) и Hellstrom, I et al., Proc. Nat'l Acad. Sci. USA 82: 1499-1502 (1985); 5821337 (см. Bruggemann, M. et al., J. Exp.Med. 166: 1351-1361 (1987)). В качестве альтернативы можно использовать нерадиоактивные методы анализа (см., например, нерадиоактивный анализ цитотоксичности для проточной цитометрии ACTI™ (CellTechnology, Inc., Маунтин-Вью, Калифорния; и нерадиоактивный анализ цитотоксичности CytoTox 96® (Promega, Мадисон, Висконсин). Эффекторные клетки, подходящие для такого анализа, включают мононуклеарные клетки периферической крови (РВМС) и естественные клетки-киллеры (NK). В качестве альтернативы или в качестве дополнения ADCC-активность молекулы, представляющей интерес, можно оценить в условиях in vivo, например, в модели на животных, такой как та, которая раскрыта в Clynes et al. Proc. Nat'I Acad. Sci. USA 95: 652-656 (1998). Также можно проводить анализ связывания C1q, чтобы подтвердить, что антитело не способно связывать C1q и, следовательно, у него отсутствует CDC-активность. См., например, анализ связывания C1q и С3с в ELISA в WO 2006/029879 и WO 2005/100402. Для оценки активации комплемента можно проводить анализ CDC (см., например, Gazzano-Santoro et al., J. Immunol. Methods 202: 163 (1996); Cragg, M.S. et al., Blood 101: 1045-1052 (2003); и Cragg, M.S. and M.J. Glennie, Blood 103: 2738-2743 (2004)). Определение связывания с FcRn и in vivo клиренса/времени полужизни также может быть осуществлено с применением способов, известных в данной области техники (см., например, Petkova, S.B. et al, Int'l. Immunol. 18 (12): 1759-1769 (2006)).
Антитела со сниженной эффекторной функцией включают антитела с заменой одного или более остатков Fc-области 238, 265, 269, 270, 297, 327 и 329 (патент США №6737056). Такие Fc-мутанты включают Fc-мутантов с заменами в двух или более аминокислотных положениях 265, 269, 270, 297 и 327, включая так называемый Fc-мутант «DANA» с заменой остатков 265 и 297 аланином (патент США №7332581).
Описаны некоторые варианты антител с улучшенным или уменьшенным связыванием с FcR. (См., например, патент США №6737056, WO 2004/056312 и Shields et al., J. Biol. Chem. 9 (2): 6591-6604 (2001).)
В определенных вариантах осуществления вариант антитела содержит Fc-область с одной или более аминокислотными заменами, которые улучшают ADCC, например, с заменами в положениях 298, 333 и/или 334 Fc-области (нумерация остатков согласно EU-системе).
В некоторых вариантах осуществления в Fc-области проведены изменения, которые приводят к изменению (т.е. к улучшению или снижению) связывания C1q и/или комплементзависимой цитотоксичности (CDC), например, как описано в патенте США №6194551, WO 99/51642 и Idusogie et al. J. Immunol. 164: 4178-4184 (2000).
Антитела с увеличенным периодом полужизни и улучшенным связыванием с неонатальным рецептором Fc (FcRn), который отвечает за перенос материнских IgG плоду (Guyer et al., J. Immunol. 117: 587 (1976) и Kim et al., J. Immunol. 24: 249 (1994)), описаны в US 2005/0014934 A1 (Hinton et al.). Эти антитела содержат Fc-домен с одной или более заменами, которые улучшают связывание Fc-области с FcRn. Такие Fc-варианты включают варианты с заменами в одном или более остатках Fc-области: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, замену аминокислотного остатка 434 в Fc-области (патент США №7371826).
См. также Duncan & Winter, Nature 322: 738-40 (1988); патент США №5648260; патент США №5624821; и WO 94/29351 в отношении других примеров вариантов Fc-области.
г) Цистеин-сконструированные варианты антител
В определенных вариантах осуществления может быть желательно создать антитела, сконструированные с использованием цистеина, например, «thioMAb,» в которых один или более остатков замещены остатками цистеина. В конкретных вариантах осуществления замещенные остатки находятся в доступных сайтах антитела. При замене этих остатков цистеином реакционноспособные тиоловые группы располагаются в доступных сайтах антитела и могут использоваться для конъюгации антитела с другими фрагментами, такими как лекарственные фрагменты или фрагменты линкер-лекарственный препарат, для создания иммуноконъюгата, как дополнительно описано в данном документе. В определенных вариантах осуществления цистеином может быть замещен любой из следующих остатков: V205 (нумерация согласно Kabat) легкой цепи; А118 (EU-нумерация) тяжелой цепи; и S400 (EU-нумерация) Fc-области тяжелой цепи. Антитела, сконструированные с использованием остатков цистеина, можно получить, как описано, например, в патенте США №7521541.
д) Производные антител
В определенных вариантах осуществления антитело, представленное в данном документе, можно дополнительно модифицировать путем введения дополнительных небелковых фрагментов, известных в данной области техники и легкодоступных. Фрагменты, подходящие для дериватизации антитела, включают без ограничения водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают без ограниченя полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (гомополимеры или статистические сополимеры), декстран или поли (н-винилпирролидон)полиэтиленгликоль, гомополимеры пропропиленгликоля, сополимеры оксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Пропиональдегид полиэтиленгликоля может иметь преимущества в производстве благодаря своей стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Количество молекул полимера, присоединенных к антителу, может варьироваться и, в случае присоединении более чем одного полимера, они могут представлять собой одинаковые или разные молекулы. В целом, количество и/или тип полимеров, применяемых для дериватизации, можно определить на основании соображений, которые включают без ограничения конкретные свойства или функции антитела, которые нужно улучшить, то, будет ли производное антитела применяться в терапии в определенных условиях и т.д.
В другом варианте осуществления представлены конъюгаты антитела и небелкового компонента, которые можно избирательно нагревать путем облучения. В одном варианте осуществления небелковый фрагмент представляет собой углеродную нанотрубку (Kam et al., Proc. Natl. Acad. Sci. USA 102: 11600-11605 (2005)). Излучение может иметь любую длину волны и включает без ограничения длины волн, которые не вредят обычным клеткам, но которые нагревают небелковый компонент до температуры, при которой погибают клетки, расположенные вблизи конъюгата антитело небелковый компонент.
Б. Рекомбинантные методы и композиции
Антитела можно получать, используя рекомбинантные методы и композиции, например, как описано в патенте США №4816567. В одном варианте осуществления представлены выделенная нуклеиновая кислота, кодирующая селективное в отношении изоформы антитело к TGFβ, описанное в данном документе. Такая нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, легкую и/или тяжелую цепи антитела). В дополнительном варианте осуществления представлены один или более векторов (например, векторов экспрессии), содержащих такую нуклеиновую кислоту. В дополнительном варианте осуществления предложена клетка-хозяин, содержащая такую нуклеиновую кислоту. В одном таком варианте осуществления клетка-хозяин содержит (например, была трансформирована): (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела. В одном варианте осуществления клетка-хозяин является эукариотической, например, клетка яичника китайского хомяка (СНО) или лимфоидная клетка (например, клетка Y0, NS0, Sp20). В одном варианте осуществления представлен способ получения антитела к TGFβ, при этом способ включает культивирование клетки-хозяина, содержащей нуклеиновую кислоту, кодирующую антитело, как представлено выше, в условиях, подходящих для экспрессии антитела, и, необязательно выделение антитела из клетки-хозяина (или культуральной среды клетки-хозяина).
Для рекомбинантного получения селективного в отношении изоформы антитела к TGFβ нуклеиновая кислота, кодирующая антитело, например, как описано выше, является выделенной и вставлена в один или более векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Такая нуклеиновая кислота может быть легко выделена и секвенирована с использованием традиционных процедур (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела).
Подходящие клетки-хозяева для клонирования или экспрессии кодирующих антитела векторов включают прокариотические или эукариотические клетки, описанные в данном документе. Например, антитела могут продуцироваться в бактериях, в частности, если не требуется гликозилирование и Fc-эффекторная функция. Экспрессию фрагментов антитела и полипептидов в бактериях см., например, в патентах США №5648237, 5789199 и 5840523. (См. также Charlton, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ, 2003), стр. 245-254, где описана экспрессия фрагментов антитела в Е. coli). После экспрессии антитело можно выделять из бактериальной клеточной пасты в растворимую фракцию и дополнительно очищать.
Помимо прокариот, эукариотические микроорганизмы, такие как нитевидные грибы или дрожжи, являются подходящими хозяевами для клонирования или экспрессии кодирующих антитела векторов, включая штаммы грибов и дрожжей, пути гликозилирования которых были «гуманизированы», что приводит к получению антитела с частично или полностью человеческим профилем гликозилирования. См. Gerngross, Nat. Biotech. 22: 1409-1414 (2004), и Li et al., Nat. Biotech. 24: 210-215 (2006).
Клетки-хозяева, подходящие для экспрессии гликозилированного антитела, также получают из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы, которые можно использовать в сочетании с клетками насекомых, в частности, для трансфекции клеток Spodoptera frugiperda.
Культуры клеток растений также можно использовать в качестве хозяев. См., например, патенты США №№ 5959177, 6040498, 6420548, 7125978 и 6417429 (в которых описан способ PLANTIBODIES™ для получения антител в трансгенных растениях).
Клетки позвоночных также можно использовать в качестве хозяев. Например, можно использовать линии клеток млекопитающих, которые адаптированы для роста в суспензии. Другими примерами примениых линий клеток-хозяев млекопитающих являются линия CV1 почки обезьяны, трансформированная SV40 (COS-7); линия эмбриональных клеток почки человека (293 или клетки 293, как описано, например, в Graham et al., J. Gen Virol. 36: 59 (1977); клетки почек детенышей хомяка (ВНК); клетки Сертоли мыши (клетки ТМ4, как описано, например, в Mather, Biol. Reprod. 23: 243-251 (1980)); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK; клетки печени крысы баффало (BRL 3А); клетки легких человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); клетки TRI, как описано, например, в Mather et al., Annals N.Y. Acad. Sci. 383: 44-68 (1982); клетки MRC 5; и клетки FS4. Другие подходящие линии клеток-хозяев млекопитающих включают клетки яичника китайского хомяка (СНО), включая DHFR" клетки СНО (Urlaub et al., Proc. Natl. Acad. Sci. USA 77: 4216 (1980)); и линии клеток миеломы, такие как YO, NS0 и Sp2/0. Обзор некоторых линий клеток-хозяев млекопитающих, подходящих для продуцирования антител, см., например, в Yazaki and Wu, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ), pp. 255-268 (2003).
В. Анализы
Идентификацию, скрининг или изучение характеристик селективных в отношении изоформы антител к TGFβ, представленных в данном документе, в отношении их физических/химических свойств и/или видов биологической активности можно проводить с помощью различных анализов, известных в данной области техники, и могут быть определены их кристаллические решетки.
6. Анализы анализа связывания и другие анализы
В одном аспекте антитело по настоящему изобретению исследуют в отношении его антигенсвязывающей активности, например, известными методами, такими как ELISA, вестерн-блоттинг и т.д.
В одном аспекте связывание антител определяют с помощью иммуноферментного анализа. Эта методика хорошо известна в данной области техники. См. Schuurs and VanWeemen, Clinica Chimica Acta, 81: 1-40 (1977). В этой методике ферменты наносят в качестве меток на антиген или антитела для идентификации и локализации иммунореагентов. Любой способ, в котором измеряют степень связывания меченого ферментом антигена или антитела, меченного ферментом, с его иммунореагентом, включен в настоящее изобретение. Разнообразие используемых ферментов, способы связывания ферментов с иммунологическими компонентами, очистка конъюгатов, а также различные принципы и способы анализа хорошо описаны Schuurs и VanWeemen, см. выше.
В одном аспекте аффинность связывания антител определяют с помощью измерения поверхностного плазмонного резонанса (SPR) с помощью прибора BIAcore™-Т200. В этом способе биосенсорный чип серии S СМ5 с белком А применяется для захвата антитела для достижения примерно 100 единиц ответа (RU) на каждую проточную кювету с последующей инъекцией 3-кратных серийных разведений антигена-мишени в соответствующем буфере (например, буфере HBS-EP) при 25°С со скоростью потока 50 мкл/мин. для измерения кинетики. Скорости ассоциации (kon) и скорости диссоциации (koff) могут быть рассчитаны с использованием простой модели связывания Ленгмюра один к одному (оценочное программное обеспечение BIAcore Т200, версия 2.0) для определения равновесной константы диссоциации (KD), рассчитанной как отношение koff/kon.
В другом аспекте конкурентные анализы можно использовать для идентификации антитела, которое конкурирует со специфичным в отношении изоформы антителом к TGFβ, описанным в данном документе. В определенных вариантах осуществления такое конкурирующее антитело связывается с тем же эпитопом (например, линейным или конформационным эпитопом), который связывается с антителами 4А11, 6F12 или 2А10 и их гуманизированными вариантами, описанными в данном документе. Подробные иллюстративные способы картирования эпитопа, с которым связывается антитело, представлены в Morris (1996) "Epitope Mapping Protocols," in Methods in Molecular Biology vol. 66 (Humana Press, Totowa, NJ).
В иллюстративном конкурентном анализе иммобилизованную изоформу TGFβ (например, TGFβ1, TGFβ2 или TGFβ3) инкубируют в растворе, содержащем первое меченое антитело, которое связывается с изоформой TGFβ (например, антитело 4А11, 6F12 или 2А10, или его гуманизированный вариант, например, как показано в таблицах 5, 6, 7, 12, 13, 14, 19, 20, 23 и 24), и второе немеченое антитело, которое тестируют в отношении его способности конкурировать с первым антителом за связывание с изоформой TGFβ.Второе антитело может присутствовать в гибридомном супернатанте. В качестве контроля иммобилизованную форму TGFβ инкубируют в растворе, содержащем первое меченное антитело, но не содержащем второе немеченое антитело. После инкубации в условиях, обеспечивающих связывание первого антитела с изоформой TGFβ, удаляют избыток несвязанного антитела и измеряют количество метки, связанной с иммобилизованной изоформой TGFβ.Если количество метки, связанной с иммобилизованной изоформой TGFβ, существенно снижается в исследуемом образце по сравнению с контрольным образцом, то это указывает на то, что второе антитело конкурирует с первым антителом за связывание с изоформой TGFβ.См. Harlow and Lane (1988) Antibodies: A Laboratory Manual ch.14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY).
В другом примере связывание антител и/или конкуренцию связывания анализируют с использованием радиоиммуноанализа (RIA). RIA представляет собой хорошо известную методику. См., например, Chard, "An Introduction to Radioimmunoassy and Related Techniques", North-Holland Publishing Company, 1978. Можно использовать любой из многих вариаций RIA, например, гомогенный фазовый RIA, гетерогенный или твердофазный RIA, методы с одним антителом или методы «двойного» антитела, а также непосредственные (прямые) или обратные сэндвич-анализы. Особенно предпочтительными являются твердофазные системы, в которых антитело ковалентно связано с нерастворимой подложкой, так что и антитело, и связанный комплекс после инкубации могут быть легко отделены от растворимой свободной фракции. Описан широкий спектр твердофазных носителей, которые включают частицы декстрана, целлюлозы, непрерывные поверхности, такие как диски из полистирола или полипропилена, стенки пластмассовых пробирок, стеклянные диски, стеклянные частицы и т.п.Твердые фазы в виде частиц широко используются для множества различных анализов и включены в настоящее изобретение. Антитела присоединяют к частицам с помощью любого из ряда способов, предназначенных для создания необратимой ковалентной или нековалентной связи между белком и частицей, например, непосредственно или путем активации бромцианом. Другими альтернативами являются использование антител, захваченных в порах полиакриламидного геля или связанных с магнитными частицами. В аналитическую пробирку помещают образец или стандарт, индикатор и соответствующее количество твердофазного антитела, а также детергент для предупреждения агрегации частиц и неспецифического поглощения индикатора. После инкубационного периода, в течение которого пробирки непрерывно перемешивают, твердую фазу осаждают центрифугированием; супернатант удаляют, а твердую фазу подвергают двум или более промывкам буфером для удаления свободного индикатора, захваченного внутри и между частицами. Затем измеряют количество твердой фазы (связанной фракции). Также можно использовать иммунорадиометрические анализы, Chards, см. выше, с. 423.
7. Анализы активности
В одном аспекте представлены анализы для идентификации селективных в отношении изоформы антител к TGFβ, обладающих биологической активностью. Биологическая активность может включать, например, способность связываться и нейтрализовать одну или несколько изоформ TGFβ in vitro и/или in vivo.
В некоторых вариантах осуществления антитело по настоящему изобретению тестируют в отношении такой биологической активности в анализе in vitro. В одном варианте осуществления клетки HEK-Blue™ TGFβ (InvivoGen, Сан-Диего, Калифорния), стабильно экспрессирующие белки TGFβRI, Smad3, Smad4 человека и репортерный ген секретируемой щелочной фосфатазы (SEAP) под контролем минимального промотора β-глобина, слитого с тремя Smad3/4-связывающими элементами (SBE), можно использовать для оценки блокирующей активности антител к TGFβ.
В другом варианте осуществления биологическую активность антител к TGFβ по настоящему изобретению можно оценить с помощью анализа, описанного в Abe et al. Analytical Biochemistry (1994); 216 (2): 276-284. Вкратце, в этом анализе используется анализ блокирования TGFβ на основе клеток эпителия легких норки (MLE), в котором клетки MLE трансфицируют плазмидой, содержащей ген люциферазы ниже промотора ингибитора активатора плазминогена 1. Антитела инкубируют с каждой изоформой TGFβ (PeproTech, Роки-Хилл, Нью-Джерси) при 25°С в течение 1 часа перед добавлением в 4 × 104 репортерных клеток MLE, высеянных в 96-луночный планшет в 75 мкл буфера для анализа (DMEM с высоким содержанием глюкозы с 0,5% инактивированной нагреванием FBS, 2 мМ L-глутамина и 0,5% пенициллин-стрептомицина). Конечная концентрация антитела и TGFβ находится в диапазоне 0,001-200 нМ и 20 пМ соответственно. Планшет инкубируют при 37°С и 5% CO2 в течение 20 часов. Активность люциферазы в клетках определяют путем добавления субстрата люциферазы Firefly в соответствии с рекомендациями производителя (Dual-Luciferase Reporter Assay System, Promega).
Г. Иммуноконъюгаты
В настоящем изобретении также представлены иммуноконъюгаты, содержащие селективное в отношении изоформы антитело к TGFβ в данном документе, конъюгированное с одним или более цитотоксическими средствами, такими как химиотерапевтические средства или лекарственные средства, средства, ингибирующие рост, токсины (например, белковые токсины, ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, или их фрагменты), или радиоактивные изотопы.
В одном варианте осуществления иммуноконъюгат представляет собой конъюгат антитело-лекарственное средство (ADC), в котором антитело конъюгировано с одним или более лекарственными средствами, включая без ограничения майтанзиноид (см. патенты США №№5208020, 5416064 и европейский патент ЕР 0 425 235 В1); ауристатин, такой как лекарственный фрагменты монометилауристатина DE и DF (ММАЕ и MMAF) (см. патенты США №№5635483, 5780588 и 7498298); доластатин; калихеамицин или его производное (см. патенты США №№5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001 и 5877296; Hinman et al., Cancer Res. 53: 3336-3342 (1993); и Lode et al., Cancer Res. 58: 2925-2928 (1998)); антрациклин, такой как дауномицин или доксорубицин (см. Kratz et al., Current Med. Chem. 13: 477-523 (2006); Jeffrey et al., Bioorganic & Med. Chem. Letters 16: 358-362 (2006); Torgov et al., Bioconj. Chem. 16: 717-721 (2005); Nagy et al., Proc. Natl. Acad. Sci. USA 97: 829-834 (2000); Dubowchik et al., Bioorg. & Med. Chem. Letters 12: 1529-1532 (2002); King et al., J. Med. Chem. 45: 4336-4343 (2002); и патент США №6630579); метотрексат; виндезин; таксан, такой как доцетаксел, паклитаксел, ларотаксел, тезетаксел и ортатаксел; трихотецен; и СС1065.
В другом варианте осуществления иммуноконъюгат содержит антитело, описанное в данном документе, конъюгированное с ферментативно активным токсином или его фрагментом, включая без ограничения цепь А дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, цепь А экзотоксина (из Pseudomonas aeruginosa), цепь А рицина, цепь А абрина, цепь А модецина, альфа-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор из момордики харантской, курцин, кротин, ингибитор из Saponaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены.
В другом варианте осуществления иммуноконъюгат содержит антитело, описанное в данном документе, конъюгированное с радиоактивным атомом, с образованием радиоконъюгата. Для получения радиоконъюгатов доступен ряд радиоактивных изотопов. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb 212 и радиоактивные изотопы Lu. Если радиоконъюгат используют в целях обнаружения, он может содержать радиоактивный атом для сцинтиграфических исследований, например, tc99m или 1123, или спиновую метку для ядерного магнитного резонанса (ЯМР) (также известного как магнито-резонансная томография, МРТ), такую как снова йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Конъюгаты антитела и цитотоксического средства можно создавать, используя ряд бифункциональных агентов, связывающих белок, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие, как дисукцинимидилсуберат), альдегиды (такие, как глутаральдегид), бис-азидо соединения (такие, как бис (п-азидобензоил)-гександиамин), производные бис-диазония (такие, как бис (п-диазоний бензоил)-этилендиамин), диизоцианаты (такие, как толуол 2,6-диизоцианат) и бис-активные соединения фтора (такие, как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин на основе рицина можно получать, как описано в Vitetta et al., Science 238: 1098 (1987). Меченая углеродом-14 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) представляет собой иллюстративное хелатирующее средство для конъюгации радионуклида с антителом. См. WO 94/11026. Линкер может представлять собой «расщепляемый линкер», облегчающий высвобождение цитотоксического лекарственного средства в клетке. Например, можно использовать кислотно-лабильный линкер, чувствительный к пептидазе линкер, фотолабильный линкер, диметиловый линкер или дисульфид содержащий линкер (Chari et al., Cancer Res. 52: 127-131 (1992); патент США №5208020).
В контексте данного документа под иммуноконъюгатами или ADC прямо подразумевают, без ограничения конъюгатами, полученными с помощью перекрестно-сшивающих реагентов, помимо прочего, BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB, а также SVSB (сукцинимидил- (4-винилсульфон)бензоат), которые являются коммерчески доступными (например, от Pierce Biotechnology, Inc., Рокфорд, Иллинойс, США).
Д. Способы и композиции для диагностики и обнаружения
В определенных вариантах осуществления любое из селективных в отношении изоформы антител к TGFβ, представленных в данном документе, применимо для обнаружения присутствия одной или более изоформ TGFβ (например, TGFβl, TGFβ2 или TGFβ3) в биологическом образце. Термин «обнаружение», используемый в данном документе, включает количественное или качественное обнаружение. В определенных вариантах осуществления биологический образец содержит клетку или ткань, такую как клетка или ткань легких, почек, сердца, поджелудочной железы, печени или кожи.
В одном варианте осуществления предложено селективное в отношении изоформы антитело к TGFβ для применения в способе диагностики или обнаружения. В еще одном аспекте представлен способ обнаружения присутствия одной или более изоформ TGFβ (например, TGFβ1, TGFβ2 или TGFβ3) в биологическом образце. В определенных вариантах осуществления способ включает приведение биологического образца в контакт с селективным в отношении изоформы антителом к TGFβ, описанным в данном документе, в условиях, допускающих связывание селективного в отношении изоформы антитела к TGFβ с одной или более изоформами TGFβ (например, TGFβ1, TGFβ2, и/или TGFβ3), и определение того, образуется ли комплекс между селективным в отношении изоформы антителом к TGFβ и изоформой (ами) TGFβ.Такой способ может представлять собой in vitro или in vivo способ. В одном варианте осуществления антитело к TGFβ используют для отбора субъектов, подходящих для терапии селективным в отношении изоформы антителом к TGFβ, например в случае, когда изоформа TGFβ является биомаркером для отбора пациентов.
Например, в приведенном ниже примере 4 показано, что экспрессия TGFβ3 заметно повышена у пациентов с SSc по сравнению со здоровым контролем, и, кроме того, в анализе объемной биопсии легких пациентов с идиопатическим легочным фиброзом (IPF) методом кПЦР обнаруживается TGFβ3, и, в меньшей степени, TGFβ2, были экспрессированы при значительно более высоких уровнях по сравнению с легкими контроля, тогда как экспрессия TGFβl была в среднем немного ниже при IPF по сравнению с контролем. Таким образом, в некоторых вариантах осуществления уровни экспрессии изоформ TGFβ, например, в конкретной ткани, можно использовать для диагностики состояния этой ткани у пациента.
В некоторых вариантах осуществления уровень экспрессии TGFβ2 и/или TGFβ3 можно использовать для выявления пациентов, которые, по-видимому, ответят на лечение селективным в отношении изоформы антителом к TGFβ по настоящему изобретению, например, антителом к TGFβ2, антителом к TGFβ2/3 или антителом к TGFβ3, описанным в данном документе. В некоторых вариантах осуществления определено, что пациент с большей вероятностью ответит на лечение селективным в отношении изоформы антителом к TGFβ по настоящему изобретению, если у пациента повышен исходный уровень (до лечения селективным в отношении изоформы антителом к TGFβ) по сравнению с эталонным уровнем (например, у здорового или контрольного субъекта) одного или более или всех биомаркеров, выбранных из TGFβ2, TGFβ3, периостина (POSTN) и олигомерного матриксного белка хряща (СОМР). В некоторых вариантах осуществления такому пациенту ранее вводили средство терапии на основе ингибитора IL-6, и у него недостаточный, минимальный или отсутствует ответ на указанное средство терапии. В некоторых таких вариантах осуществления ингибитор IL-6 представляет собой тоцилизумаб.
В некоторых вариантах осуществления уровни биомаркеров (например, TGFβ2, TGFβ3, СОМР и/или POSTN) в сыворотке крови количественно оценивают, чтобы определить, может ли пациент ответить на лечение селективным в отношении изоформы антителом к TGFβ.В некоторых вариантах осуществления пациента выбирают для лечения, если уровень одного или нескольких биомаркеров (например, TGFβ2, TGFβ3, СОМР и/или POSTN) в сыворотке крови повышен/повышены на исходном уровне по сравнению с контрольным уровнем (например, у здорового субъекта). В дополнительных вариантах осуществления пациенту, у которого определена вероятность ответа на лечение селективным в отношении изоформы антитела к TGFβ по настоящему изобретению и/или который выбран для лечения, вводят селективное в отношении изоформы антитело к TGFβ, например, в терапевтически эффективном количестве.
В некоторых вариантах осуществления уровни биомаркеров (например, TGFβ2, TGFβ3, СОМР и/или POSTN) в сыворотке крови количественно оценивают, чтобы определить, что пациент с меньшей вероятностью будет испытывать улучшения, если пациенту проводят лечение, включающее терапевтическое средство, отличное от селективного в отношении изоформы антитела к TGFβ по настоящему изобретению.
Специалистам в данной области техники хорошо известно измерение уровней биомаркеров в сыворотке крови, таких как, например, TGFβ2, TGFβ3, POSTN и СОМР. Последовательности изоформ TGFβ были представлены выше. Нуклеиновые и аминокислотные последовательности СОМР и POSTN известны в данной области техники, и для справки ниже приведены иллюстративные аминокислотные последовательности:
Каноническая аминокислотная последовательность POSTN человека представляет собой:


Последовательности нуклеиновых кислот и другие вариантные аминокислотные последовательности, кодирующие POSTN человека, известны в данной области техники и могут быть найдены в базе данных Uniprot, доступной в World Wide Web по адресу uniprot.org/uniprot/Q15063 для ID Q15063 в Uniprot.
Каноническая аминокислотная последовательность СОМР человека представляет собой:

Последовательности нуклеиновых кислот и другие вариантные аминокислотные последовательности, кодирующие СОМР человека, известны в данной области техники и могут быть найдены в базе данных Uniprot, доступной в World Wide Web по адресу uniprot.org/uniprot/P49747 для ID Р49747 в Uniprot.
Уровни POSTN и СОМР в сыворотке крови можно определить любым подходящим способом, известным в данной области техники, например, ELISA, микроматричный анализ или ПЦР. В другом варианте осуществления уровни белков POSTN и СОМР можно определить с помощью специфических иммуноанализов на основе IMPACT, например, как описано ниже в примере 4. Например, цельную кровь можно собрать у субъекта или пациента в соответствии с подходящими способами в данной области техники, дать ей свернуться и затем центрифугировать. Уровни биомаркеров в сыворотке крови определяют с помощью специфических иммуноанализов на основе IMPACT (Roche Diagnostics, Пенцберг, Германия). Вкратце, образцы можно инкубировать с чипом IMPACT, покрытым биотинилированными фрагментами антител F (ab')2, специфичными к белку-мишени, представляющему интерес (например, биомаркеру, такому как, например, TGFβ2, TGFβ3, СОМР или POSTN). После этапов промывки добавляют и инкубируют специфичные для биомаркеров антитела для обнаружения, меченые дигоксигенином. После промывки добавляли флуоресцентный конъюгат антидигоксигенин-латекс, и специфически связанную флуоресцентную метку можно было обнаружить с помощью детекторной камеры с зарядовой связью. Испускаемая флуоресценция преобразуется в интенсивность сигнала с использованием стандартной кривой, состоящей из стандартов рекомбинантных белков (R&D Systems, Миннеаполис, Миннесота, США) и специального программного обеспечения (Roche Diagnostics).
Нуклеиновая кислота, в соответствии с любым из способов, описанных в данном документе, может представлять собой РНК, транскрибированную из геномной ДНК, или кДНК, созданную из РНК. Нуклеиновая кислота может быть получена от позвоночного, например млекопитающего. Считается, что нуклеиновая кислота «получена из» определенного источника, если она получена непосредственно из этого источника или если она является копией нуклеиновой кислоты, обнаруженной в этом источнике.
Нуклеиновая кислота включает копии нуклеиновой кислоты, например, копии, полученные в результате амплификации. Амплификация может быть желательной в некоторых случаях, например, для получения требуемого количества материала для обнаружения вариаций. Затем ампликоны могут быть подвергнуты способу обнаружения вариаций, такому как описанные ниже, для определения экспрессии определенных генов.
Микроматричный анализ представляет собой мультиплексный способ, в котором обычно используется набор из тысяч зондов нуклеиновых кислот для гибридизации, например, с образцом кДНК или кРНК в условиях высокой жесткости. Гибридизация зонд-мишень обычно обнаруживается и количественно оценивается путем обнаружения меченных флуорофором, серебром или хемилюминесценцией мишеней для определения относительного количества последовательностей нуклеиновых кислот в мишени. В типичных микроматрицах зонды прикрепляются к твердой поверхности ковалентной связью с химической матрицей (посредством эпоксисилана, аминосилана, лизина, полиакриламида или других). Твердая поверхность представляет собой, например, стекло, кремниевый чип или микроскопические гранулы. Коммерчески доступны различные микрочипы, в том числе производимые, например, компаниями Affymetrix, Inc. и Illumina, Inc.
В другом варианте осуществления уровни биомаркеров в сыворотке крови могут быть измерены с использованием версии для клинических испытаний иммунологического анализа Elecsys (Roche Diagnostics) POSTN, предназначенного для использования на Cobas е601 (Roche Diagnostics).
В некоторых вариантах осуществления представлен способ определения груза заболевания и/или прогноза у субъекта, у которого имеется фиброзное заболевание. В некоторых неограничивающих вариантах осуществления фиброзное состояние представляет собой SSc или IPF. Способ может включать определение уровней POSTN и/или СОМР в сыворотке периферической крови субъекта.
Способы определения уровней белков известны в данной области техники. Уровни экспрессии белков можно определить в образцах цельной крови, плазмы крови или сыворотки крови. В данной области техники известны различные способы определения уровней экспрессии белка в таких биологических образцах, включая различные способы иммуноанализа. Ранее был описан широкий спектр методик иммуноанализа, см., например, патенты США №№4016043, 4424279 и 4018653. Они включают как односайтовые, так и двусайтовые или «сэндвич»-анализы неконкурентного типа, а также традиционные конкурентные анализы связывания. Эти анализы также включают прямое связывание меченого антитела с биомаркером-мишенью.
Сэндвич-анализы являются одними из наиболее применимых и часто используемых анализов. Существует ряд вариаций методики сэндвич-анализа, и предполагается, что все они охватываются настоящим изобретением. Вкратце, в типичном прямом анализе немеченое антитело иммобилизуют на твердой подложке, а тестируемый образец приводят в контакт со связанной молекулой. После подходящего периода инкубации в течение периода времени, достаточного для образования комплекса антитело-антиген, затем добавляют второе антитело, специфическое к антигену, меченое репортерной молекулой, способной давать детектируемый сигнал, и инкубируют, давая время, достаточное для образования другого комплекса антитело-антиген-меченое антитело. Любой непрореагировавший материал смывается, и присутствие антигена определяется по наблюдению за сигналом, производимым репортерной молекулой. Результаты могут быть либо качественными, путем простого наблюдения за видимым сигналом, либо могут быть оценены количественно путем сравнения с контрольным образцом, содержащим известные количества биомаркера.
В других вариантах осуществления наличие повышенных уровней одной или нескольких изоформ TGFβ можно использовать в диагностических целях для определения наличия или исходного наличия патологий, возникающих в результате нарушения, связанного с TGFβ (например, фиброза или рака). Например, иммуноанализы с использованием антител к TGFβ представляют такой диагностический тест. Различные форматы таких анализов доступны и хорошо известны специалистам в данной области техники, включая RIA, ELISA и иммунофлуоресценцию. См. в целом Ruoslahti et al., М. Enz., 82: 803-831 (1982). В качестве альтернативы для той же цели можно использовать зонды нуклеиновых кислот для обнаружения и количественного определения мРНК TGFβ.Такие способы также хорошо известны в данной области техники.
В некоторых вариантах осуществления за субъектом наблюдают в динамике, и начальные уровни биомаркеров в сыворотке крови сравнивают с уровнями в более поздние временные точки во время лечения, чтобы оценить, изменяется ли груз заболевания субъекта и/или улучшается или ухудшается прогноз субъекта. В некоторых вариантах осуществления состояние и/или прогноз субъекта с меньшей вероятностью улучшатся, если уровень экспрессии одного или обоих POSTN и СОМР повышен по сравнению со здоровым контролем или эталонным стандартом, или по сравнению с уровнями POSTN или СОМР субъекта в начале или до начала схемы лечения заболевания, связанного с TGFβ, например, IPF или SSc.
В некоторых аспектах способ идентификации субъекта как субъекта с высокой активностью TGFβ и/или субъекта, на которого, вероятно, лечение ингибитором изоформы TGFβ, например, селективным в отношении изоформы антителом к TGFβ по настоящему изобретению (например, антителом к TGFβ2, антителом к TGFβ2/3 или антителом к TGFβ3, описанными в данном документе) будет оказывать благоприятное действие. В некоторых аспектах у субъекта имеется SSc. В некоторых аспектах у субъекта имеется IPF. В некоторых аспектах способ включает определение уровней экспрессии одного или более генов наборе сигнатур 18 генов со следующими генами: PRSS23, PXDN, COL8A1, COL6A3, SERPINE2, TNC, СОМР, THBS1, COL11A1, COL1A1, COL5A2, COL1A2, COL4A1, COL4A2, SFRP4, ALPK2, COL5A1, TAGLN. Гены набора сигнатур 18 генов описаны в таблице 2 ниже. В некоторых аспектах способ включает идентификацию субъекта как субъекта с высокой активностью TGFβ и/или с вероятностью того, что лечение ингибитором изоформы TGFβ окажет благоприятное действие, если уровни 1 или более, 2 или более, 3 или более, 4 или более, 5 или более, 6 или более, 7 или более, 8 или более, 9 или более, 10 или более, 11 или более, 12 или более, 13 или более, 14 или более, 15 или более, 16 или более, 17 или более или всех 18 генов в наборе сигнатур 18 генов повышены по сравнению с уровнями гена в контрольном наборе от здорового субъекта (например, эталонным значением). В одном аспекте способ включает идентификацию субъекта как субъекта с высокой активностью TGFβ и/или субъекта, на которого, вероятно, лечение ингибитором изоформы TGFβ окажет благоприятное действие, если уровень каждого и всех 18 генов в наборе сигнатур 18 генов повышен по сравнению с уровнями соответствующего гена в контрольном наборе от здорового субъекта.

