Настоящее изобретение относится к способу получения сопряженных диенов, прежде всего, летучих диенов, с использованием биологического процесса. Более конкретно, настоящее изобретение относится к способу получения бутадиена, изопрена или диметилбутадиена из низших алкенолов с использованием ферментативной дегидратации, прежде всего, с использованием алкенол-дегидратазы, такой как линалоол-дегидратаза (ЕС 4.2.1.127).
Сопряженные диены, например 1,3-диены, такие как бутадиен или изопрен, являются важными промышленными соединениями. Изопрен (2-метил-1,3-бутадиен) представляет собой сопряженный диен, характеризующийся формулой C5H8. Он является основным соединением для шинной промышленности, а также широко применяется для получения адгезивов. Указанное соединение получают химическим способом с использованием нескольких методик:
- экстракционная дистилляция нефти (фракция C5),
- дегидратация изоамилена,
- двойная дегидратация изопентана,
- реакция изобутена с формальдегидом,
- реакция ацетона с ацетиленом,
- димеризация пропилена.
В заявке WO 2009/076676 описан метаболический путь синтеза изопрена. Указанный путь основан на дефосфорилировании-дегидратации переработанных промежуточных продуктов мевалонатного пути, т.е. изопренил-пирофосфата или пренил-пирофосфата. Недостатком указанного способа является необходимость прохождения всего мевалонатного пути, включающего двойное фосфорилирование мевалоната, последующее декарбоксилирование-дегидратацию с образованием изопренил-пирофосфата, дальнейшую изомеризацию с образованием пренил-пирофософата и, наконец, двойное дефосфорилирование/дегидратацию с образованием изопрена.
Бутадиен (1,3-бутадиен) представляет собой сопряженный диен, характеризующийся формулой C4H6. Он является важным промышленным химическим реагентом, используемым в качестве мономера при синтезе синтетического каучука, нейлона, акрилонитрил-бутадиен-стирола (АБС), пластиков, латекса. Существуют различные способы получения бутадиена. Бутадиен, например, получают в качестве побочного продукта при проведении парового крекинга, используемого для получения этилена и других олефинов. В указанном процессе бутадиен присутствует в потоке C4, и его обычно отделяют от других побочных продуктов экстракцией в апротонный полярный растворитель, такой как ацетонитрил, из которого его затем отгоняют. Бутадиен также можно получить каталитической дегидратацией нормального бутана, или его можно получить из этанола. В последнем случае используют два различных процесса. В ходе одностадийного процесса этанол превращают в бутадиен, водород и воду при нагревании при температуре 400-450°С над катализатором из оксида металла (Kirshenbaum I., Butadiene, энциклопедия «Encyclopedia of Chemical Technology», под ред. Grayson M., 3е изд., Нью-Йорк, Wiley & Sons, т. 4, сс. 313-337 (1978)). При проведении двухстадийного процесса этанол окисляют до ацетальдегида, который взаимодействует с дополнительным количеством этанола над пористым катализатором на основе кремнезема, активированного танталом, при температуре 325-350°С, при этом получают бутадиен (Kirshenbaum I. (1978), см. выше). Бутадиен также можно получить каталитической дегидратацией нормальных бутенов.
В течение двух последних десятилетий технологии генной инженерии обеспечили возможность модификации метаболизма микроорганизмов, и в связи с этим применение микроорганизмов для получения важных соединений, которые другим способом получают с низким выходом. За счет повышения эффективности распространенных в природе метаболических путей указанные технологии открывают новые пути для получения множества соединений промышленного значения биологическим способом. Некоторые промышленные соединения, такие как аминокислоты для корма животных, биодеградабельные пластики или текстильные волокна, в настоящее время обычно получают с использованием генетически модифицированных микроорганизмов.
До сих пор существует необходимость обеспечения экологически безопасных, экономически эффективных и простых способов получения указанных выше соединений.
Настоящее изобретение направлено на разработку способа решения указанной проблемы, который описан в вариантах осуществления настоящего изобретения, а также в пунктах формулы изобретения.
Настоящее изобретение основано на разработке нового биокатализатора для синтеза летучих диеновых соединений, прежде всего, сопряженных диенов, таких как 1,3-диены, и основано на превращении низших алкенолов, прежде всего, ферментативной дегидратацией низших алкенолов. В основе настоящего изобретения лежит демонстрация возможности проведения указанного превращения биологическим способом с использованием фермента, катализирующего реакцию дегидратации. Настоящее изобретение можно осуществлять in vitro, в системах, не содержащих клеток, или с использованием организмов, прежде всего микроорганизмов. Настоящее изобретение также относится к получению сопряженных диенов, таких как 1,3-диены, из углеродного источника, прежде всего из углевода (прежде всего, глюкозы), полиола (прежде всего, глицерина), биодеградабельного полимера (прежде всего, крахмала, целлюлозы, поли-3-гидроксиалкеноата), при этом в присутствии микроорганизма углеродный источник превращается в низший алкенол, который затем превращается в сопряженный диен, такой как 1,3-диен.
Более конкретно, настоящее изобретение относится к способу получения сопряженного диена, отличающегося тем, что указанный способ включает стадию ферментативного превращения соединения общей формулы CnH2nO в соединение общей формулы CnH2n-2+H2O, где 3<n<7, с использованием алкенол-дегидратазы. Указанное превращение представляет собой дегидратацию.
Соединение формулы CnH2nO, где 3<n<7, в контексте настоящего изобретения называется низшим алкенолом.
В одном предпочтительном варианте осуществления настоящего изобретения индекс п равен 4. В этом случае низший алкенол, который используют в качестве субстрата в способе по настоящему изобретению, характеризуется формулой C4H8O. Соединениями, которые характеризуются указанной формулой, являются бут-2-ен-1-ол (кротиловый спирт), бут-3-ен-2-ол и бут-3-ен-1-ол (изокротиловый спирт). Диеном, который получают в результате превращения указанных соединений способом по настоящему изобретению, является бутадиен. В прежде всего предпочтительном варианте осуществления настоящего изобретения низшим алкенолом, который используют в качестве субстрата в способе по настоящему изобретению, является бут-2-ен-1-ол (кротиловый спирт) или бут-3-ен-2-ол, а получаемым диеном является бутадиен.
В другом предпочтительном варианте индекс n равен 5. В этом случае низший алкенол, который используют в качестве субстрата в способе по настоящему изобретению, характеризуется формулой C5H10O. Соединениями, которые характеризуются указанной формулой, являются 2-метилбут-2-ен-1-ол, 3-метилбут-2-ен-1-ол (пренол), 3-метилбут-3-ен-2-ол, 2-метилбут-3-ен-2-ол, 2-метилбут-3-ен-1-ол и 3-метилбут-3-ен-1-ол (изопренол). Диеном, который получают в результате превращения указанных соединений способом по настоящему изобретению, является изопрен. В более предпочтительном варианте низшим алкенолом, который используют в качестве субстрата в способе по настоящему изобретению, является 2-метилбут-2-ен-1-ол, 3-метилбут-2-ен-1-ол (пренол), 3-метилбут-3-ен-2-ол, 2-метилбут-3-ен-2-ол или 3-метилбут-3-ен-1-ол (изопренол), а получаемым диеном является изопрен. В еще более предпочтительном варианте низшим алкенолом, который используют в качестве субстрата в способе по настоящему изобретению, является 3-метилбут-2-ен-1-ол (пренол), 3-метилбут-3-ен-2-ол, 2-метилбут-3-ен-2-ол или 3-метилбут-3-ен-1-ол (изопренол), а получаемым диеном является изопрен. Прежде всего, в предпочтительном варианте низшим алкенолом, который используют в качестве субстрата в способе по настоящему изобретению, является 3-метилбут-2-ен-1-ол (пренол) или 2-метилбут-3-ен-2-ол, а получаемым диеном является изопрен.
В еще одном предпочтительном варианте индекс n равен 6. В этом случае низший алкенол, который используют в качестве субстрата в способе по настоящему изобретению, характеризуется формулой С6Н12О. Соединениями, которые характеризуются указанной формулой, являются 2,3-диметилбут-2-ен-1-ол, 2,3-диметилбут-3-ен-2-ол и 2,3-диметилбут-3-ен-1-ол. Диеном, который получают в результате превращения указанных соединений способом по настоящему изобретению, является диметилбутадиен. В прежде всего предпочтительном варианте низшим алкенолом, который используют в качестве субстрата в способе по настоящему изобретению, является 2,3-диметилбут-2-ен-1-ол или 2,3-диметилбут-2-ен-1-ол, а получаемым диеном является диметилбутадиен.
Соединения общей формулы CnH2nO, где 3<n<7, можно подразделить на три группы, а именно:
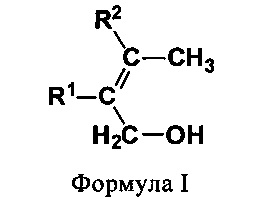
(I) первичные аллиловые спирты (ПРА) формулы I:
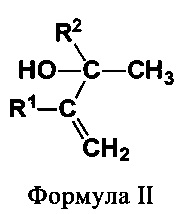
(II) вторичные или третичные аллиловые спирты (ВТА) формулы II:
и
(III) первичные гомоалиловые спирты (ПГА) формулы III:

где R1 и R2 независимо выбирают из Н и CH3.
В одном предпочтительном варианте соединением общей формулы CnH2nO, где 3<n<7, является первичный аллиловый спирт (ПРА) формулы I:

где R1 и R2 независимо выбирают из Н и СН3. Соединениями, характеризующимися указанной формулой, являются бут-2-ен-1-ол (кротиловый спирт), 2-метилбут-2-ен-1-ол, 3-метилбут-2-ен-1-ол (пренол) и 2,3-диметилбут-2-ен-1-ол (см. фиг. 1). В одном предпочтительном варианте первичным аллиловым спиртом является бут-2-ен-1-ол (кротиловый спирт), а получаемым диеном является бутадиен. В другом предпочтительном варианте первичным аллиловым спиртом является 3-метилбут-2-ен-1-ол (пренол), а получаемым диеном является изопрен.
В еще одном предпочтительном варианте соединением общей формулы CnH2nO, где 3<n<7, является вторичный или третичный аллиловый спирт (ВТА) формулы II:
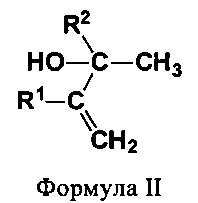
где R1 и R2 независимо выбирают из Н и СН3. Соединениями, характеризующимися указанной формулой, являются бут-3-ен-2-ол, 3-метилбут-3-ен-2-ол, 2-метилбут-3-ен-2-ол и 2,3-диметилбут-3-ен-2-ол (см. фиг. 2).
В одном предпочтительном варианте ВТА является бут-3-ен-2-ол, а получаемым диеном является бутадиен. В другом предпочтительном варианте ВТА является 2-метилбут-3-ен-2-ол, а получаемым диеном является изопрен.
В еще одном предпочтительном варианте соединением общей формулы CnH2nO, где 3<n<7, является первичный гомоаллиловый спирт (ПГА) формулы III:
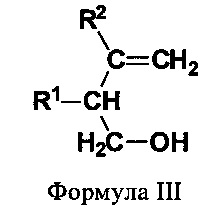
где R1 и R2 независимо выбирают из Н и СН3. Соединениями, характеризующимися указанной формулой, являются бут-3-ен-1-ол (изокротиловый спирт), 2-метилбут-3-ен-1-ол, 3-метилбут-3-ен-1-ол (изопренол) и 2,3-диметилбут-3-ен-1-ол (см. фиг.3).
В одном предпочтительном варианте гомоаллиловым спиртом является 3-метилбут-3-ен-1-ол (изопренол), а получаемым диеном является изопрен.
На фиг. 4 представлена схема превращения упомянутых выше соединений ПРА, ПГА и ВТА в сопряженный диен способом по настоящему изобретению.
Если в контексте настоящего изобретения упоминается соединение, которое существует в форме стереоизомеров, например, за счет Z/E-изомеризации в положении двойных связей атомов C=C (sp2-гибридизация), или за счет R/S-изомеризации в положении хиральных центров атомов углерода (sp3-гибридизация), ссылка на указанное соединение включает все указанные стереоизомеры. Например, при упоминании бут-2-ен-1-ола (кротилового спирта) включены его цис- (Z), а также транс- (Е) стереоизомеры. Аналогичным образом, при упоминании 2-метилбут-2-ен-1-ола включены его цис- (Z), а также транс- (Е) стереоизомеры. Кроме того, при упоминании бут-3-ен-2-ола, 3-метилбут-3-ен-2-ола, 2-метилбут-3-ен-1-ола или 2,3-диметилбут-3-ен-1-ола включены оба R- и S-изомеры указанных соединений.
В одном предпочтительном варианте ссылка на бут-2-ен-1-ол (кротиловый спирт) включает цис-(Z) стереоизомер. В другом предпочтительном варианте ссылка на бут-2-ен-1-ол (кротиловый спирт) включает транс-(Е) стереоизомер. В еще одном предпочтительном варианте ссылка на бут-2-ен-1-ол (кротиловый спирт) включает смесь, содержащую оба стереоизомера. Если любое из указанных соединений используют в качестве субстрата в способе по настоящему изобретению, продуктом является бутадиен.
В одном предпочтительном варианте ссылка на 2-метилбут-2-ен-1-ол включает цис-(Z) стереоизомер. В другом предпочтительном варианте ссылка на 2-метилбут-2-ен-1-ол включает транс-(Е) стереоизомер. В еще одном предпочтительном варианте ссылка на 2-метилбут-2-ен-1-ол включает смесь, содержащую оба стереоизомера. Если любое из указанных соединений используют в качестве субстрата в способе по настоящему изобретению, продуктом является изопрен.
В одном предпочтительном варианте ссылка на бут-3-ен-2-ол включает R-изомер. В другом предпочтительном варианте ссылка на бут-3-ен-2-ол включает S-изомер. В еще одном предпочтительном варианте ссылка на бут-3-ен-2-ол включает смесь, содержащую оба изомера. Если любое из указанных соединений используют в качестве субстрата в способе по настоящему изобретению, продуктом является бутадиен.
В одном предпочтительном варианте ссылка на 3-метилбут-3-ен-2-ол включает R-изомер. В другом предпочтительном варианте ссылка на 3-метилбут-3-ен-2-ол включает S-изомер. В еще одном предпочтительном варианте ссылка на 3-метилбут-3-ен-2-ол включает смесь, содержащую оба изомера. Если любое из указанных соединений используют в качестве субстрата в способе по настоящему изобретению, продуктом является изопрен.
В одном предпочтительном варианте ссылка на 2-метилбут-3-ен-1-ол включает R-изомер. В другом предпочтительном варианте ссылка на 2-метилбут-3-ен-1-ол включает S-изомер. В еще одном предпочтительном варианте ссылка на 2-метилбут-3-ен-1-ол включает смесь, содержащую оба изомера. Если любое из указанных соединений используют в качестве субстрата в способе по настоящему изобретению, продуктом является изопрен.
