ОБЛАСТЬ ИЗОБРЕТЕНИЯ
В целом, настоящее изобретение относится к вакцинной композиции, содержащей фрагменты связанного с раком белка Her2/neu, способам изготовления такой композиции и ее применениям для предупреждения или лечения рака, характеризующегося экспрессией или сверхэкспрессией белка Her2/neu.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Опухолевый антиген Her2/neu, кодируемый протоонкогеном erbB2/neu, представляет собой белок массой 185 кДа, принадлежащий к семейству человеческого рецептора эпидермального фактора роста. Он состоит из богатого цистеином внеклеточного домена (extracellular domain, ECD; аминокислоты 23-652) с несколькими сайтами гликозилирования, гидрофобного трансмембранного домена (аминокислоты 653-675) и внутриклеточного тирозинкиназного домена (аминокислоты 676-1255). Рецептор Her2/neu экспрессируется на клеточной мембране множества типов эпителиальных клеток и регулирует аспекты роста и деления клеток посредством связывания с определенными факторами роста.
Her2/neu экспрессируется на низких уровнях во многих нормальных клетках, но сверхэкспрессирован при многих видах рака, включая рак молочной железы, яичника, эндометрия, желудка, поджелудочной железы, предстательной железы и слюнной железы. Например, показано, что при метастатическом раке молочной железы Her2/neu сверхэкспрессирован приблизительно в 30% случаев. Эта сверхэкспресеия связана с неблагоприятным прогнозом для пациента с раком молочной железы, поскольку соответствует сокращению безрецидивных периодов и времени выживания. В настоящее время наиболее распространенные формы лечения рака молочной железы включают хирургическое вмешательство, химиотерапию и/или лучевую терапию. Если рак не ограничен определенной областью, хирургическое вмешательство само по себе не позволяет устранить рак. Более того, лучевая терапия, а также химиотерапия могут оказывать тяжелые негативные побочные эффекты.
Ввиду недостатков, связанных с лечением, применяемым в настоящее время, были предприняты попытки найти дополнительные способы лечения пролиферативных расстройств, таких как рак молочной железы, включая иммунотерапию.
Клинические последствия сверхэкспрессии Her2/neu при опухолях сделали Her2/neu привлекательной мишенью для антитело-опосредованной иммунотерапии, самой по себе или дополнительно к обычной химиотерапии. Например, было показано, что моноклональное антитело (monoclonal antibody, mAb) 4D5 снижает рост опухолей, экспрессирующих Her2/neu, у мышей посредством как прямых, так и непрямых механизмов, таких как апоптоз, антитело-зависимая клеточно-опосредованная цитотоксичность (antibody-dependent cell-mediated cytotoxicity, ADCC) или комплемент-зависимая цитотоксичность (complement-dependent cytotoxicity, CDC). На основании этих результатов, в клинических исследованиях была проанализирована гуманизированная форма этого антитела, трастузумаб (Herceptin®). Общая выживаемость у пациентов с опухолями молочной железы, сверхэкспрессирующими Her2/neu, после цитотоксического лечения с Herceptin® была повышена по сравнению с химиотерапией или трастузумабом самими по себе. В настоящее время Herceptin® применяют в качестве монотерапии, но он демонстрирует еще более высокую эффективность в комбинации с цитотоксической химиотерапией. Тем не менее, следует отметить, что трастузумаб обычно эффективен только при раке молочной железы со сверхэкспрессией рецептора Her2/neu. Кроме того, обычно необходимы многократные инфузии, что повышает стоимость лечения.
Альтернативный способ лечения или предупреждения видов рака, связанных с Her2/neu, с применением пассивной иммунотерапии моноклональными антителами, такими как трастузумаб, основан на индукции опухоль-специфичных гуморальных и/или клеточных иммунных ответов и идентификации антигенов, распознаваемых человеческими В- и Т-лимфоцитами. Например, иммунизацией мышей клетками, экспрессирующими Her2/neu, было получено множество антител, направленных против внеклеточного домена (ECD) Her2/neu. По-видимому, биологический эффект этих антител эпитоп-специфичен; то есть, он основан на специфичном распознавании короткой подпоследовательности в ECD Her2/neu. Тем не менее, некоторые антитела не оказывают эффекта или даже активно стимулируют рост опухоли.
Такая вакцинная иммунотерапия рака основана на антигенах, против которых направлены гуморальные и/или клеточные ответы. В идеальном случае эти антигены должны быть экспрессированы или сверхэкспрессированы исключительно в опухолевых клетках, и их часто называют опухоль-ассоциированными антигенами (tumour-associated antigens, ТАА). Одним из первых ТАА, описанных для рака молочной железы, был Her2/neu. В то же время, было изучено множество других ТАА, представляющих различные эпитопы, но до настоящего времени ни один из них не был успешно применен в клинической практике.
Обычно в зависимости от предполагаемого типа иммунного ответа (то есть, В-клеточный или Т-клеточный ответ) применяют различные способы. Например, для индукции В-клеточного (то есть гуморального) ответа антигены должны содержать В-клеточный эпитоп.Как общеизвестно в данной области, В-клеточный эпитоп является частью антигена, распознаваемого и связываемого В-клеточным рецептором. Липиды, полисахариды и белки/пептиды могут содержать В-клеточные эпитопы, которые, при их введении в выбранный организм, приводят к выработке В-клетками антител, специфично связывающихся с введенным эпитопом.
В данной области известны отдельные фрагменты ECD Her2/neu, включая В-клеточные эпитопы. Например, в WO 2002/068474 описана вакцина, содержащая пептид из 9-25 аминокислот, последовательность которых присутствует во внеклеточной части белка Her2/neu. Кроме того, в WO 2007/118660 описана мультипептидная вакцина, содержащая определенную комбинацию пептидов, представляющих различные аминокислотные последовательности, присутствующие во внеклеточной части белка Her2/neu. Эти пептиды могут быть введены по отдельности или в комбинации в форме нескольких отдельных пептидов, каждый из которых предпочтительно отдельно конъюгирован с системой доставки. В еще одном примере в WO 2011/020604 описаны слитые пептиды, содержащие несколько В-клеточных эпитопов Her2/neu, в сочетании с виросомной системой доставки. Было показано, что эти виросомы обеспечивают более высокий титр антител против одного В-клеточного эпитопа по сравнению с теми же слитыми пептидами в сочетании с Montanide™ или системой доставки на основе иммуностимулирующих комплексов (ISCOM).
Несмотря на ряд попыток разработать подходящую вакцину для индукции иммунитета против Her2/neu, до настоящего времени не существует эффективной вакцины для клинического применения. Задачей настоящего изобретения является обеспечение улучшенной композиции, подходящей для применения в качестве вакцины для лечения или предупреждения состояния, такого как рак, характеризующегося экспрессией или сверхэкспрессией Her2/neu.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В настоящем описании описана вакцинная композиция, содержащая: адъювант; и
по меньшей мере один слитый пептид, конъюгированный с белком-носителем,
где белок-носитель представляет собой вариант дифтерийного токсина CRM197 (регистрационный номер в GenBank 1007216А; SEQ ID NO: 61) и где по меньшей мере один слитый пептид содержит два или более несмежных В-клеточных эпитопа Her2/neu, выбранных из группы, состоящей из SEQ ID NO: 1-7 и 15-60 и аминокислотных последовательностей, по меньшей мере на 85% идентичных любой из последовательностей, указанных выше.
В настоящем описании также описана фармацевтическая композиция, содержащая вакцинную композицию, как описано здесь, и фармацевтически приемлемый носитель.
В настоящем описании также описан способ лечения или предупреждения рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu, у пациента, нуждающегося в этом, включающий стадию введения указанному пациенту эффективного количества вакцинной композиции, как описано здесь, или фармацевтической композиции, как описано здесь.
В настоящем описании также описано применение вакцинной композиции, как описано здесь, в изготовлении лекарственного средства для лечения или предупреждения рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu, у пациента, нуждающегося в этом.
В настоящем описании также раскрыта вакцинная композиция, как описано здесь, или фармацевтическая композиция, как описано здесь, для применения в лечении или предупреждении рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu, у пациента, нуждающегося в этом.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На Фиг. 1 показаны титры антител против Р4 в образцах сыворотки, полученных после четвертой иммунизации виросомами, содержащими слитые пептиды Р467 или Р647, или конъюгатами в форме слитых белков с CRM197 (CRM). Данные представлены как значения оптической плотности (OD).
На Фиг. 2 показаны титры антител против Р6 в образцах сыворотки, полученных после четвертой иммунизации виросомами, содержащими слитые пептиды Р467 или Р647, или конъюгатами в форме слитых белков с CRM197 (CRM). Данные представлены как значения OD.
На Фиг. 3 показаны титры антител против Р7 в образцах сыворотки, полученных после четвертой иммунизации виросомами, содержащими слитые пептиды Р467 или Р647, или конъюгатами в форме слитых белков с CRM197 (CRM). Данные представлены как значения OD.
На Фиг. 4 показаны титры антител против Р467 в образцах сыворотки, полученных после четвертой иммунизации виросомами, содержащими слитые пептиды Р467 или Р647, или конъюгатами в форме слитых белков с CRM197 (CRM). Данные представлены как значения OD.
На Фиг. 5 показаны титры антител против Р647 в образцах сыворотки, полученных после четвертой иммунизации виросомами, содержащими слитые пептиды Р467 или Р647, или конъюгатами в форме слитых белков с CRM197 (CRM). Данные представлены как значения OD.
На Фиг. 6 показаны титры антител против Р467 в образцах сыворотки, полученных через две недели после второй (ВА1; 1-й забор крови), третьей (ВА2; 2-й забор крови) и четвертой (ВА3; 3-й забор крови) иммунизации виросомами, содержащими слитые пептиды Р467 или Р647, или конъюгатами в форме слитых белков с CRM 197 (CRM). Данные представлены как значения OD.
На Фиг. 7 показаны титры антител против Р647 в образцах сыворотки, полученных через две недели после второй (ВА1; 1-й забор крови), третьей (ВА2; 2-й забор крови) и четвертой (ВА3; 3-й забор крови) иммунизации виросомами, содержащими слитые пептиды Р467 или Р647, или конъюгатами в форме слитых белков с CRM197 (CRM). Данные представлены как значения OD.
На Фиг. 8 показано, что антитела из образцов сыворотки, полученных через две недели после второй (ВА1; 1-й забор крови) и третьей (ВА2; 2-й забор крови) иммунизации виросомными композициями, содержащими слитые пептиды Р467 или Р647 (15, 30, 50 мкг), или конъюгатами в форме слитых белков с CRM197 (CRM; 30 мкг), способны связываться с рекомбинантным ECD Her2/neu. Данные представлены как значения OD.
На Фиг. 9 показаны антитела, специфичные в отношении рекомбинантного Her2/neu, из образцов сыворотки, полученных через две недели после второй (ВА1; 1-й забор крови) иммунизации виросомными композициями, содержащими слитые пептиды Р467 или Р647 (15, 30, 50 мкг), или конъюгатами в форме слитых белков с CRM197 (CRM; 30 мкг). Данные представлены как значения OD.
На Фиг. 10 показаны антитела, специфичные в отношении рекомбинантного Her2/neu, из образцов сыворотки, полученных через две недели после четвертой (ВА3; 3-й забор крови) иммунизации виросомными композициями, содержащими слитые пептиды Р467 или Р647 (15, 30, 50 мкг), или конъюгатами в форме слитых белков с CRM197 (CRM; 30 мкг). Данные представлены как значения OD.
На Фиг. 11 показана динамика титров антител, специфичных в отношении рекомбинантного Her2/neu, после второй, третьей и четвертой иммунизаций конъюгатами в форме слитых белков с CRM 197 (CRM; 30 мкг), содержащими слитые пептиды Р467 или Р647. Сыворотку анализировали в разведении 1:4000. Данные представлены как значения OD.
На Фиг. 12 показано прямое сравнение титров антител после 2-й, 3-й и 4-й иммунизаций слитым пептидом Р467, конъюгированным с виросомами или с CRM 197 в концентрации 30 мкг.Слева направо в каждой группе: предварительный пул (кровь, полученная до иммунизации); TIRIV (пустые виросомы); Р467 - 30 мг на мышь (конъюгированный с виросомами); Р467 -30 мг на мышь с CRM (конъюгированный с CRM197). Все образцы сыворотки анализировали в одном и том же разведении. Данные представлены как значения OD.
На Фиг. 13 показано прямое сравнение титров антител после 2-й, 3-й и 4-й иммунизации слитым пептидом Р647, конъюгированным с виросомами или с CRM 197 в концентрации 30 мкг. Слева направо в каждой группе: предварительный пул (кровь, полученная до иммунизации); TIRIV (пустые виросомы); Р647 - 30 мг на мышь (конъюгированный с виросомами); Р647 - 30 мг на мышь с CRM (конъюгированный с CRM197). Все образцы сыворотки анализировали в одном и том же разведении. Данные представлены как значения OD.
На Фиг. 14 показано, что иммунизация виросомами, содержащими слитые пептиды Р467 или Р647, или конъюгатами в форме слитых белков с CRM197 (CRM) приводит к выработке антител, способных связываться с нативным Her2/neu, экспрессированным на поверхности клеток рака молочной железы SKBR-3. Данные представлены как значения OD.
На Фиг. 15А показаны титры антител IgG против Р467 у мышей после трех введений слитого белка P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили сам по себе (P467-CRM), с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Титры антител сравнивали с уровнями, отмеченными у контрольных животных, которым вводили пептид Р467 сам по себе (Р467; 25 мкг), Alum от Brenntag (Alum (Brenntag)) или CRM197 сам по себе (CRM). Данные представлены как значения OD; *р<0,05, **р<0,01 и ***р<0,001.
На Фиг. 15В показаны титры антител IgG1 против Р467 у мышей после трех введений слитого белка P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили сам по себе (P467-CRM), с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Титры антител сравнивали с уровнями, отмеченными у контрольных животных, которым вводили пептид Р467 сам по себе. Данные представлены как значения OD; *р<0,05, **р<0,01 и ***р<0,001.
На Фиг. 15С показаны титры антител IgG2a против Р467 у мышей после трех введений слитого белка P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили сам по себе (P467-CRM), с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Титры антител сравнивали с уровнями, отмеченными у контрольных животных, которым вводили пептид Р467 сам по себе (Р467; 25 мкг), Alum от Brenntag (Alum (Brenntag)) или CRM197 сам по себе (CRM). Данные представлены как значения OD; *р<0,05, **р<0,01 и ***р<0,001.
На Фиг. 16А показаны титры антител IgG против внеклеточного Her2/neu у мышей после трех введений слитого белка P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили сам по себе (P467-CRM), с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Титры антител сравнивали с уровнями, отмеченными у контрольных животных, которым вводили пептид Р467 сам по себе (Р467; 25 мкг), Alum от Brenntag (Alum (Brenntag)) или CRM197 сам по себе (CRM). Данные представлены как значения OD; *р<0,05, **р<0,01 и ***р<0,001.
На Фиг. 16В показаны титры антител IgG1 против внеклеточного Her2/neu у мышей после трех введений слитого белка P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили сам по себе (P467-CRM), с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Титры антител сравнивали с уровнями, отмеченными у контрольных животных, которым вводили пептид Р467 сам по себе (Р467; 25 мкг), Alum от Brenntag (Alum (Brenntag)) или CRM 197 сам по себе (CRM). Данные представлены как значения OD; *р<0,05, **р<0,01 и ***р<0,001.
На Фиг. 17А показано образование IL-2 CRM197-активированными спленоцитами, имеющими происхождение от мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили сам по себе (P467-CRM), с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Уровни IL-2 (пг/мл) сравнивали с уровнями IL-2, отмеченными у CRM197-активированных спленоцитов, имеющих происхождение от контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM197 сам по себе (CRM). Данные представлены в пг/мл; **р<0,01; ns - статистически незначимо.
На Фиг. 17В показано образование IL-2 Р467-активированными спленоцитами, имеющими происхождение от мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили сам по себе (P467-CRM), с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Уровни IL-2 (пг/мл) сравнивали с уровнями IL-2, отмеченными у Р467-активированных спленоцитов, имеющих происхождение от контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM197 сам по себе (CRM). Данные представлены в пг/мл; *р<0,05; **р<0,01; ns - статистически незначимо.
На Фиг. 18А показано образование IFNγ CRM197-активированными спленоцитами, имеющими происхождение от мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили сам по себе (P467-CRM), с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Уровни IFNγ (пг/мл) сравнивали с уровнями IFNγ, отмеченными у CRM197-aKTHBnpoBaHHbix спленоцитов, имеющих происхождение от контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM197 сам по себе (CRM). Данные представлены в пг/мл; *р<0,05; ns - статистически незначимо.
На Фиг. 18В показано образование IFNγ Р467-активированными спленоцитами, имеющими происхождение от мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили сам по себе (P467-CRM), с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Уровни IFNγ (пг/мл) сравнивали с уровнями IFNγ, отмеченными у Р467-активированных спленоцитов, имеющих происхождение от контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM197 сам по себе (CRM). Данные представлены в пг/мл; ns - статистически незначимо.
На Фиг. 19А показано образование IL-5 СКМ197-активированными спленоцитами, имеющими происхождение от мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили сам по себе (P467-CRM), с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Уровни IL-5 (пг/мл) сравнивали с уровнями IL-5, отмеченными у CRM197-активированных спленоцитов, имеющих происхождение от контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM 197 сам по себе (CRM). Данные представлены в пг/мл; *р<0,05; **р<0,01; ns - статистически незначимо.
На Фиг. 19В показано образование IL-5 Р467-активированными спленоцитами, имеющими происхождение от мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили сам по себе (P467-CRM), с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Уровни IL-5 (пг/мл) сравнивали с уровнями IL-5, отмеченными у Р467-активированных спленоцитов, имеющих происхождение от контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM197 сам по себе (CRM). Данные представлены в пг/мл; ns - статистически незначимо.
На Фиг. 20А показано распределение CD3+CD4+ спленоцитов (Т-лимфоцитов) при умерщвлении мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Процент CD3+CD4+ спленоцитов сравнивали с процентом CD3+CD4+ спленоцитов у контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM 197 сам по себе (CRM). Данные представлены как процент от общего числа клеток; ns - статистически незначимо.
На Фиг. 20В показано распределение CD3+CD8+ спленоцитов (Т-лимфоцитов) при умерщвлении мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Процент CD3+CD8+ спленоцитов сравнивали с процентом CD3+CD8+ спленоцитов у контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM 197 сам по себе (CRM). Данные представлены как процент от общего числа клеток; ns - статистически незначимо.
На Фиг. 20С показано распределение CD3-CD19+ спленоцитов (В-клеток) при умерщвлении мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Процент CD3-CD19+ спленоцитов сравнивали с процентом CD3-CD19+ спленоцитов у контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM197 сам по себе (CRM). Данные представлены как процент от общего числа клеток; ns - статистически незначимо.
На Фиг. 20D показано распределение CD335 + спленоцитов (натуральных клеток-киллеров (NK-клеток)) при умерщвлении мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Процент CD335 + спленоцитов сравнивали с процентом CD335 + спленоцитов у контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM 197 сам по себе (CRM). Данные представлены как процент от общего числа клеток; *р<0.05; ns - статистически незначимо.
На Фиг. 21А показано распределение CD3+CD4+IFNγ+ спленоцитов (Т-клеток, образующих IFNγ) при умерщвлении мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Процент CD3 + CD4 + IFNγ + спленоцитов сравнивали с процентом CD3+CD4+IFNγ+ спленоцитов у контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM 197 сам по себе (CRM). Данные представлены как процент от общего числа клеток; ns - статистически незначимо.
На Фиг. 21В показано распределение CD3+CD8+IFNγ+ спленоцитов (Т-клеток, образующих IFNγ) при умерщвлении мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Процент CD3+CD8+IFNγ+ спленоцитов сравнивали с процентом CD3+CD8+IFNγ+ спленоцитов у контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM 197 сам по себе (CRM). Данные представлены как процент от общего числа клеток; *р<0.05; ns - статистически незначимо.
На Фиг. 21С показано распределение CD335+IFNγ+ спленоцитов (Т-клеток, образующих IFNγ) при умерщвлении мышей, иммунизированных слитым белком P467-CRM197 в дозах 10 мкг, 25 мкг и 50 мкг, который вводили с Alum от Brenntag (P467-CRM + Alum), с Alum от Serva (P467-CRM + Alum (Serva)) или с Montanide (P467-CRM + Montanide). Процент CD335+IFNγ+ спленоцитов сравнивали с процентом CD335+IFNγ+ спленоцитов у контрольных животных, которым вводили Alum сам по себе (Alum (Brenntag)), Montanide сам по себе (Montanide) или CRM197 сам по себе (CRM). Данные представлены как процент от общего числа клеток; *р<0.05; ns - статистически незначимо.
На Фиг. 22 показаны титры антител IgG, специфичных в отношении пептида Р467, через 3 недели (ВА2), 8 недель (ВА3), 16 недель (ВА4) и 6 месяцев (ВА5) после последнего введения пептидной конструкции P467-CRM, которую вводили в дозах 10 мкг, 25 мкг и 50 мкг с Alum (P467-CRM + Alum) или Montanide (P467-CRM + Montanide). Данные представлены как значения OD. Слева направо в каждой группе: ВА2, ВА3, ВА4, ВА5 и до забора крови.
На Фиг. 23 показаны титры антител IgG, специфичных в отношении Her2/neu, через 3 недели (ВА2), 8 недель (ВА3), 16 недель (ВА4) и 6 месяцев (ВА5) после последнего введения пептидной конструкции P467-CRM, которую вводили в дозах 10 мкг, 25 мкг и 50 мкг с Alum (P467-CRM + Alum) или Montanide (P467-CRM + Montanide). Данные представлены как значения OD. Слева направо в каждой группе: ВА2, ВА3, ВА4, ВА5 и до забора крови.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Терминология, использованная здесь, приведена лишь с целью описания определенных воплощений и не является ограничивающей. Если не определено иное, все технические и научные термины, использованные здесь, имеют то же значение, в котором их обычно понимает специалист в области, к которой относится настоящее изобретение. Для практического использования настоящего изобретения могут быть применены любые материалы и методы, сходные с описанными здесь или эквивалентные им. За определениями и терминами данной области и другими методами, известными специалисту в данной области, специалисты-практики могут обращаться к Sambrook et al., (1989) Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Press, Plainsview, N.Y., и Ausubel et al. (1999) Current Protocols in Molecular Biology (Supplement 47), John Wiley & Sons, New York, Murphy et al. (1995) Virus Taxonomy Springer Verlag: 79-87.
Если контекст не требует иного, в данном описании слово «содержать/включать» или варианты, такие как «содержит/включает» или «содержащий/включающий», следует понимать как подразумевающие включение указанного элемента, или объекта/признака, или группы элементов или объектов/признаков, но не исключение какого-либо другого элемента, или объекта/признака, или группы элементов или объектов/признаков.
Под «состоящим из» подразумевают включающий только то, что следует за фразой «состоящий из». Таким образом, фраза «состоящий из» указывает на то, что перечисленные элементы необходимы или обязательны и присутствие других элементов невозможно. Под «состоящим по существу из» подразумевают включающий любые элементы, перечисленные после этой фразы, с ограничением другими элементами, которые не препятствуют или способствуют активности или действию, указанным в описании для перечисленных элементов. Таким образом, фраза «состоящий по существу из» указывает на то, что перечисленные элементы необходимы или обязательны, но другие элементы не являются обязательными и могут присутствовать или не присутствовать в зависимости от того, влияют ли они на активность или действие перечисленных элементов.
Если контекстом не продиктовано иное, при использовании здесь формы единственного числа включают множественное число. Таким образом, например, указание «клетки» включает одну клетку, а также две или более клетки; указание «организма» включает один организм, а также два или более организма; и так далее.
Для указания нуклеотидных и аминокислотных последовательностей используют идентификационные номера последовательностей (SEQ ID NO:). SEQ ID NO: численно соответствуют идентификаторам последовательностей <400>1, <400>2 и так далее. Здесь представлена сводка по идентификаторам последовательностей.
Настоящее изобретение основано, по меньшей мере отчасти, на неожиданном обнаружении авторами изобретения того, что вакцинная композиция, содержащая (1) адъювант и (2) слитый пептид нескольких В-клеточных эпитопов, имеющих происхождение от внеклеточного домена (ECD) Her2/neu, конъюгированных с нетоксичным вариантом дифтерийного токсина CRM197 (регистрационный номер в GenBank 1007216А), может вызывать антиген-специфичный гуморальный иммунный ответ, который намного превосходит ответ, достигаемый с использованием такой системы доставки, как виросомы.
Таким образом, в одном аспекте предложена вакцинная композиция, содержащая:
адъювант; и
по меньшей мере один слитый пептид, конъюгированный с белком-носителем,
где белок-носитель представляет собой вариант дифтерийного токсина CRM197 (регистрационный номер в GenBank 1007216А; SEQ ID NO: 61) и где по меньшей мере один слитый пептид содержит два или более несмежных В-клеточных эпитопа Her2/neu, выбранных из группы, состоящей из SEQ ID NO: 1-7 и 15-60 и аминокислотных последовательностей, по меньшей мере на 85% идентичных любой из последовательностей, указанных выше.
В-клеточные эпитопы
При использовании здесь термин «В-клеточный эпитоп» относится к части молекулы, распознаваемой В-клеточным рецептором (антителом). Таким образом, «В-клеточный эпитоп» следует понимать как небольшую подпоследовательность антигена, где указанная подпоследовательность эпитопа может быть распознана антителом. Антиген может содержать множество В-клеточных эпитопов, и поэтому с ним может связываться множество различных антител, но любой заданный эпитопный фрагмент данного антигена будет обычно связываться только с одним антителом.
При использовании здесь термины «пептид» и «полипептид» использованы в их наиболее широком смысле для обозначения молекулы из двух или более аминокислотных остатков или аналогов аминокислот. Аминокислотные остатки могут быть связаны пептидными связями или, альтернативно, другими связями, например, сложноэфирными, простыми эфирными и так далее, но в большинстве случаев будут связаны пептидными связями.
При использовании здесь термины «аминокислота» или «аминокислотный остаток» включают как природные, так и искусственные или синтетические аминокислоты, включая как D-, так и L-формы, и аналоги аминокислот. «Аналог аминокислоты» следует понимать как аминокислоту, не встречающуюся в природе, которая отличается от соответствующей ей аминокислоты, встречающейся в природе, по одному или более чем одному атому. Например, аналогом аминокислоты цистеина может быть гомоцистеин.
Специалистам в данной области будет ясно, как определить, является ли пептид В-клеточным эпитопом Her2/neu. В наглядном примере можно с большой степенью точности определить, является ли пептид В-клеточным эпитопом или содержит ли он В-клеточный эпитоп, используя известные компьютерные программы, сравнивающие последовательность рассматриваемого пептида с базой данных известных последовательностей и/или частичных последовательностей, о которых известно, что их распознают антитела, кодируемые человеческой или мышиной зародышевой линией. Альтернативно, В-клеточный эпитоп данного пептида или полипептида можно определить анализом при помощи компьютера с использованием различных комбинаций коррелятов антигенности, таких как доступность поверхности, гибкость цепи, профили гидрофобности/гидрофильности, предполагаемая вторичная структура и так далее. Альтернативно, для определения того, является ли рассматриваемый пептид В-клеточным эпитопом или содержит ли он В-клеточный эпитоп, можно провести по меньшей мере одну иммунизацию животного рассматриваемым пептидом, дать развиться иммунному ответу и затем проанализировать сыворотку животного на предмет антител, специфично связывающихся по меньшей мере с частью введенного пептида с применением, например, твердофазного иммуноферментного анализа (enzyme linked immunosorbant assay, ELISA), радиоиммунного анализа, вестерн-блот-анализа или спот-блот-анализа. Более подробное описание того, как определить, является ли рассматриваемый пептид В-клеточным эпитопом, приведено ниже в Примерах 6 и 7.
В Таблице 1 ниже представлены аминокислотные последовательности В-клеточных эпитопов внеклеточного домена Her2/neu, включая их производные.

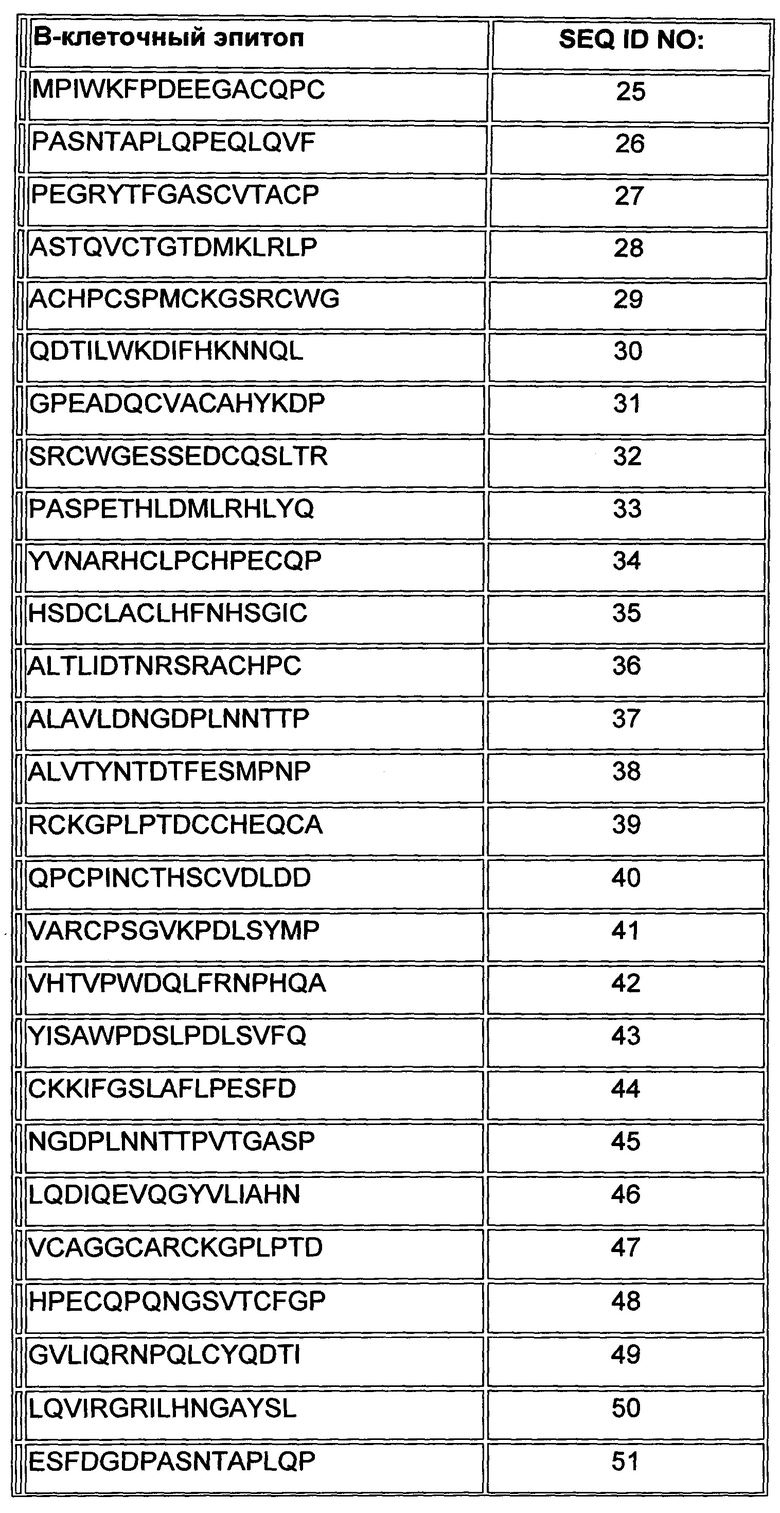
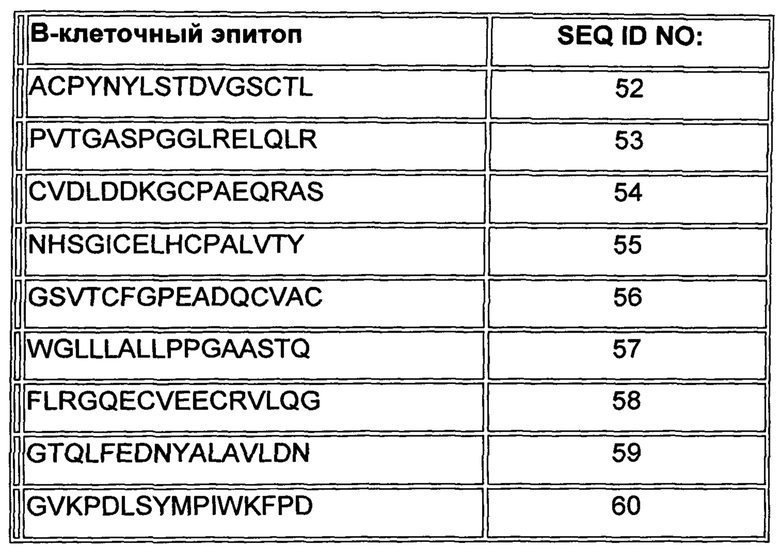
В одном воплощении, раскрытом здесь, два или более В-клеточных эпитопа выбраны из группы, состоящей из SEQ ID NO: 1-7 и 15-60, как показано в Таблицах 1 и 2, и аминокислотных последовательностей, по меньшей мере на 85% идентичных любой из последовательностей, указанных выше (то есть последовательностей, по меньшей мере на 85% идентичных любой из SEQ ID NO: 1-7 и 15-60). В другом воплощении, раскрытом здесь, два или более В-клеточных эпитопа выбраны из группы, состоящей из SEQ ID NO: 1-7 и аминокислотных последовательностей, по меньшей мере на 85% идентичных любой из последовательностей, указанных выше; то есть последовательностей, по меньшей мере на 85% идентичных любой из SEQ ID NO: 1-7. Следует отметить, что никакие из этих В-клеточных эпитопов не являются смежными в нативном Her2/neu.
