Данное изобретение относится к молекуле биспецифического (моноклонального) антитела, у которой первый связывающий домен, связывает антиген на CD8+ Т-клетках, не существующий в естественных условиях в и/или на CD8+ Т-клетках, и второй связывающий домен, связывает опухолеспецифический антиген, существующий в естественных условиях на поверхности опухолевых клеток. Кроме того, предложена последовательность нуклеиновой кислоты, кодирующей молекулу (моноклонального) биспецифического антитела по изобретению. Другими аспектами изобретения являются векторы и клетки-хозяева, содержащие указанную последовательность нуклеиновой кислоты, способ получения молекулы (биспецифического) антитела по изобретению и лекарственный препарат/композиция, содержащие указанную молекулу (биспецифического) антитела. Кроме того, изобретение относится к трансдуцированным CD8+ Т-клеткам, содержащим антиген, который не существует в естественных условиях в и/или на CD8+ Т-клетках, и/или Т-клеточный рецептор. В изобретении также предложено применение указанных молекул (биспецифических) антител в способе лечения конкретных заболеваний, а также фармацевтическая композиция/лекарственный препарат, содержащие указанные молекулы (биспецифических) антител, в которых указанные молекулы (биспецифических) антител вводят в составе определенного режима терапии в комбинации с трансдуцированными CD8+ Т-клетками, содержащими антиген, который не существует в естественных условиях в и/или на CD8+ Т-клетках, и/или Т-клеточный рецептор. В изобретении также предложен способ лечения конкретных заболеваний и набор, содержащий молекулу (биспецифического) антитела по изобретению.
ПРЕДШЕСТВУЮШИЙ УРОВЕНЬ ТЕХНИКИ
Исследовали возможность лечения рака и хронических инфекционных заболеваний при помощи трансфузии Т-клеток (т.е. Т-лимфоцитов), или адоптивной Т-клеточной терапии. Адоптивная Т-клеточная терапия способна усиливать противоопухолевый иммунитет, повышать эффективность вакцин и ослаблять реакцию «трансплантат против хозяина». В качестве источника клеток для адоптивной Т-клеточной терапии используют в том числе цитотоксические Т-клетки (ЦТЛ), или инфильтрирующие опухоль лимфоциты (ИОЛ). Биспецифические антитела можно применять для "вооружения" (активированных) Т-клеток для образования мостика между ними и поверхностным антигеном опухолевых клеток. Биспецифические антитела, нацеленные с одной стороны на поверхностный маркер/антиген опухолевых клеток и с другой стороны на другой маркер/антиген, который естественным образом/эндогенно экспрессируется в или на клетках, описаны, например, Glorius et al., Blood 116 (2010), 1173; Rothe et al., Blood 118 (2011), 1585; Zhengxing et al., Blood 111 (2007), 2211-2219, Herrmann et al., Cancer Research 68 (2008), 1221-1227; Singer et al., Journal of Immunotherapy 33 (2010), 599-608; Brandl et al., Experimental Hematology 27 (1999), 1264-1270; James et al., European Journal of Cancer 35 (1999), S343-S344; Chen et al., Clinical Cancer Research 1 (1995), 1319-1325; Valera et al., Molecular Cancer Therapeutics 9 (2010), 1872-1883; Gelderman et al., European Journal of Immunology 36 (2006), 977-984; Schweizer et al., Cancer Immunology Immunotherapy 51 (2002), 621-629; Friedman et al., Biotechnology and Applied Biochemistry 54 (2009), 121-131; Schaefer et al., Cancer Cell 20 (2011), 472-486 и Kazuhiko et al., International Journal of Molecular Medicine 25 (2010), 209-215.
Известно, что антиген-специфические цитотоксические Т-клетки (ЦТЛ) обладают способностью уничтожать человеческие опухолевые клетки, что показывает регрессия опухоли после адоптивного переноса пациентам с меланомой размноженных ex-vivo инфильтрирующих опухоль лимфоцитов (ИОЛ) или Т-клеток, трансфецированных геном Т-клеточного рецептора (Leen et al., Annu. Rev. Immunol. 115 (2007), 98-104). Известным альтернативным подходом является применение биспецифических антител для перенацеливания большого количества эндогенных Т-клеток. Эти биспецифические антитела, некоторые типы которых обозначаются BiTE (от англ "bispecific T-cell engager") были сконструированы так, что с одной стороны они направлены против поверхностного маркера CD3 (существующего в естественных условиях/эндогенно экспрессирующегося Т-клетками), а с другой стороны - против поверхностного антигена на опухолевых клетках (который естественным образом/эндогенно экспрессируется на поверхности опухолевых клеток). Кроме того, в более ранних работах было показано, что биспецифические антитела данного класса, направленные против CD3 и целевого антигена, обладают исключительно высокой активностью и способны заставить CD8+ Т-клетки и CD4+ Т-клетки лизировать опухолевые клетки при очень низком соотношении эффекторных клеток и клеток-мишеней (Е:Т). В настоящее время проходят клинические исследования двух антител BiTE: блинатумомаб (Blinatumomab, также обозначается МТ103) является биспецифическим антителом к CD3 и CD19. В настоящее время он проходит исследования I фазы у пациентов с рецидивирующей неходжкинской лимфомой (NHL) поздней стадии (Bargou et al., Science 321 (2008), 974-977) и исследования II фазы у пациентов с В-клеточным острым лимфобластным лейкозом (B-ALL) (Торр et al., Blood 112 (2008), 1926). Вторым антителом BiTE, проходящим исследования I фазы, является МТ-110 (Micromet Inc), распознающее карцинома-ассоциированный антиген, адгезивную молекулу эпителиальных клеток (ЕрСАМ, или CD326) и CD3 (Brischwein et al., Mol. Immunol. 43 (2006), 1129-1143). Одно биспецифическое антитело (катумаксомаб [Ремоваб®]; специфическое к CD3 и ЕрСАМ человека) получило разрешение на продажу в Европе в 2009.
In vitro и в модели на мышах биспецифические антитела способны соединять Т-клетки и опухолевые клетки за счет одновременного связывания CD3 и целевого антигена, что запускает активацию Т-клеток, включая слияние цитотоксических гранул и транзиторное высвобождение цитокинов и гранзимов. Однако, активация большого числа Т-клеток (независимо от специфичности Т-клеточного антигена) и/или эффект "свидетеля" лизиса опухолевых клеток представляют серьезные проблемы при использовании биспецифических антител у человека, например, в составе терапевтического режима.
Одной из таких проблем является так называемый "синдром высвобождения цитокинов" (СВЦ), который в модели на мышах обычно не вызывает побочных эффектов, но может вызывать катастрофические побочные эффекты у людей (Suntharalingam et al., The New England Journal of Medicine 355 (2006), 1018-1028). СВЦ включает головную боль, миалгию, тошноту, диарею, эритему, вазодиляцию и гипотензию. Наиболее серьезные формы приводят к образованию инфильтрата в легких, повреждению легких, почечной недостаточности и диссеминированному внутрисосудистому свертыванию (Suntharalingam et al., The New England Journal of Medicine 355 (2006), 1018-1028). Даже если СВЦ не связан с применением биспецифических антител, при введении биспецифических антител наблюдалось значительное количество других побочных эффектов (частота возникновения нежелательных явлений третьей степени и выше составила более 80%) (Торр et al., Journal of Clinical Oncology 29 (2011), 2093-2098). Такие побочные эффекты связывают с вовлечением Т-клеток, они включают лимфопению, изменение биохимических показателей крови и развитие неврологической симптоматики.
Вследствие большого количества побочных эффектов у биспецифических антител невозможно применять типы антител с длительным периодом полужизни, поскольку в случае СВЦ длительный период полужизни является нежелательным.
Таким образом, технической проблемой, решение которой предложено в данном изобретении, является разработка способов и средств для лечения онкологических заболеваний, таких как злокачественные новообразования эпителиального, эндотелиального или мезотелиального происхождения и злокачественные новообразования кроветворной ткани, за счет индукции иммунного ответа, опосредованного Т-клетками.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Упомянутые выше способы и средства для лечения онкологических заболеваний, таких как злокачественные новообразования эпителиального, эндотелиального или мезотелиального происхождения и злокачественные новообразования кроветворной ткани за счет индукции иммунного ответа, опосредованного Т-клетками, должны позволить преодолеть указанные выше недостатки терапии, основанной на использовании известных биспецифических антител.
Решение указанной технической проблемы достигается благодаря воплощениям, описанным в формуле изобретения.
Настоящее изобретение относится к трансдукции CD8+ Т-клеток маркерным белком, который не существует в естественных условиях в и/или на поверхности CD8+ Т-клеток, и их направленной мобилизации в опухоль с помощью молекул биспецифических антител (см. Фиг. 7 и 21). В контексте настоящего изобретения трансдукцию CD8+ Т-клеток можно осуществлять с использованием ретровирусных систем, как описано ниже. Настоящее изобретение относится к молекуле биспецифического антитела, содержащей первый связывающий домен, специфически связывающийся с антигеном на CD8+ Т-клетках, не существующим в естественных условиях в и/или на CD8+ Т-клетках, и второй связывающий домен, связывающийся с опухолеспецифическим антигеном, существующим в естественных условиях на поверхности опухолевых клеток, при этом указанные CD8+ Т-клетки были трансдуцированы антигеном, не существующим в естественных условиях в и/или на CD8+ Т-клетках. В контексте настоящего изобретения молекула биспецифического антитела, содержащая первый связывающий домен, специфически связывающийся с антигеном на CD8+ Т-клетках, не существующим в естественных условиях в и/или на CD8+ Т-клетках, и второй связывающий домен, связывающийся с опухолеспецифическим антигеном, существующим в естественных условиях на поверхности опухолевых клеток, представляет собой молекулу (моноклонального) антитела. Как показано в приведенных ниже Примерах, в качестве доказательства концепции изобретения, было создано биспецифическое антитело, у которого первый связывающий домен взаимодействует/связывается с (человеческим) EGFR (представляющим собой антиген, не встречающийся в естественных условиях в или на Т-клетках (CD8+ Т-клетках) и второй связывающий домен взаимодействует/связывается с ЕрСАМ (представляющим собой опухолеспецифический антиген, существующий в естественных условиях на поверхности опухолевых клеток). Лечение опухолей с использованием комбинации этого биспецифического антитела и трансдуцированных опухолеспецифических Т-клеток (CD8+ Т-клеток), экспрессирующих (человеческий) белок del-hEGFR, значительно увеличивает выживаемость-мышей-гю-сравнению-с-контрольными-экспериментальными животными (см. Фиг. 12 и 14). Неожиданно было обнаружено, что Т-клетки (CD8+ Т-клетки), трансдуцированные антигеном (в приведенных ниже Примерах (подтверждающих концепцию) - (человеческим) белком del-hEGFR с последовательностью, показанной в SEQ ID NO: 12 (кодируемой кДНК, последовательность которой показана в SEQ ID NO: 11)), не существующим в естественных условиях в и/или на поверхности этих клеток, могут быть специфически мобилизованы с использованием молекулы биспецифического антитела, связывающейся первым связывающимся доменом с антигеном, не встречающимся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках) (человеческим) hEGFR), который был введен в указанные Т-клетки (CD8 Т-клетки) и связывающейся вторым связывающим доменом с опухолеспецифическим антигеном (ЕрСАМ), существующим в естественных условиях на поверхности опухолевых клеток.
В этом контексте термин "биспецифическая связывающая конструкция", используемый в данном документе, в частности относится к молекуле биспецифического антитела, способной связываться с антигеном, в естественных условиях/эндогенно не экспрессирующимся в или на CD8+ Т-клетках, и способной индуцировать элиминацию/лизис клеток-мишеней (за счет второго связывающего домена, связывающегося с опухолеспецифическим антигеном, существующим в естественных условиях (то есть эндогенно экспрессирующимся) на поверхности опухолевых клеток). Связывание антигена, не существующего в естественных условиях в и/или на CD8+ Т-клетках (например, антитела, производных антитела или фрагментов антитела), опосредованное через биспецифическую связывающую конструкцию (молекулу биспецифического антитела), обеспечивает физический контакт между опухолеспецифическими Т-клетками (CD8+ Т-клетками) и опухолевой клеткой (см. Фиг. 7 и 21). На нетрансдуцированные или эндогенные Т-клетки (CD8+ Т-клетки) влияние биспецифической связывающей конструкции (молекулы биспецифического антитела) не распространяется. Следовательно, молекула биспецифического антитела по изобретению способна вызывать лизис клеток-мишеней in vivo и/или in vitro. Соответствующие клетки-мишени охватывают клетки, экспрессирующие поверхностную молекулу, которая распознается вторым (lg-производным) связывающим доменом молекул биспецифического антитела по изобретению. Такие поверхностные молекулы описаны ниже.
Лизис клеток-мишеней можно детектировать способами, известными в области техники. Такие способы включают, среди прочих, физиологические исследования in vitro. Такие физиологические исследования позволяют отслеживать гибель клеток, например, по потере целостности мембран (например, метод на основе FACS-анализа с использованием пропидия иодида, окрашивание трипановым синим, фотометрическое исследование высвобождения ферментов (ЛДГ), радиометрическое исследование высвобождения 51Cr, флуориметрическое исследование высвобождения Европия и высвобождение кальцеина-АМ). Другие исследования включают определение жизнеспособности клеток, например, фотометрическое исследование с использованием МТТ, ХТТ, WST-1 и аламарового синего, радиометрическое исследование включения 3H-тимидина, клоногенные исследования, позволяющие зарегистрировать клеточные деления, а также флуориметрические исследования с использованием родамина123, позволяющие измерить трансмембранный потенциал митохондрий. Кроме того, можно оценивать апоптоз, например, регистрируя экспонирование фосфатидилсерина методом FACS, с помощью метода TUNEL на основе ИФА, определяя активность каспазы (фотометрически, флуориметрически или в формате ИФА) или анализируя изменения клеточной морфологии (сморщивание, «вспенивание» мембран (блеббинг)).
Термин "связывание с" в контексте данного изобретения обозначает связывание (взаимодействие) друг с другом по меньшей мере двух "сайтов, взаимодействующих с антигеном". Термин "сайт, взаимодействующий с антигеном" обозначает в данном изобретении мотив или полипептид, обладающий способностью специфически взаимодействовать со специфическим антигеном или группой специфических антигенов. Указанное связывание/взаимодействие также подразумевает "специфическое распознавание". Термин "специфически распознающий" означает в данном изобретении, что конструкция антитела способна специфически взаимодействовать и/или связываться по меньшей мере с двумя аминокислотами каждой из человеческих молекул-мишеней, согласно описанию в данном документе. Антитела могут распознавать, взаимодействовать и/или связываться с различными эпитопами одной и той же молекулы-мишени. Данный термин относится к специфичности молекулы антитела, т.е, к его способности различать специфические участки человеческой молекулы-мишени, согласно описанию в данном документе. Специфическое взаимодействие сайтов, взаимодействующих с антигенами, с их специфическим антигенами может инициировать сигнал, например, вследствие изменения конформации антигена, олигомеризации антигена и т.д. Таким образом, специфический мотив аминокислотной последовательности сайта, взаимодействующего с антигеном, и антиген связываются друг с другом благодаря своей первичной, вторичной или третичной структуре, а также в результате вторичных модификаций указанной структуры.
Термин "специфическое взаимодействие", используемый в данном изобретении, означает, что биспецифическая связывающая конструкция (молекула биспецифического антитела) по изобретению перекрестно не реагирует или по существу перекрестно не реагирует с (поли)пептидами схожих структур. Следовательно, биспецифическая конструкция по изобретению специфически связывается/взаимодействует с опухолевыми маркерами, маркерами клеточной поверхности, антигенами, которые не существуют в естественных условиях в и/или на CD8+ Т-клетках, и способна благодаря своему второму (lg-производному) домену взаимодействовать с другими избранными специфическими соединениями, антигенами, маркерами клеточной поверхности, опухолевыми маркерами и т.д., которые не существуют в естественных условиях на поверхности опухолевых клеток. Конкретные примеры таких молекул, против которых направлены указанные первый и второй lg-производные домены, приведены ниже.
Перекрестную реактивность панели исследуемых конструкций можно исследовать, например, оценивая связывание конструкций биспецифических антител из указанной панели в стандартных условиях (см., например, Harlow andLane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, (1988) and Using Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, (1999)) с представляющим интерес (поли)пептидом, а также с рядом более или менее (структурно и/или функционально) близкородственных (поли)пептидов. Только те конструкции (т.е. антитела, (биспецифические) scFv и т.п.), которые связываются с (поли)пептидом/белком, представляющим интерес, но не связываются или по существу не связываются с каким-либо другим (поли)пептидом, экспрессирующимся в той же ткани, что и (поли)пептид, представляющий интерес, например, в клетках опухолевой ткани, считаются специфическими в отношении (поли)пептида/белка, представляющего интерес, и отбираются для дальнейших исследований в соответствии со способом, приведенным в данном документе. Эти способы могут включать, среди прочих, исследование связывания, блокирования и конкуренции структурно и/или функционально близкородственных молекул. Эти исследования связывания также включают FACS-анализ, поверхностный плазмонный резонанс (SPR, например, BIAcore®), аналитическое ультрацентрифугирование, изотермическую титрационную калориметрию, анизотропию флуоресценции, флуоресцентную спектроскопию или исследования связывания с радиоактивными лигандами. Кроме того, можно выполнять физиологические исследования, например, цитотоксические исследования и исследования, упомянутные выше. Таким образом, примеры такого специфического взаимодействия между сайтом, взаимодействующим с антигеном, и специфическим антигеном могут включать специфичность лиганда к своему рецептору. Указанное определение в частности охватывает взаимодействие лигандов, генерирующее сигнал при связывании с их специфическим рецептором. Примеры соответствующих лигандов включают цитокины, которые взаимодействуют/связываются со своими специфическими цитокиновыми рецепторами. Указанное определение также охватывает связывание сайта, взаимодействующего с антигеном, с антигенами, например, антигенами из семейства селектинов, интегринами и антигенами семейства ростовых факторов, например, EGF. Другим примером указанного взаимодействия, охватываемого указанным определением, является взаимодействие антигенной детерминанты (эпитопа) с антигенсвязывающим сайтом антитела.
Термин "связывание с" относится не только к линейному эпитопу, но может также относиться к конформационному эпитопу, структурному эпитопу или эпитопу прерывистого типа, состоящему из двух участков человеческих молекул-мишеней или их частей. В контексте данного изобретения конформационный эпитоп характеризуется двумя или более отдельными последовательностями аминокислот, разделенными в первичной последовательности, которые оказываются вблизи друг от друга на поверхности молекулы при сворачивании полипептида в нативный белок (Sela, Science 166 (1969), 1365 и Laver, Cell 61 (1990), 553-536). Кроме того, термин "связывание с" используется взаимозаменяемо в контексте данного изобретения с термином "взаимодействие с".
Таким образом, специфичность можно определять экспериментально способами, известными в области техники, и способами, приведенными в данном описании. Такие способы включают вестерн-блоттинг, ИФА, РИА, электрохемилюминесценцию, иммунорадиометрическое исследование и пептидное сканирование.
Термин (Ig-производный) "первый связывающий домен" относится к "домену, являющемуся производным иммуноглобулина", в частности, к антителу или его фрагменту, к одноцепочечным антителам, к синтетическим антителам, к фрагментам антител, таким как фрагменты Fab, F(ab2)ʹ, Fv или scFv и т.д., или химически модифицированным производным любого из них. Молекулы таких антител могут происходить от различных биологических видов или могут быть химерными. В контексте данного изобретения (как проиллюстрировано в приведенных Примерах), указанный (lg-производный) первый домен, содержащийся в молекуле биспецифического антитела по изобретению, может представлять собой (моноклональное) антитело, с которым слит второй (lg-производный) "связывающий домен".
Термин (lg-производный) "второй связывающий домен" относится к домену, являющемуся производным иммуноглобулина, в частности, к антителу или его фрагменту, к одноцепочечным антителам, к синтетическим антителам, к фрагментам антител, таким как фрагменты Fab, F(ab2)ʹ, Fv или scFv и т.д., или химически модифицированным производным любого из них. Молекулы таких антител могут происходить от различных биологических видов или могут быть химерными. В контексте данного изобретения (как проиллюстрировано в приведенных Примерах), указанный (Ig-производный) второй домен, содержащийся в молекуле биспецифического антитела по изобретению, может представлять собой scFv.
Молекулы биспецифических антител по изобретению представляют собой (моноклональные) биспецифические антитела, которые имеют связывающие детерминанты по меньшей мере для двух различных сайтов и могут относиться к любому типу. В последнее время было разработано большое разнообразие типов рекомбинантных антител, например, бивалентные, тривалентные или тетравалентные биспецифические антитела. Примеры включают слияние антител типа IgG и одноцепочечных доменов (различные типы описаны, например, Coloma, M.J., et al., Nature Biotech 15 (1997), 159-163; WO 2001/077342; Morrison, S.L., Nature Biotech 25 (2007), 1233-1234; Holliger, P., et. al, Nature Biotech. 23 (2005), 1126-1136; Fischer, N., and Léger, O., Pathobiology 74 (2007), 3-14; Shen, J., et. al., J. Immunol. Methods 318 (2007), 65-74; Wu, C, et al., Nature Biotech. 25 (2007), 1290-1297). Биспецифическое антитело или его фрагменты также охватывают бивалентные, тривалентные или тетравалентные биспецифические антитела, описанные в WO 2009/080251; WO 2009/080252; WO 2009/080253; WO 2009/080254; WO 2010/112193; WO 2010/115589; WO 2010/136172; WO 2010/145792; WO 2010/145793 и WO 2011/117330.
"Антитела" по данному изобретению имеют два или более связывающих домена и являются биспецифическими. Это означает, что антитела могут быть биспецифическими даже в случаях, когда у них имеется более двух связывающих доменов (т.е. антитело является тривалентным или мультивалентным). Биспецифические антитела по изобретению включают, например, мультивалентные одноцепочечные антитела, диатела и триатела, а также антитела, имеющие константные домены со структурой полноразмерных антител, с которыми другие антигенсвязывающие домены (например, одноцепочечные Fv, VH домен и/или VL домен, Fab или (Fab)2,) связаны при помощи одного или более пептидных линкеров. Антитела могут быть полноразмерными, происходящими от одного биологического вида, или быть химерными или гуманизированными. У антитела с более чем двумя антигенсвязывающими доменами, некоторые связывающие домены могут быть идентичными, при условии, что белок имеет связывающие домены для двух различных антигенов.
Термин "валентный" в данном описании обозначает присутствие определенного числа связывающих доменов в молекуле антитела. Таким образом, термины "бивалентный", "тетравалентный" и "гексавалентный" обозначают присутствие в молекуле антитела двух связывающих доменов, четырех связывающих доменов и шести связывающих доменов, соответственно. Биспецифические антитела согласно изобретению по меньшей мере являются "бивалентными" и могут быть "тривалентными" или "мультивалентными" (например, "тетравалентными" или "гексавалентными"). Предпочтительно, биспецифическое антитело по изобретению является бивалентным, тривалентным или тетравалентным. Соответственно, в контексте данного изобретения указанное биспецифическое антитело является бивалентным. В контексте данного изобретения указанное биспецифическое антитело является тривалентным. В контексте данного изобретения указанное биспецифическое антитело является тетравалентным.
Как указано выше (и показано на Фиг. 1), наиболее предпочтительно, молекула биспецифического антитела по изобретению, включает (Ig-производный) второй домен, который может представлять собой scFv. Таким образом, в воплощении данного изобретения, приведенном в качестве примера для подтверждения концепции, молекула биспецифического антитела имеет одну детерминанту для (человеческого) EGFR (в первом связывающем домене) и другую детерминанту, присутствующую во втором scFv, направленную против/способную взаимодействовать с другой молекулой/соединением. Эти другие молекулы/соединения могут содержать молекулы клеточной поверхности, опухолевые маркеры, опухолевые антигены и т.п. Примеры этих других соединений/молекул приведены ниже.
Следовательно, биспецифические связывающие молекулы в контексте данного изобретения могут относиться к молекулам антител, содержащим два связывающих домена, являющихся производными антитела, причем один связывающий домен может представлять собой scFv. Один из указанных связывающих доменов имеет в своем составе вариабельные участки (или их части) антитела, фрагмента антитела или его производного, способные специфически связываться/взаимодействовать с первой (человеческой) молекулой-мишенью, не встречающейся в естественных условиях в и/или на CD8+ Т-клетках (согласно описанию ниже). Второй связывающий домен содержит вариабельные участки (или их части) антитела, фрагмента антитела или его производного, способные специфически связываться/взаимодействовать с другим (человеческим) антигеном (вторая молекула-мишень), согласно описанию ниже. Следовательно, указанный второй связывающий домен согласно данному изобретению представляет собой (Ig-производный) второй домен, описанный выше, содержащий сайт, взаимодействующий с антигеном, специфический в отношении молекулы клеточной поверхности, существующей в естественных условиях на поверхности опухолевых клеток, или в отношении опухолеспецифического маркера (антигена), существующего в естественных условиях на поверхности опухолевых клеток. Предпочтительно, два указанных домена/участка молекулы биспецифического антитела ковалентно связаны друг с другом. Такое соединение может быть либо непосредственным (первый домен [специфический в отношении первой (человеческой) молекулы-мишени, не встречающейся в естественных условиях в или на CD8+ Т-клетках, включающий CDR-области или CDR-области и каркасные области, в соответствии с описанием выше] - второй домен [специфический в отношении молекулы клеточной поверхности и/или опухолеспецифического маркера] или первый домен [специфический в отношении молекулы клеточной поверхности и/или опухолеспецифического маркера] - второй домен [специфический в отношении первой (человеческой) молекулы-мишени, не встречающейся в естественных условиях в и/или на CD8+ Т-клетках, включая CDR-области или CDR-области и каркасные области, согласно описанию выше) или опосредованным дополнительной полипептидной линкерной последовательностью (первый домен - линкерная последовательность - второй домен). В случае использования линкера этот линкер в контексте данного изобретения имеет длину и последовательность, достаточные для того, чтобы каждый из первого и второго доменов независимо друг от друга сохранял различную специфичность связывания. В контексте данного изобретения дополнительная полипептидная линкерная последовательность может также представлять собой фрагмент антитела, которое само может быть, например, Fc-частью или одним или несколькими константными доменами антитела.
В контексте данного изобретения первый связывающий домен также может быть частью первого плеча антитела, а второй связывающий домен также может быть частью второго плеча антитела, или наоборот, при этом оба плеча антитела связаны в области контакта. Первое плечо антитела содержит вариабельные участки (или их части) антитела, фрагмента антитела или его производного, способные специфически связываться/взаимодействовать с первой (человеческой) молекулой-мишенью, не существующей в естественных условиях в или на CD8+ Т-клетках, согласно описанию ниже. Второе плечо антитела содержит вариабельные участки (или их части) антитела, фрагмента антитела или его производного, способные специфически связываться/взаимодействовать с молекулой клеточной поверхности, существующей в естественных условиях на поверхности опухолевой клетки, или с опухолеспецифическим антигеном, существующим в естественных условиях на поверхности опухолевой клетки. "Область контакта" содержит аминокислотные остатки (или другие группы, не являющиеся аминокислотами, например, углеводные группы) первого плеча антитела, взаимодействующие с одним или более "контактирующих" аминокислотных остатков (или другие группы, не являющиеся аминокислотами) в области контакта со вторым плечом антитела. Предпочтительной областью контакта является домен иммуноглобулина, например, константный домен (или его участки) тяжелых цепей антитела, причем связывание/взаимодействие с областью контакта обеспечивает гетеродимеризацию двух плечей антитела (см., например, Ridgway, J.В., et al., Protein Eng. 9 (1996), 617-621; WO 96/027011; Merchant, A.M., et al., Nature Biotech. 16 (1998), 677-681; Atwell, S., et al., J. Mol. Biol. 270 (1997), 26-35; ЕР 1 870 459 A1; WO 2007/147901; WO 2009/089004(A1) и WO 2010/129304).
Антитела, конструкции антител, молекулы биспецифических антител, фрагменты антител, производные антител (все являющиеся производными Ig), применяемые в соответствии с изобретением, или их соответствующие иммуноглобулиновые цепи можно также модифицировать с использованием стандартных методик, известных в области техники, например, при помощи делеции, вставки, замены, добавления аминокислот и/или рекомбинации и/или любых других модификаций, известных в области техники, как по-отдельности, так и в комбинации. Способы введения таких модификаций в последовательность ДНК, определяющую последовательность аминокислот цепи иммуноглобулина, хорошо известны специалисту в данной области техники; см., например, Sambrook (1989), loc. cit. Термин "домен, являющийся производным Ig" в частности относится к конструкциям (поли)пептидов, содержащим по меньшей мере один CDR. Фрагменты или производные указанных доменов, являющихся производными Ig, характеризуют (поли)пептиды, которые являются частями указанных выше молекул антител и/или которые модифицированы с использованием химических/биохимических или молекулярно-биологических способов. Соответствующие способы известны в области техники и описаны, в том числе, в лабораторных руководствах (см. Sambrook et al., Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press, 2 издание (1989) и 3 издание (2001); Gerhardt et al., Methods for General and Molecular Bacteriology ASM Press (1994); Lefkovits, Immunology Methods Manual: The Comprehensive Sourcebook of Techniques; Academic Press (1997); Golemis, Protein-Protein Interactions: A Molecular Cloning Manual Cold Spring Harbor Laboratory Press (2002)).
Термин "CDR", используемый в данном документе, относится к "участкам, определяющим комплементарность", хорошо известным в области техники. CDR являются частью иммуноглобулинов, определяющей специфичность указанных молекул и контактирующей со специфическим лигандом. CDR представляют собой наиболее вариабельные части молекулы и обусловливают разнообразие этих молекул. В каждом V домене находится три участка CDR: CDR1, CDR2 и CDR3. CDR-H обозначает участок CDR вариабельного домена тяжелой цепи, a CDR-L обозначает участок CDR вариабельного домена легкой цепи. VH означает вариабельный домен тяжелой цепи, a VL означает вариабельный домен легкой цепи. Участки CDR домена, являющегося производным Ig, могут быть идентифицированы, как описано Kabat "Sequences of Proteins of Immunological Interest", 5th edit. NIH Publication no. 91-3242 U.S. Department of Health and Human Services (1991); Chothia J. Mol. Biol. 196 (1987), 901-917 или Chothia Nature 342 (1989), 877-883.
Таким образом, в контексте данного изобретения молекула антитела, например, биспецифического антитела, описанная выше, выбрана из группы, состоящей из полноразмерного антитела (иммуноглобулина, например IgG1, IgG2, IgG2b, IgG3, IgG4, IgA, IgM, IgD или IgE), F(ab)-, Fabʹ-SH-, Fv-, Fabʹ-, F(abʹ)2-фрагмента, химерного антитела, антитела с пересаженными CDR, полностью человеческого антитела, конструкции бивалентного антитела, гибридного антитела, синтетического антитела, бивалентного одноцепочечного антитела, тривалентного одноцепочечного антитела и мультивалентного одноцепочечного антитела.
Термин "полностью человеческое антитело" в данном документе означает антитело, содержащее только белковые последовательности иммуноглобулина человека. Полностью человеческое антитело может содержать углеводные цепи мышиного происхождения, если оно синтезировано в организме мыши, мышиной клетке или гибридоме, полученной из мышиной клетки. Аналогично, "мышиное антитело" или "антитело мыши" обозначает антитело, которое содержит только белковые последовательности иммуноглобулина мыши. В альтернативном случае, "полностью человеческое антитело" может содержать углеводные цепи крысиного происхождения, если оно синтезировано в организме крысы, в крысиной клетке или гибридоме, полученной из крысиной клетки. Аналогично, термин "крысиное антитело" обозначает антитело, которое содержит только последовательности иммуноглобулина крысы. Полностью человеческие антитела можно получать, например, методом фагового дисплея, который представляет собой широко распространенную методику скрининга, позволяющую производить и выполнять скрининг полностью человеческих антител. Анитела, полученные методом фагового дисплея, могут также использоваться в контексте данного изобретения. Методы фагового дисплея описаны, например, в документах US 5,403,484, US 5,969,108 и US 5,885,793. Другая методика, позволяющая создавать полностью человеческие антитела, включает модификацию технологии получения мышиных гибридом. Создают трансгенных мышей, у которых собственные мышиные гены заменены локусами иммуноглобулинов человека (см., например, US 5,877,397).
Термин антитело, используемый в данном документе, также охватывает химерные антитела. Термин "химерные антитела" обозначает антитело, содержащее вариабельный участок человеческого происхождения или не являющийся человеческим, химеризованный с участком антитела (например, константным участком) человека или другого биологического вида (например, мыши, лошади, кролика, собаки, коровы, цыпленка).
Термин антитело также обозначает рекомбинантные человеческие антитела, гетерологичные антитела и гетерогибридные антитела. Термин "рекомбинантное человеческое антитело" охватывает все антитела с человеческой последовательностью, которые получают, экспрессируют, создают или выделяют рекомбинантными способами, например, антитела, выделенные из животного (например, мыши), являющегося трансгенным в отношении генов иммуноглобулинов человека; антитела, экспрессируемые с использованием рекомбинантных экспрессирующих векторов, которыми трансфецируют клетку-хозяина, антитела, выделенные из рекомбинантной комбинаторной библиотеки человеческих антител, или антитела, полученные, экспрессируемые, созданные или выделенные любым другим способом, включающим сплайсинг (сшивание) последовательностей генов иммуноглобулинов человека с другими последовательностями ДНК. В таких рекомбинантных человеческих антителах вариабельные и константные участки (если таковые существуют) получены на основе последовательности генов иммуноглобулинов зародышевой линии человека. При этом, такие антитела могут быть подвергнуты мутагенезу in vitro (или при использовании трансгенного животного с последовательностями иммуноглобулинов человека, соматическому мутагенезу in vivo) и, таким образом, аминокислотные последовательности участков VH и VL рекомбинантных антител представляют собой последовательности, которые, будучи полученными на основе и родственными последовательностям VH и VL в зародышевой конфигурации, могут в естественных условиях не встречаться в составе репертуара первичных антител человека in vivo.
"Гетерологичное антитело" относится к отличному от человека трансгенному организму, продуцирующему такое антитело. Данный термин обозначает антитело, у которого аминокислотная последовательность или кодирующая его нуклеотидная последовательность соответствуют последовательности, обнаруживаемой у организма, который не является отличным от человека трансгенным животным, и как правило, происходит от видов, отличных от трансгенного животного, не являющегося человеком.
Термин "гетерогибридное антитело" обозначает антитело, имеющее легкую и тяжелую цепи, происходящие из различных организмов. Например, антитело, у которого человеческая тяжелая цепь связана с мышиной легкой цепью, представляет собой гетерогибридное антитело. Примеры гетерогибридных антител включают химерные и гуманизированные антитела.
Термин антитело также относится к гуманизированным антителам. "Гуманизированные" формы антител, не являющихся человеческими, (например, мышиных или кроличьих) представляют собой химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (например, Fv, Fab, Fabʹ, F(abʹ)2 или другие антигенсвязывающие последовательности антител), которые содержат минимальные последовательности, полученные из иммуноглобулинов, не являющихся человеческими. Зачастую гуманизированные антитела представляют собой человеческие иммуноглобулины (антитела-реципиенты), в которых остатки участка, определяющего комплементарность (CDR) заменены остатками CDR биологического вида, отличного от человека (антитело-донор), например, мыши, крысы или кролика, обладающими необходимой специфичностью, аффинностью и активностью. В некоторых случаях каркасные остатки Fv иммуноглобулина человека заменяют соответствующими остатками, не являющимися человеческими. Кроме того, гуманизированное антитело может содержать остатки, которые не обнаруживаются ни в составе антитела-реципиента, ни в импортированных последовательностях CDR или каркасных последовательностях. Данные модификации осуществляют для дальнейшего улучшения и оптимизации свойств антитела. Как правило, гуманизированное антитело содержит по существу все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или по существу все CDR участки соответствуют CDR участкам иммуноглобулинов, не являющихся человеческими, и все или по существу все FR участки представляют собой FR участки консенсусной последовательности иммуноглобулина человека. Гуманизированное антитело может также содержать по меньшей мере часть константного участка иммуноглобулина (Fc), обычно иммуноглобулина человека. Более подробно это описано в публикациях Jones Nature 321 (1986), 522-525; Reichmann Nature 332 (1998), 323-327 и Presta Curr Op Struct Biol 2 (1992), 593-596.