В одном предпочтительном варианте ссылка на 2,3-диметилбут-3-ен-1-ол включает R-изомер. В другом предпочтительном варианте ссылка на 2,3-диметилбут-3-ен-1-ол включает S-изомер. В еще одном предпочтительном варианте ссылка на 2,3-диметилбут-3-ен-1-ол включает смесь, содержащую оба изомера. Если любое из указанных соединений используют в качестве субстрата в способе по настоящему изобретению, продуктом является диметилбутадиен.
Как описано выше, способ по настоящему изобретению заключается в том, что превращение соединения общей формулы CnH2nO, в соединение общей формулы CnH2n-2+H2O, где 3<n<7, достигается при использовании алкенол-дегидратазы. Алкенол-дегидратаза представляет собой фермент, который может катализировать дегидратацию алкенола, предпочтительно фермент, который может катализировать дегидратацию, по крайней мере, одного соединения общей формулы CnH2nO, где 3<n<7, и при этом продуктом реакции является соединение общей формулы CnH2n-2+Н2О. Указанную активность можно определить методами анализа, описанными в прилагаемых примерах. Пример алкенол-дегидратазы, которую используют в способе по настоящему изобретению, включает алкенол-дегидратазу, которая была названа "линалоол-дегидратаза-изомераза" (ЕС 4.2.1.127), и которая была идентифицирована в штамме 65 Phen Castellaniella defragrans (ранее Alcaligenes defragrans) (Brodkorb и др., J. Biol. Chem., 285, cc. 30436-30442 (2010)). Линалоол-дегидратаза-изомераза представляет собой бифункциональный фермент, который принимает участие в анаэробной деградации монотерпенов. Было установлено, что нативный фермент характеризуется молекулярной массой 160 кДа, и, как полагают, представляет собой гомотетрамер, состоящий из субъединиц с молекулярной массой 40 кДа. Указанный фермент катализирует in vitro две реакции в обоих направлениях в зависимости от термодинамических движущих сил. С одной стороны, фермент катализирует изомеризацию первичного аллилового спирта - гераниола, с образованием его стереоизомера - линалоола, который включает фрагмент третичного аллилового спирта. С другой стороны, фермент катализирует отщепление воды (дегидратацию) от третичного спирта - линалоола, при этом образуется соответствующий ациклический монотерпен - β-мирцен, молекула, включающая фрагмент сопряженного диена. На фиг. 5 представлены реакции, катализируемые in vitro линалоол-дегидратазой-изомеразой в анаэробных условиях. Бактерия Castellaniella defragrans экспрессирует белок в виде белка-предшественника, содержащего один сигнальный пептид, локализованный в периплазме, и который отщепляется после переноса через мембрану. Классификационный номер этого фермента - ЕС 4.2.1.127. Линалоол-дегидратаза-изомераза способна катализировать следующую реакцию в анаэробных условиях:
Линалоол <=> мирцен + Н2О
Метод определения указанной активности включает, например, метод анализа, описанный в статье Brodkorb и др. (цитированной выше). Этот анализ проводят следующим образом: флаконы предварительно нагревают до 35°С, в них переносят раствор аноскигенного белка и добавляют дитиотреит (ДТТ) до концентрации 2 мМ. Реакционные смеси герметично закрывают мембраной из бутилкаучука и пространство над раствором продувают смесью CO2/N2 (10:90 (об./об.)). Реакцию инициируют при добавлении линалоола до определенной концентрации и смесь инкубируют при 35°С. Превращение линалоола в мирцен оценивают по образованию мирцена, например, методом газовой хроматографии.
В предпочтительном варианте линалоол-дегидратаза-изомераза также способна катализировать изомеризацию гераниола с образованием линалоола в анаэробных условиях:
Гераниол <=> линалоол
Метод определения указанной активности включает, например, метод анализа, описанный в статье Brodkorb и др. (цитированной выше). Анализ проводят следующим образом: флаконы предварительно нагревают при 35°С, в них переносят раствор аноскигенного белка и добавляют ДТТ до концентрации 2 мМ. Реакционные смеси герметично закрывают мембраной из бутилкаучука и пространство над раствором продувают смесью CO2/N2 (10:90 (об./об.)). Реакцию инициируют при добавлении гераниола до определенной концентрации и смесь инкубируют при 35°С. Превращение гераниола в линалоол оценивают по образованию мирцена, т.е. продукта второй реакции, катализируемой ферментом, например, методом газовой хроматографии.
Гераниол, линалоол и мирцен являются продуцируемыми растениями ациклическими С10-терпеноидами, принадлежащими к классу аллиловых спиртов и углеводородов, соответственно. В статье  и Harder (Z. Naturforsch., 66 с, сс. 409-412 (2011)) описана высокая субстратная специфичность линалоол-дегидратазы-изомеразы. Авторами настоящего изобретения неожиданно было установлено, что линалоол-дегидратаза-изомераза может действовать на соединения формулы CnH2nO, где 3<n<7, и превращать их в сопряженные диены. В прилагаемых примерах указанное превращение описано на примере превращения бут-2-ен-1-ола (кротилового спирта) в бутадиен, бут-3-ен-2-ола в бутадиен, 3-метилбут-2-ен-1-ола (пренола) в изопрен, 3-метилбут-3-ен-1-ола (изопренола) в изопрен и 2-метилбут-3-ен-2-ола в изопрен. Таким образом, авторами настоящего изобретения установлено, что линалоол-дегидратаза-изомераза неожиданно способна также превращать алкенолы, которые характеризуются значительно меньшей длиной по сравнению с природными субстратами линалоол-дегидратазы-изомеразы несмотря на известную высокую субстратную специфичность указанного фермента. Таким образом, в одном варианте, алкенол-дегидратазой, используемой в способе по настоящему изобретению, является линалоол-дегидратаза (ЕС 4.2.1.127).
и Harder (Z. Naturforsch., 66 с, сс. 409-412 (2011)) описана высокая субстратная специфичность линалоол-дегидратазы-изомеразы. Авторами настоящего изобретения неожиданно было установлено, что линалоол-дегидратаза-изомераза может действовать на соединения формулы CnH2nO, где 3<n<7, и превращать их в сопряженные диены. В прилагаемых примерах указанное превращение описано на примере превращения бут-2-ен-1-ола (кротилового спирта) в бутадиен, бут-3-ен-2-ола в бутадиен, 3-метилбут-2-ен-1-ола (пренола) в изопрен, 3-метилбут-3-ен-1-ола (изопренола) в изопрен и 2-метилбут-3-ен-2-ола в изопрен. Таким образом, авторами настоящего изобретения установлено, что линалоол-дегидратаза-изомераза неожиданно способна также превращать алкенолы, которые характеризуются значительно меньшей длиной по сравнению с природными субстратами линалоол-дегидратазы-изомеразы несмотря на известную высокую субстратную специфичность указанного фермента. Таким образом, в одном варианте, алкенол-дегидратазой, используемой в способе по настоящему изобретению, является линалоол-дегидратаза (ЕС 4.2.1.127).
Пример последовательности алкенол-дегидратазы, которую можно использовать в способе по настоящему изобретению, включает последовательность SEQ ID NO: 1 (фиг. 6). Последовательность алкенол-дегидратазы также доступна в базе данных UniProtKB/TrEMBL под номером доступа E1XUJ2. Указанные последовательности представляют алкенол-дегидратазу, которую классифицируют как линалоол-дегидратазу-изомеразу. В предпочтительном варианте осуществления способа по настоящему изобретению используют алкенол-дегидратазу, включающую аминокислотную последовательность SEQ ID NO: 1, или последовательность, идентичность которой с последовательностью SEQ ID NO: 1 составляет, по крайней мере, x%, и которая способна катализировать превращение соединения общей формулы CnH2nO в соединение общей формулы CnH2n-2+Н2О, где 3<n<7, при этом x равен целому числу от 30 до 100, предпочтительно 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99.
В связи с этим использованный в данном контексте термин "алкенол-дегидратаза" относится к ферменту, степень идентичности последовательности которого с последовательностью SEQ ID NO:1 составляет указанную выше величину, и который способен катализировать превращение соединения общей формулы CnH2nO в соединение общей формулы CnH2n-2+H2O, где 3<n<7. Специалист в данной области химии может с использованием последовательности SEQ ID NO:1 или соответствующих кодирующих нуклеотидных последовательностей идентифицировать другие алкенол-дегидратазы, которые способны катализировать указанное выше превращение.
Указанные варианты алкенол-дегидратазы, например, линалоол-дегидратазы (ЕС 4.2.1.127), также включают укороченные варианты с делениями в N- или С-концевых фрагментах, предпочтительно в С-концевом фрагменте, как описано ниже. В прилагаемых примерах показано, что такие укороченные варианты сохраняют способность катализировать описанные выше превращения.
Предпочтительно, степень идентичности определяют при сравнении соответствующей последовательности с аминокислотной последовательностью SEQ ID NO:1. Если последовательности, которые сравнивают, характеризуются различной длиной, степень идентичности предпочтительно относится либо к проценту аминокислотных остатков в более короткой последовательности, которые идентичны аминокислотным остаткам в более длинной последовательности, или к проценту аминокислотных остатков в более длинной последовательности, которые идентичны аминокислотным остаткам в более короткой последовательности. Степень идентичности можно определить способами, известными в данной области химии, предпочтительно с использованием пригодных компьютерных алгоритмов, таких как CLUSTAL.
При использовании метода Clustal при сравнении аминокислотных последовательностей для определения степени идентичности конкретной последовательности с последовательностью сравнения, например, равной 80%, можно использовать параметры по умолчанию или, предпочтительно, задать следующие параметры: Matrix: blosum 30 (матрица блоков аминокислотных замен), Open gap penalty (штраф за внесение делеции): 10,0, Extend gap penalty (штраф за продолжение делеций): 0,05, Delay divergent (отсрочка различающихся последовательностей): 40, Gap separation distance (пространственное разнесение): 8. Для сравнения нуклеотидных последовательностей параметр Extend gap penalty предпочтительно устанавливают равным 5,0.
Предпочтительно, степень идентичности рассчитывают по всей длине последовательности.
Более того, если в контексте настоящего изобретения используют термин "гомология", указанный термин предпочтительно обозначает "идентичность последовательности".
Как описано выше, алкенол-дегидратаза, которая известна как "линалоол-дегидратаза-изомераза", идентифицированная в бактерии Castellaniella defragrans (ранее Alcaligenes defragrans), включает сигнальный пептид, обеспечивающий перенос в периплазму. В предпочтительном варианте в способе по настоящему изобретению используют фермент, который не включает такую сигнальную последовательность. В разделе Примеры показано, что удаление сигнального пептида при включении гистидиновой метки не влияет на экспрессию фермента в Е. coli и вызывает внутриклеточное продуцирование активного белка.
Алкенол-дегидратазой, такой как линалоол-дегидратаза-изомераза, используемой в способе по настоящему изобретению, является нативная алкенол-дегидратаза или алкенол-дегидратаза, которую получают из нативной алкенол-дегидратазы, такой как линалоол-дегидратаза-изомераза, например, при введении мутаций или других изменений, которые, например, изменяют или повышают ферментативную активность, стабильность, прежде всего термическую стабильность и т.п.
В прилагаемых примерах также показано, что укороченные варианты линалоол-дегидратазы (ЕС 4.2.1.127), предпочтительно укороченные варианты, которые характеризуются делециями в С-концевом фрагменте, способны эффективно катализировать описанные выше превращения. Таким образом, термин "алкенол-дегидратаза" также включает ферменты, которые получают из алкенол-дегидратазы, такой как линалоол-дегидратаза (ЕС 4.2.1.127), за счет делеций, прежде всего, делеций в С-концевом фрагменте. Более предпочтительно, такой белок представляет собой фермент, который получают из фермента с аминокислотной последовательностью SEQ ID NO:1, за счет делеции, по крайней мере, аминокислотных остатков 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 в С-концевом фрагменте.
Термин "линалоол-дегидратаза-изомераза" или "белок/фермент, обладающий активностью линалоол-дегидратазы-изомеразы" в контексте настоящего изобретения также включает ферменты, которые получают из линалоол-дегидратазы-изомеразы, которые способны катализировать превращение соединения общей формулы CnH2nO в соединение общей формулы CnH2n-2+H2O, где 3<n<7, но которые характеризуются лишь низкой аффинностью к своим природным субстратам, т.е. гераниолу, линалоолу и/или мирцену, или которые уже не связываются со своими природными субстратами. Такая модификация предпочтительного субстрата позволяет повысить эффективность превращения соединения общей формулы CnH2nO в соединение общей формулы CnH2n-2+H2O, где 3<n<7, а также снизить продуцирование возможных нежелательных побочных продуктов. Способы модификации и/или повышения требуемых ферментативных активностей белков известны специалистам в данной области химии и включают, например, неспецифический мутагенез или сайт-направленный мутагенез и последующую селекцию ферментов, обладающих требуемыми свойствами, или подходы так называемой "направленной эволюции", шаффлинга ДНК или эволюции in vivo.
Например, для генной инженерии в прокариотических клетках молекулу нуклеиновой кислоты, кодирующую линалоол-дегидратазу-изомеразу, можно включить в плазмиды, для которых разрешен мутагенез или модификация последовательностей при рекомбинации последовательностей ДНК. Стандартные способы (см. учебник Sambrook и Russell, Molecular Cloning: А Laboratory Manual, CSH Press, Cold Spring Harbor, Нью-Йорк, США (2001)) позволяют осуществлять замены оснований или включать природные или синтетические последовательности. Фрагменты ДНК можно присоединять один к другому с применением адаптеров или линкеров для фрагментов. Более того, можно использовать генно-инженерные способы, которые обеспечивают пригодные сайты рестрикции или удаляют избыточную ДНК или сайты рестрикции. В таких случаях, когда возможны вставки, делеции или замены, можно использовать мутагенез in vitro, "репарацию праймеров", рестрикцию или легирование. В основном, в качестве аналитических методов используют секвенирование, рестрикционный анализ и другие методы биохимии и молекулярной биологии. Затем определяют ферментативную активность полученных в результате вариантов линалоол-дегидратазы-изомеразы и, прежде всего, их способность предпочтительно использовать в качестве субстрата соединение общей формулы CnH2nO, где 3<n<7, по сравнению, например, с гераниолом, линалоолом и/или мирценом.
Указанные методы идентификации вариантов с улучшенными ферментативными свойствами в отношении продуцирования сопряженного диенового соединения можно также осуществлять в присутствии кофактора, который обеспечивает стерическую и/или электронную комплементарность в каталитическом участке фермента за счет более короткого соединения общей формулы CnH2nO, где 3<n<7, по сравнению с более длинными природными субстратами.
В предпочтительном варианте алкенол-дегидратаза, которую используют в способе по настоящему изобретению, характеризуется высокой термической стабильностью. Такой фермент можно получить стандартными способами, включая, например, неспецифическую мутацию нуклеотидной последовательности, кодирующей алкенол-дегидратазу, и скрининг полученных мутантных вариантов в отношении более высокой термической стабильности. Предпочтительно, алкенол-дегидратаза является стабильной и ферментативно активной при температуре 68°С или выше. В связи с тем что температура кипения диметилбутадиена при атмосферном давлении составляет 68°С, использование указанного фермента и осуществление способа по настоящему изобретению при температуре 68°С или выше представляет преимущество, т.к. диметилбутадиен при этом испаряется из реакционной смеси, и его можно легко извлекать из газообразной фазы.