Аминокислотные последовательности, по меньшей мере на 85% идентичные любой из SEQ ID NO: 1-7 и 15-60, включают производные, являющиеся результатом замены по меньшей мере одной аминокислоты в нативном (то есть встречающемся в природе) В-клеточном эпитопе ECD Her2/neu другой аминокислотой, не присутствующей в этом положении Her2/neu, таким образом, что аминокислотная замена остается консервативной. Производные могут быть получены для повышения стабильности слитого пептида в промежуточных и/или конечных продуктах, или для повышения растворимости слитого пептида в промежуточных и/или конечных продуктах, или для повышения иммуногенности слитых пептидов. Способы получения подходящих производных будут известны специалистам в данной области. Наглядные примеры включают синтез производных или их рекомбинантное получение с использованием молекулы мутантной нуклеиновой кислоты. Кроме того, производное будет обычно сохранять свои свойства В-клеточного эпитопа, как описано здесь. Таким образом, здесь также раскрыты «функциональные» производные, в которых аминокислотная замена в нативной последовательности не приводит к потере или не приводит к полной потере способности производного функционировать в качестве В-клеточного эпитопа.
Определение дополнительных или оптимизированных иммуностимулирующих слитых пептидов может также включать стадию сравнения стимуляции В-клеток слитым пептидом и стимуляции В-клеток производным как определение эффективности стимуляции иммунных эффекторных клеток производным. Сравнивая производное с известным слитым пептидом, можно получить пептиды со свойствами повышенной стимуляции иммунных клеток.
При использовании здесь «консервативная замена» относится к изменению аминокислотной идентичности в заданном положении с заменой на аминокислоту приблизительно такого же размера, заряда и/или полярности. Примеры естественных консервативных замен аминокислот включают замены в пределах следующих 8 групп (обозначения в традиционном однобуквенном коде): (1) М, I, L, V; (2) F, Y, W; (3) K, R, (4) A, G; (5) S, Т; (6) Q, N; (7) Е, D; и (8) С, S.
Производное может также быть результатом аминокислотных замен, являющихся функционально эквивалентными. При использовании здесь их следует понимать как аминокислотные замены, которые, при их проведении, приводят к слитому пептиду, обеспечивающему идентичный или сопоставимый (например, в пределах 10%) результат ELISA сыворотки животного, которому был введен слитый пептид, содержащий дериватизированный эпитопный фрагмент или дериватизированные эпитопные фрагменты, по сравнению со слитым пептидом без соответствующей дериватизации. Антигенность слитого пептида может, например, быть определена измерением титра антител, полученного иммунизацией животных, посредством ELISA, как описано, например, в Примерах 6 и 7. Аналогичный процесс может быть применен для анализа функциональной эквивалентности консервативной или иной аминокислотной замены. Например, иммунный ответ, вызванный слитым пептидом, содержащим недериватизированный «исходный» фрагмент, сравнивают, с применением того же анализа, с ответом, вызванным слитым пептидом, содержащим дериватизированный фрагмент. Если иммунный ответ, вызванный слитым пептидом, содержащим дериватизированный фрагмент, настолько же интенсивен, как ответ, вызванный слитым пептидом, содержащим недериватизированный фрагмент, то аминокислотную замену можно считать функционально эквивалентной. Если иммунный ответ после дериватизации превышает ответ без дериватизации, то аминокислотную замену можно считать улучшенной.
При использовании здесь термины «нативный», «природный» и «естественный» относятся к форме молекулы, обычно встречающейся в природе. Как таковая, «нативная» последовательность ECD Her2/neu относится к аминокислотным остаткам 23-652 опубликованной ранее аминокислотной последовательности Her2/neu (регистрационный номер в базе данных Swiss-Prot Р04626; ERBB2_HUMAN). Наоборот, «ненативная» последовательность, включая «ненативный линкер», представляет собой любую аминокислотную последовательность, не принадлежащую к нативной последовательности ECD Her2/neu. Таким образом, в одном воплощении, раскрытом здесь, пептидный «ненативный линкер» не является продолжением фрагментов Her2/neu, с которыми он соединен, на смежную нативную последовательность Her2/neu.
Слитые пептиды
Иммунопревентивные и/или иммунотерапевтические вакцинные композиции для применения при видах рака, экспрессирующих Her2/neu или сверхэкспрессирующих Her2/neu, на основе нескольких эпитопов/пептидов подразумевают введение таких пептидов в форме раздельных (то есть отдельных) пептидов. Это приводит к определенным недостаткам. Например, одновременное введение нескольких пептидов в одной композиции приводит к риску агрегации этих пептидов со снижением их предполагаемой доступности для иммунной системы хозяина. В предельных случаях растворимость таких агрегатов может снижаться настолько, что агрегаты выпадают в осадок, становясь недоступными для иммунной системы хозяина. В то же время, раздельное введение таких пептидов в разных растворах и в разных временных точках снижает вероятность полезного сочетания иммуногенных эффектов таких пептидов.
Дополнительные трудности возникают при использовании нескольких отдельных эпитопов с определенными типами систем доставки, такими как виросомы, липосомы или вирусоподобные частицы (VLP). Для обеспечения возможности воспроизводимого изготовления вакцин предпочтительно присутствие эпитопов в определенных концентрациях. Тем не менее, при сочетании (то есть ковалентном связывании) нескольких пептидных фрагментов с одной системой доставки, такой как виросома или липосома, трудно обеспечить одинаковое количество эпитопов, связанных с каждой системой доставки. Неизбежно возникают колебания числа эпитопов, связанных с одной системой доставки. Несмотря на то, что можно с определенной уверенностью полагать, что каждая жизнеспособная система доставки будет связана, скажем, с каждым из эпитопов А и В, некоторые системы доставки будут связаны с несколько большим числом эпитопов А, чем предполагалось, в то время как в других будет несколько повышено число эпитопов В. В то время как середина гауссова распределения эпитопов А и В будет стремиться к предполагаемому соотношению фрагментов А:В, любое гауссово распределение по определению содержит выбросы, и именно эти выбросы могут снижать максимальную иммуногенную эффективность, и по мере повышения строгости требований по поддержанию заданного соотношения эпитопов в системе доставки их удаление становится все более дорогостоящим и трудоемким. Сходные проблемы возникают при сочетании систем доставки, таких как виросомы, связанные с различными эпитопными фрагментами, при попытке получить желаемое отношение одного фрагмента ECD Her2/neu к другому.
Здесь раскрыт способ, позволяющий преодолеть или по меньшей мере частично уменьшить указанные выше недостатки. Связывание по меньшей мере двух несмежных В-клеточных эпитопов ECD Her2/neu или их производных в одном слитом пептиде позволяет получить однородную композицию, в которой будет присутствовать слитый пептид только одного типа. Элементы слитого пептида (то есть эпитопы ECD Her2/neu, являющиеся несмежными в нативном ECD) одинаковы в каждом пептиде и могут быть выбраны (или выбраны и модифицированы) таким образом, чтобы минимизировать нежелательные взаимодействия внутри полипептидов и между ними.
В одном воплощении, описанном здесь, вакцинная композиция будет содержать слитый пептид одного типа. Соответственно, соотношение присутствующих В-клеточных эпитопов в конечном счете продиктовано соотношением этих фрагментов в слитом пептиде. Это означает, что любое желаемое соотношение для получения иммуногенного ответа, может быть легко и надежно зафиксировано на уровне конструирования и разработки слитого пептида.
В завершение, при использовании нетоксичного варианта дифтерийного токсина CRM 197 в качестве белка-носителя для доставки слитых пептидов, описанных здесь, вариабельность, связанная с относительным распределением каждого В-клеточного эпитопа Her2/neu в вакцинной композиции может быть минимизирована или устранена, так как отношение одного В-клеточного эпитопа Her2/neu к его любому другому В-клеточному эпитопу, несмежному в нативном состоянии, остается постоянным независимо от того, сколько слитых пептидов связано с белком-носителем, поскольку, как указано выше, это отношение определено на уровне отдельного слитого пептида.
В других воплощениях, описанных здесь, вакцинная композиция будет содержать слитый пептид более чем одного типа.
Вакцинная композиция, раскрытая здесь, вызывает антиген-специфичный гуморальный иммунный ответ у хозяина, которому ее вводят. Неожиданно, антиген-специфичный ответ, вызванный вакцинной композицией, на порядки превосходит иммунный ответ, который можно получить с использованием В-клеточных эпитопов Her2/neu по отдельности или с использованием альтернативных система доставки, таких как виросомы. Таким образом, настоящее изобретение позволяет устранить или по меньшей мере минимизировать недостатки сложных способов изготовления с включением пептидов в виросомы, вариабельность, часто сопутствующую попыткам включить несколько отдельных пептидов в систему доставки, и возможные нежелательные взаимодействия между отдельными В-клеточными эпитопными фрагментами, которые могут снижать эффективность полученной вакцинной композиции.
При использовании здесь термин «слитый пептид» относится к ненативному пептиду, составленному из двух или более несмежных В-клеточных эпитопов ECD Her2/neu (например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или более несмежных В-клеточных эпитопов ECD Her2/neu), связанных друг с другом, например, пептидными (амидными) связями. В одном воплощении, раскрытом здесь, два или более В-клеточных эпитопа ECD Her2/neu являются несмежными в их нативном состоянии, то есть в ECD нативного Her2/neu.
В одном воплощении, раскрытом здесь, слитый пептид содержит по меньшей мере три несмежных В-клеточных эпитопа ECD Her2/neu. В одном воплощении слитый пептид содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 8-14 и аминокислотных последовательностей, по меньшей мере на 85% идентичных любой из последовательностей, указанных выше.
В Таблице 2 ниже представлен перечень выбранных В-клеточных эпитопов, а также слитых пептидов по некоторым из воплощений, раскрытых здесь.

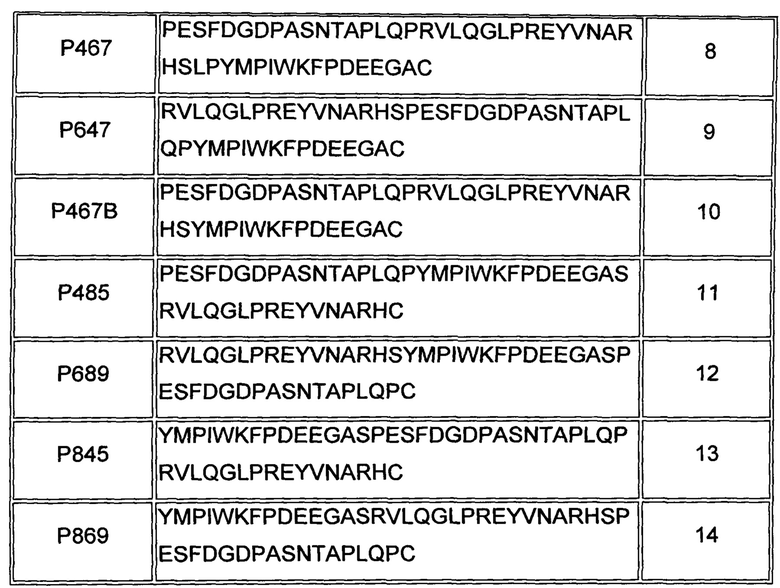
В одном воплощении, раскрытом здесь, слитый пептид содержит аминокислотную последовательность SEQ ID NO: 8 (обозначенную как слитый пептид Р467) или SEQ ID NO: 9 (обозначенную как слитый пептид Р647) или аминокислотную последовательность, по меньшей мере на 85% идентичную SEQ ID NO: 8 или SEQ ID NO: 9, как показано в Таблице 2 выше.
Здесь также предполагаются слитые пептиды, содержащие два или более В-клеточных эпитопа ECD Her2/neu, выбранных из группы, состоящей из SEQ ID NO: 1-7 и 15-60 или аминокислотных последовательностей, по меньшей мере на 85% идентичных любой из последовательностей, указанных выше, соединенные два или более раза в тандемном повторе. Например, слитые пептиды, предполагаемые здесь, могут содержать два или более тандемных повтора SEQ ID NO: 1, два или более тандемных повтора SEQ ID NO: 2, два или более тандемных повтора SEQ ID NO: 3, два или более тандемных повтора SEQ ID NO: 4, два или более тандемных повтора SEQ ID NO: 5 и так далее. Тем не менее, следует понимать, что включение двух или более разных В-клеточных эпитопов в слитый пептид повысит вероятность получения более полезного иммунного ответа, приводя к выработке антител, специфично распознающих несколько В-клеточных эпитопов ECD Her2/neu. Таким образом, в одном воплощении, раскрытом здесь, слитый пептид содержит по меньшей мере два разных В-клеточных эпитопа ECD Her2/neu, выбранных из группы, состоящей из SEQ ID NO: 1-7 и 15-60 или аминокислотных последовательностей, по меньшей мере на 85% идентичных любой из последовательностей, указанных выше.
В одном воплощении, раскрытом здесь, слитый пептид состоит из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 8-14. В еще одном воплощении слитый пептид состоит из аминокислотной последовательности SEQ ID NO: 8 (обозначенной как слитый пептид Р467) или SEQ ID NO: 9 (обозначенной как слитый пептид Р647).
В отдельных В-клеточных эпитопах SEQ ID NO: 3 и 4, как показано выше в Таблице 2, SEQ ID NO: 4 содержит дополнительный С-концевой линкер с Cys на конце (то есть GGGGGC), которого нет в SEQ ID NO: 3. В конце этого С-концевого линкера в самой SEQ ID NO: 4 расположен Cys, который позволяет этому отдельному эпитопу связываться с белком-носителем, как проиллюстрировано в некоторых Примерах, изложенных здесь ниже. При использовании в качестве части слитого пептида по изобретению В-клеточный эпитоп SEQ ID NO: 4 присутствует в этом слитом пептиде в форме SEQ ID NO: 3. В экспериментах, в которых использован В-клеточный эпитоп SEQ ID NO: 4, в качестве В-клеточного эпитопа функционирует та часть SEQ ID NO: 4, которая соответствует SEQ ID NO: 3.
Подходящие способы получения слитого пептида, как описано здесь, будут знакомы специалистам в данной области. Наглядный пример включает пептидный синтез, включающий последовательное образование пептидных связей, связывающих каждый В-клеточный эпитоп с его соответствующим соседним В-клеточным эпитопом, как описано здесь, и получение указанного слитого пептида. Подходящие способы пептидного синтеза известны в данной области. Наглядные примеры включают способы, описанные в "Amino Acid and Peptide Synthesis" (Oxford Chemistry Primers; by John Jones, Oxford University Press). Синтетические пептиды могут также быть получены жидкофазным синтезом или твердофазным пептидным синтезом (solid-phase peptide synthesis, SPPS) на различных твердых подложках (например, полистироле, полиамиде или полиэтиленгликоле (PEG)). SPPS может включать использование Fmoc (9Н-флуорен-9-илметоксикарбонил) или t-Boc (трет-бутоксикарбонил). Пептиды также можно заказать у ряда коммерческих производителей.
Альтернативно, слитый пептид может быть получен по рекомбинантной методологии. Например, молекулой нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты, кодирующую слитый белок, можно трансфицировать подходящую клетку-хозяина, способную экспрессировать указанную последовательность нуклеиновой кислоты, и указанную клетку-хозяина можно инкубировать в условиях, подходящих для экспрессии указанной последовательности нуклеиновой кислоты, с последующим выделением указанного слитого пептида. Подходящие способы получения молекулы нуклеиновой кислоты, кодирующей интересующий слитый пептид (пептиды), будут также известны специалистам в данной области на основе знания генетического кода, возможно, включая кодоны, оптимизированные, исходя из природы клетки-хозяина (например, микроорганизма), используемой для экспрессии и/или секреции рекомбинантного слитого пептида (пептидов). Специалистам в данной области будут также известны подходящие клетки-хозяева, наглядные примеры которых включают прокариотические клетки (например, Е. coli) и эукариотические клетки (например, P. pastoris). См. "Short Protocols in Molecular Biology, 5th Edition, 2 Volume Set: A Compendium of Methods from Current Protocols in Molecular Biology" (by Frederick M. Ausubel (author, editor), Roger Brent (editor), Robert E. Kingston (editor), David D. Moore (editor), J. G. Seidman (editor), John A. Smith (editor), Kevin Struhl (editor), J Wiley & Sons, London).
При использовании здесь термины «кодировать», «кодирующий» и тому подобные относятся к способности нуклеиновой кислоты обеспечивать получение другой нуклеиновой кислоты или полипептида. Например, говорят, что последовательность нуклеиновой кислоты «кодирует» полипептид, если она может быть транскрибирована и/или транслирована, обычно в клетке-хозяине, с получением полипептида или если она может быть процессирована с образованием формы, которая может быть транскрибирована и/или транслирована с получением полипептида. Такая последовательность нуклеиновой кислоты может содержать кодирующую последовательность или как кодирующую последовательность, так и некодирующую последовательность. Таким образом, термины «кодировать», «кодирующий» и тому подобные включают РНК-продукт, являющийся результатом транскрипции молекулы ДНК, белок, являющийся результатом трансляции молекулы РНК, белок, являющийся результатом транскрипции молекулы ДНК с образованием РНК-продукта и последующей трансляции РНК-продукта, или белок, являющийся результатом транскрипции молекулы ДНК с образованием РНК-продукта, процессинга РНК-продукта с образованием процессированного РНК-продукта (например, матричной РНК (мРНК)) и последующей трансляции процессированного РНК-продукта. В некоторых воплощениях кодоны последовательности нуклеиновой кислоты, кодирующей В-клеточные эпитопы или слитые пептиды, описанные здесь, оптимизированы для экспрессии в подходящей клетке-хозяине. Например, там, где слитый пептид предназначен для использования для индукции иммунного ответа против Her2/neu у субъекта-человека, кодоны последовательностей нуклеиновых кислот могут быть оптимизированы для человека. Подходящие способы оптимизации кодонов будут известны специалистам в данной области, например, использование опции "Reverse Translation" инструмента "Gene Design" в "Software Tools" на сайте Build a Genome университета Джона Хопкинса.
Два или более несмежных В-клеточных эпитопа Her2/neu, как описано здесь, могут быть связаны друг с другом в слитом пептиде любым способом, известным специалистам в данной области. Термины «связь» и «связанный» включают прямое связывание двух несмежных В-клеточных эпитопов Her2/neu пептидной связью, то есть, С-конец одного эпитопа Her2/neu ковалентно связан пептидной связью с N-концом другого нативно несмежного эпитопа. Значение терминов «связь» и «связанный» также включает связывание двух нативно несмежных эпитопов Her2/neu расположенным между ними линкерным элементом.
В одном воплощении, раскрытом здесь, по меньшей мере два указанных В-клеточных эпитопа или их производных связаны друг с другом пептидной последовательностью ненативного линкера.
При использовании здесь термин «линкер» относится к короткой полипептидной последовательности, расположенной между какими-либо двумя соседними В-клеточными эпитопами Her2/neu или их производными в слитом полипептиде. В одном воплощении линкер представляет собой полипептидный линкер из 1-10, предпочтительно 1, 2, 3, 4 или 5 встречающихся в природе или невстречающихся в природе аминокислот.В одном воплощении линкер представляет собой углеводный линкер. Подходящие углеводные линкеры будут известны специалистам в данной области. В другом воплощении, раскрытом здесь, слитый пептид содержит один или более чем один пептидный или полипептидный линкер (линкеры) вместе с одним или более чем одним непептидным или неполипептидным линкером (линкерами). Кроме того, по необходимости в тот же слитый пептид могут быть включены линкеры других типов, пептидные или непептидные. В случае использования пептидного или полипептидного линкера для соединения двух соответствующих В-клеточных эпитопных фрагментов ECD Her2/neu линкер будет предпочтительно включен таким образом, что его N-конец будет связан пептидной связью с С-концом одного эпитопного фрагмента и его С-конец будет связан пептидной связью с N-концом другого фрагмента. К отдельным В-клеточным эпитопным фрагментам в слитом пептиде могут также быть добавлены одна или более чем одна аминокислота на одном или обоих концах, предпочтительно на С-конце. Так, например, к N- и/или С-концу пептидов могут быть добавлены линкерные или спейсерные аминокислоты для связывания несмежных пептидов и для обеспечения возможности ковалентного связывания пептидов друг с другом и/или с системой доставки, такой как виросома, через молекулу липида в виросоме, используемую в качестве якоря. Наглядным примером подходящего пептидного линкера является LP (лейцин-пролин), как показано, например, в SEQ ID NO: 8.
Специалистам в данной области будет ясно, что при связывании слитого пептида с белком-носителем через линкер предпочтительно проводить такое линкер-опосредованное связывание с С-конца слитого пептида, поскольку связывание с линкером с N-конца может, в некоторых случаях, оказывать негативное влияние на желаемый иммунный ответ, который необходимо вызвать.
Идентичность последовательностей и сходство последовательностей
Ссылка на «по меньшей мере 85%» включает идентичность или сходство последовательностей на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%, например, после оптимального выравнивания или анализа наилучшего соответствия. Таким образом, в предпочтительных формах настоящего изобретения последовательность по меньшей мере на 85%, предпочтительно по меньшей мере на 86%, предпочтительно по меньшей мере на 87%, предпочтительно по меньшей мере на 88%, предпочтительно по меньшей мере на 89%, предпочтительно по меньшей мере на 90%, предпочтительно по меньшей мере на 91%, предпочтительно по меньшей мере на 92%, предпочтительно по меньшей мере на 93%, предпочтительно по меньшей мере на 94%, предпочтительно по меньшей мере на 95%, предпочтительно по меньшей мере на 96%, предпочтительно по меньшей мере на 97%, предпочтительно по меньшей мере на 98%, предпочтительно по меньшей мере на 99%, предпочтительно по меньшей мере на 100% идентична или гомологична последовательностям, указанным здесь, например, после оптимального выравнивания или анализа наилучшего соответствия.
При использовании здесь термины «идентичность», «сходство», «идентичность последовательностей», «сходство последовательностей», «гомология», «гомология последовательностей» и тому подобные означают, что в любом конкретном аминокислотном положении выравниваемой последовательности аминокислотные остатки выравниваемых последовательностей идентичны. При использовании здесь термин «сходство» или «сходство последовательностей» означает, что в любом конкретном положении выравниваемых последовательностей аминокислотные остатки последовательностей принадлежат к одному типу. Например, лейцин может быть заменен на изолейциновый или валиновый остаток. Это может быть названо консервативной заменой. В одном воплощении аминокислотные последовательности могут быть модифицированы посредством консервативной замены любого из содержащихся в них аминокислотных остатков, таким образом, что модификация не оказывает эффекта на специфичность связывания или функциональную активность модифицированного полипептида при сравнении с немодифицированным полипептидом.
В некоторых воплощениях идентичность последовательностей применительно к полипептиду относится к проценту аминокислотных остатков в последовательности-кандидате, идентичных остаткам соответствующего полипептида после выравнивания последовательностей и, по необходимости, введения разрывов для достижения максимального процента гомологии, не учитывая никакие консервативные замены как часть идентичности последовательностей. N- или С-концевые удлиняющие сегменты и вставки не рассматривают как снижение идентичности или гомологии последовательностей. Способы и компьютерные программы для проведения выравнивания двух или более аминокислотных последовательностей и определения их идентичности или гомологии хорошо известны специалистам в данной области. Например, процент идентичности или сходства двух аминокислотных последовательностей может легко быть рассчитан с применением алгоритмов, например, BLAST, FASTA или алгоритма Смита-Уотермана (Smith-Waterman). Настоящее изобретение включает последовательности, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 93%, 95%, 96%, 97%, 98% или 99% идентичные или гомологичные последовательностям, указанным здесь, и применение этих последовательностей в способах, описанных здесь.
В некоторых воплощениях идентичность последовательностей применительно к полинуклеотиду относится к проценту нуклеотидов в последовательности-кандидате, идентичных нуклеотидам соответствующего полинуклеотида после выравнивания последовательностей и, по необходимости, введения разрывов для достижения максимального процента гомологии, не учитывая никакие консервативные замены как часть идентичности последовательностей. Способы и компьютерные программы для проведения выравнивания двух или более полинуклеотидных последовательностей или последовательностей нуклеиновых кислот и определения их идентичности или гомологии хорошо известны специалистам в данной области.
Методики определения «сходства» аминокислотных последовательностей хорошо известны специалистам в данной области. Обычно «сходство» означает точное сравнение аминокислот двух или более полипептидов или сравнение в подходящем месте, где аминокислоты идентичны или обладают сходными химическими и/или физическими свойствами, такими как заряд или гидрофобность. Затем может быть определен так называемый «процент сходства» сравниваемых полипептидных последовательностей. Методики определения идентичности последовательности нуклеиновой кислоты и аминокислотной последовательности также хорошо известны в данной области м включают определение нуклеотидной последовательности мРНК данного гена (обычно через промежуточную кДНК), определение кодируемой им аминокислотной последовательности и ее сравнение со второй аминокислотной последовательностью. Обычно «идентичность» относится к точному соответствию нуклеотидов или аминокислот двух полинуклеотидов или полипептидных последовательностей, соответственно.
Две или более полинуклеотидные последовательности можно также сравнивать, определяя их «процент идентичности». Сходным образом, две или более аминокислотные последовательности можно сравнивать, определяя их «процент идентичности». Процент идентичности двух последовательностей, либо последовательностей нуклеиновых кислот, либо пептидных последовательностей, можно описать как отношение числа точных совпадений между двумя выровненными последовательностями к длине более короткой последовательности, умноженное на 100. Приближенное выравнивание для последовательностей нуклеиновых кислот обеспечивает алгоритм поиска локальной гомологии Смита и Уотермана (Smith and Waterman, Advances in Applied Mathematics 2:482-489 (1981)). Этот алгоритм может быть адаптирован для применения с пептидными последовательностями с использованием оценочной матрицы, разработанной Dayhoff, Atlas of Protein Sequences and Structure, M.O. Dayhoff ed., 5 suppl. 3:353-358, National Biomedical Research Foundation, Washington, D.C., USA, и нормализованной Gribskov, Nucl. Acids Res. 14(6).6745-6763 (1986). Подходящие программы для вычисления процента идентичности или сходства последовательностей общеизвестны в данной области.
В наглядном примере определения идентичности последовательностей нуклеиновых кислот рассматриваемую последовательность нуклеиновой кислоты используют для поиска по базе данных последовательностей нуклеиновых кислот, такой как база данных GenBank (доступная по электронному адресу http://www.ncbi.nln.nih.gov/blast/), с применением программы BLASTN версии 2.1 (на основе Altschul et al. (1997) Nucleic Acids Research 25:3389-3402). Эту программу можно использовать в режиме без разрывов. Для удаления гомологии последовательностей, обусловленной областями низкой сложности, применяют фильтрацию по умолчанию. Можно использовать параметры BLASTN по умолчанию. В наглядном примере определения идентичности аминокислотных последовательностей аминокислотную последовательность используют для поиска по базе данных белковых последовательностей, такой как база данных GenBank (доступная на сайте http://www.ncbi.nln.nih.gov/blast/), с применением программы BLASTP. Программу можно использовать в режиме без разрывов. Для удаления гомологии последовательностей, обусловленной областями низкой сложности, применяют фильтрацию по умолчанию. Используют параметры BLASTP по умолчанию. Для фильтрации последовательностей низкой сложности может быть использована программа SEG.
Оптимальное выравнивание последовательностей для выравнивания окна сравнения может быть проведено компьютеризированным применением алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics Software Package Release 7.0, Genetics Computer Group, 575 Science Drive Madison, WI, USA) или обследованием и наилучшим выравниванием (то есть приводящим к наибольшему проценту гомологии в окне сравнения), полученным любым выбранным способом. См. также семейство программ BLAST, например, как раскрыто Altschul et al., 1997, Nucl. Acids Res. 25:3389. Подробное обсуждение анализа последовательностей можно найти в разделе 19.3 Ausubel et al., "Current Protocols in Molecular Biology", John Wiley & Sons Inc, 1994-1998, Chapter 15.
Белок-носитель
CRM197 (регистрационный номер в GenBank 1007216А; SEQ ID NO: 61) представляет собой ферментативно неактивную и нетоксичную форму дифтерийного токсина, содержащую одну аминокислотную замену (Gly-Glu) по аминокислотному остатку 52. CRM197 отличается от токсина дикого типа единственной мутацией GCA, приводящей к замене Glu52. Отсутствие токсичности у CRM197, по-видимому, обусловлено потерей ферментативной активности его фрагмента А, который в токсине дикого типа катализирует химическую модификацию фактора элонгации 2 (транслоказы) в инфицированных клетках, необходимого для синтеза белка. Это свойство отсутствия токсичности делает CRM 197 подходящим белком-носителем для изготовления конъюгированных вакцин.