Распространенный способ гуманизации антител включает пересаживание CDR, в котором функциональный антигенсвязывающий сайт антитела-донора, не являющегося человеческим, пересаживают на человеческое антитело-акцептор. Способы пересаживания CDR известны в области техники и описаны, например, в публикациях US 5,225,539, US 5,693,761 и US 6,407,213. Другой схожий способ представляет собой получение гуманизированных антител с использованием трансгенных животных, генетически модифицированных таким образом, что они имеют один или более гуманизированных локусов иммуноглобулинов, способных претерпевать перестройку (реаранжировку) генных сегментов и конверсию генов (см., например, US 7,129,084).
Таким образом, в контексте данного изобретения термин "антитело" охватывает целые молекулы иммуноглобулинов, а также части таких молекул иммуноглобулинов. Кроме того, как обсуждалось выше, термин охватывает модифицированные и/или измененные молекулы антител. Термин также охватывает антитела, полученные рекомбинантным способом или при помощи синтеза. Термин также охватывает интактные антитела, а также фрагменты антител, например, отдельные легкие и тяжелые цепи, Fab, Fab/c, Fv, Fabʹ, F(abʹ)2. Термин "антитело" также охватывает бифункциональные антитела, трифункциональные антитела, полностью человеческие антитела, химерные антитела, гуманизированные антитела и конструкции антител, такие как одноцепочечные Fv (scFv) или гибридные антитела.
В контексте данного изобретения "одноцепочечные Fv" или "scFv" фрагменты антитела имеют VH и VL домены антитела, причем эти домены находятся в составе одной полипептидной цепи. Как правило, полипептид scFv также включает полипептидный линкер, расположенный между VH и VL доменами, что позволяет scFv образовывать структуру, необходимую для связывания антигена. Методики получения одноцепочечных антител описаны, например, Plückthun in The Pharmacology of Monoclonal Antibodies, Rosenburg and Moore eds. Springer-Verlag, N.Y. 113 (1994), 269-315.
"Fab фрагмент" в данном описании состоит из одной легкой цепи, а также СН1 и вариабельного участка одной тяжелой цепи. Тяжелая цепь молекулы Fab не может образовывать дисульфидной связи с другой тяжелой цепью молекулы.
Участок "Fc" имеет в своем составе фрагменты двух тяжелых цепей, содержащие СН2 и СН3 домены антитела. Фрагменты двух тяжелых цепей удерживаются благодаря двум или более дисульфидным связям и гидрофобным взаимодействиям между СН3 доменами.
"Fabʹ фрагмент" имеет в своем составе одну легкую цепь и часть одной тяжелой цепи, содержащую VH домен и СН1 домен, а также участок между СН1 и СН2 доменами, что позволяет формироваться дисульфидной связи между двумя тяжелыми цепями двух Fabʹ фрагментов с образованием молекулы F(abʹ)2.
"F(abʹ)2 фрагмент" имеет в своем составе две легкие цепи и две тяжелые цепи, содержащие часть константного участка, расположеную между CH1 и CH2 доменами, что позволяет формироваться межцепочечной дисульфидной связи между двумя тяжелыми цепями. Таким образом, F(abʹ)2 фрагмент состоит из двух Fabʹ фрагментов, соединенных между собой дисульфидной связью между двумя тяжелыми цепями.
"Fv участок" содержит вариабельные участки тяжелой и легкой цепи, но не имеет константных участков.
Следует отметить, что молекула биспецифического антитела по изобретению может содержать, кроме описанных здесь первого (Ig-производного) домена и второго (Ig-производного) домена дополнительный(е) домен(ы), например, необходимые для выделения и/или получения рекомбинантных конструкций.
Следует отметить, что согласно данному изобретению, можно модифицировать не только описанный выше первый домен молекулы или конструкции по изобретению (т.е. молекулы биспецифического антитела, описанной в данном документе), специфически взаимодействующий/связывающийся с (человеческим) антигеном на CD8+ Т-клетках, не существующим в естественных условиях в и/или на CD8+ Т-клетках.
Также предусматриваются модификации первого (Ig-производного) домена, второго (Ig-производного) домена и/или соединяющего(их) их линкерного(ых) участка(ков), например, гуманизированного антитела, антитела с пересаженными CDR или полностью человеческого антитела.
"Принципы гуманизации" хорошо известны в области техники и, в частности, описаны для молекул антител, например, молекул, являющихся производными иммуноглобулинов (Ig). Термин "гуманизированное" обозначает гуманизированные формы антител, не являющихся человеческими (например, мышиных), или их фрагментов (таких как Fv, Fab, Fabʹ, F(abʹ), scFvs или других последовательностей антигенсвязывающих частей антител), имеющих в своем составе некоторую часть последовательности, полученную из антитела, не являющегося человеческим. Гуманизированные антитела включают человеческие иммуноглобулины, в которых остатки участков, определяющих комплементарность (CDR), иммуноглобулина человека замещены остатками CDR видов, отличных от человека, таких как мышь, крыса или кролик, обладающими желаемой специфичностью, аффинностью и активностью. Как правило, гуманизированное антитело будет содержать по существу все из по меньшей мере одного, и обычно двух вариабельных доменов, в которых все или по существу все участки CDR соответствуют участкам CDR иммуноглобулина, не являющегося человеческим, и все или по существу все участки FR представляют собой участки FR консенсусной последовательности иммуноглобулина человека. Гуманизированное антитело может также содержать по меньшей мере часть константного участка иммуноглобулина (Fc), обычно человеческого иммуноглобулина; см. в том числе Jones et al., Nature 321 (1986),522-525, Presta, Curr. Op. Struct. Biol. 2 (1992), 593-596. Способы гуманизации антител, не являющихся человеческими, хорошо известны в области техники. Как правило, гуманизированное антитело имеет одну или несколько введенных в него аминокислот, происходящих из источника, не являющегося человеком, сохраняющих исходную связывающую активность антитела. Способы гуманизации антител/молекул антител более подробно описаны Jones et al., Nature 321 (1986), 522-525; Reichmann et al., Nature 332 (1988), 323-327 и Verhoeyen et al., Science 239 (1988), 1534-1536. В области техники известны конкретные примеры гуманизированных антител, например, антитела, направленные против ЕрСАМ (см., например, LoBuglio, Proceedings of the American Society of Clinical Oncology Abstract (1997), 1562 и Khor, Proceedings of the American Society of Clinical Oncology Abstract (1997), 847).
Таким образом, в контексте данного изобретения в частности предложены молекулы биспецифических антител, которые прошли гуманизацию и способны успешно применяться в фармацевтических композициях. В контексте изобретения описанные в данном документе молекулы (гуманизированного) биспецифического антитела могут использоваться в составе набора, как описано ниже.
В контексте данного изобретения первый (Ig-производный) домен описанной здесь молекулы биспецифического антитела содержит сайт взаимодействия с антигеном, специфический в отношении антигена, не существующего в естественных условиях в и/или на CD8+ Т-клетках.
Термин "антиген, не встречающийся в естественных условиях в и/или на CD8+ Т-клетках", используемый в данном документе, относится к молекулам, внедренным в CD8+ Т-клетки, которые не презентируются в естественных условиях в и/или на поверхности CD8+ Т-клеток и (эндогенно) не экспрессируются в или на нормальных (нетрансдуцированных) CD8+ Т-клетках. Таким образом, антиген/маркер, не существующий в естественных условиях в и/или на CD8+ Т-клетках, искусственным образом введен в CD8+ Т-клетки. В контексте данного изобретения указанные CD8+ Т-клетки выделены/получены у субъекта, подлежащего лечению согласно данному описанию. В контексте данного изобретения антигенные пептиды, существующие в естественных условиях/эндогенно экспрессируемые на Т-клеточном рецепторе CD8+ Т-клеток, исключаются из указанного выше термина "антиген, не существующий в естественных условиях на CD8+ Т-клетках". Аналогично, эти молекулы, искусственным образом введенные и впоследствии презентируемые в и/или на поверхности указанных CD8+ Т-клеток, содержат домены или эпитопы, доступные (in vitro или in vivo) для (lg-производных) связывающих доменов, преимущественно антител, фрагментов антител или производных, которые не присутствуют в естественных условиях в и/или на CD8+ Т-клетках. В контексте данного изобретения эти искусственно введенные молекулы презентируются в и/или на поверхности указанных CD8+ Т-клеток после (ретровирусной) трансдукции в соответствии с описанием ниже.
В контексте данного изобретения термин "антиген, не существующий в естественных условиях в и/или на CD8+ Т-клетках" обозначает антиген/маркер, который не существует в естественных условиях/не экспрессируется эндогенно в и/или на CD8+ Т-клетках в количестве, превышающем 500, 550, 600, 650, 700, 750, 800, 850, 900, 950 или 1000 молекул антигена на CD8+ Т-клетку. Таким образом, антиген/маркер не существует в естественных условиях/не экспрессируется эндогенно в и/или на CD8+ Т-клетках в концентрации, превышающей 1,0, 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9 или 2,0 0/00 (промилле) в популяции нормальных (нетрансдуцированных) CD8+ Т-клеток. Наличие и количество антигена/маркера, который существует в естественных условиях в и/или на CD8+ Т-клетках, можно определять способами, известными в области техники, например, такими как FACS-анализ, ИФА, конфокальная микроскопия, аналитическая ВЭЖХ и т.п.
Примеры таких молекул включают неимуногенные белки, предпочтительно имеющие человеческое происхождение. В альтерантивном случае, указанные молекулы могут быть либо функционально инертными белковыми молекулами, как таковые, либо станут функционально инертными в результате использования методов генетической рекомбинации, известных в области техники (примерами могут быть белковые молекулы, в которых делеция внутриклеточного сигнального домена (как проиллюстрировано в приведенных Примерах в случае молекулы (человеческого) EGFR, лишенного внутриклеточного сигнального домена, обозначаемой в данном документе конструкцией del-hEGFR (SEQ ID NOs: 11 и 12)) или инактивирующие точечные мутации внеклеточного домена делают молекулу функционально инертной). Другим примером мутированной версии (человеческого) EGFR является конструкция del-hGFRvIII (последовательность ДНК представлена в SEQ ID NO: 17, а (кодируемая) аминокислотная последовательность представлена в SEQ ID NO: 18), представленная в приведенных Примерах. hEGFRvIII представляет собой мутантный вариант рецептора эпидермального фактора роста человека, обнаруживаемый в глиобластоме и карциноме молочной железы, яичников и легких. Мутантный рецептор имеет делецию во внеклеточном домене (Lorimer et al., Proc. Natl. Acad. Sci USA 93:14815-14820 (1996)).
Ниже приведены примеры маркеров, удовлетворяющие указанным выше критериям и включающие белок Cripto (альтернативное название «cryptic family protein»), представителей семейства (не-Т-клеточных) молекул CD (кластера дифференцировки, от англ. cluster of differentiation), EGFR или TSH-R, но не ограничивающиеся ими.
В контексте данного изобретения, (а) молекулы биспецифических антител, описанные в данном документе, связываются с антигеном, не существующим в естественных условиях в и/или на CD8+ Т-клетках, выбранным из группы, состоящей из белка Cripto (cryptic family protein), представителей семейства (не-Т-клеточных) молекул CD (кластера дифференцировки), EGFR и TSH-R. Таким образом, молекулы биспецифического антитела, описанные в данном документе, взаимодействуют/связываются с представителями семейства CD, которые (исключительно) не существуют в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках) (поскольку обозначаются термином "не-Т-клеточные"), Cripto, EGFR или TSH-R. В контексте данного изобретения молекулы биспецифического антитела, описанные в данном документе, взаимодействуют/связываются с представителями семейства CD, которые не экспрессируются эндогенно в и/или на поверхности Т-клеток (CD8+ Т-клеток) (поскольку обозначаются термином "не-Т-клеточные"), Cripto, EGFR или TSH-R.
Последовательности (человеческих) белков Cripto (cryptic family protein), представителей семейства (не-Т-клеточных) молекул CD (кластера дифференцировки), EGFR или TSH-R представлены в базе данных UniProtKB/Swiss-Prot и могут быть найдены по ссылке http://www.uniprot.org/uniprot/?query=reviewed%3Ayes. Эти (белковые) последовательности также охватывают аннотированные модифицированные последовательности. В данном изобретении также предложены методы и способы, в которых используются гомологичные последовательности, а также аллельные варианты генов и т.п. конкретных последовательностей, описанных в данном документе. Предпочтительно, используют такие "варианты" и т.п. конкретных последовательностей, описанных в данном документе. Предпочтительно, такие "варианты" представляют собой варианты генов. Специалист может без особого труда найти соответствующий кодирующий участок этих (белковых) последовательностей в базе данных, которые могут также содержать записи последовательностей геномной ДНК, а также мРНК/кДНК.
Термин "семейство (не-Т-клеточных) молекул CD (кластера дифференцировки)", используемый в данном документе в отношении "антигена, не встречающегося в естественных условиях/эндогенно не экспрессируемого в и/или на CD8+ Т-клетках" относится к любой из последовательностей CD, выбранных из группы, состоящей из CD9, CD10, CD11, CD12, CD13, CD14, CD15, CD16, CD17, CD18, CD19, CD20, CD21, CD22, CD23, CD24, CD25, CD26, CD27, CD28, CD29, CD30, CD31, CD32, CD33, CD34, CD35, CD36, CD37, CD38, CD39, CD40, CD41, CD43, CD46, CD48, CD49, CD50, CD51, CD54, CD55, CD56, CD57, CD59, CD61, CD63, CD64, CD66, CD67, CD68, CD70, CD72, CD74, CD75, CD76, CD77, CD79, CD81, CD82, CD83, CD84, CD87, CD88, CD89, CD90, CD91, CD92, CD93, CD94, CD95, CD97, CD98, CD99, CD100, CD101, CD102, CD103, CD104, CD105, CD106, CD107, CD108, CD109, CD110, CD111, CD112, CD113, CD114, CD115, CD116, CD117, CD118, CD119, CD121, CD123, CD124, CD125, CD126, CD130, CD131, CD133, CD134, CD135, CD136, CD137, CD138, CD140, CD141, CD142, CD143, CD144, CD146, CD147, CD148, CD151, CD153, CD155, CD156, CD157, CD158, CD159, CD160, CD161, CD162, CD163, CD164, CD166, CD167, CD168, CD169, CD170, CD171, CD172, CD177, CD178, CD179, CD180, CD181, CD182, CD183, CD184, CD185, CD186, CD191, CD192, CD193, CD200, CD201, CD204, CD206, CD207, CD208, CD209, CD217, CD218, CD220, CD221, CD222, CD223, CD224, CD225, CD226, CD227, CD228, CD230, CD231, CD232, CD233, CD234, CD236, CD238, CD239, CD241, CD242, CD243, CD244, CD246, CD248, CD249, CD252, CD253, CD254, CD256, CD257, CD258, CD261, CD262, CD263, CD264, CD265, CD266, CD267, CD268, CD269, CD270, CD271, CD276, CD277, CD280, CD281, CD282, CD283, CD284, CD286, CD288, CD289, CD290, CD292, CD294, CD295, CD296, CD297, CD298, CD299, CD300, CD301, CD302, CD303, CD304, CD305, CD306, CD309, CD312, CD314, CD315, CD316, CD317, CD318, CD319, CD320, CD321, CD322, CD324, CD325, CD326, CD327, CD328, CD329, CD331, CD332, CD333, CD334, CD335, CD336, CD337, CD338, CD339, CD340, CD344, CD349, CD350, CD351, CD352, CD353, CD354, CD355, CD357, CD358, CD360, CD361, CD362 и CD363.
Последовательность(и) (человеческого) CD9 (антиген CD9) находи(я)тся в базе данных Swiss-Prot под номером Р21926 (версия записи 123, версия последовательности 4); последовательность(и) (человеческого) CD10 (неприлизин) находи(я)тся в базе данных Swiss-Prot под номером Р08473 (версия записи 151, версия последовательности 2); последовательность(и) (человеческого) CD11 (интегрин альфа-D) находи(я)тся в базе данных Swiss-Prot под номером Q13349 (версия записи 110, версия последовательности 2); последовательность(и) (человеческого) CD13 (аминопептидаза N) находи(я)тся в базе данных Swiss-Prot под номером Р15144 (версия записи 145, версия последовательности 4); последовательность(и) (человеческого) CD14 (дифференцировочный антиген моноцитов CD14) находи(я)тся в базе данных Swiss-Prot под номером Р08571 (версия записи 131, версия последовательности 2); последовательность(и) (человеческого) CD16 (Fc-гамма рецептор IIIb) находи(я)тся в базе данных Swiss-Prot под номером Q9ULV2 (версия записи 51, версия последовательности 1); последовательность(и) (человеческого) CD18 (интегрин бета-2) находи(я)тся в базе данных Swiss-Prot под номером Р05107 (версия записи 162, версия последовательности 2); последовательность(и) (человеческого) CD19 (В-лимфоцитарный антиген CD19) находи(я)тся в базе данных Swiss-Prot под номером Р15391 (версия записи 128, версия последовательности 6); последовательность(и) (человеческого) CD20 (В-лимфоцитарный антиген CD20) находи(я)тся в базе данных Swiss-Prot под номером Р11836 (версия записи 118, версия последовательности 1); последовательность(и) (человеческого) CD21 (рецептор комплемента 2 типа) находи(я)тся в базе данных Swiss-Prot под номером Р20023 (версия записи 128, версия последовательности 2); последовательность(и) (человеческого) CD22 (В-клеточный рецептор CD22) находи(я)тся в базе данных Swiss-Prot под номером Р20273 (версия записи 136, версия последовательности 2); последовательность(и) (человеческого) CD23 (низкоаффинный Fc-рецептор для IgE) находи(я)тся в базе данных Swiss-Prot под номером Р06734 (версия записи 133, версия последовательности 1); последовательность(и) (человеческого) CD24 (передатчик сигнала CD24) находи(я)тся в базе данных Swiss-Prot под номером Р25063 (версия записи 106, версия последовательности 2); последовательность(и) (человеческого) CD26 (дипептидил-пептидаза 4) находи(я)тся в базе данных Swiss-Prot под номером Р27487 (версия записи 140, версия последовательности 2); последовательность(и) (человеческого) CD27 (антиген CD27) находи(я)тся в базе данных Swiss-Prot под номером Р26842 (версия записи 119, версия последовательности 2); последовательность(и) (человеческого) CD29 (интегрин бета-1) находи(я)тся в базе данных Swiss-Prot под номером Р05556 (версия записи 154, версия последовательности 2); последовательность(и) (человеческого) CD30 (8 представитель семейства рецепоров фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Р28908 (версия записи 129; версия последовательности 1); последовательность(и) (человеческого) CD31 (тромоцитарно-эндотелиальная молекула клеточной адгезии) находи(я)тся в базе данных Swiss-Prot под номером Р16284 (версия записи 146, версия последовательности 1); последовательность(и) (человеческого) CD32 (низкоаффинный Fc-рецептор IgG тип II-b) находи(я)тся в базе данных Swiss-Prot под номером Р31994 (версия записи 138, версия последовательности 2); последовательность(и) (человеческого) CD33 (поверхностный антиген миелоидных клеток CD33) находи(я)тся в базе данных Swiss-Prot под номером Р20138 (версия записи 130, версия последовательности 2); последовательность(и) (человеческого) CD34 (антиген гемопоэтических прогениторных клеток CD34) находи(я)тся в базе данных Swiss-Prot под номером Р28906 (версия записи 108, версия последовательности 2); последовательность(и) (человеческого) CD35 (рецептор комплемента 1 типа) находи(я)тся в базе данных Swiss-Prot под номером Р17927 (версия записи 131, версия последовательности 3); последовательность(и) (человеческого) CD36 (тромбоцитарный гликопротеин 4) находи(я)тся в базе данных Swiss-Prot под номером Р16671 (версия записи 133, версия последовательности 2); последовательность(и) (человеческого) CD38 (АДФ-рибозил-циклаза 1) находи(я)тся в базе данных Swiss-Prot под номером Р28907 (версия записи 126, версия последовательности 2); последовательность(и) (человеческого) CD39 (эктонуклеозид-трифосфат-дифосфогидролаза 1) находи(я)тся в базе данных Swiss-Prot под номером Р49961 (версия записи 114, версия последовательности 1); последовательность(и) (человеческого) CD40 (5 представитель семейства рецепоров фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Р25942 (версия записи 147, версия последовательности 1); последовательность(и) (человеческого) CD41 (интегрин альфа-IIb) находи(я)тся в базе данных Swiss-Prot под номером Р08514 (версия записи 158, версия последовательности 3); последовательность(и) (человеческого) CD43 (лейкосиалин) находи(я)тся в базе данных Swiss-Prot под номером Р16150 (версия записи 110, версия последовательности 1); последовательность(и) (человеческого) CD46 (мембранный кофакторный белок) находи(я)тся в базе данных Swiss-Prot под номером Р15529 (версия записи 145, версия последовательности 3); последовательность(и) (человеческого) CD48 (антиген CD48) находи(я)тся в базе данных Swiss-Prot под номером Р09326 (версия записи 137, версия последовательности 2); последовательность(и) (человеческого) CD49 (интегрин альфа-4) находи(я)тся в базе данных Swiss-Prot под номером Р13612 (версия записи 128, версия последовательности 3); последовательность(и) (человеческого) CD50 (межклеточная молекула адгезии 3) находи(я)тся в базе данных Swiss-Prot под номером Р32942 (версия записи 128, версия последовательности 2); последовательность(и) (человеческого) CD51 (интегрин альфа-V) находи(я)тся в базе данных Swiss-Prot под номером Р06756 (версия записи 149, версия последовательности 2); последовательность(и) (человеческого) CD54 (межклеточная молекула адгезии 1) находи(я)тся в базе данных Swiss-Prot под номером Р05362 (версия записи 160, версия последовательности 2); последовательность(и) (человеческого) CD55 (фактор, ускоряющий инактивацию комплемента) находи(я)тся в базе данных Swiss-Prot под номером Р08174 (версия записи 143, версия последовательности 4); последовательность(и) (человеческого) CD56 (молекула адгезии нервных клеток 1) находится в базе данных Swiss-Prot под номером Р13591 (версия записи 132, версия последовательности 3); последовательность(и) (человеческого) CD57 (1 представитель семейства лектиноподобных рецепторов киллерных клеток типа G) находи(я)тся в базе данных Swiss-Prot под номером Q96E93 (версия записи 72, версия последовательности 1); последовательность(и) (человеческого) CD59 (гликопротеин CD59) находи(я)тся в базе данных Swiss-Prot под номером Р13987 (версия записи 139, версия последовательности 1); последовательность(и) (человеческого) CD61 (интегрин бета-3) находи(я)тся в базе данных Swiss-Prot под номером Р05106 (версия записи 175, версия последовательности 2); последовательность(и) (человеческого) CD63 (антиген CD63) находи(я)тся в базе данных Swiss-Prot под номером Р08962 (версия записи 122, версия последовательности 2); последовательность(и) (человеческого) CD64 (высокоаффинный Fc-рецептор иммуноглобулина G тип I) находи(я)тся в базе данных Swiss-Prot под номером Р12314 (версия записи 128, версия последовательности 2); последовательность(и) (человеческого) CD66 (родственная раковоэмбриональному антигену молекула клеточной адгезии 1) находи(я)тся в базе данных Swiss-Prot под номером Р13688 (версия записи 133, версия последовательности 2); последовательность(и) (человеческого) CD67 (родственная раковоэмбриональному антигену молекула клеточной адгезии 8) находи(я)тся в базе данных Swiss-Prot под номером Р31997 (версия записи 115, версия последовательности 2); последовательность(и) (человеческого) CD68 (макросиалин) находи(я)тся в базе данных Swiss-Prot под номером Р34810 (версия записи 106, версия последовательности 2); последовательность(и) (человеческого) CD70 (антиген CD70) находи(я)тся в базе данных Swiss-Prot под номером Р32970 (версия записи 101, версия последовательности 2); последовательность(и) (человеческого) CD72 (В-клеточный дифференцировочный антиген CD72) находи(я)тся в базе данных Swiss-Prot под номером Р21854 (версия записи 113, версия последовательности 1); последовательность(и) (человеческого) CD74 (гамма-цепь антигена гистосовместимости HLA класса II) находи(я)тся в базе данных Swiss-Prot под номером Р04233 (версия записи 141, версия последовательности 3); последовательность(и) (человеческого) CD75 (бета-галактозид-альфа-2,6-сиалил-трансфераза 1) находи(я)тся в базе данных Swiss-Prot под номером Р15907 (версия записи 130, версия последовательности 1); последовательность(и) (человеческого) CD77 (лактозилцерамид-4-альфа-галактозилтрансфераза) находи(я)тся в базе данных Swiss-Prot под номером Q9NPC4 (версия записи 100, версия последовательности 1); последовательность(и) (человеческого) CD79 (альфа-цепь белка, ассоциированного с комплексом В-клеточного рецептора антигена) находи(я)тся в базе данных Swiss-Prot под номером Р11912 (версия записи 120, версия последовательности 2); последовательность(и) (человеческого) CD81 (антиген CD81) находи(я)тся в базе данных Swiss-Prot под номером Р60033 (версия записи 82, версия последовательности 1); последовательность(и) (человеческого) CD82 (антиген CD82) находи(я)тся в базе данных Swiss-Prot под номером Р27701 (версия записи 98, версия последовательности 1); последовательность(и) (человеческого) CD83 (антиген CD83) находи(я)тся в базе данных Swiss-Prot под номером Q01151 (версия записи 113, версия последовательности 1); последовательность(и) (человеческого) CD84 (5 представитель семейства SLAM 5) находи(я)тся в базе данных Swiss-Prot под номером Q9UIB8 (версия записи 87, версия последовательности 1); последовательность(и) (человеческого) CD87 (поверхностный рецептор урокиназного активатора плазминогена) находи(я)тся в базе данных Swiss-Prot под номером Q03405 (версия записи 129, версия последовательности 1); последовательность(и) (человеческого) CD88 (рецептор хемотаксического анафилотоксина С5а) находи(я)тся в базе данных Swiss-Prot под номером Р21730 (версия записи 116, версия последовательности 2); последовательность(и) (человеческого) CD89 (Fc-рецептор иммуноглобулина А) находи(я)тся в базе данных Swiss-Prot под номером Р24071 (версия записи 121, версия последовательности 1); последовательность(и) (человеческого) CD90 (мембранный гликопротеин Thy-1) находи(я)тся в базе данных Swiss-Prot под номером Р04216 (версия записи 128, версия последовательности 2); последовательность(и) (человеческого) CD91 (белок родственный рецептору липопротеинов низкой плотности тип 1) находи(я)тся в базе данных Swiss-Prot под номером Q07954 (версия записи 133, версия последовательности 2); последовательность(и) (человеческого) CD92 (белок, подобный переносчику холина тип 1) находи(я)тся в базе данных Swiss-Prot под номером Q8WWI5 (версия записи 79, версия последовательности 1); последовательность(и) (человеческого) CD93 (рецептор компонента комплемента C1q) находи(я)тся в базе данных Swiss-Prot под номером Q9NPY3 (версия записи 115, версия последовательности 3); последовательность(и) (человеческого) CD94 (антиген естественных киллерных клеток CD94) находи(я)тся в базе данных Swiss-Prot под номером Q13241 (версия записи 107, версия последовательности 2); последовательность(и) (человеческого) CD95 (6 представитель семейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Р48023 (версия записи 134, версия последовательности 1); последовательность(и) (человеческого) CD97 (антиген CD97) находи(я)тся в базе данных Swiss-Prot под номером Р48960 (версия записи 125, версия последовательности 4); последовательность(и) (человеческого) CD98 (тяжелая цепь антигена клеточной поверхности 4F2) находи(я)тся в базе данных Swiss-Prot под номером Р08195 (версия записи 140, версия последовательности 3); последовательность(и) (человеческого) CD99 (антиген CD99) находи(я)тся в базе данных Swiss-Prot под номером Р14209 (версия записи 117, версия последовательности 1); последовательность(и) (человеческого) CD100 (семафорин-4D) находи(я)тся в базе данных Swiss-Prot под номером Q92854 (версия записи 125, версия последовательности 1); последовательность(и) (человеческого) CD101 (2 представитель суперсемейства иммуноглобулинов) находи(я)тся в базе данных Swiss-Prot под номером Q93033 (версия записи 89, версия последовательности 2); последовательность(и) (человеческого) CD102 (межклеточная молекула адгезии 2) находи(я)тся в базе данных Swiss-Prot под номером Р13598 (версия записи 131, версия последовательности 2); последовательность(и) (человеческого) CD103 (интегрин альфа-E) находи(я)тся в базе данных Swiss-Prot под номером Р38570 (версия записи 118, версия последовательности 3); последовательность(и) (человеческого) CD104 (интегрин бета-4) находи(я)тся в базе данных Swiss-Prot под номером Р16144 (версия записи 160, версия последовательности 5); последовательность(и) (человеческого) CD105 (эндоглин) находи(я)тся в базе данных Swiss-Prot под номером Р17813 (версия записи 133, версия последовательности 2); последовательность(и) (человеческого) CD106 (белок адгезии сосудистых клеток 1) находи(я)тся в базе данных Swiss-Prot под номером Р19320 (версия записи 158, версия последовательности 1); последовательность(и) (человеческого) CD107 (ассоциированный с мембраной лизосом гликопротеин 1) находи(я)тся в базе данных Swiss-Prot под номером Р11279 (версия записи 117, версия последовательности 3); последовательность(и) (человеческого) CD108 (семафорин-7А) находи(я)тся в базе данных Swiss-Prot под номером 075326 (версия записи 107, версия последовательности 1); последовательность(и) (человеческого) CD109 (антиген CD109) находи(я)тся в базе данных Swiss-Prot под номером Q6YHK3 (версия записи 64, версия последовательности 2); последовательность(и) (человеческого) CD110 (рецептор тромбопоэтина) находи(я)тся в базе данных Swiss-Prot под номером Р40238 (версия записи 122, версия последовательности 1); последовательность(и) (человеческого) CD111 (родственный рецептору полиовируса белок 1) находи(я)тся в базе данных Swiss-Prot под номером Q15223 (версия записи 114, версия последовательности 3); последовательность(и) (человеческого) CD112 (родственный рецептору полиовируса белок 2) находи(я)тся в базе данных Swiss-Prot под номером Q92692 (версия записи 123, версия последовательности 1); последовательность(и) (человеческого) CD113 (родственный рецептору полиовируса белок 3) находи(я)тся в базе данных Swiss-Prot под номером Q9NQS3 (версия записи 78, версия последовательности 1); последовательность(и) (человеческого) CD114 (рецептор гранулоцитарного колониестимулирующего фактора) находи(я)тся в базе данных Swiss-Prot под номером Q99062 (версия записи 129, версия последовательности 1); последовательность(и) (человеческого) CD115 (рецептор макрофагального колониестимулирующего фактора 1) находи(я)тся в базе данных Swiss-Prot под номером Р07333 (версия записи 145, версия последовательности 2); последовательность(и) (человеческого) CD116 (альфа-субъединица рецептора гранулоцитарно-макрофагального колониестимулирующего фактора) находи(я)тся в базе данных Swiss-Prot под номером Р15509 (версия записи 128, версия последовательности 1); последовательность(и) (человеческого) CD117 (рецептор фактора роста тучных и стволовых клеток Kit) находи(я)тся в базе данных Swiss-Prot под номером Р10721 (версия записи 150, версия последовательности 1); последовательность(и) (человеческого) CD118 (рецептор лейкемия-ингибирующего фактора) находи(я)тся в базе данных Swiss-Prot под номером Р42702 (версия записи 115, версия последовательности 1); последовательность(и) (человеческого) CD119 (рецептор интерферона гамма тип 1) находи(я)тся в базе данных Swiss-Prot под номером Р15260 (версия записи 140, версия последовательности 1); последовательность(и) (человеческого) CD121 (рецептор интерлейкина-1 тип 1) находи(я)тся в базе данных Swiss-Prot под номером Р14778 (версия записи 151, версия последовательности 1); последовательность(и) (человеческого) CD123 (альфа-субъединица рецептора интерлейкина-3) находи(я)тся в базе данных Swiss-Prot под номером Р26951 (версия записи 110, версия последовательности 1); последовательность(и) (человеческого) CD124 (альфа-субъединица рецептора интерлейкина-4) находи(я)тся в базе данных Swiss-Prot под номером Р24394 (версия записи 144, версия последовательности 1); последовательность(и) (человеческого) CD125 (альфа-субъединица рецептора интерлейкина-5) находи(я)тся в базе данных Swiss-Prot под номером Q01344 (версия записи 120, версия последовательности 2 последовательность(и) (человеческого) CD126 (альфа-субъединица рецептора интерлейкина-6) находи(я)тся в базе данных Swiss-Prot под номером Р08887 (версия записи 143, версия последовательности 1); последовательность(и) (человеческого) CD130 (бета-субъединица рецептора интерлейкина-6) находи(я)тся в базе данных Swiss-Prot под номером Р40189 (версия записи 142, версия последовательности 2); последовательность(и) (человеческого) CD131 (общая бета-субъединица рецепторов цитокинов) находи(я)тся в базе данных Swiss-Prot под номером Р32927 (версия записи 128, версия последовательности 2); последовательность(и) (человеческого) CD133 (проминин-1) находи(я)тся в базе данных Swiss-Prot под номером O43490 (версия записи 110, версия последовательности 1); последовательность(и) (человеческого) CD134 (4 представитель суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Р43489 (версия записи 106, версия последовательности 1); последовательность(и) (человеческого) CD135 (рецепторная тирозиновая протеинкиназа FLT3) находи(я)тся в базе данных Swiss-Prot под номером Р36888 (версия записи 119, версия последовательности 2); последовательность(и) (человеческого) CD136 (рецептор белка, стимулирующего макрофаги) находи(я)тся в базе данных Swiss-Prot под номером Q04912 (версия записи 129, версия последовательности 2); последовательность(и) (человеческого) CD137 (9 представитель суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Q07011 (версия записи 109, версия последовательности 1); последовательность(и) (человеческого) CD138 (синдекан-1) находи(я)тся в базе данных Swiss-Prot под номером Р18827 (версия записи 114, версия последовательности 3); последовательность(и) (человеческого) CD140 (рецептор тромбоцитарного фактора роста тип бета) находи(я)тся в базе данных Swiss-Prot под номером Р09619 (версия записи 154, версия последовательности 1); последовательность(и) (человеческого) CD141 (тромбомодулин) находи(я)тся в базе данных Swiss-Prot под номером Р07204 (версия записи 162, версия последовательности 2); последовательность(и) (человеческого) CD142 (тканевой фактор) находи(я)тся в базе данных Swiss-Prot под номером Р13726 (версия записи 137, версия последовательности 1); последовательность(и) (человеческого) CD143 (ангиотензин-превращающий фермент) находи(я)тся в базе данных Swiss-Prot под номером Р12821 (версия записи 157, версия последовательности 1); последовательность(и) (человеческого) CD144 (кадгерин-5) находи(я)тся в базе данных Swiss-Prot под номером Р33151 (версия записи 108, версия последовательности 5); последовательность(и) (человеческого) CD146 (гликопротеин клеточной поверхности MUC18) находи(я)тся в базе данных Swiss-Prot под номером Р43121 (версия записи 109, версия последовательности 2); последовательность(и) (человеческого) CD 147 (басигин) находи(я)тся в базе данных Swiss-Prot под номером Р35613 (версия записи 134, версия последовательности 2); последовательность(и) (человеческого) CD148 (рецепторная тирозиновая протеинфосфатаза эта) находи(я)тся в базе данных Swiss-Prot под номером Q12913 (версия записи 124, версия последовательности 3); последовательность(и) (человеческого) CD151 (антиген CD151) находи(я)тся в базе данных Swiss-Prot под номером Р48509 (версия записи 108, версия