Изобретение относится к способам получения композиций, содержащих секреторноподобный иммуноглобулин, в частности секреторноподобный IgA и/или секреторноподобный IgM, и композициям, получаемым этими способами.
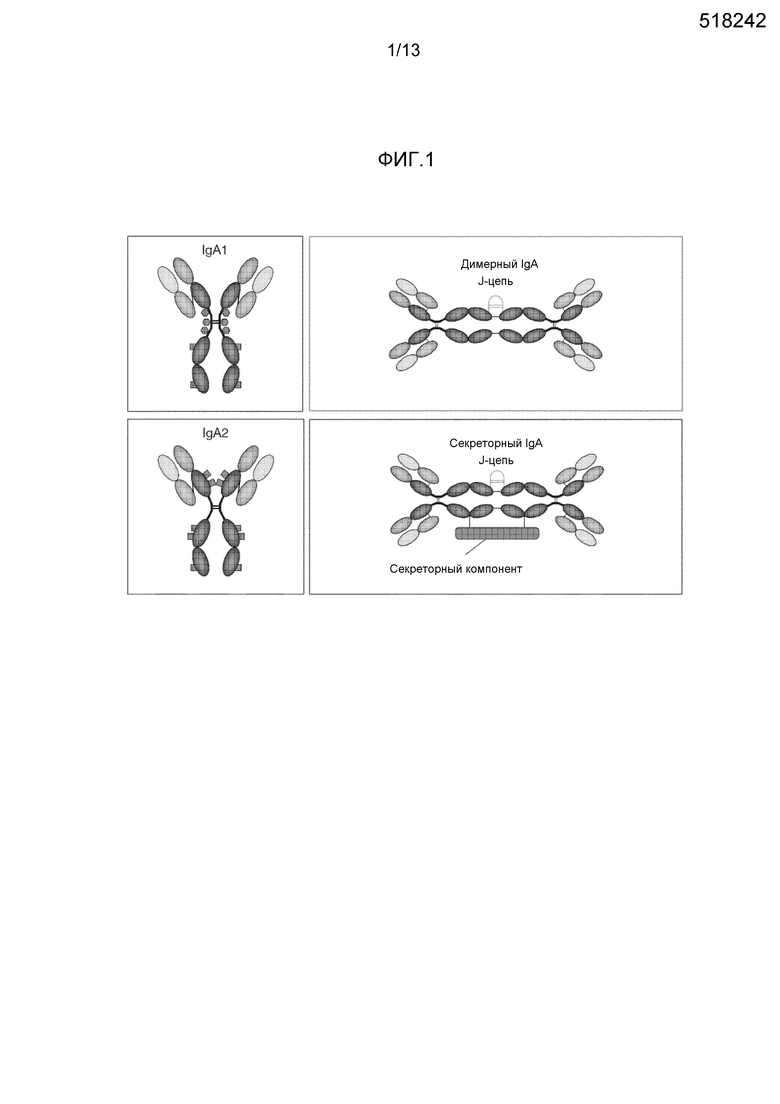
IgA является вторым наиболее широко распространенным классом Ig после IgG в плазме человека, где он находится в концентрации 0,88-4,10 г/л. Существуют два подкласса IgA, IgA1 и IgA2 (фиг. 1) (Zuercher AW et al. Plasma derived immunoglobulins. Principles of Immunopharmacology. 3rd ed. Birkhauser, 2011: p. 271-301), которые отличаются своими дисульфидными связями, соединяющими тяжелые и легкие цепи, а также своим антигенным разнообразием из-за значимых отличий в шарнирной области молекулы. Гликозилированный паттерн двух подклассов является различным: тяжелая цепь обоих подклассов является N-гликозилированной; в отличие от этого O-гликозилирование обнаруживают в IgA1, но не в IgA2, из-за укороченной шарнирной области IgA2.
IgA человека обнаруживают в двух основных формах: или циркулирующим в крови/плазме, или секретированным на поверхностях слизистых оболочек. В плазме IgA преимущественно присутствует в качестве мономеров (80-90%) и его производят плазматические клетки костного мозга; главным подклассом в плазме является IgA1.
Термин полимерный IgA (pIgA) описывает димерный, иногда тетрамерный IgA, ковалентно связанный в своих "концевых частях" посредством соединяющей цепи (J) (фиг. 1).
Молекулы IgM составляют 10% общего содержания Ig в сыворотке. Они ограничены преимущественно внутрисосудистой совокупностью и являются частью первичного, антиген-специфичного, гуморального иммунного ответа; филогенетически и онтогенетически они являются самыми ранними молекулами антител (Ab). IgM существует преимущественно в качестве пентамера, объединенного J-цепью и организованного в планарную структуру; иногда IgM можно также обнаружить в гексамерной форме, в которой отсутствует J-цепь (Zuercher AW et al. Plasma derived immunoglobulins. Principles of Immunopharmacology. 3rd ed. Birkhauser, 2011: p. 271-301).
Поверхности слизистых оболочек пищеварительных, дыхательных и мочеполовых путей, а также протоки экзокринных желез выстланы слоями эпителиальных клеток, которые образуют прочный барьер, отделяющий внутренние отделы тела от внешней среды. У людей эти обширные поверхности покрывают 400 м2, площадь, которая постоянно подвергается экзогенным патогенам (Corthesy, B. (2010) Future Microbiol. 5:817-829). Комбинация внутренних и индуцибельных клеточных и молекулярных механизмов обеспечивает защиту против колонизации и входа/инвазии микробов. У здоровых индивидуумов секреторный IgA (SIgA) является наиболее распространенным антителом (Ab), выполняющим функцию иммунной эксклюзии на люминальной стороне слизистых оболочек (Macpherson, A.J. et al. (2008) Mucosal Immunol. 1:11-22), тогда как секреторные IgM Аb принимают эту функцию на себя у пациентов с недостаточностью IgA. SIgA синтезируется плазматическими клетками собственной пластинки слизистой оболочки кишечника, верхних дыхательных путей или мочеполовых путей. SIgA состоит из димера pIgA и высокогликозилированного секреторного компонента (SC) массой приблизительно 75 кДа (фиг. 1); сходным образом пентамерный IgM, содержащий J цепь, связанный с SC, составляет SIgM. SC представляет внеклеточную часть полимерного рецептора Ig (pIgR). pIgR необходим для трансэпителиального транспорта pIgA или пентамерного IgM от места продуцирования к поверхности слизистой оболочки, где комплекс pIgR-pIgA/pIgR-IgM превращается в SIgA/SIgM посредством ферментативного расщепления (Zuercher AW et al. Plasma derived immunoglobulins. Principles of Immunopharmacology. 3rd ed. Birkhauser, 2011: p. 1-31). Связь с SC защищает IgA или IgM от протеолитического распада. SIgA является преобладающим Ig в серозно-слизистых секретах, таких как слюна, трахео-бронхеальные секреты, молозиво, молоко, слезная жидкость, кишечные секреты и мочеполовые секреты. Это самый значимый Ig, продуцируемый в слизистых оболочках (и таким образом в организме человека); приблизительно 3-5 г SIgA секретируется ежедневно в просвет кишечника. Таким образом SIgA является существенным для иммунной эксклюзии и для поддержания целостности эпителия. SIgM присутствует на более низких уровнях, но выполняет те же функции иммунной эксклюзии, что и SIgA.
Для некоторых патогенов, таких как полиовирус, сальмонелла, или грипп, защиту против инфекции слизистой оболочки можно индуцировать посредством активной иммунизации слизистой оболочки разрешенными к применению вакцинами. Однако для большинства патогенов слизистых оболочек не доступны никакие активные вакцины для слизистых оболочек. Альтернативно, защитные уровни Аb могли бы быть доставлены прямо к поверхностям слизистых оболочек посредством пассивной иммунизации. В природе это происходит физиологически у многих видов млекопитающих посредством переноса материнских антител к их потомству через молоко (Brandtzaeg, P. (2003) Vaccine 21:3382-3388). В исследованиях на человеке и животных посредством пассивной иммунизации слизистой оболочки было продемонстрировано, что молекулы антител pIgA и SIgA, введенные посредством пероральной, интраназальной, внутриматочной или легочной инстилляции, могут предотвратить, ослабить или устранить бактериальные и вирусные инфекции (Corthesy, B. (2003) Curr. Pharm. Biotechnol. 4:51-67). Однако секреторную форму IgA, в естественных условиях обнаруживаемую на поверхностях слизистых оболочек, использовали редко, и крупномасштабное получение SIgA на сегодняшний день невозможно. Конструирование SIgA с применением биотехнологических способов является сложной проблемой, но такие молекулы могли бы иметь важные клинические применения (Corthesy, B. (2002) Trends Biotechnol. 20:65-71). То же самое относится к IgM, содержащему секреторный компонент.
Сущность изобретения
Авторы изобретения с удивлением обнаружили, что возможно комбинировать иммуноглобулин, содержащий J-цепь, и полученный из плазмы, в частности, IgA и/или IgM, с секреторным компонентом без необходимости в первую очередь очищать иммуноглобулин, содержащий J-цепь. Это изобретение открывает путь к широкомасштабному получению секреторноподобного IgA и/или IgM, которые можно использовать в медицине, например, для предотвращения и лечения инфекций на поверхности слизистых оболочек у субъектов, в частности у субъектов-людей.
Одним из аспектов изобретения является способ получения композиции, содержащей секреторноподобный иммуноглобулин, в частности IgA и/или IgM, in vitro, включающий стадии
(a) получения выделенной из крови белковой композиции, содержащей иммуноглобулин, имеющий в составе J-цепь, в частности IgA и/или IgM, в неочищенном виде; и
(b) смешивания композиции стадии (a) с секреторным компонентом.
Предпочтительно, секреторноподобный иммуноглобулин является секреторноподобным IgA. Предпочтительно, композиция на этапе (a) содержит по меньшей мере 5% IgA, содержащего J-цепь, более предпочтительно по меньшей мере 10%, более предпочтительно по меньшей мере 20%, даже более предпочтительно по меньшей мере 30%, наиболее предпочтительно она содержит по меньшей мере 50% IgA, содержащего J-цепь. Предпочтительно, композицию на этапе (a) получают из крови человека, например плазмы человека или ее фракций, обогащенных IgA или даже IgA, содержащим J-цепь, но никакой очистки IgA, содержащего J-цепь, не требуется.
Предпочтительно, секреторноподобный иммуноглобулин является секреторноподобным IgM. Предпочтительно, композиция на этапе (a) содержит по меньшей мере 5% IgM, содержащего J-цепь, более предпочтительно по меньшей мере 10%, более предпочтительно по меньшей мере 20%, даже более предпочтительно по меньшей мере 30%, наиболее предпочтительно она содержит по меньшей мере 50% IgM, содержащего J-цепь. Предпочтительно, композицию на этапе (a) получают из крови человека, например из плазмы человека или ее фракций, обогащенных IgM, или даже IgM, содержащим J-цепь, но никакой очистки IgM, содержащего J-цепь, не требуется.
Секреторный компонент, применяемый на этапе (b), является предпочтительно рекомбинантным секреторным компонентом, более предпочтительно рекомбинантным секреторным компонентом человека, предпочтительно полученным посредством клеточной линии млекопитающих. Однако секреторный компонент из природного источника также можно использовать, такой как секреторный компонент, выделенный из молока, слюны, слизи или сходных источников.
Молярное отношение между секреторным компонентом и J-цепью внутри димеров/полимеров IgA или пентамеров IgM в композиции на этапе (a) находится в диапазоне между 1:10 и 10:1, предпочтительно между 1:5 и 5:1, более предпочтительно между 1:2 и 2:1.
В другом аспекте изобретения молярное отношение между секреторным компонентом и J-цепью внутри композиции на этапе a) находится в диапазоне между 1:10 и 10:1, предпочтительно между 1:5 и 5:1, более предпочтительно между 1:2 и 2:1.
Другом аспектом изобретения является композиция, содержащая секреторноподобный IgA и/или секреторноподобный IgM или их сочетания, получаемые посредством способа по изобретению. Композиция может дополнительно содержать фармацевтически приемлемый носитель или эксципиент.
Дополнительным аспектом изобретения является композиция, описанная выше для медицинского использования.