SEQ ID NO:61 (CRM197; регистрационный номер в GenBank 1007216А)

Способы, посредством которых слитый пептид может быть связан с CRM 197, известны специалистам в данной области.
Наглядные примеры включают примеры, описанные Chang et al. (1998, FEBS Letters, 427:362-366) и Berti ef al. (2004, Biophysical Journal, 86:3-9), a также приведенные здесь в Примере 1 ниже.
Конъюгацию слитого пептида, как описано здесь, с CRM197 обычно проводят посредством активации лизиновых остатков с использованием подходящих агентов для перекрестного связывания. Поскольку CRM197 содержит 40 лизиновых остатков и многие из них доступны для перекрестного связывания, конечные продукты конъюгации с CRM 197 всегда неоднородны. Независимо от теории или конкретного механизма действия, обычно принято считать, что отношение «слитый пептид: белок-носитель» зависит от размера или молекулярной массы слитого пептида. Например, если слитый пептид относительно небольшой (например, приблизительно 10 аминокислот в длину), возможно получение белка-носителя, конъюгированного с 20-39 слитыми пептидами. В случае большого пептида, наоборот, белок-носитель может быть конъюгирован с 20 или меньшим числом слитых пептидов. В одном воплощении, описанном здесь, белок-носитель содержит от 2 до 39 слитых пептидов. В другом воплощении белок-носитель содержит по меньшей мере 20 слитых пептидов. В еще одном воплощении белок-носитель содержит от 6 до 12 слитых пептидов.
Слитый пептид обычно связан с белком-носителем ковалентной связью. Тем не менее, в некоторых воплощениях слитый пептид может быть связан с белком-носителем нековалентной связью. Там, где слитый пептид связан с белком-носителем нековалентно, нековалентная связь будет обычно включать электромагнитное взаимодействие одного или более чем одного атома слитого пептида с одним или более чем одним атомом белка-носителя. Наглядные примеры включают ионную связь (то есть притяжение двух противоположно заряженных ионов, благодаря их противоположному заряду), силы Ван-дер-Ваальса (то есть силы между постоянными и/или индуцированными диполями существующих ковалентных связей в слитом пептиде и белке-носителе) и/или гидрофобные взаимодействия (то есть силы, являющиеся результатом склонности гидрофобных/алифатических областей слитого пептида (пептидов), описанного здесь, связываться с гидрофобными областями белка-носителя).
Адъюванты
При использовании здесь термин «адъювант» обычно относится к классу веществ, которые могут повышать интенсивность иммунного ответа, вызванного конъюгатом слитого пептида с белком-носителем, до значений, превышающих те, которые можно было бы ожидать при использовании слитого пептида самого по себе или конъюгата слитого пептида с белком-носителем, как описано здесь, без адъюванта.
Подходящие адъюванты будут известны специалистам в данной области. Неограничивающие примеры подходящих адъювантов включают соли алюминия (например, гидрат окиси алюминия, фосфат алюминия и сульфат калия и алюминия (также называемые Alum)), липосомы, виросомы, эмульсии типа «вода в масле» или «масло в воде» (например, адъювант Фрейнда, Montanide®, MF59® и AS03), 3-O-деацил-4'-монофосфориллипид A (MPL), адъюванты, содержащие MPL (например, AS01, AS02 и AS04), и адъюванты на основе сапонинов. Адъюванты на основе сапонинов содержат сапонины или производные сапонинов, например, из Quillaja saponaria, Panax ginseng, Panax notoginseng, Panax quinquefolium, Platycodon grandiflorum, Polygala senega, Polygala tenuifolia, Quillaja brasiliensis, Astragalus membranaceus и Achyranthes bidentata. Типичные адъюванты на основе сапонинов включают иммуностимулирующие комплексы (ISCOMS), ISCOM-матрицу, адъювант ISCOMATRIX™, адъювант Matrix М™, адъювант Matrix С™, адъювант Matrix Q™, адъювант AblSCO®-100, адъювант AblSCO®-300, ISCOPREP™, производное ISCOPREP™, адъювант, содержащий ISCOPREP™ или производное ISCOPREP™, QS-21, производное QS-21 и адъювант, содержащий QS-21 или производное QS-21. Вакцинная композиция, как описано здесь, может также быть объединена с иммуномодулирующими агентами, включая, например, цитокины, хемокины и факторы роста. Здесь также предполагают смеси двух или более адъювантов в одной вакцинной композиции.
В одном воплощении адъювант представляет собой гидрат окиси алюминия. В одном воплощении адъювант представляет собой Montanide.
В одном воплощении, раскрытом здесь, вакцинная композиция дополнительно содержит ингибитор контрольных точек. Как будет ясно специалистам в данной области, термин «ингибитор контрольных точек» обозначает молекулы, которые ингибируют, снижают или иным образом влияют или модулируют один или более чем один из белков контрольных точек, полностью или частично. Наглядные примеры подходящих ингибиторов контрольных точек включают антитела и их антиген-связывающие фрагменты (например, Fab-фрагменты). Подходящие белки контрольных точек будут известны специалистам в данной области, и наглядные примеры таких белков включают CTLA-4 и его лиганды CD80 и CD86; PDI и его лиганды PDLI и PDL2; ОХ40 и его лиганд OX40L; LAG-3 и его лиганд МНС класса I или II; TIM-3 и его лиганд GAL-9; и В- и Т-лимфоцитарный аттенюатор (В- and T-lymphocyte attenuator, BTLA) и его лиганд медиатор проникновения вируса герпеса (herpes virus entry mediator, HVEM).
Фармацевтическая композиция
В настоящем описании также описана фармацевтическая композиция, содержащая вакцинную композицию, как описано здесь, и фармацевтически приемлемый носитель. Подходящие фармацевтически приемлемые носители (например, эксципиенты, разбавители и так далее) будут известны специалистам в данной области. Например, могут быть использованы множество водных (фармацевтически приемлемых) носителей, такие как забуференная вода, 0,4%-й солевой раствор, 0,3%-й глицин, гиалуроновая кислота и тому подобное. Композиции могут быть стерилизованы обычными хорошо известными методиками стерилизации или могут быть стерилизованы фильтрацией. Полученные водные растворы могут быть упакованы для использования, как есть, или лиофилизированы, при этом лиофилизированные препараты смешивают со стерильным раствором перед введением. Композиции могут дополнительно содержать фармацевтически приемлемые вспомогательные вещества, необходимые для создания условий, близких к физиологическим, такие как агенты для коррекции рН и буферные агенты, агенты для коррекции тоничности, смачивающие агенты и тому подобное, например, среди многих других, ацетат натрия, лактат натрия, хлорид натрия, хлорид калия, хлорид кальция, сорбитанмонолаурат, триэтаноламинолеат, сахарозу или другие углеводы. Подходящие способы получения соединений для парентерального введения будут известны или очевидны специалистам в данной области и описаны более подробно, например, в A. Gennaro (2000) "Remington: The Science and Practice of Pharmacy", 20th edition, Lippincott, Williams, & Wilkins; Pharmaceutical Dosage Forms and Drug Delivery Systems (1999) H.C. Ansel et al., eds 7.sup.th ed., Lippincott, Williams, & Wilkins; and Handbook of Pharmaceutical Excipients (2000) A.H. Kibbe et al., eds., 3.sup.rd ed. Amer. Pharmaceutical Assoc.
Фармацевтическая композиция может быть в форме, подходящей для парентерального введения (например, подкожной, внутримышечной или внутривенной инъекции) или в аэрозольной форме, подходящей для введения ингаляцией, такой как интраназальная ингаляция или пероральная ингаляция.
Фармацевтические композиции, описанные здесь, могут также быть представлены в наборе. Набор может содержать дополнительные компоненты, способствующие осуществлению способов, как описано здесь, такие как устройство (устройства) для введения, эксципиент (эксципиенты) и/или разбавитель (разбавители). Наборы могут содержать контейнеры для размещения различных компонентов и инструкции по применению компонентов набора в таких способах.
В одном воплощении, раскрытом здесь, фармацевтическая композиция дополнительно содержит ингибитор контрольных точек, как описано здесь. Применения и способы лечения или предупреждения рака
Здесь также раскрыт способ лечения или предупреждения рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu, у пациента, нуждающегося в этом, включающий стадию введения указанному пациенту эффективного количества вакцинной композиции, как описано здесь, или фармацевтической композиции, как описано здесь.
Настоящее описание также включает применение вакцинной композиции, как описано здесь, в изготовлении лекарственного средства для лечения или предупреждения рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu, у пациента, нуждающегося в этом.
Настоящее описание также включает вакцину, как описано здесь, или фармацевтическую композицию, как описано здесь, для применения в лечении или предупреждении рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu, у пациента, нуждающегося в этом.
Специалистам в данной области будут известны типы рака, характеризующиеся экспрессией или сверхэкспрессией белка Her2/neu. Наглядные примеры включают рак молочной железы, рак яичника, рак эндометрия, рак желудка, рак поджелудочной железы, рак предстательной железы и рак слюнной железы. В одном воплощении рак представляет собой рак молочной железы. В другом воплощении рак представляет собой рак желудка.
Вакцинные или фармацевтические композиции, как описано здесь, обычно вводят в «эффективном количестве», то есть количестве, эффективном для оказания любого одного или более из, среди прочего, терапевтического или профилактического эффекта. Специалисты в данной области смогут, посредством рутинных исследований, определить эффективное нетоксичное количество, которое следует включить в фармацевтическую композицию или ввести для получения желаемого результата. Обычно вакцинные и/или фармацевтические композиции, как раскрыто здесь, могут быть введены способом, совместимым с путем введения и физическими характеристиками реципиента (включая состояние здоровья), и таким образом, что это окажет желаемый эффект (эффекты) (то есть терапевтически эффективный, иммуногенный и/или защитный). Например, подходящая доза композиции может зависеть от множества факторов, включая, без ограничения, физические характеристики субъекта (например, возраст, массу тела, пол), применение композиции в качестве единственного агента или как части комбинированной терапии, прогрессирование (то есть патологическое состояние) какого-либо основного вида рака и другие факторы, которые могут быть определены специалистами в данной области. Другие наглядные примеры общих соображений, которые можно принимать во внимание при определении, например, подходящей дозы композиций, обсуждаются в Gennaro (2000), "Remington: The Science and Practice of Pharmacy", 20th edition, Lippincott, Williams, & Wilkins; и Gilman et al., (Eds), (1990), "Goodman And Gilman's: The Pharmacological Bases of Therapeutics", Pergamon Press.
Ожидают, что это количество будет входить в относительно широкий диапазон, который может быть определен способами, известными специалистам в данной области, с учетом некоторых из изложенных выше соображений.
Эффективное количество слитого пептида (конъюгированного с белком-носителем), как описано здесь, будет обычно входить в диапазон от приблизительно 5 мкг до приблизительно 1,0 мг слитого пептида на субъекта, от приблизительно 10 мкг до приблизительно 500 мкг слитого пептида на субъекта или от приблизительно 15 мкг до приблизительно 60 мкг слитого пептида на субъекта. Эффективное количество может быть определено, например, стандартными способами, включающими измерение титров антиген-специфичных антител. Можно проводить мониторинг иммунитета, обеспеченного композициями, описанными здесь, для определения наличия или отсутствия необходимости повторного введения. Например, после оценки титра антиген-специфичных антител в сыворотке, обычно через несколько дней или недель после первого введения композиции субъекту, могут быть необходимы и/или желательны возможные повторные иммунизации. Применение нескольких введений повысит вероятность получения высоких титров антиген-специфичных антител, как показано в Примерах ниже.
Вакцинные и/или фармацевтические композиции, как описано здесь, могут быть введены субъекту, нуждающемуся в этом, отдельно или в комбинации с дополнительным терапевтическим агентом (агентами), то есть, как часть комбинированной терапии. В контексте комбинированной терапии введение может быть одновременным или последовательным; то есть вакцинную и/или фармацевтическую композицию вводят первой с последующим введением дополнительного терапевтического и/или профилактического агента (агентов), или вакцинную и/или фармацевтическую композицию вводят после введения дополнительного терапевтического агента (агентов). Таким образом, при введении субъекту двух или более агентов «в комбинации» они могут быть введены в одной композиции в одно и то же время, или в раздельных композициях в одно и то же время, или в раздельных композициях с разделением во времени.
Дополнительный терапевтический агент (агенты) может включать ингибитор контрольных точек, как описано здесь. Таким образом, в одном воплощении способы, раскрытые здесь, дополнительно включают стадию введения указанному пациенту эффективного количества ингибитора контрольных точек, как описано здесь.
Специалистам в данной области будет ясно, что оптимальное число отдельных введений и интервал между ними, в случае их необходимости для получения желаемого иммунного ответа, могут быть определены, например, формой, путем и местом введения и характеристиками конкретного субъекта, у которого будут проводить лечение, как описано здесь. Оптимальные условия могут быть определены с применением обычных методик, известных специалистам в данной области.
В некоторых случаях может быть желательным проводить несколько или множество введений вакцинных и/или фармацевтических композиций, как описано здесь. Например, композиции могут быть введены 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более раз. Введения могут быть проведены с интервалом от приблизительно одних суток до приблизительно двенадцати недель и, в определенных воплощениях, с интервалом от приблизительно одной до приблизительно четырех недель. Периодические повторные введения могут быть необходимы для достижения желаемого терапевтического результата, такого как уменьшение размера опухоли и/или снижение частоты метастазов. Специалистам в данной области также будет очевидно, что оптимальный курс введения может быть определен с применением обычного курса лечения, или тестов по определению эффективности или иммунного статуса.
Следует понимать, что «индукция» иммунного или антиген-специфичного гуморального иммунного ответа, как подразумевают здесь, включает получение или стимуляцию иммунного ответа и/или усиление ранее существовавшего иммунного ответа с получением желаемого физиологического эффекта, такого как уменьшение размера опухоли и/или снижение частоты метастазов. Эффект может быть профилактическим в смысле полного или частичного предупреждения развития рака, полного или частичного предупреждения развития метастазов и/или полного или частичного предупреждения симптома, связанного с указанным раком.
При использовании здесь термин «введение» обычно относится к стадии введения вакцинных и/или фармацевтических композиций, как описано здесь, в организм пациента, таким образом, что иммунная система пациента формирует ответ на несколько В-клеточных эпитопов Her2/neu, присутствующих в слитом пептиде (пептидах). При использовании здесь «пациент, нуждающийся в этом», включает индивида, у которого диагностирован рак, где раковые клетки экспрессируют или сверхэкспрессируют белок Her2/neu. Он также включает индивидов, у которых еще не диагностирован рак, таких как индивиды, у которых нет никаких симптомов, но у которых рак может быть в наследственном анамнезе, и, следовательно, у которых предполагают наличие генетической предрасположенности к развитию рака. Таким образом, в его наиболее широком смысле термин «пациент, нуждающийся в этом», включает индивидов, которые уже нуждаются в этом, а также индивидов с риском развития Her2/neu-ассоциированного рака.
При использовании здесь лекарственное средство, обеспечивающее «предупреждение» рака, будет снижать риск развития рака у индивида, в идеальном случае до нуля. Кроме того, данный термин также относится к предупреждению повторного развития рака, например, после хирургического удаления первичной опухоли. При использовании здесь лекарственное средство, обеспечивающее «лечение» рака, будет в идеальном случае устранять заболевание полностью посредством устранения его основной причины, таким образом, что после прекращения введения композиции не будет происходить повторного развития заболевания, но будет сохраняться ремиссия. При использовании здесь лекарственное средство, обеспечивающее «уменьшение интенсивности» рака, не устраняет основную причину заболевания, но уменьшает тяжесть заболевания, оцениваемую по какой-либо принятой системе оценки и/или по улучшению самочувствия пациента, например, уменьшению боли и/или дискомфорта.
«Эффективное количество» представляет собой количество фармацевтического препарата, которое само по себе или вместе с дополнительными дозами в соответствии с установленной схемой введения обеспечивает желаемое предупреждение, лечение или уменьшение интенсивности, как обсуждено здесь.
Способ и схема введения могут варьировать в зависимости от стадии или тяжести состояния, подлежащего лечению, и должны быть определены квалифицированным практикующим врачом. Например, вакцинная или фармацевтическая композиция по настоящему изобретению, как описано здесь, может быть введена в форме для подкожного, внутрикожного, интралимфатического, местного введения, введения через слизистую оболочку (интраназального) или внутримышечного введения. Все эти формы хорошо известны специалистам в области фармацевтики.
В определенных воплощениях композиции, описанные здесь, могут быть введены внутримышечным, подкожным, интралимфатическим или интраназальным способом введения, как известно специалистам в данной области.
Как обсуждено здесь, схему введения с использованием композиций, как описано здесь, выбирают в соответствии с множеством фактором, включая, например, возраст, массу тела и состояние здоровья пациента, стадию или тяжесть состояния, подлежащего лечению, и конкретный слитый пептид (пептиды), которые предполагается вводить. Квалифицированный врач легко сможет определить и установить эффективное количество, например, белка-носителя (содержащего связанный с ним слитый пептид (пептиды)), необходимое для предупреждения, лечения или уменьшения интенсивности прогрессирования или тяжести злокачественной опухоли. Схема введения будет также учитывать количество адъюванта, которое необходимо будет вводить и которое, как ожидают, будет варьировать в зависимости по меньшей мере от некоторых факторов, определяющих количество белка-носителя / слитого пептида (пептидов), необходимое для введения субъекту, таких как возраст, масса тела и состояние здоровья пациента. Для оптимальной точности достижения концентрации белка-носителя / слитого пептида (пептидов) и/или адъюванта в диапазоне, обеспечивающем эффективность (то есть эффективного количества) без токсичности или с не более чем приемлемой токсичностью, необходима схема введения, основанная, по меньшей мере частично, на кинетике доступности белка-носителя / слитого пептида (пептидов) и адъюванта в целевой области (областях). Этот процесс будет обычно включать анализ распределения, равновесия и выведения белка-носителя / слитого пептида (пептидов) и адъюванта, который может быть проведен специалистами в данной области.
Все патенты, заявки на патенты и публикации, упомянутые здесь, полностью включены посредством ссылки.
Специалистам в данной области будет ясно, что возможны другие варианты и модификации изобретения, описанного здесь, помимо тех, что описаны конкретно. Следует понимать, что сущность и объем изобретения включает все такие варианты и модификации. Изобретение также включает все стадии, признаки, композиции и соединения, упомянутые или указанные в данном описании, по отдельности или совместно, и все и любые комбинации любых двух или более из указанных стадий или признаков.
Далее определенные воплощения изобретения будут описаны со ссылкой на следующие примеры, приведенные исключительно для наглядности и не ограничивающие объем применимости, описанный выше.
ПРИМЕРЫ
Следующие примеры направлены на конъюгацию двух слитых пептидов (несмежных В-клеточных эпитопов, имеющих происхождение от Her2/neu) с рекомбинантным детоксифицированным дифтерийным токсином CRM197. Начальный анализ конъюгатов слитых пептидов с белком-носителем проводили посредством электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) и иммуноблоттинга. В завершение, конъюгаты слитых пептидов с белком-носителем отправляли в Австрию для начальных исследований иммуногенности у животных.
Пример 1 - Конъюгация пептидных антигенов Her2/neu с рекомбинантным детоксифицированным вариантом дифтерийного токсина CRM197
Вакцина против рака молочной железы на основе виросом PEV6A/B была успешно испытана в клиническом исследовании I фазы (Wiedermann et al., Breast Cancer Res Treat. 2010). Тем не менее, у этой вакцинной композиции были существенные технические ограничения по стабильности и совместимости компонентов, самым наглядным проявлением которых являлся факт неизбежности двух раздельных инъекций.
PEV6C позволила преодолеть технические ограничения PEV6A/B. PEV6C сочетает все три антигена (Р6, Р7 и Р4; SEQ ID NO: 1-3; соответственно) в одной пептидной цепи (см. также WO 2011/020604 А1; Multi-epitope vaccine for her2/neu-associated cancers (мультиэпитопная вакцина против her2/neu-ассоциированных видов рака)) и может быть изготовлена в стабильной лиофилизируемой форме для однократного введения.
Получение и результаты
Для конъюгации с белком-носителем CRM 197 использовали следующие слитые белки:
пептид 1: Р467 (до конъюгации):
ацетатная соль H-Pro-Glu-Ser-Phe-Asp-Gly-Asp-Pro-Ala-Ser-Asn-Thr-Ala-Pro-Leu-Gln-Pro-Arg-Val-Leu-Gln-Gly-Leu-Pro-Arg-Glu-Tyr-Val-Asn-Ala-Arg-His-Ser-Leu-Pro-Tyr-Met-Pro-Ile-Trp-Lys-Phe-Pro-Asp-Glu-Glu-Gly-Ala-Cys (SEQ ID NO: 8); и
пептид 2: P647 (до конъюгации):
ацетатная соль H-Arg-Val-Leu-Gln-Gly-Leu-Pro-Arg-Glu-Tyr-Val-Asn-Ala-Arg-His-Ser-Pro-Glu-Ser-Phe-Asp-Gly-Asp-Pro-Ala-Ser-Asn-Thr-Ala-Pro-Leu-Gln-Pro-Tyr-Met-Pro-Ile-Trp-Lys-Phe-Pro-Asp-Glu-Glu-Gly-Ala-Cys (SEQ ID NO: 9).
Конъюгацию проводили в PBS-буфере при рН 7,4 с использованием агента для перекрестного связывания сульфосукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилата (сульфо-SMCC). Этот реагент взаимодействует сначала с первичными аминами (то есть лизиновыми остатками) белка-носителя CRM197 перед тем, как ему позволяют взаимодействовать с тиоловыми группами (то есть цистеиновыми остатками). В CRM197 есть 39 лизиновых остатков, теоретически доступных для взаимодействия с агентом для перекрестного связывания. Для получения хорошего распределения пептидного антигена на белке-носителе, отношение «слитый пептид: белок-носитель» корректировали для конъюгации в среднем 20 молекул слитого пептида на одну молекулу CRM197. Взаимодействие проводили в течение 45 мин при комнатной температуре (к.т.) следующим образом.
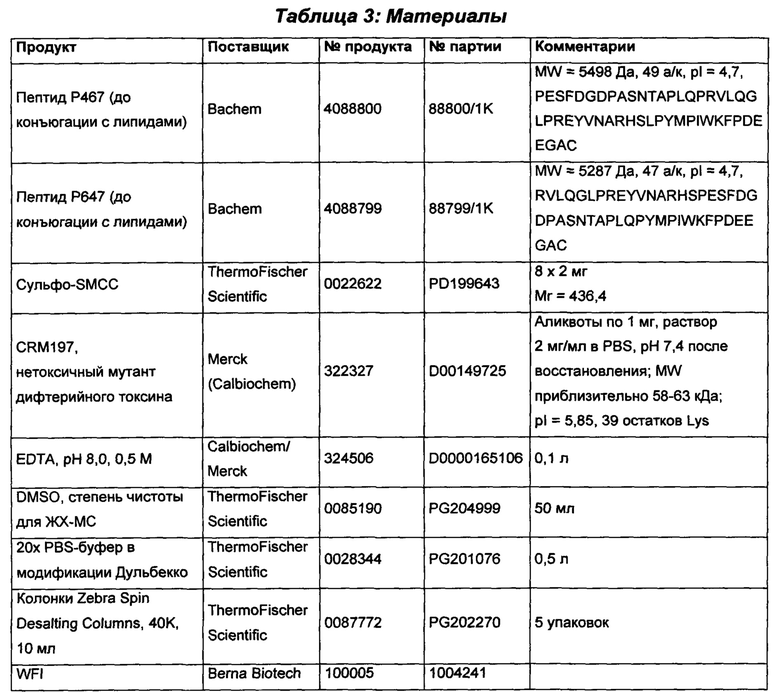
Методика
1. Достать слитые пептиды из -20°С и оставить при комнатной температуре (к.т.) приблизительно на 10 мин перед открытием флаконов.
2. Поместить навески слитых пептидов в стеклянные флаконы объемом 3 мл:
(а) Р467, 7,8 мг, ресуспендированный в 780 мкл воды для инъекций (WFI);
(б) Р467, 7,1 мг, ресуспендированный в 710 мкл диметилсульфоксида (DMSO);
(в) Р647, 6,9 мг, ресуспендированный в 690 мкл WFI;
(г) Р647, 9,0 мг, ресуспендированный в 900 мкл DMSO.
Все растворы легко ресуспендировали без видимых агрегатов или мутности.
3. Развести 20х забуференный фосфатом физиологический раствор (PBS) до 1х PBS водой Milli-Q и добавить этилендиаминтетрауксусную кислоту (EDTA) до конечной концентрации 1 мМ.
4. Восстановить 6 флаконов CRM197 в 0,5 мл WFI до 2 мг/мл. Объединить по 3 флакона с получением 2 партий и хранить при 4°С (1,5 мл на партию, по 3 мг в каждой).
5. Взять один флакон с сульфо-SMCC (2 мг), добавить в него 200 мкл PBS/1 мМ ETDA и растворить (исходный раствор 10 мг/мл; раствор оставался мутным).
6. Добавить 75 мкл сульфо-SMCC (10 мг/мл) к каждому флакону с 3 мг CRM197. Общий объем: 1,575 мл.
7. Инкубировать флаконы в течение 45 мин при к.т.с медленным перемешиванием (вращение, 25 об/мин). В это время уравновесить 2 колонки Zeba Spin Desalting Columns PBS/EDTA-буфером.
8. После инкубации на стадии (7) реакционную смесь переносят в уравновешенную колонку Zeba Spin Column (1,5 мл) и добавляют 200 мкл PBS/EDTA, после чего проводят центрифугирование при 1000 g в течение 4 мин.
9. Получали по 1,8 мл каждого раствора.
10. Внести в новые флаконы объемом 3 мл следующим образом:
(а) флакон 1: 600 мкл Р467 (10 мг/мл в WFI); и
(б) флакон 2: 600 мкл Р647 (10 мг/мл в WFI).
11. Добавить по 50 мкл 20х PBS в каждый флакон для повышения рН раствора приблизительно до 7.
12. Добавить по 1,8 мл обессоленного CRM197-SMCC в каждый флакон.
13. Инкубировать флаконы в течение 45 мин при к.т.с медленным перемешиванием (вращение), затем хранить флаконы при при 4°С до анализа. Во время инкубации во флаконе 2 был отмечен видимый осадок. Тем не менее, осадок исчез после нагревания раствора.
Заключение
Пептид P467-CRM197, приблизительно 2,449 мг/мл Р467, приблизительно 1,22 мг/мл CRM197, приблизительно 2450 мкл; в 1x PBS/1 мМ EDTA, партия 140829-1_am.
Пептид P647-CRM197, приблизительно 2,449 мг/мл Р647, приблизительно 1,22 мг/мл CRM197, приблизительно 2450 мкл; в 1x PBS/1 мМ EDTA, партия 140829-2_am.
Была отмечена склонность пептидного конъюгата P647-CRM197 к образованию агрегатов в процессе взаимодействия. В случае пептидного конъюгата P467-CRM197 видимой глазом агрегации не было.
Пример 2 - Окрашивание кумасси голубым и иммуноблот конъюгатов CRH/1197-антиген с использованием мышиной антисыворотки Е
Конъюгатные композиции анализировали посредством SDS-PAGE и оценивали окрашиванием геля кумасси голубым и иммуноблоттингом с использованием полученной ранее мышиной сыворотки против комбинации пептидных антигенов Р4, Р6 и Р7, конъюгированных со столбнячным анатоксином (антисыворотка Е).
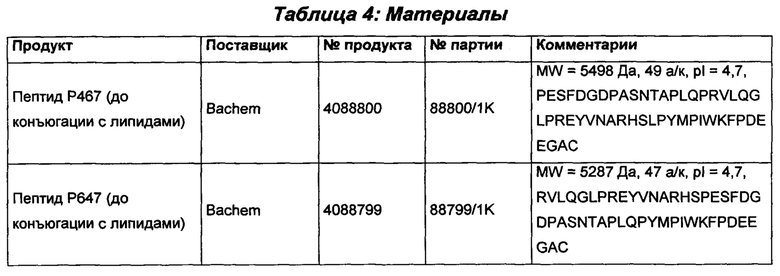
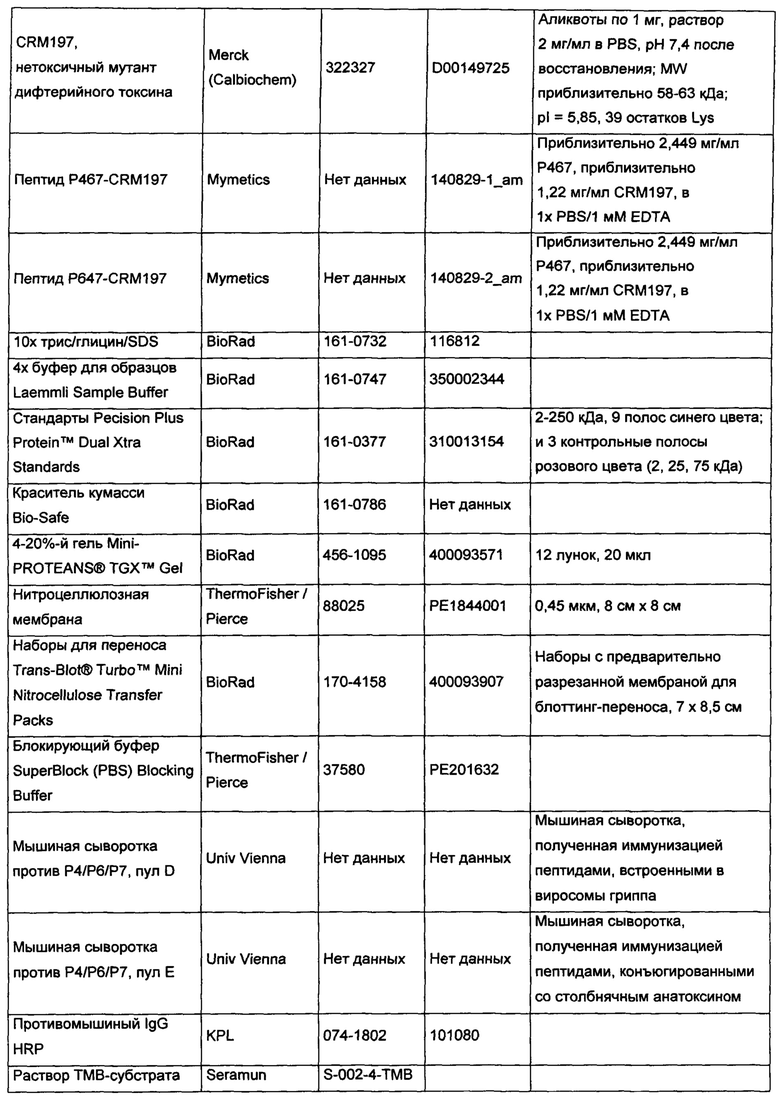

Методика
Получали два полиакриламидных геля с додецилсульфатом натрия (SDS-PA) следующим образом:
- исходный раствор CRM197 (2 мг/мл) разводили 1:10 в PBS, рН 7,4;
- антиконъюгатные растворы разводили 1:10 в PBS, рН 7,4;
- неконъюгированные слитые пептиды готовили в форме исходных растворов, 10 мг/мл, и разводили 1:20 в PBS, рН 7,4.