Модифицированный вариант алкенол-дегидратазы, например вариант, связывающийся или предпочтительно связывающийся с соединением общей формулы CnH2nO, где 3<n<7, в качестве субстрата, но характеризующийся низкой аффинностью к своим природным субстратам или не связывающийся со своими природными субстратами, или вариант, характеризующийся повышенной термической стабильностью, можно получить из природной алкенол-дегидратазы, такой как линалоол-дегидратаза-изомераза, или из уже модифицированной, оптимизированной или полученной синтетическим способом алкенол-дегидратазы.
Способ по настоящему изобретению можно проводить in vitro, например, в присутствии выделенного фермента или клеточных лизатов, содержащих фермент, или частично очищенных ферментных препаратов. Термин «in vitro» предпочтительно обозначает систему, не содержащую клеток.
В одном варианте фермент, используемый в способе, применяют в очищенной форме, при этом соединение общей формулы CnH2nO превращают в соединение общей формулы CnH2n-2+H2O, где 3<n<7. Однако такой способ может оказаться дорогостоящим в связи с высокой себестоимостью получения фермента и субстрата.
В связи с этим в другом предпочтительном варианте ферменты, используемые в способе, присутствуют в реакционной смеси в виде неочищенного экстракта, или, в другом варианте, в форме нелизированных бактерий, при этом можно экономить на затратах, связанных с очисткой белков. Однако, затраты, связанные с указанным способом, все еще могут оставаться достаточно высокими из-за себестоимости получения и очистки субстратов.
При проведении реакции in vitro ферменты, нативные или рекомбинантные, очищенные или не очищенные, инкубируют в присутствии субстрата в физико-химических условиях, обеспечивающих активное состояние ферментов, и инкубацию проводят в течение достаточного периода времени, чтобы обеспечить продуцирование диена. При завершении инкубации необязательно определяют присутствие диенового соединения с использованием любой системы детектирования, известной специалисту в данной области химии, такой как газовая хроматография или колориметрические методы анализа для определения образования указанных соединений.
Прежде всего, в предпочтительном варианте осуществления настоящего изобретения способ проводят in vitro, а фермент иммобилизуют. Специалисту в данной области химии известны способы иммобилизации ферментов на различных подложках.
В другом предпочтительном варианте способ по настоящему изобретению осуществляют в клеточной культуре в присутствии организма, предпочтительно микроорганизма, продуцирующего фермент. Таким образом, в указанном варианте осуществления настоящего изобретения используют организм, предпочтительно микроорганизм, который продуцирует алкенол-дегидратазу, такую как линалоол-дегидратаза-изомераза. В предпочтительном варианте (микро)организм является рекомбинантным, при этом организм хозяина продуцирует фермент, который является гетерологичным относительно хозяина-продуцента. В связи с этим способ можно осуществлять непосредственно в культуральной среде, без необходимости отделения или очистки ферментов. В прежде всего предпочтительном варианте используют (микро)организм, характеризующийся природным или искусственно приобретенным свойством эндогенно продуцировать соединение общей формулы CnH2nO, где 3<n<7, а также экспрессирующий или сверхэкспрессирующий алкенол-дегидратазу, такую как линалоол-дегидратаза-изомераза, природная или модифицированная, и таким образом продуцирующий диеновое соединение напрямую из углеродного источника, присутствующего в растворе.
Например, способ по настоящему изобретению можно осуществлять с использованием микроорганизмов, которые продуцируют соединение общей формулы CnH2nO, где 3<n<7. Например, в статье Perez и др., Phytochemistry, 19, сс. 183-187 (1980) описаны ферменты из апельсинового дерева Citrus sinensis, которые способны гидролизовать аллилфосфаты, например, пренилдифосфотаза (ЕС 3.1.7.1), которая способна превращать пренолдифосфат в пренол и дифосфат. Последовательности нуклеиновых кислот, кодирующие указанные ферменты, можно вводить в микроорганизмы, которые продуцируют соответствующий субстрат, и таким образом способны продуцировать пренол. Кроме того, в статье Withers и др., Appl. Environ. Microbiol. 73, сс. 6277-6283 (2007) описаны, например, клетки Е. coli, которые преобразованы генно-инженерным способом по изопентилпирофосфатному биосинтетическому пути на основе мевалоната, и которые также экспрессируют ген nudF штамма Bacillus subtilis 6,051. Белок, кодируемый геном nudF, оказывает непосредственное действие на предшественники пренилдифосфата и вызывает продуцирование изопентенола (изопренола).
В связи с этим в одном предпочтительном варианте осуществления способа по настоящему изобретению предпочтительно используют микроорганизм, который способен продуцировать соединение общей формулы CnH2nO, где 3<n<7, и который преобразован генно-инженерным способом таким образом, что указанный микроорганизм сверхэкспрессирует алкенол-дегидратазу, причем указанная алкенол-дегидратаза предпочтительно получена из организма, отличающегося от микроорганизма-хозяина. Генетическая модификация может включать, например, вставку соответствующего гена, кодирующего алкенол-дегидратазу, в хромосому, экспрессирующую фермент из плазмиды, содержащей расположенный против хода транскрипции промотор последовательности, кодирующей фермент, причем промотор и кодирующую последовательность предпочтительно получают из различных организмов, или любой другой способ, известный специалисту в данной области химии. В другом варианте, можно выбрать другие бактерии или дрожжи, характеризующиеся специфическими преимуществами. Например, можно использовать дрожжи, такие как Saccharomyces cerevisiae, экстремофильную бактерию, такую как Thermus thermophilus, или анаэробные бактерии из семейства Clostridiae, микроводоросли или фотосинтетические бактерии.
Можно также выделять гены, кодирующие белки, которые ответственны за синтез соединения общей формулы CnH2nO, где 3<n<7, и вводить указанные гены в другие организмы, прежде всего, в микроорганизм, такой как, например, Е. coli, Saccharomyces или Pichia, экстремофильную бактерию, такую как Thermus thermophilus, или в анаэробные бактерии из семейства Clostridiae, микроводоросли или фотосинтетические бактерии.
В предпочтительном варианте (микро)организмом, используемым в способе по настоящему изобретению, является (микро)организм, который генетически модифицирован, и содержит молекулу нуклеиновой кислоты, кодирующую алкенол-дегидратазу. Указанную молекулу нуклеиновой кислоты, кодирующую алкенол-дегидратазу, как описано выше, можно использовать в отдельности или в качестве части вектора. Молекулы нуклеиновых кислот дополнительно включают последовательности, контролирующие экспрессию, функционально связанные с полинуклеотидом, включенным в молекулу нуклеиновой кислоты. Использованный в данном контексте термин "функционально связанный" обозначает линкер, расположенный между одной или более последовательностями, контролирующими экспрессию, и кодирующей областью в полинуклеотиде, подлежащем экспрессии таким образом, чтобы экспрессия происходила в условиях, совместимых с последовательностью, контролирующей экспрессию.
Экспрессия включает транскрипцию гетерологичной последовательности ДНК, предпочтительно, в транслируемую мРНК. Регуляторные элементы, обеспечивающие экспрессию в грибах, а также в бактериях, известны специалистам в данной области химии. Они включают промоторы, энхансеры, стоп-кодоны, направленные кодоны и т.п. Примеры приведены ниже в связи с разъяснениями, касающимися векторов.
Промоторы, предназначенные для применения в сочетании с молекулой нуклеиновой кислоты, являются гомологичными или гетерологичными в отношении их происхождения и/или в отношении гена, подлежащего экспрессии. Пригодными промоторами являются, например, промоторы, которые сами по себе пригодны для конститутивной экспрессии. Однако можно также использовать промоторы, которые активируются только во время, определяемое внешними факторами. В данном контексте можно также использовать искусственно и/или химически индуцируемые промоторы.
Векторы включают, кроме того, последовательности, контролирующие экспрессию, функционально связанные с указанными полинуклеотидами, содержащимися в векторах. Указанные последовательности, контролирующие экспрессию, можно модифицировать таким образом, чтобы обеспечить транскрипцию и синтез транслируемой РНК в бактериях или грибах.
Кроме того, можно вводить различные мутации в полинуклеотиды способами, стандартно используемыми в молекулярной биологии (см., например, учебник Sambrook and Russell, Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, Нью-Йорк, США (2001)), что приводит к синтезу полипептидов и к возможной модифицикации биологических свойств. Точечные мутации можно вводить в те участки, где модификация аминокислотной последовательности оказывает влияние, например, на биологическую активность или регуляцию полипептида.
Более того, можно получить мутантные формы, включающие модифицированный субстрат или обладающие модифицированной специфичностью продукта. Предпочтительно, такие мутантные формы характеризуются повышенной активностью. Кроме того, вставка мутантных вариантов в полинуклеотиды, кодирующие фермент, как определено выше, обеспечивает оптимизацию степени экспрессии генов и/или активности ферментов, кодируемых указанными полинуклеотидами, например, в отношении термической стабильности.
В случае генетически модифицированных бактерий или грибов, полинуклеотиды, кодирующие фермент, как определено выше, или фрагменты указанных молекул можно вставлять в плазмиды, которые допускают мутагенез или модификацию последовательности при рекомбинации последовательностей ДНК. Стандартные способы (см. учебник Sambrook и Russell, Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, Нью-Йорк, США (2001)) позволяют проводить замены оснований или добавлять природные или синтетические последовательности. Фрагменты ДНК можно соединять один с другим с использованием адаптеров и линкеров к фрагментам. Более того, можно использовать генно-инженерные способы, которые обеспечивают пригодные участки рестрикции или удаление избыточной ДНК или участков рестрикции. В указанных случаях, когда возможны вставки, делеции или замещения, можно использовать мутагенез in vitro, "репарацию праймера", рестрикцию или лигирование. В основном, в качестве методов анализа используют секвенирование, анализ рестрикции и другие методы, известные в биохимии и молекулярной биологии.
Экспрессия полинуклеотида, введенного в (микро)организм, происходит таким образом, что при этом вызывает продуцирование полипептида, обладающего описанной выше активностью. Обзор различных экспрессионных систем приведен, например, в книге Methods in Enzymology, 153, сс. 385-516 (1987), в главе Bitter и др., Methods in Enzymology, 153, сс. 516-544 (1987), а также в статьях Sawers и др., Applied Microbiology and Biotechnology, 46, сс. 1-9 (1996), Billman-Jacobe, Current Opinion in Biotechnology, 7, 500-504 (1996), Hockney, Trends in Biotechnology, 12, cc. 456-463 (1994), Griffiths и др., Methods in Molecular Biology, 75, cc. 427-440 (1997). Обзор экспрессионных систем на основе дрожжей приведен, например, в статьях Hensing и др., Antonie van Leuwenhoek, 67, сс. 261-279 (1995), Bussineau и др., Developments in Biological Standardization, 83, cc. 13-19 (1994), Gellissen и др., Antonie van Leuwenhoek, 62, cc. 79-93 (1992), Fleer, Current Opinion in Biotechnology, 3, cc. 486-496 (1992), Vedvick, Current Opinion in Biotechnology, 2, cc. 742-745 (1991) и Buckholz, Bio/Technology, 9, cc. 1067-1072 (1991).
Векторы экспрессии широко описаны в литературе. Как правило, они включают не только ген селективного маркера, но и источник репликации, обеспечивающий репликацию в выбранном хозяине, а также бактериальный или вирусный промотор и в большинстве случаев стоп-кодон транскрипции. Между промотором и стоп-кодоном присутствует обычно, по крайней мере, один сайт рестрикции или полилинкер, который обеспечивает вставку кодирующей последовательности ДНК. Последовательность ДНК, в естественных условиях контролирующую транскрипцию соответствующего гена, можно использовать в качестве последовательности промотора, если она является активной в выбранном организме хозяина. Однако указанную последовательность также можно заменять на другие последовательности промотора. Можно использовать промоторы, обеспечивающие конститутивную экспрессию гена, и индуцируемые промоторы, которые допускают преднамеренный контроль экспрессии гена. Последовательности бактериальных и вирусных промоторов, характеризующиеся указанными свойствами, подробно описаны в литературе. Регуляторные последовательности для экспрессии в микроорганизмах (например, Е. coli, S. cerevisiae) достаточно подробно описаны в литературе. Промоторами, допускающими, прежде всего, высокую экспрессию расположенной по ходу транскрипции последовательности, являются, например, промотор Т7 (Studier и др., Methods in Enzymology, 185, сс. 60-89 (1990)), промоторы lacUV5, trp, trp-lacUV5 (DeBoer и др., в книге Rodriguez and Chamberlin (изд.), Promoters, Structure and Function, Praeger, Нью-Йорк, cc. 462-481 (1982), DeBoer и др., Proc. Natl. Acad. Sci., USA, cc. 21-25 (1983)), lpl, rac (Boros и др., Gene, 42, cc. 97-100 (1986)). Индуцируемые промоторы предпочтительно используют для синтеза полипептидов. Указанные промоторы в большинстве случаев обеспечивают более высокие выходы полипептидов по сравнению с конститутивными промоторами. Для получения оптимального количества полипептида в большинстве случаев используют двухстадийный процесс. Сначала клетки хозяина культивируют в оптимальных условиях, при этом получают относительно высокую плотность клеток. На второй стадии индуцируют транскрипцию в зависимости от типа используемого промотора. В этой связи, прежде всего пригодным является промотор tac, который индуцируется лактозой или ИПТГ (изопропил-β-D-тиогалактопиранозидом) (deBoer и др., Proc. Natl. Acad. Sci., USA, 80, cc. 21-25 (1983)). Стоп-кодоны для транскрипции также описаны в литературе.
Участок, кодирующий алкенол-дегидратазу, можно модифицировать способами, известными специалисту в данной области химии. Так, например, можно вставлять маркеры, которые упрощают очистку белка, такие как гистидиновая метка (см. пример 1). Кроме того, можно также удалять или разрушать сигнальную последовательность фермента, которая обеспечивает локализацию в периплазме, таким образом обеспечивая внутриклеточное продуцирование белка. Можно также присоединять к кодирующей области сигнал секреции, обеспечивающий секрецию белка в культуральной среде.
Можно также экспрессировать алкенол-дегидратазу в виде гибридного белка, при этом алкенол-дегидратазу присоединяют к другому полипептидному фрагменту, например, к другому ферменту.
Трансформацию клетки хозяина полинуклеотидом или вектором по настоящему изобретению можно проводить стандартными способами, например, как описано в учебниках Sambrook и Russell, Molecular Cloning, A Laboratory Manual, CSH Press, Cold Spring Harbor, Нью-Йорк, USA (2001), Methods in Yeast Genetics, A Laboratory Course Manual, Cold Spring Harbor Laboratory Press (1990). Клетку хозяина культивируют в питательной среде, удовлетворяющей требованиям конкретной используемой клетки хозяина, прежде всего, при соответствующем значении рН, температуры, соответствующей концентрации соли, соответствующей аэрации, соответствующей концентрации антибиотиков, витаминов, следовых количеств элементов и т.п.