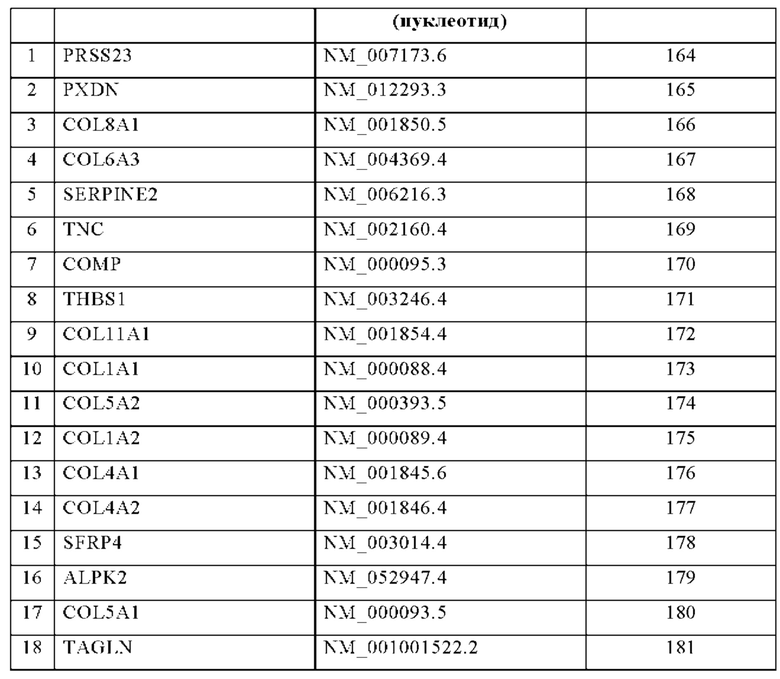
В конкретном варианте осуществления субъект идентифицируется как субъект с высокой активностью TGFβ и/или с вероятностью того, что лечение ингибитором изоформы TGFβ окажет благоприятное действие, если уровни одного или более генов, выбранных из SERPINE1, COL1A1, COL1A2, и COL3A1, повышены по сравнению с уровнем соответствующего гена в контрольном наборе от здорового субъекта (например, эталонным значением). Гены этого набора генов более подробно описаны в таблице 3 ниже. В некоторых аспектах способ включает идентификацию субъекта как субъекта с высокой активностью TGFβ и/или с вероятностью того, что лечение ингибитором изоформы TGFβ окажет благоприятное действие, если уровни 1 или более, 2 или более, 3 или более или всех 4 генов SERPINE1, COL1A1, COL1A2 и COL3A1 повышены по сравнению с уровнем соответствующего гена в контрольном наборе от здорового субъекта (например, эталонным значением).
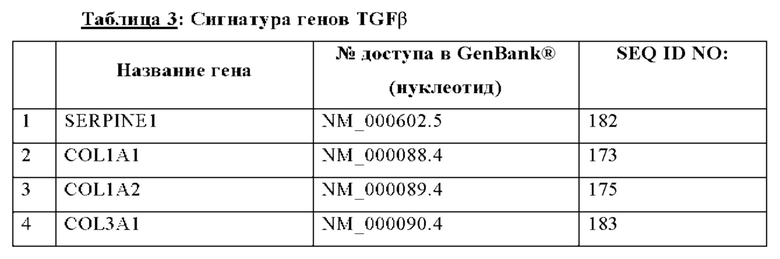
В некоторых вариантах осуществления вышеуказанных способов уровень экспрессии гена считается повышенным, если повышение его экспрессии по сравнению с контролем или эталонным уровнем гена является статистически значимым. Способы определения статистической значимости хорошо известны в данной области техники. В качестве неограничивающего примера статистическую значимость можно определить с использованием, например, непарного двустороннего t-критерия Стьюдента. В других вариантах осуществления уровень экспрессии гена считается повышенным, если повышение его экспрессии по сравнению с контролем или эталонным уровнем гена происходит по меньшей мере в 2 раза, по меньшей мере в 3 раза или по меньшей мере в 4 раза.
В других аспектах представлены способы мониторинга ответа на лечение субъекта ингибитором TGFβ, таким как, например, антитело к TGFβ2 и/или антитело к TGFβ3, описанные в данном документе. Способ включает определение уровня экспрессии одного или более TGFβ-индуцируемых генов, выбранных из PRSS23, PXDN, COL8A1, COL6A3, SERPINE2, TNC, СОМР, THBS1, COL11A1, COL1A1, COL5A2, COL1A2, COL4A1, COL4A2, SFRP4, ALPK2, COL5A1, TAGLN, и/или одного или более генов, выбранных из SERPINE1, COL1A1, COL1A2 и COL3A1. В некоторых вариантах осуществления способ дополнительно включает прекращение лечения ингибитором TGFβ, если уровни экспрессии одного или более генов, выбранных из PRSS23, PXDN, COL8A1, COL6A3, SERPINE2, TNC, СОМР, THBS1, COL11A1, COL1A1, COL5A2, COL1A2, COL4A1, COL4A2, SFRP4, ALPK2, COL5A1, TAGLN, и/или одного или более генов, выбранных из SERPINE1, COL1A1, COL1A2 и COL3A1, не снижены по сравнению с соответствующим уровнем в более ранней временной точке, например, до или в начале лечения ингибитором TGFβ.
В другом варианте осуществления представлен способ выбора субъекта для лечения ингибитором TGFβ, таким как, например, антитело к TGFβ2 и/или антитело к TGFβ3, описанные в данном документе. Способ может включать определение уровня одного или обоих POSTN и СОМР в сыворотке крови у субъекта и введение субъекту ингибитора TGFβ, если уровни одного или обоих POSTN и СОМР в сыворотке крови повышены по сравнению с контрольным уровнем у здорового субъекта. В некоторых вариантах осуществления ингибитор TGFβ представляет собой антитело к TGFβ3, например, антитело 2А10 или его гуманизированный вариант, например, как изображено на Фиг. 12-16 или представлено в таблицах 5-7 в примере 1 ниже.
В некоторых вариантах осуществления способ идентификации субъекта как субъекта с высокой активностью TGFβ и/или субъекта, на которого, вероятно, лечение ингибитором изоформы TGFβ окажет благоприятное действие, и/или выбора субъекта для лечения ингибитором TGFβ включает введение средства терапии субъекта. В некоторых вариантах осуществления средство терапии предназначено для лечения нарушения, связанного с TGFβ, например, как описано в данном документе. В некоторых вариантах осуществления средство терапии предназначено для лечения фиброзного состояния, например, как описано в данном документе. В некоторых вариантах осуществления средство терапии включает введение антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3, описанных в данном документе. В предпочтительном варианте осуществления средство терапии включает введение антитела к TGFβ3, описанного в данном документе, например, выделенного антитела к TGFβ3, где указанное антитело селективно нейтрализует TGFβ3, и при этом антитело характеризуется одним или более из следующих свойств: (а) антитело к TGFβ3 связывается со шпилечной областью бета6/бета7 TGFβ3; (б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3; (в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFβR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3; (г) антитело к TGFβ3 контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека; (д) антитело к TGFβ3 контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFJ33 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи; (е) изоформная селективность антитела к TGFβ3 к TGFβ3 по сравнению с TGFβ1 достигается за счет прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека); (ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается за счет прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека); (и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 к TGFβ; (к) антитело к TGFP3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 к TGFβ; (л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; (м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; (н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом CAT-192 к TGFβl; (п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3; (р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -НЗ, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9; (с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, 1388, L389, Y391, V392, G393, R394, Р396, K397 и V398 TGFβ3 человека; и (т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3.
В некоторых аспектах вышеуказанных вариантов осуществления антитело к TGFβ3 содержит пару VH/VL, где пара VH/VL содержит аминокислотные последовательности (соответственно), выбранные из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9). В некоторых вариантах осуществления антитело к TGFβ3 содержит пару VH/VL, содержащую аминокислотные последовательности (соответственно) SEQ ID NO: 52/36, или аминокислотные последовательности (соответственно) SEQ ID NO: 51/36, или SEQ ID NO: 55/54 или аминокислотные последовательности (соответственно) SEQ ID NO: 53/36 или SEQ ID NO: 57/56. В некоторых вариантах осуществления антитело к TGFβ3 содержит пару VH/VL, содержащую аминокислотные последовательности (соответственно) SEQ ID NO: 57/56.
В некоторых аспектах вышеуказанных вариантов осуществления антитело к TGFβ3 содержит полную пару H/L-цепей, при этом полная пара H/L-цепей содержит аминокислотные последовательности (соответственно), выбранные из группы SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.l), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.6), SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8), SEQ ID NO: 72/58 (v2.9), или аминокислотные последовательности (соответственно) SEQ ID NO: 74/58, или аминокислотные последовательности (соответственно) SEQ ID NO: 73/58 или SEQ ID NO: 77/76, или аминокислотные последовательности (соответственно) SEQ ID NO: 75/58 или SEQ ID NO: 79/78. В некоторых аспектах антитело к TGFβ3 содержит полную пару H/L-цепей, содержащую аминокислотные последовательности (соответственно) SEQ ID NO: 79/78.
В некоторых аспектах вышеуказанных вариантов осуществления средство терапии включает введение антитела к TGFβ2/3, описанного в данном документе, например, выделенного антитела к TGFβ2/3, где указанное антитело селективно нейтрализует TGFβ2 и TGFβ3, и при этом антитело характеризуется одним или более из следующих свойств: (а) селективность антитела к TGFβ2/3 в отношении TGFβ2 и TGFβ3 по сравнению с TGFβ1 человека в отношении селективной нейтрализации достигается посредством прямого контакта антигенсвязывающего домена антитела с аминокислотным остатком Е373 TGFβ2 или TGFβ3 (нумерация TGFβ2 человека); (б) антитело к TGFβ2/3 нейтрализует TGFβ2 и/или TGFβ3 посредством аллостерического механизма; (в) антитело к TGFβ2/3 индуцирует конформационные изменения в гомодимере TGFβ2; (г) антитело к TGFβ2/3 индуцирует конформационное изменение в гомодимере TGFβ2, где конформационное изменение включает совместное сжатие двух мономеров на несколько градусов; (д) антитело к TGFβ2/3 представляет собой двухвалентное антитело или одновалентное антитело; (е) антитело к TGFP2/3 содержит (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 15; (ж) антитело к TGFβ2/3 связывается с гомодимером TGFβ2, при этом гомодимер TGFβ2 имеет первый и второй мономер TGFβ2, и при этом антитело к TGFβ2/3 содержит антигенсвязывающий домен, который непосредственно контактирует с (i) аминокислотными остатками V313, Q314, D315, R320, L322, Y323, R328, D329, F345 и А347 первого мономера TGFβ2, и (ii) аминокислотными остатками N368, Т369, 1370, N371, Р372, Е373, А374, S375, А376 и S377 второго мономера TGFβ2 (нумерация TGFβ2 человека); (и) антитело к TGFβ2/3, как и в (ж), где антигенсвязывающий домен находится в пределах 5 ангстрем от аминокислотных остатков TGFβ2 и/или TGFβ3; (к) при этом антитело к TGFβ2/3 связывается с тем же эпитопом TGFβ3, что и в (ж); и (л) антитело к TGFβ2/3 не нейтрализует TGFβ2 и/или TGFβ3 в одноплечевой форме.
В некоторых аспектах вышеуказанных вариантов осуществления антитело к TGFβ2/3 содержит пару VH/VL, при этом пара VH/VL содержит аминокислотные последовательности (соответственно), выбранные из SEQ ID NO: 27/26 (4А11 кролика), SEQ ID NO: 81/80 (v1), SEQ ID NO: 81/82 (v2), SEQ ID NO: 83/80 (v3), SEQ ID NO: 83/82 (v4), SEQ ID NO: 81/84 (v5), SEQ ID NO: 81/85 (v6), SEQ ID NO: 83/84 (v7), SEQ ID NO: 86/84 (v7/1), SEQ ID NO: 87/84 (v7.2), SEQ ID NO: 88/84 (v7.3), SEQ ID NO: 83/89 (v7.4), SEQ ID NO: 83/90 (v7.5), SEQ ID NO: 83/91 (v7.6), SEQ ID NO: 83/92 (v7.7), SEQ ID NO: 93/84 (v7.8), SEQ ID NO: 94/84 (v7.9), SEQ ID NO: 95/84 (v7.10), SEQ ID NO: 96/84 (v7.11), SEQ ID NO: 97/84 (v7.12), SEQ ID NO: 98/84 (v7.13), SEQ ID NO: 99/84 (v7.14), SEQ ID NO: 100/84 (v7.15), SEQ ID NO: 102/101 (v7.16), SEQ ID NO: 103/101 (v7.17), SEQ ID NO: 104/101 (v7.18), SEQ ID NO: 105/101 (v7.19) и SEQ ID NO: 83/85 (v8).
В некоторых аспектах вышеуказанных вариантов осуществления антитело к TGFP2/3 содержит полную пару H/L-цепей, при этом полная пара H/L-цепей содержит аминокислотные последовательности (соответственно), выбранные из SEQ ID NO: 32/33 (4А11 кролика), SEQ ID NO: 107/106 (v1), SEQ ID NO: 107/108 (v2), SEQ ID NO: 109/106 (v3), SEQ ID NO: 109/108 (v4), SEQ ID NO: 107/110 (v5), SEQ ID NO: 107/111 (v6), SEQ ID NO: 109/110 (v7), SEQ ID NO: 112/110 (v7.1), SEQ ID NO: 113/110 (v7.2), SEQ ID NO: 114/110 (v7.3), SEQ ID NO: 114/115 (v7.4), SEQ ID NO: 114/116 (v7.5), SEQ ID NO: 114/117 (v7.6), SEQ ID NO: 114/118 (v7.7), SEQ ID NO: 119/110 (v7.8), SEQ ID NO: 120/110 (v7.9), SEQ ID NO: 121/110 (v7.10), SEQ ID NO: 122/110 (v7.11), SEQ ID NO: 123/110 (v7.12), SEQ ID NO: 124/110 (v7.13), SEQ ID NO: 125/110 (v7.14), SEQ ID NO: 126/110 (v7.15), SEQ ID NO: 127/186 (v7.16), SEQ ID NO: 128/186 (v7.17), SEQ ID NO: 129/186 (v7.18), SEQ ID NO: 130/186 (v7.19) и SEQ ID NO: 114/111 (v8).
В некоторых аспектах вышеуказанных вариантов осуществления средство терапии включает введение антитела к TGFβ2, описанного в данном документе, например, выделенного антитела к TGFβ2, где антитело содержит: (a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 18; и (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21. В некоторых аспектах антитело селективно нейтрализует TGFβ2. В некоторых аспектах антитело к TGFβ2 имеет одно или более из следующих свойств: (а) селективно нейтрализует TGFJ32; (б) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ; (в) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ; (г) имеет сниженную токсичность у грызунов по сравнению с антителом 1D11 ко всем видам TGFβ; (д) имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; и/или (е) имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом.
В некоторых аспектах вышеуказанных вариантов осуществления антитело к TGFP2 содержит пару VH/VL, при этом пара VH/VL содержит аминокислотные последовательности (соответственно), выбранные из SEQ ID NO: 25/24 (6F12 кролика), SEQ ID NO: 132/131 (v1), SEQ ID NO: 132/133 (v1.l), SEQ ID NO: 132/134 (v1.2), SEQ ID NO: 132/135 (v1.3), SEQ ID NO: 132/136 (v1.4), SEQ ID NO: 132/137 (v1.5), SEQ ID NO: 138/131 (v1.6), SEQ ID NO: 139/131 (v1.7), SEQ ID NO: 140/131 (v1.8), SEQ ID NO: 141/131 (v1.9), SEQ ID NO: 142/131 (v2), SEQ ID NO: 132/143 (v3) и SEQ ID NO: 142/144 (v4).
В некоторых аспектах вышеуказанных вариантов осуществления антитело к TGFβ2 содержит полную пару H/L-цепей, при этом полная пара H/L-цепей содержит аминокислотные последовательности (соответственно), выбранные из SEQ ID NO: 31/30 (6F12 кролика), SEQ ID NO: 146/145 (v1), SEQ ID NO: 146/147 (v1.1), SEQ ID NO: 146/148 (v1.2), SEQ ID NO: 146/149 (v1.3), SEQ ID NO: 146/150 (v1.4), SEQ ID NO: 146/151 (v1.5), SEQ ID NO: 152/145 (v1.6), SEQ ID NO: 153/145 (v1.7), SEQ ID NO: 154/145 (v1.8), SEQ ID NO: 155/145 (v1.9), SEQ ID NO: 156/145 (v2), SEQ ID NO: 146/157 (v3) и SEQ ID NO: 156/158 (v4).
Иллюстративные состояния и нарушения, которые можно диагностировать и/или лечить в соответствии с настоящим изобретением, например, как описано выше, включают нарушения, связанные с TGFβ, например, заболевания, характеризующиеся накоплением внеклеточного матрикса, заболевания, вызванные циркулирующим TGFβ или TGFβ, активированным в локальном участке, включая одну или более изоформ TGFβ, состояния, вызванные подавлением иммунной системы вследствие эндогенного продуцирования TGFβ, формы острой иммунной недостаточности, возникающие в результате тяжелых травм, ожогов и заболеваний, таких как вирусные или бактериальные инфекции, полиорганные системные заболевания вследствие продуцирования или сверхпродуцирования TGFβ, и опухоли, продуцирующие TGFβ. В некоторых вариантах осуществления нарушения, которые можно диагностировать с помощью селективного в отношении изоформы антитела к TGFβ по настоящему изобретению, включают фиброзное заболевание, например, описанное в данном документе. В некоторых вариантах осуществления фиброзное заболевание представляет собой фиброзное нарушение легких, печени, сердца, почек, поджелудочной железы, глаз и/или кожи. В некоторых вариантах осуществления фиброз представляет собой фиброзное состояние или нарушение, описанные в разделе Ж ниже.
В некоторых вариантах осуществления нарушение, связанное с TGFβ, которое можно диагностировать с помощью антитела по настоящему изобретению, диагностируют с помощью одного или более антител 4А11, 6F12 и 2А10 или их гуманизированного варианта, например, как описано в данном документе, например, на Фиг. 12-16 и 18-24.
В определенных вариантах осуществления представлены меченые селективные в отношении изоформы антитела к TGFβ, например, антитела, описанные выше. Метки включают, но не ограничиваются этим, метки или фрагменты, выявляемые прямым образом (например, флуоресцентные, хромофорные, электронноплотные, хемилюминесцентные и радиоактивные метки), а также фрагменты, такие как ферменты или лиганды, выявляемые непрямым образом, например, посредством ферментативной реакции или молекулярного взаимодействия. Иллюстративные метки включают без ограничения радиоактивные изотопы 32Р, 14С, 125I, 3Н и 131I, флуорофоры, такие как хелаты редкоземельных металлов или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например, люциферазу светляков и бактериальную люциферазу (патент США №4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизозим, сахаридооксидазы, например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфат дегидрогеназу, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, сопряженные с ферментом, который использует пероксид водорода для окисления предшественника красителя, такой как HRP, лактопероксидазу или микропероксидазу, биотин/авидин, спиновые метки, бактериофаговые метки, стабильные свободные радикалы и т.п.
Е. Фармацевтические составы
Фармацевтические составы селективного в отношении изоформы антитела к TGFβ, описанного в данном документе, готовят, смешивая такое антитело, имеющее необходимую степень чистоты, с одним или более необязательными фармацевтически приемлемыми носителями (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)) в форме лиофилизированных составов или водных растворов. Фармацевтически приемлемые носители в общем случае являются нетоксичными для реципиентов в применяемых дозировках и концентрациях и включают без ограничения: буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензил аммония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол; бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (содержащие менее 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; металлсодержащие комплексы (например, комплексы Zn-белок) и/или неионные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ). Иллюстративные фармацевтически приемлемые носители в данном документе дополнительно включают средства для диспергирования лекарственных средств в интерстициальном пространстве, такие как растворимые нейтрально-активные гликопротеины гиалуронидазы (sHASEGP), например, растворимые гликопротеины гиалуронидазы РН-20 человека, такие как rHuPH20 (HYLENEX, Baxter International, Inc.). Некоторые иллюстративные sHASEGP и способы их применения, включая rHuPH20, описаны в патентных публикациях США №№2005/0260186 и 2006/0104968. В одном аспекте sHASEGP комбинируют с одной или более дополнительными гликозаминогликаназами, такими как хондроитиназы.
Иллюстративные лиофилизированные составы антител описаны в патенте США №6267958. Водные составы антител включают описанные в патенте США №6171586 и WO 2006/044908, причем последние составы содержат гистидин-ацетатный буфер.
Состав, представленный в данном документе, также может содержать более одного активного ингредиента, если это необходимо для конкретного показания, которое лечат, предпочтительно те, которые обладают дополняющими видами активности и не оказывают отрицательного влияния друг на друга. Например, может быть желательно дополнительно предоставить терапевтическое средство, выбранное из пирфенидона, нинтеданиба, микофенилатмофетила, ингибитора IL-6 (например, тоцилизумаб, см., например, патент США №9539263, сарилумаб), антитело к CTFG (например, FG-3019), ингибитор аутотаксина, ингибитор JAK, ингибитор IL-11 и сывороточный амилоид Р (РТХ2). (См. Raghu et al. European Respiratory Journal 2012 40: 2819) (FG-309, Ninou et al. (2018) Front Med (Lausanne); 5: 180 (ингибиторы аутотаксина).) Такие активные ингредиенты приемлемо присутствуют в комбинации в количествах, которые эффективны для предполагаемой цели.
Активные ингредиенты также могут быть заключены в микрокапсулах, полученных, например, с помощью методик коацервации или путем межфазной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли (метилметакрилатные) микрокапсулы, соответственно, в коллоидных системах для доставки лекарственных средств (например, липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие методики описаны в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Можно получать препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело, причем матрицы находятся в виде формованных изделий, например, пленок или микрокапсул.
Составы, предназначенные для введения in vivo, в общем случае являются стерильными. Стерильность можно легко обеспечить, например, путем фильтрации через стерильные фильтровальные мембраны.
Ж. Терапевтические способы и композиции
В настоящее время обнаружено, что селективные в отношении изоформы антитела к TGFβ, описанные в данном документе, применимы в терапевтических способах. TGFβ долгое время считался ключевым медиатором патологического фиброза, но его разнообразные гомеостатические функции создают проблемы для разработки терапевтических ингибиторов TGFβ с приемлемым терапевтическим индексом. В то время как три изоформы TGFβ способны опосредовать аналогичные сигнальные активности в отношении клеток-мишеней, их отличающиеся LAP-домены и паттерны экспрессии предполагают уникальные роли для каждой изоформы, что подтверждается генетическими доказательствами потери функции у мышей и людей. Не намереваясь ограничиваться какой-либо одной конкретной теорией или механизмом действия, в данном документе обнаружено, что пороги и механизмы активации латентных TGFβ2 и TGFβ3 отличаются от таковых для TGFβ1 (см., например, пример 14); TGFβ2 и TGFβ3, в отличие от TGFβ1, экспрессируются при повышенных уровнях в фиброзной ткани человека и в мышиных моделях легких и печени; и селективное в отношении изоформы терапевтическое ингибирование TGFβ2 и TGFβ3 с помощью мощного аллостерического антитела может улучшить экспериментальный фиброз in vivo. Эти данные подтверждают селективное в отношении изоформы ингибирование TGFβ2 и/или TGFβ3 в качестве терапевтической стратегии для пациентов с хроническими фиброзными нарушениями. Например, в приведенном ниже примере 4 показано, что уровни экспрессии TGFβ2 и/или TGFβ3 повышены в фиброзной ткани человека, а экспрессия TGFβ3 высоко коррелирует с передачей сигналов TGFβR при IPF и SSc, двух различных фиброзных нарушениях, и в примере 5, описывающем мышиную модель индуцированного блеомицином фиброза легких, в которой нейтрализация TGFβ2 с помощью 6F12, нейтрализация TGFβ3 с помощью 2А10 и нейтрализация как TGFβ2, так и TGFβ3 либо комбинацией антител 6F12+2А10, либо монотерапией антителом 4А11 с двойной специфичностью (антитело к TGFβ2/3) снижали синтез нового коллагена. Кроме того, в биоптатах кожи, взятых на исходном уровне в исследовании FaSScinate (Khanna et al. (2016) The Lancet; vol. 387: 10038, p. 2630-2640), в настоящее время продемонстрировано, что экспрессия TGFβ3, а также экспрессия определенного набора генов в сигнатуре TGFβ заметно активируются у пациентов с SSc по сравнению со здоровым контролем, и что экспрессия TGFβ3 значительно и положительно коррелирует с сигнатурой TGFβ, что подтверждает ингибирование TGp3 для лечения SSc с использованием антитела к TGFβ3, описанного в данном документе. Кроме того, данные в примере 16 показывают, что лечение нейтрализующими антителами к TGFβ2 и TGFβ3 может уменьшить фиброз легких in vivo, а данные в примере 17 показывают, что ингибирование TGFβ2 and TGFβ3 может уменьшить фиброз печени in vivo. Таким образом, эти данные подтверждают ингибирование TGFβ2 и/или TGFβ3 для лечения нарушений, связанных с TGFβ, таких как без ограничения фиброзные состояния.
Таким образом, в некоторых аспектах селективные в отношении изоформы антитела к TGFβ применимы для лечения нарушений, связанных с TGFβ. Неограничивающие конкретные примеры нарушений, связанных с TGFβ, включают нейрональные, глиальные, астроцитарные, гипоталамические и другие железистые, макрофагальные, эпителиальные, стромальные и бластоцелические нарушения, фиброз, рубцевание, повреждение тканей, например, вызванное радиацией, и адгезию во время заживления ран, фиброзные кожные заболевания, такие как склеродермия, рубцовая ткань при патологии ЦНС, рубцевание кожи, келоидное рубцевание и рубцевание нервов, фиброзные заболевания брюшной полости, легких, печени и почек, такие как хронический фиброз печени, острое повреждение печени, интерстициальный фиброз легких и почек, и цирроз печени, кистозный фиброз, сосудистые заболевания, например, фиброз сердца, повреждение артерий, такое как атеросклероз и артериосклероз, доброкачественные и злокачественные опухоли, некоторые лейкозы, не ингибируемые TGFβ, и злокачественные новообразования (например, саркомы, карциномы и меланомы), включая предстательную железу, фиброз, рак яичников, злокачественную меланому, рак молочной железы, легких, толстой кишки, прямой кишки, колоректальный рак или рак шейки матки и метастатический рак, а также нейроэндокринные опухоли пищеварительной системы и глиобластомы, ангиопатии, васкулопатии, нефропатии, системный склероз, инфекции, такие как инфекции, вызываемые макрофагами, и вирусные инфекции, такие как гепатит С и ВИЧ, иммунологические, ангиогенные и воспалительные нарушения и дефициты, такие как ревматоидный артрит, заболевание глаз, особенно связанное с фиброзом глаз, включая пролиферативную ретинопатию, отслоение сетчатки и дренирующее хирургическое вмешательство после глаукомы, такие как нейрональная сетчатка, пигментный эпителий сетчатки, хориоидея и стекловидное тело человеческого глаза, а также катаракта, остеопороз, респираторный дистресс-синдром взрослых, состояние после инфаркта миокарда, рестеноз после ангиопластики, гломерулонефрит, состояние, связанное с диабетом, такие как гипергликемия, диабет, диабетическая нефропатия, диабетическая невропатия или ретинопатия и заболевания с недостаточностью макрофагов.
Предпочтительно нарушение, связанное с TGFβ, представляет собой фиброз, повреждение артерии, инфекцию, ревматоидный артрит, диабет или диабетическое состояние или злокачественное новообразование, такое как рак, который экспрессирует TGFβ, более предпочтительно, при котором рак характеризуется избыточной активацией TGFβ. Такой рак может сверхэкспрессировать TGFβ или в качестве альтернативы не характеризоваться сверхэкспрессией TGFβ.
В дополнительных аспектах селективные в отношении изоформы антитела к TGFβ применимы для лечения фиброзных заболеваний, например, фиброзных заболеваний легких, печени, почек, кожи, сердца, поджелудочной железы, органов желудочно-кишечного тракта и мочеполовых органов; сосудистой ткани; и ткани глаз, такой как ткань роговицы, ткань сетчатки и ткани слезной железы. В некоторых вариантах осуществления селективные в отношении изоформы антитела к TGFβ применимы для лечения заболеваний и нарушений, связанных с аберрантной регуляцией и функцией TGFβ, например без ограничения фиброза легких, гломерулонефрита и диабетического заболевания почек, застойной сердечной недостаточности, цирроза печени, синдрома Марфана, гипертрофических рубцов и SSc. В определенных вариантах осуществления фиброзное состояние связано с аутоиммунным заболеванием. В других вариантах осуществления фиброзное состояние возникает после травмы, включая облучение, щелочной ожог, физический ожог, хирургическое вмешательство, физическую травму или их комбинацию. См. Niessen et al., "On the nature of hypertrophic scars and keloids: a review," Plast. Reconstr. Surg. 104:1435-1458 (1999); Friedlander M., "Fibrosis and diseases of the eye," J. Clin. Invest. 117:576-586 (2007); Noble et al., "Pulmonary fibrosis: patterns and perpetrators," J. Clin. Invest. 122:2756-2762 (2012); Bahn R.S., "Graves' ophthalmopathy," N. Engl. J. Med. 362:726-738 (2010); Hinz В., "Formation and function of the myofibroblast during tissue repair," J. Invest. Dermatol. 127:526-537 (2007); Gauglitz et al., "Hypertrophic scarring and keloids: pathomechanisms and current and emerging treatment strategies," Mol. Med. 17:113-125 (2011); Lehmann et al., "Immune mechanisms in thyroid eye disease," Thyroid: official journal of the American Thyroid Association 18:959-965 (2008); Phan S.H., "The myofibroblast in pulmonary fibrosis;" Chest 122:286S-289S (2002); Kuriyan et al., "The eye and thyroid disease," Curr. Opin. Ophthalmol. 19:499-506 (2008).
Иллюстративные фиброзные состояния, которые можно лечить селективными в отношении изоформы антителами к TGFβ, описанными в данном документе, более подробно описаны ниже.
8. Фиброз легких
Иллюстративные фиброзные состояния легких (т.е. фиброз легких) включают без ограничения идиопатический фиброз легких (IPF); идиопатический фиброз верхних долей легких (болезнь Амитани); семейный фиброз легких; фиброз легких, вторичный по отношению к системным воспалительным заболеваниям, таким как ревматоидный артрит, склеродермия, волчанка, криптогенный фиброзирующий альвеолит, хроническая обструктивная болезнь легких (COPD) или хроническая астма; муковисцидоз; неспецифическую интерстициальную пневмонию (NSIP); криптогенную организующуюся пневмонию (СОР); прогрессирующий массивный фиброз; осложнение пневмокониоза угольщиков; склеродермию/системный склероз (SSc, включая ограниченную кожную (IcSSc) и диффузную кожную (dcSSc) формы и связанное с SSc интерстициальное заболевание легких (SSc-ILD)); облитерирующий бронхиолит с организующейся пневмонией, ILD, связанное с заболеванием соединительной ткани (CT-ILD), прогрессирующее фиброзирующее ILD, гиперчувствительный пневмонит, легочную гипертензию, туберкулез, силикоз, асбестоз легких; острое повреждение легких; и острый респираторный дистресс (ARD; включая вызванный бактериальной пневмонией, вызванный повреждением и вызванный вирусной пневмонией, вызванный искусственной вентиляцией легких, вызванный нелегочным сепсисом).
9. Фиброз печени
Иллюстративные фиброзные состояния печени (т.е. фиброз печени) включают без ограничения цирроз печени любой этиологии; врожденный фиброз печени; ожирение; жировую печень; алкогольный фиброз печени; неалкогольный стеатогепатит (NASH); повреждение желчных протоков; первичный билиарный цирроз (РВС), первичный склерозирующий холангит (PSC); фиброз печени, вызванный инфекцией или вирусом (например, хронические вирусные инфекции гепатита В и С); муковисцидоз; аутоиммунный гепатит; некротический гепатит; первичный склерозирующий холангит; гемохроматоз; нарушения желчевыводящих путей; нарушение функции печени, связанное с инфекциями.
10. Фиброз сердца
Иллюстративные фиброзные состояния сердца и/или перикарда (т.е. фиброз сердца или перикарда или фиброз связанной сосудистой сети) включают без ограничения эндомиокардиальный фиброз; васкулопатия сердечного аллотрансплантата (CAV); инфаркт миокарда; фиброз предсердий; хроническую сердечную недостаточность; артериосклероз; атеросклероз; стеноз сосудов; миокардит; застойную кардиомиопатию; коронарные инфаркты; варикозное расширение вен; стеноз коронарных артерий и другие постишемические состояния; идиопатический ретроперитонеальный фиброз.
11. Фиброз почек
Иллюстративные фиброзные состояния почек (т.е. фиброз почек) включают без ограничения гломерулонефрит (включая мембранопролиферативные, диффузно-пролиферативные, быстро прогрессирующие или склерозирующие, постинфекционные и хронические формы); диабетический гломерулосклероз; фокально-сегментарный гломерулосклероз; IgA-нефропатию; диабетическую нефропатию; ишемическую нефропатию; тубулоинтерстициальный фиброз почек, связанную с ВИЧ нефропатию; мембранную нефропатию; гломерулонефрит, вторичный по отношению к системным воспалительным заболеваниям, таким как волчанка, склеродермия и диабетический гломерулонефрит; идиопатический мембранопролиферативный гломерулонефрит; мезангиальный пролиферативный гломерулонефрит; серповидный гломерулонефрит; амилоидоз (который среди других тканей поражает почки); аутоиммунный нефрит; почечный тубулоинстерстиальный фиброз; почечный атеросклероз; синдром Альпорта; нефротический синдром; хроническую почечную недостаточность; хроническое заболевание почек, перигломерулярный фиброз/атубулярных клубочков; синдром сочетанной апикальной эмфиземы и базального фиброза (синдрома эмфиземы/фиброза); гломерулярную гипертензию; нефрогенную фиброзирующую дерматопатию; поликистозную болезнь почек; болезни Фабри и почечную гипертензию.
12. Фиброз поджелудочной железы
Иллюстративные фиброзные состояния поджелудочной железы (т.е. фиброз поджелудочной железы) включают без ограничения ремоделирующий стромальный панкреатит и стромальный фиброз.
Иллюстративные фиброзные состояния желудочно-кишечного тракта (т.е. фиброз желудочно-кишечного тракта) включают без ограничения болезнь Крона; язвенный колит; коллагеновый колит; колоректальный фиброз; атрофия ворсинок; гиперплазию крипт; образование полипов; заживление язвы желудка; и микроскопический колит.
13. Фиброз глаз
Иллюстративные фиброзные состояния глаз включают без ограничения окулярный фиброз, фиброз глаз, пролиферативную витреоретинопатию; витреоретинопатию любой этиологии; фиброза, связанный с дисфункцией сетчатки; фиброз, связанный с влажной или сухой макулодистрофией; рубцевание роговицы и конъюнктивы; фиброз эндотелия роговицы; переднюю субкапсулярную катаракту и помутнение задней капсулы хрусталика; фиброзные заболевания переднего сегмента глаза; фиброз стромы роговицы (например, связанный с помутнением роговицы); фиброз трабекулярной сети (например, связанный с глаукомой); фиброзные заболевания заднего сегмента глаза; фиброваскулярное рубцевание (например, в сетчатке или хориоидальной сосудистой сети глаз); фиброз сетчатки; эпиретинальный фиброз; глиоз сетчатки; субретинальный фиброз (например, связанный с возрастной макулодистрофией); тракционную отслойку сетчатки в сочетании с сокращением ткани при диабетической ретинопатии; врожденный фиброз глазницы; фиброз слезных желез; субэпителиальный фиброз роговицы и офтальмопатию Грейвса.
14. Дополнительные фиброзные нарушения
Дополнительные фиброзные нарушения или фиброз, возникающие в результате любого из вышеупомянутых состояний, включают без ограничения повреждение/фиброз спинного мозга или фиброз центральной нервной системы, такие как фиброз после инсульта, мышечная дистрофия Дюшенна, контрактура Дюпюитрена, фиброз, связанный с нейродегенеративным нарушением, таким как болезнь Альцгеймера или рассеянный склероз; рестеноз сосудов; фиброз матки; эндометриоз; фиброиды яичников; болезнь Пейрони; синдром поликистозных яичников, апикальный фиброз легких, связанный с заболеванием анкилозирующим спондилитом; послеоперационные спайки, рубцевание и фиброз, возникающий при микробных инфекциях (например, бактериальных, вирусных, паразитарных, грибковых и т.д.). Фиброз кожи может включать, например, без ограничения склеродермию, келоиды и морфею.
Способы лечения
Используемое в данном документе лечение фиброза или фиброзных состояний, например, описанных выше, включает в себя нарушение фиброзных процессов для остановки прогрессирования фиброзного состояния, замедления прогрессирования фиброзного состояния или индукции регрессии фиброзного состояния (т.е. улучшения самочувствия пациента в отношении степени фиброза в пораженной ткани или органе). В определенных вариантах осуществления, когда лечение предшествует возникновению фиброзного состояния, т.е. лечение проводят до известного или иным образом ожидаемого начала фиброза, тогда такое лечение может включать предупреждение развития или возникновения фиброзного состояния. Таким образом, введение различных активных средств можно проводить в течение подходящей продолжительности либо для контроля, либо для остановки прогрессирования фиброзного состояния, либо для предупреждения его возникновения.
Таким образом, в некоторых аспектах настоящее изобретение относится к способу лечения фиброза, где фиброз представляет собой фиброзное состояние легких, печени, сердца, почек, поджелудочной железы, глаз и/или кожи. В одном аспекте фиброз представляет собой фиброз легких, выбранный из идиопатического фиброза легких (IPF); идиопатического фиброза верхних долей легких (болезнь Амитани); семейного фиброза легких; фиброза легких, вторичного по отношению к системным воспалительным заболеваниям, таким как ревматоидный артрит, склеродермия, волчанка, криптогенный фиброзирующий альвеолит, хроническая обструктивная болезнь легких (COPD) или хроническая астма; муковисцидоза; неспецифической интерстициальной пневмонии (NSIP); криптогенной организующейся пневмонии (СОР); прогрессирующего массивного фиброза; осложнения пневмокониоза угольщиков; склеродермии/системного склероза (SSc, включая ограниченную кожную (IcSSc) и диффузную кожную (dcSSc) формы и связанное с SSc интерстициальное заболевание легких (SSc-ILD)); облитерирующего бронхиолита с организующейся пневмонией, ILD, связанного с заболеванием соединительной ткани (CT-ILD), прогрессирующеего фиброзирующего ILD, гиперчувствительного пневмонита, легочной гипертензии, туберкулеза, силикоза, асбестоза легких; острого повреждения легких; и острого респираторного дистресса (ARD; включая вызванный бактериальной пневмонией, вызванный повреждением и вызванный вирусной пневмонией, вызванный искусственной вентиляцией легких, вызванный нелегочным сепсисом). В другом аспекте фиброз представляет собой фиброзное состояние печени, выбранное из цирроза печени вследствие всех этиологии; врожденного фиброза печени; ожирения; жировой печени; алкогольного фиброза печени; неалкогольного стеатогепатита (NASH); повреждения желчных протоков; первичного билиарного цирроза (РВС); первичного склерозирующего холангита (PSC); инфекционного или вирусного фиброза печени (например, хронический инфекций, вызванных вирусом гепатита В и С); муковисцидоза, аутоиммунного гепатита; некротизирующего гепатита; первичного склерозирующего холангита; гемохроматоза; нарушений системы желчных протоков; печеночной дисфункции, обусловленной инфекциями. В другом аспекте фиброз представляет собой фиброзное состояние сердца и/или перикарда (т.е. фиброз сердца или перикарда или фиброз связанной сосудистой сети), выбранное из эндомиокардиального фиброза; васкулопатии сердечного аллотрансплантата (CAV); инфаркта миокарда; фиброза предсердий; хронической сердечной недостаточности; артериосклероза; атеросклероза; стеноза сосудов; миокардита; застойной кардиомиопатии; коронарных инфарктов; варикозного расширения вен; стеноза коронарных артерий и других постишемических состояний; идиопатического ретроперитонеального фиброза. В другом аспекте фиброз представляет собой фиброзное состояние почек, выбранное из гломерулонефрита (включая мембранопролиферативные, диффузно-пролиферативные, быстро прогрессирующие или склерозирующие, постинфекционные и хронические формы); диабетического гломерулосклероза; фокально-сегментарного гломерулосклероза; IgA-нефропатии; диабетической нефропатии; ишемической нефропатии; тубулоинтерстициального фиброза почек; связанной с ВИЧ нефропатии; мембранной нефропатии; гломерулонефрита, вторичного по отношению к системным воспалительным заболеваниям, таким как волчанка, склеродермия и диабетический гломерулонефрит; идиопатического мембранопролиферативного гломерулонефрита; мезангиального пролиферативного гломерулонефрита; серповидного гломерулонефрита; амилоидоза (который среди других тканей поражает почки); аутоиммунного нефрита; почечного тубулоинстерстиального фиброза; почечного атеросклероза; синдрома Альпорта; нефротического синдрома; хронической почечной недостаточности; хронического заболевания почек; перигломерулярного фиброза/атубулярных клубочков; синдрома сочетанной апикальной эмфиземы и базального фиброза (синдрома эмфиземы/фиброза); гломерулярной гипертензии; нефрогенной фиброзирующей дерматопатии; поликистозной болезни почек; болезни Фабри и почечной гипертензии. В другом аспекте фиброз представляет собой фиброзное состояние поджелудочной железы, выбранное из стромального ремоделирующего панкреатита и стромального фиброза. В другом аспекте фиброз представляет собой фиброзное состояние желудочно-кишечного тракта (например, фиброз желудочно-кишечного тракта), выбранное из болезни Крона; язвенного колита; коллагенозного колита; колоректального фиброза; атрофии ворсинок; гиперплазии крипт; образования полипов; заживления язвы желудка; и микроскопического колита. В другом аспекте фиброз представляет собой фиброзное состояние глаз, выбранное из окулярного фиброза, фиброза глаз, пролиферативной витреоретинопатии; витреоретинопатии любой этиологии; фиброза, связанного с дисфункцией сетчатки; фиброза, связанного с влажной или сухой макулодистрофией; рубцевания роговицы и конъюнктивы; фиброза эндотелия роговицы; передней субкапсулярной катаракты и помутнения задней капсулы хрусталика; фиброзных заболеваний переднего сегмента глаза; фиброза стромы роговицы (например, связанного с помутнением роговицы); фиброза трабекулярной сети (например, связанного с глаукомой); фиброзных заболеваний заднего сегмента глаза; фиброваскулярного рубцевания (например, в сетчатке или хориоидальной сосудистой сети глаз); фиброза сетчатки; эпиретинального фиброза; глиоза сетчатки; субретинального фиброза (например, связанного с возрастной макулодистрофией); тракционной отслойки сетчатки в сочетании с сокращением ткани при диабетической ретинопатии; врожденного фиброза глазницы; фиброза слезных желез; субэпителиального фиброза роговицы; и офтальмопатии Грейвса. В другом аспекте фиброз выбран из фиброза, возникающего в результате повреждения/фиброза спинного мозга или фиброза центральной нервной системы, таких как фиброз после инсульта, мышечная дистрофия Дюшенна, контрактура Дюпюитрена, фиброз, связанный с нейродегенеративным нарушением, таким как болезнь Альцгеймера или рассеянный склероз; рестеноз сосудов; фиброз матки; эндометриоз; фиброиды яичников; болезнь Пейрони; синдром поликистозных яичников; апикальный фиброз легких, связанный с заболеванием анкилозирующим спондилитом; рубцевание; полиорганная дисфункция соединительной ткани, включающая ткани кожи, легких, сердца, почек и/или кишечника, например, системный склероз (SSc), SSc кожи и фиброз, возникающий при микробных инфекциях (например, бактериальных, вирусных, паразитарных, грибковых и т.д.).
В одном варианте осуществления в настоящем изобретении представлен способ лечения нарушения, связанного с TGFβ, где способ включает введение субъекту, у которого имеется такое нарушение, связанное с TGFβ, эффективного количества селективного в отношении изоформы антитела к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3). В одном таком варианте осуществления способ дополнительно включает введение субъекту эффективного количества по меньшей мере одного дополнительного терапевтического средства, как описано ниже. В другом варианте осуществления в настоящем изобретении представлен способ фиброза (например, фиброзного состояния, описанного выше), где способ включает введение субъекту, у которого имеется такой фиброз, эффективного количества селективного в отношении изоформы антитела к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3). В одном таком варианте осуществления способ дополнительно включает введение субъекту эффективного количества по меньшей мере одного дополнительного терапевтического средства, как описано ниже. В дополнительно варианте осуществления в настоящем изобретении представлен способ ингибирования TGFBR-зависимой передачи сигналов SMAD, ингибирования сборки сигнальных комплексов TGFβ-TGFBR, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченного эндоглином, и/или ингибирования синтеза нового коллагена у субъекта. В другом варианте осуществления способ включает введение субъекту эффективного количества селективного в отношении изоформы антитела к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или для ингибирования синтеза нового коллагена. Иллюстративные варианты осуществления с использованием селективных в отношении изоформы антител к TGFβ по настоящему изобретению представлены ниже.
Способы лечения идиопатического легочного фиброза
В настоящем изобретении представлены способы лечения идиопатического легочного фиброза (IPF). Способы обычно включают введение индивидууму, у которого имеется IPF, эффективного количества селективного в отношении изоформы антитела к TGFβ, описанного в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3). В некоторых вариантах осуществления дозировку и эффективность лечения можно контролировать по обращению или замедлению прогрессирования обычной интерстициальной пневмонии (UIP) при гистопатологической оценке легочной ткани, полученной с помощью хирургической биопсии. Критерии диагностики IPF известны. Ryu et al. (1998) Mayo Clin. Proc. 73:1085-1101. В других вариантах осуществления диагноз IPF представляет собой окончательный или вероятный IPF, поставленный с помощью компьютерной томографии высокого разрешения (HRCT). При диагностике с помощью HRCT отмечают наличие следующих признаков: (1) наличие ретикулярной аномалии и/или тракционных бронхоэктазов с преобладанием базальных и периферических отделов; (2) наличие сот с базальным и периферическим преобладанием; и (3) отсутствие атипичных признаков, таких как микроузелки, перибронховаскулярные узелки, консолидация, изолированные (несотовые) кисты, затухание по типу матового стекла (или, если оно присутствует, менее обширное, чем ретикулярное затемнение) и медиастинальная лимфаденопатия (или, если присутствует, недостаточно обширная, чтобы быть видимой на рентгенограмме грудной клетки). Диагноз окончательного IPF ставится, если соблюдаются характеристики (1), (2) и (3). Диагноз вероятного IPF ставится, если соблюдаются характеристики (1) и (3). В некоторых предпочтительных вариантах осуществления лечение приводит к увеличению, например, к статистически значимому увеличению легочной функции. Значения легочной функции хорошо известны в данной области техники. Ниже приведен пример значений легочной функции, которые можно использовать. Предполагается, что другие показатели легочной функции или их комбинации входят в объем настоящего изобретения. Значения включают без ограничения FEV (объем форсированного выдоха), FVC (форсированная жизненная емкость легких), FEF (форсированный выдох), Vmax (максимальный поток), PEFR (пиковая скорость выдоха), FRC (функциональная остаточная емкость легких), RV (остаточный объем), TLC (общая емкость легких). FEV измеряет объем воздуха, выдыхаемого за заранее определенный период времени, путем форсированного выдоха сразу после полного вдоха. FVC измеряет общий объем выдыхаемого воздуха сразу после полного вдоха. Поток форсированного выдоха измеряет объем воздуха, выдыхаемого во время FVC, деленный на время в секундах. Vmax представляет собой максимальный поток, измеренный во время FVC. PEFR измеряет максимальную скорость потока во время форсированного выдоха, начиная с полного вдоха. RV представляет собой объем воздуха, остающийся в легких после полного выдоха.
Способы лечения фиброза печени
В настоящем изобретении представлены способы лечения фиброза печени, включая уменьшение клинического фиброза печени, уменьшение вероятности возникновения фиброза печени и уменьшение параметра, связанного с фиброзом печени. Особый интерес во многих вариантах осуществления представляет лечение человека. Фиброз печени является предшественником осложнений, связанных с циррозом печени, таких как портальная гипертензия, прогрессирующая печеночная недостаточность и гепатоцеллюлярная карцинома. Таким образом, уменьшение фиброза печени снижает частоту таких осложнений. Соответственно, в настоящем изобретении дополнительно представлены способы снижения вероятности того, что у индивидуума разовьются осложнения, связанные с циррозом печени, путем совместной терапии, включающей введение эффективного количества селективного в отношении изоформы антитела к TGFβ, описанного в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3). Является ли лечение эффективным количеством селективного в отношении изоформы антитела к TGFβ, описанного в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) эффективным для уменьшения фиброза печени, определяется любым из ряда хорошо зарекомендовавших себя методик измерения фиброза печени и функции печени. Уменьшается ли фиброз печени, определяется путем анализа образца биопсии печени. Анализ биопсии печени включает оценку двух основных компонентов: некровоспаления, оцениваемого по «степени» в качестве показателя тяжести и продолжающейся активности заболевания, и поражениям фиброза и паренхиматозного или сосудистого ремоделирования, оцениваемых по «стадии» как отражающих длительное течение заболевания. См., например, Brunt (2000) Hepatol. 31:241-246; and METAVIR (1994) Hepatology 20:15-20. На основании анализа биопсии печени присваивается балл. Существует ряд стандартизированных систем оценки, которые обеспечивают количественную оценку степени и тяжести фиброза. К ним относятся системы подсчета баллов METAVIR, Knodell, Scheuer, Ludwig и Ishak. Система оценки METAVIR основана на анализе различных признаков биопсии печени, включая фиброз (портальный фиброз, центрилобулярный фиброз и цирроз); некроз (частичный и дольковый некроз, ацидофильная ретракция и баллонирующая дегенерация); воспаление (воспаление портального тракта, портальные лимфоидные агрегаты и распространение портального воспаления); изменения желчных протоков; и индекс Knodell (баллы перипортального некроза, лобулярного некроза, портального воспаления, фиброза и общей активности заболевания). Определения каждого этапа в системе METAVIR следующие: балл: 0, фиброз отсутствует; балл: 1, звездчатое расширение портального тракта, но без образования перегородок; балл: 2, расширение портального тракта с редким образованием перегородок; балл: 3, многочисленные перегородки без цирроза; и балл: 4, цирроз. Система оценивания Knodell, также называемая индексом активности гепатита, классифицирует образцы на основе оценок по четырем категориям гистологических признаков: I. Перипортальный и/или мостиковый некроз; II. Внутридольковая дегенерация и очаговый некроз; III. Портальное воспаление; и IV. Фиброз. В системе стадирования Knodell баллы являются следующими: балл: 0, фиброз отсутствует; балл: 1, легкий фиброз (фиброзное портальное расширение); балл: 2, умеренный фиброз; балл: 3, тяжелый фиброз (мостиковый фиброз); и балл: 4, цирроз. Чем выше балл, тем тяжелее поражение ткани печени. Knodell (1981) Hepatol. 1:431. В системе стадирования Scheuer баллы являются следующими: балл: 0, фиброз отсутствует; балл: 1, увеличенные, фиброзированные портальные тракты; балл: 2, перипортальные или портально-портальные перегородки, но неповрежденная архитектура; балл: 3, фиброз с искажением архитектуры, но без явного цирроза печени; балл: 4, вероятный или окончательный цирроз. Scheuer (1991) J. Hepatol. 13:372. Система оценивания Ishak описана в Ishak (1995) J. Hepatol. 22:696-699. Стадия 0, фиброз отсутствует; стадия 1, фиброзное расширение некоторых портальных областей с короткими фиброзными перегородками или без них; стадия 2, фиброзное расширение большинства портальных зон с короткими фиброзными перегородками или без них; стадия 3, фиброзное расширение большинства портальных областей с редкими портально-портальным мостиковыми образованиями (Р--Р); стадия 4, фиброзное расширение портальных зон с выраженным мостиковым образованием (Р--Р), а также портально-центральными мостиковыми образованиями (Р--С); стадия 5, выраженные мостиковые образования (Р-- и/или Р--С) с редкими узелками (неполный цирроз); стадия 6, цирроз, вероятный или окончательный. Польза антифиброзной терапии также может быть измерена и оценена с помощью шкалы Чайлда-Пью, которая включает многокомпонентную систему баллов, основанную на аномалиях уровня билирубина в сыворотке крови, уровне альбумина в сыворотке крови, протромбиновом времени, наличии и тяжести асцита и наличии и степени тяжести энцефалопатии. В зависимости от наличия и тяжести отклонений этих параметров пациенты могут быть отнесены к одной из трех категорий по нарастающей тяжести клинического заболевания: А, В или С.
Способы лечения почечного фиброза
Почечный фиброз характеризуется избыточным накоплением компонентов внеклеточного матрикса (ЕСМ). Считается, что сверхпродуцирование TGFD лежит в основе фиброза тканей, вызванного избыточным отложением ЕСМ, что приводит к заболеванию. Фиброгенное действие TGFD является результатом одновременной стимуляции синтеза белка матрикса, ингибирования деградации матрикса и повышенной экспрессии интегрина, что облегчает сборку ЕСМ. В настоящем изобретении представлены способы лечения идиопатического почечного фиброза. Способы обычно включают введение индивидууму, у которого имеется почечный фиброз, селективного в отношении изоформы антитела к TGFβ, описанного в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3). Используемый в данном документе термин «эффективное количество» селективного в отношении изоформы антитела к TGFβ, описанного в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3), представляет собой дозу, которая эффективна для уменьшения почечного фиброза; и/или которая эффективна для снижения вероятности того, что у индивидуума разовьется почечный фиброз; и/или которая эффективна для снижения параметра, связанного с почечным фиброзом; и/или которая эффективна для уменьшения нарушения, связанного с фиброзом почек. В одном варианте осуществления эффективное количество селективного в отношении изоформы антитела к TGFβ, описанного в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3), представляет собой количество, достаточное для уменьшения почечного фиброза на по меньшей мере приблизительно 10%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50% по сравнению со степенью почечного фиброза у индивидуума до лечения селективным в отношении изоформы антителом к TGFβ. Уменьшается ли фиброз в почках, определяют с помощью любого известного способа. Например, выполняется гистохимический анализ образцов биопсии почки в отношении степени отложения ЕСМ и/или фиброза.
Другие способы известны в данной области техники. См., например, Masseroli et al. (1998) Lab. Invest. 78:511-522; патент США №6214542. В некоторых вариантах осуществления эффективное количество селективного в отношении изоформы антитела к TGFβ, описанного в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3), представляет собой количество, которое эффективно для улучшения функции почек на по меньшей мере приблизительно 10%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50% по сравнению с исходным уровнем функции почек у индивидуума до лечения селективным в отношении изоформы антителом к TGFβ (например, антителом к TGFβ2, антителом к TGFβ2/3 или антителом к TGFβ3) по настоящему изобретению. В некоторых вариантах осуществления эффективное количество селективного в отношении изоформы антитела к TGFβ представляет собой количество, которое эффективно для замедления снижения функции почек на по меньшей мере приблизительно 10%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50% по сравнению со снижением функции почек, которое могло бы произойти при отсутствии лечения с помощью селективного в отношении изоформы антитела к TGFβ по настоящему изобретению. Функцию почек можно измерить с помощью любого известного анализа, включая без ограничения уровень креатинина в плазме крови (где нормальные уровни обычно находятся в диапазоне от приблизительно 0,6 до приблизительно 1,2 мг/дл); клиренс креатинина (где нормальный диапазон клиренса креатинина составляет от приблизительно 97 до приблизительно 137 мл/мин, у мужчин и от приблизительно 88 до приблизительно 128 мл/мин, у женщин); скорость клубочковой фильтрации (рассчитанная или полученная из клиренса инулина или другими методами), азот мочевины крови (где нормальный диапазон составляет от приблизительно 7 до приблизительно 20 мг/дл); и уровень белка в моче. В настоящем изобретении также представлен способ лечения почечного фиброза у индивидуума, включающий введение индивидууму селективного в отношении изоформы антитела к TGFβ в количестве, эффективном для профилактики или лечения почечного фиброза у индивидуума, например, увеличение времени до удвоения уровня креатинина в сыворотке крови, увеличение времени до терминальной стадии почечной недостаточности, требующей заместительной почечной терапии (например, диализа или трансплантации), повышение вероятности выживания, снижение риска смерти, облегчение груза заболевания или замедление прогрессирования заболевания у индивидуума, уменьшая при этом частоту или тяжесть одного или более побочных эффектов, которые обычно возникают без такого лечения.
Антитела к TGFβ2
В некоторых аспектах вышеуказанных способов лечения нарушения, связанного с TGFβ, такого как без ограничения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), способ включает введение субъекту, у которого имеется такое нарушение, связанное с TGFβ, эффективного количества антитела к TGFβ2 по настоящему изобретению, имеющему одно или более из следующих свойств:
антитело к TGFβ2:
(а) селективно нейтрализует TGFβ2;
(б) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ;
(в) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ;
(г) имеет сниженную токсичность у грызунов по сравнению с антителом 1D11 ко всем видам TGFβ;
(д) имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; и/или
(е) имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом.
В другом аспекте в настоящем изобретении представлен способ лечения фиброзного состояния, например, такого как фиброзное состояние, описанное выше, причем способ включает введение субъекту, у которого имеется такой фиброз, эффективного количества селективного в отношении изоформы антитела к TGFβ, имеющего одно или более из следующих свойств:
антитело к TGFβ2 селективно нейтрализует TGFβ2; антитело к TGFβ2 связывается с TGFβ2 с kd, составляющей менее 10 пМ, и/или имеет клеточную IC50, составляющую менее 250 пМ; связывается с TGFβ2 с kd, составляющей менее или равную приблизительно 5 пМ, приблизительно 4 пМ, приблизительно 3 пМ, приблизительно 2 пМ или приблизительно 1 пМ; антитело к TGFβ2 по настоящему изобретению имеет клеточную IC50; составляющую менее или равную приблизительно 250 пМ, приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ, приблизительно 50 пМ или приблизительно 40 пМ.
В одном аспекте в настоящем изобретении представлен способ лечения фиброза, при этом способ включает введение субъекту, у которого имеется такой фиброз, эффективного количества селективного в отношении изоформы антитела к TGFβ, выбранного из следующего:
(i) антитело к TGFβ2, содержащее по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 17; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 19; (д) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 20; и (е) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21;
(ii) антитело к TGFβ2, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (а) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 17; и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 18. В другом аспекте в настоящем изобретении представлено антитело, содержащее (а) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18;
(iii) антитело к TGFβ2, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (а) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 19; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 20; и (в) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21. В одном варианте осуществления антитело содержит (a) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 19; (б) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 20; и (в) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21;
(iv) антитело к TGFβ2 содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18, и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21. В дополнительном варианте осуществления антитело к TGFβ2 содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18, HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17;
(v) антитело к TGFβ2, содержащее (а) домен VH, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 16, (ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 17, и (iii) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 18; и (б) домен VL, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VL, выбранные из (i) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 19, (ii) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 20, и (iii) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21; и
(vi) антитело к TGFβ2, содержащее (а) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 19; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 20; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21.
В любом из вышеуказанных вариантов осуществления способов лечения фиброза антитело к TGFβ2 может быть гуманизировано. В одном варианте осуществления селективное в отношении изоформы антитело к TGFβ содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область. В любом из вышеуказанных вариантов осуществления гуманизированное антитело к TGFβ2 содержит одну или более мутаций в каркасной области VH, выбранных из группы, состоящей из 37V или 37I, 48М или 48L, 49G или 49А, 67L, 71K и 78V, и 105Р или 105R. В некоторых вариантах осуществления антитело к TGFβ2 содержит VH, где VH содержит аминокислотную последовательность SEQ ID NO: 25, при этом VH содержит набор мутаций каркасной области, выбранный из группы, состоящей из: (i) 37V, 48М и 49G в FR2 и 105Р в FR4 (h6F12.v1 и h6F12.v3); (ii) 37V и 48М в FR2, 67L, 71K и 78V в FR3 и 105Р в FR4 (h6F12.v2 и h6F12.v4); (iii) 37I в FR2 (h6F12.v1.6); (iv) 48L в FR2 (h6F12.v1.7); (v) 49А в FR2 (h6F12.v1.8); (vi) 105R в FR4 (h6F12.v1.9); (vii) 37V, 48М и 49G в FR2 и 105Р в FR4 (h6F12.v1 и h6F12.v3); (viii) 37V и 48М в FR2, 67L, 71K и 78V в FR3 и 105Р в FR4 (6F12.v2 и h6F12.v4); (ix) 37I в FR2 (h6F12.v1.6); (х) 48L в FR2 (h6F12.v1.7); (xi) 49А в FR2 (h6F12.v1.8); и (xii) 105R в FR4 (h6F12.v1.9). В любом из вышеуказанных вариантов осуществления гуманизированное антитело к TGFβ2 содержит одну или более мутаций в каркасной области VL, выбранных из группы, состоящей из 43S или 43А, 66G, 69Т, 71F и 87Y. В некоторых вариантах осуществления антитело к TGFβ2 содержит VL, где VL содержит аминокислотную последовательность SEQ ID NO: 24, при этом VL содержит набор мутаций каркасной области, выбранный из группы, состоящей из: (i) 43S в FR2 и 66Е, 69Р, 71Y и 87F в FR3 (h6F12.v1 и h6F12.v2); (ii) 43S в FR2 и 58V, 66Е, 69Р, 71Y и 87F в FR3 (h6F12.v3 и h6F12.v4); (iii) 43А в FR2 (h6F12.v1.1); (iv) 66G в FR3 (h6F12.v1.2); (v) 69Т в FR3 (h6F12.v1.3); (vi) 71F в FR3 (h6F12.v1.4); и (vii) 87Y в FR3 (h6F12.v1.5).
В дополнительном аспекте в настоящем изобретении представлен способ лечения фиброза, при этом способ включает введение субъекту, у которого имеется такой фиброз, эффективного количества селективного в отношении изоформы антитела к TGFβ, выбранного из следующего:
антитело к TGFβ2, содержащее последовательность вариабельного домена тяжелой цепи (VH), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 132 и 138-142; или VH содержит одну, две или три HVR, выбранные из: (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 17; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 18; и антитело содержит вариабельный домен легкой цепи (VL), имеющий по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 24, 131, 133-137, 143 и 144, или VL содержит один, два или три HVR, выбранных из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 19; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 20; и (в) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21;
антитело к TGFβ2, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 25/24 (6F12 кролика), SEQ ID NO: 132/131 (v1), SEQ ID NO: 132/133 (v1.1), SEQ ID NO: 132/134 (v1.2), SEQ ID NO: 132/135 (v1.3), SEQ ID NO: 132/136 (v1.4), SEQ ID NO: 132/137 (v1.5), SEQ ID NO: 138/131 (v1.6), SEQ ID NO: 139/131 (v1.7), SEQ ID NO: 140/131 (v1.8), SEQ ID NO: 141/131 (v1.9), SEQ ID NO: 142/131 (v2), SEQ ID NO: 132/143 (v3) и SEQ ID NO: 142/144 (v4); или
антитело к TGFβ2, содержащее полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 31, 146 и 152-156, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 30, 145, 147-151, 157 и 158;
антитело к TGFβ2, содержащее полную пару H/L-цепей, при этом полная пара H/L-цепей (соответственно) содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 31/30 (6F12 кролика), SEQ ID NO: 146/145 (v1), SEQ ID NO: 146/147 (v1.1), SEQ ID NO: 146/148 (v1.2), SEQ ID NO: 146/149 (v1.3), SEQ ID NO: 146/150 (v1.4), SEQ ID NO: 146/151 (v1.5), SEQ ID NO: 152/145 (v1.6), SEQ ID NO: 153/145 (v1.7), SEQ ID NO: 154/145 (v1.8), SEQ ID NO: 155/145 (v1.9), SEQ ID NO: 156/145 (v2), SEQ ID NO: 146/157 (v3) и SEQ ID NO: 156/158 (v4); или
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ2, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 19; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 20; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21.
Антитела к TGFβ2/3
В некоторых аспектах вышеуказанных способов лечения нарушения, связанного с TGFβ, такого как без ограничения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), способ включает введение субъекту, у которого имеется такое нарушение, связанное с TGFβ (например, фиброз), эффективного количества селективного в отношении изоформы антитела к TGFβ2/3 по настоящему изобретению, которое селективно нейтрализует TGFβ2 и TGFβ3, и имеет одно или более из следующих свойств:
(а) селективность антитела к TGFβ2/3 в отношении TGFβ2 и TGFβ3 по сравнению с TGFβ1 человека в отношении селективной нейтрализации достигается посредством прямого контакта антигенсвязывающего домена антитела с аминокислотным остатком Е373 TGFβ2 или TGFβ3 (нумерация TGFβ2 человека);
(б) антитело к TGFβ2/3 нейтрализует TGFβ2 и/или TGFβ3 посредством аллостерического механизма;
(в) антитело к TGFβ2/3 индуцирует конформационные изменения в гомодимере TGFβ2;
(г) антитело к TGFβ2/3 индуцирует конформационное изменение в гомодимере TGFβ2, при этом конформационное изменение включает совместное сжатие двух мономеров на несколько градусов;
(д) антитело к TGFβ2/3 представляет собой двухвалентное антитело или одновалентное антитело;
(е) антитело к TGFβ2/3 содержит (i) CDR тяжелой цепи, содержащую CDR-H1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащую CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 15;
(ж) антитело к TGFβ2/3 связывается с гомодимером TGFβ2, при этом гомодимер TGFβ2 имеет первый и второй мономер TGFβ2, и при этом антитело к TGFβ2/3 содержит антигенсвязывающий домен, который непосредственно контактирует с (i) аминокислотными остатками V313, Q314, D315, R320, L322, Y323, R328, D329, F345 и А347 первого мономера TGFβ2, и (ii) аминокислотными остатками N368, Т369, I370, N371, Р372, Е373, А374, S375, А376 и S377 второго мономера TGFβ2 (нумерация TGFβ2 человека);
(и) антитело к TGFβ2/3, как в (ж), где антигенсвязывающий домен находится в пределах 5 ангстрем от аминокислотных остатков TGFβ2 и/или TGFβ3;
(к) при этом антитело к TGFβ2/3 связывается с эпитопом, по сути, сходным с эпитопом, как в (ж) в TGFβ3; и
(л) антитело к TGFβ2/3 не нейтрализует TGFβ2 и/или TGFβ3 в форме с одним плечом.
В дополнительном аспекте в настоящем изобретении представлен способ лечения фиброза, при этом способ включает введение субъекту, у которого имеется такой фиброз, эффективного количества антитела к TGFβ2/3, которое имеет одно или более из следующих свойств:
селективно нейтрализует TGFβ2 и TGFβ3; связывается с TGFβ2 и/или TGFβ3 с kd, составляющей менее 10 пМ, и/или имеет клеточную IC50, составляющую менее 250 пМ;
связывается с TGFβ2 и/или TGFβ3 с kd, составляющей менее приблизительно 10 пМ, приблизительно 9 пМ, приблизительно 8 пМ, приблизительно 7 пМ, приблизительно 6 пМ, приблизительно 5 пМ, приблизительно 4 пМ, приблизительно 3 пМ, приблизительно 2 пМ или приблизительно 1 пМ;
имеет клеточную IC50 для ингибирования TGFβ2, составляющую приблизительно 250 пМ, приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ или приблизительно 50 пМ;
имеет клеточную IC50 для ингибирования TGFβ3, составляющую приблизительно 250 пМ, приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ, приблизительно 50 пМ, приблизительно 40 пМ, приблизительно 30 пМ или менее чем приблизительно 30; или
имеет клеточную IC50 для ингибирования TGFβ2, составляющую приблизительно 250 пМ, и клеточную IC50 для ингибирования (нейтрализации) TGFβ3, составляющую приблизительно 30 пМ.
В дополнительном аспекте в настоящем изобретении представлен способ лечения фиброза, при этом способ включает введение субъекту, у которого имеется такой фиброз, эффективного количества антитела к TGFβ2/3, выбранного из следующего:
антитело, содержащее по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащей аминокислотную последовательность антитела к TGFβ2/3 SEQ ID NO: 11; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 12; (г) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 13; (д) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 14; и (е) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 15;
антитело, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (а) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащей аминокислотную последовательность антитела к TGFβ2/3 SEQ ID NO: 11; и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 12; и по меньшей мере одну, по меньшей мере две или все три последовательности HVR VL, выбранные из (а) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 13; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 14; и (в) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 15;
антитело, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 11; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 12; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 13; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 14; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 15.
В любом из вышеуказанных вариантов осуществления способов лечения фиброза антитело к TGFβ2/3 может быть гуманизировано. В одном варианте осуществления антитело к TGFβ2/3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область. В некоторых вариантах осуществления антитело к TGFβ2/3 содержит VH, где VH содержит аминокислотную последовательность SEQ ID NO: 27, при этом VH содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 2Q и 24V в FR1, 48I и 49G в FR2, 71K, 73S, 78V и 91F в FR3 и 105Р в FR4 (h4A11.v1, h4A11.v2, h4A11.v5, h4A11.v6); (ii) 2Q в FR1, 37V в FR2, 67F, 71K, 73S, 78V и 91F в FR3 и 105Р в FR4 (h4A11.v3, h4A11.v4, 4A11.v7, h4A11.v8); (iii) делеции IE в FR1 (h4A11.v7.1); (iv) делеции 75K и 76N в FR3 (h4A11.v7.2); (v) делеции IE в FR1 и 75K76N в FR3 (h4A11.v7.3); (vi) 2V в FR1 (h4A11.v7.8); (vi) 37I в FR2 (h4A11.v7.9); (vii) 67V в FR3 (h4A11.v7.10); (viii) 71V в FR3 (h4A11.v7.11); (ix) 73T в FR3 (h4A11.v7.12); (x) 78F в FR3 (h4A11.v7.13); (xi) 91Y в FR3 (h4A11.v7.14); (xii) 105Q в FR4 (h4A11.v7.15); (xiii) 2V в FR1, 37I в FR2, 67V, 73T, 78F в FR3, 105Q в FR4 (h4A11.v7.16); (xiv) 2V в FR1, 37I в FR2, 67V, 73T, 91Y в FR3, 105Q в FR4 (h4A11.v7.17); (xv) 2V в FR1, 37I в FR2, 67V, 73T в FR3, 105Q в FR4 (h4A11.v7.18); и (xvi) 2V в FR1, 37I в FR2, 67V, 73T, делеции 75K и 76N в FR3, 105Q в FR4 (h4A11.v7.19). В другом варианте осуществления антитело к TGFβ2/3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VL, содержащую модификации FR, выбранные из группы, состоящей из: 2А или 2I, 4L, 36F или 36Y, 43Р или 43А, и 58V или 58I отношению к аминокислотной последовательности VL SEQ ID NO: 26. В некоторых вариантах осуществления антитело к TGFβ2/3 содержит VL, где VL содержит аминокислотную последовательность SEQ ID NO: 26, при этом VL содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 2A и 4L в FR1 и 36F в FR2 (h4A11.v1 и h4A11.v3); (ii) 2A и 4L в FR1 и 36F и 43Р в FR2 (h4A11.v2 и h4A11.v4); (iii) 2A в FR1, 36F и 43Р в FR2 и 58V в FR3 (h4A11.v5 и h4A11.v7); (iv) 2A и 4L в FR1 и 36F в FR2 (h4A11.v6 и h4A11.v8); (v) 21 в FR1 (h4A11.v7.4); (vi) 36Y в FR2 (h4A11.v7.5); (vii) 43A в FR2 (h4A11.v7.6); (viii) 58I в FR3 (h4A11.v7.7); и (ix) 21 в FR1, 43A в FR2, 58I в FR3 (h4A11.v7.16.19).
В дополнительном аспекте в настоящем изобретении представлен способ лечения фиброза, при этом способ включает введение субъекту, у которого имеется такой фиброз, эффективного количества антитела к TGFβ2/3, выбранного из следующего:
антитело, содержащее последовательность вариабельного домена тяжелой цепи (VH), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 27, 81, 83, 86-88, 93-100 и 102-105, и вариабельного домена легкой цепи (VL), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 26, 80, 82, 84, 85, 89-92 и 101;
антитело, содержащее последовательности VH и VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 27/26 (4А11 кролика), SEQ ID NO: 81/80 (v1), SEQ ID NO: 81/82 (v2), SEQ ID NO: 83/80 (v3), SEQ ID NO: 83/82 (v4), SEQ ID NO: 81/84 (v5), SEQ ID NO: 81/85 (v6), SEQ ID NO: 83/84 (v7), SEQ ID NO: 86/84 (v7/1), SEQ ID NO: 87/84 (v7.2), SEQ ID NO: 88/84 (v7.3), SEQ ID NO: 83/89 (v7.4), SEQ ID NO: 83/90 (v7.5), SEQ ID NO: 83/91 (v7.6), SEQ ID NO: 83/92 (v7.7), SEQ ID NO: 93/84 (v7.8), SEQ ID NO: 94/84 (v7.9), SEQ ID NO: 95/84 (v7.10), SEQ ID NO: 96/84 (v7.11), SEQ ID NO: 97/84 (v7.12), SEQ ID NO: 98/84 (v7.13), SEQ ID NO: 99/84 (v7.14), SEQ ID NO: 100/84 (v7.15), SEQ ID NO: 102/101 (v7.16), SEQ ID NO: 103/101 (v7.17), SEQ ID NO: 104/101 (v7.18), SEQ ID NO: 105/101 (v7.19), SEQ ID NO: 83/85 (v8);
антитело, содержащее полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 33, 107, 109, 112-114 и 119-130, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 32, 106, 108, 110, 111, 115-118 и 186;
антитело, содержащее полную пару H/L-цепей, при этом полная пара H/L-цепей (соответственно) содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 32/33 (4А11 кролика), SEQ ID NO: 107/106 (v1), SEQ ID NO: 107/108 (v2), SEQ ID NO: 109/106 (v3), SEQ ID NO: 109/108 (v4), SEQ ID NO: 107/110 (v5), SEQ ID NO: 107/111 (v6), SEQ ID NO: 109/110 (v7), SEQ ID NO: 112/110 (v7.1), SEQ ID NO: 113/110 (v7.2), SEQ ID NO: 114/110 (v7.3), SEQ ID NO: 114/115 (v7.4), SEQ ID NO: 114/116 (v7.5), SEQ ID NO: 114/117 (v7.6), SEQ ID NO: 114/118 (v7.7), SEQ ID NO: 119/110 (v7.8), SEQ ID NO: 120/110 (v7.9), SEQ ID NO: 121/110 (v7.10), SEQ ID NO: 122/110 (v7.11), SEQ ID NO: 123/110 (v7.12), SEQ ID NO: 124/110 (v7.13), SEQ ID NO: 125/110 (v7.14), SEQ ID NO: 126/110 (v7.15), SEQ ID NO: 127/186 (v7.16), SEQ ID NO: 128/186 (v7.17), SEQ ID NO: 129/186 (v7.18), SEQ ID NO: 130/186 (v7.19) и SEQ ID NO: 114/111 (v8);
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ2/3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 11; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 12; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 13; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 14; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 15;
антитело, которое связывается с эпитопом, охватывающим гомодимер TGFβ2, при этом гомодимер TGFβ2 имеет первый и второй мономер TGFβ2, при этом антитело к TGFβ2/3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками V313, Q314, D315, R320, L322, Y323, R328, D329, F345 и А347 первого мономера TGFβ2, и аминокислотными остатками N368, Т369, I370, N371, Р372, Е373, А374, S375, А376 и S377 второго мономера TGFβ2, и при этом в некоторых вариантах осуществления антитело к TGFβ2/3 связывается с тем же эпитопом в TGFβ3.
Антитела к TGFβ3
В некоторых аспектах вышеуказанных способов лечения нарушения, связанного с TGFβ, такого как без ограничения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), способ включает введение субъекту, у которого имеется такое нарушение, связанное с TGFβ, эффективного количества антитела к TGFβ3, которое селективно нейтрализует TGFβ3 по настоящему изобретению, имеющему одно или более из следующих свойств:
(а) антитело к TGFβ3 связывается со шпилечной областью бета6/бета7 TGFβ3;
(б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3;
(в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFBR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3;
(г) антитело к TGFβ3 непосредственно связывает аминокислотный остаток R394 в шпилечной области бета6/бета7 TGFβ3 человека;
(д) антитело к TGFβ3 непосредственно связывает аминокислотный остаток R394 в шпилечной области бета6/бета7 TGFβ3 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи;
(е) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ1 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека);
(ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека);
(и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ;
(к) антитело к TGFβ3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 ко всем видам TGFβ;
(л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом CAT-192 к TGFβ1;
(п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3;
(р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9;
(с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, I388, L389, Y391, V392, G393, R394, Р396, K397 и V398 TGFβ3 человека; и
(т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3.
В дополнительном аспекте в настоящем изобретении представлен способ лечения фиброза, при этом способ включает введение субъекту, у которого имеется такой фиброз, эффективного количества селективного в отношении изоформы антитела к TGFβ, имеющего одно или более из следующих свойств:
антитело, которое селективно нейтрализует TGFβ3;
антитело, которое связывается с TGFβ3 с kd, составляющей менее 10 пМ, и/или имеет клеточную IC50, составляющую менее 250 пМ;
антитело, которое связывается с TGFβ3 с kd, составляющей менее приблизительно 5 пМ, приблизительно 4 пМ, приблизительно 3 пМ или менее приблизительно 2 пМ;
антитело, которое имеет клеточную IC50, составляющую менее приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ или приблизительно 50;
антитело; представленное в данном документе, которое имеет клеточную IC50 для ингибирования (нейтрализации) TGFβ3, составляющую менее приблизительно 20;
антитело, которое селективно нейтрализует TGFβ3 и имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ;
антитело, которое селективно нейтрализует TGFβ3 и имеет улучшенный профиль безопасности.
В дополнительном аспекте в настоящем изобретении представлен способ лечения фиброза, при этом способ включает введение субъекту, у которого имеется такой фиброз, эффективного количества антитела к TGFβ3, выбранного из следующего:
антитело, содержащее по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащей аминокислотную последовательность, выбранную из SEQ ID NO: 5, 34, 35 или 159; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 9;
антитело, содержащее (а) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9;
антитело, содержащее (а) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 34; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9;
антитело, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 35; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9; и
антитело, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 159; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9.
В определенных вариантах осуществления любая одна или более аминокислот антитела к TGFβ3, как указано выше, замещены в следующих положениях HVR:
- в HVR-H2 (SEQ ID NO: 5): в положении N54 (например, N54S, N54Q) или Т56 (например, Т56А).
В некоторых вариантах осуществления замены представляют собой консервативные замены, как представлено в данном документе.
В любом из вышеуказанных вариантов осуществления способов лечения фиброза антитело к TGFβ3 может быть гуманизировано. В одном варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область. В другом варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VH, содержащую модификации FR, выбранные из группы, состоящей из 47L или 47W; 49А, 49S или 49G; 73D или 73N; и 76N, 78D или 78L, 78А или 78V, по отношению к VH, имеющей аминокислотную последовательность SEQ ID NO: 23. В некоторых вариантах осуществления антитело к TGFβ3 содержит VH, где VH содержит аминокислотную последовательность SEQ ID NO: 23, при этом VH содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 47L, 49А в FR2, 78V в FR3 (h2A10.v1); (ii) 47L, 49А в FR2, 73D, 76S, 78V в FR3 (h2A10.v2); (iii) 47W в FR2 (h2A10.v1.5); (iv): 49G в FR2 (h2A10.v1.6); (v) 78A в FR3 (h2A10.v1.7); (vi) 47W в FR2 (h2A10.v2.5); (vii) 49S в FR2 (h2A10.v2.6); (viii) 73N в FR3 (h2A10.v2.7); (ix) 76N в FR3 (h2A10.v2.8); (x) 78L в FR3 (h2A10.v2.9); и (xi) 49S в FR2, 76N, 78L в FR3 (h2A10.v3 и h2A10.v4). В другом варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VL, содержащую модификации FR, выбранные из группы, состоящей из: 4L или 4М, 38Н или 38Q, 43А или 43Q и 58V, по отношению к аминокислотной последовательности VL SEQ ID NO: 22. В другом варианте осуществления антитело содержит VL SEQ ID NO: 22, где VL содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 4L в FR1, 38Н и 43Q в FR2, 58I в FR3 (h2A10.v1 и h2A10.v2); (ii) 4M в FR1 (h2A10.v1.1 и h2A10.v2.1); (iii) 38Q в FR2 (h2A10.v1.2 и h2A10.v2.2); (iv) 43A в FR2 (h2A10.v1.3 и h2A10.v2.3); (v) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); (vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4); (vii) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); и (vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4).
В дополнительном аспекте в настоящем изобретении представлен способ лечения фиброза, при этом способ включает введение субъекту, у которого имеется такой фиброз, эффективного количества антитела к TGFβ3, выбранного из следующего:
антитело, содержащее последовательность вариабельного домена тяжелой цепи (VH), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37 и 42-50, и вариабельного домена легкой цепи (VL), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9);
антитело, содержащее последовательность вариабельного домена тяжелой цепи (VH), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37, 42-50, 55 и 57, и вариабельного домена легкой цепи (VL), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41, 54 и 56;
антитело, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8), SEQ ID NO: 50/36 (v2.9), SEQ ID NO: 55/54 (v3) и SEQ ID NO: 57/56 (v4);
антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 59, 64 и 65-72, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело, содержащее полную пару H/L-цепей, при этом полная пара H/L-цепей содержит (соответственно) аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.6), SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8) и SEQ ID NO: 72/58 (v2.9);
антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29; 59, 64, 65-72, 77 и 79, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 28, 58, 60-63, 76 и 78;
антитело, содержащее полную пару H/L-цепей, при этом полная пара H/L-цепей (соответственно) содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.6), SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8), SEQ ID NO: 72/58 (v2.9), SEQ ID NO: 77/76 (v3) и SEQ ID NO: 79/78;
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 52, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело, содержащее последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 52/36 (h2A10.v2.N54Q);
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74, и полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело, содержащее полные последовательности H/L-цепи (соответственно) AN аминокислотные последовательности SEQ ID NO: 74/58 (h2A10.v2.N54Q);
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 51 или 55, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 51 или 55, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41 и 54;
антитело, содержащее последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 51/36 (h2A10.v2.N54S) или SEQ ID NO: 55/54 (h2A10.v3);
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 73 или 77, и/или антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 73 или 77, и/или антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58, 60-63 и 76;
антитело, содержащее полные последовательности H/L-цепи (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 73/58 (h2A10.v2.N54S) или SEQ ID NO: 77/76 (h2A10.v3);
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 53 или 57, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 53 или 57, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41 и 56;
антитело, содержащее последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 53/36 (h2A10.v2.T56A) или SEQ ID NO: 57/56 (h2A10.v4);
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74 или 79, и/или полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74 или 79, и/или полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58, 60-63 и 78;
антитело, содержащее полные последовательности H/L-цепи (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 75/58 (h2A10.v2.T56A) или SEQ ID NO: 79/78 (h2A10.v4);
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 34; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 35; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (а) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 159; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 53/36 (h2A10.v2.t56A), SEQ ID NO: 57/56 (h2A10.v4), SEQ ID NO: 51/36 (h2A10.v2.N54S), SEQ ID NO: 55/54 (h2A10.v3), SEQ ID NO: 52/36 (h2A10.v2.N54Q), SEQ ID NO: 23/22 (2A10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9);
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 53/36 (h2A10.v2.t56A), SEQ ID NO: 57/56 (h2A10.v4), SEQ ID NO: 51/36 (h2A10.v2.N54S), SEQ ID NO: 55/54 (h2A10.v3), SEQ ID NO: 52/36 (h2A10.v2.N54Q), SEQ ID NO: 23/22 (2A10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8), SEQ ID NO: 50/36 (v2.9), SEQ ID NO: 55/54 (v3) и SEQ ID NO: 57/56 (v4);
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно) SEQ ID NO: 57/56 (h2A10.v4);
антитело, содержащее антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, I388, L389, Y391, V392, G393, R394, Р396, K397 и V398 на TGFβ3 человека, где антитело к TGFβ3 селективно нейтрализует TGFβ3;
антитело, которое селективно нейтрализует TGFβ3 и имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ;
антитело, которое имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ в дозе 50 мг/кг.
В некоторых аспектах вышеуказанных способов лечения нарушения, связанного с TGFβ, способ включает введение субъекту, у которого имеется системная склеродермия (SSc), эффективного количества антитела к TGFβ3, которое селективно нейтрализует TGFβ3, имеющего одно или более из следующих свойств:
(а) антитело к TGFβ3 связывается со шпилечной областью бета6/бета7 TGFβ3;
(б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3;
(в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFBR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3;
(г) антитело к TGFβ3 непосредственно связывает аминокислотный остаток R394 в шпилечной области бета6/бета7 TGFβ3 человека;
(д) антитело к TGFβ3 непосредственно связывает аминокислотный остаток R394 в шпилечной области бета6/бета7 TGFβ3 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи;
(е) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ1 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека);
(ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека);
(и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ;
(к) антитело к TGFβ3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 ко всем видам TGFβ;
(л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом CAT-192 к TGFβ1;
(п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3;
(р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9;
(с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, I388, L389, Y391, V392, G393, R394, Р396, K397 и V398 TGFβ3 человека; и
(т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3.
В дополнительном аспекте в настоящем изобретении представлен способ лечения SSc, при этом способ включает введение субъекту, у которого имеется SSc, эффективного количества селективного в отношении изоформы антитела к TGFβ, имеющего одно или более из следующих свойств:
антитело, которое селективно нейтрализует TGFβ3;
антитело, которое связывается с TGFβ3 с kd, составляющей менее 10 пМ, и/или имеет клеточную IC50, составляющую менее 250 пМ;
антитело, которое связывается с TGFβ3 с kd, составляющей менее приблизительно 5 пМ, приблизительно 4 пМ, приблизительно 3 пМ или менее приблизительно 2 пМ;
антитело, которое имеет клеточную IC50, составляющую менее приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ или приблизительно 50;
антитело, представленное в данном документе, которое имеет клеточную IC50 для ингибирования (нейтрализации) TGFβ3, составляющую менее приблизительно 20;
антитело, которое селективно нейтрализует TGFβ3 и имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ;
антитело, которое селективно нейтрализует TGFβ3 и имеет улучшенный профиль безопасности.
В дополнительном аспекте в настоящем изобретении представлен способ лечения SSc, при этом способ включает введение субъекту, у которого имеется SSc, эффективного количества антитела к TGFβ3, выбранного из следующего:
антитело, содержащее по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащей аминокислотную последовательность, выбранную из SEQ ID NO: 5, 34, 35 или 159; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 9;
антитело, содержащее (а) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9;
антитело, содержащее (а) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 34; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9;
антитело, содержащее (а) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 35; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9; и
антитело, содержащее (а) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 159; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9.
В определенных вариантах осуществления любая одна или более аминокислот антитела к TGFβ3, как указано выше, замещены в следующих положениях HVR:
- в HVR-H2 (SEQ ID NO: 5): в положении N54 (например, N54S, N54Q) или Т56 (например, Т56А).
В некоторых вариантах осуществления замены представляют собой консервативные замены, как представлено в данном документе.
В любом из вышеуказанных вариантов осуществления способов лечения SSc антитело к TGFβ3 может быть гуманизировано. В одном варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область. В другом варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VH, содержащую модификации FR, выбранные из группы, состоящей из 47L или 47W; 49А, 49S или 49G; 73D или 73N; и 76N, 78D или 78L, 78А или 78V. В некоторых вариантах осуществления антитело к TGFβ3 содержит VH, где VH содержит аминокислотную последовательность SEQ ID NO: 23, при этом VH содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 47L, 49А в FR2, 78V в FR3 (h2A10.v1); (ii) 47L, 49А в FR2, 73D, 76S, 78V в FR3 (h2A10.v2); (iii) 47W в FR2 (h2A10.v1.5); (iv): 49G в FR2 (h2A10.v1.6); (v) 78А в FR3 (h2A10.v1.7); (vi) 47W в FR2 (h2A10.v2.5); (vii) 49S в FR2 (h2A10.v2.6); (viii) 73N в FR3 (h2A10.v2.7); (ix) 76N в FR3 (h2A10.v2.8); (х) 78L в FR3 (h2A10.v2.9); и (xi) 49S в FR2, 76N, 78L в FR3 (h2A10.v3 и h2A10.v4). В другом варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VL, содержащую модификации FR, выбранные из группы, состоящей из: 4L или 4М, 38Н или 38Q, 43А или 43Q и 58V. В другом варианте осуществления антитело содержит VL SEQ ID NO: 22, где VL содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 4L в FR1, 38Н и 43Q в FR2, 58I в FR3 (h2A10.v1 и h2A10.v2); (ii) 4M в FR1 (h2A10.v1.1 и h2A10.v2.1); (iii) 38Q в FR2 (h2A10.v1.2 и h2A10.v2.2); (iv) 43A в FR2 (h2A10.v1.3 и h2A10.v2.3); (v) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); (vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4); (vii) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); и (vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4).
В дополнительном аспекте в настоящем изобретении представлен способ лечения SSc, при этом способ включает введение субъекту, у которого имеется такая SSc, эффективного количества антитела к TGFβ3, выбранного из следующего:
антитело, содержащее последовательность вариабельного домена тяжелой цепи (VH), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37 и 42-50, и вариабельного домена легкой цепи (VL), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9);
антитело, содержащее последовательность вариабельного домена тяжелой цепи (VH), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37, 42-50, 55 и 57, и вариабельного домена легкой цепи (VL), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41, 54 и 56;
антитело, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8), SEQ ID NO: 50/36 (v2.9), SEQ ID NO: 55/54 (v3) и SEQ ID NO: 57/56 (v4);
антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 59, 64 и 65-72, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело, содержащее полную пару H/L-цепей, при этом полная пара H/L-цепей содержит (соответственно) аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.6), SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8) и SEQ ID NO: 72/58 (v2.9);
антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 59, 64, 65-72, 77 и 79, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 28, 58, 60-63, 76 и 78;
антитело, содержащее полную пару H/L-цепей, при этом полная пара H/L-цепей (соответственно) содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.6), SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8), SEQ ID NO: 72/58 (v2.9), SEQ ID NO: 77/76 (v3) и SEQ ID NO: 79/78 (v4);
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 52, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело, содержащее последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 52/36 (h2A10.v2.N54Q);
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74, и полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело, содержащее полные последовательности H/L-цепи (соответственно) AN аминокислотные последовательности SEQ ID NO: 74/58 (h2A10.v2.N54Q);
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 51 или 55, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 51 или 55, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41 и 54;
антитело, содержащее последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 51/36 (h2A10.v2.N54S) или SEQ ID NO: 55/54 (h2A10.v3);
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 73 или 77, и/или антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 73 или 77, и/или антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58, 60-63 и 76;
антитело, содержащее полные последовательности H/L-цепи (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 73/58 (h2A10.v2.N54S) или SEQ ID NO: 77/76 (h2A10.v3);
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 53 или 57, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 53 или 57, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41 и 56;
антитело, содержащее последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 53/36 (h2A10.v2.T56A) или SEQ ID NO: 57/56 (h2A10.v4);
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74 или 79, и/или полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74 или 79, и/или полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58, 60-63 и 78;
антитело, содержащее полные последовательности H/L-цепи (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 75/58 (h2A10.v2.T56A) или SEQ ID NO: 79/78 (h2A10.v4);
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 34; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 35; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 159; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 53/36 (h2A10.v2.t56A), SEQ ID NO: 57/56 (h2A10.v4), SEQ ID NO: 51/36 (h2A10.v2.N54S), SEQ ID NO: 55/54 (h2A10.v3), SEQ ID NO: 52/36 (h2A10.v2.N54Q), SEQ ID NO: 23/22 (2A10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9);
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 53/36 (h2A10.v2.t56A), SEQ ID NO: 57/56 (h2A10.v4), SEQ ID NO: 51/36 (h2A10.v2.N54S), SEQ ID NO: 55/54 (h2A10.v3), SEQ ID NO: 52/36 (h2A10.v2.N54Q), SEQ ID NO: 23/22 (2A10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (vl.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.1) , SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8), SEQ ID NO: 50/36 (v2.9), SEQ ID NO: 55/54 (v3) и SEQ ID NO: 57/56 (v4);
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно) SEQ ID NO: 57/56 (h2A10.v4);
антитело, содержащее антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, 1388, L389, Y391, V392, G393, R394, Р396, K397 и V398 на TGFβ3 человека, где антитело к TGFβ3 селективно нейтрализует TGFβ3;
антитело, которое селективно нейтрализует TGFβ3 и имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ;
антитело, которое имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ в дозе 50 мг/кг.
В некоторых аспектах вышеуказанных способов лечения нарушения, связанного с TGFβ, способ включает введение субъекту, у которого имеется идиопатический легочный фиброз (IPF), эффективного количества антитела к TGFβ3, которое селективно нейтрализует TGFβ3, имеющего одно или более из следующих свойств:
(а) антитело к TGFβ3 связывается со шпилечной областью бета6/бета7 TGFβ3;
(б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3;
(в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFBR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3;
(г) антитело к TGFβ3 непосредственно связывает аминокислотный остаток R394 в шпилечной области бета6/бета7 TGFβ3 человека;
(д) антитело к TGFβ3 непосредственно связывает аминокислотный остаток R394 в шпилечной области бета6/бета7 TGFβ3 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи;
(е) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ1 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека);
(ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека);
(и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ;
(к) антитело к TGFβ3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 ко всем видам TGFβ;
(л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом CAT-192 к TGFβ1;
(п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3;
(р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9;
(с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, 1388, L389, Y391, V392, G393, R394, Р396, K397 и V398 TGFβ3 человека; и
(т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3.
В дополнительном аспекте в настоящем изобретении представлен способ лечения IPF, при этом способ включает введение субъекту, у которого имеется такой IPF, эффективного количества селективного в отношении изоформы антитела к TGFβ, имеющего одно или более из следующих свойств:
антитело, которое селективно нейтрализует TGFβ3;
антитело, которое связывается с TGFβ3 с KD, составляющей менее 10 пМ, и/или имеет клеточную IC50, составляющую менее 250 пМ;
антитело, которое связывается с TGFβ3 с KD, составляющей менее приблизительно 5 пМ, приблизительно 4 пМ, приблизительно 3 пМ или менее приблизительно 2 пМ;
антитело, которое имеет клеточную IC50, составляющую менее приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ или приблизительно 50;
антитело, представленное в данном документе, которое имеет клеточную IC50 для ингибирования (нейтрализации) TGFβ3, составляющую менее приблизительно 20;
антитело, которое селективно нейтрализует TGFβ3 и имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ;
антитело, которое селективно нейтрализует TGFβ3 и имеет улучшенный профиль безопасности.
В дополнительном аспекте в настоящем изобретении представлен способ лечения IPF, при этом способ включает введение субъекту, у которого имеется такой IPF, эффективного количества антитела к TGFβ3, выбранного из следующего:
антитело, содержащее по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащей аминокислотную последовательность, выбранную из SEQ ID NO: 5, 34, 35 или 159; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 9;
антитело, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9;
антитело, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 34; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: б; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9;
антитело, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 35; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9; и
антитело, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 159; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9.
В определенных вариантах осуществления любая одна или более аминокислот антитела к TGFβ3, как указано выше, замещены в следующих положениях HVR:
- в HVR-H2 (SEQ ID NO: 5): в положении N54 (например, N54S, N54Q) или Т56 (например, Т56А).
В некоторых вариантах осуществления замены представляют собой консервативные замены, как представлено в данном документе.
В любом из вышеуказанных вариантов осуществления способов лечения IPF антитело к TGFβ3 может быть гуманизировано. В одном варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область. В другом варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VH, содержащую модификации FR, выбранные из группы, состоящей из 47L или 47W; 49А, 49S или 49G; 73D или 73N; и 76N, 78D или 78L, 78А или 78V. В некоторых вариантах осуществления антитело к TGFβ3 содержит VH, где VH содержит аминокислотную последовательность SEQ ID NO: 23, при этом VH содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 47L, 49А в FR2, 78V в FR3 (h2A10.v1); (ii) 47L, 49А в FR2, 73D, 76S, 78V в FR3 (h2A10.v2); (iii) 47W в FR2 (h2A10.v1.5); (iv): 49G в FR2 (h2A10.v1.6); (v) 78A в FR3 (h2A10.v1.7); (vi) 47W в FR2 (h2A10.v2.5); (vii) 49S в FR2 (h2A10.v2.6); (viii) 73N в FR3 (h2A10.v2.7); (ix) 76N в FR3 (h2A10.v2.8); (x) 78L в FR3 (h2A10.v2.9); и (xi) 49S в FR2, 76N, 78L в FR3 (h2A10.v3 и h2A10.v4). В другом варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VL, содержащую модификации FR, выбранные из группы, состоящей из: 4L или 4М, 38Н или 38Q, 43А или 43Q и 58V. В другом варианте осуществления антитело содержит VL SEQ ID NO: 22, где VL содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 4L в FR1, 38Н и 43Q в FR2, 581 в FR3 (h2A10.v1 и h2A10.v2); (ii) 4М в FR1 (h2A10.v1.1 и h2A10.v2.1); (iii) 38Q в FR2 (h2A10.v1.2 и h2A10.v2.2); (iv) 43А в FR2 (h2A10.v1.3 и h2A10.v2.3); (v) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); (vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4); (vii) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); и (vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4).
В дополнительном аспекте в настоящем изобретении представлен способ лечения IPF, при этом способ включает введение субъекту, у которого имеется такой IPF, эффективного количества антитела к TGFβ3, выбранного из следующего:
антитело, содержащее последовательность вариабельного домена тяжелой цепи (VH), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37 и 42-50, и вариабельного домена легкой цепи (VL), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9);
антитело, содержащее последовательность вариабельного домена тяжелой цепи (VH), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37, 42-50, 55 и 57, и вариабельного домена легкой цепи (VL), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41, 54 и 56;
антитело, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (vl.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.1) , SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8), SEQ ID NO: 50/36 (v2.9), SEQ ID NO: 55/54 (v3) и SEQ ID NO: 57/56 (v4);
антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 59, 64 и 65-72, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело, содержащее полную пару H/L-цепей, при этом полная пара H/L-цепей содержит (соответственно) аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (vl.5), SEQ ID NO: 65/58 (v1.1) , SEQ ID NO: 66/58 (vl.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8) и SEQ ID NO: 72/58 (v2.9);
антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 59, 64, 65-72, 77 и 79, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 28, 58, 60-63, 76 и 78;
антитело, содержащее полную пару H/L-цепей, при этом полная пара H/L-цепей (соответственно) содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (vl.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.1) , SEQ ID NO: 66/58 (vl.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8), SEQ ID NO: 72/58 (v2.9), SEQ ID NO: 77/76 (v3) и SEQ ID NO: 79/78;
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 52, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело, содержащее последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 52/36 (h2A10.v2.N54Q);
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74, и полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело, содержащее полные последовательности H/L-цепи (соответственно) AN аминокислотные последовательности SEQ ID NO: 74/58 (h2A10.v2.N54Q);
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 51 или 55, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 51 или 55, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41 и 54;
антитело, содержащее последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 51/36 (h2A10.v2.N54S) или SEQ ID NO: 55/54 (h2A10.v3);
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 73 или 77, и/или антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 73 или 77, и/или антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58, 60-63 и 76;
антитело, содержащее полные последовательности H/L-цепи (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 73/58 (h2A10.v2.N54S) или SEQ ID NO: 77/76 (h2A10.v3);
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 53 или 57, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 53 или 57, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41 и 56;
антитело, содержащее последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 53/36 (h2A10.v2.T56A) или SEQ ID NO: 57/56 (h2A10.v4);
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74 или 79, и/или полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74 или 79, и/или полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58, 60-63 и 78;
антитело, содержащее полные последовательности H/L-цепи (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 75/58 (h2A10.v2.T56A) или SEQ ID NO: 79/78 (h2A10.v4);
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 34; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 35; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 159; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 53/36 (h2A10.v2.t56A), SEQ ID NO: 57/56 (h2A10.v4), SEQ ID NO: 51/36 (h2A10.v2.N54S), SEQ ID NO: 55/54 (h2A10.v3), SEQ ID NO: 52/36 (h2A10.v2.N54Q), SEQ ID NO: 23/22 (2A10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (vl.3), SEQ ID NO: 37/41 (vl.4), SEQ ID NO: 42/36 (vl.5), SEQ ID NO: 43/36 (v1.1) , SEQ ID NO: 44/36 (vl.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9);
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 53/36 (h2A10.v2.t56A), SEQ ID NO: 57/56 (h2A10.v4), SEQ ID NO: 51/36 (h2A10.v2.N54S), SEQ ID NO: 55/54 (h2A10.v3), SEQ ID NO: 52/36 (h2A10.v2.N54Q), SEQ ID NO: 23/22 (2A10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.1) , SEQ ID NO: 37/40 (vl.3), SEQ ID NO: 37/41 (vl.4), SEQ ID NO: 42/36 (vl.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (vl.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8), SEQ ID NO: 50/36 (v2.9), SEQ ID NO: 55/54 (v3) и SEQ ID NO: 57/56 (v4);
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно) SEQ ID NO: 57/56 (h2A10.v4);
антитело, содержащее антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, I388, L389, Y391, V392, G393, R394, Р396, K397 и V398 на TGFβp3 человека, где антитело к TGFβ3 селективно нейтрализует TGFβ3;
антитело, которое селективно нейтрализует TGFβ3 и имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ;
антитело, которое имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ в дозе 50 мг/кг.
Селективное в отношении изоформы антитело к TGFβ в соответствии с любым из вышеуказанных аспектов способов лечения фиброза может представлять собой моноклональное антитело, включая химерное, гуманизированное или человеческое антитело. В одном варианте осуществления селективное в отношении изоформы антитело к TGFβ представляет собой фрагмент антитела, например, фрагмент Fv, Fab, Fab', scFv, диатело или фрагмент F(ab')2. В другом варианте осуществления антитело представляет собой полноразмерное антитело, например, интактное антитело на основе IgG1 или IgG4, предпочтительно, IgG1, или другой класс или изотип антитела, определенный в данном документе.
В некоторых аспектах представленных способов лечения нарушения, связанного с TGFβ, способ включает введение субъекту, у которого имеется нарушение, связанное с TGFβ, эффективного количества антитела к TGFβ3, содержащего: CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 9. В некоторых вариантах осуществления CDR-H2 имеет аминокислотную последовательность SEQ ID NO:
35. В определенных вариантах осуществления выделенное антитело к TGFβ3 дополнительно содержит VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 57. В некоторых вариантах осуществления антитело к TGFβ3 дополнительно содержит VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 56. В определенных вариантах осуществления антитело к TGFβ3 дополнительно содержит полную Н-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 79. В других вариантах осуществления антитело к TGFβ3 дополнительно содержит полную L-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 78. В некоторых вариантах осуществления нарушение представляет собой фиброз. В определенных вариантах осуществления нарушение представляет собой SSc. В некоторых вариантах осуществления нарушение представляет собой IPF. В определенных вариантах осуществления антитело к TGFβ3 представляет собой моноклональное антитело, такое как химерное, гуманизированное или человеческое антитело. В некоторых вариантах осуществления антитело к TGFp3 представляет собой фрагмент антитела, например, фрагмент Fv, Fab, Fab', scFv, диатело или фрагмент F(ab')2. В других вариантах осуществления антитело к TGFβ3 представляет собой полноразмерное антитело, например, интактное антитело на основе IgG1 или IgG4, такое как IgG1, или другой класс или изотип антитела, определенный в данном документе.
Лекарственные препараты и варианты применения антител Используемые в данном документе лекарственные препараты, содержащие селективное в отношении изоформы антитело к TGFβ по настоящему изобретению, производство таких лекарственных препаратов и варианты применения лекарственных препаратов и селективных в отношении изоформы антител к TGFβ по настоящему изобретению для лечения нарушения, связанного с TGFβ, может включать нарушение процессов, обусловленных TGFβ для того, чтобы остановить прогрессирование нарушения, связанного с TGFβ, замедлить прогрессирование нарушения, связанного с TGFβ, или вызвать регресс нарушения, связанного с TGFβ (т.е. улучшить состояние здоровья субъекта (пациента) в зависимости от степени нарушения, связанного с TGFβ, в пораженной ткани или органе). В некоторых вариантах осуществления, если такие варианты применения и лечение предшествуют возникновению нарушения, связанного с TGFβ, т.е. применение или лечение проводят до известного или иным образом ожидаемого начала нарушения, связанного с TGFβ, тогда такое применение или лечение может включать предупреждение развития или начала нарушения, связанного с TGFβ. Таким образом, введение различных активных средств можно проводить в течение подходящей продолжительности либо для контроля, либо для остановки прогрессирования нарушения, связанного с TGFβ, либо для предупреждения его возникновения.
Используемые в данном документе лекарственные препараты, содержащие селективное в отношении изоформы антитело к TGFβ по настоящему изобретению, производство таких лекарственных препаратов и варианты применения лекарственных препаратов и селективных в отношении изоформы антител к TGFβ по настоящему изобретению для лечения фиброза или фиброзных состояний, включают нарушение фиброзных процессов для того, чтобы остановить прогрессирование фиброзного состояния, замедлить прогрессирование фиброзного состояния или вызвать регрессию фиброзного состояния (т.е. улучшить состояние здоровья субъекта (пациента) в зависимости от степени фиброза в пораженной ткани или органе). В определенных вариантах осуществления такие варианты применения и средства лечения предшествуют возникновению фиброзного состояния, т.е. применение или лечение проводят до известного или иным образом ожидаемого начала фиброза, тогда такое лечение может включать предупреждение развития или возникновения фиброзного состояния. Таким образом, введение различных активных средств можно проводить в течение подходящей продолжительности либо для контроля, либо для остановки прогрессирования фиброзного состояния, либо для предупреждения его возникновения.
Таким образом, в некоторых аспектах в настоящем изобретении представлен лекарственный препарат, содержащий селективное в отношении изоформы антитело к TGFβ по настоящему изобретению, и/или варианты применения такого лекарственного препарата или антитела для лечения фиброза, где фиброз представляет собой фиброзное состояние легких, печени, сердца, почек, поджелудочной железы, глаз и/или кожи. В одном аспекте фиброз представляет собой фиброз легких, выбранный из идиопатического фиброза легких (IPF); идиопатического фиброза верхних долей легких (болезнь Амитани); семейного фиброза легких; фиброза легких, вторичного по отношению к системным воспалительным заболеваниям, таким как ревматоидный артрит, склеродермия, волчанка, криптогенный фиброзирующий альвеолит, хроническая обструктивная болезнь легких (COPD) или хроническая астма; муковисцидоза; неспецифической интерстициальной пневмонии (NSIP); криптогенной организующейся пневмонии (СОР); прогрессирующего массивного фиброза; осложнения пневмокониоза угольщиков; склеродермии/системного склероза (SSc, включая ограниченную кожную (lcSSc) и диффузную кожную (dcSSc) формы и связанное с SSc интерстициальное заболевание легких (SSc-ILD)); облитерирующего бронхиолита с организующейся пневмонией, ILD, связанного с заболеванием соединительной ткани (CT-ILD), прогрессирующеего фиброзирующего ILD, гиперчувствительного пневмонита, легочной гипертензии, туберкулеза, силикоза, асбестоза легких; острого повреждения легких; и острого респираторного дистресса (ARD; включая вызванный бактериальной пневмонией, вызванный повреждением и вызванный вирусной пневмонией, вызванный искусственной вентиляцией легких, вызванный нелегочным сепсисом). В другом аспекте фиброз представляет собой фиброзное состояние печени, выбранное из цирроза печени вследствие всех этиологии; врожденного фиброза печени; ожирения; жировой печени; алкогольного фиброза печени; неалкогольного стеатогепатита (NASH); повреждения желчных протоков; первичного билиарного цирроза (РВС); первичного склерозирующего холангита (PSC); инфекционного или вирусного фиброза печени (например, хронический инфекций, вызванных вирусом гепатита В и С); муковисцидоза, аутоиммунного гепатита; некротизирующего гепатита; первичного склерозирующего холангита; гемохроматоза; нарушений системы желчных протоков; печеночной дисфункции, обусловленной инфекциями. В другом аспекте фиброз представляет собой фиброзное состояние сердца и/или перикарда (т.е. фиброз сердца или перикарда или фиброз связанной сосудистой сети), выбранное из эндомиокардиального фиброза; васкулопатии сердечного аллотрансплантата (CAV); инфаркта миокарда; фиброза предсердий; хронической сердечной недостаточности; артериосклероза; атеросклероза; стеноза сосудов; миокардита; застойной кардиомиопатии; коронарных инфарктов; варикозного расширения вен; стеноза коронарных артерий и других постишемических состояний; идиопатического ретроперитонеального фиброза. В другом аспекте фиброз представляет собой фиброзное состояние почек, выбранное из гломерулонефрита (включая мембранопролиферативные, диффузно-пролиферативные, быстро прогрессирующие или склерозирующие, постинфекционные и хронические формы); диабетического гломерулосклероза; фокально-сегментарного гломерулосклероза; IgA-нефропатии; диабетической нефропатии; ишемической нефропатии; тубулоинтерстициального фиброза почек; связанной с ВИЧ нефропатии; мембранной нефропатии; гломерулонефрита, вторичного по отношению к системным воспалительным заболеваниям, таким как волчанка, склеродермия и диабетический гломерулонефрит; идиопатического мембранопролиферативного гломерулонефрита; мезангиального пролиферативного гломерулонефрита; серповидного гломерулонефрита; амилоидоза (который среди других тканей поражает почки); аутоиммунного нефрита; почечного тубулоинстерстиального фиброза; почечного атеросклероза; синдрома Альпорта; нефротического синдрома; хронической почечной недостаточности; хронического заболевания почек; перигломерулярного фиброза/атубулярных клубочков; синдрома сочетанной апикальной эмфиземы и базального фиброза (синдрома эмфиземы/фиброза); гломерулярной гипертензии; нефрогенной фиброзирующей дерматопатии; поликистозной болезни почек; болезни Фабри и почечной гипертензии. В другом аспекте фиброз представляет собой фиброзное состояние поджелудочной железы, выбранное из стромального ремоделирующего панкреатита и стромального фиброза. В другом аспекте фиброз представляет собой фиброзное состояние желудочно-кишечного тракта (например, фиброз желудочно-кишечного тракта), выбранное из болезни Крона; язвенного колита; коллагенозного колита; колоректального фиброза; атрофии ворсинок; гиперплазии крипт; образования полипов; заживления язвы желудка; и микроскопического колита. В другом аспекте фиброз представляет собой фиброзное состояние глаз, выбранное из окулярного фиброза, фиброза глаз, пролиферативной витреоретинопатии; витреоретинопатии любой этиологии; фиброза, связанного с дисфункцией сетчатки; фиброза, связанного с влажной или сухой макулодистрофией; рубцевания роговицы и конъюнктивы; фиброза эндотелия роговицы; передней субкапсулярной катаракты и помутнения задней капсулы хрусталика; фиброзных заболеваний переднего сегмента глаза; фиброза стромы роговицы (например, связанного с помутнением роговицы); фиброза трабекулярной сети (например, связанного с глаукомой); фиброзных заболеваний заднего сегмента глаза; фиброваскулярного рубцевания (например, в сетчатке или хориоидальной сосудистой сети глаз); фиброза сетчатки; эпиретинального фиброза; глиоза сетчатки; субретинального фиброза (например, связанного с возрастной макулодистрофией); тракционной отслойки сетчатки в сочетании с сокращением ткани при диабетической ретинопатии; врожденного фиброза глазницы; фиброза слезных желез; субэпителиального фиброза роговицы; и офтальмопатии Грейвса. В другом аспекте фиброз выбран из фиброза, возникающего в результате повреждения/фиброза спинного мозга или фиброза центральной нервной системы, таких как фиброз после инсульта, мышечная дистрофия Дюшенна, контрактура Дюпюитрена, фиброз, связанный с нейродегенеративным нарушением, таким как болезнь Альцгеймера или рассеянный склероз: рестеноз сосудов; фиброз матки; эндометриоз; фиброиды яичников; болезнь Пейрони; синдром поликистозных яичников; апикальный фиброз легких, связанный с заболеванием анкилозирующим спондилитом; рубцевание; и фиброз, возникающий при микробных инфекциях (например, бактериальных, вирусных, паразитарных, грибковых и т.д.).
В одном варианте осуществления в настоящем изобретении представлен лекарственный препарат, содержащий селективное в отношении изоформы антитело к TGFβ по настоящему изобретению, и/или варианты применения такого лекарственного препарата или антитела для лечения нарушения, связанного с TGFβ, где применение включает введение субъекту, у которого имеется такое нарушение, связанное с TGFβ, эффективного количества селективного в отношении изоформы антитела к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3). В одном таком варианте осуществления применение дополнительно включает введение субъекту эффективного количества по меньшей мере одного дополнительного терапевтического средства, описанного ниже.
В одном варианте осуществления в настоящем изобретении представлен лекарственный препарат, содержащий селективное в отношении изоформы антитело к TGFβ по настоящему изобретению, и/или варианты применения такого лекарственного препарата или антитела для лечения фиброза (например, фиброзного состояния, описанного выше), где применение включает введение субъекту, у которого имеется такой фиброз, эффективного количества селективного в отношении изоформы антитела к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3). В одном таком варианте осуществления применение дополнительно включает введение субъекту эффективного количества по меньшей мере одного дополнительного терапевтического средства, описанного ниже.
В дополнительном аспекте представлено селективное в отношении изоформы антитело к TGFβ, описанное в данном документе (например, антитело к TGFβ2, антитело к TGFβ2/3 или антитело к TGFβ3) для применения в качестве лекарственного препарата. В дополнительных аспектах представлено селективное в отношении изоформы антитело к TGFβ (например, антитело к TGFβ2, антитело к TGFβ2/3 или антитело к TGFβ3) для применения в лечении нарушения, связанного с TGFβ (например, фиброза, например, фиброза легких, например, IPF, печени, поджелудочной железы, сердца, почек, глаз и/или кожи, например, SSc). В определенных вариантах осуществления представлено селективное в отношении изоформы антитело к TGFβ (например, антитело к TGFβ2, антитело к TGFβ2/3 или антитело к TGFβ3) для применения в способе лечения. В определенных вариантах осуществления в настоящем изобретении представлено селективное в отношении изоформы антитело к TGFβ (например, антитело к TGFβ2, антитело к TGFβ2/3 или антитело к TGFβ3) для применения в способе лечения субъекта, у которого имеется фиброз, включающего введение индивидууму эффективного количества селективного в отношении изоформы антитела к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3). В одном таком варианте осуществления применение дополнительно включает введение индивидууму эффективного количества по меньшей мере одного дополнительного терапевтического средства, например, как описано ниже. В дополнительных вариантах осуществления в настоящем изобретении представлено селективное в отношении изоформы антитело к TGFβ (например, антитело к TGFβ2, антитело к TGFβ2/3 или антитело к TGFβ3) для применения в ингибировании TGFBR-зависимой передачи сигналов SMAD, ингибировании сборки сигнальных комплексов TGFβ-TGFBR, ингибировании передачи сигналов TGFβ посредством комплекса TGFBR1/R2, ингибировании передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или для ингибирования синтеза нового коллагена у субъекта. В определенных вариантах осуществления в настоящем изобретении представлено селективное в отношении изоформы антитело к TGFβ (например, антитело к TGFβ2, антитело к TGFβ2/3 или антитело к TGFβ3) для применения в способе ингибирования TGFBR-зависимой передачи сигналов SMAD, ингибирования сборки сигнальных комплексов TGFβ-TGFBR, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или ингибирования синтеза нового коллагена у субъекта, включающем введение субъекту эффективного количества селективного в отношении изоформы антитела к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или для ингибирования синтеза нового коллагена у субъекта. Субъект в соответствии с любым из вышеуказанных вариантов осуществления предпочтительно является человеком.
В дополнительном аспекте в настоящем изобретении представлено применение селективного в отношении изоформы антитела к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в производстве или приготовлении лекарственного препарата. В одном варианте осуществления лекарственный препарат предназначен для лечения нарушения, связанного с TGFβ, например, фиброза, такого как без ограничения фиброз, например, фиброз легких, например, IPF, печени, поджелудочной железы, сердца, почек, глаз и/или кожи, например, SSc. В дополнительном варианте осуществления лекарственный препарат предназначен для применения в способе лечения фиброза, включающем введение субъекту, у которого имеется фиброз, эффективного количества лекарственного препарата. В одном таком варианте осуществления применение дополнительно включает введение субъекту эффективного количества по меньшей мере одного дополнительного терапевтического средства, например, как описано ниже. В дополнительно варианте осуществления лекарственный препарат предназначен для ингибирования TGFBR-зависимой передачи сигналов SMAD, ингибирования сборки сигнальных комплексов TGFβ-TGFBR, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченного эндоглином, и/или ингибирования синтеза нового коллагена у субъекта. В дополнительном варианте осуществления лекарственный препарат предназначен для применения в способе ингибирования TGFBR-зависимой передачи сигналов SMAD, ингибирования сборки сигнальных комплексов TGFβ-TGFBR, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или ингибирования синтеза нового коллагена у субъекта, включающий введение индивидууму эффективного количества лекарственного препарата для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или для ингибирования синтеза нового коллагена у субъекта.
Антитела к TGFβ2
В любом из вышеуказанных аспектов, в отношении селективных в отношении изоформы антител к TGFβ, описанных в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в качестве лекарственного препарата (и их производства, приготовления или применения) и/или для применения в способе лечения, и/или для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или для ингибирования синтеза нового коллагена у субъекта, например, для лечения фиброза (например, фиброза легких, например, IPF, печени, поджелудочной железы, сердца, почек, глаз и/или кожи, например, SSc), антитело к TGFβ2 по настоящему изобретению может иметь одно или более из следующих свойств:
антитело к TGFβ2:
(а) селективно нейтрализует TGFβ2;
(б) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ;
(в) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ;
(г) имеет сниженную токсичность у грызунов по сравнению с антителом 1D11 ко всем видам TGFβ;
(д) имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; и/или
(е) имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ; и
при этом в некоторых вариантах осуществления применение, лекарственный препарат или способ предназначены для лечения фиброза, и при этом фиброз представляет собой SSc или IPF.
В дополнительных аспектах, в отношении селективных в отношении изоформы антител к TGFβ, описанных в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в качестве лекарственного препарата (и их производства, приготовления или применения) и/или для применения в способе лечения, и/или для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, у субъекта, и/или для ингибирования синтеза нового коллагена, например, для лечения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), антитело к TGFβ2 по настоящему изобретению может иметь одно или более из следующих свойств:
антитело к TGFβ2 селективно нейтрализует TGFβ2; антитело к TGFβ2 связывается с TGFβ2 с KD, составляющей менее 10 пМ, и/или имеет клеточную IC50, составляющую менее 250 пМ; связывается с TGFβ2 с KD, составляющей менее или равную приблизительно 5 пМ, приблизительно 4 пМ, приблизительно 3 пМ, приблизительно 2 пМ или приблизительно 1 пМ; антитело к TGFβ2 по настоящему изобретению имеет клеточную IC50, составляющую менее или равную приблизительно 250 пМ, приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ, приблизительно 50 пМ или приблизительно 40 пМ.
В любом из вышеуказанных аспектов, в отношении селективных в отношении изоформы антител к TGFβ, описанных в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в качестве лекарственного препарата (и их производства, приготовления или применения) и/или для применения в способе лечения, и/или для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, у субъекта, и/или для ингибирования синтеза нового коллагена, например, для лечения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), антитело к TGFβ по настоящему изобретению может иметь одно или более из следующих свойств:
антитело к TGFβ2, содержащее по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 17; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 19; (д) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 20; и (е) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21;
антитело к TGFβ2, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 17; и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 18. В другом аспекте в настоящем изобретении представлено антитело, содержащее (a) HVR-Н1, содержащую аминокислотную последовательность SEQ ID NO: 16; (б) HVR-Н2, содержащую аминокислотную последовательность SEQ ID NO: 17; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18;
антитело к TGFβ2, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 19; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 20; и (в) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21. В одном варианте осуществления антитело содержит (a) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 19; (б) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 20; и (в) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21;
антитело к TGFβ2 содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18, и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21. В дополнительном варианте осуществления антитело к TGFβ2 содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18, HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17;
антитело к TGFβ2, содержащее (а) домен VH, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 16, (ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 17, и (iii) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 18; и (б) домен VL, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности HVR VL, выбранные из (i) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 19, (ii) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 20, и (iii) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21; и
антитело к TGFβ2, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 19; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 20; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21.
В любом из вышеуказанных аспектов селективное в отношении изоформы антитело к TGFβ может быть гуманизировано. В одном варианте осуществления селективное в отношении изоформы антитело к TGFβ содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область. В любом из вышеуказанных вариантов осуществления гуманизированное антитело к TGFβ2 содержит одну или более мутаций в каркасной области VH, выбранных из группы, состоящей из 37V или 37I, 48М или 48L, 49G или 49А, 67L, 71K и 78V, и 105Р или 105R. В некоторых вариантах осуществления антитело к TGFβ2 содержит VH, где VH содержит аминокислотную последовательность SEQ ID NO: 25, при этом VH содержит набор мутаций каркасной области, выбранный из группы, состоящей из: (i) 37V, 48М и 49G в FR2 и 105Р в FR4 (h6F12.v1 и h6F12.v3); (ii) 37V и 48М в FR2, 67L, 71K и 78V в FR3 и 105Р в FR4 (h6F12.v2 и h6F12.v4); (iii) 371 в FR2 (h6F12.v1.6); (iv) 48L в FR2 (h6F12.v1.7); (v) 49A в FR2 (h6F12.v1.8); (vi) 105R в FR4 (h6F12.v1.9); (vii) 37V, 48M и 49G в FR2 и 105P в FR4 (h6F12.v1 и h6F12.v3); (viii) 37V и 48M в FR2, 67L, 71K и 78V в FR3 и 105Р в FR4 (6F12.v2 и h6F12.v4); (ix) 371 в FR2 (h6F12.v1.6); (х) 48L в FR2 (h6F12.v1.7); (xi) 49А в FR2 (h6F12.v1.8); и (xii) 105R в FR4 (h6F12.v1.9). В любом из вышеуказанных вариантов осуществления гуманизированное антитело к TGFβ2 содержит одну или более мутаций в каркасной области VL, выбранных из группы, состоящей из 43S или 43А, 66G, 69Т, 71F и 87Y, по отношению к VL, имеющей аминокислотную последовательность SEQ ID NO: 24. В некоторых вариантах осуществления антитело к TGFβ2 содержит VL, где VL содержит аминокислотную последовательность SEQ ID NO: 24, при этом VL содержит набор мутаций каркасной области, выбранный из группы, состоящей из: (i) 43S в FR2 и 66Е, 69Р, 71Y и 87F в FR3 (h6F12.v1 и h6F12.v2); (ii) 43S в FR2 и 58V, 66Е, 69Р, 71Y и 87F в FR3 (h6F12.v3 и h6F12.v4); (iii) 43А в FR2 (h6F12.v1.1); (iv) 66G в FR3 (h6F12.v1.2); (v) 69T в FR3 (h6F12.v1.3); (vi) 71F в FR3 (h6F12.v1.4); и (vii) 87Y в FR3 (h6F12.v1.5).
В любом из вышеуказанных аспектов, в отношении селективных в отношении изоформы антител к TGFβ, описанных в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в качестве лекарственного препарата (и их производства, приготовления или применения) и/или для применения в способе лечения, и/или для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, у субъекта, и/или для ингибирования синтеза нового коллагена, например, для лечения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), селективное в отношении изоформы антитело к TGFβ по настоящему изобретению может иметь одно или более из следующих свойств:
антитело к TGFβ2, содержащее последовательность вариабельного домена тяжелой цепи (VH), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 132 и 138-142; или VH содержит одну, две или три HVR, выбранные из: (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO: 17; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 18; и антитело содержит вариабельный домен легкой цепи (VL), имеющий по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 24, 131, 133-137, 143 и 144, или VL содержит один, два или три HVR, выбранных из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 19; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 20; и (в) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 21;
антитело к TGFβ2, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 25/24 (6F12 кролика), SEQ ID NO: 132/131 (v1), SEQ ID NO: 132/133 (v1.1) , SEQ ID NO: 132/134 (v1.1) , SEQ ID NO: 132/135 (v1.3), SEQ ID NO: 132/136 (v1.4), SEQ ID NO: 132/137 (v1.5), SEQ ID NO: 138/131 (v1.6), SEQ ID NO: 139/131 (vl.7), SEQ ID NO: 140/131 (v1.8), SEQ ID NO: 141/131 (v1.9), SEQ ID NO: 142/131 (v2), SEQ ID NO: 132/143 (v3) и SEQ ID NO: 142/144 (v4); или
антитело к TGFβ2, содержащее полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 31, 146 и 152-156, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 30, 145, 147-151, 157 и 158;
антитело к TGFβ2, содержащее полную пару H/L-цепей, при этом полная пара H/L-цепей (соответственно) содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 31/30 (6F12 кролика), SEQ ID NO: 146/145 (v1), SEQ ID NO: 146/147 (v1.1) , SEQ ID NO: 146/148 (v1.1) , SEQ ID NO: 146/149 (v1.3), SEQ ID NO: 146/150 (v1.4), SEQ ID NO: 146/151 (v1.5), SEQ ID NO: 152/145 (v1.1) , SEQ ID NO: 153/145 (v1.7), SEQ ID NO: 154/145 (vl.8), SEQ ID NO: 155/145 (vl.9), SEQ ID NO: 156/145 (v2), SEQ ID NO: 146/157 (v3) и SEQ ID NO: 156/158 (v4);
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ2, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 16; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 19; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 20; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 21.
Антитела к TGFβ2/3
В любом из вышеуказанных аспектов, в отношении селективных в отношении изоформы антител к TGFβ, описанных в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в качестве лекарственного препарата (и их производства, приготовления или применения) и/или для применения в способе лечения, и/или для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, у субъекта, и/или для ингибирования синтеза нового коллагена, например, для лечения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), антитело к TGFβ2/3, которое селективно нейтрализует TGFβ2 и TGFβ3, описанные в данном документе, может иметь одно или более из следующих свойств:
(а) селективность антитела к TGFβ2/3 в отношении TGFβ2 и TGFβ3 по сравнению с TGFβ1 человека в отношении селективной нейтрализации достигается посредством прямого контакта антигенсвязывающего домена антитела с аминокислотным остатком Е373 TGFβ2 или TGFβ3 (нумерация TGFβ2 человека);
(б) антитело к TGFβ2/3 нейтрализует TGFβ2 и/или TGFβ3 посредством аллостерического механизма;
(в) антитело к TGFβ2/3 индуцирует конформационные изменения в гомодимере TGFβ2;
(г) антитело к TGFβ2/3 индуцирует конформационное изменение в гомодимере TGFβ2, при этом конформационное изменение включает совместное сжатие двух мономеров на несколько градусов;
(д) антитело к TGFβ2/3 представляет собой двухвалентное антитело или одновалентное антитело;
(е) антитело к TGFβ2/3 содержит (i) CDR тяжелой цепи, содержащую CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащую CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 15;
(ж) антитело к TGFβ2/3 связывается с гомодимером TGFβ2, при этом гомодимер TGFβ2 имеет первый и второй мономер TGFβ2, и при этом антитело к TGFβ2/3 содержит антигенсвязывающий домен, который непосредственно контактирует с (i) аминокислотными остатками V313, Q314, D315, R320, L322, Y323, R328, D329, F345 и А347 первого мономера TGFβ2, и (ii) аминокислотными остатками N368, Т369, 1370, N371, Р372, Е373, А374, S375, А376 и S377 второго мономера TGFβ2 (нумерация TGFβ2 человека);
(и) антитело к TGFβ2/3, как в (ж), где антигенсвязывающий домен находится в пределах 5 ангстрем от аминокислотных остатков TGFβ2 и/или TGFβ3;
(к) при этом антитело к TGFβ2/3 связывается с эпитопом, по сути, сходным с эпитопом, как в (ж) в TGFβ3; и
(л) антитело к TGFβ2/3 не нейтрализует TGFβ2 и/или TGFβ3 в форме с одним плечом; и
при этом в некоторых вариантах осуществления применение, лекарственный препарат или способ предназначены для лечения фиброза, и при этом фиброз представляет собой SSc или IPF.
В любом из вышеуказанных аспектов, в отношении селективных в отношении изоформы антител к TGFβ, описанных в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в качестве лекарственного препарата (и их производства, приготовления или применения) и/или для применения в способе лечения, и/или для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, у субъекта, и/или для ингибирования синтеза нового коллагена, например, для лечения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), антитело к TGFβ2/3 по настоящему изобретению может иметь одно или более из следующих свойств:
селективно нейтрализует TGFβ2 и TGFβ3; связывается с TGFβ2 и/или TGFβ3 с KD, составляющей менее 10 пМ, и/или имеет клеточную IC50, составляющую менее 250 пМ;
связывается с TGFβ2 и/или TGFβ3 с KD, составляющей менее приблизительно 10 пМ, приблизительно 9 пМ, приблизительно 8 пМ, приблизительно 7 пМ, приблизительно 6 пМ, приблизительно 5 пМ, приблизительно 4 пМ, приблизительно 3 пМ, приблизительно 2 пМ или приблизительно 1 пМ;
имеет клеточную IC50 для ингибирования TGFβ2, составляющую приблизительно 250 пМ, приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ или приблизительно 50 пМ;
имеет клеточную IC50 для ингибирования TGFβ3, составляющую приблизительно 250 пМ, приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ, приблизительно 50 пМ, приблизительно 40 пМ, приблизительно 30 пМ или менее чем приблизительно 30; или
имеет клеточную IC50 для ингибирования TGFβ32, составляющую приблизительно 250 пМ, и клеточную IC50 для ингибирования (нейтрализации) TGFβ3, составляющую приблизительно 30 пМ.
В любом из вышеуказанных аспектов в отношении применения селективного в отношении изоформы антитела к TGFβ, описанного в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в качестве лекарственного препарата (и его производства или приготовления), и/или для применения в способе лечения, например, для лечения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), селективное в отношении изоформы антитело к TGFβ может быть выбрано из:
антитела к TGFβ2/3, содержащего по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащей аминокислотную последовательность антитела к TGFβ2/3 SEQ ID NO: 11; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 12; (г) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 13; (д) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 14; и (е) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 15;
антитела к TGFβ2/3, содержащего по меньшей мере одну, по меньшей мере две или все три последовательности HVR VH, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащей аминокислотную последовательность антитела к TGFβ2/3 SEQ ID NO: 11; и (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 12; и по меньшей мере одну, по меньшей мере две или все три последовательности HVR VL, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 13; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 14; и (в) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 15;
антитела к TGFβ2/3, содержащего (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 11; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 12; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 13; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 14; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 15.
В любом из вышеуказанных вариантов применения антитело к TGFβ2/3 может быть гуманизировано. В одном варианте осуществления антитело к TGFβ2/3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область. В некоторых вариантах осуществления антитело к TGFβ2/3 содержит VH, где VH содержит аминокислотную последовательность SEQ ID NO: 27, при этом VH содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 2Q и 24V в FR1, 48I и 49G в FR2, 71K, 73S, 78V и 91F в FR3 и 105Р в FR4 (h4A11.v1, h4A11.v2, h4A11.v5, h4A11.v6); (ii) 2Q в FR1, 37V в FR2, 67F, 71K, 73S, 78V и 91F в FR3 и 105P в FR4 (h4A11.v3, h4A11.v4, 4A11.v7, h4A11.v8); (iii) делеции 1E в FR1 (h4A11.v7.1); (iv) делеции 75K и 76N в FR3 (h4A11.v7.2); (v) делеции IE в FR1 и 75K76N в FR3 (h4A11.v7.3); (vi) 2V в FR1 (h4A11.v7.8); (vi) 371 в FR2 (h4A11.v7.9); (vii) 67V в FR3 (h4A11.v7.10); (viii) 71V в FR3 (h4A11.v7.11); (ix) 73T в FR3 (h4A11.v7.12); (x) 78F в FR3 (h4A11.v7.13); (xi) 91Y в FR3 (h4A11.v7.14); (xii) 105Q в FR4 (h4A11.v7.15); (xiii) 2V в FR1, 37I в FR2, 67V, 73T, 78F в FR3, 105Q в FR4 (h4A11.v7.16); (xiv) 2V в FR1, 37I в FR2, 67V, 73T, 91Y в FR3, 105Q в FR4 (h4A11.v7.17); (xv) 2V в FR1, 37I в FR2, 67V, 73T в FR3, 105Q в FR4 (h4A11.v7.18); и (xvi) 2V в FR1, 371 в FR2, 67V, 73T, делеции 75K и 76N в FR3, 105Q в FR4 (h4A11.v7.19). В другом варианте осуществления антитело к TGFβ2/3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VL, содержащую модификации FR, выбранные из группы, состоящей из: 2А или 2I, 4L, 36F или 36Y, 43Р или 43А, и 58V или 58I отношению к VL, имеющей аминокислотную последовательность SEQ ID NO: 26. В некоторых вариантах осуществления антитело к TGFβ2/3 содержит VL, где VL содержит аминокислотную последовательность SEQ ID NO: 26, при этом VL содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 2А и 4L в FR1 и 36F в FR2 (h4A11.v1 и h4A11.v3); (ii) 2А и 4L в FR1 и 36F и 43Р в FR2 (h4A11.v2 и h4A11.v4); (iii) 2А в FR1, 36F и 43Р в FR2 и 58V в FR3 (h4A11.v5 и h4A11.v7); (iv) 2А и 4L в FR1 и 36F в FR2 (h4A11.v6 и h4A11.v8); (v) 21 в FR1 (h4A11.v7.4); (vi) 36Y в FR2 (h4A11.v7.5); (vii) 43A в FR2 (h4A11.v7.6); (viii) 58I в FR3 (h4A11.v7.7); и (ix) 2I в FR1, 43A в FR2, 58I в FR3 (h4A11.v7.16-19).
В любом из вышеуказанных аспектов, в отношении селективных в отношении изоформы антител к TGFβ, описанных в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в качестве лекарственного препарата (и их производства, приготовления или применения) и/или для применения в способе лечения, и/или для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, у субъекта, и/или для ингибирования синтеза нового коллагена, например, для лечения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), антитело к TGFβ2/3 по настоящему изобретению может иметь одно или более из следующих свойств:
антитело к TGFβ2/3, содержащее последовательность вариабельного домена тяжелой цепи (VH), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 27, 81, 83, 86-88, 93-100 и 102-105, и вариабельного домена легкой цепи (VL), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 26, 80, 82, 84, 85, 89-92 и 101;
антитело к TGFβ2/3, содержащее последовательности VH и VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 27/26 (4А11 кролика), SEQ ID NO: 81/80 (v1), SEQ ID NO: 81/82 (v2), SEQ ID NO: 83/80 (v3), SEQ ID NO: 83/82 (v4), SEQ ID NO: 81/84 (v5), SEQ ID NO: 81/85 (v6), SEQ ID NO: 83/84 (v7), SEQ ID NO: 86/84 (v7/1), SEQ ID NO: 87/84 (v7.2), SEQ ID NO: 88/84 (v7.3), SEQ ID NO: 83/89 (v7.4), SEQ ID NO: 83/90 (v7.5), SEQ ID NO: 83/91 (v7.6), SEQ ID NO: 83/92 (v7.7), SEQ ID NO: 93/84 (v7.8), SEQ ID NO: 94/84 (v7.9), SEQ ID NO: 95/84 (v7.10), SEQ ID NO: 96/84 (v7.11), SEQ ID NO: 97/84 (v7.12), SEQ ID NO: 98/84 (v7.13), SEQ ID NO: 99/84 (v7.14), SEQ ID NO: 100/84 (v7.15), SEQ ID NO: 102/101 (v7.16), SEQ ID NO: 103/101 (v7.17), SEQ ID NO: 104/101 (v7.18), SEQ ID NO: 105/101 (v7.19), SEQ ID NO: 83/85 (v8);
антитело к TGFβ2/3, содержащее полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 33, 107, 109, 112-114 и 119-130, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 32, 106, 108, 110, 111, 115-118 и 186;
антитело к TGFβ2/3, содержащее полную пару H/L-цепей, при этом полная пара H/L-цепей (соответственно) содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 32/33 (4А11 кролика), SEQ ID NO: 107/106 (v1), SEQ ID NO: 107/108 (v2), SEQ ID NO: 109/106 (v3), SEQ ID NO: 109/108 (v4), SEQ ID NO: 107/110 (v5), SEQ ID NO: 107/111 (v6), SEQ ID NO: 109/110 (v7), SEQ ID NO: 112/110 (v7.1), SEQ ID NO: 113/110 (v7.2), SEQ ID NO: 114/110 (v7.3), SEQ ID NO: 114/115 (v7.4), SEQ ID NO: 114/116 (v7.5), SEQ ID NO: 114/117 (v7.6), SEQ ID NO: 114/118 (v7.7), SEQ ID NO: 119/110 (v7.8), SEQ ID NO: 120/110 (v7.9), SEQ ID NO: 121/110 (v7.10), SEQ ID NO: 122/110 (v7.11), SEQ ID NO: 123/110 (v7.12), SEQ ID NO: 124/110 (v7.13), SEQ ID NO: 125/110 (v7.14), SEQ ID NO: 126/110 (v7.15), SEQ ID NO: 127/186 (v7.16), SEQ ID NO: 128/186 (v7.17), SEQ ID NO: 129/186 (v7.18), SEQ ID NO: 130/186 (v7.19) и SEQ ID NO: 114/111 (v8);
антитело, которое связывается с тем же эпитопом, что и антитело к TGFβ2/3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 10; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 11; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 12; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 13; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 14; и (e) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 15;
антитело к TGFβ2/3, которое связывается с эпитопом в фрагменте гомодимера TGFβ2, при этом гомодимер TGFβ2 имеет первый и второй мономер TGFβ2, при этом антитело к TGFβ2/3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками V313, Q314, D315, R320, L322, Y323, R328, D329, F345 и А347 первого мономера TGFβ2, и аминокислотными остатками N368, Т369, I370, N371, Р372, Е373, А374, S375, А376 и S377 второго мономера TGFβ2, и при этом в некоторых вариантах осуществления антитело к TGFβ2/3 связывается с той же областью и/или эпитопом в TGFβ3.
Антитела к TGFβ3
В любом из вышеуказанных аспектов, в отношении селективных в отношении изоформы антител к TGFβ, описанных в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в качестве лекарственного препарата (и их производства, приготовления или применения) и/или для применения в способе лечения, и/или для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, у субъекта, и/или для ингибирования синтеза нового коллагена, например, для лечения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), антитело к TGFβ3, которое селективно нейтрализует TGFβ3, описанное в данном документе, может иметь одно или более из следующих свойств:
(а) антитело к TGFβ3 связывается со шпилечной областью бета6/бета7 TGFβ3;
(б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3;
(в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFBR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3;
(г) антитело к TGFβ3 непосредственно связывает аминокислотный остаток R394 в шпилечной области бета6/бета7 TGFβ3 человека;
(д) антитело к TGFβ3 непосредственно связывает аминокислотный остаток R394 в шпилечной области бета6/бета7 TGFβ3 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи;
(е) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ1 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека);
(ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека);
(и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ;
(к) антитело к TGFβ3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 ко всем видам TGFβ;
(л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом CAT-192 к TGFβ1;
(п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3;
(р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9;
(с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, I388, L389, Y391, V392, G393, R394, Р396, K397 и V398 TGFβ3 человека; и
(т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3; и
при этом в некоторых вариантах осуществления применение, лекарственный препарат или способ предназначены для лечения фиброза, и при этом фиброз представляет собой SSc или IPF.
В дополнительных аспектах, в отношении селективных в отношении изоформы антител к TGFβ, описанных в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в качестве лекарственного препарата (и их производства, приготовления или применения) и/или для применения в способе лечения, и/или для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, у субъекта, и/или для ингибирования синтеза нового коллагена, например, для лечения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), селективное в отношении изоформы антитело к TGFβ по настоящему изобретению может иметь одно или более из следующих свойств:
антитело к TGFβ3, которое селективно нейтрализует TGFβ3;
антитело к TGFβ3, которое связывается с TGFβ3 с KD, составляющей менее 10 пМ, и/или имеет клеточную IC50, составляющую менее 250 пМ;
антитело к TGFβ3, которое связывается с TGFβ3 с KD, составляющей менее приблизительно 5 пМ, приблизительно 4 пМ, приблизительно 3 пМ или менее приблизительно 2 пМ;
антитело к TGFβ3, которое имеет клеточную IC50, составляющую менее приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 75 пМ или приблизительно 50;
антитело к TGFβ3, представленное в данном документе, которое имеет клеточную IC50 для ингибирования (нейтрализации) TGFβ3, составляющую менее приблизительно 20;
антитело к TGFβ3, которое селективно нейтрализует TGFβ3 и имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ; и
антитело к TGFβ3, которое селективно нейтрализует TGFβ3 и имеет улучшенный профиль; и
при этом в некоторых вариантах осуществления применение, лекарственный препарат или способ предназначены для лечения фиброза, и при этом фиброз представляет собой SSc или IPF.
В любом из вышеуказанных аспектов, в отношении селективных в отношении изоформы антител к TGFβ, описанных в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в качестве лекарственного препарата (и их производства, приготовления или применения) и/или для применения в способе лечения, и/или для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, у субъекта, и/или для ингибирования синтеза нового коллагена, например, для лечения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), антитело к TGFβ3 по настоящему изобретению может иметь одно или более из следующих свойств:
антитело к TGFβ3, содержащее по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащей аминокислотную последовательность, выбранную из SEQ ID NO: 5, 34, 35 или 159; (в) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO: 9;
антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9;
антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 34; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9;
антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 35; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9;
антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 159; (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6; (г) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7; (д) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO: 8; и (е) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9.
В определенных вариантах осуществления любая одна или более аминокислот антитела к TGFβ3, как указано выше, замещены в следующих положениях HVR:
- в HVR-H2 (SEQ ID NO: 5): в положении N54 (например, N54S, N54Q) или Т56 (например, Т56А).
В некоторых вариантах осуществления замены представляют собой консервативные замены, как представлено в данном документе.
В любом из вышеуказанных вариантов осуществления антитело к TGFβ3 может быть гуманизировано. В одном варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область. В другом варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VH, содержащую модификации FR, выбранные из группы, состоящей из 47L или 47W; 49А, 49S или 49G; 73D или 73N; и 76N, 78D или 78L, 78А или 78V. В некоторых вариантах осуществления антитело к TGFβ3 содержит VH, где VH содержит аминокислотную последовательность SEQ ID NO: 23, при этом VH содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 47L, 49А в FR2, 78V в FR3 (h2A10.v1); (ii) 47L, 49А в FR2, 73D, 76S, 78V в FR3 (h2A10.v2); (iii) 47W в FR2 (h2A10.v1.5); (iv): 49G в FR2 (h2A10.v1.6); (v) 78A в FR3 (h2A10.v1.7); (vi) 47W в FR2 (h2A10.v2.5); (vii) 49S в FR2 (h2A10.v2.6); (viii) 73N в FR3 (h2A10.v2.7); (ix) 76N в FR3 (h2A10.v2.8); (x) 78L в FR3 (h2A10.v2.9); и (xi) 49S в FR2, 76N, 78L в FR3 (h2A10.v3 и h2A10.v4). В другом варианте осуществления антитело к TGFβ3 содержит HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно содержит VL, содержащую модификации FR, выбранные из группы, состоящей из: 4L или 4М, 38Н или 38Q, 43А или 43Q и 58V, по отношению к аминокислотной последовательности VL SEQ ID NO: 22. В другом варианте осуществления антитело содержит VL SEQ ID NO: 22, где VL содержит набор модификаций каркасной области, выбранный из группы, состоящей из: (i) 4L в FR1, 38Н и 43Q в FR2, 58I в FR3 (h2A10.v1 и h2A10.v2); (ii) 4М в FR1 (h2A10.v1.1 и h2A10.v2.1); (iii) 38Q в FR2 (h2A10.v1.2 и h2A10.v2.2); (iv) 43А в FR2 (h2A10.vl.3 и h2A10.v2.3); (v) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); (vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4); (vii) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); и (vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4).
В любом из вышеуказанных аспектов, в отношении селективных в отношении изоформы антител к TGFβ, описанных в данном документе (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в качестве лекарственного препарата (и их производства, приготовления или применения) и/или для применения в способе лечения, и/или для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, у субъекта, и/или для ингибирования синтеза нового коллагена, например, для лечения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи), селективное в отношении изоформы антитело к TGFβ по настоящему изобретению может иметь одно или более из следующих свойств:
антитело к TGFβ3, содержащее последовательность вариабельного домена тяжелой цепи (VH), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37 и 42-50, и вариабельного домена легкой цепи (VL), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9);
антитело к TGFβ3, содержащее последовательность вариабельного домена тяжелой цепи (VH), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37, 42-50, 55 и 57, и вариабельного домена легкой цепи (VL), имеющего по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41, 54 и 56;
антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8), SEQ ID NO: 50/36 (v2.9), SEQ ID NO: 55/54 (v3) и SEQ ID NO: 57/56 (v4);
антитело к TGFβ3 содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 59, 64 и 65-72, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело к TGFβ3, содержащее полную пару H/L-цепей, при этом полная пара H/L-цепей содержит (соответственно) аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1), SEQ ID NO: 59/61 (v1.1) , SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (vl.5), SEQ ID NO: 65/58 (v1.6), SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8) и SEQ ID NO: 72/58 (v2.9);
антитело к TGFβ3, содержащее полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95% идентичности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 59, 64, 65-72, 77 и 79, и/или полную аминокислотную последовательность L-цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 28, 58, 60-63, 76 и 78;
антитело к TGFβ3, содержащее полную пару H/L-цепей, при этом полная пара H/L-цепей (соответственно) содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1) , SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.1) , SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8), SEQ ID NO: 72/58 (v2.9), SEQ ID NO: 77/76 (v3) и SEQ ID NO: 79/78;
антитело к TGFβ3, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 52, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 52/36 (h2A10.v2.N54Q);
антитело к TGFβ3, содержащее полную Н-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74, и полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело к TGFβ3, содержащее полные последовательности H/L-цепи (соответственно) AN аминокислотные последовательности SEQ ID NO: 74/58 (h2A10.v2.N54Q);
антитело к TGFβ3, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 51 или 55, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело к TGFβ3, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 51 или 55, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41 и 54;
антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 51/36 (h2A10.v2.N54S) или SEQ ID NO: 55/54 (h2A10.v3);
антитело к TGFβ3, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 73 или 77, и/или антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело к TGFβ3, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 73 или 77, и/или антитело к TGFβ3 содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58, 60-63 и 76;
антитело к TGFβ3, содержащее полные последовательности H/L-цепи (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 73/58 (h2A10.v2.N54S) или SEQ ID NO: 77/76 (h2A10.v3);
антитело к TGFβ3, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 53 или 57, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41;
антитело к TGFβ3, содержащее VH, имеющее по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности SEQ ID NO: 53 или 57, и аминокислотную последовательность VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41 и 56;
антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 53/36 (h2A10.v2.T56A) или SEQ ID NO: 57/56 (h2A10.v4);
антитело к TGFβ3, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74 или 79, и/или полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63;
антитело к TGFβ3, содержащее полную Н-цепь, имеющую по меньшей мере 95% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74 или 79, и/или полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58, 60-63 и 78;
антитело к TGFβ3, содержащее полные последовательности H/L-цепи (соответственно), содержащие аминокислотные последовательности SEQ ID NO: 75/58 (h2A10.v2.T56A) или SEQ ID NO: 79/78 (h2A10.v4);
антитело к TGFβ3, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 5; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело к TGFβ3, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 34; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело к TGFβ3, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 35; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело к TGFβ3, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4; (б) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO: 159; и (в) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6;
антитело к TGFβ3, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 53/36 (h2A10.v2.t56A), SEQ ID NO: 57/56 (h2A10.v4), SEQ ID NO: 51/36 (h2A10.v2.N54S), SEQ ID NO: 55/54 (h2A10.v3), SEQ ID NO: 52/36 (h2A10.v2.N54Q), SEQ ID NO: 23/22 (2A10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9);
антитело к TGFβ3, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно), выбранные из группы, состоящей из SEQ ID NO: 53/36 (h2A10.v2.t56A), SEQ ID NO: 57/56 (h2A10.v4), SEQ ID NO: 51/36 (h2A10.v2.N54S), SEQ ID NO: 55/54 (h2A10.v3), SEQ ID NO: 52/36 (h2A10.v2.N54Q), SEQ ID NO: 23/22 (2A10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8), и SEQ ID NO: 50/36 (v2.9), SEQ ID NO: 55/54 (v3) и SEQ ID NO: 57/56 (v4);
антитело к TGFβ3, которое связывается с тем же эпитопом, что и антитело к TGFβ3, содержащее последовательности VH/VL (соответственно) SEQ ID NO: 57/56 (h2A10.v4);
антитело к TGFβ3, содержащее антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, I388, L389, Y391, V392, G393, R394, Р396, K397 и V398 на TGFβ3 человека, где антитело к TGFβ3 селективно нейтрализует TGFβ3;
антитело к TGFβ3, которое селективно нейтрализует TGFβ3 и имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ;
антитело к TGFβ3, которое имеет сниженную токсичность у мышей по сравнению с антителом 1D11 ко всем видам TGFβ в дозе 50 мг/кг.
Селективное в отношении изоформы антитело к TGFβ в соответствии с любым из вышеуказанных вариантов применения (например, в качестве лекарственного препарата для лечения фиброза) может представлять собой моноклональное антитело, включая химерное, гуманизированное или человеческое антитело. В одном варианте осуществления селективное в отношении изоформы антитело к TGFβ представляет собой фрагмент антитела, например, фрагмент Fv, Fab, Fab', scFv, диатело или фрагмент F(ab')2. В другом варианте осуществления антитело представляет собой полноразмерное антитело, например, интактный IgG1 или IgG4, предпочтительно, антитело на основе IgG1, или другой класс или изотип антитела, определенный в данном документе.
В любом из аспектов, представленных в данном документе, для применения антитела к TGFβ3 в качестве лекарственного препарата (и его производства, приготовления или применения) и/или для применения в способе лечения, и/или для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, у субъекта, и/или для ингибирования синтеза нового коллагена, например, для лечения фиброза (например, фиброза легких, печени, поджелудочной железы, сердца, почек, глаз и/или кожи) антитело к TGFβ3 представляет собой антитело, которое селективно нейтрализует TGFβ3, как описано в данном документе, и содержит: CDR тяжелой цепи, содержащие CDR- H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-Н3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8, a CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9. В некоторых вариантах осуществления CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35. В определенных вариантах осуществления антитело к TGFβ3 дополнительно содержит VH, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 57. В некоторых вариантах осуществления антитело к TGFβ3 дополнительно содержит VL, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 56. В определенных вариантах осуществления антитело к TGFβ3 дополнительно содержит полную Н-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 79. В других вариантах осуществления антитело к TGFβ3 дополнительно содержит полную L-цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 78. В некоторых вариантах осуществления нарушение представляет собой фиброз. В определенных вариантах осуществления нарушение представляет собой SSc. В некоторых вариантах осуществления нарушение представляет собой IPF. В определенных вариантах осуществления антитело к TGFβ3 представляет собой моноклональное антитело, такое как химерное, гуманизированное или человеческое антитело. В некоторых вариантах осуществления антитело к TGFβ3 представляет собой фрагмент антитела, например, фрагмент Fv, Fab, Fab', scFv, диатело или фрагмент F(ab')2. В других вариантах осуществления антитело к TGFβ3 представляет собой полноразмерное антитело, например, интактное антитело на основе IgG1 или IgG4, такое как IgG1, или другой класс или изотип антитела, определенный в данном документе.
Варианты применения фармацевтических составов
В дополнительном аспекте в настоящем изобретении представлены фармацевтические составы, содержащие любое из селективных в отношении изоформы антител к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3), представленных в данном документе, например, для применения в любом из вышеуказанных терапевтических способов, варианты применения и лекарственные препараты. В одном варианте осуществления фармацевтический состав содержит любое из селективных в отношении изоформы антител к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела TGFβ3), представленных в данном документе, и фармацевтически приемлемый носитель. В другом варианте осуществления фармацевтический состав содержит любое из селективных в отношении изоформы антител к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3, например, антитело, имеющее аминокислотную последовательность VL и VH, как изображено на любой из Фиг. 12-16 и 18-24 или в таблицах 5, 6, 7, 12, 13, 14, 19, 20, 23 и 24), представленных в данном документе, и по меньшей мере одно дополнительное терапевтическое средство, например, как описано ниже.
Комбинированная терапия
Антитела по настоящему изобретению можно применять в терапии отдельно или в комбинации с другими средствами. Например, антитело по настоящему изобретению можно вводить совместно с по меньшей мере одним дополнительным терапевтическим средством. В некоторых вариантах осуществления терапевтическое средство выбрано из пирфенидона, нинтеданиба, микофенилата мофетила, ингибитора IL-6 (например, тоцилизумаба, сарилумаба), антитела к CTFG (например, FG-3019), ингибитора аутотаксина, ингибитора JAK, ингибитора IL-11 и сывороточного амилоида P (РТХ2). См. Raghu et al. European Respiratory Journal 2012 40: 2819) (FG-309, Ninou et al. (2018) Front Med (Lausanne); 5: 180 (ингибиторы аутотаксина). В предпочтительном варианте осуществления субъекту, нуждающемуся в этом, вводят антитело к TGFβ (например, антитело к TGFβ2, антитело к TGFβ2/3 или антитело к TGFβ3) в комбинации с тоцилизумабом. В предпочтительном варианте осуществления субъект, нуждающийся в этом, представляет собой пациента, страдающего SSc. В некоторых вариантах осуществления антитело к TGFβ представляет собой антитело к TGFβ3, которое селективно нейтрализует TGFβ3. Терапевтические дозировки тоцилизумаба, которые можно вводить субъекту, например, в комбинации с антителом к TGFβ3 (или антителом к TGFβ2 или антителом к TGFβ2/3) в соответствии с настоящим изобретением, описаны в патенте США №9539263, и включают, например, дозу, составляющую приблизительно 162 мг на дозу, вводимую, например, каждую неделю или каждые две недели, при этом предпочтительным путем введения является подкожный.
В определенных вариантах осуществления субъекту вводят антитело к TGFβ по настоящему изобретению в комбинации с терапевтическим средством для лечения IPF. Некоторые терапевтические средства ранее были описаны в качестве кандидатов или средств для лечения IPF. Они описаны в опубликованной литературе и рассмотрены, например, в Rafli et al., J. Thorac. Dis (2013) 5(l):48-73. Такие средства включают средства, которые имеют антиоксидантную, иммуносупрессорную и/или противовоспалительную активность, такие как N-ацетилцистеин; средства, которые имеют антифибротическую, противовоспалительную и/или антиоксидантную активность, такие как пирфенидон, пиридин для перорального введения, одобренный для клинического применения в лечении IPF; или антитело к интегрину αvβ6 (например, STX-100); средства, которые ингибируют фактор роста соединительной ткани (CTGF), такие как антитело к CTGF (например, FG-3019); средства, которые ингибируют рецепторы соматостатина, такие как аналоги соматостатина (например, SOM230, октреотид); средства, которые ингибируют IL-13, IL-4 и CCL2, такие как антитело к IL13 (например, QAX576, тралокинумаб, лебрикизумаб), антитело к IL4, комбинированное средство, направленное против Il13/против IL4 (например, биспецифическое антитело к IL13/к IL4, такое как SAR156597), ингибитор антитела к IL-6 (например, тоцилизумаб, сарилумаб), антитело к CCL2 (например, CNTO888); средства, которые имеют антиангиогенную, иммуномодулирующую и/или противовоспалительную активность, такие как талидомид или миноциклин; средства, которые ингибируют фермент, подобный лизилоксидазе 2 (LOXL2), такие как антитело к LOXL2 (например, GS-6624 [симтузумаб]); средства, которые ингибируют ангиогенез, такие как ингибитор тирозинкиназы, BIBF 1120, тетратиомолибдат; средства, которые ингибируют отложение внеклеточного матрикса и/или нарушают отложение коллагена, такие как доксициклин; средства, которые воздействуют на ренин-ангиотензиновую систему, такие как лозартан; и другие средства, которые имеют антипролиферативную и/или антифиброзную активность, такие как монооксид углерода.
Такие комбинированные виды терапии, отмеченные выше, включают комбинированное введение (когда два или более терапевтических средства включены в один или в отдельные составы), а также раздельное введение, в случае чего введение антитела по настоящему изобретению может происходить до, одновременно и/или после введения дополнительного терапевтического средства или средств. В одном варианте осуществления введение селективного в отношении изоформы антитела к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) и введение дополнительного терапевтического средства (например, ингибитора антитела к IL-6 (например, тозилумаба, сарилумаба) происходят в течение приблизительно одного месяца, или в течение приблизительно одной, двух или трех недель, или в течение приблизительно одного, двух, трех, четырех, пяти или шести дней, отделенных друг от друга.
В конкретном варианте осуществления комбинированная терапия включает введение субъекту, нуждающемуся в такой комбинированной терапии, например, пациенту, у которого диагностировано нарушение, связанное с TGFβ, например, фиброз, например, IPF, COPD, PF-ILD (например, SSc), фиброз печени (например, цирроз печени или хронический фиброз печени или другое фиброзное состояние или рак, описанные в данном документе), терапевтически эффективное количество антитела к TGFβ по настоящему изобретению (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) в комбинации со средством, выбранным из группы, состоящей из: N-ацетилцистеина; средств, которые имеют антифибротическую, противовоспалительную и/или антиоксидантную активность, таких как пирфенидон, пиридин для перорального введения, одобренный для клинического применения в лечении IPF; или антитела к интегрину αvβ6 (например, STX-100); средств, которые ингибируют фактор роста соединительной ткани (CTGF), такие как антитело к CTGF (например, FG-3019); средств, которые ингибируют рецепторы соматостатина, такие как аналоги соматостатина (например, SOM230, октреотид); средств, которые ингибируют IL-13, IL-4 и CCL2, таких как антитело к IL13 (например, QAX576, тралокинумаб, лебрикизумаб), антитела к IL4, комбинированного средства, направленного против Il13/против IL4 (например, биспецифического антитела к IL13/к IL4, такого как SAR156597), ингибитора антитела к IL-6 (например, тоцилизумаба, сарилумаба), антитела к CCL2 (например, CNTO888); средств, которые имеют антиангиогенную, иммуномодулирующую и/или противовоспалительную активность, таких как талидомид или миноциклин; средств, которые ингибируют фермент, подобный лизилоксидазе 2 (LOXL2), таких как антитело к LOXL2 (например, GS-6624 [симтузумаб]); средств, которые ингибируют ангиогенез, таких как ингибитор тирозинкиназы, BIBF 1120, тетратиомолибдат; средств, которые ингибируют отложение внеклеточного матрикса и/или нарушают отложение коллагена, такие как доксициклин; средств, которые воздействуют на ренин-ангиотензиновую систему, таких как лозартан; и других средств, которые имеют антипролиферативную и/или антифиброзную активность, таких как монооксид углерода.
Введение и составление
Антитело по настоящему изобретению (и любое дополнительное терапевтическое средство) можно вводить любым подходящим способом, включая парентеральное, внутрилегочное и интраназальное, и, если это необходимо для местного лечения, внутриочаговое введение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Введение дозы можно проводить любым подходящим способом, например, путем инъекций, таких как внутривенные или подкожные инъекции, частично в зависимости от того, является ли введение кратковременным или постоянным. В предпочтительных вариантах осуществления введение антитела к TGFβ, описанного в данном документе, осуществляется подкожно или внутривенно. В данном документе предусмотрены различные схемы введения доз, включая без ограничения однократное или многократное введение в различные временные точки, болюсное введение и импульсную инфузию.
Антитела по настоящему изобретению составляют, дозируют и вводят способом, соответствующим надлежащей медицинской практике. Факторы, которые необходимо учитывать в этом контексте, включают конкретное нарушение, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние отдельного пациента, причину нарушения, место доставки средства, применение введения, график введения и другие факторы, известные практикующим врачам. Антитело необязательно составляют с одним или более средствами, применяемыми в настоящее время для предупреждения или лечения рассматриваемого нарушения. Эффективное количество таких других средств зависит от количества антитела, присутствующего в составе, типа нарушения или лечения и других факторов, которые обсуждались выше. Другие средства обычно применяют в таких же дозировках и вводят путями, описанными в данном документе, или в дозировках, составляющих от около 1 до 99% дозировок, описанных в данном документе, или в любой дозировке и любым путем, которые эмпирически/клинически определены как подходящие.
Для предупреждения или лечения заболевания соответствующая дозировка антитела по настоящему изобретению (применяемого отдельно или в комбинации с одним или более другими дополнительными терапевтическими средствами) зависит от типа заболевания, подлежащего лечению, типа антитела, тяжести и течения заболевания, того, вводится ли антитело с профилактической или терапевтической целью, предыдущей терапии, истории болезни пациента и его ответа на антитело, а также решения лечащего врача. Антитело предпочтительно вводить пациенту за один раз или в течение серии курсов лечения. В зависимости от типа и тяжести заболевания, начальная кандидатная дозировка антитела для введения пациенту может составлять от приблизительно 1 мкг/кг до 15 мг/кг (например, 0,1 мг/кг - 10 мг/кг), например, за одно или более отдельных введений или в виде непрерывной инфузии. Одна типичная суточная дозировка может варьироваться от приблизительно 1 мкг/кг до 100 мг/кг или больше, в зависимости от факторов, упомянутых выше. В случае повторных введений в течение нескольких дней или более, в зависимости от состояния, лечение в общем случае проводят до достижения необходимой степени подавления симптомов заболевания. Одна типовая дозировка антитела находится в диапазоне от приблизительно 0,05 мг/кг до приблизительно 10 мг/кг. Таким образом, пациенту можно вводить одну или более доз, составляющих приблизительно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любую их комбинацию). Такие дозы можно вводить с интервалами, например, раз в неделю или раз в три недели (например, чтобы пациент получал от приблизительно двух до приблизительно двадцати или, например, приблизительно шести доз антитела). Можно вводить начальную более высокую нагрузочную дозу, за которой следует одна или более меньших доз. При этом можно применять другие схемы введения доз. Эффективность такой терапии легко контролировать с помощью традиционных методик и анализов.
«Субъект» или «индивидуум» в соответствии с любым из вышеуказанных вариантов осуществления может быть человеком. Следует понимать, что любой из вышеуказанных составов или терапевтических способов можно осуществлять, используя иммуноконъюгат по настоящему изобретению вместо селективного в отношении изоформы антитела к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) или в дополнение к нему.
И. Готовые изделия
В другом аспекте настоящего изобретения представлено готовое изделие, содержащее материалы, применяемые для лечения, предупреждения и/или диагностики вышеуказанных нарушений. Готовое изделие содержит контейнер и этикетку или вкладыш в упаковку на нем или связанные с ним. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, пакеты с раствором для внутривенного введения и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластмасса. Контейнер содержит композицию, которая сама по себе или в комбинации с другой композицией эффективна для лечения, предупреждения и/или диагностики патологического состояния, и может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет для внутривенного раствора или флакон с пробкой, прокалываемой гиподермической иглой для инъекций). По меньшей мере одно активное средство в композиции представляет собой антитело по настоящему изобретению. На этикетке или на вкладыше в упаковку указано, что композицию применяют для лечения выбранного патологического состояния. Кроме того, готовое изделие может содержать (а) первый контейнер с содержащейся в нем композицией, причем указанная композиция содержит антитело по настоящему изобретению; и (б) второй контейнер с содержащейся в нем композицией, причем указанная композиция содержит дополнительное цитотоксическое или иное терапевтическое средство. Готовое изделие в этом варианте осуществления может дополнительно содержать вкладыш в упаковку, на котором указано, что композиции можно применять для лечения конкретного патологического состояния. В качестве альтернативы или в качестве дополнения готовое изделие может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), фосфатно-солевой буферный раствор, раствор Рингера и раствор декстрозы. Оно также может содержать другие материалы, необходимые с коммерческой и пользовательской точки зрения, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Следует понимать, что любое из вышеуказанных готовых изделий может включать иммуноконъюгат по настоящему изобретению вместо селективного в отношении изоформы антитела к TGFβ (например, антитела к TGFβ2, антитела к TGFβ2/3 или антитела к TGFβ3) или в дополнение к нему.
III. ПРОНУМЕРОВАННЫЕ ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ
Вариант осуществления 1. Выделенное антитело к фактору некроза опухоли бета 3 (TGFβ3), где антитело селективно нейтрализует TGFβ3, и при этом антитело характеризуется одним или более из следующих свойств:
(а) антитело к TGFβ3 специфически связывается со шпилечной областью бета6/бета7 TGFβ3;
(б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3;
(в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFBR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3;
(г) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека;
(д) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи;
(е) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ1 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека);
(ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека);
(и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ;
(к) антитело к TGFβ3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 ко всем видам TGFβ;
(л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом CAT-192 к TGFβ1;
(п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3;
(р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9;
(с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, I388, L389, Y391, V392, G393, R394, Р396, K397 и V398 TGFβ3 человека; и
(т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3.
Вариант осуществления 2. Выделенное антитело к TGFβ3, где антитело содержит: (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9.
Вариант осуществления 3. Антитело к TGFβ3 по варианту осуществления 1 или варианту осуществления 2, где антитело содержит CDR-H2, имеющую аминокислотную последовательность SEQ ID NO: 5.
Вариант осуществления 4. Антитело к TGFβ3 по любому из вариантов осуществления 1-3, где антитело содержит аминокислотную последовательность вариабельной области тяжелой цепи (VH), имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37 и 42-50.
Вариант осуществления 5. Антитело к TGFβ3 по любому из вариантов осуществления 1-4, где антитело содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 23, 37 и 42-50.
Вариант осуществления 6. Антитело к TGFβ3 по любому из вариантов осуществления 1-5, где антитело содержит полную аминокислотную последовательность тяжелой (Н) цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 59, 64 и 65-72.
Вариант осуществления 7. Антитело к TGFβ3 по любому из вариантов осуществления 1-6, где антитело содержит полную аминокислотную последовательность тяжелой (Н) цепи, выбранную из группы, состоящей из SEQ ID NO: 29, 59, 64 и 65-72.
Вариант осуществления 8. Антитело к TGFβ3 по любому из вариантов осуществления 1-7, где антитело содержит аминокислотную последовательность вариабельной области легкой цепи (VL), имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41.
Вариант осуществления 9. Антитело к TGFβ3 по любому из вариантов осуществления 1-8, где антитело содержит полную аминокислотную последовательность легкой (L) цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63.
Вариант осуществления 10. Антитело к TGFβ3 по любому из вариантов осуществления 1-9, где антитело содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63.
Вариант осуществления 11. Антитело к TGFβ3 по любому из вариантов осуществления 1-10, где антитело содержит пару VH/VL, при этом пара VH/VL содержит аминокислотные последовательности (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8) и SEQ ID NO: 50/36 (v2.9).
Вариант осуществления 12. Антитело к TGFβ3 по любому из вариантов осуществления 1-11, где антитело содержит полную пару H/L-цепей, при этом полная пара H/L-цепей содержит аминокислотные последовательности (соответственно), выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.6), SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8) и SEQ ID NO: 72/58 (v2.9).
Вариант осуществления 13. Антитело к TGFβ3 по любому из вариантов осуществления 1-10, где антитело содержит VL SEQ ID NO: 22, содержащую одну или более модификаций каркасной области, выбранных из группы, состоящей из: 4L или 4М, 38Н или 38Q, 43А или 43Q и 58V.
Вариант осуществления 14. Антитело к TGFβ3 по любому из вариантов осуществления 1-10, где VL содержит набор модификаций каркасной области, выбранный из группы, состоящей из:
(i) 4L в FR1, 38Н и 43Q в FR2, 581 в FR3 (h2A10.v1 и h2A10.v2);
(ii) 4М в FR1 (h2A10.v1.1 и h2A10.v2.1);
(iii) 38Q в FR2 (h2A10.v1.2 и h2A10.v2.2);
(iv) 43А в FR2 (h2A10.v1.3 и h2A10.v2.3);
(v) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4);
(vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4);
(vii) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); и
(vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4).
Вариант осуществления 15. Антитело к TGFβ3 по любому из вариантов осуществления 1-10, 13 или 14, где антитело содержит VH SEQ ID NO: 23, содержащую одну или более модификаций каркасной области, выбранных из группы, состоящей из: 47L или 47W; 49А, 49S или 49G; 73D или 73N; и 76N, 78D или 78L, 78А или 78V.
Вариант осуществления 16. Антитело к TGFβ3 по варианту осуществления 15, где VH содержит набор модификаций каркасной области, выбранный из группы, состоящей из:
(i) 47L, 49А в FR2, 78V в FR3 (h2A10.v1);
(ii) 47L, 49A в FR2, 73D, 76S, 78V в FR3 (h2A10.v2);
(iii) 47W в FR2 (h2A10.v1.5);
(iv): 49G в FR2 (h2A10.v1.6);
(v) 78A в FR3 (h2A10.v1.7);
(vi) 47W в FR2 (h2A10.v2.5);
(vii) 49S в FR2 (h2A10.v2.6);
(viii) 73N в FR3 (h2A10.v2.7);
(ix) 76N в FR3 (h2A10.v2.8);
(x) 78L в FR3 (h2A10.v2.9); и
(xi) 49S в FR2, 76N, 78L в FR3 (h2A10.v3 и h2A10.v4).
Вариант осуществления 17. Антитело по варианту осуществления 13 или варианту осуществления 15, содержащее VL, где VL сохраняет лейцин (L) в положении 4 в каркасной области I и лейцин (L) в положении 47 в каркасной области II.
Вариант осуществления 18. Антитело по варианту осуществления 15 или варианту осуществления 17, где VH сохраняет D в положении 73 в каркасной области III VH из r2A10.
Вариант осуществления 19. Антитело к TGFβ3 по варианту осуществления 1 или варианту осуществления 2, где антитело к TGFβ3 содержит CDR-H2, имеющую аминокислотную последовательность SEQ ID NO: 159.
Вариант осуществления 20. Антитело к TGFβ3 по варианту осуществления 19, где антитело содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 52.
Вариант осуществления 21. Антитело к TGFβ3 по варианту осуществления 19 или варианту осуществления 20, где антитело содержит аминокислотную последовательность VH SEQ ID NO: 52.
Вариант осуществления 22. Антитело к TGFβ3 по вариантам осуществления 19-21, где антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74.
Вариант осуществления 23. Антитело к TGFβ3 по любому из вариантов осуществления 19-22, где антитело содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 74.
Вариант осуществления 24. Антитело к TGFβ3 по любому из вариантов осуществления 19-23, где антитело содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41.
Вариант осуществления 25. Антитело к TGFβ3 по любому из вариантов осуществления 19-24, где антитело содержит аминокислотную последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 22, 36 и 38-41.
Вариант осуществления 26. Антитело к TGFβ3 по любому из вариантов осуществления 19-25, где антитело содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63.
Вариант осуществления 27. Антитело к TGFβ3 по любому из вариантов осуществления 19-26, где антитело содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63.
Вариант осуществления 28. Антитело к TGFβ3 по варианту осуществления 19, где антитело содержит пару VH/VL, содержащую аминокислотные последовательности (соответственно) SEQ ID NO: 52/36.
Вариант осуществления 29. Антитело к TGFβ3 по варианту осуществления 19, где антитело содержит полную пару H/L-цепей, при этом пара H/L-цепей содержит аминокислотные последовательности (соответственно) SEQ ID NO: 74/58.
Вариант осуществления 30. Антитело к TGFβ3 по варианту осуществления 1 или варианту осуществления 2, где антитело содержит CDR-H2, имеющую аминокислотную последовательность SEQ ID NO: 34.
Вариант осуществления 31. Антитело к TGFβ3 по варианту осуществления 30, где антитело содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 51 или 55.
Вариант осуществления 32. Антитело к TGFβ3 по варианту осуществления 30 или варианту осуществления 31, где антитело содержит аминокислотную последовательность VH SEQ ID NO: 51 или 55.
Вариант осуществления 33. Антитело к TGFβ3 по вариантам осуществления 30-32, где антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 73 или 77.
Вариант осуществления 34. Антитело к TGFβ3 по любому из вариантов осуществления 30-33, где антитело содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 73.
Вариант осуществления 35. Антитело к TGFβ3 по любому из вариантов осуществления 30-33, где антитело содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 77.
Вариант осуществления 36. Антитело к TGFβ3 по любому из вариантов осуществления 30-35, где антитело содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41.
Вариант осуществления 37. Антитело к TGFβ3 по любому из вариантов осуществления 30-36, где антитело содержит аминокислотную последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 22, 36 и 38-41.
Вариант осуществления 38. Антитело к TGFβ3 по любому из вариантов осуществления 30-37, где антитело содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63.
Вариант осуществления 39. Антитело к TGFβ3 по любому из вариантов осуществления 30-38, где антитело содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63.
Вариант осуществления 40. Антитело к TGFβ3 по варианту осуществления 30, где антитело содержит пару VH/VL, при этом пара VH/VL содержит аминокислотные последовательности (соответственно) SEQ ID NO: 51/36 или SEQ ID NO: 55/54.
Вариант осуществления 41. Антитело к TGFβ3 по варианту осуществления 30, где антитело содержит полную пару H/L-цепей, при этом пара H/L-цепей содержит аминокислотные последовательности (соответственно) SEQ ID NO: 73/58 или SEQ ID NO: 77/76.
Вариант осуществления 42. Антитело к TGFβ3 по варианту осуществления 1 или варианту осуществления 2, где антитело содержит CDR-H2, имеющую аминокислотную последовательность SEQ ID NO: 35.
Вариант осуществления 43. Антитело к TGFβ3 по варианту осуществления 42, где антитело содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 53 или 57.
Вариант осуществления 44. Антитело к TGFβ3 по варианту осуществления 42 или варианту осуществления 43, где антитело содержит аминокислотную последовательность VH SEQ ID NO: 53.
Вариант осуществления 45. Антитело к TGFβ3 по варианту осуществления 42 или варианту осуществления 43, где антитело содержит аминокислотную последовательность VH SEQ ID NO: 57.
Вариант осуществления 46. Антитело к TGFβ3 по вариантам осуществления 42-45, где антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 75 или 79.
Вариант осуществления 47. Антитело к TGFβ3 по любому из вариантов осуществления 42-46, где антитело содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 75.
Вариант осуществления 48. Антитело к TGFβ3 по любому из вариантов осуществления 42-46, где антитело содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 79.
Вариант осуществления 49. Антитело к TGFβ3 по любому из вариантов осуществления 42-48, где антитело содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41.
Вариант осуществления 50. Антитело к TGFβ3 по любому из вариантов осуществления 42-49, где антитело содержит аминокислотную последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 22, 36 и 38-41.
Вариант осуществления 51. Антитело к TGFβ3 по любому из вариантов осуществления 42-50, где антитело содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63.
Вариант осуществления 52. Антитело к TGFβ3 по любому из вариантов осуществления 42-51, где антитело содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63.
Вариант осуществления 53. Антитело к TGFβ3 по варианту осуществления 42, где антитело содержит пару VH/VL, содержащую аминокислотные последовательности (соответственно) SEQ ID NO: 53/36 или SEQ ID NO: 57/56.
Вариант осуществления 54. Антитело к TGFβ3 по варианту осуществления 42, где антитело содержит полную пару H/L-цепей, при этом пара H/L-цепей содержит аминокислотные последовательности (соответственно) SEQ ID NO: 75/58 или SEQ ID NO: 79/78.
Вариант осуществления 55. Антитело к TGFβ3, содержащее:
(а) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9;
(б) пару VH/VL, при этом VH пары VH/VL содержит аминокислотную последовательность SEQ ID NO: 57 и VL пары VH/VL содержит аминокислотную последовательность SEQ ID NO: 56; или
(в) полную пару H/L-цепей, при этом Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79 и L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78.
Вариант осуществления 56. Выделенное антитело к TGFβ2/3, где антитело селективно нейтрализует TGFβ2 и TGFβ3, и при этом антитело характеризуется одним или более из следующих свойств:
(а) селективность антитела к TGFβ2/3 в отношении TGFβ2 и TGFβ3 по сравнению с TGFβ1 человека в отношении селективной нейтрализации достигается посредством прямого контакта антигенсвязывающего домена антитела с аминокислотным остатком Е373 TGFβ2 или TGFβ3 (нумерация TGFβ2 человека);
(б) антитело к TGFβ2/3 нейтрализует TGFβ2 и/или TGFβ3 посредством аллостерического механизма;
(в) антитело к TGFβ2/3 индуцирует конформационное изменение в гомодимере TGFβ2 и/или TGFβ3;
(г) антитело к TGFβ2/3 индуцирует конформационное изменение в гомодимере TGFβ2 и/или TGFβ3, при этом конформационное изменение включает совместное сжатие двух мономеров на несколько градусов;
(д) антитело к TGFβ2/3 представляет собой двухвалентное антитело или одновалентное антитело;
(е) антитело к TGFβ2/3 содержит (i) CDR тяжелой цепи, содержащую CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащую CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 15;
(ж) антитело к TGFβ2/3 специфически связывается с гомодимером TGFβ2, при этом гомодимер TGFβ2 имеет первый и второй мономер TGFβ2, и при этом антитело к TGFβ2/3 содержит антигенсвязывающий домен, который непосредственно контактирует с (i) аминокислотными остатками V313, Q314, D315, R320, L322, Y323, R328, D329, F345 и А347 первого мономера TGFβ2, и (ii) аминокислотными остатками N368, Т369, I370, N371, Р372, Е373, А374, S375, А376 и S377 второго мономера TGFβ2 (нумерация TGFβ2 человека);
(и) антитело к TGFβ2/3, как в (ж), где антигенсвязывающий домен находится в пределах 5 ангстрем от аминокислотных остатков TGFβ2 и/или TGFβ3;
(к) при этом антитело к TGFβ2/3 специфически связывается с тем же эпитопом на TGFβ3, что и в (ж); и
(л) антитело к TGFβ2/3 не нейтрализует TGFβ2 и/или TGFβ3 в одновалентной форме.
Вариант осуществления 57. Антитело к TGFβ2/3 по варианту осуществления 56, где антитело содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 27, 81, 83, 86-88, 93-100 и 102-105.
Вариант осуществления 58. Антитело к TGFβ2/3 по варианту осуществления 56 или варианту осуществления 57, где антитело содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 27, 81, 83, 86-88, 93-100 и 102-105.
Вариант осуществления 59. Антитело к TGFβ2/3 по любому из вариантов осуществления 56-58, где антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 33, 107, 109, 112-114 и 119-130.
Вариант осуществления 60. Антитело к TGFβ2/3 по любому из вариантов осуществления 56-59, где антитело содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 26, 80, 82, 84, 85, 89-92 и 101.
Вариант осуществления 61. Антитело к TGFβ2/3 по любому из вариантов осуществления 56-60, где антитело содержит аминокислотную последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 26, 80, 82, 84, 85, 89-92 и 101.
Вариант осуществления 62. Антитело к TGFβ2/3 по любому из вариантов осуществления 56-61, где антитело содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 32, 106, 108, 110, 111, 115-118 и 186.
Вариант осуществления 63. Антитело к TGFβ2/3 по любому из вариантов осуществления 56-62, где антитело содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 32, 106, 108, 110, 111, 115-118 и 186.
Вариант осуществления 64. Антитело к TGFβ2/3 по варианту осуществления 56, где антитело содержит пару VH/VL, при этом пара VH/VL содержит аминокислотные последовательности (соответственно), выбранные из группы, состоящей из SEQ ID NO: 27/26 (4А11 кролика), SEQ ID NO: 81/80 (v1), SEQ ID NO: 81/82 (v2), SEQ ID NO: 83/80 (v3), SEQ ID NO: 83/82 (v4), SEQ ID NO: 81/84 (v5), SEQ ID NO: 81/85 (v6), SEQ ID NO: 83/84 (v7), SEQ ID NO: 86/84 (v7.1), SEQ ID NO: 87/84 (v7.2), SEQ ID NO: 88/84 (v7.3), SEQ ID NO: 83/89 (v7.4), SEQ ID NO: 83/90 (v7.5), SEQ ID NO: 83/91 (v7.6), SEQ ID NO: 83/92 (v7.7), SEQ ID NO: 93/84 (v7.8), SEQ ID NO: 94/84 (v7.9), SEQ ID NO: 95/84 (v7.10), SEQ ID NO: 96/84 (v7.11), SEQ ID NO: 97/84 (v7.12), SEQ ID NO: 98/84 (v7.13), SEQ ID NO: 99/84 (v7.14), SEQ ID NO: 100/84 (v7.15), SEQ ID NO: 102/101 (v7.16), SEQ ID NO: 103/101 (v7.17), SEQ ID NO: 104/101 (v7.18), SEQ ID NO: 105/101 (v7.19) и SEQ ID NO: 83/85 (v8).
Вариант осуществления 65. Антитело к TGFβ2/3 по варианту осуществления 56, где антитело содержит полную пару H/L-цепей, при этом полная пара H/L-цепей содержит аминокислотные последовательности (соответственно), выбранные из группы, состоящей из SEQ ID NO: 32/33 (4А11 кролика), SEQ ID NO: 107/106 (v1), SEQ ID NO: 107/108 (v2), SEQ ID NO: 109/106 (v3), SEQ ID NO: 109/108 (v4), SEQ ID NO: 107/110 (v5), SEQ ID NO: 107/111 (v6), SEQ ID NO: 109/110 (v7), SEQ ID NO: 112/110 (v7.1), SEQ ID NO: 113/110 (v7.2), SEQ ID NO: 114/110 (v7.3), SEQ ID NO: 114/115 (v7.4), SEQ ID NO: 114/116 (v7.5), SEQ ID NO: 114/117 (v7.6), SEQ ID NO: 114/118 (v7.7), SEQ ID NO: 119/110 (v7.8), SEQ ID NO: 120/110 (v7.9), SEQ ID NO: 121/110 (v7.10), SEQ ID NO: 122/110 (v7.11), SEQ ID NO: 123/110 (v7.12), SEQ ID NO: 124/110 (v7.13), SEQ ID NO: 125/110 (v7.14), SEQ ID NO: 126/110 (v7.15), SEQ ID NO: 127/186 (v7.16), SEQ ID NO: 128/186 (v7.17), SEQ ID NO: 129/186 (v7.18), SEQ ID NO: 130/186 (v7.19) и SEQ ID NO: 114/111 (v8).
Вариант осуществления 66. Антитело к TGFβ2/3 по варианту осуществления 56, где антитело содержит VL, содержащую аминокислотную последовательность SEQ ID NO: 26, содержащую одну или более модификаций каркасной области, выбранных из группы, состоящей из: 2А или 2I, 4L, 36F или 36Y, 43Р или 43А и 58V или 58I.
Вариант осуществления 67. Антитело к TGFβ2/3 по варианту осуществления 56, где VL содержит набор модификаций каркасной области, выбранный из группы, состоящей из:
(i) 2А и 4L в FR1 и 36F в FR2 (h4A11.v1 и h4A11.v3);
(ii) 2A и 4L в FR1 и 36F и 43Р в FR2 (h4A11.v2 и h4A11.v4);
(iii) 2А в FR1, 36F и 43Р в FR2 и 58V в FR3 (h4A11.v5 и h4A11.v7);
(iv) 2А и 4L в FR1 и 36F в FR2 (h4A11.v6 и h4A11.v8);
(v) 2I в FR1 (h4A11.v7.4);
(vi) 36Y в FR2 (h4A11.v7.5);
(vii) 43A в FR2 (h4A11.v7.6);
(viii) 58I в FR3 (h4A11.v7.7); и
(ix) 2I в FR1, 43A в FR2, 58I в FR3 (h4A11.v7.16-19).
Вариант осуществления 68. Антитело к TGFβ2/3 по варианту осуществления 56, где антитело содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 27, содержащую одну или более модификаций каркасной области, выбранных из группы, состоящей из: делеции 1E, 2Q или 2V, 24V, 37V или 37I, 48I, 49G, 67F или 67V, 71K или 71V, 73S или 73Т, делеции 75K и 76N, 78V или 78F, 91F или 91Y, 105Р или 105Q.
Вариант осуществления 69. Антитело к TGFβ2/3 по варианту осуществления 56 или варианту осуществления 68, где VL содержит набор модификаций каркасной области, выбранный из группы, состоящей из:
(i) 2Q и 24V в FR1, 481 и 49G в FR2, 71K, 73S, 78V и 91F в FR3 и 105Р в FR4 (h4A11.v1, h4A11.v2, h4A11.v5, h4A11.v6);
(ii) 2Q в FR1, 37V в FR2, 67F, 71K, 73S, 78V и 91F в FR3 и 105P в FR4 (h4A11.v3, h4A11.v4, h4A11.v7, h4A11.v8);
(iii) делеции 1E в FR1 (h4A11.v7.1);
(iv) делеции 75K и 76N в FR3 (h4A11.v7.2);
(v) делеции 1E в FR1 и 75K76N в FR3 (h4A11.v7.3);
(vi) 2V в FR1 (h4A11.v7.8);
(vi) 37I в FR2 (h4A11.v7.9);
(vii) 67V в FR3 (h4A11.v7.10);
(viii) 71V в FR3 (h4A11.v7.11);
(ix) 73T в FR3 (h4A11.v7.12);
(x) 78F в FR3 (h4A11.v7.13);
(xi) 91Y в FR3 (h4A11.v7.14);
(xii) 105Q в FR4 (h4A11.v7.15);
(xiii) 2V в FR1, 37I в FR2, 67V, 73T, 78F в FR3, 105Q в FR4 (h4A11.v7.16);
(xiv) 2V в FR1, 37I в FR2, 67V, 73T, 91Y в FR3, 105Q в FR4 (h4A11.v7.17);
(xv) 2V в FR1, 37I в FR2, 67V, 73T в FR3, 105Q в FR4 (h4A11.v7.18); и
(xvi) 2V в FR1, 37I в FR2, 67V, 73T, делеции 75K и 76N в FR3, 105Q в FR4 (h4A11.v7.19).
Вариант осуществления 70. Выделенное антитело к TGFβ2, где антитело содержит:
(a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17, и CDR -Н3 имеет аминокислотную последовательность SEQ ID NO: 18; и
(б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20, и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21.
Вариант осуществления 71. Антитело к TGFβ2 по варианту осуществления 70, где антитело селективно нейтрализует TGFβ2.
Вариант осуществления 72. Антитело к TGFβ2 по варианту осуществления 71, где антитело:
(а) имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ;
(б) имеет сниженную токсичность у грызунов по сравнению с антителом 1D11 ко всем видам TGFβ;
(в) имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом; и/или
(г) имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом.
Вариант осуществления 73. Антитело к TGFβ2 по варианту осуществления 70 или варианту осуществления 71, где антитело содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 132 и 138-142.
Вариант осуществления 74. Антитело к TGFβ2 по варианту осуществления 73, где антитело содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 25, 132 и 138-142.
Вариант осуществления 75. Антитело к TGFβ2 по варианту осуществления 70 или варианту осуществления 71, где антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 31, 146 и 152-156.
Вариант осуществления 76. Антитело к TGFβ2 по варианту осуществления 75, где антитело содержит полную аминокислотную последовательность Н-цепи, выбранную из группы, состоящей из SEQ ID NO: 31, 146 и 152-156.
Вариант осуществления 77. Антитело к TGFβ2 по варианту осуществления 70 или варианту осуществления 71, где антитело содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 24, 131, 133-137, 143 и 144.
Вариант осуществления 78. Антитело к TGFβ2 по варианту осуществления 77, где антитело содержит аминокислотную последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 24, 131, 133-137, 143 и 144.
Вариант осуществления 79. Антитело к TGFβ2 по варианту осуществления 70 или варианту осуществления 71, где антитело содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 30, 145, 147-151, 157 и 158.
Вариант осуществления 80. Антитело к TGFβ2 по варианту осуществления 79, где антитело содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 30, 145, 147-151, 157 и 158.
Вариант осуществления 81. Антитело к TGFβ2 по варианту осуществления 70 или варианту осуществления 71, где антитело содержит пару VH/VL, при этом пара VH/VL содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 25/24 (6F12 кролика), SEQ ID NO: 132/131 (v1), SEQ ID NO: 132/133 (v1.1), SEQ ID NO: 132/134 (v1.2), SEQ ID NO: 132/135 (v1.3), SEQ ID NO: 132/136 (v1.4), SEQ ID NO: 132/137 (v1.5), SEQ ID NO: 138/131 (v1.6), SEQ ID NO: 139/131 (v1.7), SEQ ID NO: 140/131 (v1.8), SEQ ID NO: 141/131 (v1.9), SEQ ID NO: 142/131 (v2), SEQ ID NO: 132/143 (v3) и SEQ ID NO: 142/144 (v4).
Вариант осуществления 82. Антитело к TGFβ2 по варианту осуществления 70 или варианту осуществления 71, где антитело содержит полную пару H/L-цепей, при этом полная пара H/L-цепей содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 31/30 (6F12 кролика), SEQ ID NO: 146/145 (v1), SEQ ID NO: 146/147 (v1.1), SEQ ID NO: 146/148 (v1.2), SEQ ID NO: 146/149 (v1.3), SEQ ID NO: 146/150 (v1.4), SEQ ID NO: 146/151 (v1.5), SEQ ID NO: 152/145 (v1.6), SEQ ID NO: 153/145 (v1.7), SEQ ID NO: 154/145 (v1.8), SEQ ID NO: 155/145 (v1.9), SEQ ID NO: 156/145 (v2), SEQ ID NO: 146/157 (v3) и SEQ ID NO: 156/158 (v4).
Вариант осуществления 83. Антитело к TGFβ2 по варианту осуществления 70 или варианту осуществления 71, где антитело содержит VL, содержащую аминокислотную последовательность SEQ ID NO: 24, содержащую одну или более мутаций каркасной области, выбранных из группы, состоящей из 43S или 43А, 66G, 69Т, 71F и 87Y.
Вариант осуществления 84. Антитело к TGFβ2 по варианту осуществления 83, где VL содержит набор мутаций каркасной области, выбранный из группы, состоящей из:
(i) 43S в FR2 и 66Е, 69Р, 71Y и 87F в FR3 (h6F12.v1 и h6F12.v2);
(ii) 43S в FR2 и 58V, 66Е, 69Р, 71Y и 87F в FR3 (h6F12.v3 и h6F12.v4);
(iii) 43А в FR2 (h6F12.v1.1);
(iv) 66G в FR3 (h6F12.v1.2);
(v) 69T в FR3 (h6F12.v1.3);
(vi) 71F в FR3 (h6F12.v1.4); и
(vii) 87Y в FR3 (h6F12.v1.5).
Вариант осуществления 85. Антитело к TGFβ2 по варианту осуществления 70 или варианту осуществления 71, где антитело содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 25, содержащую одну или более мутаций каркасной области, выбранных из группы, состоящей из 37V или 37I, 48М или 48L, 49G или 49А, 67L, 71K и 78V, и 105Р или 105R.
Вариант осуществления 86. Антитело к TGFβ2 по варианту осуществления 85, где VH содержит набор мутаций каркасной области, выбранный из группы, состоящей из:
(i) 37V, 48М и 49G в FR2 и 105Р в FR4 (h6F12.v1 и h6F12.v3);
(ii) 37V и 48М в FR2, 67L, 71K и 78V в FR3 и 105Р в FR4 (h6F12.v2 и h6F12.v4);
(iii) 37I в FR2 (h6F12.v1.6);
(iv) 48L в FR2 (h6F12.v1.7);
(v) 49A в FR2 (h6F12.v1.8);
(vi) 105R в FR4 (h6F12.v1.9);
(vii) 37V, 48M и 49G в FR2 и 105P в FR4 (h6F12.v1 и h6F12.v3);
(viii) 37V и 48M в FR2, 67L, 71K и 78V в FR3 и 105Р в FR4 (6F12.v2 и h6F12.v4);
(ix) 37I в FR2 (h6F12.v1.6);
(x) 48L в FR2 (h6F12.v1.7);
(xi) 49A в FR2 (h6F12.v1.8); и
(xii) 105R в FR4 (h6F12.v1.9).
Вариант осуществления 87. Антитело к TGFβ3 по любому из вариантов осуществления 1-55 или антитело к TGFβ2/3 по любому из вариантов осуществления 56-68, где антитело специфически связывается с TGFβ3 человека.
Вариант осуществления 88. Антитело к TGFβ3 по любому из вариантов осуществления 1-55, где антитело специфически связывается как с незрелой, так и со зрелой формами TGFβ3.
Вариант осуществления 89. Антитело к TGFβ2/3 по любому из вариантов осуществления 56-68 или антитело к TGFβ2 по любому из вариантов осуществления 70-83, где антитело специфически связывается с TGFβ2 человека.
Вариант осуществления I-1. Выделенное антитело к фактору некроза опухоли бета 3 (TGFβ3), где антитело селективно нейтрализует TGFβ3, и при этом антитело характеризуется одним или более из следующих свойств:
(р) антитело к TGFβ3 содержит: (i) CDR тяжелой цепи, содержащие CDR-Н1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9;
(а) антитело к TGFβ3 специфически связывается со шпилечной областью бета6/бета7 TGFβ3;
(б) связывание антитела к TGFβ3 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3;
(в) связывание антитела к TGFβ3 с TGFβ3 блокирует связывание TGFBR2 и ингибирует связывание сигнальных рецепторов TGFBR1/TGFBR2 с TGFβ3;
(г) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека;
(д) антитело к TGFβ3 непосредственно контактирует с аминокислотным остатком R394 в шпилечной области бета6/бета7 TGFβ3 человека, а остаток R394 TGFβ3 образует ионный солевой мостик с антителом к TGFβ3 в CDR2 тяжелой цепи;
(е) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ1 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками Т387, L389 и Т395 TGFβ3 (нумерация TGFβ3 человека);
(ж) изоформная селективность антитела к TGFβ3 в отношении TGFβ3 по сравнению с TGFβ2 достигается посредством прямого контакта антигенсвязывающего домена антитела к TGFβ3 с аминокислотными остатками R325, R394 и V398 TGFβ3 (нумерация TGFβ3 человека);
(и) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом 1D11 ко всем видам TGFβ;
(к) антитело к TGFβ3 имеет сниженную токсичность у грызунов или яванских макаков по сравнению с антителом 1D11 ко всем видам TGFβ;
(л) антитело к TGFβ3 имеет сниженную токсичность по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(м) антитело к TGFβ3 имеет сниженную токсичность у грызунов по сравнению с низкомолекулярным ингибитором всех видов TGFβ галунисертибом;
(н) антитело к TGFβ3 имеет сниженную токсичность по сравнению с антителом CAT-192 к TGFβ1;
(п) антитело к TGFβ3 имеет сниженную токсичность по сравнению с селективным в отношении изоформы антителом к TGFβ2 и/или антителом к TGFβ2/3;
(с) антитело к TGFβ3 содержит антигенсвязывающий домен, который непосредственно контактирует с аминокислотными остатками R325, K331, W332, Н334, Е335, Т387, I388, L389, Y391, V392, G393, R394, Р396, K397 и V398 TGFβ3 человека; и
(т) антитело к TGFβ3, как в (с), где антигенсвязывающий домен находится в пределах 15-8, 8, 8-5, 7-5, 6-5 или 5 ангстрем от аминокислотных остатков TGFβ3.
Вариант осуществления I-2. Выделенное антитело к TGFβ3, где антитело содержит: (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9.
Вариант осуществления I-3. Антитело к TGFβ3 по варианту осуществления I-1 или варианту осуществления I-2 где антитело содержит аминокислотную последовательность вариабельной области тяжелой цепи (VH), имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 23, 37, 42-53, 55 и 57.
Вариант осуществления I-4. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-3, где антитело содержит аминокислотную последовательность VH, выбранную из группы, состоящей из SEQ ID NO: 23, 37, 42-53, 55 и 57.
Вариант осуществления I-5. Антитело к TGFβ3 по любому из варианту осуществления I-1-I-4, где антитело содержит полную аминокислотную последовательность тяжелой (Н) цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 59, 64, 65-75, 77 и 79.
Вариант осуществления I-6. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-5, где антитело содержит полную аминокислотную последовательность тяжелой (Н) цепи, выбранную из группы, состоящей из SEQ ID NO: 29, 59, 64, 65-75, 77 и 79.
Вариант осуществления I-7. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-6, где антитело содержит аминокислотную последовательность вариабельной области легкой цепи (VL), имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41, 54 и 56.
Вариант осуществления I-8. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-7, где антитело содержит аминокислотную последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 22, 36, 38-41, 54 и 56.
Вариант осуществления I-9. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-8, где антитело содержит полную аминокислотную последовательность легкой (L) цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58, 60-63, 76 и 78.
Вариант осуществления I-10. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-9, где антитело содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63, 76 и 78.
Вариант осуществления I-11. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-10, где антитело содержит пару VH/VL, при этом пара VH/VL содержит аминокислотные последовательности (соответственно), выбранные из группы, состоящей из SEQ ID NO: 23/22 (2А10 крысы), SEQ ID NO: 37/36 (v1), SEQ ID NO: 37/38 (v1.1), SEQ ID NO: 37/39 (v1.2), SEQ ID NO: 37/40 (v1.3), SEQ ID NO: 37/41 (v1.4), SEQ ID NO: 42/36 (v1.5), SEQ ID NO: 43/36 (v1.6), SEQ ID NO: 44/36 (v1.7), SEQ ID NO: 45/36 (v2), SEQ ID NO: 45/38 (v2.1), SEQ ID NO: 45/39 (v2.2), SEQ ID NO: 45/40 (v2.3), SEQ ID NO: 45/41 (v2.4), SEQ ID NO: 46/36 (v2.5), SEQ ID NO: 47/36 (v2.6), SEQ ID NO: 48/36 (v2.7), SEQ ID NO: 49/36 (v2.8), SEQ ID NO: 50/36 (v2.9), SEQ ID NO: 51/36 (h2A10.v2.N54S), SEQ ID NO: 52/36 (h2A10.v2.N54Q), SEQ ID NO: 53/36 (h2A10.v2.T56A), SEQ ID NO: 55/54 (v3) и SEQ ID NO: 57/56 (v4).
Вариант осуществления I-12. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-11, где антитело содержит полную пару H/L-цепей, при этом полная пара H/L-цепей содержит аминокислотные последовательности (соответственно), выбранные из группы, состоящей из SEQ ID NO: 29/28 (2А10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.6), SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8), SEQ ID NO: 72/58 (v2.9), SEQ ID NO: 73/58 (h2A10.v2.N54S), SEQ ID NO: 74/58 (h2A10.v2.N54Q), SEQ ID NO: 75/58 (h2A10.v2.T56A), SEQ ID NO: 77/76 (v3) и SEQ ID NO: 79/78 (v4).
Вариант осуществления I-13. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-10, где антитело содержит VL SEQ ID NO: 22, содержащую одну или более модификаций каркасной области, выбранных из группы, состоящей из: 4L или 4М, 38Н или 38Q, 43А или 43Q и 58V.
Вариант осуществления I-14. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-10, где VL содержит набор модификаций каркасной области, выбранный из группы, состоящей из:
(i) 4L в FR1, 38Н и 43Q в FR2, 58I в FR3 (h2A10.v1 и h2A10.v2);
(ii) 4М в FR1 (h2A10.v1.1 и h2A10.v2.1);
(iii) 38Q в FR2 (h2A10.v1.2 и h2A10.v2.2);
(iv) 43А в FR2 (h2A10.v1.3 и h2A10.v2.3);
(v) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4);
(vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4);
(vii) 58V в FR3 (h2A10.v1.4 и h2A10.v2.4); и
(vi) 38Q, 43A в FR2, 58V в FR3 (h2A10.v3 и h2A10.v4).
Вариант осуществления I-15. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-10, варианту осуществления I-13 и варианту осуществления I-14, где антитело содержит VH SEQ ID NO: 23, содержащую одну или более модификаций каркасной области, выбранных из группы, состоящей из: 47L или 47W; 49A, 49S или 49G; 73D или 73N; и 76N, 78D или 78L, 78А или 78V.
Вариант осуществления I-16. Антитело к TGFβ3 по варианту осуществления I-15, где VH содержит набор модификаций каркасной области, выбранный из группы, состоящей из:
(i) 47L, 49А в FR2, 78V в FR3 (h2A10.v1);
(ii) 47L, 49А в FR2, 73D, 76S, 78V в FR3 (h2A10.v2);
(iii) 47W в FR2 (h2A10.v1.5); (iv): 49G в FR2 (h2A10.v1.6);
(v) 78A в FR3 (h2A10.v1.7);
(vi) 47W в FR2 (h2A10.v2.5);
(vii) 49S в FR2 (h2A10.v2.6);
(viii) 73N в FR3 (h2A10.v2.7);
(ix) 76N в FR3 (h2A10.v2.8);
(x) 78L в FR3 (h2A10.v2.9); и
(xi) 49S в FR2, 76N, 78L в FR3 (h2A10.v3 и h2A10.v4).
Вариант осуществления I-17. Антитело по варианту осуществления I-13 или варианту осуществления I-15, содержащее VL, где VL сохраняет лейцин (L) в положении 4 в каркасной области I и лейцин (L) в положении 47 в каркасной области II.
Вариант осуществления I-18. Антитело по варианту осуществления I-15 или варианту осуществления I-17, где VH сохраняет D в положении 73 в каркасной области III VH из r2A10.
Вариант осуществления I-19. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-12 где антитело содержит CDR-H2, имеющую аминокислотную последовательность SEQ ID NO: 5.
Вариант осуществления I-20. Антитело к TGFβ3 по любому из варианту осуществления I-1-I-12 где антитело к TGFβ3 содержит CDR-H2, имеющую аминокислотную последовательность SEQ ID NO: 159.
Вариант осуществления I-21. Антитело к TGFβ3 по варианту осуществления I-20, где антитело содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 52.
Вариант осуществления I-22. Антитело к TGFβ3 по варианту осуществления I-20 или варианту осуществления I-21, где антитело содержит аминокислотную последовательность VH SEQ ID NO: 52.
Вариант осуществления I-23. Антитело к TGFβ3 по вариантам осуществления I-20-I-22, где антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 74.
Вариант осуществления I-24. Антитело к TGFβ3 по любому из вариантов осуществления I-20-I-23, где антитело содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 74.
Вариант осуществления I-25. Антитело к TGFβ3 по любому из вариантов осуществления I-20-I-24, где антитело содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 22, 36 и 38-41.
Вариант осуществления I-26. Антитело к TGFβ3 по любому из вариантов осуществления I-20-I-25, где антитело содержит аминокислотную последовательность VL, выбранную из группы, состоящей из SEQ ID NO: 22, 36 и 38-41.
Вариант осуществления I-27. Антитело к TGFβ3 по любому из вариантов осуществления I-20-I-26, где антитело содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 28, 58 и 60-63.
Вариант осуществления I-28. Антитело к TGFβ3 по любому из вариантов осуществления I-20-I-27, где антитело содержит полную аминокислотную последовательность L-цепи, выбранную из группы, состоящей из SEQ ID NO: 28, 58 и 60-63.
Вариант осуществления I-29. Антитело к TGFβ3 по варианту осуществления I-20, где антитело содержит пару VH/VL, содержащую аминокислотные последовательности (соответственно) SEQ ID NO: 52/36.
Вариант осуществления I-30. Антитело к TGFβ3 по варианту осуществления I-20, где антитело содержит полную пару H/L-цепей, при этом пара H/L-цепей содержит аминокислотные последовательности (соответственно) SEQ ID NO: 74/58.
Вариант осуществления I-31. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-12, где антитело содержит CDR-H2, имеющую аминокислотную последовательность SEQ ID NO: 34.
Вариант осуществления I-32. Антитело к TGFβ3 по варианту осуществления I-31, где антитело содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 55.
Вариант осуществления I-33. Антитело к TGFβ3 по варианту осуществления I-31 или варианту осуществления I-32, где антитело содержит аминокислотную последовательность VH SEQ ID NO: 55.
Вариант осуществления I-34. Антитело к TGFβ3 по вариантам осуществления I-31-I-33, где антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 77.
Вариант осуществления I-35. Антитело к TGFβ3 по любому из вариантов осуществления I-31-I-34, где антитело содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 77.
Вариант осуществления I-36. Антитело к TGFβ3 по вариантам осуществления I-31-I-35, где антитело содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 54.
Вариант осуществления I-37. Антитело к TGFβ3 по любому из вариантов осуществления I-31-I-37, где антитело содержит аминокислотную последовательность VL SEQ ID NO: 54.
Вариант осуществления I-38. Антитело к TGFβ3 по вариантам осуществления I-31-I-37, где антитело содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 76.
Вариант осуществления I-39. Антитело к TGFβ3 по любому из вариантов осуществления I-31-I-38, где антитело содержит полную аминокислотную последовательность L-цепи SEQ ID NO: 76.
Вариант осуществления I-40. Антитело к TGFβ3 по варианту осуществления I-31, где антитело содержит пару VH/VL, при этом пара VH/VL содержит аминокислотные последовательности (соответственно) SEQ ID NO: 55/54.
Вариант осуществления I-41. Антитело к TGFβ3 по варианту осуществления I-31, где антитело содержит полную пару H/L-цепей, при этом пара H/L-цепей содержит аминокислотные последовательности (соответственно) SEQ ID NO: 77/76.
Вариант осуществления I-42. Антитело к TGFβ3 по любому из вариантов осуществления I-1-I-12, где антитело содержит CDR-H2, имеющую аминокислотную последовательность SEQ ID NO: 35.
Вариант осуществления I-43. Антитело к TGFβ3 по варианту осуществления I-42, где антитело содержит аминокислотную последовательность VH, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 57.
Вариант осуществления I-44. Антитело к TGFβ3 по варианту осуществления I-42 или варианту осуществления I-43, где антитело содержит аминокислотную последовательность VH SEQ ID NO: 57.
Вариант осуществления I-45. Антитело к TGFβ3 по вариантам осуществления I-42-I-44, где антитело содержит полную аминокислотную последовательность Н-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 79.
Вариант осуществления I-46. Антитело к TGFβ3 по любому из вариантов осуществления I-42-I-45, где антитело содержит полную аминокислотную последовательность Н-цепи SEQ ID NO: 79.
Вариант осуществления I-47. Антитело к TGFβ3 по любому из вариантов осуществления I-42-I-46, где антитело содержит аминокислотную последовательность VL, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 56.
Вариант осуществления I-48. Антитело к TGFβ3 по любому из вариантов осуществления I-42-I-47, где антитело содержит аминокислотную последовательность VL SEQ ID NO: 56.
Вариант осуществления I-49. Антитело к TGFβ3 по вариантам осуществления I-42-I-48, где антитело содержит полную аминокислотную последовательность L-цепи, имеющую по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности последовательности аминокислотной последовательности SEQ ID NO: 76.
Вариант осуществления I-50. Антитело к TGFβ3 по любому из вариантов осуществления I-42-I-49, где антитело содержит полную аминокислотную последовательность L-цепи SEQ ID NO: 76.
Вариант осуществления I-51. Антитело к TGFβ3 по варианту осуществления I-42, где антитело содержит пару VH/VL, содержащую аминокислотные последовательности (соответственно) SEQ ID NO: 57/56.
Вариант осуществления I-52. Антитело к TGFβ3 по варианту осуществления I-42, где антитело содержит полную пару H/L-цепей, при этом пара H/L-цепей содержит аминокислотные последовательности (соответственно) SEQ ID NO: 79/78.
Вариант осуществления I-53. Антитело к TGFβ3, содержащее:
(а) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9;
(б) пару VH/VL, при этом VH пары VH/VL содержит аминокислотную последовательность SEQ ID NO: 57 и VL пары VH/VL содержит аминокислотную последовательность SEQ ID NO: 56; или
(в) полную пару H/L-цепей, при этом Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79 и L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78.
Вариант осуществления 90. Антитело по любому из вариантов осуществления 1-89 или вариантов осуществления I-1-I-53, которое представляет собой моноклональное антитело.
Вариант осуществления 91. Антитело по любому из вариантов осуществления 1-89 или вариантов осуществления I-1-I-53, которое представляет собой человеческое, гуманизированное или химерное антитело.
Вариант осуществления 92. Антитело по любому из вариантов осуществления 1-89 или вариантов осуществления I-1-I-53, которое представляет собой фрагмент антитела.
Вариант осуществления 93. Антитело по любому из вариантов осуществления 1-89 или вариантов осуществления I-1-I-53, содержащее Fc-область человека, которая относится к изотипу IgG1 или IgG4.
Вариант осуществления 94. Антитело по варианту осуществления 93, содержащее Fc-область человека, которая относится к изотипу IgG1.
Вариант осуществления 95. Антитело по варианту осуществления 93 или варианту осуществления 94, где Fc-область содержит модификацию, которая приводит к удалению сайта гликозилирования в положении аминокислотного остатка N297 (EU-нумерация согласно Kabat).
Вариант осуществления 96. Антитело по варианту осуществления 95, где модификация представляет собой мутацию, выбранную из N297G или N297A.
Вариант осуществления 97. Антитело по варианту осуществления 96, где модификация представляет собой мутацию N297G.
Вариант осуществления 98. Антитело по любому из вариантов осуществления 93-97, где Fc-область модифицирована с удалением эффекторной функции.
Вариант осуществления 99. Антитело по любому из вариантов осуществления 1-98, вариантов осуществления I-1-I-53, где антитело имеет Cmax, составляющую приблизительно 230-260 мкг/мл, и/или период полужизни (t½), составляющий от приблизительно 15 до 16 дней.
Вариант осуществления 100. Выделенная нуклеиновая кислота, кодирующая антитело по любому из вариантов осуществления 1-99 или вариантов осуществления I-1-I-53.
Вариант осуществления 101. Клетка-хозяин, содержащая нуклеиновую кислоту по варианту осуществления 100.
Вариант осуществления 102. Способ получения антитела, включающий культивирование клетки-хозяина по варианту осуществления 101, таким образом, чтобы получить антитело.
Вариант осуществления 103. Способ по варианту осуществления 102, дополнительно включающий выделение антитела из клетки-хозяина.
Вариант осуществления 104. Антитело, полученное с помощью способа по варианту осуществления 102 или варианту осуществления 103.
Вариант осуществления 105. Иммуноконъюгат, содержащий антитело по варианту осуществления 1 и цитотоксическое средство.
Вариант осуществления I-105. Иммуноконъюгат, содержащий антитело по любому из вариантов осуществления 1-89 или вариантов осуществления I-1-I-53 и цитотоксическое средство.
Вариант осуществления 106. Фармацевтический состав, содержащий антитело по любому из вариантов осуществления 1-99 или вариантов осуществления I-1-I-53, или иммуноконъюгат по варианту осуществления 105, или иммуноконъюгат по варианту осуществления I-105, и фармацевтически приемлемый носитель.
Вариант осуществления 107. Фармацевтический состав, содержащий антитело по варианту осуществления 106, дополнительно содержащий дополнительное терапевтическое средство.
Вариант осуществления 108. Фармацевтический состав по варианту осуществления 107, где дополнительное терапевтическое средство выбрано из группы, состоящей из пирфенидона, нинтеданиба, микофенилата мофетила, ингибитора IL-6 (например, тоцилизумаба), антитела к CTFG (например, FG-3019), ингибитора аутотаксина, ингибитора JAK, ингибитора IL-11 и РТХ2.
Вариант осуществления 109. Антитело по любому из вариантов осуществления 1-99 или вариантов осуществления I-1-I-53 для применения в качестве лекарственного средства.
Вариант осуществления 110. Антитело по любому из вариантов осуществления 1-99 или вариантов осуществления I-1-I-53 для применения в лечении нарушения, связанного с TGFβ.
Вариант осуществления 111. Антитело по варианту осуществления 1 и антитело по варианту осуществления 70 для применения в комбинации для лечения нарушения, связанного с TGFβ.
Вариант осуществления 112. Антитела по варианту осуществления 110 или варианту осуществления 111, где нарушение, связанное с TGFβ, представляет собой фиброз.
Вариант осуществления 113. Антитела по варианту осуществления 112, где фиброз представляет собой фиброзное состояние легких, печени, сердца, почек, поджелудочной железы, глаз и/или кожи.
Вариант осуществления 114. Антитело по любому из вариантов осуществления 1-99 или вариантов осуществления I-1-I-53 для применения в производстве лекарственного препарата для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или для ингибирования синтеза нового коллагена.
Вариант осуществления 115. Применение антитела по любому из вариантов осуществления 1-99 или вариантов осуществления I-1-I-53 в производстве лекарственного препарата для лечения у субъекта нарушения, связанного с TGFβ.
Вариант осуществления 116. Применение по варианту осуществления 115, где нарушение, связанное с TGFβ, представляет собой фиброз, при этом необязательно фиброз представляет собой фиброзное состояние легких, печени, сердца, почек, поджелудочной железы, глаз и/или кожи.
Вариант осуществления 117. Применение по любому из вариантов осуществления 1-99 или вариантов осуществления I-1-I-53 в производстве лекарственного препарата для ингибирования TGFBR-зависимой передачи сигналов SMAD, для ингибирования сборки сигнальных комплексов TGFβ-TGFBR, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, для ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или для ингибирования синтеза нового коллагена.
Вариант осуществления 118. Способ лечения субъекта, у которого имеется нарушение, связанное с TGFβ, при этом способ включает введение субъекту, нуждающемуся в этом, эффективного количества антитела по любому из вариантов осуществления 1-99 или вариантов осуществления I-1-I-53, или введение субъекту фармацевтического состава по варианту осуществления 106.
Вариант осуществления 119. Способ ингибирования TGFBR-зависимой передачи сигналов SMAD, ингибирования сборки сигнальных комплексов TGFβ-TGFBR, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или ингибирования синтеза нового коллагена у субъекта, включающий введение субъекту, нуждающемуся в этом, эффективного количества антитела по любому из вариантов осуществления 1-99 или вариантов осуществления I-1-I-53 для ингибирования TGFBR-зависимой передачи сигналов SMAD, ингибирования сборки сигнальных комплексов TGFβ-TGFBR, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR1/R2, ингибирования передачи сигналов TGFβ посредством комплекса TGFBR2/ALK1, облегченной эндоглином, и/или ингибирования синтеза нового коллагена у субъекта.
Вариант осуществления 120. Способ по варианту осуществления 118 или варианту осуществления 119, дополнительно включающий введение субъекту дополнительного терапевтического средства.
Вариант осуществления 121. Способ по варианту осуществления 120, где дополнительное терапевтическое средство выбрано из группы, состоящей из пирфенидона, нинтеданиба, микофенилата мофетила, ингибитора IL-6 (например, тоцилизумаба, сарилумаба), антитела к CTFG (например, FG-3019), ингибитора аутотаксина и РТХ2.
Вариант осуществления 122. Способ по любому из вариантов осуществления 118-121, включающий введение субъекту эффективного количества антитела к TGFβ3 по п. 1 и эффективного количества антитела к TGFβ2 по варианту осуществления 70.
Вариант осуществления 123. Способ по любому из вариантов осуществления 118-122, где у субъекта имеется нарушение, связанное с TGFβ, которое представляет собой фиброз.
Вариант осуществления 124. Способ по варианту осуществления 123, где фиброз представляет собой фиброз легких, выбранный из группы, состоящей из идиопатического фиброза легких (IPF), идиопатического фиброза верхних долей легких (болезнь Амитани), семейного фиброза легких, фиброза легких (например, фиброза легких, вторичного по отношению к системным воспалительным заболеваниям, таким как ревматоидный артрит, склеродермия, волчанка, криптогенный фиброзирующий альвеолит, хроническая обструктивная болезнь легких (COPD) или хроническая астма), муковисцидоз, неспецифическая интерстициальная пневмония (NSIP), криптогенная организующаяся пневмония (СОР), прогрессирующий массивный фиброз, склеродермия/системный склероз (SSc, включая ограниченную кожную (lcSSc) и диффузную кожную (dcSSc) формы и связанное с SSc интерстициальное заболевание легких (SSc-ILD)), облитерирующий бронхиолит с организующейся пневмонией, ILD, связанное с заболеванием соединительной ткани (CT-ILD), гиперчувствительный пневмонит, легочная гипертензия, туберкулез, силикоз, асбестоз легких, острое повреждение легких и острый респираторный дистресс (ARD, включая вызванный бактериальной пневмонией, вызванный повреждением и вызванный вирусной пневмонией, вызванный искусственной вентиляцией легких, вызванный нелегочным сепсисом).
Вариант осуществления 125. Способ по варианту осуществления 123, где фиброз представляет собой фиброзное состояние печени, выбранное из группы, состоящей из цирроза печени, врожденного фиброза печени, ожирения, жировой печени, алкогольного фиброза печени, неалкогольного стеатогепатита (NASH), повреждения желчных протоков, первичного билиарного цирроза (РВС), первичного склерозирующего холангита (PSC), инфекционного или вирусного фиброза печени (например, хронических инфекций, вызванных вирусом гепатита В и С), муковисцидоза, аутоиммунного гепатита, некротизирующего гепатита, первичного склерозирующего холангита, гемохроматоза, нарушений системы желчных протоков и печеночной дисфункции, обусловленной инфекциями.
Вариант осуществления 126. Способ по варианту осуществления 123, где фиброз представляет собой фиброзное состояние сердца и/или перикарда, выбранное из группы, состоящей из эндомиокардиального фиброза, васкулопатии сердечного аллотрансплантата (CAV), инфаркта миокарда, фиброза предсердий, застойной сердечной недостаточности, артериосклероза, атеросклероза, стеноза сосудов, миокардита, застойной кардиомиопатии, коронарных инфарктов, варикозного расширения вен, стеноза коронарных артерий и других постишемических состояний, и идиопатического ретроперитонеального фиброза.
Вариант осуществления 127. Способ по варианту осуществления 123, где фиброз представляет собой фиброзное состояние почек, выбранное из группы, состоящей из гломерулонефрита (включая мембранопролиферативные, диффузно-пролиферативные, быстро прогрессирующие или склерозирующие, постинфекционные и хронические формы), диабетического гломерулосклероза, фокально-сегментарного гломерулосклероза, IgA-нефропатии, диабетической нефропатии, ишемической нефропатии, тубулоинтерстициального фиброза почек, связанной с ВИЧ нефропатии, мембранной нефропатии, гломерулонефрита, вторичного по отношению к системным воспалительным заболеваниям, таким как волчанка, склеродермия и диабетический гломерулонефрит, идиопатического мембранопролиферативного гломерулонефрита, мезангиального пролиферативного гломерулонефрита, серповидного гломерулонефрита, амилоидоза (который среди других тканей поражает почки), аутоиммунного нефрита, почечного тубулоинстерстиального фиброза, почечного атеросклероза, синдрома Альпорта, нефротического синдрома, хронической почечной недостаточности, хронического заболевания почек, перигломерулярного фиброза/атубулярных клубочков, синдрома сочетанной апикальной эмфиземы и базального фиброза (синдрома эмфиземы/фиброза), гломерулярной гипертензии, нефрогенной фиброзирующей дерматопатии, поликистозной болезни почек, болезни Фабри и почечной гипертензии.
Вариант осуществления 128. Способ по варианту осуществления 123, где фиброз представляет собой фиброзное состояние поджелудочной железы, выбранное из группы, состоящей из стромального ремоделирующего панкреатита и стромального фиброза.
Вариант осуществления 129. Способ по варианту осуществления 123, где фиброз представляет собой фиброзное состояние желудочно-кишечного тракта, выбранное из группы, состоящей из болезни Крона, язвенного колита, коллагенозного колита, колоректального фиброза, атрофии ворсинок, гиперплазии крипт, образования полипов, заживления язвы желудка и микроскопического колита.
Вариант осуществления 130. Способ по варианту осуществления 123, где фиброз представляет собой фиброзное состояние глаз, выбранное из группы, состоящей из окулярного фиброза, фиброза глаз, пролиферативной витреоретинопатии, витреоретинопатии любой этиологии, фиброза, связанного с дисфункцией сетчатки, фиброза, связанного с влажной или сухой макулодистрофией, рубцевания роговицы и конъюнктивы, фиброза эндотелия роговицы, передней субкапсулярной катаракты и помутнения задней капсулы хрусталика, фиброзных заболеваний переднего сегмента глаза, фиброза стромы роговицы (например, связанного с помутнением роговицы), фиброза трабекулярной сети (например, связанного с глаукомой), фиброзных заболеваний заднего сегмента глаза, фиброваскулярного рубцевания (например, в сетчатке или хориоидальной сосудистой сети глаза), фиброза сетчатки, эпиретинального фиброза, глиоза сетчатки, субретинального фиброза (например, связанного с возрастной макулодистрофией), тракционной отслойки сетчатки в сочетании с сокращением ткани при диабетической ретинопатии, врожденного фиброза глазницы, фиброза слезных желез, субэпителиального фиброза роговицы и офтальмопатии Грейвса.
Вариант осуществления 131. Способ по варианту осуществления 123, где фиброз выбран из фиброза, возникающего в результате повреждения/фиброза спинного мозга или фиброза центральной нервной системы, таких как фиброз после инсульта, мышечная дистрофия Дюшенна, фиброз, связанный с нейродегенеративным нарушением, таким как болезнь Альцгеймера или рассеянный склероз, рестеноз сосудов, фиброз матки, эндометриоз, фиброиды яичников, болезнь Пейрони, синдром поликистозных яичников, апикальный фиброз легких, связанный с заболеванием анкилозирующим спондилитом, рубцевание и фиброз, возникающие при микробных инфекциях (например, бактериальных, вирусных, паразитарных, грибковых).
Вариант осуществления 132. Способ по варианту осуществления 123, где фиброз представляет собой SSc.
Вариант осуществления 133. Способ по варианту осуществления 123, где фиброз представляет собой IPF.
Вариант осуществления 134. Способ по варианту осуществления 123, где фиброз представляет собой хроническую обструктивную болезнь легких (COPD).
Вариант осуществления 135. Способ по варианту осуществления 123, где фиброз представляет собой прогрессирующее фиброзирующее интерстициальное заболевание легких (PF-ILD).
Вариант осуществления 136. Способ по варианту осуществления 135, где PF-ILD представляет собой заболевание или состояние, выбранное из группы, состоящей из неспецифической интерстициальной пневмонии (NSIP), криптогенной организующейся пневмонии (СОР), прогрессирующего массивного фиброза, осложнения пневмокониоза шахтеров, склеродермии/системной склеродермии, облитерирующего бронхиолита с организующейся пневмонией; ILD, связанного с заболеванием соединительной ткани (CT-ILD), и гиперчувствительного пневмонита.
Вариант осуществления 137. Способ по варианту осуществления 123, где фиброз представляет собой цирроз печени или хронический фиброз печени.
Вариант осуществления 138. Способ по варианту осуществления 123, где фиброз представляет собой фиброз желудочно-кишечного тракта, например, фиброз кишечника, необязательно выбранный из группы, состоящей из фиброза, связанного с болезнью Крона, язвенным колитом, коллагенозным колитом, колоректального фиброза, атрофии ворсинок, гиперплазии крипт, образования полипов, заживления язвы желудка и микроскопического колита.
Вариант осуществления 139. Способ по варианту осуществления 123, где фиброз представляет собой фиброзное состояние глаз, фиброз, возникающий в результате повреждения спинного мозга, фиброз или фиброз центральной нервной системы, или фиброз, связанный с нейродегенеративным нарушением.
Вариант осуществления 140. Способ по любому из варианту осуществления 118-139, где антитело представляет собой антитело к TGFβ2.
Вариант осуществления 141. Способ по варианту осуществления 140, где антитело к TGFβ2 содержит:
(a) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 16, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 17 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 18; и (б) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 19; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 20 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 21.
Вариант осуществления 142. Способ по любому из вариантов осуществления 118-139, где антитело представляет собой антитело к TGFβ3.
Вариант осуществления 143. Способ по варианту осуществления 142, где антитело к TGFβ3 содержит:
(а) (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 35 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9;
(б) пару VH/VL, при этом VH пары VH/VL содержит аминокислотную последовательность SEQ ID NO: 57 и VL пары VH/VL содержит аминокислотную последовательность SEQ ID NO: 56; или
(в) полную пару H/L-цепей, при этом Н-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 79 и L-цепь пары H/L-цепей содержит аминокислотную последовательность SEQ ID NO: 78.
Вариант осуществления 144. Способ по любому из вариантов осуществления 118-139, где антитело представляет собой антитело к TGFβ2/3.
Вариант осуществления 145. Способ по варианту осуществления 144, где антитело к TGFβ2/3 содержит: (i) CDR тяжелой цепи, содержащие CDR-H1, -Н2 и -Н3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 10, CDR-H2 имеет аминокислотную последовательность SEQ ID NO: 11 и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 12; и (ii) CDR легкой цепи, содержащие CDR-L1, -L2 и -L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 13; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 14 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 15.
Вариант осуществления 146. Способ диагностики SSc у субъекта, при этом способ включает определение уровней экспрессии генов в наборе сигнатур 18 генов, состоящей из PRSS23, PXDN, COL8A1, COL6A3, SERPINE2, TNC, СОМР, THBS1, COL11A1, COL1A1, COL5A2, COL1A2, COL4A1, COL4A2, SFRP4, ALPK2, COL5A1 и TAGLN, и установление диагноза SSc субъекту, если установлено, что уровни генов повышены по сравнению с уровнями генов в контрольном наборе от здорового субъекта или сигнатуре эталонных генов.
Вариант осуществления 147. Способ по варианту осуществления 146, где уровень гена повышается, если повышение экспрессии по сравнению с контрольным набором от здорового субъекта или сигнатурой эталонных генов является статистически значимым, необязательно по меньшей мере в два раза более высоким, или по меньшей мере в три раза более высоким, или по меньшей мере в четыре раза более высоким по сравнению с контрольным набором от здорового субъекта или сигнатурой эталонных генов.
Вариант осуществления 148. Способ мониторинга ответа на лечение субъекта антителом к TGFβ2 и/или антителом к TGFβ3, причем способ включает определение уровня экспрессии одного или более индуцируемых TGFβ генов, выбранных из группы, состоящей из serpinel, col1a1, col1a2 и col3a1 в образце от субъекта, где субъект получал одно или более введений антитела к TGFβ2 и/или антитела к TGFβ3.
Вариант осуществления 149. Способ по варианту осуществления 148, где у субъекта определяют наличие ответа на лечение антителом к TGFβ2 и/или антителом к TGFβ3, если уровень экспрессии одного или более индуцируемых TGFβ генов, значительно снижен по сравнению с уровнями одного или более индуцируемых TGFβ генов до лечения, при этом необязательно способ дополнительно включает осуществление дополнительного введения антитела к TGFβ2 и/или антитела к TGFβ3, если установлено, что уровень экспрессии одного или более индуцируемых TGFβ генов, значимо снижен.
Вариант осуществления 150. Способ по варианту осуществления 148 или варианту осуществления 149, где субъекту вводят антитело к TGFβ2 в виде монотерапии.
Вариант осуществления 151. Способ по варианту осуществления 148 или варианту осуществления 149, где субъекту вводят антитело к TGFβ3 в виде монотерапии.
Вариант осуществления 152. Способ по варианту осуществления 148, где уровень экспрессии одного или более генов, индуцируемых TGFβ, определяют с помощью кПЦР или микроматричного анализа.
Вариант осуществления 153. Способ или применение по любому из вариантов осуществления 100-152, где субъектом является человек.
Вариант осуществления 154. Набор, содержащий антитело по любому из вариантов осуществления 1-99 или вариантов осуществления I-1-I-53.
Вариант осуществления I-155. Способ лечения нарушения, связанного с TGFβ, у субъекта, нуждающегося в этом, при этом способ включает:
определение того, что образец от субъекта содержит повышенный уровень одного или более биомаркеров; и
введение субъекту эффективного количества селективного в отношении изоформы антитела к TGFβ.
Вариант осуществления I-156. Способ выбора средства терапии для субъекта с аутоиммунным или воспалительным заболеванием, включающий:
определение уровня одного или более биомаркеров субъекта; и
выбор лекарственного препарата на основе уровня одного или более биомаркеров, при этом выбранный лекарственный препарат представляет собой селективное в отношении изоформы антитело к TGFβ, если уровень одного или более биомаркеров повышен.
Вариант осуществления I-157. Способ лечения нарушения, связанного с TGFp, у субъекта, нуждающегося в этом, включающий введение субъекту эффективного количества селективного в отношении изоформы антитела к TGFβ, при этом до начала введения субъект имеет повышенный уровень одного или более биомаркеров.
Вариант осуществления I-158. Способ по любому из вариантов осуществления I-155-I-157, где один или более биомаркеров выбраны из группы, состоящей из TGFβ2, TGFβ3, периостина (POSTN) и олигомерного матриксного белка хряща (СОМР).
Вариант осуществления I-159. Способ по любому из вариантов осуществления I-155-I-157, где один или более биомаркеров выбраны из группы, состоящей из PRSS23, PXDN, COL8A1, COL6A3, SERPINE2, TNC, СОМР, THBS1, COL11A1, COL1A1, COL5A2, COL1A2, COL4A1, COL4A2, SFRP4, ALPK2, COL5A1 и TAGLN.
Вариант осуществления I-160. Способ по любому из вариантов осуществления I-155-I-157, где один или более биомаркеров выбраны из группы, состоящей из SERPINE1, COL1A1, COL1A2 и COL3A1.
Вариант осуществления I-161. Способ по любому из вариантов осуществления I-155-I-160, где уровень одного или более биомаркеров повышен в образце, взятом у субъекта, при этом образец представляет собой образец кожи, легких или крови.
Вариант осуществления I-162. Способ по варианту осуществления I-158, где образец представляет собой образец сыворотки крови.
Вариант осуществления I-160. Способ по любому из вариантов осуществления I-155-I-159, где селективное в отношении изоформы антитело к TGFβ представляет собой антитело к TGFβ2, антитело к TGFβ2/3 или антитело к TGFβ3.
Вариант осуществления I-161. Способ по любому из вариантов осуществления I-155-I-160, где повышенные биомаркеры представляют собой один или оба из периостина и СОМР.
Вариант осуществления I-162. Способ по любому из вариантов осуществления I-155-I-161, где нарушение представляет собой фиброзное нарушение.
Вариант осуществления I-163. Способ по варианту осуществления I-162, где фиброзное нарушение представляет собой фиброз легких, фиброзное заболевание печени, фиброзное состояние сердца и/или перикарда, фиброзное состояние почек, фиброзное состояние поджелудочной железы, фиброзное заболевание состояние желудочно-кишечного тракта, фиброзное состояние глаз или фиброз в результате повреждения/фиброза спинного мозга или фиброз центральной нервной системы.
Вариант осуществления I-164. Способ по варианту осуществления I-162, где фиброзное заболевание выбрано из группы, состоящей из IPF, SSc, COPD и PF-ILD.
Вариант осуществления I-165. Способ в соответствии с любым из вариантов осуществления I-155-I-164, где повышенный уровень относится к здоровому индивидууму или контрольной группе, и при этом повышение является статистически значимым, необязательно по меньшей мере двукратным повышением или, по меньшей мере трехкратным повышением, или по меньшей мере четырехкратным повышением по сравнению со здоровым индивидуумом или контрольной группой.
Вариант осуществления I-166. Способ по любому из вариантов осуществления I-155-I-164, где субъект не отвечал на терапию ингибитором IL-6.
Вариант осуществления I-167. Способ по варианту осуществления I-166, где средство терапии на основе антитела к IL-6 представляла собой тоцилизумаб.
Вариант осуществления I-168. Способ по любому из вариантов осуществления I-155-I-167, где селективное в отношении изоформы антитело к TGFβ представляет собой антитело в соответствии с любым из вариантов осуществления 1-99 или в соответствии с вариантами осуществления I-1-I-53.
IV. ПРИМЕРЫ
Ниже приведены примеры способов и композиций по настоящему изобретению. Следует понимать, что на практике можно реализовать ряд других вариантов осуществления с учетом общего описания, приведенного выше.
Пример 1: Получение и скрининг селективных в отношении изоформы антител к TGFβ
Материалы и методы:
Получение моноклональных кроличьих антител к TGFβ человека
Новозеландских белых кроликов иммунизировали человеческими изоформами TGFβ2 и TGFβ3 (Peprotech) и выделяли одну В-клетку с использованием модифицированного протокола, относящегося к опубликованной литературе {Offner et al. PLoS ONE 9(2), 2014). Вкратце, мононуклеарные клетки периферической крови кролика (РВМС) выделяли путем центрифугирования плотности крови (разведение 1:1 с PBS), взятой из ушной артерии, с использованием Lympholyte®-M (Cedarlane Labs). После промывания PBS РВМС ресуспендировали в культуральной среде RPMI с добавкой и переносили в 6-луночный планшет для удаления макрофагов и моноцитов за счет неспецифической адгезии к планшету. Затем неадгезивные клетки собирали для обогащения В-клетками с помощью иммунофлуоресцентного окрашивания биотинилированным антителом к CD11b кролика, антителом к Т-лимфоцитам кролика (AbD Serotec, BioRad) и антителом к IgM кролика (BD Bioscience) для дальнейшей деплеции кроличьих миелоидных клеток, Т-клеток и IgM В-клеток через колонку MACS (Miltenyl Biotec) в соответствии с инструкциями производителя. Следующий рабочий процесс включал использование FITC-меченого козьего антитела к IgG кролика (Serotec, BioRad), АРС-меченого TGFβ1 человека (Innova Biosciences) и RPE-меченого TGFβ2/3 человека (Innova Biosciences) для сортировки кроличьих IgG+и человеческих TGFβ2/3+-реактивных В-клеток в отдельные лунки. Супернатанты В-клеточных культур подвергали первичному скринингу методом ELISA в отношении связывания изоформ TGFβ2/3 человека, В-клетки лизировали и хранили при -80°С. При молекулярном клонировании вариабельные области легкой и тяжелой цепей В-клеток, специфичных к TGFβ2/3 человека, амплифицировали с помощью ПЦР и клонировали в векторы экспрессии, как описано в опубликованной литературе {Offner et al. PLoS ONE 9(2), 2014). Каждое рекомбинантное кроличье моноклональное антитело экспрессировали в клетках Expi293 (Thermo Fisher Scientific, Уолтем, Массачусетс) и затем очищали с помощью белка А. Очищенные антитела к TGFβ2/3 человека затем подвергали функциональной характеристике по блокированию и определению аффинности.
Разработка крысиных гибридомных антител к TGFβ человека
Крыс Sprague Dawley (Charles River, Холлистер, Калифорния) иммунизировали каждые две недели 100 мкг белков TGFβ2 или TGFβ3 человека (Peprotech), смешанных с адъювантом RIBI (Sigmal, Сент-Луис, Миссури), и вводили в основание хвоста или смешивали с неполным адъювантом Фрейнда (BD) и разделяли между несколькими участками: LP., S.C. у основания хвоста, s.c. на затылке и s.c. в обоих пятках. Несколько лимфатических узлов собирали через три дня после последней иммунизации. IgM-отрицательные В-клетки этих крыс очищали от лимфоцитов с помощью магнитного разделения (Miltenyi Biotec, Сан-Диего, Калифорния) и сливали с клетками миеломы мыши Sp2ab (Enzo Life Sciences, Фармингдейл, Нью-Йорк) посредством электрослияния (Harvard Apparatus, Холлистон, Массачусетс). Слитые клетки инкубировали при 37°С, 7% СО2 в течение ночи в среде С (StemCell Technologies, Ванкувер, Британская Колумбия, Канада) перед посевом в 6-луночные планшеты, содержащие среду Е (StemCell Technologies, Ванкувер, Британская Колумбия, Канада) с добавлением HAT (Sigma- Aldrich, Сент-Луис, Миссури) и инкубировали при 37°С, 7% СО2 в течение трех дней. Гибридомы TGFβ2 собирали и сортировали с помощью антитела к IgG крысы (Sigma-Aldrich), а гибридомы TGFβ3 сортировали с помощью антитела к IgG крысы (Sigma-Aldrich) и метили TGFβ3-PE человека с помощью набора для мечения антител Lightning-Link R-PE (Novus, Сентенниал, Колорадо).
Анализ блокирования TGFβ на основе клеток с применением HEKBlue
Клетки HEK-Blue™ TGFβ (InvivoGen, Сан-Диего, Калифорния), стабильно экспрессирующие белки TGFβRI, Smad3, Smad4 человека и репортерный ген секретируемой щелочной фосфатазы (SEAP) под контролем минимального промотора β-глобина, слитого с тремя Smad3/4-связывающими элементами (SBE) использовали для оценки блокирующей способности моноклональных антител, полученных от крыс и кроликов, иммунизированных зрелыми рецептор-связывающими доменами TGFβ1, TGFβ2 и TGFβ3 человека. Антитела инкубировали с каждой изоформой TGFβ (PeproTech, Роки-Хилл, Нью-Джерси) при 25°С в течение 1 часа перед добавлением к 4,5×104 клеток HEK-Blue™ TGF-β, высеянных в 96-луночный планшет в 100 мкл буфера для анализа (DMEM с высоким содержанием глюкозы с 10% инактивированной нагреванием FBS, 2 мМ L-глутамина и 0,5% пенициллин-стрептомицина). Конечная концентрация антитела и TGFβ составляла 0,001-200 нМ и 20 пМ соответственно. Планшет инкубировали при 37°С и 5% СО2 в течение 18-22 часов. Уровень SEAP в супернатанте клеток определяли с помощью анализа QUANTI-Blue™ в соответствии с инструкциями производителя (InvivoGen). mAb ко всем видам TGFβ нейтрализует TGFβ1, 2 и 3 при низких клеточных потенциях (IC50), составляющих 0,72, 2,2 и 0,026 нМ соответственно.
Скрининг ELISA
Для скрининга связывания клонов-кандидатов с TGFβ1, TGFβ2 и TGFβ3 использовали твердофазный иммуноферментный анализ (ELISA). Сначала каждую изоформу TGFβ (Peprotech) иммобилизовали в лунки 384-луночных планшетов для ELISA Grenier Bio-One с буфером для покрытия (50 мМ карбоната, рН 9,6) в концентрации 1 мкг/мл; 50 мкл/лунка для инкубации в течение ночи при 4°С.Затем покрытые планшеты для ELISA промывали 3 раза буфером для разведения ELISA по 100 мкл на лунку (PBS/0,5% Tween20). Затем планшеты для ELISA блокировали буфером для разведения ELISA 100 мкл/лунка в течение одного часа. После блокирования буфер удаляли, а образцы и контроля добавляли из 96-луночных планшетов в 384-луночные планшеты (50 мкл/лунка). Затем планшеты для ELISA с образцом инкубировали в течение 30 минут при комнатной температуре при встряхивании планшетов на шейкере. Затем планшеты трижды промывали промывочным буфером (PBS/0,05% TWEEN 20, 20Х Stock Hyclone SH3A649-01) 100 мкл/нед. Затем разведенный конъюгат HRP (конъюгированное пероксидазой AffiniPure козье антитело к крысиному IgG (H+L), Jackson ImmunoResearch Laboratories, INC. 112-035-003, разбавленное 1:10000, 50 мкл/лунка) добавляли в каждую лунку и планшеты для ELISA инкубировали в течение 30 минут при комнатной температуре. После инкубации планшеты промывали 3 раза промывочным буфером (100 мкл/лунка), а затем добавляли субстрат (BIOFX, субстрат ТМВ, TMBW-1000-01, 50 мкл/лунка). После 5-минутной инкубации при комнатной температуре в течение 5 минут добавляли стоп-раствор (стоп-реагент BIOFX, LBSP-1000-01, 50 мкл/лунка). Наконец, планшеты считывали при 659 нм в отношения поглощения.
Результаты:
Моноклональные антитела, полученные от крыс и кроликов, иммунизированных зрелыми рецептор-связывающими доменами TGFβ1, TGFβ2 и TGFβ3 человека, подвергали скринингу в отношении связывания с помощью прямого ELISA и поверхностного плазмонного резонанса (SPR), а также в отношении их способности ингибировать TGFβR-зависимую передачу сигналов, индуцированную зрелым рецептор-связывающим доменом в репортерной клеточной линии (репортер HEKBlue SMAD2/3 SEAP), как описано выше. Из 96 исходных кроличьих клонов, которые демонстрировали связывание с TGFβ2, и 85 клонов, которые связывались с TGFβ3, 48 клонов отбирали для дальнейшего скрининга. Касательно касается кроличьих антител, 1995 гибридом IgG+TGFβ2 и 2660 гибридом IgG+TGFβ3+ сортировали по одной клетке с использованием сортера FACSAria Fusion (BD Biosciences, Франклин-Лейке, Нью-Джерси) в 96-луночные планшеты. Через семь дней после посева супернатанты подвергали скринингу с помощью ELISA в отношении белков TGFβ2 или TGFβ3 человека. Собирали супернатанты от 28 гибридом TGFβ2 и 48 TGFβ3, демонстрирующих связывание с их соответствующими антигенами с помощью ELISA, и очищали с помощью белка G (GammaBind Plus, GE Healthcare, Питтсбург, Пенсильвания). Среди 28 клонов TGFβ2+ 4 показали функциональную блокировку против TGFβ2, включая 6F12. Среди 48 клонов TGFβ3+ 28 клонов показали функциональную блокировку против TGFβ3 человека, включая 2А10.
Высокоэффективные ингибирующие антитела (KD<10 пМ и клеточная IC50<250 пМ) идентифицировали с селективностью в отношении TGFβ2 (mAb 6F12 (крысы)), TGFβ3 (mAb 16С10 (кролика) и 2А10 (крысы)) и антитела с двойной специфичностью, которое связывалось и ингибировало активность как TGFβ2, так и TGFβ3 (mAb 4А11 (кролика)), но ни одно из этих антител не связывалось с TGFβ1 или не ингибировало его (см. таблицу 4 ниже).
Относительная активность этих антител в отношении выбранных изоформ TGFβ была сопоставима или превосходила активность антитела «ко всем видам TGFβ», 1D11 (Dasch, JR et al. J. Immunol. 142: 1536-41 (1989)) (см. таблицу 4, ниже). Примечательно, что TGFβ2-специфическое антитело 6F12 было почти в 100 раз более эффективным, чем 1D11, в блокировании TGFβ2-зависимой репортерной активности в клетках (IC50 40 пМ по сравнению с 3,6 нМ).
Кроме того, между зрелым TGFβ2 человека и мыши имеются три различия в аминокислотах, a 6F12 и 4А11 связываются с обоими пептидами с одинаковой аффинностью (Фиг. 55А) и обладают одинаковой ингибирующей активностью в отношении обоих (Фиг. 55).
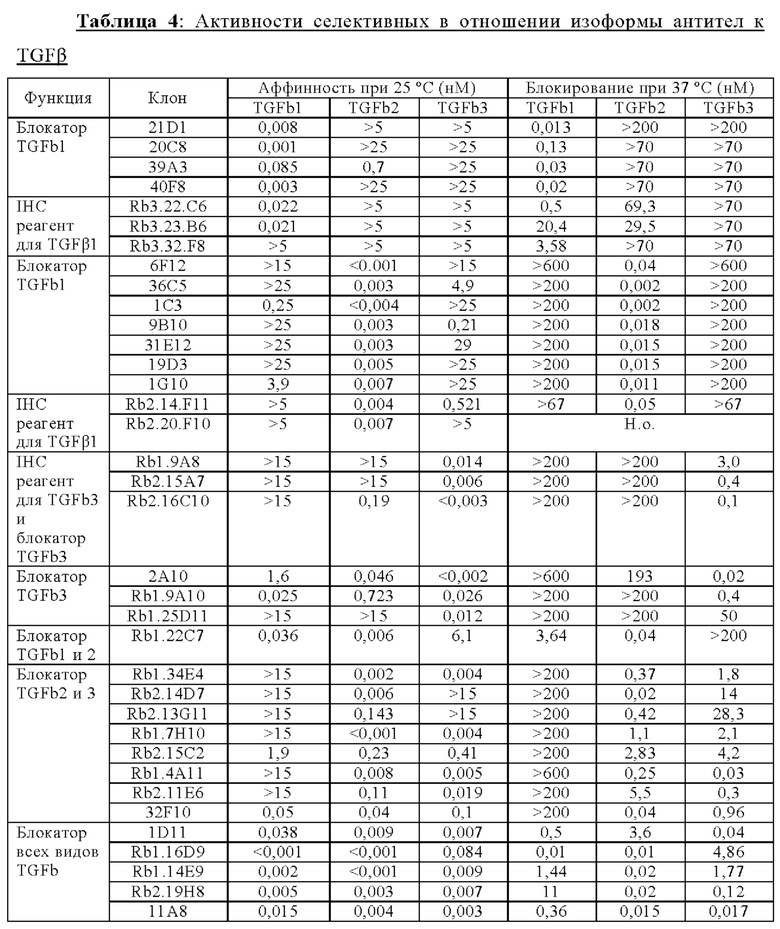
Последовательности моноклональных антител представлены в таблицах 5-7 ниже.