В общей сложности готовили по 36 мкл каждого образца и вносили по 15 мкл на каждую дорожку. Получали два одинаковых геля. Гель 1 окрашивали кумасси голубым и гель 2 использовали для вестерн-блота. Расположение образцов в геле 1 и геле 2 указано в Таблице 5 ниже.
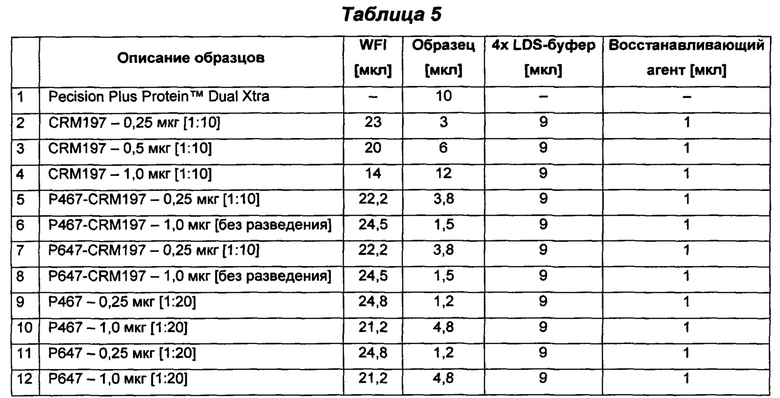
Результаты Гель 1 - окрашивание кумасси голубым
Гель 1 погружали в краситель кумасси голубой Bio-Safe на 1 ч при к.т.с покачиванием, после чего промывали водой Milli-Q в течение 1 ч при к.т.
Гель 2 - Вестерн-блот
После электрофореза гель 2 извлекали из кассеты и проводили короткую промывку в воде Milli-Q. Затем готовили сэндвич для переноса, следуя инструкциям изготовителя (BioRad), и устанавливали его в прибор Turbo Transfer Blot. После переноса нитроцеллюлозную мембрану проверяли на предмет полного присутствия предварительно окрашенных маркерных белков. Лишь минимальные количества маркеров оставались видны на SDS-PA-геле. Затем мембрану инкубировали в буфере SuperBlock в течение 1 ч при к.т. Мышиную сыворотку пула Е разводили 1:500 в SuperBlock и инкубировали на мембране в течение 2 ч при к.т. Затем мембрану промывали три раза с использованием 1x PBS, по 5 мин каждый раз. После этого мембрану инкубировали с противомышиным IgG (KPL, 074-1802; Ab20), разведенным 1:5000 в SuperBlock, в течение 1 ч при к.т. Затем мембрану промывали три раза с использованием 1х PBS, по 5 мин каждый раз, и проявляли 3,3',5,5'-тетраметилбензидином (ТМВ) в течение 5 мин при к.т.
Результаты продемонстрировали явное изменение молекулярной массы для каждого конъюгата CRM197 в сторону большей молекулярной массы, указывая на выраженную конъюгацию слитых пептидов с CRM197. Широкие полосы конъюгатов, отмеченные в обоих случаях, также указывают на относительно большой разброс по слитым пептидам на белке-носителе, как следовало ожидать после реакции конъюгации. Результаты также указывают на отсутствие (или минимальное количество) оставшегося неконъюгированного белка CRM197. С другой стороны, была отмечена незначительная фракция слитых пептидов, составлявшая, согласно оценкам, менее 10% от общего количества использованных слитых пептидов.
Выводы
Было отмечено явное смещение конъюгатов CRM197, указывавшее на успешную конъюгацию слитых пептидов Р467 и Р647 с белком-носителем CRM197.
Значимых различий между конъюгатами P467-CRM197 и P647-CRM197 отмечено не было.
В обеих реакциях конъюгации наблюдали незначительное количество свободных слитых пептидов.
Иммуноблот с использованной антисывороткой продемонстрировал некоторую перекрестную реактивность с дифтерийным токсином CRM197, несмотря на то, что она была получена с использованием конъюгатов пептидов со столбнячным анатоксином. Он подтверждает смещение молекул CRM197 после конъюгации, но, поскольку он не является специфичным для двух рассматриваемых слитых пептидов, не позволяет подтвердить их конъюгацию с белком-носителем. Антисыворотка окрашивает свободные неконъюгированные слитые пептиды, Р467 несколько лучше, чем Р647.
Пример 3 - Иммуноблот конъюгатов CRH/1197-антиген с использованием мышиной антисыворотки D
Конъюгатные композиции анализировали посредством SDS-PAGE и оценивали иммуноблоттингом с использованием полученной ранее мышиной сыворотки против комбинации пептидных антигенов Р4, Р6 и Р7, конъюгированных с виросомами гриппа (антисыворотка D). Присутствие слитых пептидов Р467 и Р647 на белке-носителе CRM197 подтверждали по отдельности иммуноблотом с использованием антисыворотки D.
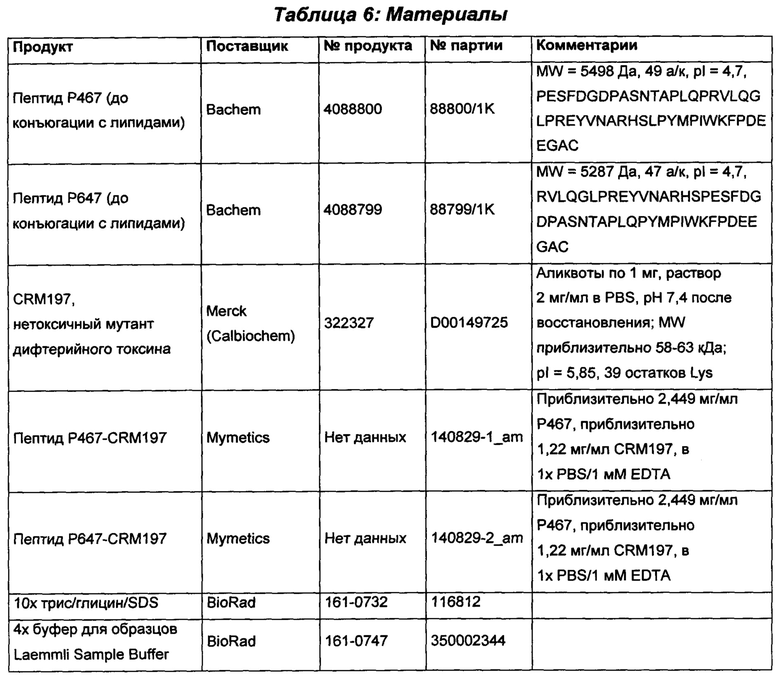
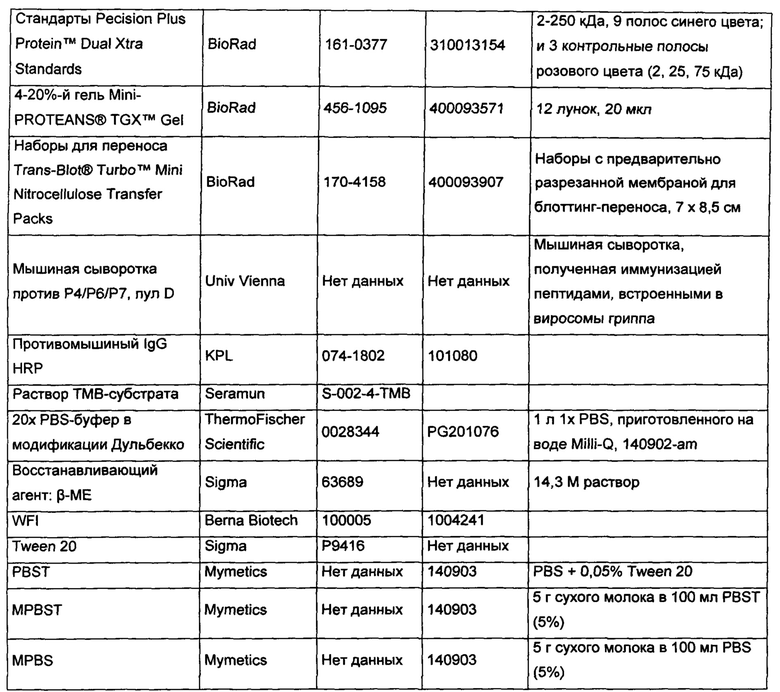
Методика
Получали один SDS-PA-гель следующим образом.
Все растворы, содержащие CRM197, конъюгаты слитых пептидов с CRM197 или неконъюгированные слитые пептиды разводили до 1 мг/мл в PBS при рН 7,4. В общей сложности готовили по 36 мкл каждого образца и вносили по 15 мкл на каждую дорожку. Расположение образцов в геле 1 и геле 2 указано в Таблице 7 ниже.
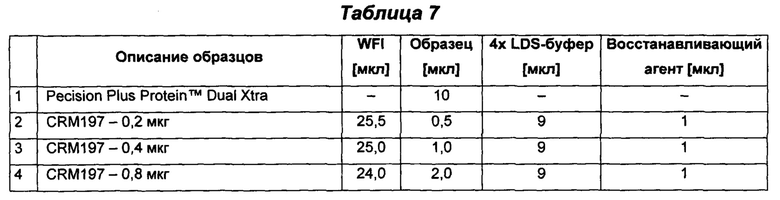
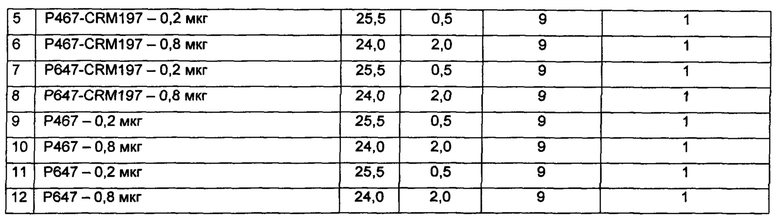
Результаты
Вестерн-блот. После электрофореза гель извлекали из кассеты и проводили короткую промывку в воде Milli-Q. Готовили сэндвич для переноса, следуя инструкциям изготовителя (BioRad), и устанавливали его в прибор Turbo Transfer Blot. После переноса нитроцеллюлозную мембрану проверяли на предмет полного присутствия предварительно окрашенных маркерных белков. Лишь минимальные количества маркеров оставались видны на SDS-PA-геле. Затем нитроцеллюлозную мембрану инкубировали в 5%-м молоке в забуференном фосфатом физиологическом растворе с Tween 20 (MPBST) в течение 1 ч при к.т.
Мышиную сыворотку пула D разводили 1:500 в MPBST и инкубировали на мембране в течение 1 ч при к.т. Затем мембрану промывали три раза с использованием 1х MPBST, по 5 мин каждый раз, после чего инкубировали с противомышиным IgG (KPL, 074-1802; Ab20), разведенным 1:5000 в MPBST, в течение 30 мин при к.т.Затем мембрану промывали три раза с использованием 1х MPBST, по 5 мин каждый раз, и проявляли раствором ТМВ в течение 10 мин при к.т.
Выводы
Иммуноблот с использованной антисывороткой D продемонстрировал реактивность только в отношении конъюгатов CRM197 со слитыми пептидами. Перекрестной реактивности в отношении неконъюгированного CRM 197 отмечено не было. Эти результаты показывают, что значительное количество слитых пептидов было конъюгировано с белком-носителем, и подтверждают, что смещение в миграции по гелю для конъюгатов CRM197 со слитыми пептидами обусловлено конъюгацией слитых пептидов с белком-носителем, а не межмолекулярным перекрестным связыванием CRM197. Сигнала, соответствующего свободным неконъюгированным слитым пептидам, выявлено не было, либо из-за того, что антисыворотка с ними не взаимодействует, либо из-за того, что слитые пептиды мигрировали до конца геля и не были перенесены на нитроцеллюлозную мембрану.
Антисыворотка D взаимодействовала с конъюгатом P647-CRM197 несколько сильнее, чем с конъюгатом P467-CRM197. Результаты показали, что с конъюгированными слитыми пептидами антисыворотка взаимодействовала сильнее, чем со свободными слитыми пептидами.
Пример 4 - Спот-блот P467-CRM197 и P647-CRM197
Присутствие слитых пептидов Р467 и Р647 на белке-носителе CRM 197 также подтверждали в спот-блот-эксперименте с использованием антисыворотки D.
Образцы
- CRM 197, 1 мг/мл в PBS, рН 7,4;
- антиген Р467, 1 мг/мл в PBS, рН 7,4;
- антиген Р647, 1 мг/мл в PBS, рН 7,4;
- P467-CRM197, 1 мг/мл в PBS, рН 7,4 (расчетная концентрация); и
- P647-CRM197, 1 мг/мл в PBS, рН 7,4 (расчетная концентрация).
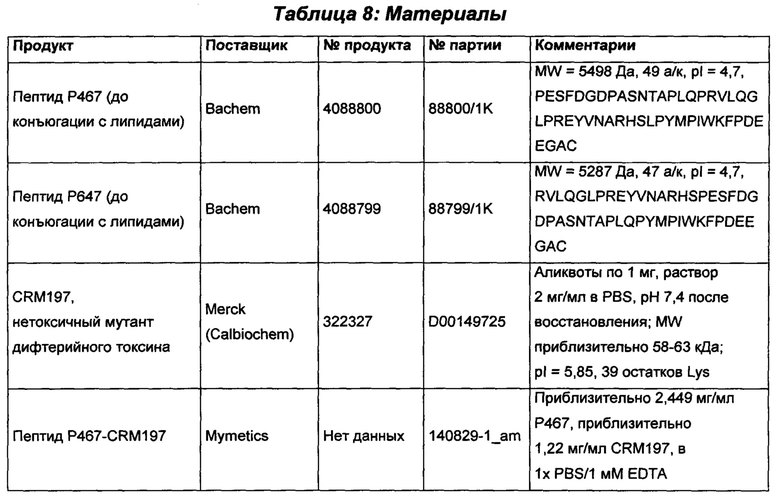
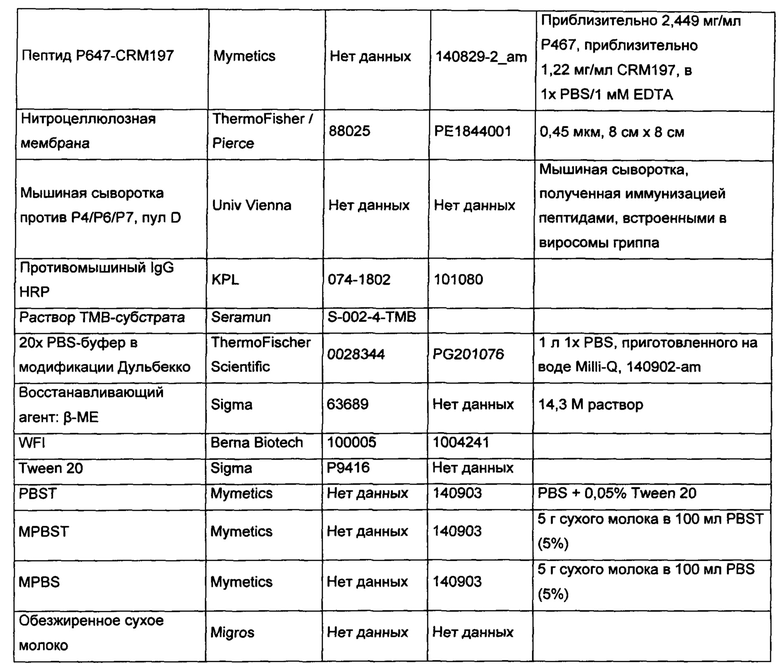
Методика
Использовали предварительно разрезанную нитроцеллюлозную мембрану и дорожки делали карандашом. Проводили двукратные разведения каждого образца в PBS при рН 7,4 в пробирках типа Эппендорф, начиная с 1 мг/мл (1 мкг/мкл). Теоретические концентрации в каждой точке (1 мкл) составляли 1 мкг / 500 нг / 250 нг / 125 нг / 62,5 нг / 31,25 нг. В каждой точке приблизительно 1 мкл каждого образца переносили на мембрану (в двух повторах), как указано ниже:
- дорожка А - CRM 197;
- дорожка В - антиген Р467 (свободный слитый пептид без конъюгации с липидом);
- дорожка С - антиген Р647 (свободный слитый пептид без конъюгации с липидом);
- дорожка D P467-CRM197 (начиная с 1 мкг слитого пептида и 0,5 мкг CRM197); и
- дорожка Е P647-CRM197 (начиная с 1 мкг слитого пептида и 0,5 мкг CRM197).
Мембране давали высохнуть на воздухе в течение 10 мин при к.т. и затем блокировали 5%-м молоком в PBST (MPBST) в течение 30 мин при к.т. После этого мембрану инкубировали с первичным антителом (мышиной антисывороткой пула D, разведенной 1:500 в MPBST) в течение 1 ч при к.т. Затем мембрану промывали три раза с использованием MPBST, по 5 мин каждый раз, после чего инкубировали с вторичным антителом (противомышиным IgG; KPL, 074-1802; Ab20; разведенным 1:5000 в MPBST) в течение 30 мин при к.т. После этой инкубации мембрану промывали три раза с использованием MPBST, по 5 мин каждый раз, и затем инкубировали с раствором ТМВ при к.т. в течение приблизительно 3 мин с последующей промывкой водой Milli-Q перед сушкой.
Выводы
Спот-блот с мышиной антисывороткой D продемонстрировал реактивность только в отношении конъюгатов CRM197 со слитыми пептидами. Перекрестная реактивность в отношении неконъюгированного белка CRM197 была минимальна. Поскольку количество CRM197 в конъюгатах составляло приблизительно 50% по сравнению с количеством слитого пептида, минимальный фон в этом образце был, вероятно, обусловлен перекрестной реактивностью в отношении варианта дифтерийного токсина.
Свободные (неконъюгированные) слитые пептиды (слитые пептиды без липидного конъюгата) давали сигнал с антисывороткой только в случае Р647, но не Р467. Причина этого не вполне ясна. Тем не менее, в первых разведениях образца Р647 сигнал был несколько слабее, чем в последующих разведениях. Это может указывать на присутствие ингибитора сайта связывания с антителом или на уменьшение связывания с нитроцеллюлозной мембраной. Природа этого ингибитора неизвестна, но он может быть связан с DMSO, который был исходно использован для получения высоко концентрированного исходного раствора слитого пептида (10 мг/мл).
Мышиная антисыворотка D очень сильно взаимодействовала с конъюгатами P467-CRM197 и P647-CRM197, подтверждая конъюгацию большей части слитых пептидов с белком-носителем.
Из-за разной реактивности сыворотки в отношении свободных слитых пептидов и конъюгатов слитых пептидов с CRM197 определить количество антигена было невозможно. Тем не менее, ввиду отсутствия стадий очистки после реакции конъюгации, его можно рассчитать по теоретическим начальным концентрациям слитых пептидов.
В завершение, результаты показывают, что с конъюгированными слитыми пептидами антисыворотка взаимодействовала сильнее, чем со свободными слитыми пептидами.
Пример 5 - Получение образцов для исследований по титрованию дозы у мышей
Далее описано получение конъюгатов слитых пептидов с CRM197 для экспериментов по титрованию дозы у мышей.
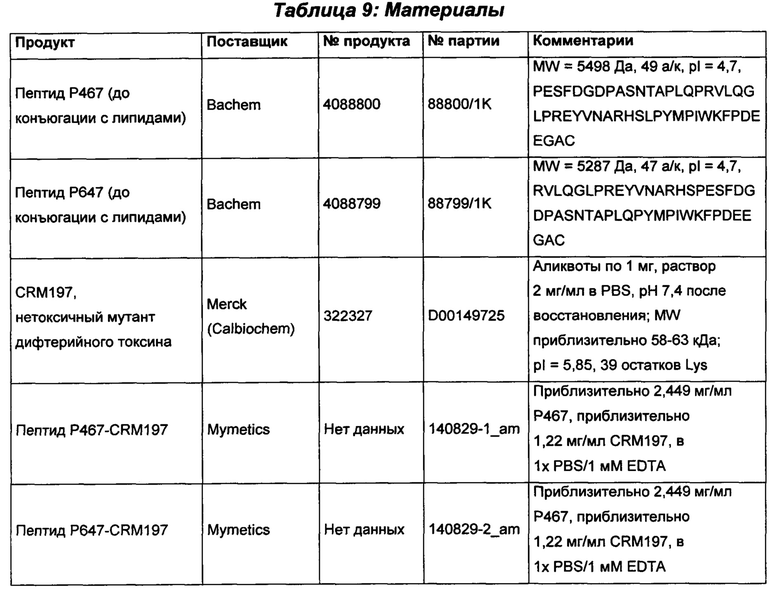
Методика
Для получения образцов для исследований у животных реакционные растворы конъюгатов разводили до определенной целевой концентрации. Предполагали, что для следующих препаратов эффективность конъюгации пептида с белком составляла по меньшей мере 90%.
Получение образцов для групп высокой дозы (группа 1)
Предполагаемое содержание конъюгированного слитого пептида в этих образцах составляло приблизительно 420 мкг на пробирку (1,2 мг/мл).
Пробирка 1:
- 1045 мкл P467-CRM197;
- 880 мкл (1х) PBS, рН 7,4.
Этот раствор тщательно перемешивали, переносили по 350 мкл в пять стеклянных флаконов (по 2,5 мл каждый) и отмечали следующим образом: P467-CRM197; группа 1 - высокая доза; расчетная концентрация Р467 приблизительно 1,2 мг/мл; партия 140903-1_am.
Пробирка 2:
- 1045 мкл P647-CRM197 (интенсивно перемешанный на вихревой мешалке перед пипетированием);
- 880 мкл (1х) PBS, рН 7,4.
Этот раствор тщательно перемешивали, переносили по 350 мкл в пять стеклянных флаконов (по 2,5 мл каждый) и отмечали следующим образом: P647-CRM197; группа 1 - высокая доза; расчетная концентрация Р647 приблизительно 1,2 мг/мл; партия 140903-4_am.
Получение образцов для групп средней дозы (группа 2)
Предполагаемое содержание конъюгированного слитого пептида в этих образцах составляло приблизительно 210 мкг на пробирку (0,6 мг/мл).
Пробирка 1:
- 525 мкл P467-CRM197;
- 1400 мкл (1х) PBS, рН 7,4.
Этот раствор тщательно перемешивали, переносили по 350 мкл в пять стеклянных флаконов объемом 2,5 мл и отмечали следующим образом:P467-CRM 197; группа 2 - средняя доза; расчетная концентрация Р467 приблизительно 0,6 мг/мл; партия 140903-2_am.
Пробирка 2:
- 525 мкл P647-CRM197 (интенсивно перемешанный на вихревой мешалке перед пипетированием);
- 1400 мкл (1х) PBS, рН 7,4.
Этот раствор тщательно перемешивали на вихревой мешалке, переносили по 350 мкл в пять стеклянных флаконов объемом 2,5 мл и отмечали следующим образом: P647-CRM197; группа 2 - средняя доза; расчетная концентрация Р647 приблизительно 0,6 мг/мл; партия 140903-5_am.
Получение образцов для групп низкой дозы (группа 3)
Предполагаемое содержание конъюгированного слитого пептида в этих образцах составляло приблизительно 105 мкг на пробирку (0,3 мг/мл).
Пробирка 1:
- 262 мкл P467-CRM197;
- 1663 мкл (1х) PBS, рН 7,4.
Этот раствор тщательно перемешивали, переносили по 350 мкл в пять стеклянных флаконов объемом 2,5 мл и отмечали следующим образом: P467-CRM197; группа 3 - низкая доза; расчетная концентрация Р467 приблизительно 0,3 мг/мл; партия 140903-3_am.
Пробирка 2:
- 262 мкл P647-CRM197 (интенсивно перемешанный на вихревой мешалке перед пипетированием);
- 1663 мкл (1х) PBS, рН 7,4.
Этот раствор тщательно перемешивали на вихревой мешалке, переносили по 350 мкл в пять стеклянных флаконов объемом 2,5 мл и отмечали следующим образом: P647-CRM197; группа 3 - низкая доза; расчетная концентрация Р647 приблизительно 0,3 мг/мл; партия 140903-6_am.
Пример 6 - Иммунизационные исследования Пептидные антигены
Два слитых пептида Р467 (SEQ ID NO: 8) и Р647 (SEQ ID NO: 9) были синтезированы в Bachem (Швейцария). Оба гибрида были связаны с виросомами (вирусными оболочками инактивированного вируса гриппа A/Brisbane/59/2007), являющимися подходящей системой доставки антигенов, не требующей дополнительных адъювантов. Анализировали три концентрации, рассчитанные по количеству слитого пептида: 15 мкг, 30 мкг и 50 мкг на инъекцию. Для сравнения оба пептида были связаны с CRM197 и использованы в параллельном эксперименте в концентрации 30 мкг на инъекцию. Все процедуры конъюгации (с CRM и виросомами) были проведены в Mymetics (Швейцария).
Виросомные композиции доставляли замороженными, и они образовывали мутноватую суспензию при размораживании.
После связывания конъюгат CRM-P647 получали в форме осажденной композиции, в то время как CRM-P467 оставался растворимым в водном буфере.
Три В-клеточных эпитопа, имеющих происхождение от Her2/neu, Р4 (SEQIDNO:3), Р6 (SEQ ID NO: 1) и Р7 (SEQ ID NO: 2), были синтезированы и связаны с KLH (гемоцианином фиссуреллы, keyhole limpet hemocyanin) в piChem (Австрия), и они были использованы в качестве сорбируемых антигенов для оценки гуморальных иммунных ответов на отдельные пептиды посредством ELISA.
Для оценки гуморальных иммунных ответов, направленных на неконъюгированные слитые пептиды, в качестве сорбируемых антигенов в ELISA-анализах использовали Р467 и Р647.
Протокол иммунизации
Самок мышей Balb/C иммунизировали подкожно с 2-3-недельными интервалами в соответствии со следующим планом исследования.
Группы, получавшие конъюгаты с виросомами, примировали однократно инактивированным вирусом гриппа (инактивированным A/Brisbane/59/2007). Иммунизации конъюгированными конструкциями начинали через 16 дней после примирования. Все виросомные композиции доставляли готовыми к применению и использовали без каких-либо добавок по 100 мкл на инъекцию.
Перед введением инъекцией исходные растворы конъюгатов CRM со слитыми пептидами разбавляли раствором NaCl и смешивали с гелевой суспензией гидрата окиси алюминия. CRM-P647 (осажденный) перемешивали на вихревой мешалке и суспензию использовали так же, как CRM-P467.
Контрольные группы получали пустые виросомы (TIRIV) или раствор NaCI с гидратом окиси алюминия.
Животные
60 самок мышей Balb/C, 6-8 недель на дату доставки (29 октября 2014 г.), источник: Charles River, Германия.
10 групп: n = 5 мышей на группу. Виварий: General Hospital Vienna/Biomedical Unit.
Группы
- Группа А - пустые виросомы (TIRIV) - 100 мкл на мышь;
- группа В - Р467-виросомы - 50 мкг на мышь в 100 мкл;
- группа С - Р467-виросомы - 30 мкг на мышь в 100 мкл;
- группа D - Р467-виросомы - 15 мкг на мышь в 100 мкл;
- группа Е - Р647-виросомы - 50 мкг на мышь в 100 мкл;
- группа F - Р647-виросомы - 30 мкг на мышь в 100 мкл;
- группа G - Р647-виросомы - 15 мкг на мышь в 100 мкл;
- группа Н - NaCl + гидрат окиси алюминия (контроль) - 100 мкл на мышь;
- группа I - P467-CRM + гидрат окиси алюминия - 30 мкг на мышь в 150 мкл; и
- группа J - P467-CRM + гидрат окиси алюминия - 30 мкг на мышь в 150 мкл.
Забор крови проводили перед иммунизацией, через три недели после 2-й иммунизации, через две недели после 3-й иммунизации и через две недели после 4-й иммунизации. Затем образцы крови анализировали на предмет:
- титров пептид-специфичных антител (IgG);
- титров антител, специфичных в отношении конструкций (IgG);
- титров антител, специфичных в отношении рекомбинантного внеклеточного Her2/neu (IgG); и
- специфичности в отношении нативного Her2/neu, экспрессированного в раковых клетках SKBR-3.
Протоколы ELISA
(1) Пептид-специфичный ELISA
На микротитрационные планшеты сорбировали конъюгаты KLH со слитыми пептидами в количестве 0,5 мкг на лунку. Для оценки фона на лунки сорбировали KLH сам по себе. После блокировки добавляли разведенные сыворотки. Связанный IgG выявляли антителом, меченным пероксидазой хрена (HRP) (кроличий противомышиный IgG POX, специфичный в отношении Fc-фрагмента, №315-035-008; Jackson Immuno Research) с последующим окрашиванием ТМВ. Планшеты считывали после добавления останавливающего раствора при 450-630 нм.
(2) ELISA, специфичный в отношении слитых пептидов
В качестве сорбируемых антигенов использовали немодифицированные слитые пептиды Р467 и Р647 в концентрации 0,5 мкг на лунку в карбонатном буфере. Затем добавляли разведенные сыворотки и анализ проводили, как описано в пептид-специфичном протоколе выше.
(3) ELISA. специфичный в отношении внеклеточного Her2/neu
В качестве сорбируемых антигенов использовали слитый белок, содержащий рекомбинантный внеклеточный домен человеческого Her2/neu (аминокислотные остатки 23-652 Her2/neu) и Fc-область человеческого lgG1 (химерный ErbB2/Fc, номер по каталогу # 1129-ER, R&D Systems). На планшеты сорбировали белок, 0,1 мкг на лунку, и 0,1%-й бычий сывороточный альбумин (BSA) для оценки фона. Затем добавляли разведенные сыворотки и проводили анализ, как описано выше, вычитая сигнал фона, полученный в лунках, где был только BSA.
(4) Клеточный анализ, специфичный в отношении Her2/neu, с использованием клеток SKBR-3
В качестве источника нативного белка Her2/neu использовали клеточную линию рака молочной железы человека SKBR-3 (НТВ30, АТСС, USA), сверхэкспрессирующую Her2/neu. Клетки собирали через три дня после пассажа и хранили при -80°С. Лизис клеток проводили, как описано ранее в Godell V and Disis ML J. Immunol. Meth (2005), с некоторыми незначительными изменениями. Вариант 1: на планшеты сорбировали гуманизированное антитело против Her2/neu герцептин (предоставленное General Hospital Vienna). Вариант 2: на планшеты сорбировали гуманизированное антитело против Her2/neu перьета (предоставленное General Hospital Vienna). Содержание белка в клеточных лизатах анализировали количественно с использованием набора Pierce ВСА Protein Assay Kit # 23227 непосредственно перед применением. Лизаты разбавляли до концентрации 100 мкг на лунку 1% BSA/PBS. После блокировки клеточные лизаты добавляли в лунки. Для контроля индивидуального фона добавляли BSA/PBS. Для выявления IgG использовали связанный с HRP овечий противомышиный IgG (Amersham/GE Healthcare). Окрашивание ТМВ проводили, как описано выше. Для каждого образца сыворотки значение OD рассчитывали как разность показателя OD SKBR-3-лунок и лунок с сорбированным BSA.