Организмами, используемыми в настоящем изобретении, являются прокариоты или эукариоты, предпочтительно они представляют собой микроорганизмы. Использованный в данном контексте термин "микроорганизм" относится к бактериям и к грибам, таким как дрожжи, а также к водорослям и архебактериям. Термин "микроорганизм" также включает клетки растений или клетки животных. Прежде всего, в предпочтительном варианте микроорганизмами являются бактерии. Предпочтительными бактериями, которые следует использовать в способе по настоящему изобретению, являются бактерии рода Bacillus, Clostridium, Corynebacterium, Pseudomonas, Zymomonas, Methylobacter или Escherichia. Прежде всего, в предпочтительном варианте бактерия относится к роду Escherichia, а еще более предпочтительно к виду Escherichia coli. В другом предпочтительном варианте бактерия относится к виду Pseudomonas putida или к виду Zymomonas mobilis или к виду Corynebacterium glutamicum.
В другом предпочтительном варианте микроорганизмами являются рекомбинантные бактерии, предпочтительно рода Escherichia, модифицированные таким образом, чтобы эндогенно продуцировать соединение общей формулы CnH2nO, где 3<n<7, и превращать его в диеновое соединение, как описано выше.
Использованный в данном контексте термин "микроорганизм" относится к бактериям и к грибам, таким как дрожжи, а также к водорослям и архебактериям. В одном предпочтительном варианте микроорганизмом является бактерия. В принципе, можно использовать любую бактерию. Предпочтительными бактериями, которые следует использовать в способе по настоящему изобретению, являются бактерии рода Bacillus, Clostridium, Corynebacterium, Pseudomonas, Zymomonas или Escherichia. В прежде всего предпочтительном варианте бактерия принадлежит к виду Escherichia, и даже еще более предпочтительно к виду Escherichia coli. В другом предпочтительном варианте бактерия принадлежит к виду Pseudomonas putida или к виду Zymomonas mobilis или к виду Corynebacterium glutamicum.
В другом предпочтительном варианте микроорганизмом является гриб, более предпочтительно гриб рода Saccharomyces, Schizosaccharomyces, Aspergillus, Trichoderma, Kluyveromyces или Pichia и еще более предпочтительно вида Saccharomyces cerevisiae, Schizosaccharomyces pombe, Aspergillus niger, Trichoderma reesei, Kluyveromyces marxianus, Kluyveromyces lactis или Pichia pastoris.
В прежде всего предпочтительном варианте микроорганизмом является рекомбинантный гриб, предпочтительно дрожжи, продуцирующие соединение общей формулы CnH2nO, где 3<n<7, и превращающие его в диеновое соединение, как описано выше.
В другом предпочтительном варианте способ по настоящему изобретению включает применение фотосинтетического микроорганизма, экспрессирующего алкенол-дегидратазу. Предпочтительно, микроорганизмом является фотосинтетическая бактерия или микроводоросли. Еще даже более предпочтительно указанный микроорганизм характеризуется природным или искусственным свойством эндогенно продуцировать соединение общей формулы CnH2nO, где 3<n<7. В указанном случае микроорганизм способен напрямую продуцировать диен из СО2, присутствующего в растворе.
В еще одном предпочтительном варианте способ по настоящему изобретению включает применение многоклеточного организма, экспрессирующего алкенол-дегидратазу. Примеры указанных организмов включают растения или животных.
В одном варианте способ включает культивирование микроорганизмов в стандартных условиях культивирования (30-37°С при давлении 1 атм., в ферментере, обеспечивающем анаэробный рост бактерий). Температура кипения бутадиена и изопрена составляет -4°С и 34°С, соответственно, и при выбранной температуре культивирования, равной 34°С или выше, указанные соединения уже находятся в газообразном состоянии. В предпочтительном варианте способ включает культивирование микроорганизмов в нестандартных условиях, предпочтительно при более высокой температуре по сравнению с температурой, соответствующей условиям культивирования термофильных организмов. Преимущество указанного варианта заключается в том, что даже такие диены, которые характеризуются более высокой температурой кипения, прежде всего диметилбутадиен (температура кипения которого составляет 68°С), будут выделяться из культуры в газообразном состоянии, и их можно легко собрать из газообразной фазы. В связи с этим прежде всего в таких вариантах осуществления способа по настоящему изобретению, в которых продуцируется диметилбутадиен, микроорганизмом является термофильный микроорганизм, который можно культивировать при температуре 68°С или выше.
В другом предпочтительном варианте в способе по настоящему изобретению применение микроорганизма осуществляют при иммобилизации микроорганизма на подложке.
В еще одном предпочтительном варианте способ по настоящему изобретению осуществляют в микроаэрофильных условиях. Это значит, что количество подаваемого воздуха ограничивают таким образом, чтобы свести к минимуму остаточную концентрацию кислорода в газообразных потоках, содержащих получаемое диеновое соединение.
В другом предпочтительном варианте способ по настоящему изобретению осуществляют в условиях, обеспечивающих выделение диена в газообразном состоянии из реакционной смеси. Преимущество такого способа заключается в том, что термодинамическое равновесие реакции смещается в сторону продуцирования сопряженного диена. Предпочтительно, способ, кроме того, включает стадию отбора газообразного диена. Таким образом, в предпочтительном варианте способ осуществляют в присутствии системы, предназначенной для отбора продуцируемого диена в ходе реакции в газообразной форме.
В предпочтительном варианте способ также включает детектирование продуцируемого диена (бутадиена, изопрена или диметилбутадиена), который присутствует в газообразной фазе. Присутствие диена, который необходимо получить, в окружающей воздушной среде или в среде другого газа, даже в малых количествах, можно детектировать с использованием различных методик и, прежде всего, с использованием систем газовой хроматографии с инфракрасными детекторами или с пламенно-ионизационными детекторами, или в сочетании с масс-спектрометрией.
Настоящее изобретение также относится к применению организма, который продуцирует алкенол-дегидратазу, такую как линалоол-дегидратаза-изомераза, для превращения соединения общей формулы CnH2nO в соединение общей формулы CnH2n-2+Н2О, где 3<n<7, как описано выше в связи со способом по настоящему изобретению. В предпочтительном варианте таким организмом является рекомбинантный организм, при этом подразумевается, что указанный организм генетически модифицирован за счет вставки, по крайней мере, одной молекулы нуклеиновой кислоты, кодирующей алкенол-дегидратазу, такую как линалоол-дегидратаза-изомераза. Предпочтительно, такая молекула нуклеиновой кислоты является гетерогенной в отношении организма, что означает отсутствие указанной молекулы в указанном организме в природных условиях.
В предпочтительном варианте таким организмом является организм, который продуцирует соединение общей формулы CnH2nO, где 3<n<7.
Настоящее изобретение также относится к применению алкенол-дегидратазы, такой как линалоол-дегидратаза-изомераза, для превращения соединения общей формулы CnH2nO в соединение общей формулы CnH2n-2+H2O, где 3<n<7, как описано выше в данном контексте в связи со способом по настоящему изобретению.
Более того, настоящее изобретение также относится к композиции, включающей организм, который продуцирует алкенол-дегидратазу, и соединение общей формулы CnH2nO, где 3<n<7. Настоящее изобретение также относится к композиции, включающей алкенол-дегидратазу, такую как линалоол-дегидратаза-изомераза, а также соединение общей формулы CnH2nO, где 3<n<7.
В отношении предпочтительных вариантов различных перечисленных компонентов, те же самые принципы, описанные выше, относятся к способам по настоящему изобретению.
На фиг. 1 схематически показаны первичные аллиловые спирты (ПРА) общей формулы CnH2nO, где 3<n<7. Прежде всего показаны:
субстрат/систематическое название/формула/категория/R1/R2/продукт.
На фиг. 2 схематически показаны вторичный и третичный аллиловые спирты (ВТА) общей формулы CnH2nO, где 3<n<7.
Субстрат/систематическое название/формула/категория/R1/R2/продукт.
На фиг. 3 схематически показаны первичные гомоаллиловые спирты (ПГА) общей формулы CnH2nO, где 3<n<7.
Субстрат/систематическое название/формула/категория/R1/R2/продукт.
На фиг. 4 схематически показано превращение упомянутых выше соединений ПРА, ПГА и ВТА в сопряженный диен по настоящему изобретению.
На фиг. 5 схематически показаны основные реакции, катализируемые линалоол-дегидратазой-изомеразой.
На фиг. 6 представлена аминокислотная последовательность линалоол-дегидратазы-изомеразы из Castellaniella defragrans (ранее Alcaligenes defragrans).
На фиг. 7 показаны хроматограммы, полученные с применением газового хроматографа с пламенно-ионизационным детектором (ГХ/ПИД), при проведении ферментативного (линия черного цвета) и неферментативного (линия красного цвета) анализов с использованием 80 мМ транс-кротилового спирта после инкубации в течение 22 ч.
На фиг. 8 показаны хроматограммы ГХ/ПИД, полученные при проведении ферментативного (линия черного цвета) и неферментативного (линия красного цвета) анализов с использованием 80 мМ 3-бутен-2-ола после инкубации в течение 22 ч.
На фиг. 9 показаны хроматограммы ГХ/ПИД, полученные при проведении ферментативного (линия черного цвета) и неферментативного (линия синего цвета) анализов с использованием 80 мМ изопренола после инкубации в течение 22 ч.
На фиг. 10 показаны хроматограммы ГХ/ПИД, полученные при проведении ферментативного (линия красного цвета) и неферментативного (линия черного цвета) анализов с использованием 80 мМ 2-метил-3-бутен-2-ола после инкубации в течение 22 ч.
На фиг. 11 представлена кинетика продуцирования 1,3-бутадиена для ряда концентраций транс-кротилового спирта, как описано в примере 7.
На фиг. 12 представлена кинетика продуцирования 1,3-бутадиена для ряда концентраций бут-3-ен-2-ола, как описано в примере 8.
На фиг. 13 показаны хроматограммы ГХ/ПИД, полученные при проведении ферментативного (линия красного цвета) и неферментативного (линия черного цвета) анализов с использованием 50 мМ бут-3-ен-1-ола после инкубации в течение 18 ч.
На фиг. 14 представлены типичные хроматограммы ГХ/ПИД, полученные при проведении ферментативного (линия красного цвета) и неферментативного (линия черного цвета) анализов с использованием 160 мМ 2-метилбут-3-ен-2-ола после инкубации в течение 6 ч.
На фиг. 15 представлены типичные хроматограммы ГХ/ПИД, полученные при проведении ферментативного (линия красного цвета) и неферментативного (линия черного цвета) анализов с использованием 50 мМ 3-метилбут-3-ен-2-ола после инкубации в течение 1 ч.
На фиг. 16 представлена активность укороченных вариантов линалоол-дегидратазы для полной реакции превращения кротилового спирта в 1,3-бутадиен, а также для реакции дегидратации бут-3-ен-2-ола, приводящей к образованию 1,3-бутадиена, соответственно.
На фиг. 17 продемонстрирована способность рекомбинантной линалоол-дегидратазы-изомеразы катализировать превращение цис- или транс-кротилового спирта в 1,3-бутадиен.
Другие объекты и преимущества настоящего изобретения описаны в следующих примерах, которые приведены в целях иллюстрации, и не ограничивают объем настоящего изобретения.
Примеры
Пример 1
Клонирование и экспрессия в E. coli гена линалоол-дегидратазы-изомеразы
Клонирование и бактериальная культура
Последовательность линалоол-дегидратазы-изомеразы, выведенную из генома Castellaniella defragrans (ранее Alcaligenes defragrans), получали конкатенацией олигонуклеотидов, при этом получали последовательность, пригодную для использования кодонов Е. coli. Удлинение из 6 гистидиновых кодонов вставляли после инициирующего кодона метионина, при этом получали аффинную метку для очистки. Полученный таким образом ген клонировали в экспрессионный вектор pET25b(+) (вектор конструировали с использованием GeneArt AG). Компетентные клетки Е. coli BL21 (DE3) (фирмы Novagen) трансформировали указанным вектором по методике теплового шока. В качестве отрицательного контроля штамм Е. coli BL21 (DE3) трансформировали пустым вектором. Трансформированные клетки выращивали при встряхивании (160 об/мин) в автоиндукционной среде ZYM-5052 (Studier FW, Prot. Exp. Pur., 41, cc. 207-234 (2005)) при 37°C в течение 6 ч и экспрессию белка продолжали при 18°С в течение ночи (приблизительно 12 ч). Клетки собирали центрифугированием при 4°С, 10000 об/мин в течение 20 мин и осадки замораживали при - 80°С.
Приготовление клеточного лизата
Осадки из 100 мл культивированных клеток оттаивали на льду и ресуспендировали в 4 мл 50 мМ Трис-HCl, рН 7,5. Затем добавляли 10 мкл лизоназы (фирмы Novagen). Клетки инкубировали при комнатной температуре в течение 10 мин и затем снова помещали в лед на 20 мин. Концентрацию белка определяли методом Брэдфорда (фирмы Biorad).
Пример 2
Получение 1,3-бутадиена из (2Е)-2-бутен-1-ола (транс-кротилового спирта)
Ферментативные анализы проводили в следующих условиях:
50 мМ Трис-HCl, рН 7,5,
2 мМ D,L-дитиотреит,
0-80 мМ (2Е)-2-бутен-1-ол (транс-кротиловый спирт).
Значение рН доводили до значения 7,5.
Клеточный лизат (0,25 мл), содержащий рекомбинантную линалоол-дегидратазу-изомеразу, добавляли в 0,5 мл реакционной смеси. Параллельно проводили контрольную реакцию в отсутствие фермента с использованием лизата клеток Е. Coli, трансформированных пустым вектором. Анализируемые смеси инкубировали при 37°С в течение 1-22 ч в герметично закрытом стеклянном флаконе (объемом 2 мл, фирмы Interchim) при встряхивании. Затем отбирали 1 мл верхней фазы и вкалывали в газовый хроматограф Varian 450-GC, снабженный пламенно-ионизационным детектором (ПИД). В качестве газа-носителя использовали азот при скорости потока 1,5 мл/мин. Летучие компоненты разделяли хроматографически на колонке Rt-Alumina Bond/Na2SO4 (фирмы Restek) в изотермическом режиме при 130°С. Продукт ферментативной реакции идентифицировали при сравнении со стандартом - 1,3-бутадиеном (фирмы Sigma). В указанных условиях проведения газовой хроматографии (ГХ) время удерживания бутадиена составляло 7,6 мин. Значительное продуцирование 1,3-бутадиена наблюдали при проведении ферментативного анализа в присутствии линалоол-дегидратазы-изомеразы. При проведении контрольного анализа в отсутствие фермента сигнала, соответствующего бутадиену, не наблюдали (фиг. 8). Число оборотов для указанного превращения достигало приблизительно 3×10-5×с-1 молекул субстрата/активный участок фермента.
Пример 3
Получение 1,3-бутадиена из 3-бутен-2-ола
Ферментативные анализы проводили в следующих условиях:
50 мМ Трис-HCl, рН 7,5,
2 мМ D,L-дитиотреит,
0-80 мМ 3-бутен-2-ол.
Значение рН доводили до значения 7,5.