последовательности 3); последовательность(и) (человеческого) CD153 (8 представитель суперсемейства фактора некроза опухоли (лиганд)) находи(я)тся в базе данных Swiss-Prot под номером Р32971 (версия записи 90, версия последовательности 1); последовательность(и) (человеческого) CD155 (рецептор полиовируса) находи(я)тся в базе данных Swiss-Prot под номером Р15151 (версия записи 132, версия последовательности 2); последовательность(и) (человеческого) CD156 (белок, содержащий дизинтегриновый и металлопротеиназный домены, тип 8) находи(я)тся в базе данных Swiss-Prot под номером Р78325 (версия записи 115, версия последовательности 1); последовательность(и) (человеческого) CD157 (АДФ-рибозил-циклаза 2) находи(я)тся в базе данных Swiss-Prot под номером Q10588 (версия записи 116, версия последовательности 2); последовательность(и) (человеческого) CD158 (иммуноглобулин-подобный рецептор киллерных клеток 3DL3) находи(я)тся в базе данных Swiss-Prot под номером Q8N743 (версия записи 91, версия последовательности 2); последовательность(и) (человеческого) CD159 (интегральный мембранный белок NKG2-A/NKG2-B типа II) находи(я)тся в базе данных Swiss-Prot под номером Р26715 (версия записи 116, версия последовательности 2); последовательность(и) (человеческого) CD160 (антиген CD160) находи(я)тся в базе данных Swiss-Prot под номером O95971 (версия записи 98, версия последовательности 1); последовательность(и) (человеческого) CD161 (1 представитель подсемейства лектин-подобных рецепторов киллерных клеток типа В) находи(я)тся в базе данных Swiss-Prot под номером Q12918 (версия записи 81, версия последовательности 1); последовательность(и) (человеческого) CD162 (гликопротеиновый лиганд P-селектина) находи(я)тся в базе данных Swiss-Prot под номером Q14242 (версия записи 103, версия последовательности 1); последовательность(и) (человеческого) CD163 (богатый цистеином скевенджер-рецептор 1 типа белок M130) находи(я)тся в базе данных Swiss-Prot под номером Q86VB7 (версия записи 77, версия последовательности 2); последовательность(и) (человеческого) CD164 (белок ядра сиаломуцина 24) находи(я)тся в базе данных Swiss-Prot под номером Q04900 (версия записи 89), версия последовательности 2); последовательность(и) (человеческого) CD166 (антиген CD166) находи(я)тся в базе данных Swiss-Prot под номером Q13740 (версия записи 111, версия последовательности 2); последовательность(и) (человеческого) CD167 (рецептор, содержащий дискоидиновый домен, тип 2) находи(я)тся в базе данных Swiss-Prot под номером Q16832 (версия записи 120, версия последовательности 2); последовательность(и) (человеческого) CD168 (рецептор для клеток с опосредованной гиалуроновой кислотой подвижностью) находи(я)тся в базе данных Swiss-Prot под номером O75330 (версия записи 99, версия последовательности 2); последовательность(и) (человеческого) CD169 (сиалоадгезин) находи(я)тся в базе данных Swiss-Prot под номером Q9BZZ2 (версия записи 103, версия последовательности 2); последовательность(и) (человеческого) CD170 (связывающий сиаловую кислоту иммуноглобулин-подобный лектин 5) находи(я)тся в базе данных Swiss-Prot под номером O15389 (версия записи 106, версия последовательности 1); последовательность(и) (человеческого) CD171 (молекула адгезии нервных клеток L1) находи(я)тся в базе данных Swiss-Prot под номером Р32004 (версия записи 139, версия последовательности 2); последовательность(и) (человеческого) CD172 (сигнальный регуляторный белок бета-1) находи(я)тся в базе данных Swiss-Prot под номером O00241 (версия записи 112, версия последовательности 5); последовательность(и) (человеческого) CD177 (антиген CD177) находи(я)тся в базе данных Swiss-Prot под номером Q8N6Q3 (версия записи 65, версия последовательности 2); последовательность(и) (человеческого) CD178 (6 представитель суперсемейства фактора некроза опухоли (лиганд)) находи(я)тся в базе данных Swiss-Prot под номером Р48023 (версия записи 134, версия последовательности 1); последовательность(и) (человеческого) CD179 (йота цепь иммуноглобулинов) находи(я)тся в базе данных Swiss-Prot под номером Р12018 (версия записи 115, версия последовательности 2); последовательность(и) (человеческого) CD180 (антиген CD180) находи(я)тся в базе данных Swiss-Prot под номером Q99467 (версия записи 101, версия последовательности 2); последовательность(и) (человеческого) CD181 (рецептор С-Х-С-хемокинов типа 1) находи(я)тся в базе данных Swiss-Prot под номером Р25024 (версия записи 125, версия последовательности 2); последовательность(и) (человеческого) CD182 (рецептор С-Х-С-хемокинов типа 2) находи(я)тся в базе данных Swiss-Prot под номером Р25025 (версия записи 123, версия последовательности 2); последовательность(и) (человеческого) CD183 (рецептор С-Х-С-хемокинов типа 3) находи(я)тся в базе данных Swiss-Prot под номером Р49682 (версия записи 118, версия последовательности 2); последовательность(и) (человеческого) CD184 (рецептор С-Х-С-хемокинов типа 4) находи(я)тся в базе данных Swiss-Prot под номером Р61073 (версия записи 95, версия последовательности 1); последовательность(и) (человеческого) CD185 (рецептор С-Х-С-хемокинов типа 5) находи(я)тся в базе данных Swiss-Prot под номером Р32302 (версия записи 109, версия последовательности 1); последовательность(и) (человеческого) CD186 (рецептор С-Х-С-хемокинов типа 6) находи(я)тся в базе данных Swiss-Prot под номером O00574 (версия записи 104, версия последовательности 1); последовательность(и) (человеческого) CD191 (рецептор С-С-хемокинов типа 1) находи(я)тся в базе данных Swiss-Prot под номером Р32246 (версия записи 106, версия последовательности 1); последовательность(и) (человеческого) CD192 (рецептор С-С-хемокинов типа 2) находи(я)тся в базе данных Swiss-Prot под номером Р41597 (версия записи 128, версия последовательности 1); последовательность(и) (человеческого) CD193 (рецептор С-С-хемокинов типа 3) находи(я)тся в базе данных Swiss-Prot под номером Р51677 (версия записи 112, версия последовательности 1); последовательность(и) (человеческого) CD200 (мембранный гликопротеин ОХ-2) находи(я)тся в базе данных Swiss-Prot под номером Р41217 (версия записи 110, версия последовательности 4); последовательность(и) (человеческого) CD201 (эндотелиальный рецептор протеина С) находи(я)тся в базе данных Swiss-Prot под номером Q9UNN8 (версия записи 110, версия последовательности 1); последовательность(и) (человеческого) CD204 (макрофагальные скевенджер-рецепторы типов I и II) находи(я)тся в базе данных Swiss-Prot под номером Р21757 (версия записи 122, версия последовательности 1); последовательность(и) (человеческого) CD206 (макрофагальный рецептор для маннозы 1) находи(я)тся в базе данных Swiss-Prot под номером Р22897 (версия записи 138, версия последовательности 1); последовательность(и) (человеческого) CD207 (представитель К 4 семейства содержащих лектиновый домен типа С) находи(я)тся в базе данных Swiss-Prot под номером Q9UJ71 (версия записи 85, версия последовательности 2); последовательность(и) (человеческого) CD208 (ассоциированный с мембраной лизосом гликопротеин 3) находи(я)тся в базе данных Swiss-Prot под номером Q9UQV4 (версия записи 69, версия последовательности 3); последовательность(и) (человеческого) CD209 (антиген CD209) находи(я)тся в базе данных Swiss-Prot под номером Q9NNX6 (версия записи 103, версия последовательности 1); последовательность(и) (человеческого) CD217 (рецептор А интерлейкина-17) находи(я)тся в базе данных Swiss-Prot под номером Q96F46 (версия записи 94, версия последовательности 2); последовательность(и) (человеческого) CD218 (рецептор 1 интерлейкина-18) находи(я)тся в базе данных Swiss-Prot под номером Q13478 (версия записи 104, версия последовательности 1); последовательность(и) (человеческого) CD220 (рецептор инсулина) находи(я)тся в базе данных Swiss-Prot под номером Р06213 (версия записи 175, версия последовательности 4); последовательность(и) (человеческого) CD221 (рецептор инсулиноподобного фактора роста 1) находи(я)тся в базе данных Swiss-Prot под номером Р08069 (версия записи 145, версия последовательности 1); последовательность(и) (человеческого) CD222 (катион-независимый рецептор маннозо-6-фосфата) находи(я)тся в базе данных Swiss-Prot под номером Р11717 (версия записи 137, версия последовательности 3); последовательность(и) (человеческого) CD223 (белок гена активации лимфоцитов 3) находи(я)тся в базе данных Swiss-Prot под номером Р18627 (версия записи 108, версия последовательности 5); последовательность(и) (человеческого) CD224 (гамма-глутамил-транспептидаза 1) находи(я)тся в базе данных Swiss-Prot под номером Р19440 (версия записи 137, версия последовательности 2); последовательность(и) (человеческого) CD225 (индуцируемый интерфероном трансмембранный белок 1) находи(я)тся в базе данных Swiss-Prot под номером Р13164 (версия записи 101, версия последовательности 3); последовательность(и) (человеческого) CD226 (антиген CD226) находи(я)тся в базе данных Swiss-Prot под номером Q15762 (версия записи 89, версия последовательности 2); последовательность(и) (человеческого) CD227 (муцин-1) находи(я)тся в базе данных Swiss-Prot под номером Р15941 (версия записи 136, версия последовательности 3); последовательность(и) (человеческого) CD228 (меланотрансферрин) находи(я)тся в базе данных Swiss-Prot под номером Р08582 (версия записи 124, версия последовательности 2); последовательность(и) (человеческого) CD230 (прионовый белок) находи(я)тся в базе данных Swiss-Prot под номером Р04156 (версия записи 161, версия последовательности 1); последовательность(и) (человеческого) CD231 (тетраспанин-7) находи(я)тся в базе данных Swiss-Prot под номером Р41732 (версия записи 115, версия последовательности 2); последовательность(и) (человеческого) CD232 (плексин-С1) находи(я)тся в базе данных Swiss-Prot под номером O60486 (версия записи 80, версия последовательности 1); последовательность(и) (человеческого) CD233 (анионный транспортный белок полосы 3 эритроцитов) находи(я)тся в базе данных Swiss-Prot под номером Р02730 (версия записи 167, версия последовательности 3); последовательность(и) (человеческого) CD234 (антиген Duffy/рецептор хемокинов) находи(я)тся в базе данных Swiss-Prot под номером Q16570 (версия записи 114, версия последовательности 3); последовательность(и) (человеческого) CD236 (гликофорин-С) находи(я)тся в базе данных Swiss-Prot под номером Р04921 (версия записи 116, версия последовательности 1); последовательность(и) (человеческого) CD238 (гликопротеин группы крови Kell) находи(я)тся в базе данных Swiss-Prot под номером Р23276 (версия записи 124, версия последовательности 2); последовательность(и) (человеческого) CD239 (молекула адгезии базальной поверхности клеток) находи(я)тся в базе данных Swiss-Prot под номером Р50895 (версия записи 117, версия последовательности 2); последовательность(и) (человеческого) CD241 (переносчик аммония антиген Rh тип А) находи(я)тся в базе данных Swiss-Prot под номером Q02094 (версия записи 98, версия последовательности 2); последовательность(и) (человеческого) CD242 (молекула межклеточной адгезии 4) находи(я)тся в базе данных Swiss-Prot под номером Q14773 (версия записи 106, версия последовательности 1); последовательность(и) (человеческого) CD243 (белок множественной лекарственной устойчивости 1) находи(я)тся в базе данных Swiss-Prot под номером Р08183 (версия записи 146, версия последовательности 3; последовательность(и) (человеческого) CD244 (рецептор естественных киллерных клеток 2 В4) находи(я)тся в базе данных Swiss-Prot под номером Q9BZW8 (версия записи 94, версия последовательности 2); последовательность(и) (человеческого) CD246 (тирозинкиназный рецептор ALK) находи(я)тся в базе данных Swiss-Prot под номером Q9UM73 (версия записи 120, версия последовательности 3); последовательность(и) (человеческого) CD248 (эндосиалин) находи(я)тся в базе данных Swiss-Prot под номером Q9HCU0 (версия записи 87, версия последовательности 1); последовательность(и) (человеческого) CD249 (глутамил-аминопептидаза) находи(я)тся в базе данных Swiss-Prot под номером Q07075 (версия записи 121, версия последовательности 3); последовательность(и) (человеческого) CD252 (представитель 4 суперсемейства фактора некроза опухоли (лиганд)) находи(я)тся в базе данных Swiss-Prot под номером Р23510 (версия записи 101, версия последовательности 1); последовательность(и) (человеческого) CD253 (10 представитель суперсемейства фактора некроза опухоли (лиганд)) находи(я)тся в базе данных Swiss-Prot под номером Р50591 (версия записи 118, версия последовательности 1); последовательность(и) (человеческого) CD254 (11 представитель суперсемейства фактора некроза опухоли (лиганд)) находи(я)тся в базе данных Swiss-Prot под номером O14788 (версия записи 110, версия последовательности 1); последовательность(и) (человеческого) CD256 (13 представитель суперсемейства фактора некроза опухоли (лиганд)) находи(я)тся в базе данных Swiss-Prot под номером O75888 (версия записи 111, версия последовательности 1); последовательность(и) (человеческого) CD257 (представитель 13B суперсемейства фактора некроза опухоли (лиганд)) находи(я)тся в базе данных Swiss-Prot под номером Q9Y275 (версия записи 127, версия последовательности 1); последовательность(и) (человеческого) CD258 (14 представитель суперсемейства фактора некроза опухоли (лиганд)) находи(я)тся в базе данных Swiss-Prot под номером O43557 (версия записи 117, версия последовательности 2); последовательность(и) (человеческого) CD261 (представитель 10А суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером O00220 (версия записи 112, версия последовательности 3); последовательность(и) (человеческого) CD262 (представитель 10 В суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером O14763 (версия записи 133, версия последовательности 2); последовательность(и) (человеческого) CD263 (представитель 10С суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером O14798 (версия записи 99, версия последовательности 3); последовательность(и) (человеческого) CD264 (представитель 10D суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Q9UBN6 (версия записи 109, версия последовательности 1); последовательность(и) (человеческого) CD265 (представитель 11А суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Q9Y6Q6 (версия записи 100, версия последовательности 1); последовательность(и) (человеческого) CD266 (представитель 12А суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Q9NP84 (версия записи 89, версия последовательности 1); последовательность(и) (человеческого) CD267 (представитель 13 В суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером O14836 (версия записи 102, версия последовательности 1); последовательность(и) (человеческого) CD268 (представитель 13С суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Q96RJ3 (версия записи 91, версия последовательности 1); последовательность(и) (человеческого) CD269 (17 представитель суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Q02223 (версия записи 125, версия последовательности 2); последовательность(и) (человеческого) CD270 (14 представитель суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Q92956 (версия записи 134, версия последовательности 3); последовательность(и) (человеческого) CD271 (16 представитель суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Р08138 (версия записи 135, версия последовательности 1); последовательность(и) (человеческого) CD276 (антиген CD276) находи(я)тся в базе данных Swiss-Prot под номером Q5ZPR3 (версия записи 71, версия последовательности 1); последовательность(и) (человеческого) CD277 (представитель А1 подсемейства 3 бутирофилина) находи(я)тся в базе данных Swiss-Prot под номером O00481 (версия записи 102, версия последовательности 3); последовательность(и) (человеческого) CD280 (маннозный рецептор С-типа 2) находи(я)тся в базе данных Swiss-Prot под номером Q9UBG0 (версия записи 79, версия последовательности 2); последовательность(и) (человеческого) CD281 (Toll-подобный рецептор 1) находи(я)тся в базе данных Swiss-Prot под номером Q15399 (версия записи 125, версия последовательности 3); последовательность(и) (человеческого) CD282 (Toll-подобный рецептор 2) находи(я)тся в базе данных Swiss-Prot под номером O60603 (версия записи 129, версия последовательности 1); последовательность(и) (человеческого) CD283 (Toll-подобный рецептор 3) находи(я)тся в базе данных Swiss-Prot под номером O15455 (версия записи 120, версия последовательности 1); последовательность(и) (человеческого) CD284 (Toll-подобный рецептор 4) находи(я)тся в базе данных Swiss-Prot под номером O00206 (версия записи 125, версия последовательности 2); последовательность(и) (человеческого) CD286 (Toll-подобный рецептор 6) находи(я)тся в базе данных Swiss-Prot под номером Q9Y2C9 (версия записи 108, версия последовательности 2); последовательность(и) (человеческого) CD288 (Toll-подобный рецептор 8) находи(я)тся в базе данных Swiss-Prot под номером Q9NR97 (версия записи 103, версия последовательности 1); последовательность(и) (человеческого) CD289 (Toll-подобный рецептор 9) находи(я)тся в базе данных Swiss-Prot под номером Q9NR96 (версия записи 107, версия последовательности 2); последовательность(и) (человеческого) CD290 (Toll-подобный рецептор 10) находи(я)тся в базе данных Swiss-Prot под номером Q9BXR5 (версия записи 105, версия последовательности 2); последовательность(и) (человеческого) CD292 (рецептор костного морфогенетического белка типа-1А) находи(я)тся в базе данных Swiss-Prot под номером Р36894 (версия записи 146, версия последовательности 2); последовательность(и) (человеческого) CD294 (предполагаемый сопряженный с G-белком рецептор 44) находи(я)тся в базе данных Swiss-Prot под номером Q9Y5Y4 (версия записи 91, версия последовательности 3); последовательность(и) (человеческого) CD295 (рецептор лептина) находи(я)тся в базе данных Swiss-Prot под номером Р48357 (версия записи 132, версия последовательности 2); последовательность(и) (человеческого) CD296 (GPI-связанная NAD(P)(+)-аргинин-АДФ-рибозилтрансфераза 1) находи(я)тся в базе данных Swiss-Prot под номером Р52961 (версия записи 96, версия последовательности 2); последовательность(и) (человеческого) CD297 (экто-АДФ-рибозилтрансфераза 4) находи(я)тся в базе данных Swiss-Prot под номером Q93070 (версия записи 106, версия последовательности 2); последовательность(и) (человеческого) CD298 (субъединица бета-3 Na/K-АТФазы) находи(я)тся в базе данных Swiss-Prot под номером Р54709 (версия записи 102, версия последовательности 1); последовательность(и) (человеческого) CD299 (представитель M 4 семейства содержащих лектиновый домен типа С) находи(я)тся в базе данных Swiss-Prot под номером Q9H2X3 (версия записи 108, версия последовательности 1); последовательность(и) (человеческого) CD300 (CMRF35-подобная молекула 9) находи(я)тся в базе данных Swiss-Prot под номером Q6UXG3 (версия записи 67, версия последовательности 2); последовательность(и) (человеческого) CD301 (представитель А семейства 10 содержащих лектиновый домен типа С) находи(я)тся в базе данных Swiss-Prot под номером Q8IUN9 (версия записи 80, версия последовательности 1); последовательность(и) (человеческого) CD302 (антиген CD302) находи(я)тся в базе данных Swiss-Prot под номером Q8IX05 (версия записи 64, версия последовательности 1); последовательность(и) (человеческого) CD303 (представитель С семейства 4 содержащих лектиновый домен типа С) находи(я)тся в базе данных Swiss-Prot под номером Q8WTT0 (версия записи 82, версия последовательности 1); последовательность(и) (человеческого) CD304 (нейропилин-1) находи(я)тся в базе данных Swiss-Prot под номером 014786 (версия записи 129, версия последовательности 3); последовательность(и) (человеческого) CD305 (ассоциированный с лейкоцитами иммуноглобулин-подобный рецептор 1) находи(я)тся в базе данных Swiss-Prot под номером Q6GTX8 (версия записи 70, версия последовательности 1); последовательность(и) (человеческого) CD306 (ассоциированный с лейкоцитами иммуноглобулин-подобный рецептор 2) находи(я)тся в базе данных Swiss-Prot под номером Q6ISS4 (версия записи 63, версия последовательности 1); последовательность(и) (человеческого) CD309 (2 рецептор фактора роста сосудистого эндотелия) находи(я)тся в базе данных Swiss-Prot под номером Р35968 (версия записи 138, версия последовательности 2); последовательность(и) (человеческого) CD312 (содержащий EGF-подобный модуль муциноподобный подобный рецептору гормона 2) находи(я)тся в базе данных Swiss-Prot под номером Q9UHX3 (версия записи 113, версия последовательности 2); последовательность(и) (человеческого) CD314 (интегральный мембранный белок NKG2-D типа II) находи(я)тся в базе данных Swiss-Prot под номером Р26718 (версия записи 117, версия последовательности 1); последовательность(и) (человеческого) CD315 (негативный регулятор рецептора простагландина F2) находи(я)тся в базе данных Swiss-Prot под номером Q9P2B2 (версия записи 98, версия последовательности 2); последовательность(и) (человеческого) CD316 (8 представитель суперсемейства иммуноглобулинов) находи(я)тся в базе данных Swiss-Prot под номером Q969P0 (версия записи 81, версия последовательности 1); последовательность(и) (человеческого) CD317 (костномозговой стромальный антиген 2) находи(я)тся в базе данных Swiss-Prot под номером Q10589 (версия записи 95, версия последовательности 1); последовательность(и) (человеческого) CD318 (содержащий домен CUB белок 1) находи(я)тся в базе данных Swiss-Prot под номером Q9H5V8 (версия записи 78, версия последовательности 3); последовательность(и) (человеческого) CD319 (7 представитель семейства SLAM) находи(я)тся в базе данных Swiss-Prot под номером Q9NQ25 (версия записи 92, версия последовательности 1); последовательность(и) (человеческого) CD320 (антиген CD320) находи(я)тся в базе данных Swiss-Prot под номером Q9NPF0 (версия записи 86, версия последовательности 1); последовательность(и) (человеческого) CD321 (адгезионная молекула А межклеточных контактов) находи(я)тся в базе данных Swiss-Prot под номером Q9Y624 (версия записи 124, версия последовательности 1); последовательность(и) (человеческого) CD322 (адгезионная молекула В межклеточных контактов) находи(я)тся в базе данных Swiss-Prot под номером Р57087 (версия записи 107, версия последовательности 1); последовательность(и) (человеческого) CD324 (кадгерин-1) находи(я)тся в базе данных Swiss-Prot под номером Р12830 (версия записи 157, версия последовательности 3); последовательность(и) (человеческого) CD325 (кадгерин-2) находи(я)тся в базе данных Swiss-Prot под номером Р19022 (версия записи 118, версия последовательности 4), последовательность(и) (человеческого) CD326 (адгезивная молекула эпителиальных клеток) находи(я)тся в базе данных Swiss-Prot под номером Р16422 (версия записи 118, версия последовательности 2); последовательность(и) (человеческого) CD327 (связывающий сиаловые кислоты иммуноглобулин-подобный лектин 6) находи(я)тся в базе данных Swiss-Prot под номером O43699 (версия записи 107, версия последовательности 2); последовательность(и) (человеческого) CD328 (связывающий сиаловые кислоты иммуноглобулин-подобный лектин 7) находи(я)тся в базе данных Swiss-Prot под номером Q9Y286 (версия записи 111, версия последовательности 1); последовательность(и) (человеческого) CD329 (связывающий сиаловые кислоты иммуноглобулин-подобный лектин 8) находи(я)тся в базе данных Swiss-Prot под номером Q9NYZ4 (версия записи 100, версия последовательности 2); последовательность(и) (человеческого) CD331 (рецептор 1 фактора роста фибробластов) находи(я)тся в базе данных Swiss-Prot под номером Р11362 (версия записи 169, версия последовательности 3); последовательность(и) (человеческого) CD332 (рецептор 2 фактора роста фибробластов) находи(я)тся в базе данных Swiss-Prot под номером Р21802 (версия записи 165, версия последовательности 1); последовательность(и) (человеческого) CD333 (рецептор 3 фактора роста фибробластов) находи(я)тся в базе данных Swiss-Prot под номером Р22607 (версия записи 161, версия последовательности 1); последовательность(и) (человеческого) CD334 (рецептор 4 фактора роста фибробластов) находи(я)тся в базе данных Swiss-Prot под номером Р22455 (версия записи 136, версия последовательности 2); последовательность(и) (человеческого) CD335 (рецептор естественной цитотоксичности 1) находи(я)тся в базе данных Swiss-Prot под номером O76036 (версия записи 98, версия последовательности 1); последовательность(и) (человеческого) CD336 (рецептор естественной цитотоксичности 2) находи(я)тся в базе данных Swiss-Prot под номером O95944 (версия записи 86, версия последовательности 2); последовательность(и) (человеческого) CD337 (рецептор естественной цитотоксичности 3) находи(я)тся в базе данных Swiss-Prot под номером O14931 (версия записи 103, версия последовательности 1); последовательность(и) (человеческого) CD338 (2 представитель подсемейства транспортеров АТФ-связывающих кассет) находи(я)тся в базе данных Swiss-Prot под номером Q9UNQ0 (версия записи 120, версия последовательности 3); последовательность(и) (человеческого) CD339 (белок jagged-1) находи(я)тся в базе данных Swiss-Prot под номером Р78504 (версия записи (версия записи 129; версия последовательности 3); последовательность(и) (человеческого) CD340 (рецепторная тирозиновая протеинкиназа erbB-2) находи(я)тся в базе данных Swiss-Prot под номером Р04626 (версия записи 162, версия последовательности 1); последовательность(и) (человеческого) CD344 (Frizzled-4) находи(я)тся в базе данных Swiss-Prot под номером Q9ULV1 (версия записи 107, версия последовательности 2); последовательность(и) (человеческого) CD349 (Frizzled-9) находи(я)тся в базе данных Swiss-Prot под номером O00144 (версия записи 103, версия последовательности 1); последовательность(и) (человеческого) CD350 (Frizzled-10) находи(я)тся в базе данных Swiss-Prot под номером Q9ULW2 (версия записи 100, версия последовательности 1); последовательность(и) (человеческого) CD351 (высокоаффинный Fc-рецептор IgA и IgM) находи(я)тся в базе данных Swiss-Prot под номером Q8WWV6 (версия записи 65, версия последовательности 1); последовательность(и) (человеческого) CD352 (6 представитель семейства SLAM) находи(я)тся в базе данных Swiss-Prot под номером Q96DU3 (версия записи 93, версия последовательности 3); последовательность(и) (человеческого) CD353 (8 представитель семейства SLAM) находи(я)тся в базе данных Swiss-Prot под номером Q9P0V8 (версия записи 80, версия последовательности 1); последовательность(и) (человеческого) CD354 (инициирующий рецептор 1, экспрессируемый на миелоидных клетках) находи(я)тся в базе данных Swiss-Prot под номером Q9NP99 (версия записи 93, версия последовательности 1); последовательность(и) (человеческого) CD355 (молекула цитотоксических и регуляторных Т-клеток) находи(я)тся в базе данных Swiss-Prot под номером 095727 (версия записи 81, версия последовательности 2); последовательность(и) (человеческого) CD357 (18 представитель суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером Q9Y5U5 (версия записи 103, версия последовательности 1); последовательность(и) (человеческого) CD358 (21 представитель суперсемейства рецепторов фактора некроза опухоли) находи(я)тся в базе данных Swiss-Prot под номером 075509 (версия записи 110, версия последовательности 1); последовательность(и) (человеческого) CD360 (рецептор интерлейкина-21) находи(я)тся в базе данных Swiss-Prot под номером Q9HBE5 (версия записи 104, версия последовательности 1); последовательность(и) (человеческого) CD361 (белок EVI2B) находи(я)тся в базе данных Swiss-Prot под номером Р34910 (версия записи 87, версия последовательности 2); последовательность(и) (человеческого) CD362 (синдекан-2) находи(я)тся в базе данных Swiss-Prot под номером Р34741 (версия записи 105, версия последовательности 2); последовательность(и) (человеческого) CD363 (рецептор 1 сфингозин-1-фосфата) находи(я)тся в базе данных Swiss-Prot под номером Р21453 (версия записи 116, версия последовательности 2); последовательность(и) (человеческого) белка Cripto (Criptic family protein 1-B) находи(я)тся в базе данных Swiss-Prot под номером P0CG36 (версия записи 12, версия последовательности 1); последовательность(и) (человеческого) рецептора тиротропина (TSHR) находи(я)тся в базе данных Swiss-Prot под номером Р16473 (версия записи 152, версия последовательности 2); или последовательность(и) (человеческого) рецептора эпидермального фактора роста (EGFR) находи(я)тся в базе данных Swiss-Prot под номером Р00533 (версия записи 178, версия последовательности 2).
Как упоминалось выше, второй (lg-производный) домен описанной выше молекулы биспецифического антитела может содержать сайт, взаимодействующий с антигеном, специфический в отношении молекулы клеточной поверхности, существующей в естественных условиях на опухолевых клетках.
Термин "молекула клеточной поверхности, существующая в естественных условиях на опухолевых клетках", используемый в данном документе, также обозначает молекулы, презентируемые на поверхности опухолевых клеток. Термин "существует в естественных условиях" относится к молекулам, которые эндогенно экспрессируются на поверхности опухолевых клеток. Термин "молекула клеточной поверхности" относится к молекулам, которые (естественным образом/эндогенно) экспрессируются/презентируются на поверхности клеток и содержат домены или эпитопы, доступные (in vitro или in vivo) для (lg-производных) связывающих доменов, предпочтительно антител, фрагментов антител или производных. Как показано выше, указанный второй Ig-производный связывающий домен может представлять собой scFv. Примерами указанных молекул клеточной поверхности являются мембранные и трансмембранные белки, молекулы, адаптированные к указанным белкам или клеточной поверхности и т.д. Таким образом, в контексте изобретения указанные молекулы клеточной поверхности представляют собой опухолеспецифические маркеры. В контексте изобретения указанный опухолеспецифический маркер относится к маркерам, которые обычно эндогенно экспрессируются на поверхности опухолевых клеток.
В контексте данного изобретения термин "опухолеспецифический маркер" относится к молекулам, которые естественным образом/эндогенно презентируются и/или локализуются на поверхности опухолевых клеток или которые экспрессируются повсеместно, но являются доступными только для связывания с антителами, фрагментами антител или производными антител на поверхности опухолевых клеток. "Опухолеспецифический маркер" в данном описании обозначает белок, экспрессирующийся преимущественно или исключительно на опухолевых клетках. Преимущественно означает относительно более высокий уровень экспрессии на опухолевых клетках по сравнению с нормальными соматическими клетками, тогда как исключительно означает экспрессию белка на опухолевых клетках, не обнаруживаемого на соматических клетках стандартными способами определения белка, известными специалистам. Белки, удовлетворяющие данным критериям, можно, например, идентифицировать путем вычитающей и сравнительной гибридизации, хорошо известных в области техники. Степень опухолеспецифической экспрессии, необходимую для применения способа лечения по данному изобретению, можно определить при помощи анализа с использованием клеток, в котором клетки, экспрессирующие антиген, представляющий интерес, и Т-клетки, специфические в отношении этого антигена, инкубируют вместе и определяют специфическую цитотоксическую активность.
"Преимущественная экспрессия" относится к белкам, для которых характерна повышенная экспрессия опухолевыми клетками по сравнению с нормальными клетками вследствие сверхэкспрессии белка в результате амплификации гена, усиления транскрипции или стабилизации мРНК или в результате мутации, влияющей на обмен таких белков. "Преимущественно" также относится к белкам, которые экспрессируются на опухолевых клетках и на нормальных клетках, но при этом нормальные клетки обычно являются недоступными для Т-клеток или антител, например, в иммунологически привилегированных зонах организма человека. Кроме того, в контексте лечения данное определение охватывает белки, экспрессирующиеся на опухолевых клетках, но не экспрессирующиеся на нормальных клетках, например, белки, которые экспрессируются исключительно при эмбриональном развитии.
"Исключительная экспрессия" относится к белкам, которые в ходе лечения обнаруживаются только на опухолевых клетках. Предпочтительно, такие белки экспонируются на клеточной поверхности и несут точечные мутации или делеции в своей внеклеточной части, отсутствующие у нормальных клеток. Аналогично, к этой категории относятся неоэпитопы, образующиеся в результате опухолеспецифической активности шеддаз. "Исключительная экспрессия" также включает аномальные гликоструктуры, обнаруживаемые исключительно у опухолевых, но не у нормальных клеток.
В контексте данного изобретения первый связывающий домен, описанный в данном документе, и второй связывающий домен, описанный в данном документе, принадлежащие описанной здесь молекуле биспецифического антитела, связываются с различными антигенами.
Примеры опухолевых маркеров, которые присутствуют на поверхности опухолевых клеток, приведены ниже и включают ЕрСАМ, HER-1, HER-2, HER-3, CD20, CD22, CD33, CD52, СА-12-5, HLA-DR, MUC-1 (муцин), А33-антиген, PSMA (проста-специфический мембранный антиген), рецептор трансферрина, тенасцин или CA-IX, но не ограничиваются ими.
Таким образом, в контексте данного изобретения, описанные в данном документе молекулы биспецифических антител включают антиген/маркер, который существует в естественных условиях на поверхности опухолевых клеток, выбранный из ЕрСАМ, HER-1, HER-2, HER-3, CD20, CD22, CD33, CD52, СА-12-5, HLA-DR, MUC-1 (муцина), А33-антигена, PSMA (простат-специфического мембранного антигена), рецептора трансферрина, тенасцина и CA-IX. В контексте данного изобретения описанные здесь молекулы биспецифических антител включают антиген/маркер, который эндогенно экспрессируется на поверхности опухолевых клеток, выбранный из группы, состоящей из ЕрСАМ, HER-1, HER-2, HER-3, CD20, CD22, CD33, CD52, СА-12-5, HLA-DR, MUC-1 (муцина), А33-антигена, PSMA (простат-специфического мембранного антигена), рецептора трансферрина, тенасцина и CA-IX.
Последовательность(и) (человеческих) ЕрСАМ, HER-1, HER-2, HER-3, CD20, CD22, CD33, CD52, СА-12-5, HLA-DR, MUC-1 (муцина), А33-антигена, PSMA (простат-специфического мембранного антигена), рецептора трансферрина, тенасцина или CA-IX содержатся в базе данных UniProtKB/Swiss-Prot и доступ к ним может быть получен по ссылке http://www.uniprot.org/uniprot/?query=reviewed%3Ayes. Данные (белковые) последовательности также охватывают аннотированные модифицированные последовательности. В данном изобретении также предложены методы и способы, в которых используются гомологичные последовательности, а также аллельные варианты генов и т.п. приведенных в данном документе конкретных последовательностей. Предпочтительно, используются такие "варианты" и т.п. конкретных последовательностей. Предпочтительно, такие "варианты" представляют собой варианты генов. Специалист в данной области техники легко может найти соответствующую кодирующую область таких (белковых) последовательностей среди записей в базах данных, которые могут также включать записи последовательностей геномной ДНК, а также мРНК/кДНК.