Подробное описание изобретения
Как уже упоминалось выше, авторы изобретения к удивлению обнаружили, что является возможным сочетать полученные из плазмы иммуноглобулины, содержащие J-цепь, в частности IgA, содержащий J-цепь, и/или IgM, содержащий J-цепь, с секреторным компонентом без необходимости сначала очищать иммуноглобулин, содержащий J-цепь. Это изобретение открывает путь к широкомасштабному получению секреторноподобного IgA и/или секреторноподобного IgM, которые можно использовать в медицине, например, для предотвращения и лечения инфекций на поверхностях слизистых оболочек у субъекта, в частности у субъектов-людей.
Одним из аспектов изобретения является способ получения композиции, содержащей секреторноподобный иммуноглобулин, в частности секреторноподобный IgA или секреторноподобный IgM, in vitro, включающий стадии
(a) получения из крови белковой композиции, содержащей IgA, имеющий в составе J-цепь, или IgM, имеющий в составе J-цепь, в неочищенном виде,
(b) смешивания или сочетание композиции стадии (a) с секреторным компонентом.
Предпочтительно, композиция на этапе (a) содержит по меньшей мере 5% (масс./масс.) IgA, содержащего J-цепь, более предпочтительно по меньшей мере 10%, более предпочтительно по меньшей мере 20%, даже более предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 50%, наиболее предпочтительно она содержит по меньшей мере 70% IgA, содержащего J-цепь. Предпочтительно, композиция на этапе (a) получена из крови человека, например из плазмы человека или ее фракций, обогащенных IgA, или даже IgA, содержащего J-цепь, но никаких специфических этапов очистки димеров или полимеров IgA, содержащего J-цепь, не требуется. Таким образом, IgA, содержащий J-цепь, также находится в композиции, содержащей другие белки, такие как мономерный IgA, IgM или IgG. Например, композиция может содержать более 10% мономерного IgA, и/или более чем 10% IgG, и/или более чем 10% IgM. В то время как обогащение димерным IgA, содержащим J-цепь, может происходить как часть обработки фракций плазмы человека, для очистки белков плазмы, таких как IgG, альбумин, альфа-1 антитрипсин и факторы свертывания, никакой стадии очистки, специфически спланированной для отделения димерного IgA, содержащего J-цепь, или полимеров из других белков, таких как аффинная хроматография или эксклюзия на основе размера и отбор фракции с релевантной молекулярной массой, для получения композиции на этапе (a), не требуется.
В другом предпочтительном аспекте по изобретению, композиция на этапе (a) содержит по меньшей мере 5% (масс./масс.) IgM, содержащего J-цепь, более предпочтительно по меньшей мере 10%, более предпочтительно по меньшей мере 20%, даже более предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 50%, наиболее предпочтительно она содержит по меньшей мере 70% IgM, содержащего J-цепь. Предпочтительно, композиция на этапе (a) получена из крови человека, например плазмы человека или ее фракций, обогащенных IgM или даже IgM, содержащим J-цепь, но никаких специфических этапов очистки IgM, содержащего J-цепь, не требуется. Таким образом, IgM, содержащий J-цепь, будет находиться в композиции, также содержащей другие белки, такие как IgA или IgG. Например, композиция может содержать более чем 10% IgA, и/или более чем 10% IgG. В то время как обогащение IgM, содержащим J-цепь, может происходить как часть обработки фракций плазмы человека, для очистки белков плазмы, таких как IgG, альбумин, альфа-1 антитрипсин и факторы свертывания, никакой стадии очистки, специфически спланированной для отделения IgM, содержащего J-цепь, от других белков, таких как аффинная хроматография или эксклюзия на основе размера и отбор фракции с релевантной молекулярной массой, для получения композиции на этапе (a), не требуется.
Как применяют в настоящем документе, термин "секреторный компонент" относится к белку, который специфически связывается с иммуноглобулином, содержащим J-цепь, и относится к или получается из идентичного внеклеточной части рецептора полимерного иммуноглобулина (pIgR), предпочтительно pIgR млекопитающих, более предпочтительно pIGR приматов, наиболее предпочтительно pIgR человека. Предпочтительно, секреторный компонент придает увеличенную стабильность иммуноглобулину, содержащему J-цепь. Секреторный компонент, содержащийся в композиции, может быть рекомбинантным секреторным компонентом, предпочтительно секреторным компонентом, полученным в клеточной линии млекопитающих. Секреторный компонент в его традиционном узком значении (упоминаемый как "естественный секреторный компонент" в настоящем документе) является внеклеточной частью рецептора полимерного иммуноглобулина (pIgR), который, как правило, соединяется во время секреции с димерным или полимерным IgA или пентамерным IgM, содержащим J-цепь. IgA/IgM, содержащий J-цепь, связывается с рецептором полимерного иммуноглобулина на базолатеральной поверхности эпителиальных клеток и захватывается в клетку посредством трансцитоза. Этот рецепторный комплекс затем переходит через клеточные компартменты, перед тем как он транспортируется на люминальную поверхность эпителиальных клеток. Трансцитозный комплекс IgA/IgM-pIgR затем высвобождается посредством протеолиза, и часть рецептора полимерного иммуноглобулина (pIgR), обозначаемая как естественный секреторный компонент, остается связанной с IgA/IgM, содержащим J-цепь, высвобождая секреторный IgA/IgM. Однако существует свидетельство того, что также может иметь место обратный трансцитоз IgA, т.е. от люминальной поверхности к базолатеральной поверхности.
pIgR человека клонируют и секвенируют, его последовательность доступна в качестве SwissProt entry P01833, и представлена в Seq ID NO: 1. pIgR человека является гликопротеином с 764 аминокислотными остатками, содержащими сигнальный пептид (остатки от 1 до 18), внеклеточную часть (остатки от 19 до 638), трансмембранную область (остатки от 639 до 661) и цитоплазматическую область (остатки от 662 до 764). Предполагают, что остатки от 19 до 603 связаны с IgA, содержащим J-цепь, как описано выше, и эту часть этого гликопротеина, как правило, обозначают как секреторный компонент (в настоящем документе упоминается как "естественный секреторный компонент").
Секреторный компонент, применяемый в композиции по изобретению, может содержать любую внеклеточную последовательность pIgR, которая способна соединяться с IgA, содержащим J-цепь. Например, секреторный компонент может содержать внеклеточные домены pIgR из источников млекопитающих, например из приматов, крупного рогатого скота, лошадей, кошек, собак, кроликов, морских свинок, крыс или мышей, или их вариантов. Функциональные гибриды внеклеточных доменов из нескольких видов млекопитающих или их вариантов тоже рассматриваются для применения в изобретении, например, полученные посредством слияния иммуноглобулиноподобных доменов из различных видов в секреторный компонентоподобный белок. Функциональный секреторный компонент может также быть составлен посредством слияния отбора в норме присутствующих иммуноглобулиноподобных доменов, например, секреторный компонент кролика является функционально составленным только из доменов 1, 4 и 5. Предпочтительно, однако, применяют секреторный компонент человека или его функциональные варианты.
Таким образом, секреторный компонент, применяемый в композиции по изобретению, предпочтительно содержит остатки от 19 до 603 в SEQ ID NO: 1 или их функциональные варианты. Функциональные варианты могут включать делеции, вставки и/или замены, предпочтительно замены являются консервативными заменами, например, основный аминокислотный остаток заменен на другую основную аминокислоту, гидрофобная аминокислота заменена на другую гидрофобную аминокислоту и т.д. Вариантный секреторный компонент по меньшей мере на 50% идентичен в последовательности к остаткам от 19 до 603 SEQ ID NO: 1, предпочтительно по меньшей мере на 55%, 60%, 65%, 70%, 75%, 80%, более предпочтительно по меньшей мере на 85% или даже 90%, даже более предпочтительно по меньшей мере 92%, 94%, 95%, 97%, 98%, или даже на 99% идентичен остаткам от 19 до 603 SEQ ID NO: 1. Предпочтительно, секреторный компонент содержит внеклеточную часть pIgR, более предпочтительно внеклеточную часть pIgR человека, наиболее предпочтительно секреторный компонент содержит или даже состоит из остатков от 19 до 603 SEQ ID NO: 1.
Специалист хорошо знает, как получать секреторный компонент посредством рекомбинантных методов. Пример экспрессии секреторного компонента человека в клетках CHO был описан Phalipon et al (Phalipon A et al. (2002) Immunity 17:107-115), но это изобретение не ограничено секреторным компонентом, полученным посредством этой системы. Например, желаемую последовательность кДНК можно получать синтетически или клонировать посредством ПЦР-РВ, с применением РНК, выделенной из клеток или ткани, экспрессирующей pIgR, в качестве образца. кДНК затем может быть вставлена в экспрессирующий вектор млекопитающих, такой как pcDNA3 - доступно много альтернативных экспрессирующих векторов. Рекомбинантный экспрессирующий вектор затем будет введен в подходящую клеточную линию-хозяина, такую как CHO, Cos, HEK293 или BHK. Другие клеточные линии доступны и также могут быть применены. Способы введения таких векторов в клеточную линию включают липофекцию, электропорацию и другие методы, хорошо известные специалисту. Как правило, клетки, содержащие экспрессирующий вектор и экспрессирующие белок, представляющий интерес, затем отбирают и клонируют. Также можно использовать вирусные экспрессирующие системы, например вирус коровьей оспы можно использовать для экспрессии белков на высоких уровнях в клетках млекопитающих, экспрессирующие системы бакуловируса можно использовать для экспрессии белков в клетках насекомых. Экспрессирующие системы дрожжей или бактерий также могут быть рассмотрены, и такие экспрессирующие системы известны специалисту. Таким же образом, экспрессирующие системы растений также могут быть рассмотрены, и такие системы известны специалисту.
Секреторный компонент или его вариант, применяемый в композиции по изобретению, также может содержать метку, такую как гексагистидиновую метку, которая может помочь в очистке полученного белка. Если такая метка прикреплена посредством расщепляемого линкера, метка может быть расщеплена до использования в изобретении. Сходным образом, секреторный компонент можно получать в качестве белка слияния. Снова расщепляемый линкер можно использовать, чтобы партнер слияния мог быть отщеплен от секреторного компонента до использования в изобретении.
Специалист может затем очистить экспрессированный белок с применением стандартных методов. Рекомбинантный секреторный компонент можно очищать до высокого уровня чистоты с применением подходящего способа, например, эксклюзионной и/или ионообменной хроматографии. Предпочтительно окончательное получение рекомбинантного секреторного компонента будет существенным образом свободным от загрязнений, особенно белков клетки-хозяина. Однако секреторный компонент может также специфичным образом соединяться с иммуноглобулином, содержащим J-цепь, в неочищенном виде, таким образом, очистка перед объединением с иммуноглобулином, содержащим J-цепь, не существенна.
Секреторный компонент может также быть получен из природного источника, предпочтительно из молока, слюны или слизи. Предпочтительно секреторный компонент происходит из человеческого источника, но секреторный компонент других видов может также быть применен в изобретении.
Молярное отношение между секреторным компонентом и J-цепью в димерах/полимерах IgA или пентамерах IgM в композиции стадии (a) находится в диапазоне между 1:10 и 10:1, предпочтительно между 1:5 и 5:1, более предпочтительно между 1:2 и 2:1.
Молярное отношение между секреторным компонентом и J-цепью в композиции на стадии (a) находится между 1:10 и 10:1, предпочтительно между 1:5 и 5:1, более предпочтительно между 1:2 и 2:1.
Количество секреторного компонента, применяемого на стадии (b), может составлять по меньшей мере 1 часть (по массе) секреторного компонента к 50 частям (по массе) от белка в композиции стадии (a), предпочтительно по меньшей мере 1 часть к 40, 30, 20, 15, 10, наиболее предпочтительно по меньшей мере 1 часть секреторного компонента к 5 частям белка в композиции стадии (a).
Другим аспектом изобретения является композиция, содержащая секреторноподобный IgA, получаемый способом по изобретению. Еще другим аспектом изобретения является композиция, содержащая секреторноподобный IgM, получаемый способом по изобретению. Еще дополнительным аспектом является композиция, содержащая секреторноподобный IgA и секреторноподобный IgM, получаемый способом по изобретению, например, в молярном отношении между 10:1 и 1:10, предпочтительно между 5:1 и 1:5, более предпочтительно между 2:1 и 1:2. В другом аспекте изобретения комбинированное содержание IgA и IgM в композиции превышает 50%, предпочтительно 60%, более предпочтительно 70%, даже более предпочтительно 80%, даже более предпочтительно 90%, наиболее предпочтительно оно составляет 100%.