Пример 7 - Пептид-специфичные антитела
Использовали сыворотки, полученные при последнем заборе крови (то есть после четырех иммунизаций). Сыворотки мышей, иммунизированных виросомами и CRM197, анализировали в разных разведениях, как указано ниже:
- группы, которым вводили виросомные композиции: разведение 1:5000;
- группы, которым вводили конъюгаты слитых пептидов с CRM: разведение 1:100000.
Данные показаны как значения OD. Вычитали фон, отмеченный в лунках с сорбированным KLH (см. Фиг. 1-3).
Результаты
Антитела против всех трех отдельных пептидов были получены во всех группах введения (группах B-G и I-J). В контрольных группах (группах А и Н) пептид-специфичных антител выявлено не было. Были отмечены существенные различия по титрам антител.
После четырех иммунизаций конъюгатами слитых пептидов с CRM титры антител ко всем трем отдельным пептидам были приблизительно в десять раз выше, чем после четырех иммунизаций соответствующими виросомными композициями.
Для обеих конструкций (Р467 и Р647), независимо от партнера по связыванию, наиболее высоким был титр антител против В-клеточного эпитопа Р7 (SEQ ID NO: 2), в то время как наиболее низким был титр антител против В-клеточного эпитопа Р4 (SEQ ID NO: 3).
Как показано на Фиг. 4 и 5, также были получены антитела, связывавшиеся со слитыми пептидами. Эти результаты были получены в экспериментах, в которых использовали сыворотки, полученные при последнем заборе крови после четвертой иммунизации. Для групп, иммунизированных виросомами, сыворотки разводили 1:50000. Для групп, иммунизированных конъюгатами слитых пептидов с CRM 197, сыворотки разводили 1:100000. Данные, показанные на Фиг. 4 и 5, представлены как значения OD. Аналогично пептид-специфичным результатам, показанным на Фиг. 1-3, в группах, иммунизированных конъюгатами слитых пептидов с CRM197, титры антител были приблизительно в десять раз выше, чем в группах, иммунизированных соответствующими виросомными композициями. Обычно для Р647 наблюдали более высокие титры, чем для Р467.
Пример 8 - Динамика антител во время иммунизации CRM-конъюгатами
Забор крови проводили через две недели после 2-й, 3-й и 4-й иммунизации и определяли антитела, специфичные в отношении обоих слитых пептидов (Р467 и Р647) (сыворотки разводили 1:100000). Как показано на Фиг. 6 и 7, титры антител, специфичных в отношении слитых белков, были достаточно высокими уже после 2-й иммунизации и несколько возрастали после 3-й и 4-й иммунизации. Малый рост титров антител, специфичных в отношении слитых белков, после 3-й и 4-й иммунизации указывает на то, что двух или трех иммунизаций (то есть двух или трех последовательных введений) конъюгатом слитого пептида с CRM197 может быть достаточно для получения эффективного иммунного ответа. (ВА1 - забор крови после двух иммунизаций; ВА2 - забор крови после трех иммунизаций; ВА3 - забор крови после четырех иммунизаций.)
Пример 9 - Антитела, специфичные в отношении внеклеточного Her2/neu
Забор крови проводили в двух временных точках: ВА1 (забор крови после двух иммунизаций) и ВА2 (забор крови после трех иммунизаций). Сыворотки анализировали в разных разведениях - сыворотки от групп, иммунизированных виросомными композициями, разводили 1:400, и сыворотки от групп, иммунизированных конъюгатами слитых пептидов с CRM, разводили 1:2000. Данные, показанные на Фиг. 8, представляют собой значения OD.
Как показано на Фиг. 8, обе конструкции со слитыми пептидами (конъюгированными с виросомами или CRM197) приводили к выработке антител, связывавшихся с рекомбинантным внеклеточным белком Her2/neu. Существенной реактивности в отношении Her2/neu не было отмечено ни у мышей, получавших отрицательный контроль, ни в сыворотках, полученных до иммунизации. Согласно результатам анализов, специфичных в отношении отдельных пептидов и слитых пептидов, Her2/neu-специфичные титры, отмеченные в группах, иммунизированных конъюгатами слитых пептидов с CRM, были значительно выше, чем при использовании виросомных композиций при концентрации 30 мкг. Значимый титр антител также наблюдали после третьей иммунизации виросомами. У мышей, которых иммунизировали конъюгатами слитых пептидов с CRM, титры антител были высокими уже после 2-й иммунизации без существенного повышения после 3-й иммунизации.
Анализ титров антител в сыворотках после 2-й, 3-й и 4-й иммунизации выявил разную динамику гуморальных иммунных ответов при использовании виросом и CRM-конъюгатов. После 2-й иммунизации (через 6 недель после начала иммунизации) у животных, иммунизированных конъюгатами слитых пептидов с CRM, были отмечены более высокие титры антител, чем при использовании виросомных конъюгатов (см. Фиг. 9). После 3-й иммунизации виросомным конъюгатом наблюдали существенный рост титра антител (см. Фиг. 8). После 4-й иммунизации титры антител, специфичных в отношении Her2/neu, у мышей, иммунизированных виросомами и CRM, были сопоставимы (см. Фиг. 10). У мышей, иммунизированных конъюгатами слитых пептидов с CRM, рост титра антител между 3-й и 4-й иммунизацией был незначительным при сильной иммуногенности после 2-й иммунизации с еще более сильной иммуногенностью после 3-й иммунизации (см. Фиг. 11).
Как показано на Фиг. 12, сравнение динамики гуморальных иммунных ответов против рекомбинантного Her2/neu после 2-й, 3-й и 4-й иммунизации с использованием 30 мкг виросомного конъюгата Р467 или 30 мкг конъюгата P467-CRM демонстрирует, что даже после 2-й иммунизации титр антител против Her2/neu после иммунизации CRM-конъюгатом (четвертый столбик в каждой группе) был существенно выше титра антител после иммунизации соответствующим виросомным конъюгатом (третий столбик в каждой группе). После 4-й иммунизации оба конъюгата приводили к сопоставимым титрам антител против рекомбинантного Her2/neu.
Как показано на Фиг. 13, сравнение динамики гуморальных иммунных ответов против рекомбинантного Her2/neu после 2-й, 3-й и 4-й иммунизации с использованием 30 мкг виросомного конъюгата Р647 или 30 мкг конъюгата P647-CRM демонстрирует, что даже после 2-й иммунизации титр антител против Her2/neu после иммунизации CRM-конъюгатом (четвертый столбик в каждой группе) был существенно выше титра антител после иммунизации соответствующим виросомным конъюгатом (третий столбик в каждой группе).
Эти результаты демонстрируют разительное отличие динамики титров Her2/neu-специфичных антител при использовании виросом в качестве системы доставки слитых пептидов от соответствующей динамики при использовании CRM197. Повышение титров антител, отмеченное при использовании виросомных конъюгатов, было значительно медленнее, чем при использовании CRM-конъюгатов, и для достижения уровней, сходных с соответствующими CRM-иммунизациями, обычно было необходимо четыре иммунизации (применительно к слитому пептиду Р467). Применительно к слитому пептиду Р647, виросомные конъюгаты уступали CRM-конъюгатам, поскольку не обеспечивали сопоставимых с ними титров антител даже после 4-й иммунизации (Фиг. 13).
Пример 10 - Специфичность в отношении нативного белка Her2/neu
Анализировали сыворотки, полученные при последнем заборе крови после четырех иммунизаций. Данные представлены как значения OD, и все сыворотки разводили 1:100.
Как показано на Фиг. 14, полученные антитела распознавали нативную форму Her2/neu, сверхэкспрессированную в клетках рака молочной железы SKBR-3. Малые разведения сыворотки были необходимы для работы в ELISA-диапазоне с неочищенным лизатом клеток и соответствовали данным, полученным авторами изобретения ранее. Несмотря на существенно меньшие титры антител против пептидов и рекомбинантного внеклеточного белка Her2/neu в группах, иммунизированных виросомными конъюгатами, значения OD, полученные для конъюгатов слитых пептидов с виросомами и CRM197, были сопоставимыми. Эти результаты можно объяснить реактивностью антител, выработанных против липидной связи или некоторых частей виросом, в отношении компонентов SKBR-3-клеточных лизатов (например, клеточных мембран). В конъюгатах слитых пептидов с CRM197 такая связь отсутствует.
Пример 11 - Оценка иммунных ответов после иммунизации пептидом P467-CRM с Alum/Al(OH)3 или Montanide
Подготовка реагентов
Слитый белок Р467 (49 аминокислот в длину; синтезированный в Bachem, Швейцария) конъюгировали с CRM197 (мутантным дифтерийным токсином; PiChem/Грац/Австрия) и поставляли в исходной концентрации 0,4 мг/мл. Иммунный ответ, опосредованный P467-CRM, оценивали у мышей в присутствии или в отсутствие трех адъювантов:
1) ALHYDROGEL®"85" (гидрат окиси алюминия от Brenntag, Дания; содержание алюминия 10 мг/мл, партия №85563; использовался в разведении 1/10 в соответствии с инструкциями изготовителя);
2) суспензия Alu-Gel-S (гидрат окиси алюминия от Serva, Германия; степень чистоты 1,3%, стерильный, содержание алюминия 5,9-7,1 мг/мл, № по каталогу 12261, партия №111589; использовался в разведении 1/3 в соответствии с инструкциями изготовителя); и
3) Montanide ISA 51VG (от Seppic, Франция, №по каталогу 36362ZFL2R3; партия №2423395).
Протокол иммунизации описан ниже в разделе «Протокол иммунизации». Кратко, анализировали три концентрации пептидной конструкции: 10 мкг, 25 мкг и 50 мкг на инъекцию. Гидрат окиси алюминия от Serva, который был использован в исследованиях на животных, описанных в Примерах выше, анализировали в двух группах мышей (см. раздел «Протокол иммунизации») и сравнивали с использованием гидрата окиси алюминия от Brenntag. Количество P467-CRM, использованное с гидратом окиси алюминия от Serva в этих двух группах, составляло 10 мкг и 25 мкг.
Эмульгирование пептидной конструкции с использованием Montanide
Эмульгирование P467-CRM, PBS или NaCl с использованием Montanide проводили в соответствии с инструкциями изготовителя (Seppic, Франция).
Пептидную конструкцию P467-CRM смешивали с PBS (для гидрата окиси алюминия, Brenntag) или NaCl (для гидрата окиси алюминия, Serva). Затем в раствор добавляли адъюванты и тщательно перемешивали при комнатной температуре. Затем смеси держали при комнатной температуре в течение по меньшей мере 45 минут перед введением.
Протокол иммунизации
Самок мышей Balb/C (Charles River Germany, 6-8 недель на дату доставки) иммунизировали подкожно с 3-недельными интервалами (n = 8 мышей на группу; в контрольных группах n = 4).
Группы мышей, от А до О, иммунизировали следующим образом.
Группа A (P467-CRM, 10 мкг на инъекцию): 250 мкл P467-CRM с 1250 мкл NaCl (1500 мкл). 150 мкл на мышь (10 мкг на мышь).
Группа В (P467-CRM, 25 мкг на инъекцию): 625 мкл P467-CRM с 875 мкл NaCl (1500 мкл). 150 мкл на мышь (25 мкг на мышь).
Группа С (P467-CRM, 50 мкг на инъекцию): 1250 мкл P467-CRM с 250 мкл NaCl (1500 мкл). 150 мкл на мышь (50 мкг на мышь).
Группа D (P467-CRM - Alum Brenntag, 10 мкг на инъекцию): 250 мкл P467-CRM со 175 мкл Alum Brenntag + 1325 мкл PBS (1750 мкл). 175 мкл на мышь (10 мкг на мышь).
Группа Е (P467-CRM - Alum Brenntag, 10 мкг на инъекцию): 625 мкл P467-CRM со 175 мкл Alum Brenntag + 950 мкл PBS (1750 мкл). 175 мкл на мышь (25 мкг на мышь).
Группа F (P467-CRM - Alum Brenntag, 50 мкг на инъекцию): 1250 мкл P467-CRM со 175 мкл Alum Brenntag + 325 мкл PBS (1750 мкл). 175 мкл на мышь (50 мкг на мышь).
Группа G (P467-CRM - Alum Serva, 10 мкг на инъекцию): 250 мкл P467-CRM с 1000 мкл Alum Serva + 250 мкл NaCl (1500 мкл). 150 мкл на мышь (10 мкг на мышь).
Группа Н (P467-CRM - Alum Serva, 25 мкг на инъекцию): 625 мкл P467-CRM с 1000 мкл Alum Serva (1625 мкл). 160 мкл на мышь (10 мкг на мышь).
Группа I (P467-CRM-Montanide, 10 мкг на инъекцию): 300 мкл P467-CRM с 300 мкл NaCl + 600 мкл Montanide. 100 мкл на мышь.
Группа J (P467-CRM-Montanide, 25 мкг на инъекцию): 700 мкл P467-CRM с 700 мкл Montanide. 126 мкл на мышь.
Группа K (P467-CRM-Montanide, 50 мкг на инъекцию): 1300 мкл P467-CRM с 1300 мкл Montanide. 252 мкл на мышь.
Г руппа L (Р467-мономер, 25 мкг на инъекцию): 250 мкл P467-CRM с 1250 мкл NaCl. 150 мкл на мышь (25 мкг на мышь).
Группа М (Montanide): 1000 мкл NaCl + 1000 мкл Montanide. 252 мкл на мышь.
Группа N (Alum Brenntag): 1575 мкл PBS + 175 мкл Alum Brenntag. 175 мкл на мышь.
Группа О (CRM, 50 мкг на инъекцию): Исходная концентрация: 2 мг/мл. Разведение 1:4 в NaCl (150+450 NaCl). 100 мкл на мышь.
Контрольные группы получали CRM, гидрат окиси алюминия (Brenntag) или Montanide сами по себе.
Забор крови у животных проводили перед первым введением (ВА0), через 20 дней после 2-й иммунизации (ВА1) и через 18 дней после 3-й иммунизации (ВА2). Для оценки динамики титров антител также проводили забор крови через 8 недель после 3-й иммунизации (ВА3).
У оставшихся 4 мышей из каждой группы (за исключением контрольных групп М, N и О, где умерщвляли всех мышей) титры антител дополнительно оценивали через 8 и 16 недель после 3-й иммунизации (ВА4 и ВА5, соответственно) для определения антител в динамике и необходимых интервалов повторной иммунизации.
Получение и культивирование клеток селезенки
Мышей (n = 4 на группу) умерщвляли, в асептических условиях извлекали селезенки, измельчали и фильтровали их через стерильные фильтры. Суспензии клеток получали, как описано ранее (Wiedermann et al, Int. Immunol. 1999; Oct; 11 (10): 1717-24). Спленоциты высевали в 96-луночные круглодонные планшеты в концентрации 0,5×106 на лунку и стимулировали с использованием CRM или Р467 в концентрации 20 мкг/мл в течение 72 часов. Супернатанты храним при -20°С до анализа. В качестве контроля проводили стимуляцию с использованием ConA.
Протоколы ELISA
(1) Измерение уровней пептид-специфичных антител
В качестве сорбируемого антигена использовали неконъюгированный слитый пептид Р467 (Bachem, Швейцария), разведенный в карбонатном буфере, в концентрации 0,5 мкг на микротитрационную лунку. Сыворотки (n = 8 на группу) разводили, добавляли в лунки и связанные антитела IgG выявляли меченным пероксидазой хрена (HRP) кроличьим противомышиным IgG POX (специфичным в отношении Fc-фрагмента, номер по каталогу 315-035-008; Jackson Immuno Research) с последующим добавлением 3,3',5,5'-тетраметилбензидина (ТМВ). Добавляли останавливающий раствор и планшеты считывали при 450-630 нм.
Для выявления IgG-подтипов IgG1 и IgG2a в лунки микротитрационного планшета с сорбированным антигеном вносили разведенные сыворотки с последующим добавлением крысиного антитела против мышиного IgG1 или крысиного антитела против мышиного IgG2a. Затем для выявления использовали вторичное HRP-меченное мышиное антитело против крысиного IgG с последующим добавлением ТМВ. Добавляли останавливающий раствор и планшеты считывали при 450-630 нм.
(2) Измерение уровней Her2/neu-специфичных антител
На лунки микротитрационного планшета сорбировали слитый белок, содержащий рекомбинантный внеклеточный домен человеческого Her2/neu (аминокислотные остатки 23-652), конъюгированный с Fc-областью человеческого IgG1. На каждую лунку сорбировали 0,1 мкг слитого белка. Затем анализировали разведенные мышиные сыворотки, как описано выше.
Для выявления IgG-подтипов IgG1 и IgG2a в лунки с сорбированным антигеном вносили разведенные сыворотки с последующим добавлением крысиного антитела против мышиного IgG1 или крысиного антитела против мышиного IgG2a. Для выявления использовали вторичное HRP-меченное мышиное антитело против крысиного IgG (H+L).
(3) Измерение образования цитокинов IL-2, IL-5 и IFNγ in vitro
Получали супернатанты спленоцитов, стимулированных CRM или Р467, и концентрацию IL-2, IL-5 и IFNγ в них анализировали посредством ELISA, следуя инструкциям изготовителя (Affymetrix eBioscience, США). Супернатанты использовали без разведения или с разведением 1:10-1:20 перед анализом. Анализы с флуоресцентно-активированной сортировкой клеток (fluorescence-activated cell sorting, FACS) FACS проводили на свежевыделенных клетках селезенки (2,5×106 клеток/мл в 24-луночных плоскодонных планшетах). Клетки инкубировали в течение 2 часов в присутствии форболмиристатацетата (РМА; 10 нг/мл) и иономицина (1,25 мкМ) и в течение еще 4 часов при 37°С в присутствии брефелдина А (10 мкг/мл). Затем клетки выдерживали при 4°С в течение ночи и окрашивали на следующее утро противомышиными антителами к следующим мишеням.
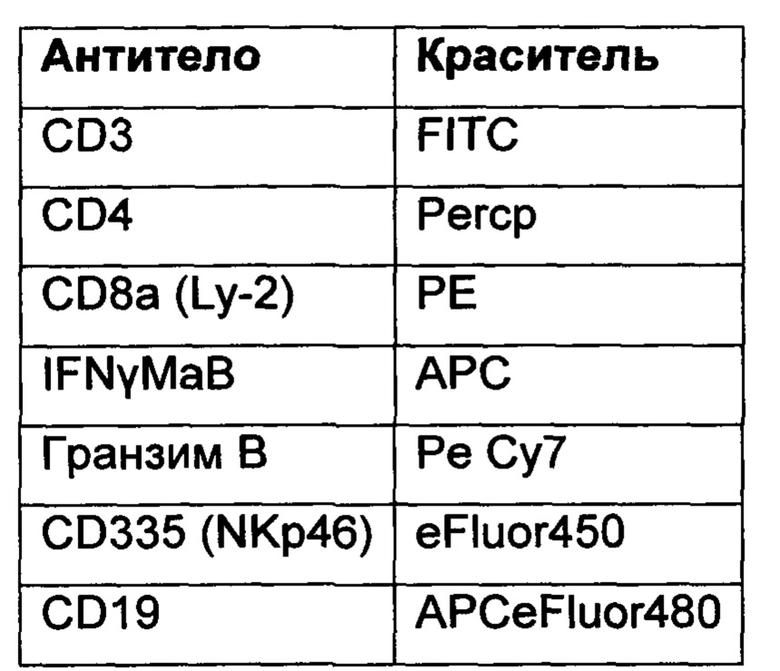
(1) Протокол окрашивания
Клетки собирали и разделяли по микропробиркам (1× клеток на пробирку). Для каждого мышиного образца проводили по одному окрашиванию с использованием указанной выше смеси антител.
Затем клетки отмывали, блокировали Fc-блокирующим реагентом (антителом против мышиного CD16/32) и окрашивали антителами к CD3, CD4, CD8, CD19 и CD335, с последующей фиксацией/пермеабилизацией и внутриклеточным окрашиванием на IFNγ и гранзим В.
Проводили анализы для описания популяций спленоцитов - Т-клеток (CD3+CD4+ и CD3+CD8+), В-клеток (CD3-CD19+) и NK-клеток (CD3-CD335+). Анализы также включали выявление внутриклеточного белка IFNγ.
Статистический анализ
Для анализа результатов ELISA применяли Prism с непарным критерием Стьюдента (двусторонний, доверительные интервалы 95%). Значимые различия отмечены одной или несколькими звездочками (* Р-значение <0,05, ** 0,01 < Р-значение <0,001, *** Р-значение <0,001). Отсутствие значимости обозначено «ns». При сравнении более чем двух групп применяли Prism с односторонним дисперсионным анализом (analysis of variance, ANOVA) Для анализа результатов FACS применяли Prism с критерием Манна-Уитни (двусторонний, доверительные интервалы 95%).
Результаты
Гуморальные ответы (1)
Уровни пептид-специфичных антител
Сыворотки, полученные при последнем заборе крови после трех иммунизаций, а также сыворотки, полученные до иммунизации, разводили 1:100000 (для IgG и IgG1) или 1:100 (для IgG2a). Титры антител для общих IgG, IgG1 и IgG2a сыворотки показаны на Фиг. 15А-С, соответственно.
На этих графических материалах показаны сравнение и статистическая оценка следующих групп (в рамке):
- D, Е, F: Alum: 10 мкг, 25 мкг, 50 мкг.
- I, J, K: Montanide: 10 мкг, 25 мкг, 50 мкг.
Полученные данные демонстрируют, что антитела против Р467 были получены у всех мышей, иммунизированных P467-CRM. У контрольных животных и у мышей, иммунизированных Р467 самим по себе, гуморального иммунного ответа получено не было. По сравнению с Alum (Brenntag), уровни антител IgG, IgG1 и IgG2a были значительно выше при введении P467-CRM с Montanide. Титры IgG2a были обычно ниже, чем у IgG1, несмотря на то, что у животных, иммунизированных P467-CRM + Montanide, были отмечены существенно более высокие титры IgG2a.
(2) Уровни Her2/neu-специФичных антител
Сыворотки, полученные при последнем заборе крови после трех иммунизаций, а также сыворотки, полученные до иммунизации, разводили 1:2500 (до 10000) (IgG), 1:10000 (IgG1) и 1:100 (IgG2a). Титры антител для IgG, IgG1 и IgG2a сыворотки показаны на Фиг. 16А-С, соответственно.
Высокие титры антител IgG и IgG1 против внеклеточного Her2/neu были очевидны у всех животных, иммунизированных P467-CRM. У контрольных животных и у мышей, иммунизированных Р467 самим по себе, очевидного гуморального иммунного ответа против Her2/neu.не было. По сравнению с Alum (Brenntag), титры антител IgG и IgG1 были значительно выше при введении пептидной конструкции P467-CRM с Montanide.
Образование цитокинов in vitro (IL-2, IFNγ, IL-5)
(1) Образование IL-2 после стимуляции CRM или P467
Уровни IL-2 в CRM-стимулированных спленоцитах, имеющих происхождение от мышей, иммунизированных P467-CRM в присутствии Montanide, были выше, чем в CRM-стимулированных спленоцитах, имеющих происхождение от мышей, иммунизированных P467-CRM в присутствии Alum (см. Фиг. 17А и 17В).
(2) Образование IFNγ после стимуляции CRM или Р467
На Фиг. 18А и 18В показана концентрация IFNγ в супернатанте спленоцитов, культивированных in vitro в присутствии CRM (Фиг. 18А) или Р467 (Фиг. 18В). У мышей, иммунизированных с использованием 25 мкг P467-CRM с Montanide, уровень IFNγ был вне определяемого диапазона. Данные этой мыши были исключены, и был проведен дополнительный анализ, как показано на Фиг. 18С (стимуляция CRM) и Фиг. 18D (стимуляция Р467).
Стимуляция CRM приводила к образованию IFNγ в спленоцитах, имеющих происхождение от всех мышей, иммунизированных P467-CRM в присутствии Alum или Montanide. При использовании Montanide (в группе 10 мкг) уровни IFNγ были выше, чем в соответствующей группе, иммунизированной P467-CRM в присутствии Alum (Brenntag). При стимуляции P467-CRM наиболее высокими уровни IFNγ были в группе Montanide, иммунизированной с использованием 50 мкг P467-CRM.
(3) Образование IL-5 после стимуляции CRM или Р467
На Фиг. 19А и 19B показана концентрация IL-5 в супернатанте спленоцитов, культивированных in vitro в присутствии CRM (Фиг. 19А) или Р467 (Фиг. 19В).
Уровни IL-5 были существенно выше в CRM-стимулированных спленоцитах, имеющих происхождение от мышей, иммунизированных P467-CRM в присутствии Montanide. Стимуляция пептидной конструкцией P467-CRM приводила к образованию некоторого количества IL-5 у мышей, иммунизированных с Montanide, однако уровень IL-5 был приблизительно в 10 раз ниже, чем после стимуляции CRM.
Описание спленоцитов посредством FACS-анализа; внутриклеточное окрашивание образованного IFNγ
Для оценки возможного влияния применения различных адъювантов (то есть Alum против Montanide) на распределение лимфоцитов, в частности CD8+ и CD4+ лимфоцитов, проводили FACS-анализ спленоцитов после иммунизации различными концентрациями Р467 в присутствии Montanide или Alum. У мышей выделяли спленоциты и анализировали их на предмет процента Т-клеток (CD3+CD4+, Фиг. 20А, и CD3+CD8+, Фиг. 20В), В-клеток (CD3-CD19+, Фиг. 20С) и NK-клеток (CD3-CD335+, Фиг. 20D).
Полученные данные демонстрируют значительное снижение NK-клеток в спленоцитах, имеющих происхождение от мышей, иммунизированных более высокими дозами P467-CRM (то есть 25 мг и 50 мг) с Montanide по сравнению с Alum (Brenntag).
Выявление спленоцитов, образующих IFNγ, посредством FACS-анализа
Для определения того, будет ли иммунизация с использованием различных адъювантов приводть к изменению распределения CD4+ или CD8+ клеток, образующих IFNγ, у мышей из всех групп выделяли спленоциты и измеряли внутриклеточный IFNγ в Т-клетках (CD4+, Фиг. 21 А, и CD8+, Фиг. 21В) и NK-клетках (CD3-CD335+, Фиг. 21С).
Полученные данные демонстрируют, что CD4+ Т-клетки образовывали IFNγ, что было выражено у мышей, иммунизированных возрастающими концентрациями P467-CRM. Все контрольные группы (и неиммунизированные мыши) продемонстрировали сходные уровни IFNγ. В отличие от этого, уровни IFNγ были повышены в CD8+ Т-клетках, имеющих происхождение от мышей, иммунизированных с использованием 25 мкг P467-CRM с Montanide. Полученные данные также демонстрируют образование IFNγ NK-клетками, имеющими происхождение от мышей, иммунизированных P467-CRM + Montanide.
Динамика гуморальных иммунных ответов против Her2 и пептида Р467
Для оценки динамики гуморальных иммунных ответов получали образцы крови через 8 недель, 16 недель и 6 месяцев после последней иммунизации.
Затем в образцах крови анализировали титры антител, специфичных в отношении Her2/neu и пептида Р467, в соответствии со способами, раскрытыми здесь.
Как показано на Фиг. 22, титры антител IgG, специфичных в отношении пептида Р467, оставались повышенными через 8 недель (ВА3), 16 недель (ВА4) и 6 месяцев (ВА5) после последнего введения пептидной конструкции P467-CRM с Alum или Montanide. Сходным образом, как показано на Фиг. 23, титры антител IgG, специфичных в отношении Her2/neu, оставались повышенными через 8 недель (ВА3), 16 недель (ВА4) и 6 месяцев (ВА5) после последнего введения пептидной конструкции P467-CRM с Alum или Montanide.
Заключение
Как конъюгаты слитых пептидов с CRM 197, так и виросомные композиции, содержащие слитые пептиды, приводили к получению антител, специфичных в отношении отдельных В-клеточных эпитопов (Р4, Р6 и Р7), а также слитых пептидов (Р467 и Р647). Кроме того, было отмечено, что эти антитела также связывались с рекомбинантным внеклеточным доменом Her2/neu и с нативным белком Her2/neu, экспрессированным на клетках рака молочной железы SKBR-3.
Конъюгаты слитых пептидов с CRM197 стимулировали выработку антител, специфичных в отношении В-клеточных эпитопов и Her2/neu, эффективнее; то есть, по сравнению с соответствующими виросомными композициями, были получены значительно (приблизительно в 10-20 раз) более высокие титры антител.
По титрам антител против рекомбинантного Her2/neu было явное различие в динамике гуморальных иммунных ответов в процессе иммунизации, и полученные результаты демонстрируют, что CRM-конъюгаты приводят к более раннему повышению титров антител (уже после 2-й иммунизации с пиком после 3-й иммунизации), в то время как при использовании виросомных конъюгатов повышение титров антител было более медленным, при этом для достижения уровней, отмеченных у мышей, иммунизированных соответствующими CRM-конъюгатами, было необходимо четыре иммунизации.
Обычно виросомы являются подходящей системой доставки антигенов. Тем не менее, результаты раскрытые здесь, ясно демонстрируют задержку роста титров антител после иммунизации виросомными конъюгатами, а также необходимость четырех иммунизаций для достижения титров, сходных с наблюдаемыми у животных, иммунизированных соответствующими CRM-конъюгатами. Как время, так и дозы для получения оптимальных гуморальных иммунных ответов являются важными факторами, которые необходимо учитывать для клинического применения вакцинных композиций Her2/neu. Таким образом, эти результаты показывают, что CRM197 является более подходящим белком-носителем для слитых пептидов Her2/neu в контексте доставки вакцины.
Было показано, что как Alum (Brenntag), так и Montanide дополнительно усиливают указанные выше гуморальные иммунные ответы на пептидную конструкцию P467-CRM. У всех мышей, иммунизированных с использованием Montanide (во всех дозах), титры специфичных антител IgG, а также IgG1 против Р467, были значительно выше, чем при иммунизации с использованием Alum. Титры антител IgG2a против Р467 у мышей, иммунизированных с использованием Montanide (во всех дозах), также были выше, чем при использовании Alum. Титры антител IgG и IgG1 против Her2/neu были значительно выше у мышей, иммунизированных с использованием Montanide, вводимого вместе с пептидными конструкциями, во всех исследованных дозах. В отличие от антител, специфичных в отношении пептида Р467, антитела IgG2a против Her2/neu были выявлены в небольшом количестве или не были обнаружены.