Последовательность(и) ЕрСАМ (адгезивной молекулы эпителиальных клеток) (человека) находи(я)тся в базе данных Swiss-Prot под номером Р16422 (версия записи 117, версия последовательности 2); последовательность(и) (человеческого) HER-1 (рецептора эпидермального фактора роста) находи(я)тся в базе данных Swiss-Prot под номером Р00533 (версия записи 177, версия последовательности 2); последовательность(и) (человеческого) HER-2 (рецептора тирозиновой протеинкиназы erbB-2) находи(я)тся в базе данных Swiss-Prot под номером Р04626 (версия записи 161, версия последовательности 1); последовательность(и) (человеческого) HER-3 (рецептора тирозиновой протеинкиназы erbB-3) находи(я)тся в базе данных Swiss-Prot под номером Р21860 (версия записи 140, версия последовательности 1); последовательность(и) (человеческого) CD20 (В-лимфоцитарный антиген CD20) находи(я)тся в базе данных Swiss-Prot под номером Р11836 (версия записи 117, версия последовательности 1); последовательность(и) (человеческого) CD22 (В-лимфоцитарный антиген CD22) находи(я)тся в базе данных Swiss-Prot под номером Р20273 (версия записи 135, версия последовательности 2); последовательность(и) (человеческого) CD33 (В-лимфоцитарный антиген CD33) находи(я)тся в базе данных Swiss-Prot под номером Р20138 (версия записи 129, версия последовательности 2); последовательность(и) (человеческого) СА-12-5 (муцин 16) находи(я)тся в базе данных Swiss-Prot под номером Q8WXI7 (версия записи 66, версия последовательности 2); последовательность(и) (человеческого) HLA-DR находи(я)тся в базе данных Swiss-Prot под номером Q29900 (версия записи 59, версия последовательности 1); последовательность(и) (человеческого) MUC-1 (муцин-1) находи(я)тся в базе данных Swiss-Prot под номером Р15941 (версия записи 135, версия последовательности 3); последовательность(и) (человеческого) А33 (антиген клеточной поверхности А33) находи(я)тся в базе данных Swiss-Prot под номером Q99795 (версия записи 104, версия последовательности 1); последовательность(и) (человеческого) PSMA (глутамат-карбоксипептидаза 2) находи(я)тся в базе данных Swiss-Prot под номером Q04609 (версия записи 133, версия последовательности 1), последовательность(и) (человеческого) рецептора трансферрина находи(я)тся в базе данных Swiss-Prot под номером Q9UP52 (версия записи 99, версия последовательности 1) и Р02786 (версия записи 152, версия последовательности 2); последовательность (человеческого) тенасцина находи(я)тся в базе данных Swiss-Prot под номером Р24821 (версия записи 141, версия последовательности 3); или последовательность(и) (человеческого) CA-IX (карбоангидразы 9) находи(я)тся в базе данных Swiss-Prot под номером Q16790 (версия записи 115, версия последовательности 2).
В контексте данного изобретения описано биспецифическое антитело, которое содержит первый связывающий домен, связывающийся с/направленный против/взаимодействующий с (человеческим) белком Cripto, и второй домен, связывающийся с/направленный против/взаимодействующий с ЕрСАМ (человека).
Предложенные здесь молекулы или конструкции (т.е. описанные в данном документе молекулы биспецифических антител) особенно подходят для применения в медицинских учреждениях. Например, с помощью описанных здесь биспецифических конструкций можно осуществлять лечение онкологических заболеваний. В контексте данного изобретения онкологическое заболевание может представлять собой злокачественное новообразование эпителиального, эндотелиального или мезотелиального происхождения или злокачественное новообразование кроветворной ткани. В контексте данного изобретения злокачественное новообразование выбрано из группы, состоящей из рака желудочно-кишечного тракта, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи, рака полости рта, рака желудка, рака шейки матки, В и Т-клеточной лимфомы, миелолейкоза, рака яичников, лейкоза, лимфолейкоза, носоглоточной карциномы, рака толстой кишки, рака предстательной железы, почечноклеточного рака, злокачественных опухолей головы и шеи, рака кожи (меланомы), рака мочеполового тракта, например, рака яичка, рака яичников, эндотелиомы, рака шейки матки и рака почки, рака желчевыводящих путей, рака пищевода, рака слюнных желез и рака щитовидной железы или других злокачественных заболеваний, таких как гемобластозы, глиомы, саркомы или остеосаркомы.
Предложенные здесь молекулы или конструкции (т.е. описанные в данном документе молекулы биспецифических антител) особенно подходят для применения в медицинских учреждениях. Например, можно осуществлять лечение опухолевых заболеваний и/или лимфом с помощью биспецифических конструкций, предназначенных для борьбы с этими заболеваниями. Показанием для назначения (молекулы) биспецифического антитела является экспрессия опухолевого антигена. Опухолевый антиген, экспрессируемый в организме, можно скомбинировать практически с любым упомянутым выше Т-клеточным маркером (представляющим антиген, существующий в естественных условиях/эндогенно экспрессируемый на поверхности опухолевых клеток). Например, можно осуществлять лечение рака ЖКТ, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи и/или рака полости рта с использованием биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифических антител), у которых второй связывающий домен направлен против ЕрСАМ (человека) (как опухолеспецифического антигена, существующего в естественных условиях на поверхности опухолевых клеток), и содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающимся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), Таким образом, в контексте данного изобретения конструкцию биспецифического антитела, распознающего ЕрСАМ (человека) (посредством второго связывающего домена) и содержащего первый связывающий домен, направленный против/связывающийся с/взаимодействующий с белком Cripto, можно использовать для лечения рака ЖКТ, например, аденокарциномы ЖКТ. Можно осуществлять лечение рака ЖКТ, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи и/или рака полости рта с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) HER1 (как опухолеспецифического антигена, существующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение рака желудка, рака молочной железы и/или рака шейки матки с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) HER2 (как опухоле-специфического антигена, существующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение рака желудка и/или рака легких с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) HER3 (как опухоле-специфического антигена, существующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение В-клеточной лимфомы и/или Т-клеточной лимфомы с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) CD20 (как опухоле-специфического антигена, существующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение В-клеточной лимфомы и/или Т-клеточной лимфомы с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) CD22 (как опухоле-специфического антигена, существующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение миелолейкоза с помощью биспецифических конструкций, у которых второй связывающий домен направлен против (человеческого) CD33 (как опухоле-специфического антигена, существующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение рака яичников, рака легких, рака молочной железы и/или рака ЖКТ с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) СА12-5 (как опухоле-специфического антигена, существующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение рака ЖКТ, лейкоза и/или носоглоточной карциномы с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) HLA-DR (как опухоле-специфического антигена, существующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение рака толстой кишки, рака молочной железы, рака яичников, рака легких и/или рака поджелудочной железы с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) MUC-1 (как опухолеспецифического антигена, существующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение рака толстой кишки с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) А33 (как опухоле-специфического антигена, присутствующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение рака предстательной железы с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) PSMA (как опухоле-специфического антигена, существующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение рака ЖКТ, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи и/или рака полости рта с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) рецептора трансферрина (как опухоле-специфического антигена, существующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение рака поджелудочной железы, рака легких и/или рака молочной железы с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) рецептора трансферрина (как опухоле-специфического антигена, существующего в естественных условиях на поверхности опухолевых клеток), и содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках). Можно осуществлять лечение рака почки с помощью биспецифических молекул или конструкций (т.е. описанных здесь молекул биспецифического антитела), у которых второй связывающий домен направлен против (человеческого) CA-IX (как опухоле-специфического антигена, существующего в естественных условиях на поверхности опухолевых клеток), содержащих первый связывающий домен, направленный против/связывающийся с/взаимодействующий с одним из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках).
Кроме того, в Примерах, приведенных в качестве подтверждения концепции данного изобретения, молекула специфического биспецифического антитела по изобретению содержит указанный выше первый (lg-производный) домен, связывающийся с/направленный против/взаимодействующий с (человеческим) EGFR, и второй (lg-производный) домен, связывающийся с/направленный против/взаимодействующий с ЕрСАМ (человека).
Адгезивная молекула эпителиальных клеток (ЕрСАМ, также обозначаемая 17-1А антиген, KSA, EGP40, GA733-2, ks1-4, или esa) представляет собой 40-кДа мембранный гликопротеин, состоящий из 314 аминокислот, специфически экспрессирующийся в эпителии и при различных видах карцином человека (см. обзор Balzar, J. Mol. Med. (1999), 77, 699-712). ЕрСАМ открыли и впоследствии клонировали благодаря распознаванию мышиным моноклональным антителом 17-1А/эдреколомабом (Goettlinger, Int J Cancer 38 (1986), 47-53 и Simon, Proc. Natl. Acad. Sci. USA 87 (1990), 2755-2759). ЕрСАМ обеспечивает ориентированную и высоко упорядоченную адгезию эпителиальных клеток (Litvinov, J Cell Biol. 139 (1997), 1337-1348). При злокачественной трансформации эпителиальных клеток быстро растущие опухолевые клетки теряют упорядоченное расположение, характерное для клеток эпителия. Вследствие этого поверхностное распределение ЕрСАМ становится менее локализованным, молекулы лучше экспонируются на опухолевых клетках и оказываются доступными для связывания с антителами, фрагментами антител или производными антител на поверхности опухолевых клеток. Опухолевые клетки большинства карцином продолжают экспрессировать ЕрСАМ на своей поверхности, поскольку происходят из эпителиальных клеток.
In vivo наблюдается зависимость между экспрессией ЕрСАМ и усиленной пролиферацией эпителия и обратная корреляция с клеточной дифференцировкой (см. обзор Balzar, J. Mol. Med. 77 (1999), 699-712). Экспрессия ЕрСАМ наблюдается при большинстве карцином (см. обзор Balzar, J. Mol. Med. 77 (1999), 699-712 или публикации De Bree, Nucl Med Commun. 15 (1994), 613-27; Zhang, Clin Cancer Res. 4 (1998), 295-302). Благодаря такой распространенной экспрессии ЕрСАМ считают "карцинома-ассоциированным" антигеном. Во многих случаях наблюдалась более высокая экспрессия ЕрСАМ в опухолевых клетках по сравнению с эпителиальными клетками, от которых они произошли, или с менее агрессивными формами указанных типов рака. Например, повышенная экспрессия ЕрСАМ является ранним событием при развитии рака предстательной железы (Poczatek, J. Urol. 162 (1999), 1462-1644). Кроме того, при большинстве плоскоклеточных карцином и аденокарцином шейки матки выраженная экспрессия ЕрСАМ коррелирует с усиленной пролиферацией и исчезновением маркеров терминальной дифференцировки (Litvinov, Am. J. Pathol. 148 (1996), 865-75). При раке молочной железы сверхэкспрессия ЕрСАМ в опухолевых клетках является предиктором выживаемости (Gastl, Lancet 356 (2000), 1981-1982). ЕрСАМ является маркером, используемым для детекции диссеминированных опухолевых клеток у пациентов с плоскоклеточной карциномой головы, шеи и легких (Chaubal, Anticancer Res. 19 (1999), 2237-2242 и Piyathilake, Hum. Pathol. 31 (2000), 482-487). В нормальном плоском эпителии, обнаруживаемом в эпидермисе, ротовой полости, надгортаннике, глотке, гортани и пищеводе, на наблюдается выраженной экспрессии ЕрСАМ (Quak, Hybridoma 9 (1990), 377-387). Было показано, что ЕрСАМ экспрессирует большинство клеток первичного, метастазирующего и диссеминированного НМКРЛ (немелкоклеточного рака легких (Passlick, Int J Cancer 87 (2000), 548-552)), аденокарцином желудка и пищеводно-желудочного перехода (Martin, J. Clin. Pathol. 52 (1999), 701-4) и клеточных линий, ведущих свое происхождение от карцином толстой кишки, поджелудочной железы и молочной железы (Szala, Proc. Natl. Acad. Sci. USA 87 (1990), 3542-6 и Packeisen, Hybridoma 18(1999), 37-40).
Как проиллюстрировано Примерами, приведенными для подтверждения концепции данного изобретения, антитело к (человеческому) EGFR скомбинировали с антителом к ЕрСАМ (мыши) (G8.8) для получения биспецифической конструкции MAb225_scFv_G8.8. Аминокислотная последовательность легкой цепи антитела к (человеческому) EGFR приведена ниже (обозначена SEQ ID NO: 1):

Аминокислотная последовательность тяжелой цепи антитела к (человеческому) EGFR приведена ниже (обозначена SEQ ID NO: 2):


Аминокислотная последовательность легкой цепи антитела к ЕрСАМ (мыши) (G8.8) приведена ниже (обозначена SEQ ID NO: 3):

Аминокислотная последовательность тяжелой цепи антитела к ЕрСАМ (мыши) (G8.8) приведена ниже (обозначена SEQ ID NO: 4):
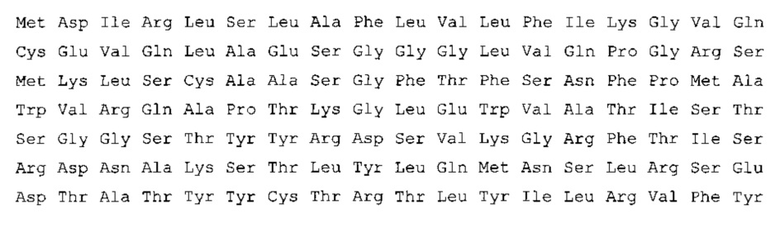

Аминокислотная последовательность легкой цепи биспецифического продукта (MAb225_scFv_G8.8) приведена ниже (обозначена SEQ ID NO: 5):

Аминокислотная последовательность тяжелой цепи биспецифического продукта (MAb225_scFv_G8.8) приведена ниже (обозначена SEQ ID NO: 6):


Кроме того, как показано на Фиг. 20 и 21, в качестве (дополнительного) подтверждения концепции данного изобретения, было создано биспецифическое антитело (бсАт) "EpCAM-EGFRvIII, MR1.1", имеющее антигенсвязывающий сайт для (человеческого) del-hEGFRvIII на одном плече и для (мышиного) ЕрСАМ на другом плече; см. Пример 4. Аминокислотная последовательность легкой цепи биспецифического продукта (EpCAM-EGFRvIII, MR1.1) показана на Фиг. 23А (обозначена SEQ ID NO: 15). Аминокислотная последовательность тяжелой цепи биспецифического продукта (EpCAM-EGFRvIII, MR1.1) показана на Фиг. 23 В (обозначена SEQ ID NO: 16).
В изобретении также предложены последовательности нуклеиновых кислот, кодирующих молекулу биспецифического антитела по изобретению.
Для специалиста в данной области техники является очевидным, что к молекуле по изобретению можно присоединить регуляторные последовательности. Например, можно использовать промоторы, энхансеры транскрипции и/или последовательности, позволяющие индуцировать экспрессию полинуклеотида по изобретению. Подходящей индуцибельной системой является, например, тетрациклин-регулируемая экспрессия генов, описанная, например, Gossen and Bujard Proc. Natl. Acad. Sci. USA 89 (1992), 5547-5551 и Gossen et al. Trends Biotech. 12 (1994), 58-62, или дексаметазон-индуцируемая система экспрессии генов, описанная, например, Crook ЕМВО J. 8 (1989), 513-519.
Кроме того, предусматривается, что молекула нуклеиновой кислоты может содержать, например, тиоэфирные связи и/или аналоги нуклеотидов. Указанные модификации могут придать молекуле нуклеиновой кислоты стабильность к воздействию эндо- и/или экзонуклеаз в клетке. Транскрипция указанных молекул нуклеиновой кислоты может осуществляться благодаря соответствующим векторам, содержащим химерный ген, обеспечивающий транскрипцию указанной молекулы нуклеиновой кислоты в клетке. При этом следует также учитывать, что данный полинуклеотид можно использовать для "направленного воздействия на ген" или "генной терапии". В другом воплощении указанные молекулы нуклеиновой кислоты являются меченными. Способы детекции нуклеиновых кислот хорошо известны в области техники, например, саузерн блоттинг и нозерн-блоттинг, ПЦР или удлинение праймеров. Данное воплощение может найти применение в способах скрининга для верификации успешного введения описанных выше молекул нуклеиновой кислоты при генной терапии.
Указанные молекулы нуклеиновых кислот могут представлять собой полученные рекомбинантным способом химерные молекулы нуклеиновых кислот, содержащие любую из указанных выше молекул нуклеиновых кислот по отдельности или в комбинации. В контексте данного изобретения молекула нуклеиновой кислоты представляет собой часть вектора.
Таким образом, данное изобретение также относится к вектору, содержащему молекулу нуклеиновой кислоты, описанную в данном изобретении.
Специалистам в молекулярной биологии известны многие подходящие векторы, выбор которых будет зависеть от желаемой функции, и включают плазмиды, космиды, вирусы, бактериофаги и другие векторы, обычно используемые в генной инженерии. Способы, хорошо известные специалистам, можно применять для конструирования различных плазмид и векторов, см., например, методы, описанные Sambrook et al. (loc cit.) и Ausubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y. (1989), (1994). В альтернативном случае, полинуклеотиды и векторы по изобретению можно встраивать в липосомы для доставки в клетки-мишени. Как подробно обсуждается ниже, для выделения отдельных последовательностей ДНК использовали клонирующий вектор. Когда требуется экспрессия конкретного полипептида, соответствующие последовательности можно встраивать в экспрессирующие векторы. Стандартные клонирующие векторы включают pBluescript SK, pGEM, pUC9, pBR322, pGA18 и pGBT9. Стандартные экспрессирующие векторы включают pTRE, pCAL-n-EK, pESP-1, pOP13CAT.
Изобретение также относится к вектору, содержащему последовательность нуклеиновой кислоты, которая представляет собой регуляторную последовательность, функционально сязанную с указанной последовательностью нуклеиновой кислоты, кодирующей конструкцию (молекулу) биспецифического антитела, описанную в данном документе.
Такие регуляторные последовательности (контролирующие элементы) известны специалистам и могут включать промотор, сплайс-кассету, кодон инициации трансляции и сайт интеграции для встраивания вставки в вектор. В контексте данного изобретения указанная молекула нуклеиновой кислоты функционально связана с указанной последовательностью, контролирующей экспрессию, что обеспечивает экспрессию в эукариотических или прокариотических клетках.
Предусматривается, что указанный вектор представляет собой экспрессирующий вектор, содержащий молекулу нуклеиновой кислоты, кодирующую конструкцию (молекулу) биспецифического антитела, описанную в данном документе.
Термин "регуляторная последовательность" обозначает последовательности ДНК, необходимые для экспрессии кодирующих последовательностей, с которыми они лигированы. Природа таких контролирующих последовательностей различается в зависимости от организма-хозяина. У прокариот контролирующие последовательности обычно включают промотор, сайт связывания рибосом и терминаторы. У эукариот контролирующие последовательности обычно включают промоторы, терминаторы и, в некоторых случаях, энхансеры, трансактиваторы или факторы транскрипции. Термин "контролирующая последовательность" охватывает по меньшей мере все компоненты, присутствие которых необходимо для экспрессии, и может также охватывать дополнительные предпочтительные компоненты.
Термин "функционально связан" означает смежное расположение, при котором взаимодействие описанных компонентов позволяет им функционировать надлежащим образом. Контролирующая последовательность, "функционально связанная" с кодирующей последовательностью, лигирована таким образом, что экспрессия кодирующей последовательности достигается в условиях, подходящих для контролирующей последовательности. В случае, если контролирующая последовательность является промотором, для специалиста очевидно, что предпочтительно использовать двуцепочечную нуклеиновую кислоту.
В контексте данного изобретения указанный вектор является экспресирующим вектором. "Экспрессирующий вектор" представляет собой конструкцию, которую можно использовать для трансформации выбранного хозяина и обеспечивать экспрессию кодирующей последовательности в выбранном хозяине. Экспрессирующие векторы могут быть, например, клонирующими векторами, бинарными векторами или интегрирующими векторами. Экспрессия включает транскрипцию молекулы нуклеиновой кислоты, предпочтительно транслируемой в мРНК. Регуляторные элементы, обеспечивающие экспрессию в прокариотических и/или эукариотических клетках, хорошо известны специалистам в данной области техники. В случае эукариотических клеток они обычно содержат промоторы, обеспечивающие инициацию транскрипции, и возможно, сигналы полиаденилирования, обеспечивающие терминацию транскрипции и стабилизацию транскрипта. Возможные регуляторные элементы, обеспечивающие экспрессию в прокариотических клетках-хозяевах, включают, например, PL, lac, trp или tac промотор Е. coli, а примерами регуляторных элементов, обеспечивающих экспресию в эукариотических клетках-хозяевах, являются промоторы АОХ1 или GAL1 для дрожжей или промоторы CMV-, SV40-, RSV (вируса саркомы Рауса), CMV-энхансер, SV40-энхансер или интрон глобина для клеток млекопитающих и других животных.
Помимо элементов, отвечающих за инициацию транскрипции, регуляторные элементы могут также включать сигналы терминации транскрипции, такие как сайт полиаденилирования SV40 или сайт полиаденилирования tk, расположенные в 3ʹ направлении от полинуклеотида. Кроме того, в зависимости от используемой системы экспрессии, к кодирующей последовательности упомянутой последовательности нуклеиновой кислоты могут быть добавлены лидерные последовательности, хорошо известные в области техники, способные направлять полипептид в клеточные компартменты или его секрецию в среду, см., например, приведенные Примеры.
Лидерные последовательности находятся в одной рамке считывания с транслируемыми, инициирующими и терминирующими последовательностями и предпочтительно, с лидерной последовательностью, способной направлять секрецию транслированного белка или его части в периплазматическое пространство или внеклеточную среду. Как вариант, гетерологичная последовательность может кодировать гибридный белок с N-концевым сигнальным пептидом, придающим желаемые характеристики экспрессируемому рекомбинантному продукту, например, для стабилизации или упрощения очистки; см. выше. В данном контексте в области техники известны подходящие векторы, такие как экспрессирующий вектор кДНК Okayama-Berg pcDV1 (Pharmacia), pCDM8, pRc/CMV, pcDNA1, pcDNA3 (Invitrogene), pEF-DHFR, pEF-ADA или pEF-neo (Raum et al. Cancer Immunol Immunother50 (2001), 141-150) или pSPORT1 (GIBCO BRL).
В контексте данного изобретения последовательности, контролирующие экспрессию, предствляют собой системы эукариотических промоторов в векторах, способных трансформировать или трансфецировать эукариотические клетки-хозяева, однако могут применяться и контролирующие последовательности для прокариотических хозяев. После введения вектора в соответствующего хозяина, хозяина поддерживают в условиях, подходящих для эффективной экспрессии нуклеотидных последовательностей, возможно с последующим выделением и очисткой полипептида по изобретению, как описано, например, в приведенных Примерах.
Альтернативной экспрессирующей системой, которую можно использовать для экспрессии белка, регулирующего клеточный цикл, является система клеток насекомых. В одной из таких систем в качестве вектора используют вирус ядерного полиэдроза совки калифорнийской люцерновой (Autographa californica) (AcNPV) для экспрессии чужеродных генов в клетках Spodoptera frugiperda или Trichoplusia larvae. Кодирующую последовательность упомянутой молекулы нуклеиновой кислоты можно клонировать в несущественную область генома вируса, такую как ген полиэдрина (полигедрина), и поместить под контроль промотора полиэдрина. Успешное встраивание указанной кодирующей последовательности инактивирует ген полиэдрина и образуется рекомбинантный вирус, не содержащий белка оболочки. Затем рекомбинантные вирусы используют для инфицирования клеток S. frugiperda или Trichoplusia larvae, в которых экспрессируется белок по изобретению (Smith, J. Virol. 46 (1983), 584; Engelhard, Proc. Nat. Acad. Sci. USA 91 (1994), 3224-3227).
Дополнительные регуляторные элементы могут включать энхансеры транскрипции и трансляции. Предпочтительно, описанные выше векторы по изобретению содержат селективный маркер и/или маркер для подсчета трансформантов.
Селективные маркерные гены, придающие устойчивость к антиметаболитам и подходящие для отбора трансформированных клеток и, например, тканей растений и растений, хорошо известны специалистам в данной области и включают, например, ген dhfr, обеспечивающий устойчивость к метотрексату (Reiss, Plant Physiol. (Life Sci. Adv.) 13 (1994), 143-149), npt, обеспечивающий устойчивость к аминогликозидам неомицину, канамицину и паромицину (Herrera-Estrella, ЕМВО J. 2 (1983), 987-995) и hygro, обеспечивающий устойчивость к гигромицину (Marsh, Gene 32 (1984), 481-485). Описаны дополнительные селективные гены, в частности, trpB, позволяющий клеткам использовать индол вместо триптофана; hisD, позволяющий клеткам использовать гистинол вместо гистидина (Hartman, Proc. Natl. Acad. Sci. USA 85 (1988), 8047); гены, кодирующие маннозо-6-фосфат-изомеразу, позволяющую клеткам использовать маннозу (WO 94/20627), и ODC (орнитин-декарбоксилазу), придающую резистентность к ингибитору орнитин-декарбоксилазы 2-(дифторметил)-DL-орнитину, DFMO (McConlogue, 1987, In: Current Communications in Molecular Biology, Cold Spring Harbor Laboratory ed.), или дезаминазу Aspergillus terreus, придающую резистентность к бластицидину S (Tamura, Biosci. Biotechnol. Biochem. 59 (1995), 2336-2338).
Специалистам в данной области также известны и доступны для приобретения подходящие маркеры для подсчета трансформантов. Предпочтительно, указанный маркер представляет собой ген, кодирующий люциферазу (Giacomin, PI. Sci. 116 (1996), 59-72; Scikantha, J. Bact. 178 (1996), 121), зеленый флуоресцентный белок (Gerdes, FEBS Lett. 389 (1996), 44-47) или β-глюкуронидазу (Jefferson, EMBO J. 6 (1987), 3901-3907). Данное воплощение особенно подходит для простого и быстрого скрининга клеток, тканей и организмов, содержащих упомянутый вектор.
Как описано выше, упомянутую молекулу нуклеиновой кислоты можно использовать по-отдельности или в составе вектора для экспрессии кодируемой биспецифической конструкции в клетках, например, для очистки, а также для целей генной терапии, предпочтительно в комбинации с трансдуцированными CD8+ Т-клетками. Молекулы нуклеиновой кислоты или векторы, содержащие последовательности ДНК, кодирующие любую из описанных выше биспецифических конструкций, вводят в клетки, которые в свою очередь продуцируют полипептид, представляющий интерес.Генная терапия, основанная на введении терапевтических генов в клетки при помощи ex-vivo или in-vivo методик, представляет собой одно из наиболее важных применений переноса генов. Подходящие векторы, способы или системы доставки генов для генной терапии in-vitro или in-vivo описаны в литературе и известны специалистам в данной области техники; см., например, Giordano, Nature Medicine 2 (1996), 534-539; Schaper, Cira Res. 79 (1996), 911-919; Anderson, Science 256 (1992), 808-813; Verma, Nature 389 (1994), 239; Isner, Lancet 348 (1996), 370-374; Muhlhauser, Cira Res. 77 (1995), 1077-1086; Onodera, Blood 91 (1998), 30-36; Verma, Gene Ther. 5 (1998), 692-699; Nabel, Ann. N.Y. Acad. Sci. 811 (1997), 289-292; Verzeletti, Hum. Gene Ther. 9 (1998), 2243-51; Wang, Nature Medicine 2 (1996), 714-716; WO 94/29469; WO 97/00957; US 5,580,859; US 5,589,466 или Schaper, Current Opinion in Biotechnology 7 (1996), 635-640. Упомянутые молекулы нуклеиновой кислоты и векторы могут быть созданы для направленного введения в клетку или введения в клетку при помощи липосом или вирусных векторов (например, аденовирусных, ретровирусных). В контексте данного изобретения указанная клетка представляет собой клетку зародышевой линии, эмбриональную клетку, яйцеклетку или полученную из них клетку, наиболее предпочтительно указанная клетка представляет собой стволовую клетку. Примером эмбриональной стволовой клетки может быть, в том числе, стволовая клетка, описанная Nagy, Proc. Natl. Acad. Sci. USA 90 (1993), 8424-8428.
В соответствии с вышесказанным, данное изобретение относится к способам получения векторов, в частности, плазмид, космид и бактериофагов, обычно используемых в генной инженерии, включающих молекулу нуклеиновой кислоты, кодирующую последовательность конструкции биспецифического антитела, описанную в данном документе. В контексте данного изобретения указанный вектор представляет собой экспрессирующий вектор и/или переносящий ген или направляющий вектор. Для доставки упомянутых полинуклеотидов или векторов в целевые клеточные популяции можно использовать экспрессирующие векторы, полученные на основе вирусов, например, ретровирусов, вирусов vaccinia, аденоассоциированных вирусов, вирусов герпеса или вируса папилломы крупного рогатого скота.
Способы, хорошо известные специалистам в данной области, можно использовать для конструирования рекомбинантных векторов; см., например, методики, описанные Sambrook et al. (loc cit.), Ausubel (1989, loc cit.) или другие стандартные руководства. В альтернативном случае, упомянутые молекулы нуклеиновой кислоты и векторы можно встраивать в липосомы для доставки в клетки-мишени. Векторы, содержащие молекулы нуклеиновой кислоты по изобретению, можно переносить в клетки-хозяева с помощью хорошо известных способов, варьирующихся в зависимости от типа клетки-хозяина. Например, трансфекцию с помощью хлорида кальция часто используют для прокариотических клеток, тогда как обработку фосфатом кальция или электропорацию можно использовать для других клеток-хозяев; см. Sambrook, выше. Указанным вектором может быть в том числе pEF-DHFR, pEF-ADA или pEF-neo. Векторы pEF-DHFR, pEF-ADA и pEF-neo описаны в области техники, например, Mack et al. Proc. Natl. Acad. Sci. USA 92 (1995), 7021-7025 и Raum et al. Cancer Immunol Immunother 50 (2001), 141-150.
В изобретении также предложены хозяева, трансформированные или трансфецированные вектором, согласно данному описанию. Такой хозяин может быть получен путем введения в хозяина по меньшей мере одного из описанных выше векторов или по меньшей мере одной из описанных выше молекул нуклеиновых кислот. Присутствие по меньшей мере одного указанного вектора или по меньшей мере одной молекулы нуклеиновой кислоты в хозяине может обеспечить экспрессию гена, кодирующего описанные выше молекулы или конструкции биспецифического антитела (т.е. описанной в данном документе молекулы биспецифического антитела).
Описанная молекула нуклеиновой кислоты или вектор, введенный в клетку-хозяина, может либо интегрироваться в геном хозяина, либо находиться внехромосомно.
Хозяин может быть прокариотической или эукариотической клеткой.
Термин "прокариотические" охватывает все бактерии, которые могут быть трансформированы, трансдуцированы или трансфецированы молекулами ДНК или РНК для экспрессии белка по изобретению. Прокариотические хозяева включают грам-отрицательные, а также грам-положительные бактерии, например, такие как Е. coli, S. typhimurium, Serratia marcescens и Bacillus subtilis. Термин "эукариотические" охватывает дрожжи, высшие растения и, предпочтительно, клетки млекопитающих. В зависимости от использованного на этапе рекомбинантной продукции хозяина, белок, кодируемый полинуклеотидом по данному изобретению, может быть гликозилированным или негликозилированным. Наиболее предпочтительным является использование плазмид или вирусов, содержащих кодирующую последовательность полипептида по изобретению с присоединенной к нему N-концевой последовательностью FLAG-tag и/или С-концевой последовательностью His-tag. Предпочтительно, длина указанного эпитопа FLAG-tag составляет приблизительно от 4 до 8 аминокислот, наиболее предпочтительно 8 аминокислот. Описанный выше полинуклеотид можно использовать для трансформации или трансфекции хозяина, с помощью любой методики, известной специалистам в данной области техники. Кроме того, способы получения слитых, функционально связанных генов и их экспрессии, например, в клетках млекопитающих и бактерий хорошо известны в области техники (Sambrook, loc cit.).
В контексте данного изобретения (клетка) хозяин представляет собой бактериальную клетку, клетку насекомого, гриба, растения или животного.
В частности, предусматривается, что упомянутый хозяин может представлять собой клетку млекопитающего, более предпочтительно, человеческую клетку или линию клеток человека.
Наиболее предпочтительные клетки-хозяева включают клетки HEK293, СНО, COS, линии миеломных клеток, такие как SP2/0 или NS/0. Как проиллюстрировано в приведенных Примерах, наиболее предпочтительными хозяевами являются клетки HEK293.
В следующем воплощении данное изобретение относится к способу продукции молекулы или конструкции биспецифического антитела (т.е. описанных здесь молекул биспецифического антитела), описанных выше, включающему культивирование (поддержание в культуре) клетки и/или клетки-хозяина по изобретению в условиях, обеспечивающих экспрессию молекулы или конструкции биспецифического антитела (т.е. описанных здесь молекул биспецифического антитела) и выделение молекулы или конструкции (т.е. описанных здесь молекул биспецифического антитела) из клетки и/или культуральной среды.
Трансформированных хозяев можно выращивать в ферментерах и культивировать в соответствии с методиками, известными в области техники, для достижения оптимального роста клеток. Полипептид по изобретению можно затем выделять из культуральной среды. Выделение и очистку, например, экспрессированных в микроорганизмах полипептидов по изобретению, можно осуществлять любыми стандартными способами, например, такими как препаративное хроматографическое разделение и иммунологическое разделение, например, с применением моноклональных или поликлональных антител, направленных, например, против эпитопа полипептида по изобретению, как описано в приведенных Примерах.
Кроме того, в изобретении предложена композиция (лекарственный препарат), содержащая молекулу биспецифического (моноклонального) антитела по изобретению, описанную в данном документе или молекулу (человеческого) биспецифического антитела, полученную способами, описанными выше, молекулу нуклеиновой кислоты по изобретению, вектор или хозяина по изобретению, трансдуцированные CD8+ Т-клетки. В контексте данного изобретения указанная композиция представляет собой фармацевтическую композицию, возможно дополнительно содержащую подходящие носители, стабилизаторы и/или эксципиенты.
Кроме того, в изобретении предложена молекула биспецифического антитела, описанная в данном документе, для применения в качестве лекарственного препарата, при этом молекулу антитела следует вводить перед, одновременно или после введения трансдуцированных CD8+ Т-клеток, содержащих антиген, который не существует в естественных условиях в или на CD8+ Т-клетках, при этом указанные CD8+ Т-клетки получают у субъекта, подлежащего лечению.
В контексте данного изобретения предложена фармацевтическая композиция/лекарственный препарат, содержащая молекулу биспецифического антитела, описанную выше, которую следует вводить в комбинации с трансдуцированными CD8+ Т-клетками, содержащими антиген, который не существует в естественных условиях в или на CD8+ Т-клетках, при этом указанную молекулу биспецифического антитела следует вводить перед, одновременно или после введения трансдуцированных CD8+ Т-клеток, содержащих антиген, который не существует в естественных условиях в или на CD8+ Т-клетках, и при этом указанные CD8+ Т-клетки получают у субъекта, подлежащего лечению.
В контексте данного изобретения Т-клетки трансдуцированы антигеном, не существующим в естественных условиях/эндогенно не экспрессируемым в и/или на Т-клетках, как описано выше. Изобретение также относится к CD8+ Т-клеткам, которые трансдуцированы антигеном, не существующим в естественных условиях/эндогенно не экспрессируемым в и/или на CD8+ Т-клетках, как описано выше. В контексте данного изобретения эти трансдуцированные Т-клетки (CD8+ Т-клетки) также содержат Т-клеточный рецептор (TCR). Эти трансдуцированные Т-клетки (CD8+ Т-клетки) являются опухолеспецифическими, либо поскольку эти Т-клетки были выделены из естественного аутологичного пула Т-клеток и способны лизировать опухолевые клетки, либо поскольку эти Т-клетки были ко-трансдуцированы опухолеспецифическим Т-клеточным рецептором (TCR). В контексте данного изобретения Т-клетки (CD8+ Т-клетки) также охватывают клетки, способные распознавать комплекс при помощи Т-клеточного рецептора, комплекс представляет собой конъюгат молекулы главного комплекса гистосовместимости (который здесь обозначен "ГКГС") (молекулы МНС; у человека он обозначается "человеческий лейкоцитарный антиген" (HLA)) и специфического для Т-клеточного рецептора пептидного антигена (который (структурно) отличается от антигена, не существующего в естественных условиях в CD8+ Т-клетках, как описано выше). Таким образом, в контексте данного изобретения, для развития цитотоксической реакции может быть необходимо (i) существование Т-клетки (CD8+ Т-клетки), имеющей Т-клеточный рецептор, специфический для HLA-типа клетки-мишени (обозначающей опухолевую клетку субъекта, подлежащего лечению) и (ii) антигенного пептида, чтобы комплекс, образующийся при связывании молекулы HLA, мог быть распознан TCR.