Дополнительным аспектом изобретения является композиция, описанная выше, для медицинского применения. Например, композиции по изобретению можно использовать преимущественно для лечения некротического энтероколита и, как правило, инфекций на поверхности слизистых оболочек.
Композиция может дополнительно содержать один или несколько фармацевтически приемлемых носителей или эксципиентов, и/или стабилизаторов. Композицию можно формулировать в жидкой форме, в виде сиропа, лосьона, мази, порошка, который может быть разведен жидкостью перед введением, капсулы, пилюли, геля, крема, желе, состава с контролируемым высвобождением или любого другого состава, подходящего для предполагаемого медицинского применения. Например, для лечения заболеваний ЖКТ композицию можно формулировать с защитным покрытием, которое растворяется в желаемой области ЖКТ для высвобождения композиции. Композицию можно принимать перорально, вводить местно, энтерально, посредством ингаляции или любым другим подходящим способом для назначаемого применения. Для перорального введения можно совместно вводить ингибиторы протонной помпы.
Белки в композиции могут быть концентрированы, например, с использованием диа/ультрафильтрации или других стандартных способов, предшествующих созданию состава. Кроме того, композиция может быть лиофилизирована, а затем растворена в подходящем растворе перед применением.
Определения
Термин "секреторноподобный IgA" или "секреторноподобный IgA плазмы" предназначен охватывать IgA(плазма), содержащий J-цепь, комбинированный с белком, который является секреторным компонентом или его функциональным вариантом, который служит для предоставления некоторой защиты от протеолитического расщепления. Как правило, IgA, содержащий J-цепь, содержит два или четыре, или даже больше мономеров IgA. Как правило, IgA, содержащий J-цепь, смешивают с секреторным компонентом или его вариантом in vitro, т.е. связь между секреторным компонентом и IgA, содержащим J-цепь, имеет место in vitro скорее, чем во время трансцитоза.
Термин "секреторноподобный IgM" предназначен охватывать IgM (плазма), содержащий J-цепь, комбинированный с секреторным компонентом или его функциональным вариантом in vitro. Предпочтительно, IgM, содержащий J-цепь, является пентамерным IgM.
"Специфические стадии очистки димеров или полимеров IgA, содержащего J-цепь" относятся к стадиям очистки, которые были бы включены в процесс очистки, специфически спланированный, чтобы отделить димеры или полимеры IgA, содержащего J-цепь, от других белков, таких как мономерный IgA, другие иммуноглобулины и другие белки плазмы. Такие специфические стадии очистки могут, например, включать аффинную хроматографию с применением лиганда, специфически связывающегося с J-цепью, или эксклюзионную хроматографию, отбирающую фракции, содержащие белки с молекулярной массой, соответствующей димерам IgA, содержащего J-цепь. В то время как процесс получения композиции из плазмы может содержать способы, которые ведут к обогащению IgA или даже IgA, содержащего J-цепь, такие как ионообменная хроматография, IgA, содержащий J-цепь, не является специфически очищенным. Таким образом, "IgA, содержащий J-цепь, в неочищенном виде" относится к композиции, содержащей менее чем 80% IgA, содержащего J-цепь, как правило, менее чем 70%, 60% или 50% IgA, содержащего J-цепь, он может даже сдержать менее чем 40%, 30%, 25%, 20%, 15% или даже 10% IgA, содержащего J-цепь.
"Специфические стадии очистки IgM, содержащего J-цепь" относятся к стадиям очистки, которые были бы включены в процесс очистки, специфически спланированный, чтобы отделить IgM, содержащий J-цепь, от других белков, таких как другие иммуноглобулины, и другие белки плазмы. Такие специфические стадии очистки могут, например, включать аффинную хроматографию с лигандом, специфически связывающимся с IgM или с J-цепью, или эксклюзионную хроматографию, отбирающую фракции, содержащие белки молекулярной массы, соответствующей пентамерам IgM, содержащего J-цепь. В то время как процесс получения композиции из плазмы может содержать способы, которые ведут к обогащению IgM, содержащего J-цепь, такие как ионообменная хроматография, IgM, содержащий J-цепь, не является специфически очищенным. Таким образом "IgM, содержащий J-цепь, в неочищенном виде" относится к композиции, содержащей менее чем 80% IgM, содержащего J-цепь, как правило, менее чем 70%, 60%, или 50% IgM, содержащего J-цепь, он может даже содержать менее чем 40%, 30%, 25%, 20%, 15% или даже 10% IgM, содержащего J-цепь.
Как используют в настоящем документе, термин "секреторный компонент" относится к белку, который специфически связывается с иммуноглобулином, содержащим J-цепь, и относится к или его получают из, или он идентичен внеклеточной части полимерного рецептора иммуноглобулина (pIgR), предпочтительно pIgR млекопитающих, более предпочтительно pIGR приматов, наиболее предпочтительно pIgR человека. Предпочтительно, секреторный компонент придает увеличенную стабильность иммуноглобулину, содержащему J-цепь. Как было подробно описано выше, наиболее предпочтительным секреторным компонентом является секреторный компонент человека, например, соответствующий остаткам от 19 до 603 SEQ ID NO: 1. Однако аминокислотные делеции, вставки, замены могут быть включены при условии, что они приводят к функциональному белку, т.е. такому, который все еще способен объединяться с IgA, содержащим J-цепь, и предпочтительно придающий ему защиту от протеолитического расщепления. Гомологи из других видов млекопитающих также включены, поскольку они являются химерными белками, содержащими части из различных видов.
Термин "% [процент]", когда его используют для описания содержания иммуноглобулина в композиции/препарате, означает массу на массу белка.
Список чертежей
Изобретение будет проиллюстрировано следующими неограничивающими примерами, касающимися следующих чертежей и списка последовательностей:
На фигуре 1 представлена диаграмма структуры мономерного, димерного и содержащего J-цепь секреторного IgA.
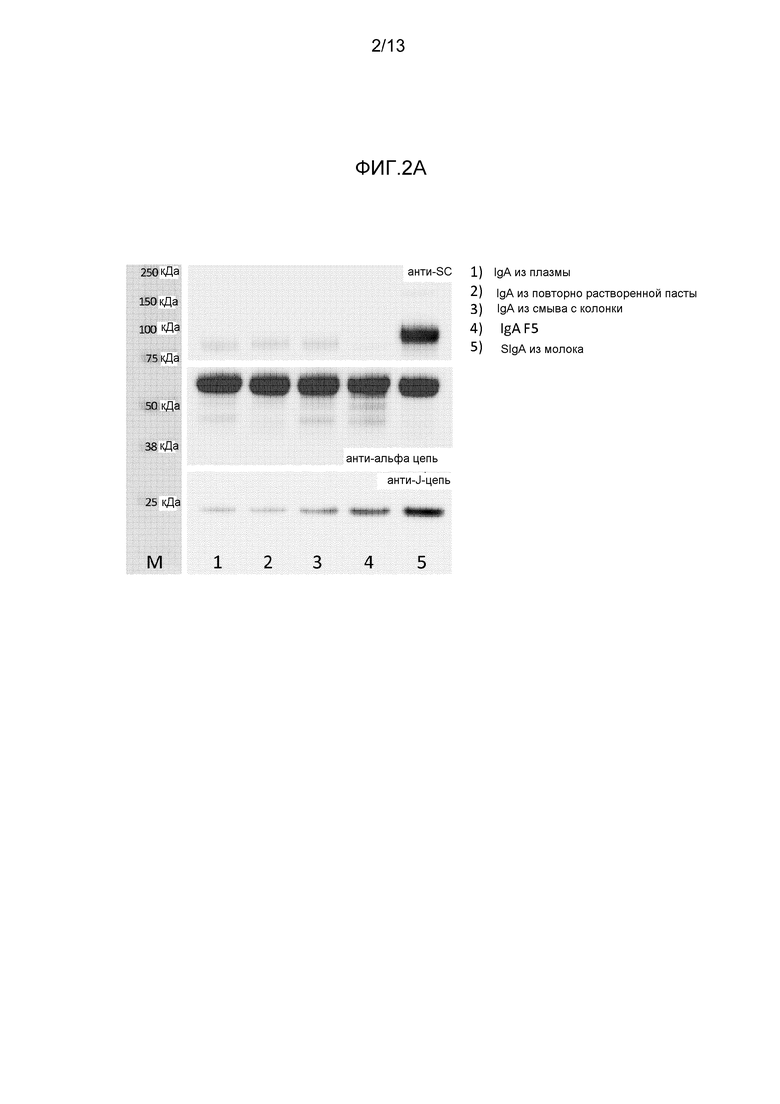
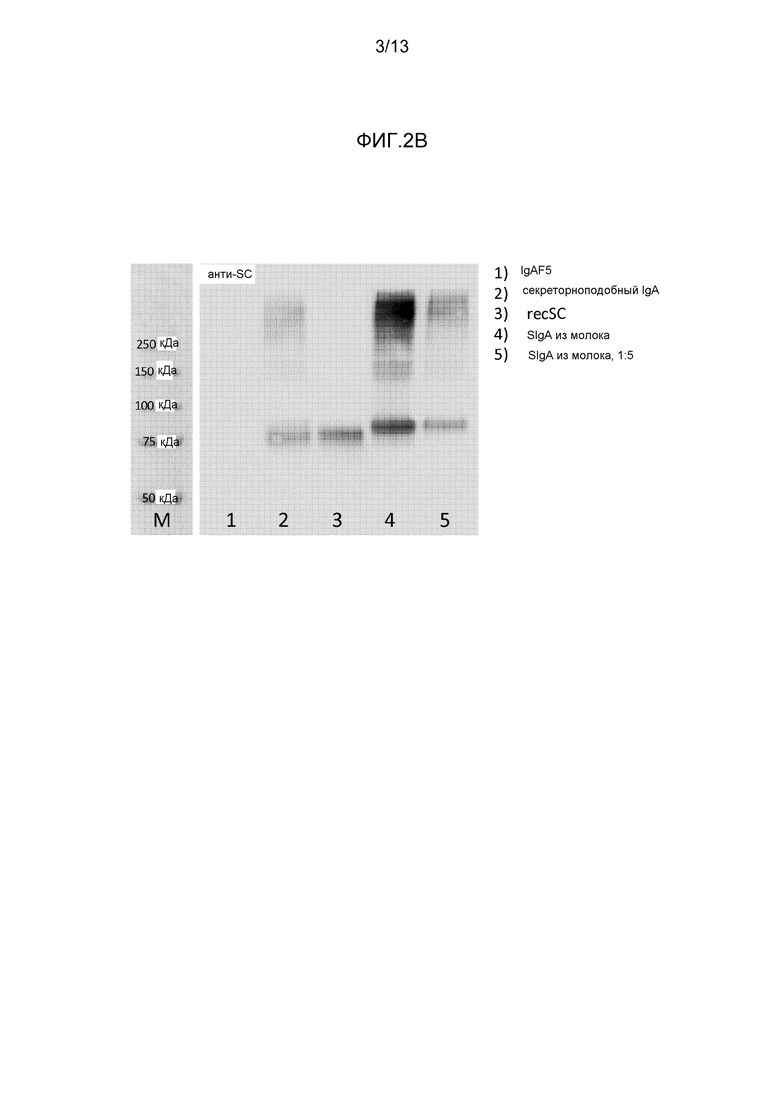
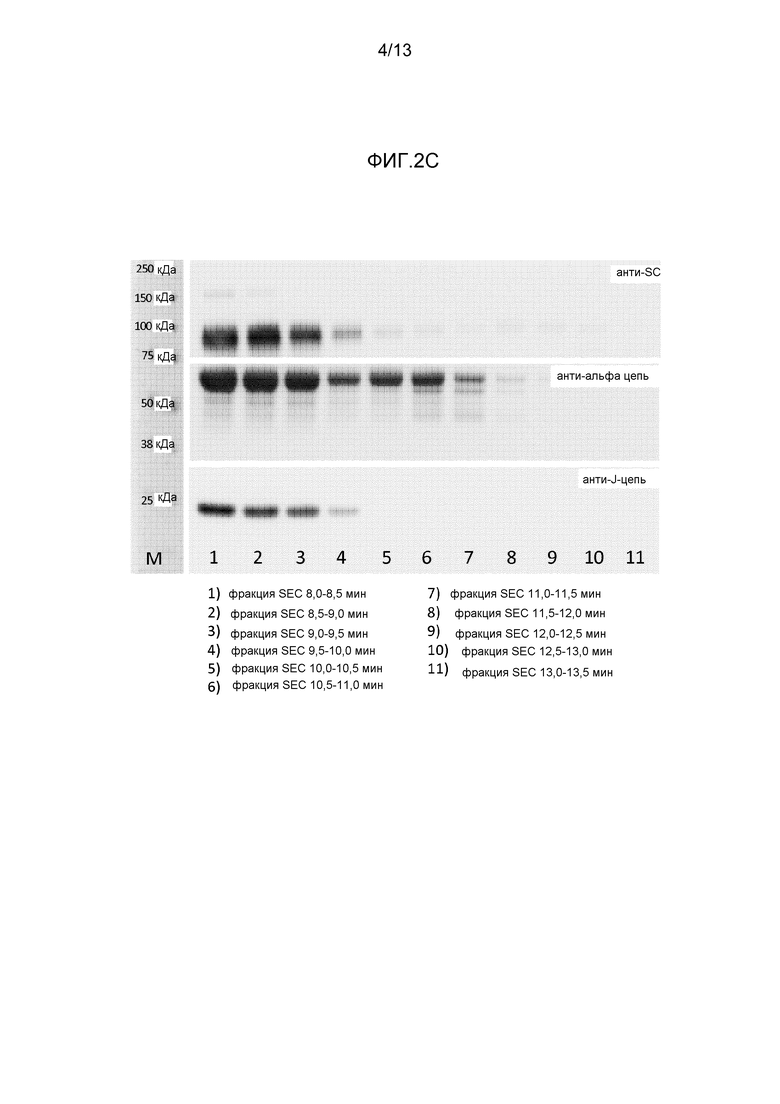
На фигуре 2 представлены вестерн-блоттинги различных препаратов IgA, полученных с различными антителами:
- На фиг. 2A представлен вестерн-блоттинг различных препаратов IgA, полученных с применением антитела к α-цепи, J-цепи или секреторному компоненту.