Образование IFNγ спленоцитами было значительно интенсивнее при культивировании клеток in vitro в присутствии CRM, по сравнению с клетками, культивированными в присутствии Р467. После стимуляции CRM уровни IFNγ в группах, которым вводили низкие дозы пептидных конструкций вместе с Montanide, были значительно выше, чем в группах, получавших те же дозы конструкций вместе с Alum (Brenntag). Полученные данные демонстрируют, что Montanide позволяет получить более высокие титры антител, а также более высокие уровни цитокинов, чем при использовании Alum.
| название | год | авторы | номер документа |
|---|---|---|---|
| САХАРИД-КОНЪЮГАТНЫЕ ВАКЦИНЫ | 2005 |
|
RU2454244C2 |
| СИНТЕТИЧЕСКАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ (ВАРИАНТЫ), ЭКСПРЕССИОННЫЙ ВЕКТОР ДЛЯ КЛЕТОК МЛЕКОПИТАЮЩЕГО, КЛЕТКА МЛЕКОПИТАЮЩЕГО ХОЗЯИНА И СПОСОБ ЭКСПРЕССИИ ЧЕЛОВЕЧЕСКОГО БЕЛКА АНТИГЕНА ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА-2 (HER2/neu) ИЛИ ЕГО УКОРОЧЕННОЙ ФОРМЫ | 2004 |
|
RU2397249C2 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ | 2011 |
|
RU2608905C2 |
| ИММУНОГЕННЫЙ ПЕПТИД ПРОТИВ СТРЕПТОКОККОВ ГРУППЫ А | 2018 |
|
RU2775621C2 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ АНТИГЕНОВ STAPHYLOCOCCUS AUREUS | 2010 |
|
RU2536981C9 |
| КОНЪЮГАТ ПОЛИСАХАРИД-БЕЛОК ДЛЯ ИНДУЦИРОВАНИЯ ИММУННОГО ОТВЕТА И ЗАЩИТЫ ПРОТИВ ИНФЕКЦИИ Staphylococcus aureus, СПОСОБЫ ПОЛУЧЕНИЯ КОНЪЮГАТА (ВАРИАНТЫ), КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ КОНЪЮГАТ И СПОСОБЫ ИНДУЦИРОВАНИЯ ИММУННОГО ОТВЕТА И ПРЕДОТВРАЩЕНИЯ ИНФЕКЦИИ Staphylococcus aureus | 2010 |
|
RU2531234C2 |
| Плазмидная ДНК-матрица pVAX-RBD, молекул мРНК-RBD, полученная с использованием ДНК-матрицы pVAX-RBD, обеспечивающая синтез и секрецию белка RBD SARS-CoV 2 в клетках организма млекопитающих и комплекс в виде наночастиц, содержащих молекулы мРНК-RBD, индуцирующих SARS-CoV-специфические антитела, обладающие вируснейтрализующей активностью | 2021 |
|
RU2754230C1 |
| ПРИМЕНЕНИЕ ПЕПТИДА, УСИЛИВАЮЩЕГО СЕКРЕЦИЮ СОМАТОТРОПНОГО ГОРМОНА, В КАЧЕСТВЕ АДЪЮВАНТА ВАКЦИНЫ | 2017 |
|
RU2768134C2 |
| СПОСОБ ВАКЦИНАЦИИ ПРОТИВ АУТОАНТИГЕНА У ПАЦИЕНТА-ЧЕЛОВЕКА | 2016 |
|
RU2734924C2 |
| СЛИТЫЕ БЕЛКИ КАРЦИНОЭМБРИОНАЛЬНОГО АНТИГЕНА | 2005 |
|
RU2380375C2 |

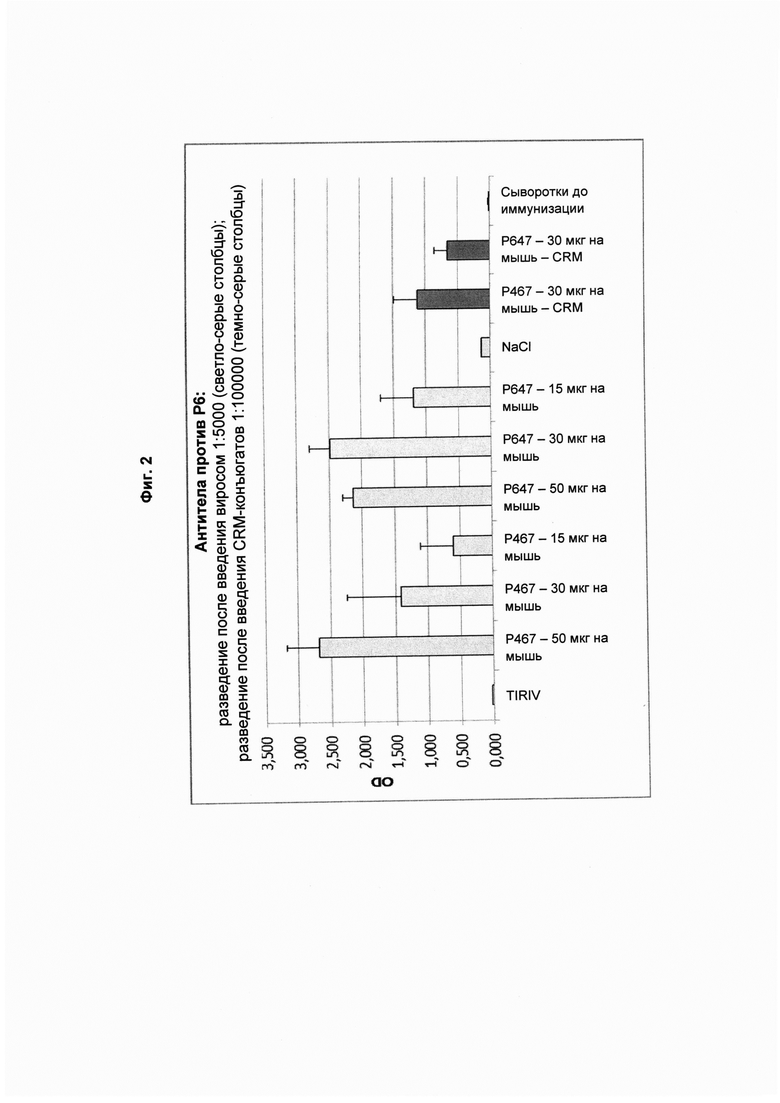
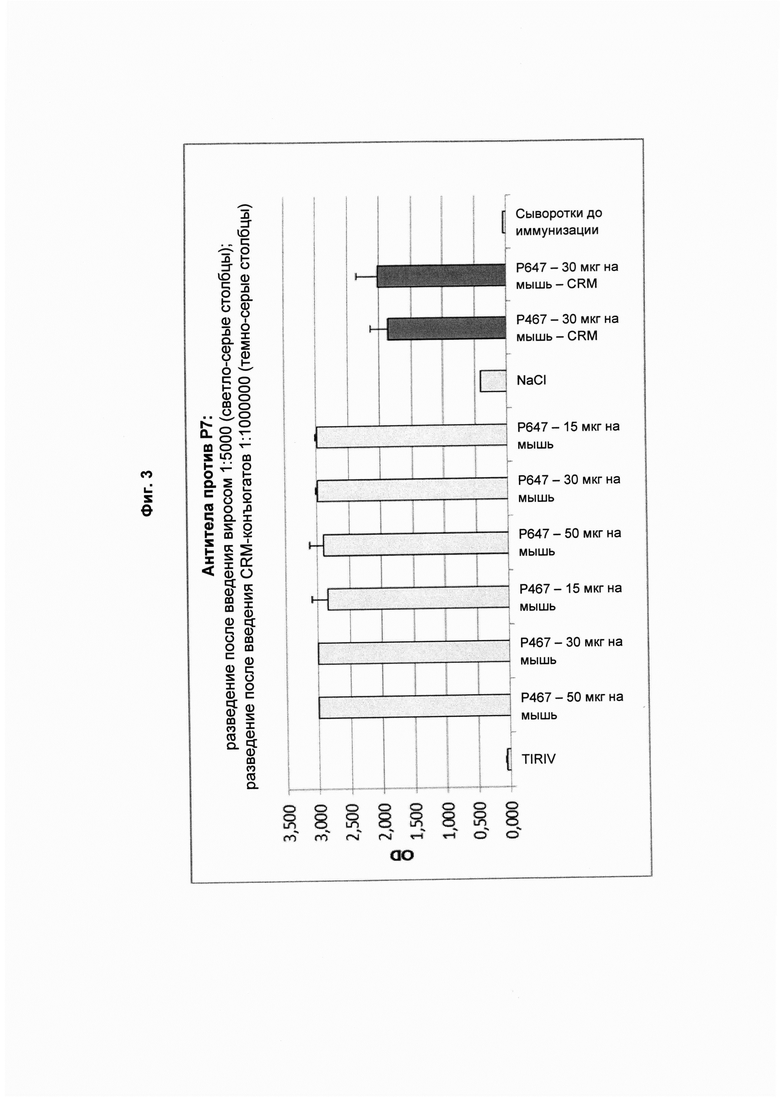
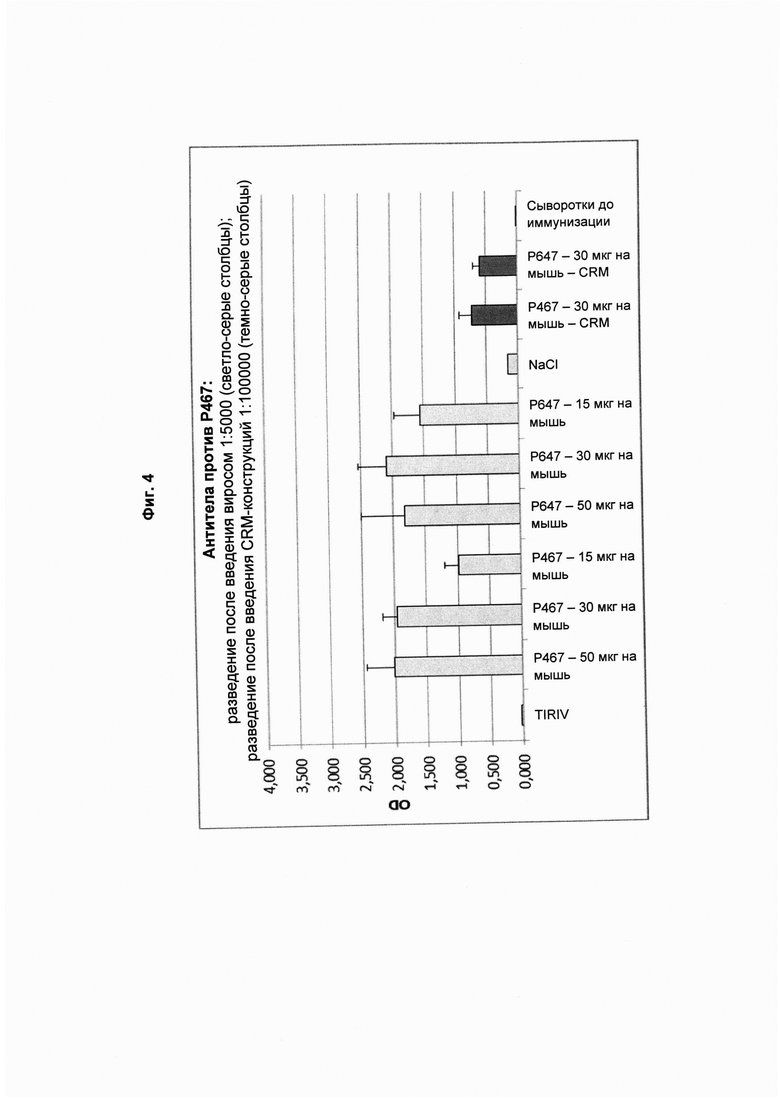
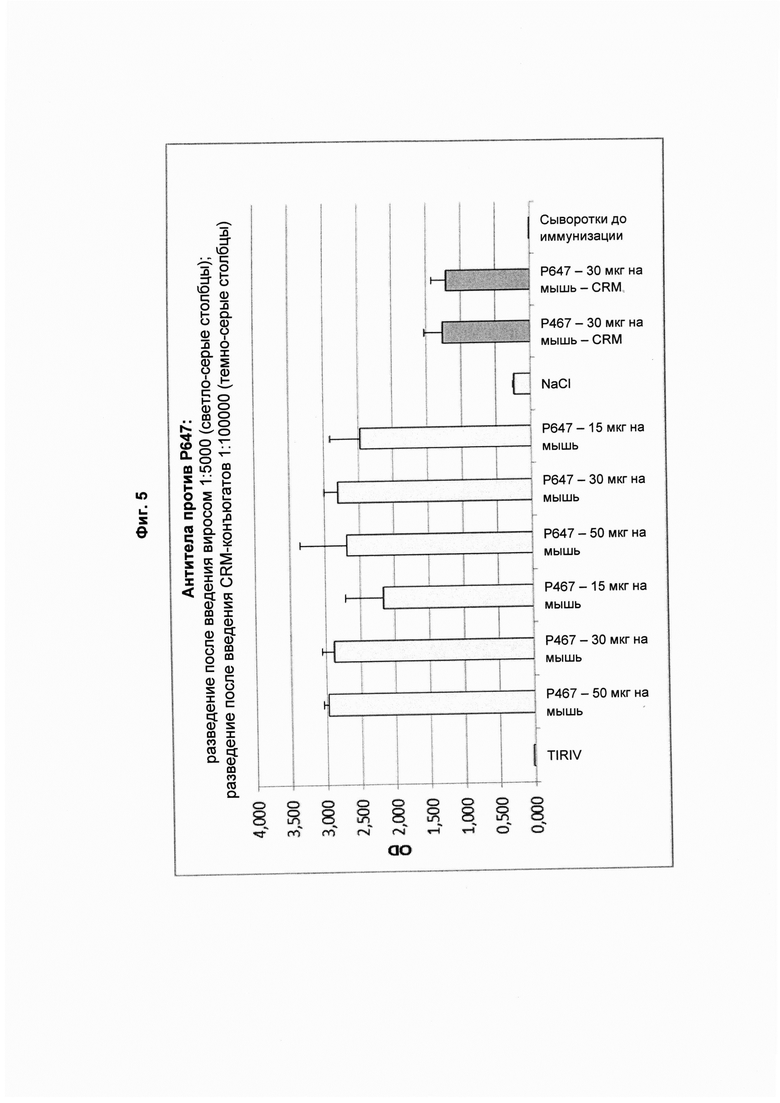
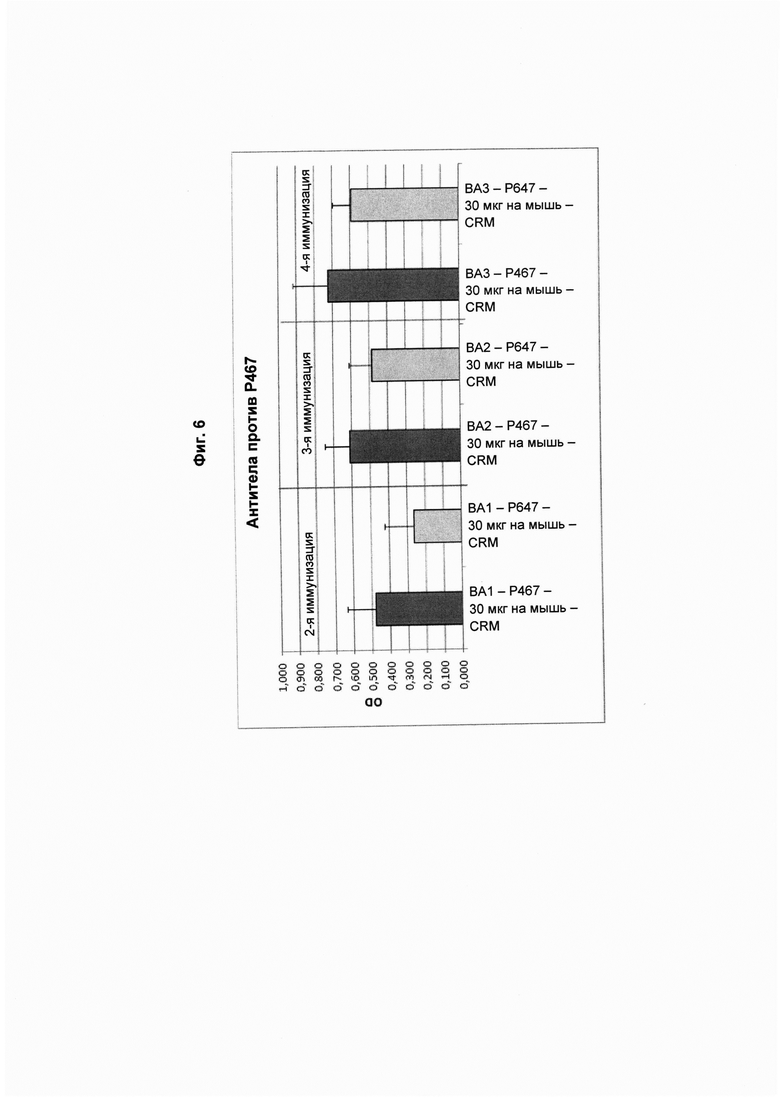
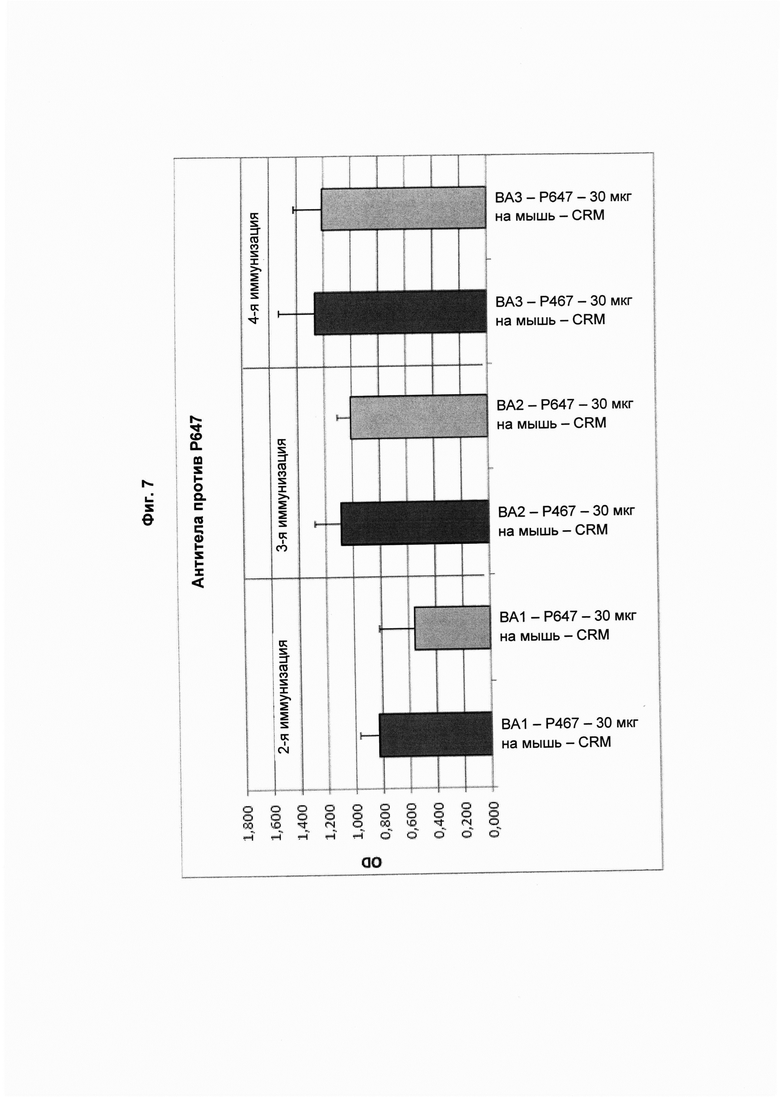
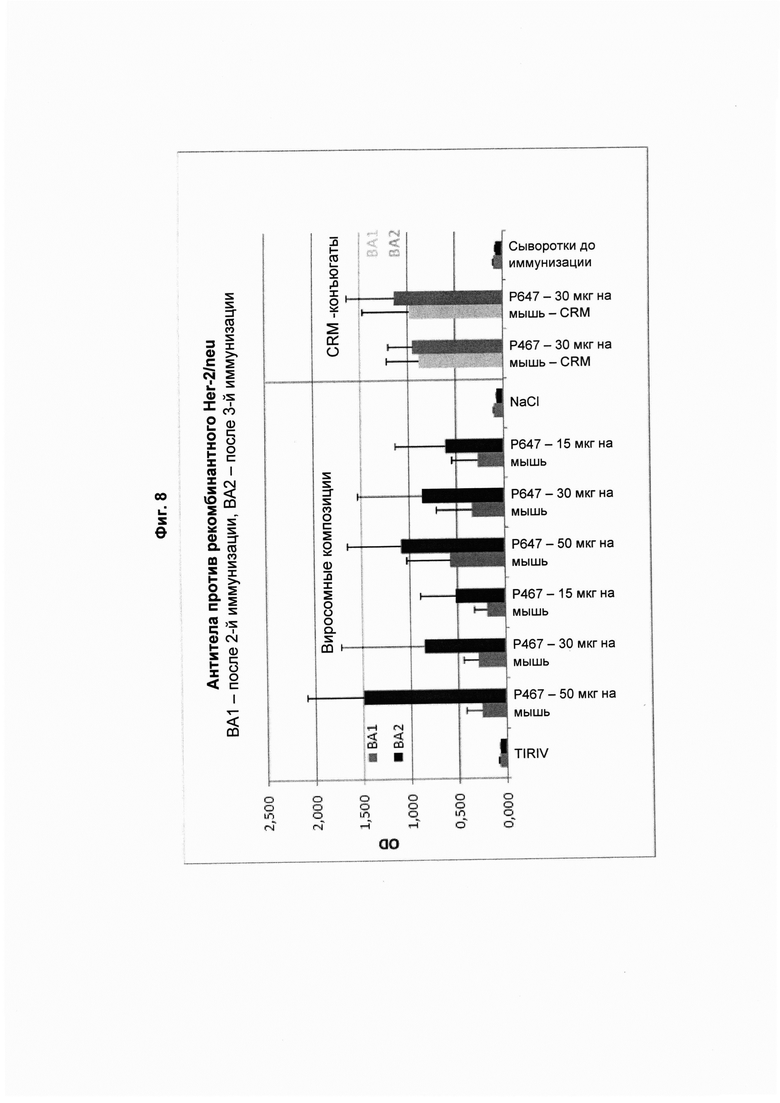

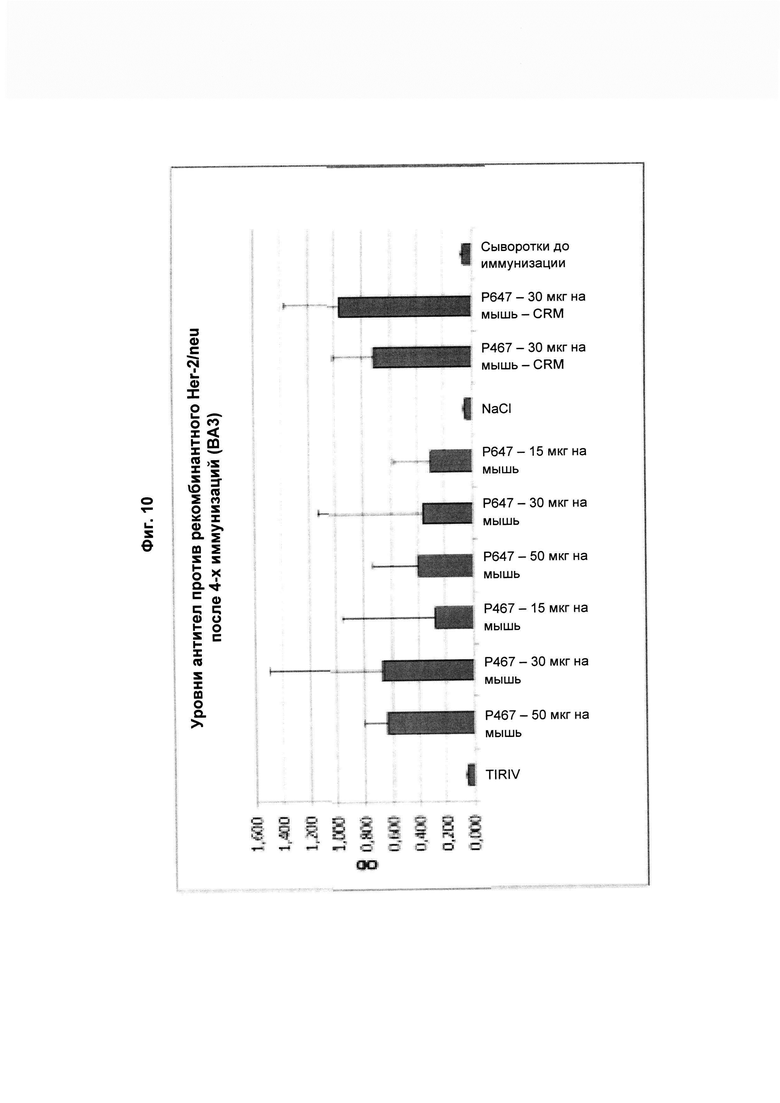