Клеточный лизат (0,25 мл), содержащий рекомбинантную линалоол-дегидратазу-изомеразу, добавляли в 0,5 мл реакционной смеси. Параллельно проводили контрольную реакцию в отсутствие фермента с использованием лизата клеток Е. Coli, трансформированных пустым вектором. Анализируемые смеси инкубировали при 37°С в течение 1-22 ч в герметично закрытом стеклянном флаконе (объемом 2 мл, фирмы Interchim) при встряхивании. Продуцирование 1,3-бутадиена анализировали методом ГХ/ПИД, как описано в примере 2. Значительное продуцирование 1,3-бутадиена наблюдали при проведении ферментативного анализа с использованием линалоол-дегидратазы-изомеразы. При проведении контрольного анализа в отсутствие фермента сигнала, соответствующего бутадиену, не наблюдали (фиг. 8). Число оборотов для указанного превращения достигало приблизительно 10-4×с-1 молекул субстрата/активный участок фермента.
Аналогичные эксперименты также проводили с использованием R- и S-энантиомерно чистого бут-3-ен-2-ола, при этом получали аналогичные результаты.
Пример 4
Продуцирование 2-метил-1,3-бутадиена (изопрена) из 3-метил-2-бутен-1-ола (пренола)
Ферментативные анализы проводили в следующих условиях:
50 мМ Трис-HCl, рН 7,5,
2 мМ D,L-дитиотреит,
0-80 мМ 3-метил-2-бутен-1-ол (пренол).
Значение рН доводили до значения 7,5.
Клеточный лизат (0,25 мл), содержащий рекомбинантную линалоол-дегидратазу-изомеразу, добавляли в 0,5 мл реакционной смеси. Параллельно проводили контрольную реакцию в отсутствие фермента с использованием лизата клеток Е. Coli, трансформированных пустым вектором. Анализируемые смеси инкубировали при 37°С в течение 1-22 ч в герметично закрытом стеклянном флаконе (объемом 2 мл, фирмы Interchim) при встряхивании. Затем отбирали 100 мкл верхней фазы и вкалывали в газовый хроматограф Varian 450-GC, снабженный ПИД. Летучие компоненты из верхней фазы разделяли на колонке Rtx-1 (фирмы Restek) с использованием азота в качестве газа-носителя при скорости потока 1,5 мл/миг. Каждый образец нагревали в печи в течение 4 мин при 100°С, температуру повышали со скоростью 20°С/мин до температуры 130°С и выдерживали в течение 1,5 мин при 130°С. Полное время анализа составляло 7 мин. Продукт ферментативной реакции идентифицировали при сравнении с изопреновым стандартом (фирмы Sigma). В указанных условиях ГХ время удерживания изопрена составляло 3,08 мин. Значительное продуцирование изопрена наблюдали в ходе ферментативного анализа в присутствии линалоол-дегидратазы-изомеразы. При проведении контрольного анализа в отсутствие фермента регистрировали незначительный сигнал, соответствующий изопрену, из-за самопроизвольного разложения пренола (табл. 1). Число оборотов для указанного превращения достигало приблизительно 3×10-4×с-1 молекул субстрата/активный участок фермента.

Пример 5
Продуцирование 2-метил-1,3-бутадиена (изопрена) из 3-метил-3-бутен-1-ола(изопренола)
Ферментативные анализы проводили в следующих условиях:
50 мМ Трис-HCl, рН 7,5,
2 мМ D,L-дитиотреит,
0-80 мМ 3-метил-3-бутен-1-ол (изопренол).
Значение рН доводили до значения 7,5.
Клеточный лизат (0,25 мл), содержащий рекомбинантную линалоол-дегидратазу-изомеразу, добавляли в 0,5 мл реакционной смеси. Параллельно проводили контрольную реакцию в отсутствие фермента с использованием лизата клеток Е. Coli, трансформированных пустым вектором. Анализируемые смеси инкубировали при 37°С в течение 1-22 ч в герметично закрытом стеклянном флаконе (объемом 2 мл, фирмы Interchim) при встряхивании. Продуцирование изопрена анализировали методом ГХ/ПИД, как описано в примере 4. Значительное продуцирование изопрена наблюдали при проведении ферментативного анализа в присутствии линалоол-дегидратазы-изомеразы. При проведении контрольного анализа в отсутствие фермента сигнала, соответствующего изопрену, не наблюдали (фиг. 9). Число оборотов для указанного превращения достигало приблизительно 3×10-4×с-1 молекул субстрата/активный участок фермента.
Пример 6
Продуцирование 2-метил-1,3-бутадиена (изопрена) из 2-метил-3-бутен-2-ола
Ферментативные анализы проводили в следующих условиях:
50 мМ Трис-HCl, рН 7,5,
2 мМ D,L-дитиотреит,
0-80 мМ 2-метил-3-бутен-2-ол.
Значение рН доводили до значения 7,5.
Клеточный лизат (0,25 мл), содержащий рекомбинантную линалоол-дегидратазу-изомеразу, добавляли в 0,5 мл реакционной смеси. Параллельно проводили контрольную реакцию в отсутствие фермента с использованием лизата клеток Е. Coli, трансформированных пустым вектором. Анализируемые смеси инкубировали при 37°С в течение 1-22 ч в герметично закрытом стеклянном флаконе (объемом 2 мл, фирмы Interchim) при встряхивании. Продуцирование изопрена анализировали методом ГХ/ПИД, как описано в примере 4. Значительное продуцирование изопрена наблюдали при проведении ферментативного анализа в присутствии линалоол-дегидратазы-изомеразы. При проведении контрольного анализа в отсутствие фермента сигнала, соответствующего изопрену, не наблюдали (фиг. 10). Число оборотов для указанного превращения достигало приблизительно 10-3×с-1 молекул субстрата/активный участок фермента.
Пример 7
Кинетика продуцирования 1,3-бутадиена из (Е)-бут-2-ен-1-ола (транс-кротилового спирта)
Лизис клеток и приготовление супернатанта
Клеточный лизат получали по методике, описанной в примере 1, но при следующих изменениях. Осадок, полученный из 200 мл культивированных клеток, оттаивали на льду и ресуспендировали в 3 мл Трис-HCl (50 мМ, рН 7,5), содержащем 4 мМ D,L-дитиотреит, 20 мМ глутатион, 25 мМ MgCb и 25 мМ KCl. Затем добавляли 10 мкл лизоназы (фирмы Merck). Клетки инкубировали при комнатной температуре в течение 10 мин и затем снова помещали в лед на 20 мин. Клеточный лизат осветляли центрифугированием при 13000 об/мин при 4°С в течение 10 мин. Собранный супернатант концентрировали в ячейке для ультрафильтрации Amicon Ultra-4, 10 кДа (фирмы Millipore). Уровень экспрессии линалоол-дегидратазы-изомеразы в супернатанте оценивали методом электрофореза в полиакриламидном геле в додецилсульфате натрия (ПААГ-ДСН). Общую концентрацию белка определяли методом Брэдфорда (фирмы Biorad).
Ферментативный анализ
Ферментативные анализы проводили в следующих условиях:
50 мМ Трис-HCl, рН 7,5,
0-160 мМ транс-кротиловый спирт (фирмы Alfa Aesar).
Супернатант (0,2-0,4 мл), содержащий рекомбинантную линалоол-дегидратазу-изомеразу, добавляли в 0,5 мл реакционной смеси. Концентрация линалоол-дегидратазы-изомеразы при проведении указанного анализа составляла приблизительно 1,2 мг/мл. Параллельно проводили контрольные реакции в отсутствие фермента, содержащие супернатант клеток E. coli, трансформированных пустым вектором. Реакционные смеси инкубировали при 37°С в герметично закрытом стеклянном флаконе (объемом 2 мл, фирмы Interchim) при встряхивании. Реакцию останавливали при замораживании аналитических пробирок при -80°С. Продуцирование бутадиена определяли, анализируя аликвотные части образцов, отобранные после инкубации в течение 5,5 ч. Количество полученного 1,3-бутадиена определяли методом ГХ по методике, описанной в примере 2. На фиг. 11 представлена кинетика продуцирования 1,3-бутадиена для ряда концентраций транс-кротилового спирта.
Пример 8
Кинетика продуцирования 1,3-бутадиена из бут-3-ен-2-ола
Осветленный клеточный лизат, содержащий рекомбинантную линалоол-дегидратазу-изомеразу, получали по методике, описанной в примере 7.
Ферментативный анализ
Ферментативные анализы проводили в следующих условиях:
50 мМ Трис-HCl, рН 7,5,
0-160 мМ бут-3-ен-2-ол (фирмы Sigma-Aldrich).
Супернатант (0,2-0,4 мл), содержащий рекомбинантную линалоол-дегидратазу-изомеразу, добавляли в 0,5 мл реакционной смеси. Концентрация линалоол-дегидратазы-изомеразы в указанном анализе составляла приблизительно 1,2 мг/мл. Параллельно проводили контрольные реакции в отсутствие фермента, содержащие супернатант клеток E. coli, трансформированных пустым вектором. Реакционные смеси инкубировали при 37°С в герметично закрытом стеклянном флаконе (объемом 2 мл, фирмы Interchim) при встряхивании. Реакцию останавливали при замораживании аналитических пробирок при -80°С. Продуцирование бутадиена определяли, анализируя аликвотные части образцов, отобранные после инкубации в течение 5,5 ч. Количество полученного 1,3-бутадиена определяли методом ГХ по методике, описанной в примере 2. На фиг. 12 представлена кинетика продуцирования 1,3-бутадиена для ряда концентраций бут-3-ен-2-ола.
Пример 9
Получение 1,3-бутадиена из бут-3-ен-1-ола (изокротилового спирта)
Приготовление клеточного лизата
Клеточный лизат получали по методике, описанной в примере 1, при следующих изменениях. Осадок, полученный из 200 мл культивированных клеток, оттаивали на льду и ресуспендировали в 3 мл Трис-HCl (50 мМ, рН 7,5), содержащем 4 мМ D,L-дитиотреит, 20 мМ глутатион, 25 мМ MgCl2 и 25 мМ KCl. Затем добавляли 10 мкл лизоназы (фирмы Merck). Клетки инкубировали при комнатной температуре в течение 10 мин и затем снова помещали в лед на 20 мин.
Ферментативный анализ
Ферментативные анализы проводили в следующих условиях:
50 мМ Трис-HCl, рН 7,5,
50 мМ бут-3-ен-1-ол (фирмы Sigma).
Клеточный лизат (0,4 мл), содержащий рекомбинантную линалоол-дегидратазу-изомеразу, добавляли в 0,5 мл реакционной смеси. Параллельно проводили контрольные реакции в отсутствие фермента в присутствии лизата клеток E. coli, трансформированных пустым вектором. Реакционные смеси инкубировали при 37°С в течение 18 ч в герметично закрытом стеклянном флаконе (объемом 2 мл, фирмы Interchim) при встряхивании. Затем отбирали 1 мл верхней фазы и вкалывали в газовый хроматограф Varian 450-GC, снабженный ПИД. В качестве газа-носителя использовали азот при скорости потока 6 мл/мин. Летучие компоненты разделяли на колонке J&W GS-Alumina (длина 30 м×внутренний диаметр 0,53 мм, фирмы Restek) в изотермическом режиме при 130°С. Продукт ферментативной реакции идентифицировали при сравнении с 1,3-бутадиеновым стандартом (фирмы Sigma-Aldrich). В указанных условиях проведения ГХ время удерживания бутадиена составляло 3,05 мин. 1,3-Бутадиен получали при проведении ферментативного анализа в присутствии линалоол-дегидратазы-изомеразы, при проведении контрольного анализа в отсутствие фермента сигнал, соответствующий бутадиену, не наблюдали (фиг. 13).
Пример 10
Продуцирование 2-метил-1,3-бутадиена (изопрена) из 2-метилбут-3-ен-2-ола
При проведении другой серии экспериментов воспроизводили реакцию, исследованную в примере 6, и подтверждали результат. Анализ проводили в следующих условиях.
Осветленный клеточный лизат, содержащий рекомбинантную линалоол-дегидратазу-изомеразу, получали по методике, описанной в примере 7.
Ферментативный анализ
Ферментативные анализы проводили в следующих условиях:
50 мМ Трис-HCl, рН 7,5,
160 мМ 2-метилбут-3-ен-2-ол (фирмы Sigma).
Супернатант (0,2-0,4 мл), содержащий рекомбинантную линалоол-дегидратазу-изомеразу, добавляли в 0,5 мл реакционной смеси. Концентрация линалоол-дегидратазы-изомеразы в указанном анализе составляла приблизительно 1,2 мг/мл. Параллельно проводили контрольные реакции в отсутствие фермента в присутствии супернатанта клеток E. coli, трансформированных пустым вектором. Реакционные смеси инкубировали при 37°С в течение 6 ч в герметично закрытом стеклянном флаконе (объемом 2 мл, фирмы Interchim) при встряхивании. Затем реакцию останавливали при замораживании аналитических пробирок при -80°С.
Затем отбирали 1 мл верхней фазы и вкалывали в газовый хроматограф Varian 450-GC, снабженный ПИД. В качестве газа-носителя использовали азот при скорости потока 1,5 мл/мин. Летучие компоненты разделяли на колонке Rt-Alumina Bond7Na2SO4 (фирмы Restek) в изотермическом режиме при 155°С. Продукт ферментативной реакции идентифицировали при сравнении с изопреновым стандартом (фирмы Sigma-Aldrich). В указанных условиях проведения ГХ время удерживания изопрена составляло 7,5 мин.
Значительное продуцирование изопрена наблюдали при проведении ферментативного анализа в присутствии линалоол-дегидратазы-изомеразы. При проведении контрольного анализа в отсутствие фермента сигнала, соответствующего изопрену, не наблюдали (фиг. 14).
Пример 11
Продуцирование 2-метил-1,3-бутадиена (изопрена) из 3-метилбут-3-ен-2-ола
Приготовление клеточного лизата
Клеточный лизат получали по методике, описанной в примере 1, при следующих изменениях. Осадок, полученный из 200 мл культивированных клеток, оттаивали на льду и ресуспендировали в 3 мл Трис-HCl (50 мМ, рН 7,5), содержащем 4 мМ D,L-дитиотреит, 20 мМ глутатион, 25 мМ MgCl2 и 25 мМ KCl. Затем добавляли 10 мкл лизоназы (фирмы Merck). Клетки инкубировали при комнатной температуре в течение 10 мин и затем снова помещали в лед на 20 мин.
Ферментативный анализ
Ферментативные анализы проводили в следующих условиях:
50 мМ Трис-HCl, рН 7,5,
50 мМ 3-метилбут-3-ен-2-ол (фирмы Sigma-Aldrich).