- На фиг. 2B представлен вестерн-блоттинг различных препаратов секреторноподобного и секреторного IgA, полученного с применением антитела к антисекреторному компоненту.
- На фиг. 2C представлен вестерн-блоттинг различных фракций эксклюзионной хроматографии секреторноподобного IgAF5, полученного с применением антитела к секреторному компоненту, α-цепи и J-цепи.
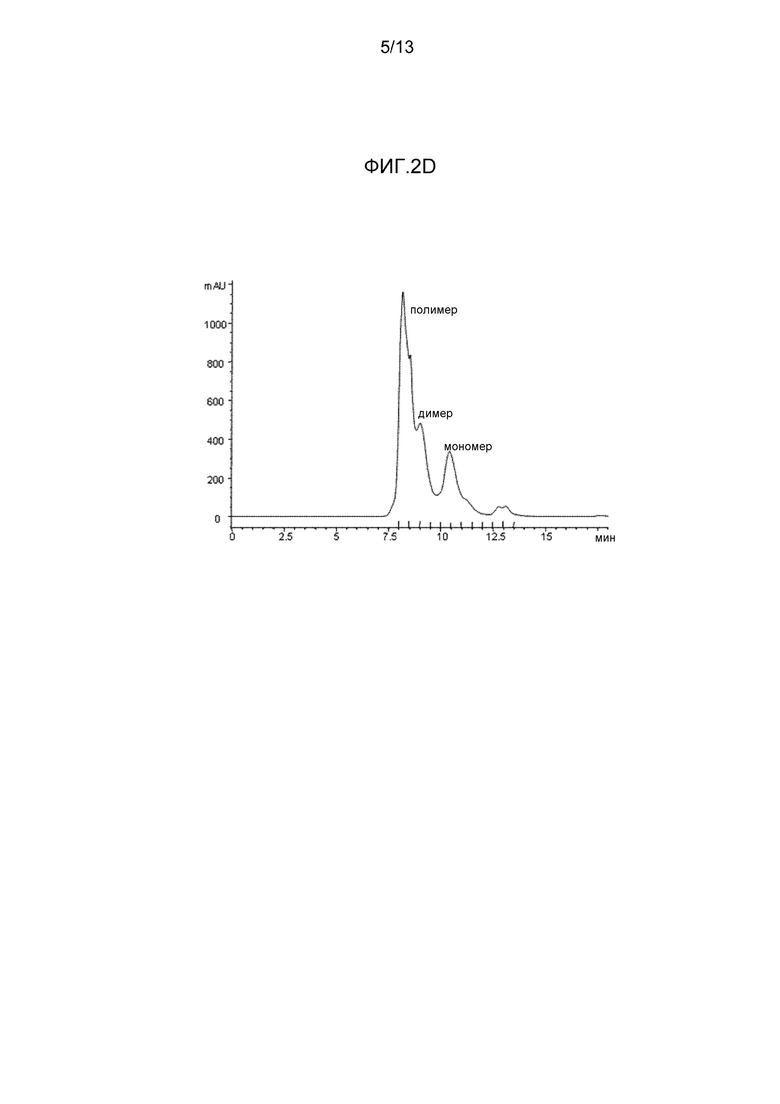
- На фиг. 2D представлена хроматограмма цикла эксклюзионной хроматографии секреторноподобного IgAF5.
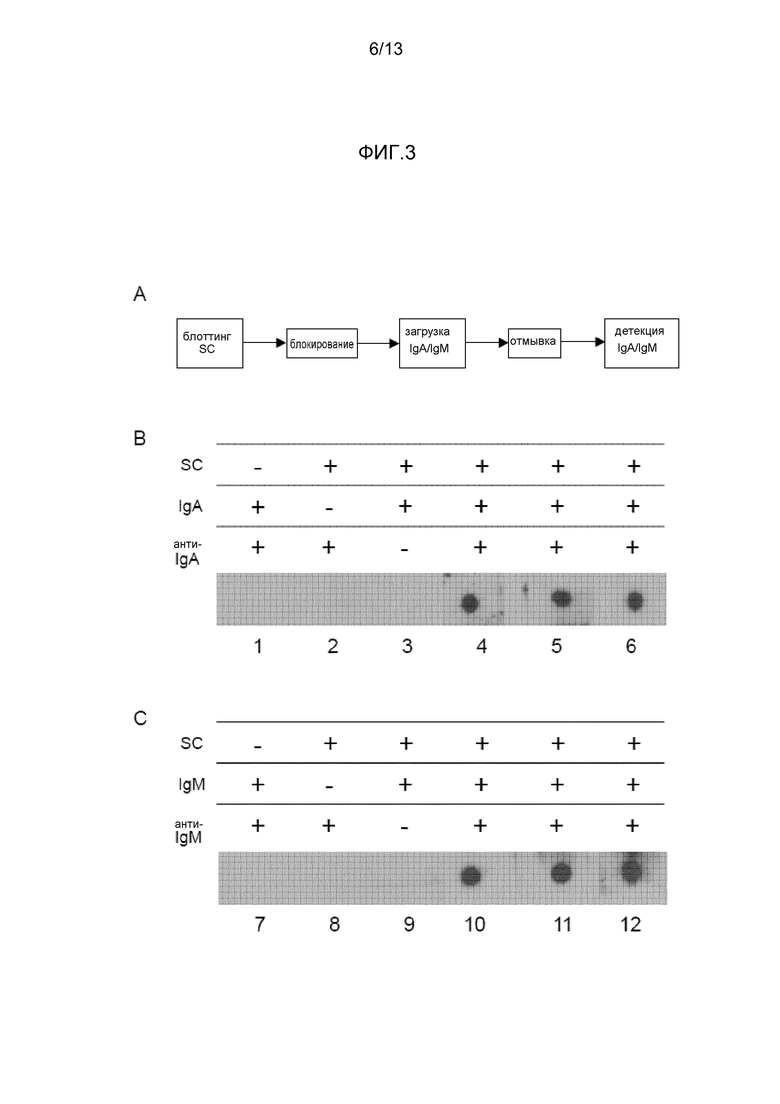
На фигуре 3 представлены точечные блоты с применением иммобилизованного секреторного компонента:
- На фиг. 3A представлена блок-схема того, как налаживают анализ.
- На фиг. 3B представлен SC, захватывающий IgA, содержащий J-цепь.
- На фиг. 3C представлен SC, захватывающий IgM, содержащий J-цепь.
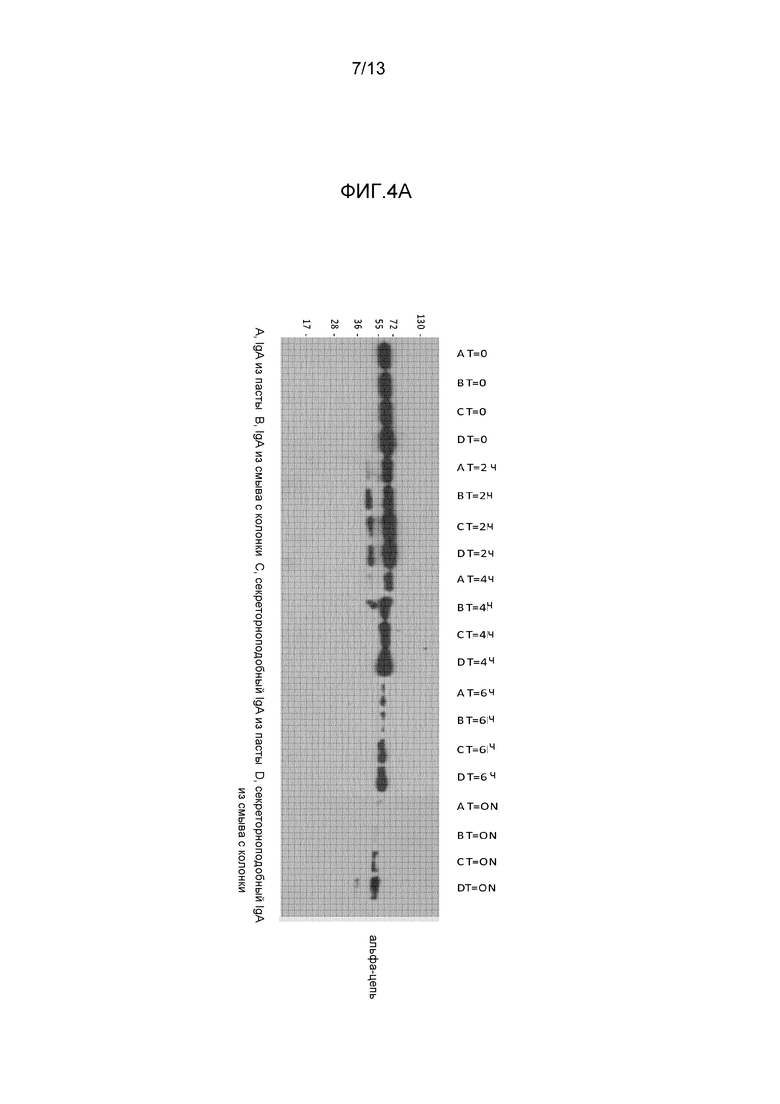
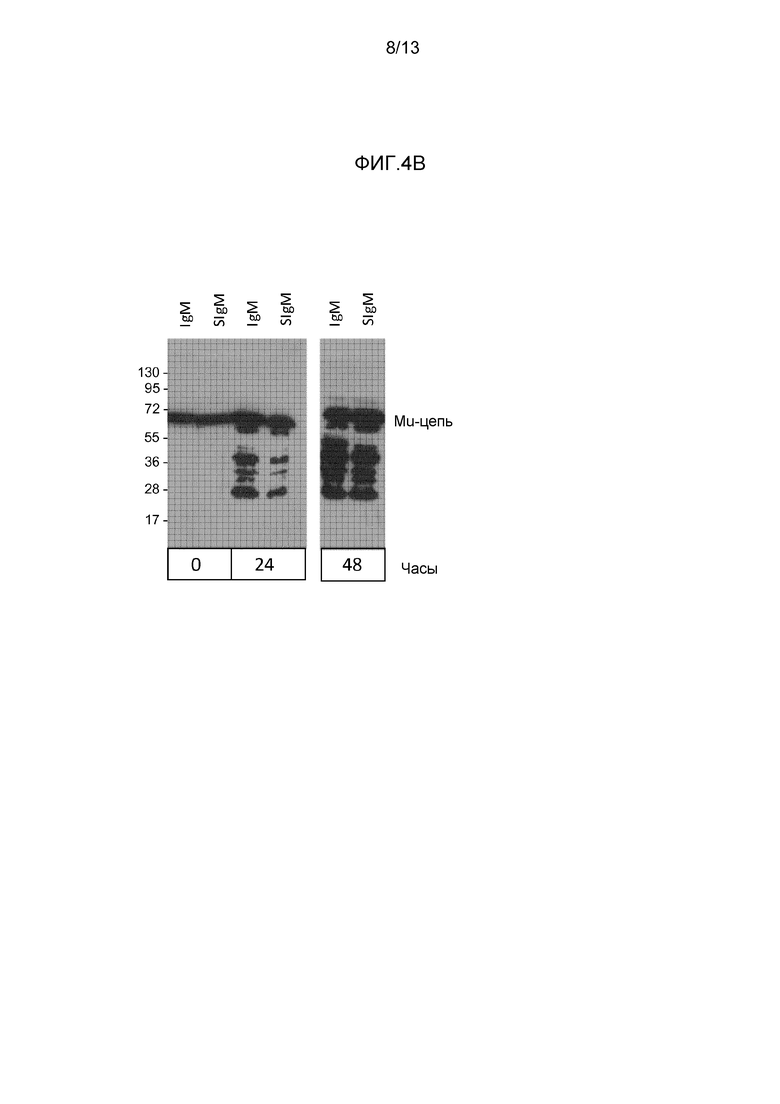
На фигуре 4 представлены вестерн-блоттинги экспериментов по определению зависимости от времени различных препаратов IgA (A) или препаратов IgM (B), инкубированных с кишечными смывами. Блоты получали с применением антител к тяжелой цепи.