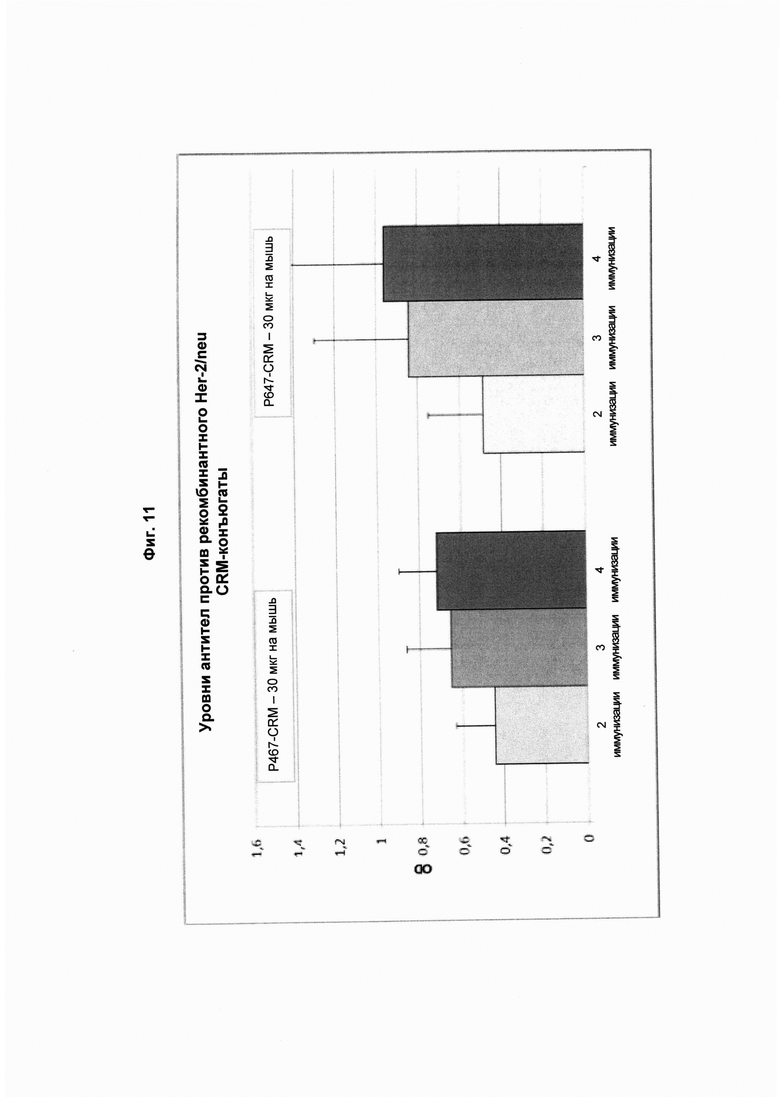
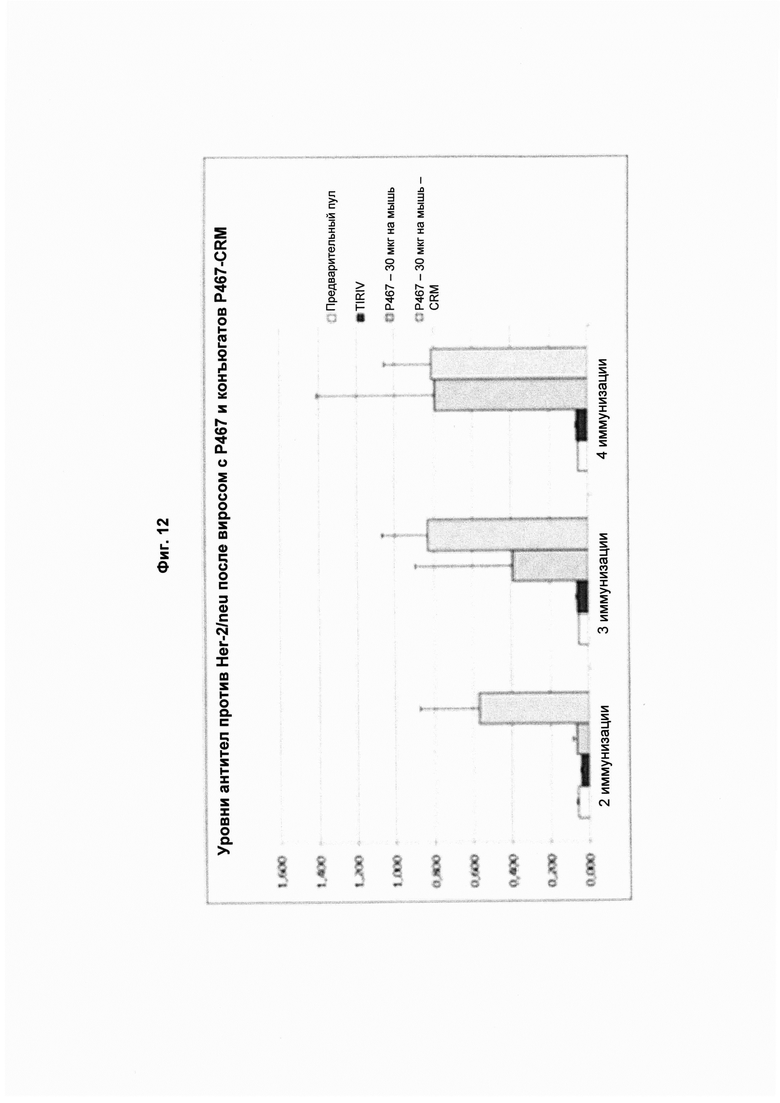
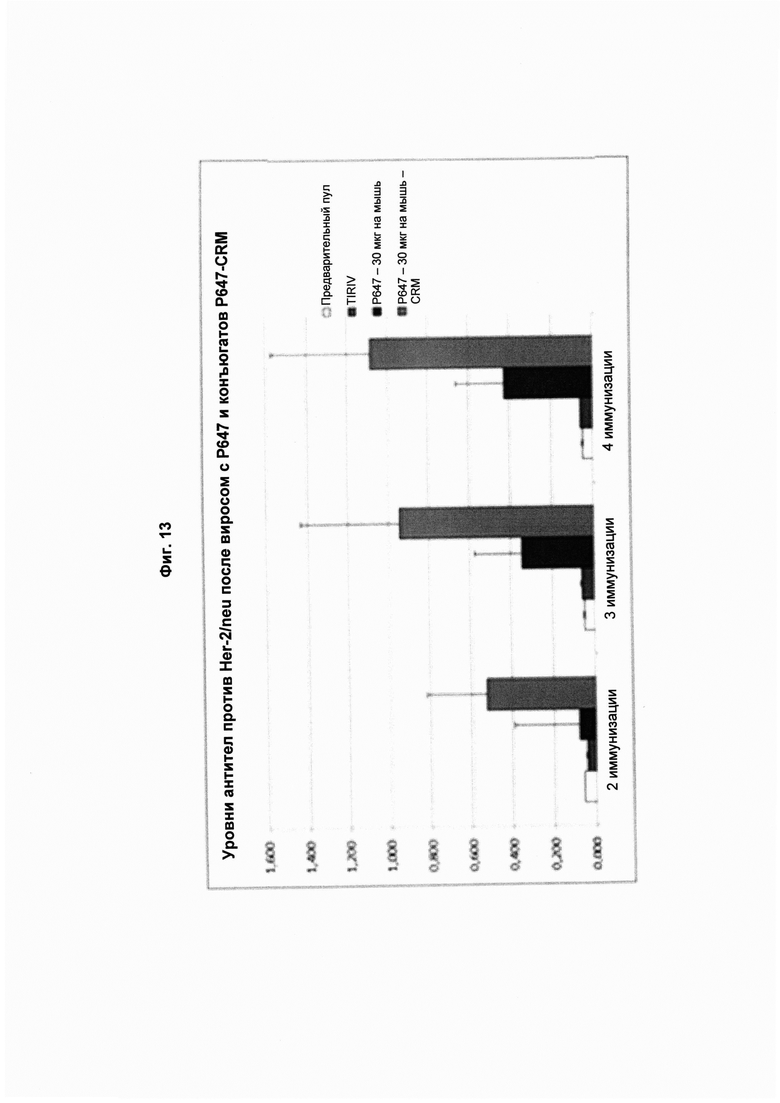
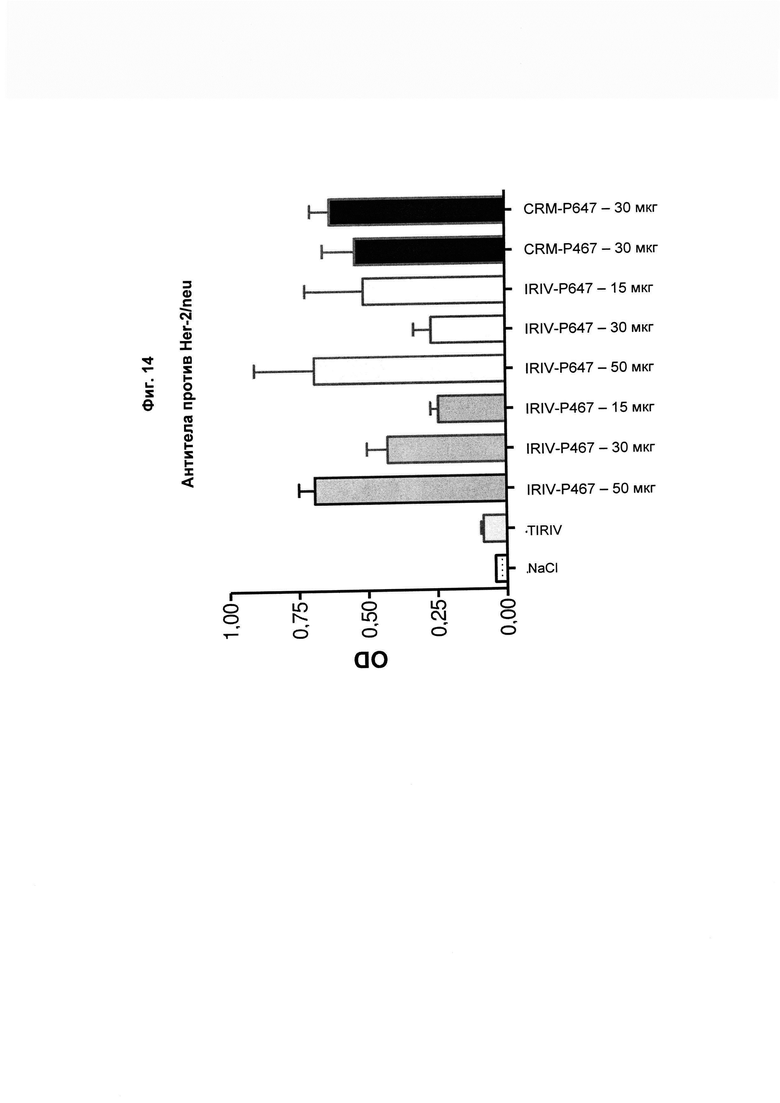
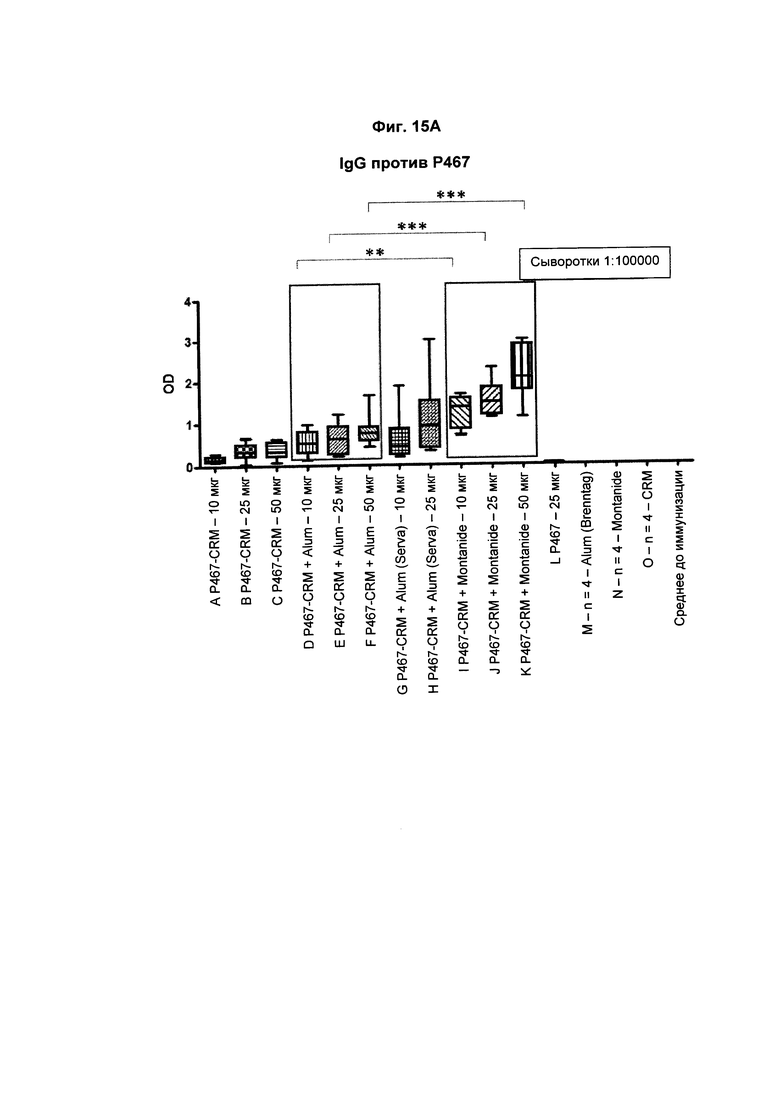
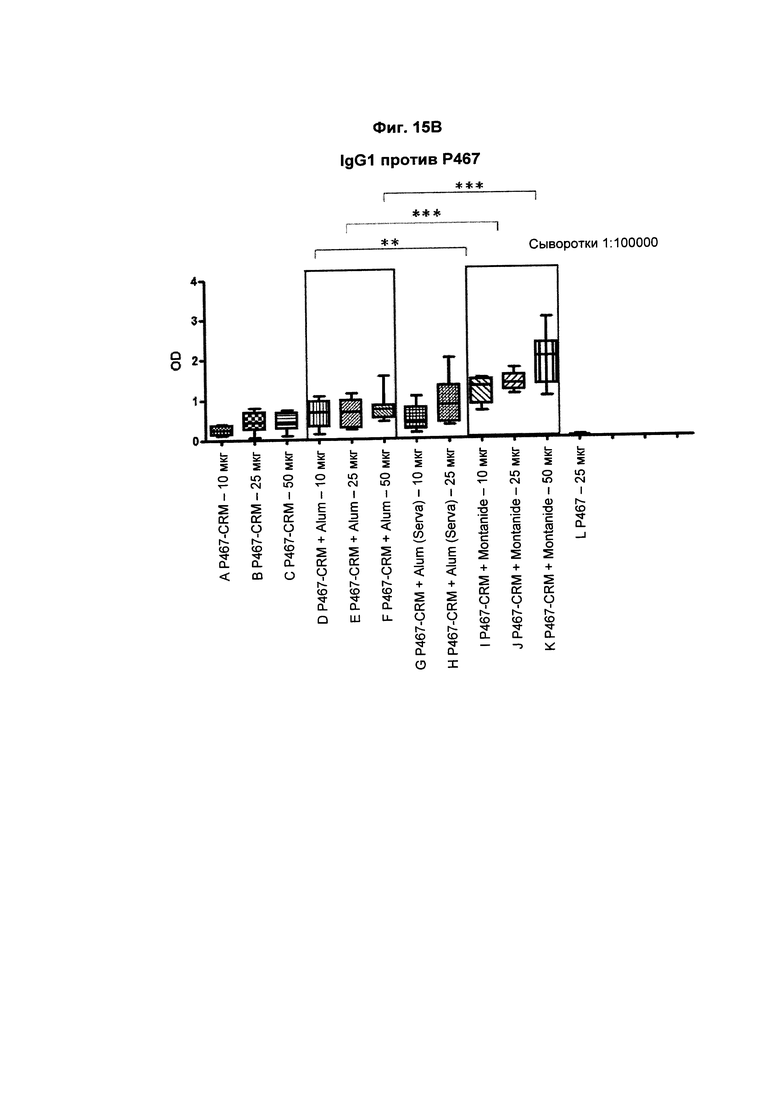
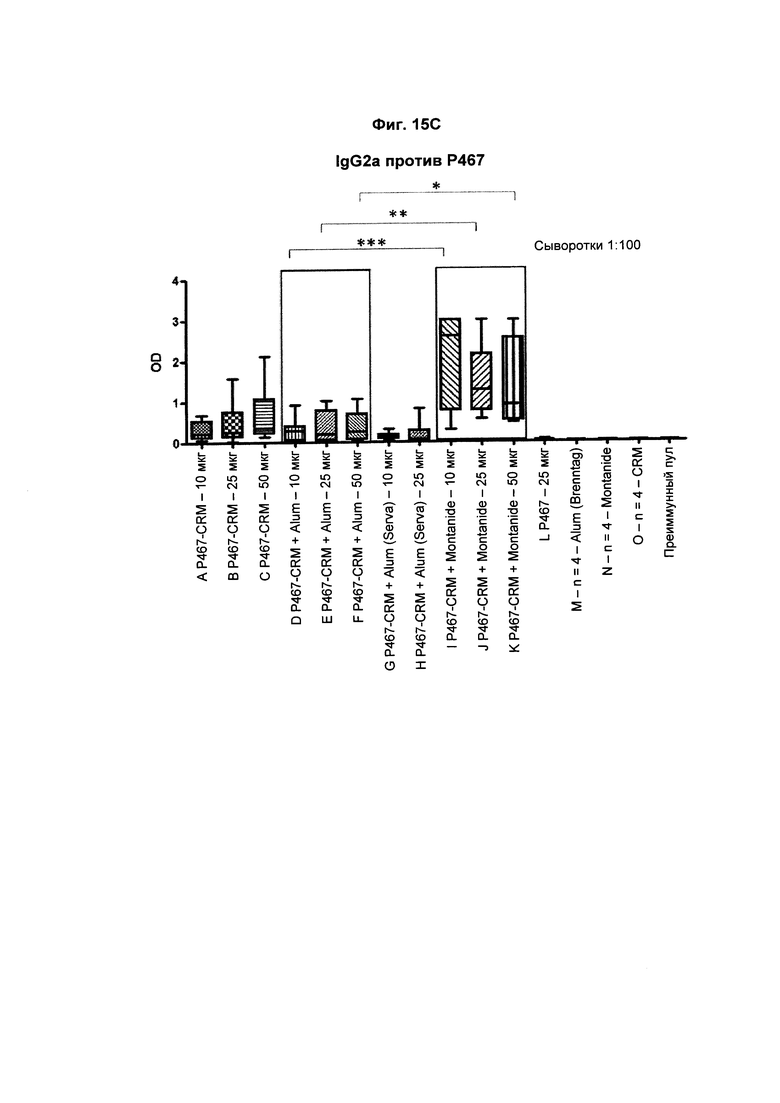
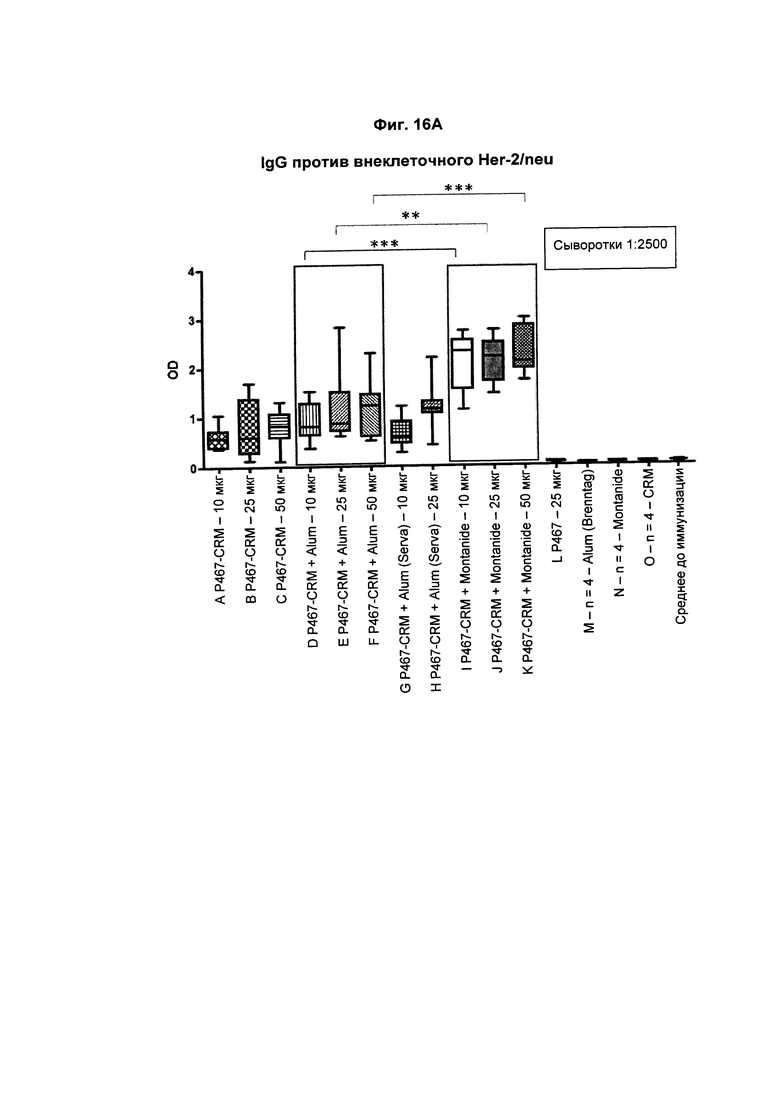
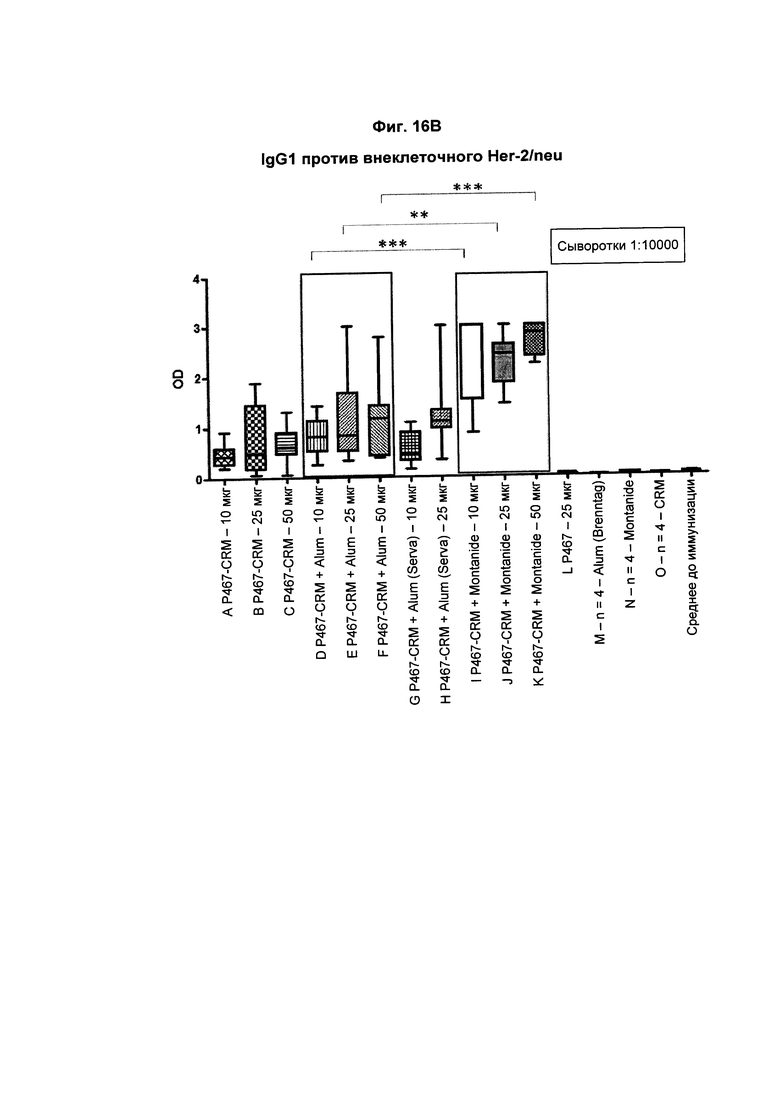
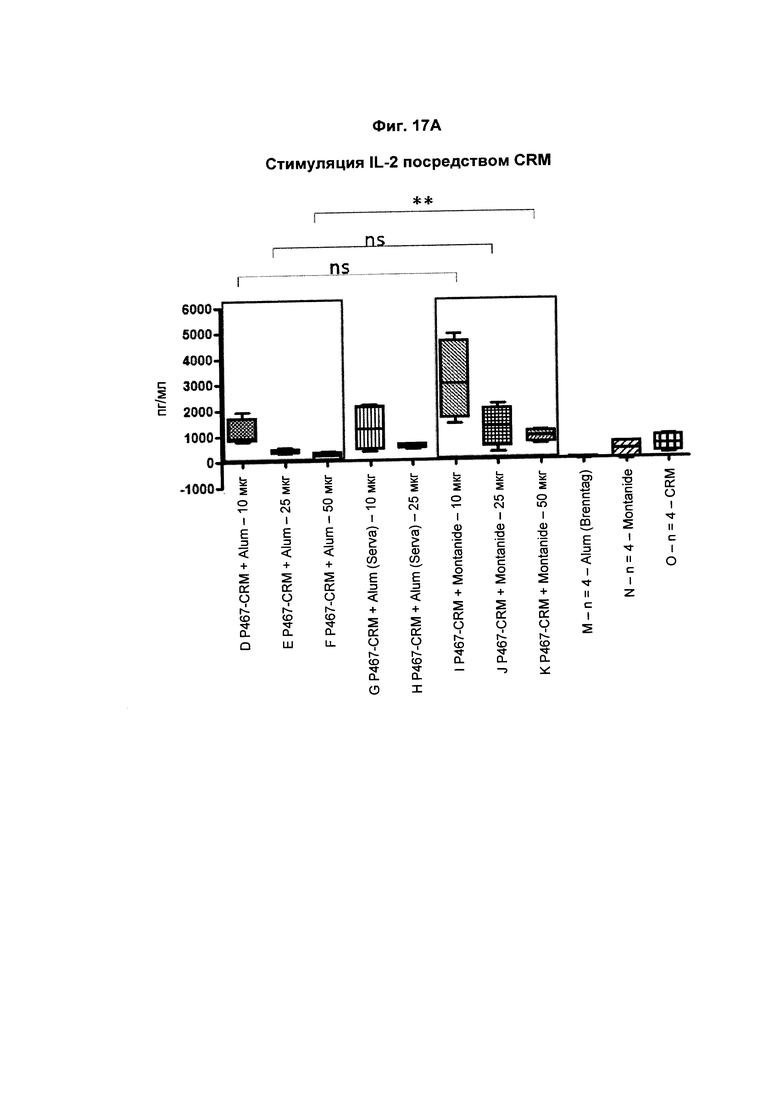
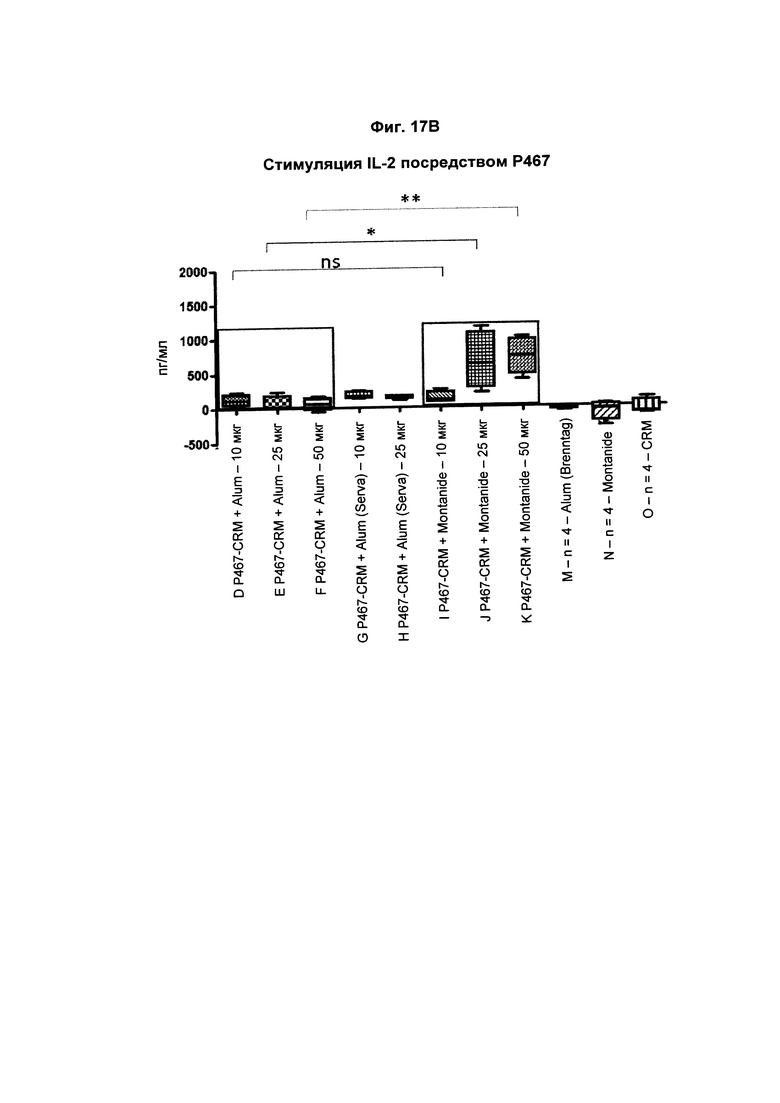
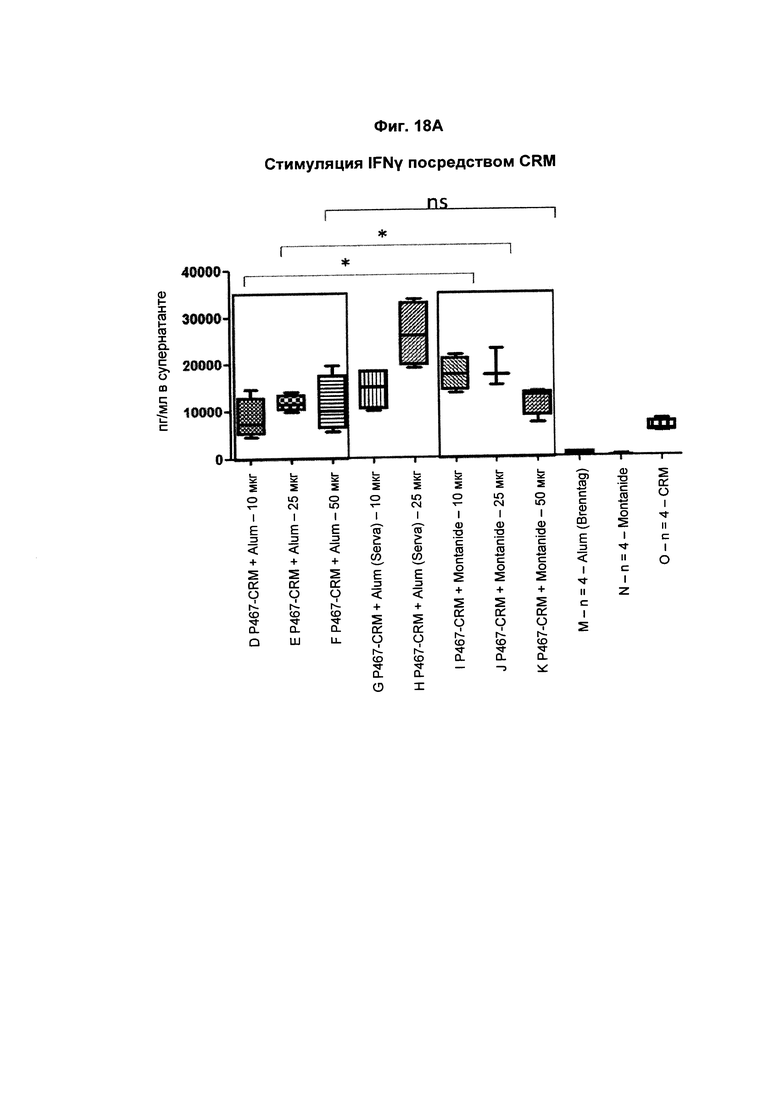
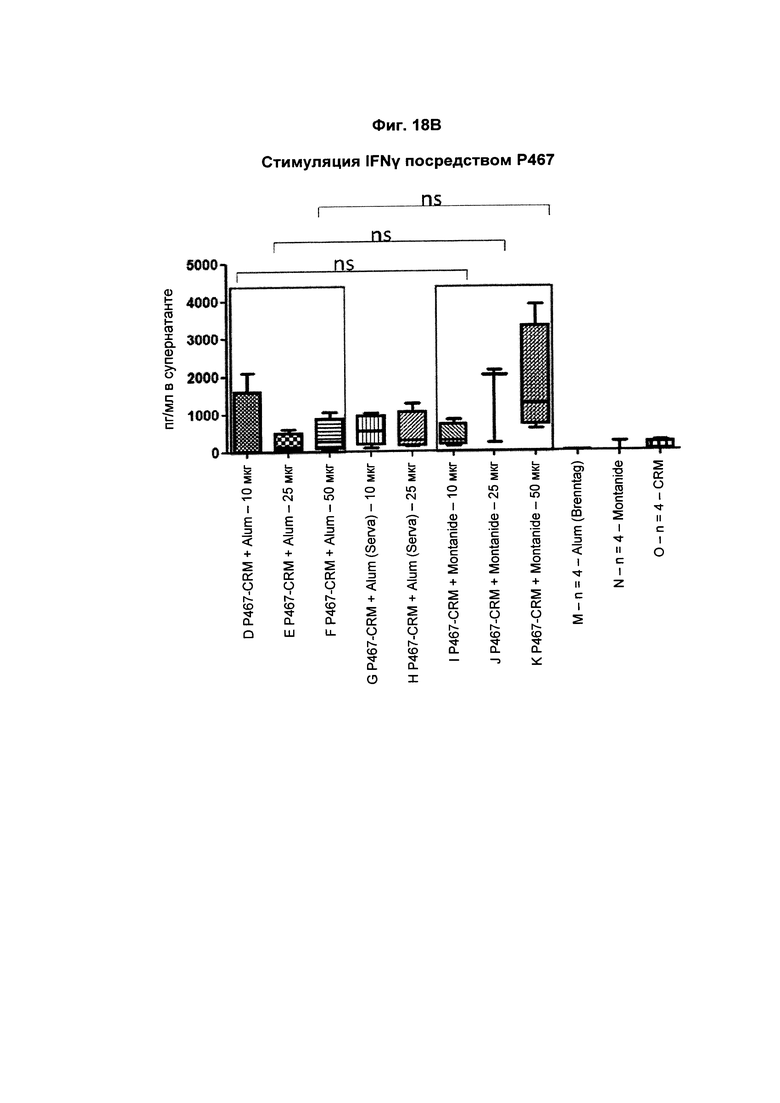
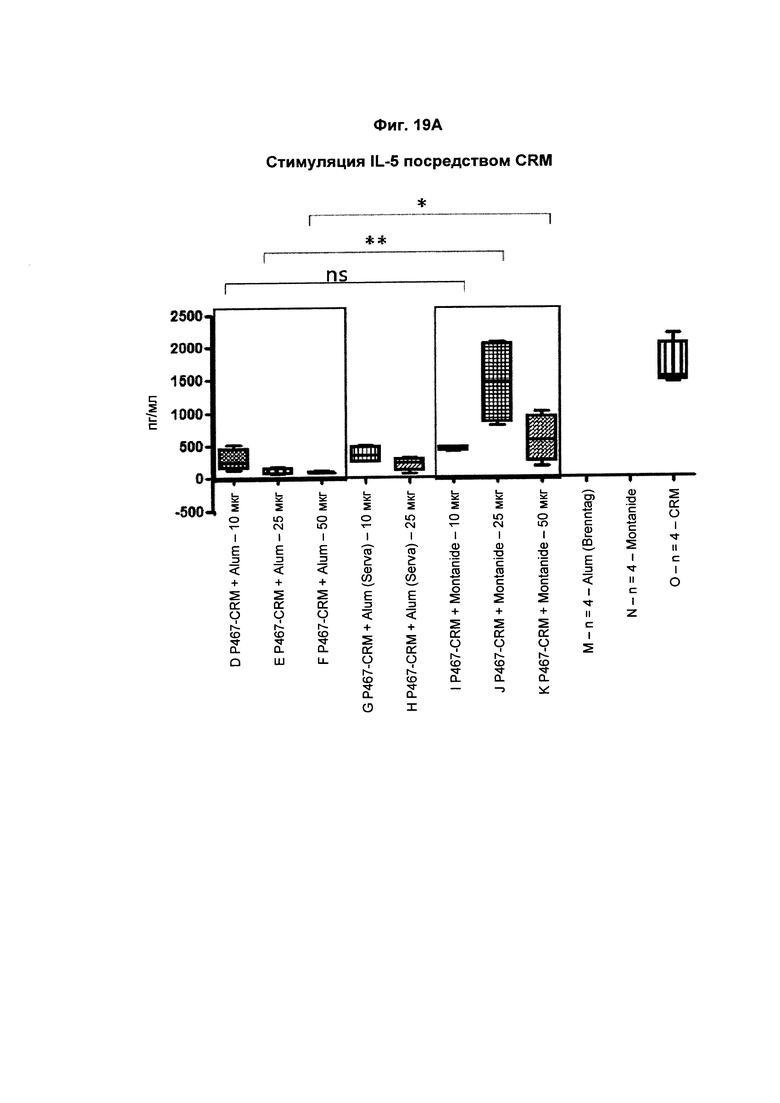
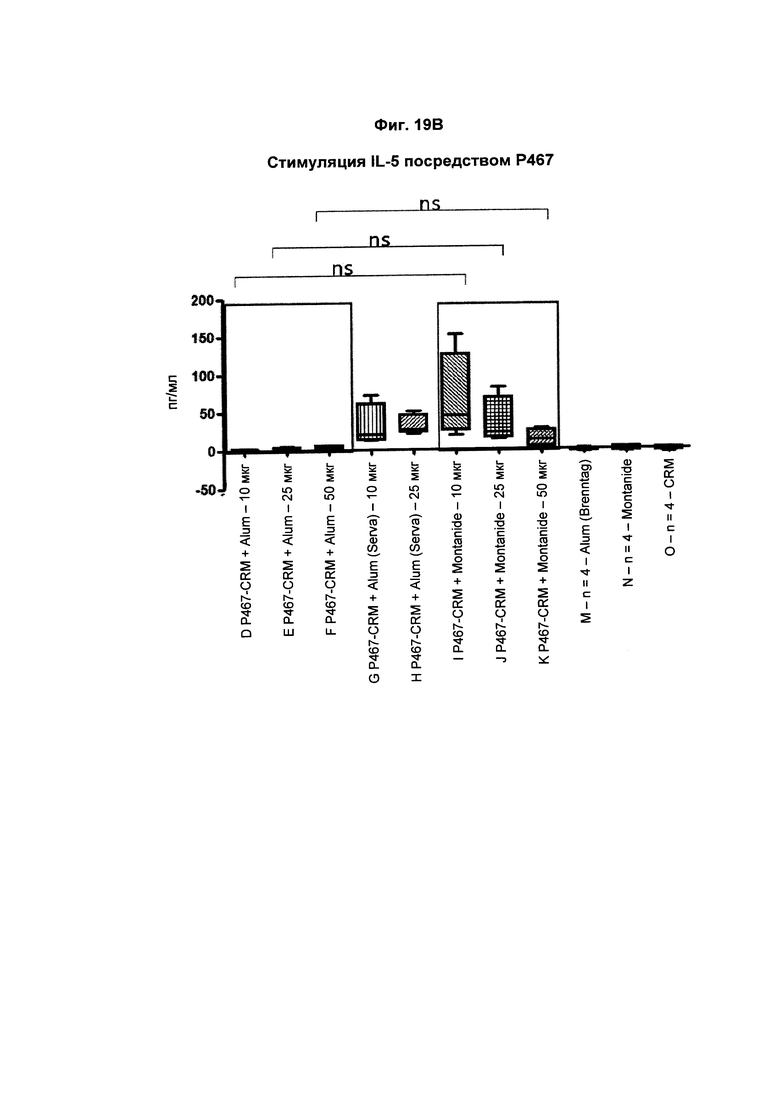
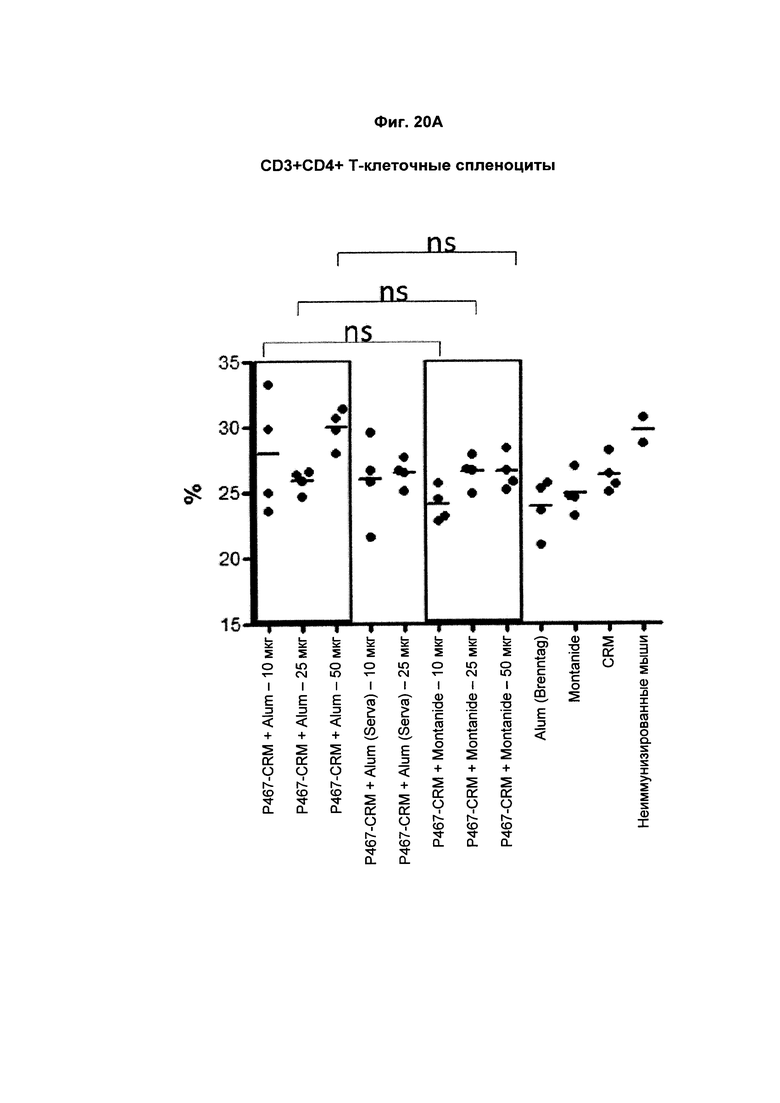
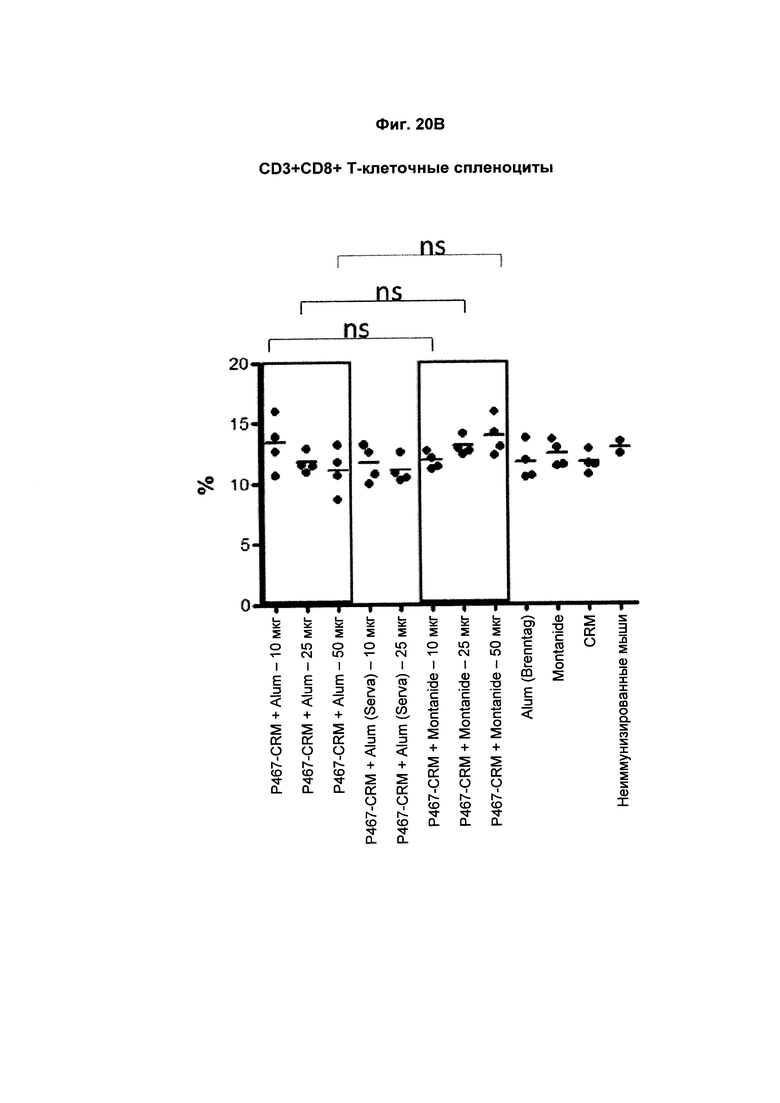
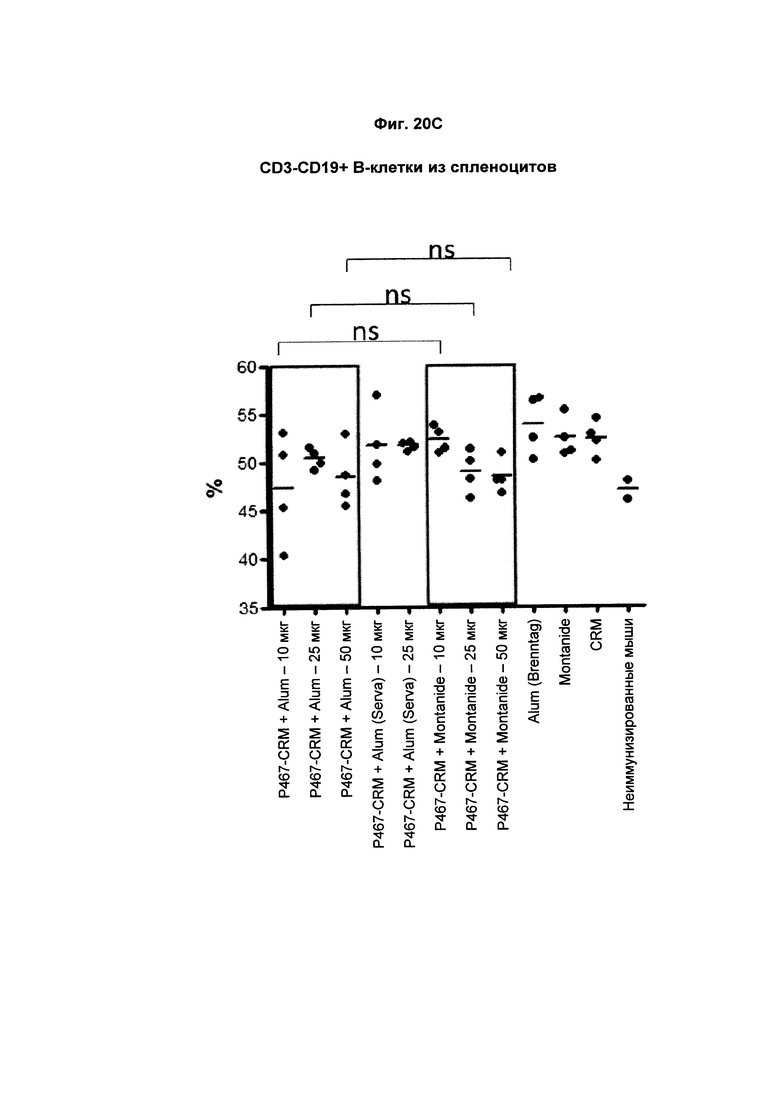
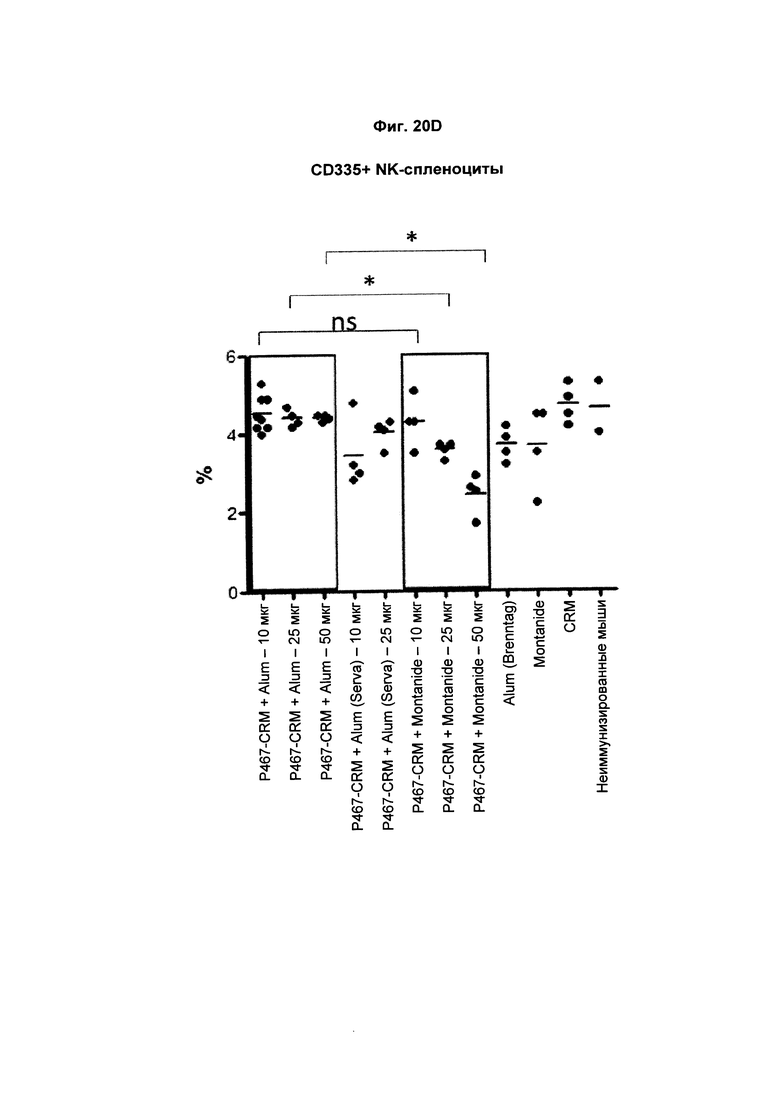
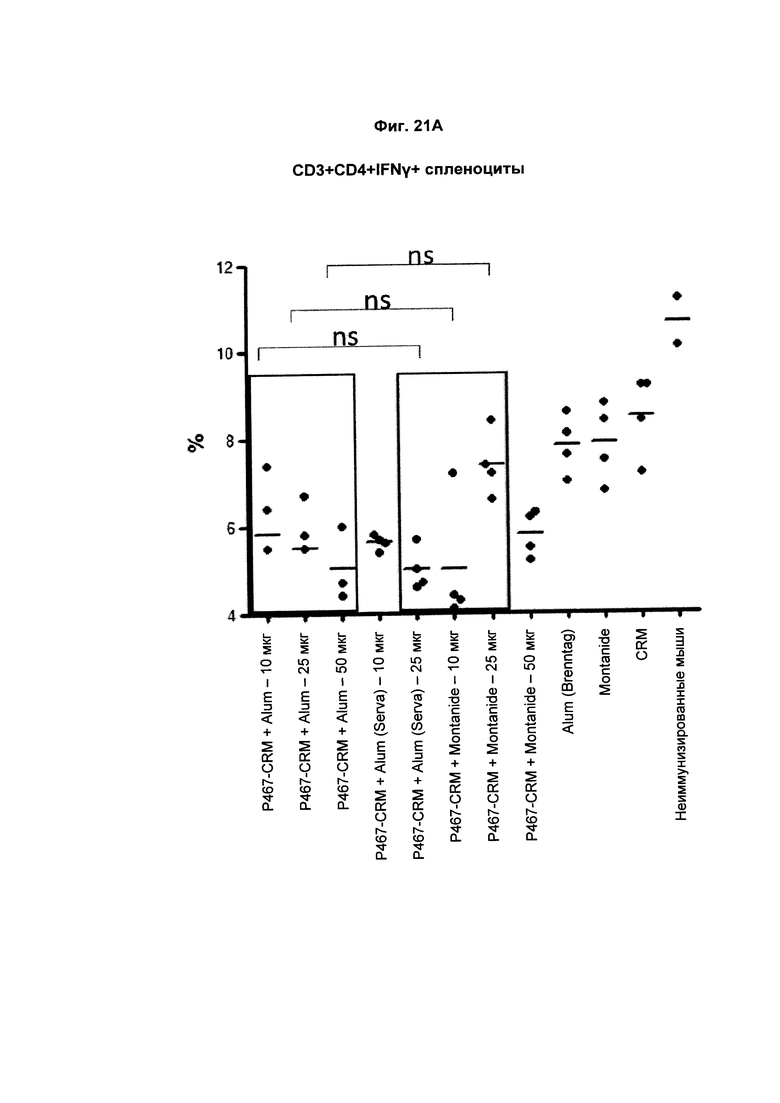
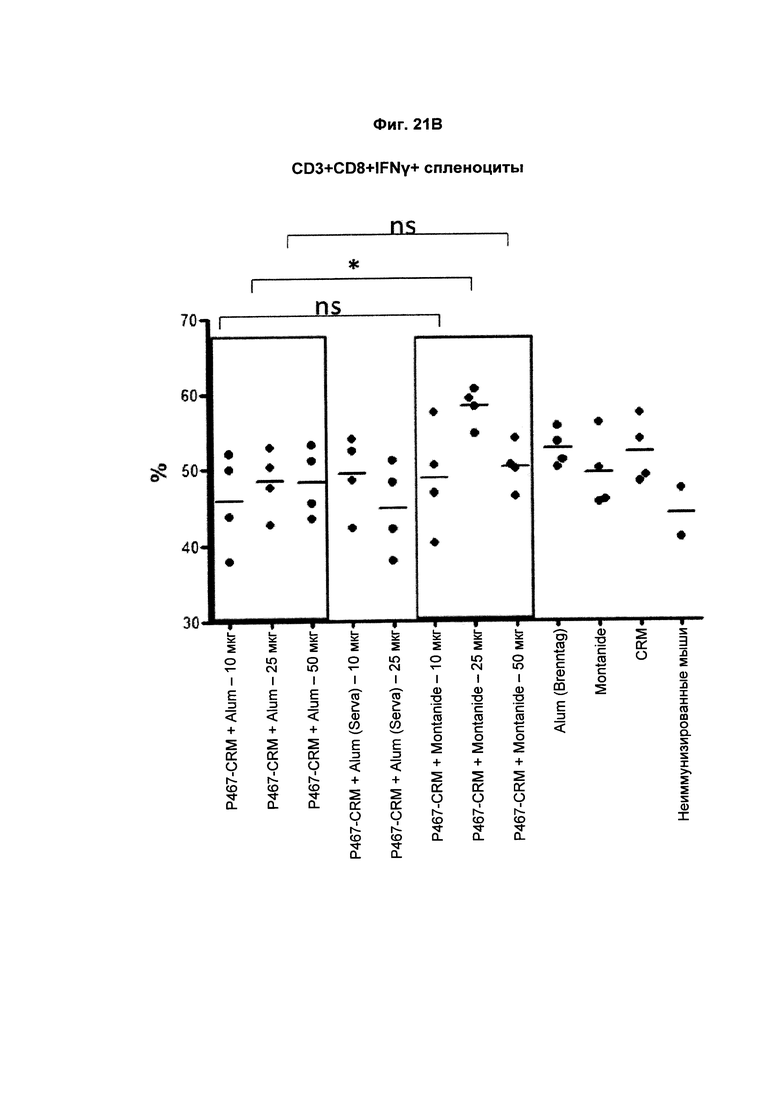
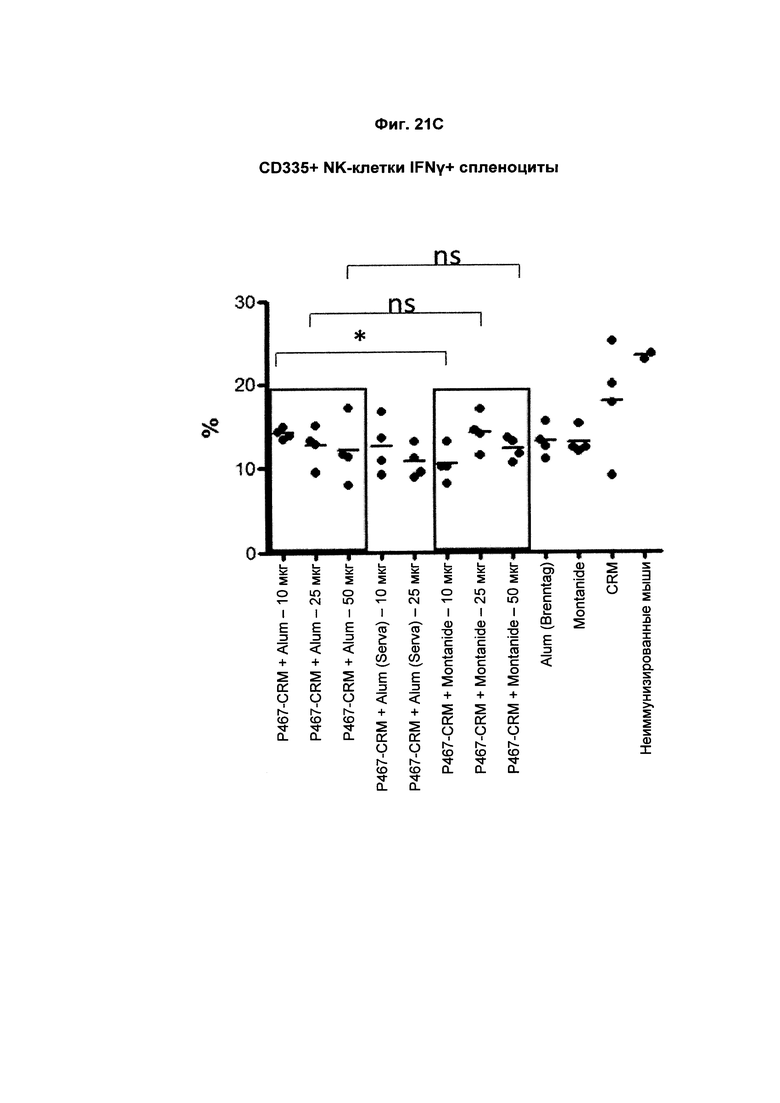
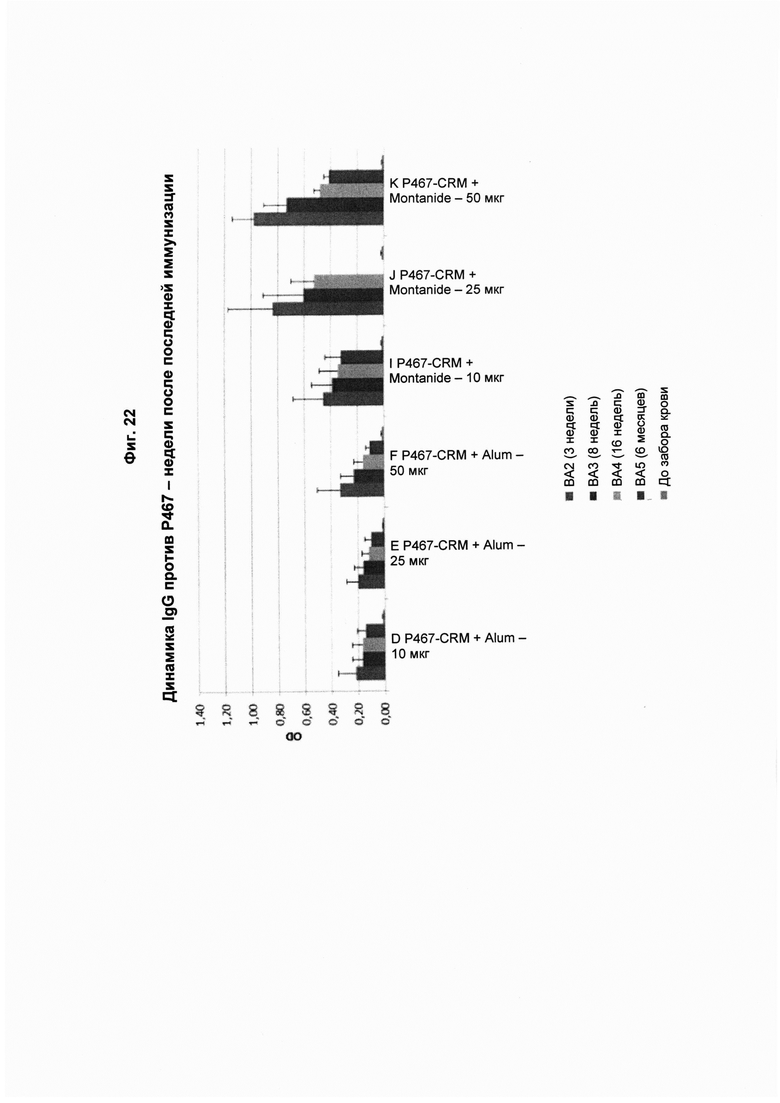
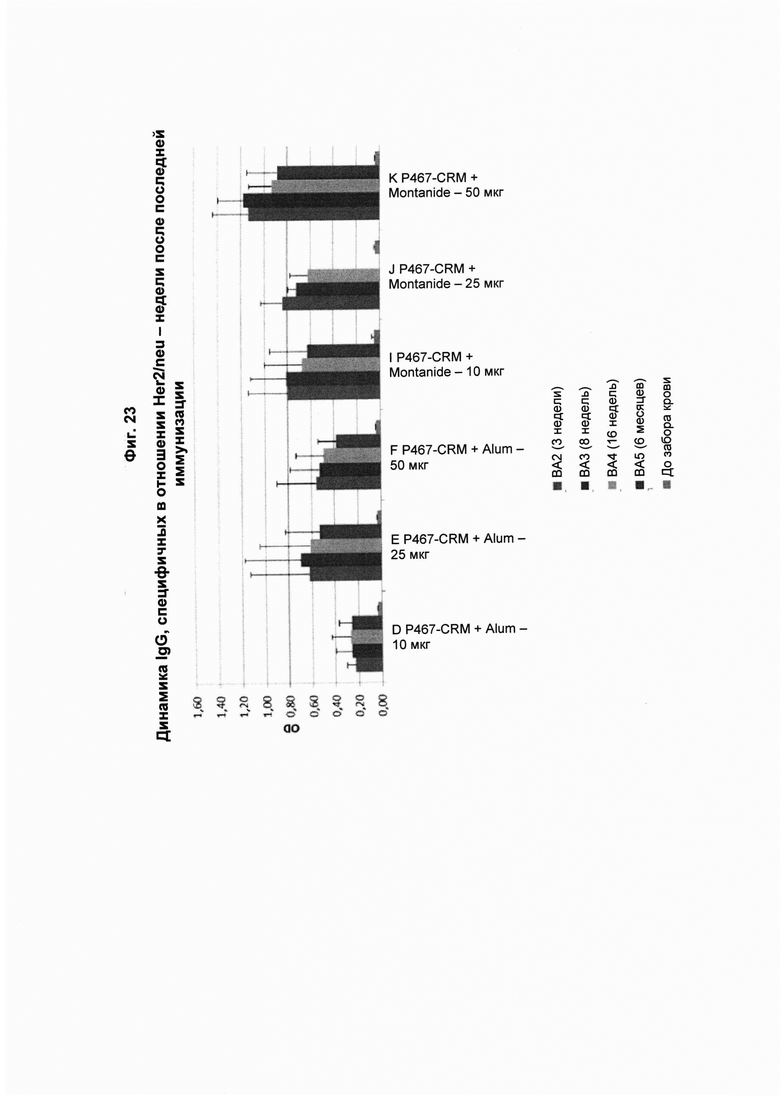
Группа изобретений относится к медицине, а именно к онкологии, и может быть использована для получения вакцинной композиции для лечения рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu. Вакцинная композиция содержит: (1) адъювант, представляющий собой эмульсию типа «вода в масле», которая представляет собой Montanide; и (2) по меньшей мере один слитый пептид, конъюгированный с белком-носителем, где белок-носитель представляет собой вариант дифтерийного токсина CRM197, имеющий последовательность SEQ ID NO: 61, и где по меньшей мере один слитый пептид содержит два или более несмежных B-клеточных эпитопа Her2/neu, выбранных из группы, состоящей из SEQ ID NO:1-7. Группа изобретений также включает фармацевтические композиции и способы применения вакцинных и/или фармацевтических композиций для лечения или предупреждения рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu, у пациента, нуждающегося в этом. Использование данной группы изобретений позволяет получить усиленный гуморальный иммунный ответ, стимулируя выработку антител, специфичных в отношении В-клеточных эпитопов и Her2/neu, при введении указанных пептидов, конъюгированных с CRM-197, и эмульсии типа «вода в масле». 4 н. и 14 з.п. ф-лы, 23 ил., 9 табл., 11 пр.
1. Вакцинная композиция, содержащая:
(1) адъювант, представляющий собой эмульсию типа «вода в масле»,
которая представляет собой Montanide; и (2) по меньшей мере один слитый пептид, конъюгированный с белком-носителем, где белок-носитель представляет собой вариант дифтерийного токсина CRM197, имеющий последовательность SEQ ID NO: 61, и где по меньшей мере один слитый пептид содержит два или более несмежных B-клеточных эпитопа