Термин "Т-клеточный рецептор" в данном документе обозначает любой Т-клеточный рецептор, при условии соблюдения трех критериев: (i) опухолевая специфичность, (ii) распознавание (большинства) опухолевых клеток, что означает, что антиген-мишень должны экспрессировать (большинство) опухолевые(х) клетки(ок) и (iii) что TCR совместим с HLA-типом субъекта, подлежащего лечению.
В этом контексте подходящие Т-клеточные рецепторы, отвечающие трем указанным выше критериям, известны в области техники, например, WT1 (специфический антиген опухоли Вильмса типа 1; последовательность описана Sugiyama H., Japanese Journal of Clinical Oncology 40 (2010), 377-87), MAGE (последовательность описана в WO 2007/032255 и PCT/US 2011/57272), SSX (предварительная заявка на патент США 61/388,983), NY-ESO-1 (последовательность описана в PCT/GB 2005/001924) и/или HER2neu (последовательность описана в WO 2011/0280894).
В контексте данного изобретения Т-клетки (CD8+ Т-клетки) выделяют/получают у субъекта. Способы выделения/получения Т-клеток (CD8+ Т-клеток) пациентов хорошо известны в области техники и включают лейкаферез, выделение/получение клеток с использованием прибора FACSort, отделение живых клеток от мертвых в свежих биопсийных образцах, содержащих живые клетки, вручную или с использованием микроманипулятора (Dudley et al., J. Immunother. 26 (2003), 332-342; Robbins et al., J. Clin. Oncol. 29 (2011), 917-924 и Leisegang, J. Mol. Med. 86 (2008), 573-58), но не ограничиваются ими. Термин "свежий биопсийный материал пациента" обозначает опухолевую ткань, удаленную у субъекта хирургическим или любым другим известным способом, а также линии опухолевых клеток или клетки (выделенные) из опухолевой ткани/опухолевые клетки. Выделенные/полученные Т-клетки впоследствии культивируют и осуществляют их экспансию с использованием антитела к CD3, с использованием моноклональных антител к CD3 и CD28 и/или с использоваием антитела к CD3, к CD28 и IL-2 (Dudley et al., J. Immunother. 26 (2003), 332-342; Dudley et al., J. Clin. Oncol. 26 (2008), 5233-5239).
В контексте данного изобретения эти выделенные/полученные Т-клетки являются CD8+ Т-клетками. Способы детекции существующих в естественных условиях/эндогенно экспрессируемых антигенов/маркеров на поверхности известны в области техники и включают проточную цитометрию (Koch et al., Immunity & Ageing 5 (2008), 6), полимеразную цепную реакцию (Fernandes S., Clinical and Diagnostic Laboratory Immunology 12 (2005), 477-483) и конфокальную микроскопию (Kenny E. et al., Immunology 101 (2000), 178-184), но не ограничиваются ими.
На следующем этапе Т-клетки искусственным образом/генетически модифицируют/трансдуцируют способами, известными в области техники (Francois M. Lemoine et al., J Gene Med 6 (2004), 374-386). Способы трансдукции Т-клеток известны в области техники и включают, в случае если осуществляют трансдукцию нуклеиновой кислотой или рекомбинантной нуклеиновой кислотой, например, электропорацию, кальций-фосфатный способ, способ с использованием катионных липидов или липосомный способ, но не ограничиваются ими. Трансдукцию нуклеиновой кислотой можно осуществить стандартным и эффективным способом с использованием доступных для приобретения реагентов для трансфекции, например, липофектамина (выпускается компанией Invitrogene). В случае, когда используется вектор, трансдукцию вектором можно осуществить способом, описанным выше для нуклеиновой кислоты, при условии, что вектор представляет собой плазмидный вектор. В контексте данного изобретения способы трансдукции Т-клеток включают ретровирусную или лентивирусную трансдукцию Т-клеток, а также трансфекцию мРНК.
В данном контексте, в области техники известны (ретровирусные) векторы, подходящие для трансдукции (человеческих) Т-клеток, например, SAMEN CMV/SRa (Clay et al., J. Immunol. 163 (1999), 507-513), LZRS-id3-IHRES (Heemskerk et al., J. Exp. Med. 186 (1997), 1597-1602), FeLV (Neil et al., Nature 308 (1984), 814-820), SAX (Kantoff et al., Proc. Natl. Acad. Sci. USA 83 (1986), 6563-6567), pDOL (Desiderio, J. Exp. Med. 167 (1988), 372-388), N2 (Kasid et al., Proc. Natl. Acad. Sci. USA 87 (1990), 473-477), LNL6 (Tiberghien et al., Blood 84 (1994), 1333-1341), pZipNEO (Chen et al., J. Immunol. 153 (1994), 3630-3638), LASN (Mullen et al., Hum. Gene Ther. 7 (1996), 1123-1129), pGIXsNa (Taylor et al., J. Exp. Med. 184 (1996), 2031-2036), LCNX и LXSN (Sun et al., Hum. Gene Ther. 8 (1997), 1041-1048), SFG (Gallardo et al., Blood 90 (1997), 952-957), HMB-Hb-Hu (Vieillard et al., Proc. Natl. Acad. Sci. USA 94 (1997), 11595-11600), pMV7 (Cochlovius et al., Cancer Immunol. Immunother. 46 (1998), 61-66), pSTITCH (Weitjens et al., Gene Ther 5 (1998), 1195-1203), pLZR (Yang et al., Hum. Gene Ther. 10 (1999), 123-132), pBAG (Wu et al., Hum. Gene Ther. 10 (1999), 977-982), rKat.43.267bn (Gilham et al., J. Immunother. 25 (2002), 139-151), pLGSN (Engels et al., Hum. Gene Ther. 14 (2003), 1155-1168), pMP71 (Engels et al., Hum. Gene Ther. 14 (2003), 1155-1168), pGCSAM (Morgan et al., J. Immunol. 171 (2003), 3287-3295), pMSGV (Zhao et al., J. Immunol. 174 (2005), 4415-4423), pMX (de Witte et al., J. Immunol. 181 (2008), 5128-5136).
Согласно данному изобретению, термин "лекарственный препарат" используется взаимозаменяемо с термином "фармацевтическая композиция" и относится к композиции для введения пациенту, предпочтительно пациенту-человеку. В контексте данного изобретения данный лекарственный препарат/фармацевтическую композицию следует вводить пациенту, у которого были выделены/получены CD8+ Т-клетки. В контексте данного изобретения пациент обозначает пациента-человека. Кроме того, в контексте данного изобретения этот пациент страдает заболеванием, при этом указанное заболевание представляет собой онкологическое заболевание, в частности, злокачественное новообразование эпителиального, эндотелиального или мезотелиального происхождения или злокачественное новообразование кроветворной ткани. В контексте данного изобретения злокачественное новообразование выбрано из группы, состоящей из рака ЖКТ, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи, рака полости рта, рака желудка, рака шейки матки, В и Т-клеточной лимфомы, миелолейкоза, рака яичников, лейкоза, лимфолейкоза, носоглоточной карциномы, рака толстой кишки, рака предстательной железы, почечноклеточного рака, раковых образований головы и шеи, рака кожи (меланомы), рака мочеполового тракта, например, рака яичка, эндотелиомы, рака шейки матки и рака почки, рака желчевыводящих путей, рака пищевода, рака слюнных желез и рака щитовидной железы или других злокачественных заболеваний, таких как гемобластозы, глиомы, саркомы или остеосаркомы.
В предпочтительном воплощении фармацевтическая композиция/лекарственный препарат содержит молекулу биспецифического антитела, описанную в данном документе, для парентерального, трансдермального, внутрипросветного, внутриартериального, внутриоболочечного введения или инъекции непосредственно в ткань или опухоль. В контексте данного изобретения композиция/лекарственный препарат содержит молекулу биспецифического антитела, описанную в данном документе, которую следует вводить перед, одновременно или после введения трансдуцированных CD8+ Т-клеток, содержащих антиген, который эндогенно не экспрессируется/не существует в естественных условиях в и/или на поверхности Т-клеток. В контексте данного изобретения фармацевтическая композиция/лекарственный препарат, содержащие молекулу биспецифического антитела, описанную в данном документе, следует вводить в комбинации с трансдуцированными CD8+ Т-клетками, содержащими антиген, который в естественных условиях не существует в или на CD8+ Т-клетках, при этом указанные CD8+ Т-клетки получают у субъекта, подлежащего лечению.
Использование термина "в комбинации" не ограничивает порядок, в котором субъекту следует вводить составляющие терапевтического режима. Таким образом, описанные здесь фармацевтическая композиция/лекарственный препарат, охватывают введение молекулы биспецифического антитела, описанной ранее в данном документе, одновременно с или после введения трансдуцированных CD8+ Т-клеток, содержащих антиген, который в естественных условиях не существует/эндогенно не экспрессируется в или на CD8+ Т-клетках. "В комбинации" в данном описании также не ограничивает время между введением молекулы биспецифического антитела, описанной ранее в данном документе, и трансдуцированных CD8+ Т-клеток, содержащих антиген, который в естественных условиях не существует/эндогенно не экспрессируется в или на CD8+ Т-клетках. Таким образом, когда два компонента не вводят одновременно/параллельно, введение может разделять 1 минута, 5 минут, 15 минут, 30 минут, 45 минут, 1 час, 2 часа, 4 часа, 6 часов, 12 часов, 24 часа, 48 часов или 72 часа или любой подходящий временной интервал, легко определяемый специалистом в данной области и/или описанный в данном документе.
В контексте данного изобретения термин "в комбинации" также охватывает ситуации, когда молекулу биспецифического антитела, описанную в данном документе, и трансдуцированные CD8+ Т-клетки, содержащие антиген, который в естественных условиях не существует/эндогенно не экспрессируется в и/или на CD8+ Т-клетках, предварительно инкубируют вместе перед введением субъекту. Так, два компонента можно преинкубировать перед введением, например, в течение 1 минуты, 5 минут, 10 минут, 15 минут, 30 минут, 45 минут или 1 часа или в течение любого подходящего интервала времени, легко определяемого специалистом в данной области. В другом воплощении изобретение относится к терапевтическому режиму, в котором молекулу биспецифического антитела, описанную в данном документе, и трансдуцированные CD8+ Т-клетки, содержащие антиген/маркер, который в естественных условиях не существует/эндогенно не экспрессируется в и/или на CD8+ Т-клетках, следует вводить одновременно/параллельно. В контексте данного изобретения молекулу биспецифического антитела, описанную в данном документе, можно вводить после введения трансдуцированных CD8+ Т-клеток, содержащих антиген, который естественным образом не существует/эндогенно не экспрессируется в и/или на CD8+ Т-клетках.
Кроме того, термин "в комбинации", используемый в данном документе, не ограничивает заявленные терапевтические режимы введением молекулы биспецифического антитела, описанной в данном документе, и трансдуцированных CD8+ Т-клеток, содержащих антиген, который в естественных условиях не существует/эндогенно не экспрессируется в или на CD8+ Т-клетках, непосредственно друг за другом (т.е. введением одного из двух компонентов, за которым (через некоторый промежуток времени) следует введение другого, без введения и/или выполнения какого-либо иного протокола лечения между ними. Таким образом, данные терапевтические режимы также охватывают раздельное введение молекулы биспецифического антитела, описанной в данном документе, и трансдуцированных CD8+ Т-клеток, содержащих антиген, который в естественных условиях не существует/эндогенно не экспрессируется в или на CD8+ Т-клетках, при этом введения разделены одним или несколькими протоколами лечения, необходимыми и/или подходящими для лечения или предупреждения заболевания или его симптома. Примеры таких промежуточных протоколов лечения включают введение обезболевающих препаратов, введение химиотерапевтических препаратов, хирургическое лечение заболевания или его симптома, но не ограничиваются ими. Таким образом, описанные в данном документе терапевтические режимы охватывают введение молекулы биспецифического антитела, описанной в данном документе, и трансдуцированных CD8+ Т-клеток, содержащих антиген, который в естественных условиях не существует/эндогенно не экспрессируется в или на CD8+ Т-клетках, в сочетании ни с одним, с одним или несколькими протоколами лечения, подходящими для лечения или предупреждения заболевания или его симптома, как описано в данном документе или известно из области техники.
В частности, предусматривается, что указанную фармацевтическую композицию/лекарственный препарат следует вводить пациенту посредством инфузии или инъекции. В контексте данного изобретения трансдуцированные CD8+ Т-клетки, содержащие антиген, который в естественных условиях не существует в или на CD8+ Т-клетках, следует вводить посредством инфузии или инъекции. Введение подходящих композиций/лекарственных препаратов можно осуществлять различными способами, путем внутривенного, внутрибрюшинного, подкожного, внутримышечного, местного или внутрикожного введения.
Фармацевтическая композиция/лекарственный препарат по настоящему изобретению может также содержать фармацевтически приемлемый носитель. Примеры подходящих фармацевтических носителей хорошо известны в области техники и включают растворы фосфатного солевого буфера, воду, эмульсии, такие как эмульсии типа масло/вода, различные типы смачивающих агентов, стерильные растворы и т.д. Композиции, содержащие такие носители, могут быть приготовлены при помощи хорошо известных стандартных способов. Эти фармацевтические композиции можно вводить субъекту в подходящей дозе. Режим дозирования определяют лечащие врачи в зависимости от клинических факторов. Как известно в медицине, дозировки для пациентов зависят от множества факторов, включая вес пациента, площадь поверхности тела, возраст, конкретное соединение, которое необходимо ввести, пол, время и путь введения, общее состояние здоровья, а также необходимость одновременного введения других препаратов. Как правило, режим введения может представлять собой регулярное введение фармацевтической композиции в диапазоне от 1 мкг до 5 г единиц в сутки. Однако, более предпочтительная дозировка при непрерывном введении может находиться в диапазоне от 0,01 мкг до 2 мг, предпочтительно от 0,01 мкг до 1 мг, более предпочтительно от 0,01 мкг до 100 мкг, еще более предпочтительно от 0,01 мкг до 50 мкг и наиболее предпочтительно от 0,01 мкг до 10 мкг единиц на килограмм веса тела в час. Наиболее предпочтительные дозировки указаны в данном документе ниже. Динамику можно отслеживать путем периодической оценки. Дозировка может варьироваться, однако предпочтительная дозировка для внутривенного введения ДНК составляет приблизительно от 106 до 1012 копий молекул ДНК. Композиции по изобретению можно вводить местно или системно. Введение как правило бывает парентеральным, например, внутривенным; ДНК можно также вводить непосредственно в целевой участок, например, посредством биолистической доставки во внутренний или поверхностный участок-мишень или через катетер в область артерии. Препараты для парентерального введения включают стерильные водные или неводные растворы, суспензии и эмульсии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъекционные органические эфиры, такие как этилолеат. Водные носители включают воду, спиртовые/водные растворы, эмульсии или суспензии, включая солевые и забуференные среды. Растворители для парентерального введения включают раствор натрия хлорида, раствор декстрозы в растворе Рингера, раствор декстрозы в растворе натрия хлорида, раствор Рингер-лактат или нелетучие масла. Растворители для внутривенного введения включают растворы для восполнения дефицита жидкости и питательных веществ, растворы для восполнения дефицита электролитов (например, на основе раствора декстрозы в растворе Рингера) и т.п. Также могут присутствовать консерванты или другие добавки, например, такие как противомикробные агенты, антиоксиданты, хелатирующие агенты и инертные газы и т.п. Кроме того, фармацевтическая композиция по данному изобретению может содержать белковые носители, например, сывороточный альбумин или иммуноглобулин, предпочтительно человеческого происхождения. Предусматривается, что кроме конструкций белкового биспецифического антитела или молекул нуклеиновой кислоты или кодирующих их векторов (описанных в данном изобретении), фармацевтическая композиция по изобретению может содержать другие биологически активные агенты, в зависимости от предполагаемого применения фармацевтической композиции. Такие агенты могут представлять собой лекарства, действующие на желудочно-кишечный тракт, лекарства, действующие как цитостатики, лекарства, предупреждающие гиперурикемию, лекарства, ингибирующие иммунные реакции (например, кортикостероиды), лекарства, действующие на систему кровообращения и/или такие агенты, как костимулирующие молекулы Т-клеток или цитокины, известные в области техники.
Возможными показаниями к применению композиции(й)/лекарственного(ых) препарата(ов) по изобретению являются онкологические заболевания, в частности злокачественные новообразования эпителиального происхождения, такие как рак молочной железы, рак толстой кишки, рак предстательной железы, злокачественные опухоли головы и шеи, рак кожи (меланома), рак мочеполового тракта, например, рак яичника, рак яичка, эндотелиома, рак шейки матки и рака почки, рак легких, рак желудка, рак желчевыводящих путей, рак пищевода, рак слюнных желез и рак щитовидной железы или другие злокачественные заболевания, такие как гемобластозы, глиомы, саркомы или остеосаркомы.
Изобретение также предусматривает совместное введение с другими соединениями, например, молекулами, способными обеспечить активирующий сигнал эффекторным клеткам иммунной системы, клеточную пролиферацию или стимуляцию клеток. Указанные молекулы могут, например, обеспечивать дополнительный первичный активирующий сигнал для Т-клеток (например, представлять собой дополнительные костимулирующие молекулы: молекулы семейства В7, Ox40L, 4.1 BBL, CD40L, анти-CTLA-4, анти-PD-1) или представлять собой интерлейкин из суперсемейства цитокинов (например, IL-2).
Композиция по изобретению, описанная выше, может также представлять собой диагностическую композицию, возможно также включать средства и способы детекции.
Биспецифические связывающие молекулы или конструкции (т.е. описанные здесь молекулы биспецифического антитела), предложенные в данном документе, также подходят для применения в иммунологических анализах, где их можно использовать в жидкой фазе или связанными с твердофазными носителями. Примерами иммунологических анализов, в которых можно использовать полипептид по изобретению, являются прямые или непрямые конкурентные или неконкурентные иммунологические анализы. Примерами таких иммунологических анализов являются твердофазный иммуноферментный анализ (твердофазный ИФА), иммуноферментный анализ (ИФА), радиоиммунный анализ (РИА), иммунометрический анализ сэндвич-типа и вестерн-блоттинг.
Биспецифические связывающие молекулы или конструкции (т.е. описанные здесь молекулы биспецифического антитела) по изобретению могут быть связаны с множеством различных носителей и использоваться для выделения клеток, специфически связанных с указанными полипептидами. Примеры известных носителей включают стекло, полистирол, поливинил-хлорид, полипропилен, полиэтилен, поликарбонат, декстран, нейлон, амилозу, естественную и модифицированную целлюлозу, полиакриламид, агарозу и магнетит.Для целей изобретения носитель может быть растворимым или нерастворимым, например, представлять собой гранулы.
Существует множество меток и способов введения метки, известных специалистам в области техники. Типы меток, которые можно использовать в данном изобретении, включают ферменты, радиоизотопы, коллоидные металлы, флуоресцентные соединения, хемилюминесцентные соединения и биолюминесцентные соединения.
В наиболее предпочтительном воплощении данного изобретения предусмотрено применение конструкций/молекул биспецифического антитела в качестве лекарственных препаратов. В контексте данного изобретения описано применение молекул биспецифического антитела в качестве лекарственного препарата, при котором указанные молекулы биспецифического антитела следует вводить перед, одновременно или после введения трансдуцированных CD8+ Т-клеток, содержащих антиген, который в естественных условиях не существует на CD8+ Т-клетках, и при котором указанные CD8+ Т-клетки, получают у субъекта, подлежащего лечению. Указанный лекарственный препарат можно использовать в способе лечения онкологического заболевания, в частности, злокачественного новообразования эпителиального, эндотелиального или мезотелиального происхождения или злокачественного новообразования кроветворной ткани. В контексте данного изобретения злокачественное новообразование выбрано из группы, состоящей из рака ЖКТ, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи, рака полости рта, рака желудка, рака шейки матки, В и Т-клеточной лимфомы, миелолейкоза, рака яичников, лейкоза, лимфолейкоза, носоглоточной карциномы, рака толстой кишки, рака предстательной железы, почечноклеточного рака, злокачественных опухолей головы и шеи, рака кожи (меланомы), рака мочеполового тракта, например, рака яичка, рака яичника, эндотелиомы, рака шейки матки и рака почки, рака желчевыводящих путей, рака пищевода, рака слюнных желез и рака щитовидной железы или других злокачественных заболеваний, таких как гемобластозы, глиомы, саркомы или остеосаркомы.
Кроме того, в контексте данного изобретения предусматривается применение описанной здесь молекулы биспецифического антитела, содержащей (i) первый связывающий домен, связывающийся с антигеном на CD8+ Т-клетках, не существующим в естественных условиях в или на CD8+ Т-клетках; и (ii) второй связывающий домен, связывающийся с опухолеспецифическим антигеном, присутствующим в естественных условиях на поверхности опухолевых клеток, в способе лечения онкологического заболевания, где указанную молекулу биспецифического антитела следует вводить перед, одновременно или после введения трансдуцированных CD8+ Т-клеток, содержащих антиген, который в естественных условиях не присутствует в или на CD8+ Т-клетках, и где указанные CD8+ Т-клетки были получены у субъекта, подлежащего лечению.
Кроме того, в контексте данного изобретения способ лечения онкологического заболевания включает введение молекулы биспецифического антитела по данному изобретению нуждающемуся в этом субъекту, которая включает (i) первый связывающий домен, связывающийся с антигеном на CD8+ Т-клетках, не существующим в естественных условиях в или на CD8+ Т-клетках; и (ii) второй связывающийся домен, связывающийся с опухолеспецифическим антигеном, существующим в естественных условиях на поверхности опухолевых клеток; при этом указанную молекулу биспецифического антитела следует вводить перед, одновременно или после введения трансдуцированных CD8+ Т-клеток указанного субъекта, содержащих антиген, который в естественных условиях не существует в или на CD8+ Т-клетках. В контексте данного изобретения злокачественное новообразование выбрано из группы, состоящей из рака ЖКТ, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи, рака полости рта, рака желудка, рака шейки матки, В и Т-клеточной лимфомы, миелолейкоза, рака яичников, лейкоза, лимфолейкоза, носоглоточной карциномы, рака толстой кишки, рака предстательной железы, почечноклеточного рака, раковых образований головы и шеи, рака кожи (меланомы), рака мочеполового тракта, например, рака яичка, рака яичника, эндотелиомы, рака шейки матки и рака почки, рака желчевыводящих путей, рака пищевода, рака слюнных желез и рака щитовидной железы или других злокачественных заболеваний, таких как гемобластозы, глиомы, саркомы или остеосаркомы.
Кроме того, согласно изобретению, молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую ЕрСАМ (человека) (как опухоле-специфический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не существующий в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно использовать в способе лечения рака ЖКТ, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи и/или рака полости рта. В контексте данного изобретения молекулу биспецифического антитела, направленного против ЕрСАМ (человека) (вторым связывающим доменом) и содержащую первый связывающий домен, направленный против/связывающийся с/взаимодействующий с белком Cripto, можно применять в лечении рака ЖКТ, например, аденокарциномы желудочно-кишечного тракта. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) HER1 (как опухолеспецифический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения рака ЖКТ, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи и/или рака полости рта. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) HER2 (как опухолеспецифический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения рака желудка, рака молочной железы и/или рака шейки матки. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) HER3 (как опухолеспецифический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения рака желудка и/или рака легких. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) CD20 как опухолеспецифический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток) и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения В-клеточной лимфомы и/или Т-клеточной лимфомы. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) CD22 (как опухолеспецифический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения В-клеточной лимфомы и/или Т-клеточной лимфомы. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) CD33 (как опухолеспецифический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения миелолейкоза. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) СА12-5 (как опухолеспецифический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения рака яичников, рака легких, рака молочной железы и/или рака ЖКТ. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) HLA-DR (как опухолеспецифический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения рака ЖКТ, лейкоза и/или носоглоточной карциномы. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) MUC-1 (как опухолеспецифический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения рака толстой кишки, рака молочной железы, рака яичников, рака легких и/или рака поджелудочной железы. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) А33 (как опухолеспецифический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения рака толстой кишки. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) PSMA (как опухолеспецифический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения рака предстательной железы. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) рецептор трансферрина (как опухоле-специфический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения рака ЖКТ, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи и/или рака полости рта. Молекулу или конструкцию (т.е. описанную здесь молекулу биспецифического антитела), содержащую (человеческий) CA-IX (как опухолеспецифический антиген, присутствующий в естественных условиях на поверхности опухолевых клеток), и один из описанных здесь антигенов, не встречающихся в естественных условиях в и/или на Т-клетках (CD8+ Т-клетках), можно применять в способе лечения рака почки.
Изобретение также относится к способу лечения онкологического заболевания, такого как злокачественное новообразование эпителиального, эндотелиального или мезотелиального происхождения и/или злокачественное новообразование кроветворной ткани. Такие заболевания будут среди следующих: рак пищевода, желудка, толстой кишки, тонкой кишки, печени, поджелудочной железы, молочной железы, легких, мозга, почки, яичка, рак кожи, лейкозы и/или лимфомы, предусматривающие введение субъекту трансдуцированных CD8+ Т-клеток.
В контексте данного изобретения, указанный субъект является человеком. В контексте данного изобретения способ лечения заболевания отличается тем, что включает этапы
(a) выделения CD8+ Т-клеток у субъекта;
(b) трансдукции указанных выделенных CD8+ Т-клеток антигеном, который в естественных условиях не существует в CD8+ Т-клетках, как описано выше в данном документе; и
(c) введения трансдуцированных CD8+ Т-клеток указанному субъекту.
В контексте данного изобретения указанные трансдуцированные CD8+ Т-клетки вводят указанному субъекту путем внутривенной инфузии.
Кроме того, в данном изобретении предложен способ лечения заболевания, включающий этапы
(a) выделения CD8+ Т-клеток у субъекта;
(b) трансдукции указанных выделенных CD8+ Т-клеток антигеном, который в естественных условиях не присутствует в CD8+ Т-клетках, как описано выше в данном документе;
(c) ко-трансдукции указанных выделенных CD8+ Т-клеток Т-клеточным рецептором;
(d) экспансии CD8+ Т-клеток с использованием анти-CD3 и анти-CD28 антител; и
(e) введения трансдуцированных CD8+ Т-клеток указанному субъекту.
Данное изобретение относится к выделенным CD8+ Т-клеткам, анализ которых осуществляется способами, позволяющими установить, что (опухолевый) антиген, существующий в естественных условиях на выделенных CD8+ Т-клетках, идентичен опухолевому антигену, с которым описанное здесь биспецифическое антитело связывается своим вторым связывающим доменом. В контексте данного изобретения выделенные/полученные CD8+ Т-клетки (содержащие антиген, присутствующий в естественных условиях на поверхности выделенных CD8+ Т-клеток) искусственным образом модифицированы введением антигена/маркера, не встречающегося в естественных условиях/эндогенно не экспрессируемого в и/или на CD8+ Т-клетках. В контексте данного изобретения искусственная модификация выделенных/полученных CD8+ Т-клеток относится к описанным в данном документе способам трансдукции. Таким образом, в контексте данного изобретения субъект, подлежащий лечению, относится к субъекту, отличающемуся наличием заболевания, отличающегося наличием опухолеспецифического антигена, присутствующего в естественных условиях на поверхности опухолевой клетки, согласно описанию выше в данном документе. В контексте данного изобретения введение трансдуцированных CD8+ Т-клеток, полученых/выделенных у субъекта, подлежащего лечению, осуществляют путем внутривенной инфузии.
В следующем воплощении данное изобретение относится к способу лечения заболевания, включающему этапы
(a) выделения инфильтрирующих опухоль лимфоцитов (ИОЛ) из удаленной опухоли пациента;
(b) культивирования и трансдукции ИОЛ антигеном, который в естественных условиях не присутствует в CD8+ Т-клетках, согласно описанию выше в данном документе;
(c) селекции культур ИОЛ по способности распознавать опухолевые клетки;
(d) экспансии ИОЛ с использованием анти-CD3 и/или анти-CD28 антител; и
(e) введения трансдуцированных CD8+ Т-клеток указанному субъекту.
Термин функциональное исследование "способности распознавать опухолевые клетки" означает совместное культивирование ИОЛ либо с аутологичными, например, принадлежащими пациенту, опухолевыми клетками, либо с клеточной линией идентичного HLA-типа. Определяют цитотоксическую активность в отношении опухолевых клеток (ЛДГ, высвобождение кальцеина). Кроме того, можно определять секрецию цитокинов, исследовать внутриклеточные цитокины Т-клеток методом проточной цитометрии, проводить анализ ELISPOT.
Упомянутый выше этап (d) (относящийся к этапу экспансии ИОЛ с использованием анти-CD3 и/или анти-CD28 антител) можно также осуществлять в присутствии (активирующих) цитокинов, таких как интерлейкин-2 и/или интерлейкин-15 (IL-15). В контексте данного изобретения упомянутый выше этап (d) (относящийся к этапу экспансии ИОЛ с использованием анти-CD3 и/или анти-CD28 антител) можно также осуществлять в присутствии интерлейкина-12 (IL-12), интерлейкина-7 (IL-7) и/или интерлейкина-21 (IL-21).
Способ лечения может дополнительно включать введение биспецифического (моноклонального) антитела по данному изобретению. Указанное (моноклональное) биспецифическое антитело можно вводить перед, одновременно или после введения трансдуцированных CD8+ Т-клеток. В контексте данного изобретения введение трансдуцированных CD8+ Т-клеток осуществляют путем внутривенной инфузии. В контексте данного изобретения трансдуцированные CD8+ Т-клетки выделяют/получают у субъекта, подлежащего лечению.
В изобретении предложен набор, содержащий биспецифическое (моноклональное) антитело, молекулу нуклеиновой кислоты, вектор или хозяина согласно изобретению. Указанный набор в частности может найти применение в приготовлении фармацевтической композиции по данному изобретению и помимо прочего может содержать контейнер, пригодный для инъекций или инфузий. Предпочтительно, набор по данному изобретению также содержит, как вариант, (а) буфер(ы), исходные растворы и/или другие реагенты или материалы, необходимые для медицинских или научных целей. Данное изобретение относится к набору, содержащему (А) молекулу биспецифического антитела, содержащую (i) первый связывающий домен, связывающийся с антигеном на CD8+ Т-клетках, не встречающимся в естественных условиях в или на CD8+ Т-клетках; и второй связывающий домен, связывающийся с опухолеспецифиеским антигеном, присутствующим в естественных условиях на поверхности опухолевых клеток; и (В) материал, необходимый для трансдукции CD8+ Т-клеток, выделенных/полученных у субъекта, подлежащего лечению, антигеном, не встречающимся в естественных условиях в или на указанных CD8+ Т-клетках.
В контексте данного изобретения "материал, необходимый для трансдукции CD8+ Т-клеток" выделенных/полученных у субъекта, подлежащего лечению, по меньшей мере представляет собой нуклеиновую кислоту, кодирующую антиген, не существующий в естественных условиях/эндогенно не экспрессируемый в и/или на CD8+ Т-клетках. Нуклеиновая кислота, кодирующая антиген, не существующий в естественных условиях/эндогенно не экспрессируемый в и/или на CD8+ Т-клетках, может быть функционально связана с (а) регуляторной(ыми) последовательностью(ями), которая(ые) обычно находятся в составе вектора (например, плазмиды или вирусной ДНК), включая последовательности, необходимые для селекции in vitro и амплификации вектора в бактериях. Вектор, обеспечивающий экспрессию антигена, не встречающегося в естественных условиях/эндогенно не экспрессируемого в CD8+ Т-клетках, в данном документе обозначается "экспрессирующий вектор". Таким образом, другой подходящий "материал, необходимый для трансдукции CD8+ Т-клеток" выделенных/полученных у субъекта, подлежащего лечению, может представлять собой вектор/экспрессирующий вектор, содержащий по меньшей мере нуклеиновую кислоту, кодирующую антиген, не встречающийся в естественных условиях в и/или на CD8+ Т-клетках. В этом контексте подходящие векторы для трансдукции Т-клеток/CD8+ Т-клеток охватывают вектор, выбранный из группы, состоящей из SAMEN CMV/SRa (Clay et al., J. Immunol. 163 (1999), 507-513), LZRS-id3-IHRES (Heemskerk et al., J. Exp. Med. 186 (1997), 1597-1602), FeLV (Neil et al., Nature 308 (1984), 814-820), SAX (Kantoff et al., Proc. Natl. Acad. Sci. USA 83 (1986), 6563-6567), pDOL (Desiderio, J. Exp. Med. 167 (1988), 372-388), N2 (Kasid et al., Proc. Natl. Acad. Sci. USA 87 (1990), 473-477), LNL6 (Tiberghien et al., Blood 84 (1994), 1333-1341), pZipNEO (Chen et al., J. Immunol. 153 (1994), 3630-3638), LASN (Mullen et al., Hum. Gene Ther. 7 (1996), 1123-1129), pGIXsNa (Taylor et al., J. Exp. Med. 184 (1996), 2031-2036), LCNX и LXSN (Sun et al., Hum. Gene Ther. 8 (1997), 1041-1048), SFG (Gallardo et al., Blood 90 (1997), 952-957), HMB-Hb-Hu (Vieillard et al., Proc. Natl. Acad. Sci. USA 94 (1997), 11595-11600), pMV7 (Cochlovius et al., Cancer Immunol. Immunother. 46 (1998), 61-66), pSTITCH (Weitjens et al., Gene Ther 5 (1998), 1195-1203), pLZR (Yang et al., Hum. Gene Ther. 10 (1999), 123-132), pBAG (Wu et al., Hum. Gene Ther. 10 (1999), 977-982), rKat.43.267bn (Gilham et al., J. Immunother. 25 (2002), 139-151), pLGSN (Engels et al., Hum. Gene Ther. 14 (2003), 1155-1168), pMP71 (Engels et al., Hum. Gene Ther. 14 (2003), 1155-1168), pGCSAM (Morgan et al., J. Immunol. 171 (2003), 3287-3295), pMSGV (Zhao et al., J. Immunol. 174 (2005), 4415-4423) и pMX (de Witte et al., J. Immunol. 181 (2008), 5128-5136). "Материал, необходимый для трансдукции CD8+ Т-клеток", может также охватывать клетки-хозяева, трансформированные или трансфецированне вектором, как средство для экспрессии антигена, не встречающегося в естественных условиях/эндогенно не экспрессируемого в и/или на CD8+ Т-клетках. Указанные клетки-хозяева могут быть получены путем введения в клетку-хозяина по меньшей мере одного описанного выше вектора или по меньшей мере одной молекулы нуклеиновой кислоты, кодирующей антиген, не встречающийся в естественных условиях/эндогенно не экспрессируемый в и/или на CD8+ Т-клетках. Присутствие по меньшей мере одного указанного вектора или по меньшей мере одной нуклеиновой кислоты в клетке-хозяине может обеспечивать экспрессию гена, кодирующего описанный здесь антиген, не встречающийся в естественных условиях/эндогенно не экспрессируемый в и/или на CD8+ Т-клетках.
Материал, необходимый для трансдукции CD8+ Т-клеток может также охватывать подробную информацию и/или оборудование, необходимое для генетической/искусственной модификации/трансдукции этих выделенных/полученных CD8+ Т-клеток. Если для трансдукции CD8+ Т-клеток используют нуклеиновую кислоту или рекомбинантную нуклеиновую кислоту, кодирующую антиген, не встречающийся в естественных условиях/эндогенно не экспрессируемый в и/или на CD8+ Т-клетках, в наборе может быть предложена информация и/или оборудование, необходимые для электропорации, кальций-фосфатного метода, метода с использованием катионных липидов, липосомного метода, а также реагент(ы) для трансфекции, буфер(ы) и/или другие реагенты, необходимые для трансдукции.
Материал, необходимый для трансдукции CD8+ Т-клеток, может также охватывать информацию и/или оборудование, необходимое для культивирования и экспансии выделенных/полученных CD8+ Т-клеток, например, антитело к CD3, моноклональное антитело к CD3 и CD28 и/или антитело к CD3, антитело к CD28 и IL-2.