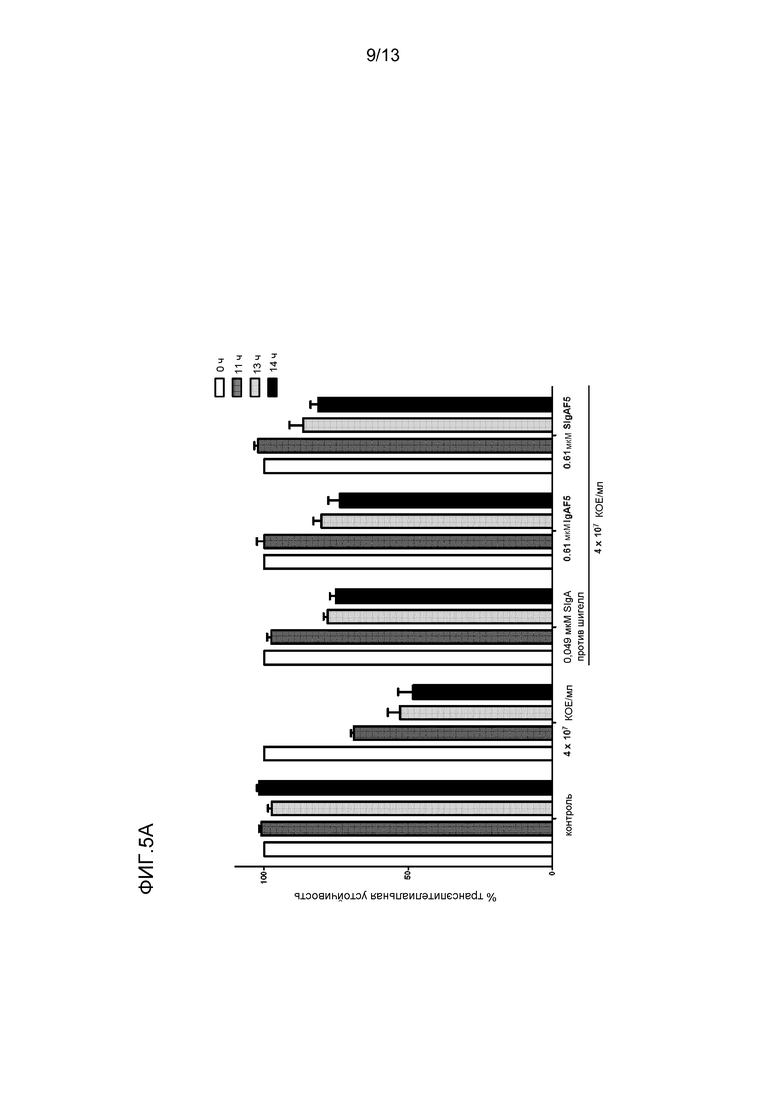
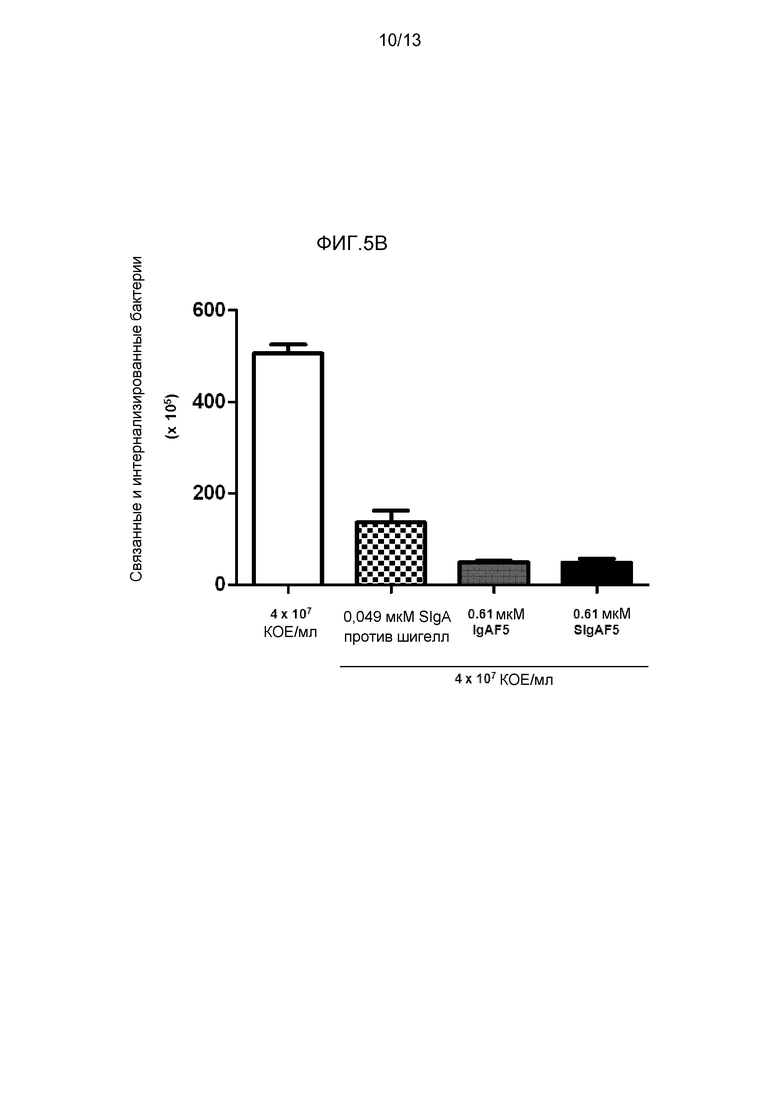
На фигуре 5 представлена защита от инфекции шигелл посредством различных препаратов IgA:
- На фиг. 5A представлено снижение трансэпителиальной устойчивости шигелл к поляризованным монослоям Caco-2, и защита от такого уменьшения в TER посредством SIgA, IgAF5 и SIgAF5 против шигелл (см. Phalipon A et al (1995) J. Exp. Med. 182: 769-778).
- На фиг. 5B представлено снижение численности связанных и интернализированных бактерий посредством SIgA, IgAF5 и SIgAF5 к шигеллам.
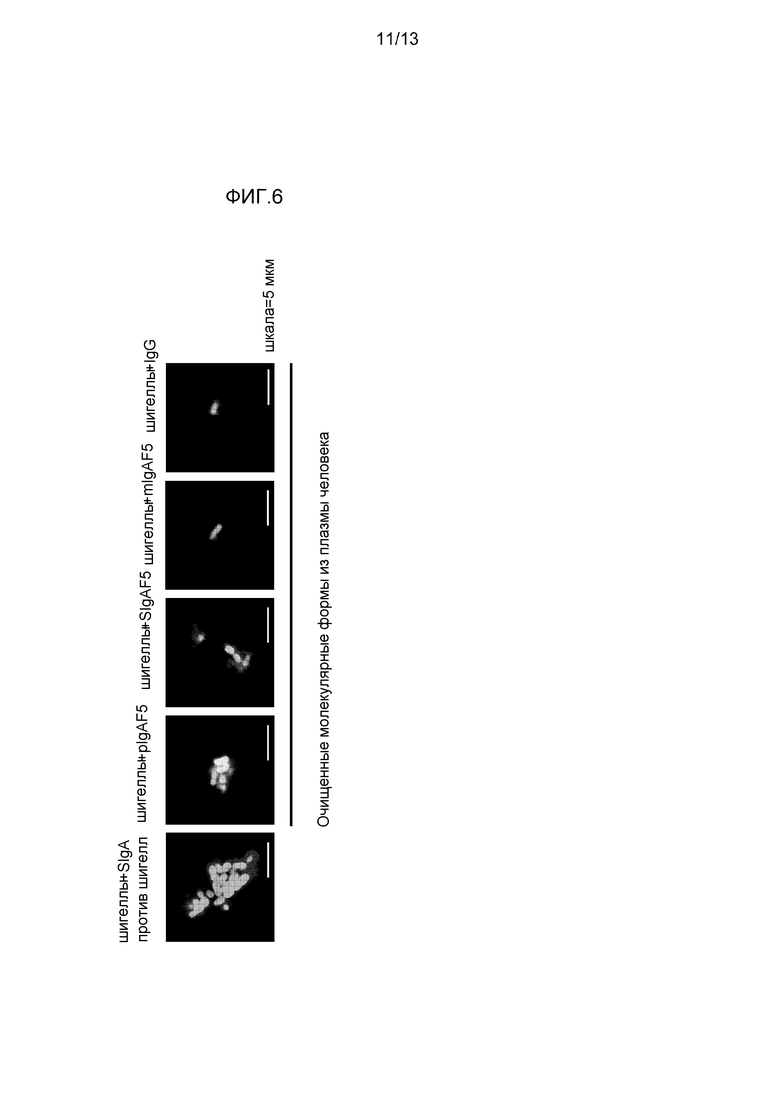
На фигуре 6 представлены снимки шигелл в иммунных комплексах, полученных после инкубации с SIgA(SIgAC5), против шигелл, примененных в качестве положительного контроля, и полученные из плазмы полимерные IgAF5, SIgAF5, мономерные IgAF5 и IgG.
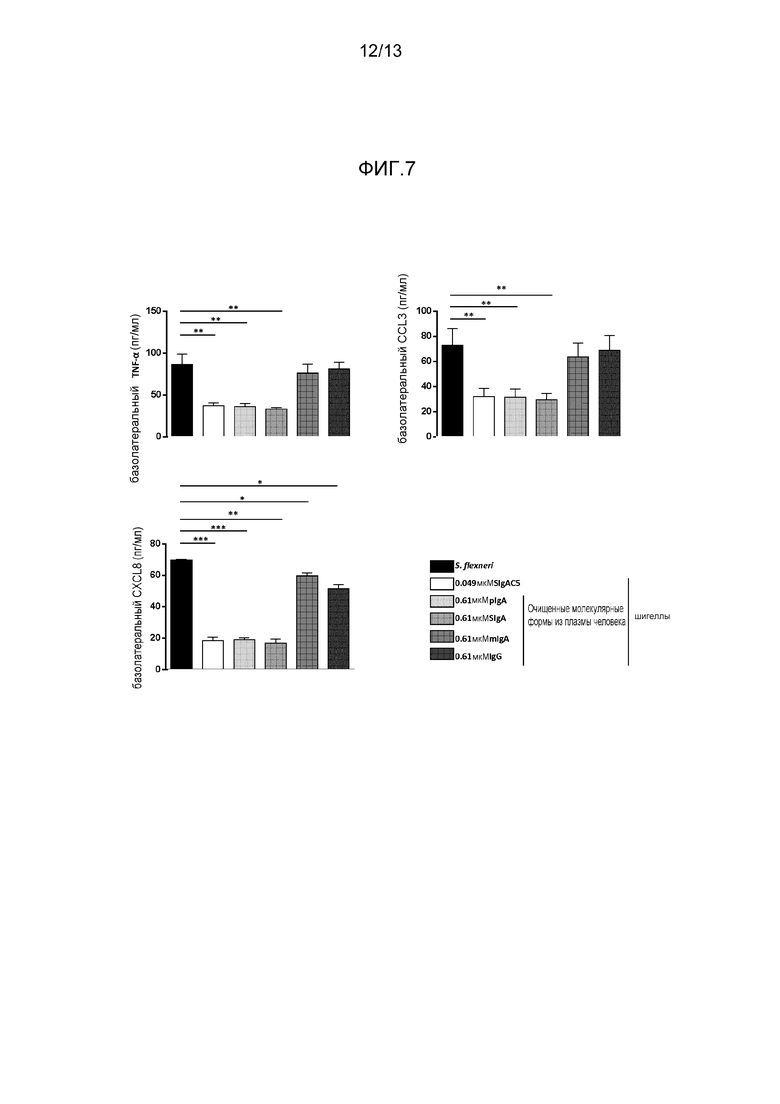
На фигуре 7 представлена секреция цитокина TNF-α и хемокинов CXCL8 и CCL3 посредством поляризованных монослоев эпителиальных клеток Caco-2, подвергнутых воздействию шигелл в отдельности или в комплексе с различными Аb.