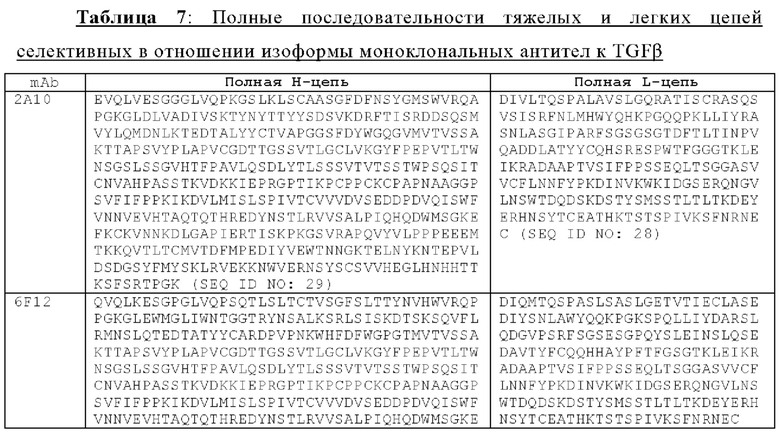

Пример 2: Различные профили токсичности для ингибирования TGFβ2 и TGFβ3 селективными в отношении изоформы моноклональными антителами к TGFβ
Конститутивные нокауты TGFβ2 и TGFβ3 зародышевой линии не выживают в перинатальный период вследствие дефектов развития, включая расщелину неба у обоих, и сердечно-сосудистые дефекты у нокаутов TGFβ2, а ингибиторы всех видов TGFβ имеют значительную токсичность, как подробно описано выше. Для оценки потенциальной токсичности ингибирования TGFβ2, TGFβ3 или обоих проводили 4-недельные токсикологические исследования с использованием 6F12, 2А10 и 4А11 следующим образом:
TGFβ3 (2А10):
В пилотном токсикологическом исследовании mAb 2А10 вводили три раза в неделю (TIW) в течение 4 недель (всего 12 доз) мышам CD-1 в дозах 0 (контроль), 10 и 50 мг/кг I.P. TIW и 50 мг/кг I.V. TIW в несущей среде, состоящей из фосфатно-солевого буфера. Самцов и самок (n=5/пол/группа) относили к группам токсичности при всех уровнях доз и планировали их вскрытие в конце периода введения дозы (день 29). Кровь для токсикокинетической оценки собирали у дополнительных мышей в контрольной группе (n=6 самцов/группа) и в группах, обработанных mAb 2А10 (n=9 самцов/группа). Критерии оценки включали следующие параметры: клинические наблюдения, вес тела, клиническую патологию (гематологию и клиническую биохимию), анатомическую патологию и токсикокинетику.
Воздействие 2А10 на AUC возрастало примерно пропорционально дозе с повышением дозы от 10 до 50 мг/кг.
Все животные дожили до запланированного вскрытия, и какие-либо клинические наблюдения, связанные с 2А10, влияния на вес тела или изменения клинической патологии отсутствовали. Результаты, связанные с 2А10, были ограничены метафизарной дисплазией, наблюдаемой у подгруппы мышей при введении высокой дозы (6 мышей при введении 50 мг/кг). В заключение, введение mAb 2А10 мышам CD-1 в дозах 10 и 50 мг/кг TIW в течение 4 недель хорошо переносилось, а результат, связанный с 2А10, заключался в ограниченной метафизарной дисплазии при введении высокой дозы.
TGFβ2 (6F12):
В пилотном токсикологическом исследовании mAb 6F12 вводили три раза в неделю (TIW) в течение 4 недель (всего 12 доз) мышам CD-1 в дозах 0 (контроль), 10 и 50 мг/кг LP. в несущей среде, состоящей из фосфатно-солевого буфера. Самцов и самок (n=5/пол/группа) относили к группам токсичности при всех уровнях доз и планировали их вскрытие в конце периода введения дозы (день 29). Кровь для токсикокинетической оценки собирали у дополнительных мышей в группах, обработанных mAb 6F12 (n=9 самцов/группа). Критерии оценки включали следующие параметры: клинические наблюдения, вес тела, клиническую патологию (гематологию и клиническую биохимию), анатомическую патологию и токсикокинетику.
Воздействие 6F12 на AUC возрастало примерно пропорционально дозе с повышением дозы от 10 до 50 мг/кг.
Все животные выживали до запланированного вскрытия при введении дозы 10 мг/кг, и какие-либо клинические наблюдения, связанные с 6F12, при введении доз 10 и 50 мг/кг отсутствовали.
Результаты, связанные с 6F12, при введении дозы 10 и 50 мг/кг, включали минимальное или умеренное увеличение нейтрофилов и лимфоцитов у самок с гистопатологической корреляцией воспаления брюшины, указывающей на хроническое воспаление, и повышение ALT и AST без корреляции с гистопатологией. Дополнительные результаты, связанные с 6F12 при введении дозы 50 мг/кг, включали раннюю эвтаназию 3 мышей в токсикокинетической группе вследствие снижения веса тела на более чем 20% (наблюдаемое в день 8), а также от минимального до умеренного повышения уровня глобулина с соответствующим снижением отношение альбумина к глобулину у обоих полов.
Все мыши в группах изучения токсичности дожили до запланированного вскрытия в день 29, и никаких изменений веса тела не наблюдалось.
В заключение, при введении мышам CD-1 mAb 6F12 в дозах 10 и 50 мг/кг TIW в течение 4 недель результаты, согласующиеся с хроническим воспалением, наблюдались при введении доз 10 и 50 мг/кг.
TGFβ2/3 (4All):
В пилотном токсикологическом исследовании mAb 4А11 вводили три раза в неделю (TIW) в течение 4 недель (всего 12 доз) мышам CD-1 в дозах 0 (контроль), 10 и 50 мг/кг LP., а также 50 мг/кг I.V. в несущей среде, состоящей из фосфатно-солевого буфера. Самцов и самок (n=5/пол/группа) относили к группам токсичности при всех уровнях доз и планировали их вскрытие в конце периода введения дозы (день 29). Кровь для токсикокинетической оценки собирали у дополнительных мышей в контрольной группе (n=6 самцов/группа) и в группах, обработанных mAb 4А11 (n=9 самцов/группа). Критерии оценки включали следующие параметры: клинические наблюдения, вес тела, клиническую патологию (гематологию и клиническую биохимию), анатомическую патологию и токсикокинетику.
Основные токсикологические данные при введении доз 10 и 50 мг/кг включали смертность у 3 мышей (1 при дозе 10 мг/кг и 2 при дозе 50 мг/кг) и паховые/мошоночные грыжи. Дополнительные основные результаты при введении дозы 50 мг/кг включали клиническое наблюдение потери функции задних конечностей, патологические признаки периректального серозного кровоизлияния и гемоабдомена, периартериит/артериит и пристеночные и интерстициальные кровоизлияния в сердечно-сосудистой системе (восходящей аорте и миокарде), а также дегенерацию и некроз скелетной мышцы.
Таким образом, введение mAb 4А11 мышам CD-1 в дозах 10 и 50 мг/кг TIW в течение 4 недель приводило к смертности в дозах 10 и 50 мг/кг и неблагоприятным патологическим проявлениям грыж при всех дозах и в сердечно-сосудистой системе в дозе 50 мг/кг.
Результаты исследования токсичности обобщены в таблице 8 ниже.
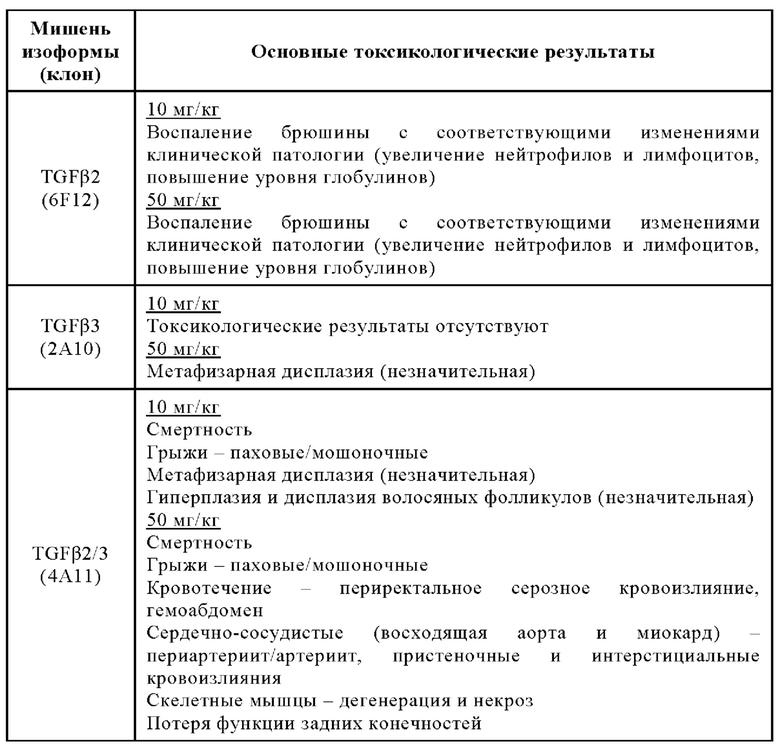
Пример 3: ФК исследование мышей с использованием моноклональных антител к TGFβ
Материалы и методы:
Fab-области антител к TGFβ сливали с Fc-областью мышиного IgG2a с мутацией LALA-PG для ослабления эффекторной функции в исследованиях на мышах in vivo (см. Lo, М. et al. Journal of Biological Chemistry 292, 3900-3908) (2017); Schlothauer, T. et al. Protein Engineering Design and Selection 29, 457-466 (2016)).
Результаты:
Fab-области антител к TGFβ сливали с Fc-областью мышиного IgG2a с мутацией LALA-PG для ослабления эффекторной функции для исследований у мышей in vivo. В исследованиях фармакокинетики однократной дозы у мышей С57/В6 эти химерные антитела демонстрировали линейную фармакокинетику с дозозависимым накоплением и отсутствием признаков образования антител к лекарственным средствам. Все антитела с наиболее высокой эффективностью in vitro (6F12 (антитело к TGFβ2); 2А10 (антитело к TGFβ3) и 4А11 (антитело к TGFβ2/3 с двойной специфичностью) показали биодоступность in vivo -95% и клиренс приблизительно 4 мл/день/кг, в то время как 16С10, несколько менее активное антитело к TGFβ3, демонстрировало клиренс приблизительно 2 мл/день/кг (Фиг. 2).
Пример 4: Уровни экспрессии TGFβ2 и TGFβ3 повышены в фиброзной ткани человека, а экспрессия TGFβ3 высоко коррелирует с передачей сигналов TGFβR при IPF и SSc
Материалы и методы:
Выделение тканей (IPF) и анализ РНК
Анализы РНК тканей человека описывали ранее (Depianto, DJ, et. al. Thorax. 2015 Jan;70(l):48-56; Chandriani, S, et. al. J Immunol. 2014 Jul 1;193(1):111-9). Вкратце, ткани человека получали в Калифорнийском университете в Центре легких Сан-Франциско от пациентов с IPF во время биопсии или трансплантации легких. Здоровые нормальные ткани легких получали из легких, не используемых Сетью доноров трансплантации Северной Калифорнии. РНК выделяли из быстрозамороженных образцов легких. Микроматричный анализ выполняли в соответствии с рекомендациями производителя (Agilent, Санта-Клара, Калифорния).
Получение набора генов TGFβ, релевантных для кожи при SSc, и оценка сигнатур:
TGFβ-чувствительные гены идентифицировали путем секвенирования РНК на первичных фибробластах легких человека, стимулированных 5 нг/мл TGFβ1 человека (RnD systems) или контрольной средой в течение 24 часов. Данные секвенирования обрабатывали с использованием языка программирования R и пакетов из проекта Bioconductor (см. всемирную сеть на сайте bioconductor.org), чтобы выразить их на генном уровне в виде нормализованных считываемых значений на килобазу на миллион считываемых значений (nRPKM). Пилотный набор данных микроматричного анализа биоптатов при SSc и нормальной контрольной кожи (GSE58095) загружали из NCBI Gene Expression Omnibus. Для идентификации набора генов, которые активируются в коже при Ssc и потенциально могут быть связаны с активностью TGFβ в фибробластах, проводили 4-стороннее сравнение, в котором сравнивали гены со значительной дисрегуляцией in vitro (≥4-кратное повышение TGFβ) и при Ssc по сравнению с НС (≥50% увеличение заболеваемости). Этот набор кандидатных генов изображен на Фиг. 5Б. Набор генов оценивали в наборе данных из микроматичного анализы биоптатов кожи FaSScinate (см. Khanna et al. (2016) The Lancet; vol. 387:10038, p. 2630-2640 для описания исследования FaSSCinate) и сокращали до набора основных сигнатур 18 генов, как показано в таблице 2 выше, так что все гены вносят однородный вклад в первый основной компонент (>0,7 для нагрузки РС1). Для этого набора генов определяли составную оценку экспрессии генов с использованием загрузки первого основного компонента, чтобы уменьшить совокупную экспрессию сигнатуры 18 генов до одной непрерывной переменной для каждого субъекта.
ScRNAseq
Подготовка ткани легких человека:
Эксплантированные ткани легких получали от пациентов с патологическим диагнозом обычной интерстициальной пневмонии и согласованным клиническим диагнозом IPF, установленным в результате мультидисциплинарного обсуждения и обзора клинических материалов. После бронхоальвеолярного лаважа свежую ткань эксплантата легких хранили в полной среде на влажном льду в течение ночи. Ткань промывали в HBSS, а затем тщательно измельчали в буфере для расщепления (HBSS, 2,5 мг/мл коллагеназы D, 100 мкг/мл ДНКазы). Измельченную ткань встряхивали 45 минут при 37°С. Остаточный тканевый материал переносили в свежий буфер для расщепления и встряхивали еще 45 минут при 37°С. Отдельные клетки из обоих раундов расщепления объединяли и использовали для последующих анализов. Оставшуюся ткань обрабатывали с помощью диссоциатора gentleMACS (Miltenyi Biotec) для высвобождения дополнительных клеток в суспензию отдельных клеток. Этот препарат затем фильтровали и добавляли к ранее выделенным клеткам.
Выделение клеточных популяций с помощью FACS:
Общее количество клеток определяли и ресуспендировали в соответствующем объеме 1×PBS с добавлением 2 мМ EDTA и 2% фетальной бычьей сыворотки (FBS). Блокирование FcR осуществляли с использованием блокирующего реагента Miltenyi Biotec FcR в соответствии с протоколом производителя. Затем клетки окрашивали смесью антител, включая CD45-BUV395, ЕРСАМ-РЕ, CD31-BV605 и Live/Dead-Efluor 780, на льду в течение 30 минут, а затем 3 раза промывали добавленным буфером PBS. Собирали 5 популяций клеток для нисходящего РНК-секвенирования отдельных клеток: Несортированные живые клетки, CD45+, CD45-/EPCAM+, CD45-/CD31+ и CD45-/CD31-/EPCAM- (трижды негативные).
РНК-секвенирование отдельных клеток:
РНК-секвенирование отдельных клеток выполняли на платформе 10х Genomics с использованием библиотеки Chromium Single Cell 3' и набора гелевых гранул v2 в соответствии с руководством пользователя производителя (10х Genomics). Плотность клеток использовали для определения объема суспензии отдельных клеток, необходимого для мастер-микса для обратной транскрипции (ОТ), с целью достижения ~ 6000 клеток на образец. кДНК и библиотеки готовили в соответствии с руководством пользователя производителя (10х Genomics). Библиотеки профилировали с помощью набора ДНК высокой чувствительности Bioanalyzer (Agilent Technologies) и количественно определяли с использованием набора для количественного анализа библиотек Kapa (Kapa Biosystems). Каждую библиотеку секвенировали на одной дорожке HiSeq4000 (Illumina) в соответствии со спецификацией производителя (10х Genomics).
Обработка данных секвенирования:
Cell Ranger v2.1.0 (10Х Genomics, Плезантон, Калифорния) использовали для сопоставления необработанных считываемых значений секвенирования с эталоном человека GRCh38, подсчета признаков генов, фильтрации пустых капель, кластеризации клеток и идентификации кластерных маркерных генов. Визуализацию результатов tSNE из Cell Ranger выполняли с использованием браузера Loupe Cell Browser v3.1 (10X Genomics, Плезантон, Калифорния).
Иммуногистохимический анализ pSMAD3 и гибридизация in situ TGFβ3
Иммуногистохимический анализ фосфо-SMAD3 проводили на фиксированных формалином и залитых парафином срезах с использованием кроличьего моноклонального антитела (EP823Y, 0,76 мкг/мл; Abcam). Вкратце, срезы депарифинизировали и использовали для иммуногистохимического анализа на автоматизированной платформе для окрашивания (Bond RX; Leica Biosystems) с использованием запатентованных реагентов для поиска антигена (ER2; Leica Biosystems), обнаружения связанных первичных антител (конъюгированное с HR-полимером антитело к кроличьему антителу, PowerVision; Leica Biosystems). Сигнал визуализировали с помощью DAB.
Количественное определение сывороточного периостина и уровней СОМР
Цельную кровь собирали у пациентов и у 25 здоровых контрольных лиц соответствующего возраста и пола с помощью стандартной методики венепункции в пробирки для отделения сыворотки крови, давали ей свернуться в течение 30 минут и центрифугировали при 4°С в течение 10 минут. Сыворотку собирали и хранили при -70°С до анализа. Уровни хрящевого олигомерного матриксного белка (СОМР) определяли с использованием специфических иммуноанализов на основе IMPACT (Roche Diagnostics, Пенцберг, Германия). Вкратце, образцы инкубировали с чипом IMPACT, покрытым биотинилированными фрагментами антител к F(ab')2 СОМР. После этапов промывки добавляли и инкубировали специфичные для биомаркеров антитела для обнаружения, меченые дигоксигенином. После промывки добавляли флуоресцентный конъюгат антидигоксигенин-латекс, и специфически связанную флуоресцентную метку обнаруживали с помощью детекторной камеры с зарядовой связью. Испускаемую флуоресценцию преобразовывали в интенсивность сигнала с использованием стандартной кривой, состоящей из стандартов рекомбинантных белков (R&D Systems, Миннеаполис, Миннесота, США) и специального программного обеспечения (Roche Diagnostics).
Чувствительность для СОМР составляла 0 25 нг/мл. Уровни периостина в сыворотке крови определяли с использованием клинической версии иммуноанализа периостина Elecsys (Roche Diagnostics), предназначенного для использования на Cobas е601 (Roche Diagnostics).
Результаты:
При кПЦР-анализе объемных образцов биопсии легких пациентов с идиопатическим легочным фиброзом (IPF) TGFβ3 и, в меньшей степени, TGFβ2 экспрессировались при значительно более высоких уровнях по сравнению с контрольными легкими, тогда как экспрессия TGFβ1 была в среднем немного ниже при IPF по сравнению с контролем (Фиг. 3А). Иммуногистохимический анализ выявил значительное увеличение ядерного окрашивания фосфо-SMAD2/3 в ткани при IPF по сравнению с контролем, особенно в очагах фибробластов, областях активированных фибробластов и миофибробластов, что является патогномоничным для гистологического диагноза обычной интерстициальной пневмонии (UIP), отличительной черты IPF (Фиг. 3Б). Поскольку различия в экспрессии генов в объемной ткани легких при IPF могут отражать цитологический состав образца, затем предпринимали попытку охарактеризовать клеточное происхождение изоформ TGFβ на основе одной клетки с использованием секвенирования РНК отдельных клеток (scRNAseq). Глобальные паттерны экспрессии генов при scRNAseq, полученные из легких при IPF, различают несколько клеточных линий, включая гемопоэтические (макрофаги, лимфоциты, плазмобласты, тучные клетки), эпителиальные (АЕС2 и базальные, реснитчатые и секреторные бронхиальные эпителиальные клетки), мезенхимальные (фибробласты, миофибробласты, гладкомышечные) и сосудистые (эндотелиальные) типы клеток.
TGFβ1 значительно экспрессировался при наиболее высоких уровнях в гемопоэтических клетках, эндотелиальных клетках и миофибробластах. TGFβ2 был более ограниченным и в основном обнаруживался в эпителиальных клетках. Экспрессия TGFβ3 была строго ограничена мезенхимальными клетками (фибробластами, миофибробластами и гладкомышечными клетками). Экспрессия TGFβR-зависимых генов-мишеней выявила различные закономерности: SERPINE1 экспрессируется в фибробластах, макрофагах и эндотелиальных клетках, периостин (POSTN) в миофибробластах и эндотелиальных клетках, a COL1A1 и СОМР в миофибробластах, в значительной степени перекрываясь с TGFβ3 (Фиг. 4А). Двойной иммуногистохимический анализ для pSMAD3 и гибридизация in situ для TGFβ3 выявила многочисленные миофибробласты в фибробластных очагах, коэкспрессирующих TGFβ3 и ядерный pSMAD3 (Фиг. 4Б), что позволяет предположить, что при поражениях UIP TGFβ3 может управлять TGFβR-зависимой передачей сигналов аутокринным или паракринным образом.
В биоптатах кожи, взятых на исходном уровне в исследовании FaSScinate, экспрессия TGFβ3 заметно повышается у пациентов с SSc по сравнению со здоровым контролем, тогда как уровни TGFβ1 и TGFβ2 сопоставимы (Фиг. 5А). Для оценки потенциального вклада передачи сигналов TGFβ в дифференциально экспрессируемые гены в коже при SSc, получали набор генов, который связан как с фибробластами, отвечающими на передачу сигналов TGFβ, так и с нарушением регуляции в коже при SSc. Учитывая поразительную гетерогенность в профилях экспрессии генов, которые, как известно, существуют в биоптатах кожи пациентов с SSc, было высказано предположение, что в любой момент времени только поднабор когорты с SSc будет демонстрировать высокий профиль активности TGFβ.
Используя сигнатуру 18 генов TGFβ, проводили кластеризацию пациентов с SSc методом k-средних, и идентифицировали два кластера - кластер «TGFβ-высокий», который демонстрирует более высокие уровни экспрессии для многих членов набора генов кластера «TGFβ-низкий», в котором большинство членов набора генов зарегистрировали более низкое значение экспрессии.
Экспрессия TGFβ3 значительно и положительно коррелировала с TGFβ-индуцируемой сигнатурой (коэффициент корреляции ро Спирмена=0,92, р<0,0001), в то время как две другие изоформы TGFβ не коррелировали с оценкой (Фиг. 6А и Фиг. 6Б).
Сигнатура TGFβ-индуцируемого гена кожи также высоко коррелировала с уровнями экспрессии двух генов, кодирующих растворимые секретируемые компоненты ЕСМ, POSTN и СОМР. POSTN и СОМР, белки, кодируемые этими генами, обнаруживаются в сыворотке периферической крови в качестве растворимых биомаркеров, и их уровни удивительно хорошо коррелируют с сигнатурой генов TGFβ биоптата кожи в одновременно взятых образцах крови (Фиг. 6Б). Кроме того, уровни сывороточного периостина и СОМР в значительной степени коррелировали с модифицированной шкалой оценки кожи Роднана (MRSS), клиническим показателем системного фиброза кожи, используемым в исследованиях SSc (Фиг. 6В). Для оценки взаимосвязи между активностью TGFβ кожи и прогрессированием заболевания, оценивали изменение MRSS в течение 48-недельного курса исследования, и обнаружили, что у пациентов с «высокой» сигнатурой TGFβ было меньше шансов на улучшение по сравнению с пациентами с «низкой» сигнатурой TGFβ независимо от назначенного лечения (в исследовании не было выявлено значительного преимущества тоцилизумаба по сравнению с плацебо по показателю результатов MRSS; Фиг. 7). В совокупности эти данные позволяют предположить, что экспрессия TGFβ3 и сигнальная активность в выделенном биоптате кожи отражают общий груз заболевания и прогноз при SSc. Таким образом, эти данные подтверждают ингибирование TGFβ3 в качестве мишени для лечения IPF и SSc, а также использование уровней периостина в сыворотке и белка СОМР или уровней экспрессии генов сигнатуры TGFβ в тканях в качестве биомаркеров активности TGFβ3.
Пример 5: Активность селективных в отношении изоформы антител к TGFβ2 и TGFβ3 in vivo в модели фиброза легких
Материалы и методы:
Инстилляции блеомицина
Для исследования с использованием блеомицина взрослых мышей (>12 недель) рандомизировали в зависимости от их веса перед исследованием для сведения к минимуму различия между экспериментальной и контрольной группами. Для интратрахеального (I.T.) введения дозы всех мышей слегка анестезировали изофлураном в индукционной камере. После анестезии животных удаляли из камеры, удерживали вручную, открывали рот животного и отводили язык. Шприц объемом 1 мл с 50 мкл стерильного изотонического раствора для инъекций или блеомицина (0,70 ЕД/кг (DNC №0703-3155-01; TEVA) в 50 мкл стерильного изотонического раствора) присоединяли к игле зонда 24 размера. Иглу через зонд вводили в трахею и интратрахеально вводили дозу либо несущей среды, либо блеомицина. После доставки за животными непрерывно наблюдали до тех пор, пока они полностью не проснулись и не начали двигаться.
Для анализа экспрессии профибротического гена животных забирали через 14 дней после введения блеомицина. РНК собирали из цельных легких мышей и определяли экспрессию гена с помощью ОТ-кПЦР.
Для измерения отложения коллагена в легких мышей забирали через 24 дня после введения блеомицина. Мечение дейтерированной водой использовали для оценки нового синтеза коллагена в исследованиях (Blaauboer ME, et. al. Matrix Biol. 2013;32(7-8):424-31.). В наших исследованиях мечение начинали в день 9 после введения блеомицина с помощью I.P. введения дейтерированной воды (кат. №DLM-4-99.8-1000; Cambridge Isotope Laboratories) в разделенных дозах по 35 мл/кг в 2 приема с интервалом 4 часа. После этого 8% дейтерированной воды в питьевой воде предоставляли без ограничений в бутылке с водой до конца исследования.
Включение дейтерированной воды в гидроксипролин анализировали, как описано ранее (Gardner JL, et al. Am J Physiol Gastrointest Liver Physiol. 2007;292(6):G1695-705). Масс-спектрометрия и анализ выполняли в Metabolic Solutions. Содержание нового гидроксипролина выражали в «мкг на легкое».
Экспрессия генов
Анализы экспрессии генов в модели фиброза легких у мышей, индуцированного блеомицином, выполняли путем анализа опубликованного набора данных, доступ к которому можно получить с помощью номера доступа серии GEO GSE37635 (см. всемирную сеть на сайте ncbi.nlm.nih.gov/geo/query/acc.cgiacc=GSE37635) (см. также Blaauboer, 2013, Matrix Biol 32, 424-431).
РНК-секвенирование
Для проведения исследования на основе РНК-секвенирования на различных популяциях клеток легких в мышиной модели блеомицина целые легкие мыши собирали в день 14 после введения блеомицина. В контрольной группе, получавшей солевой раствор, было 5 мышей, а в группе, получавшей блеомицин, было 8 мышей. Отдельные клетки выделяли после расщепления протеазой в соответствии с нашим ранее опубликованным протоколом (Sun Т, et al., 2019, JCI Insight; Jun 18;5(14):e 128674). После промывания клетки блокировали очищенным крысиным антителом к CD16/CD32 мыши (кат.№553142; BD Biosciences) и окрашивали антителами к CD45 (кат.№17-0451-82, eBioscience), EpCAM (кат. №11-5791-82, eBioscience) и CD31 (кат. №25-0311-82, eBioscience). Четыре клеточные популяции от каждой мыши сортировали на основе их маркеров клеточной поверхности: лейкоциты (CD45+; EpCAM-); эпителиальные клетки (CD45-, ЕрСАМ+); эндотелиальные клетки (CD45-; EpCAM-; CD31+) и фибробласты (трижды негативные). РНК очищали из отсортированных клеток с использованием набора RNeasy Mini (кат.№74106; Qiagen). 0,5 мкг общей РНК использовали в качестве исходного материала для подготовки библиотеки с использованием набора для подготовки образцов РНК TruSeq v2 (Illumina, Сан-Диего, Калифорния), качество которого контролировали с помощью анализатора фрагментов и набора для количественного определения библиотеки (KAPA Biosystems, Уилмингтон, Массачусетс). Библиотеки мультиплексировали, а затем секвенировали на Illumina HiSeq2500 с полученимем 30 млн. считываемых значений с одним концом из 50 пар оснований. Прочитанные значения секвенирования сопоставляли с эталонным геномом мыши (GRCm38) с использованием выравнивателя коротких прочитанных значений GSNAP (Wu and Nacu, 2010) и сопоставляли с моделями генов RefSeq. Экспрессию измеряли в нормализованном количестве прочитанных значений на килобазу на миллион общих прочитанных значений (nRPKM).
Условные нокауты
Две нацеливающие конструкции нацеливали на экзон 6 в локусе Tgfb2 или Tgfb3 соответственно. Адено-FLP использовали для обработки положительных клонов ES для удаления маркера селекции неомицина PGK на стадии клеток ES. Создавали два сайта loxP, фланкирующие экзон 6 в локусах TGFβ2 или β3 у соответствующих мышей cKO.
кПЦР
Геномные ДНК легких выделяли у мышей, получавших тамоксифен, и оценивали с помощью количественной ПЦР (по сравнению с Tert). Экзон 3 расположен вне фланкированного локуса loxP и его использовали в качестве экспериментального контроля.
Результаты:
Интратрахеальная (I.Т.) стимуляция блеомицином является часто используемой моделью фиброза легких у грызунов in vivo. Блеомицин избирательно повреждает альвеолярные эпителиальные клетки, вызывая врожденный иммунный ответ с последующей дифференцировкой фибробластов в миофибробласты и отложением ЕСМ. В отличие от хронического прогрессирующего состояния, такого как IPF, модель блеомицина имеет определенные фазы: повреждение, воспаление, фиброз и, у молодых животных, разрешение. Для определения того, имеет ли значение экспрессия изоформы TGFβ в этой модели и на каких фазах, экспрессию генов легких у мышей оценивали в различные временные точки после i.t. инсталляции блеомицина.
Для оценки кинетики экспрессии изоформы TGFβ в этой модели, экспрессию генов легких оценивали у мышей в различные временные точки после i.t. инсталляция блеомицина на основе опубликованного набора транскриптомных данных (Blaauboer, см. выше). В то время как экспрессия Tgfb1 не была существенно повышена у мышей, которым вводили блеомицин, в любой временной точке, и фактически немного снижалась в день 21 и 28, экспрессия Tgfb2 и Tgfb3 существенно увеличивалась после введения блеомицина в день 7 и 14 после стимуляции (Фиг. 8А, Фиг. 8Б, Фиг. 8В). Этот паттерн хорошо перекрывается с индукцией генов, связанных с фиброгенезом и передачей сигналов TGFβ, таких как Serpinel, Fn1 и Col1a1 (Фиг. 8Г, Фиг. 8Д, Фиг. 8Е).
Значительная активация Tgfb2 и Tgfb3 во время фиброзной фазы модели совпадала с паттерном экспрессии, наблюдаемым в легких при IPF (Фиг. 3А). Для дополнительной характеристики конкретных типов клеток, ответственных за экспрессию каждой изоформы TGFβ в этой модели, РНК-секвенирование выполняли на отсортированных популяциях клеток легких через 14 дней после введения блеомицина, когда достигали пика экспрессии множественные связанные с фиброзом гены. В соответствии с данными scRNAseq из ткани легких при IPF человека (Фиг. 4А), лейкоциты (CD45+) имели наиболее высокие уровни экспрессии Tgfb1. Tgfb2 и Tgfb3 преимущественно экспрессировались в фибробластах (CD45-, ЕрСАМ- CD31-); Tgfb2 также индуцировался в эпителиальных клетках, а TGFβR-индуцируемые гены Col1a1 и Serpinel индуцировались главным образом в фибробластах после обработки блеомицином. Хотя существенной активизации Tgfb3 не наблюдалось ни в одной популяции отдельных клеток после обработки блеомицином, значительная активация Tgfb3 в объемной легочной ткани (Фиг. 8А, Фиг. 8Б, Фиг. 8В) была связана с увеличением доли фибробластов, экспрессирующих Tgfb3, в эти временные точки.
Поскольку конститутивная делеция Tgfb2 или Tgfb3 приводит к эмбриональной или перинатальной летальности, создавали индуцируемые нокауты этих генов для изучения их роли в индуцированном блеомицином фиброзе легких у взрослых животных путем введения сайтов loxP в Tgfb2 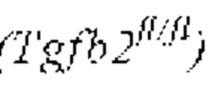 и Tgfb3 (
и Tgfb3 (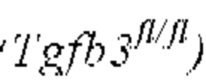 соответственно,
соответственно, 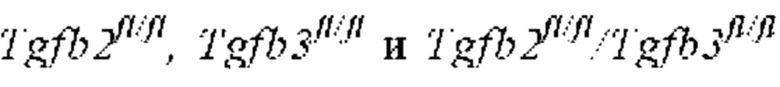 скрещивали с ROSA-CreERT2, и мышам в возрасте 12-16 недель вводили тамоксифен для удаления соответствующих аллелей TGFβ (одиночные условно нокаутированные мыши будут обозначаться как TGFβ2.cKO и TGFβ3.cKO, а мыши с двойной мишенью будут обозначаться как TGFβ2/3.cDKO). Никаких грубых аномалий или заболеваний не наблюдали у животных cKO в течение 6 недель после обработки тамоксифеном. Для оценки роли TGFβ2 и TGFβ3 в фиброзе легких взрослых животных cKO обрабатывали тамоксифеном за 2 недели до введения блеомицина. С дня 9 до 24 после введения блеомицина вводили дейтерированную питьевую воду для определения синтеза нового коллагена во всех легких путем измерения дейтерированного гидроксипролина. Как у мышей TGFβ2.cKO, так и у мышей TGFβ3.cKO синтез нового коллагена был значительно снижен, а также у мышей TGFβ2/3.cDKO, что свидетельствует о том, что и TGFβ2, и TGFβ3 вносят вклад в фиброгенез в этой модели (Фиг. 52А и Фиг. 52Б).
скрещивали с ROSA-CreERT2, и мышам в возрасте 12-16 недель вводили тамоксифен для удаления соответствующих аллелей TGFβ (одиночные условно нокаутированные мыши будут обозначаться как TGFβ2.cKO и TGFβ3.cKO, а мыши с двойной мишенью будут обозначаться как TGFβ2/3.cDKO). Никаких грубых аномалий или заболеваний не наблюдали у животных cKO в течение 6 недель после обработки тамоксифеном. Для оценки роли TGFβ2 и TGFβ3 в фиброзе легких взрослых животных cKO обрабатывали тамоксифеном за 2 недели до введения блеомицина. С дня 9 до 24 после введения блеомицина вводили дейтерированную питьевую воду для определения синтеза нового коллагена во всех легких путем измерения дейтерированного гидроксипролина. Как у мышей TGFβ2.cKO, так и у мышей TGFβ3.cKO синтез нового коллагена был значительно снижен, а также у мышей TGFβ2/3.cDKO, что свидетельствует о том, что и TGFβ2, и TGFβ3 вносят вклад в фиброгенез в этой модели (Фиг. 52А и Фиг. 52Б).
Значительная активация TGFβ2 и TGFβ3 во время «фиброзной» фазы модели отражала распределение изоформ, наблюдаемое в легких при IPF. Таким образом, индуцированная TGFβR экспрессия гена-мишени в день 14 и накопление коллагена в день 24 были приоритетными в качестве ключевых показаний в контексте профилактической обработки антителами к TGFβ в ходе исследования для оценки их активности и эффективности in vivo (схема, Фиг. 9А).
При дозе 10 мг/кг три раза в неделю однократное ингибирование TGFβ2 с помощью 6F12, TGFβ3 с помощью 2А10 или обоих с помощью комбинации 6F12 +2А10 или монотерапией с помощью 4А11 снижали синтез нового коллагена, что отражалось во включении дейтерия в гидроксипролин легких у животных, которым вводили D2O с питьевой водой с дня 9 до 23 после введения блеомицина (Фиг. 9Б). В качестве более близкого фармакодинамического показателя у отдельной когорты животных экспрессия гена фибронектина-1 (fn1) в легких в день 14 после введения блеомицина была значительно снижена под действием 6F12, 2А10, комбинации 6F12+2А10 или 4А11. Однако 16С10, менее активное антитело к TGFβ3, не влияло на экспрессию fn1 в легких (Фиг. 9В). Сходные эффекты наблюдали для других генов, индуцируемых TGFβ, включая serpinel, col1a1, col1a2 и col3a1. В то время как однократное введение либо антитела к TGFβ2, либо антитела к TGFβ3 снижало экспрессию генов и синтез коллагена, комбинация обоих путем одновременного введения 6F12+2A10 или однократного введения 4А11 давала численное превосходство в нескольких экспериментах, предполагая, что как TGFβ2, так и TGFβ3 способствует фиброгенезу в этой модели, и что антитела 6F12, 2А10 и 4А11 могут ингибировать изоформы TGFβ in vivo (Фиг. 9Г и Фиг. 9Д).
Пример 6: Гуманизация mAb 2А10 к TGFβ3
Материалы и методы:
Измерение аффинности антител с помощью BIAcore
Для определения аффинности связывания всех гуманизированных антител 2А10 использовали измерение на основе поверхностного плазмонного резонанса (SPR) с помощью прибора BIAcore™-Т200. Вкратце, биосенсорный чип серии S СМ5 с белком А сначала применяли для захвата каждого гуманизированного варианта 2А10 для достижения приблизительно 100 единиц ответа (RU) на каждой проточной кювете с последующей инъекцией 3-кратных серийных разведений TGFβ1, TGFβ2 или TGFβ3 человека (от 0,02 нМ до 5 нМ) в буфере HBS-EP (0,01 М HEPES рН 7,4, 0,15 М NaCl, 3 мМ EDTA и 0,05% об./об. поверхностно-активного вещества Р20) при 25°С со скоростью потока 50 мкл/мин. для измерения кинетики. Скорости ассоциации (kon) и скорости диссоциации (koff) рассчитывали с использованием простой модели связывания Ленгмюра один к одному (оценочное программное обеспечение BIAcore Т200, версия 2.0) для определения равновесной константы диссоциации (KD), которую рассчитывали как отношение koff/kon.
Анализ блокирования TGFβ на основе клеток с применением HEKBlue
Клетки HEK-Blue™ TGFβ (InvivoGen, Сан-Диего, Калифорния), стабильно экспрессирующие белки TGFβRI, Smad3, Smad4 человека и репортерный ген секретируемой щелочной фосфатазы (SEAP) под контролем минимального промотора β-глобина, слитого с тремя Smad3/4-связывающими элементами (SBE), использовали для оценки блокирующей активности 2А10. 2А10 инкубировали с каждой изоформой TGFβ (PeproTech, Роки-Хилл, Нью-Джерси) при 25°С в течение 1 часа перед добавлением к 4,5×104 клеток HEK-Blue™ TGF-β, высеянных в 96-луночный планшет в 100 мкл буфера для анализа (DMEM с высоким содержанием глюкозы с 10% инактивированной нагреванием FBS, 2 мМ L-глутамина и 0,5% пенициллин-стрептомицина). Конечная концентрация антитела и TGFβ составляла 0,001-200 нМ и 20 пМ соответственно. Планшет инкубировали при 37°С и 5% СО2 в течение 18-22 часов. Уровень SEAP в супернатанте клеток определяли с помощью анализа QUANTI-Blue™ в соответствии с инструкциями производителя (InvivoGen). mAb ко всем видам TGFβ нейтрализует TGFβ1, 2 и 3 при низких клеточных потенциях (IC50), составляющих 0,72, 2,2 и 0,026 нМ соответственно.
Результаты: Гуманизация 2А10:
Гуманизация 2А10. шаг 1: Привитие CDR 2А10 крысы в человеческие каркасные области:
Моноклональное антитело 2А10 гуманизировали, как описано ниже. Номера остатков представлены согласно Kabat et al. {Sequences of proteins of immunological interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)). Для гуманизации антитела 2A10 гипервариабельные области антитела 2А10 крысы (r2A10) конструировали в акцепторные каркасные области зародышевой линии человека IGKV1D-39 и IGHV3-49 для создания гуманизированной версии 1 антитела 2А10 (h2A10.v1) и акцепторные каркасные области зародышевой линии человека IGKV1D-39 и IGHV3-23 для создания гуманизированной версии 2 2А10 (h2A10.v2). В частности, из вариабельного домена легкой цепи (VL) r2A10 положения 24-34 (L1), 50-56 (L2) и 89-97 (L3) прививали в IGKV1D-39. Из вариабельного домена тяжелой цепи (VH) r2A10 положения 26-35 (H1), 50-65 (Н2) и 95-102 (Н3) прививали в IGHV3-49 и IGHV3-23 соответственно.
Кроме того, положение 4 в каркасной области I, положение 38 и 43 в каркасной области II, положение 58 в каркасной области III VL, положение 47 и 49 в каркасной области II и положение 78 в каркасной области III VH были сохраняли из крысиной последовательности в h2A10.v1. Аналогично h2A10.v1, два дополнительных положения (73 и 76) в каркасной области III VH также сохраняли из крысиной последовательности в h2A10.v2. Было обнаружено, что эти остатки являются частью остатков каркасной области, выступающих в качестве «зоны Верньера», которая может регулировать структуру CDR и точно настраивать соответствие антигена. См., например, Foote and Winter, J. Mol. Biol. 224: 487-499 (1992). Эти определения CDR включают положения, определяемые гипервариабельностью их последовательностей (Wu, Т.Т. & Kabat, Е.А. (1970)), их структурным расположением (Chothia, С.& Lesk, A.M. (1987)) и их участием в контактах антиген-антитело (MacCallum et at. J. Mol. Biol. 262: 732-745 (1996)).
Обе первичные гуманизированные версии подвергали связыванию изоформ TGFβ человека в BIAcore, и результаты показали, что h2A10.v2 более сопоставимо с r2A10, чем h2A10.v1 в сохранении связывания TGFβ3 человека (таблица 9). Интересно, что после гуманизации обе гуманизированные версии показали снижение связывания TGFβ2 человека по сравнению с r2A10.
Гуманизация 2А10, шаг 2: Завершающая очистка каркасных областей: При завершающей очистке каркасных областей гуманизированного 2А10 более гуманизированные варианты 2А10 создавали путем замены каждого крысиного остатка гуманизированного 2А10 в зоне Верньера соответствующим человеческим остатком из последовательности зародышевой линии. Затем каждый вариант оценивали в отношении связывания изоформ TGFβ человека в BIAcore и блокирующей активности в HEKBlue для выяснения его важности.
В h2A10.v1 этими вариантами каркасных областей с завершающей очисткой являются h2A10.v1.1 (VL: L4M), h2A10.v1.2 (VL: H38Q), h2A10.v1.3 (VL: Q43A), h2A10.v1.4 (VL: I58V), h2A10.v1.5 (VH: L47W), h2A10.v1.6 (VH: A49G), h2A10.v1.7 (VH: V78A) (таблица 9). При очистке некоторые из вариантов каркасных областей с завершающей очисткой в h2A10.v1 имеют тенденцию к проблемам с экспрессией (h2A10.v1.l) и агрегацией (мономер<95% для h2A10.v1.3, h2A10.v1.4, h2A10.v1.6). Поэтому было решено перейти к вариантам h2A10.v2.
В h2A10.v2 этими вариантами каркасных областей с завершающей очисткой являются h2A10.v2.1 (VL: L4M), h2A10.v2.2 (VL: H38Q), h2A10.v2.3 (VL: Q43A), h2A10.v2.4 (VL: I58V), h2A10.v2.5 (VH: L47W), h2A10.v2.6 (VH: A49S), h2A10.v2.7 (VH: D73N), h2A10.v2.8 (VH: S76N), h2A10.v2.9 (VH: V78L). В отличие от вариантов h2A10.v1, все варианты h2A10.v2 экспрессировались и хорошо себя проявляли при очистке (мономер>99%). В исследовании связывания BIAcore большинство вариантов оказались такими же, как исходное h2A10.v2, за исключением вариантов h2A10.v2.1 (VL: L4M) и h2A10.v2.7 (VH: D73N) с незначительным снижением связывания TGFβ3 (в 2-3 раза) и варианта h2A10.v2.5 (VH: L47W) с более значительным снижением связывания TGFβ3 (в 385 раз) (таблица 9). Удивительно, но в клеточном исследовании блокирования с применением HEKBlue TGFβ как h2A10.v2.5 (VH: L47W), так и h2A10.v2.7 (VH: D73N) показал значительную потерю блокировки TGFβ3 (более чем в 10 раз), но не для h2A10.v2.1 (VL: L4M) (таблица 9). В целом было установлено, что все крысиные остатки в зоне Верньера h2A10.v2 могут быть заменены остатками зародышевой линии человека, за исключением положения 4 в каркасе I VL, положения 47 в каркасе II и положения 73 в каркасе III VH.
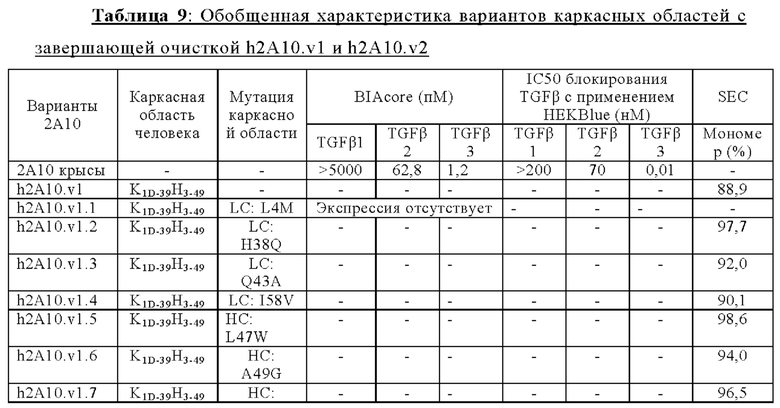
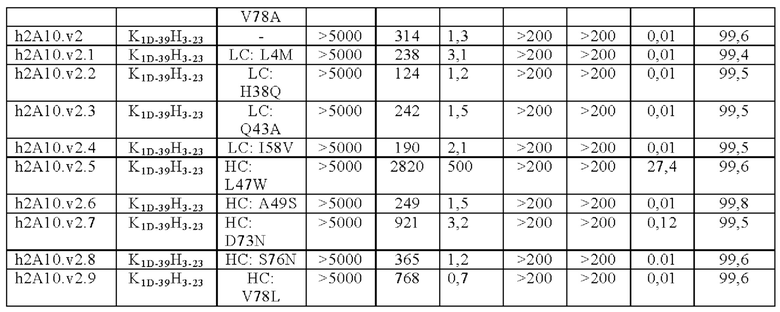
Гуманизация 2А10, шаг 3: Фиксация сайта N-связанного гликозилирования CDR-H2 (N54Y55T56)
В h2A10.v2 потенциальный сайт N-связанного гликозилирования (N54Y55T56) в CDR-H2 идентифицировали в виде потенциальной помехи. Для решения этой проблемы применяли два подхода, один из которых заключался в замене Asn в положении 54 на нуклеофильный Ser (h2A10.v2.N54S) или его гомолог Gln (h2A10.v2.N54Q), а другой заключался в сохранении Asn в положении 54, но замене Thr в положении 56 на меньший остаток Ala (h2A10.v2.T56A). Все три варианта экспрессировались и хорошо себя проявляли при очистке (мономер>99%) (таблица 10). Однако в анализе BIAcore только вариант h2A10.v2.T56A сохранил аналогичное связывание TGFβ3 человека, в то время как два других варианта показали незначительное снижение (<2 раза) по сравнению с исходным h2A10.v2. Однако, несмотря на различия в связывании, не наблюдалось никаких различий в отношении блокирования TGFβ3 человека для всех трех вариантов.
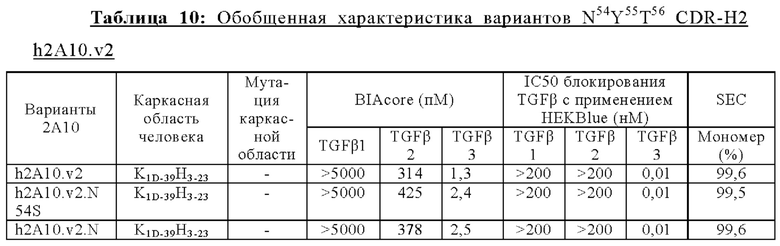

Гуманизация 2А10, шаг 4: Создание конечной версии
В результате объединения результатов завершающей очистки каркасных областей и репарации сайта N-связанного гликозилирования CDR-H2 создавали две последние гуманизированные версии 2А10, h2A10.v3 и h2A10.v4, на основе дальнейшего конструирования h2A10.v2. Обе версии сохраняли L в положении 4 каркаса I VL и L в положении 47 каркаса II, D в положении 73 каркаса III VH из r2A10. Остальные шесть остатков каркасных областей, полученные из r2A10 в зоне Верньера, заменяли соответствующими остатками зародышевой линии человека, такими как Q в положении 38 и А в положении 43 в каркасе II, V в положении 58 в каркасе III VL и S в положении 49 в каркасе II, N в положении 76 и L в положении 78 в каркасе III VH. Единственное различие между двумя версиями заключалось в сайте N-связанного гликозилирования CDR-H2, которое представляло собой S54Y55T56 в h2A10.v3 и N54Y55A56 в h2A10.v4. Однако это различие было значительным с функциональной точки зрения, поскольку в анализах связывания BIAcore TGFβ3 и блокирования с применением HEKBlue TGFβ3 h2A10.v4 оказалось лучше, чем h2A10.v3, и сопоставимо с исходным r2A10 (таблица 11).
В целом, процесс гуманизации 2А10 был довольно сложным. Проблемы при гуманизации крысиного 2А10 включали неожиданные трудности с экспрессией антител на 1-м этапе (привитие CDR 2А10 на ближайшую пару VH/VL человеческих каркасных областей) и проблемы со стабильностью вариантов на 2-м этапе завершающей очистки каркасных областей (см. Фиг. 10, Фиг. 11).
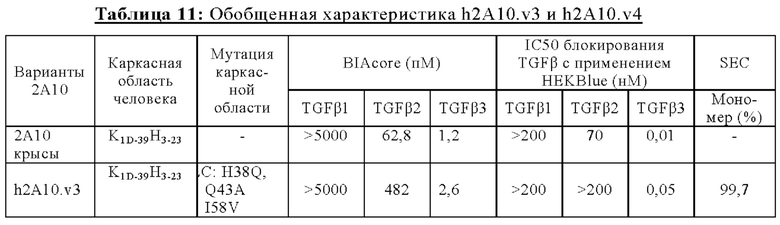
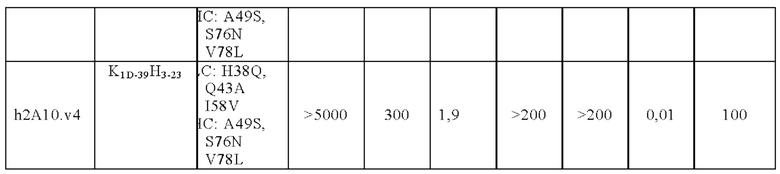
Последовательности вариантов гуманизированного антитела 2А10 (CDR выбранных вариантов, вариабельные области и полные последовательности) показаны в таблице 12, таблице 13 и таблице 14 ниже, а выравнивание последовательностей изображено на Фиг. 12, Фиг. 13, Фиг. 14, Фиг. 15 и Фиг. 16.




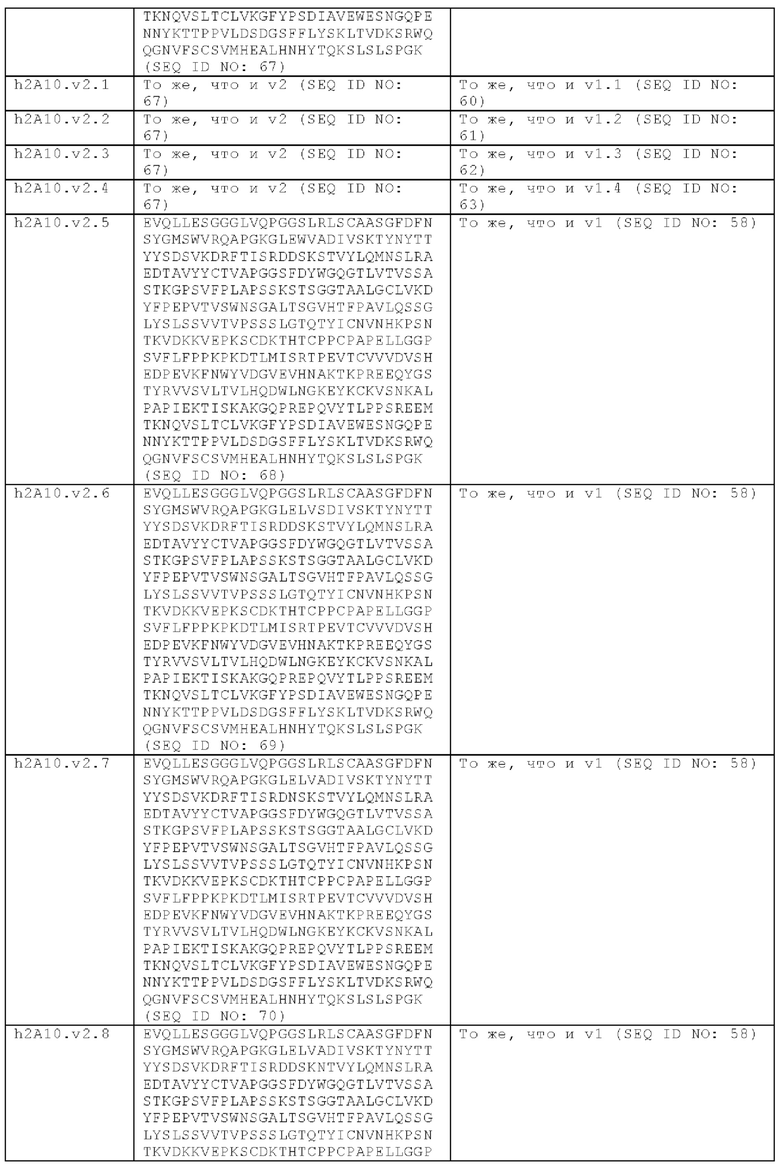


Пример 7: ФК исследование гуманизированных вариантов 2А10 у яванских макаков
Материалы и методы:
Восемь экспериментально наивных самок яванских макаков (Масаса fascicularis, камбоджийского происхождения) относили к одной из двух групп (n=4/группа). В начале (день - 1) исследования животным было примерно 4 года, и они весили примерно 2,5-4 кг, и для исследования использовали только животных, которые казались здоровыми и не имели явных аномалий. Животным в группах 1 или 2 вводили однократно I.V. болюсную дозу 10 мг/кг антитела h2A10.v3 или h2A10.v4 к TGFβ3 соответственно. Дозы вводили через головную вену на предплечье. В различные временные точки вплоть до 43 дней после введения дозы образцы сыворотки крови (n=4/момент времени) собирали и анализировали в отношении концентраций тестируемого образца в сыворотке крови методами ELISA.
Концентрации в сыворотке крови животных, которым вводили 10 мг/кг вариантов антител h2A10.v3 и h2A10.v4 к TGFβ3, определяли методом ELISA. 384-луночные планшеты покрывали 0,5 мкг/мл овечьего антитела к IgG человека (Binding Site, Сан-Диего, Калифорния, США), разведенного в буфере для покрытия (0,05 М карбонат/бикарбонатный буфер, рН 9,6) и инкубировали в течение ночи при 4°С. Планшеты промывали 3 раза промывочным буфером (0,5% Tween-20 в буфере PBS, рН 7,4) и обрабатывали блокирующим буфером (PBS/0,5% BSA/15 ppm Proclin, рН 7,4) в течение 1-2 часов при комнатной температуре (к.т.). Планшеты повторно промывали 3 раза промывочным буфером, а затем добавляли образцы, разведенные в растворителе для образцов (PBS/0,5% BSA/0,05% Tween 20/5 мМ EDTA/0,25% CHAPS/0,35M NaCl/15 ppm Proclin, рН 7,4) в лунки и инкубировали в течение 2 часов при к.т. На следующий день планшеты промывали 6 раз промывочным буфером. Детектирующее антитело, конъюгированное с пероксидазой хрена (HRP) козье антитело к IgG (H+L) человека (Bethyl Laboratories, Inc., Монтгомери, Техас, США), разведенное до 100 нг/мл в буфере для анализа (PBS/0,5% BSA/15 ppm Proclin/0,05% Tween 20, рН 7,4), добавляли в лунки и инкубировали на шейкере в течение 1 часа при к.т. Планшеты промывали 6 раз промывочным буфером и проявляли с использованием субстрата пероксидазы ТМВ (Moss Inc., Пасадена, Мериленд) в течение 20 минут, а затем 1 М фосфорной кислоты для остановки реакции. Поглощение измеряли при 450 нм относительно эталонной длины волны 620 нм. Концентрацию образцов экстраполировали на основе 4-параметрической аппроксимации стандартной кривой. Отчетный диапазон анализа составлял 0,156-10 нг/мл. Минимальное необходимое разведение составляло 1:100 с минимальной количественной концентрацией 16 нг/мл.
Результаты:
После одной I.V. болюсной дозы 10 мг/кг антитела h2A10.v3 к TGFβ3, средний клиренс (CL) составлял 4,35 мл/день/кг, Cmax составляла 238 мкг/мл, концентрация (AUCinf) составляла 2400 дней • мкг/мл, конечный период полужизни (t1/2, λz) составлял 15,1 дней, объем распределения в равновесном состоянии (Vss) составлял 88,2 мл/кг. h2A10.v3 продемонстрировало кажущееся двухфазное распределение, характеризующееся быстрой фазой начального распределения, за которой следует более медленная фаза элиминации (Фиг. 17).
После одной I.V. болюсной дозы 10 мг/кг антитела h2A10.v4 к TGFβ3, средний клиренс (CL) составлял 4,21 мл/день/кг, Cmax составлял 252 мкг/мл, концентрация (AUCinf) составляла 2470 дней ⋅ мкг/мл, конечный период полужизни (t1/2, λz) составлял 16,0 дней, объем распределения в равновесном состоянии (Vss) составлял 79,1 мл/кг соответственно. h2A10.v4 продемонстрировало кажущееся двухфазное распределение, характеризующееся быстрой фазой начального распределения, за которой следует более медленная фаза элиминации.
Фармакокинетика вариантов антител h2A10.v3 и h2A10.v4 к TGFβ3 после однократного I.V. болюсного введения в дозе 10 мг/кг была ожидаемой для моноклонального антитела на основе IgG1 человека у яванских макаков и аналогичной введению типичных моноклональных антител на основе IgG1 и IgG4.
ФК данные обобщены в таблице 15 ниже.
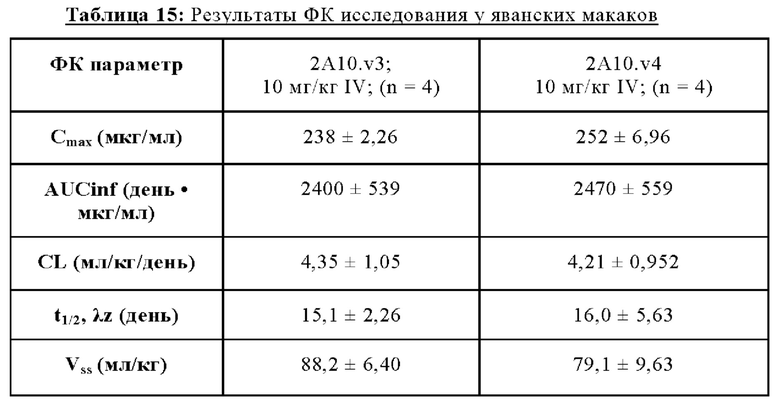
Пример 8: Гуманизация 4А11 (антитела к TGFβ2/3 с двойной специфичностью)
Материалы и методы:
Измерение аффинности антител с помощью BIAcore
Определение аффинности связывания всех гуманизированных антител 4А11 проводили с использованием измерения SPR с помощью прибора BIAcore™-Т200, как описано выше в примере 6 (пример Ab 2А11).
Анализ блокирования TGFβ на основе клеток с применением HEKBlue
Анализ блокирования TGFβ3 на основе клеток с применением HEKBlue проводили, как описано выше в примере 6.
Результаты:
Гуманизация 4АН:
Гуманизация 4А11, шаг 1: Привитие CDR 4А11 кролика на человеческие каркасные области:
Полученное из В-клеток кролика моноклональное антитело 4А11 гуманизировали, как описано ниже. Номера остатков представлены согласно Kabat et al., Sequences of proteins of immunological interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991).
Вариабельные области антитела 4A11 кролика (4А11 кролика) сначала выравнивали по отношению ко всем зародышевым линиям человека. Четыре человеческие зародышевые линии легкой цепи (IGKV4-1, IGKV1D-39, IGKV3-20, IGKV2-24) и две человеческие зародышевые линии тяжелой цепи (IGHV3-48, IGHV4-59) идентифицировали в качестве наиболее близких акцепторных каркасных областей для привития легкой цепи и гипервариабельных областей тяжелой цепи (CDR) 4А11 кролика соответственно. В частности, из вариабельного домена (VL) легкой цепи 4А11 кролика положения 24-34 (L1), 50-56 (L2) и 89-97 (L3) прививали в IGKV4-1, IGKV1D-39, IGKV3-20 и IGKV2-24. Из вариабельного домена тяжелой цепи (VH) 4А11 кролика положения 26-35 (H1), 50-65 (Н2) и 95-102 (Н3) прививали в IGHV3-48 и IGHV4-59 соответственно.
Восемь гуманизированных вариантов создавали путем шаффлинга различных акцепторных каркасных областей для привития: гуманизированной версии 1 4А11 (h4A11.v1) посредством IGKV4-1 и IGHV3-48; гуманизированной версии 2 4А11 (h4A11.v2) посредством IGKV1D-39 и IGHV3-48; гуманизированной версии 3 4А11 (h4A11.v3) посредством IGKV4-1 и IGHV4-59; гуманизированной версии 4 4А11 (h4A11.v4) посредством IGKV1D-39 и IGHV4-59; гуманизированной версии 5 4А11 (h4A11.v5) посредством IGKV3-20 и IGHV3-48; гуманизированной версии 6 4А11 (h4A11.v6) посредством IGKV2-24 и IGHV3-48; гуманизированной версии 7 4А11 (h4A11.v7) посредством IGKV3-20 и IGHV4-59; гуманизированной версии 8 4А11 (h4A11.v8) посредством IGKV2-24 и IGHV4-59 (таблица 16).
В дополнение к CDR, привитым на ближайшие акцепторные каркасные области, описанные выше, выполняли следующие модификации:
Каркасная область легкой цепи:
h4A11.v1 и h4A11.v3 использовали одну и ту же акцепторную каркасную область IGKV4-1, сохраняли кроличьи остатки в положении 2 и 4 в каркасной области I и в положении 36 в каркасной области II VL из 4А11 кролика.
h4A11.v2 и h4A11.v4 использовали одну и ту же акцепторную каркасную область IGKV1D-39, сохраняли кроличьи остатки в положении 2 и 4 в каркасной области I и в положении 36 и 43 в каркасной области II VL из 4А11 кролика.
h4A11.v5 и h4A11.v7 использовали одну и ту же акцепторную каркасную область IGKV3-20, сохраняли кроличьи остатки в положении 2 в каркасной области I, в положении 36 и 43 в каркасной области II и в положении 58 в каркасной области III VL из 4А11 кролика.
h4A11.v6 и h4A11.v8 использовали одну и ту же акцепторную каркасную область IGKV2-24, сохраняли кроличьи остатки в положении 2 и 4 в каркасной области I и в положении 36 в каркасной области II VL из 4А11 кролика.
Каркасная область тяжелой цепи:
h4A11.v1, h4A11.v2, h4A11.v5, h4A11.v6 использовали одну и ту же акцепторную каркасную область IGHV3-48, сохраняли кроличьи остатки в положениях 2 и 24 в каркасной области I, в положении 48 и 49 в каркасной области II, в положении 71, 73, 78 и 91 в каркасной области III и в положении 105 в каркасной области IV VH из 4А11 кролика.
h4A11.v3, h4A11.v4, h4A11.v7, h4A11.v8 использовали одну и ту же акцепторную каркасную область IGHV4-59, сохраняли кроличьи остатки в положении 2 в каркасной области I, в положении 37 в каркасной области II, в положении 67, 71, 73, 78 и 91 в каркасной области III и в положении 105 в каркасной области IV VH из 4А11 кролика.
Было обнаружено, что все из этих остатков являются частью остатков каркасной области, выступающих в качестве «зоны Верньера», которая может регулировать структуру CDR и точно настраивать соответствие антигена. См., например, Foote and Winter, J. Mol. Biol. 224: 487-499 (1992). Эти определения CDR включают положения, определяемые гипервариабельностью их последовательностей (Wu, Т.Т. & Kabat, Е.А. (1970)), их структурным расположением (Chothia, С. & Lesk, A.M. (1987)) и их участием в контактах антиген-антитело (MacCallum et al. J. Mol. Biol. 262: 732-745 (1996)).
Все восемь гуманизированных вариантов 4A11 дополнительно подвергали анализу BIAcore для определения аффинности связывания изоформ TGFβ человека и анализу на основе клеток с применением HEKBlue для оценки эффективности блокирования изоформ TGFβ человека. Хотя наблюдали некоторое падение (в 3-10 раз) связывания TGFβ2 и 3 человека для большинства вариантов, степень снижения блокирования TGFβ2 и 3 человека была более выраженной и значительной (таблица 16). Во всех восьми гуманизированных версиях только h4A11.v7 сохраняло сопоставимую блокировку TGFβ2 и 3 человека по сравнению с 4А11 кролика, несмотря на снижение связывания TGFβ2 и 3 человека в несколько раз. Поэтому этот клон переносили на следующий этап гуманизации, завершающую очистку каркасных областей.
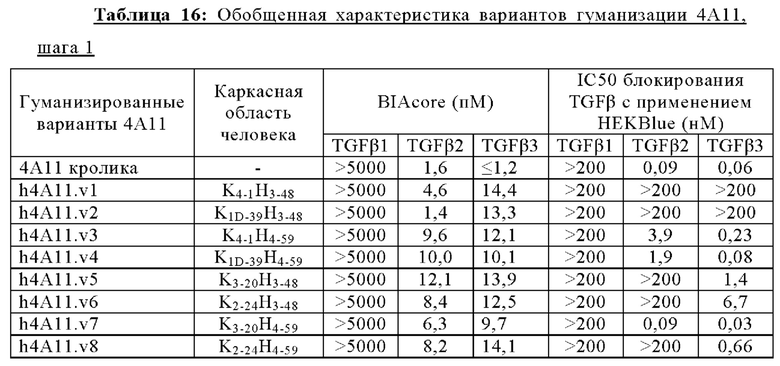
Гуманизация 4А11, шаг 2: завершающая очистка каркасных областей h4A11.v7:
При завершающей очистке каркасных областей h4A11.v7 создавали три варианта (v7.1 - v7.3) с удалением аминокислотной вставки, введенной при гуманизации, шаге 1. Затем создавали другие гуманизированные варианты путем замены каждого кроличьего остатка h4A11.v7 в зоне Верньера соответствующими остатками зародышевой линии человека (v7.4 v7.15). Для определения того, какие остатки необходимо сохранить, связывание с изоформами TGFβ каждого варианта затем оценивали с помощью BIAcore, а эффективность блокирования оценивали с помощью анализа на основе клеток с применением HEKBlue.
В h4A11.v7 этими вариантами каркасных областей с завершающей очисткой являются h4A11.v7.1 (VH: делеция Е1), h4A11.v7.2 (VH: делеция K75 и N76), h4A11.v7.3 (VH: делеция E1, K75 и N76), h4A11.v7.4 (VL: A2I), h4A11.v7.5 (VL: F36Y), h4A11.v7.6 (VL: P43A), h4A11.v7.7 (VL: V58I), h4A11.v7.8 (VH: Q2V), h4A11.v7.9 (VH: V37I), h4A11.v7.10 (VH: F67V), h4A11.v7.11 (VH: K71V), h4A11.v7.12 (VH: S73T), h4A11.v7.13 (VH: V78F), h4A11.v7.14 (VH: F91Y), h4A11.v7.15 (VH: P105Q) (таблица 17).
Большинство вариантов оказались сходными или слегка улучшенными по сравнению с исходным h4A11.v7 в анализе связывания и активности, за исключением значительного влияния на h4A11.v7.11 (VH: K71V) с некоторым снижением связывания TGFβ2 (в 6 раз) и связывания TGFβ3 (в 2 раза) и значительно большей потерей в блокировании TGFβ2 (>2000 раз) и блокировании TGFβ3 (в 118 раз). Имел место один вариант с минорным влиянием: h4A11.v7.5 (VL: F36Y) со снижением как связывания, так и блокирования TGFβ2 почти в 2 раза, и два варианта с субминорным влиянием: h4A11.v7.13 (VH: V78F) со снижением связывания TGFβ2 в 2,5 раза; h4A11.v7.14 (VH: F91Y) со снижением связывания TGFβ2 в 4 раза (таблица 17). Таким образом, было установлено, что все кроличьи остатки в зоне Верньера h4A11.v7 могут быть заменены остатками зародышевой линии человека, за исключением основного положения 71 в каркасной области III VH, минорного положения 36 в каркасной области II VL и/или субминорного положения 78 и 91 в каркасной области III VH.
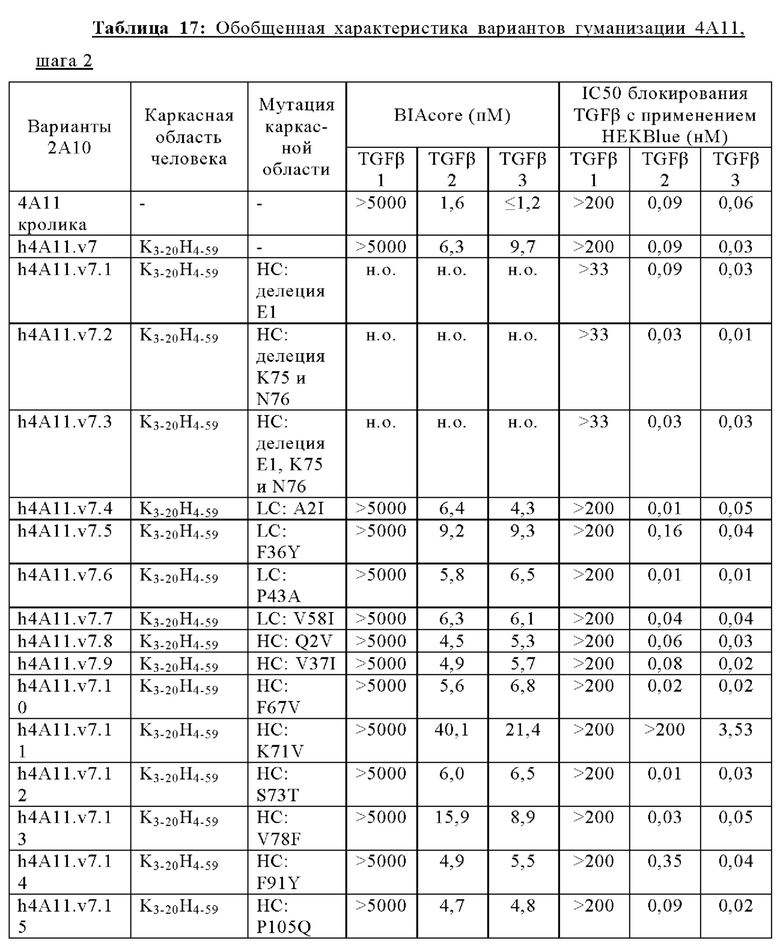
Гуманизация 4А11, шаг 3: Создание и отбор конечных версий
В соответствии с результатами завершающей очистки каркасных областей h4A11.v7, сначала получали h4A11.v7.18 путем замены некритических остатков каркасной области h4A11.v7 в зоне Верньера соответствующими остатками зародышевой линии человека, которые представляли собой А - I в положении 2 в каркасной области, Р - А в положении 43 в каркасной области II и V - I в положении 58 в каркасной области III VL и Q - V в положении 2 в каркасной области I, V - I в положении 37 в каркасной области II, F - V в положении 67 в каркасной области III, S - Т в положении 73 в каркасной области III, Р - Q в положении 105 в каркасной области IV VH (таблица 18). Вместе со всеми вышеуказанными заменами два других субминорных остатка каркасной области в h4A11.v7, V78 или F91 в каркасной области III VH также заменяли соответствующими остатками зародышевой линии человека для создания 2 других гуманизированных версий, h4A11.v7.16. (VH: V78F) и h4A11.v7.17 (VH: F91Y). Еще одну гуманизированную версию, h4A11.v7.19, создавали путем включения делеции K75 и N76 в каркасную область III VH, что способствовало улучшению блокировки TGFβ2 и 3 при завершающей очистке каркасной области h4A11.v7 (таблица 17, выше: h4A11.v7.2 и h4A11.v7.3).
Интересно, что при сравнении всех 4 окончательных гуманизированных версий только h4A11.v7.19 превосходит h4A11.v7 и сопоставимо с исходным 4А11 кролика не только в отношении связывания TGFβ2 и TGFβ3 человека, но также и в отношении блокирования TGFβ2 и TGFβ3 человека.
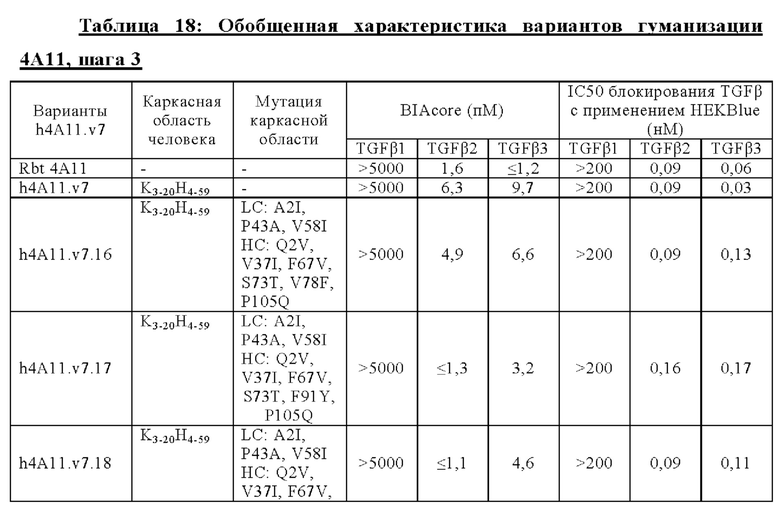

Аминокислотные последовательности вариантов 4А11 представлены в таблице 19 и таблице 20 ниже, а выравнивание последовательностей изображено на Фиг. 18, Фиг. 19, Фиг. 20 и Фиг. 21.
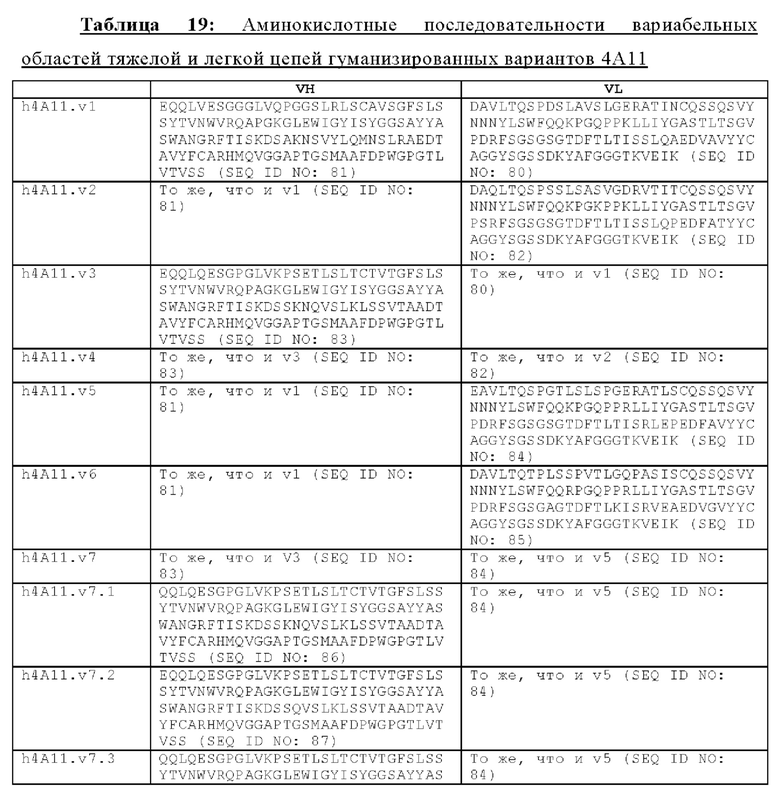
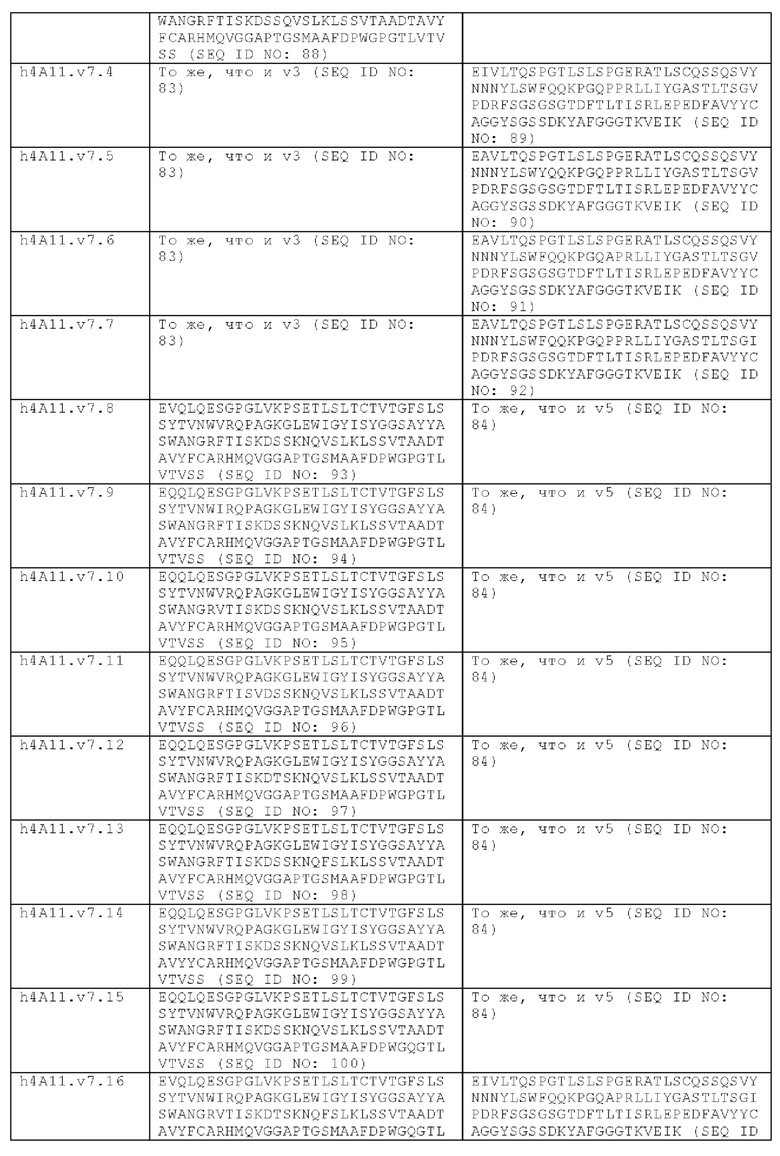

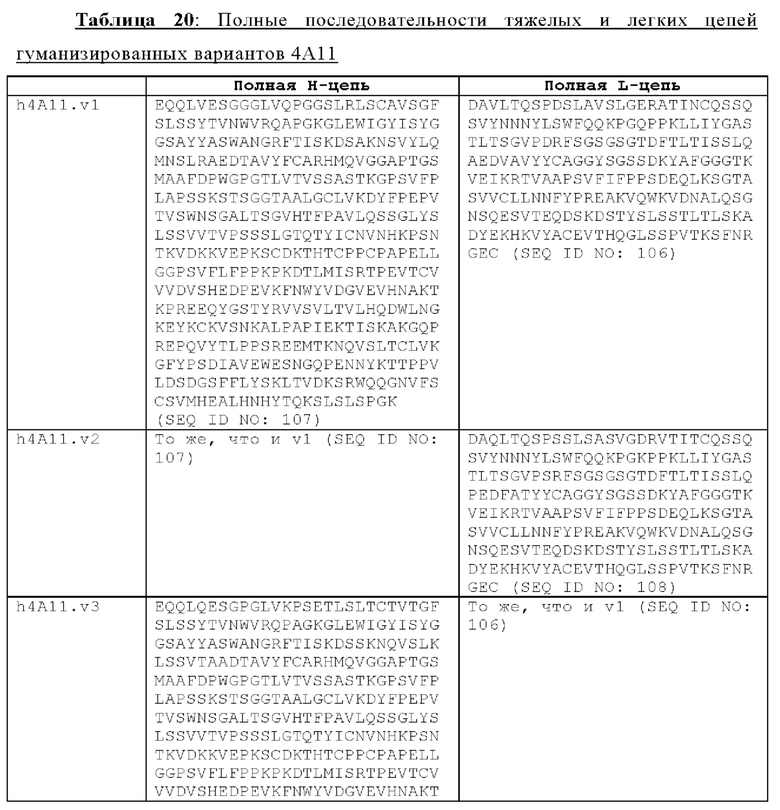
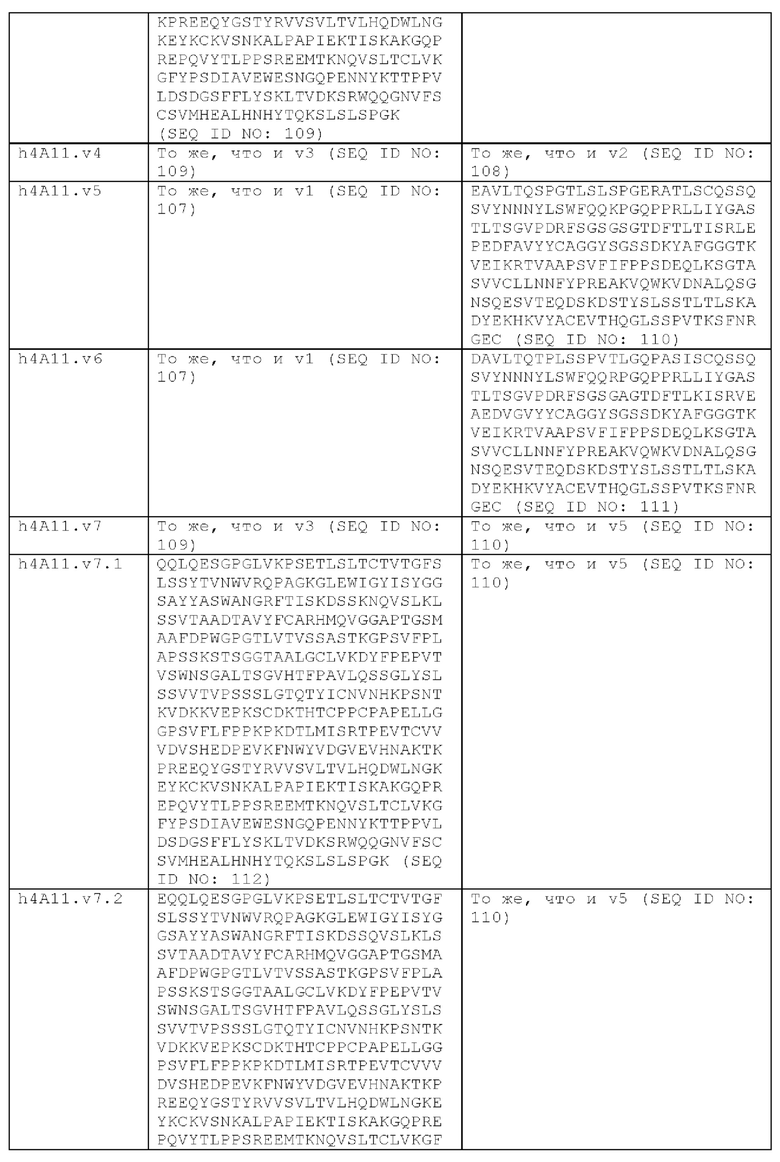
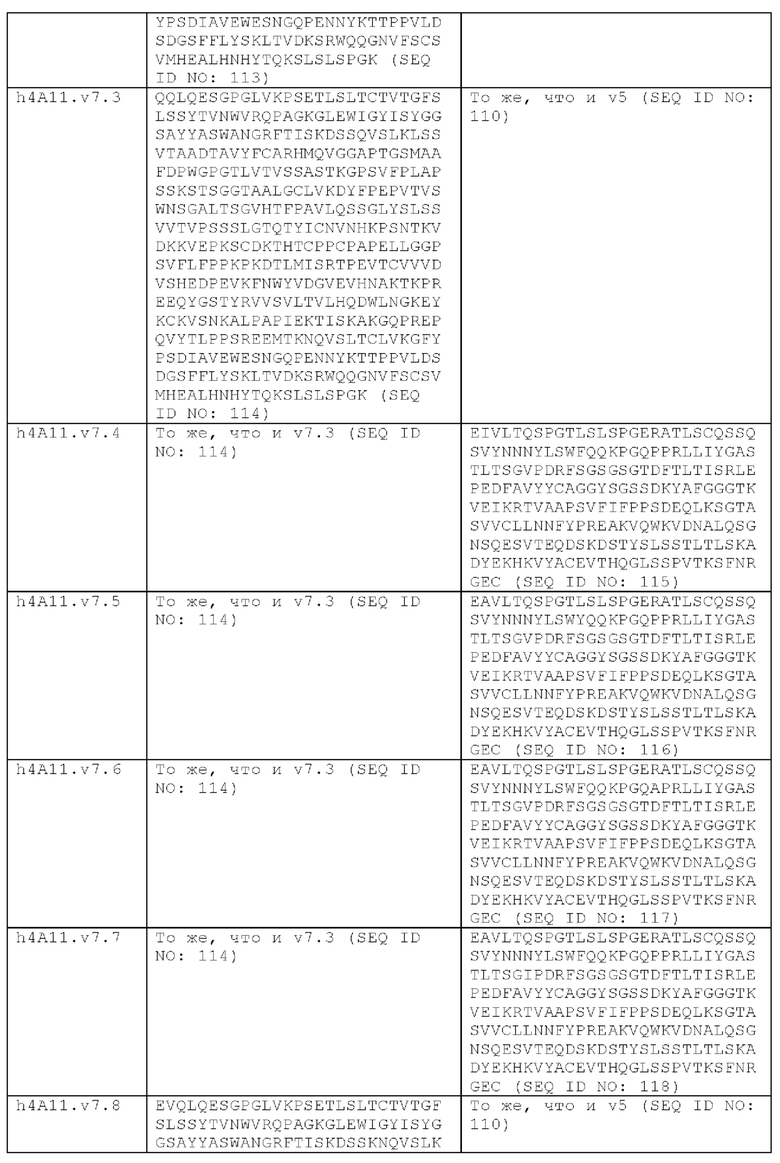




Пример 9: Гуманизация 6F12 (антитела к TGFβ2)
Материалы и методы:
Измерение аффинности антител с помощью BIAcore
Определение аффинности связывания всех гуманизированных антител 4А11 проводили с использованием измерения SPR с помощью прибора BIAcore™-Т200, как описано выше в примере 6 (пример Ab 2А11).
Анализ блокирования TGFβ на основе клеток с применением HEKBlue
Анализ блокирования TGFβ3 на основе клеток с применением HEKBlue проводили, как описано выше в примере 6.
Результаты:
Гуманизация 6F12, шаг 1: Привитие CDR 6F12 крысы в человеческие каркасные области
Моноклональное антитело 6F12 гуманизировали, как описано ниже. Номера остатков представлены согласно Kabat et al., см. выше.
Вариабельные области антитела 6F12 крысы (r6F12) сначала сравнивали по отношению ко всем зародышевым линиями человека. Две человеческие зародышевые линии легкой цепи (IGKV1-5, IGKV3-15) и две человеческие зародышевые линии тяжелой цепи (IGHV2-26, IGHV4-59) идентифицировали в качестве наиболее близких акцепторных каркасных областей для привития легкой цепи и гипервариабельных областей тяжелой цепи (CDR) r6F12 соответственно. В частности, из вариабельного домена (VL) легкой цепи r6F12 положения 24-34 (L1), 50-56 (L2) и 89-97 (L3) прививали в IGKV1-5 и IGKV3-15. Из вариабельного домена тяжелой цепи (VH) r6F12 положения 26-35 (H1), 50-65 (Н2) и 95-102 (Н3) прививали в IGHV2-26 и IGHV4-59 соответственно.
Кроме того, все четыре гуманизированных варианта создавали путем шаффлинга различных акцепторных каркасных областей для привития: гуманизированной версии 1 6F12 (h6F12.v1) посредством IGKV1-5 и IGHV2-26; гуманизированной версии 2 6F12 (h6F12.v2) посредством IGKV1-5 и IGHV4-59; гуманизированной версии 3 6F12 (h6F12.v3) посредством IGKV3-15 и IGHV2-26; гуманизированной версии 4 6F12 (h6F12.v4) посредством IGKV3-15 и IGHV4-59. В дополнение к CDR, привитым на ближайшие акцепторные каркасные области, описанные выше, для каркасной области легкой цепи, h6F12.v1 и h6F12.v2, использовали одну и ту же акцепторную каркасную область IGKV1-5, сохраняли крысиные остатки в положении 43 в каркасной области II и положениях 66, 69, 71 и 87 в каркасной области III VL из r6F12; h6F12.v3 и h6F12.v4 использовали одну и ту же акцепторную каркасную область IGKV3-15, сохраняли крысиные остатки в положении 43 в каркасной области II и в положениях 58, 66, 69, 71 и 87 в каркасной области III VL из r6F12. Для каркасной области тяжелой цепи, h6F12.v1 и h6F12.v3, использовали одну и ту же акцепторную каркасную область IGHV2-26, сохраняли крысиные остатки в положениях 37, 48 и 49 в каркасной области II и в положении 105 в каркасной области IV VH из r6F12; h6F12.v2 и h6F12.v4 использовали одну и ту же акцепторную каркасную область IGHV4-59, сохраняли крысиные остатки в положениях 37 и 48 в каркасной области II, в положениях 67, 71 и 78 в каркасной области III и в положении 105 в каркасной области IV VH из r6F12. Было обнаружено, что все из этих остатков являются частью остатков каркасной области, выступающих в качестве «зоны Верньера», которая может регулировать структуру CDR и точно настраивать соответствие антигена. См., например, Foote and Winter, J. Mol. Biol. 224: 487-499 (1992). Эти определения CDR включают положения, определяемые гипервариабельностью их последовательностей (Wu, Т.Т. & Kabat, Е.А. (1970)), их структурным расположением (Chothia, С.& Lesk, A.M. (1987)) и их участием в контактах антиген-антитело (MacCallum et al. J. Mol. Biol. 262: 732-745 (1996)).
Все 4 гуманизированные варианта 6F12 дополнительно подвергали анализу BIAcore для определения аффинности связывания изоформ TGFβ человека и анализу на основе клеток с применением HEKBlue для оценки эффективности блокирования изоформ TGFβ человека. В анализе BIAcore несмотря на то, что все 4 гуманизированных варианта 6F12 продемонстрировали селективное и сходное связывание с TGFβ2 человека, сопоставимое с исходным r6F12, аффинность связывания была за пределом обнаружения прибора, и поэтому результаты нельзя было использовать для определения того, какой вариант следует продвигать далее. Интересно, что в анализе активности на основе клеток с применением HEKBlue результаты показали снижение блокирования TGFβ2 человека приблизительно в 2,5-5 раза для всех вариантов, за исключением h6F12.v3 (таблица 21).
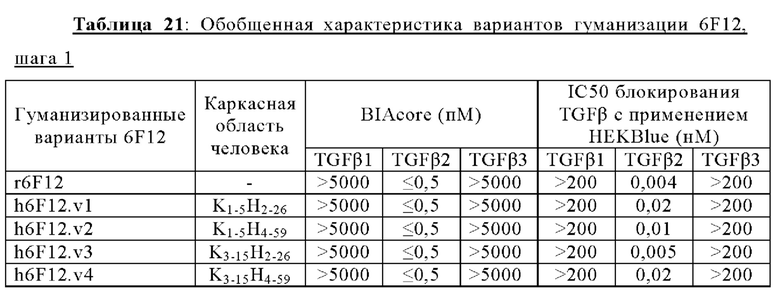
Гуманизация 6F12, шаг 2: завершающая очистка каркасных областей h6F12.v1
При завершающей очистке каркасных областей гуманизированного 6F12 более гуманизированные варианты 6F12 создавали путем замены каждого крысиного остатка h6F12.v1 в зоне Верньера соответствующим человеческим остатком из последовательности зародышевой линии. Каждый вариант оценивали не с помощью BIAcore вследствие ограничений прибора, а с помощью анализа на основе клеток с применением HEKBlue для определения эффективности блокирования изоформ TGFβ человека.
Этими вариантами каркасных областей с завершающей очисткой являются h6F12.v1.1 (VL: S43A), h6F12.v1.2 (VL: E66G), h6F12.v1.3 (VL: P69T), h6F12.v1.4 (VL: Y71F), h6F12.v1.5 (VL: F87Y), h6F12.v1.6 (VH: V37I), h6F12.v1.7 (VH: M48L), h6F12.v1.8 (VH: G49A) и h6F12.v1.9 (VH: P105R).
Большинство вариантов оказались сходными или слегка улучшенными по сравнению с исходным h6F12.v1 в анализе активности, за исключением значительного влияния на h6F12.v1.6 (VH: V37I) и h6F12.v1.9 (VH: P105R) с некоторым снижением блокирования TGFβ2 в приблизительно 5-8 раз. Имели место два варианта с минорным влиянием: h6F12.v1.5 (VL: F87Y) со снижением в 2 раза и h6F12.v1.8 (VH: G49A) со снижением в 4 раза блокирования TGFβ2 соответственно (таблица 22). Таким образом, было установлено, что все крысиные остатки в зоне Верньера h6F12.v1 могут быть заменены остатками зародышевой линии человека, за исключением основного положения 37 в каркасной области II и положения 105 в каркасной области IV VH, и минорного положения 87 в каркасной области III VL и положения 49 в каркасной области II VH.
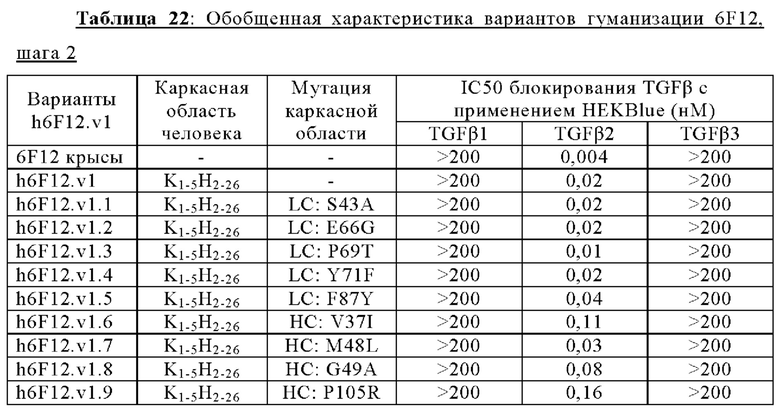
Последовательности вариантов 6F12 представлены в таблице 23 и таблице 24 ниже, а выравнивание последовательностей изображено на Фиг. 22, Фиг. 23 и Фиг. 24.
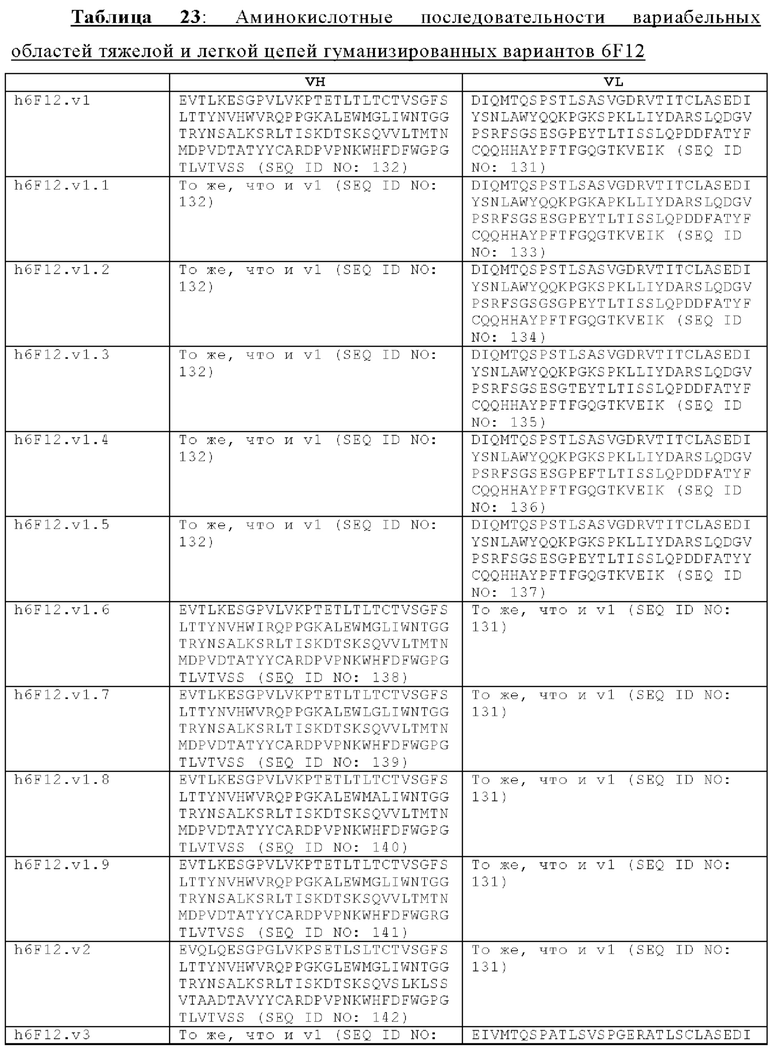

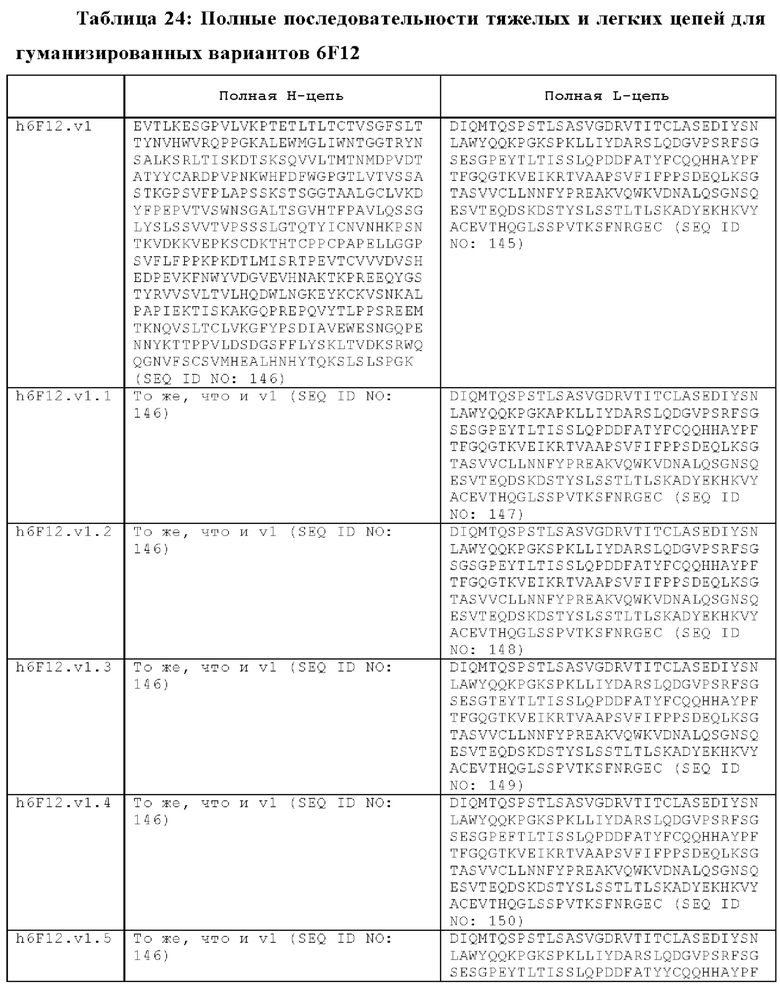


Пример 10: Структурная характеристика селективных в отношении изоформы антител к TGFβ
Материалы и методы
Fab экспрессировали из клеток СНО. TGFβ2 и TGFβ3 человека приобретали в Peprotech (Роки-Хилл, Нью-Джерси). Fab и TGFβ2/3 смешивали в соотношении 1:1,5 на льду в течение 30 минут, комплексы выделяли из колонки HiLoad Superdex S200 16/600 (GE Life Health Science) в 25 мМ Tris, рН 7,5, и 50 мМ NaCl. Образцы концентрировали до 10 мг/мл для кристаллизации. Для выращивания кристаллов использовали метод диффузии паров. 1 мкл белка смешивали с 1 мкл маточной жидкости на 500 мкл луночного раствора. 2А10 TGFβ3 кристаллизовали в 25% ПЭГ 1000 и 0,1 М Hepes, рН 7,5. Комплекс 4A11v2/TGFβ2 кристаллизовали в 15% ПЭГ 4000 и 0,1 М какодилата натрия, рН 6. Комплекс 4A11v7 и TGFβ2 кристаллизовали в 30% ПЭГ 1000 и 0,1 М Tris, рН 8,5. Осуществляли криопротекцию кристаллов 20% глицерином и замораживали в жидком азоте перед сбором данных.
Результаты:
2А10/β3
Кристаллическую структуру Fab 2A10v4 (экспрессируемого в СНО) и зрелой димерной формы TGFβ3 человека (A301-S412, экспрессируемой в Е. coli, приобретенной в Peprotech (Роки-Хилл, Нью-Джерси)) определяли при 2,5 Å с уточненной статистикой R/Rfree 22,6% и 27,1% соответственно (таблица 25).