Клеточный лизат (0,45 мл), содержащий рекомбинантную линалоол-дегидратазу-изомеразу, добавляли в 0,5 мл реакционной смеси. Параллельно проводили контрольную реакцию в отсутствие фермента в присутствии лизата клеток E. coli, трансформированных пустым вектором. Реакционные смеси инкубировали при 37°С в течение 1 ч в герметично закрытом стеклянном флаконе (объемом 2 мл, фирмы Interchim) при встряхивании. Затем отбирали 1 мл верхней фазы и вкалывали в газовый хроматограф Varian 450-GC, снабженный ПИД. В качестве газа-носителя использовали азот при скорости потока 6 мл/мин. Летучие компоненты разделяли на колонке J&W GS-Alumina (длина 30 м × внутренний диаметр 0,53 мм, фирмы Restek) в изотермическом режиме при 150°С. Продукт ферментативной реакции идентифицировали при сравнении с изопреновым стандартом (фирмы Sigma-Aldrich). В указанных условиях проведения ГХ время удерживания изопрена составляло 3,85 мин. Значительное продуцирование изопрена наблюдали при проведении ферментативного анализа с использованием линалоол-дегидратазы-изомеразы, при проведении контрольного анализа в отсутствие фермента регистрировали незначительный сигнал, соответствующий изопрену, из-за самопроизвольного разложения 3-метилбут-3-ен-2-ола (фиг. 15, табл. 2).

Пример 12
Укороченные варианты линалоол-дегидратазы: активность в отношении полной реакции превращения кротилового спирта в 1,3-бутадиен и реакции дегидратации бут-3-ен-2-ола. приводящей к образованию 1,3-бутадиена
В ходе различных экспериментов, направленных на повышение активности и растворимости линалоол-дегидратазы-изомеразы, был идентифицирован набор из 9 вариантов фермента, укороченных по С-концевому фрагменту. Полноразмерный фермент дикого типа соответствовал Ml-К397 и указан в последовательности (SEQ ID NO: 1). Наблюдаемые укороченные варианты были обозначены следующим образом: M1-L385, M1-R386, М1-Р388, М1-Р389, М1-А391, М1-К393, M1-L394, М1-А395 и M1-G396, как показано в последовательности SEQ ID NO: 1. В указанных укороченных по С-концевому фрагменту вариантах была изменена только длина цепи белка, остальная часть последовательности белка оставалась неизменной. Идентичность наиболее короткого варианта (M1-L385) составляла 96,9%.
Активность определяли при проведении следующего анализа.
Указанный анализ проводили следующим образом. Набор вариантов линалоол-дегидратазы-изомеразы, укороченных по С-концевому фрагменту, клонированных в коммерческий экспрессионный вектор pET25b+, трансформировали в компетентные клетки BL21 (DE3). Выделенные клоны использовали для посева в 1 мл автоиндукционной среды (Studier F.W., Prot. Exp. Pur., 41, cc. 207-234 (2005)) и культивировали при 30°С в течение ночи (20-22 ч) в инкубаторе-качалке при 700 об/мин и 85% влажности. Клетки осаждали центрифугированием и выдерживали при -80°С в течение ночи. Указанные клеточные осадки, которые содержали экспрессированные варианты рекомбинантной линалоол-дегидратазы-изомеразы, ресуспендировали в реакционной смеси, содержащей 50 мМ Трис-HCl (рН 7,5), 25 мМ KCl, 25 мМ MgCl2, 4 мМ ДТТ, 10 мМ глутатион, а также 50 мМ транс-кротиловый спирт (фирмы Alfa Aesar) или 50 мМ бут-3-ен-2-ол (фирмы Sigma Aldrich). Контрольные реакции проводили с использованием бактериальных клонов, содержащих пустой экспрессионный вектор peT25b+ или экспрессионный вектор, экспрессирующий фермент дикого типа. Указанную реакционную смесь инкубировали при 37°С в течение 16 ч, и реакцию останавливали при инкубации в течение 5 мин при 80°С. Затем определяли количество полученного 1,3-бутадиена методом ГХ. Для анализа верхней фракции методом ГХ газообразную верхнюю фракцию (300 мкл) вкалывали в систему Bruker GC450, снабженную колонкой Restek RT-Alumina (5 м×0,32 мм) и системой ПИД. Анализ методом ГХ, использованный для детектирования 1,3-бутадиена, проводили при постоянной температуре в печи, равной 140°С, температуре порта инжектора, равной 200°С, при делении потока 1:4 и температуре ПИД, равной 250°С. В качестве газа-носителя использовали азот (при постоянной скорости потока 1,25 мл/мин), а для снабжения системы детектора ПИД использовали смесь воздуха (при скорости потока воздуха 300 мл/мин), азота (при скорости потока 28 мл/мин) и водорода (скорость потока 30 мл/мин).
Активность описанных выше укороченных вариантов линалоол-дегидратазы для полной реакции превращения кротилового спирта в 1,3-бутадиен, а также соответственно для реакции дегидратации бут-3-ен-2-ола, приводящей к образованию 1,3-бутадиена, показана на фиг.16.
Пример 13
Рекомбинантная линалоол-дегидратаза-изомераза катализирует превращение цис- и транс-стереоизомеров кротилового спирта в 1,3-бутадиен