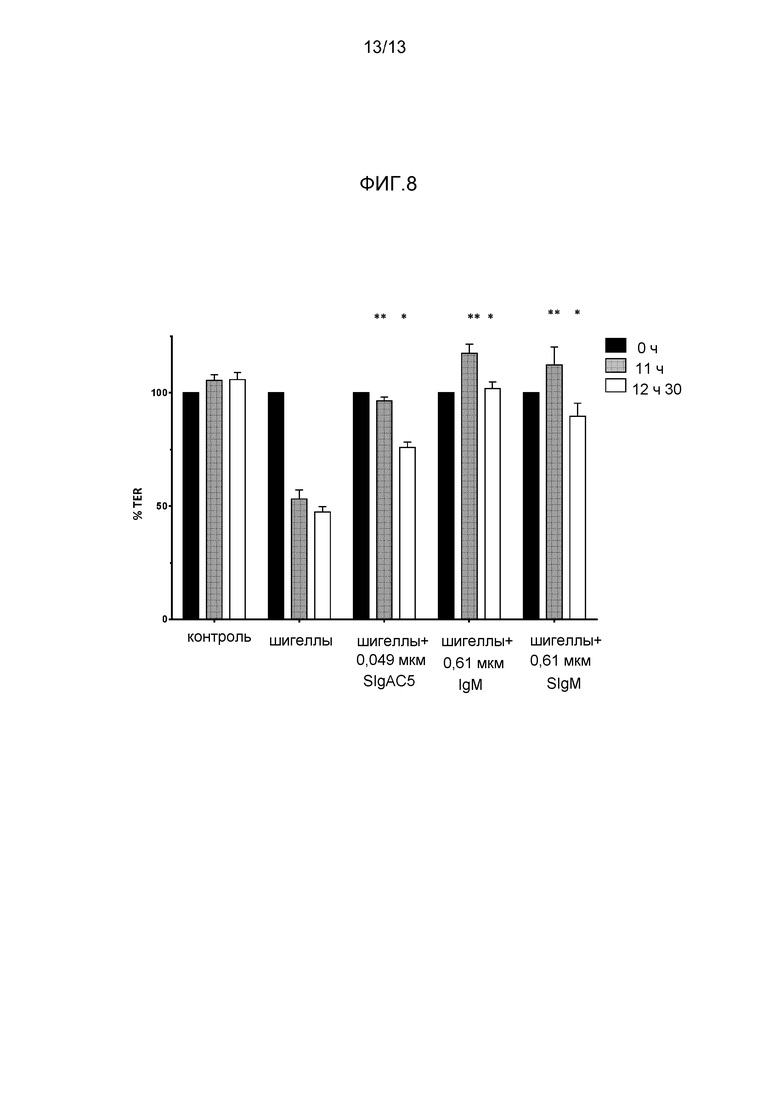
На фигуре 8 представлена защита от инфицирования шигеллой посредством препаратов пентамерного IgM и SIgM.
- На фиг 8 представлено снижение трансэпителиальной устойчивости шигелл на поляризованных монослоях Caco-2, и защита от такого снижения в TER посредством SIgA, IgM и SIgM против шигелл (см. Phalipon A et al (1995) J. Exp. Med. 182: 769-778).
В SEQ ID NO: 1 представлена белковая последовательность pIgR человека.
Примеры
Пример 1: Вестерн-блоттинг препаратов IgA из плазмы, смешанных с рекомбинантным секреторным компонентом
Материалы и методы
1.1 Получение IgA из плазмы посредством аффинной хроматографии и/или посредством последовательной элюции колонки MPHQ
IgA плазмы человека очищали посредством аффинной хроматографии с применением смолы CaptureSelect Human IgA (Bioaffinity Company BAC, Naarden, Netherlands) в соответствии с протоколом производителя смолы с применением 3 различных источников IgA плазмы в качестве исходного вещества, а именно криообедненной плазмы, повторно растворенной пасты, фракционированной в холодном этаноле, или фракции смыва из анионно-обменной (AIEX) хроматографической колонки, полученные посредством вычищения указанной колонки в соответствии с коммерчески применимым процессом очистки IgG CSL Behring AG (Berne, Switzerland). В кратком изложении, криообедненный пул плазмы, повторно растворенную пасту или фракцию смыва AIEX разбавляли в фосфатно-солевом буфере (PBS) до концентрации IgA приблизительно 1 мг/мл, а затем загружали в уравновешенную PBS колонку CaptureSelect Human IgA, без превышения способности колонки к связыванию IgA. После загрузки колонку отмывали PBS, и IgA элюировали с применением глицинового буфера при pH 3. Элюат доводили до pH 4,5 с применением 0,5M Tris (pH 8) и концентрировали до 16 мг/мл белка в PBS. SIgA из молока человека очищали тем же способом.
Из стадии хроматографии AIEX производственного процесса IVIg CSL Behring AG (Berne, Switzerland), фракцию F4 получали после пост-промывания колонки Macro-Prep High Q (Bio-Rad, Hercule, CA) с применением 10 мМ фосфата/30 мМ ацетата при pH 6,5 посредством элюции с применением 55 мМ тартрата/5 мМ ацетата при pH 7,6. Затем фракцию F5 элюировали с применением 50 мМ фосфата/25 мМ цитрата при pH 5,0. F4 и F5 приводили к приблизительно 1 мг/мл в PBS посредством ультра-/диафильтрации, а затем изымали IgG посредством аффинной хроматографии с применением смолы IgSelect (GE Healthcare, Glattbrugg, Switzerland). IgAF4 был напрямую собран в потоке IgSelect хроматографии загрузки F4. Для получения IgAF5, из IgSelect поток загрузки F5 извлекали IgM посредством аффинной хроматографии с применением смолы CaptureSelect Human IgM (Bioaffinity Company BAC). IgAF4 и IgAF5 приводили к конечным концентрациям посредством ультра-/диафильтрации.
1.2 Получение IgM из плазмы посредством аффинной хроматографии
IgM плазмы человека очищали посредством аффинной хроматографии с применением смолы CaptureSelect Human IgM (Bioaffinity Company BAC, Naarden, Netherlands) в соответствии с протоколом производителя смолы с применением тех же 3 различных источников в качестве исходного вещества, как описано в разделе 1.1 для IgA, а именно криообедненной плазмы, повторно растворенной пасты, фракционированной в холодном этаноле, или фракции смыва из анионо-обменной (AIEX) хроматографической колонки, полученной посредством извлечения из указанной колонки, в соответствии с коммерчески примененным процессом очистки IgG CSL Behring AG (Berne, Switzerland). В кратком изложении, криообедненный пул плазмы, повторно растворенную пасту или фракцию смыва AIEX разбавляли в фосфатно-солевом буфере (PBS) до концентрации IgM приблизительно 1 мг/мл, а затем загружали на уравновешенную PBS колонку CaptureSelect Human IgM, без превышения способности колонки связывать IgM. После загрузки колонку отмывали с применением PBS, и IgM элюировали с применением глицинового буфера при pH 3. Элюат был доведен до pH 4,5 с применением 0,5M Tris (pH 8) и сконцентрирован до 10 мг/мл белка в PBS.
1.3. Вестерн-блоттинги
SDS-PAGE и электроблоттинг на нитроцеллюлозных мембранах (NC) проводили с применением системы мини-клеток из Invitrogen (Carlsbad, CA) по протоколам производителя. В кратком изложении, образцы были денатурированы в буфере для образцов в восстановительных или невосстановительных условиях, соответственно, и электрофоретически отделены на градиентных гелях заводского изготовления, NuPAGE Novex Bis-Tris 4-12% 1,0 мм 10-луночный, с применением NuPAGE MOPS Electrophoresis Buffer (Invitrogen). Мокрый перенос на NC мембраны (0,2 мкм) проводили с применением XCell II Blot Module (Invitrogen) и NuPAGE Transfer Buffer. Мембраны затем блокировали в течение 30 мин в растворе PBS-0,5% Tween 20 (PBS-T), содержащем 4% порошковое молоко Rapilait skim (Migros, Switzerland). Для иммуноблоттинга поликлональных антител кролика применяли: 1) альфа-цепь кролика против человека (Dako, конъюгированный с пероксидазой хрена (HRP): разведение 1/5000); 2) J-цепь кролика против человека (BioGenex, Fremont, CA; разведение 1/300), с последующей вторичной антисывороткой против кролика, конъюгированной с HRP (Sigma; разведение 1/10000); 3) SC кролика против человека (Dako; разведение 1/5000), с последующей вторичной антисывороткой против кролика, конъюгированной с HRP (Sigma; разведение 1/10000). Все инкубации проводили в PBS-T, содержащем 4% порошковое молоко, при температуре окружающей среды в течение 1-2 часов. После окончательной промывки с применением PBS-T иммунологический анализ на мембране был выявлен посредством хемилюминесценции и записан в цифровом виде в системе ImageQuant LAS 4000 (GE Healthcare Lifesciences).
1.4 Объединение IgA, полученного из плазмы, с рекомбинантным секреторным компонентом
Секреторноподобный IgA получали посредством комбинирования in vitro 100 мг IgAF5 с 4 мг рекомбинантного секреторного компонента человека (recSC). Объединение проводили в PBS в течение 30 мин при комнатной температуре, как описано ранее в (Crottet, P., и Corthesy, B. (1998) J. Immunol. 161:5445-5453).
1.5 Фракционирование посредством эксклюзионной хроматографии (SEC)
IgAF5, содержащий секреторноподобный IgA (объединенный in vitro с recSC), инъецировали при 200 мкг/20 мкл в Agilent Technologies 1050 HPLC system для эксклюзионной хроматографии при скорости потока 1,5 мл/мин на TSKgel G3000SWXL 7,8 мм ID Ч30 см колонке (Tosoh Bioscience). Фракции 0,75 мл собирали между 8,0 и 13,5 мин времени удержания с интервалами в 30 сек.
Результаты
Результаты представлены на фигуре 2. На фигуре 2A представлено сравнение IgA, очищенного посредством аффинной хроматографии от плазмы, от повторно растворенной пасты и от фракции смыва AIEX или посредством последовательной элюции для получения IgAF5, как описано в 1.1 с SIgA из молока человека. Секреторный компонент был обнаружен в SIgA из молока, но ни в одной из фракций IgA, очищенной от плазмы человека. Все препараты содержали то же самое количество тяжелой цепи/альфа-цепи IgA. Как и ожидалось, SIgA из молока содержал наивысшее количество J-цепи, поскольку существенным образом ожидается, что все молекулы IgA присутствуют в качестве димеров, содержащих J-цепь. Количество J-цепи и, таким образом, димеров IgA, содержащих J-цепь, в IgA, очищенном от плазмы, было низким. Ожидается, что только малая часть IgA в плазме присутствует в димерной форме. Сходное содержание J-цепи наблюдали в IgA, очищенном от повторно растворенной пасты. Удивительно, что увеличенная фракция IgA присутствовала в качестве димеров, содержащих J-цепь, во фракции смыва с колонки, о чем свидетельствует повышенное количество J-цепи. Удивительно, что оно было дополнительно увеличено в IgAF5. Это накопление произошло без применения специфической стадии процесса для обогащения.