Структура на Фиг. 25 показывает, что каждый димер TGFβ3 связан двумя Fab 2А10. 2А10 связывается со шпилечной областью бета6/бета7 TGFβ3. Сравнение структуры 2A10/TGFβ3 со структурой TGFβ1, связанного с TGFBR1 и TGFBR2 (Фиг. 26) (см. также Radaev JBC 2011, PDB 3KFD), показывает, что Fab 2А10 стерически блокирует способность TGFBR2, но не TGFBR1, связывать TGFβ3. Поскольку известно, что сигнальный комплекс TGFβ с TGFBR1 и TGFBR2 собирается совместно, сначала путем связывания TGFβ с TGFBR2, а затем рекрутинга TGFBR1, ожидается, что блокирования связывания TGFBR2 достаточно, чтобы полностью предупредить связывание сигнальных рецепторов R1/R2 с TGFβ3. Способ связывания 2А10 отличается от антитела фрезолимумаба ко всем видам TGFβ, которое связывается со всеми изоформами TGFβ посредством эпитопа, который блокирует связывание как TGFBR1, так и TGFBR2 (PDB ID 4KV5, 4KXZ, 3EO1, 3KFD).
Эпитоп на TGFβ3, связанный 2А10, изображен на Фиг. 28 и Фиг. 29 и обобщен в перечне контактов, встречающихся в таблице 26. Остатки TGFB3, контактирующие с 2А10 (в пределах 5,0 Å), представляют собой R325, K331, W332, Н334, Е335, Т387, 1388, L389, Y391, V392, G393, R394, Р396, K397 и V398. Как видно из Фиг. 28, основная часть эпитопа представлена нитями бета6 и бета7 TGFβ3, включая шпилечную бета-область. Эта бета-шпилька содержит R394, который образует наибольшее количество контактов с 2А10 (см. таблицу 26), а также образует ионный солевой мостик с Asp50 в CDR2 тяжелой цепи 2А10. Эпитоп TGFβ3 изображен под другим углом на Фиг. 29, на которой лучше видны контакты между 2А10 и петлей 325-335 в TGFβ3.
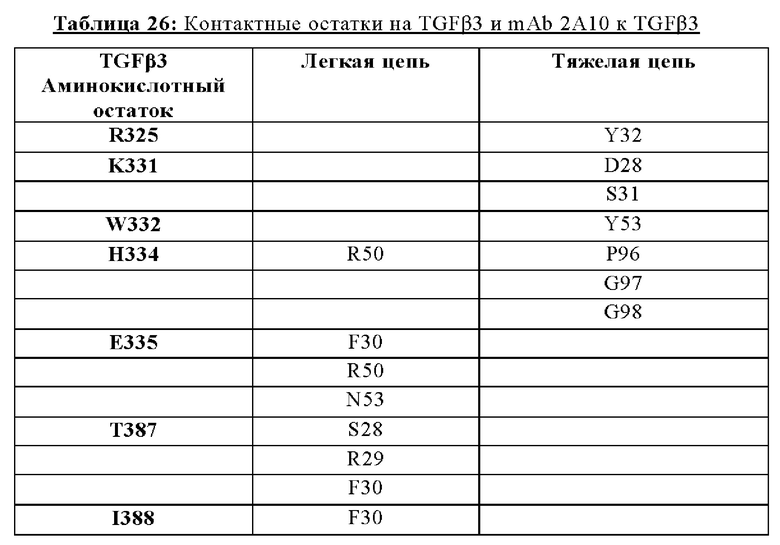
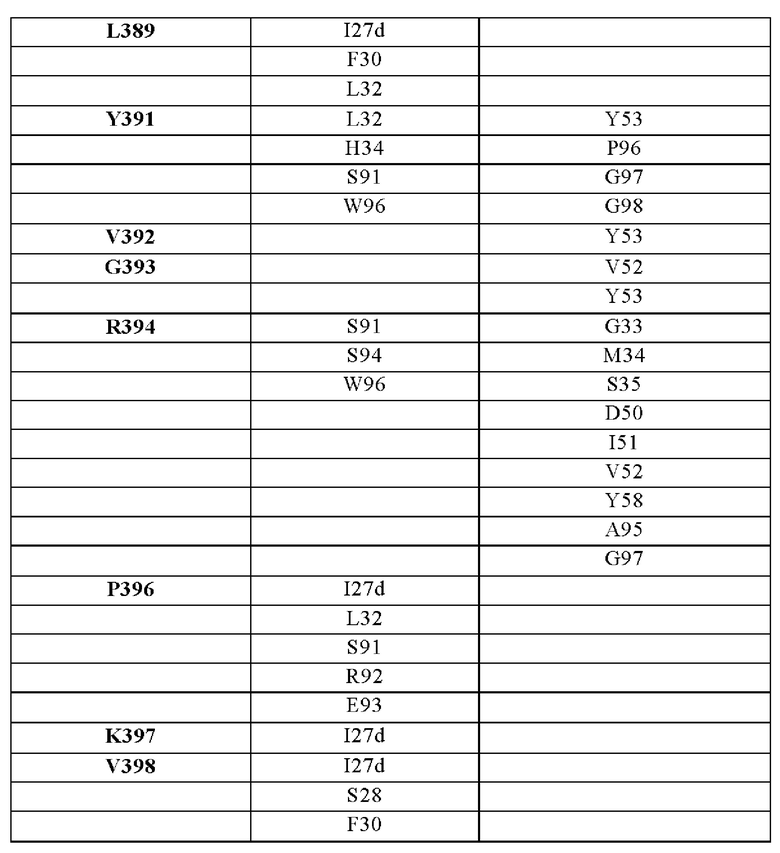
Паратоп 2А10 изображен на Фиг. 30. Как тяжелая, так и легкая цепи обеспечивают относительно одинаковую площадь поверхности для связывания TGFβ3. ~885 Å2 скрытой поверхности (BSA) приходится на легкую цепь, а ~805 Å2 BSA приходится на тяжелую цепь, всего ~1690 Å2 на границе раздела между 2А10 и TGFβ3 (среднее значение двух Fab, связанных с димером в структуре). Учитывая вогнутую поверхность паратопа антитела в комбинации с удлиненным эпитопом бета6/бета7 на TGFβ3, все шесть CDR контактируют с TGFβ3 (Фиг. 30 и таблица 26). TGFβ3 Arg394, который осуществляет основной контакт с Asp50 в тяжелой цепи 2А10, почти полностью погружен в центр паратопа антитела, контактируя с CDR Н3 легкой цепи 2А10 и со всеми тремя CDR тяжелой цепи. Удлиненная CDR L1 легкой цепи из 15 остатков вносит основной вклад во взаимодействие с цепями бета6/бета7, а короткая богатая глицином CDR Н3 способствует созданию пространства для шпильки бета6/бета7, чтобы приблизиться и взаимодействовать с CDR Н2 D50. Эти уникальные взаимодействия помогают объяснить тот факт, что BSA легкой цепи больше, чем BSA тяжелой цепи, что несколько необычно для взаимодействий антитело-антиген.
Селективность в отношении TGFβ3 по сравнению с TGFβ1 и TGFβ2, вероятно, достигается разными путями для каждой из этих изоформ. Что касается TGFβ1 человека, между TGFβ1 и TGFβ3 имеется три различия в аминокислотах вблизи эпитопа 2А10 или внутри него. Эти остатки, Т387Р (что означает, что остаток представляет собой «Т» в TGFβ3 (нумерация TGFβ3 человека) и Р в TGFβ1 (Р365 в TGFβ1)), L389V и T395K позволяют предположить, что остатки вдоль цепей бета6 (Т387Р, L389V) играют большую роль в связывании 2А10 (Фиг. 31). В TGFβ3 Т395 не вступает в прямое взаимодействие с 2А10 в кристаллической структуре, поэтому эффект замены лизина в этом остатке неясен.
Для TGFβ2 имеется четыре замены аминокислот между TGFβ2 и TGFβ3, которые лежат внутри или рядом с эпитопом 2А10. Из этих четырех остатков (R325K (что означает остаток «R» в TGFβ3 и K в TGFβ2 (нумерация TGFβ3 человека), V392I, R394K, V398I) три могут нарушать продуктивный контакт с 2А10 (Фиг. 32). TGFβ3 R325 образует водородную связь с остатком тяжелой цепи Y32, которая будет изменена при замене на лизин. Замена изолейцина в положении V398 может вызвать стерическое столкновение с 2А10. Что наиболее важно, замена остатка R394, находящегося в сильном контакте (на лизин в TGFβ2), вероятно, также изменит химию взаимодействия с 2А10 неблагоприятным образом. Не ожидается, что замена изолейцином валина в положении 392 окажет значительное влияние на связывание, учитывая его положение в эпитопе и консервативный характер замены. В целом, структура 2А10, связанного с TGFβ3, дает четкое представление о том, как 2А10 достигает высокой селективности между изоформами TGFβ.
4A11/TGFβ2
Чтобы лучше понять причины функциональных различий между гуманизированными клонами 4А11, структуры двух версий 4А11, v2 и v7, связанных со зрелой формой TGFβ2 человека, расшифровывали с использованием способов, описанных выше в отношении 2А10. V2 и v7 гуманизировали до каркасных областей VL1VH3 и VL3VH4 соответственно. На Фиг. 33 изображены две структуры. 4A11v2/TGFβ2 определяли при 2,9 Å с помощью статистики R/Rfree 23/30%, в то время как 4A11v7/TGFβ2 определяли при 1,91 Å с помощью статистики R/Rfree 20,5/24,4% (таблица 27). Каждый димер TGFβ2 связан двумя Fab 4А11, которые имеют почти идентичный эпитоп. В то время как общая архитектура комплексов была очень похожей, 4A11v7 кристаллизовалось в другой пространственной группе, и имели место некоторые небольшие различия в углах изгиба для второго Fab, связанного с TGFβ2.
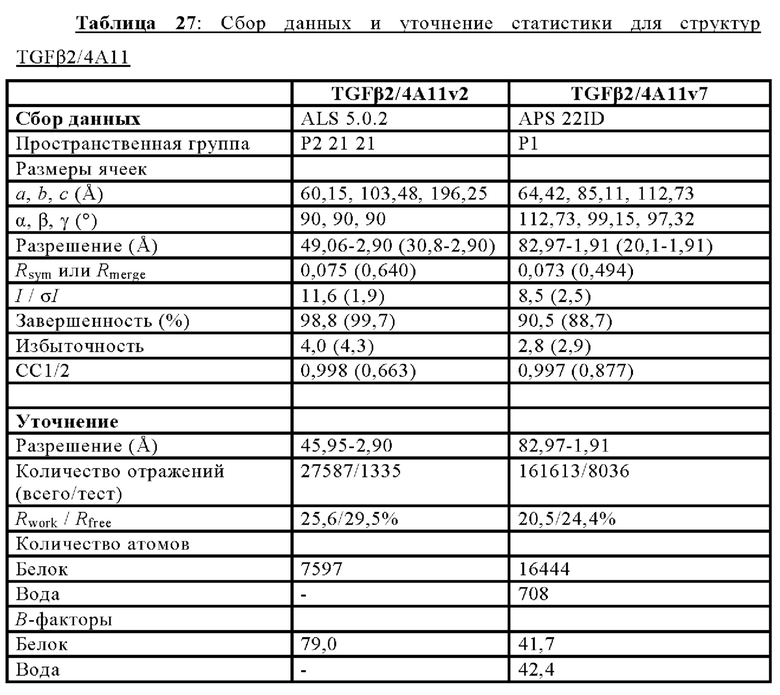
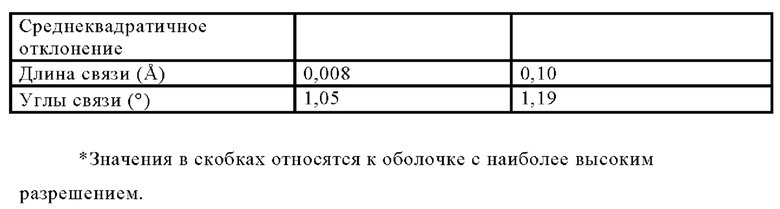
Эпитоп для 4А11 находится на нижней стороне димера TGFβ2, при этом два Fab-фрагмента связаны с двумя идентичными эпитопами, как и ожидалось, для симметричного гомодимера (Фиг. 34). Эпитоп для 4А11 включает остатки TGFβ2 V313, Q314, D315, R320, L322, Y323, R328, D329, F345 и А347 в одном мономере TGFβ2, и N368, Т369, I370, N371, Р372, Е373, А374, S375, А376. и S377 во втором мономере TGFβ2 (Фиг. 35 и таблицы 28 и 29). На структуру 4A11v2/TGFβ2 приходится ~1760 Å2 скрытой площади поверхности, а на структуру 4A11v7/TGFβ2 приходится ~1950 Å2 скрытой площади поверхности. Эпитоп 4А11 находится в центре верхушки альфа-спирали (остатки 359-372). Внутри эпитопа наблюдается значительная консервативность последовательности между TGFβ1/2/3, но одно основное различие, обнаруженное в TGFβ1, но не в TGFβ2/3, заключается в замене глутаминовой кислоты на глицин в положении 373 (нумерация TGFβ2 человека) (Фиг. 35). Этот остаток глутаминовой кислоты находится в центре эпитопа 4А11 и образует полярный контакт с Т33 CDR-H1. Замена глутаминовой кислоты на глицин разрушает это полярное взаимодействие, а также удаляет большую часть площади поверхности, введенной боковой цепью.
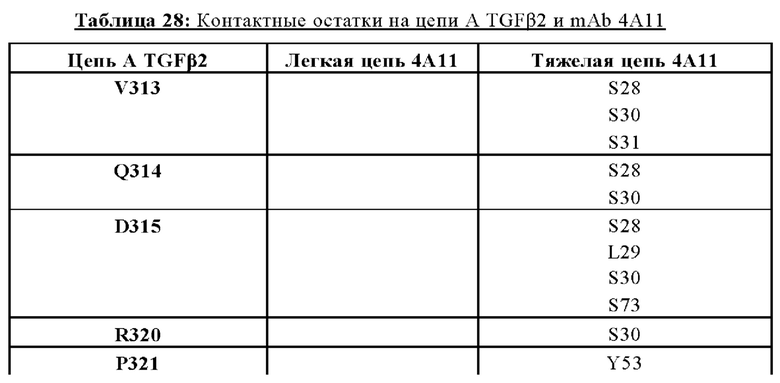
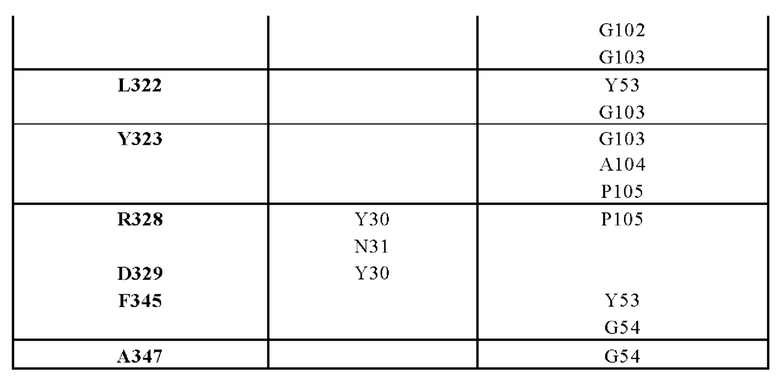
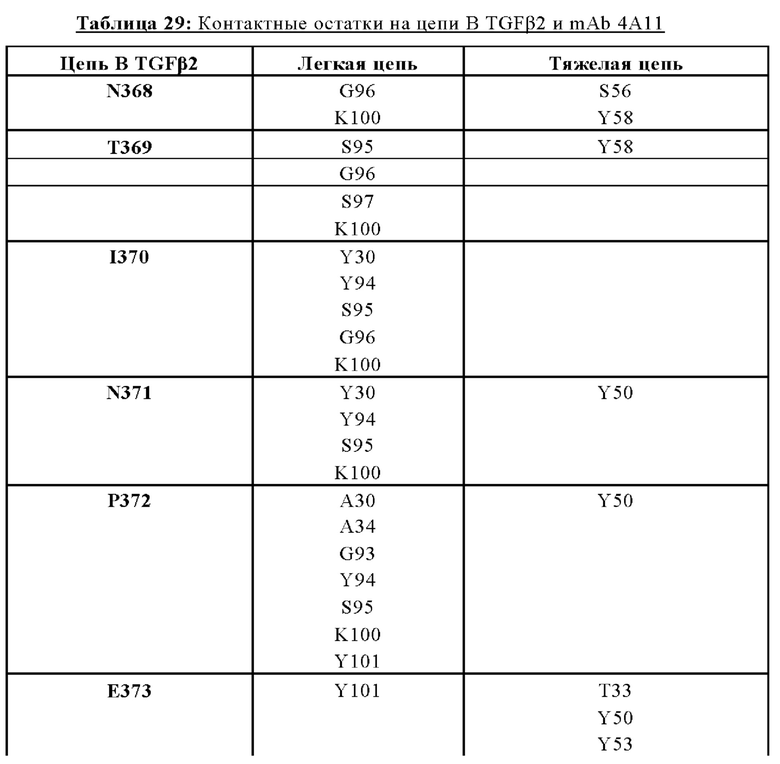
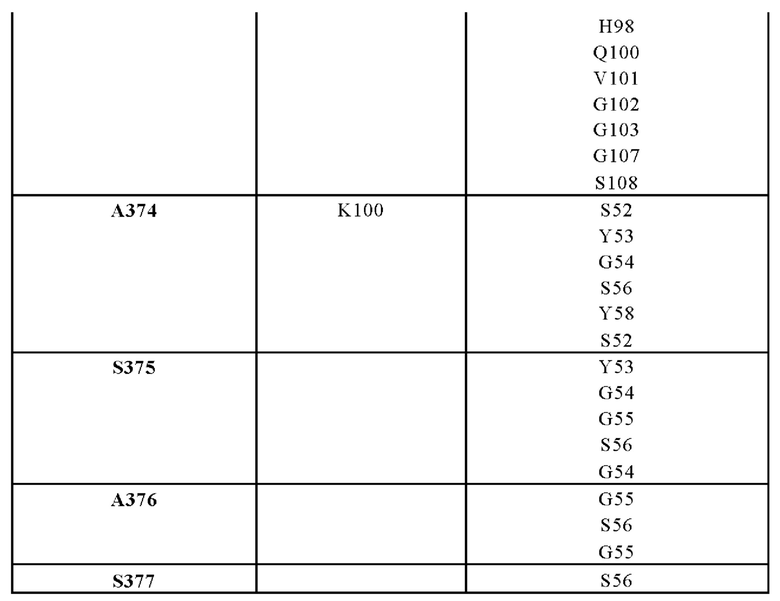
Как показано на Фиг. 34, эпитоп 4А11 находится вдали от сайтов связывания TGFBR1/TGFBR2, и ни одна молекула 4А11 не конкурирует стерически за связывание с рецептором. Вместо этого Fab 4А11, по-видимому, индуцируют конформационные изменения в TGFβ2, вызывая «сжатие» двух мономеров друг по отношению к другу на несколько градусов (Фиг. 36). Сравнение связанного с 4А11 TGFβ2 с TGFβ3, связанного с TGFBR1 и TGFBR2, предполагает, что рецептор-связывающий сайт по меньшей мере для одного из мономеров TGFβ2 нарушен этим конформационным изменением (Фиг. 37). Эти изменения, проявляющиеся в кристаллической структуре, указывают на то, что 4А11 использует аллостерический механизм для ингибирования передачи сигналов TGFβ2/3 путем конформационного нарушения одного или обоих рецептор-связывающих сайтов. Важно отметить, что двухвалентное связывание молекулы 4А11 на основе IgG необходимо для индукции этого конформационного изменения в TGFβ2/3 и, следовательно, для ингибирования передачи сигналов TGFβ2/3. В отличие от блокирующего рецептор антитела, где блокирование одного рецептор-связывающего сайта R1/R2 на димере TGFβ может блокировать передачу сигнала, маловероятно, что связывание одного плеча 4А11 будет достаточным для ингибирования передачи сигнала. Учитывая расположение эпитопов, двухвалентное связывание полной молекулы 4А11 на основе IgG может также стерически предупреждать приближение молекулы TGFβ2/3, связанной с IgG, достаточно близко к мембране к любому из рецепторов TGFB.
Во время процесса гуманизации 4А11 было отмечено, что ряд последовательностей каркаса антитела вызывали разрыв между аффинностью антитела и IC50 в анализе на основе клеток. Например, 4A11v2 имел пикомолярную аффинность в отношении TGFβ2, но утрачивал всю активность в клеточных анализах, в то время как 4A11v7 имел аффинность in vitro, подобную v2, но сохранял очень высокую активность в клеточных анализах. Как упоминалось ранее, V2 и v7 гуманизировали до каркасных областей VL1VH3 и VL3VH4 соответственно, которые имеют значительные различия в последовательностях. Поэтому было более тщательно исследовано, как остатки каркаса могут влиять на способность 4А11 образовывать двухвалентный комплекс с димером TGFβ2 (Фиг. 38).
Структуры 4A11v2 и v7 с TGFβ2 показывают, что каркасы тяжелых цепей располагаются близко друг к другу в структуре и действительно взаимодействуют, образуя поверхность раздела. Таким образом, остатки в этих каркасных областях могут влиять на способность молекулы 4А11 на основе IgG образовывать двухвалентный комплекс. Тщательное изучение этой поверхности раздела каркасной области НС/НС позволяет предположить, что остатки каркасной области v2 представляют собой менее допустимую поверхность раздела, чем поверхность раздела для v7 (Фиг. 39). Например, в положении 19 НС представлен аргинин в v2 и серии в v7, а в положении 81 НС представлен глутамин в v2 и лизин в v7. В v2 R19 и Q81 сближаются, образуя то, что выглядит как энтропийно неблагоприятный контакт без стабилизирующего полярного взаимодействия. В v7 остаток S19 освобождает место для K81, чтобы протянуться и образовать Н-связь с Ser79. Кроме того, v2 имеет два крупных объемных остатка (Y79 и N82a), не образующих явных продуктивных контактов. Важно отметить, что эти положения являются меньшими остатками в v7 (S79 и S82a), что, вероятно, позволяет поверхности раздела каркасной области НС объединяться более допустимым образом. Хотя эти контакты каркасной области НС в v7, по-видимому, не являются особенно комплементарными, они, по-видимому, более допустимы (менее неблагоприятны), чем контакты каркасной области между двумя тяжелыми цепями v2. Эти взаимодействия кажутся неблагоприятными как вследствие стерической объемности, так и вследствие потенциального энтропийного штрафа за скрытие больших гибких боковых цепей.
Дальнейшее подтверждение приведенного выше анализа, расчет скрытой площади поверхности для взаимодействий каркасных областей НС/НС в структурах v2 и v7 предполагает, что контакт v7 НС/НС является более комплементарным. В структуре 4A11v2/TGFβ2 поверхность раздела НС/НС (цепь D и цепь F) покрывает приблизительно 660 Å2 площади поверхности. Для структуры 4A11v7/TGFβ2 имеется два комплекса 2:2 в асимметричной ячейке кристалла, а между двумя наборами взаимодействий НС/НС каркасной области 1050 Å2 (цепь D/F) и 730 Å2 (цепь В/Н) скрыты. Большая разница здесь между двумя комплексами 2:2, вероятно, вызвана артефактами упаковки кристаллов, но общая тенденция предполагает, что поверхность раздела v7 НС/НС больше, что предполагает более комплементарную контактную область. Также важно отметить, что в контексте эксперимента по кристаллизации концентрации белка очень высоки (>10 мг/мл), и при таких концентрациях можно преодолеть отталкивающие эффекты некомплементарных взаимодействий каркасных областей НС/НС, и образовывался двухвалентный комплекс. В контексте эксперимента на основе раствора с более низкой концентрацией (например, анализ на основе клеток или in vivo) отталкивающие эффекты, вызванные каркасной областью, могут привести к тому, что одновалентное связывание будет предпочтительнее двухвалентного связывания.
Последствия этих различий каркасных областей важны для объяснения различий в клеточной активности 4A11v2 по сравнению с v7 (Фиг. 40). Если при низкой концентрации v2 не может связываться с димером TGFβ2/3 двухвалентной связью, это антитело не будет индуцировать необходимые конформационные изменения в TGFβ, чтобы заблокировать второй сайт рецептора и ингибировать передачу сигнала. Для v7 контакты каркасной области допускают двухвалентное связывание с димером TGFβ2/3, и, следовательно, это антитело может индуцировать конформационные изменения, которые блокируют образование тетрамера рецептора, и, следовательно, может ингибировать передачу сигнала. Все эти данные указывают на новый механизм аллостерического ингибирования TGFβ2/3 антителом 4А11, который не основан на стерическом блокировании (интерференции) рецептора, а вместо этого требует двухвалентного связывания с димером TGFβ для блокирования передачи сигнала.
Пример 11: Сравнение безопасности с антителом 1D11 ко всем видам TGFβ
В этом примере mAb 2А10, антитело, имеющее вариабельную область 2A10.v4 с Fc-областью мышиного IgG1, или антитело 1D11 ко всем видам TGFβ вводят три раза в неделю (TIW) в течение 4 недель (всего 12 доз) мышам CD-1 в дозах 0 (контроль), 10 и 50 мг/кг I.P. TIW и 50 мг/кг I.V. TIW в несущей среде, состоящей из фосфатно-солевого буфера. Самцов и самок относили к группам токсичности при всех уровнях доз и планировали их вскрытие в конце периода введения дозы (день 29). Кровь для токсикокинетической оценки собирали у дополнительных мышей в контрольной группе и группе, обработанной mAb 2A10/2A10v4 (n=9 самцов/группа). Критерии оценки включают следующие параметры: клинические наблюдения, вес тела, клиническую патологию (гематологию и клиническую биохимию), анатомическую патологию и токсикокинетику.
Определяют воздействия AUC, и ожидается, что они будут увеличиваться примерно пропорционально дозе при увеличении дозы от 10 до 50 мг/кг.
Не ожидается никаких клинических наблюдений, связанных с 2А10 или 2A10.v4, влияния на вес тела или изменений клинической патологии, в то время как такие связанные клинические наблюдения, влияние на массу тела или изменения клинической патологии, указывающие на повышенную токсичность, ожидаются для мышей, обработанных антителом 1D11, например, гистологические поражения, потеря массы, неопухолевая кистозная гиперплазия эпителия и воспаление языка, а также дисплазия зубов и гиперплазия эпителия десен и пищевода, как описано в Lonning et al. (2011) Current Pharmaceutical Biotechnology, 12, 2176-2189.
Пример 12: Сравнение безопасности с селективными антителами к TGFβ1
В этом примере каждое из mAb 2А10, антитела, имеющего вариабельную область 2A10.v4, слитую с Fc-областью IgG1 мыши, вариабельную область антитела CAT-192 к TGFβ1 (метелимумаба), слитую с Fc IgG1 мыши, вводят три раза в неделю (TIW) в течение 4 недель (всего 12 доз) мышам CD-1 в дозах 0 (контроль), 10 и 50 мг/кг I.P. TIW и 50 мг/кг I.V. TIW в несущей среде, состоящей из фосфатно-солевого буфера. Самцов и самок относили к группам токсичности при всех уровнях доз и планировали их вскрытие в конце периода введения дозы (день 29). Кровь для токсикокинетической оценки собирали у дополнительных мышей в контрольной группе и группе, обработанной mAb 2A10/2A10v4 (n=9 самцов/группа). Критерии оценки включают следующие параметры: клинические наблюдения, вес тела, клиническую патологию (гематологию и клиническую биохимию), анатомическую патологию и токсикокинетику.
Определяют воздействия AUC, и ожидается, что они будут увеличиваться примерно пропорционально дозе при увеличении дозы от 10 до 50 мг/кг.
Никаких клинических наблюдений, связанных с 2А10 или 2A10.v4, влияния на вес тела или изменений клинической патологии не ожидается, в то время как такие связанные клинические наблюдения, влияние на вес тела или изменения клинической патологии, указывающие на повышенную токсичность, ожидаются для мышей, получавших CAT-192, например, как описано в Denton A&R 56: 323 (2007).
Пример 13: Кондиционированная среда с полноразмерным TGFβ3 ингибирует образование альвеолосфер in vitro
Материалы и методы
Выделение клеток легких
Клетки легких выделяли в соответствии с ранее опубликованным протоколом (см. Rock et al., 2011 Proceedings of the National Academy of Sciences of the United States of America 108, E1475-1483). Ткани разрушали и собирали отдельные клетки после того, как легкие надували и расщепляли смесью растворов протеаз (5 ед./мл диспазы, 450 ЕД/мл коллагеназы типа I, 4 ЕД/мл эластазы и 0,33 ЕД/мл ДНКазы I в DMEM/F12) в течение 45 минут при 37°С при частом встряхивании. Затем клетки промывали с помощью DMEM с 10% фетальной бычьей сывороткой (FBS, Invitrogen) и ресуспендировали в лизисном буфере ACK в течение 3 минут для лизиса эритроцитов. После промывания клетки блокировали очищенным крысиным антителом к CD16/CD32 мыши (кат. №553142; BD Biosciences) и окрашивали антителами к CD45 (кат. №17-0451-82, eBioscience), ЕрСАМ (кат. №11-5791-82, eBioscience). Клетки АЕС2 сортировали на клеточном сортере BD FACSAria Fusion (BD Biosciences) на основе следующих маркеров: CD45-; ЕрСАМ+; и tdTomato+. Данные анализировали с помощью программного обеспечения Flowjo.
3-Мерная культура органоидов АЕС
Культуру органоидов АЕС поддерживали в соответствии с ранее опубликованным протоколом с незначительными изменениями (см. Barkauskas, 2013 JCI 123, 3025-3036; Sun Т, et al, 2019, JCI Insight; Jun 18; 5 (14): e128674). Отсортированные клетки AEC2 смешивали со стромальными клетками человека NHLF (кат. № СС-2512; Lonza Walkersville Inc. USA) в соотношении 1:10. Клетки ресуспендировали в предварительно охлажденной смешанной среде 1:1 MTEC/Plus и Matrigel с пониженным содержанием фактора роста (кат. №356231; BD Biosciences). 104 клеток АЕС2 и 105 клеток NHLF помещали в 90 мкл смешанной среды в 24-луночный 0,4-мкм вкладыш Transwell (кат .№3470; Costar). 500 мкл среды MTEC/Plus добавляли в нижнюю камеру. Среду меняли через день. Органоиды оценивали через 12-14 дней культивирования с помощью инвертированного флуоресцентного микроскопа.
Микроскопия
Все изображения культуры органоидов получали на инвертированном микроскопе Nikon Ti-E Perfect Focus. Крупные изображения создавали путем автоматического сшивания нескольких смежных кадров из многоточечного сбора данных с использованием моторизованного предметного столика. Изображения анализировали с помощью программного обеспечения ImageJ.
Скрининг библиотеки Secretome
Библиотека Secretome представляет собой набор плазмид, экспрессирующих индивидуальную кДНК человека на векторах экспрессии млекопитающих, содержащих ~1700 отдельных полноразмерных генов, кодирующих секретируемые белки. Для идентификации факторов, которые могут быть вовлечены в биологию АЕС в системе альвеолярных органоидов, библиотеку сначала разделяли на 96 подбиблиотек, каждая из которых содержала около 10-20 отдельных плазмид. Каждый пул плазмид подбиблиотеки временно трансфицировали в клетки 293Т с использованием липофектамина 2000 (кат. №11668019, ThermoFisher) в соответствии с рекомендациями производителя. Кондиционированные среды собирали через два дня и хранили при 4°С до использования в культуре альвеолосфер. Положительные подбиблиотеки идентифицировали в конце культивирования. Во время второго раунда скрининга создавали и тестировали кондиционированные среды из отдельных плазмид в каждой положительной подбиблиотеке. В конечном итоге идентифицировали положительные отдельные гены, ответственные за фенотипы культур органоидов.
Результаты
Альвеолярный эпителий легких поддерживается и восстанавливается за счет пролиферации и гиперплазии альвеолярных эпителиальных клеток типа 2 (АЕС2) с последующей дифференцировкой в альвеолярные эпителиальные клетки типа 1 (АЕС1). Для исследования биологии АЕС использовали 3-мерную систему культивирования альвеолосферных органоидов, в которой АЕС2 с картами зачатков выделяли от мышей Sftpc-CreERT2; Rosa26R-tdTm после индукции тамоксифеном и кокультивировали с фибробластами легких в матригеле. Через -14 дней формируются сфероиды, происходящие из отдельных АЕС2, с внешним слоем АЕС2 и внутренним просветом с АЕС1, которые дифференцировались от предшественников АЕС2. Эта систему ранее использовали для идентификации TAZ как регулятора дифференцировки АЕС2-АЕС1 во время восстановления альвеол (см. Sun, Т. et al. (2019). JCI Insight; Jun 18; 5 (14): e128674). Для выявления потенциальных секретируемых факторов, влияющих на пролиферацию альвеолярных эпителиальных клеток, проводили скрининг с использованием библиотеки экспрессионных плазмид, содержащей примерно 1700 отдельных генов человека, кодирующих секретируемые белки. Этой плазмидной библиотекой временно трансфицировали клетки 293Т, сначала в пулах (10-20 отдельных плазмид/пул), а затем по отдельности, через два дня собирали кондиционированные среды и добавляли кондиционированные среды к культурам альвеолосфер. Кондиционированные среды из клеток, трансфицированных конструкцией, кодирующей полноразмерный TGFβ3, полностью ингибировали образование альвеолосфер. В отличие от этого, кондиционированные среды из клеток, трансфицированных полноразмерным TGFβ1, не оказывали никакого эффекта, несмотря на использование тех же условий трансфекции и векторов экспрессии. Предыдущие исследования показали, что рецептор-связывающий домен («зрелая» форма) TGFβ1 может способствовать апоптозу первичных альвеолярных эпителиальных клеток типа 2 (АЕС2) in vitro и in vivo. Действительно, рекомбинантные зрелые формы TGFβ1 или TGFβ3 ингибировали образование альвеолосфер в этих экспериментах. Таким образом, в используемых условиях культивирования полноразмерный LAP-ассоциированный TGFβ1 неактивен, но полноразмерный LAP-ассоциированный TGFβ3 активен, что позволяет предположить, что «латентный» TGFβ3 имеет внутреннюю активность и/или может быть активирован с помощью уникального механизма, представленного в данной культуральной системе, отличной от описанных для TGFβ1.
Пример 14: Полноразмерные TGFβ2 и TGFβ3 могут активировать TGFβR-зависимую передачу сигналов, независимую от связывания интегрина in vitro
Поскольку связывание интегрина является установленным механизмом активации TGFβ1, а дифференциальная активность TGFβ1 и TGFβ3 наблюдали в альвеолосферной модели, затем предпринимали попытку определить зависимость активации TGFβ3 от интегрина. Использовали 2-клеточную репортерную систему in vitro, в которой векторы экспрессии, кодирующие полноразмерный TGFβ1 или TGFβ3, котрансфицировали в клетки 293Т с векторами экспрессии, кодирующими интегрины αv и β6, и кокультивировали с репортерной клеточной линией, управляющей экспрессией люциферазы ниже от SMAD-зависимого промоторного элемента («MLEC») (см. Abe et al., 1994; Anal Biochem, 216, 276-284). В то время как полноразмерный TGFβ1 требовал коэкспрессии интегрина αvβ6 для индукции репортерной активности, было обнаружено, что полноразмерный TGFβ3 надежно индуцирует репортерную активность без котрансфекции конструкций интегрина αv и β6. Коэкспрессия αvβ6 дополнительно повышала полноразмерную TGFβ3-зависимую репортерную активность, и это дальнейшее повышение ослаблялось мутацией интегрин-связывающего мотива RGD в LAP TGFβ3 в RGE (Фиг. 41).
В соответствии с наблюдаемыми эффектами эксперимента с альвеолосферами, описанными выше, супернатанты клеток, трансфицированных полноразмерным TGFβ3, но не TGFβ1, значительно индуцировали репортерную активность (Фиг. 42). Сходную репортерную индукцию наблюдали с супернатантами клеток, трансфицированных полноразмерным TGFβ2; поскольку в TGFβ2 отсутствует мотив RGD (Фиг. 1), его активация, вероятно, не зависит от интегрина (Фиг. 42). Общая активность TGFβ в этих трех супернатантах была сопоставима после стадии подкисления, которая высвобождает рецептор-связывающий домен TGFβ из его латентного комплекса (Фиг. 42). Для исключения возможности того, что эти эффекты были специфичны для используемых нами репортерных клеточных линий, клетки HEK-Blue (InvivoGen) и SCOS-7 оценивали как альтернативные репортерные линии TGFβ и реципиентные клеточные линии соответственно. В обоих случаях наблюдали высокую «внутреннюю» активность TGFβ2 и/или TGFβ3 (Фиг. 43А и Фиг. 43Б).
Напряжение цитоскелета необходимо для передачи сил во время опосредованной интегрином αvβ6 активации латентного TGFβ1. Для исключения возможного участия других зависимых от цитоскелета активностей интегринов, эту культуральную систему обрабатывали цитохалазином D для ингибирования полимеризации актина, что уменьшало опосредованную интегрином αvβ6 активацию TGFβ1 и TGFβ3. Однако «внутренняя» активность TGFβ3 оставалась неизменной (Фиг. 44), что свидетельствует о том, что эта активность не зависит от интегрина.
В аппарате Гольджи пропептид TGFβ1 (про-TGFβ1) протеолитически расщепляется фурином, что приводит к образованию SLC. Это протеолитическое расщепление необходимо для создания функционального TGFβ1. Все три изоформы TGFβ имеют консервативные сайты расщепления фурином (RKKR или RHRR, Фиг. 1). Для оценки потребности в этом расщеплении протеазой для активности TGFβ2/3 в нашей системе, сайты расщепления фурином в каждой изоформе TGFβ мутировали. Мутированные полноразмерные TGFβ2 и β3 полностью утратили свою внутреннюю активность (Фиг. 45). Таким образом, подобно TGFβ1, расщепление фурином необходимо для активности TGFβ2/3, несмотря на различные внеклеточные механизмы активации SLC. Для оценки кинетики «внутренней» активности TGFβ2 и TGFβ3, фосфо-SMAD2 оценивали в нормальных фибробластах легких человека (NHLF) после обработки супернатантами для трансфекции, и наблюдали кинетику, сходную с кинетикой клеток, обработанных рекомбинантными рецептор-связывающими доменами TGFβ, с пиком pSMAD2 через приблизительно 1 ч после стимуляции (Фиг. 46).
Для исключения возможности того, что другие факторы, присутствующие в кондиционированных средах трансфицированных клеток, могут способствовать активации TGFβ2 и TGFβ3, создавали рекомбинантные полноразмерные изоформы TGFβ. Для улучшения экспрессии и стабильности, а также для облегчения очистки выполняли N-концевые слияния в рамке считывания константного домена IgG1 человека («Fc») с каждой изоформой (Fc-FL-TGFβ1, 2 и 3). В соответствии с эффектами, наблюдаемыми в кондиционированных средах, очищенный рекомбинантный Fc-FL-TGFβ1 не активировал репортерную активность в MLEC. Однако Fc-FL-TGFβ2 и Fc-FL-TGFβ3 устойчиво индуцировали репортерную активность (Фиг. 47), которая ингибировалась их соответствующими специфичными в отношении изоформы антителами (Фиг. 48).
Предыдущие исследования показали, что рекомбинантный LAP из TGFβ1 (LAP1) может значительно ингибировать рецептор-связывающие домены TGFβ1, 2 и 3 in vitro и in vivo, предположительно путем их преобразования в SLC. В то время как рецептор-связывающие домены изоформ TGFβ в высокой степени гомологичны, домены LAP более разнообразны (Фиг. 49). Таким образом, постулировалось, что LAP из TGFβ1, 2 и 3 (LAP1, 2 и 3 соответственно) могут иметь различную ингибирующую способность. Создавали рекомбинантные N-концевые слитые белки IgG1 человека с LAP1, 2 и 3 человека (Fc-LAP1, 2 и 3). Для каждого Fc-LAP наблюдали существенно разные способности к нейтрализации. В соответствии с наблюдениями с кондиционированными средами и рекомбинантными полноразмерными белками TGFβ, Fc-LAP1 более сильно ингибировал рецептор-связывающие домены всех трех изоформ TGFβ, чем Fc-LAP2 или Fc-LAP3 (Фиг. 50А, 50Б и 50В и Фиг. 51). В совокупности эти данные показывают, что относительная способность LAP1, 2 и 3 ингибировать активность TGFβ различается и может объяснить «внутреннюю» активность полноразмерных TGFβ2 и TGFβ3, не наблюдаемую для TGFβ1.
В совокупности эти данные показывают, что FL-TGFβ2 и TGFβ3 могут иметь механизмы активации и пороговые значения, отличные от таковых для TGFβ1. Хотя FL-TGFβ1 может быть активирован за счет эктопической экспрессии интегрина αvβ6, а активность FL-TGFβ3 может быть увеличена за счет связывания интегрина αvβ6, как FL-TGFβ2, так и FL-TGFβ3, по-видимому, обладают «внутренней» активностью и/или активируются с помощью еще не установленных механизмов. Биохимическая ингибирующая активность LAP1, 2 и 3 различается, при этом биохимическая IC50 LAP1 в отношении каждой рецептор-связывающейся изоформы на несколько порядков ниже, чем у LAP2 и LAP3. Учитывая, что латентный TGFβ1 имеет гораздо более высокий порог активации, чем латентный TGFβ2 или TGFβ3, эти механистические наблюдения предполагают, что экспрессия генов TGFβ2 и TGFβ3 с большей вероятностью, чем экспрессия гена TGFβ1, отражает их активность в ткани.
Рецептор-связывающие домены TGFβ1, 2 и 3 в высокой степени гомологичны, при этом аминокислотное сходство составляет 83-90%. Димерные рецептор-связывающие домены всех трех изоформ связываются с парами гетеродимерных рецепторных комплексов TGFβR1 и TGFβR2; затем тетрамерный рецепторный комплекс активирует внутриклеточную передачу сигналов посредством активности рецепторной тирозинкиназы (RTK) TGFβR1, также известной как ALK5. В канонической передаче сигналов TGFβR ALK5 фосфорилирует SMAD2 и SMAD3, которые затем ассоциируются с SMAD4, транслоцируются в ядро и направляют транскрипцию генов. Дополнительные сигнальные пути, не зависящие от SMAD, такие как МАР-киназы, AKT, JAK-STAT и NFκB, могут активироваться ниже TGFβR в определенных контекстах (Derynck, 2019 Science signaling 12). Биохимические и структурные исследования показали, что сборка сигнальных комплексов TGFβ-TGFβR имеет тонкие специфические в отношении изоформы различия: TGFβ1 и TGFβ3 сильнее связываются с TGFβR2 и образуют сильные взаимодействия только с TGFβR1 в комплексе с TGFβR2, в то время как TGFβ2 слабо связывается как с TGFβR1, так и с TGFβR2 по отдельности, и авидность может стимулировать образование полного комплекса (Radaev, 2010, JBC 285, 14806-14814). TGFβ1 и TGFβ2 кристаллизуются в «закрытые» структуры, которые облегчают связывание с TGFβR1 и TGFβR2, в то время как TGFβ3 может принимать аналогичную «закрытую» или менее упорядоченную «открытую» структуру в кристаллической форме, что может приводить к различиям в авидности сборки комплекса лиганд-рецептор. Кроме того, TGFβR3 (бетагликан), не передающий сигнал рецептор, может способствовать связыванию TGFβ2 с комплексами TGFβR1/2, но, по-видимому, не играет аналогичной роли в связывании рецепторов TGFβ1 или TGFβ3 (del Re, 2004 JBC 279, 22765-22772). Структурный анализ аллостерического ингибирующего механизма 4А11, продемонстрированный в данном документе предполагает, что двухвалентное антитело, связывающееся с димером TGFβ2 или TGFβ3, ограничивает TGFβ в форме, которая исключает одновременное связывание с двумя наборами гетеродимеров рецептора (Фиг. 7). В конечном итоге, несмотря на эти различия в сборке сигнальных комплексов, все рекомбинантные рецептор-связывающие домены TGFβ1, 2 и 3 способны индуцировать TGFβR-зависимую передачу сигналов SMAD в одинаковой степени в клеточных анализах in vitro. Следовательно, различия в активности эндогенных изоформ TGFβ in vivo скорее связаны с различиями в их паттернах экспрессии и механизмах высвобождения из SLC или LLC, чем с различиями в их рецептор-связывающих доменах.
Обсуждение
Домены LAP TGFβ1, 2 и 3 значительно менее гомологичны, чем рецептор-связывающиеся домены, при этом аминокислотное сходство составляет лишь 42-56%. Все три изоформы имеют консервативные сайты расщепления фурином (RKKR или RHRR) между LAP и рецептор-связывающим доменом, а также консервативную последовательность «застежки» (YYAKE). TGFβ1 и TGFβ3 имеют последовательность связывания интегрина RGD, но TGFβ2 имеет замену R-S в этом сайте, и не было показано, что он активируется посредством связывания интегрина. Структуры SLC, включая LAP и рецептор-связывающий домен, определяли для TGFβ1, но не для TGFβ2 или 3. «Пальцевая» область рецептор-связывающего домена TGFβ1, которая взаимодействует с TGFβR2, защищена «латентным лассо», петлей между α1 и α2 спиралями LAP, которая связана с последовательностью застежки. Во всех трех изоформах С-концевая последовательность спирали LAP α1 высококонсервативна, с остатком лизина (GQILSKL), который взаимодействует с двумя консервативными тирозинами (YYAKE) в застежке TGF1; кроме того, аланин (YYAKE) взаимодействует с аргинином в спирали α5 рядом с С-концом LAP, который сохраняется во всех трех изоформах (Shi et al. 2011; Nature 474, 343-349).
Латентные лассо трех изоформ TGFβ LAP демонстрируют некоторое разнообразие последовательностей: 6 пролинов в TGFβ1, 5 в TGFβ2 и 4 в TGFβ3, а длина лассо на 1 и 2 остатка короче в TGFβ2 и TGFβ3 соответственно, чем в TGFβ1. Существуют значительные различия в последовательностях между тремя изоформами вокруг сайта связывания интегрина RGD: TGFβ2 имеет последовательность SGD, вряд ли связывает интегрины и на 9 остатков длиннее TGFβ1 между β9 и β10 цепями; TGFβ3 имеет RGD и на 3 остатка длиннее TGFβ1 между β9 и β10 цепями; и петли между цепями β4 и β5 и цепями β7 и β8 существенно длиннее в TGFβ2 и TGFβ3, чем в TGFβ1. Хотя структурно-функциональные отношения для этих областей еще не выяснены, возможно, что они могут способствовать различным способам активации и/или ингибирующей активности доменов LAP TGFβ2 и TGFβ3 по сравнению с TGFβ1. Гетерозиготные мутации с потерей функции были описаны в генах TGFB2 и TGFB3, связанных с синдромом Лоеса-Дитца (см. Lindsay, 2012 Nature Genetics; 44, 922-927; Bertoli-Avella, 2015 J Am Coll Cardiol 65, 1324-1336; Schepers, 2018 Human Mutation 39, 621-634) в ключевых остатках в доменах LAP, что указывает на сайт расщепления фурином как в застежке в TGFβ2, так и в сайте связывания интегрина RGD в TGFβ3, что позволяет предположить, что эти консервативные мотивы важны по меньшей мере для некоторых функций TGFβ2 и TGFβ3. Хотя в данном документе показано, что полноразмерные TGFβ2 и TGFβ3 имеют «внутреннюю» активность, эта активность остается зависимой от расщепления фурином и, в случае TGFβ3, может усиливаться за счет связывания интегрина. В соответствии с представлением о том, что LAP, а не рецептор-связывающий домен, придает специфичность активации, в данном документе показано, что LAP TGFβ1 может значительно ингибировать все три изоформы, тогда как LAP TGFβ2 и TGFβ3 являются значительно менее ингибирующими.
Пример 15: Фармакокинетика специфичных в отношении изоформы антител к TGFβ у мышей C57BL/6
Фармакокинетику (PK) трех химерных антител (6F2, 2А10 и 4А11) характеризовали после однократного внутривенного (IV) введения дозы 1 или 10 мг/кг или внутрибрюшинного (IP) введения 10 мг/кг самкам мышей C57BL/6. Все три антитела продемонстрировали благоприятные ФК профили с очевидным двухфазным распределением, характеризующимся короткой фазой распределения, за которой следует более длительная фаза элиминации. Линейные и пропорциональные дозе (Cmax и AUC; площадь под кривой концентрация-время) ФК параметры наблюдали в диапазоне от 1 до 10 мг/кг, при этом клиренс составлял от 3,5 до 4,9 мл/день/кг во всех когортах. После однократного IP введения 6F2 и 4А11 в дозе 10 мг/кг AUClast составляла ≥97%, а Cmax составляла ~50-60% при сравнении AUClast и Cmax после IV введения в дозе 10 мг/кг. Данные показаны в таблице 30 ниже.
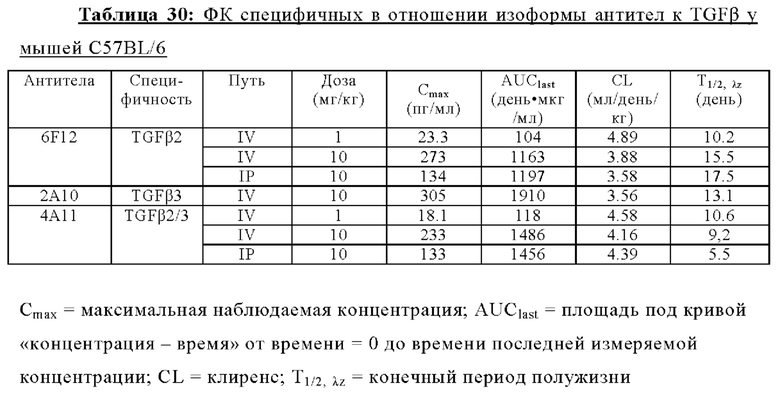
Пример 16: Активность селективных в отношении изоформы антител к TGFβ2 и TGFβ3 in vivo в модели фиброза легких
Материалы и методы
Выделение геномной ДНК легких и количественная ПЦР
Геномную ДНК выделяли из целых легких мыши с использованием набора для анализа крови и ткани с помощью DNeasy (кат. №69506; Qiagen). Уровни ДНК TGFβ определяли количественно с помощью кПЦР с использованием анализов для оценки количества копий Taqman (Applied Biosystems). Пороговые значения цикла (Ct) нормализовали к внутреннему эталону, Tert (ΔСТ). Уровни геномной ДНК рассчитывали методом 2-ΔΔСТ. Использовали следующие анализы Taqman от Applied Biosystems:
экзон 3 Tgfb2 (Mm00035180_cn)
экзон 6 Tgfb2 (Mm00035822_cn)
экзон 3 Tgfb3 (Mm00381025_cn)
экзон 6 Tgfb3 (Mm00381036_cn)
и Tert (4458373).
Гистологический анализ фиброза легких мышей в модели, индуцированной блеомицином
Фиксированные в формалине образцы легких мышей (5 долей) заливали целиком и обрабатывали по 1 предметному стеклу на животное, окрашивая гематоксилин-эозином или трихромом Массона. В качестве альтернативы фиброз оценивали с помощью иммуногистохимического анализа с использованием антитела к коллагену III (кат. № ab7778, Abeam). Степень легочного фиброза оценивали в соответствии со следующими критериями:
(A) паттерн интерстициального фиброза - количество очагов: 0, ничего не обнаружено; 1, ≤10; 2, ≤15; 3, >15/ все срезы, но отдаленные; 4, мультифокально коалесцирующие или локально обширные; 5, диффузный
(Б) интерстициальный фиброз - размер очагов: 0, ничего не обнаружено; 1, наиболее крупный очаг ≤ площади ~2 альвеолярных пространств; 2, наиболее крупный очаг ≤ площади ~4 альвеолярных пространств; 3, коалесцирующие (>4 открытых альвеолярных промежутков); 4, локально обширный (60-90% всей доли); 5, диффузный (>90% всей доли);
(B) Общие индексы: количество очагов х размер очагов
Результаты
Для оценки in vivo антифиброгенных эффектов антитела к TGFβ2 (6F12) и антитела к TGFβ3 (2А10) мышей превентивно обрабатывали и исследовали в отношении TGFβ-индуцированной экспрессии генов в легких в день 14 после введения блеомицина, когда экспрессия этих генов достигала пика. Экспрессия Serpinel, Fnl и Collal была значительно снижена у мышей, обработанных любым антителом (Фиг. 53А и 53Б). Антитело 4А11 значительно ингибировало активность как изоформы TGFβ2, так и изоформы TGFβ3 in vitro (таблица 30 выше). In vivo обработка 4А11 снижала накопление нового коллагена в день 24 по сравнению с комбинацией 6F12 и 2А10 (Фиг. 53В). Выполняли подбор дозы 4А11 и наблюдали сопоставимую эффективность при дозах 10 мг/кг три раза или один раз в неделю, но не при дозе 3 мг/кг/неделя (Фиг. 53В). Патологическое исследование легких в день 24 показало значительно более низкие показатели заболевания и накопление коллагена в группе, получавшей высокие дозы 4А11, по сравнению с контрольной группой (Фиг. 53Г и 53Д). В совокупности эти данные показывают, что обработка нейтрализующими антителами к TGFβ2 и TGFβ3 может уменьшить фиброз легких in vivo.
Пример 17: TGFβ2 и TGFβ3 способствуют фиброзу печени
Материалы и методы
Модель с использованием CCl4
Самцам мышей C57BL/6J в возрасте 8-10 недель (Jackson West) вводили в дозе 0,5 мл/кг веса тела IP четыреххлористый углерод (Sigma-Aldrich, №319961, 1:6 об./об., разбавленный в кукурузном масле) или кукурузное масло (Sigma-Aldrich) три раза в неделю в течение 6 недель. Мыши получали антитело к TGFb2/3 (4А11)-mIgG2a-LALAPG или контрольное антитело (антитело к gp120-mIgG2a-DANA) три раза в неделю в дозе 10 мг/кг в течение 6 недель. Животных умерщвляли через 72 часа после последней инъекции CCl4 и брали целую печень и сыворотку крови для гистологического и молекулярного анализа.
Выделение РНК и количественная ОТ-ПЦР на основе тканей мыши
Выделяли целые легкие мыши (исследования с использованием блеомицина) или доли печени мыши (исследования с использованием CCl4). Ткани обрабатывали с использованием диссоциатора gentleMACS (Miltenyi Biotec) для гомогенизации тканей. РНК экстрагировали из гомогенатов тканей с использованием набора RNeasy Mini (кат. №74106; Qiagen). Уровни экспрессии генов количественно определяли с помощью ОТ-кПЦР с использованием набора Taqman RNA-to-СТ 1-Step (кат. №4392938; Applied Biosystems). Реакции проводили в ПЦР-системе в реальном времени QuantStudio 6 Flex (ThermoFisher). Значения порогового цикла (Ct) нормализовали к гену домашнего хозяйства GAPDH (ΔСТ). Относительные уровни экспрессии генов рассчитывали методом 2-ΔΔCT.
Использовали следующие наборы для анализа экспрессии гена Taqman от Applied Biosystems:
Serpine (Mm00435858_m1)
Fn1 (Mm01256744_m1)
Colla1 (Mm00801666_g1)
Col3a1 (Mm01254476_m1)
Tgfb1 (Mm01178820_ml)
Tgfb2 (Mm00436955_ml)
Tgfb3 (Mm00436960_ml)
Gapdh мыши (кат. №4352661).
Гистологический анализ фиброза печени мышей в модели с использованием CCl4
Обрабатывали фиксированные в формалине образцы печени мышей. Для каждого животного один срез окрашивали гематоксилин-эозином, а один срез окрашивали трихромом. Поражения оценивают вслепую по групповой идентичности по степени нарушения архитектоники и степени фиброза на основе ранее опубликованного метода (Zhao et al, 2008 Pathol Int 58, 580-588).
Результаты
Для расширения наблюдений примера 16, приведенного выше, за пределы легочного фиброза, экспрессию гена изоформы TGFβ в образцах пациентов с NAFLD/NASH человека оценивали по опубликованному набору данных, сравнивая фиброз легкой и распространенной степени (GSE49541, (Murphy et al., 2013 Gastroenterology 145, 1076-1087; Moylan et al, 2014 Hepatology 59, 471-482)). Аналогично ткани легких при IPF, уровни экспрессии TGFB1 были сопоставимы в зависимости от тяжести заболевания, в то время как TGFB2, так и TGFB3 были значительно повышены в печени пациентов с более распространенным фиброзом (Фиг. 54А). В модели повреждения и фиброза печени, вызванных четыреххлористым углеродом (CCl4), у мышей после 6 недель воздействия CCl4 наблюдали активацию Tgfb2 и Tgfb3, но не экспрессию Tgfb1 (Фиг. 54Б). Обработка антителом к TGFβ2/3 (4А11) значительно ослабляла экспрессию гена коллагена и гистологические индексы заболевания у мышей, на которых воздействовали CCl4 (Фиг. 54В и 54Г). Гистологическое исследование показало умеренное уменьшение фиброза в группе, обработанной 4А11, по сравнению с контролем. В совокупности эти данные свидетельствуют о том, что ингибирование TGFβ2 и TGFβ3 может уменьшать фиброз печени in vivo.
Пример 18: Блокада TGFβ2 и TGFβ3 не способствует усилению иммунных ответов, связанных с блокадой антитела ко всем видам TGFβ в модели колита
Материалы и методы
Мышей C57BL/6 в возрасте 8-10 недель (Envigo) инокулировали перорально 0,2 мл 1×109 КОЕ/мл суспензии Helicobacter hepaticus в течение двух последовательных дней. Мышей обрабатывали антителом к IL10R (кат. № ВЕ0050, BioXCell) еженедельно в дозе 1 мг/мышь для индуцирования колита. Для проверки эффектов антител к TGFβ мышам вводили антитело ко всем видам TGFβ (1D11), антитело к TGFβ2/3 (4А11) или контрольное антитело три раза в неделю в дозе 10 мг/кг до конца исследования. Животных подвергали эвтаназии в день 24, и кишечник мышей выделяли, взвешивали и обрабатывали, как указано.
Собственную пластинку кишечника мыши выделяли из слепой кишки и проксимальных отделов толстой кишки в соответствии с рекомендацией производителя (кат. №130-097-410, Miltenyi). Определяли общее количество клеток, и клетки окрашивали антителами к CD3e (кат. №553064, BD Biosciences), CD4 (кат. №553729, BD Biosciences), CD8 (кат. №553036, BD Biosciences), CD11b (кат. №553312, BD Biosciences), Ly6G (кат. №551460, BD Biosciences), Ly6C (кат. №1760-09, Southern Biotech). Данные анализировали с помощью программного обеспечения Flowjo.
На Фиг. 56Г представлено количество иммунных клеток в собственной пластинке у остальных мышей в каждой группе (3-5/группа), определенное с помощью проточной цитометрии с использованием их поверхностных маркеров: Т-клетки (CD3+; CD4+ или CD8+); гранулоциты («гранулоциты», CD11b+; Ly6G+); моноциты («моноциты», CD11b+; Ly6C+). *Р<0,05, **Р<0,01, н. зн., Р >0,05 согласно ANOVA с применением критерия Даннета. Данные представляют собой средние значения ±SEM. Для ясности столбцы статистики не показаны на графиках, на которых отсутствуют номинально значимые различия между экспериментальными группами.
Результаты
Передача сигналов TGFβ играет ключевую роль в регуляции иммунных ответов. Предыдущие исследования с генетической или фармакологической блокадой передачи сигналов TGFβ показали избыточное воспаление и инфильтрацию лейкоцитами во многих органах, что приводило к серьезной побочной токсичности (М.J. Anderton et al, Toxicol Pathol 39, 916-924 (2011); M.S. Mitra et al, Tox Sci, 175, 24-34 (2020); J.C. Marie, et al., Immunity 25, 441-454 (2006); M.O. Li, et al., Immunity 25, 455-471 (2006). Для оценки вклада блокады TGFβ2 и TGFβ3 в системные иммунные ответы использовали модель колита, вызванного блокадой IL-10R вследствие инфицирования Helicobacter hepaticus (Фиг. 56А). Эта модель часто используется для оценки участия нерегулируемых иммунных реакций и кишечной микробиоты, двух основных компонентов воспалительных заболеваний кишечника (IBD) (М.С. Kullberg et al., Infection and immunity 66, 5157-5166 (1998); M.C. Kullberg et al., Infection and immunity 69, 4232-4241 (2001); M.C. Kullberg et al, JEM 203, 2485-2494 (2006)). В соответствии с предыдущими результатами, у мышей, инфицированных Н. hepaticus, которых обрабатывали антителом к IL10R, развивался колит. Обработка антителом ко всем видам TGFβ (1D11) способствовало усилению воспаления, о чем свидетельствует увеличение веса толстой кишки, экспрессия воспалительных цитокинов и инфильтрация иммунными клетками (Фиг. 56Б-56Г). У мышей, обработанных TGFβ1-сберегающим антителом к TGFβ2/3 (4А11), не наблюдалось усиления воспаления по этим параметрам. Эти результаты позволяют предположить, что селективная блокада TGFβ2/3 не будет вызывать токсических воспалительных реакций, наблюдаемых при блокаде всех видов TGFβ.
Хотя вышеизложенное изобретение было подробно описано посредством иллюстрации и примеров в целях ясности понимания, описания и примеры не следует воспринимать как ограничивающие объем настоящего изобретения. Раскрытие всех патентов и научной литературы, цитируемых в данном документе, в полном объеме и явным образом включено посредством ссылки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ДЖЕНЕНТЕК, ИНК.
<120> СЕЛЕКТИВНЫЕ В ОТНОШЕНИИ ИЗОФОРМЫ АНТИТЕЛА К TGF-БЕТА
И СПОСОБЫ ИХ ПРИМЕНЕНИЯ
<130> P35791-WO
<140>
<141>
<150> 63/044478
<151> 26 июня 2020 г.
<150> 62/991806
<151> 19 марта 2020 г.
<160> 188
<170> PatentIn, версия 3.5
<210> 1
<211> 390
<212> Белок
<213> Homo sapiens
<400> 1
Met Pro Pro Ser Gly Leu Arg Leu Leu Pro Leu Leu Leu Pro Leu Leu
1 5 10 15
Trp Leu Leu Val Leu Thr Pro Gly Arg Pro Ala Ala Gly Leu Ser Thr
20 25 30
Cys Lys Thr Ile Asp Met Glu Leu Val Lys Arg Lys Arg Ile Glu Ala
35 40 45
Ile Arg Gly Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser Pro Pro Ser
50 55 60
Gln Gly Glu Val Pro Pro Gly Pro Leu Pro Glu Ala Val Leu Ala Leu
65 70 75 80
Tyr Asn Ser Thr Arg Asp Arg Val Ala Gly Glu Ser Ala Glu Pro Glu
85 90 95
Pro Glu Pro Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr Arg Val Leu
100 105 110
Met Val Glu Thr His Asn Glu Ile Tyr Asp Lys Phe Lys Gln Ser Thr
115 120 125
His Ser Ile Tyr Met Phe Phe Asn Thr Ser Glu Leu Arg Glu Ala Val
130 135 140
Pro Glu Pro Val Leu Leu Ser Arg Ala Glu Leu Arg Leu Leu Arg Leu
145 150 155 160
Lys Leu Lys Val Glu Gln His Val Glu Leu Tyr Gln Lys Tyr Ser Asn
165 170 175
Asn Ser Trp Arg Tyr Leu Ser Asn Arg Leu Leu Ala Pro Ser Asp Ser
180 185 190
Pro Glu Trp Leu Ser Phe Asp Val Thr Gly Val Val Arg Gln Trp Leu
195 200 205
Ser Arg Gly Gly Glu Ile Glu Gly Phe Arg Leu Ser Ala His Cys Ser
210 215 220
Cys Asp Ser Arg Asp Asn Thr Leu Gln Val Asp Ile Asn Gly Phe Thr
225 230 235 240
Thr Gly Arg Arg Gly Asp Leu Ala Thr Ile His Gly Met Asn Arg Pro
245 250 255
Phe Leu Leu Leu Met Ala Thr Pro Leu Glu Arg Ala Gln His Leu Gln
260 265 270
Ser Ser Arg His Arg Arg Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser
275 280 285
Thr Glu Lys Asn Cys Cys Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys
290 295 300
Asp Leu Gly Trp Lys Trp Ile His Glu Pro Lys Gly Tyr His Ala Asn
305 310 315 320
Phe Cys Leu Gly Pro Cys Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr
325 330 335
Ser Lys Val Leu Ala Leu Tyr Asn Gln His Asn Pro Gly Ala Ser Ala
340 345 350
Ala Pro Cys Cys Val Pro Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr
355 360 365
Tyr Val Gly Arg Lys Pro Lys Val Glu Gln Leu Ser Asn Met Ile Val
370 375 380
Arg Ser Cys Lys Cys Ser
385 390
<210> 2
<211> 414
<212> Белок
<213> Homo sapiens
<400> 2
Met His Tyr Cys Val Leu Ser Ala Phe Leu Ile Leu His Leu Val Thr
1 5 10 15
Val Ala Leu Ser Leu Ser Thr Cys Ser Thr Leu Asp Met Asp Gln Phe
20 25 30
Met Arg Lys Arg Ile Glu Ala Ile Arg Gly Gln Ile Leu Ser Lys Leu
35 40 45
Lys Leu Thr Ser Pro Pro Glu Asp Tyr Pro Glu Pro Glu Glu Val Pro
50 55 60
Pro Glu Val Ile Ser Ile Tyr Asn Ser Thr Arg Asp Leu Leu Gln Glu
65 70 75 80
Lys Ala Ser Arg Arg Ala Ala Ala Cys Glu Arg Glu Arg Ser Asp Glu
85 90 95
Glu Tyr Tyr Ala Lys Glu Val Tyr Lys Ile Asp Met Pro Pro Phe Phe
100 105 110
Pro Ser Glu Asn Ala Ile Pro Pro Thr Phe Tyr Arg Pro Tyr Phe Arg
115 120 125
Ile Val Arg Phe Asp Val Ser Ala Met Glu Lys Asn Ala Ser Asn Leu
130 135 140
Val Lys Ala Glu Phe Arg Val Phe Arg Leu Gln Asn Pro Lys Ala Arg
145 150 155 160
Val Pro Glu Gln Arg Ile Glu Leu Tyr Gln Ile Leu Lys Ser Lys Asp
165 170 175
Leu Thr Ser Pro Thr Gln Arg Tyr Ile Asp Ser Lys Val Val Lys Thr
180 185 190
Arg Ala Glu Gly Glu Trp Leu Ser Phe Asp Val Thr Asp Ala Val His
195 200 205
Glu Trp Leu His His Lys Asp Arg Asn Leu Gly Phe Lys Ile Ser Leu
210 215 220
His Cys Pro Cys Cys Thr Phe Val Pro Ser Asn Asn Tyr Ile Ile Pro
225 230 235 240
Asn Lys Ser Glu Glu Leu Glu Ala Arg Phe Ala Gly Ile Asp Gly Thr
245 250 255
Ser Thr Tyr Thr Ser Gly Asp Gln Lys Thr Ile Lys Ser Thr Arg Lys
260 265 270
Lys Asn Ser Gly Lys Thr Pro His Leu Leu Leu Met Leu Leu Pro Ser
275 280 285
Tyr Arg Leu Glu Ser Gln Gln Thr Asn Arg Arg Lys Lys Arg Ala Leu
290 295 300
Asp Ala Ala Tyr Cys Phe Arg Asn Val Gln Asp Asn Cys Cys Leu Arg
305 310 315 320
Pro Leu Tyr Ile Asp Phe Lys Arg Asp Leu Gly Trp Lys Trp Ile His
325 330 335
Glu Pro Lys Gly Tyr Asn Ala Asn Phe Cys Ala Gly Ala Cys Pro Tyr
340 345 350
Leu Trp Ser Ser Asp Thr Gln His Ser Arg Val Leu Ser Leu Tyr Asn
355 360 365
Thr Ile Asn Pro Glu Ala Ser Ala Ser Pro Cys Cys Val Ser Gln Asp
370 375 380
Leu Glu Pro Leu Thr Ile Leu Tyr Tyr Ile Gly Lys Thr Pro Lys Ile
385 390 395 400
Glu Gln Leu Ser Asn Met Ile Val Lys Ser Cys Lys Cys Ser
405 410
<210> 3
<211> 412
<212> Белок
<213> Homo sapiens
<400> 3
Met Lys Met His Leu Gln Arg Ala Leu Val Val Leu Ala Leu Leu Asn
1 5 10 15
Phe Ala Thr Val Ser Leu Ser Leu Ser Thr Cys Thr Thr Leu Asp Phe
20 25 30
Gly His Ile Lys Lys Lys Arg Val Glu Ala Ile Arg Gly Gln Ile Leu
35 40 45
Ser Lys Leu Arg Leu Thr Ser Pro Pro Glu Pro Thr Val Met Thr His
50 55 60
Val Pro Tyr Gln Val Leu Ala Leu Tyr Asn Ser Thr Arg Glu Leu Leu
65 70 75 80
Glu Glu Met His Gly Glu Arg Glu Glu Gly Cys Thr Gln Glu Asn Thr
85 90 95
Glu Ser Glu Tyr Tyr Ala Lys Glu Ile His Lys Phe Asp Met Ile Gln
100 105 110
Gly Leu Ala Glu His Asn Glu Leu Ala Val Cys Pro Lys Gly Ile Thr
115 120 125
Ser Lys Val Phe Arg Phe Asn Val Ser Ser Val Glu Lys Asn Arg Thr
130 135 140
Asn Leu Phe Arg Ala Glu Phe Arg Val Leu Arg Val Pro Asn Pro Ser
145 150 155 160
Ser Lys Arg Asn Glu Gln Arg Ile Glu Leu Phe Gln Ile Leu Arg Pro
165 170 175
Asp Glu His Ile Ala Lys Gln Arg Tyr Ile Gly Gly Lys Asn Leu Pro
180 185 190
Thr Arg Gly Thr Ala Glu Trp Leu Ser Phe Asp Val Thr Asp Thr Val
195 200 205
Arg Glu Trp Leu Leu Arg Arg Glu Ser Asn Leu Gly Leu Glu Ile Ser
210 215 220
Ile His Cys Pro Cys His Thr Phe Gln Pro Asn Gly Asp Ile Leu Glu
225 230 235 240
Asn Ile His Glu Val Met Glu Ile Lys Phe Lys Gly Val Asp Asn Glu
245 250 255
Asp Asp His Gly Arg Gly Asp Leu Gly Arg Leu Lys Lys Gln Lys Asp
260 265 270
His His Asn Pro His Leu Ile Leu Met Met Ile Pro Pro His Arg Leu
275 280 285
Asp Asn Pro Gly Gln Gly Gly Gln Arg Lys Lys Arg Ala Leu Asp Thr
290 295 300
Asn Tyr Cys Phe Arg Asn Leu Glu Glu Asn Cys Cys Val Arg Pro Leu
305 310 315 320
Tyr Ile Asp Phe Arg Gln Asp Leu Gly Trp Lys Trp Val His Glu Pro
325 330 335
Lys Gly Tyr Tyr Ala Asn Phe Cys Ser Gly Pro Cys Pro Tyr Leu Arg
340 345 350
Ser Ala Asp Thr Thr His Ser Thr Val Leu Gly Leu Tyr Asn Thr Leu
355 360 365
Asn Pro Glu Ala Ser Ala Ser Pro Cys Cys Val Pro Gln Asp Leu Glu
370 375 380
Pro Leu Thr Ile Leu Tyr Tyr Val Gly Arg Thr Pro Lys Val Glu Gln
385 390 395 400
Leu Ser Asn Met Val Val Lys Ser Cys Lys Cys Ser
405 410
<210> 4
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 4
Ser Tyr Gly Met Ser
1 5
<210> 5
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 5
Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp Ser
1 5 10 15
Val Lys Asp
<210> 6
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 6
Ala Pro Gly Gly Ser Phe Asp Tyr
1 5
<210> 7
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 7
Arg Ala Ser Gln Ser Val Ser Ile Ser Arg Phe Asn Leu Met His
1 5 10 15
<210> 8
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 8
Arg Ala Ser Asn Leu Ala Ser
1 5
<210> 9
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 9
Gln His Ser Arg Glu Ser Pro Trp Thr
1 5
<210> 10
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 10
Ser Tyr Thr Val Asn
1 5
<210> 11
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 11
Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn Gly
1 5 10 15
<210> 12
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 12
His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe Asp
1 5 10 15
Pro
<210> 13
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 13
Gln Ser Ser Gln Ser Val Tyr Asn Asn Asn Tyr Leu Ser
1 5 10
<210> 14
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 14
Gly Ala Ser Thr Leu Thr Ser
1 5
<210> 15
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 15
Ala Gly Gly Tyr Ser Gly Ser Ser Asp Lys Tyr Ala
1 5 10
<210> 16
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 16
Thr Tyr Asn Val His
1 5
<210> 17
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 17
Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys Ser
1 5 10 15
<210> 18
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 18
Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe
1 5 10
<210> 19
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 19
Leu Ala Ser Glu Asp Ile Tyr Ser Asn Leu Ala
1 5 10
<210> 20
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 20
Asp Ala Arg Ser Leu Gln Asp
1 5
<210> 21
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 21
Gln Gln His His Ala Tyr Pro Phe Thr
1 5
<210> 22
<211> 110
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 22
Asp Ile Val Leu Thr Gln Ser Pro Ala Leu Ala Val Ser Leu Gly Gln
1 5 10 15
Arg Ala Thr Ile Ser Cys Arg Ala Ser Gln Ser Val Ser Ile Ser Arg
20 25 30
Phe Asn Leu Met His Trp Tyr Gln His Lys Pro Gly Gln Gln Pro Lys
35 40 45
Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg
50 55 60
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Pro
65 70 75 80
Val Gln Ala Asp Asp Leu Ala Thr Tyr Tyr Cys Gln His Ser Arg Glu
85 90 95
Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 23
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 23
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Lys Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Asp Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Gln Ser Met
65 70 75 80
Val Tyr Leu Gln Met Asp Asn Leu Lys Thr Glu Asp Thr Ala Leu Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Val Met Val Thr Val Ser Ser
115
<210> 24
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 24
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Glu Thr Val Thr Ile Glu Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Gln Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Gln Tyr Ser Leu Glu Ile Asn Ser Leu Gln Ser
65 70 75 80
Glu Asp Ala Val Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 25
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 25
Gln Val Gln Leu Lys Glu Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Ser Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Arg Met Asn Ser Leu Gln Thr Glu Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Pro Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 26
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 26
Ala Ala Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Ser Ile Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 27
<211> 122
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 27
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Tyr Thr
20 25 30
Val Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Ala Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg His Met
85 90 95
Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe Asp Pro Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 28
<211> 217
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 28
Asp Ile Val Leu Thr Gln Ser Pro Ala Leu Ala Val Ser Leu Gly Gln
1 5 10 15
Arg Ala Thr Ile Ser Cys Arg Ala Ser Gln Ser Val Ser Ile Ser Arg
20 25 30
Phe Asn Leu Met His Trp Tyr Gln His Lys Pro Gly Gln Gln Pro Lys
35 40 45
Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg
50 55 60
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Pro
65 70 75 80
Val Gln Ala Asp Asp Leu Ala Thr Tyr Tyr Cys Gln His Ser Arg Glu
85 90 95
Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Ala
100 105 110
Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu
115 120 125
Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro
130 135 140
Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn
145 150 155 160
Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His
180 185 190
Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile
195 200 205
Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 29
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 29
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Lys Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Asp Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Gln Ser Met
65 70 75 80
Val Tyr Leu Gln Met Asp Asn Leu Lys Thr Glu Asp Thr Ala Leu Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Val Met Val Thr Val Ser Ser Ala Lys Thr Thr Ala Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Val Cys Gly Asp Thr Thr Gly Ser Ser Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Leu Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Thr Ser Ser
180 185 190
Thr Trp Pro Ser Gln Ser Ile Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Glu Pro Arg Gly Pro Thr Ile Lys
210 215 220
Pro Cys Pro Pro Cys Lys Cys Pro Ala Pro Asn Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Ile Lys Asp Val Leu Met Ile Ser
245 250 255
Leu Ser Pro Ile Val Thr Cys Val Val Val Asp Val Ser Glu Asp Asp
260 265 270
Pro Asp Val Gln Ile Ser Trp Phe Val Asn Asn Val Glu Val His Thr
275 280 285
Ala Gln Thr Gln Thr His Arg Glu Asp Tyr Asn Ser Thr Leu Arg Val
290 295 300
Val Ser Ala Leu Pro Ile Gln His Gln Asp Trp Met Ser Gly Lys Glu
305 310 315 320
Phe Lys Cys Lys Val Asn Asn Lys Asp Leu Gly Ala Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Pro Lys Gly Ser Val Arg Ala Pro Gln Val Tyr Val
340 345 350
Leu Pro Pro Pro Glu Glu Glu Met Thr Lys Lys Gln Val Thr Leu Thr
355 360 365
Cys Met Val Thr Asp Phe Met Pro Glu Asp Ile Tyr Val Glu Trp Thr
370 375 380
Asn Asn Gly Lys Thr Glu Leu Asn Tyr Lys Asn Thr Glu Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Tyr Phe Met Tyr Ser Lys Leu Arg Val Glu Lys
405 410 415
Lys Asn Trp Val Glu Arg Asn Ser Tyr Ser Cys Ser Val Val His Glu
420 425 430
Gly Leu His Asn His His Thr Thr Lys Ser Phe Ser Arg Thr Pro Gly
435 440 445
Lys
<210> 30
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 30
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Glu Thr Val Thr Ile Glu Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Gln Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Gln Tyr Ser Leu Glu Ile Asn Ser Leu Gln Ser
65 70 75 80
Glu Asp Ala Val Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg Ala Asp Ala Ala
100 105 110
Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser Gly
115 120 125
Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp Ile
130 135 140
Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val Leu
145 150 155 160
Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met Ser
165 170 175
Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser Tyr
180 185 190
Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys Ser
195 200 205
Phe Asn Arg Asn Glu Cys
210
<210> 31
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 31
Gln Val Gln Leu Lys Glu Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Ser Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Arg Met Asn Ser Leu Gln Thr Glu Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Pro Gly
100 105 110
Thr Met Val Thr Val Ser Ser Ala Lys Thr Thr Ala Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Val Cys Gly Asp Thr Thr Gly Ser Ser Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Leu Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Thr Ser Ser
180 185 190
Thr Trp Pro Ser Gln Ser Ile Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Glu Pro Arg Gly Pro Thr Ile Lys
210 215 220
Pro Cys Pro Pro Cys Lys Cys Pro Ala Pro Asn Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Ile Lys Asp Val Leu Met Ile Ser
245 250 255
Leu Ser Pro Ile Val Thr Cys Val Val Val Asp Val Ser Glu Asp Asp
260 265 270
Pro Asp Val Gln Ile Ser Trp Phe Val Asn Asn Val Glu Val His Thr
275 280 285
Ala Gln Thr Gln Thr His Arg Glu Asp Tyr Asn Ser Thr Leu Arg Val
290 295 300
Val Ser Ala Leu Pro Ile Gln His Gln Asp Trp Met Ser Gly Lys Glu
305 310 315 320
Phe Lys Cys Lys Val Asn Asn Lys Asp Leu Gly Ala Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Pro Lys Gly Ser Val Arg Ala Pro Gln Val Tyr Val
340 345 350
Leu Pro Pro Pro Glu Glu Glu Met Thr Lys Lys Gln Val Thr Leu Thr
355 360 365
Cys Met Val Thr Asp Phe Met Pro Glu Asp Ile Tyr Val Glu Trp Thr
370 375 380
Asn Asn Gly Lys Thr Glu Leu Asn Tyr Lys Asn Thr Glu Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Tyr Phe Met Tyr Ser Lys Leu Arg Val Glu Lys
405 410 415
Lys Asn Trp Val Glu Arg Asn Ser Tyr Ser Cys Ser Val Val His Glu
420 425 430
Gly Leu His Asn His His Thr Thr Lys Ser Phe Ser Arg Thr Pro Gly
435 440 445
Lys
<210> 32
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 32
Ala Ala Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Ser Ile Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
Gly Asp Pro Val Ala Pro Thr Val Leu Ile Phe Pro Pro Ala Ala Asp
115 120 125
Gln Val Ala Thr Gly Thr Val Thr Ile Val Cys Val Ala Asn Lys Tyr
130 135 140
Phe Pro Asp Val Thr Val Thr Trp Glu Val Asp Gly Thr Thr Gln Thr
145 150 155 160
Thr Gly Ile Glu Asn Ser Lys Thr Pro Gln Asn Ser Ala Asp Cys Thr
165 170 175
Tyr Asn Leu Ser Ser Thr Leu Thr Leu Thr Ser Thr Gln Tyr Asn Ser
180 185 190
His Lys Glu Tyr Thr Cys Lys Val Thr Gln Gly Thr Thr Ser Val Val
195 200 205
Gln Ser Phe Asn Arg Gly Asp Cys
210 215
<210> 33
<211> 445
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 33
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Tyr Thr
20 25 30
Val Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Ala Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg His Met
85 90 95
Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe Asp Pro Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Gly Gln Pro Lys Ala Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Cys Gly Asp Thr Pro Ser Ser Thr
130 135 140
Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Leu Pro Glu Pro Val Thr
145 150 155 160
Val Thr Trp Asn Ser Gly Thr Leu Thr Asn Gly Val Arg Thr Phe Pro
165 170 175
Ser Val Arg Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Ser
180 185 190
Val Thr Ser Ser Ser Gln Pro Val Thr Cys Asn Val Ala His Pro Ala
195 200 205
Thr Asn Thr Lys Val Asp Lys Thr Val Ala Pro Ser Thr Cys Ser Lys
210 215 220
Pro Thr Cys Pro Pro Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Ile
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Asp Asp Pro Glu Val Gln
260 265 270
Phe Thr Trp Tyr Ile Asn Asn Glu Gln Val Arg Thr Ala Arg Pro Pro
275 280 285
Leu Arg Glu Gln Gln Phe Asn Ser Thr Ile Arg Val Val Ser Thr Leu
290 295 300
Pro Ile Ala His Gln Asp Trp Leu Arg Gly Lys Glu Phe Lys Cys Lys
305 310 315 320
Val His Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Arg Gly Gln Pro Leu Glu Pro Lys Val Tyr Thr Met Gly Pro Pro
340 345 350
Arg Glu Glu Leu Ser Ser Arg Ser Val Ser Leu Thr Cys Met Ile Asn
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Lys Asn Gly Lys
370 375 380
Ala Glu Asp Asn Tyr Lys Thr Thr Pro Ala Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Tyr Phe Leu Tyr Asn Lys Leu Ser Val Pro Thr Ser Glu Trp Gln
405 410 415
Arg Gly Asp Val Phe Thr Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Ile Ser Arg Ser Pro Gly Lys
435 440 445
<210> 34
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 34
Asp Ile Val Ser Lys Thr Tyr Ser Tyr Thr Thr Tyr Tyr Ser Asp Ser
1 5 10 15
Val Lys Asp
<210> 35
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 35
Asp Ile Val Ser Lys Thr Tyr Asn Tyr Ala Thr Tyr Tyr Ser Asp Ser
1 5 10 15
Val Lys Asp
<210> 36
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 36
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln His Lys Pro Gly Lys Gln Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Ile Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 37
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 37
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Pro
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 38
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 38
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln His Lys Pro Gly Lys Gln Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Ile Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 39
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 39
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Gln Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Ile Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 40
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 40
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln His Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Ile Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 41
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 41
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln His Lys Pro Gly Lys Gln Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 42
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 42
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Pro
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 43
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 43
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Pro
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Gly Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 44
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 44
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Pro
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 45
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 45
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 46
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 46
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 47
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 47
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ser Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 48
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 48
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 49
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 49
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 50
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 50
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 51
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 51
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Ser Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 52
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 52
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Gln Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 53
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 53
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Ala Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 54
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 54
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 55
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 55
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ser Asp Ile Val Ser Lys Thr Tyr Ser Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 56
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 56
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 57
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 57
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ser Asp Ile Val Ser Lys Thr Tyr Asn Tyr Ala Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 58
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 58
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln His Lys Pro Gly Lys Gln Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Ile Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 59
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 59
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Pro
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 60
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 60
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln His Lys Pro Gly Lys Gln Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Ile Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 61
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 61
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Gln Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Ile Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 62
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 62
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln His Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Ile Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 63
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 63
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln His Lys Pro Gly Lys Gln Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 64
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 64
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Pro
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 65
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 65
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Pro
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Gly Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 66
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 66
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Pro
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 67
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 67
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 68
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 68
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 69
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 69
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ser Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 70
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 70
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 71
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 71
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 72
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 72
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 73
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 73
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Ser Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 74
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 74
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Gln Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 75
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 75
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Asp Ile Val Ser Lys Thr Tyr Asn Tyr Ala Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 76
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 76
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 77
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 77
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ser Asp Ile Val Ser Lys Thr Tyr Ser Tyr Thr Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 78
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 78
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Ser Ile Ser
20 25 30
Arg Phe Asn Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 79
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 79
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Asn Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ser Asp Ile Val Ser Lys Thr Tyr Asn Tyr Ala Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Val Ala Pro Gly Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 80
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 80
Asp Ala Val Leu Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 81
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 81
Glu Gln Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ala Lys Asn Ser Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 82
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 82
Asp Ala Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Lys Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 83
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 83
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 84
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 84
Glu Ala Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 85
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 85
Asp Ala Val Leu Thr Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val
65 70 75 80
Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 86
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 86
Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu Thr
1 5 10 15
Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr Thr
20 25 30
Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn Gly
50 55 60
Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu Lys
65 70 75 80
Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala Arg
85 90 95
His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe Asp
100 105 110
Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 87
<211> 123
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 87
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Gln Val Ser Leu Lys Leu
65 70 75 80
Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala Arg His
85 90 95
Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe Asp Pro
100 105 110
Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 88
<211> 122
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 88
Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu Thr
1 5 10 15
Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr Thr
20 25 30
Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn Gly
50 55 60
Arg Phe Thr Ile Ser Lys Asp Ser Ser Gln Val Ser Leu Lys Leu Ser
65 70 75 80
Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala Arg His Met
85 90 95
Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe Asp Pro Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 89
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 89
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 90
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 90
Glu Ala Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 91
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 91
Glu Ala Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 92
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 92
Glu Ala Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Ile Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 93
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 93
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 94
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 94
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Ile Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 95
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 95
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Val Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 96
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 96
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Val Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 97
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 97
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 98
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 98
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 99
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 99
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 100
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 100
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 101
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 101
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Ile Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 102
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 102
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Ile Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Val Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 103
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 103
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Ile Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Val Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 104
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 104
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Ile Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Val Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 105
<211> 123
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 105
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Ile Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Val Thr Ile Ser Lys Asp Thr Ser Gln Val Ser Leu Lys Leu
65 70 75 80
Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala Arg His
85 90 95
Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe Asp Pro
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 106
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 106
Asp Ala Val Leu Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 107
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 107
Glu Gln Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ala Lys Asn Ser Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 108
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 108
Asp Ala Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Lys Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 109
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 109
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 110
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 110
Glu Ala Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 111
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 111
Asp Ala Val Leu Thr Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val
65 70 75 80
Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 112
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 112
Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu Thr
1 5 10 15
Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr Thr
20 25 30
Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn Gly
50 55 60
Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu Lys
65 70 75 80
Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala Arg
85 90 95
His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe Asp
100 105 110
Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 113
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 113
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Gln Val Ser Leu Lys Leu
65 70 75 80
Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala Arg His
85 90 95
Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe Asp Pro
100 105 110
Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 114
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 114
Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu Thr
1 5 10 15
Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr Thr
20 25 30
Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn Gly
50 55 60
Arg Phe Thr Ile Ser Lys Asp Ser Ser Gln Val Ser Leu Lys Leu Ser
65 70 75 80
Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala Arg His Met
85 90 95
Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe Asp Pro Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 115
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 115
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 116
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 116
Glu Ala Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 117
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 117
Glu Ala Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 118
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 118
Glu Ala Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Ile Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 119
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 119
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 120
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 120
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Ile Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 121
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 121
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Val Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 122
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 122
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Val Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 123
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 123
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 124
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 124
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 125
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 125
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 126
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 126
Glu Gln Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ser Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 127
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 127
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Ile Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Val Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 128
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 128
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Ile Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Val Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 129
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 129
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Ile Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Val Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Arg His Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe
100 105 110
Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 130
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 130
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Thr Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Thr Val Asn Trp Ile Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Tyr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Val Thr Ile Ser Lys Asp Thr Ser Gln Val Ser Leu Lys Leu
65 70 75 80
Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala Arg His
85 90 95
Met Gln Val Gly Gly Ala Pro Thr Gly Ser Met Ala Ala Phe Asp Pro
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 131
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 131
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 132
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 132
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Val Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Met
35 40 45
Gly Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 133
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 133
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 134
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 134
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 135
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 135
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Thr Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 136
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 136
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 137
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 137
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 138
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 138
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Met
35 40 45
Gly Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 139
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 139
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Val Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu
35 40 45
Gly Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 140
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 140
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Val Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Met
35 40 45
Ala Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 141
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 141
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Val Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Met
35 40 45
Gly Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Arg Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 142
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 142
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 143
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 143
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ala Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 144
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 144
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ala Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 145
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 145
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 146
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 146
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Val Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Met
35 40 45
Gly Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 147
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 147
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 148
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 148
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 149
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 149
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Thr Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 150
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 150
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 151
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 151
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 152
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 152
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Met
35 40 45
Gly Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 153
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 153
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Val Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu
35 40 45
Gly Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 154
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 154
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Val Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Met
35 40 45
Ala Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 155
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 155
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 156
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 156
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Tyr
20 25 30
Asn Val His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Ile Trp Asn Thr Gly Gly Thr Arg Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Pro Val Pro Asn Lys Trp His Phe Asp Phe Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 157
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 157
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ala Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 158
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 158
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Leu Ala Ser Glu Asp Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Arg Ser Leu Gln Asp Gly Val Pro Ala Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Pro Glu Tyr Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln His His Ala Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 159
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 159
Asp Ile Val Ser Lys Thr Tyr Gln Tyr Thr Thr Tyr Tyr Ser Asp Ser
1 5 10 15
Val Lys Asp
<210> 160
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 160
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Ile Phe Ile Thr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Phe Pro Ala Ser Gly Ser Thr Asn Tyr Asn Glu Met Phe
50 55 60
Glu Gly Lys Ala Thr Leu Thr Val Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Asp Gly Asn Tyr Ala Leu Asp Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 161
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 161
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Ser Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asp
65 70 75 80
Pro Val Glu Ala Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Asn Asn
85 90 95
Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 162
<211> 836
<212> Белок
<213> Homo sapiens
<400> 162
Met Ile Pro Phe Leu Pro Met Phe Ser Leu Leu Leu Leu Leu Ile Val
1 5 10 15
Asn Pro Ile Asn Ala Asn Asn His Tyr Asp Lys Ile Leu Ala His Ser
20 25 30
Arg Ile Arg Gly Arg Asp Gln Gly Pro Asn Val Cys Ala Leu Gln Gln
35 40 45
Ile Leu Gly Thr Lys Lys Lys Tyr Phe Ser Thr Cys Lys Asn Trp Tyr
50 55 60
Lys Lys Ser Ile Cys Gly Gln Lys Thr Thr Val Leu Tyr Glu Cys Cys
65 70 75 80
Pro Gly Tyr Met Arg Met Glu Gly Met Lys Gly Cys Pro Ala Val Leu
85 90 95
Pro Ile Asp His Val Tyr Gly Thr Leu Gly Ile Val Gly Ala Thr Thr
100 105 110
Thr Gln Arg Tyr Ser Asp Ala Ser Lys Leu Arg Glu Glu Ile Glu Gly
115 120 125
Lys Gly Ser Phe Thr Tyr Phe Ala Pro Ser Asn Glu Ala Trp Asp Asn
130 135 140
Leu Asp Ser Asp Ile Arg Arg Gly Leu Glu Ser Asn Val Asn Val Glu
145 150 155 160
Leu Leu Asn Ala Leu His Ser His Met Ile Asn Lys Arg Met Leu Thr
165 170 175
Lys Asp Leu Lys Asn Gly Met Ile Ile Pro Ser Met Tyr Asn Asn Leu
180 185 190
Gly Leu Phe Ile Asn His Tyr Pro Asn Gly Val Val Thr Val Asn Cys
195 200 205
Ala Arg Ile Ile His Gly Asn Gln Ile Ala Thr Asn Gly Val Val His
210 215 220
Val Ile Asp Arg Val Leu Thr Gln Ile Gly Thr Ser Ile Gln Asp Phe
225 230 235 240
Ile Glu Ala Glu Asp Asp Leu Ser Ser Phe Arg Ala Ala Ala Ile Thr
245 250 255
Ser Asp Ile Leu Glu Ala Leu Gly Arg Asp Gly His Phe Thr Leu Phe
260 265 270
Ala Pro Thr Asn Glu Ala Phe Glu Lys Leu Pro Arg Gly Val Leu Glu
275 280 285
Arg Ile Met Gly Asp Lys Val Ala Ser Glu Ala Leu Met Lys Tyr His
290 295 300
Ile Leu Asn Thr Leu Gln Cys Ser Glu Ser Ile Met Gly Gly Ala Val
305 310 315 320
Phe Glu Thr Leu Glu Gly Asn Thr Ile Glu Ile Gly Cys Asp Gly Asp
325 330 335
Ser Ile Thr Val Asn Gly Ile Lys Met Val Asn Lys Lys Asp Ile Val
340 345 350
Thr Asn Asn Gly Val Ile His Leu Ile Asp Gln Val Leu Ile Pro Asp
355 360 365
Ser Ala Lys Gln Val Ile Glu Leu Ala Gly Lys Gln Gln Thr Thr Phe
370 375 380
Thr Asp Leu Val Ala Gln Leu Gly Leu Ala Ser Ala Leu Arg Pro Asp
385 390 395 400
Gly Glu Tyr Thr Leu Leu Ala Pro Val Asn Asn Ala Phe Ser Asp Asp
405 410 415
Thr Leu Ser Met Asp Gln Arg Leu Leu Lys Leu Ile Leu Gln Asn His
420 425 430
Ile Leu Lys Val Lys Val Gly Leu Asn Glu Leu Tyr Asn Gly Gln Ile
435 440 445
Leu Glu Thr Ile Gly Gly Lys Gln Leu Arg Val Phe Val Tyr Arg Thr
450 455 460
Ala Val Cys Ile Glu Asn Ser Cys Met Glu Lys Gly Ser Lys Gln Gly
465 470 475 480
Arg Asn Gly Ala Ile His Ile Phe Arg Glu Ile Ile Lys Pro Ala Glu
485 490 495
Lys Ser Leu His Glu Lys Leu Lys Gln Asp Lys Arg Phe Ser Thr Phe
500 505 510
Leu Ser Leu Leu Glu Ala Ala Asp Leu Lys Glu Leu Leu Thr Gln Pro
515 520 525
Gly Asp Trp Thr Leu Phe Val Pro Thr Asn Asp Ala Phe Lys Gly Met
530 535 540
Thr Ser Glu Glu Lys Glu Ile Leu Ile Arg Asp Lys Asn Ala Leu Gln
545 550 555 560
Asn Ile Ile Leu Tyr His Leu Thr Pro Gly Val Phe Ile Gly Lys Gly
565 570 575
Phe Glu Pro Gly Val Thr Asn Ile Leu Lys Thr Thr Gln Gly Ser Lys
580 585 590
Ile Phe Leu Lys Glu Val Asn Asp Thr Leu Leu Val Asn Glu Leu Lys
595 600 605
Ser Lys Glu Ser Asp Ile Met Thr Thr Asn Gly Val Ile His Val Val
610 615 620
Asp Lys Leu Leu Tyr Pro Ala Asp Thr Pro Val Gly Asn Asp Gln Leu
625 630 635 640
Leu Glu Ile Leu Asn Lys Leu Ile Lys Tyr Ile Gln Ile Lys Phe Val
645 650 655
Arg Gly Ser Thr Phe Lys Glu Ile Pro Val Thr Val Tyr Thr Thr Lys
660 665 670
Ile Ile Thr Lys Val Val Glu Pro Lys Ile Lys Val Ile Glu Gly Ser
675 680 685
Leu Gln Pro Ile Ile Lys Thr Glu Gly Pro Thr Leu Thr Lys Val Lys
690 695 700
Ile Glu Gly Glu Pro Glu Phe Arg Leu Ile Lys Glu Gly Glu Thr Ile
705 710 715 720
Thr Glu Val Ile His Gly Glu Pro Ile Ile Lys Lys Tyr Thr Lys Ile
725 730 735
Ile Asp Gly Val Pro Val Glu Ile Thr Glu Lys Glu Thr Arg Glu Glu
740 745 750
Arg Ile Ile Thr Gly Pro Glu Ile Lys Tyr Thr Arg Ile Ser Thr Gly
755 760 765
Gly Gly Glu Thr Glu Glu Thr Leu Lys Lys Leu Leu Gln Glu Glu Val
770 775 780
Thr Lys Val Thr Lys Phe Ile Glu Gly Gly Asp Gly His Leu Phe Glu
785 790 795 800
Asp Glu Glu Ile Lys Arg Leu Leu Gln Gly Asp Thr Pro Val Arg Lys
805 810 815
Leu Gln Ala Asn Lys Lys Val Gln Gly Ser Arg Arg Arg Leu Arg Glu
820 825 830
Gly Arg Ser Gln
835
<210> 163
<211> 757
<212> Белок
<213> Homo sapiens
<400> 163
Met Val Pro Asp Thr Ala Cys Val Leu Leu Leu Thr Leu Ala Ala Leu
1 5 10 15
Gly Ala Ser Gly Gln Gly Gln Ser Pro Leu Gly Ser Asp Leu Gly Pro
20 25 30
Gln Met Leu Arg Glu Leu Gln Glu Thr Asn Ala Ala Leu Gln Asp Val
35 40 45
Arg Glu Leu Leu Arg Gln Gln Val Arg Glu Ile Thr Phe Leu Lys Asn
50 55 60
Thr Val Met Glu Cys Asp Ala Cys Gly Met Gln Gln Ser Val Arg Thr
65 70 75 80
Gly Leu Pro Ser Val Arg Pro Leu Leu His Cys Ala Pro Gly Phe Cys
85 90 95
Phe Pro Gly Val Ala Cys Ile Gln Thr Glu Ser Gly Ala Arg Cys Gly
100 105 110
Pro Cys Pro Ala Gly Phe Thr Gly Asn Gly Ser His Cys Thr Asp Val
115 120 125
Asn Glu Cys Asn Ala His Pro Cys Phe Pro Arg Val Arg Cys Ile Asn
130 135 140
Thr Ser Pro Gly Phe Arg Cys Glu Ala Cys Pro Pro Gly Tyr Ser Gly
145 150 155 160
Pro Thr His Gln Gly Val Gly Leu Ala Phe Ala Lys Ala Asn Lys Gln
165 170 175
Val Cys Thr Asp Ile Asn Glu Cys Glu Thr Gly Gln His Asn Cys Val
180 185 190
Pro Asn Ser Val Cys Ile Asn Thr Arg Gly Ser Phe Gln Cys Gly Pro
195 200 205
Cys Gln Pro Gly Phe Val Gly Asp Gln Ala Ser Gly Cys Gln Arg Arg
210 215 220
Ala Gln Arg Phe Cys Pro Asp Gly Ser Pro Ser Glu Cys His Glu His
225 230 235 240
Ala Asp Cys Val Leu Glu Arg Asp Gly Ser Arg Ser Cys Val Cys Ala
245 250 255
Val Gly Trp Ala Gly Asn Gly Ile Leu Cys Gly Arg Asp Thr Asp Leu
260 265 270
Asp Gly Phe Pro Asp Glu Lys Leu Arg Cys Pro Glu Arg Gln Cys Arg
275 280 285
Lys Asp Asn Cys Val Thr Val Pro Asn Ser Gly Gln Glu Asp Val Asp
290 295 300
Arg Asp Gly Ile Gly Asp Ala Cys Asp Pro Asp Ala Asp Gly Asp Gly
305 310 315 320
Val Pro Asn Glu Lys Asp Asn Cys Pro Leu Val Arg Asn Pro Asp Gln
325 330 335
Arg Asn Thr Asp Glu Asp Lys Trp Gly Asp Ala Cys Asp Asn Cys Arg
340 345 350
Ser Gln Lys Asn Asp Asp Gln Lys Asp Thr Asp Gln Asp Gly Arg Gly
355 360 365
Asp Ala Cys Asp Asp Asp Ile Asp Gly Asp Arg Ile Arg Asn Gln Ala
370 375 380
Asp Asn Cys Pro Arg Val Pro Asn Ser Asp Gln Lys Asp Ser Asp Gly
385 390 395 400
Asp Gly Ile Gly Asp Ala Cys Asp Asn Cys Pro Gln Lys Ser Asn Pro
405 410 415
Asp Gln Ala Asp Val Asp His Asp Phe Val Gly Asp Ala Cys Asp Ser
420 425 430
Asp Gln Asp Gln Asp Gly Asp Gly His Gln Asp Ser Arg Asp Asn Cys
435 440 445
Pro Thr Val Pro Asn Ser Ala Gln Glu Asp Ser Asp His Asp Gly Gln
450 455 460
Gly Asp Ala Cys Asp Asp Asp Asp Asp Asn Asp Gly Val Pro Asp Ser
465 470 475 480
Arg Asp Asn Cys Arg Leu Val Pro Asn Pro Gly Gln Glu Asp Ala Asp
485 490 495
Arg Asp Gly Val Gly Asp Val Cys Gln Asp Asp Phe Asp Ala Asp Lys
500 505 510
Val Val Asp Lys Ile Asp Val Cys Pro Glu Asn Ala Glu Val Thr Leu
515 520 525
Thr Asp Phe Arg Ala Phe Gln Thr Val Val Leu Asp Pro Glu Gly Asp
530 535 540
Ala Gln Ile Asp Pro Asn Trp Val Val Leu Asn Gln Gly Arg Glu Ile
545 550 555 560
Val Gln Thr Met Asn Ser Asp Pro Gly Leu Ala Val Gly Tyr Thr Ala
565 570 575
Phe Asn Gly Val Asp Phe Glu Gly Thr Phe His Val Asn Thr Val Thr
580 585 590
Asp Asp Asp Tyr Ala Gly Phe Ile Phe Gly Tyr Gln Asp Ser Ser Ser
595 600 605
Phe Tyr Val Val Met Trp Lys Gln Met Glu Gln Thr Tyr Trp Gln Ala
610 615 620
Asn Pro Phe Arg Ala Val Ala Glu Pro Gly Ile Gln Leu Lys Ala Val
625 630 635 640
Lys Ser Ser Thr Gly Pro Gly Glu Gln Leu Arg Asn Ala Leu Trp His
645 650 655
Thr Gly Asp Thr Glu Ser Gln Val Arg Leu Leu Trp Lys Asp Pro Arg
660 665 670
Asn Val Gly Trp Lys Asp Lys Lys Ser Tyr Arg Trp Phe Leu Gln His
675 680 685
Arg Pro Gln Val Gly Tyr Ile Arg Val Arg Phe Tyr Glu Gly Pro Glu
690 695 700
Leu Val Ala Asp Ser Asn Val Val Leu Asp Thr Thr Met Arg Gly Gly
705 710 715 720
Arg Leu Gly Val Phe Cys Phe Ser Gln Glu Asn Ile Ile Trp Ala Asn
725 730 735
Leu Arg Tyr Arg Cys Asn Asp Thr Ile Pro Glu Asp Tyr Glu Thr His
740 745 750
Gln Leu Arg Gln Ala
755
<210> 164
<211> 3713
<212> ДНК
<213> Homo sapiens
<400> 164
gcactcggct gtgcggcggg gcaggcatgg gagccgcgcg ctctctcccg gcgcccacac 60
ctgtctgagc ggcgcagcga gccgcggccc gggcgggctg ctcggcgcgg aacagtgctc 120
ggcatggcag ggattccagg gctcctcttc cttctcttct ttctgctctg tgctgttggg 180
caagtgagcc cttacagtgc cccctggaaa cccacttggc ctgcataccg cctccctgtc 240
gtcttgcccc agtctaccct caatttagcc aagccagact ttggagccga agccaaatta 300
gaagtatctt cttcatgtgg accccagtgt cataagggaa ctccactgcc cacttacgaa 360
gaggccaagc aatatctgtc ttatgaaacg ctctatgcca atggcagccg cacagagacg 420
caggtgggca tctacatcct cagcagtagt ggagatgggg cccaacaccg agactcaggg 480
tcttcaggaa agtctcgaag gaagcggcag atttatggct atgacagcag gttcagcatt 540
tttgggaagg acttcctgct caactaccct ttctcaacat cagtgaagtt atccacgggc 600
tgcaccggca ccctggtggc agagaagcat gtcctcacag ctgcccactg catacacgat 660
ggaaaaacct atgtgaaagg aacccagaag cttcgagtgg gcttcctaaa gcccaagttt 720
aaagatggtg gtcgaggggc caacgactcc acttcagcca tgcccgagca gatgaaattt 780
cagtggatcc gggtgaaacg cacccatgtg cccaagggtt ggatcaaggg caatgccaat 840
gacatcggca tggattatga ttatgccctc ctggaactca aaaagcccca caagagaaaa 900
tttatgaaga ttggggtgag ccctcctgct aagcagctgc cagggggcag aattcacttc 960
tctggttatg acaatgaccg accaggcaat ttggtgtatc gcttctgtga cgtcaaagac 1020
gagacctatg acttgctcta ccagcaatgc gatgcccagc caggggccag cgggtctggg 1080
gtctatgtga ggatgtggaa gagacagcag cagaagtggg agcgaaaaat tattggcatt 1140
ttttcagggc accagtgggt ggacatgaat ggttccccac aggatttcaa cgtggctgtc 1200
agaatcactc ctctcaaata tgcccagatt tgctattgga ttaaaggaaa ctacctggat 1260
tgtagggagg ggtgacacag tgttccctcc tggcagcaat taagggtctt catgttctta 1320
ttttaggaga ggccaaattg ttttttgtca ttggcgtgca cacgtgtgtg tgtgtgtgtg 1380
tgtgtgtgta aggtgtctta taatctttta cctatttctt acaattgcaa gatgactggc 1440
tttactattt gaaaactggt ttgtgtatca tatcatatat catttaagca gtttgaaggc 1500
atacttttgc atagaaataa aaaaaatact gatttggggc aatgaggaat atttgacaat 1560
taagttaatc ttcacgtttt tgcaaacttt gatttttatt tcatctgaac ttgtttcaaa 1620
gatttatatt aaatatttgg catacaagag atatgaattc ttatatgtgt gcatgtgtgt 1680
tttcttctga gattcatctt ggtggtgggt ttttttgttt ttttaattca gtgcctgatc 1740
tttaatgctt ccataaggca gtgttcccat ttaggaactt tgacagcatt tgttaggcag 1800
aatattttgg atttggaggc atttgcatgg tagtctttga acagtaaaat gatgtgttga 1860
ctatactgat acacatatta aactatacct tatagtaaac cagtatccca agctgctttt 1920
agttccaaaa atagtttctt ttccaaaggt tgttgctcta ctttgtagga agtctttgca 1980
tatggccctc ccaactttaa agtcatacca gagtggccaa gagtgtttat cccaaccctt 2040
ccatttaaca ggatttcact cacatttctg gaactagcta tttttcagaa gacaataatc 2100
agggcttaat tagaacaggc tgtatttcct cccagcaaac agttgtggcc acactaaaaa 2160
caatcatagc attttacccc tggattatag cacatctcat gttttatcat ttggatggag 2220
taatttaaaa tgaattaaat tccagagaac aatggaagca ttgcctggca gatgtcacaa 2280
cagaataacc acttgtttgg agcctggcac agtcctccag cctgatcaaa aattattctg 2340
catagttttc agtgtgcttt ctgggagcta tgtacttctt caatttggaa acttttctct 2400
ctcatttata gtgaaaatac ttggaagtta ctttaagaaa accagtgtgg cctttttccc 2460
tctagcttta aaagggccgc ttttgctgga atgctctagg ttatagataa acaattaggt 2520
ataatagcaa aaatgaaaat tggaagaatg caaaatggat cagaatcatg ccttccaata 2580
aaggccttta cacatgtttt atcaatatga ttatcaaatc acagcatata cagaaaagac 2640
ttggacttat tgtatgtttt tattttatgg ctctcggcct aagcacttct ttctaaatgt 2700
atcggagaaa aaatcaaatg gactacaagc acgtgtttgc tgtgcttgca ccccaggtaa 2760
acctgcattg tagcaatttg taaggatatt cagatggagc actgtcactt agacattctc 2820
tgggggattt tctgcttgtc tttcttgagc tttttggaag gataattctg ataaggcact 2880
caagaaacgt acaaccacag tgctttcttc aaatcatatg agaaatacta tgcatagcaa 2940
ggagatgcag agccgccagg aaaattctga gttccagcac aattttcttt ggaatctaac 3000
aggaatctag cctgaggaag aagggaggtc tccatttcta tgtctggtat ttgggggttt 3060
tgtttgtttt tgctttagct tggtgaaaaa aagttcactg aacaccaaga ccagaatgga 3120
tttttttaaa aaaatagatg ttccttttgt gaagcacctt gattccttga ttttgatttt 3180
ttgcaaagtt agacaatggc acaaagtcaa aatgaaatca atgtttagtt cacaagtaga 3240
tgtaatttac taaagaatga tacacccata tgctatatac agcttaactc acagaactgt 3300
aaaagaaaat tataaaataa ttcaacatgt ccatcttttt agtgataata aaagaaagca 3360
tggtattaaa ctatcataga agtagacaga aaaagaaaaa aggactcatg gcattattaa 3420
tataattagt gctttacatg tgttagttat acatattaga agcatatttg cctagtaagg 3480
ctagtagaac cacatttccc aaagtgtgct ccttaaacac tcatgcctta tgattttcta 3540
ccaaaagtaa aaagggttgt attaagtcag aggaagatgc ctctccattt tccctctctt 3600
tatcagaggt tcacatgcct gtctgcacat taaaagctct gggaagacct gttgtaaagg 3660
gacaagttga ggttgtaaaa tctgcattta aataaacatc tttgatcaca aaa 3713
<210> 165
<211> 6817
<212> ДНК
<213> Homo sapiens
<400> 165
gcagtcggga gccggccgtg gtggctccgt gcgtccgagc gtccgtccgc gccgtcggcc 60
atggccaagc gctccagggg ccccgggcgc cgctgcctgt tggcgctcgt gctgttctgc 120
gcctggggga cgctggccgt ggtggcccag aagccgggcg cagggtgtcc gagccgctgc 180
ctgtgcttcc gcaccaccgt gcgctgcatg catctgctgc tggaggccgt gcccgccgtg 240
gcgccgcaga cctccatcct agatcttcgc tttaacagaa tcagagagat ccaacctggg 300
gcattcaggc ggctgaggaa cttgaacaca ttgcttctca ataataatca gatcaagagg 360
atacctagtg gagcatttga agacttggaa aatttaaaat atctctatct gtacaagaat 420
gagatccagt caattgacag gcaagcattt aagggacttg cctctctaga gcaactatac 480
ctgcacttta atcagataga aactttggac ccagattcgt tccagcatct cccgaagctc 540
gagaggctat ttttgcataa caaccggatt acacatttag ttccagggac atttaatcac 600
ttggaatcta tgaagagatt gcgactggac tcaaacacac ttcactgcga ctgtgaaatc 660
ctgtggttgg cggatttgct gaaaacctac gcggagtcgg ggaacgcgca ggcagcggcc 720
atctgtgaat atcccagacg catccaggga cgctcagtgg caaccatcac cccggaagag 780
ctgaactgtg aaaggccccg gatcacctcc gagccccagg acgcagatgt gacctcgggg 840
aacaccgtgt acttcacctg cagagccgaa ggcaacccca agcctgagat catctggctg 900
cgaaacaata atgagctgag catgaagaca gattcccgcc taaacttgct ggacgatggg 960
accctgatga tccagaacac acaggagaca gaccagggta tctaccagtg catggcaaag 1020
aacgtggccg gagaggtgaa gacgcaagag gtgaccctca ggtacttcgg gtctccagct 1080
cgacccactt ttgtaatcca gccacagaat acagaggtgc tggttgggga gagcgtcacg 1140
ctggagtgca gcgccacagg ccaccccccg ccgcggatct cctggacgag aggtgaccgc 1200
acacccttgc cagttgaccc gcgggtgaac atcacgcctt ctggcgggct ttacatacag 1260
aacgtcgtac agggggacag cggagagtat gcgtgctctg cgaccaacaa cattgacagc 1320
gtccatgcca ccgctttcat catcgtccag gctcttcctc agttcactgt gacgcctcag 1380
gacagagtcg ttattgaggg ccagaccgtg gatttccagt gtgaagccaa gggcaacccg 1440
ccgcccgtca tcgcctggac caagggaggg agccagctct ccgtggaccg gcggcacctg 1500
gtcctgtcat cgggaacact tagaatctct ggtgttgccc tccacgacca gggccagtac 1560
gaatgccagg ctgtcaacat catcggctcc cagaaggtcg tggcccacct gactgtgcag 1620
cccagagtca ccccagtgtt tgccagcatt cccagcgaca caacagtgga ggtgggcgcc 1680
aatgtgcagc tcccgtgcag ctcccagggc gagcccgagc cagccatcac ctggaacaag 1740
gatggggttc aggtgacaga aagtggaaaa tttcacatca gccctgaagg attcttgacc 1800
atcaatgacg ttggccctgc agacgcaggt cgctatgagt gtgtggcccg gaacaccatt 1860
gggtcggcct cggtgagcat ggtgctcagt gtgaatgttc ctgacgtcag tcgaaatgga 1920
gatccgtttg tagctacctc catcgtggaa gcgattgcga ctgttgacag agctataaac 1980
tcaacccgaa cacatttgtt tgacagccgt cctcgttctc caaatgattt gctggccttg 2040
ttccggtatc cgagggatcc ttacacagtt gaacaggcac gggcgggaga aatctttgaa 2100
cggacattgc agctcattca ggagcatgta cagcatggct tgatggtcga cctcaacgga 2160
acaagttacc actacaacga cctggtgtct ccacagtacc tgaacctcat cgcaaacctg 2220
tcgggctgta ccgcccaccg gcgcgtgaac aactgctcgg acatgtgctt ccaccagaag 2280
taccggacgc acgacggcac ctgtaacaac ctgcagcacc ccatgtgggg cgcctcgctg 2340
accgccttcg agcgcctgct gaaatccgtg tacgagaatg gcttcaacac ccctcggggc 2400
atcaaccccc accgactgta caacgggcac gcccttccca tgccgcgcct ggtgtccacc 2460
accctgatcg ggacggagac cgtcacaccc gacgagcagt tcacccacat gctgatgcag 2520
tggggccagt tcctggacca cgacctcgac tccacggtgg tggccctgag ccaggcacgc 2580
ttctccgacg gacagcactg cagcaacgtg tgcagcaacg accccccctg cttctctgtc 2640
atgatccccc ccaatgactc ccgggccagg agcggggccc gctgcatgtt cttcgtgcgc 2700
tccagccctg tgtgcggcag cggcatgact tcgctgctca tgaactccgt gtacccgcgg 2760
gagcagatca accagctcac ctcctacata gacgcatcca acgtgtacgg gagcacggag 2820
catgaggccc gcagcatccg cgacctggcc agccaccgcg gcctgctgcg gcagggcatc 2880
gtgcagcggt ccgggaagcc gctgctcccc ttcgccaccg ggccgcccac ggagtgcatg 2940
cgggacgaga acgagagccc catcccctgc ttcctggccg gggaccaccg cgccaacgag 3000
cagctgggcc tgaccagcat gcacacgctg tggttccgcg agcacaaccg cattgccacg 3060
gagctgctca agctgaaccc gcactgggac ggcgacacca tctactatga gaccaggaag 3120
atcgtgggtg cggagatcca gcacatcacc taccagcact ggctcccgaa gatcctgggg 3180
gaggtgggca tgaggacgct gggagagtac cacggctacg accccggcat caatgctggc 3240
atcttcaacg ccttcgccac cgcggccttc aggtttggcc acacgcttgt caacccactg 3300
ctttaccggc tggacgagaa cttccagccc attgcacaag atcacctccc ccttcacaaa 3360
gctttcttct ctcccttccg gattgtgaat gagggcggca tcgatccgct tctcaggggg 3420
ctgttcgggg tggcggggaa aatgcgtgtg ccctcgcagc tgctgaacac ggagctcacg 3480
gagcggctgt tctccatggc acacacggtg gctctggacc tggcggccat caacatccag 3540
cggggccggg accacgggat cccaccctac cacgactaca gggtctactg caatctatcg 3600
gcggcacaca cgttcgagga cctgaaaaat gagattaaaa accctgagat ccgggagaaa 3660
ctgaaaaggt tgtatggctc gacactcaac atcgacctgt ttccggcgct cgtggtggag 3720
gacctggtgc ctggcagccg gctgggcccc accctgatgt gtcttctcag cacacagttc 3780
aagcgcctgc gagatgggga caggttgtgg tatgagaacc ctggggtgtt ctccccggcc 3840
cagctgactc agatcaagca gacgtcgctg gccaggatcc tatgcgacaa cgcggacaac 3900
atcacccggg tgcagagcga cgtgttcagg gtggcggagt tccctcacgg ctacggcagc 3960
tgtgacgaga tccccagggt agacctccgg gtgtggcagg actgctgtga agactgtagg 4020
accagggggc agttcaatgc cttttcctat catttccgag gcagacggtc tcttgagttc 4080
agctaccagg aggacaagcc gaccaagaaa acaagaccac ggaaaatacc cagtgttggg 4140
agacaggggg aacatctcag caacagcacc tcagccttca gcacacgctc agatgcatct 4200
gggacaaatg acttcagaga gtttgttctg gaaatgcaga agaccatcac agacctcaga 4260
acacagataa agaaacttga atcacggctc agtaccacag agtgcgtgga tgccgggggc 4320
gaatctcacg ccaacaacac caagtggaaa aaagatgcat gcaccatttg tgaatgcaaa 4380
gacgggcagg tcacctgctt cgtggaagct tgcccccctg ccacctgtgc tgtccccgtg 4440
aacatcccag gggcctgctg tccagtctgc ttacagaaga gggcggagga aaagccctag 4500
gctcctggga ggctcctcag agtttgtctg ctgtgccatc gtgagatcgg gtggccgatg 4560
gcagggagct gcggactgca gaccaggaaa cacccagaac tcgtgacatt tcatgacaac 4620
gtccagctgg tgctgttaca gaaggcagtg caggaggctt ccaaccagag catctgcgga 4680
gaaggaggca cagcaggtgc ctgaagggaa gcaggcagga gtcctagctt cacgttagac 4740
ttctcaggtt tttatttaat tcttttaaaa tgaaaaattg gtgctactat taaattgcac 4800
agttgaatca tttaggcgcc taaattgatt ttgcctccca acaccatttc tttttaaata 4860
aagcaggata cctctatatg tcagccttgc cttgttcaga tgccaggagc cggcagacct 4920
gtcacccgca ggtggggtga gtcttggagc tgccagaggg gctcaccgaa atcggggttc 4980
catcacaagc tatgtttaaa aagaaaattg gtgtttggca aacggaacag aacctttgat 5040
gagagcgttc acagggacac tgtctggggg tgcagtgcaa gcccccggcc tcttccctgg 5100
gaacctctga actcctcctt cctctgggct ctctgtaaca tttcaccaca cgtcagcatc 5160
taatcccaag acaaacattc ccgctgctcg aagcagctgt atagcctgtg actctccgtg 5220
tgtcagctcc ttccacacct gattagaaca ttcataagcc acatttagaa acaggtttgc 5280
tttcagctgt cacttgcaca catactgcct agttgtgaac caaatgtgaa aaaacctcct 5340
tcatcccatt gtgtatctga tacctgccga gggccaaggg tgtgtgttga caacgccgct 5400
cccagccggc cctggttgcg tccacgtcct gaacaagagc cgcttccgga tggctcttcc 5460
caagggagga ggagctcaag tgtcgggaac tgtctaactt caggttgtgt gagtgcgtta 5520
aaaaaaaaaa aaaaaaagaa tccctatacc tcatttgtat ttttaaaatg cgtgatgttt 5580
tatgaaattg tgtccatttt ttaggtatta gatatggcag aaaaaccatt tccactatgc 5640
aaagttcttt tagacgtcag tgaaaatcaa ctctcatacc tcatggtctc tctttaattg 5700
accaaaacct tccatttttc tctaaataca aagcgatctg tgttctgagc aacctttccc 5760
cgaacacaca gcttcagtgc agcacgctga cctgagtatc caccatgtgc caggcacagt 5820
gctgggcaca cgaggcacca aggtccgggc cacctgcccg cagcaaggcc cagctgaggt 5880
ggtggaggga gcccctgagg tcaggggccg tttcggttca gggtggcagg tgtccagcac 5940
tggggtatgg cgtcgaggct tccatggggt gggggaggcc agcttccttc tgacaggatg 6000
ggcgcataca gtgcctggtg tgatttgtgc acaacccgtg ttccaggtgc acatcctccc 6060
aaggagacac ccagaccctt ccagcacggg ccggccaagt tgctgcggcg gaggcagcat 6120
ttcagctgtg aggaaggtca ttggattcat gtgttttatc tgtaaaaatg gttgtcttaa 6180
cttcttaacc tcatattggt aagtgattga taaaaattgg ttggtgtttc atgacatgtg 6240
gacttctttt gaaatagcaa gtcaaatgta gtgaccaaat tgtggaagag atttctgtca 6300
aataggaaat gtgtaagttc gtctaaaagc tgatggttat gtaagttgct caggcactca 6360
gatgacagca gattctgggt tctgggagtg ttctgtgcct cttacatgcc ctggaggcct 6420
catggtctca gtgctgaggc ggcacacctg tagcacacct gcgtaatgtg cggtctgggc 6480
cagtcacaag gaattgtgtt gtctaagcca aagggggaag ctgactgtga tttaccaaaa 6540
aaaattctgt aattcaaacc aaaatgtctg cggaatcacc agtttgatac tctctgtaat 6600
cagaacagtg ggcagtgcct gggtgaacgt gtctagcagc cactgtgcgg gatcgctgta 6660
acaggagtgg aatgtacata tttatttact tttctaactg ctccaacagc caaatgcctt 6720
ttttatgacc attgtattca gttcattacc aaagaaatgt ttgcactttg taatgatgcc 6780
tttcagttca aataaatggg tcacattttc aaatgga 6817
<210> 166
<211> 5577
<212> ДНК
<213> Homo sapiens
<400> 166
ggatcacagc ccttccccga tcctctccgt gggagccagc gagcctctct ccctgatctt 60
acgtgctcaa gggagctcac acgttcacca actcaccctt gaagtcatct caagaacaaa 120
agacaactga aagaagctgt tgtgaaggca gagcagcatc tgctgaagag acagaaacca 180
gccccagagg tgtcacagga aggcaccagc aaggacattg gtctttgatt tgattcagca 240
gtcctgtcaa gtataaatgt gatggctgtg ctgcctggcc ctctgcagct gctgggagtg 300
ctgcttacca tttccctgag ttccatcagg ctcattcagg ctggtgccta ctatgggatc 360
aagccgctgc cacctcaaat tcctcctcag atgccaccac aaattccaca ataccagccc 420
ctgggtcagc aagtacctca catgcctttg gccaaagatg gccttgccat gggcaaggag 480
atgccccact tgcagtatgg caaagagtat ccacacctac cccaatatat gaaggaaatt 540
caaccggcgc caagaatggg caaggaagcc gtacccaaga aaggcaaaga aataccatta 600
gccagtttac gaggggaaca aggtccccgt ggagagcctg gcccaagagg accacctggg 660
ccccctggtt tgccaggtca tgggatacct ggaattaaag gaaaaccagg gccacaggga 720
tatccaggag ttggaaagcc aggtatgcct ggaatgccag ggaagccagg agccatgggc 780
atgcctgggg caaaaggaga aattggacag aaaggggaaa ttgggcctat ggggatccca 840
ggaccacaag gacctccagg gcctcatgga cttcctggca ttgggaagcc aggtgggcca 900
gggttaccag ggcaaccagg accaaagggt gatcgaggac ccaaaggact accaggacct 960
caaggccttc ggggtcctaa aggagacaag ggcttcggga tgccaggtgc gccaggtgta 1020
aaggggcctc cagggatgca cggccctccc ggccctgttg gactgccagg agtgggcaaa 1080
ccaggagtga caggcttccc tgggccccag ggccccctgg gaaagccagg ggctccagga 1140
gaacctgggc cacaaggccc tattggggta ccgggggttc aaggacctcc tgggataccc 1200
ggaattggaa agccaggcca ggatgggatc ccaggccagc caggatttcc aggtggcaaa 1260
ggggagcaag gactgccagg gctaccagga cccccaggcc ttccagggat tgggaaacca 1320
ggcttcccag gacccaaagg tgaccggggc atgggaggtg ttcctggggc tcttggacca 1380
agaggggaga aaggaccaat aggtgcccca ggaatagggg gtcctccagg agagccaggc 1440
ctgcctggaa tcccaggtcc tatgggccct ccaggtgcta ttggttttcc tggacccaaa 1500
ggagaaggtg ggattgtagg gccacagggg ccaccaggtc ccaagggtga gccagggctt 1560
caaggcttcc caggaaagcc aggtttcctt ggtgaagtag ggcctcctgg catgaggggt 1620
ttgccaggtc ccatagggcc caagggggaa gctgggcaaa aaggtgtacc aggactccct 1680
ggtgttccag ggcttctcgg acctaaggga gagccaggaa tcccagggga tcagggttta 1740
cagggccccc caggtatccc agggattggg ggccctagtg gccccattgg accacctggg 1800
attccaggcc ccaaagggga gccgggcctc ccagggcccc ctgggttccc tggtataggg 1860
aaacccggag tggcaggact tcatggcccc ccagggaagc ctggtgccct tggtcctcaa 1920
ggccagcctg gccttccagg acccccaggc cctccaggac ctccaggacc cccagctgtg 1980
atgcccccta caccaccacc ccagggagag tatctgccag atatggggct gggaattgat 2040
ggcgtgaaac ccccccatgc ctacggggct aagaaaggca agaatggagg gccagcctat 2100
gagatgcctg catttaccgc cgagctaacc gcacctttcc caccggtggg ggccccagtg 2160
aagtttaaca aactgctgta taacggcaga cagaactaca acccgcagac aggcatcttc 2220
acctgtgagg tccctggtgt ctactacttt gcataccacg ttcactgcaa ggggggcaac 2280
gtgtgggttg ctctattcaa gaacaacgag cccgtgatgt acacgtacga cgagtacaaa 2340
aagggcttcc tggaccaggc atctgggagt gcagtgctgc tgctcaggcc cggagaccgg 2400
gtgttcctcc agatgccctc agaacaggct gcaggactgt atgccgggca gtatgtccac 2460
tcctcctttt caggatattt attgtatccc atgtaaaaac aaaaaaacaa aaaacaaaga 2520
aaagaaagag attttataga agaaaatgac acaccaaaaa atccaaatga aaaacataat 2580
tgcttcaaaa cacttacaca gttggaaagt tatatgtaag tgaaaatttg gaccattgtg 2640
tacaaataaa aactaagatg catgtttaat actccacaca gcagcctgta attgcgaatg 2700
atgggataga gttatgtatc aagtactgac acttggttgt acccactgga atcatattag 2760
ctgttttatg ttatatgctt ccacagtaac ctgcttattc agatcagtca aaatatatca 2820
gtatgaaaga tcatagctaa tgaaaggcac tcactcatat tgtttacttt aaaatattta 2880
taaatatgcc ttaaagaaat acaaatgata acaattacat accgtattta cttgcttaat 2940
ttcctctgta tttgtgtaga tactttgaca tggaatatat ggtggggaga cccgtagtgt 3000
taccgcccca gtgggagggg gccctgggga ccctggtaat gctttagtca aagggatatc 3060
tctcttgtat cagaggctgt gtcttttagt aacaggagtc ctcgtcagaa ttgcgtgtct 3120
gttgtctcta aaagaatggg tgaaccaatc ggcctttgtg aatttattca gtgccttctc 3180
tgtaccaagc actgggtaag gcacttttgt ggagcattag acagtaaccc tcaaggagct 3240
agagaaccgg atgggagaca tgagcagtaa ttaactcact tgttccccag agtttctatt 3300
tgttttgatt ttctttttct gtgacttatt ttcctatttt ctttcctcca tgtaattttc 3360
actatggccc aactaatata aacacctgga aattacaagg aaaaaaaatt cttcctctaa 3420
taactttcca aatttgtgga atatttattt gtaatagcag ttatcagtta tgcttatata 3480
gcattaaaaa ttctcctcct ttgactacac acacaaccac agtgtggttc taatcatgga 3540
gatatcagta atttttagta actgaatttt gaggacattt ctctgtttag catgtatgca 3600
aactgatatg taatctgagg ttccaaagtc aatttttttc tttttttttt gagatggagt 3660
cttactctgt cacccaggct ggagtgcagt agcacgatct tggcttactg caacctctac 3720
ctcctaggtt caagcaattg tcctgtctca gcctcccgag tactgggact actgtcttgc 3780
gccaccatgc ctggctaatt tttgtatatt tagtaaagat gggttttcgc catgttggcc 3840
aggctggtct caaactcctg gcctcaagtg atgcacctgc ctcagccttc caaagtgctg 3900
ggattacagg catgagccac cgtgcccggc caaagtcagc tttcaaaatc caagccataa 3960
ttggtgaggg gggagtttca gaattacata gaaaaattaa tatttgaaaa aataattctg 4020
aaatttcgaa tttaaaaaca gatgtgctgc ttctgggtgt aggtagtaaa agtataggaa 4080
aagaactgtt tccttagaag cggactgtgg aagggctatg tagaatgtca aagggcaaca 4140
agagcctgtg tttttaatgt catcctgtac tcggcacaaa tcaaaggcca atacaagtct 4200
gaaaagcaga aataaatatt tttccaggtt tttgcttggg cacatactaa ctgctttggg 4260
cattctaatc tggtctccaa acaccaaaga cccatttcga gcctgctatt agcctgctgc 4320
tgactctatc acttggagca ataatgtggg gttatggtgg tggaatcttg tatatttttg 4380
tcaaaaataa aaccatgagt taaggggata gatatagatg gaaaaataca caaataaata 4440
cggtatgaaa acacatggaa atgtgtcttt gtcaaatctg aatcattatt accatcacaa 4500
aaattcttct cttggccaat atctcatttc cctatatagt atacaagcac catttcttct 4560
caatttttaa gaagagaaat tagtccatta ccacaggggt tcttgtcact actaattata 4620
caacaatctt ttcccaacaa aaagatgtcc tccacaacct ttgttttcaa agcagacagc 4680
atctatgtgg ccaaatatac tttgggttgt tcttgaggat actggttttg ggctgatgac 4740
tatggtgggc agcatggatc cattgggctc cttctgctaa acagccacat tgaaatggtt 4800
taaaagcaag tcagatcagg tgatttgtaa aattgtattt atctgtacat gtatgggctt 4860
ttaattccca ccaagaaaga gagaaattat ctttttagtt aaaaccaaat ttcacttttc 4920
aaaatatctt ccaacttatt tattggttgt cactcaattg cctatatata tatatatatg 4980
tgtgtgtgtg tgtgtgtgcg cgtgagcgca cgtgtgtgta tgcgtgcgca tgtgtgtgta 5040
tgtgtattat cagacatagg tttctaactt ttagatagaa gaggagcaac atctatgcca 5100
aatactgtgc attctacaat ggtgctaatc tcagacctaa atgatactcc atttaattta 5160
aaaaagagtt ttaaataatt atctatgtgc ctgtatttcc cttttgagtg ctgcacaaca 5220
tgttaacata ttagtgtaaa agcagatgaa acaaccacgt gttctaaagt ctagggattg 5280
tgctataatc cctatttagt tcaaaattaa ccagaattct tccatgtgaa atggaccaaa 5340
ctcatattat tgttatgtaa atacagagtt ttaatgcagt atgacatccc acaggggaaa 5400
agaatgtctg tagtgggtga ctgttatcaa atattttata gaatacaatg aacggtgaac 5460
agactggtaa cttgtttgag ttcccatgac agatttgaga cttgtcaata gcaaatcatt 5520
tttgtattta aatttttgta ctgatttgaa aaacatcatt aaatatcttt aaaagta 5577
<210> 167
<211> 10532
<212> ДНК
<213> Homo sapiens
<400> 167
agtcttccac caaaggccgt tcagttctcc tgggctccag cctcctgcaa ggactgcaag 60
agttttcctc cgcagctctg agtctccact tttttggtgg agaaaggctg caaaaagaaa 120
aagagacgca gtgagtggga aaagtatgca tcctattcaa acctaattga atcgaggagc 180
ccagggacac acgccttcag gtttgctcag gggttcatat ttggtgctta gacaaattca 240
aaatgaggaa acatcggcac ttgcccttag tggccgtctt ttgcctcttt ctctcaggct 300
ttcctacaac tcatgcccag cagcagcaag cagatgtcaa aaatggtgcg gctgctgata 360
taatatttct agtggattcc tcttggacca ttggagagga acatttccaa cttgttcgag 420
agtttctata tgatgttgta aaatccttag ctgtgggaga aaatgatttc cattttgctc 480
tggtccagtt caacggaaac ccacataccg agttcctgtt aaatacgtat cgtactaaac 540
aagaagtcct ttctcatatt tccaacatgt cttatattgg gggaaccaat cagactggaa 600
aaggattaga atacataatg caaagccacc tcaccaaggc tgctggaagc cgggccggtg 660
acggagtccc tcaggttatc gtagtgttaa ctgatggaca ctcgaaggat ggccttgctc 720
tgccctcagc ggaacttaag tctgctgatg ttaacgtgtt tgcaattgga gttgaggatg 780
cagatgaagg agcgttaaaa gaaatagcaa gtgaaccgct caatatgcat atgttcaacc 840
tagagaattt tacctcactt catgacatag taggaaactt agtgtcctgt gtgcattcat 900
ccgtgagtcc agaaagggct ggggacacgg aaacccttaa agacatcaca gcacaagact 960
ctgctgacat tattttcctt attgatggat caaacaacac cggaagtgtc aatttcgcag 1020
tcattctcga cttccttgta aatctccttg agaaactccc aattggaact cagcagatcc 1080
gagtgggggt ggtccagttt agcgatgagc ccagaaccat gttctccttg gacacctact 1140
ccaccaaggc ccaggttctg ggtgcagtga aagccctcgg gtttgctggt ggggagttgg 1200
ccaatatcgg cctcgccctt gatttcgtgg tggagaacca cttcacccgg gcagggggca 1260
gccgcgtgga ggaaggggtt ccccaggtgc tggtcctcat aagtgccggg ccttctagtg 1320
acgagattcg ctacggggtg gtagcactga agcaggctag cgtgttctca ttcggccttg 1380
gagcccaggc cgcctccagg gcagagcttc agcacatagc taccgatgac aacttggtgt 1440
ttactgtccc ggaattccgt agctttgggg acctccagga gaaattactg ccgtacattg 1500
ttggcgtggc ccaaaggcac attgtcttga aaccgccaac cattgtcaca caagtcattg 1560
aagtcaacaa gagagacata gtcttcctgg tggatggctc atctgcactg ggactggcca 1620
acttcaatgc catccgagac ttcattgcta aagtcatcca gaggctggaa atcggacagg 1680
atcttatcca ggtggcagtg gcccagtatg cagacactgt gaggcctgaa ttttatttca 1740
atacccatcc aacaaaaagg gaagtcataa ccgctgtgcg gaaaatgaag cccctggacg 1800
gctcggccct gtacacgggc tctgctctag actttgttcg taacaaccta ttcacgagtt 1860
cagccggcta ccgggctgcc gaggggattc ctaagctttt ggtgctgatc acaggtggta 1920
agtccctaga tgaaatcagc cagcctgccc aggagctgaa gagaagcagc ataatggcct 1980
ttgccattgg gaacaagggt gccgatcagg ctgagctgga agagatcgct ttcgactcct 2040
ccctggtgtt catcccagct gagttccgag ccgccccatt gcaaggcatg ctgcctggct 2100
tgctggcacc tctcaggacc ctctctggaa cccctgaagt tcactcaaac aaaagggata 2160
tcatctttct tttggatgga tcagccaacg ttggaaaaac caatttccct tatgtgcgcg 2220
actttgtaat gaacctagtt aacagccttg atattggaaa tgacaatatt cgtgttggtt 2280
tagtgcaatt tagtgacact cctgtaacgg agttctcttt aaacacatac cagaccaagt 2340
cagatatcct tggtcatctg aggcagctgc agctccaggg aggttcgggc ctgaacacag 2400
gctcagccct aagctatgtc tatgccaacc acttcacgga agctggcggc agcaggatcc 2460
gtgaacacgt gccgcagctc ctgcttctgc tcacagctgg gcagtctgag gactcctatt 2520
tgcaagctgc caacgccttg acacgcgcgg gcatcctgac tttttgtgtg ggagctagcc 2580
aggcgaataa ggcagagctt gagcagattg cttttaaccc aagcctggtg tatctcatgg 2640
atgatttcag ctccctgcca gctttgcctc agcagctgat tcagccccta accacatatg 2700
ttagtggagg tgtggaggaa gtaccactcg ctcagccaga gagcaagcga gacattctgt 2760
tcctctttga cggctcagcc aatcttgtgg gccagttccc tgttgtccgt gactttctct 2820
acaagattat cgatgagctc aatgtgaagc cagaggggac ccgaattgcg gtggctcagt 2880
acagcgatga tgtcaaggtg gagtcccgtt ttgatgagca ccagagtaag cctgagatcc 2940
tgaatcttgt gaagagaatg aagatcaaga cgggcaaagc cctcaacctg ggctacgcgc 3000
tggactatgc acagaggtac atttttgtga agtctgctgg cagccggatc gaggatggag 3060
tgcttcagtt cctggtgctg ctggtcgcag gaaggtcatc tgaccgtgtg gatgggccag 3120
caagtaacct gaagcagagt ggggttgtgc ctttcatctt ccaagccaag aacgcagacc 3180
ctgctgagtt agagcagatc gtgctgtctc cagcgtttat cctggctgca gagtcgcttc 3240
ccaagattgg agatcttcat ccacagatag tgaatctctt aaaatcagtg cacaacggag 3300