Таким образом, в заключение, "материал, необходимый для трансдукции CD8+ Т-клеток", выделенных/полученных у субъекта, подлежащего лечению, представляет собой по меньшей мере нуклеиновую кислоту, кодирующую антиген, не встречающийся в естественных условиях в и/или на CD8+ Т-клетках, вектор/экспрессирующий вектор, содержащий по меньшей мере нуклеиновую кислоту, кодирующую антиген, не встречающийся в естественных условиях в и/или на CD8+ Т-клетках, реагент(ы) для трансфекции, буфер(ы) и/или материал, необходимый для трансдукции и/или культивирования CD8+ Т-клеток, выделенных/полученных у субъекта, подлежащего лечению с использованием антигена, не встречающегося в естественных условиях в и/или на указанных CD8+ Т-клетках.
Кроме того, компоненты набора по изобретению могут быть упакованы в отдельные флаконы или бутыли или находиться вместе в контейнерах или мультиконтейнерах. Кроме того, набор по данному изобретению содержит (закрытый) мешок для инкубации клеток, в котором клетки пациента, предпочтительно Т-клетки, описанные в данном документе выше, можно трансдуцировать и инкубировать в условиях, соответствующих стандарту GMP (надлежащей производственной практики, описанному в руководстве по надлежащей производственной практике, опубликованном комиссией Евросоюза на сайте http://ec.europa.eu/health/documents/eudralex/index_en.htm). Кроме того, набор по данному изобретению содержит (закрытый) мешок для инкубации клеток, в котором выделенные/полученные у пациента CD8+ Т-клетки можно трансдуцировать и инкубировать в условиях, соответствующих стандарту GMP. Кроме того, в контексте данного изобретения набор может также содержать молекулу нуклеиновой кислоты, кодирующую антиген, не встречающийся в естественных условиях в CD8+ Т-клетках, описанный здесь выше, и/или молекулу нуклеиновой кислоты, кодирующую Т-клеточный рецептор, описанный здесь выше. Набор по данному изобретению может предпочтительно применяться в том числе для осуществления способа по изобретению и может использоваться в различных приложениях, упомянутых в данном документе, например, в качестве средства для исследований или медицинского средства. Изготовление наборов предпочтительно осуществляется в соответствии со стандартными процедурами, известными специалисту в данной области техники.
Эти и другие воплощения изложены и содержатся в описании данного изобретения и приведенных Примерах. Дополнительная литература, касающаяся любого из антител, способов, применений и соединений, используемых в соответствии с данным изобретением, может быть найдена в публичных библиотеках и базах данных, например, с помощью электронных устройств. Например, можно использовать открытую базу данных "Medline", доступ к которой можно получить по ссылке http://www.ncbi.nlm.nih.gov/PubMed/medline.html. Специалистам в данной области известны другие базы данных и адреса, например, такие как http://www.ncbi.nlm.nih.gov, http://www.infobiogen.fr/, http://www.fmi.ch/biology/research_tools.html, http://www.tigr.org/, и доступ к ним может быть получен, например, по ссылке http://www.lycos.com.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1: Репрезентативное биспецифическое антитело, специфичное к EGFR человека и ЕрСАМ мыши (MAb225_scFv_G8.8)
Mab225 с каркасом мышиного IgG2a гибридизовано с одноцепочечным (se) Fv фрагментом С-концами тяжелых цепей.
Фиг. 2: Репрезентативные изображения, полученные при исследовании антитела Mab225, специфического к EGFR человека, методом эксклюзионной хроматографии и ДСН-ПАГ
Антитело, очищенное с использованием белка А, подвергали эксклюзионной хроматографии. (А) Профиль элюирования с колонки HiLoad Superdex 200. Пиковые фракции объединяли и оценивали чистоту белка при помощи ДСН-ПАГ. (В) Исследование биспецифического антитела при помощи ДСН-ПАГ в невосстанавливающих (NR) и восстанавливающих (R) условиях.
Фиг. 3: Репрезентативные изображения, полученные при исследовании антитела G8.8, специфического к ЕрСАМ мыши, методом эксклюзионной хроматографии и ДСН-ПАГ
Антитело, очищенное с использованием белка А, подвергали эксклюзионной хроматографии. (А) Профиль элюирования с колонки HiLoad Superdex 200. Пиковые фракции объединяли и оценивали чистоту белка при помощи ДСН-ПАГ. (В) Исследование биспецифического антитела при помощи ДСН-ПАГ в невосстанавливающих (NR) и восстанавливающих (R) условиях.
Фиг. 4: Репрезентативные изображения, полученные при исследовании биспецифического антитела MAb225_scFv_G8.8 методом эксклюзионной хроматографии и ДСН-ПАГ
(А) Профиль элюирования с колонки HiLoad Superdex 200. Номерами обозначены фракция биспецифического антитела (1) и фракция агрегатов (2). Пиковые фракции (1) объединяли и оценивали чистоту белка при помощи ДСН- ПАГ. (В) Исследование биспецифического антитела при помощи ДСН-ПАГ в невосстанавливающих (NR) и восстанавливающих (R) условиях.
Фиг. 5: Аффинная хроматография и ДСН-ПАГ эктодомена ЕрСАМ мыши
Культуральный супернатант, содержащий ЕрСАМ мыши с гистидиновым эпитопом (His-tag) на С-конце, очищали с использованием Ni-хелатной хроматографии. (А) Профиль элюирования с колонки HisTrap FF. (В) Анализ диализованного белка при помощи ДСН-ПАГ в восстанавливающих условиях. Гель окрашивали красителем Coomassie Brilliant Blue.
Фиг. 6: Взаимодействие эктодомена рекомбинантного ЕрСАМ мыши с антителами, специфическими к ЕрСАМ
Рекомбинантный мышиный ЕрСАМ взаимодействует с G8.8 или биспецифическим антителом MAB225_scFv_G8.8. (А) Иммуннопреципитацию рекомбинантного ЕрСАМ осуществляли с помощью G8.8. Звездочкой обозначен ЕрСАМ. (В) Иммуннопреципитацию рекомбинантного ЕрСАМ осуществляли с помощью MAB225_scFv_G8.8. Звездочкой обозначен ЕрСАМ.
Фиг. 7: Схематическое изображение нового принципа лечения при помощи мобилизации опухолеспецифических Т-клеток в опухоль с использованием биспецифического антитела
Т-клетки (в данном случае Т-клетки трансгенных мышей TCR-I; Т-клетки TCR tg) несут Т-клеточный рецептор к иммунодоминантному эпитопу 1 большого Т-антигена (Т спец. TCR). Эти Т-клетки дополнительно трансдуцированы маркерным антигеном (del-hEGFR; SEQ ID NOs: 11 и 12). Опухолевые клетки-мишени естественным образом экспрессируют большой Т-антиген, презентируемый главным комплексом гистосовместимости (ГКГС), и опухолевый антиген (ЕрСАМ). Биспецифическое антитело (MAb225_scFv_G8.8) с антигенсвязывающим сайтом для del-hEGFR на одном плече и для ЕрСАМ (мыши) на другом плече присоединяет клетки обоих типов. Специфический к опухолевому пептиду TCR, опухолевый пептид и МНС образуют "иммунный синапс".
Фиг. 8: Эффективность трансдукции первичных Т-клеток с использованием del-hEGFR
Выполняли ретровирусную трансдукцию первичных мышиных Т-клеток с использованием del-hEGFR (SEQ ID NOs: 11 и 12). Анализ при помощи проточной цитометрии показал эффективную трансдукцию с использованием del-hEGFR (темная кривая) по сравнению с нетрансдуцированными Т-клетками (светлая кривая).
Фиг. 9: Связывание трансдуцированных Т-клеток и опухолевых клеток при помощи биспецифического антитела
Клетки 4Т1, экспрессирующие ЕрСАМ, высаживали и выращивали до конфлюентности. Т-клетки B3Z, трансдуцированные Del-hEGFR (стабильная клеточная линия, флуоресцентно меченные), предварительно нагружали биспецифическим антителом (MAb225_scFv_G8.8) и затем инкубировали в чашке с адгезированными 4Т1. После тщательной отмывки оставшиеся клетки лизировали и замеряли остаточную флуоресценцию. Добавляли биспецифическое антитело, направленное против ЕрСАМ и hEFGR, или моноспецифические антитела (анти-ЕрСАМ и анти-hEGFR, служившие контролем). Биспецифическое антитело удерживало во флаконе значительно больше трансдуцированных клеток по сравнению с любым из контролей. ** указывает p<0,01 для всех сравнений.
Фиг. 10: Перенацеливание Т-клеток на лизис опухолевых клеток с помощью биспецифического антитела
Опухолеспецифические Т-клетки (TCR-I, Т-клетки, трансгенные по Т-клеточному рецептору, распознающему иммунодоминантный эпитоп большого Т-антигена) трансдуцировали del-hEGFR (SEQ ID NOs: 11 и 12) и преинкубировали с биспецифическим антителом (MAb225_scFv_G8.8 (SEQ ID NOs: 5 и 6)). В опухолевые клетки mGC8 (стабильная клеточная линия), экспрессирующие ЕрСАМ и большой Т-антиген, или в опухолевые клетки В16, не экспрессирующие ни один из них, вводили флуоресцентную метку (кальцеин). Клетки культивировали совместно при указанном соотношении опухолевых и Т-клеток (Т:Е) в течение ночи и количественно определяли лизис клеток по интенсивности испускаемой флуоресценции. Индукция лизиса, зависящего от соотношения клеток-мишеней и эффекторных клеток, наблюдалась в случае клеток mGC8, но не клеток В16.
Фиг. 11: Обработка развившихся опухолей комбинацией биспецифического антитела и трансдуцированных опухолеспецифических Т-клеток
Мышам вводили подкожно опухолевые клетки (клеточная линия mGC8). Мыши получали (внутривенно - при введении клеток; внутривенно и внурибрюшинно - при введении антитела) в указанные моменты времени либо ФСБ, только моноспецифическое антитело к ЕрСАМ (ЕрСАМ G8.8 (SEQ ID NOs: 3 и 4)), трансдуцированные Т-клетки (CD8+ Т-клетки) с моноспецифическим антителом к ЕрСАМ (ЕрСАМ G8.8 (SEQ ID NOs: 3 и 4)) и моноспецифическим антителом к EGFR (EGFR Mab225 (SEQ ID NOs: 1 и 2)), либо трансдуцированные Т-клетки (CD8+ Т-клетки) с биспецифическим антителом (MAb225_scFv_G8.8; специфическое к ЕрСАМ и EGFR (SEQ ID NOs: 5 и 6)). Это комбинированное воздействие вызывало значительное уменьшение и замедление опухолевого роста при сравнении с любым другим воздействием. Различия в объемах опухолей становились значимыми на 31 сутки при сравнении группы 1 с группой 4, на 40 сутки при сравнении группы 2 с группой 4 и на 50 сутки при сравнении группы 3 с группой 4 (p<0,001 для всех сравнений).
Фиг. 12: Обработка развившихся опухолей комбинацией биспецифического антитела и трансдуцированных опухолеспецифических Т-клеток значительно увеличивает выживаемость
Кривые выживаемости мышей в группах воздействия показаны на Фиг. 11. Исследование прекращали на 72 сутки. Комбинированное воздействие значительно увеличивало выживаемость мышей по сравнению со всеми тремя контрольными воздействиями. Различия в выживаемости были значимыми при сравнении группы 1 с группой 4, группы 2 с группой 4, и группы 3 с группой 4.
Фиг. 13: Обработка развившихся опухолей комбинацией биспецифического антитела и трансдуцированных опухолеспецифических Т-клеток (подтверждающее исследование)
Мышам вводили подкожно опухолевые клетки (mGC8), как и в исследовании, показанном на Фиг. 11. В указанные моменты времени мыши получали либо ФСБ, только моноспецифическое антитело к ЕрСАМ (ЕрСАМ G8.8 (SEQ ID NOs: 3 и 4)), биспецифическое антитело (MAb225_scFv_G8.8; специфическое к ЕрСАМ и EGFR (SEQ ID NOs: 5 и 6)), трансдуцированные Т-клетки (CD8+ Т-клетки) с моноспецифическим антителом к ЕрСАМ (ЕрСАМ G8.8 (SEQ ID NOs: 3 и 4)) и моноспецифическим антителом к EGFR (MAb 225 (SEQ ID NOs: 1 и 2)), либо трансдуцированные Т-клетки (CD8+ Т-клетки) с биспецифическим антителом (MAb225_scFv_G8.8; специфическое к ЕрСАМ и EGFR (SEQ ID NOs: 5 и 6)) в двух различных концентрациях. Это комбинированное воздействие вызывало значительное снижение и замедление опухолевого роста по сравнению с любым другим воздействием. Различия в объемах опухолей становились значимыми на 36 сутки при сравнении группы 1 с группой 5, на 47 сутки при сравнении группы 2 с группой 5, на 54 сутки при сравнении группы 3 с группой 5 и на 63 сутки при сравнении группы 4 с группой 5 (р<0,001 для всех сравнений).
Фиг. 14: Обработка развившихся опухолей комбинацией биспецифического антитела и трансдуцированных опухолеспецифических Т-клеток значительно увеличивает выживаемость (подтверждающее исследование)
Кривые выживаемости мышей в группах воздействия показаны на Фиг. 13. Дополнительная группа из трех мышей, не имеющих опухолей, получала комбинированное воздействие (как контроль на токсичность, обозначенная "7-без опухоли"). Комбинированное воздействие значительно увеличивало выживаемость мышей по сравнению с любым контрольным воздействием. Различия в выживаемости были значимыми при сравнении группы 1 с группой 5, группы 2 с группой 5, группы 3 с группой 5 и группы 4 с группой 5.
Фиг. 15: Белковая последовательность ЕрСАМ мыши, несущая His- Avitag
Белковая последовательность эктодомена ЕрСАМ, несущая на С-конце эпитоп His-Avi (соответствующая последовательность SEQ ID NO: 10), кодируемая последовательностью кДНК, показана в SEQ ID NO: 9.
Фиг. 16: Последовательность кДНК ЕрСАМ мыши
Последовательность кДНК эктодомена ЕрСАМ, несущая на С-конце эпитоп His-Avi (SEQ ID NO: 7).
Фиг. 17: Карта вектора, кодирующего эктодомен ЕрСАМ
Схематическая карта плазмидного вектора для экспрессии в эукариотических клетках, кодирующего эктодомен (ECD) ЕрСАМ.
Фиг. 18: Карта вектора, кодирующего легкую цепь МАВ225
Схематическая карта плазмидного вектора для экспрессии в эукариотических клетках, кодирующего легкую цепь МАВ225 (SEQ ID NO: 1).
Фиг. 19: Карта вектора, кодирующего тяжелую цепь МАВ225 с гибридизованным scFv G8.8
Схематическая карта плазмидного вектора для экспрессии в эукариотических клетках, кодирующего тяжелую цепь МАВ225, гибридизованную С- концом с scFv G8.8 (SEQ ID NO: 6).
Фиг. 20: Репрезентативные изображения, полученные при исследовании биспецифического антитела EpCAM-EGFRvIII, MR1.1 методом эксклюзионной хроматографии и ДСН-ПАГ
(А) Аналитическая эксклюзионная хроматография биспецифического антитела, направленного против EGFRvIII (человека) и ЕрСАМ (мыши), т.е. бсАт EpCAM-EGFRvIII, MR1.1. (В) Исследование указанного биспецифического антитела EpCAM-EGFRvIII, MR1.1 при помощи ДСН-ПАГ в невосстанавливающих (NR) и восстанавливающих (R) условиях. Окраска кумасси синим.
Фиг. 21: Схематическое изображение нового принципа лечения при помощи мобилизации опухолеспецифических Т-клеток в опухоль с использованием биспецифического антитела EpCAM-EGFRvIII, MR1.1
Схематическое изображение нового принципа лечения при помощи мобилизации опухолеспецифических Т-клеток к опухоли с использованием биспецифического антитела: Т-клетки (в данном случае Т-клетки трансгенных мышей OT-I) несут Т-клеточный рецептор, специфический к овальбумину. Эти Т-клетки дополнительно трансдуцированы маркерным антигеном (del-hEGFRvIII; SEQ ID NOs: 17 и 18). Опухолевые клетки-мишени (например, клетки меланомы В16, экспрессирующие овальбумин (OVA)), естественным образом экспрессируют большой Т-антиген, презентируемый главным комплексом гистосовместимости (ГКГС), и опухолевый антиген (ЕрСАМ). МНС на опухолевых клетках-мишенях в данном примере презентирует пептидный фрагмент SIINFEKL овальбумина. Биспецифическое антитело (EpCAM-EGFRvIII, MR1.1) с антигенсвязывающим сайтом для del-hEGFRvIII (SEQ ID NOs: 17 и 18) на одном плече и для ЕрСАМ (мыши) на другом плече присоединяет клетки обоих типов. Специфический к опухолевому пептиду TCR, опухолевый пептид и МНС образуют "иммунный синапс".
Фиг. 22: Связывание трансдуцированных Т-клеток и опухолевых клеток при помощи биспецифического антитела
Клетки меланомы В16, экспрессирующие ЕрСАМ (меченные GFP), высаживали и выращивали до конфлюентности. Т-клетки B3Z, трансдуцированные Del-hEGFRvIII (стабильная клеточная линия), предварительно нагружали биспецифическим антителом (EpCAM-EGFRvIII, MR1.1) и затем инкубировали в планшете с адгезированными В16 (столбец 5). После тщательной отмывки (столбцы 1-4) оставшиеся клетки трипсинизировали и подсчитывали флуоресцентные и нефлуоресцентные клетки. Биспецифическое антитело (EpCAM-EGFRvIII, MR1.1) после отмывки удерживало во флаконе больше трансдуцированных клеток (столбец 1) по сравнению с любым из контролей (столбцы 2-4).
Фиг. 23: Аминокислотные последовательности биспецифического антитела EpCAM-EGFRvIII, MR1.1
(А) Аминокислотная последовательность легкой цепи биспецифического антитела EpCAM-EGFRvIII, MR1.1 (без лидерной последовательности), обозначенная SEQ ID NO: 15. (В) Аминокислотная последовательность тяжелой цепи биспецифического продукта EpCAM-EGFRvIII, MR1.1 (без лидерной последовательности), обозначенная SEQ ID NO: 16.
Фиг. 24: Последовательности del-hEGFRvIII
(А) Последовательность ДНК del-hEGFRvIII (кодирующая белковую последовательность, изображенную на Фиг. 24(B)) показана в SEQ ID NO: 17. (В) Белковая последовательность del-hEGFRvIII; соответствующая SEQ ID NO: 18.
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Для иллюстрации правильности концепции, в следующих ниже примерах антитело к EGFR человека (Mab225; SEQ ID NOs: 1 и 2) скомбинировали с антителом к ЕрСАМ мыши (G8.8; SEQ ID NOs: 3 и 4) для образования биспецифического продукта (MAb225_scFv_G8.8; SEQ ID NOs: 5 и 6). Кроме того, как показано на Фиг. 20 и 21, было сконструировано биспецифическое антитело "бсАт EpCAM-EGFRvIII, MR1.1" (SEQ ID NOs: 15 и 16), имеющее антигенсвязывающий сайт для del-hEGFRvIII (SEQ ID NOs: 17 и 18) на одном плече и для ЕрСАМ (мыши) на другом плече; см. Пример 4.
Пример 1: Клонирование и экспрессия биспецифического антитела MAb225_scFv_G8.8
Технология рекомбинантной ДНК
Для работы с ДНК использовали стандартные методы, описанные Sambrook, J. et al., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Реагенты для молекулярной биологии использовали согласно инструкциям производителей.
Анализ последовательностей ДНК и белка и обработка данных
Общая информация о нуклеотидных последовательностях легких и тяжелых цепей иммуноглобулинов человека приведена Kabat, Е.А. et al., (1991) Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242. Аминокислоты цепей антитела пронумерованы в соответствии с системой нумерации EU (Edelman, G.M., et al., PNAS 63 (1969) 78-85; Kabat, E.A., et al., (1991) Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242). Для получения, картирования, анализа, аннотирования и иллюстрирования последовательностей использовали пакет программ GCGʹs (Genetics Computer Group, Madison, Wisconsin) вариант 10.2 и программу Vector NTI Advance suite вариант 11.5, разработанную компанией Infomax.
Секвенирование ДНК
Последовательности ДНК определяли путем секвенирования двуцепочечной матрицы, которое выполнялось компаниями SequiServe (Vaterstetten, Германия) и Geneart AG (Regensburg, Германия).
Синтез генов
Необходимые генные сегменты были получены в компании Geneart AG (Regensburg, Германия) из синтетических олигонуклеотидов и продуктов ПЦР при помощи автоматизированного синтеза генов. Генные сегменты, фланкированные уникальными сайтами расщепления рестрикционными эндонуклеазами, клонировали в плазмиды pGA18 (ampR). Плазмидную ДНК получали из трансформированных бактерий и определяли концентрацию при помощи УФ спектроскопии. Последовательности ДНК субклонированных генных фрагментов верифицировали путем секвенирования ДНК. Последовательности ДНК, кодирующие легкие цепи, содержали вариабельные и константные участки легкой цепи, фланкированные сайтами рестрикции эндонуклеаз Narl на 5ʹ-конце и Nhel на 3ʹ-конце. Последовательности ДНК, кодирующие вариабельные участки тяжелых цепей и фрагмент участка СН1 были фланкированы сайтами рестрикции Kpnl на 5ʹ-конце и BamHI на 3ʹ-конце. Последовательности ДНК, кодирующие конструкцию scFv, сожержали фрагмент домена СН3, фланкированный сайтами рестрикции для эндонуклеаз BsrGI на 5ʹ-конце и Xbal на 3ʹ-конце. Последовательность ДНК, кодирующая аминокислоты 1-267 ЕрСАМ мыши (SEQ ID NOs: 7 (последовательность кДНК) и 8 (аминокислотная последовательность)) имела сайты BamHI на 5ʹ-конце и NotI на 3ʹ-конце. Все конструкции имели 5ʹ-концевую последовательность ДНК, кодирующую лидерный пептид, обеспечивающий секрецию белков в эукариотических клетках.
1.1 Конструирование экспрессионных плазмид
Для конструирования всех цепей антитела использовали экспрессирующий вектор. Вектор состоит из следующих элементов (как проиллюстрировано на Фиг. 18 на примере легкой цепи Mab225 (SEQ ID NO: 1)):
- ориджин (точка начала репликации) oriP, из вируса Эпштейна-Барр (EBV),
- ориджин репликации из вектора pUC18 для репликации данной плазмиды в Е. coli
- ген бета-лактамазы, придающий резистентность Е. coli к ампициллину,
- предранний энхансер и промотор цитомегаловируса человека (HCMV),
- сигнальная последовательность для полиаденилирования ("поли-А") из бычьего гормона роста, и
- уникальные сайты расщепления рестрикционными эндонуклеазами KpnI, BamHI, BsrGI и XbaI (вектор для тяжелой цепи), или
- уникальные сайты расщепления рестрикционными эндонуклеазами NarI и NheI (легкая цепь).
Экспрессирующий вектор, кодирующий конструкцию тяжелой цепи, содержал следующие дополнительные элементы (схематически изображенные на Фиг. 19 для тяжелой цепи Mab225, гибридизованной с scFv G8.8 (SEQ ID NO: 6)):
- кассета резистентности к неомицину (neor),
- ранний промотор вируса обезьян 40, и
- сигнальная последовательность для полиаденилирования ("поли-А") из вируса обезьян 40.
Плазмиды pG18 (ampR), несущие синтезированные сегменты ДНК, кодирующие антитело, и экспрессирующий вектор расщепляли либо рестрикционными ферментами KpnI и BamHI в случае экспрессирующего вектора для тяжелой цепи, либо NarI и NheI в случае вектора для легкой цепи. Аналогично, плазмиды pG18, несущие фрагмент scFv, разрезали BsrGI и XbaI, как и вектор для тяжелой цепи. Все полученные фрагменты подвергали электрофорезу в агарозном геле. Очищенные сегменты ДНК затем лигировали с выделенными фрагментами экспрессирующего вектора KpnI/BamHI, NarI/NheI или BsrGI/XbaI с получением результирующих экспрессирующих векторов. Результирующими экспрессирующими векторами трансформировали клетки Е. coli, выделяли плазмидную ДНК (Miniprep) и проводили рестрикционный анализ и секвенирование ДНК. Клоны с правильной последовательностью ДНК выращивали в 150 мл среды LB-Amp, вновь выделяли плазмидную ДНК (Maxiprep) и использовали в последующих экспериментах.
Для конструирования эктодомена рекомбинантного ЕрСАМ мыши (соответствующая аминокислотная последовательность обозначена SEQ ID NO: 10 (кодируется последовательностью кДНК, приведенной в SEQ ID NO: 9) использовали экспрессирующий вектор, тогда как последовательность ориджина была найдена в PubMed под номером NM_008532 (NM_008532.2, GI:112293274). Вектор (см. Фиг. 17) состоял из следующих элементов:
- ориджин репликации oriP, из вируса Эпштейна-Барр (EBV),
- ориджин репликации из вектора pUC18 для репликации данной плазмиды в Е. coli
- ген бета-лактамазы, придающий резистентность Е. coli к ампициллину,
- кассета, кодирующая гигромицин-b-фосфотрансферазу,
- ранний промотор вируса обезьян 40,
- сигнальная последовательность для полиаденилирования ("поли-А") из вируса обезьян 40,
- предранний энхансер и промотор цитомегаловируса человека (HCMV),
- сайт PreScission Plus, за которым следовал эпитоп His-Avitag для N-концевого сшивания представляющих интерес кДНК,
- сигнальная последовательность для полиаденилирования ("поли-А") из бычьего гормона роста, и
- уникальные сайты для рестрикционных эндонуклеаз BamHI и NotI.
Плазмиды pG18 (ampR), несущие синтезированный сегмент ДНК, кодирующий эктодомен ЕрСАМ, и экспрессирующий вектор (см. Фиг. 17) расщепляли BamHI и NotI. Все полученные фрагменты подвергали электрофорезу в геле агарозы. Очищенные сегменты ДНК затем лигировали с выделенным фрагментом экспрессирующего вектора Roche, расщепленного BamHI/NotI, с получением результирующего экспрессирующего вектора. Результирующим экспрессирующим вектором трансформировали клетки E. coli, выделяли плазмидную ДНК (Miniprep) и проводили рестрикционный анализ и секвенирование ДНК. Клоны с правильной последовательностью ДНК выращивали в 150 мл среды LB-Amp, вновь выделяли плазмидную ДНК (Maxiprep) и использовали в последующих экспериментах.
Пример 2: Экспрессия и очистка эктодомена ЕрСАМ, демонстрирующего высокую аффинность связывания
2.1 Транзиторная экспрессия вариантов иммуноглобулина в эмбриональных клетках почки человека HEK293
Варианты рекомбинантных иммуноглобулинов (МАВ225, MAB225_scFv_G8.8 и G8.8) экспрессировали при транзиторной трансфекции эмбриональных клеток почки человека 293-F с использованием экспрессирующей системы Freestyle™ 293 согласно инструкциям производителя (Invitrogen, США). Вкратце, суспензию клеток FreeStyle™ 293-F культивировали в среде для экспрессии Freestyle™ 293 при 37°С/8% CO2. За день до трансфекции клетки высаживали в свежую среду при плотности 1×106 жизнеспособных клеток/мл. Комплексы DNA-293fectin™ готовили в среде Opti-MEM® I (Invitrogen, США) с использованием 665 мкл реагента для трансфекции 293-Free™ (Novagen, EMD, США) и 500 мкг общей ДНК (включая плазмидную ДНК, кодирующую легкую цепь, и плазмидную ДНК, кодирующую тяжелую цепь, в молярном соотношении 1:1) в общем объеме среды 500 мл. Надосадочную культуральную жидкость, содержащую антитела, собирали через 6 дней после трансфекции путем центрифугирования при 3500 об/мин в течение 15 минут при комнатной температуре (RT) и фильтровали через стерильный фильтр (0,22 мкм). Надосадочную жидкость хранили при -80°С до очистки. Процедура получения эктодомена ЕрСАМ мыши была аналогична процедуре получения вариантов иммуноглобулинов, а для трансфекции использовали 500 мкг ДНК в конечном объеме среды 500 мл.
2.2 Очистка антител
Антитела выделяли из культуральной надосадочной жидкости путем аффинной хроматографии с применением Protein A-Sepharose™ (GE Healthcare, Швеция) и эксклюзионной хроматографии на колонках Superdex200. Вкратце, стерилизованную фильтрованием надосадочную культуральную жидкость наносили на колонку MabSelect Xtra (GE Healthcare) объемом 5 мл, уравновешенную ФСБ (10 мМ Na2HPO4, 1 мМ KH2PO4, 137 мМ NaCl и 2,7 мМ KCl, рН 7,4). Несвязанные белки отмывали уравновешивающим буфером. Антитело и варианты антитела элюировали 0,1 M цитратным буфером, pH 3,0, а фракции, содержащие белок, нейтрализовали 0,2 M Tris, pH 9,0. Затем элюированные фракции, содержащие белок, объединяли, концентрировали с помощью ультрафильтрационных устройств Amicon Ultra (пористость мембран: 30 K, Millipore) до объема 3 мл и загружали в колонку для гель-фильтрации Superdex200 HiLoad 120 мл 16/60 или 26/60 (GE Healthcare, Швеция), уравновешенную 20 мМ гистидина, 140 мМ NaCl, pH 6,0. Фракции, содержащие очищенные антитела с процентным содержанием высокомолекулярных агрегатов менее 5%, объединяли и хранили в аликвотах с приблизительной концентрацией 1,0 мг/мл при -80°C.
Эктодомен ЕрСАМ мыши (SEQ ID NO: 9) выделяли из культуральной надосадочной жидкости при помощи аффинной хроматографии с использованием Ni-хелатной хроматографии. Вкратце, стерилизованную фильтрованием культуральную надосадочную жидкость наносили на колонку HisTrap FF (GE Healthcare, Швеция), уравновешенную 20 мМ NaPO4, 0,5 M NaCl, 20 мМ имидазола, pH 7,40. Несвязанные белки отмывали уравновешивающим буфером. Эктодомен ЕрСАМ (SEQ ID NO: 9) элюировали буфером, содержащим 20 мМ NaP04, 0,5 M NaCl, 500 мМ имидазола, pH 7,40. Пиковые фракции объединяли и диализовали в течение ночи против буфера, содержащего 20 мМ гистидина, 140 мМ NaCl, pH 6,70. Белок хранили в аликвотах с концентрацией 17,5 мг/мл при -80°C.
2.3 Анализ очищенных белков
Концентрацию белка в очищенных образцах белка определяли, измеряя оптическую плотность (OD) при 280 нм, используя молярный коэффициент экстинкции, рассчитанный по аминокислотной последовательности. Степень чистоты и молекулярный вес антител анализировали с помощью ДСН-ПАГ в присутствии и в отсутствие восстанавливающего агента (5 мМ 1,4-дитиотрейтол) и окрашивания кумасси бриллиантовым синим. Системы гелей NuPAGE® Pre-Cast (Invitrogen, США) использовали согласно инструкциям производителя (4-12% бис-трис гели). Содержание агрегатов в образцах антител анализировали при помощи высокоэффективной эксклюзионной хроматографии с использованием колонок TSKgel G3000SW (TOSOH Bioscience, США) в проточном буфере 50 мМ KPO4, pH 7,5, 300 мМ NaCl при 25°C. В колонку вкалывали 25 мкг белка при скорости 0,8 мл/мин и элюировали в изократическом режиме в течение 20 минут. Целостность аминокислотного скелета цепей восстановленных вариантов иммуноглобулинов верифицировали при помощи масс-спектрометрии NanoElectrospray Q-TOF после удаления N-гликанов ферментативной обработкой пептид-N-гликозидазой F (Roche Molecular Biochemicals).
2.4 Иммунопреципитация рекомбинантного ЕрСАМ мыши
Иммунопреципитацию выполняли для проверки способности антитела G8.8 (SEQ ID NOs: 3 и 4) и биспецифического антитела MAB225_G8.8 (SEQ ID NOs: 5 и 6) связываться с эктодоменом (ECD) рекомбинантного ЕрСАМ мыши (SEQ ID NO: 10). Приблизительно 15 мкг ЕрСАМ ECD и 10 мкг антитела разводили буфером TBS с добавлением 0,1% Tween до конечного объема 1,5 мл и инкубировали в течение 10 мин при комнатной температуре. Параллельно проводили обработку антитела в отсутствие ЕрСАМ, используемое в качестве отрицательного контроля, для идентификации полос, соответствующих тяжелой и легкой цепи. Добавляли 50 мкл 50% суспензии белка А и инкубировали образцы в течение 40 мин при помешивании при комнатной температуре, отмывали TBS-T, элюировали цитратным буфером с рН 3,0 и вносили в 4-12% бис-трис гель. В качестве контроля использовали эктодомен рекомбинантного ЕрСАМ в количестве 4 мкг. Гель окрашивали кумасси бриллиантовым синим.
2.5 Масс-спектрометрия
Массу целых антител (в дегликозилированной форме) определяли и подтверждали при помощи масс-спектрометрии с ионизацией электрораспылением (ESI-MS). Вкратце, 100 мкг очищенных антител дегликозилировали, обрабатывая N-гликозидазой F (PNGaseF, ProZyme) в количестве 50 мЕд в 100 мМ KH2PO4/K2HPO4, pH 7 при 37°C в течение 12-24 ч при концентрации белка до 2 мг/мл и затем обессоливали при помощи ВЭЖХ на колонке Sephadex G25 (GE Healthcare). Массу соответствующих цепей антител определяли при помощи ESI-MS после дегликозилирования и восстановления. Вкратце, 50 мкг антител в 115 мкл инкубировали в 60 мкл 1М ТСЕР и 50 мкл 8 M гуанидин-HCl с последующим обессоливанием. Массу целых антител и массу цепей антител после восстановления определяли при помощи ESI-MS с использованием системы Q-Star Elite MS, оборудованной источником ионизации NanoMate. Диапазон измеряемых масс зависит от молекулярного веса образцов. Как правило, для восстановленных антител задавали диапазон масс 600-2000 m/z, а для невосстановленных антител 1000-3600 m/z.
2.6 Поверхностный плазмонный резонанс
Связывающие свойства EGFR-специфического Mab225 (SEQ ID NOs: 1 и 2) и ЕрСАМ-специфического антитела G8.8 (SEQ ID NOs: 3 и 4), а также биспецифического антитела MAb225_scFv_G8.8 определяли при помощи технологии поверхностного плазмонного резонанса (SPR) с использованием инструмента Biacore (Biacore, GE-Healthcare, Uppsala). Данная система хорошо подходит для изучения молекулярных взаимодействий. Она позволяет непрерывно отслеживать в реальном времени связывание лиганд/аналит и таким образом определять константы скорости ассоциации (ka), константы скорости диссоциации (kd), и равновесные константы (KD) в различных условиях экспериментов. Технология SPR основана на измерении показателя преломления в поверхностном слое покрытого золотом чипа биосенсора. Изменения показателя преломления указывают на изменения массы на поверхности, вызванные взаимодействием иммобилизованного лиганда с аналитом в инжектируемом растворе. Если молекулы связываются с иммобилизованным лигандом на поверхности, масса увеличивается, в случае диссоциации масса уменьшается. В качестве захватывающего антитела на поверхности чипа биосенсора СМ5 иммобилизовали антитело козы к IgG мыши при помощи связывания через аминогруппы согласно инструкциям производителя. Проточные ячейки активировали смесью 0,1 M N-гидроксисукцинимида и 0,4 M 1-этил-3-(3-диметиламинопропил)-карбодиимид-гидрохлорида в соотношении 1:1 при скорости потока 5 мкл/мин при 25°С. Антитело к IgG мыши инжектировали в 10 мМ ацетате натрия, pH 4,5, при концентрации 10 мкг/мл. Проточную ячейку, служившую референтным контролем, обрабатывали аналогичным образом, но вместо захватывающего антитела использовали буфер. Поверхности блокировали, пропуская 1 M этаноламин/HCl pH 8,5. Изучаемые антитела разводили (30 нМ MAb225_scFv_G8.8, 5 нМ Mab225, 8 нМ G8.8) в ФСБ-Т+0,1% бычьего сывороточного альбумина (буфер для разведения) и для захвата в качестве лигандов их инжектировали в отдельных циклах со скоростью 5 мкл/мин в течение 60 сек. Взаимодействие происходило при 37°C, при использовании ФСБ-Т в качестве проточного буфера. Аналиты инжектировали в сериях с трехкратно увеличивающимися концентрациями (EGFR ECD 4,12 - 1000 нМ, ЕрСАМ ECD 0,91 - 2000 нМ в ФСБ-Т + 0,1% бычьего сывороточного альбумина) при скорости потока 50 мкл/мин, время ассоциации составляло 180 сек, время диссоциации 1200 сек. Для регенерации захватывающего антитела в конце каждого цикла пропускали 10 мМ глицин, pH 2,0 при скорости потока 30 мкл/мин в течение 60 сек. Сигналы регистрировали со скоростью один сигнал в секунду.