На фигуре 2B представлено, что в IgAF5 не присутствует никакого секреторного компонента, содержащего IgA. После объединения с recSC были обнаружены свободный recSC (75 кДа) и димерный IgA, объединенный с recSC. Фактически секреторноподобный IgA плазмы оказался сходным с SIgA из молока. Было определено, что в препарате IgAF5, примененном в представленном эксперименте, содержание IgAF5, содержащего J-цепь, составляло приблизительно 20%. Фактически, сила сигнала, наблюдаемого в дорожке 2, была сравнима с сигналом SIgA молока человека, разведенного 1:5.
На фигуре 2C представлено содержание SC, альфа-цепи и J-цепи IgA во фракциях, полученных посредством эксклюзионной хроматографии секреторноподобного IgAF5. recSC наблюдали в ранних фракциях, соответствующих формам с высокой молекулярной массой IgA-подобных полимерных и димерных форм. Появление SC совпадало с появлением J-цепи, указывая на то, что фактически фракция IgAF5, содержащая SC, была димерной фракцией, содержащей J-цепь. Кроме того, альфа-цепь IgA обнаружили во фракциях с более маленькой молекулярной массой, вероятно содержащей мономерную фракцию IgAF5; эти фракции были лишены SC и J-цепи. Эти данные продемонстрировали, что recSC, смешанный с IgA, полученным из плазмы, содержащим мономерные и димерные формы IgA, специфически объединен с димерными формами IgA, содержащим J-цепь.
На фигуре 2D представлена хроматограмма цикла SEC, в течение которого фракции собирали между временем удержания 8,0 мин и 13,5 мин. Пики, представляющие полимеры, димеры и мономеры IgA, указаны.
Пример 2: Анализ точечного блота реассоциации (DORA)
Анализ точечного блота реассоциации применяли, чтобы продемонстрировать объединение иммобилизованного секреторного компонента с IgA или IgM, полученными из плазмы, in vitro. В кратком изложении, как представлено на фигуре 3A, секреторный компонент был точечно нанесен на мембраны блоттинга; участки неспецифического связывания блокировали. После этого IgA, полученный из плазмы (фигура 3B), или IgM, полученный посредством аффинной хроматографии из плазмы (фигура 3C) (дорожка 4), из повторно растворенной пасты (дорожка 5) или из фракции смыва AIEX (дорожка 6), полученной, как описано в 1.1 и 1.2, наносили на мембрану. После отмывания несвязанных IgA или IgM, связанный IgA/IgM обнаруживали, как описано в кратком изложении ниже.
DORA проводили существенным образом, как описано (Rindisbacher, L. et al. (1995) J. Biol. Chem. 270:14220-14228), со следующими модификациями: мембраны для блоттинга состояли из полимера фторида поливинилидона (PVDF), блокирующий раствор был фосфатно-солевой буфер-0,05% Tween-20 (PBS-T), содержащий 1% бычий сывороточный альбумин (BSA), сырьевые препараты, обогащенные IgA, применяли для оверлейной инкубации в 200 мкл PBS-T, содержащем 0,1% BSA, и обнаружение антител было непосредственно соединено с HRP.
Результаты для IgA представлены на фигуре 3B. Иммобилизованный секреторный компонент был способен захватывать IgA, полученный из плазмы. Сходно с тем, что представлено на фигуре 2C, это демонстрирует, что recSC объединен с димерами IgA, полученными из плазмы.
Результаты для IgM представлены на фигуре 3C. Иммобилизованный секреторный компонент был способен захватывать IgM, полученный из плазмы. Это демонстрирует, что recSC связан с IgM, полученным из плазмы.
Пример 3: Расщепление секреторноподобного IgA и секреторноподобного IgM с применением кишечных смывов
Чтобы доказать функциональное преимущество объединения очищенного секреторного компонента с IgA и IgM, содержащими J-цепь, соответственно, IgA и IgM получали, как описано в параграфе 1.1 и 1.2. Секреторноподобный IgA получали посредством обогащения IgA, содержащего J-цепь, с применением эксклюзионной хроматографии и комбинирования in vitro их 10 мкг с 2,5 мкг рекомбинантного секреторного компонента человека (hSCrec). Секреторноподобный IgM получали посредством обогащения пентамерного IgM с применением эксклюзионной хроматографии и комбинирования in vitro их 25 мкг с 2,5 мкг рекомбинантного секреторного компонента человека (hSCrec). Объединение проводили в PBS в течение 30 мин при комнатной температуре, как описано ранее (Crottet, P., и Corthesy, B. (1998) J. Immunol. 161:5445-5453). Целостность и надлежащая сборка молекул в возможно ковалентные комплексы проверяли посредством SDS-PAGE в невосстановительных и восстановительных условиях, с последующим вестерн-блоттингом и иммунологическим анализом с применением антисыворотки, специфичной для hSC, как указано выше.
Сбор кишечных смывов из мышей BALB/c (возрастом 4-6 недель) проводили в соответствии с опубликованной процедурой (Crottet, P., и Corthesy, B. (1998) J. Immunol. 161:5445-5453). Для расщепления in vitro 120 нг очищенного IgA, содержащего J-цепь, и повторно растворенного секреторноподобного IgA смешивали (или нет) с 1 или 2 мкл кишечных смывов в окончательно объеме 20 мкл PBS и инкубировали при 37°C в течение различных периодов времени, как показано на фигуре 4 (T=время в часах). Для расщепления IgM in vitro 250 нг очищенного IgM и секреторноподобного IgM смешивали с 4 мкл кишечных смывов. Реакции останавливали посредством добавления 2 мкл смеси ингибиторов протеазы Complete™ (Roche Applied Science, Rotkreuz, Switzerland) и держали в замороженном состоянии до проведения анализа посредством вестерн-блоттинга, детектирующего восстановленную форму тяжелой цепи антитела.
Результаты представлены на фигуре 4A и B. IgA из повторно растворенной пасты и со смыва с колонки, полученный, как описано в 1.1, и IgM со смыва с колонки, как описано в 1.2, были либо применены как таковые, либо оба после объединения с recSC для формирования секреторноподобного IgA (фиг. 4A) или секреторноподобного IgM (фиг. 4B). Для IgA после 4 часов расщепления с применением кишечных ферментов сигнал необъединенного IgA начал уменьшаться, указывая на протеолитическое расщепление; эффект был сильнее через 6 часов и после расщепления в течение ночи, никакой интактной альфа-цепи IgA не было выявлено посредством вестерн-блоттинга. В отличие от этого, секреторноподобный IgA был гораздо менее чувствителен к расщеплению посредством кишечных протеаз и даже после воздействия в течение ночи значительная часть альфа-цепи IgA внутри секреторноподобного IgA оставалась интактной.
Для IgM (фиг. 4B) представлено сравнение IgM и только что объединенного секреторноподобного IgM (SIgM) с теми же препаратами после 24 часов и 48 часов расщепления. Появление деградированных фрагментов mu-цепей произошло быстрее и интенсивнее для IgM по сравнению с SIgM, подтверждая для IgM - сходно как для IgA, - что объединение с recSC обеспечивало улучшенную структурную стабильность.
В общем, это демонстрирует, что специфическое объединение с recSC обеспечивало улучшенную структурную стабильность и создаст склонные к расщеплению молекулы IgA плазмы, подходящие для применения на слизистой оболочке, например, посредством перорального пути введения.
Пример 4: Shigella flexneri
Эпителиальную клеточную линию аденокарциномы кишечника человека Caco-2 (American Type Tissue Collection) высевали на полиэстеровые фильтры Snapwell (диаметр 12 мм; размер пор 0,4 мкм; Corning Costar), как описано (Crottet, S., Corthesy-Theulaz, I., Spertini, F., и Corthesy, B. (2002) J. Biol. Chem. 277:33978-33986). Целостность поляризованного клеточного монослоя Caco-2 проверяли посредством измерения трансэпителиального электрического сопротивления (TER) с применением устройства Millicell-ERS (Millipore). Значения TER хорошо дифференцированных монослоев варьировали между 450 и 550 ΩЧсм2.
2×107 бактерий (Phalipon A. et al. (1995) J. Exp. Med. 182:769-778) смешивали со 100 мкг IgA, 125 мкг секреторноподобного IgA, 275 мкг IgM или 300 мкг SIgM в окончательном объеме 500 мкл простого DMEM (P-DMEM: DMEM, дополненный 10 мМ HEPES, 20 мкг/мл трансферрина, 2 мМ глутамином, 1% заменимых аминокислот, 1 мМ пируватом натрия) и инкубировали в течение 1 часа при КТ при легком перемешивании. Смеси ресуспендировали в P-DMEM для инфицирования поляризованных клеточных монослоев Caco-2.
За 1 час перед применением поляризованных клеточных монослоев Caco-2 C-DMEM заменяли на P-DMEM в обоих апикальном и базолатеральном компартментах. Апикальную среду затем заменяли посредством 500 мкл бактериальных суспензий (2×107 бактерий) отдельно или в комбинации с антителом. Значения TER измеряли в выбранных временных точках от начала инфекционного периода.
Для определения количества бактерий, которые адгезировались и инфицировали клетки, клетки Caco-2 в фильтрах отмывали три раза с применением PBS, клетки инкубировали в 500 мкл холодного лизирующего буфера [10 мМ Tris-HCl (pH 7), 0,2% Nonidet P-40, 50 мМ NaCl, 2 мМ ЭДТА (pH 8)] в течение 5 мин на льду и лизировали посредством пипетирования. Серийные разведения лизатов клеток (10-2-10-6) наносили на чашки с агаром LB и через 24 часа инкубации при 37°C, колониеобразующие единицы (КОЕ) определяли, подсчитывая на глаз дубликатные чашки.
Для изучения целостности клеточных монослоев Caco-2 отмывали Snapwells с применением PBS с последующей фиксацией в течение ночи с 5 мл 4% параформальдегида при 4°C. После промывания с применением PBS фильтры пермеабилизировали и неспецифически связывающие участки были блокированы с применением PBS, содержащим 5% ЭТС и 0,2% Triton X-100 (PBS-Tr), в течение 30 мин при КТ. Все антитела разбавляли в PBS-Tr. Фильтры инкубировали с антителами кролика против человека ZO-1 (1/200, Invitrogen) в течение 2 часов при КТ, промывали в PBS, с последующей обработкой антителами IgG козы против кролика, конъюгированными с Alexa Fluor® 647 (1/100, Invitrogen), в течение 90 мин при КТ. Для визуализации клеток фильтры в конце инкубировали с 100 нг/мл 4ʹ,6-диамидино-2-фенилиндолом (DAPI) в PBS (Invitrogen) в течение 30 мин. Фильтры вынимали из их подставок и покрывали раствором Vectashield для наблюдения с применением конфокального микроскопа Zeiss LSM 710 Meta (Carl Zeiss, Germany), оснащенного объективом 10x или 40x. Изображения обрабатывали с применением программного обеспечения Zeiss ZEN 2009 light.
Для изучения комплексов Shigella-IgA применяли бактерии, постоянно экспрессирующие зеленый флуоресцентный белок, и образование иммунных комплексов верифицировали после инкубации с биотинилированными антителами IgA1/IgA2 мыши против человека (1/10, BD) в течение 30 мин при КТ при легком перемешивании, с последующей обработкой стрептовидином, конъюгированным с цианином 5 (1/400, GE HealthCare), в течение 30 мин при КТ при легком перемешивании. Три промывки с применением PBS проводили между каждым этапом, и все антитела разбавляли с применением PBS/5% FCS. Меченые иммунные комплексы наносили на предметные стекла (Thermo Scientific), готовили препарат и немедленно визуализировали с применением конфокального микроскопа Zeiss LSM 710 Meta (Carl Zeiss, Germany), оснащенного 63x объективом. Изображения обрабатывали с применением программного обеспечения Zeiss ZEN 2009 light.
CXCL8 (IL-8), TNF-α, и CCL3 (MIP-3a) человека в базолатеральном компартменте поляризованных клеточных монослоев Caco-2 количественно определяли посредством ELISA с применением коммерческих наборов (BD Biosciences и R&D Systems, соответственно).
Результаты
Воздействие шигелл на поляризованный монослой эпителиальных клеток кишечника привел к разрыву целостности монослоя, о чем свидетельствовало снижение TER (фигура 5A), к массивной инвазии, о которой свидетельствовали повышенная численность бактерий, обнаруженная при объединении с эпителиальными клетками (фигура 5B), и посредством заметно нарушенного клеточного монослоя, как наблюдали посредством лазерного сканирующего конфокального микроскопа. Добавление секреторноподобного IgA задерживало и частично ингибировало разрушение монослоя, о чем свидетельствовало значительное ингибирование уменьшения TER (фигура 5A), посредством снижения количества бактерий, связанных с эпителиальными клетками (фигура 5B), и посредством более сохраненной целостности клеточного монослоя при анализе посредством конфокальной микроскопии.