caccagcacc agtttcaggt gaaaaggacg tggtgtttct gcttgatggc tctgagggcg 3360
tcaggagcgg cttccctctg ttgaaagagt ttgtccagag agtggtggaa agcctggatg 3420
tgggccagga ccgggtccgc gtggccgtgg tgcagtacag cgaccggacc aggcccgagt 3480
tctacctgaa ttcatacatg aacaagcagg acgtcgtcaa cgctgtccgc cagctgaccc 3540
tgctgggagg gccgaccccc aacaccgggg ccgccctgga gtttgtcctg aggaacatcc 3600
tggtcagctc tgcgggaagc aggataacag aaggtgtgcc ccagctgctg atcgtcctca 3660
cggccgacag gtctggggat gatgtgcgga acccctccgt ggtcgtgaag aggggtgggg 3720
ctgtgcccat tggcattggc atcgggaacg ctgacatcac agagatgcag accatctcct 3780
tcatcccgga ctttgccgtg gccattccca cctttcgcca gctggggacc gtccaacagg 3840
tcatctctga gagggtgacc cagctcaccc gcgaggagct gagcaggctg cagccggtgt 3900
tgcagcctct accgagccca ggtgttggtg gcaagaggga cgtggtcttt ctcatcgatg 3960
ggtcccaaag tgccgggcct gagttccagt acgttcgcac cctcatagag aggctggttg 4020
actacctgga cgtgggcttt gacaccaccc gggtggctgt catccagttc agcgatgacc 4080
ccaaggtgga gttcctgctg aacgcccatt ccagcaagga tgaagtgcag aacgcggtgc 4140
agcggctgag gcccaaggga gggcggcaga tcaacgtggg caatgccctg gagtacgtgt 4200
ccaggaacat cttcaagagg cccctgggga gccgcattga agagggcgtc ccgcagttcc 4260
tggtcctcat ctcgtctgga aagtctgacg atgaggtgga cgacccggcg gtggagctca 4320
agcagtttgg cgtggcccct ttcacgatcg ccaggaacgc agaccaggag gagctggtga 4380
agatctcgct gagccccgaa tatgtgttct cggtgagcac cttccgggag ctgcccagcc 4440
tggagcagaa actgctgacg cccatcacga ccctgacctc agagcagatc cagaagctct 4500
tagccagcac tcgctatcca cctccagcag ttgagagtga tgctgcagac attgtctttc 4560
tgatcgacag ctctgaggga gttaggccag atggctttgc acatattcga gattttgtta 4620
gcaggattgt tcgaagactc aacatcggcc ccagtaaagt gagagttggg gtcgtgcagt 4680
tcagcaatga tgtcttccca gaattctatc tgaaaaccta cagatcccag gccccggtgc 4740
tggacgccat acggcgcctg aggctcagag gggggtcccc actgaacact ggcaaggctc 4800
tcgaatttgt ggcaagaaac ctctttgtta agtctgcggg gagtcgcata gaagacgggg 4860
tgccccaaca cctggtcctg gtcctgggtg gaaaatccca ggacgatgtg tccaggttcg 4920
cccaggtgat ccgttcctcg ggcattgtga gtttaggggt aggagaccgg aacatcgaca 4980
gaacagagct gcagaccatc accaatgacc ccagactggt cttcacagtg cgagagttca 5040
gagagcttcc caacatagaa gaaagaatca tgaactcgtt tggaccctcc gcagccactc 5100
ctgcacctcc aggggtggac acccctcctc cttcacggcc agagaagaag aaagcagaca 5160
ttgtgttcct gttggatggt tccatcaact tcaggaggga cagtttccag gaagtgcttc 5220
gttttgtgtc tgaaatagtg gacacagttt atgaagatgg cgactccatc caagtggggc 5280
ttgtccagta caactctgac cccactgacg aattcttcct gaaggacttc tctaccaaga 5340
ggcagattat tgacgccatc aacaaagtgg tctacaaagg gggaagacac gccaacacta 5400
aggtgggcct tgagcacctg cgggtaaacc actttgtgcc tgaggcaggc agccgcctgg 5460
accagcgggt ccctcagatt gcctttgtga tcacgggagg aaagtcggtg gaagatgcac 5520
aggatgtgag cctggccctc acccagaggg gggtcaaagt gtttgctgtt ggagtgagga 5580
atatcgactc ggaggaggtt ggaaagatag cgtccaacag cgccacagcg ttccgcgtgg 5640
gcaacgtcca ggagctgtcc gaactgagcg agcaagtttt ggaaactttg catgatgcga 5700
tgcatgaaac cctttgccct ggtgtaactg atgctgccaa agcttgtaat ctggatgtga 5760
ttctggggtt tgatggttct agagaccaga atgtttttgt ggcccagaag ggcttcgagt 5820
ccaaggtgga cgccatcttg aacagaatca gccagatgca cagggtcagc tgcagcggtg 5880
gccgctcgcc caccgtgcgt gtgtcagtgg tggccaacac gccctcgggc ccggtggagg 5940
cctttgactt tgacgagtac cagccagaga tgctcgagaa gttccggaac atgcgcagcc 6000
agcaccccta cgtcctcacg gaggacaccc tgaaggtcta cctgaacaag ttcagacagt 6060
cctcgccgga cagcgtgaag gtggtcattc attttactga tggagcagac ggagatctgg 6120
ctgatttaca cagagcatct gagaacctcc gccaagaagg agtccgtgcc ttgatcctgg 6180
tgggccttga acgagtggtc aacttggagc ggctaatgca tctggagttt gggcgagggt 6240
ttatgtatga caggcccctg aggcttaact tgctggactt ggattatgaa ctagcggagc 6300
agcttgacaa cattgccgag aaagcttgct gtggggttcc ctgcaagtgc tctgggcaga 6360
ggggagaccg cgggcccatc ggcagcatcg ggccaaaggg tattcctgga gaagacggct 6420
accgaggcta tcctggtgat gagggtggac ccggtgagcg tggtccgcct ggtgtgaacg 6480
gcactcaagg tttccagggc tgcccgggcc agagaggagt aaagggctct cggggattcc 6540
caggagagaa gggcgaagta ggagaaattg gactggatgg tctggatggt gaagatggag 6600
acaaaggatt gcctggttct tctggagaga aagggaatcc tggaagaagg ggtgataaag 6660
gacctcgagg agagaaagga gaaagaggag atgttgggat tcgaggggac ccgggtaacc 6720
caggacaaga cagccaggag agaggaccca aaggagaaac cggtgacctc ggccccatgg 6780
gtgtcccagg gagagatgga gtacctggag gacctggaga aactgggaag aatggtggct 6840
ttggccgaag gggacccccc ggagctaagg gcaacaaggg cggtcctggc cagccgggct 6900
ttgagggaga gcaggggacc agaggtgcac agggcccagc tggtcctgct ggtcctccag 6960
ggctgatagg agaacaaggc atttctggac ctcggggaag cggaggtgcc gctggtgctc 7020
ctggagaacg aggcagaacc ggtccactgg gaagaaaggg tgagcccgga gagccaggac 7080
caaaaggagg aatcgggaac cggggccctc gtggggagac gggagatgac gggagagacg 7140
gagttggcag tgaaggacgc agaggcaaaa aaggagaaag aggattccct ggatacccag 7200
gaccaaaggg taacccaggt gaacctgggc taaatggaac aacaggaccc aaaggcatca 7260
gaggccgaag gggaaattcg ggacctccag ggatagttgg acagaaggga gaccctggct 7320
acccaggacc agctggtccc aagggcaaca ggggcgactc catcgatcaa tgtgccctca 7380
tccaaagcat caaagataaa tgcccttgct gttacgggcc cctggagtgc cccgtcttcc 7440
caacagaact agcctttgct ttagacacct ctgagggagt caaccaagac actttcggcc 7500
ggatgcgaga tgtggtcttg agtattgtga atgacctgac cattgctgag agcaactgcc 7560
cacggggggc ccgggtggct gtggtcacct acaacaacga ggtgaccacg gagatccggt 7620
ttgctgactc caagaggaag tcggtcctcc tggacaagat taagaacctt caggtggctc 7680
tgacatccaa acagcagagt ctggagactg ccatgtcgtt tgtggccagg aacacattta 7740
agcgtgtgag gaacggattc ctaatgagga aagtggctgt tttcttcagc aacacaccca 7800
caagagcatc cccacagctc agagaggctg tgctcaagct ctcagatgcg gggatcaccc 7860
ccttgttcct tacaaggcag gaagaccggc agctcatcaa cgctttgcag atcaataaca 7920
cagcagtggg gcatgcgctt gtcctgcctg cagggagaga cctcacagac ttcctggaga 7980
atgtcctcac gtgtcatgtt tgcttggaca tctgcaacat cgacccatcc tgtggatttg 8040
gcagttggag gccttccttc agggacagga gagcggcagg gagcgatgtg gacatcgaca 8100
tggctttcat cttagacagc gctgagacca ccaccctgtt ccagttcaat gagatgaaga 8160
agtacatagc gtacctggtc agacaactgg acatgagccc agatcccaag gcctcccagc 8220
acttcgccag agtggcagtt gtgcagcacg cgccctctga gtccgtggac aatgccagca 8280
tgccacctgt gaaggtggaa ttctccctga ctgactatgg ctccaaggag aagctggtgg 8340
acttcctcag caggggaatg acacagttgc agggaaccag ggccttaggc agtgccattg 8400
aatacaccat agagaatgtc tttgaaagtg ccccaaaccc acgggacctg aaaattgtgg 8460
tcctgatgct gacgggcgag gtgccggagc agcagctgga ggaggcccag agagtcatcc 8520
tgcaggccaa atgcaagggc tacttcttcg tggtcctggg cattggcagg aaggtgaaca 8580
tcaaggaggt atacaccttc gccagtgagc caaacgacgt cttcttcaaa ttagtggaca 8640
agtccaccga gctcaacgag gagcctttga tgcgcttcgg gaggctgttg ccatccttcg 8700
tcagcagtga aaatgctttt tacttgtccc cagatatcag gaaacagtgt gattggttcc 8760
aaggggacca acccacaaag aaccttgtga agtttggtca caaacaagta aatgttccga 8820
ataacgttac ttcaagtcct acatccaacc cagtgacgac aacgaagccg gtgactacga 8880
cgaagccggt gaccaccaca acaaagcctg taaccaccac aacaaagcct gtgactatta 8940
taaatcagcc atctgtgaag ccagccgctg caaagccggc ccctgcgaaa cctgtggctg 9000
ccaagcctgt ggccacaaag atggccactg ttagaccccc agtggcggtg aagccagcaa 9060
cggcagcgaa gcctgtagca gcaaagccag cagctgtaag accccccgct gctgctgctg 9120
caaaaccagt ggcgaccaag cctgaggtcc ctaggccaca ggcagccaaa ccagctgcca 9180
ccaagccagc caccactaag cccatggtta agatgtcccg tgaagtccag gtgtttgaga 9240
taacagagaa cagcgccaaa ctccactggg agagggctga gccccccggt ccttattttt 9300
atgacctcac cgtcacctca gcccatgatc agtccctggt tctgaagcag aacctcacgg 9360
tcacggaccg cgtcattgga ggcctgctcg ctgggcagac ataccatgtg gctgtggtct 9420
gctacctgag gtctcaggtc agagccacct accacggaag tttcagtaca aagaaatctc 9480
agcccccacc tccacagcca gcaaggtcag cttctagttc aaccatcaat ctaatggtga 9540
gcacagaacc attggctctc actgaaacag atatatgcaa gttgccgaaa gacgaaggaa 9600
cttgcaggga tttcatatta aaatggtact atgatccaaa caccaaaagc tgtgcaagat 9660
tctggtatgg aggttgtggt ggaaacgaaa acaaatttgg atcacagaaa gaatgtgaaa 9720
aggtttgcgc tcctgtgctc gccaaacccg gagtcatcag tgtgatggga acctaagcgt 9780
gggtggccaa catcatatac ctcttgaaga agaaggagtc agccatcgcc aacttgtctc 9840
tgtagaagct ccgggtgtag attcccttgc actgtatcat ttcatgcttt gatttacact 9900
cgaactcggg agggaacatc ctgctgcatg acctatcagt atggtgctaa tgtgtctgtg 9960
gaccctcgct ctctgtctcc aggcagttct ctcgaatact ttgaatgttg tgtaacagtt 10020
agccactgct ggtgtttatg tgaacattcc tatcaatcca aattccctct ggagtttcat 10080
gttatgcctg ttgcaggcaa atgtaaagtc tagaaaataa tgcaaatgtc acggctactc 10140
tatatacttt tgcttggttc attttttttc ccttttagtt aagcatgact ttagatggga 10200
agcctgtgta tcgtggagaa acaagagacc aactttttca ttccctgccc ccaatttccc 10260
agactagatt tcaagctaat tttctttttc tgaagcctct aacaaatgat ctagttcaga 10320
aggaagcaaa atcccttaat ctatgtgcac cgttgggacc aatgccttaa ttaaagaatt 10380
taaaaaagtt gtaatagaga atatttttgg cattcctcta atgttgtgtg tttttttttt 10440
gtgtgtgctg gagggagggg atttaatttt aattttaaaa tgtttaggaa atttatacaa 10500
agaaactttt taataaagta tattgaaagt tt 10532
<210> 168
<211> 2259
<212> ДНК
<213> Homo sapiens
<400> 168
cgataaagcc cccgccgccg cggcagccag cttgcgctgt ggggctgccc gggctgcgcg 60
gcgtctgcag gcgccaccgc tgcctctttc cggctgtgac cctcctcgcc gccgccgctt 120
ggctgcgtcc tccgactccc cgcgccgccg agaccaggct cccgctccgg ttgcggccgc 180
accgccctcc gcggccgccc cctggggatc cagcgagcgc ggtcgtcctt ggtggaagga 240
accatgaact ggcatctccc cctcttcctc ttggcctctg tgacgctgcc ttccatctgc 300
tcccacttca atcctctgtc tctcgaggaa ctaggctcca acacggggat ccaggttttc 360
aatcagattg tgaagtcgag gcctcatgac aacatcgtga tctctcccca tgggattgcg 420
tcggtcctgg ggatgcttca gctgggggcg gacggcagga ccaagaagca gctcgccatg 480
gtgatgagat acggcgtaaa tggagttggt aaaatattaa agaagatcaa caaggccatc 540
gtctccaaga agaataaaga cattgtgaca gtggctaacg ccgtgtttgt taagaatgcc 600
tctgaaattg aagtgccttt tgttacaagg aacaaagatg tgttccagtg tgaggtccgg 660
aatgtgaact ttgaggatcc agcctctgcc tgtgattcca tcaatgcatg ggttaaaaat 720
gaaaccaggg atatgattga caatctgctg tccccagatc ttattgatgg tgtgctcacc 780
agactggtcc tcgtcaacgc agtgtatttc aagggtctgt ggaaatcacg gttccaaccc 840
gagaacacaa agaaacgcac tttcgtggca gccgacggga aatcctatca agtgccaatg 900
ctggcccagc tctccgtgtt ccggtgtggg tcgacaagtg cccccaatga tttatggtac 960
aacttcattg aactgcccta ccacggggaa agcatcagca tgctgattgc actgccgact 1020
gagagctcca ctccgctgtc tgccatcatc ccacacatca gcaccaagac catagacagc 1080
tggatgagca tcatggtgcc caagagggtg caggtgatcc tgcccaagtt cacagctgta 1140
gcacaaacag atttgaagga gccgctgaaa gttcttggca ttactgacat gtttgattca 1200
tcaaaggcaa attttgcaaa aataacaaca gggtcagaaa acctccatgt ttctcatatc 1260
ttgcaaaaag caaaaattga agtcagtgaa gatggaacca aagcttcagc agcaacaact 1320
gcaattctca ttgcaagatc atcgcctccc tggtttatag tagacagacc ttttctgttt 1380
ttcatccgac ataatcctac aggtgctgtg ttattcatgg ggcagataaa caaaccctga 1440
agagtataca aaagaaacca tgcaaagcaa cgactacttt gctacgaaga aagactcctt 1500
tcctgcatct ttcatagttc tgttaaatat ttttgtacat cgcttctttt tcaaaactag 1560
ttcttaggaa cagactcgat gcaagtgttt ctgttctggg aggtattgga gggaaaaaac 1620
aagcaggatg gctggaacac tgtactgagg aatgaataga aaggcttcca gatgtctaaa 1680
agattcttta aactactgaa ctgttaccta ggttaacaac cctgttgagt atttgctgtt 1740
tgtccagttc aggaattttt gttttgtttt gtctatatgt gcggcttttc agaagaaatt 1800
taatcagtgt gacagaaaaa aaaatgtttt atggtagctt ttacttttta tgaaaaaaaa 1860
attatttgcc ttttaaattc ttttccccca tccccctcca aagtcttgat agcaagcgtt 1920
attttggggg tagaaacggt gaaatctcta gcctctttgt gtttttgttg ttgttgttgt 1980
tgttgtttta tataatgcat gtattcacta aaataaaatt taaaaaactc ctgtcttgct 2040
agacaaggtt gctgttgtgc agtgtgcctg tcactactgg tctgtactcc ttggatttgc 2100
atttttgtat tttgtacaaa gtaaaaataa actgttatga gtagtaaaaa taaagctatt 2160
tctctgctat ttgaaaatac aatagaagaa actgagcctt ttagacattc gtcagcctct 2220
tctaataaac ctttgtacta tgtaaacatc aggaaattc 2259
<210> 169
<211> 8500
<212> ДНК
<213> Homo sapiens
<400> 169
ggccacagcc tgcctactgt cacccgcctc tcccgcgcgc agatacacgc ccccgcctcc 60
gtgggcacaa aggcagcgct gctggggaac tcgggggaac gcgcacgtgg gaaccgccgc 120
agctccacac tccaggtact tcttccaagg acctaggtct ctcgcccatc ggaaagaaaa 180
taattctttc aagaagatca gggacaactg atttgaagtc tactctgtgc ttctaaatcc 240
ccaattctgc tgaaagtgag ataccctaga gccctagagc cccagcagca cccagccaaa 300
cccacctcca ccatgggggc catgactcag ctgttggcag gtgtctttct tgctttcctt 360
gccctcgcta ccgaaggtgg ggtcctcaag aaagtcatcc ggcacaagcg acagagtggg 420
gtgaacgcca ccctgccaga agagaaccag ccagtggtgt ttaaccacgt ttacaacatc 480
aagctgccag tgggatccca gtgttcggtg gatctggagt cagccagtgg ggagaaagac 540
ctggcaccgc cttcagagcc cagcgaaagc tttcaggagc acacagtgga tggggaaaac 600
cagattgtct tcacacatcg catcaacatc ccccgccggg cctgtggctg tgccgcagcc 660
cctgatgtta aggagctgct gagcagactg gaggagctgg agaacctggt gtcttccctg 720
agggagcaat gtactgcagg agcaggctgc tgtctccagc ctgccacagg ccgcttggac 780
accaggccct tctgtagcgg tcggggcaac ttcagcactg aaggatgtgg ctgtgtctgc 840
gaacctggct ggaaaggccc caactgctct gagcccgaat gtccaggcaa ctgtcacctt 900
cgaggccggt gcattgatgg gcagtgcatc tgtgacgacg gcttcacggg cgaggactgc 960
agccagctgg cttgccccag cgactgcaat gaccagggca agtgcgtaaa tggagtctgc 1020
atctgtttcg aaggctacgc cggggctgac tgcagccgtg aaatctgccc agtgccctgc 1080
agtgaggagc acggcacatg tgtagatggc ttgtgtgtgt gccacgatgg ctttgcaggc 1140
gatgactgca acaagcctct gtgtctcaac aattgctaca accgtggacg atgcgtggag 1200
aatgagtgcg tgtgtgatga gggtttcacg ggcgaagact gcagtgagct catctgcccc 1260
aatgactgct tcgaccgggg ccgctgcatc aatggcacct gctactgcga agaaggcttc 1320
acaggtgaag actgcgggaa acccacctgc ccacatgcct gccacaccca gggccggtgt 1380
gaggaggggc agtgtgtatg tgatgagggc tttgccggtg tggactgcag cgagaagagg 1440
tgtcctgctg actgtcacaa tcgtggccgc tgtgtagacg ggcggtgtga gtgtgatgat 1500
ggtttcactg gagctgactg tggggagctc aagtgtccca atggctgcag tggccatggc 1560
cgctgtgtca atgggcagtg tgtgtgtgat gagggctata ctggggagga ctgcagccag 1620
ctacggtgcc ccaatgactg tcacagtcgg ggccgctgtg tcgagggcaa atgtgtatgt 1680
gagcaaggct tcaagggcta tgactgcagt gacatgagct gccctaatga ctgtcaccag 1740
cacggccgct gtgtgaatgg catgtgtgtt tgtgatgacg gctacacagg ggaagactgc 1800
cgggatcgcc aatgccccag ggactgcagc aacaggggcc tctgtgtgga cggacagtgc 1860
gtctgtgagg acggcttcac cggccctgac tgtgcagaac tctcctgtcc aaatgactgc 1920
catggccagg gtcgctgtgt gaatgggcag tgcgtgtgcc atgaaggatt tatgggcaaa 1980
gactgcaagg agcaaagatg tcccagtgac tgtcatggcc agggccgctg cgtggacggc 2040
cagtgcatct gccacgaggg cttcacaggc ctggactgtg gccagcactc ctgccccagt 2100
gactgcaaca acttaggaca atgcgtctcg ggccgctgca tctgcaacga gggctacagc 2160
ggagaagact gctcagaggt gtctcctccc aaagacctcg ttgtgacaga agtgacggaa 2220
gagacggtca acctggcctg ggacaatgag atgcgggtca cagagtacct tgtcgtgtac 2280
acgcccaccc acgagggtgg tctggaaatg cagttccgtg tgcctgggga ccagacgtcc 2340
accatcatcc aggagctgga gcctggtgtg gagtacttta tccgtgtatt tgccatcctg 2400
gagaacaaga agagcattcc tgtcagcgcc agggtggcca cgtacttacc tgcacctgaa 2460
ggcctgaaat tcaagtccat caaggagaca tctgtggaag tggagtggga tcctctagac 2520
attgcttttg aaacctggga gatcatcttc cggaatatga ataaagaaga tgagggagag 2580
atcaccaaaa gcctgaggag gccagagacc tcttaccggc aaactggtct agctcctggg 2640
caagagtatg agatatctct gcacatagtg aaaaacaata cccggggccc tggcctgaag 2700
agggtgacca ccacacgctt ggatgccccc agccagatcg aggtgaaaga tgtcacagac 2760
accactgcct tgatcacctg gttcaagccc ctggctgaga tcgatggcat tgagctgacc 2820
tacggcatca aagacgtgcc aggagaccgt accaccatcg atctcacaga ggacgagaac 2880
cagtactcca tcgggaacct gaagcctgac actgagtacg aggtgtccct catctcccgc 2940
agaggtgaca tgtcaagcaa cccagccaaa gagaccttca caacaggcct cgatgctccc 3000
aggaatcttc gacgtgtttc ccagacagat aacagcatca ccctggaatg gaggaatggc 3060
aaggcagcta ttgacagtta cagaattaag tatgccccca tctctggagg ggaccacgct 3120
gaggttgatg ttccaaagag ccaacaagcc acaaccaaaa ccacactcac aggtctgagg 3180
ccgggaactg aatatgggat tggagtttct gctgtgaagg aagacaagga gagcaatcca 3240
gcgaccatca acgcagccac agagttggac acgcccaagg accttcaggt ttctgaaact 3300
gcagagacca gcctgaccct gctctggaag acaccgttgg ccaaatttga ccgctaccgc 3360
ctcaattaca gtctccccac aggccagtgg gtgggagtgc agcttccaag aaacaccact 3420
tcctatgtcc tgagaggcct ggaaccagga caggagtaca atgtcctcct gacagccgag 3480
aaaggcagac acaagagcaa gcccgcacgt gtgaaggcat ccactgaaca agcccctgag 3540
ctggaaaacc tcaccgtgac tgaggttggc tgggatggcc tcagactcaa ctggaccgca 3600
gctgaccagg cctatgagca ctttatcatt caggtgcagg aggccaacaa ggtggaggca 3660
gctcggaacc tcaccgtgcc tggcagcctt cgggctgtgg acataccggg cctcaaggct 3720
gctacgcctt atacagtctc catctatggg gtgatccagg gctatagaac accagtgctc 3780
tctgctgagg cctccacagg ggaaactccc aatttgggag aggtcgtggt ggccgaggtg 3840
ggctgggatg ccctcaaact caactggact gctccagaag gggcctatga gtactttttc 3900
attcaggtgc aggaggctga cacagtagag gcagcccaga acctcaccgt cccaggagga 3960
ctgaggtcca cagacctgcc tgggctcaaa gcagccactc attataccat caccatccgc 4020
ggggtcactc aggacttcag cacaacccct ctctctgttg aagtcttgac agaggaggtt 4080
ccagatatgg gaaacctcac agtgaccgag gttagctggg atgctctcag actgaactgg 4140
accacgccag atggaaccta tgaccagttt actattcagg tccaggaggc tgaccaggtg 4200
gaagaggctc acaatctcac ggttcctggc agcctgcgtt ccatggaaat cccaggcctc 4260
agggctggca ctccttacac agtcaccctg cacggcgagg tcaggggcca cagcactcga 4320
ccccttgctg tagaggtcgt cacagaggat ctcccacagc tgggagattt agccgtgtct 4380
gaggttggct gggatggcct cagactcaac tggaccgcag ctgacaatgc ctatgagcac 4440
tttgtcattc aggtgcagga ggtcaacaaa gtggaggcag cccagaacct cacgttgcct 4500
ggcagcctca gggctgtgga catcccgggc ctcgaggctg ccacgcctta tagagtctcc 4560
atctatgggg tgatccgggg ctatagaaca ccagtactct ctgctgaggc ctccacagcc 4620
aaagaacctg aaattggaaa cttaaatgtt tctgacataa ctcccgagag cttcaatctc 4680
tcctggatgg ctaccgatgg gatcttcgag acctttacca ttgaaattat tgattccaat 4740
aggttgctgg agactgtgga atataatatc tctggtgctg aacgaactgc ccatatctca 4800
gggctacccc ctagtactga ttttattgtc tacctctctg gacttgctcc cagcatccgg 4860
accaaaacca tcagtgccac agccacgaca gaggccctgc cccttctgga aaacctaacc 4920
atttccgaca ttaatcccta cgggttcaca gtttcctgga tggcatcgga gaatgccttt 4980
gacagctttc tagtaacggt ggtggattct gggaagctgc tggaccccca ggaattcaca 5040
ctttcaggaa cccagaggaa gctggagctt agaggcctca taactggcat tggctatgag 5100
gttatggtct ctggcttcac ccaagggcat caaaccaagc ccttgagggc tgagattgtt 5160
acagaagccg aaccggaagt tgacaacctt ctggtttcag atgccacccc agacggtttc 5220
cgtctgtcct ggacagctga tgaaggggtc ttcgacaatt ttgttctcaa aatcagagat 5280
accaaaaagc agtctgagcc actggaaata accctacttg cccccgaacg taccagggac 5340
ataacaggtc tcagagaggc tactgaatac gaaattgaac tctatggaat aagcaaagga 5400
aggcgatccc agacagtcag tgctatagca acaacagcca tgggctcccc aaaggaagtc 5460
attttctcag acatcactga aaattcggct actgtcagct ggagggcacc cacagcccaa 5520
gtggagagct tccggattac ctatgtgccc attacaggag gtacaccctc catggtaact 5580
gtggacggaa ccaagactca gaccaggctg gtgaaactca tacctggcgt ggagtacctt 5640
gtcagcatca tcgccatgaa gggctttgag gaaagtgaac ctgtctcagg gtcattcacc 5700
acagctctgg atggcccatc tggcctggtg acagccaaca tcactgactc agaagccttg 5760
gccaggtggc agccagccat tgccactgtg gacagttatg tcatctccta cacaggcgag 5820
aaagtgccag aaattacacg cacggtgtcc gggaacacag tggagtatgc tctgaccgac 5880
ctcgagcctg ccacggaata cacactgaga atctttgcag agaaagggcc ccagaagagc 5940
tcaaccatca ctgccaagtt cacaacagac ctcgattctc caagagactt gactgctact 6000
gaggttcagt cggaaactgc cctccttacc tggcgacccc cccgggcatc agtcaccggt 6060
tacctgctgg tctatgaatc agtggatggc acagtcaagg aagtcattgt gggtccagat 6120
accacctcct acagcctggc agacctgagc ccatccaccc actacacagc caagatccag 6180
gcactcaatg ggcccctgag gagcaatatg atccagacca tcttcaccac aattggactc 6240
ctgtacccct tccccaagga ctgctcccaa gcaatgctga atggagacac gacctctggc 6300
ctctacacca tttatctgaa tggtgataag gctgaggcgc tggaagtctt ctgtgacatg 6360
acctctgatg ggggtggatg gattgtgttc ctgagacgca aaaacggacg cgagaacttc 6420
taccaaaact ggaaggcata tgctgctgga tttggggacc gcagagaaga attctggctt 6480
gggctggaca acctgaacaa aatcacagcc caggggcagt acgagctccg ggtggacctg 6540
cgggaccatg gggagacagc ctttgctgtc tatgacaagt tcagcgtggg agatgccaag 6600
actcgctaca agctgaaggt ggaggggtac agtgggacag caggtgactc catggcctac 6660
cacaatggca gatccttctc cacctttgac aaggacacag attcagccat caccaactgt 6720
gctctgtcct acaaaggggc tttctggtac aggaactgtc accgtgtcaa cctgatgggg 6780
agatatgggg acaataacca cagtcagggc gttaactggt tccactggaa gggccacgaa 6840
cactcaatcc agtttgctga gatgaagctg agaccaagca acttcagaaa tcttgaaggc 6900
aggcgcaaac gggcataaat tccagggacc actgggtgag agaggaataa ggcccagagc 6960
gaggaaagga ttttaccaaa gcatcaatac aaccagccca accatcggtc cacacctggg 7020
catttggtga gagtcaaagc tgaccatgga tccctggggc caacggcaac agcatgggcc 7080
tcacctcctc tgtgatttct ttctttgcac caaagacatc agtctccaac atgtttctgt 7140
tttgttgttt gattcagcaa aaatctccca gtgacaacat cgcaatagtt ttttacttct 7200
cttaggtggc tctgggaatg ggagaggggt aggatgtaca ggggtagttt gttttagaac 7260
cagccgtatt ttacatgaag ctgtataatt aattgtcatt atttttgtta gcaaagatta 7320
aatgtgtcat tggaagccat cccttttttt acatttcata caacagaaac cagaaaagca 7380
atactgtttc cattttaagg atatgattaa tattattaat ataataatga tgatgatgat 7440
gatgaaaact aaggattttt caagagatct ttctttccaa aacatttctg gacagtacct 7500
gattgtattt tttttttaaa taaaagcaca agtacttttg agtttgttat tttgctttga 7560
attgttgagt ctgaatttca ccaaagccaa tcatttgaac aaagcgggga atgttgggat 7620
aggaaaggta agtagggata gtggtcaagt gggaggggtg gaaaggagac taaagactgg 7680
gagagaggga agcacttttt ttaaataaag ttgaacacac ttgggaaaag cttacaggcc 7740
aggcctgtaa tcccaacact ttgggaggcc aaggtgggag gatagcttaa ccccaggagt 7800
ttgagaccag cctgagcaac atagtgagaa cttgtctcta cagaaaaaaa aaaaaaaaaa 7860
aatttaatta ggcaagcgtg gtagtgcgca cctgtcgtcc cagctactca ggaggctgag 7920
gtaggaaaat cactggagcc caggagttag aggttacagt gagctatgat cacactactg 7980
cactccagcc tgggcaacag agggagaccc tgtctctaaa taaaaaaaga aaagaaaaaa 8040
aaagcttaca acttgagatt cagcatcttg ctcagtattt ccaagactaa tagattatgg 8100
tttaaaagat gcttttatac tcattttcta atgcaactcc tagaaactct atgatatagt 8160
tgaggtaagt attgttacca cacatgggct aagatcccca gaggcagact gcctgagttc 8220
aattcttggc tccaccattc ccaagttccc taacctctct atgcctcagt ttcctcttct 8280
gtaaagtagg gacactcata cttctcattt cagaacattt ttgtgaagaa taaattatgt 8340
tatccatttg aggcccttag aatggtaccc ggtgtatatt aagtgctagt acatgttagc 8400
tatcatcatt atcactttat atgagatgga ctggggttca tagaaaccca atgacttgat 8460
tgtggctact actcaataaa taatagaatt tggatttaaa 8500
<210> 170
<211> 2452
<212> ДНК
<213> Homo sapiens
<400> 170
agaaagcgag cagccaccca gctccccgcc accgccatgg tccccgacac cgcctgcgtt 60
cttctgctca ccctggctgc cctcggcgcg tccggacagg gccagagccc gttgggctca 120
gacctgggcc cgcagatgct tcgggaactg caggaaacca acgcggcgct gcaggacgtg 180
cgggagctgc tgcggcagca ggtcagggag atcacgttcc tgaaaaacac ggtgatggag 240
tgtgacgcgt gcgggatgca gcagtcagta cgcaccggcc tacccagcgt gcggcccctg 300
ctccactgcg cgcccggctt ctgcttcccc ggcgtggcct gcatccagac ggagagcggc 360
gcgcgctgcg gcccctgccc cgcgggcttc acgggcaacg gctcgcactg caccgacgtc 420
aacgagtgca acgcccaccc ctgcttcccc cgagtccgct gtatcaacac cagcccgggg 480
ttccgctgcg aggcttgccc gccggggtac agcggcccca cccaccaggg cgtggggctg 540
gctttcgcca aggccaacaa gcaggtttgc acggacatca acgagtgtga gaccgggcaa 600
cataactgcg tccccaactc cgtgtgcatc aacacccggg gctccttcca gtgcggcccg 660
tgccagcccg gcttcgtggg cgaccaggcg tccggctgcc agcggcgcgc acagcgcttc 720
tgccccgacg gctcgcccag cgagtgccac gagcatgcag actgcgtcct agagcgcgat 780
ggctcgcggt cgtgcgtgtg tgccgttggc tgggccggca acgggatcct ctgtggtcgc 840
gacactgacc tagacggctt cccggacgag aagctgcgct gcccggagcg ccagtgccgt 900
aaggacaact gcgtgactgt gcccaactca gggcaggagg atgtggaccg cgatggcatc 960
ggagacgcct gcgatccgga tgccgacggg gacggggtcc ccaatgaaaa ggacaactgc 1020
ccgctggtgc ggaacccaga ccagcgcaac acggacgagg acaagtgggg cgatgcgtgc 1080
gacaactgcc ggtcccagaa gaacgacgac caaaaggaca cagaccagga cggccggggc 1140
gatgcgtgcg acgacgacat cgacggcgac cggatccgca accaggccga caactgccct 1200
agggtaccca actcagacca gaaggacagt gatggcgatg gtatagggga tgcctgtgac 1260
aactgtcccc agaagagcaa cccggatcag gcggatgtgg accacgactt tgtgggagat 1320
gcttgtgaca gcgatcaaga ccaggatgga gacggacatc aggactctcg ggacaactgt 1380
cccacggtgc ctaacagtgc ccaggaggac tcagaccacg atggccaggg tgatgcctgc 1440
gacgacgacg acgacaatga cggagtccct gacagtcggg acaactgccg cctggtgcct 1500
aaccccggcc aggaggacgc ggacagggac ggcgtgggcg acgtgtgcca ggacgacttt 1560
gatgcagaca aggtggtaga caagatcgac gtgtgtccgg agaacgctga agtcacgctc 1620
accgacttca gggccttcca gacagtcgtg ctggacccgg agggtgacgc gcagattgac 1680
cccaactggg tggtgctcaa ccagggaagg gagatcgtgc agacaatgaa cagcgaccca 1740
ggcctggctg tgggttacac tgccttcaat ggcgtggact tcgagggcac gttccatgtg 1800
aacacggtca cggatgacga ctatgcgggc ttcatctttg gctaccagga cagctccagc 1860
ttctacgtgg tcatgtggaa gcagatggag caaacgtatt ggcaggcgaa ccccttccgt 1920
gctgtggccg agcctggcat ccaactcaag gctgtgaagt cttccacagg ccccggggaa 1980
cagctgcgga acgctctgtg gcatacagga gacacagagt cccaggtgcg gctgctgtgg 2040
aaggacccgc gaaacgtggg ttggaaggac aagaagtcct atcgttggtt cctgcagcac 2100
cggccccaag tgggctacat cagggtgcga ttctatgagg gccctgagct ggtggccgac 2160
agcaacgtgg tcttggacac aaccatgcgg ggtggccgcc tgggggtctt ctgcttctcc 2220
caggagaaca tcatctgggc caacctgcgt taccgctgca atgacaccat cccagaggac 2280
tatgagaccc atcagctgcg gcaagcctag ggaccagggt gaggacccgc cggatgacag 2340
ccaccctcac cgcggctgga tgggggctct gcacccagcc ccaaggggtg gccgtcctga 2400
gggggaagtg agaagggctc agagaggaca aaataaagtg tgtgtgcagg ga 2452
<210> 171
<211> 7789
<212> ДНК
<213> Homo sapiens
<400> 171
agccgctgcg cccgagctgg cctgcgagtt cagggctcct gtcgctctcc aggagcaacc 60
tctactccgg acgcacaggc attccccgcg cccctccagc cctcgccgcc ctcgccaccg 120
ctcccggccg ccgcgctccg gtacacacag gatccctgct gggcaccaac agctccacca 180
tggggctggc ctggggacta ggcgtcctgt tcctgatgca tgtgtgtggc accaaccgca 240
ttccagagtc tggcggagac aacagcgtgt ttgacatctt tgaactcacc ggggccgccc 300
gcaaggggtc tgggcgccga ctggtgaagg gccccgaccc ttccagccca gctttccgca 360
tcgaggatgc caacctgatc ccccctgtgc ctgatgacaa gttccaagac ctggtggatg 420
ctgtgcgggc agaaaagggt ttcctccttc tggcatccct gaggcagatg aagaagaccc 480
ggggcacgct gctggccctg gagcggaaag accactctgg ccaggtcttc agcgtggtgt 540
ccaatggcaa ggcgggcacc ctggacctca gcctgaccgt ccaaggaaag cagcacgtgg 600
tgtctgtgga agaagctctc ctggcaaccg gccagtggaa gagcatcacc ctgtttgtgc 660
aggaagacag ggcccagctg tacatcgact gtgaaaagat ggagaatgct gagttggacg 720
tccccatcca aagcgtcttc accagagacc tggccagcat cgccagactc cgcatcgcaa 780
aggggggcgt caatgacaat ttccaggggg tgctgcagaa tgtgaggttt gtctttggaa 840
ccacaccaga agacatcctc aggaacaaag gctgctccag ctctaccagt gtcctcctca 900
cccttgacaa caacgtggtg aatggttcca gccctgccat ccgcactaac tacattggcc 960
acaagacaaa ggacttgcaa gccatctgcg gcatctcctg tgatgagctg tccagcatgg 1020
tcctggaact caggggcctg cgcaccattg tgaccacgct gcaggacagc atccgcaaag 1080
tgactgaaga gaacaaagag ttggccaatg agctgaggcg gcctccccta tgctatcaca 1140
acggagttca gtacagaaat aacgaggaat ggactgttga tagctgcact gagtgtcact 1200
gtcagaactc agttaccatc tgcaaaaagg tgtcctgccc catcatgccc tgctccaatg 1260
ccacagttcc tgatggagaa tgctgtcctc gctgttggcc cagcgactct gcggacgatg 1320
gctggtctcc atggtccgag tggacctcct gttctacgag ctgtggcaat ggaattcagc 1380
agcgcggccg ctcctgcgat agcctcaaca accgatgtga gggctcctcg gtccagacac 1440
ggacctgcca cattcaggag tgtgacaaga gatttaaaca ggatggtggc tggagccact 1500
ggtccccgtg gtcatcttgt tctgtgacat gtggtgatgg tgtgatcaca aggatccggc 1560
tctgcaactc tcccagcccc cagatgaacg ggaaaccctg tgaaggcgaa gcgcgggaga 1620
ccaaagcctg caagaaagac gcctgcccca tcaatggagg ctggggtcct tggtcaccat 1680
gggacatctg ttctgtcacc tgtggaggag gggtacagaa acgtagtcgt ctctgcaaca 1740
accccacacc ccagtttgga ggcaaggact gcgttggtga tgtaacagaa aaccagatct 1800
gcaacaagca ggactgtcca attgatggat gcctgtccaa tccctgcttt gccggcgtga 1860
agtgtactag ctaccctgat ggcagctgga aatgtggtgc ttgtccccct ggttacagtg 1920
gaaatggcat ccagtgcaca gatgttgatg agtgcaaaga agtgcctgat gcctgcttca 1980
accacaatgg agagcaccgg tgtgagaaca cggaccccgg ctacaactgc ctgccctgcc 2040
ccccacgctt caccggctca cagcccttcg gccagggtgt cgaacatgcc acggccaaca 2100
aacaggtgtg caagccccgt aacccctgca cggatgggac ccacgactgc aacaagaacg 2160
ccaagtgcaa ctacctgggc cactatagcg accccatgta ccgctgcgag tgcaagcctg 2220
gctacgctgg caatggcatc atctgcgggg aggacacaga cctggatggc tggcccaatg 2280
agaacctggt gtgcgtggcc aatgcgactt accactgcaa aaaggataat tgccccaacc 2340
ttcccaactc agggcaggaa gactatgaca aggatggaat tggtgatgcc tgtgatgatg 2400
acgatgacaa tgataaaatt ccagatgaca gggacaactg tccattccat tacaacccag 2460
ctcagtatga ctatgacaga gatgatgtgg gagaccgctg tgacaactgt ccctacaacc 2520
acaacccaga tcaggcagac acagacaaca atggggaagg agacgcctgt gctgcagaca 2580
ttgatggaga cggtatcctc aatgaacggg acaactgcca gtacgtctac aatgtggacc 2640
agagagacac tgatatggat ggggttggag atcagtgtga caattgcccc ttggaacaca 2700
atccggatca gctggactct gactcagacc gcattggaga tacctgtgac aacaatcagg 2760
atattgatga agatggccac cagaacaatc tggacaactg tccctatgtg cccaatgcca 2820
accaggctga ccatgacaaa gatggcaagg gagatgcctg tgaccacgat gatgacaacg 2880
atggcattcc tgatgacaag gacaactgca gactcgtgcc caatcccgac cagaaggact 2940
ctgacggcga tggtcgaggt gatgcctgca aagatgattt tgaccatgac agtgtgccag 3000
acatcgatga catctgtcct gagaatgttg acatcagtga gaccgatttc cgccgattcc 3060
agatgattcc tctggacccc aaagggacat cccaaaatga ccctaactgg gttgtacgcc 3120
atcagggtaa agaactcgtc cagactgtca actgtgatcc tggactcgct gtaggttatg 3180
atgagtttaa tgctgtggac ttcagtggca ccttcttcat caacaccgaa agggacgatg 3240
actatgctgg atttgtcttt ggctaccagt ccagcagccg cttttatgtt gtgatgtgga 3300
agcaagtcac ccagtcctac tgggacacca accccacgag ggctcaggga tactcgggcc 3360
tttctgtgaa agttgtaaac tccaccacag ggcctggcga gcacctgcgg aacgccctgt 3420
ggcacacagg aaacacccct ggccaggtgc gcaccctgtg gcatgaccct cgtcacatag 3480
gctggaaaga tttcaccgcc tacagatggc gtctcagcca caggccaaag acgggtttca 3540
ttagagtggt gatgtatgaa gggaagaaaa tcatggctga ctcaggaccc atctatgata 3600
aaacctatgc tggtggtaga ctagggttgt ttgtcttctc tcaagaaatg gtgttcttct 3660
ctgacctgaa atacgaatgt agagatccct aatcatcaaa ttgttgattg aaagactgat 3720
cataaaccaa tgctggtatt gcaccttctg gaactatggg cttgagaaaa cccccaggat 3780
cacttctcct tggcttcctt cttttctgtg cttgcatcag tgtggactcc tagaacgtgc 3840
gacctgcctc aagaaaatgc agttttcaaa aacagactca gcattcagcc tccaatgaat 3900
aagacatctt ccaagcatat aaacaattgc tttggtttcc ttttgaaaaa gcatctactt 3960
gcttcagttg ggaaggtgcc cattccactc tgcctttgtc acagagcagg gtgctattgt 4020
gaggccatct ctgagcagtg gactcaaaag cattttcagg catgtcagag aagggaggac 4080
tcactagaat tagcaaacaa aaccaccctg acatcctcct tcaggaacac ggggagcaga 4140
ggccaaagca ctaaggggag ggcgcatacc cgagacgatt gtatgaagaa aatatggagg 4200
aactgttaca tgttcggtac taagtcattt tcaggggatt gaaagactat tgctggattt 4260
catgatgctg actggcgtta gctgattaac ccatgtaaat aggcacttaa atagaagcag 4320
gaaagggaga caaagactgg cttctggact tcctccctga tccccaccct tactcatcac 4380
ctgcagtggc cagaattagg gaatcagaat caaaccagtg taaggcagtg ctggctgcca 4440
ttgcctggtc acattgaaat tggtggcttc attctagatg tagcttgtgc agatgtagca 4500
ggaaaatagg aaaacctacc atctcagtga gcaccagctg cctcccaaag gaggggcagc 4560
cgtgcttata tttttatggt tacaatggca caaaattatt atcaacctaa ctaaaacatt 4620
ccttttctct tttttcctga attatcatgg agttttctaa ttctctcttt tggaatgtag 4680
atttttttta aatgctttac gatgtaaaat atttattttt tacttattct ggaagatctg 4740
gctgaaggat tattcatgga acaggaagaa gcgtaaagac tatccatgtc atctttgttg 4800
agagtcttcg tgactgtaag attgtaaata cagattattt attaactctg ttctgcctgg 4860
aaatttaggc ttcatacgga aagtgtttga gagcaagtag ttgacattta tcagcaaatc 4920
tcttgcaaga acagcacaag gaaaatcagt ctaataagct gctctgcccc ttgtgctcag 4980
agtggatgtt atgggattct ttttttctct gttttatctt ttcaagtgga attagttggt 5040
tatccatttg caaatgtttt aaattgcaaa gaaagccatg aggtcttcaa tactgtttta 5100
ccccatccct tgtgcatatt tccagggaga aggaaagcat atacactttt ttctttcatt 5160
tttccaaaag agaaaaaaat gacaaaaggt gaaacttaca tacaaatatt acctcatttg 5220
ttgtgtgact gagtaaagaa tttttggatc aagcggaaag agtttaagtg tctaacaaac 5280
ttaaagctac tgtagtacct aaaaagtcag tgttgtacat agcataaaaa ctctgcagag 5340
aagtattccc aataaggaaa tagcattgaa atgttaaata caatttctga aagttatgtt 5400
ttttttctat catctggtat accattgctt tatttttata aattattttc tcattgccat 5460
tggaatagat atctcagatt gtgtagatat gctatttaaa taatttatca ggaaatactg 5520
cctgtagagt tagtatttct atttttatat aatgtttgca cactgaattg aagaattgtt 5580
ggttttttct tttttttgtt ttgttttttt tttttttttt ttttgctttt gacctcccat 5640
ttttactatt tgccaatacc tttttctagg aatgtgcttt tttttgtaca catttttatc 5700
cattttacat tctaaagcag tgtaagttgt atattactgt ttcttatgta caaggaacaa 5760
caataaatca tatggaaatt tatatttata cttactgtat ccatgcttat ttgttctcta 5820
ctggctttat gtcatgaagt atatgcgtaa ataccattca taaatcaata tagcatatac 5880
aaaaataaat tacagtaagt catagcaaca ttcacagttt gtatgtgatt gagaaagact 5940
gagttgctca ggcctaggct tagaatttgc tgcgtttgtg gaataaaaga acaaaatgat 6000
acattagcct gccatatcaa aaacatataa aagagaaatt atccctaagt caagggcccc 6060
cataagaata aaatttctta ttaaggtcat tagatgtcat tgaatccttt tcaaagtgca 6120
gtatgaaaac aaagggaaaa acactgaagc acacgcaact ctcacagcga cattttctga 6180
cccacgaatg atgccttggg tgggcaacac gattgcatgt tgtggagaca cttcggaagt 6240
aaatgtggat gagggaggag ctgtccttgc aatgttgagc caagcattac agatacctcc 6300
tcttgaagaa ggaataataa gtttaatcaa aaaagaagac taaaaaatgt aaaatttgga 6360
aggaatccat aaatgcgtgt gtgtctaaat acaaattatc atgtgaagaa aaggcccaag 6420
tgtaccaata agcagacctt gatttttgga tgggctaatt atgaatgtgg aatactgacc 6480
agttaatttc cagttttaat gaaaacagat caaagaagaa attttatgag taggttaaag 6540
gtctggcttt gaggtctatt aaacactaga aaggactggc tgggtgagat aaaatcttcc 6600
ttgttgattt tcactctcat tctataaata ctcatctttc tgagtagcca tgatcacata 6660
caaatgtaaa ttgccaaatc attttatagt accaaggtga agaagcagga actagaaagt 6720
gttgataata gctgtggagt taggaaaact gatgtgaagg aaataattct ttgaaatggc 6780
aaagaattaa ataccatcat tcattatcag aagagttcaa cgtttgaagt gctgggagat 6840
aattctaatt cattcttgga tagtgaagca aaactgattg aaaataccaa gataagacag 6900
aaaaagtgac tggaaagagg agcttttctt ccaggcatgt tccagtttca ccctaagact 6960
gaccttcaaa taatcaggtt gtactgaaat aaaggacttg ttaaaaatta aaattatgtc 7020
atcgagatga tagctttttt cctcctccaa cagtttattg tcatgtgttg tgggagagct 7080
cgagtgaaga gcaataaact ccaggtctta taagaatgta catacaataa aggtggtgcc 7140
agcagttttt ttttttctaa agagtcacat gtagaaaagc ctccagtatt aagctcctga 7200
attcattcct taaataaatt ggctctctct ctcttctata atttcttttt ctttttattt 7260
ttgagatgaa gtcttgctct gtcgcccagg ctggagtgca gtgacacaat ctcggctcac 7320
tgcaacctct gcctccccgg ttcaagcaat tctccctcct gcctcagcct cccaagtagc 7380
tgggactaca agcgcccgcc accaagcctg gctaattctg tatttttagt aaagacgggg 7440
tttcaccttg ttccggacaa acactaagcc ctaaagggaa atccaaaata aaaacatcta 7500
tttttaataa cactttctat ctaaatcagg gtgacttttt aaaaaaaatc cggaagcttt 7560
ttgttgaatt acgttacaga cttagttacc agtccttgtt agagttacct tcagttgaca 7620
tgctgtgaat ggtcccacct cttttatggc agaattcatt acttaaaata actctatttt 7680
cttccccctt acctaaataa cagaaaggct cactatgtcc caaatatcat tggcagaagc 7740
aaactataaa gtcataagcc ctttgcagtg caagtctaga aataatttt 7789
<210> 172
<211> 7311
<212> ДНК
<213> Homo sapiens
<400> 172
actgacggca tgaagccttt aggggcacac agtactctca gcttgttggt ggaagcccct 60
catctgcctt cattctgaag gcagggcccg gcagaggaag gatcagaggg tcgcggccgg 120
agggtcccgg ccggtggggc caactcagag ggagaggaaa gggctagaga cacgaagaac 180
gcaaaccatc aaatttagaa gaaaaagccc tttgactttt tccccctctc cctccccaat 240
ggctgtgtag caaacatccc tggcgatacc ttggaaagga cgaagttggt ctgcagtcgc 300
aatttcgtgg gttgagttca cagttgtgag tgcggggctc ggagatggag ccgtggtcct 360
ctaggtggaa aacgaaacgg tggctctggg atttcaccgt aacaaccctc gcattgacct 420
tcctcttcca agctagagag gtcagaggag ctgctccagt tgatgtacta aaagcactag 480
attttcacaa ttctccagag ggaatatcaa aaacaacggg attttgcaca aacagaaaga 540
attctaaagg ctcagatact gcttacagag tttcaaagca agcacaactc agtgccccaa 600
caaaacagtt atttccaggt ggaactttcc cagaagactt ttcaatacta tttacagtaa 660
aaccaaaaaa aggaattcag tctttccttt tatctatata taatgagcat ggtattcagc 720
aaattggtgt tgaggttggg agatcacctg tttttctgtt tgaagaccac actggaaaac 780
ctgccccaga agactatccc ctcttcagaa ctgttaacat cgctgacggg aagtggcatc 840
gggtagcaat cagcgtggag aagaaaactg tgacaatgat tgttgattgt aagaagaaaa 900
ccacgaaacc acttgataga agtgagagag caattgttga taccaatgga atcacggttt 960
ttggaacaag gattttggat gaagaagttt ttgaggggga cattcagcag tttttgatca 1020
caggtgatcc caaggcagca tatgactact gtgagcatta tagtccagac tgtgactctt 1080
cagcacccaa ggctgctcaa gctcaggaac ctcagataga tgagtatgca ccagaggata 1140
taatcgaata tgactatgag tatggggaag cagagtataa agaggctgaa agtgtaacag 1200
agggacccac tgtaactgag gagacaatag cacagacgga ggcaaacatc gttgatgatt 1260
ttcaagaata caactatgga acaatggaaa gttaccagac agaagctcct aggcatgttt 1320
ctgggacaaa tgagccaaat ccagttgaag aaatatttac tgaagaatat ctaacgggag 1380
aggattatga ttcccagagg aaaaattctg aggatacact atatgaaaac aaagaaatag 1440
acggcaggga ttctgatctt ctggtagatg gagatttagg cgaatatgat ttttatgaat 1500
ataaagaata tgaagataaa ccaacaagcc cccctaatga agaatttggt ccaggtgtac 1560
cagcagaaac tgatattaca gaaacaagca taaatggcca tggtgcatat ggagagaaag 1620
gacagaaagg agaaccagca gtggttgagc ctggtatgct tgtcgaagga ccaccaggac 1680
cagcaggacc tgcaggtatt atgggtcctc caggtctaca aggccccact ggaccccctg 1740
gtgaccctgg cgataggggc cccccaggac gtcctggctt accaggggct gatggtctac 1800
ctggtcctcc tggtactatg ttgatgttac cgttccgtta tggtggtgat ggttccaaag 1860
gaccaaccat ctctgctcag gaagctcagg ctcaagctat tcttcagcag gctcggattg 1920
ctctgagagg cccacctggc ccaatgggtc taactggaag accaggtcct gtgggggggc 1980
ctggttcatc tggggccaaa ggtgagagtg gtgatccagg tcctcagggc cctcgaggcg 2040
tccagggtcc ccctggtcca acgggaaaac ctggaaaaag gggtcgtcca ggtgcagatg 2100
gaggaagagg aatgccagga gaacctgggg caaagggaga tcgagggttt gatggacttc 2160
cgggtctgcc aggtgacaaa ggtcacaggg gtgaacgagg tcctcaaggt cctccaggtc 2220
ctcctggtga tgatggaatg aggggagaag atggagaaat tggaccaaga ggtcttccag 2280
gtgaagctgg cccacgaggt ttgctgggtc caaggggaac tccaggagct ccagggcagc 2340
ctggtatggc aggtgtagat ggccccccag gaccaaaagg gaacatgggt ccccaagggg 2400
agcctgggcc tccaggtcaa caagggaatc caggacctca gggtcttcct ggtccacaag 2460
gtccaattgg tcctcctggt gaaaaaggac cacaaggaaa accaggactt gctggacttc 2520
ctggtgctga tgggcctcct ggtcatcctg ggaaagaagg ccagtctgga gaaaaggggg 2580
ctctgggtcc ccctggtcca caaggtccta ttggataccc gggcccccgg ggagtaaagg 2640
gagcagatgg tgtcagaggt ctcaagggat ctaaaggtga aaagggtgaa gatggttttc 2700
caggattcaa aggtgacatg ggtctaaaag gtgacagagg agaagttggt caaattggcc 2760
caagagggga agatggccct gaaggaccca aaggtcgagc aggcccaact ggagacccag 2820
gtccttcagg tcaagcagga gaaaagggaa aacttggagt tccaggatta ccaggatatc 2880
caggaagaca aggtccaaag ggttccactg gattccctgg gtttccaggt gccaatggag 2940
agaaaggtgc acggggagta gctggcaaac caggccctcg gggtcagcgt ggtccaacgg 3000
gtcctcgagg ttcaagaggt gcaagaggtc ccactgggaa acctgggcca aagggcactt 3060
caggtggcga tggccctcct ggccctccag gtgaaagagg tcctcaagga cctcagggtc 3120
cagttggatt ccctggacca aaaggccctc ctggaccacc tgggaaggat gggctgccag 3180
gacaccctgg gcaacgtggg gagactggat ttcaaggcaa gaccggccct cctgggccag 3240
ggggagtggt tggaccacag ggaccaaccg gtgagactgg tccaataggg gaacgtgggc 3300
atcctggccc tcctggccct cctggtgagc aaggtcttcc tggtgctgca ggaaaagaag 3360
gtgcaaaggg tgatccaggt cctcaaggta tctcagggaa agatggacca gcaggattac 3420
gtggtttccc aggggaaaga ggtcttcctg gagctcaggg tgcacctgga ctgaaaggag 3480
gggaaggtcc ccagggccca ccaggtccag ttggctcacc aggagaacgt gggtcagcag 3540
gtacagctgg cccaattggt ttaccagggc gcccgggacc tcagggtcct cctggtccag 3600
ctggagagaa aggtgctcct ggagaaaaag gtccccaagg gcctgcaggg agagatggag 3660
ttcaaggtcc tgttggtctc ccagggccag ctggtcctgc cggctcccct ggggaagacg 3720
gagacaaggg tgaaattggt gagccgggac aaaaaggcag caagggtgac aagggagaaa 3780
atggccctcc cggtccccca ggtcttcaag gaccagttgg tgcccctgga attgctggag 3840
gtgatggtga accaggtcct agaggacagc aggggatgtt tgggcaaaaa ggtgatgagg 3900
gtgccagagg cttccctgga cctcctggtc caataggtct tcagggtctg ccaggcccac 3960
ctggtgaaaa aggtgaaaat ggggatgttg gtcccatggg gccacctggt cctccaggcc 4020
caagaggccc tcaaggtccc aatggagctg atggaccaca aggaccccca gggtctgttg 4080
gttcagttgg tggtgttgga gaaaagggtg aacctggaga agcagggaac ccagggcctc 4140
ctggggaagc aggtgtaggc ggtcccaaag gagaaagagg agagaaaggg gaagctggtc 4200
cacctggagc tgctggacct ccaggtgcca aggggccacc aggtgatgat ggccctaagg 4260
gtaacccggg tcctgttggt tttcctggag atcctggtcc tcctggggaa cctggccctg 4320
caggtcaaga tggtgttggt ggtgacaagg gtgaagatgg agatcctggt caaccgggtc 4380
ctcctggccc atctggtgag gctggcccac caggtcctcc tggaaaacga ggtcctcctg 4440
gagctgcagg tgcagaggga agacaaggtg aaaaaggtgc taagggggaa gcaggtgcag 4500
aaggtcctcc tggaaaaacc ggcccagtcg gtcctcaggg acctgcagga aagcctggtc 4560
cagaaggtct tcggggcatc cctggtcctg tgggagaaca aggtctccct ggagctgcag 4620
gccaagatgg accacctggt cctatgggac ctcctggctt acctggtctc aaaggtgacc 4680
ctggctccaa gggtgaaaag ggacatcctg gtttaattgg cctgattggt cctccaggag 4740
aacaagggga aaaaggtgac cgagggctcc ctggaactca aggatctcca ggagcaaaag 4800
gggatggggg aattcctggt cctgctggtc ccttaggtcc acctggtcct ccaggtttac 4860
caggtcctca aggcccaaag ggtaacaaag gctctactgg acccgctggc cagaaaggtg 4920
acagtggtct tccagggcct cctgggtctc caggtccacc tggtgaagtc attcagcctt 4980
taccaatctt gtcctccaaa aaaacgagaa gacatactga aggcatgcaa gcagatgcag 5040
atgataatat tcttgattac tcggatggaa tggaagaaat atttggttcc ctcaattccc 5100
tgaaacaaga cattgagcat atgaaatttc caatgggtac tcagaccaat ccagcccgaa 5160
cttgtaaaga cctgcaactc agccatcctg acttcccaga tggtgaatat tggattgatc 5220
ctaaccaagg ttgctcagga gattccttca aagtttactg taatttcaca tctggtggtg 5280
agacttgcat ttatccagac aaaaaatctg agggagtaag aatttcatca tggccaaagg 5340
agaaaccagg aagttggttt agtgaattta agaggggaaa actgctttca tacttagatg 5400
ttgaaggaaa ttccatcaat atggtgcaaa tgacattcct gaaacttctg actgcctctg 5460
ctcggcaaaa tttcacctac cactgtcatc agtcagcagc ctggtatgat gtgtcatcag 5520
gaagttatga caaagcactt cgcttcctgg gatcaaatga tgaggagatg tcctatgaca 5580
ataatccttt tatcaaaaca ctgtatgatg gttgtgcgtc cagaaaaggc tatgaaaaga 5640
ctgtcattga aatcaataca ccaaaaattg atcaagtacc tattgttgat gtcatgatca 5700
atgactttgg tgatcagaat cagaagttcg gatttgaagt tggtcctgtt tgttttcttg 5760
gctaagatta agacaaagaa catatcaaat caacagaaaa tataccttgg tgccaccaac 5820
ccattttgtg ccacatgcaa gttttgaata aggatggtat agaaaacaac gctgcatata 5880
caggtaccat ttaggaaata ccgatgcctt tgtgggggca gaatcacatg gcaaaagctt 5940
tgaaaatcat aaagatataa gttggtgtgg ctaagatgga aacagggctg attcttgatt 6000
cccaattctc aactctcctt ttcctatttg aatttctttg gtgctgtaga aaacaaaaaa 6060
agaaaaatat atattcataa aaaatatggt gctcattctc atccatccag gatgtactaa 6120
aacagtgtgt ttaataaatt gtaattattt tgtgtacagt tctatactgt tatctgtgtc 6180
catttccaaa acttgcacgt gtccctgaat tccatctgac tctaatttta tgagaattgc 6240
agaactctga tggcaataaa tatatgtatt atgaaaaaat aaagttgtaa tttctgatga 6300
ctctaagtcc ctttctttgg ttaataataa aatgcctttg tatatattga tgttgaagag 6360
ttcaattatt tgatgtcgcc aacaaaattc tcagagggca aaaatctgga agacttttgg 6420
aagcacactc tgatcaactc ttctctgccg acagtcattt tgctgaattt cagccaaaaa 6480
tattatgcat tttgatgctt tattcaaggc tatacctcaa actttttctt ctcagaatcc 6540
aggatttcac aggatacttg tatatatgga aaacaagcaa gtttatattt ttggacaggg 6600
aaatgtgtgt aagaaagtat attaacaaat caatgcctcc gtcaagcaaa caatcatatg 6660
tatacttttt ttctacgtta tctcatctcc ttgttttcag tgtgcttcaa taatgcaggt 6720
taatattaaa gatggaaatt aagcaattat ttatgaattt gtgcaatgtt agattttctt 6780
atcaatcaag ttcttgaatt tgattctaag ttgcatatta taacagtctc gaaaattatt 6840
ttacttgccc aacaaatatt acttttttcc tttcaagata attttataaa tcatttgacc 6900
tacctaattg ctaaatgaat aacatatggt ggactgttat taagagtatt tgttttaagt 6960
cattcaggaa aatctaaact tttttttcca ctaaggtatt tactttaagg tagcttgaaa 7020
tagcaataca atttaaaaat taaaaactga attttgtatc tattttaagt aatatatgta 7080
agacttgaaa ataaatgttt tatttcttat ataaagtgtt aaattaattg ataccagatt 7140
tcactggaac agtttcaact gataatttat gacaaaagaa catacctgta atattgaaat 7200
taaaaagtga aatttgtcat aaagaatttc ttttattttt gaaatcgagt ttgtaaatgt 7260
ccttttaaga agggagatat gaatccaata aataaactca agtcttggct a 7311
<210> 173
<211> 5914
<212> ДНК
<213> Homo sapiens
<400> 173
gcagacggga gtttctcctc ggggtcggag caggaggcac gcggagtgtg aggccacgca 60
tgagcggacg ctaaccccct ccccagccac aaagagtcta catgtctagg gtctagacat 120
gttcagcttt gtggacctcc ggctcctgct cctcttagcg gccaccgccc tcctgacgca 180
cggccaagag gaaggccaag tcgagggcca agacgaagac atcccaccaa tcacctgcgt 240
acagaacggc ctcaggtacc atgaccgaga cgtgtggaaa cccgagccct gccggatctg 300
cgtctgcgac aacggcaagg tgttgtgcga tgacgtgatc tgtgacgaga ccaagaactg 360
ccccggcgcc gaagtccccg agggcgagtg ctgtcccgtc tgccccgacg gctcagagtc 420
acccaccgac caagaaacca ccggcgtcga gggacccaag ggagacactg gcccccgagg 480
cccaagggga cccgcaggcc cccctggccg agatggcatc cctggacagc ctggacttcc 540
cggacccccc ggaccccccg gacctcccgg accccctggc ctcggaggaa actttgctcc 600
ccagctgtct tatggctatg atgagaaatc aaccggagga atttccgtgc ctggccccat 660
gggtccctct ggtcctcgtg gtctccctgg cccccctggt gcacctggtc cccaaggctt 720
ccaaggtccc cctggtgagc ctggcgagcc tggagcttca ggtcccatgg gtccccgagg 780
tcccccaggt ccccctggaa agaatggaga tgatggggaa gctggaaaac ctggtcgtcc 840
tggtgagcgt gggcctcctg ggcctcaggg tgctcgagga ttgcccggaa cagctggcct 900
ccctggaatg aagggacaca gaggtttcag tggtttggat ggtgccaagg gagatgctgg 960
tcctgctggt cctaagggtg agcctggcag ccctggtgaa aatggagctc ctggtcagat 1020
gggcccccgt ggcctgcctg gtgagagagg tcgccctgga gcccctggcc ctgctggtgc 1080
tcgtggaaat gatggtgcta ctggtgctgc cgggccccct ggtcccaccg gccccgctgg 1140
tcctcctggc ttccctggtg ctgttggtgc taagggtgaa gctggtcccc aagggccccg 1200
aggctctgaa ggtccccagg gtgtgcgtgg tgagcctggc ccccctggcc ctgctggtgc 1260
tgctggccct gctggaaacc ctggtgctga tggacagcct ggtgctaaag gtgccaatgg 1320
tgctcctggt attgctggtg ctcctggctt ccctggtgcc cgaggcccct ctggacccca 1380
gggccccggc ggccctcctg gtcccaaggg taacagcggt gaacctggtg ctcctggcag 1440
caaaggagac actggtgcta agggagagcc tggccctgtt ggtgttcaag gaccccctgg 1500
ccctgctgga gaggaaggaa agcgaggagc tcgaggtgaa cccggaccca ctggcctgcc 1560
cggaccccct ggcgagcgtg gtggacctgg tagccgtggt ttccctggcg cagatggtgt 1620
tgctggtccc aagggtcccg ctggtgaacg tggttctcct ggccctgctg gccccaaagg 1680
atctcctggt gaagctggtc gtcccggtga agctggtctg cctggtgcca agggtctgac 1740
tggaagccct ggcagccctg gtcctgatgg caaaactggc ccccctggtc ccgccggtca 1800
agatggtcgc cccggacccc caggcccacc tggtgcccgt ggtcaggctg gtgtgatggg 1860
attccctgga cctaaaggtg ctgctggaga gcccggcaag gctggagagc gaggtgttcc 1920
cggaccccct ggcgctgtcg gtcctgctgg caaagatgga gaggctggag ctcagggacc 1980
ccctggccct gctggtcccg ctggcgagag aggtgaacaa ggccctgctg gctcccccgg 2040
attccagggt ctccctggtc ctgctggtcc tccaggtgaa gcaggcaaac ctggtgaaca 2100
gggtgttcct ggagaccttg gcgcccctgg cccctctgga gcaagaggcg agagaggttt 2160
ccctggcgag cgtggtgtgc aaggtccccc tggtcctgct ggtccccgag gggccaacgg 2220
tgctcccggc aacgatggtg ctaagggtga tgctggtgcc cctggagctc ccggtagcca 2280
gggcgcccct ggccttcagg gaatgcctgg tgaacgtggt gcagctggtc ttccagggcc 2340
taagggtgac agaggtgatg ctggtcccaa aggtgctgat ggctctcctg gcaaagatgg 2400
cgtccgtggt ctgactggcc ccattggtcc tcctggccct gctggtgccc ctggtgacaa 2460
gggtgaaagt ggtcccagcg gccctgctgg tcccactgga gctcgtggtg cccccggaga 2520
ccgtggtgag cctggtcccc ccggccctgc tggctttgct ggcccccctg gtgctgacgg 2580
ccaacctggt gctaaaggcg aacctggtga tgctggtgct aaaggcgatg ctggtccccc 2640
tggccctgcc ggacccgctg gaccccctgg ccccattggt aatgttggtg ctcctggagc 2700
caaaggtgct cgcggcagcg ctggtccccc tggtgctact ggtttccctg gtgctgctgg 2760
ccgagtcggt cctcctggcc cctctggaaa tgctggaccc cctggccctc ctggtcctgc 2820
tggcaaagaa ggcggcaaag gtccccgtgg tgagactggc cctgctggac gtcctggtga 2880
agttggtccc cctggtcccc ctggccctgc tggcgagaaa ggatcccctg gtgctgatgg 2940
tcctgctggt gctcctggta ctcccgggcc tcaaggtatt gctggacagc gtggtgtggt 3000
cggcctgcct ggtcagagag gagagagagg cttccctggt cttcctggcc cctctggtga 3060
acctggcaaa caaggtccct ctggagcaag tggtgaacgt ggtccccctg gtcccatggg 3120
cccccctgga ttggctggac cccctggtga atctggacgt gagggggctc ctggtgccga 3180
aggttcccct ggacgagacg gttctcctgg cgccaagggt gaccgtggtg agaccggccc 3240
cgctggaccc cctggtgctc ctggtgctcc tggtgcccct ggccccgttg gccctgctgg 3300
caagagtggt gatcgtggtg agactggtcc tgctggtccc gccggtcctg tcggccctgt 3360
tggcgcccgt ggccccgccg gaccccaagg cccccgtggt gacaagggtg agacaggcga 3420
acagggcgac agaggcataa agggtcaccg tggcttctct ggcctccagg gtccccctgg 3480
ccctcctggc tctcctggtg aacaaggtcc ctctggagcc tctggtcctg ctggtccccg 3540
aggtccccct ggctctgctg gtgctcctgg caaagatgga ctcaacggtc tccctggccc 3600
cattgggccc cctggtcctc gcggtcgcac tggtgatgct ggtcctgttg gtccccccgg 3660
ccctcctgga cctcctggtc cccctggtcc tcccagcgct ggtttcgact tcagcttcct 3720
gccccagcca cctcaagaga aggctcacga tggtggccgc tactaccggg ctgatgatgc 3780
caatgtggtt cgtgaccgtg acctcgaggt ggacaccacc ctcaagagcc tgagccagca 3840
gatcgagaac atccggagcc cagagggcag ccgcaagaac cccgcccgca cctgccgtga 3900
cctcaagatg tgccactctg actggaagag tggagagtac tggattgacc ccaaccaagg 3960
ctgcaacctg gatgccatca aagtcttctg caacatggag actggtgaga cctgcgtgta 4020
ccccactcag cccagtgtgg cccagaagaa ctggtacatc agcaagaacc ccaaggacaa 4080
gaggcatgtc tggttcggcg agagcatgac cgatggattc cagttcgagt atggcggcca 4140
gggctccgac cctgccgatg tggccatcca gctgaccttc ctgcgcctga tgtccaccga 4200
ggcctcccag aacatcacct accactgcaa gaacagcgtg gcctacatgg accagcagac 4260
tggcaacctc aagaaggccc tgctcctcca gggctccaac gagatcgaga tccgcgccga 4320
gggcaacagc cgcttcacct acagcgtcac tgtcgatggc tgcacgagtc acaccggagc 4380
ctggggcaag acagtgattg aatacaaaac caccaagacc tcccgcctgc ccatcatcga 4440
tgtggccccc ttggacgttg gtgccccaga ccaggaattc ggcttcgacg ttggccctgt 4500
ctgcttcctg taaactccct ccatcccaac ctggctccct cccacccaac caactttccc 4560
cccaacccgg aaacagacaa gcaacccaaa ctgaaccccc tcaaaagcca aaaaatggga 4620
gacaatttca catggacttt ggaaaatatt tttttccttt gcattcatct ctcaaactta 4680
gtttttatct ttgaccaacc gaacatgacc aaaaaccaaa agtgcattca accttaccaa 4740
aaaaaaaaaa aaaaaaagaa taaataaata actttttaaa aaaggaagct tggtccactt 4800
gcttgaagac ccatgcgggg gtaagtccct ttctgcccgt tgggcttatg aaaccccaat 4860
gctgcccttt ctgctccttt ctccacaccc cccttggggc ctcccctcca ctccttccca 4920
aatctgtctc cccagaagac acaggaaaca atgtattgtc tgcccagcaa tcaaaggcaa 4980
tgctcaaaca cccaagtggc ccccaccctc agcccgctcc tgcccgccca gcacccccag 5040
gccctggggg acctggggtt ctcagactgc caaagaagcc ttgccatctg gcgctcccat 5100
ggctcttgca acatctcccc ttcgtttttg agggggtcat gccgggggag ccaccagccc 5160
ctcactgggt tcggaggaga gtcaggaagg gccacgacaa agcagaaaca tcggatttgg 5220
ggaacgcgtg tcaatccctt gtgccgcagg gctgggcggg agagactgtt ctgttccttg 5280
tgtaactgtg ttgctgaaag actacctcgt tcttgtcttg atgtgtcacc ggggcaactg 5340
cctgggggcg gggatggggg cagggtggaa gcggctcccc attttatacc aaaggtgcta 5400
catctatgtg atgggtgggg tggggaggga atcactggtg ctatagaaat tgagatgccc 5460
ccccaggcca gcaaatgttc ctttttgttc aaagtctatt tttattcctt gatatttttc 5520
tttttttttt tttttttttg tggatgggga cttgtgaatt tttctaaagg tgctatttaa 5580
catgggagga gagcgtgtgc ggctccagcc cagcccgctg ctcactttcc accctctctc 5640
cacctgcctc tggcttctca ggcctctgct ctccgacctc tctcctctga aaccctcctc 5700
cacagctgca gcccatcctc ccggctccct cctagtctgt cctgcgtcct ctgtccccgg 5760
gtttcagaga caacttccca aagcacaaag cagtttttcc ccctaggggt gggaggaagc 5820
aaaagactct gtacctattt tgtatgtgta taataatttg agatgttttt aattattttg 5880
attgctggaa taaagcatgt ggaaatgacc caaa 5914
<210> 174
<211> 6829
<212> ДНК
<213> Homo sapiens
<400> 174
gcagactgtg ctggagctgg tgctgaaaaa gggggtttgc agaggctgcc ctggggctgg 60
tgctgaaaga agagcccaca gctgacttca tggtgctaca ataacctcag aatctacttt 120
tcactctcag gagaacccac atgtctaata tttagacatg atggcaaact gggcggaagc 180
aagacctctc ctcattctta ttgttttatt agggcaattt gtctcaataa aagcccagga 240
agaagacgag gatgaaggat atggtgaaga aatagcctgc actcagaatg gccagatgta 300
cttaaacagg gacatttgga aacctgcccc ttgtcagatc tgtgtctgtg acaatggagc 360
cattctctgt gacaagatag aatgccagga tgtgctggac tgtgccgacc ctgtaacgcc 420
ccctggggaa tgctgtcctg tctgttcaca aacacctgga ggtggcaata ccaattttgg 480
tagaggaaga aagggacaaa agggagaacc aggattagtg cctgttgtaa caggcatacg 540
tggtcgtcca ggaccggcag gacctccagg atcacaggga ccaagaggag agcgagggcc 600
aaaaggaaga cctggccctc gtggacctca gggaattgat ggagaaccag gtgttcctgg 660
tcaacctggt gctccaggac ctcctggaca tccgtcccac ccaggacccg atggcttgag 720
caggccgttt tcagctcaaa tggctgggtt ggatgaaaaa tctggacttg ggagtcaagt 780
aggactaatg cctggctctg tgggtcctgt tggcccaagg ggaccacagg gtttacaagg 840
acagcaaggt ggtgcaggac ctacaggacc tcctggtgaa cctggtgatc ctggaccaat 900
gggtccgatt ggttcacgtg gaccagaggg ccctcctggt aaacctgggg aagatggtga 960
acctggcaga aatggaaatc ctggtgaagt gggatttgca ggatctccgg gagctcgtgg 1020
atttcctggg gctcctggtc ttccaggtct gaagggtcac cgaggacaca aaggtcttga 1080
aggccctaaa ggtgaagttg gagcacctgg ttccaagggt gaagctggcc ccactggtcc 1140
aatgggtgcc atgggtcctc tgggtccgag gggaatgcca ggagagagag ggagacttgg 1200
gccacagggt gctcctggac aacgaggtgc acatggtatg cctggaaaac ctggaccaat 1260
gggtcctctt gggataccag gctcttctgg ttttccagga aatcctggaa tgaagggaga 1320
agcaggtcct acaggggcgc gaggccctga aggtcctcag gggcagagag gtgaaactgg 1380
gcccccaggt ccagttggct ctccaggtct tcctggtgca ataggaactg atggtactcc 1440
tggtgccaaa ggcccaacgg gctctccagg tacctctggt cctcctggct cagcagggcc 1500
tcctggatct ccaggacctc agggtagcac tggtcctcag ggaattcgag gccaaccggg 1560
tgatccagga gttccaggtt tcaaaggaga agctggccca aaaggggaac cagggccaca 1620
tggtattcag ggtccgatag gcccacccgg tgaagaaggc aaaagaggtc ccagaggtga 1680
cccaggaaca gttggtcctc cagggccagt gggagaaagg ggtgctcctg gcaatcgtgg 1740
ttttccaggc tctgatggtt tacctgggcc aaagggtgct caaggagaac ggggtcctgt 1800
aggttcttca ggacccaaag gaagccaggg ggatccagga cgtccagggg aacctgggct 1860
tccaggtgct cggggtttga caggaaatcc tggtgttcaa ggtcctgaag gaaaacttgg 1920
acctttgggt gcgccagggg aagatggccg tccaggtcct ccaggctcca taggaatcag 1980
agggcagccc gggagcatgg gccttccagg ccccaaaggt agcagtggtg accctgggaa 2040
acctggagaa gcaggaaatg ctggagttcc tgggcagagg ggagctcctg gaaaagatgg 2100
tgaagttggt ccttctggtc ctgtgggccc gccgggtcta gctggtgaaa gaggagaaca 2160
aggacctcca ggccccacag gttttcaggg gcttcctggt cctccagggc ctcctggaga 2220
aggtggaaaa ccaggtgatc aaggtgttcc tggagatccc ggagcagttg gcccgttagg 2280
acctagagga gaacgaggaa atcctgggga aagaggagaa cctgggataa ctggactccc 2340
tggtgagaag ggaatggctg gaggacatgg tcctgatggc ccaaaaggca gtccaggtcc 2400
atctgggacc cctggagata caggcccacc aggtcttcaa ggtatgccgg gagaaagagg 2460
aattgcagga actcctggcc ccaagggtga cagaggtggc ataggagaaa aaggtgctga 2520
aggcacagct ggaaatgatg gtgcaagagg tcttccaggt cctttgggcc ctccaggtcc 2580
ggcaggtcct actggagaaa agggtgaacc tggtcctcga ggtttagttg gccctcctgg 2640
ctcccggggc aatcctggtt ctcgaggtga aaatgggcca actggagctg ttggttttgc 2700
cggaccccag ggtcctgacg gacagcctgg agtaaaaggt gaacctggag agccaggaca 2760
gaagggagat gctggttctc ctggaccaca aggtttagca ggatcccctg gccctcatgg 2820
tcctaatggt gttcctggac taaaaggtgg tcgaggaacc caaggtccgc ctggtgctac 2880
aggatttcct ggttctgcgg gcagagttgg acctccaggc cctgctggag ctccaggacc 2940
tgcgggaccc ctaggggaac ccgggaagga gggacctcca ggtcttcgtg gggaccctgg 3000
ctctcatggg cgtgtgggag atcgaggacc agctggcccc cctggtggcc caggagacaa 3060
aggggaccca ggagaagatg ggcaacctgg tccagatggc ccccctggtc cagctggaac 3120
gaccgggcag agaggaattg ttggcatgcc tgggcaacgt ggagagagag gcatgcccgg 3180
cctaccaggc ccagcgggaa caccaggaaa agtaggacca actggtgcaa caggagataa 3240
aggtccacct ggacctgtgg ggcccccagg ctccaatggt cctgtagggg aacctggacc 3300
agaaggtcca gctggcaatg atggtacccc aggacgggat ggtgctgttg gagaacgtgg 3360
tgatcgtgga gaccctgggc ctgcaggtct gccaggctct cagggtgccc ctggaactcc 3420
tggccctgtg ggtgctccag gagatgcagg acaaagagga gatccgggtt ctcggggtcc 3480
tataggacca cctggtcgag ctgggaaacg tggattacct ggaccccaag gacctcgtgg 3540
tgacaaaggt gatcatggag accgaggtga cagaggtcag aagggccaca gaggctttac 3600
tggtcttcag ggtcttcctg gccctcctgg tccaaatggt gaacaaggaa gtgctggaat 3660
ccctggacca tttggcccaa gaggtcctcc aggcccagtt ggtccttcag gtaaagaagg 3720
aaaccctggg ccacttgggc caattggacc tccaggtgta cgaggcagtg taggagaagc 3780
aggacctgag ggccctcctg gtgagcctgg cccacctggc cctccgggtc cccctggcca 3840
ccttacagct gctcttgggg atatcatggg gcactatgat gaaagcatgc cagatccact 3900
tcctgagttt actgaagatc aggcggctcc tgatgacaaa aacaaaacgg acccaggggt 3960
tcatgctacc ctgaagtcac tcagtagtca gattgaaacc atgcgcagcc ccgatggctc 4020
gaaaaagcac ccagcccgca cgtgtgatga cctaaagctt tgccattccg caaagcagag 4080
tggtgaatac tggattgatc ctaaccaagg atctgttgaa gatgcaatca aagtttactg 4140
caacatggaa acaggagaaa catgtatttc agcaaaccca tccagtgtac cacgtaaaac 4200
ctggtgggcc agtaaatctc ctgacaataa acctgtttgg tatggtcttg atatgaacag 4260
agggtctcag ttcgcttatg gagaccacca atcacctaat acagccatta ctcagatgac 4320
ttttttgcgc cttttatcaa aagaagcctc ccagaacatc acttacatct gtaaaaacag 4380
tgtaggatac atggacgatc aagctaagaa cctcaaaaaa gctgtggttc tcaaaggggc 4440
aaatgactta gatatcaaag cagagggaaa tattagattc cggtatatcg ttcttcaaga 4500
cacttgctct aagcggaatg gaaatgtggg caagactgtc tttgaatata gaacacagaa 4560
tgtggcacgc ttgcccatca tagatcttgc tcctgtggat gttggcggca cagaccagga 4620
attcggcgtt gaaattgggc cagtttgttt tgtgtaaagt aagccaagac acatcgacaa 4680
tgagcaccac catcaatgac caccgccatt cacaagaact ttgactgttt gaagttgatc 4740
ctgagactct tgaagtaatg gctgatcctg catcagcatt gtatatatgg tcttaagtgc 4800
ctggcctcct tatccttcag aatatttatt ttacttacaa tcctcaagtt ttaattgatt 4860
ttaaatattt ttcaatacaa cagtttaggt ttaagatgac caatgacaat gaccaccttt 4920
gcagaaagta aactgattga ataaataaat ctccgttttc ttcaatttat ttcagtgtaa 4980
tgaaaaagtt gcttagtatt tatgaggaaa ttcttcttcc tggcaggtag cttaaagagt 5040
ggggtatata gagccacaac acatgtttat tttgcttggc tgcagttgaa aaatagaaat 5100
tagtgccctt ttgtgacctc tcattccaag attgtcaatt aaaaatgagt ttaaaatgtt 5160
taacttgtga tcgagaccta catgcatgtc ttgatattgt gtaactataa tagagactct 5220
ttaaggagaa tcttaaaaaa aaaaaaacgt ttctcactgt cttaaataga atttttaaat 5280
agtatatatt cagtggcatt ttggagaaca aagtgaattt acttcgactt cttaaatttt 5340
tgtaaaagac tataagttta gacatctttc tcattcaaat ttaaagatat ctttctcctc 5400
ttgatcaatc tatcaatatt gatagaagtc acactagtat ataccattta atacatttac 5460
actttcttat ttaagaagat attgaatgca aaataattga catatagaac tttacaaaca 5520
tatgtccaag gactctaaat tgagactctt ccacatgtac aatctcatca tcctgaagcc 5580
tataatgaag aaaaagatct agaaactgag ttgtggagct gactctaatc aaatgtgatg 5640
attggaatta gaccatttgg cctttgaact ttcataggaa aaatgaccca acatttctta 5700
gcatgagcta cctcatctct agaagctggg atggacttac tattcttgtt tatattttag 5760
atactgaaag gtgctatgct tctgttatta ttccaagact ggagataggc agggctaaaa 5820
aggtattatt atttttcctt taatgatggt gctaaaattc ttcctataaa attccttaaa 5880
aataaagatg gtttaatcac taccattgtg aaaacataac tgttagactt cccgtttctg 5940
aaagaaagag catcgttcca atgcttgttc actgttcctc tgtcatactg tatctggaat 6000
gctttgtaat acttgcatgc ttcttagacc agaacatgta ggtccccttg tgtctcaata 6060
cttttttttt cttaattgca tttgttggct ctattttaat ttttttcttt taaaataaac 6120
agctgggacc atcccaaaag acaagccatg catacaactt tggtcatgta tctctgcaaa 6180
gcatcaaatt aaatgcacgc ttttgtcatg tcagtggttt ttgttttgtg aaattccttt 6240
gaccatatta gatctatttc atttccaata gtgaaaagga gatgtggtgg tatactttgt 6300
ttgccatttg tttaaaagat acaacggata ccttctatca tgtatgtact ggcttataaa 6360
tgaaaatcta tctacaacat tacccacaaa ggcaacatga caccaattat cactgcctct 6420
gcccttaaaa atgtcagagt agtattattg ataaaaaggg caagcaatag atttttcatg 6480
actgaataaa ctgtaataat aaaacatatg tctcaaagtg tatcacatat gaatttagcc 6540
taattgtttt cagtttcatt ctcaatattt agtttacaac atcattttcc cctaaactgg 6600
ttatattttg acctgtatat cttaaatttg agtatttata tgcctaaata catgtgtgag 6660
ttttgtttga cttccaagtc caaactataa gattatataa gttcatatag atgaatcaga 6720
aatatgtggt aatactatta agtcacaaac actaacaatt tccaactata gaaataacag 6780
ttcttatttg gattttggga atgctaccaa taaaagcctg cccagacca 6829
<210> 175
<211> 5072
<212> ДНК
<213> Homo sapiens
<400> 175
agcaccacgg cagcaggagg tttcggctaa gttggaggta ctggccacga ctgcatgccc 60
gcgcccgcca ggtgatacct ccgccggtga cccaggggct ctgcgacaca aggagtctgc 120
atgtctaagt gctagacatg ctcagctttg tggatacgcg gactttgttg ctgcttgcag 180
taaccttatg cctagcaaca tgccaatctt tacaagagga aactgtaaga aagggcccag 240
ccggagatag aggaccacgt ggagaaaggg gtccaccagg ccccccaggc agagatggtg 300
aagatggtcc cacaggccct cctggtccac ctggtcctcc tggcccccct ggtctcggtg 360
ggaactttgc tgctcagtat gatggaaaag gagttggact tggccctgga ccaatgggct 420
taatgggacc tagaggccca cctggtgcag ctggagcccc aggccctcaa ggtttccaag 480
gacctgctgg tgagcctggt gaacctggtc aaactggtcc tgcaggtgct cgtggtccag 540
ctggccctcc tggcaaggct ggtgaagatg gtcaccctgg aaaacccgga cgacctggtg 600
agagaggagt tgttggacca cagggtgctc gtggtttccc tggaactcct ggacttcctg 660
gcttcaaagg cattagggga cacaatggtc tggatggatt gaagggacag cccggtgctc 720
ctggtgtgaa gggtgaacct ggtgcccctg gtgaaaatgg aactccaggt caaacaggag 780
cccgtgggct tcctggtgag agaggacgtg ttggtgcccc tggcccagct ggtgcccgtg 840
gcagtgatgg aagtgtgggt cccgtgggtc ctgctggtcc cattgggtct gctggccctc 900
caggcttccc aggtgcccct ggccccaagg gtgaaattgg agctgttggt aacgctggtc 960
ctgctggtcc cgccggtccc cgtggtgaag tgggtcttcc aggcctctcc ggccccgttg 1020
gacctcctgg taatcctgga gcaaacggcc ttactggtgc caagggtgct gctggccttc 1080
ccggcgttgc tggggctccc ggcctccctg gaccccgcgg tattcctggc cctgttggtg 1140
ctgccggtgc tactggtgcc agaggacttg ttggtgagcc tggtccagct ggctccaaag 1200
gagagagcgg taacaagggt gagcccggct ctgctgggcc ccaaggtcct cctggtccca 1260
gtggtgaaga aggaaagaga ggccctaatg gggaagctgg atctgccggc cctccaggac 1320
ctcctgggct gagaggtagt cctggttctc gtggtcttcc tggagctgat ggcagagctg 1380
gcgtcatggg ccctcctggt agtcgtggtg caagtggccc tgctggagtc cgaggaccta 1440
atggagatgc tggtcgccct ggggagcctg gtctcatggg acccagaggt cttcctggtt 1500
cccctggaaa tatcggcccc gctggaaaag aaggtcctgt cggcctccct ggcatcgacg 1560
gcaggcctgg cccaattggc ccagctggag caagaggaga gcctggcaac attggattcc 1620
ctggacccaa aggccccact ggtgatcctg gcaaaaacgg tgataaaggt catgctggtc 1680
ttgctggtgc tcggggtgct ccaggtcctg atggaaacaa tggtgctcag ggacctcctg 1740
gaccacaggg tgttcaaggt ggaaaaggtg aacagggtcc ccctggtcct ccaggcttcc 1800
agggtctgcc tggcccctca ggtcccgctg gtgaagttgg caaaccagga gaaaggggtc 1860
tccatggtga gtttggtctc cctggtcctg ctggtccaag aggggaacgc ggtcccccag 1920
gtgagagtgg tgctgccggt cctactggtc ctattggaag ccgaggtcct tctggacccc 1980
cagggcctga tggaaacaag ggtgaacctg gtgtggttgg tgctgtgggc actgctggtc 2040
catctggtcc tagtggactc ccaggagaga ggggtgctgc tggcatacct ggaggcaagg 2100
gagaaaaggg tgaacctggt ctcagaggtg aaattggtaa ccctggcaga gatggtgctc 2160
gtggtgctcc tggtgctgta ggtgcccctg gtcctgctgg agccacaggt gaccggggcg 2220
aagctggggc tgctggtcct gctggtcctg ctggtcctcg gggaagccct ggtgaacgtg 2280
gtgaggtcgg tcctgctggc cccaatggat ttgctggtcc tgctggtgct gctggtcaac 2340
ctggtgctaa aggagaaaga ggagccaaag ggcctaaggg tgaaaacggt gttgttggtc 2400
ccacaggccc cgttggagct gctggcccag ctggtccaaa tggtcccccc ggtcctgctg 2460
gaagtcgtgg tgatggaggc ccccctggta tgactggttt ccctggtgct gctggacgga 2520
ctggtccccc aggaccctct ggtatttctg gccctcctgg tccccctggt cctgctggga 2580
aagaagggct tcgtggtcct cgtggtgacc aaggtccagt tggccgaact ggagaagtag 2640
gtgcagttgg tccccctggc ttcgctggtg agaagggtcc ctctggagag gctggtactg 2700
ctggacctcc tggcactcca ggtcctcagg gtcttcttgg tgctcctggt attctgggtc 2760
tccctggctc gagaggtgaa cgtggtctac caggtgttgc tggtgctgtg ggtgaacctg 2820
gtcctcttgg cattgccggc cctcctgggg cccgtggtcc tcctggtgct gtgggtagtc 2880
ctggagtcaa cggtgctcct ggtgaagctg gtcgtgatgg caaccctggg aacgatggtc 2940
ccccaggtcg cgatggtcaa cccggacaca agggagagcg cggttaccct ggcaatattg 3000
gtcccgttgg tgctgcaggt gcacctggtc ctcatggccc cgtgggtcct gctggcaaac 3060
atggaaaccg tggtgaaact ggtccttctg gtcctgttgg tcctgctggt gctgttggcc 3120
caagaggtcc tagtggccca caaggcattc gtggcgataa gggagagccc ggtgaaaagg 3180
ggcccagagg tcttcctggc ttaaagggac acaatggatt gcaaggtctg cctggtatcg 3240
ctggtcacca tggtgatcaa ggtgctcctg gctccgtggg tcctgctggt cctaggggcc 3300
ctgctggtcc ttctggccct gctggaaaag atggtcgcac tggacatcct ggtacagttg 3360
gacctgctgg cattcgaggc cctcagggtc accaaggccc tgctggcccc cctggtcccc 3420
ctggccctcc tggacctcca ggtgtaagcg gtggtggtta tgactttggt tacgatggag 3480
acttctacag ggctgaccag cctcgctcag caccttctct cagacccaag gactatgaag 3540
ttgatgctac tctgaagtct ctcaacaacc agattgagac ccttcttact cctgaaggct 3600
ctagaaagaa cccagctcgc acatgccgtg acttgagact cagccaccca gagtggagca 3660
gtggttacta ctggattgac cctaaccaag gatgcactat ggatgctatc aaagtatact 3720
gtgatttctc tactggcgaa acctgtatcc gggcccaacc tgaaaacatc ccagccaaga 3780
actggtatag gagctccaag gacaagaaac acgtctggct aggagaaact atcaatgctg 3840
gcagccagtt tgaatataat gtagaaggag tgacttccaa ggaaatggct acccaacttg 3900
ccttcatgcg cctgctggcc aactatgcct ctcagaacat cacctaccac tgcaagaaca 3960
gcattgcata catggatgag gagactggca acctgaaaaa ggctgtcatt ctacagggct 4020
ctaatgatgt tgaacttgtt gctgagggca acagcaggtt cacttacact gttcttgtag 4080
atggctgctc taaaaagaca aatgaatggg gaaagacaat cattgaatac aaaacaaata 4140
agccatcacg cctgcccttc cttgatattg cacctttgga catcggtggt gctgaccagg 4200
aattctttgt ggacattggc ccagtctgtt tcaaataaat gaactcaatc taaattaaaa 4260
aagaaagaaa tttgaaaaaa ctttctcttt gccatttctt cttcttcttt tttaactgaa 4320
agctgaatcc ttccatttct tctgcacatc tacttgctta aattgtgggc aaaagagaaa 4380
aagaaggatt gatcagagca ttgtgcaata cagtttcatt aactccttcc cccgctcccc 4440
caaaaatttg aatttttttt tcaacactct tacacctgtt atggaaaatg tcaacctttg 4500
taagaaaacc aaaataaaaa ttgaaaaata aaaaccataa acatttgcac cacttgtggc 4560
ttttgaatat cttccacaga gggaagttta aaacccaaac ttccaaaggt ttaaactacc 4620
tcaaaacact ttcccatgag tgtgatccac attgttaggt gctgacctag acagagatga 4680
actgaggtcc ttgttttgtt ttgttcataa tacaaaggtg ctaattaata gtatttcaga 4740
tacttgaaga atgttgatgg tgctagaaga atttgagaag aaatactcct gtattgagtt 4800
gtatcgtgtg gtgtattttt taaaaaattt gatttagcat tcatattttc catcttattc 4860
ccaattaaaa gtatgcagat tatttgccca aatcttcttc agattcagca tttgttcttt 4920
gccagtctca ttttcatctt cttccatggt tccacagaag ctttgtttct tgggcaagca 4980
gaaaaattaa attgtaccta ttttgtatat gtgagatgtt taaataaatt gtgaaaaaaa 5040
tgaaataaag catgtttggt tttccaaaag aa 5072
<210> 176
<211> 6540
<212> ДНК
<213> Homo sapiens
<400> 176
aggtctccgc ttggagccgc cgcacccggg acggtgcgta gcgctggaag tccggccttc 60
cgagagctag ctgtccgccg cggcccccgc acgccgggca gccgtccctc gccgcctcgg 120
gcgcgccacc atggggcccc ggctcagcgt ctggctgctg ctgctgcccg ccgcccttct 180
gctccacgag gagcacagcc gggccgctgc gaagggtggc tgtgctggct ctggctgtgg 240
caaatgtgac tgccatggag tgaagggaca aaagggtgaa agaggcctcc cggggttaca 300
aggtgtcatt gggtttcctg gaatgcaagg acctgagggg ccacagggac caccaggaca 360
aaagggtgat actggagaac caggactacc tggaacaaaa gggacaagag gacctccggg 420
agcatctggc taccctggaa acccaggact tcccggaatt cctggccaag acggcccgcc 480
aggcccccca ggtattccag gatgcaatgg cacaaagggg gagagagggc cgctcgggcc 540
tcctggcttg cctggtttcg ctggaaatcc cggaccacca ggcttaccag ggatgaaggg 600
tgatccaggt gagatacttg gccatgtgcc cgggatgctg ttgaaaggtg aaagaggatt 660
tcccggaatc ccagggactc caggcccacc aggactgcca gggcttcaag gtcctgttgg 720
gcctccagga tttaccggac caccaggtcc cccaggccct cccggccctc caggtgaaaa 780
gggacaaatg ggcttaagtt ttcaaggacc aaaaggtgac aagggtgacc aaggggtcag 840
tgggcctcca ggagtaccag gacaagctca agttcaagaa aaaggagact tcgccaccaa 900
gggagaaaag ggccaaaaag gtgaacctgg atttcagggg atgccagggg tcggagagaa 960
aggtgaaccc ggaaaaccag gacccagagg caaacccgga aaagatggtg acaaagggga 1020
aaaagggagt cccggttttc ctggtgaacc cgggtaccca ggactcatag gccgccaggg 1080
cccgcaggga gaaaagggtg aagcaggtcc tcctggccca cctggaattg ttataggcac 1140
aggacctttg ggagaaaaag gagagagggg ctaccctgga actccggggc caagaggaga 1200
gccaggccca aaaggtttcc caggactacc aggccaaccc ggacctccag gcctccctgt 1260
acctgggcag gctggtgccc ctggcttccc tggtgaaaga ggagaaaaag gtgaccgagg 1320
atttcctggt acatctctgc caggaccaag tggaagagat gggctcccgg gtcctcctgg 1380
ttcccctggg ccccctgggc agcctggcta cacaaatgga attgtggaat gtcagcccgg 1440
acctccaggt gaccagggtc ctcctggaat tccagggcag ccaggattta taggcgaaat 1500
tggagagaaa ggtcaaaaag gagagagttg cctcatctgt gatatagacg gatatcgggg 1560
gcctcccggg ccacagggac ccccgggaga aataggtttc ccagggcagc caggggccaa 1620
gggcgacaga ggtttgcctg gcagagatgg tgttgcagga gtgccaggcc ctcaaggtac 1680
accagggctg ataggccagc caggagccaa gggggagcct ggtgagtttt atttcgactt 1740
gcggctcaaa ggtgacaaag gagacccagg ctttccagga cagcccggca tgccagggag 1800
agcgggttct cctggaagag atggccatcc gggtcttcct ggccccaagg gctcgccggg 1860
ttctgtagga ttgaaaggag agcgtggccc ccctggagga gttggattcc caggcagtcg 1920
tggtgacacc ggcccccctg ggcctccagg atatggtcct gctggtccca ttggtgacaa 1980
aggacaagca ggctttcctg gaggccctgg atccccaggc ctgccaggtc caaagggtga 2040
accaggaaaa attgttcctt taccaggccc ccctggagca gaaggactgc cggggtcccc 2100
aggcttccca ggtccccaag gagaccgagg ctttcccgga accccaggaa ggccaggcct 2160
gccaggagag aagggcgctg tgggccagcc aggcattgga tttccagggc cccccggccc 2220
caaaggtgtt gacggcttac ctggagacat ggggccaccg gggactccag gtcgcccggg 2280
atttaatggc ttacctggga acccaggtgt gcagggccag aagggagagc ctggagttgg 2340
tctaccggga ctcaaaggtt tgccaggtct tcccggcatt cctggcacac ccggggagaa 2400
ggggagcatt ggggtaccag gcgttcctgg agaacatgga gcgatcggac cccctgggct 2460
tcaggggatc agaggtgaac cgggacctcc tggattgcca ggctccgtgg ggtctccagg 2520
agttccagga ataggccccc ctggagctag gggtccccct ggaggacagg gaccaccggg 2580
gttgtcaggc cctcctggaa taaaaggaga gaagggtttc cccggattcc ctggactgga 2640
catgccgggc cctaaaggag ataaaggggc tcaaggactc cctggcataa cgggacagtc 2700
ggggctccct ggccttcctg gacagcaggg ggctcctggg attcctgggt ttccaggttc 2760
caagggagaa atgggcgtca tggggacccc cgggcagccg ggctcaccag gaccagtggg 2820
tgctcctgga ttaccgggtg aaaaagggga ccatggcttt ccgggctcct caggacccag 2880
gggagaccct ggcttgaaag gtgataaggg ggatgtcggt ctccctggca agcctggctc 2940
catggataag gtggacatgg gcagcatgaa gggccagaaa ggagaccaag gagagaaagg 3000
acaaattgga ccaattggtg agaagggatc ccgaggagac cctgggaccc caggagtgcc 3060
tggaaaggac gggcaggcag gacagcctgg gcagccagga cctaaaggtg atccaggtat 3120
aagtggaacc ccaggtgctc caggacttcc gggaccaaaa ggatctgttg gtggaatggg 3180
cttgccagga acacctggag agaaaggtgt gcctggcatc cctggcccac aaggttcacc 3240
tggcttacct ggagacaaag gtgcaaaagg agagaaaggg caggcaggcc cacctggcat 3300
aggcatccca gggctgcgag gtgaaaaggg agatcaaggg atagcgggtt tcccaggaag 3360
ccctggagag aagggagaaa aaggaagcat tgggatccca ggaatgccag ggtccccagg 3420
ccttaaaggg tctcccggga gtgttggcta tccaggaagt cctgggctac ctggagaaaa 3480
aggtgacaaa ggcctcccag gattggatgg catccctggt gtcaaaggag aagcaggtct 3540
tcctgggact cctggcccca caggcccagc tggccagaaa ggggagccag gcagtgatgg 3600
aatcccgggg tcagcaggag agaagggtga accaggtcta ccaggaagag gattcccagg 3660
gtttccaggg gccaaaggag acaaaggttc aaagggtgag gtgggtttcc caggattagc 3720
cgggagccca ggaattcctg gatccaaagg agagcaagga ttcatgggtc ctccggggcc 3780
ccagggacag ccggggttac cgggatcccc aggccatgcc acggaggggc ccaaaggaga 3840
ccgcggacct cagggccagc ctggcctgcc aggacttccg ggacccatgg ggcctccagg 3900
gcttcctggg attgatggag ttaaaggtga caaaggaaat ccaggctggc caggagcacc 3960
cggtgtccca gggcccaagg gagaccctgg attccagggc atgcctggta ttggtggctc 4020
tccaggaatc acaggctcta agggtgatat ggggcctcca ggagttccag gatttcaagg 4080
tccaaaaggt cttcctggcc tccagggaat taaaggtgat caaggcgatc aaggcgtccc 4140
gggagctaaa ggtctcccgg gtcctcctgg ccccccaggt ccttacgaca tcatcaaagg 4200
ggagcccggg ctccctggtc ctgagggccc cccagggctg aaagggcttc agggactgcc 4260
aggcccgaaa ggccagcaag gtgttacagg attggtgggt atacctggac ctccaggtat 4320
tcctgggttt gacggtgccc ctggccagaa aggagagatg ggacctgccg ggcctactgg 4380
tccaagagga tttccaggtc caccaggccc cgatgggttg ccaggatcca tggggccccc 4440
aggcacccca tctgttgatc acggcttcct tgtgaccagg catagtcaaa caatagatga 4500
cccacagtgt ccttctggga ccaaaattct ttaccacggg tactctttgc tctacgtgca 4560
aggcaatgaa cgggcccatg gccaggactt gggcacggcc ggcagctgcc tgcgcaagtt 4620
cagcacaatg cccttcctgt tctgcaatat taacaacgtg tgcaactttg catcacgaaa 4680
tgactactcg tactggctgt ccacccctga gcccatgccc atgtcaatgg cacccatcac 4740
gggggaaaac ataagaccat ttattagtag gtgtgctgtg tgtgaggcgc ctgccatggt 4800
gatggccgtg cacagccaga ccattcagat cccaccgtgc cccagcgggt ggtcctcgct 4860
gtggatcggc tactcttttg tgatgcacac cagcgctggt gcagaaggct ctggccaagc 4920
cctggcgtcc cccggctcct gcctggagga gtttagaagt gcgccattca tcgagtgtca 4980
cggccgtggg acctgcaatt actacgcaaa cgcttacagc ttttggctcg ccaccataga 5040
gaggagcgag atgttcaaga agcctacgcc gtccaccttg aaggcagggg agctgcgcac 5100
gcacgtcagc cgctgccaag tctgtatgag aagaacataa tgaagcctga ctcagctaat 5160
gtcacaacat ggtgctactt cttcttcttt ttgttaacag caacgaaccc tagaaatata 5220
tcctgtgtac ctcactgtcc aatatgaaaa ccgtaaagtg ccttatagga atttgcgtaa 5280
ctaacacacc ctgcttcatt gacctctact tgctgaagga gaaaaagaca gcgataagct 5340
ttcaatagtg gcataccaaa tggcactttt gatgaaataa aatatcaata ttttctgcaa 5400
tccaatgcac tgatgtgtga agtgagaact ccatcagaaa accaaagggt gctaggaggt 5460
gtgggtgcct tccatactgt ttgcccattt tcattcttgt attataatta attttctacc 5520
cccagagata aatgtttgtt tatatcactg tctagctgtt tcaaaattta ggtcccttgg 5580
tctgtacaaa taatagcaat gtaaaaatgg ttttttgaac ctccaaatgg aattacagac 5640
tcagtagcca tatcttccaa ccccccagta taaatttctg tctttctgct atgtgtggta 5700
ctttgcagct gcttttgcag aaatcacaat tttcctgtgg aataaagatg gtccaaaaat 5760
agtcaaaaat taaatatata tatatattag taatttatat agatgtcagc aattaggcag 5820
atcaaggttt agtttaactt ccactgttaa aataaagctt acatagtttt cttcctttga 5880
aagactgtgc tgtcctttaa cataggtttt taaagactag gatattgaat gtgaaacatc 5940
cgttttcatt gttcacttct aaaccaaaaa ttatgtgttg ccaaaaccaa acccaggttc 6000
atgaatatgg tgtctattat agtgaaacat gtactttgag cttattgttt ttattctgta 6060
ttaaatattt tcagggtttt aaacactaat cacaaactga atgacttgac ttcaaaagca 6120
acaaccttaa aggccgtcat ttcattagta ttcctcattc tgcatcctgg cttgaaaaac 6180
agctctgttg aatcacagta tcagtatttt cacacgtaag cacattcggg ccatttccgt 6240
ggtttctcat gagctgtgtt cacagacctc agcagggcat cgcatggacc gcaggagggc 6300
agattcggac cactaggcct gaaatgacat ttcactaaaa gtctccaaaa catttctaag 6360
actactaagg ccttttatgt aatttcttta aatgtgtatt tcttaagaat tcaaatttgt 6420
aataaaacta tttgtataaa aattaagctt ttattaattt gttgctagta ttgccacaga 6480
cgcattaaaa gaaacttact gcacaagctg ctaataaatt tgtaagcttt gcatacctta 6540
<210> 177
<211> 6446
<212> ДНК
<213> Homo sapiens
<400> 177
gagtgtggct gcagtgcgcc gggacaccag ggctccgcgc tccgcactca agaggctccc 60
gcgtcccaac ccctcgcgcc cgcgcgttcg cggatccagg ccgaggaccg aaaggggccg 120
cccgagcccc cggggccggc gcccagagag cccagcaagg ccggccgccc tgccggtgtg 180
ccgccggcgg gtgcttctgg aagggccaat gcgttcgggc agcagcccct gaagccgagc 240
ccgaggctaa gtgggactga ccggggccca gagtggacga accgccagca tggggagaga 300
ccagcgcgcg gtggccggcc ctgccctacg gcggtggctg ctgctgggga cagtgaccgt 360
ggggttcctc gcccagagcg tcttggcggg tgtgaagaag tttgatgtgc cgtgtggagg 420
aagagattgc agtgggggct gccagtgcta ccctgagaaa ggtggacgtg gtcagcctgg 480
gccagtgggc ccccaggggt acaatgggcc accaggatta caaggattcc cgggactgca 540
gggacgtaaa ggagacaagg gtgaaagggg agcccccgga gtaacgggac ccaagggcga 600
cgtgggagca agaggcgttt ctggattccc tggtgccgat ggaattcctg gacacccggg 660
gcaaggtggg cccaggggaa ggccgggcta cgatggctgc aacggaaccc agggagactc 720
aggtccacag gggccccccg gctctgaggg gttcaccggg cctcccgggc cccaaggacc 780
aaaagggcag aaaggtgagc cttatgcact gcctaaagag gagcgcgaca gatatcgggg 840
tgaacctgga gagcctggat tggtcggttt ccagggacct cccggccgcc ctgggcatgt 900
gggacagatg ggtccagttg gagctccagg gagaccagga ccacctggac cccctggacc 960
aaaaggacag caaggcaaca gaggacttgg tttctacgga gttaagggtg aaaagggtga 1020
cgtagggcag ccgggaccca acgggattcc atcagacacc ctccacccca tcatcgcgcc 1080
cacaggagtc accttccacc cagatcagta caagggtgaa aaaggcagtg agggggaacc 1140
aggaataaga ggcatttcct tgaagggaga agaaggaatc atgggctttc ctggactgag 1200
gggttaccct ggcttgagtg gtgaaaaagg atcaccagga cagaagggaa gccgaggcct 1260
ggatggctat caagggcctg atggaccccg gggacccaag ggagaagccg gagacccagg 1320
gccccctgga ctacctgcct actcccctca cccttcccta gcaaaaggtg ccagaggtga 1380
cccgggattc ccaggggccc aaggggagcc aggaagccag ggtgagccag gagacccggg 1440
cctcccaggt ccccctggcc tctccatcgg agatggagat cagaggagag gcctgccggg 1500
tgagatggga cccaagggct tcatcggaga ccccggcatc cctgcgctct acgggggccc 1560
acctggacct gatggaaagc gagggcctcc aggacccccc gggctccctg gaccacctgg 1620
acctgatggc ttcctgtttg ggctgaaagg agcaaaagga agagcaggct tccctgggct 1680
tcccggctcc cctggagccc gcggaccaaa ggggtggaaa ggtgacgctg gggaatgcag 1740
atgtacagaa ggcgacgaag ctatcaaagg tcttccggga ctgccaggac ccaagggctt 1800
cgcaggcatc aacggggagc cggggaggaa aggggacaga ggagaccccg gccaacacgg 1860
cctccctggg ttcccagggc tcaagggagt gcctggcaac attggtgctc ccggacccaa 1920
aggagcaaaa ggagattcca gaacaatcac aaccaaaggt gagcggggac agcccggcgt 1980
cccaggtgtg cccgggatga aaggtgacga tggcagccca ggccgcgatg ggctcgatgg 2040
attccccggc ctcccaggcc ctcccggtga tggcatcaag ggccctccag gggacccagg 2100
ctatccagga atacctggaa cgaagggtac tccaggagaa atgggccccc caggactggg 2160
ccttcccggc ctcaaaggcc aacgtggttt ccctggagac gccggcttac ctggaccacc 2220
aggcttcctg ggccctcctg gccccgcagg gaccccagga caaatagatt gtgacacaga 2280
tgtgaaaagg gccgttggag gtgacagaca ggaggccatc cagccaggtt gcataggagg 2340
gcccaaggga ttgccaggcc tgccaggacc cccaggcccc acaggtgcca aaggcctccg 2400
aggaatccca ggcttcgcag gagctgatgg aggaccaggg cccaggggct tgccaggaga 2460
cgcaggtcgt gaagggttcc caggaccccc agggttcata ggaccccgag gatccaaagg 2520
tgcagtgggc ctccctggcc cagatggatc cccaggtccc atcggcctgc cagggccaga 2580
tgggccccct ggggaaaggg gcctccctgg agaagtcctg ggagctcagc ccgggccacg 2640
gggagatgct ggtgtgcctg gacagcctgg gcttaaaggc cttcccggag acagaggccc 2700
ccctggattc agaggaagcc aagggatgcc tgggatgcca gggctgaagg gccagccagg 2760
cctcccagga ccttccggcc agccaggcct gtatgggcct ccaggactgc atggattccc 2820
aggagctcct ggccaagagg ggcccttggg gctgccagga atcccaggcc gtgaaggtct 2880
gcctggtgat agaggggacc ctggggacac aggcgctcct ggccctgtgg gcatgaaagg 2940
tctctctggt gacagaggag atgctggctt cacaggggag caaggccatc caggaagccc 3000
tggatttaaa ggaattgatg gaatgcctgg gacccccggg ctaaaaggag atagaggctc 3060
acctgggatg gatggtttcc aaggcatgcc tggactcaaa gggagacccg ggtttccagg 3120
gagcaaaggc gaggctggat ttttcggaat acccggtctg aagggtctgg ctggtgagcc 3180
aggttttaaa ggcagccgag gggaccctgg gcccccagga ccacctcctg tcatcctgcc 3240
aggaatgaaa gacattaaag gagagaaagg agatgaaggg cctatggggc tgaaaggata 3300
cctgggcgca aaaggtatcc aaggaatgcc aggcatccca gggctgtcag gaatccctgg 3360
gctgcctggg aggcccggcc acatcaaagg agtcaaggga gacatcggag tccccggcat 3420
ccccggtttg ccaggattcc ctggggtggc tggcccccct ggaattacgg gattcccagg 3480
attcatagga agccggggtg acaaaggtgc cccagggaga gcaggcctgt atggcgagat 3540
tggcgcgact ggtgatttcg gtgacatcgg ggacactata aatttaccag gaagaccagg 3600
cctgaagggg gagcggggca ccactggaat accaggtctg aagggattct ttggagagaa 3660
gggaacagaa ggtgacatcg gcttccctgg gataacaggc gtgactggag tccaaggccc 3720
tcctggactt aaaggacaaa caggctttcc agggctgact gggcctccag ggtcgcaggg 3780
agagctgggg cggattggac tgcctggtgg caaaggagat gatggctggc cgggagctcc 3840
gggcttacca ggttttccgg gactccgtgg gatccgcggc ttacacggct tgccaggcac 3900
caagggcttt ccaggatccc caggttctga catccacgga gacccaggct tcccaggccc 3960
tcctggggaa agaggtgacc caggagaggc caacaccctt ccaggccctg tgggagtccc 4020
aggacagaaa ggagaccaag gagctccagg ggaacgaggc ccacctggga gcccaggact 4080
tcaggggttc cctggtatca cacccccttc caacatctct ggggcacctg gtgacaaagg 4140
ggcgccaggg atatttggcc tgaaaggtta tcggggccca ccagggccac caggttctgc 4200
tgctcttcct ggaagcaaag gtgacacagg gaacccagga gctccaggaa ccccagggac 4260
caaaggatgg gccggggact ccgggcccca gggcaggcct ggtgtgtttg gtctcccagg 4320
agaaaaaggg cccaggggtg aacaaggctt catggggaac actggaccca ctggggcggt 4380
gggcgacaga ggccccaagg gacccaaggg agacccagga ttccctggtg cccccgggac 4440
tgtgggagcc cccgggattg caggaatccc ccagaagatt gccgtccaac cagggacagt 4500
gggtccccag gggaggcgag gcccccctgg ggcaccgggg gagatggggc cccagggccc 4560
ccccggagaa ccaggtttcc gtggggctcc agggaaagct gggccccaag gaagaggtgg 4620
tgtgtctgct gttcccggct tccggggaga tgaaggaccc ataggccacc aggggccgat 4680
tggccaagaa ggtgcaccag gccgtccagg gagcccgggc ctgccgggta tgccaggccg 4740
cagcgtcagc atcggctacc tcctggtgaa gcacagccag acggaccagg agcccatgtg 4800
cccagtgggc atgaacaaac tctggagtgg atacagcctg ctgtacttcg agggccagga 4860
gaaggcgcac aaccaggacc tggggctggc gggctcctgc ctggcgcggt tcagcaccat 4920
gcccttcctg tactgcaacc ctggtgatgt ctgctactat gccagccgga acgacaagtc 4980
ctactggctc tctaccactg cgccgctgcc catgatgccc gtggccgagg acgagatcaa 5040
gccctacatc agccgctgtt ctgtgtgtga ggccccggcc atcgccatcg cggtccacag 5100
tcaggatgtc tccatcccac actgcccagc tgggtggcgg agtttgtgga tcggatattc 5160
cttcctcatg cacacggcgg cgggagacga aggcggtggc caatcactgg tgtcaccggg 5220
cagctgtcta gaggacttcc gcgccacacc attcatcgaa tgcaatggag gccgcggcac 5280
ctgccactac tacgccaaca agtacagctt ctggctgacc accattcccg agcagagctt 5340
ccagggctcg ccctccgccg acacgctcaa ggccggcctc atccgcacac acatcagccg 5400
ctgccaggtg tgcatgaaga acctgtgagc cggcgcgtgc caggaagggc cattttggtg 5460
cttattctta acttattacc tcaggtgcca acccaaaaat tggttttatt tttttcttaa 5520
aaaaaaaaaa gtctaccaaa ggaatttgca tccagcagca gcacttagac ctgccagcca 5580
ctgtcaccga gcgggtgcaa gcactcgggg tccctggagg gcaagccctg cccacagaaa 5640
gccaggagca gccctggccc ccatcagccc tgctagacgc accgcctgaa ggcacagcta 5700
accacttcgc acacacccat gtaaccactg cactttccaa tgccacagac aactcacatt 5760
gttcaactcc cttctcgggg tgggacagac gagacaacag cacacaggca gccagccgtg 5820
gccagaggct cgaggggctc agggcctcag gcacccgtcc ccacacgagg gccccgtggg 5880
tgggcctggc cctgctttct acgccaatgt tatgccagct ccatgttctc ccaaataccg 5940
ttgatgtgaa ttattttaaa ggcaaaaccg tgctctttat tttaaaaaac actgataatc 6000
acactgcggt aggtcattct tttgccacat ccctatagac cactgggttt ggcaaaactc 6060
aggcagaagt ggagaccttt ctagacatca ttgtcagcct tgctacttga aggtacaccc 6120
catagggtcg gaggtgctgt ccccactgcc ccacgttgtc cctgagattt aacccctcca 6180
ctgctggggg tgagctgtac tcttctgact gccccctcct gtgtaacgac tacaaaataa 6240
aacttggttc tgaatatttt taaaccccga gttgttgacc gccttaatct cgtgtccata 6300
gagcaaaacg tctgctcaga tggatgcgag gcacagcgtc cgcccacgct gctgttttta 6360
atccatctca gtagagttga acccattcgt ggtattacag ccatttctcg gggaatgtgt 6420
ttgtttataa ctcactaatg cttaca 6446
<210> 178
<211> 2868
<212> ДНК
<213> Homo sapiens
<400> 178
ggctgcgctc cgagctgcgg agtccgggac tggagctgcc cgggcgggtt cgcgccccga 60
aggctgagag ctggcgctgc tcgtgccctg tgtgccagac ggcggagctc cgcggccgga 120
ccccgcggcc ccgctttgct gccgactgga gtttggggga agaaactctc ctgcgcccca 180
gaggatttct tcctcggcga agggacagcg aaagatgagg gtggcaggaa gagaagggcg 240
ctttctgtct gccggggtcg cagcgcgaga gggcagtgcc atgttcctct ccatcctagt 300
ggcgctgtgc ctgtggctgc acctggcgct gggcgtgcgc ggcgcgccct gcgaggcggt 360
gcgcatccct atgtgccggc acatgccctg gaacatcacg cggatgccca accacctgca 420
ccacagcacg caggagaacg ccatcctggc catcgagcag tacgaggagc tggtggacgt 480
gaactgcagc gccgtgctgc gcttcttcct ctgtgccatg tacgcgccca tttgcaccct 540
ggagttcctg cacgacccta tcaagccgtg caagtcggtg tgccaacgcg cgcgcgacga 600
ctgcgagccc ctcatgaaga tgtacaacca cagctggccc gaaagcctgg cctgcgacga 660
gctgcctgtc tatgaccgtg gcgtgtgcat ctcgcctgaa gccatcgtca cggacctccc 720
ggaggatgtt aagtggatag acatcacacc agacatgatg gtacaggaaa ggcctcttga 780
tgttgactgt aaacgcctaa gccccgatcg gtgcaagtgt aaaaaggtga agccaacttt 840
ggcaacgtat ctcagcaaaa actacagcta tgttattcat gccaaaataa aagctgtgca 900
gaggagtggc tgcaatgagg tcacaacggt ggtggatgta aaagagatct tcaagtcctc 960
atcacccatc cctcgaactc aagtcccgct cattacaaat tcttcttgcc agtgtccaca 1020
catcctgccc catcaagatg ttctcatcat gtgttacgag tggcgctcaa ggatgatgct 1080
tcttgaaaat tgcttagttg aaaaatggag agatcagctt agtaaaagat ccatacagtg 1140
ggaagagagg ctgcaggaac agcggagaac agttcaggac aagaagaaaa cagccgggcg 1200
caccagtcgt agtaatcccc ccaaaccaaa gggaaagcct cctgctccca aaccagccag 1260
tcccaagaag aacattaaaa ctaggagtgc ccagaagaga acaaacccga aaagagtgtg 1320
agctaactag tttccaaagc ggagacttcc gacttcctta caggatgagg ctgggcattg 1380
cctgggacag cctatgtaag gccatgtgcc ccttgcccta acaactcact gcagtgctct 1440
tcatagacac atcttgcagc atttttctta aggctatgct tcagtttttc tttgtaagcc 1500
atcacaagcc atagtggtag gtttgccctt tggtacagaa ggtgagttaa agctggtgga 1560
aaaggcttat tgcattgcat tcagagtaac ctgtgtgcat actctagaag agtagggaaa 1620
ataatgcttg ttacaattcg acctaatatg tgcattgtaa aataaatgcc atatttcaaa 1680
caaaacacgt aattttttta cagtatgttt tattaccttt tgatatctgt tgttgcaatg 1740
ttagtgatgt tttaaaatgt gatcgaaaat ataatgcttc taagaaggaa cagtagtgga 1800
atgaatgtct aaaagatctt tatgtgttta tggtctgcag aaggattttt gtgatgaaag 1860
gggatttttt gaaaaatcta gagaagtagc atatggaaaa ctataatgtg tcttttttac 1920
aatgacttca gctctgtttt tagctagaaa ctctaaaaac aaaaataata ataaagaaaa 1980
ataaataaaa aggagaggca gacaatgtct ggattcctgt tttttggtta cctgatttca 2040
tgatcatgat gcttcttgtc aacaccctct taagcagcac cagaaacagt gagtttgtct 2100
gtaccattag gagttaggta ctaattagtt ggctaatgct caagtatttt atacccacaa 2160
gagaggtatg tcactcatct tacttcccag gacatccacc ctgagaataa tttgacaagc 2220
ttaaaaatgg ccttcatgtg agtgccaaat tttgttttct tcatttaaat attttctttg 2280
cctaaataca tgtgagagga gttaaatata aatgtacaga gaggaaagtt gaggttccac 2340
ctctgaaatg agaattactt gacagttggg atactttaat cagaaaaaaa gaacttatct 2400
tgcagcattt tatcaacaaa tttcataatt gtggacaatt ggaggcattt attttaaaaa 2460
acaattttat tggccttttg ctaacacagt aagcatgtat tctctataag gcattcaata 2520
aatgcacaac gcccaaagga aataaaatcc tatctaatcc tactctccac tacacagagg 2580
taatcactat tagtattttg gcatattatt ctccaggtgt ttcttatgca cttataaaat 2640
gatttgaaca aataaaacta ggaacctgct atacatgtgt ttcataacct gcctcctttg 2700
cttggccctt tattgagata agttttcctg tcaagaaagc agaaaccatc tcatttctaa 2760
cagctgtgtt atattccata gtatgcatta ctcaacaaac tgttgtgcta ttggatactt 2820
aggtggtttc ttcactgaca atactgaata aacatctcaa tagtcaaa 2868
<210> 179
<211> 7437
<212> ДНК
<213> Homo sapiens
<400> 179
agaaacagcc ctcaagtttc atcacaggga tgctaaaata tattctccag cttcagcaat 60
gacctagttc actgaagggc ggggaaaaaa tgctcttctt ccagaacttc cccgggcatg 120
gaatttcccc tcttaggaag agatgaactt tctggaagca gacccatcta tgctttgacc 180
tgatcatgga agaccctgag ccctataaac tcctttgaat ccctgtcccg ggaacaggga 240
cacccaatca atgcctcaca cacagcctgg ggaatccaca gctgagccag ccctcctaca 300
acatttcttg gacaaagagt actgaaaaat ttggtgcggc atgaaaggat gaaagactcc 360
gaagggcccc agaggccccc gctgtgtttt ttatctacat tgctttccca gaaggttcct 420
gagaagtcag acgctgtgct tcgctgcata atatctggtc agcccaagcc agaggtaact 480
tggtataaga atggtcaggc catcgatggg agtggcatta tttccaacta tgaattcttt 540
gagaatcagt atattcatgt gttacatctc tcttgctgta ccaaaaatga tgctgctgtc 600
tatcaaatct cggctaaaaa ctcttttgga atgatctgtt gttctgcttc cgttgaggtt 660
gagtgctcat cagagaaccc acaattgtct cctaacctgg aagatgacag ggacaggggt 720
tggaaacatg aaacagggac acatgaagaa gaaagggcaa atcagattga tgagaaggaa 780
catccttata aggaagaaga aagcatctcc ccgggcactc ccaggtcagc tgactcctcc 840
ccctccaaat ccaaccattc actctccctc cagtcattgg gcaatcttga cattagtgtg 900
tccagttctg aaaatccttt gggtgttaaa ggaacaaggc acactggaga ggcttatgat 960
ccaagtaaca cagaagaaat tgcaaatggg ttgctttttc ttaattcaag tcatatttat 1020
gaaaaacaag acagatgttg ccacaagaca gtgcattcca tggcatcaaa gttcacggat 1080
ggtgacctga acaatgatgg tcctcatgat gaaggcttac gctctagtca gcaaaatccc 1140
aaagtacaga aatacattag cttcagcctc ccgctatctg aggcaactgc acacatttac 1200
ccaggtgaca gtgccgtggc caacaaacaa cccagcccac agctttccag tgaagactct 1260
gacagtgact atgaactttg cccagagata accctaacct acaccgagga gttttcagat 1320
gatgacctgg agtatctgga atgttctgat gttatgacgg attactctaa tgcagtttgg 1380
caaaggaacc tgctggggac tgagcatgtt tttttattag aaagcgatga cgaagagatg 1440
gaattcggtg agcattgcct gggtgggtgt gagcatttcc tcagtggaat gggttgtggg 1500
tctcgggtgt cgggtgacgc tgggcctatg gttgccactg ctggcttctg tggtcatcac 1560
tcacaacccc aagaagttgg ggtgaggagc agcagagtct ccaagcacgg tccctcatcc 1620
ccacaaacag ggatgactct cattttggga cctcaccagg atggaacgtc ttcagtgaca 1680
gaacagggga gatataaact ccccactgct cccgaggctg ctgaaaatga ttatccagga 1740
attcaaggag aaaccagaga cagccaccaa gcaagagagg aatttgccag tgacaatctg 1800
ctcaacatgg atgaatcagt aagagagaca gagatgaagc tcttgtctgg tgagtcagaa 1860
aactcaggga tgagccagtg ttgggagacg gcagctgaca agagagtggg gggaaaggac 1920
ttatggagca agaggggttc aaggaaatct gccagggtga ggcagccggg aatgaaggga 1980
aatcccaaga agccgaatgc caacctgaga gaaagtacaa cagaaggtac ccttcatctc 2040
tgctctgcca aagaatctgc tgagccccca ctaacccaga gtgataaaag agagacttct 2100
cacaccacag cagcagcgac tggtcggagt tcccatgctg atgcaagaga atgtgctatt 2160
tcaacccagg cagagcaaga agcaaaaacc cttcaaactt caacagactc agtctccaaa 2220
gaaggcaaca caaattgcaa gggagaaggc atgcaagtta atactctatt tgaaacaagc 2280
caggttccag actggagtga tcctcctcag gtacaagttc aggaaacagt cagagagaca 2340
atctcttgca gccagatgcc agctttctca gagcctgctg gggaggagtc cccattcact 2400
gggaccacaa caatttcctt ctcaaactta ggaggggtcc acaaggaaaa tgcatcatta 2460
gctcaacact cggaggtcaa accctgtacc tgtggtccac agcatgaaga aaaacaagac 2520
agagatggca acatacctga caatttcagg gaagacctaa aatatgagca gagcatctca 2580
gaagccaatg atgagactat gtccccaggt gtgttctcaa ggcatctccc caaggatgct 2640
cgtgctgact tcagggagcc tgtggctgtc tctgttgctt cccctgaacc cacagatact 2700
gccctcaccc tggaaaatgt gtgtgatgag ccaagggaca gagaagcagt gtgtgcaatg 2760
gagtgttttg aggctggtga ccaaggaacg tgttttgata ccatagattc tcttgttggg 2820
agaccagttg ataaatattc gcctcaagaa atttgctctg tagatacgga actggcagaa 2880
ggtcaaaaca aagtatctga tttatgttct tctaatgaca agacactgga agtctttttt 2940
cagacacaag tgtctgagac ttcagtgtct acgtgcaaaa gcagcaagga cggcaactca 3000
gtcatgtccc ctctttttac cagtactttc accttgaaca tttcacacac agctagtgaa 3060
ggtgccacag gagaaaatct agccaaggtg gagaattcca cctacccact ggcctccaca 3120
gtacatgctg gccaggagca gccaagcccc agcaactcag gagggcttga tgaaacacag 3180
ctcctttctt ctgagaacaa tcctttagtg caatttaaag aaggaggtga caagagcccc 3240
agtcctagtg ccgcagacac cacagccaca ccagccagtt atagttcaat tgtgagtttt 3300
ccttgggaga agccaacaac attaactgct aataatgagt gctttcaagc gaccagggag 3360
actgaggaca catcaactgt taccattgcc accgaagtcc acccagccaa ataccttgct 3420
gtgtcaattc ctgaggacaa gcatgcaggt ggcactgagg agaggttccc tcgtgcatcc 3480
catgaaaagg tttcccaatt tccttcccaa gtgcagttgg atcatatttt aagtggtgct 3540
accatcaaat ctacaaaaga gctactttgc agggcaccca gtgtgccagg agtcccacac 3600
catgtcctgc agctcccaga gggagagggt ttctgcagta attcccctct tcaggttgat 3660
aacctgtctg gagataagag ccagactgtg gacagagcag actttaggag ctatgaagag 3720
aatttccaag aaagaggaag tgaaacaaag cagggggtcc agcagcagag cctgtcccag 3780
cagggttctc tttctgcacc tgatttccaa caaagtttgc ctacgacatc tgctgcacaa 3840
gaggaaagaa acttggtgcc cacggcccac tcacccgcaa gctctaggga aggagcaggg 3900
cagcgctcag gttgggggac gagggtctcc gtggtggctg aaactgctgg ggaagaagac 3960
agtcaggctc tgagcaacgt tccatctctc tctgatatcc ttttggaaga gtctaaagaa 4020
tatagacctg gaaattggga ggcaggcaac aagctgaaga ttataactct agaggcttcc 4080
gcttctgaaa tctggccacc acgacaactg acaaattctg agagcaaggc atcagacggt 4140
ggtctcataa ttcctgacaa ggtctgggct gtacctgata gtctaaaggc agatgctgtt 4200
gtgcctgaat tggccccctc tgaaatagca gcattggctc acagtccaga ggatgctgag 4260
tcagcccttg ctgatagcag agaaagccat aaaggcgaag agcccaccat cagtgtacat 4320
tggagaagtc tttcttcccg gggtttcagc caacccagac tcctggagtc atccgtggac 4380
cctgtagatg aaaaggagtt atctgtcaca gattcactgt cagcggcttc tgaaactgga 4440
gggaaggaaa atgttaacaa tgtgagtcaa gaccaggagg aaaaacaact caagatggat 4500
cacactgcct tctttaaaaa gtttctgacc tgccctaaaa tcctagagtc ctctgtagat 4560
cccattgatg agataagtgt gatagagtac accagggctg gaaaaccaga gccctctgaa 4620
accacaccac agggcgccag agaagggggt caatcaaatg acggaaacat gggccacgaa 4680
gcggaaatcc agccggccat tttgcaagtt ccatgtctcc agggaaccat tctgagtgaa 4740
aatagaatca gcagaagcca agaaggcagt atgaagcagg aggctgaaca aattcaacct 4800
gaggaggcaa aaactgccat ttggcaagtc ctgcaaccca gcgaaggcgg tgaaagaatt 4860
ccaagtggat gtagcatagg ccaaatacaa gaaagcagtg atgggagctt aggggaggct 4920
gagcaaagca aaaaggacaa agcagaattg atttccccca cttcacctct ttctagttgt 4980
cttccaataa tgactcacgc ttctcttggg gttgacacgc acaactccac aggccaaatt 5040
catgacgtcc ctgaaaatga catagttgag cccagaaagc gtcagtatgt gtttcctgtt 5100
tcacagaaaa ggggaactat tgagaatgag cgtgggaaac ctttgccctc ttctcctgat 5160
cttaccaggt tcccttgtac ttcatctcct gaaggaaatg tcacagactt tttgataagc 5220
cacaaaatgg aggaacctaa aatagaggtg cttcaaattg gggaaaccaa acccccaagc 5280
tcatctagct cctcagcgaa gaccttggca tttatttcag gagaacgtga gttagagaaa 5340
gcccctaaat tactgcagga tccatgtcaa aagggcaccc tgggctgtgc gaaaaagtcc 5400
agggagagag agaagtccct ggaagcccga gcaggcaaat cgccagggac cctcacagca 5460
gtgacggggt cagaggaggt caagaggaag ccagaagccc caggcagtgg acatttagct 5520
gagggagtaa agaagaaaat tttgtccagg gtggcagcac tgaggctgaa actggaagaa 5580
aaggaaaata tcagaaagaa ctcagccttt cttaaaaaga tgcccaaact cgaaacatca 5640
ttatcacaca cagaagagaa acaagaccca aaaaagccat cttgcaaaag agaaggaaga 5700
gctccagtat tactgaaaaa gatccaagct gagatgttcc ctgaacactc tggaaatgta 5760
aaattaagct gccaatttgc agaaattcat gaagattcta ctatctgctg gacaaaagat 5820
tcaaagtcca tagcccaagt gcagagaagt gcaggggaca actccactgt ttcctttgcc 5880
atcgtgcaag ccagtccgaa ggaccaggga ctctattact gctgcatcaa gaacagctac 5940
ggaaaagtga ctgctgaatt taacctcaca gctgaagttc tcaaacagct gtcaagtcgc 6000
caggatacta aaggatgtga agagattgaa ttcagccaac tcatcttcaa agaagacttc 6060
ctccatgaca gctactttgg gggccgcctg cgtggtcaga tcgccacgga ggagctgcac 6120
tttggagaag gggttcaccg caaagccttc cgcagcacag tgatgcacgg cctcatgcct 6180
gtcttcaaac ctggccatgc ctgtgtgctt aaggtgcaca atgccattgc ctatgggacc 6240
agaaataatg atgagctcat ccaaaggaac tacaaactcg ctgcccagga atgctatgtt 6300
caaaatactg ccaggtatta tgccaagatc tacgctgctg aagcacagcc tctggaaggc 6360
tttggagaag tacctgagat cattcctatt tttcttatcc atcggcctga gaacaatatc 6420
ccgtatgcta cagtggagga ggagctgatt ggagaatttg tgaagtattc catcagggat 6480
gggaaagaaa taaacttctt gagaagagaa tcagaagctg gtcagaaatg ttgcaccttc 6540
cagcactggg tgtaccagaa aacaagtggc tgcctcctgg tgacggacat gcaaggtgta 6600
ggaatgaagc taactgacgt tggcatagca acgctggcta aagggtacaa gggatttaaa 6660
ggcaactgtt ccatgacctt cattgatcag tttaaagcac tacaccagtg taacaagtat 6720
tgcaaaatgc tgggactgaa atcccttcaa aacaacaacc agaaacagaa gcagccgagc 6780
attgggaaaa gcaaagttca aacaaactct atgacaataa agaaggcagg gcctgagacc 6840
ccaggcgaaa agaaaaccta acgtccctgg gtaacctaat ggccactggc tagcagcaca 6900
caatctcgcc agggaaaatc tgaggccaca caggagagaa tatacagcct gcagagagtg 6960
cgtggcaatc cttactccca gccgactgtg cgccaagatg cttctaaacc catcacctgc 7020
tgtcttcact caaatgattt cagaacagga tttgcgacca ggtttatggg gagattgaat 7080
caacgattgg tctcaaagac agtccattct ttatatacat gtttagcatt tttaccaacc 7140
tcacatcatg tgtatatttg tgtatttgca catggttgtg ctgtcgagga cctggtgctg 7200
agaagagtct gttcacagcc aaaattcttc ccactgtcat tcctaacctg ggatttctag 7260
acacatcctg ctgtgatgta aacagaaatc acgaattcgc tcactggatc aagttgttcc 7320
actggtgtct aatacgctat tgttgccgga ggtgggttct gtgacgtgaa gccatttccc 7380
atcattcaac agccagttac aattttctgt ttaattaaat tcatatttaa acaaaaa 7437
<210> 180
<211> 8442
<212> ДНК
<213> Homo sapiens
<400> 180
gtccgcactc tccgtccccg cggctggcgc aggacctcac tcgagcggag cgcccacggg 60
gagcgggtcg cggggcggcg gcggcgagga ggaggcgaga aggagttgga ggaggaggag 120
gaggaggcga gggcgagcta gcccagcggg gtcccggccg ccccgcgggc caaagtcgag 180
ccctcccgcc cgtgggcgag cgcgccagcc gccccttcca gaacagccgc cgccacaaag 240
aagaacgggg ggtgccgagg tccccatgac ctcctaaagt ggtgcggtcc ctgctgagtg 300
cgctgcccgg gccgtgaccc gcgcccctgt gcgtccccgc gcgcctccga gcgcccctgt 360
gcgccccggc ccgcgccccg ccggcatgga cgtccatacc cgctggaaag cgcgcagcgc 420
gctccgcccg ggcgccccgc tgctgccccc gctgctgctg ctgctgctgt gggcgccgcc 480
tccgagccgc gcagctcagc cagcagatct cctgaaggtt ctagattttc acaacttgcc 540
tgatggaata acaaagacaa caggcttttg cgccacgcgg cgatcttcca aaggcccgga 600
tgtcgcttac agagtcacca aagacgcgca gctcagcgca cccaccaagc agctgtaccc 660
tgcgtctgca tttcccgagg acttctccat cctaacaact gtgaaagcca agaaaggcag 720
ccaggccttc ctggtctcca tctacaacga gcagggtatc cagcagattg ggctggagct 780
gggccgctct cccgtcttcc tctacgagga ccacacgggg aagcctggcc cggaagacta 840
ccccctcttc cggggcatca acctgtcaga tggcaagtgg cacagaattg ctctcagcgt 900
ccacaagaaa aatgtcacct tgatcctcga ctgtaaaaag aagaccacca aattcctcga 960
ccgcagcgac caccccatga tcgacatcaa tggcatcatc gtgtttggca cccggatcct 1020
ggatgaggag gtgtttgagg gtgacatcca gcagctgctc tttgtctcgg accaccgggc 1080
agcttatgat tactgtgagc actacagccc tgactgtgac accgcagtac ctgacacccc 1140
acagtcgcag gaccccaatc cagatgaata ttacacggaa ggagacggcg agggtgagac 1200
ctattactac gaatacccct actacgaaga ccccgaagac ctagggaagg agcccacccc 1260
cagcaagaag cccgtggaag ctgccaaaga aaccacagag gtccccgagg agctgacccc 1320
gacccccacg gaagctgctc ccatgcctga aaccagtgaa ggggctggga aggaagagga 1380
cgtcggcatc ggggactatg actacgtgcc cagtgaggac tactacacgc cctcaccgta 1440
tgatgacctc acctatggcg agggggagga gaaccccgac cagcccacag acccaggcgc 1500
tggggccgaa attcccacca gcaccgccga cacctccaac tcctccaatc cagctccgcc 1560
tccaggggaa ggtgcggatg acttggaggg ggagttcact gaggaaacga tccggaacct 1620
tgacgagaac tactacgacc cctactacga ccccaccagc tccccgtcgg agatcgggcc 1680
gggaatgccg gcgaaccagg ataccatcta tgaagggatt ggaggacctc ggggcgagaa 1740
aggccaaaag ggagaaccag cgattatcga gccgggcatg ctcatcgagg gcccgcctgg 1800
cccagaaggc cccgcgggtc ttcccggacc tccaggaacc atgggtccca ctggccaagt 1860
cggggaccct ggagaaaggg gcccccctgg acgcccaggc cttcctgggg ccgatggcct 1920
gcccggtcct ccaggaacca tgctcatgct gcccttccgg tttggaggtg gcggcgatgc 1980
gggctccaaa ggccccatgg tctcagccca ggagtcccag gcgcaagcca ttctccagca 2040
ggccaggttg gcactgaggg gaccagctgg cccgatgggt ctcacaggga gacctggccc 2100
tgtgggtccc cctgggagcg gaggtttgaa gggcgagccg ggagacgtgg ggcctcaggg 2160
tcctcgaggt gtgcaaggcc cgcctggtcc ggccgggaag cccggaagac ggggtcgggc 2220
tgggagtgat ggagccagag gaatgcctgg acaaactggc cccaagggtg accggggttt 2280
cgacggcctg gctgggttgc caggcgagaa gggccacagg ggtgaccctg gtccttccgg 2340
cccaccagga cctccgggag acgatggaga aaggggtgac gacggagaag ttgggcccag 2400
ggggctgcct ggggagcccg ggccacgtgg tctgcttggg ccgaaggggc ccccaggtcc 2460
tcccggacct cccggtgtca cgggtatgga cggccagccg gggccaaaag gaaatgtggg 2520
tccccaggga gagcctggcc ccccaggaca gcagggtaat ccaggcgccc agggtcttcc 2580
aggcccccag ggtgcaattg gtcctccagg agaaaagggt cccttgggga aaccaggcct 2640
tccaggaatg cccggtgctg acggaccccc gggacaccct ggcaaagaag gccctccagg 2700
agagaaagga ggtcagggtc cacctggccc ccagggtccg attggctacc caggtcctcg 2760
aggagtcaag ggggccgatg gcatccgtgg tctgaagggc acaaagggcg agaagggtga 2820
agacggcttt cctgggttta aaggagacat gggcatcaag ggtgatcggg gggagatcgg 2880
cccacccggt cccaggggag aagatggccc tgaaggccca aagggtcgcg gaggtcccaa 2940
tggtgacccc ggtcctctgg gaccccctgg ggagaaggga aaactcggag tcccagggtt 3000
accagggtat ccaggaagac aaggaccaaa gggctctatt ggattccctg gatttcctgg 3060
cgccaatgga gagaagggcg gcagggggac ccctggaaag ccaggaccgc gggggcagcg 3120
aggcccaacg ggtccgaggg gtgaaagagg cccccggggc atcactggga agcctggccc 3180
caagggcaac tccggaggtg acggcccagc tggccctcct ggtgaacggg gacccaatgg 3240
accccaagga cccacaggat ttcctggacc aaagggcccc cctggccctc caggcaagga 3300
tggactccca ggacaccctg gacagagagg cgagactggt ttccaaggca agaccggccc 3360
tccaggcccc cccggcgtgg tcggccctca gggtcccacg ggagaaacgg gcccaatggg 3420
tgagcgtggc caccctgggc cccctggacc ccccggtgaa caggggcttc cgggccttgc 3480
tggaaaagaa gggacgaagg gtgacccagg ccctgcaggc ctccctggga aagatggccc 3540
tccaggatta cgtggtttcc ctggggaccg agggcttcct ggtccagtgg gagctcttgg 3600
actgaaaggc aatgaagggc cccctggccc accaggccct gcgggatctc caggggagag 3660
aggtccagct ggagccgctg ggcccatcgg aattccaggg agacctgggc cccagggacc 3720
cccagggccg gcaggagaga aaggggctcc tggcgagaaa ggcccacaag gcccagctgg 3780
ccgagacggt ctccaggggc ctgtggggct cccgggtcca gctggccctg tgggtccccc 3840
tggagaagac ggagataagg gagagatcgg ggagccgggg cagaaaggaa gcaaggggga 3900
caaaggagaa cagggtcctc ctgggcctac aggtcctcaa ggccccatcg gacagccagg 3960
cccctctgga gctgacggcg agccggggcc tcggggccag cagggccttt tcgggcagaa 4020
aggtgatgaa ggtcccagag gctttcctgg accccctggg ccagtggggc tgcagggttt 4080
gccaggacct ccaggcgaga agggtgagac aggagacgtg ggccagatgg gccccccggg 4140
tccccctggc ccccgaggac cctccggagc tccaggtgct gatggcccac aaggtccccc 4200
aggtggaata ggaaaccctg gtgcagtggg agagaagggc gagcctggcg aagcaggtga 4260
gcctggcctt ccgggagaag gcggcccccc gggacccaaa ggagaaaggg gagagaaggg 4320
cgagtcaggc ccttcaggtg ctgccggacc ccctggaccc aaaggccctc ccggagatga 4380
tggtcccaaa ggcagccctg gcccagtggg ttttcctgga gatcctggcc cccccggaga 4440
gcctggcccc gcgggtcaag atggtccccc tggtgacaaa ggagatgatg gtgaacccgg 4500
gcagacggga tcccccggcc ctactggtga accaggtcca tcggggcctc caggaaaaag 4560
gggtccccca ggccccgcag gccccgaagg cagacaggga gagaaagggg ccaagggaga 4620
agccggcttg gaaggccctc ctgggaagac tggccccatc ggcccccagg gggcccctgg 4680
gaagcccgga ccggatggcc ttcgagggat ccctggccct gtgggagaac aaggtctccc 4740
aggatcccca ggcccggacg gtccccccgg ccccatgggt cccccaggac ttcccggcct 4800
caaaggagat tctggtccca aaggtgaaaa gggtcatcca ggcctgatcg ggctcatcgg 4860
tcctccgggt gaacagggtg agaagggcga ccgtggtctc cctggccccc agggctcctc 4920
cggtcctaag ggagaacagg gtatcactgg tccttctggc ccgattgggc ctcctgggcc 4980
ccctggcctg ccgggtccgc ctggtccaaa aggtgctaag ggctcctcgg gtccaactgg 5040
cccgaagggt gaggcaggcc acccaggacc cccaggcccc ccgggccccc cgggagaggt 5100
catccagccc ctgccaatcc aggcatccag gacgcggcgg aacatcgacg ccagccagct 5160
gctggacgac gggaatggcg agaactacgt ggactacgcg gacggcatgg aagagatctt 5220
cggctctctc aactctctga agctggagat tgagcagatg aaacggcccc tgggcacgca 5280
gcagaacccc gcccgcacct gcaaggacct gcagctctgc caccccgact tcccagatgg 5340
tgaatactgg gtcgatccta accaaggatg ctccagggat tccttcaagg tttactgcaa 5400
cttcacagcc ggggggtcga catgcgtctt ccctgacaag aagtccgaag gggccagaat 5460
cacttcttgg cccaaagaaa acccgggctc ctggttcagt gaattcaagc gtgggaaact 5520
gctctcctat gtggacgccg agggcaaccc tgtgggtgtg gtacagatga ccttcctgcg 5580
gctgctgagc gcctctgccc accagaacgt cacctaccac tgctaccagt cagtggcctg 5640
gcaggacgca gccacgggca gctacgacaa ggccctccgc ttcctgggct ccaacgacga 5700
ggagatgtcc tatgacaaca acccctacat ccgcgccctg gtggacggct gtgctaccaa 5760
gaaaggctac cagaagacgg ttctggagat cgacaccccc aaagtggagc aggtgcccat 5820
cgtggacatc atgttcaatg acttcggtga agcgtcacag aaatttggat ttgaagtggg 5880
gccggcttgc ttcatgggct aggagccgcc gagcccgggc tcccgagagc aacctcgtga 5940
cctcagcatg ccattcgttc gtgagtgtcc cgtgcacgtc ctgaccctgg acagtgaagg 6000
cttctccctc ccctcccacc tgacttcatc tacgcctcgg caccacgggg tgtgggaccc 6060
cagcccggag agaacagagg gaaggagccg cgcccccacc tggagctgaa tcacatgacc 6120
tagctgcacc ccagcgcctg ggcccgcccc acgctctgtc cacacccacg cgccccggga 6180
gcggggccat gcctccagcc ccccagctcg cccgacccat cctgttcgtg aataggtctc 6240
aggggttggg ggagggactg ccagatttgg acactatatt tttttctaaa ttcaacttga 6300
agatgtgtat ttcccctgac cttcaaaaaa tgttccaagg taagcctcgt aaaggtcatc 6360
ccaccatcac caaagcctcc gtttttaaca acctccaaca cgatccattt agaggccaaa 6420
tgtcattctg caggtgcctt cccgatggat taaaggtgct tatgtttttg tgagttttaa 6480
gtaaatattt gtattgtatt gttataaatg ttaagtgtgc ctggctttca atcatgcacg 6540
gaaacccagt ctcagtccca cggacagaat gggcgaggca tggattctgg gttgcagtac 6600
cgttctgatt agaaatagga agtctcccca cccccgccct ggccaagaac gtgcaataaa 6660
ttggaagttt gccccggggc agcaagaatt tatgctgcca ttgaaaagca ggtaccagtg 6720
ccccttttca gacagttttt gattcgctct agactttttt tttttttaat agggaaaaaa 6780
tttgataatt ttcttttttc tacatgcact taagactaaa acacaggttt ggattaattt 6840
tatttgcttc ctttttccgc ttttcttccc gcagagcctg atgggagaat gtccagggca 6900
gggaaaccac attttttgta ggtgataact caatgaaaat tggtgcttat tttttacact 6960
tctctcttgt ggctctcttg tggtgctatc tatctgtttt aaggtctcct tgaaggcgca 7020
ctggggaccc tggccatgcc tcgttctccc tgctttcttt atcctgttat tgcctccaca 7080
gtctgttgcc aaggactcta agatcaatgc acgtcacttt cctttccact gggcaggata 7140
gccaagcaca ctccctcctg cgctctcccg ccccggtgcg tccactcccg agggctgtta 7200
tgaggactgg gttgtgccta cttgatttga aaacacacac aagcaataaa aagcctcttc 7260
ctgcattgtc tgtggtgtga ccatagcaga ttatatttgg ttcctgaatg tttgtggtgc 7320
taatttctgt gtttgttcca agccgttcag tcatgccatg cgctgcctcg gtagatggag 7380
taatgtacaa tgaactccat gagtctctcc agggctgcct gcagcacgtc ttttccaagt 7440
agcctatttg gattcccatc tcaaatgtcc tggatgcgag cgtcagcggc tccagagctc 7500
ggggcgggtg aggtcccctt tggggaaccc tttcctggcc atcgaggtcg gggggctgcc 7560
gtctgtgggc aggaggaccc gaggggcagc caggaaaggc gatctcttca ctgtgaaaag 7620
ttgcccgggt gcagcgcctt ttccttctac catgggaaat gcaggctggg cccttggggt 7680
gagcctgcgg ggctctggtg ctgtccccga cccccaccac caccagaatg cagttccagc 7740
ttaggaagcc acaaacaagc cacccaggag gaacaaaaca ccgccagcgt ggattttcca 7800
aatttccctg gaaagtaagt ctcgctcttg ccaaagaaaa gtctggcttg gagagtctct 7860
ggagcccagg atgccagcat gtgccaatga ctgtcacctt catctcttca aaagaaaagc 7920
catagccgag gactgtcccg cgacccccgt ggactgcgtc taggtcatgt gattctgttt 7980
tcatttctca tcccatccaa tttgtccttt tctcctgtca ttttcttcct ctgtggtccc 8040
ttcaaagttg ttataatttg tactgaactt caaaatgtgt cccgttctcc ccagaccact 8100
ctagccacag tatattgcaa taaaattact tcttatattt gcagaaattc ttttggtgta 8160
attttatttt ttcctctcaa tatatataat tggacaaacg ctggcaaaaa gaaaaaaatg 8220
gtaagcaaaa aacccaagat aaagtttcga ggacatcagg ccttttgaaa tacaatgtca 8280
aatgacacat tgtacggttt caaaaaatcc gctagacatg tcataagttt taactgtaat 8340
gcccaggaaa ggatatctta aaatattcta aacttgtgta acaaaggaat aattaactgt 8400
aatagttttt caataaatcg agttgggtgt ttccaccgta aa 8442
<210> 181
<211> 4260
<212> ДНК
<213> Homo sapiens
<400> 181
cgaggagcga gccagtgggg gaggctgaca tcaccacggc ggcagccctt taaacccctc 60
acccagccag cgccccatcc tgtctgtccg aacccagaca caagtcttca ctccttcctg 120
cgagccctga ggaagccttg tgagtgcatt ggctggggct tggagggaag ttgggctgga 180
gctggacagg agcagtgggt gcatttcagg caggctctcc tgaggtccca ggcgccagct 240
ccagctccct ggctagggaa acccaccctc tcagtcagca tgggggccca agctccaggc 300
agggtgggct ggatcactag cgtcctggat ctctctcaga ctgggcagcc ccgggctcat 360
tgaaatgccc cggatgactt ggctagtgca gaggaattga tggaaaccac cggggtgaga 420
gggaggctcc ccatctcagc cagccacatc cacaaggtgt gtgtaagggt gcaggcgccg 480
gccggttagg ccaaggctct actgtctgtt gcccctccag gagaacttcc aaggagcttt 540
ccccagacat ggccaacaag ggtccttcct atggcatgag ccgcgaagtg cagtccaaaa 600
tcgagaagaa gtatgacgag gagctggagg agcggctggt ggagtggatc atagtgcagt 660
gtggccctga tgtgggccgc ccagaccgtg ggcgcttggg cttccaggtc tggctgaaga 720
atggcgtgat tctgagcaag ctggtgaaca gcctgtaccc tgatggctcc aagccggtga 780
aggtgcccga gaacccaccc tccatggtct tcaagcagat ggagcaggtg gctcagttcc 840
tgaaggcggc tgaggactat ggggtcatca agactgacat gttccagact gttgacctct 900
ttgaaggcaa agacatggca gcagtgcaga ggaccctgat ggctttgggc agcttggcag 960
tgaccaagaa tgatgggcac taccgtggag atcccaactg gtttatgaag aaagcgcagg 1020
agcataagag ggaattcaca gagagccagc tgcaggaggg aaagcatgtc attggccttc 1080
agatgggcag caacagaggg gcctcccagg ccggcatgac aggctacgga cgacctcggc 1140
agatcatcag ttagagcgga gagggctagc cctgagcccg gccctccccc agctccttgg 1200
ctgcagccat cccgcttagc ctgcctcacc cacacccgtg tggtaccttc agccctggcc 1260
aagctttgag gctctgtcac tgagcaatgg taactgcacc tgggcagctc ctccctgtgc 1320
ccccagcctc agcccaactt cttacccgaa agcatcactg ccttggcccc tccctcccgg 1380
ctgcccccat cacctctact gtctcctccc tgggctaagc aggggagaag cgggctgggg 1440
gtagcctgga tgtgggccaa gtccactgtc ctccttggcg gcaaaagccc attgaagaag 1500
aaccagccca gcctgccccc tatcttgtcc tggaatattt ttggggttgg aactcaaaaa 1560
aaaaaaaaaa aaatcaatct tttctcaggc ctggctggca gagtttgatt tgccctggtc 1620
acttttgtta tggtttcaga tctgcgcagg ggacaggcag gcatctcggg cttgtatgtg 1680
ttcctggctg ccgtcctctg aaggaccctt catgtgccaa gattccagga cagtcagatt 1740
ctttcagagg aaacaggcca tagaacagga gagtgaatct tggggaccga agggaaaaag 1800
gagccactca gcagcatgca gtggctttgg ggagacagag gaaaggagag attgcgagtg 1860
gcagaattgt ttggaaggtt tttattttgg aaaagctgtg cagaaaaaaa ttcaagaaaa 1920
tgttgctgtg aaaagagaaa aacaaatctt aagcattaaa aaaaattcaa ccaactagct 1980
cagaagaggg gagaaggcca aggttgagag aagagtcaca gcctgtcagg cagatagctg 2040
gcctccggcg ggaaggccat ttccctgagc acttgcagag gaagacaggg tggtggcagg 2100
actggagtgg cagtggctcc caaggctctc tcctccaaca tgtgcatctg ccatctgctc 2160
tgcagtcctg ccgcaggatt ccctagtgaa gcagctcagg cctgggggag ccgtgtgtat 2220
cccagctgtg ccggcagcat cataccaatc gtgggggtgg tgaaggagcc aggggttcat 2280
tcatggttgg tttctgatca ggcatcttgg gaatggataa tggaggcagc cgctttcagg 2340
acaggcatgt ccaggggctc ctcccagcct ctaccccgaa gtcctcttcc caagtgaccc 2400
cagtgatgtt tccattgaga tgcgctcctg gctatggcag gcaccttctc aacttatatg 2460
tgggaagggg tcccccatgc ttgggggacc taggcagctg gcttggccca agagagatga 2520
ggatggaggc caaaccaaag gggggcgcca atcccctgtc caacaccttc tcaccaaaag 2580
ctcccgtttg gctggaggca gaccctgtgg ctctgaccag acttctctgg cagcaatcct 2640
ccaccatttg tatcttaaga aggccctcac cctctttgag tggagtcaga ggatgcctca 2700
gattccagat gtaagcatca gaactgcttt ctgtcaagag ctccctaatt ttgggaaaga 2760
agagcctgtc ccacactgtc aggccctgag gtcagcagat ctgctcctcc ttcccgtgcg 2820
gtacgtctag gtgctgatga gggcagtcca gggcgctctt gttctgggac aggctccagt 2880
ccccttgctc cagcaggtct ggggcaagga ggtcagaggt ggtgggaggg cccctgctct 2940
ctgtttccac ttcgtctgga tccttgctgc tgcaaagtgg cactgattct agctctgtcc 3000
cttcctcctt ggctttccgg ctccgatggg gccagtggca gggtccactc ctacaaactt 3060
gattggaagc cacattcctc tggctcaaat atacttccag catgtagtaa acagtccaga 3120
agacggtgaa acagcctacc agcaccaggg tctggggccg aggcaagcac ctacgtgagg 3180
cactgtttcc cgaagcctac agcctttctc agcccaggac aatgagctca gaaagtcttt 3240
ttccttctag ggactgcctt tttcacccaa actgttccct ctgccccctc ccccgacaag 3300
gcctgtcctg taagagttag gctagcaggg taggagcaat gctatgaaac cactgctggg 3360
acccaggtct tcccagtcct tcaccccgca ggtcaccctg agaaacacac taagaactcc 3420
atcccccaga caggacagtc ccatctaccc gcagcttgag tcttggccct agtaccttga 3480
gggtgttggg ggtgatggtg taaccatctt cctcaggaat tttcagcccc ggtgggcagg 3540
gcagggcgaa gtcatcgtgc aggtattttc cactcatggc actctctaac agcctgtgga 3600
gaagaaaaga ggtgaggatg ccccactgac tatggttcct ataccatctg ccctcaccca 3660
ttttcctgcc ttacaggtga agacagggaa gccccacagc aaaaaggaga ggcccagact 3720
gtgagttccc agcccggggc tccatctacc ggcttgacgc tggaactggg aagcacagct 3780
ggggttcaga gggcgatggg ctccctaggc ggcccttccc ctgtgactgg ccttccctct 3840
gacacatgcg gctttaccca cctttgtctg tccctgatgt ctactgcact ccacacagag 3900
ccatataggg tcagctgcca ttgccggagg atgccgacct ggaatgactc atcccctagg 3960
gcagcaaggg gcagctcgtg agccacgccc ctctgtcagc catgccagac agcagaggcc 4020
ctgcccgccc tgccacccaa aggggctcag gccacatggt ctgctccagg agctgcctca 4080
ctgtgtccca ctgaccccag gttctgcaga agggcctcac tgggtgccct tagggatgga 4140
aagggttgaa aggctgtact ccaaagcaga gtcttgcttt tctctcccgt attttggggg 4200
ttcagctggg attagaaaaa aatgtctttc caccaaatta aagaaagctt tgaaaaccac 4260
<210> 182
<211> 3156
<212> ДНК
<213> Homo sapiens
<400> 182
acagctgtgt ttggctgcag ggccaagagc gctgtcaaga agacccacac gcccccctcc 60
agcagctgaa ttcctgcagc tcagcagccg ccgccagagc aggacgaacc gccaatcgca 120
aggcacctct gagaacttca ggatgcagat gtctccagcc ctcacctgcc tagtcctggg 180
cctggccctt gtctttggtg aagggtctgc tgtgcaccat cccccatcct acgtggccca 240
cctggcctca gacttcgggg tgagggtgtt tcagcaggtg gcgcaggcct ccaaggaccg 300
caacgtggtt ttctcaccct atggggtggc ctcggtgttg gccatgctcc agctgacaac 360
aggaggagaa acccagcagc agattcaagc agctatggga ttcaagattg atgacaaggg 420
catggccccc gccctccggc atctgtacaa ggagctcatg gggccatgga acaaggatga 480
gatcagcacc acagacgcga tcttcgtcca gcgggatctg aagctggtcc agggcttcat 540
gccccacttc ttcaggctgt tccggagcac ggtcaagcaa gtggactttt cagaggtgga 600
gagagccaga ttcatcatca atgactgggt gaagacacac acaaaaggta tgatcagcaa 660
cttgcttggg aaaggagccg tggaccagct gacacggctg gtgctggtga atgccctcta 720
cttcaacggc cagtggaaga ctcccttccc cgactccagc acccaccgcc gcctcttcca 780
caaatcagac ggcagcactg tctctgtgcc catgatggct cagaccaaca agttcaacta 840
tactgagttc accacgcccg atggccatta ctacgacatc ctggaactgc cctaccacgg 900
ggacaccctc agcatgttca ttgctgcccc ttatgaaaaa gaggtgcctc tctctgccct 960
caccaacatt ctgagtgccc agctcatcag ccactggaaa ggcaacatga ccaggctgcc 1020
ccgcctcctg gttctgccca agttctccct ggagactgaa gtcgacctca ggaagcccct 1080
agagaacctg ggaatgaccg acatgttcag acagtttcag gctgacttca cgagtctttc 1140
agaccaagag cctctccacg tcgcgcaggc gctgcagaaa gtgaagatcg aggtgaacga 1200
gagtggcacg gtggcctcct catccacagc tgtcatagtc tcagcccgca tggcccccga 1260
ggagatcatc atggacagac ccttcctctt tgtggtccgg cacaacccca caggaacagt 1320
ccttttcatg ggccaagtga tggaaccctg accctgggga aagacgcctt catctgggac 1380
aaaactggag atgcatcggg aaagaagaaa ctccgaagaa aagaatttta gtgttaatga 1440
ctctttctga aggaagagaa gacatttgcc ttttgttaaa agatggtaaa ccagatctgt 1500
ctccaagacc ttggcctctc cttggaggac ctttaggtca aactccctag tctccacctg 1560
agaccctggg agagaagttt gaagcacaac tcccttaagg tctccaaacc agacggtgac 1620
gcctgcggga ccatctgggg cacctgcttc cacccgtctc tctgcccact cgggtctgca 1680
gacctggttc ccactgaggc cctttgcagg atggaactac ggggcttaca ggagcttttg 1740
tgtgcctggt agaaactatt tctgttccag tcacattgcc atcactcttg tactgcctgc 1800
caccgcggag gaggctggtg acaggccaaa ggccagtgga agaaacaccc tttcatctca 1860
gagtccactg tggcactggc cacccctccc cagtacaggg gtgctgcagg tggcagagtg 1920
aatgtccccc atcatgtggc ccaactctcc tggcctggcc atctccctcc ccagaaacag 1980
tgtgcatggg ttattttgga gtgtaggtga cttgtttact cattgaagca gatttctgct 2040
tccttttatt tttataggaa tagaggaaga aatgtcagat gcgtgcccag ctcttcaccc 2100
cccaatctct tggtggggag gggtgtacct aaatatttat catatccttg cccttgagtg 2160
cttgttagag agaaagagaa ctactaagga aaataatatt atttaaactc gctcctagtg 2220
tttctttgtg gtctgtgtca ccgtatctca ggaagtccag ccacttgact ggcacacacc 2280
cctccggaca tccagcgtga cggagcccac actgccacct tgtggccgcc tgagaccctc 2340
gcgccccccg cgcccctctt tttccccttg atggaaattg accatacaat ttcatcctcc 2400
ttcaggggat caaaaggacg gagtgggggg acagagactc agatgaggac agagtggttt 2460
ccaatgtgtt caatagattt aggagcagaa atgcaagggg ctgcatgacc taccaggaca 2520
gaactttccc caattacagg gtgactcaca gccgcattgg tgactcactt caatgtgtca 2580
tttccggctg ctgtgtgtga gcagtggaca cgtgaggggg gggtgggtga gagagacagg 2640
cagctcggat tcaactacct tagataatat ttctgaaaac ctaccagcca gagggtaggg 2700
cacaaagatg gatgtaatgc actttgggag gccaaggcgg gaggattgct tgagcccagg 2760
agttcaagac cagcctgggc aacataccaa gacccccgtc tctttaaaaa tatatatatt 2820
ttaaatatac ttaaatatat atttctaata tctttaaata tatatatata ttttaaagac 2880
caatttatgg gagaattgca cacagatgtg aaatgaatgt aatctaatag aagcctaatc 2940
agcccaccat gttctccact gaaaaatcct ctttctttgg ggtttttctt tctttctttt 3000
ttgattttgc actggacggt gacgtcagcc atgtacagga tccacagggg tggtgtcaaa 3060
tgctattgaa attgtgttga attgtatgct ttttcacttt tgataaataa acatgtaaaa 3120
atgtttcaaa aaaataataa aataaataaa tacgaa 3156
<210> 183
<211> 5490
<212> ДНК
<213> Homo sapiens
<400> 183
ggctgagttt tatgacgggc ccggtgctga agggcaggga acaacttgat ggtgctactt 60
tgaactgctt ttcttttctc ctttttgcac aaagagtctc atgtctgata tttagacatg 120
atgagctttg tgcaaaaggg gagctggcta cttctcgctc tgcttcatcc cactattatt 180
ttggcacaac aggaagctgt tgaaggagga tgttcccatc ttggtcagtc ctatgcggat 240
agagatgtct ggaagccaga accatgccaa atatgtgtct gtgactcagg atccgttctc 300
tgcgatgaca taatatgtga cgatcaagaa ttagactgcc ccaacccaga aattccattt 360
ggagaatgtt gtgcagtttg cccacagcct ccaactgctc ctactcgccc tcctaatggt 420
caaggacctc aaggccccaa gggagatcca ggccctcctg gtattcctgg gagaaatggt 480
gaccctggta ttccaggaca accagggtcc cctggttctc ctggcccccc tggaatctgt 540
gaatcatgcc ctactggtcc tcagaactat tctccccagt atgattcata tgatgtcaag 600
tctggagtag cagtaggagg actcgcaggc tatcctggac cagctggccc cccaggccct 660
cccggtcccc ctggtacatc tggtcatcct ggttcccctg gatctccagg ataccaagga 720
ccccctggtg aacctgggca agctggtcct tcaggccctc caggacctcc tggtgctata 780
ggtccatctg gtcctgctgg aaaagatgga gaatcaggta gacccggacg acctggagag 840
cgaggattgc ctggacctcc aggtatcaaa ggtccagctg ggatacctgg attccctggt 900
atgaaaggac acagaggctt cgatggacga aatggagaaa agggtgaaac aggtgctcct 960
ggattaaagg gtgaaaatgg tcttccaggc gaaaatggag ctcctggacc catgggtcca 1020
agaggggctc ctggtgagcg aggacggcca ggacttcctg gggctgcagg tgctcggggt 1080
aatgacggtg ctcgaggcag tgatggtcaa ccaggccctc ctggtcctcc tggaactgcc 1140
ggattccctg gatcccctgg tgctaagggt gaagttggac ctgcagggtc tcctggttca 1200
aatggtgccc ctggacaaag aggagaacct ggacctcagg gacacgctgg tgctcaaggt 1260
cctcctggcc ctcctgggat taatggtagt cctggtggta aaggcgaaat gggtcccgct 1320
ggcattcctg gagctcctgg actgatggga gcccggggtc ctccaggacc agccggtgct 1380
aatggtgctc ctggactgcg aggtggtgca ggtgagcctg gtaagaatgg tgccaaagga 1440
gagcccggac cacgtggtga acgcggtgag gctggtattc caggtgttcc aggagctaaa 1500
ggcgaagatg gcaaggatgg atcacctgga gaacctggtg caaatgggct tccaggagct 1560
gcaggagaaa ggggtgcccc tgggttccga ggacctgctg gaccaaatgg catcccagga 1620
gaaaagggtc ctgctggaga gcgtggtgct ccaggccctg cagggcccag aggagctgct 1680
ggagaacctg gcagagatgg cgtccctgga ggtccaggaa tgaggggcat gcccggaagt 1740
ccaggaggac caggaagtga tgggaaacca gggcctcccg gaagtcaagg agaaagtggt 1800
cgaccaggtc ctcctgggcc atctggtccc cgaggtcagc ctggtgtcat gggcttcccc 1860
ggtcctaaag gaaatgatgg tgctcctggt aagaatggag aacgaggtgg ccctggagga 1920
cctggccctc agggtcctcc tggaaagaat ggtgaaactg gacctcaggg acccccaggg 1980
cctactgggc ctggtggtga caaaggagac acaggacccc ctggtccaca aggattacaa 2040
ggcttgcctg gtacaggtgg tcctccagga gaaaatggaa aacctgggga accaggtcca 2100
aagggtgatg ccggtgcacc tggagctcca ggaggcaagg gtgatgctgg tgcccctggt 2160
gaacgtggac ctcctggatt ggcaggggcc ccaggactta gaggtggagc tggtccccct 2220
ggtcccgaag gaggaaaggg tgctgctggt cctcctgggc cacctggtgc tgctggtact 2280
cctggtctgc aaggaatgcc tggagaaaga ggaggtcttg gaagtcctgg tccaaagggt 2340
gacaagggtg aaccaggcgg tccaggtgct gatggtgtcc cagggaaaga tggcccaagg 2400
ggtcctactg gtcctattgg tcctcctggc ccagctggcc agcctggaga taagggtgaa 2460
ggtggtgccc ccggacttcc aggtatagct ggacctcgtg gtagccctgg tgagagaggt 2520
gaaactggcc ctccaggacc tgctggtttc cctggtgctc ctggacagaa tggtgaacct 2580
ggtggtaaag gagaaagagg ggctccgggt gagaaaggtg aaggaggccc tcctggagtt 2640
gcaggacccc ctggaggttc tggacctgct ggtcctcctg gtccccaagg tgtcaaaggt 2700
gaacgtggca gtcctggtgg acctggtgct gctggcttcc ctggtgctcg tggtcttcct 2760
ggtcctcctg gtagtaatgg taacccagga cccccaggtc ccagcggttc tccaggcaag 2820
gatgggcccc caggtcctgc gggtaacact ggtgctcctg gcagccctgg agtgtctgga 2880
ccaaaaggtg atgctggcca accaggagag aagggatcgc ctggtgccca gggcccacca 2940
ggagctccag gcccacttgg gattgctggg atcactggag cacggggtct tgcaggacca 3000
ccaggcatgc caggtcctag gggaagccct ggccctcagg gtgtcaaggg tgaaagtggg 3060
aaaccaggag ctaacggtct cagtggagaa cgtggtcccc ctggacccca gggtcttcct 3120
ggtctggctg gtacagctgg tgaacctgga agagatggaa accctggatc agatggtctt 3180
ccaggccgag atggatctcc tggtggcaag ggtgatcgtg gtgaaaatgg ctctcctggt 3240
gcccctggcg ctcctggtca tccaggccca cctggtcctg tcggtccagc tggaaagagt 3300
ggtgacagag gagaaagtgg ccctgctggc cctgctggtg ctcccggtcc tgctggttcc 3360
cgaggtgctc ctggtcctca aggcccacgt ggtgacaaag gtgaaacagg tgaacgtgga 3420
gctgctggca tcaaaggaca tcgaggattc cctggtaatc caggtgcccc aggttctcca 3480
ggccctgctg gtcagcaggg tgcaatcggc agtccaggac ctgcaggccc cagaggacct 3540
gttggaccca gtggacctcc tggcaaagat ggaaccagtg gacatccagg tcccattgga 3600
ccaccagggc ctcgaggtaa cagaggtgaa agaggatctg agggctcccc aggccaccca 3660
gggcaaccag gccctcctgg acctcctggt gcccctggtc cttgctgtgg tggtgttgga 3720
gccgctgcca ttgctgggat tggaggtgaa aaagctggcg gttttgcccc gtattatgga 3780
gatgaaccaa tggatttcaa aatcaacacc gatgagatta tgacttcact caagtctgtt 3840
aatggacaaa tagaaagcct cattagtcct gatggttctc gtaaaaaccc cgctagaaac 3900
tgcagagacc tgaaattctg ccatcctgaa ctcaagagtg gagaatactg ggttgaccct 3960
aaccaaggat gcaaattgga tgctatcaag gtattctgta atatggaaac tggggaaaca 4020
tgcataagtg ccaatccttt gaatgttcca cggaaacact ggtggacaga ttctagtgct 4080
gagaagaaac acgtttggtt tggagagtcc atggatggtg gttttcagtt tagctacggc 4140
aatcctgaac ttcctgaaga tgtccttgat gtgcatctgg cattccttcg acttctctcc 4200
agccgagctt cccagaacat cacatatcac tgcaaaaata gcattgcata catggatcag 4260
gccagtggaa atgtaaagaa ggccctgaag ctgatggggt caaatgaagg tgaattcaag 4320
gctgaaggaa atagcaaatt cacctacaca gttctggagg atggttgcac gaaacacact 4380
ggggaatgga gcaaaacagt ctttgaatat cgaacacgca aggctgtgag actacctatt 4440
gtagatattg caccctatga cattggtggt cctgatcaag aatttggtgt ggacgttggc 4500
cctgtttgct ttttataaac caaactctat ctgaaatccc aacaaaaaaa atttaactcc 4560
atatgtgttc ctcttgttct aatcttgtca accagtgcaa gtgaccgaca aaattccagt 4620
tatttatttc caaaatgttt ggaaacagta taatttgaca aagaaaaatg atacttctct 4680
ttttttgctg ttccaccaaa tacaattcaa atgctttttg ttttattttt ttaccaattc 4740
caatttcaaa atgtctcaat ggtgctataa taaataaact tcaacactct ttatgataac 4800
aacactgtgt tatattcttt gaatcctagc ccatctgcag agcaatgact gtgctcacca 4860
gtaaaagata acctttcttt ctgaaatagt caaatacgaa attagaaaag ccctccctat 4920
tttaactacc tcaactggtc agaaacacag attgtattct atgagtccca gaagatgaaa 4980
aaaattttat acgttgataa aacttataaa tttcattgat taatctcctg gaagattggt 5040
ttaaaaagaa aagtgtaatg caagaattta aagaaatatt tttaaagcca caattatttt 5100
aatattggat atcaactgct tgtaaaggtg ctcctctttt ttcttgtcat tgctggtcaa 5160
gattactaat atttgggaag gctttaaaga cgcatgttat ggtgctaatg tactttcact 5220
tttaaactct agatcagaat tgttgacttg cattcagaac ataaatgcac aaaatctgta 5280
catgtctccc atcagaaaga ttcattggca tgccacaggg gattctcctc cttcatcctg 5340
taaaggtcaa caataaaaac caaattatgg ggctgctttt gtcacactag catagagaat 5400
gtgttgaaat ttaactttgt aagcttgtat gtggttgttg atcttttttt tccttacaga 5460
cacccataat aaaatatcat attaaaattc 5490
<210> 184
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 184
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Glu Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Ile Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Glu Tyr Ser Gly Tyr Asp Thr Asp Pro Gln Tyr Ser
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
195 200 205
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
210 215 220
Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Pro Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
260 265 270
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
435 440 445
Gly Lys
450
<210> 185
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 185
Glu Ile Val Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Gly Asp Asp
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Ile Leu Leu Ile
35 40 45
Tyr Gly Thr Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Ser Asn Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 186
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 186
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Thr Ser Gly Ile Pro Asp Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu
65 70 75 80
Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ser Ser Asp Lys Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 187
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 187
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 188
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<220>
<221> МОД_ОСТ
<222> (1)..(1)
<223> Преимущественно пироглутаминовая кислота (pE)
<400> 188
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Met Leu Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ser Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВЫСОКОАФФИННЫЕ АНТИ-PD-1 И АНТИ-LAG-3 АНТИТЕЛА И ПОЛУЧЕННЫЕ ИЗ НИХ БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ | 2019 |
|
RU2782381C2 |
| МОДИФИЦИРОВАННЫЕ АНТИТЕЛА IGG, КОТОРЫЕ СВЯЗЫВАЮТСЯ С ТРАНСФОРМИРУЮЩИМ ФАКТОРОМ РОСТА БЕТА-1 С ВЫСОКОЙ АФФИННОСТЬЮ, АВИДНОСТЬЮ И СПЕЦИФИЧНОСТЬЮ | 2016 |
|
RU2728858C2 |
| АНТИТЕЛА, НАЦЕЛИВАЮЩИЕСЯ НА КОСТНУЮ ТКАНЬ | 2018 |
|
RU2801206C2 |
| ДИМЕРЫ SCFV-FC, КОТОРЫЕ СВЯЗЫВАЮТСЯ С ТРАНСФОРМИРУЮЩИМ ФАКТОРОМ РОСТА β1 С ВЫСОКОЙ АФФИННОСТЬЮ, АВИДНОСТЬЮ И СПЕЦИФИЧНОСТЬЮ | 2016 |
|
RU2733286C2 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, ОТНОСЯЩИЕСЯ К НИМ СПОСОБЫ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2822200C2 |
| АНТИТЕЛА К LY6G6D И СПОСОБЫ ПРИМЕНЕНИЯ | 2020 |
|
RU2818569C1 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, СВЯЗЫВАЮЩИЕ CD38, CD28 И CD3, А ТАКЖЕ СПОСОБЫ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ВИРУСНОЙ ИНФЕКЦИИ | 2019 |
|
RU2820351C2 |
| АНТИТЕЛА К CD38 И КОМБИНАЦИИ С АНТИТЕЛАМИ К CD3 И CD28 | 2018 |
|
RU2812910C2 |
| Антитело против FXI/FXIA, его антигенсвязывающий фрагмент и фармацевтическое применение | 2021 |
|
RU2838408C1 |
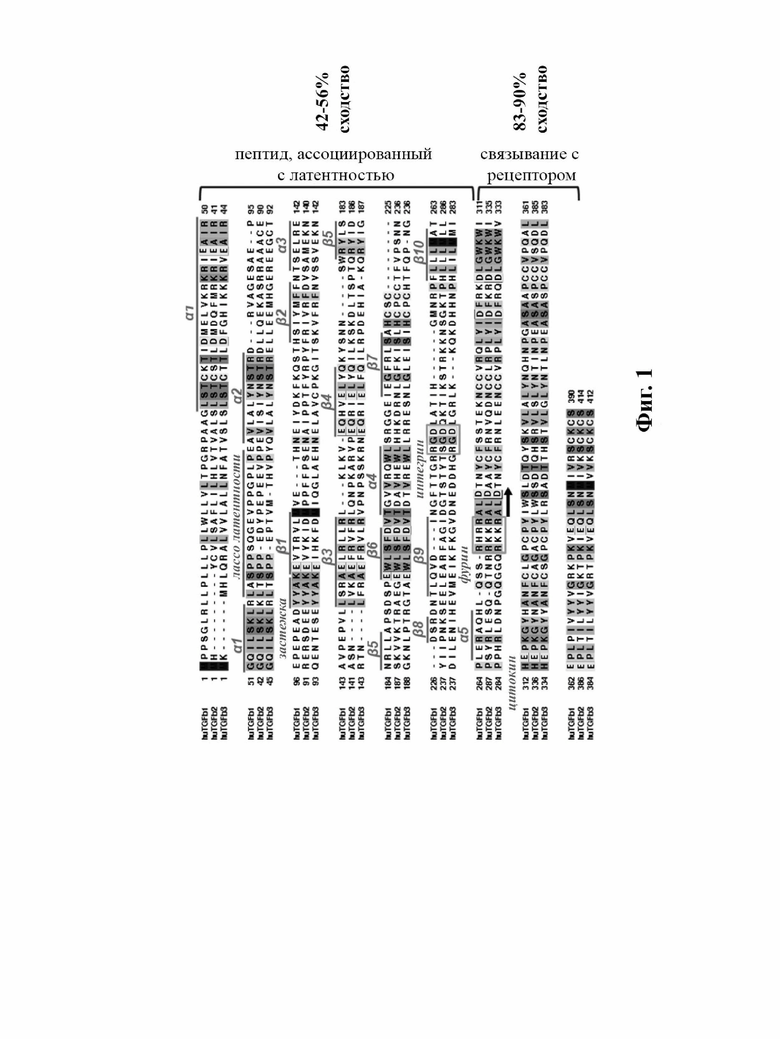
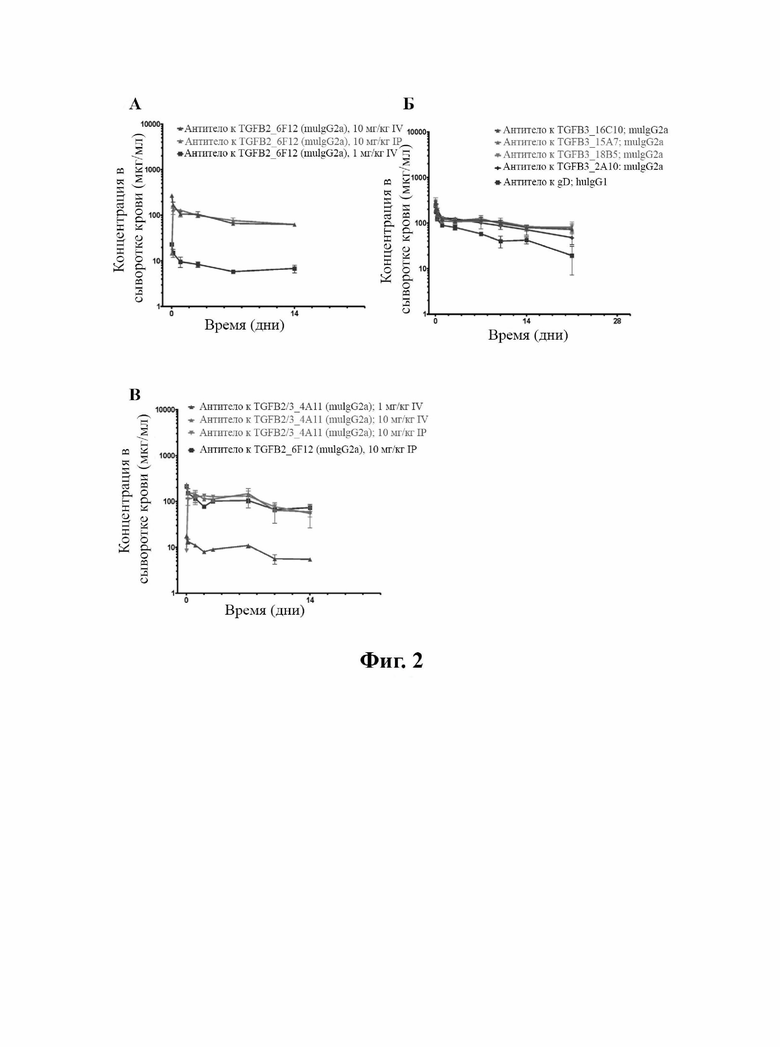