Ген, кодирующий рекомбинантную линалоол-дегидратазу-изомеразу, субклонировали в коммерческий бактериальный экспрессионный вектор реТ-25b+фирмы Novagen, трансформировали в компетентные клетки BL21 (DE3) и высеивали на чашки Петри с агаром Луриа-Бертани (LB), дополненным соответствующим антибиотиком. Компетентные клетки BL21 (DE3) также трансформировали вектором реТ-25b+, чтобы использовать в качестве отрицательного контроля в последующих ферментативных анализах. Для посева в 500 мл автоиндукционной среды (Studier F.W. (2005), цит. выше) использовали одиночные трансформанты и культуры инкубировали при 30°С в инкубаторе-качалке в течение ночи. Клеточные осадки хранили при -80°С в течение ночи, затем ресуспендировали в 7,5 мл лизирующего буфера (50 мМ Трис-HCl (рН 7,5), 4 мМ ДТТ, 25 мМ MgCl2, 25 мМ KCl), содержащего 50 мкл лизоназы фирмы Merck Novagen. Клеточные суспензии инкубировали при комнатной температуре в течение 10 мин, затем в течение 20 мин на льду. Затем клеточные лизаты осветляли центрифугированием и супернатанты концентрировали в два раза в ячейке для ультрафильтрации Amicon Ultra (фирмы Millipore). Количество рекомбинантной линалоол-дегидратазы-изомеразы, присутствующей в концентрированной растворимой фракции, оценивали методом электрофореза в полиакриламидном геле в додецилсульфате натрия (ПААГ-ДСН) при сопоставлении с калибровочной кривой для бычьего сывороточного альбумина, полученной после денситометрического сканирования геля. Ферментативные реакции проводили в стеклянных флаконах (объемом 2 мл, фирмы Interchim) с использованием 225 мкл супернатанта клеточного лизата, транс-кротилового спирта (фирмы Alfa Aesar) или цис-кротилового спирта (фирмы ChemSampCo) при концентрации в диапазоне от 0 до 100 мМ, в буфере 4 мМ ДТТ, 25 мМ MgCl2, 25 мМ KCl, 4 мМ глутатион и 50 мМ Трис-HCl (рН 7,5), при этом конечный объем реакционной смеси составлял 250 мкл. Ферментативные реакции проводили с использованием клеточного лизата, содержащего рекомбинантную линалоол-дегидратазу-изомеразу, или клеточного лизата, полученного из клеток, трансформированных пустым экспрессионным вектором. Флаконы закрывали и инкубировали при 37°С в течение 4 ч. Ферментативную реакцию останавливали при инкубации в течение 5 мин при 80°С и определяли количество 1,3-бутадиена, присутствующего в верхней фазе реакционной смеси, методом ГХ. Для анализа верхней фракции методом ГХ газообразную верхнюю фракцию (300 мкл) вкалывали в систему Bruker GC450, снабженную колонкой Restek RT-Alumina (5 м×0,32 мм) и системой ПИД. Анализ методом ГХ, использованный для детектирования 1,3-бутадиена, проводили при постоянной температуре печи, равной 140°С (изотермический режим), температуре порта инжектора, равной 200°С, при делении потока 1:4 и температуре ПИД, равной 250°С. В качестве газа-носителя использовали азот (при постоянной скорости потока 1,25 мл/мин), а для снабжения системы детектора ПИД использовали смесь воздуха (при скорости потока воздуха 300 мл/мин), азота (при скорости потока 28 мл/мин) и водорода (скорость потока 30 мл/мин). Продукт ферментативной реакции идентифицировали при сравнении с 1,3-бутадиеновым стандартом (фирмы Sigma-Aldrich). В указанных условиях проведения ГХ время удерживания бутадиена составляло 1,05 мин. На фиг.17 показана способность рекомбинантной линалоол-дегидратазы-изомеразы катализировать превращение цис- или транс-кротилового спирта в 1,3-бутадиен. При проведении контрольных реакций в отсутствие фермента сигнал, соответствующий 1,3-бутадиену, не наблюдали.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПРОИЗВОДСТВА 1,3-БУТАДИЕНА ИЗ 1,3-БУТАНДИОЛА | 2015 |
|
RU2690678C2 |
| Способ получения диенов | 2016 |
|
RU2692477C2 |
| СПОСОБ ПОЛУЧЕНИЯ АЛКЕНОЛОВ И ЕГО ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ 1,3-БУТАДИЕНА | 2015 |
|
RU2695786C2 |
| СПОСОБ ПОЛУЧЕНИЯ ДИЕНОВ | 2020 |
|
RU2826077C1 |
| МОДИФИЦИРОВАННЫЕ МИКРООРГАНИЗМЫ И СПОСОБЫ ПОЛУЧЕНИЯ БУТАДИЕНА С ИХ ПРИМЕНЕНИЕМ | 2012 |
|
RU2639564C2 |
| ТОПЛИВНЫЕ КОМПОЗИЦИИ, СОДЕРЖАЩИЕ ПРОИЗВОДНЫЕ ИЗОПРЕНА | 2010 |
|
RU2531623C2 |
| СПОСОБ ДЕГИДРАТАЦИИ КИСЛОРОДСОДЕРЖАЩИХ СОЕДИНЕНИЙ | 2016 |
|
RU2701863C2 |
| СПОСОБ ПОЛУЧЕНИЯ C5-ФРАКЦИИ "FCC", ОБОГАЩЕННОЙ ИЗОПРЕНОМ, И СЕЛЕКТИВНАЯ ПОЛИМЕРИЗАЦИЯ ИЗОПРЕНА ИЗ ДАННОЙ ФРАКЦИИ | 2004 |
|
RU2351612C2 |
| СПОСОБ ПОЛУЧЕНИЯ ОЛЕФИНА ПОСРЕДСТВОМ КАТАЛИТИЧЕСКОЙ КОНВЕРСИИ ПО МЕНЬШЕЙ МЕРЕ ОДНОГО СПИРТА | 2014 |
|
RU2660132C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПОЛУЧЕНИЯ ИЗОПРЕНА, НЕ СОДЕРЖАЩЕГО С5-УГЛЕВОДОРОДОВ В УСЛОВИЯХ НАРУШЕНИЯ ВЗАИМОСВЯЗИ МЕЖДУ ПРОДУЦИРОВАНИЕМ ИЗОПРЕНА И РОСТОМ КЛЕТОК И/ИЛИ В УСЛОВИЯХ ПРОДУЦИРОВАНИЯ ИЗОПРЕНА НА БЕЗОПАСНЫХ РАБОЧИХ УРОВНЯХ | 2009 |
|
RU2563513C2 |



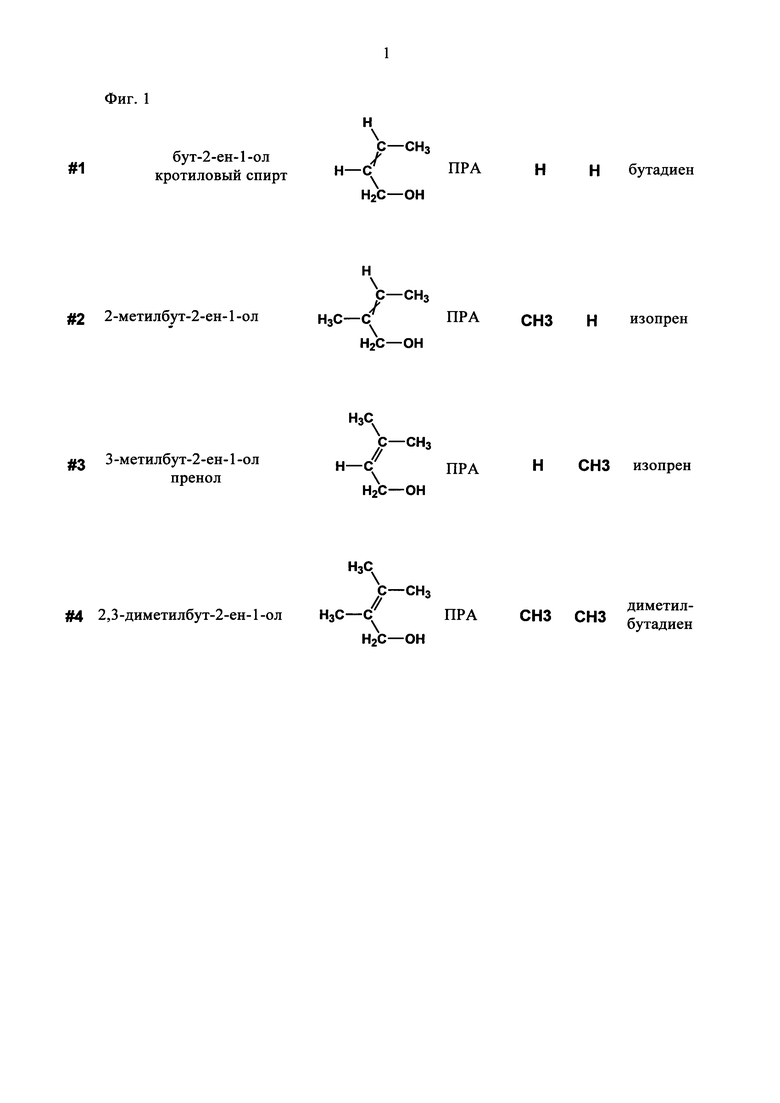
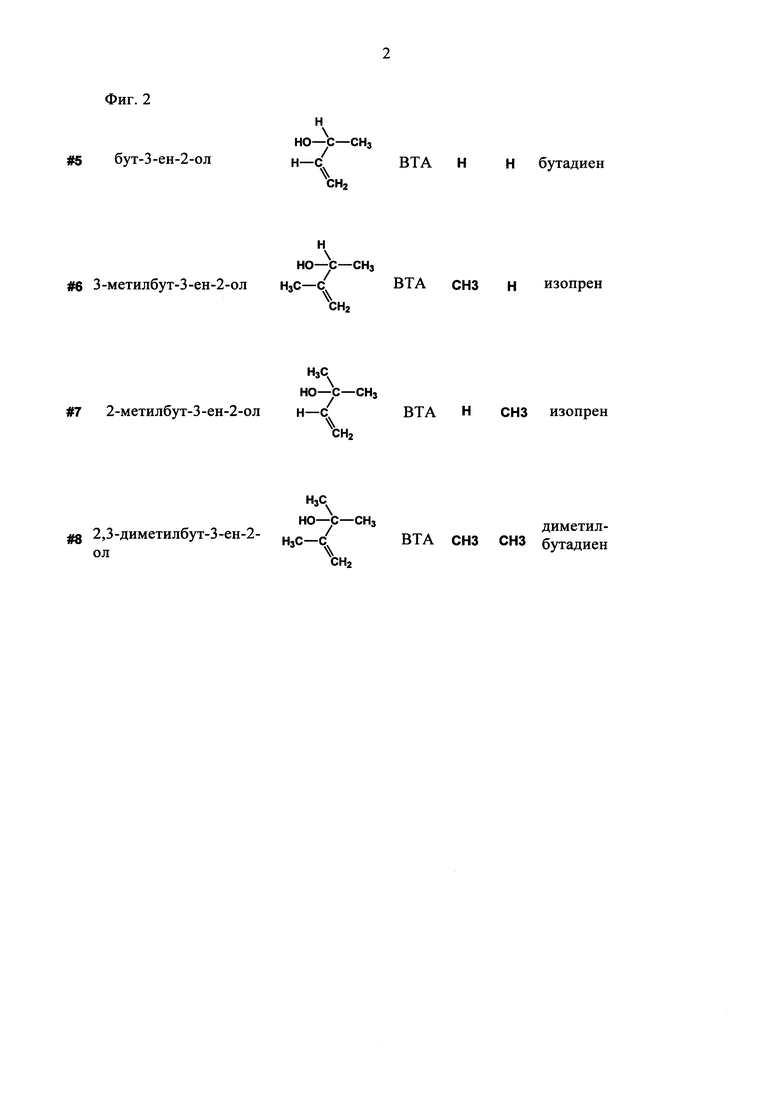
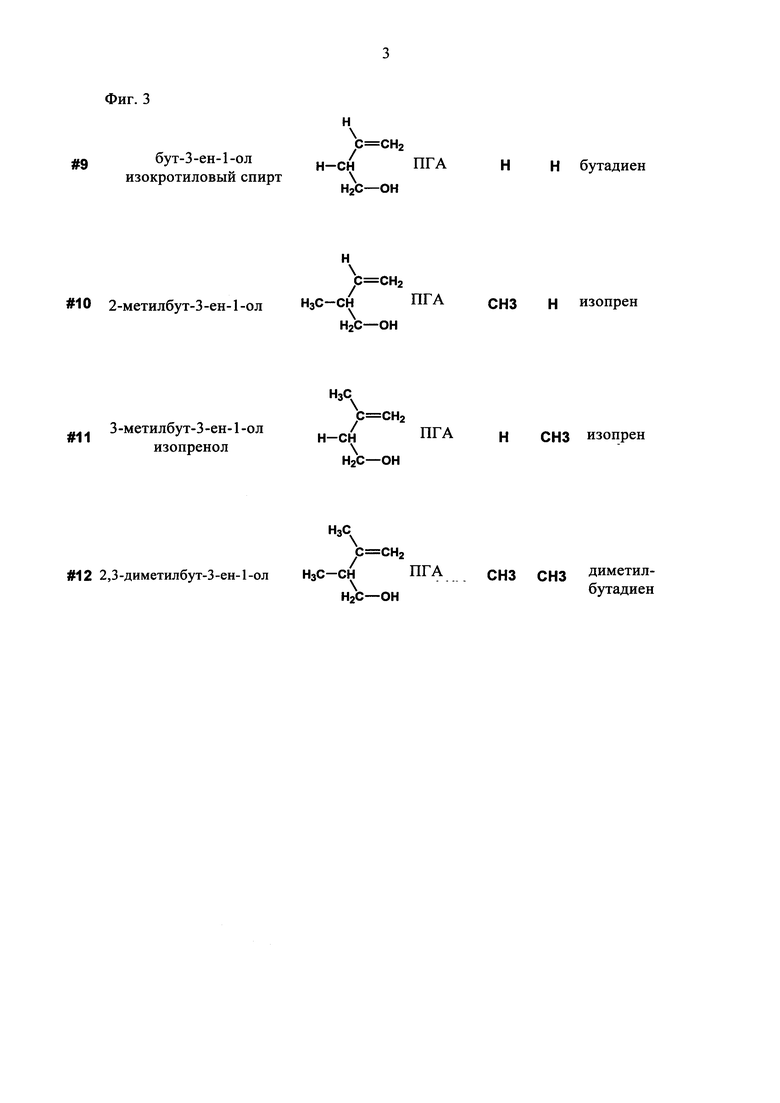




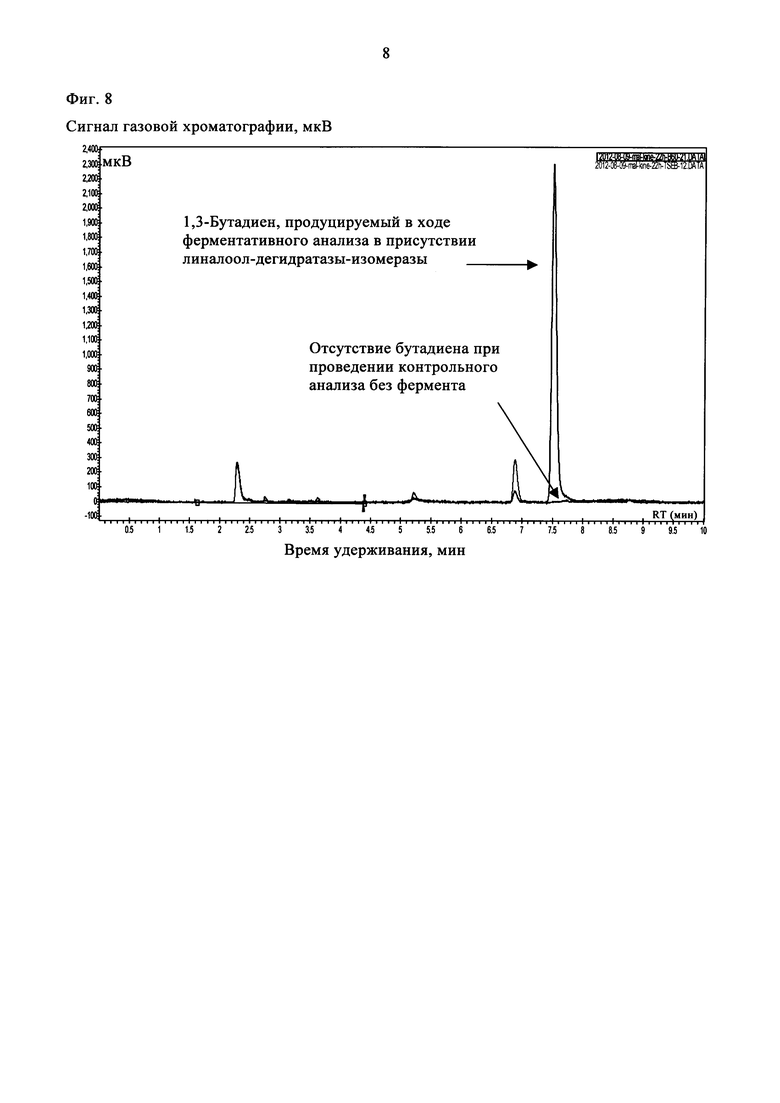

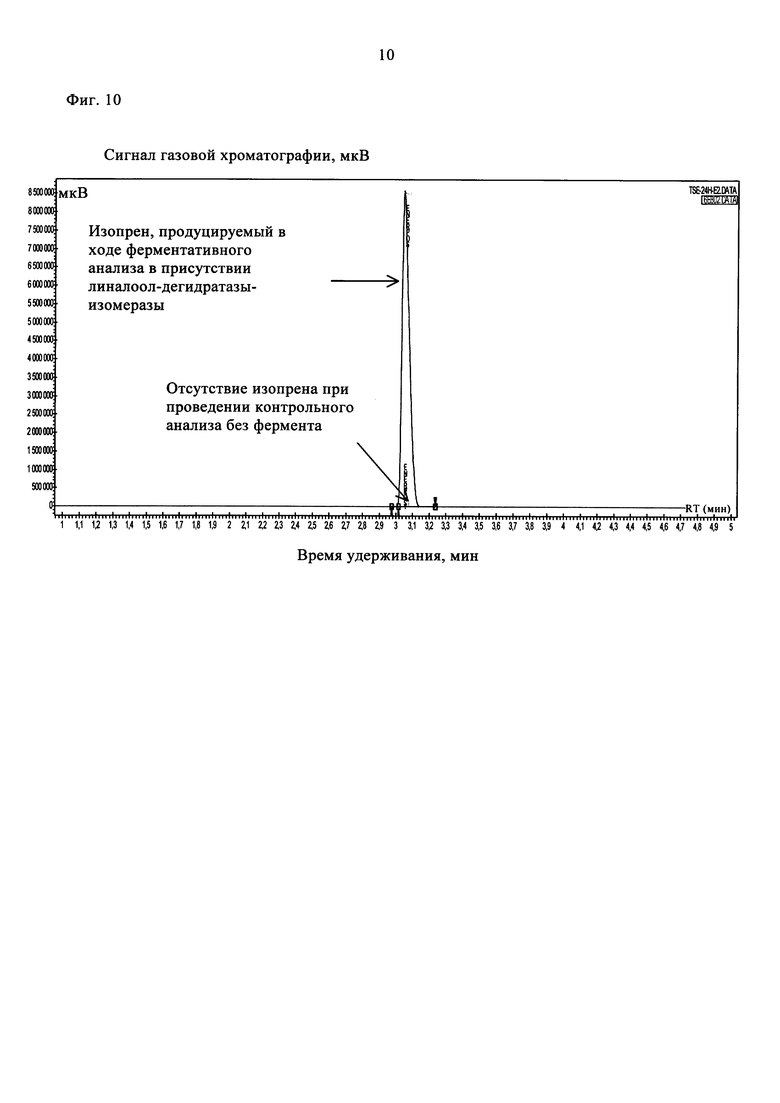
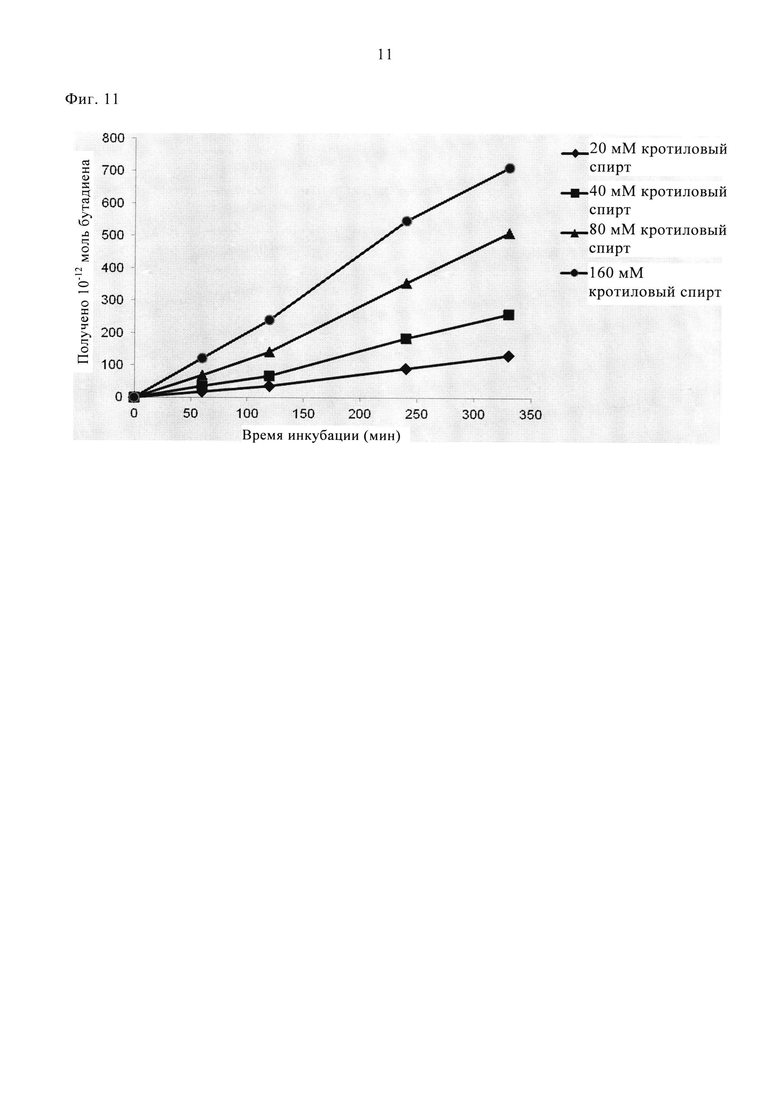

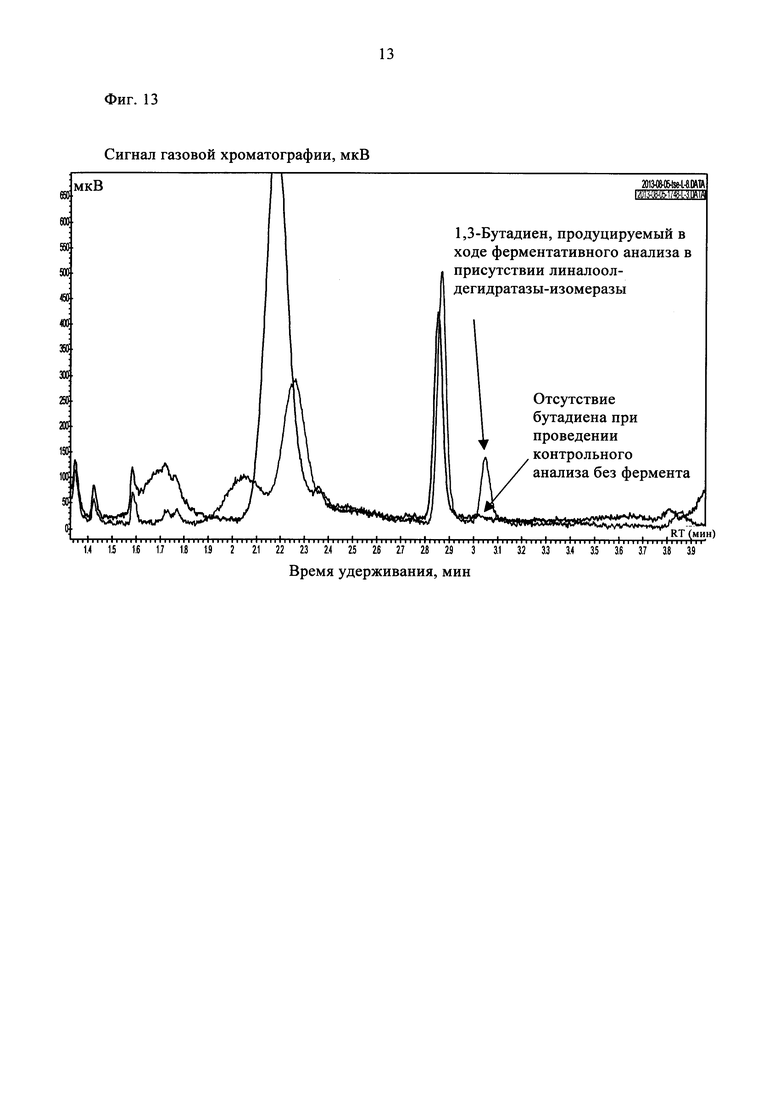
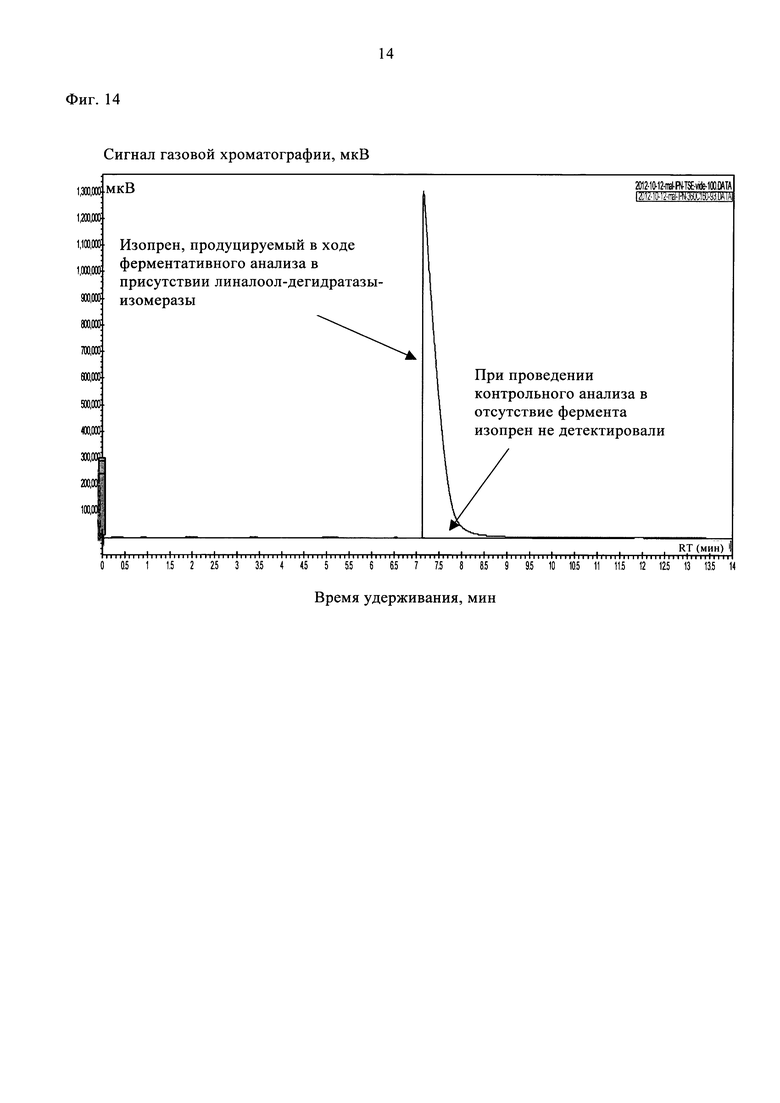
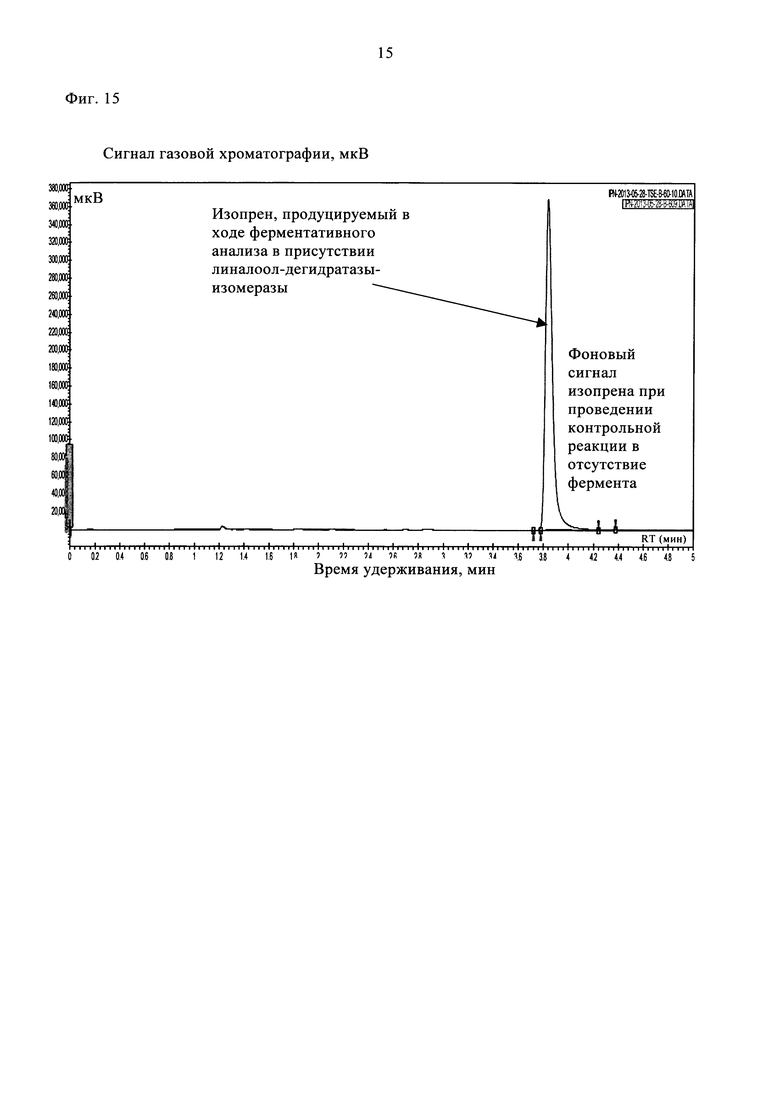


Группа изобретений относится к биотехнологии, а именно к получению сопряженных диенов методом ферментативной дегидратации. Способ получения сопряженного диена включает стадию ферментативного превращения соединения общей формулы СnН2nO в соединение общей формулы CnH2n-2+Н2О, где 3<n<7, с использованием линалоол-дегидратазы (ЕС 4.2.1.127). Указанный способ также может быть осуществлен с использованием микроорганизма, продуцирующего линалоол-дегидратазу. Также предложены композиции для получения соединения общей формулы CnH2n-2+H2O, включающие линалоол-дегидратазу или микроорганизм, который ее продуцирует, и соединение общей формулы CnH2nO, где 3<n<7. Изобретения позволяют синтезировать летучие диеновые соединения, прежде всего сопряженные диены, такие как 1,3-диены, из низших алкенолов. 4 н. и 18 з.п. ф-лы, 17 ил., 2 табл., 13 пр.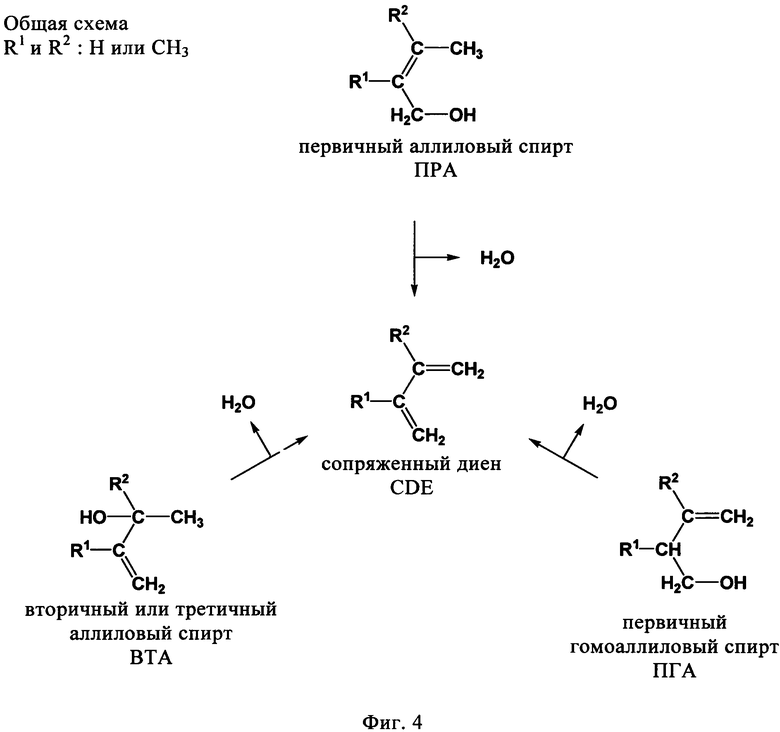