В Таблице 1 приведена краткая биохимическая характеристика взятого в качестве примера биспецифического антитела MAb225_scFv_G8.8, а также отдельных родительских антител Mab225 и G8.8, демонстрирующая, что все антитела связывались со своими соответствующими мишенями с высокой аффинностью. 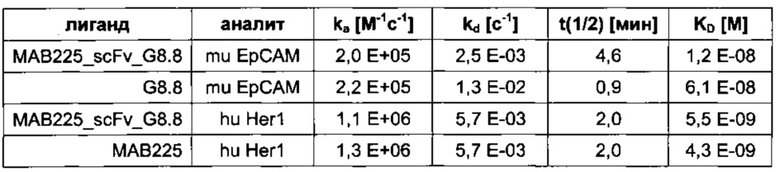
Пример 3: Трансдукция CD8+ Т-клеток и исследование цитотоксической активности
3.1 Культура клеток
Селезенку выделяли у мышей, описанных ниже в п.3.2. Суспензию одиночных клеток получали при протирании селезенки через стрейнер с размером пор 40 мкм (BD Falcon, Германия). Стрейнер промывали 3 раза чистой средой для Т-клеток и из клеточного осадка удаляли эритроциты, инкубируя с раствором, лизирующим эритроциты (BD Pharmingen, Германия), в течение 90 секунд. Первичные мышиные Т-клетки находились в среде для Т-клеток, состоящей из RPMI, 1% L-глутамина, 1% пенициллина и стрептомицина, 1% пирувата натрия, 1 мМ HEPES (все производства РАА, Германия) и 10% эмбриональной телячьей сыворотки (Gibco, США). Линию Т-клеток B3Z (Leisegang et al., J Mol Med (2008), 86(5), 573-583) поддерживали в той же среде. Пакующую клеточную линию Plat E (Cell Biolabs, Inc, США (www.cellbiolabs.com)) поддерживали в DMEM с добавлением 1% L-глутамина, 1% пенициллина-стрептомицина (все - производства РАА, Германия), 10 мкг/мл пуромицина и 1 мкг/мл бластицидина (Sigma, Германия). Линия опухолевых клеток желудка мыши была любезно предоставлена W. Zimmermann, Munich, и ее культивировали в DMEM с добавлением 1% L-глутамина, 1% пенициллина и стрептомицина, 1% пирувата натрия, 1% заменимых аминокислот и 10% эмбриональной телячьей сыворотки. Линию клеток карциномы молочной железы мыши 4Т1 (АТСС NO:°CRL-2539) высаживали в RPMI с добавлением 1% L-глутамина, 1% пенициллина и стрептомицина, 1% пирувата натрия, 1% заменимых аминокислот и 10% эмбриональной телячьей сыворотки. Линию клеток меланомы мыши В16 (АТСС NO°CRL-6322) поддерживали в DMEM с добавлением 1% L-глутамина, 1% пенициллина и стрептомицина и 10% эмбриональной телячьей сыворотки.
3.2 Мыши
Мыши C57BI/6 дикого типа были приобретены в компании Harlan laboratories (Нидерланды). Мыши, трансгенные по Т-клеточному рецептору, специфическому к иммунодоминантному эпитопу I большого Т-антигена, были приобретены в компании Jackson Laboratory, США (B6.Cg-Tg(TcraY1,TcrbY1)416Tev/J), USA). Мыши, трансгенные по Т-клеточному рецептору, специфическому к OVA (овальбумину), были приобретены в компании Jackson Laboratory, США (C57BI/6-Tg(TcraTcrb)1100Mjb/j).
3.3 Т-клетки: трансдукция с помощью вектора и клонирование
Вектором для трансдукции Т-клеток был рМР71 (Schambach et al., Mol Ther (2000), 2(5), 435-45). Укороченный человеческий рецептор эпидермального фактора роста (del-EGFR, как типичный пример иммунологически инертного белка, встроенного в мембрану, см. SEQ ID NO 11 для последовательности кДНК и SEQ ID NO: 12 для (кодируемой) аминокислотной последовательности), фланкированный рестрикционными сайтами NotI и EcoRI, был создан при помощи синтеза генов. С помощью этих двух сайтов рекомбинации del-EGFR клонировали в рМР71 путем лигирования (Т4-лигазой, Thermo scientific, Германия). Del-EGFR-рМР71 амплифицировали в клетках E.coli (DH5α) и верифицировали последовательность путем секвенирования, используя следующие последовательности праймеров: смысловой праймер: CAGCATCGTTCTGTGTTGTCT (SEQ ID NO: 13), антисмысловой праймер: CATTTAAATGTATACCCAAATCAA (SEQ ID NO: 14). ДНК выделяли с помощью набора для выделения плазмидной ДНК Qiagen plasmid maxi kit (Qiagen, Германия). Все этапы выполняли согласно инструкциям производителя.
3.4 Первичные Т-клетки и трансдукция линии Т-клеток
Трансдукцию осуществляли согласно способу, описанному Leisegang et al. (J Mol Med, (2008) 86, 573-83) с небольшими модификациями. Вкратце, пакующую клеточную линию Plat E (описанную Monta et al., Gene Ther (2000). 7, 1063-6) высаживали в 6-луночные планшеты и выращивали в течение ночи до конфлюентности 70-80%. В первый день смешивали 16 мкг ДНК и 100 мМ CaCl2 (Merck, Германия) и 126,7 мкМ хлорохина (Sigma, США). Клетки Plat-E подвергали сывороточному голоданию в течение 30 мин в среде с низким содержанием сыворотки (3%) и затем инкубировали в течение 6 ч с преципитированной ДНК. Затем среду удаляли и заменяли на культуральную среду. На второй день у мышей C57BI/6 выделяли первичные спленоциты (Harlan Laboratories, Нидерланды). Суспензию одиночных спленоцитов стимулировали в течение ночи с использованием анти-CD3 (клон 145-2с11 BD Pharmingen, США), анти-CD28 (клон 37.51, BD Pharmingen, США) и рекомбинантного мышиного IL-2 (Peprotech, Германия) в среде для Т-клеток. На третий день 24-луночные планшеты покрывали рекомбинантным ретронектином в концентрации 12,5 мкг/мл (Takara Biotech, Япония) в течение 2 ч при комнатной температуре, блокировали 2% бычьим сывороточным альбумином (Roth, Германия) в течение 30 мин при 37°C и отмывали фосфатным солевым буфером (ФСБ). Надосадочную жидкость Plat E собирали и пропускали через фильтр (40 мкм, Milipore, США). Затем к клеткам Plat E добавляли свежую среду для Т-клеток. В каждую лунку добавляли по 1 мл профильтрованной надосадочной жидкости и инокулировали путем центрифугирования в течение 2 ч при 4°C. Затем надосадочную жидкость удаляли из 24-луночного планшета. 106 Т-клеток высаживали в среде для Т-клеток с добавлением 10 Ед IL-2 и 400000 гранул, покрытых антителами к CD3 и CD28 (Invitrogen, Германия), в объеме 1 мл на лунку и инокулировали путем центрифугирования при 800 g в течение 30 мин при 32°С. На четвертый день надосадочную жидкость Plat E вновь собирали и фильтровали. Добавляли по 1 мл в каждую лунку 24-луночного планшета и инокулировали путем центрифугирования при 800 g в течение 90 мин при 32°C. Затем клетки инкубировали еще в течение 6 часов при 37°С. 1 мл надосадочной жидкости заменяли на среду для Т-клеток с IL-2. На пятый день клетки собирали, подсчитывали и вновь высаживали в плотности 106 клеток/мл в среде для Т-клеток с добавлением 10 нг/мл IL-15 (Peprotech, Германия). Т-клетки поддерживали в такой плотности до 10 дня, когда проводили анализ клеток или функциональные исследования.
3.5 Связывание клеток при помощи антитела
Клетки 4Т1, экспрессирующие ЕрСАМ, высаживали в 6-луночные планшеты и выращивали до конфлюентности. Трансдуцированные del-EGFR Т-клетки метили кальцеином (Invitrogen, Германия) в соответствии с инструкциями производителя и предварительно нагружали биспецифическим антителом к ЕрСАМ и EGFR (MAb225_scFv_G8.8) в концентрации 20 мкг/мл в течение 30 мин при 37°С. Затем клетки добавляли в культуру 4Т1 на 2 ч при 37°С. Надосадочную жидкость полностью удаляли, а планшет тщательно промывали ФСБ. Затем клетки визуализировали при помощи флуоресцентной микроскопии или лизировали и определяли высвобождение кальцеина при помощи многофункционального планшетного анализатора (Berthold, Германия).
3.6 Исследование цитолиза
Клетки mGC8 (Nöckel et al., ВМС (2006), 14, 6:57) или клетки В16 метили кальцеином в соответствии с инструкциями производителя. Трансдуцированные del EGFR Т-клетки (мышей дикого типа, ОТ-1 или TCR-I) предварительно нагружали антителом в концентрации 20 мкг/мл (биспецифическим или контрольным антителом) в течение 30 мин при 37°С. Затем Т-клетки инкубировали с клетками-мишенями при различном соотношении клеток-мишеней и эффекторных клеток в течение ночи при 37°С. Полный лизис клеток-мишеней вызывали добавлением 6% Triton X (Firma). Лизис, выраженный в процентах, рассчитывали по следующей формуле:
(MFIwieTOK, представляющих интерес - MFIфоновый уровень)/(MFIполный лизис - MFIфоновый уровень)*100
3.7 Терапия in vivo
Мышам C57BI/6 дикого типа подкожно инокулировали 5×106 клеток mGC8. Когда опухоли становились пальпируемыми (на 15 день), начинали трансдукцию Т-клеток и через 10 дней мыши получали терапию внутрибрюшинно (антитела в дозе 10 мг/кг) или внутривенно (Т-клетки или предварительно нагруженные Т-клетки в дозе 5×106 клеток на мышь). Через одну неделю процедуру повторяли. Состояние мышей контролировали каждые 2-3 дня, измеряя подкожные опухоли. Мышей умерщвляли в соответствии с установленными правилами либо в определенный момент времени, либо в соответствии с критериями, опубликованными GV SOLAS (Morton, Vet Rec (1985) 116, 431-436).
3.8. Статистический анализ
Для сравнения групп между собой использовали Т-критерий для непарных наблюдений. Объем опухолей сравнивали с использованием дисперсионного анализа, а различия в выживаемости оценивали с помощью лог-рангового критерия. Статистический анализ проводили с использованием программы GraphPad Prism (GraphPad software inc.). Результаты считали значимыми при p<0,05.
Пример 4: Клонирование и экспрессия биспецифического антитела EpCAM-EGFRvIII, MR1.1
Аналогично Примерам 1 и 2 получали, очищали и характеризовали биспецифическое антитело EpCAM-EGFRvIII, MR1.1, направленное против EGFRvIII (человека) и ЕрСАМ (мыши) (см. Фиг. 20 (А) аналитическая эксклюзионная хроматография, (В) анализ указанного биспецифического антитела с помощью ДСН-ПАГ в невосстанавливающих (NR) и восстанавливающих (R) условиях. Окрашивание кумасси синим).
Пример 5: Связывание трансдуцированных Т-клеток с клетками меланомы В16 с помощью биспецифического антитела EpCAM-EGFRvIII, MR1.1 (см. схему на Фиг. 21)
Клетки меланомы В16, экспрессирующие ЕрСАМ (меченные GFP) (клетки B16-OVA-mEpCAM), высаживали в 12-луночные планшеты и выращивали до конфлюентности в течение ночи. На следующий день трансдуцированные del-hEGFRvIII (вставка del-hEGFRvIII; см. SEQ ID NO: 17 для последовательности ДНК и SEQ ID NO: 18 для (кодируемой) аминокислотной последовательности) Т-клетки B3Z (стабильная клеточная линия) преинкубировали с биспецифическим антителом EpCAM-EGFRvIII, MR1.1 (20 мкг) в течение 1 часа при 37°С. Затем оставшееся несвязанное биспецифическое антитело отмывали. Т-клетки B3Z, предварително нагруженные биспецифическим антителом (бсАт), инкубировали с адгезированными опухолевыми клетками В16 при 37°С (столбец 5). После тщательной 4-кратной отмывки ФСБ (столбцы 1-4) оставшиеся клетки трипсинизировали и подсчитывали флуоресцентных и нефлуоресцентных клеток при помощи проточной цитометрии. Контрольные клетки меланомы В16 инкубировали с Т-клетками B3Z, трансдуцированными delEGFRvIII (без антитела EpCAM-EGFRvIII; см. столбец 2), с нетрансдуцированными Т-клетками B3Z (без антитела EpCAM-EGFRvIII, см. столбец 3) с отмывкой и без отмывки и нетрансдуцированными Т-клетками B3Z с биспецифическим антителом EpCAM-EGFRvIII (столбец 4). Биспецифическое антитело (бсАт) EpCAM-EGFRvIII удерживало во флаконе больше трансдуцированных клеток (столбец 1) по сравнению с любым из контролей при отмывке (столбцы 2-4). Результаты показаны на Фиг. 22.


















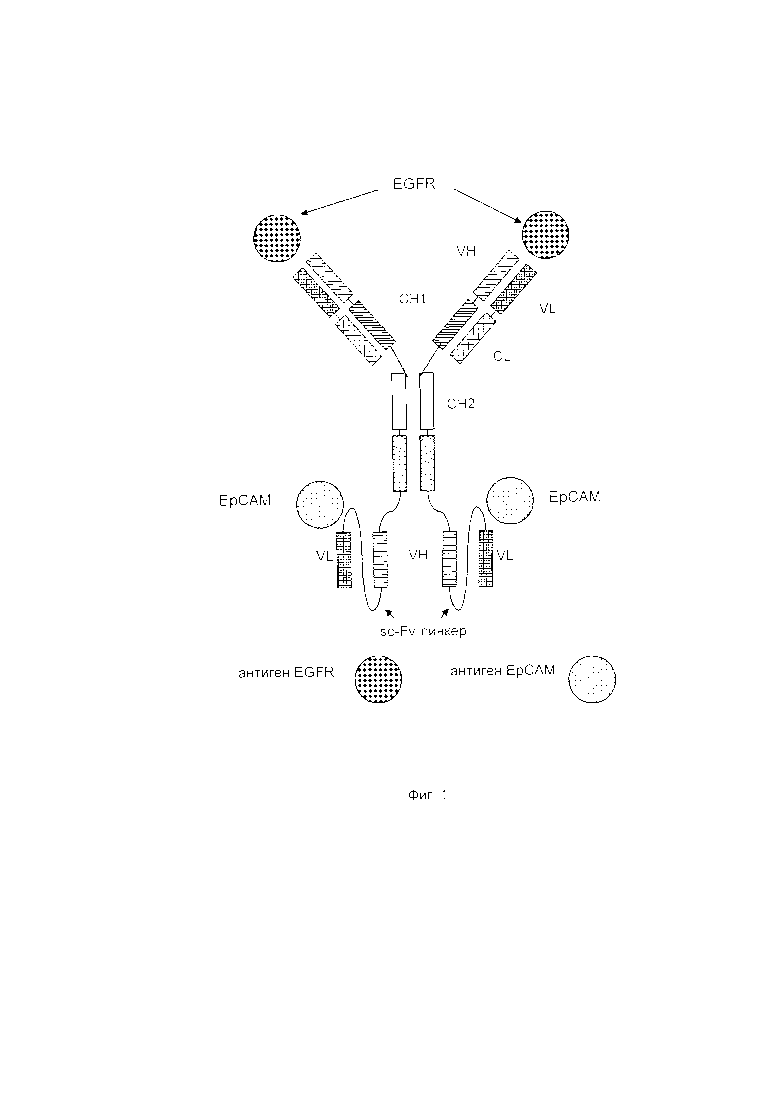
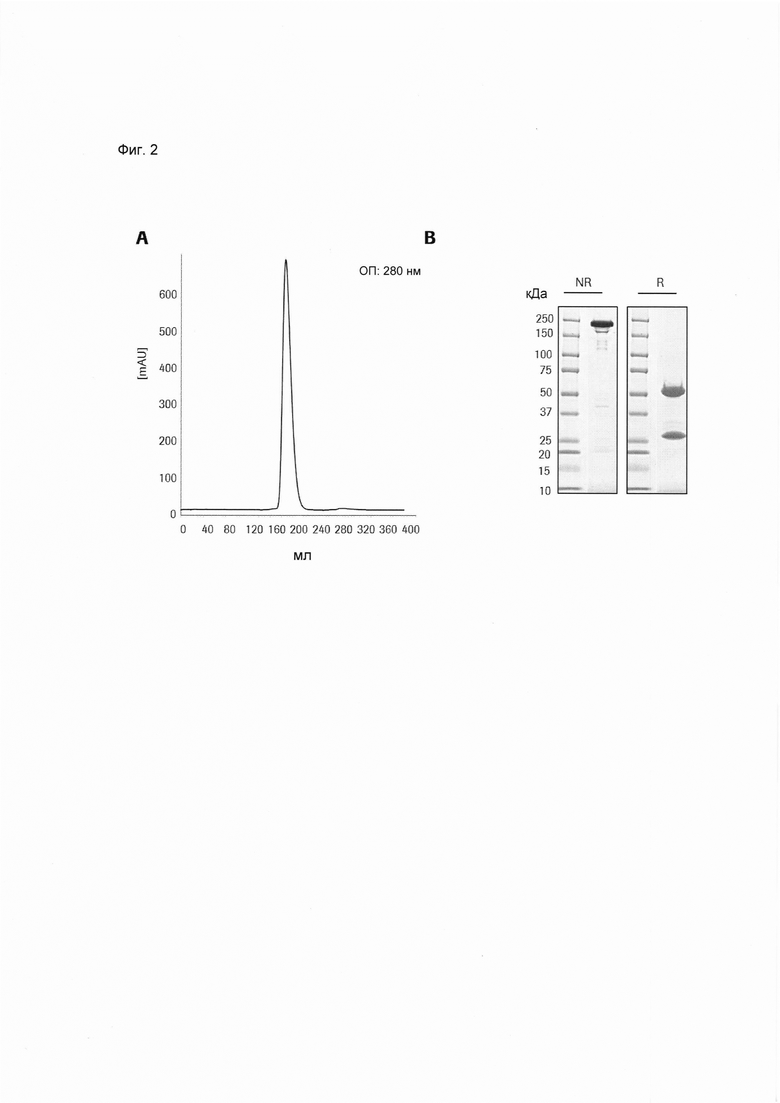
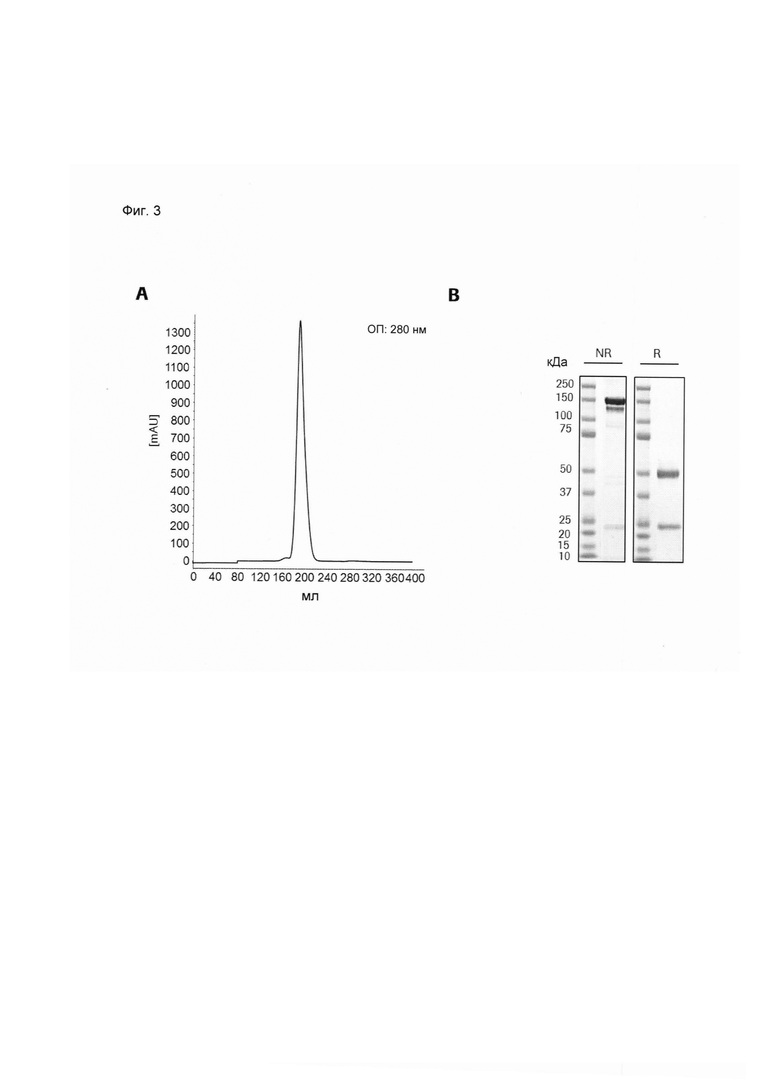
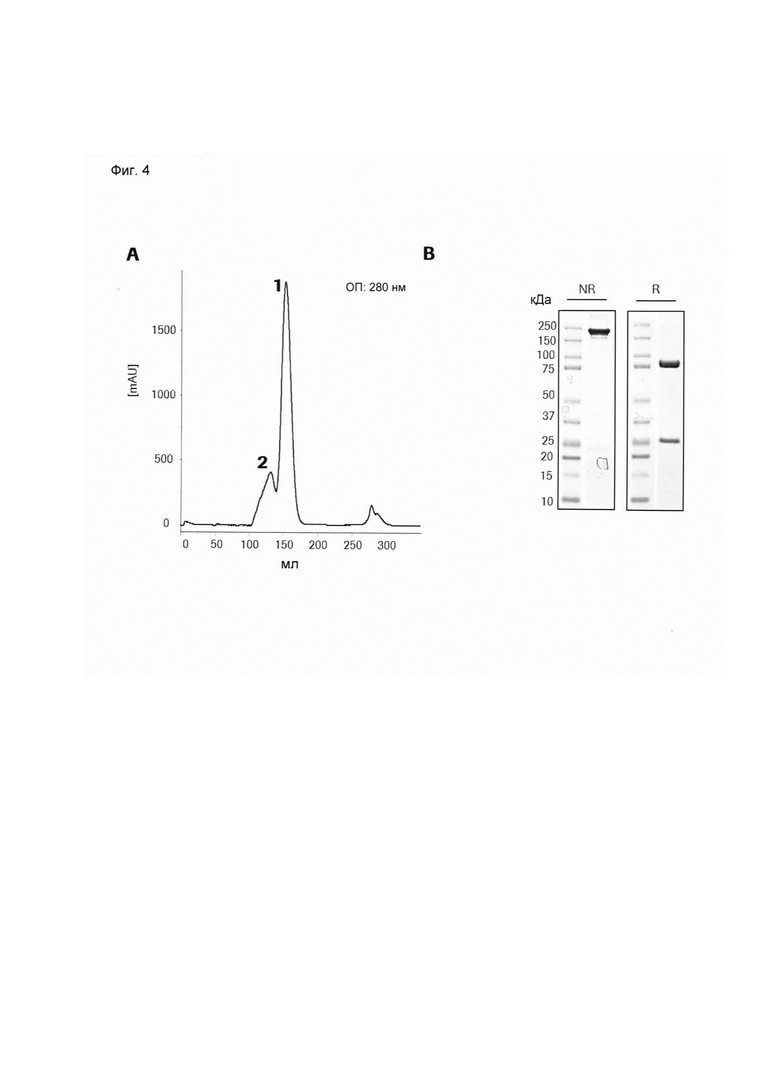
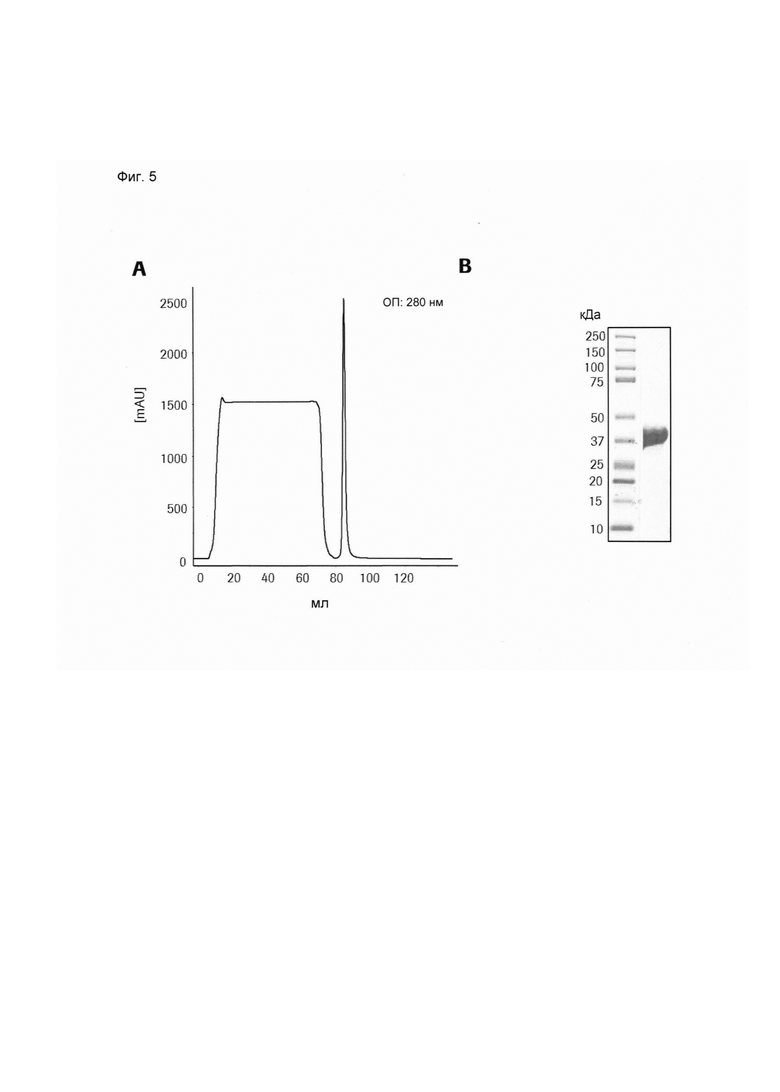