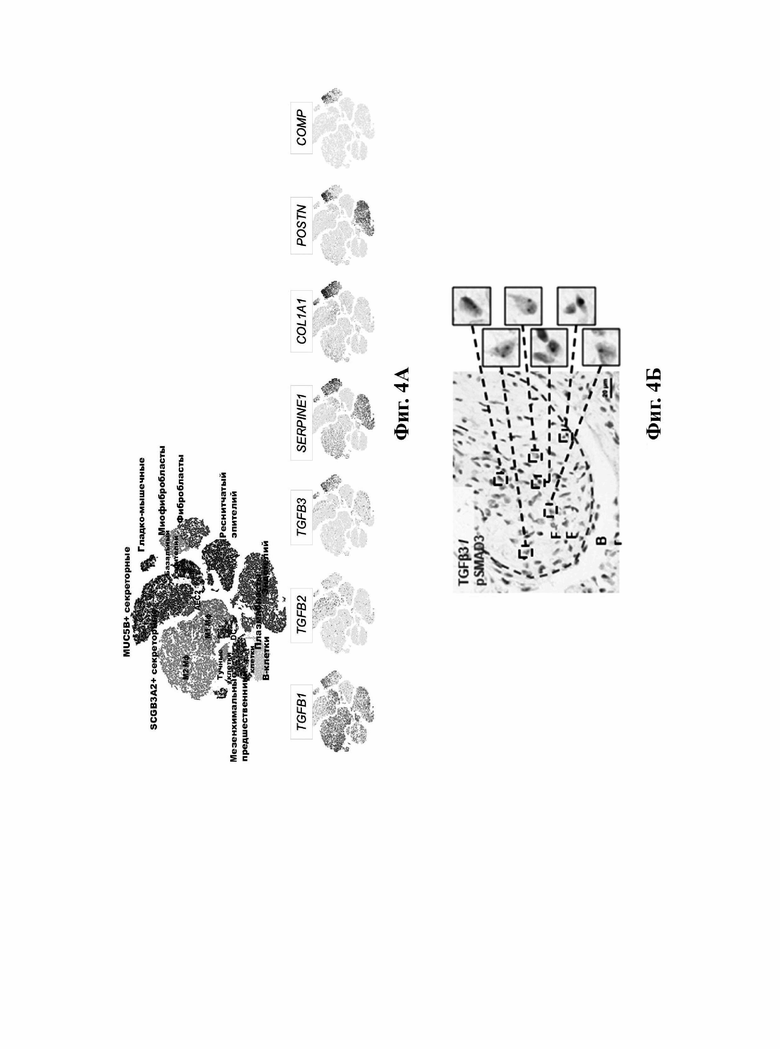
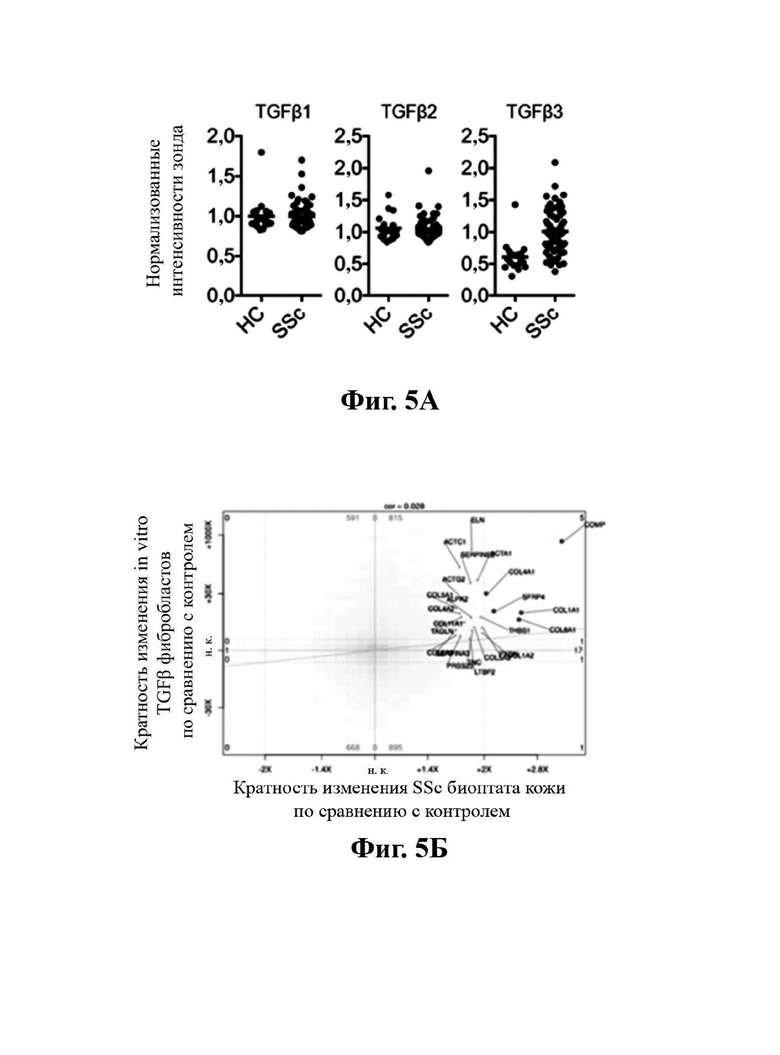