1. Способ получения сопряженного диена, отличающийся тем, что указанный способ включает стадию ферментативного превращения соединения общей формулы СnН2nO в соединение общей формулы CnH2n-2+Н2О, где 3<n<7, с использованием алкенол-дегидратазы, представляющей собой линалоол-дегидратазу (ЕС 4.2.1.127).
2. Способ по п.1, где указанным соединением общей формулы CnH2nO, где 3<n<7, является первичный аллиловый спирт (ПРА) формулы I:
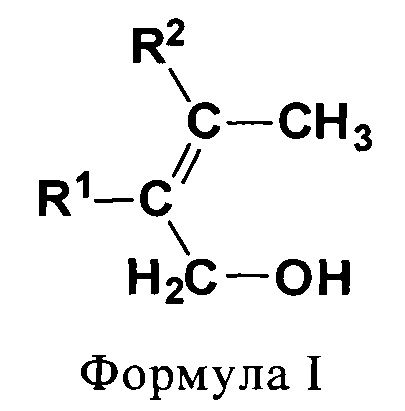
где R1 и R независимо выбирают из Н и СН3.
3. Способ по п.1, где указанным соединением общей формулы CnH2nO, где 3<n<7, является вторичный или третичный аллиловый спирт (ВТА) формулы II:
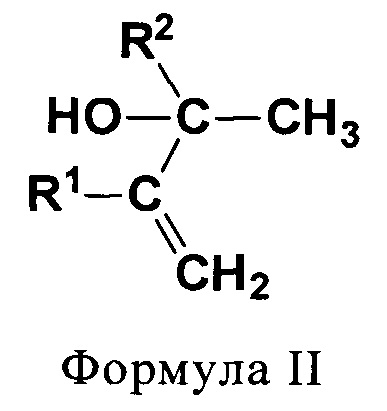
где R1 и R2 независимо выбирают из Н и СН3.
4. Способ по п.1, где указанным соединением общей формулы CnH2nO, где 3<n<7, является первичный гомоаллиловый спирт (ПГА) формулы III:
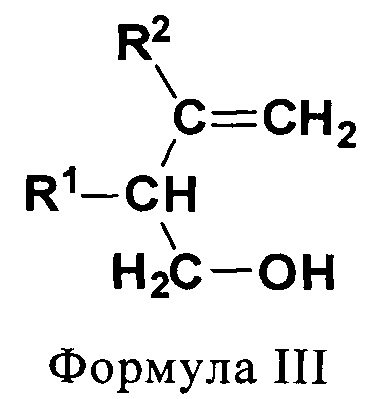
где R1 и R2 независимо выбирают из H и CH3.
5. Способ по любому из пп.1-4, который осуществляют in vitro.
6. Способ по любому из пп.1-4, отличающийся тем, что указанный способ осуществляют с использованием микроорганизма, продуцирующего алкенол-дегидратазу, представляющую собой линалоол-дегидратазу (ЕС 4.2.1.127).
7. Способ по п.6, где микроорганизм способен продуцировать соединение общей формулы CnH2nO, где 3<n<7.
8. Применение микроорганизма, продуцирующего алкенол-дегидратазу, представляющую собой линалоол-дегидратазу (ЕС 4.2.1.127), для превращения соединения общей формулы CnH2nO в соединение общей формулы CnH2n-2+H2O, где 3<n<7.
9. Композиция для получения соединения общей формулы CnH2n-2+H2O, включающая микроорганизм, который продуцирует алкенол-дегидратазу, представляющую собой линалоол-дегидратазу (ЕС 4.2.1.127), и соединение общей формулы CnH2nO, где 3<n<7.
10. Композиция для получения соединения общей формулы CnH2n-2+H2O, включающая алкенол-дегидратазу, представляющую собой линалоол-дегидратазу (ЕС 4.2.1.127) и соединение общей формулы CnH2nO, где 3<n<7.
11. Способ по любому из пп.1-4 или п.7, где алкенол-дегидратазой является белок, который включает аминокислотную последовательность, показанную в SEQ ID NO: 1, которая проявляет ферментативную активность, обеспечивающую превращение соединения общей формулы CnH2nO в соединение общей формулы CnH2n-2+H2O, где 3<n<7.
12. Применение по п.8, где алкенол-дегидратазой является белок, который включает аминокислотную последовательность, показанную в SEQ ID NO: 1, которая проявляет ферментативную активность, обеспечивающую превращение соединения общей формулы CnH2nO в соединение общей формулы CnH2n-2+H2O, где 3<n<7.
13. Композиция по п.9 или 10, где алкенол-дегидратазой является белок, который включает аминокислотную последовательность, показанную в SEQ ID NO: 1, которая проявляет ферментативную активность, обеспечивающую превращение соединения общей формулы СnН2nO в соединение общей формулы CnH2n-2+H2O, где 3<n<7.
14. Способ по любому из пп.1-4 или 7, где n равен 4, а получаемым диеновым соединением является бутадиен.
15. Применение по п.8, где n равен 4, а получаемым диеновым соединением является бутадиен.
16. Композиция по п.9 или 10, где n равен 4.
17. Способ по любому из пп.1-4 или 7, где n равен 5, а получаемым диеновым соединением является изопрен.
18. Применение по п.8, где n равен 5, а получаемым диеновым соединением является изопрен.
19. Композиция по п.9 или 10, где n равен 5
20. Способ по любому из пп.1-4 или 7, где n равен 6, а получаемым диеновым соединением является диметилбутадиен.
21. Применение по п.8, где n равен 6, а получаемым диеновым соединением является диметилбутадиен.
22. Композиция по п.9 или 10, где n равен 6.
| WO2009076676 A1, 18.06.2009 | |||
| BRODKORB ET AL | |||
| "Linalool dehydratase-isomerase, a bifunctional enzyme in the anaerobic degradation of monoterpenes", THE JOURNAL OF BIOLOGICAL CHEMISTRY, 2010, vol | |||
| ПЕРЕДВИЖНАЯ ДИАГРАММА ДЛЯ СРАВНЕНИЯ ЦЕННОСТИ РАЗЛИЧНЫХ ПРОДУКТОВ ПО ИХ КАЛОРИЙНОСТИ | 1919 |
|
SU285A1 |
| VAVILOV ET AL, "Synthesis of isoprene from 1,3-dioxolane and isobutylene", RUSSIAN JOURNAL OF APPLIED CHEMISTRY, 2010, vol | |||
| Пуговица | 0 |
|
SU83A1 |
| Аппарат для получения парогазовой смеси высокого давления | 1925 |
|
SU1598A1 |
| LUDDEKE ET AL | |||
| "Geraniol and geranial dehydrogenases induced in anaerobic monoterpene degradation by Castellaniella defragrans", APPLIED AND ENVIRONMENTAL MICROBIOLOGY, 01.2012, vol | |||
| Парный автоматический сцепной прибор для железнодорожных вагонов | 0 |
|
SU78A1 |
| Зажим для каучуковых трубок | 1921 |
|
SU2128A1 |
| ВАВИЛОВ Д.И | |||
| и др | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