Объединение специфичного к шигеллам моноклонального SIgA SIgAC5, полимерного IgA и секреторноподобного IgA привело к образованию иммунных агрегатов (фигура 6) множества бактерий, в отличие от мономерных IgA и IgG, которые покрывали только бактерию (фигура 6). Вероятно, что агрегация бактерий посредством специфического mAb и полученного из плазмы pIgA и секреторноподобного IgA содействовала уменьшению бактериальной адгезии к клеткам Caco-2, наблюдаемым на фигуре 5B. Те же три препарата антитела значимо снижали продуцирование провоспалительных цитокинов/хемокинов посредством клеток Caco-2, в то время как мономерный IgAF4 и IgG не имел (TNF-α и CCL3) или обладал слабым (CXCL8) эффектом (фигура 7). Это указывает на то, что нейтрализация шигелл посредством секреторного и полимерного IgA уменьшает способность Caco-2 клеток отвечать, окончательно приводя к общим противовоспалительным качествам IgA.
Защиту поляризованного монослоя клеток Caco-2 от инфицирования шигеллой достигали сходным образом с применением IgM и секреторноподобного IgM, до уровня, по меньшей мере, сходного с тем, который достигается, когда используется специфический SIgAC5 (фигура 8). Поддержание TER, по меньшей мере, в течение 12 ч 30 минут указывало на то, что изотип IgM обладает нейтрализующими качествами, защищающими монослой Caco-2 от повреждений, наносимых воздействием шигелл.
Пример 5: Предотвращение рецидива инфекции Clostridium difficile (CDI)
Композицию по изобретению применяют в мышиной модели инфицирования Clostridium difficile.
Мыши C57BL/6 обрабатывают с применением смеси пероральных антибиотиков (канамицина, гентамицина, колистина, метронидазола, и ванкомицина) в течение 3 суток, как описано ранее (Chen X, et al. Gastroenterology 2008 Dec;135(6):1984-92). Двумя сутками позже им дают парентеральный фосфат клиндамицина (10 мг/кг s.c.) [Сутки - 1]. Через одни сутки [Сутки 0] их испытывают посредством принудительного кормления 0,5×105 КОЕ токсиногенным штаммом 10465 C. difficile. Через от 1 до 5 суток после введения C. difficile развивается от умеренного до острого колита. Будучи нелеченным, он быстро прогрессирует в тяжелый и фатальный колит у большинства животных. Для лечения первичной инфекции животные получают ванкомицин, в течение 5 суток после испытания посредством C. difficile, и животных мониторируют на предмет смертности, а также присутствия или отсутствия тяжелого CDI с диареей. Животных, оцениваемых как находящихся в предсмертном состоянии, подвергают эвтаназии посредством единичной инъекции пентобарбиталом натрия. Для исследования рецидива CDI у животных, переживших первичное испытание C. difficile, поддерживают под наблюдением до 28 суток. Животных взвешивают 3 раза в неделю от 7 до 28 суток. После прекращения лечения ванкомицином животные получают IgA или секреторноподобный IgA (400 мг/кг массы тела посредством перорального пути введения) в течение 5 суток, начиная с суток после последней дозы ванкомицина.
Результат
Животные, получившие лечение ванкомицином, выживают после первичного инфицирования C. difficile. Однако значительная часть животных - до 70% - погибают от рецидива инфекции C. difficile в течение 3-4 суток после окончания лечения ванкомицином. В противоположность этому, рецидив инфекции предотвращают, если животных лечат секреторноподобным IgA посредством перорального пути введения. IgA плазмы в отдельности не эффективен (или, по меньшей мере, не так эффективен), как секреторноподобный IgA, при предотвращении рецидива инфекции C. difficile.
Альтернативно, композицию применяют в модели перорального воспаления слизистой оболочки, сходной с описанной в Watkins et al. (Oral Dis 2010, 16:655-660).
Соответствующим образом сформулированные препараты IgA (или раствор носителя для контроля) дают профилактически (например, начиная с -3 суток) три раза ежесуточно сирийским золотистым хомячкам в течение всей продолжительности исследования до 28 суток. В модели острого мукозита, вызванного радиацией, на 0 сутки облучали один вывернутый защечный мешок (40 Gy), другой защечный мешок оставляли нетронутым для контроля. Альтернативно, в модели мукозита, вызванного фракционированным облучением, применяли суммарную дозу 60 Gy, разделенную на восемь фракций по 7,5 Gy, как описано в (Watkins, Oral Dis 2010, 16:655-660). В еще другой модели мукозита, индуцированного комбинированным воздействием цисплатина и острого облучения, заболевание вызвано посредством комбинации цисплатина (5 мг/кг) и облучения 35 Gy на 0 сутки. Клиническая оценка перорального мукозита и мониторинг массы тела проводят ежесуточно, начиная с 6 суток и до конца исследования, как правило, на 28 сутки. Система расчета описана в (Watkins Oral Dis, 2010 16:655-660). Кроме того, образцы ткани и плазмы собирают и соответствующим образом обрабатывают в течение всего исследования для гистологического анализа, определения воспалительных маркеров в плазме и для исследований генной экспрессии в различных тканях.
Результаты
У нелеченого/леченого носителем животного развивается пероральный мукозит, с пиком заболевания приблизительно на 16-18 сутки, спонтанным излечением, о котором свидетельствует регрессия мукозита, начинающаяся приблизительно на 18-20 сутки. Животные, леченные IgA и, в частности, секреторноподобным IgA, имеют существенно более низкие показатели мукозита по сравнению с контрольными животными и теряют меньше массы, параллельно с менее тяжелыми гистологическими проявлениями и уменьшенными уровнями воспалительных маркеров (включая в качестве неограничивающих примеров воспалительные цитокины и хемокины). Уменьшение воспаления и продвижение заживления раны подтверждается на уровне экспрессии мРНК посредством методов анализа генной экспрессии.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ, СОДЕРЖАЩИЕ СЕКРЕТОРНЫЙ IgA И ПРОБИОТИКИ | 2020 |
|
RU2821998C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПРОФИЛАКТИКИ ИЛИ ЛЕЧЕНИЯ ОСТРЫХ ОСЛОЖНЕНИЙ С ИСПОЛЬЗОВАНИЕМ ПОЛИКЛОНАЛЬНОГО ИММУНОГЛОБУЛИНА | 2019 |
|
RU2806443C2 |
| РАСПЫЛЕНИЕ ИММУНОГЛОБУЛИНА | 2015 |
|
RU2729546C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИКЛОНАЛЬНОЙ МОНОСПЕЦИФИЧЕСКОЙ СЫВОРОТКИ ПРОТИВ СЕКРЕТОРНОГО ИММУНОГЛОБУЛИНА А ЧЕЛОВЕКА - SIgA | 2012 |
|
RU2499605C1 |
| АКТИВАЦИЯ CD89 В ТЕРАПИИ | 2011 |
|
RU2640082C2 |
| ЛИГАНД, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩИЙ СТВОЛ РЕЦЕПТОРА ПОЛИМЕРНОГО ИММУНОГЛОБУЛИНА (pIgR) КЛЕТКИ, СПОСОБ ИНТРОДУКЦИИ ЛИГАНДА В КЛЕТКУ И СПОСОБ ПРИСОЕДИНЕНИЯ ЛИГАНДА К КЛЕТКЕ, ЭКСПРЕССИРУЮЩЕЙ РЕЦЕПТОР ПОЛИМЕРНОГО ИММУНОГЛОБУЛИНА | 1997 |
|
RU2191781C2 |
| СРЕДСТВО, СОДЕРЖАЩЕЕ ИММУНОГЛОБУЛИНЫ И ОБЛАДАЮЩЕЕ АНТИБАКТЕРИАЛЬНОЙ И АНТИВИРУСНОЙ АКТИВНОСТЬЮ | 2010 |
|
RU2453336C1 |
| Способ получения иммуноглобулинового препарата для наружного применения | 2015 |
|
RU2616266C1 |
| ХИМЕРНЫЙ ИММУНОГЛОБУЛИНОВЫЙ ПРЕПАРАТ, ОБЛАДАЮЩИЙ СПЕЦИФИЧЕСКИМ ПРОТИВОВИРУСНЫМ ИЛИ АНТИБАКТЕРИАЛЬНЫМ ДЕЙСТВИЕМ | 2015 |
|
RU2599029C1 |
| СПОСОБ ДОСТАВКИ БИОЛОГИЧЕСКИ АКТИВНОГО СОЕДИНЕНИЯ НА АССОЦИИРОВАННУЮ СО СЛИЗИСТОЙ ОБОЛОЧКОЙ ЛИМФОРЕТИКУЛЯРНУЮ ТКАНЬ ЖИВОТНОГО, СПОСОБ ПОЛУЧЕНИЯ КОМПОЗИЦИИ И КОМПОЗИЦИЯ ДЛЯ ДОСТАВКИ БИОЛОГИЧЕСКИ АКТИВНОГО СОЕДИНЕНИЯ | 1989 |
|
RU2250102C2 |




Изобретение относится к биохимии. Описан способ получения in vitro композиции, содержащей секреторноподобный иммуноглобулин, включающий стадии получения белковой композиции, выделенной из крови, содержащей иммуноглобулин, имеющий в составе J-цепь, в неочищенном виде, и смешивания композиции стадии (а) с секреторным компонентом. Представлена композиция для применения в медицине, содержащая секреторноподобный IgA и/или секреторноподобный IgM или их сочетание, полученная описанным способом. 2 н. и 15 з.п. ф-лы, 8 ил., 5 пр.
1. Способ получения in vitro композиции, содержащей секреторноподобный иммуноглобулин, включающий стадии
(a) получения белковой композиции, выделенной из крови, содержащей иммуноглобулин, имеющий в составе J-цепь, в неочищенном виде,
(b) смешивания композиции стадии (а) с секреторным компонентом.
2. Способ по п. 1, где секреторноподобный иммуноглобулин является секреторноподобным IgA и/или секреторноподобным IgM.
3. Способ по п. 1 или 2, где композиция стадии (а) содержит по меньшей мере 5% IgA, связанного J-цепью.
4. Способ по п. 3, где композиция стадии (а) содержит по меньшей мере 10% IgA, связанного J-цепью.
5. Способ по п. 3, где композиция стадии (а) содержит по меньшей мере 20% IgA, связанного J-цепью.
6. Способ по п. 3, где композиция стадии (а) содержит по меньшей мере 30% IgA, связанного J-цепью.
7. Способ по п. 3, где композиция стадии (а) содержит по меньшей мере 50% IgA, связанного J-цепью.
8. Способ по п. 1, где композицию стадии (а) получают из крови человека.
9. Способ по п. 1, где секреторный компонент на этапе (b) является рекомбинантным секреторным компонентом.
10. Способ по п. 1, где секреторный компонент является секреторным компонентом человека.
11. Способ по п. 1, где секреторный компонент получают в клеточной линии млекопитающего.
12. Способ по п. 1, где секреторный компонент является внеклеточной частью рецептора полимерного иммуноглобулина pIgR.
13. Способ по п. 1, где на этапе (b) молярное отношение между добавленным секреторным компонентом и J-цепью в димерах/полимерах IgA варьирует между 1:10 и 10:1.
14. Способ по п. 13, где соотношение варьирует приблизительно от 1:5 до 5:1.
15. Способ по п. 14, где соотношение варьирует приблизительно от 1:2 до 2:1.
16. Композиция для применения в медицине, содержащая секреторноподобный IgA и/или секреторноподобный IgM или их сочетание, получаемые посредством способа по любому из пп. 1-15.
17. Композиция по п. 16, дополнительно содержащая фармацевтически приемлемый носитель или эксципиент.
| Устройство для контроля проскаль-зыВАНия лЕНТы ТРАНСпОРТЕРА | 1979 |
|
SU839915A1 |
| Способ профилактики и лечения острых респираторных заболеваний вирусной этиологии | 1978 |
|
SU745523A1 |
| ARMELLE PHALIPON и др | |||
| Secretory component: a new role in secretory IgA-mediated immune exclusion in vivo, Immunity | |||
| v | |||
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| Счетный сектор | 1919 |
|
SU107A1 |