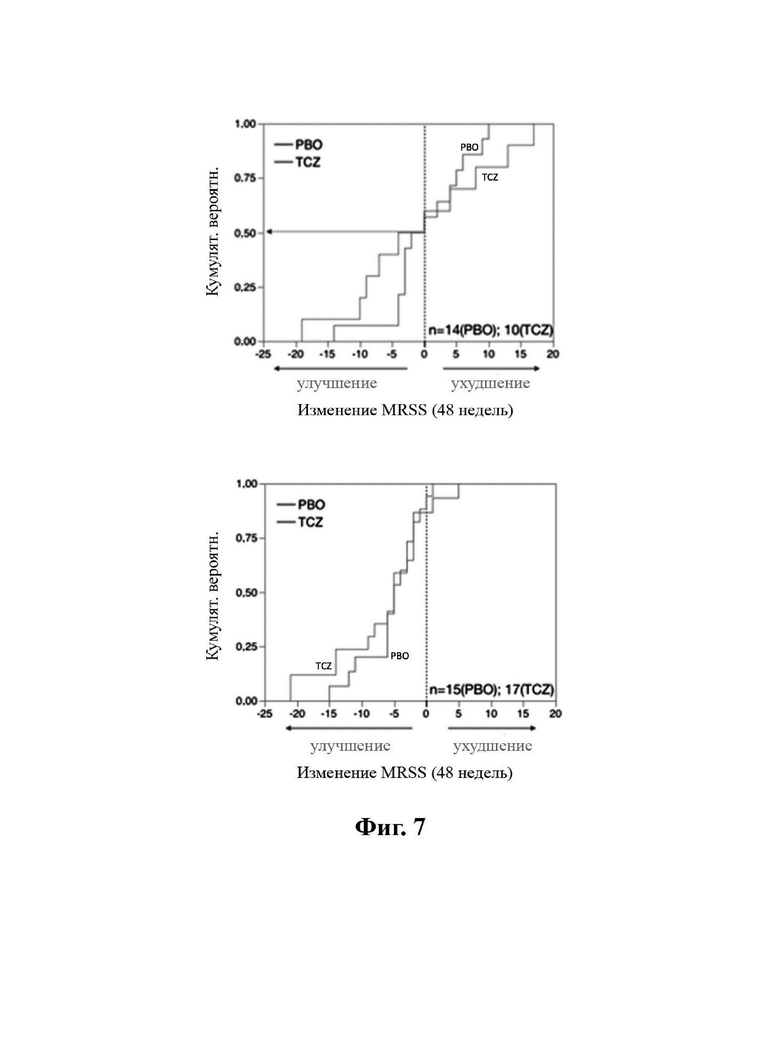
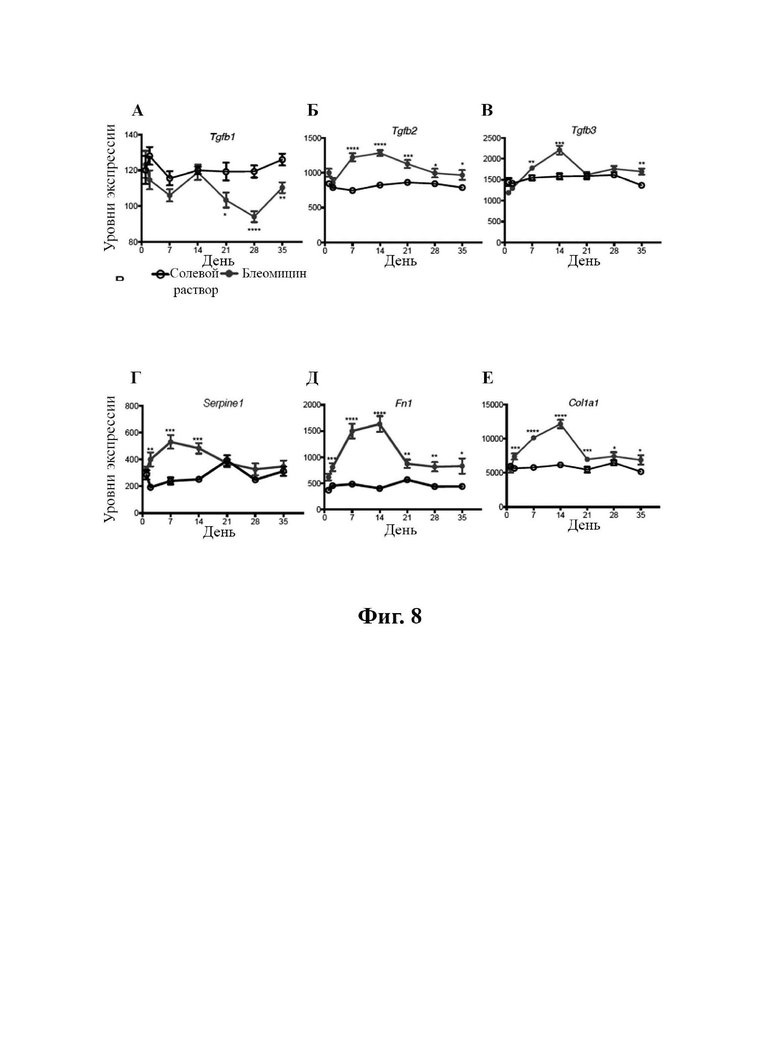
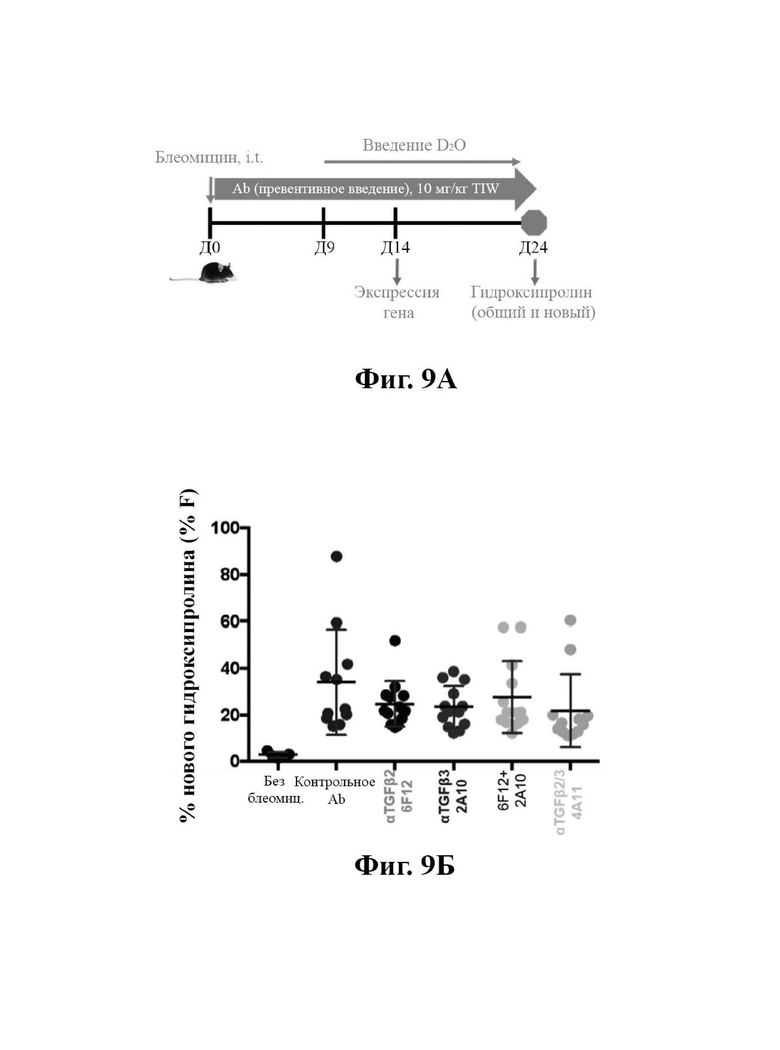
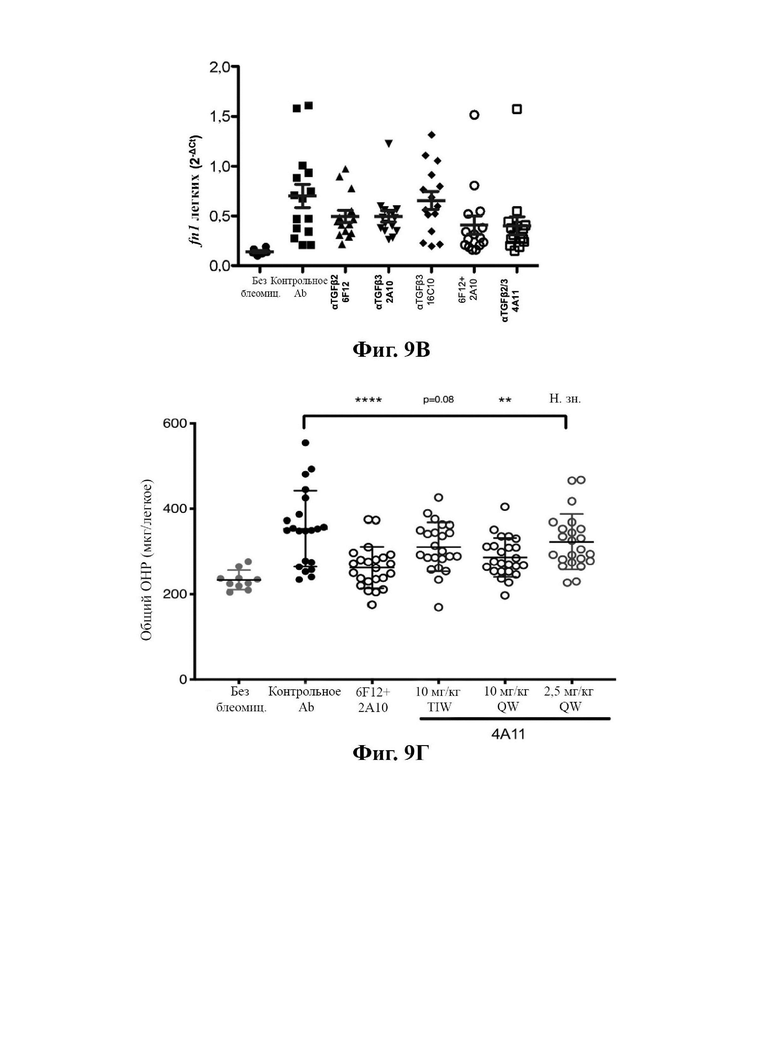
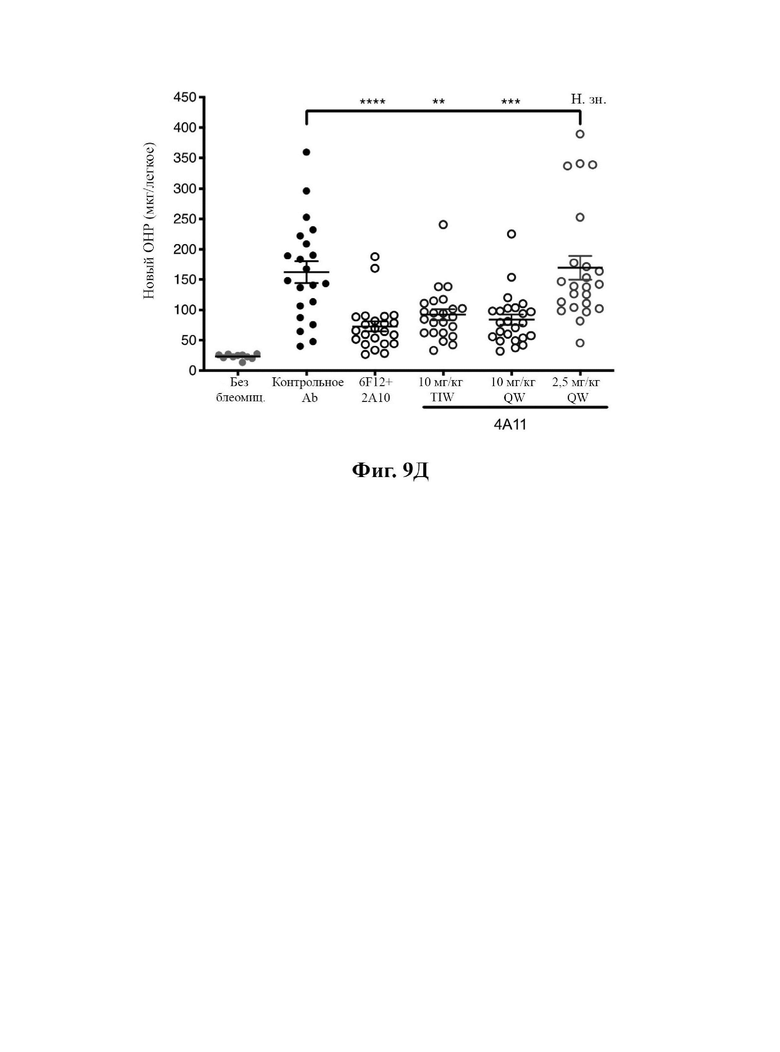
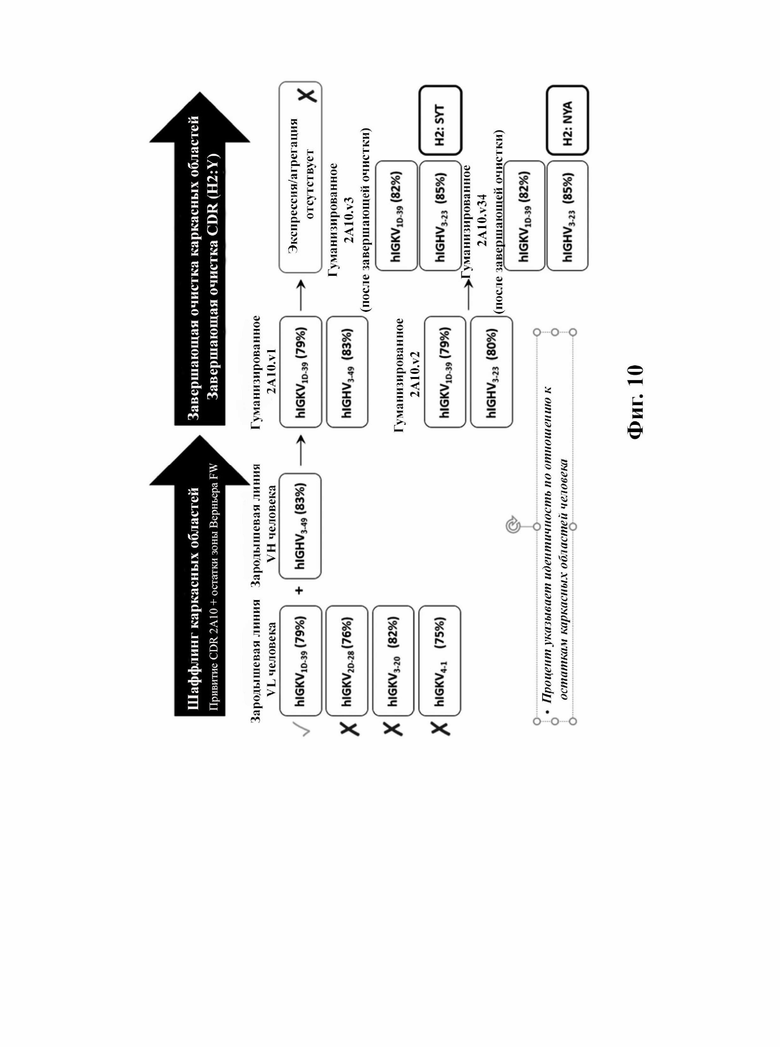
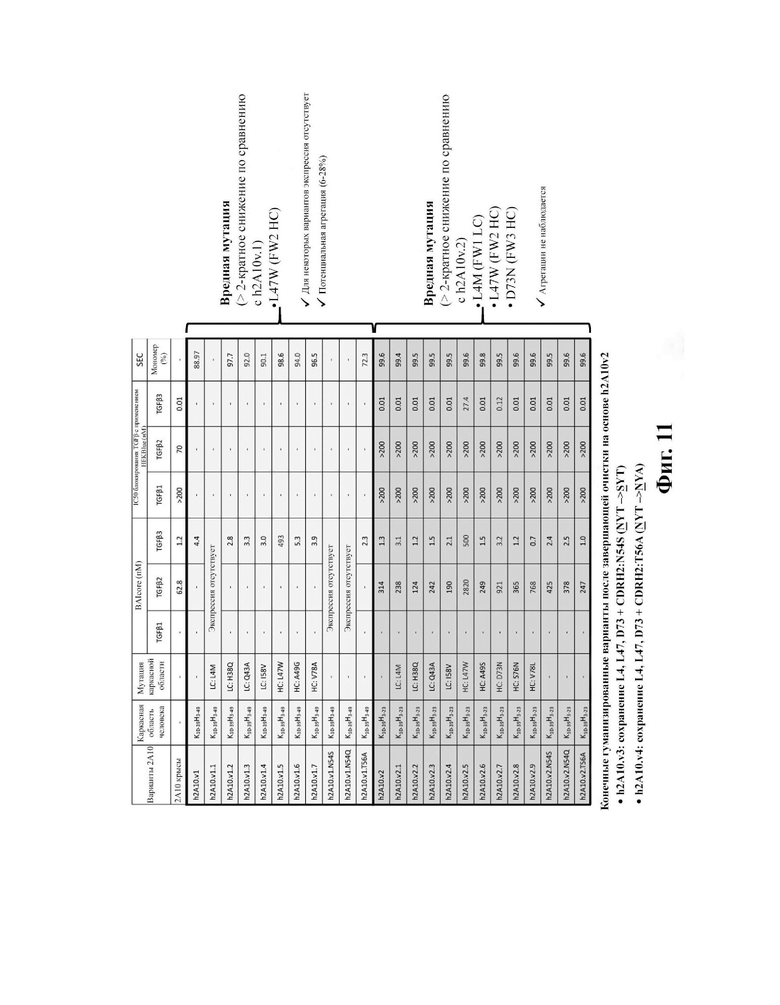
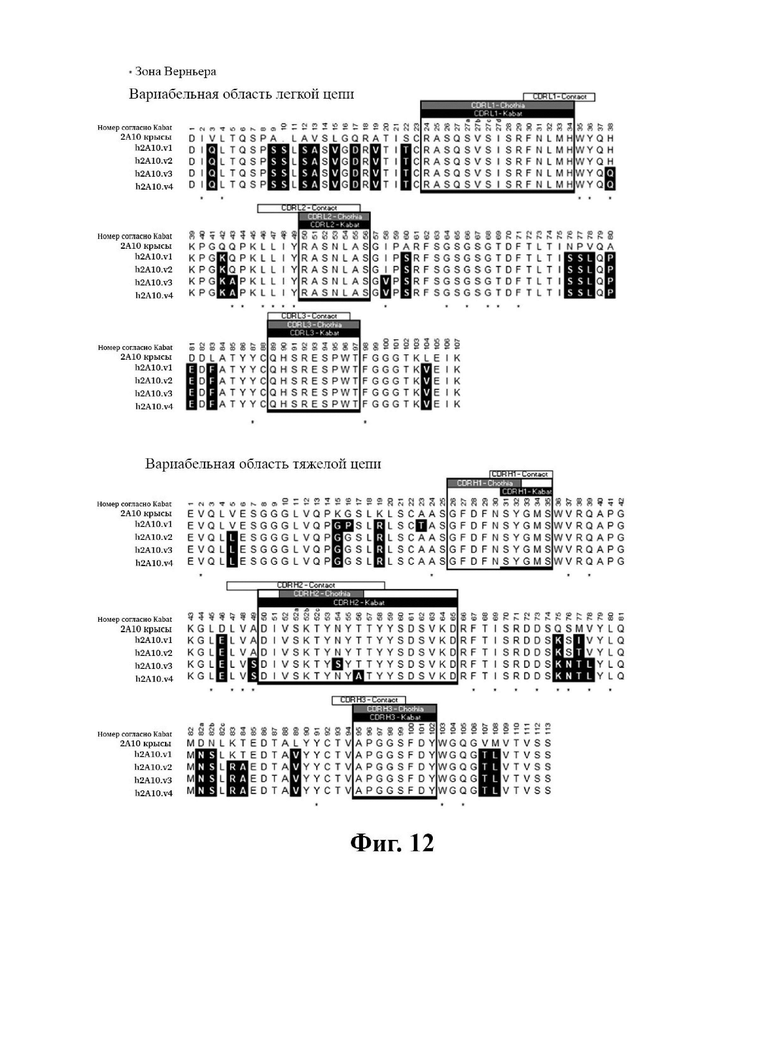