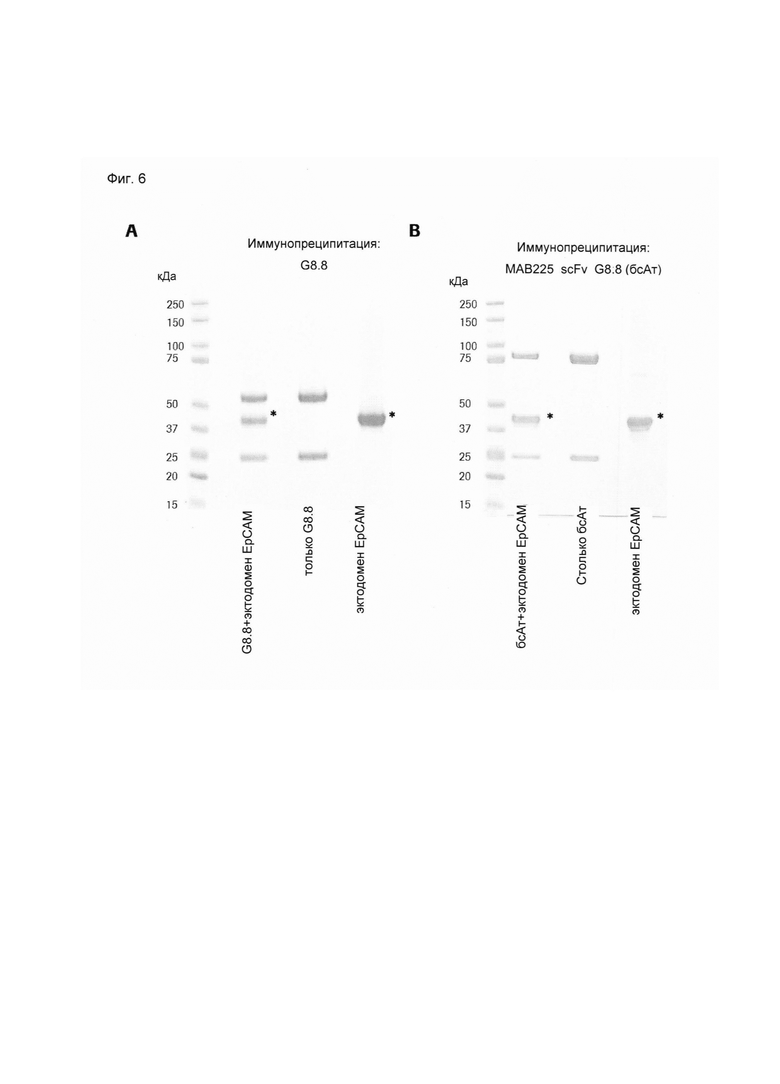
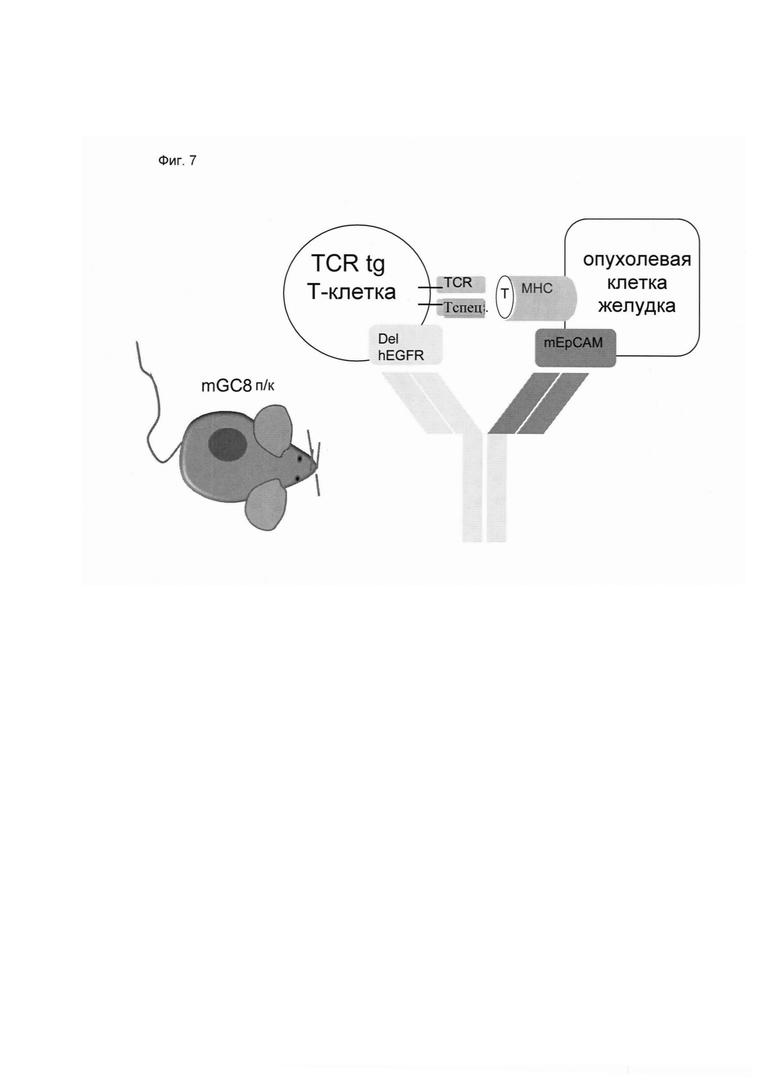
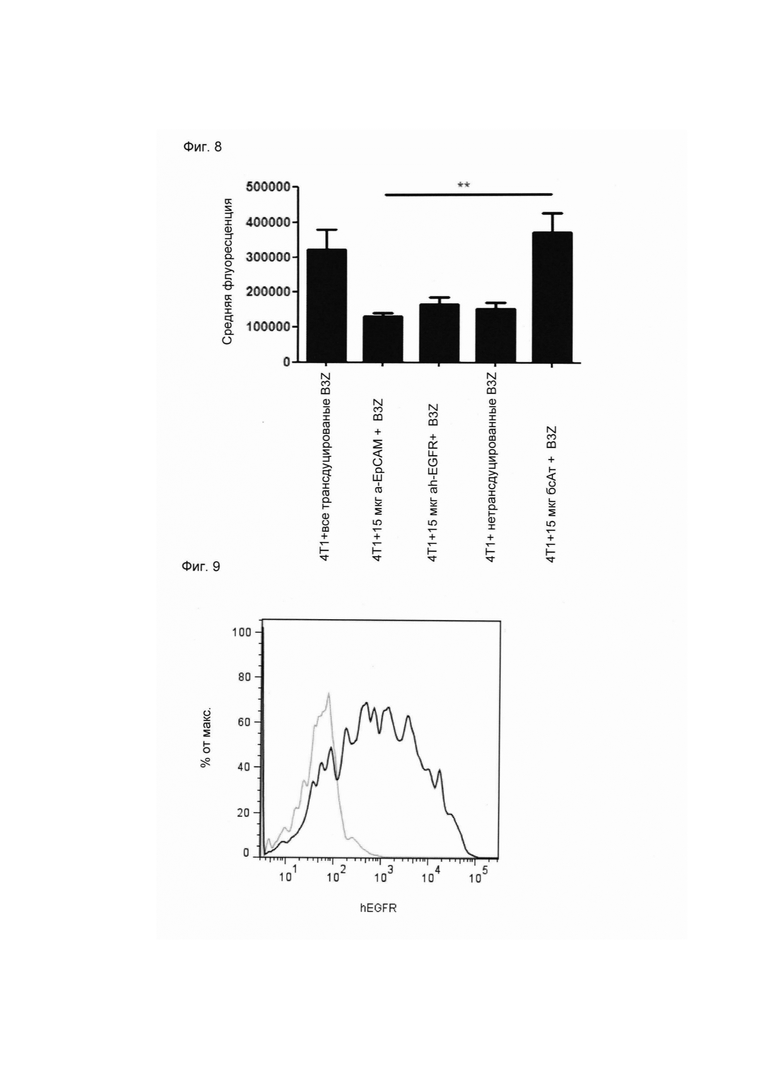
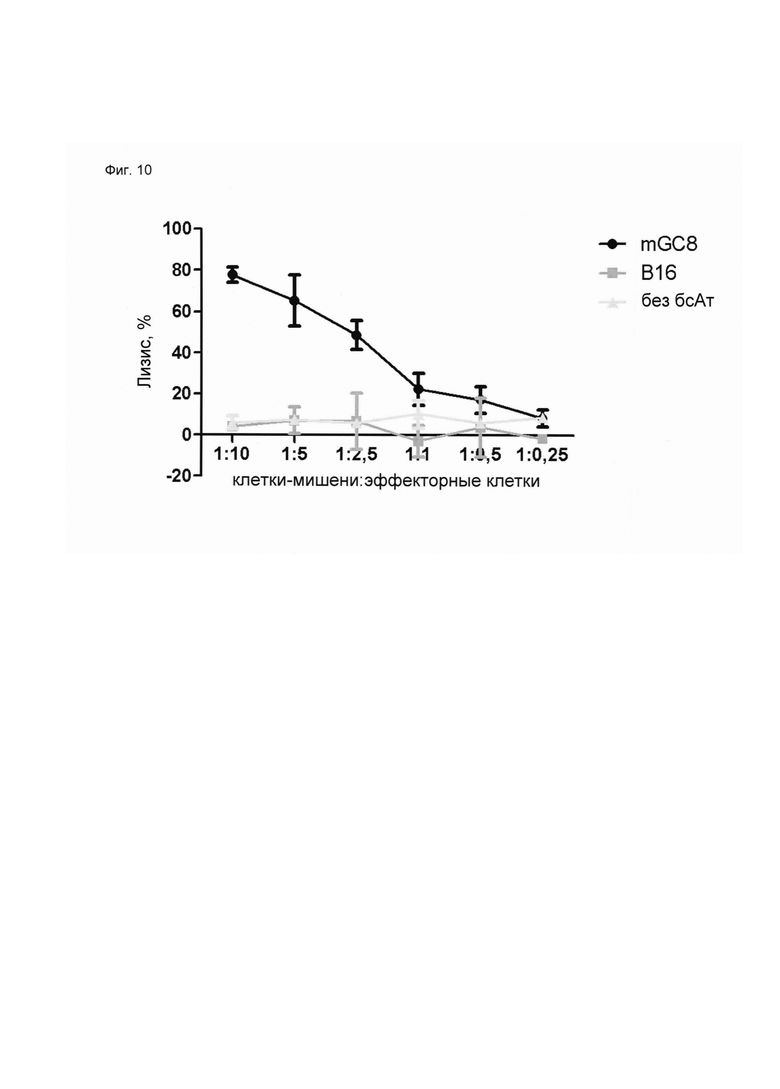
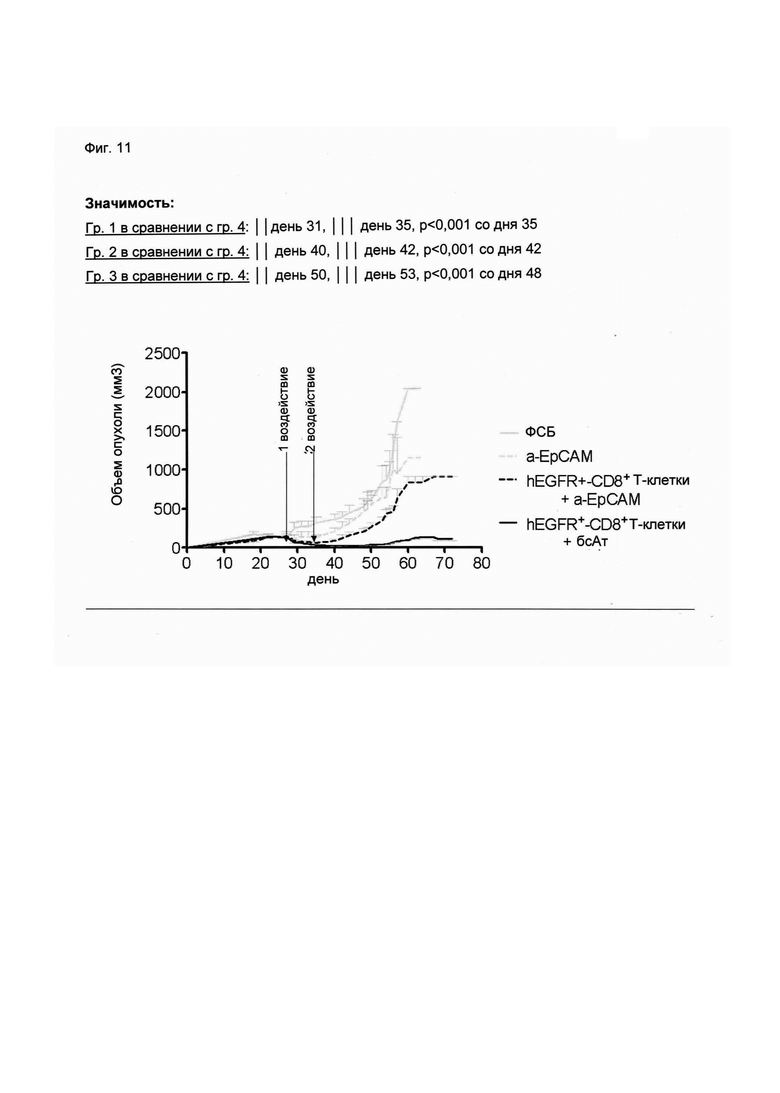
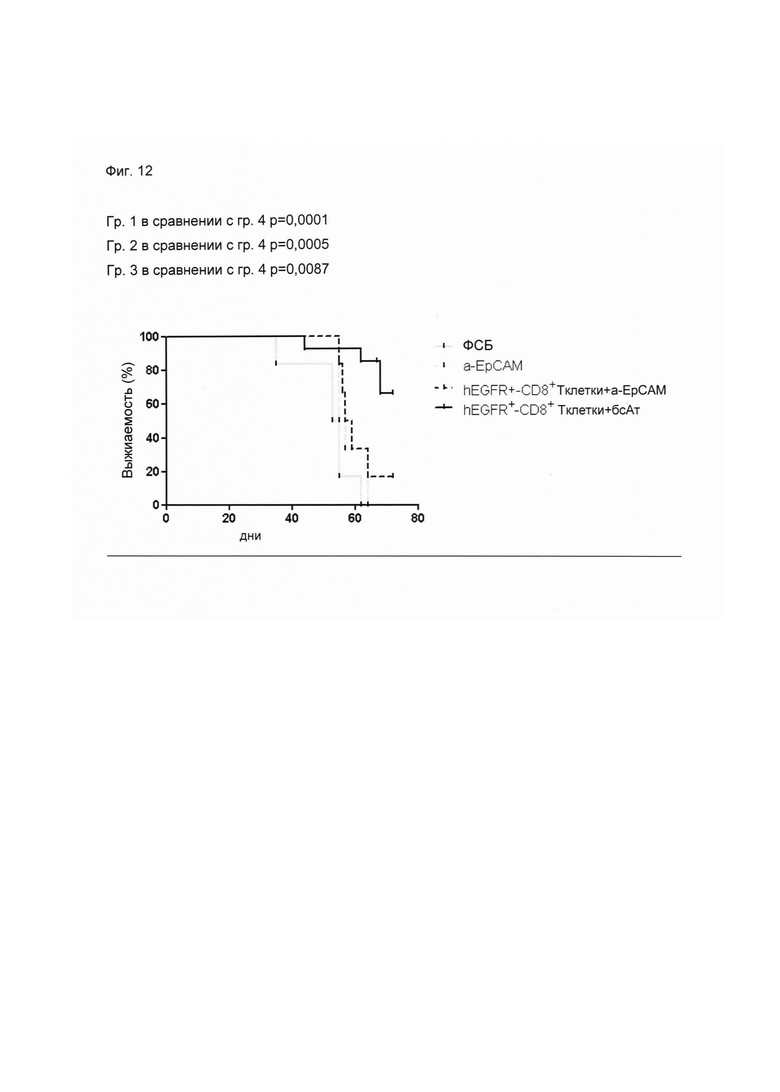
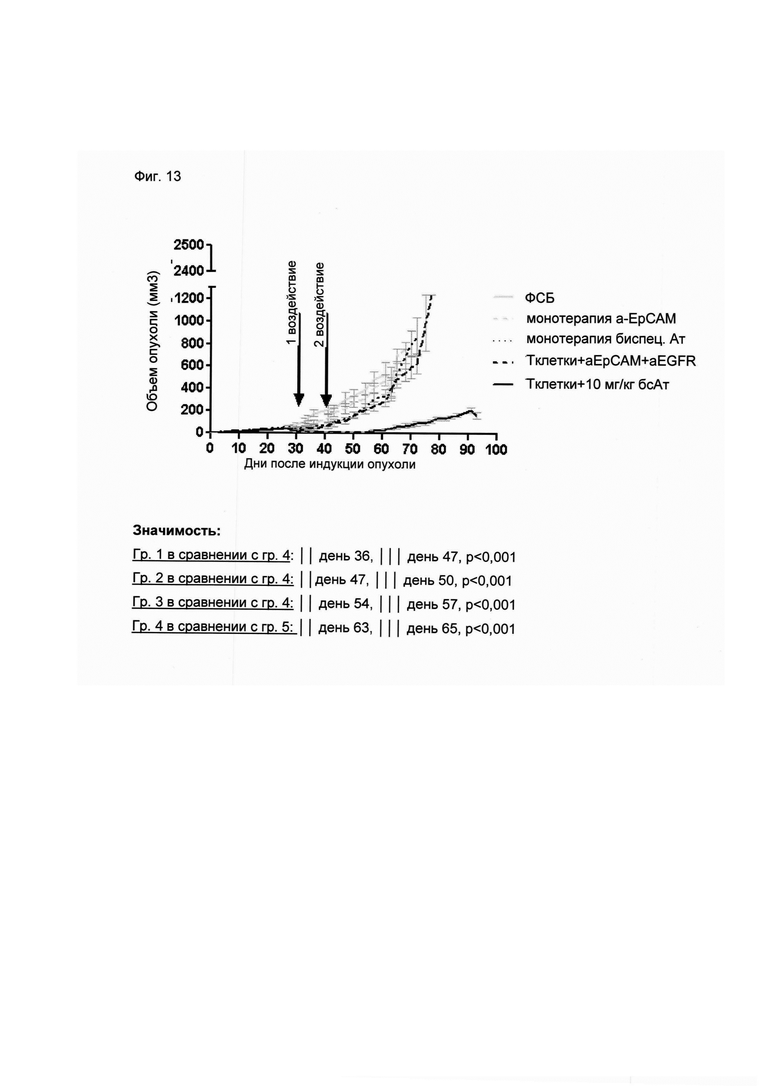
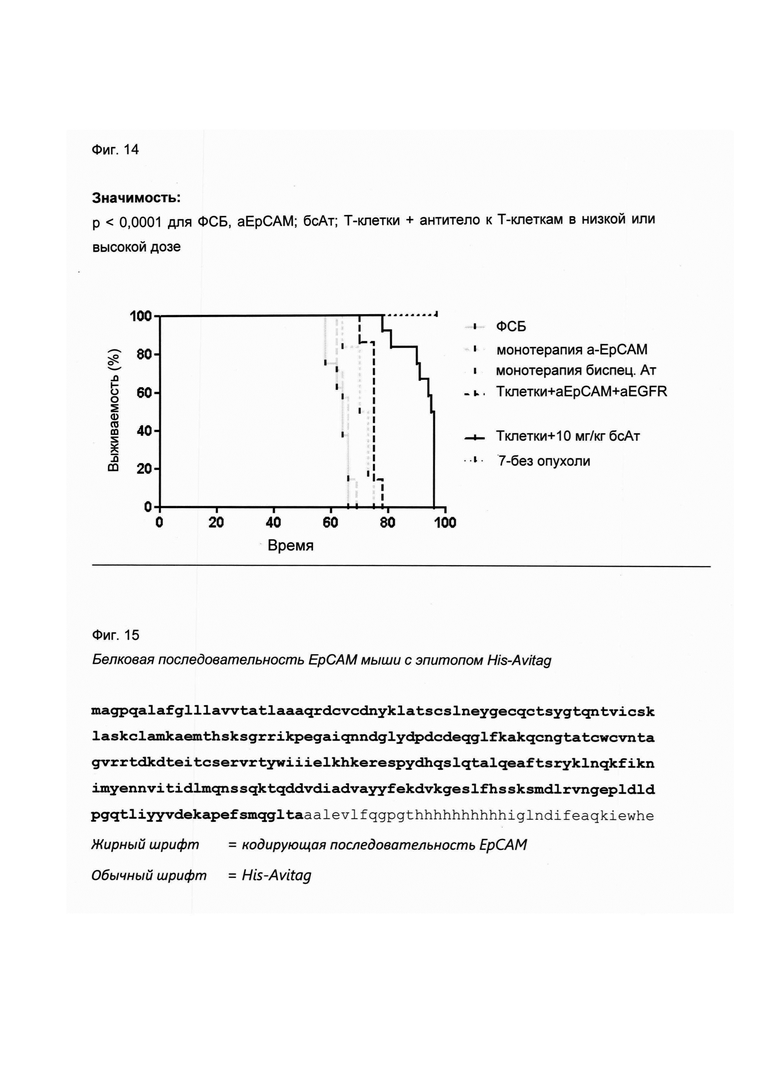

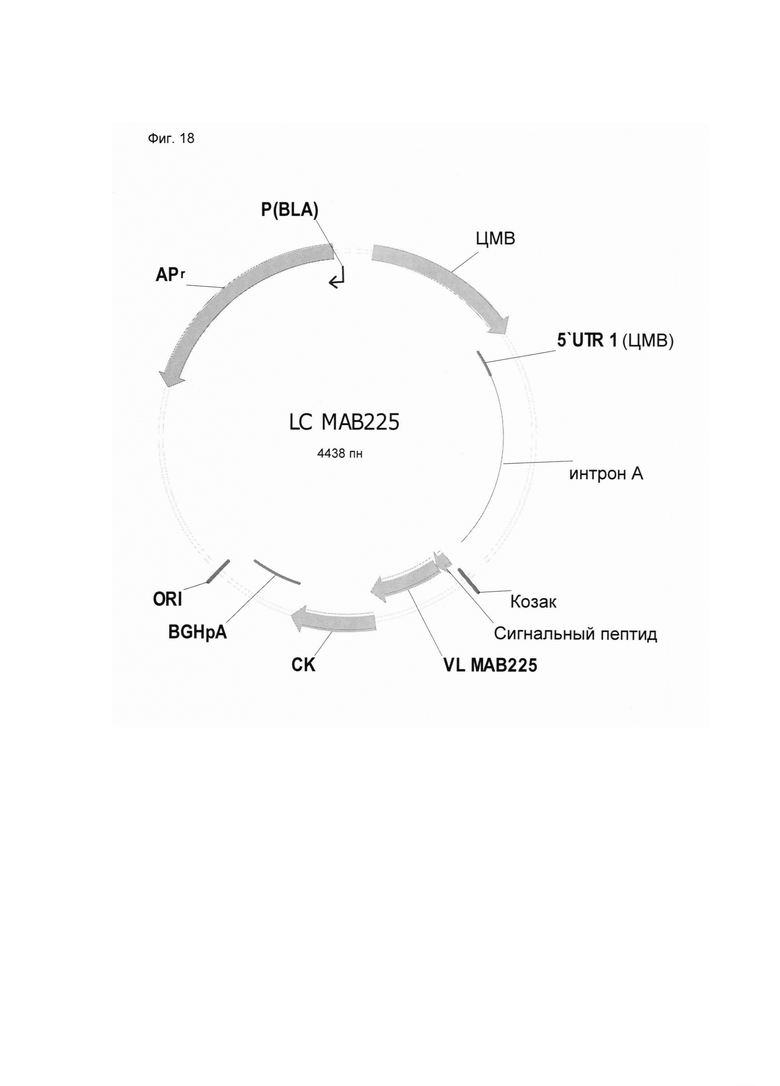
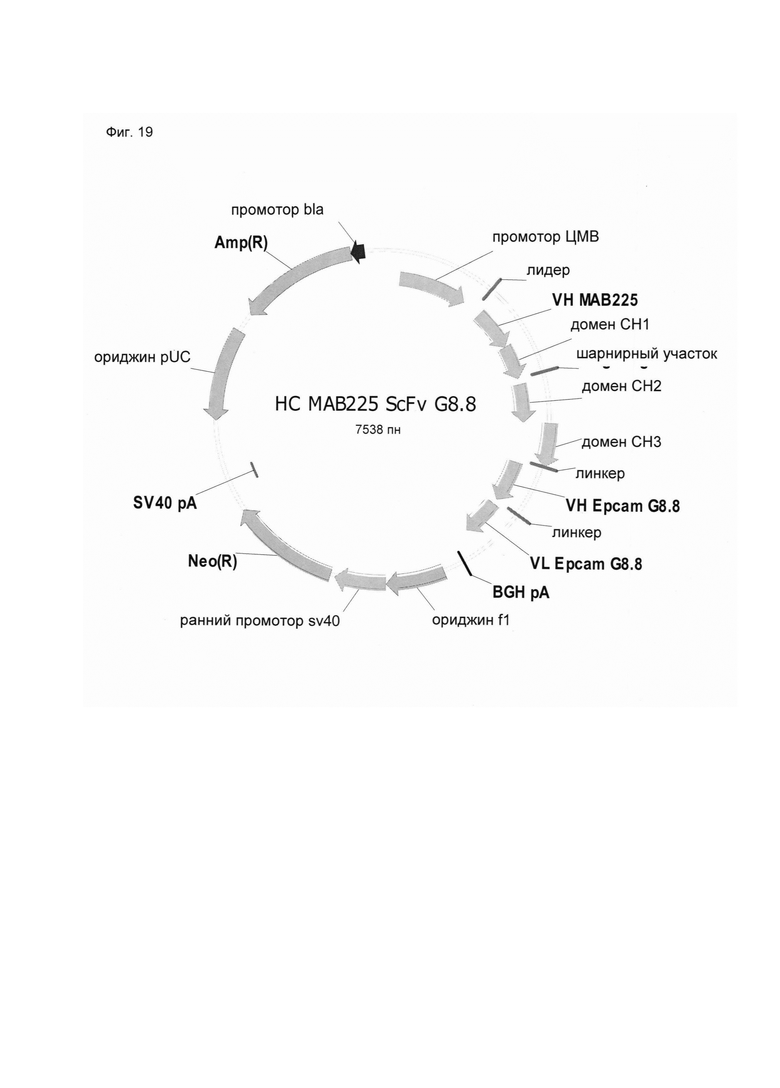
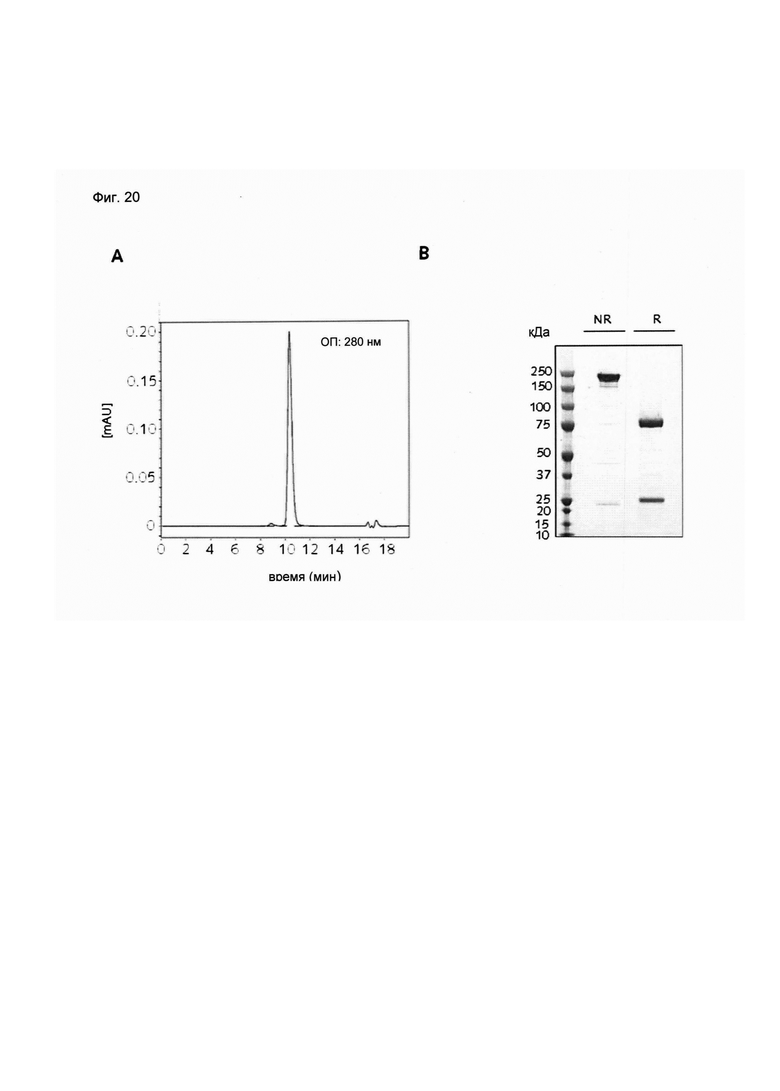
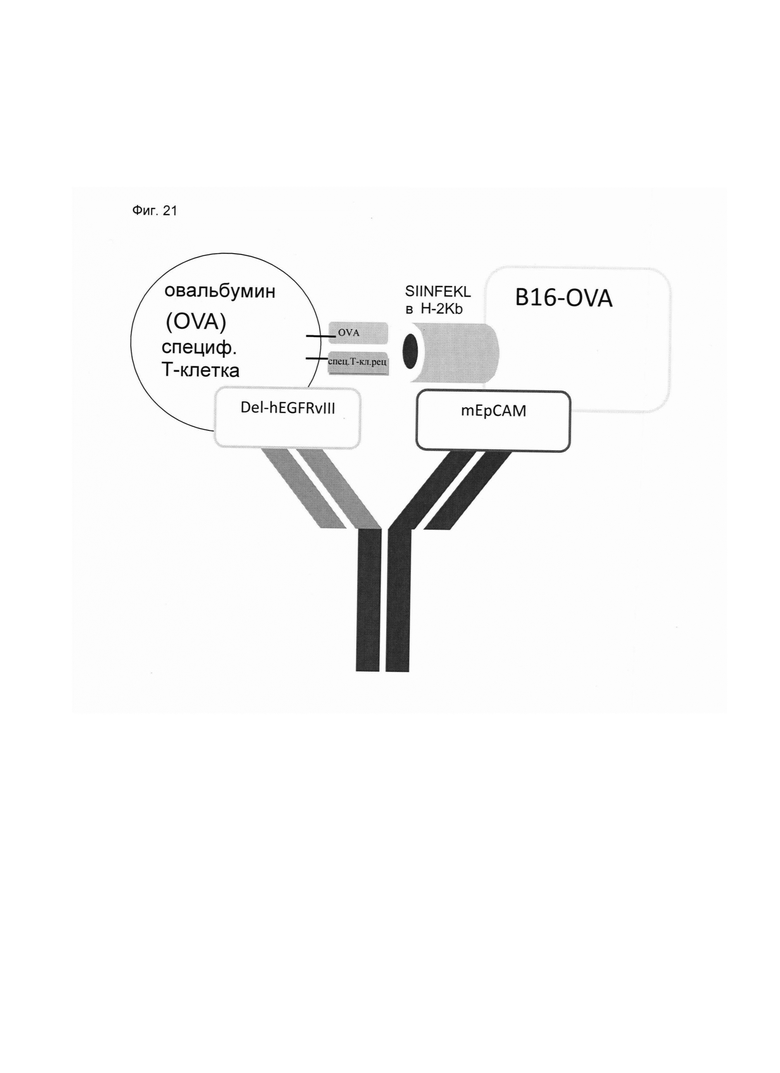
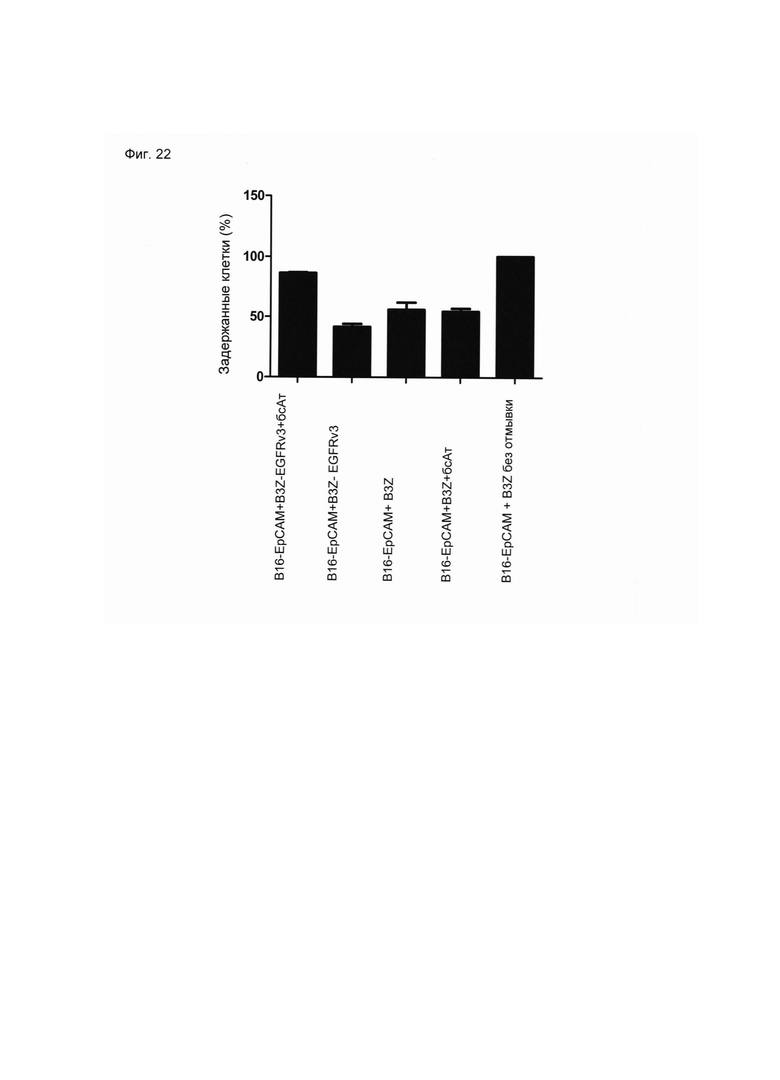


Данное изобретение относится к биотехнологии. Предложен набор, содержащий молекулу биспецифического антитела и материал, необходимый для генетической трансдукции CD8+Т-клеток, причем молекула биспецифического антитела содержит домен, связывающий антиген, трансдуцированный в CD8+Т-клетки, и домен, связывающий опухолеспецифический антиген. Также рассмотрено применение фармацевтической композиции, содержащей указанную молекулу биспецифического антитела, в комбинации с генетически трансдуцированными CD8+Т-клетками для лечения онкологического заболевания и способы лечения онкологического заболевания. Настоящее изобретение обеспечивает направленную мобилизацию CD8+Т-клеток в опухоль с помощью молекул биспецифических антител, что может найти применение в адоптивной терапии. 4 н. и 25 з.п. ф-лы, 24 ил., 1 табл., 5 пр.
1. Набор для лечения онкологического заболевания, содержащий
(A) молекулу биспецифического антитела, которая содержит
(i) первый связывающий домен, связывающий антиген, трансдуцированный в CD8+Т-клетки, где указанный антиген отсутствует в естественных условиях в или на CD8+Т-клетках; и
(ii) второй связывающий домен, связывающий опухолеспецифический антиген, существующий в естественных условиях на поверхности опухолевой клетки; и
(B) материал, необходимый для генетической трансдукции CD8+Т-клеток, полученных у субъекта, подлежащего лечению, указанным презентируемым на поверхности Т-клеток антигеном, отсутствующим в естественных условиях в или на указанных CD8+Т-клетках, где указанный материал для генетической трансдукции представляет собой экспрессионный вектор, содержащий нуклеотидную последовательность, кодирующую указанный антиген.
2. Набор по п. 1, где указанная молекула биспецифического антитела выбрана из группы, состоящей из полноразмерного антитела, химерного антитела, антитела с пересаженными CDR, полностью человеческого антитела, конструкции бивалентного антитела, гибридного антитела, синтетического антитела, двухвалентного антитела, трехвалентного антитела, четырехвалентного антитела, двухвалентного одноцепочечного антитела, трехвалентного одноцепочечного антитела и мультивалентного одноцепочечного антитела.
3. Набор по п. 1, где указанный антиген, отсутствующий в естественных условиях в или на CD8+Т-клетках, выбран из группы, состоящей из белка Cripto (cryptic family protein), представителей семейства (не Т-клеточных) молекул CD (кластера дифференцировки), EGFR и TSH-R.
4. Набор по п. 1, где указанный антиген, существующий в естественных условиях на поверхности опухолевых клеток, выбран из группы, состоящей из ЕрСАМ, HER-1, HER-2, HER-3, CD20, CD22, CD33, CD52, СА-12-5, HLA-DR, MUC-1 (муцина), А33-антигена, PSMA (простат-специфического мембранного антигена), рецептора трансферрина, тенасцина и CA-IX.
5. Набор по п. 1, где указанный первый домен и/или второй связывающий домен является человеческим и/или гуманизированным.
6. Набор по п. 1, где генетически трансдуцированные CD8+Т-клетки дополнительно содержат Т-клеточный рецептор, существующий в естественных условиях на указанных Т-клетках, и/или Т-клеточный рецептор, который был введен в указанные Т-клетки при помощи генной инженерии.
7. Набор по п. 1, где молекула биспецифического антитела кодируется последовательностью нуклеиновой кислоты.
8. Применение фармацевтической композиции для лечения онкологического заболевания, содержащей молекулу биспецифического антитела в эффективном количестве, которая содержит
(i) первый связывающий домен, связывающий антиген, трансдуцированный в CD8+Т-клетки, где указанный антиген отсутствует в естественных условиях в или на CD8+Т-клетках; и
(ii) второй связывающий домен, связывающий опухолеспецифический антиген, существующий в естественных условиях на поверхности опухолевой клетки,
в комбинации с генетически трансдуцированными CD8+Т-клетками, содержащими презентированный на поверхности Т-клеток антиген, отсутствующий в естественных условиях в или на CD8+Т-клетках, причем указанную молекулу биспецифического антитела следует вводить перед, одновременно или после введения трансдуцированных CD8+Т-клеток и причем указанные CD8+Т-клетки получены у субъекта, подлежащего лечению.
9. Применение по п. 8, где указанная молекула биспецифического антитела выбрана из группы, состоящей из полноразмерного антитела, химерного антитела, антитела с пересаженными CDR, полностью человеческого антитела, конструкции двухвалентного антитела, гибридного антитела, синтетического антитела, двухвалентного антитела, трехвалентного антитела, четырехвалентного антитела, двухвалентного одноцепочечного антитела, трехвалентного одноцепочечного антитела и мультивалентного одноцепочечного антитела.
10. Применение по п. 8, где указанный антиген, отсутствующий в естественных условиях в или на CD8+Т-клетках, выбран из группы, состоящей из белка Cripto (cryptic family protein), представителей семейства (не Т-клеточных) молекул CD (кластера дифференцировки), EGFR и TSH-R.
11. Применение по п. 8, где указанный антиген, существующий в естественных условиях на поверхности опухолевых клеток, выбран из группы, состоящей из ЕрСАМ, HER-1, HER-2, HER-3, CD20, CD22, CD33, CD52, СА-12-5, HLA-DR, MUC-1 (муцина), А33-антигена, PSMA (простат-специфического мембранного антигена), рецептора трансферрина, тенасцина и CA-IX.
12. Применение по п. 8, где указанный первый домен и/или второй связывающий домен является человеческим и/или гуманизированным.
13. Применение по п. 8, где генетически трансдуцированные CD8+Т-клетки дополнительно содержат Т-клеточный рецептор, существующий в естественных условиях на указанных Т-клетках, и/или Т-клеточный рецептор, который был введен в указанные Т-клетки при помощи генной инженерии.
14. Применение по п. 8, где молекула биспецифического антитела кодируется последовательностью нуклеиновой кислоты.
15. Способ лечения онкологического заболевания, включающий введение нуждающемуся в этом субъекту молекулы биспецифического антитела, которая содержит
(i) первый связывающий домен, связывающий антиген, трансдуцированный в CD8+Т-клетки, где указанный антиген отсутствует в естественных условиях в или на CD8+Т-клетках; и
(ii) второй связывающий домен, связывающий опухолеспецифический антиген, существующий в естественных условиях на поверхности опухолевой клетки,
и генетически трансдуцированных CD8+Т-клеток, полученных у указанного субъекта и содержащих презентируемый на поверхности Т-клеток антиген, который отсутствует в естественных условиях в или на CD8+Т-клетках, где указанную молекулу биспецифического антитела следует вводить перед, одновременно или после введения указанных CD8+Т-клеток.
16. Способ по п. 15, в котором указанное онкологическое заболевание выбрано из злокачественного новообразования эпителиального, эндотелиального или мезотелиального происхождения и злокачественного новообразования кроветворной ткани.
17. Способ по п. 15, где указанная молекула биспецифического антитела выбрана из группы, состоящей из полноразмерного антитела, химерного антитела, антитела с пересаженными CDR, полностью человеческого антитела, конструкции двухвалентного антитела, гибридного антитела, синтетического антитела, двухвалентного антитела, трехвалентного антитела, четырехвалентного антитела, двухвалентного одноцепочечного антитела, трехвалентного одноцепочечного антитела и мультивалентного одноцепочечного антитела.
18. Способ по п. 15, где указанный антиген, отсутствующий в естественных условиях в или на CD8+Т-клетках, выбран из группы, состоящей из белка Cripto (cryptic family protein), представителей семейства (не Т-клеточных) молекул CD (кластера дифференцировки), EGFR и TSH-R.
19. Способ по п. 15, где указанный антиген, существующий в естественных условиях на поверхности опухолевых клеток, выбран из группы, состоящей из ЕрСАМ, HER-1, HER-2, HER-3, CD20, CD22, CD33, CD52, СА-12-5, HLA-DR, MUC-1 (муцина), А33-антигена, PSMA (простат-специфического мембранного антигена), рецептора трансферрина, тенасцина и CA-IX.
20. Способ по п. 15, где указанный первый домен и/или второй связывающий домен является человеческим и/или гуманизированным.
21. Способ по п. 15, где генетически трансдуцированные CD8+Т-клетки дополнительно содержат Т-клеточный рецептор, существующий в естественных условиях на указанных Т-клетках, и/или Т-клеточный рецептор, который был введен в указанные Т-клетки при помощи генной инженерии.
22. Способ по п. 15, где молекула биспецифического антитела кодируется последовательностью нуклеиновой кислоты.
23. Способ лечения онкологического заболевания у субъекта, включающий этапы
(a) выделения у субъекта CD8+Т-клеток;
(b) трансдукции указанных выделенных CD8+Т-клеток антигеном, отсутствующим в естественных условиях в и/или на CD8+Т-клетках, согласно описанию в любом из пп. 1-5;
(c) введения указанных генетически трансдуцированных CD8+Т-клеток указанному субъекту; и
(d) введения указанному субъекту молекулы биспецифического антитела, описанной в любом из пп. 1-5.
24. Способ по п. 23, в котором указанные генетически трансдуцированные CD8+Т-клетки вводят указанному субъекту путем внутривенной инфузии.
25. Способ по п. 23 или 24, в котором осуществляют ко-трансдукцию указанных генетически трансдуцированных CD8+Т-клеток Т-клеточным рецептором.
26. Способ по любому из пп. 23, 24, в котором размножение указанных генетически трансдуцированных CD8+T-клеток осуществляют с использованием анти-СD3 и анти-СD28 антител.
27. Способ по п. 26, в котором размножение генетически трансдуцированных CD8+Т-клеток осуществляют в присутствии цитокинов, предпочтительно интерлейкина-2 (IL-2) и/или интерлейкина-15 (IL-15).
28. Способ лечения заболевания по любому из пп. 23, 24, в котором указанное биспецифическое антитело вводят перед, одновременно или после введения генетически трансдуцированных CD8+Т-клеток.
29. Способ лечения по п. 23 или 24, в котором указанное онкологическое заболевание выбрано из злокачественного новообразования эпителиального, эндотелиального или мезотелиального происхождения или злокачественного новообразования кроветворной ткани.
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| LAMERS C.H | |||
| et al., "Protocol for gene transduction and expansion of human T lymphocytes for clinical immunogene therapy of cancer", Cancer Gene Ther., 2002; 9(7):613-23 | |||
| GLORIUS PIA et al | |||
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| MACK MATTHIAS et al | |||
| Автоматический огнетушитель | 0 |
|
SU92A1 |