Her2/neu, выбранных из группы, состоящей из SEQ ID NO:1-7.
2. Вакцинная композиция по п. 1, где слитый пептид содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:8-14.
3. Вакцинная композиция по п. 2, где слитый пептид содержит аминокислотную последовательность SEQ ID NO:8 или SEQ ID NO:9.
4. Вакцинная композиция по п. 3, где слитый пептид состоит из аминокислотной последовательности SEQ ID NO:8 или SEQ ID NO:9.
5. Вакцинная композиция по любому из пп. 1-4, где белок-носитель содержит от 2 до 39 слитых пептидов.
6. Вакцинная композиция по п. 5, где белок-носитель содержит от 6 до 12 слитых пептидов.
7. Вакцинная композиция по любому из пп. 1-6, дополнительно содержащая ингибитор контрольных точек.
8. Фармацевтическая композиция для лечения или предупреждения рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu, где композиция содержит вакцинную композицию по любому из пп. 1-7 и фармацевтически приемлемый носитель.
9. Способ лечения или предупреждения рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu, у пациента, нуждающегося в этом, включающий стадию введения указанному пациенту эффективного количества вакцинной композиции по любому из пп. 1-7 или фармацевтической композиции по п. 8.
10. Способ по п. 9, где рак представляет собой рак молочной железы.
11. Способ по п. 9, где рак представляет собой рак желудка.
12. Применение вакцинной композиции по любому из пп. 1-7 в изготовлении лекарственного средства для лечения или предупреждения рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu, у пациента, нуждающегося в этом.
13. Применение по п. 12, где рак представляет собой рак молочной железы.
14. Применение по п. 12, где рак представляет собой рак желудка.
15. Вакцинная композиция по любому из пп. 1-7 для применения в лечении или предупреждении рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu, у пациента, нуждающегося в этом.
16. Фармацевтическая композиция по п. 8 для применения в лечении или предупреждении рака, характеризующегося экспрессией или сверхэкспрессией Her2/neu, у пациента, нуждающегося в этом.
17. Вакцинная композиция по п. 15, где рак представляет собой рак молочной железы.
18. Вакцинная композиция по п. 15, где рак представляет собой рак желудка.
| WO2011020604 A1, 24.02.2011 | |||
| Danishefsky S | |||
| et al | |||
| Development of Globo-H cancer vaccine // Acc Chem Res | |||
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| Jasinska J et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| Wagner S | |||
| et al | |||