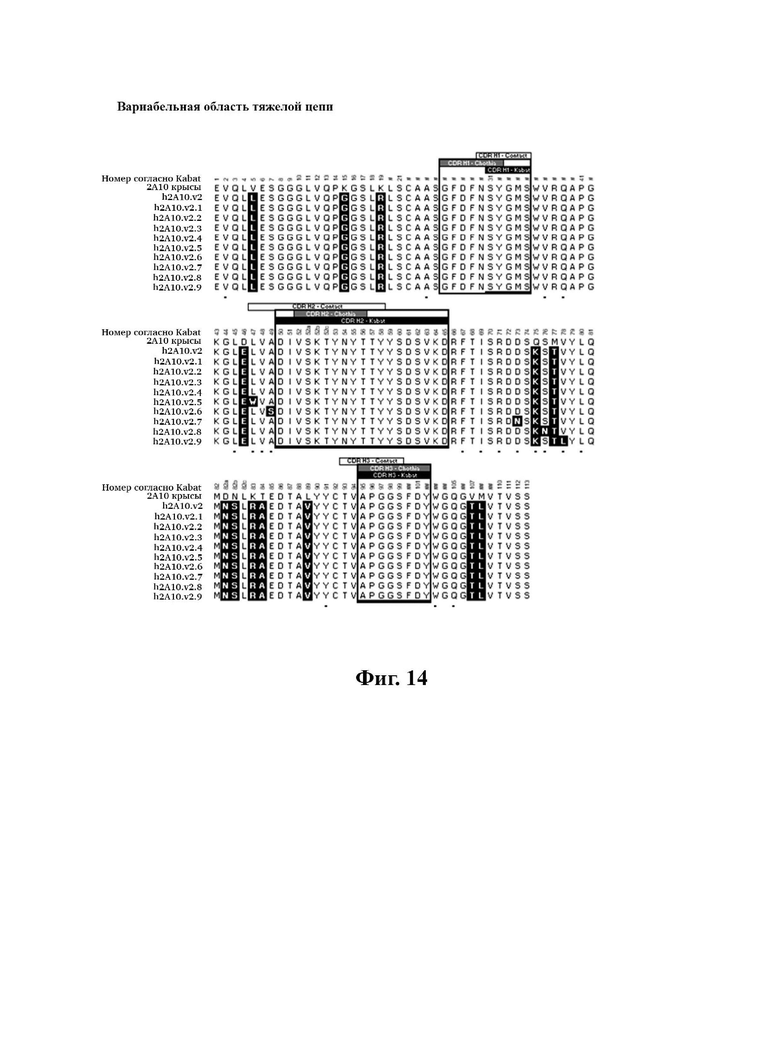
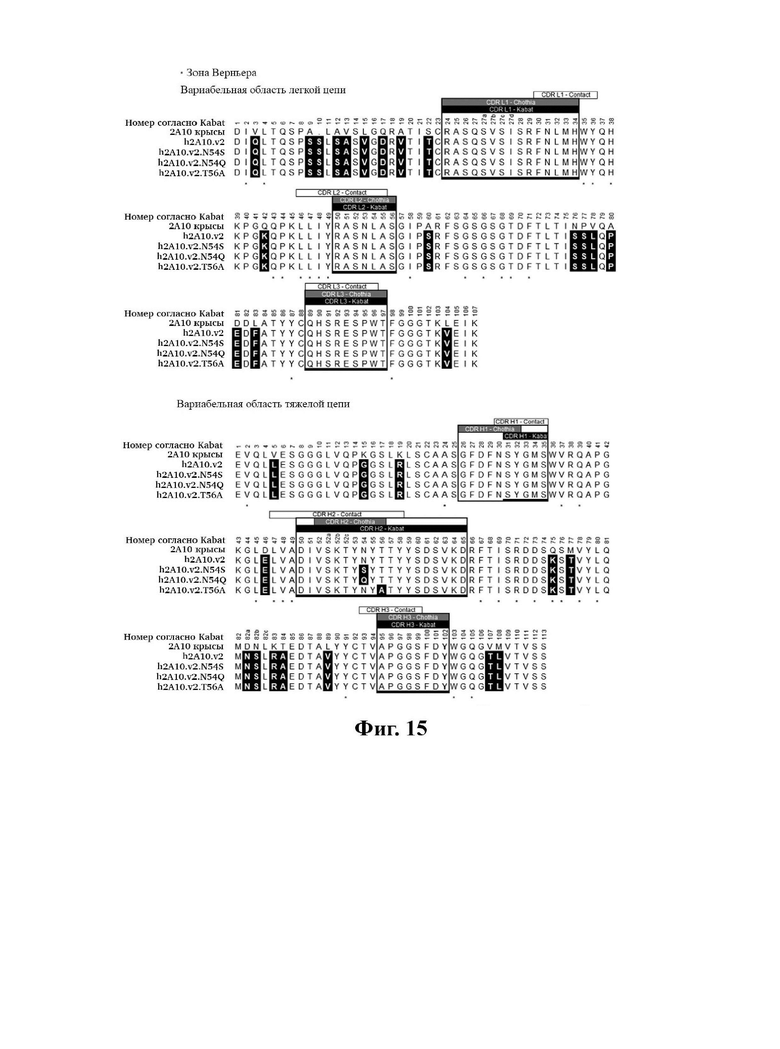
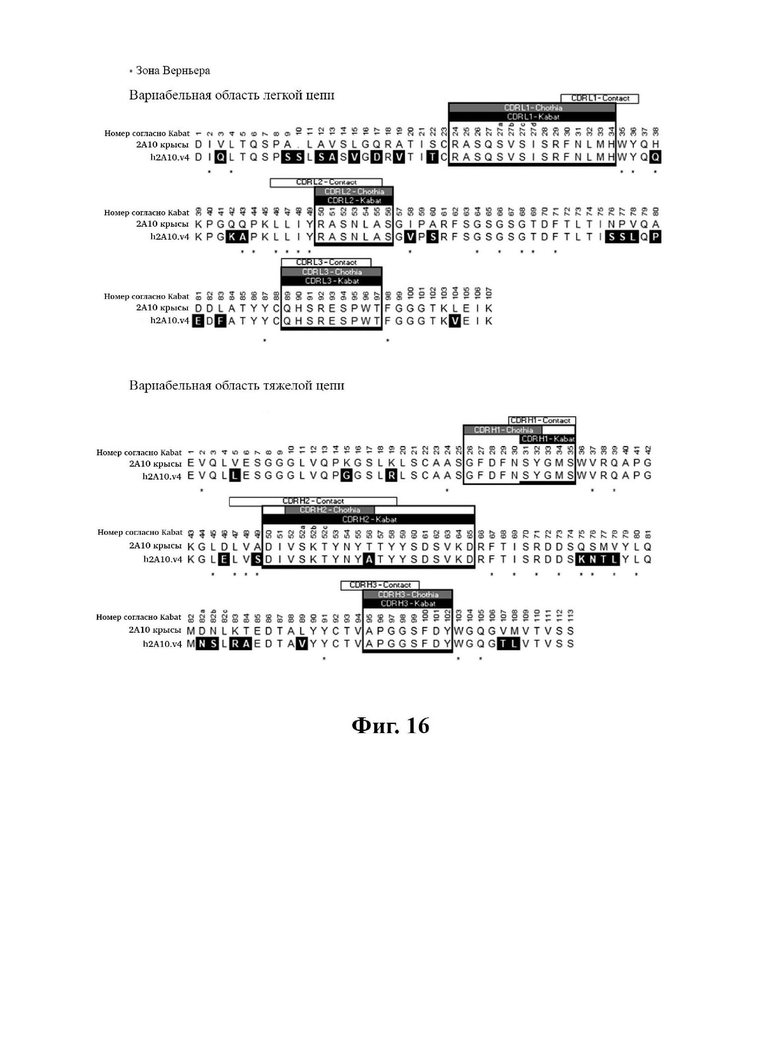
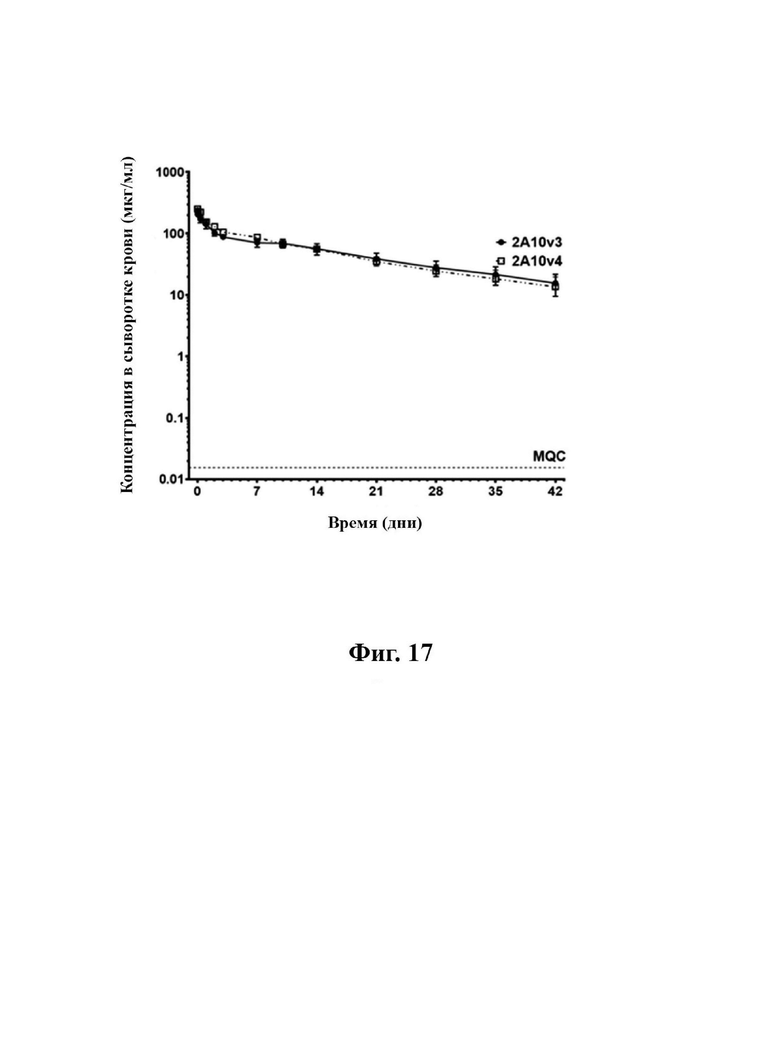


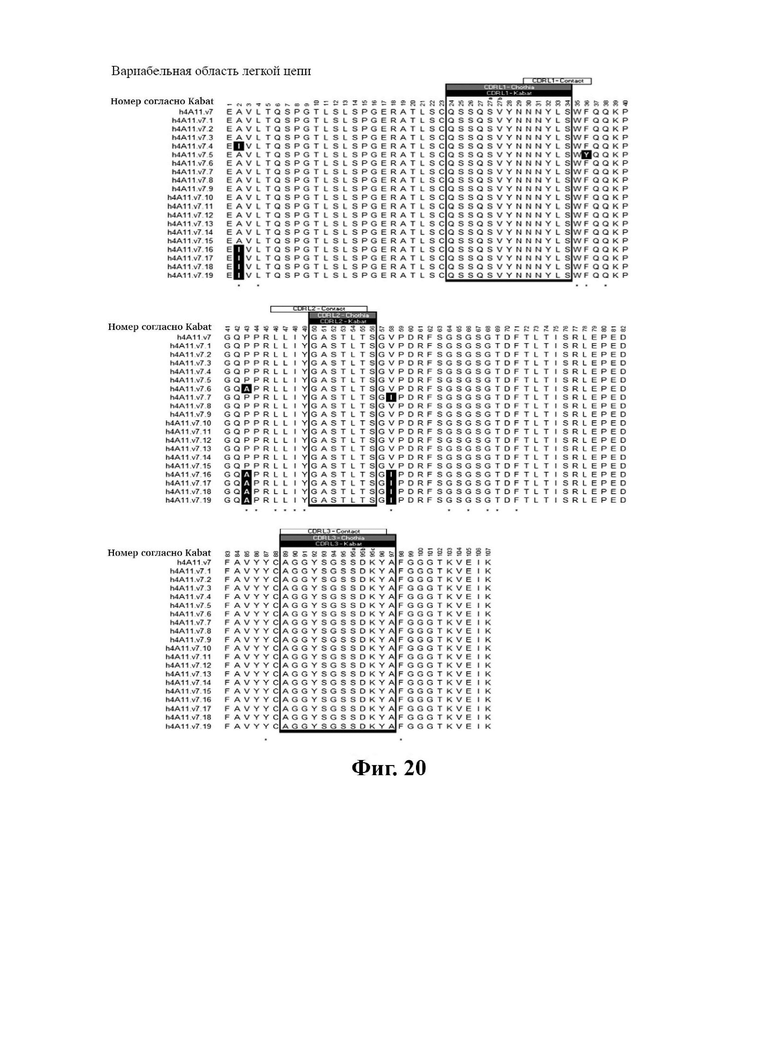
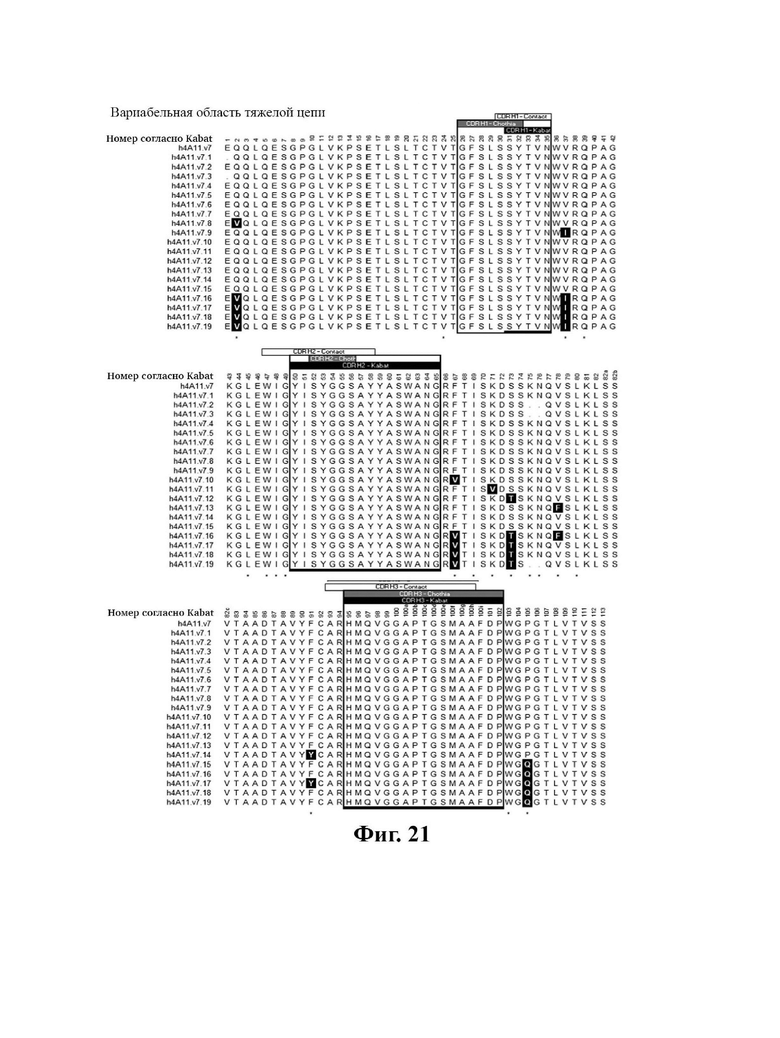
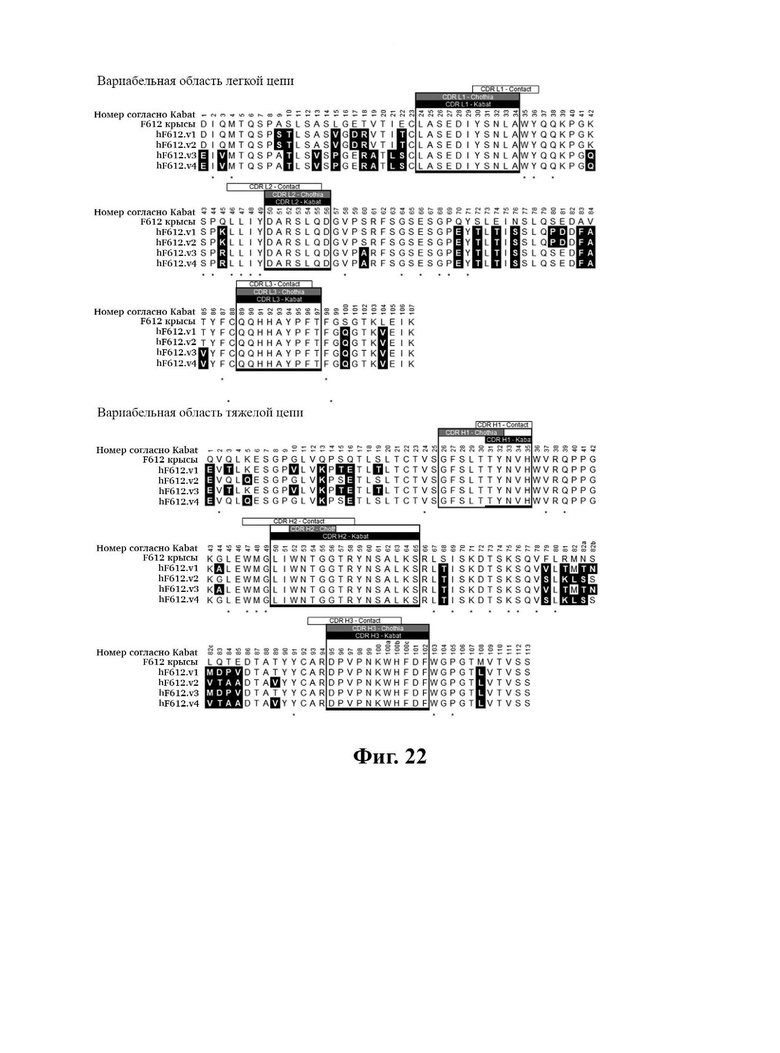
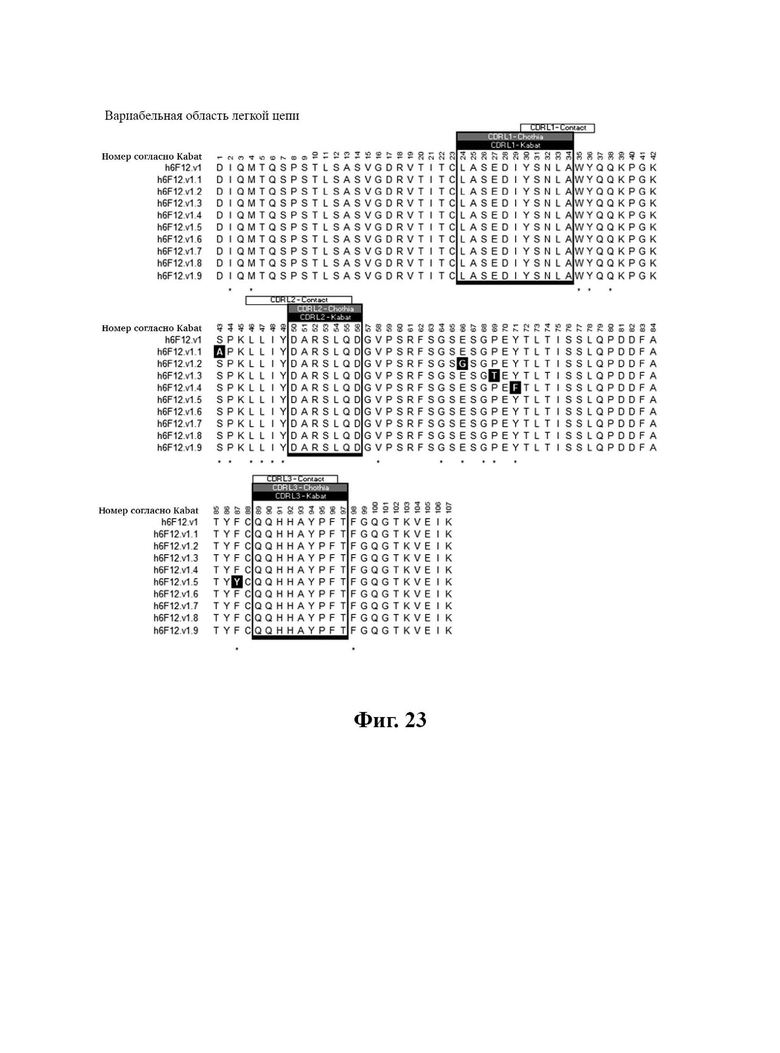
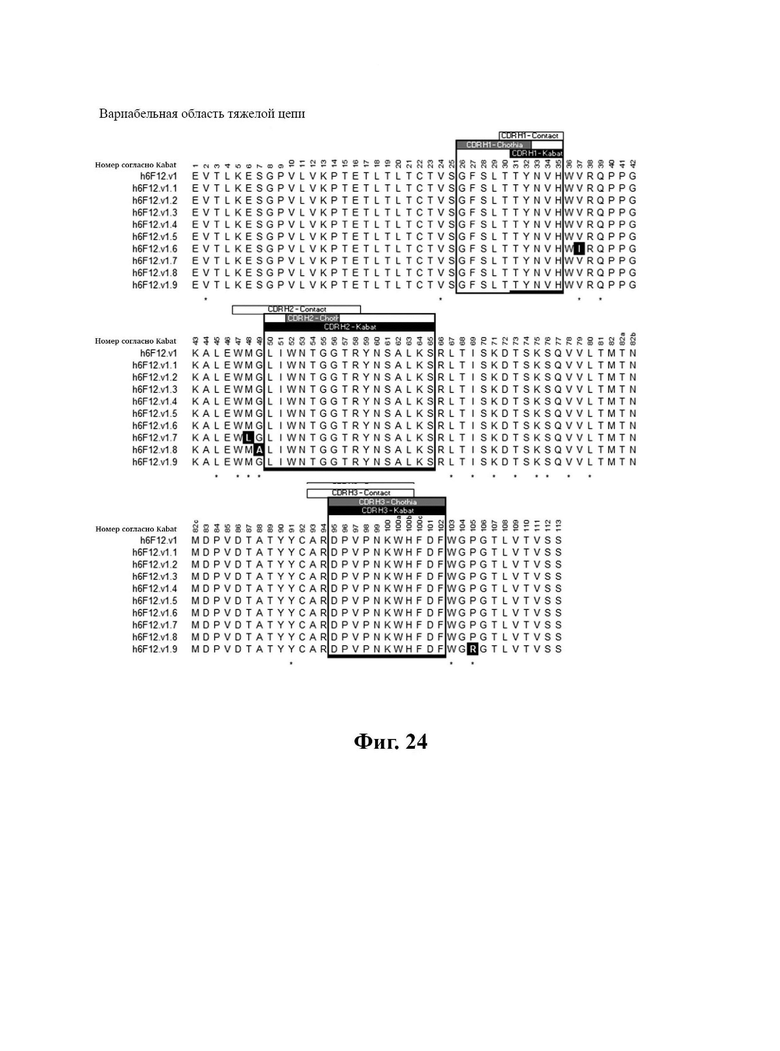
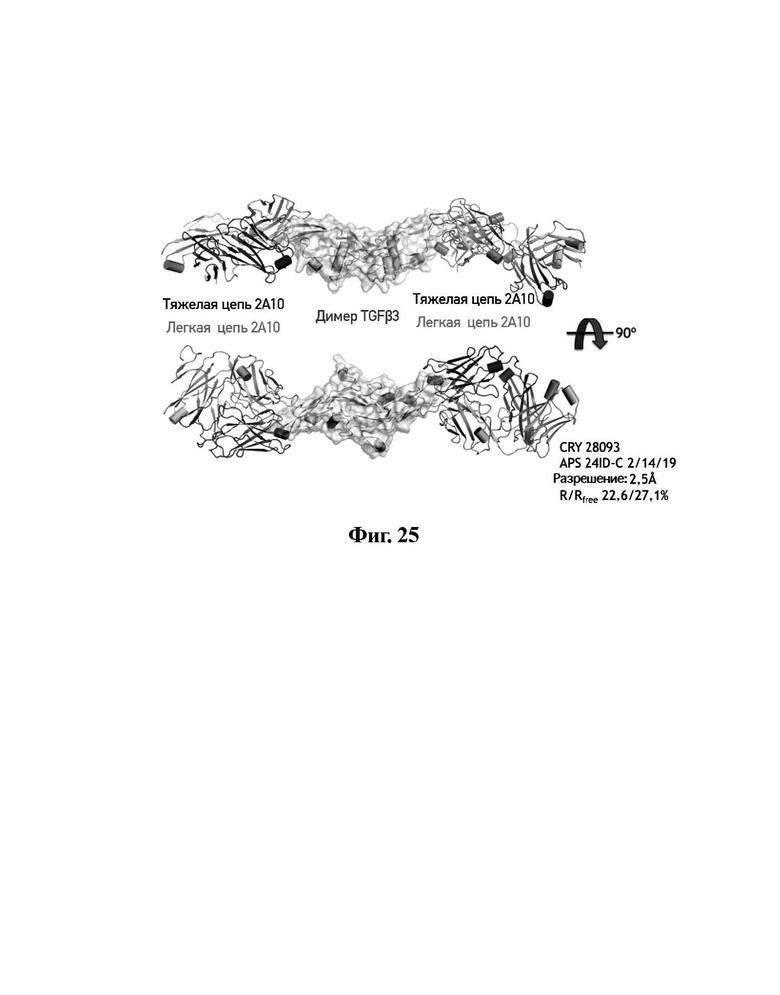

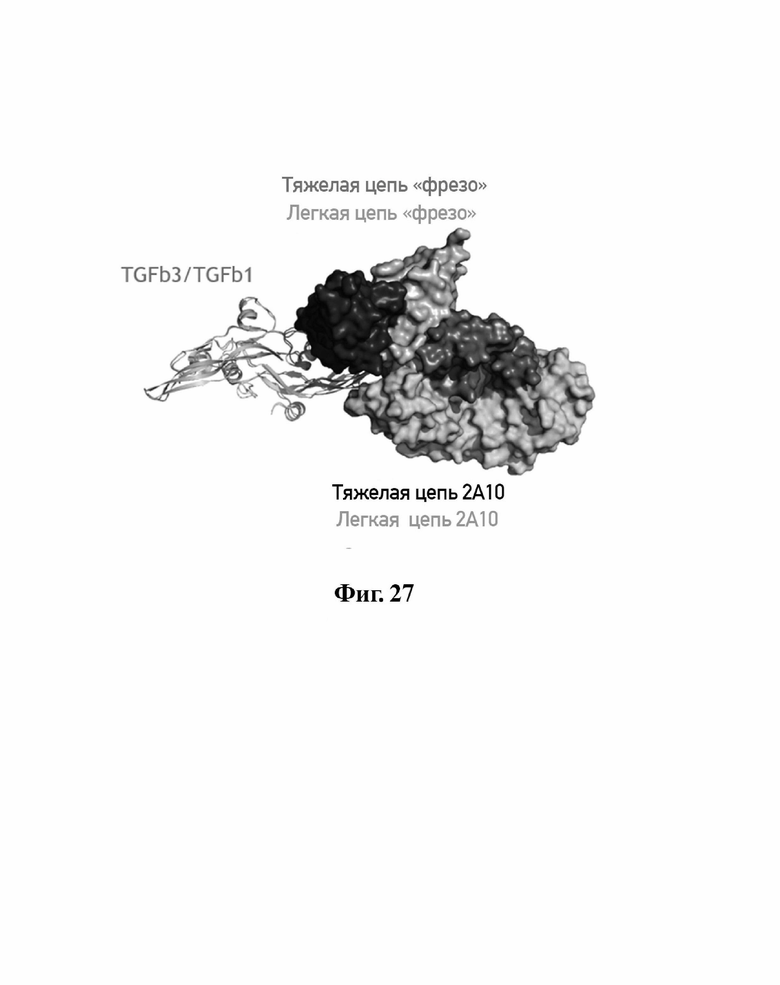
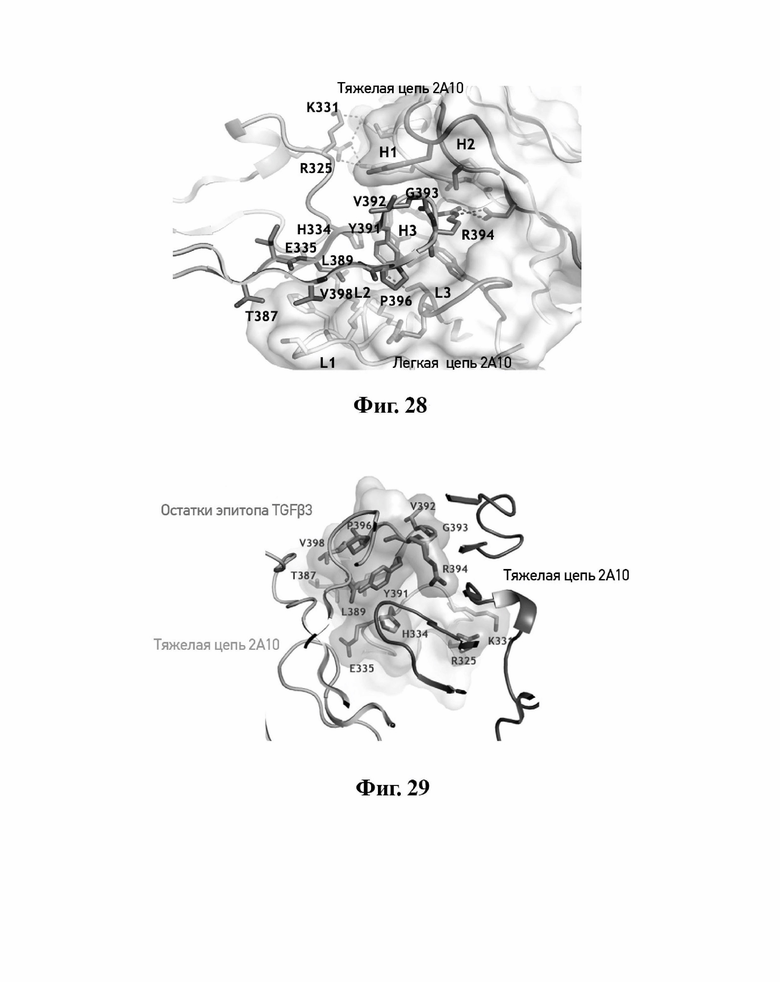

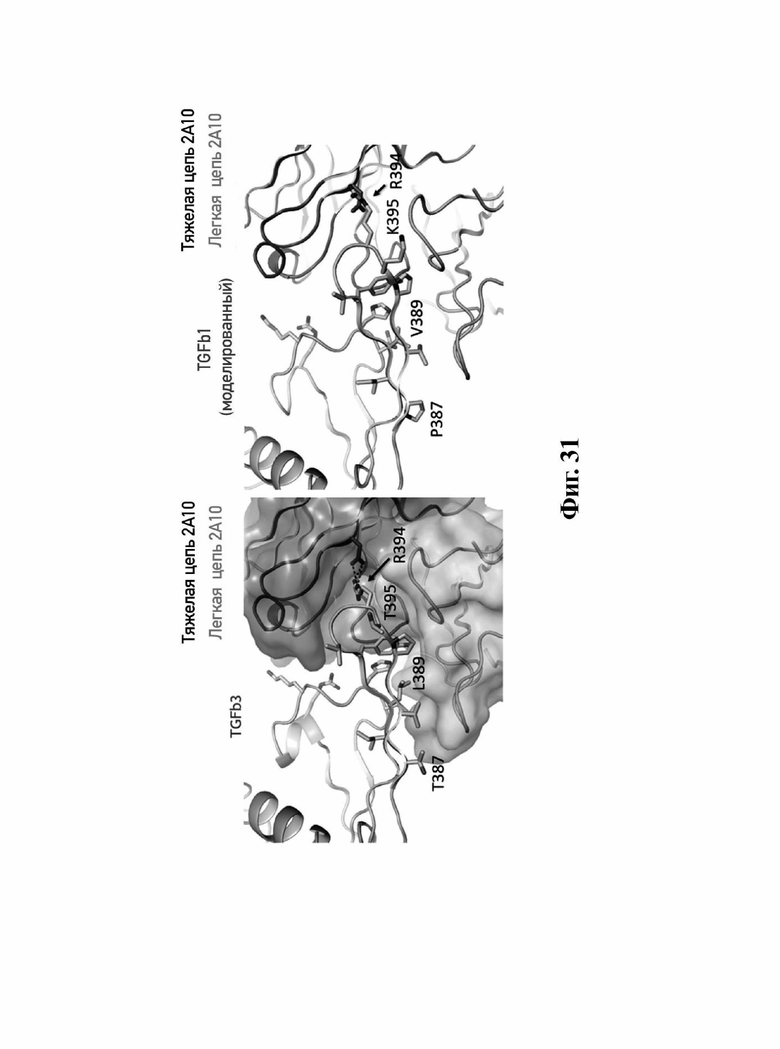
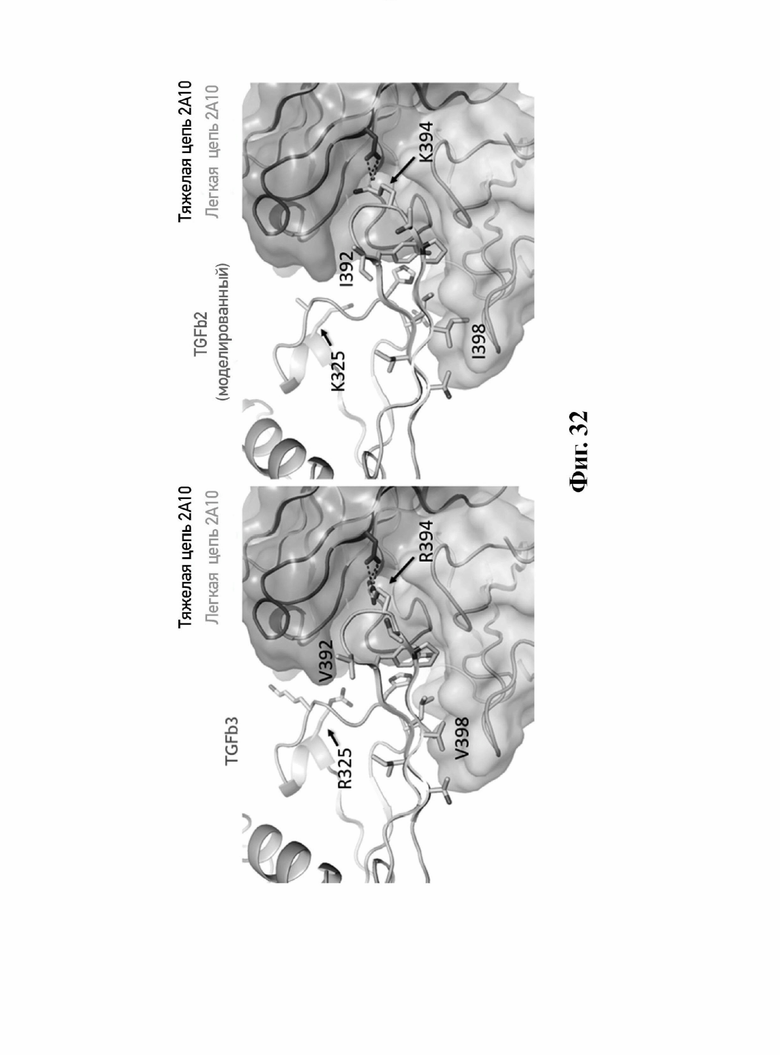
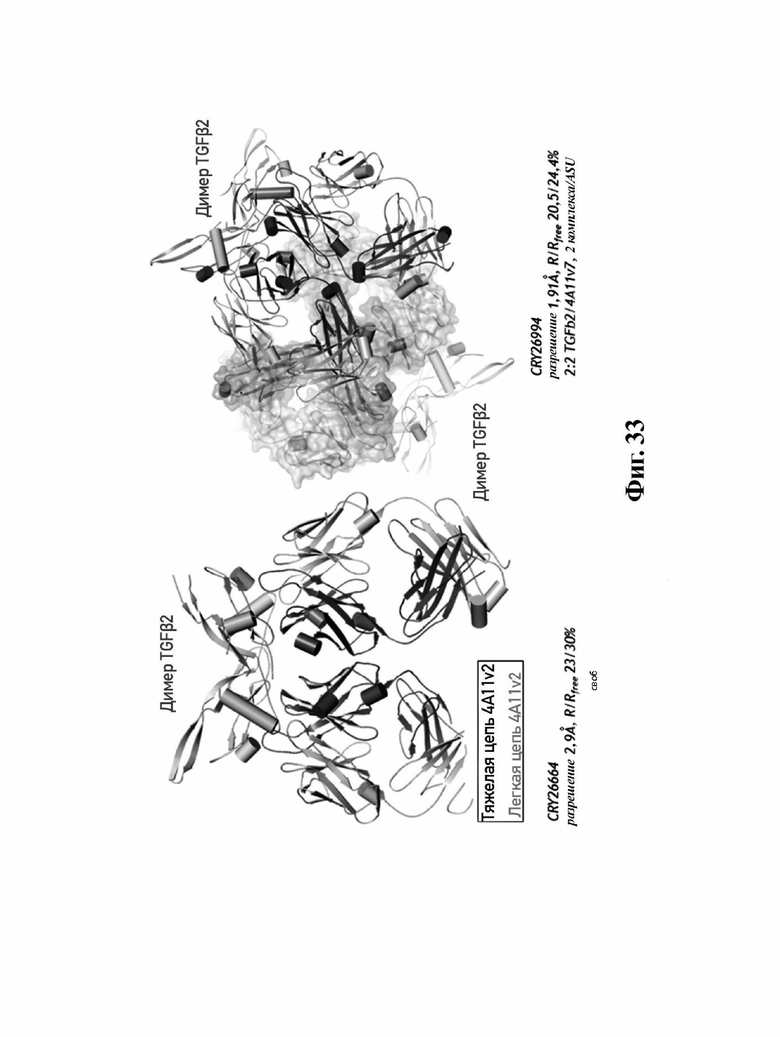
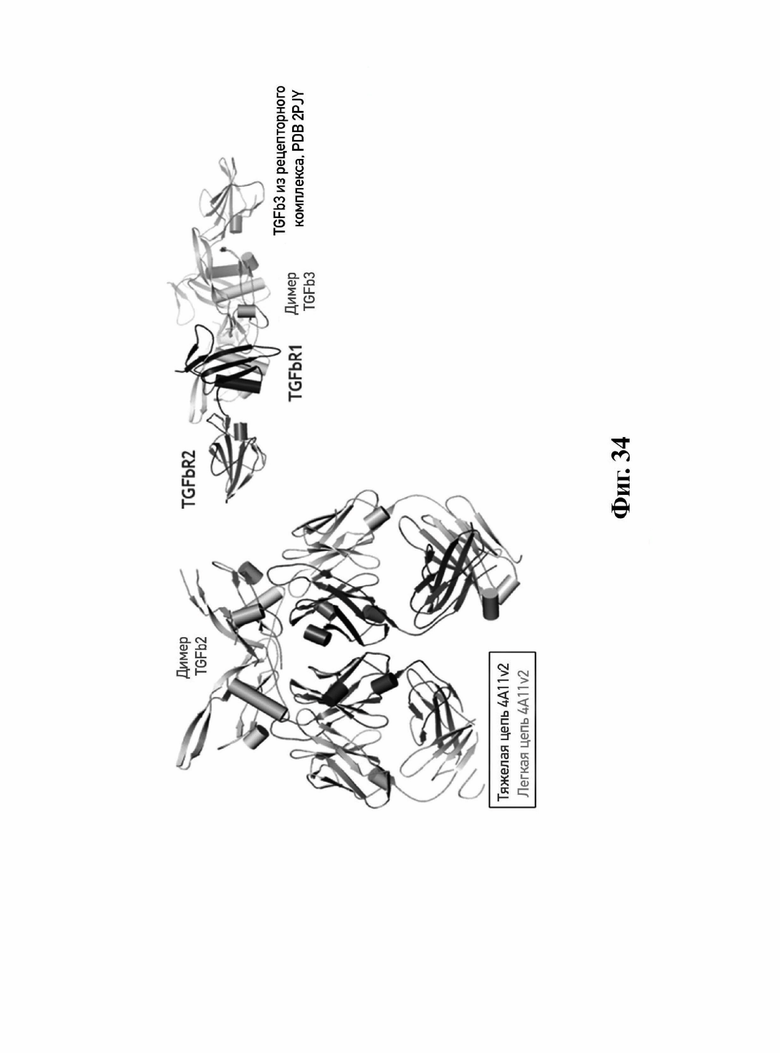
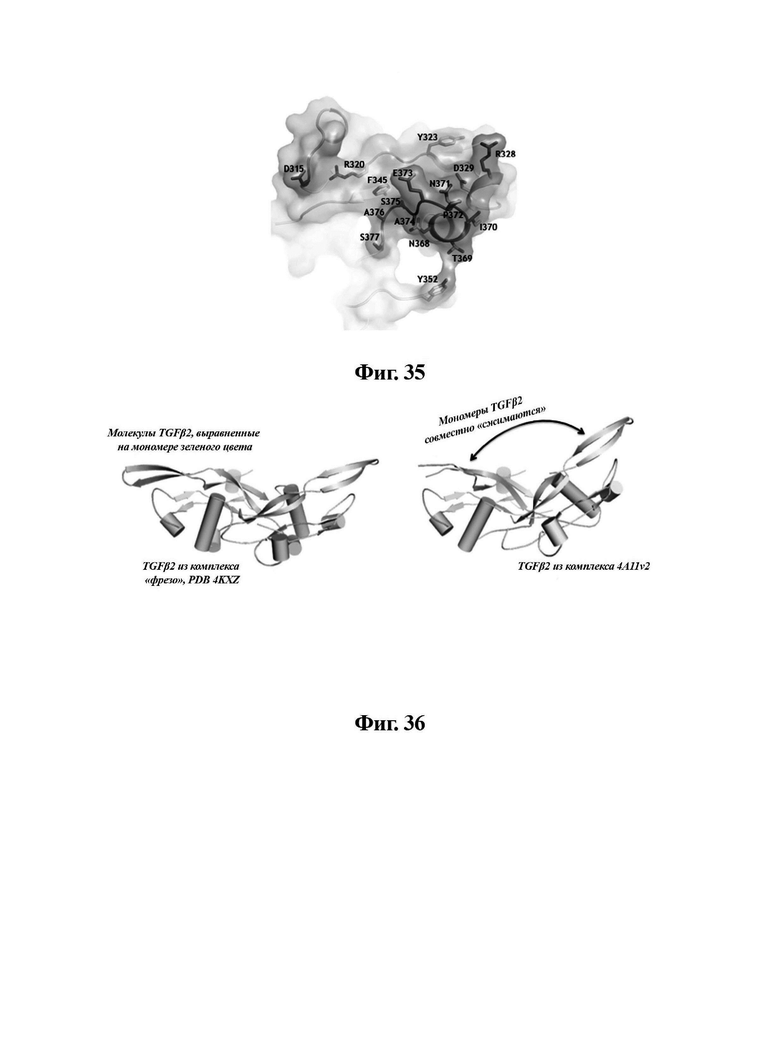
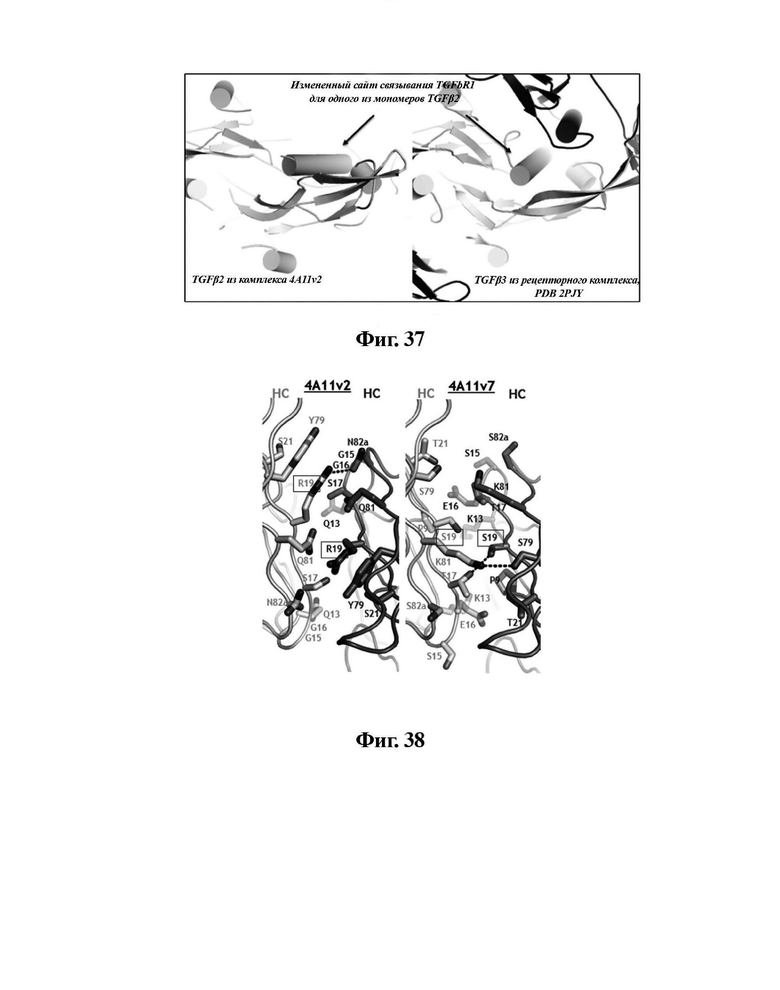
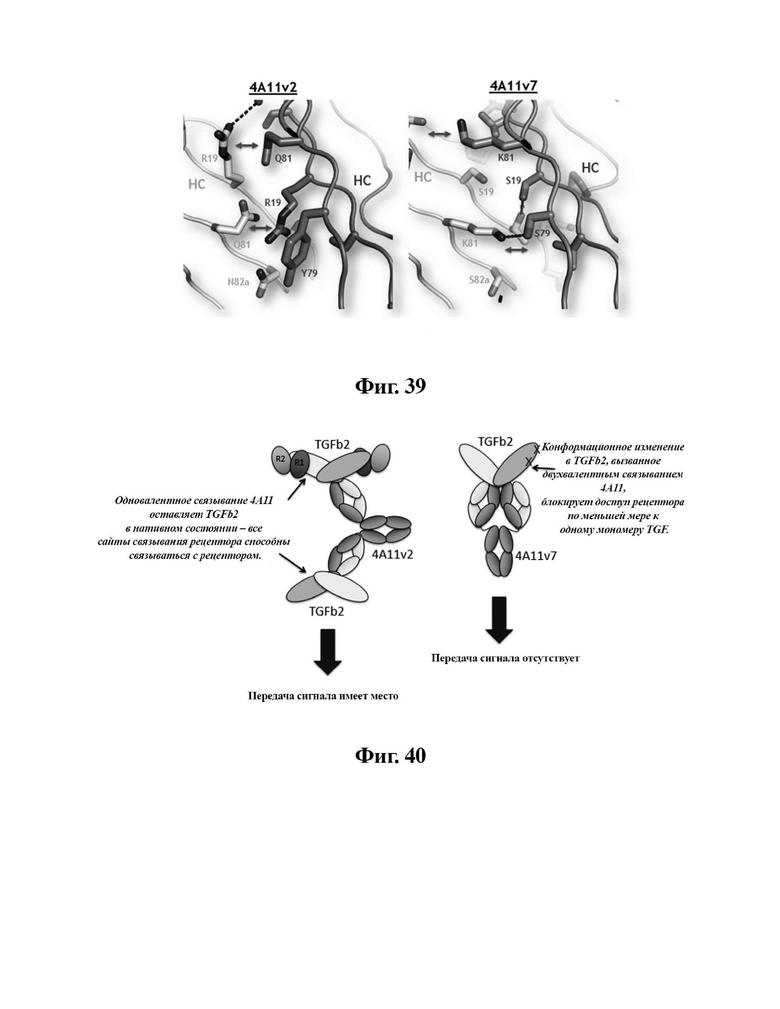
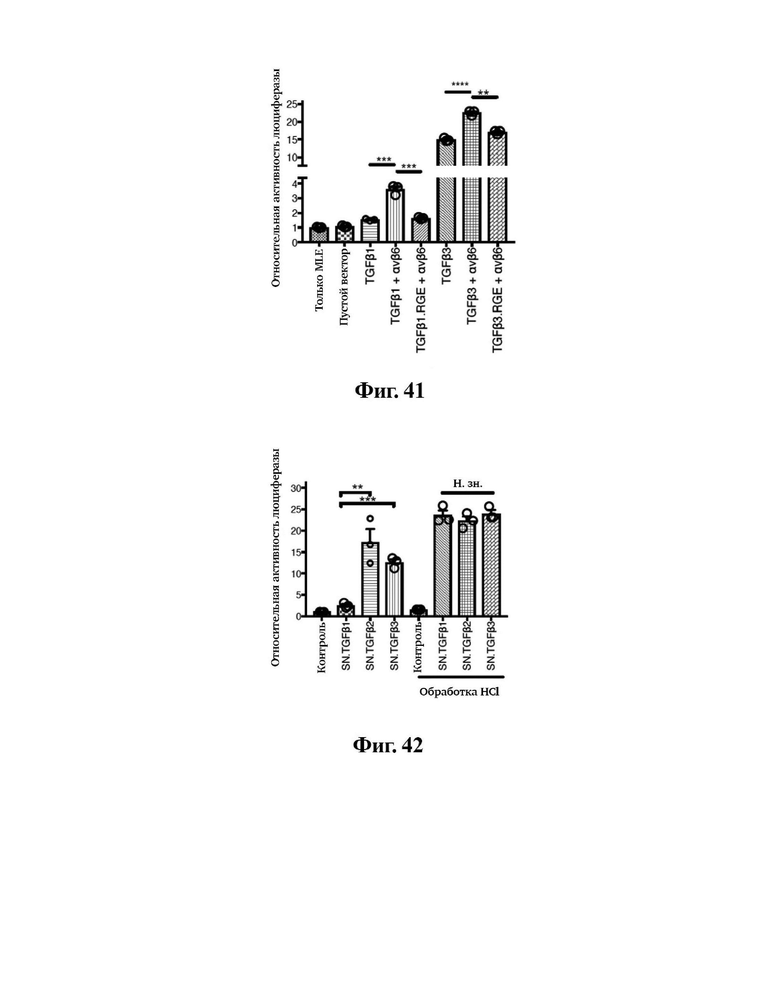
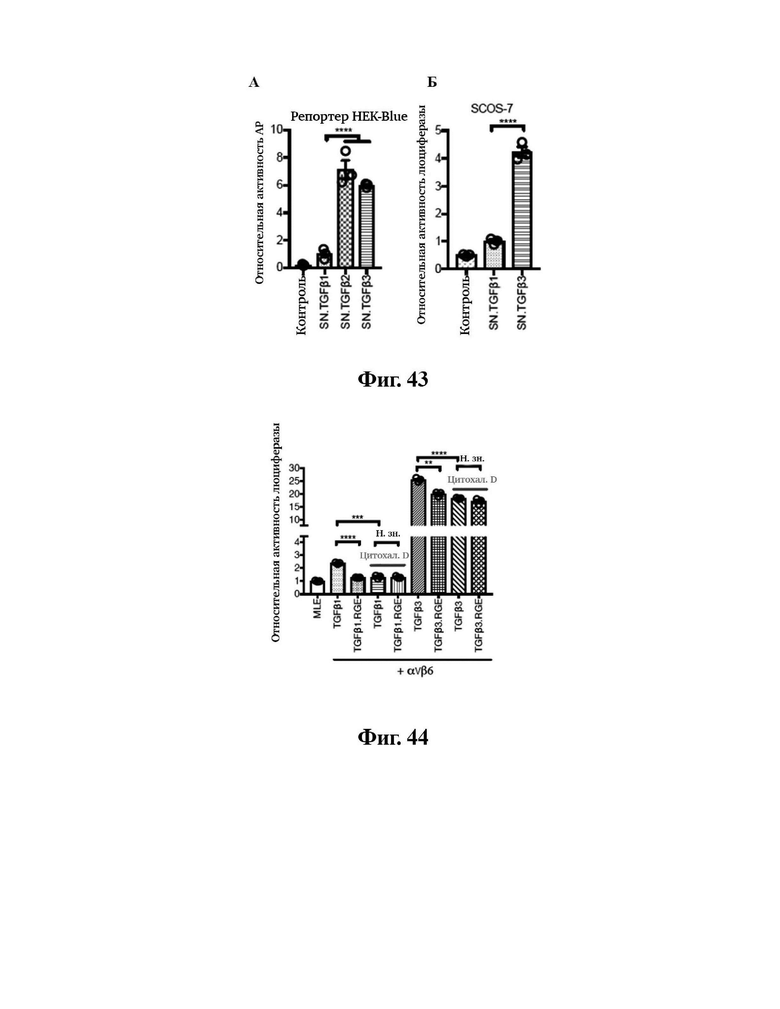
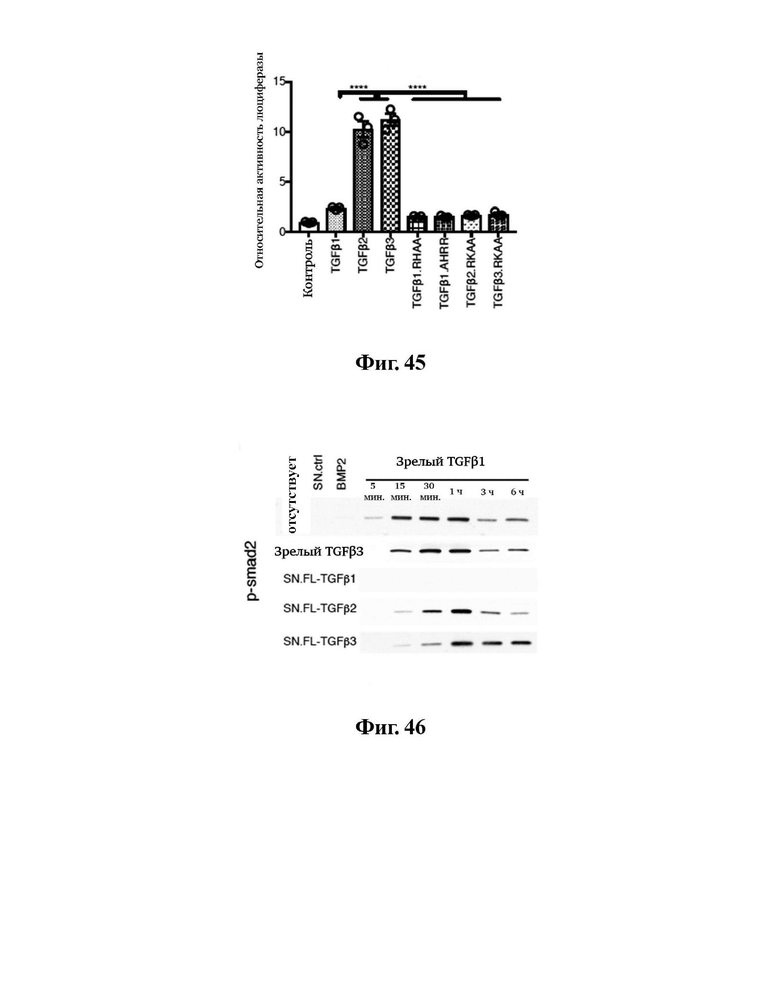
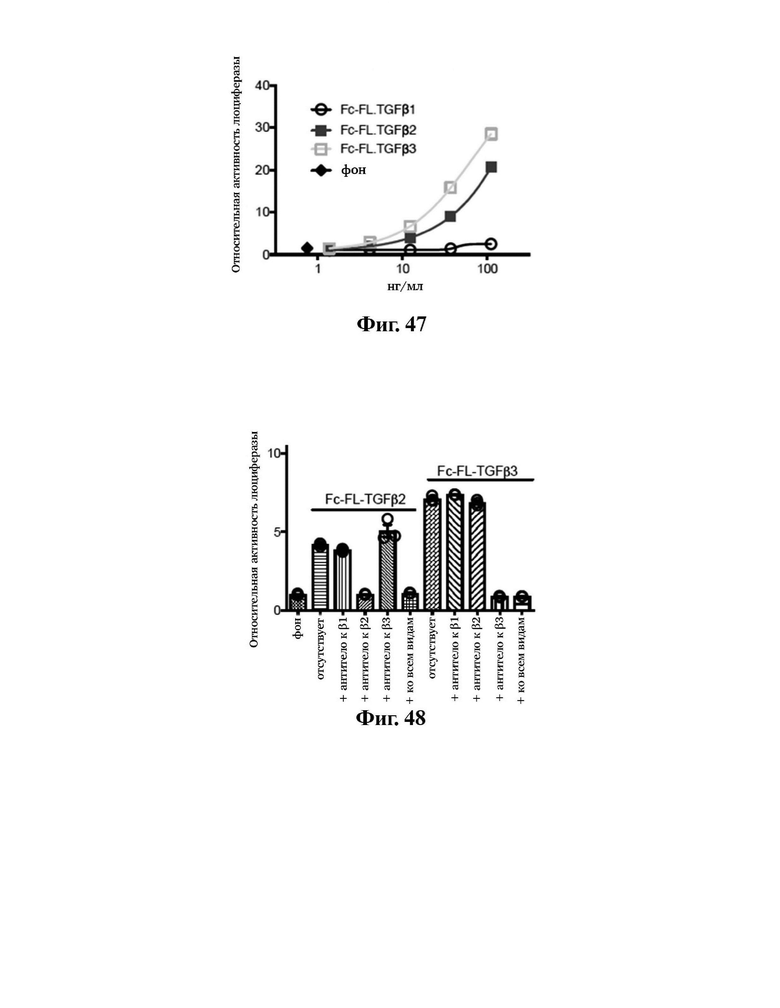
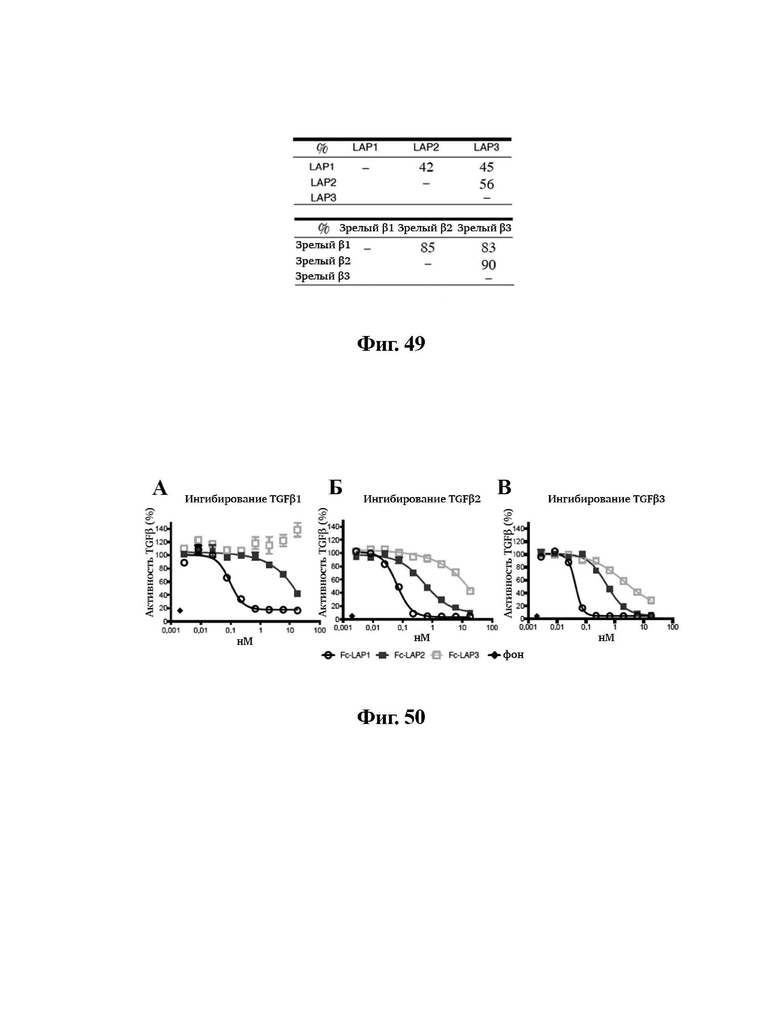
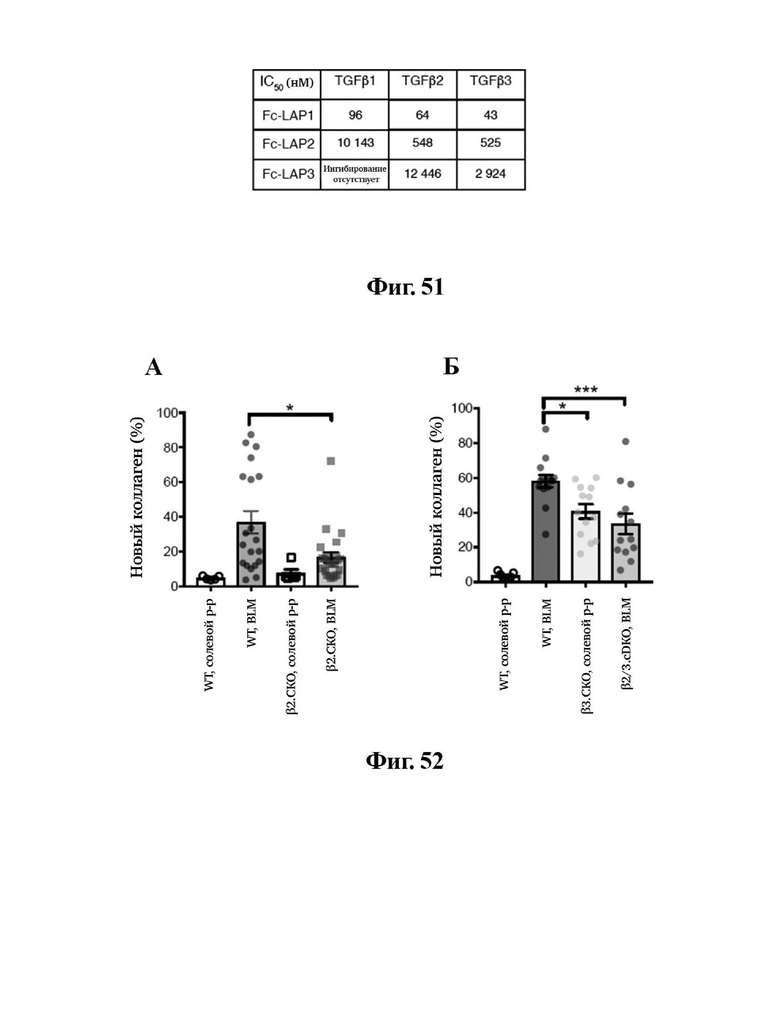
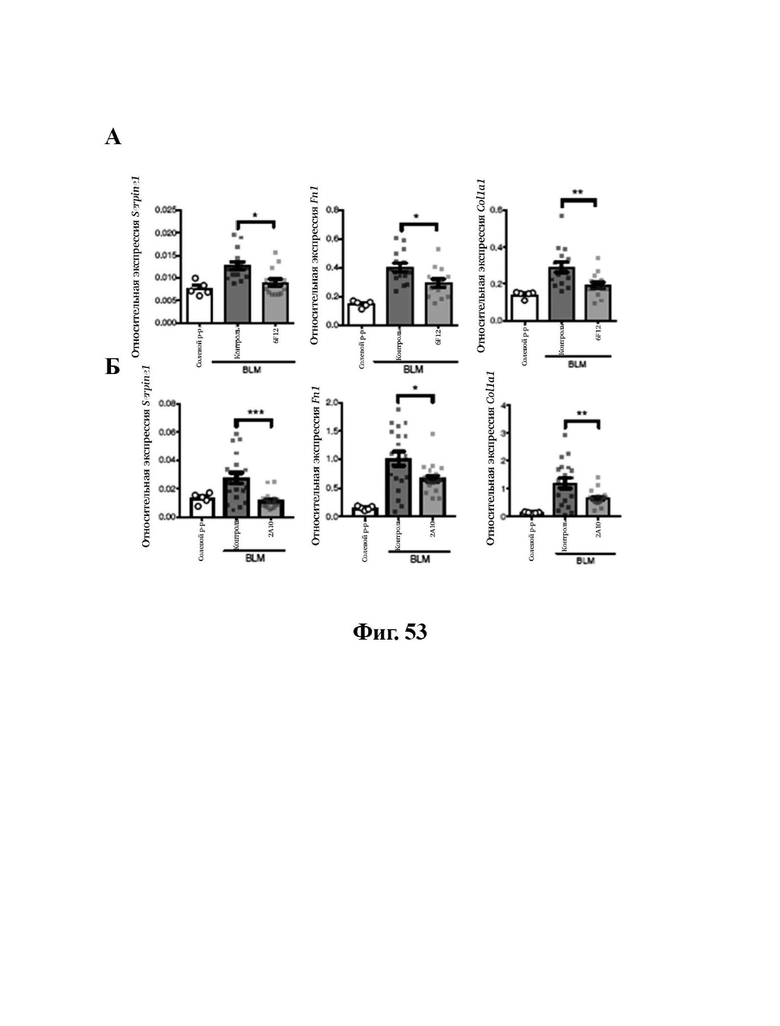
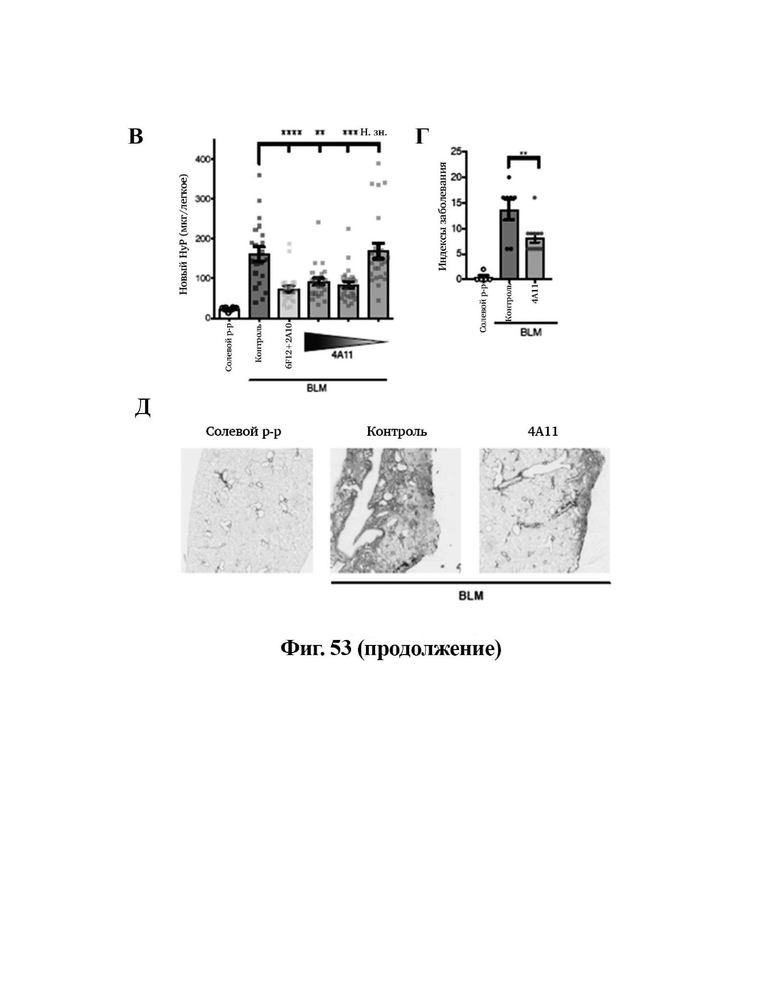
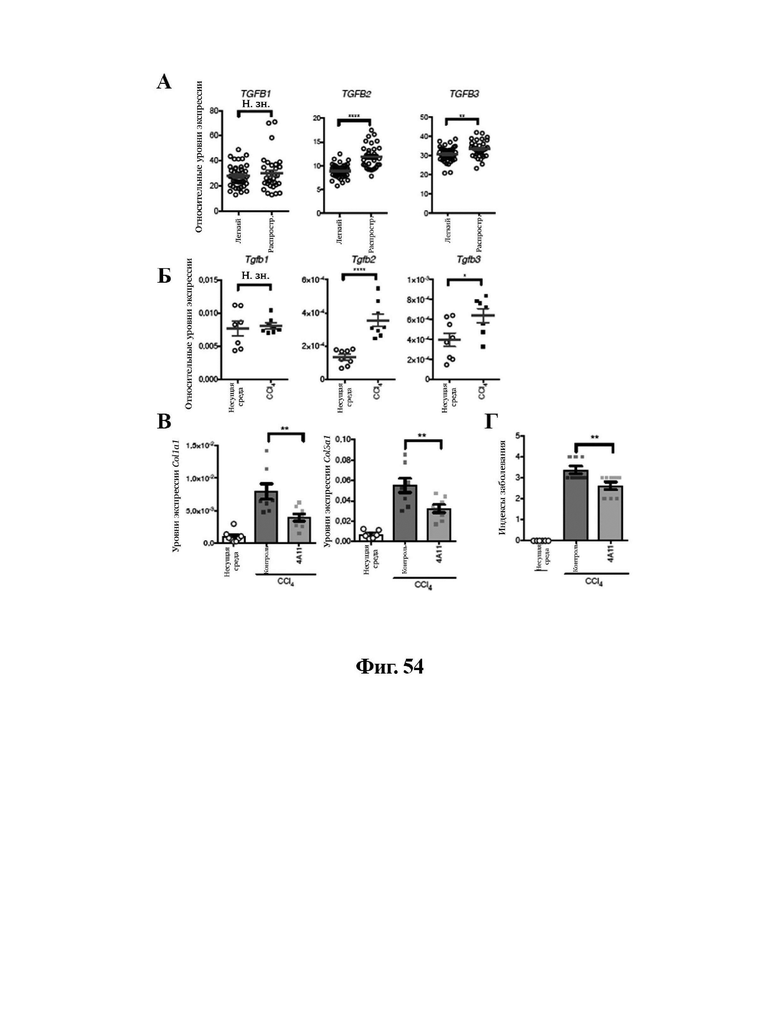
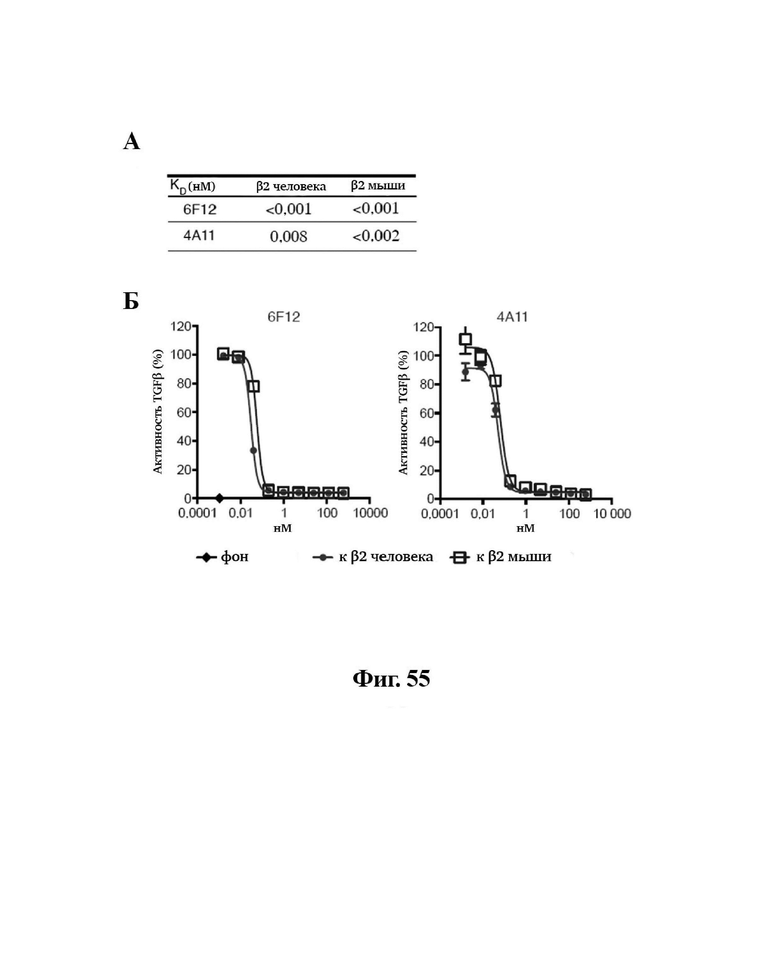
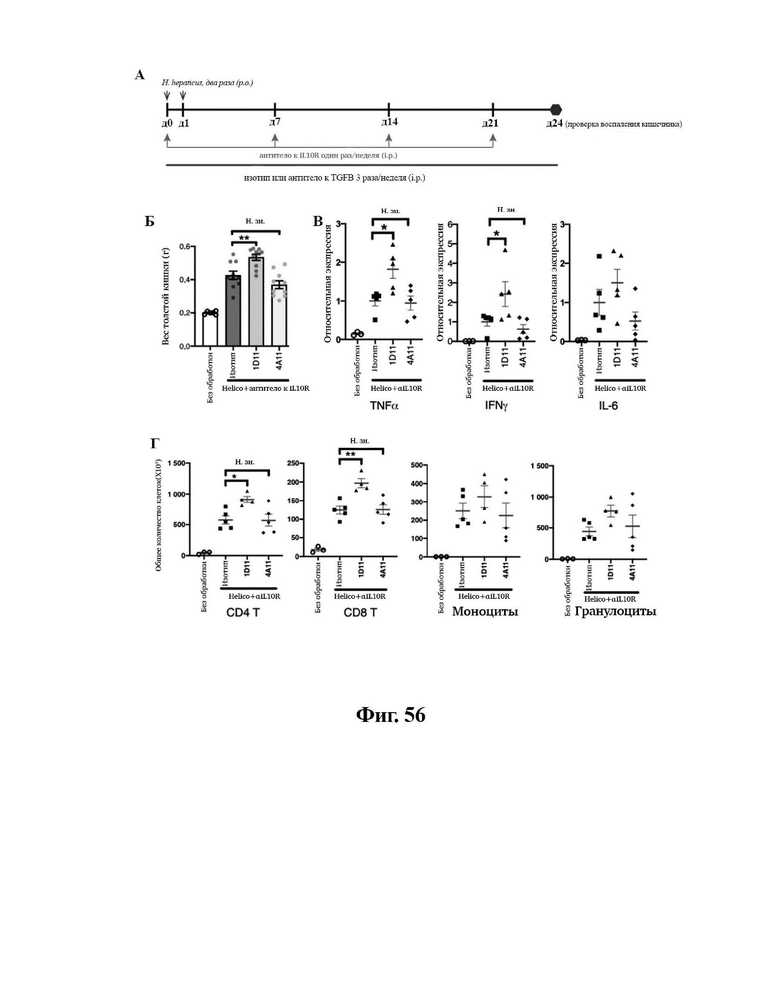
Изобретение относится к области биотехнологии. Представлены антитела к TGFβ3 и содержащие их фармацевтические составы. Изобретение может найти дальнейшее применение в терапии фиброза и других связанных с TGFβ нарушений. 8 н. и 19 з.п. ф-лы, 65 ил., 30 табл., 18 пр.
1. Выделенное антитело к фактору некроза опухоли бета 3 (TGFβ3), где антитело содержит:
(i) CDR тяжелой цепи, содержащие CDR-H1, CDR-H2 и CDR-H3, где CDR-H1 имеет аминокислотную последовательность SEQ ID NO: 4, CDR-H2 имеет аминокислотную последовательность, соответствующую одной из SEQ ID NO: 5, 34, 35 и 159, и CDR-H3 имеет аминокислотную последовательность SEQ ID NO: 6; и
(ii) CDR легкой цепи, содержащие CDR-L1, CDR-L2 и CDR-L3, где CDR-L1 имеет аминокислотную последовательность SEQ ID NO: 7; CDR-L2 имеет аминокислотную последовательность SEQ ID NO: 8 и CDR-L3 имеет аминокислотную последовательность SEQ ID NO: 9.
2. Выделенное антитело к TGFβ3 по п. 1, где антитело содержит аминокислотную последовательность вариабельной области тяжелой цепи VH, обладающую по меньшей мере 95% идентичностью с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 23, 37, 42-53, 55 и 57.
3. Выделенное антитело к TGFβ3 по п. 1, где антитело содержит аминокислотную последовательность вариабельной области легкой цепи VL, обладающую по меньшей мере 95% идентичностью с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 22, 36, 38-41, 54 и 56.
4. Выделенное антитело к TGFβ3 по п. 1, где антитело содержит тяжелую цепь (Н) и легкую цепь (L), при этом тяжелая цепь и легкая цепь содержит аминокислотные последовательности (соответственно), выбранные из группы, состоящей из SEQ ID NO: 29/28 (2A10 крысы), SEQ ID NO: 59/58 (v1), SEQ ID NO: 59/60 (v1.1), SEQ ID NO: 59/61 (v1.2), SEQ ID NO: 59/62 (v1.3), SEQ ID NO: 59/63 (v1.4), SEQ ID NO: 64/58 (v1.5), SEQ ID NO: 65/58 (v1.6), SEQ ID NO: 66/58 (v1.7), SEQ ID NO: 67/58 (v2), SEQ ID NO: 67/60 (v2.1), SEQ ID NO: 67/61 (v2.2), SEQ ID NO: 67/62 (v2.3), SEQ ID NO: 67/63 (v2.4), SEQ ID NO: 68/58 (v2.5), SEQ ID NO: 69/58 (v2.6), SEQ ID NO: 70/58 (v2.7), SEQ ID NO: 71/58 (v2.8), SEQ ID NO: 72/58 (v2.9), SEQ ID NO: 73/58 (h2A10.v2.N54S), SEQ ID NO: 74/58 (h2A10.v2.N54Q), SEQ ID NO: 75/58 (h2A10.v2.T56A), SEQ ID NO: 77/76 (v3) и SEQ ID NO: 79/78 (v4).
5. Фармацевтический состав для лечения нарушений, связанных с TGFβ, содержащий антитело по п. 1 и фармацевтически приемлемый носитель.
6. Выделенное антитело к TGFβ3, где антитело содержит:
вариабельную область тяжелой цепи, включающую (а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 4, (б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 35, и (в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 6, и вариабельную область легкой цепи, включающую (г) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 7, (д) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:8, и (е) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 9.
7. Выделенное антитело к TGFβ3 по п. 6, где (а) вариабельная область тяжелой цепи (VH) антитела содержит аминокислотную последовательность, по меньшей мере на 95% идентичную аминокислотной последовательности SEQ ID NO: 57, (б) вариабельная область легкой цепи (VL) антитела содержит аминокислотную последовательность, по меньшей мере на 95% идентичную аминокислотной последовательности SEQ ID NO: 56; или где (в) вариабельная область тяжелой цепи (VH) антитела содержит аминокислотную последовательность, по меньшей мере на 95% идентичную аминокислотной последовательности SEQ ID NO: 79, (г) вариабельная область легкой цепи (VL) антитела содержит аминокислотную последовательность, по меньшей мере на 95% идентичную аминокислотной последовательности SEQ ID NO: 78.
8. Выделенное антитело к TGFβ3 по п. 6, где антитело представляет собой моноклональное антитело.
9. Выделенное антитело к TGFβ3 по п. 6, где антитело относится к изотипу IgG1 или IgG4.
10. Выделенное антитело к TGFβ3 по п. 6, где антитело представляет собой гуманизированное или химерное антитело.
11. Выделенное антитело к TGFβ3 по п. 6, где антитело представляет собой гуманизированное антитело.
12. Выделенное антитело к TGFβ3 по п. 6, где антитело представляет собой полноразмерное антитело.
13. Выделенное антитело к TGFβ3 по п. 6, где антитело представляет собой гуманизированное моноклональное антитело изотипа IgG1.
14. Выделенное антитело к TGFβ3 по п. 6, где антитело представляет собой фрагмент антитела, который связывает TGFβ3.
15. Выделенное антитело к TGFβ3 по п. 6, где антитело представляет собой фрагмент антитела, который связывает TGFβ3, выбранный из группы, состоящей из Fab, Fab', Fab'-SH, Fv, одноцепочечный Fv (scFv), диабоди и фрагментов F(ab')2.
16. Выделенное антитело к TGFβ3 по п. 6, где Fc-область содержит модификацию, которая приводит к удалению сайта гликозилирования в положении аминокислотного остатка N297 (EU-нумерация согласно Kabat), и где модификация является мутацией N297G.
17. Фармацевтический состав для лечения нарушений, связанных с TGFβ, содержащий антитело по п. 6 и фармацевтически приемлемый носитель.
18. Выделенное антитело к TGFβ3, где антитело содержит (а) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность SEQ ID NO: 57 и (б) вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность SEQ ID NO: 56.
19. Выделенное антитело к TGFβ3 по п. 18, где антитело представляет собой моноклональное антитело.
20. Выделенное антитело к TGFβ3 по п. 18, где антитело относится к изотипу IgG1.
21. Выделенное антитело к TGFβ3 по п. 18, где антитело представляет собой полноразмерное антитело.
22. Выделенное антитело к TGFβ3 по п. 18, где антитело представляет собой гуманизированное моноклональное антитело изотипа IgG1.
23. Выделенное антитело к TGFβ3 по п. 18, где антитело представляет собой фрагмент антитела, который связывает TGFβ3, выбранный из группы, состоящей из Fab, Fab', Fab'-SH, Fv, одноцепочечный Fv (scFv), диабоди и фрагментов F(ab')2.
24. Выделенное антитело к TGFβ3 по п. 18, где Fc-область содержит модификацию, которая приводит к удалению сайта гликозилирования в положении аминокислотного остатка N297 (EU-нумерация согласно Kabat), и где модификация является мутацией N297G.
25. Фармацевтический состав для лечения нарушений, связанных с TGFβ, содержащий антитело по п. 18 и фармацевтически приемлемый носитель.
26. Выделенное антитело к TGFβ3, где антитело содержит тяжелую цепь (H), содержащую аминокислотную последовательность SEQ ID NO: 79, и легкую цепь (L), содержащую аминокислотную последовательность SEQ ID NO: 78.
27. Фармацевтический состав для лечения нарушений, связанных с TGFβ, содержащий антитело по п. 26 и фармацевтически приемлемый носитель.
| WO 2016201282 A2, 15.12.2016 | |||
| BIERNACKA A | |||
| et al., TGF-β signaling in fibrosis, Growth Factors, 2011, v.29(5), pp.196-202 | |||
| WO 2012167143 A1, 06.12.2012 | |||
| US 2019092846 A1, 28.03.2019 | |||
| ГУМАНИЗИРОВАННЫЕ АНТИ-ТФР-БЕТА-АНТИТЕЛА | 2005 |
|
RU2386638C2 |

