РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка на патент заявляет преимущество предварительной заявки на патент США № 62/128133, поданной 4 марта 2015 г., которая включена в данный документ посредством ссылки во всей своей полноте.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Область техники
Антигенсвязывающий димер, имеющий два полипептидных мономера, каждый из которых содержит молекулу одноцепочечного вариабельного фрагмента (scFv), шарнирную область и Fc-молекулу, который характеризуется высокой аффинностью и авидностью в отношении трансформирующего фактора роста β1 (TGFβ1), но не TGFβ2 или TGFβ3. Предлагаются композиции, содержащие антигенсвязывающий димер, и способы их применения для лечения заболеваний, в которые вовлечена активность TGFβ1.
Предпосылки изобретения
Многие тяжелые заболевания связаны с нарушениями функционирования TGFβ-индуцированного сигнального пути. Например, считается, что повышенный уровень TGFβ в тканях является фактором развития идиопатического легочного фиброза и миокардиального фиброза. Кроме того, высокие локальные уровни TGFβ в тканях могут способствовать поддержанию жизнедеятельности и прогрессированию опухолей из некоторых типов раковых клеток. Таким образом, подавление передачи сигналов, опосредованной TGFβ, может снижать жизнеспособность таких опухолевых клеток.
Изоформы TGFβ представляют собой гомодимерные молекулы размером ~ 25 кДа со сходным структурным каркасом, в котором два мономера ковалентно связаны посредством дисульфидного мостика. Изоформы млекопитающих характеризуются идентичностью последовательностей, составляющей 70-82%, однако обладают неперекрывающимися видами активности в развитии сосудов и регуляции функции иммунных клеток. У людей были описаны три изоформы TGFβ: TGFβ1, TGFβ2 и TGFβ3 (номера доступа в Swiss Prot P01137, P08112 и P10600 соответственно). TGFβ1 и TGFβ3 запускают каскад передачи сигнала в клетке при связывании с внеклеточными доменами двух трансмембранных рецепторов, известных как рецепторы TGFβ I и II типа. TGFβ2 может связываться с рецепторами TGFβ I и II типа, а также с рецептором TGFβ III типа.
Антитела, которые могут связываться с человеческими TGFβ1, TGFβ2 и TGFβ3, были протестированы в отношении клинического применения. Например, Grütter и соавт. раскрыли GC1008, человеческое моноклональное антитело IgG4 (Mab; т. е. GC1008), в клинических исследованиях в отношении лечения злокачественных опухолей и фиброзных заболеваний. Proc. Nat'l Acad. Sci. USA 105(51): 20251-56 (2008). GC1008 представляет собой "панспецифичное" антитело, нейтрализующее TGFβ, поскольку оно может нейтрализовать все три изоформы человеческого TGFβ. Антитела, которые избирательно нейтрализуют TGFβ1, раскрыты, например, в патенте США № 6492497 и патенте США № 7151169, которые включены в данное раскрытие посредством ссылки. Метелимумаб, также известный как CAT192 (IgG4), представляет собой человеческое моноклональное антитело IgG4, которое избирательно нейтрализует TGF-β1. См., например, патент США № 6492497. Метелимумаб тестировали в отношении лечения диффузного кожного системного склероза, также известного как склеродермия, однако он продемонстрировал недостаточную эффективность.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем раскрытии предлагаются TGFβ1-связывающие димеры scFv-Fc, которые способны избирательно нейтрализовать человеческий TGFβ1. В одном варианте осуществления димеры scFv-Fc находятся в формате белков слияния scFv-Fc, состоящих из двух полипептидных мономеров, при этом каждый мономер содержит одноцепочечную Fv-область (scFv), шарнирную область и Fc-область. VH- и VL-домены димера scFv-Fc характеризуются более высокой аффинностью и авидностью в отношении TGFβ1 и более эффективно нейтрализуют TGFβ1, чем в случае применения в формате IgG1 или IgG4.
В одном варианте осуществления scFv-компонент может состоять из таких же VH- и VL-доменов, как VH- и VL-домены метелимумаба. Вариабельные домены в scFv-компоненте могут быть связаны друг с другом с помощью линкера, например, линкера типа [G4S]3. Каждый из scFv-компонентов димеров scFv-Fc может быть слит с Fc-областью с помощью шарнирной области, например, шарнирной области человеческого IgG1 или IgG4. Мономеры димера могут быть ковалентно связаны дисульфидной связью между цистеиновыми остатками в шарнирной области. В другом варианте осуществления димеры scFv-Fc могут иметь структурные различия с метелимумабом, выражающиеся прежде всего в отсутствии CH1- и CL-доменов и наличии линкера между VH- и VL-доменами. Димеры scFv-Fc преимущественно проявляют кажущуюся аффинность в отношении TGFβ1, почти на два порядка величины превышающую таковую у scFv, содержащего такие же VH- и VL-домены (CAT191(scFv), показанного под SEQ ID NO: 12), в биологическом анализе активности с использованием клеток A549. Кроме того, димеры scFv-Fc проявляют кажущуюся аффинность в отношении TGFβ1, более чем на три порядка величины превышающую таковую у антитела в формате IgG, содержащего такие же VH- и VL-домены (например, CAT192), в биологическом анализе с использованием клеток A549. Димеры scFv-Fc также проявляют желаемые стабильность и фармакокинетические свойства. Ввиду своего относительно малого размера и удлиненного периода полувыведения из сыворотки крови димеры scFv-Fc являются особенно применимыми в терапевтических путях применения.
Соответственно, настоящее изобретение направлено на выделенный связывающий белок, содержащий вариабельный домен, который способен связываться с TGFβ1, где связывающий белок характеризуется Kd для человеческого TGFβ1, по меньшей мере на приблизительно 50% более низкой, чем Kd того же самого связывающего белка для человеческого TGFβ2, согласно измерениям по методу поверхностного плазмонного резонанса.
В другом варианте осуществления настоящее изобретение направлено на выделенный связывающий белок, содержащий вариабельный домен, который способен связываться с TGFβ1, где связывающий белок характеризуется Kd для человеческого TGFβ1, по меньшей мере на приблизительно 50% более низкой, чем Kd того же самого связывающего белка для человеческого TGFβ3, согласно измерениям по методу поверхностного плазмонного резонанса.
В дополнительном варианте осуществления настоящее изобретение направлено на выделенный связывающий белок, содержащий вариабельный домен, который способен связываться с TGFβ1, где связывающий белок характеризуется Kd для человеческого TGFβ1, по меньшей мере на приблизительно 50% более низкой, чем Kd того же самого связывающего белка для человеческого TGFβ2, и по меньшей мере на приблизительно 50% более низкой, чем Kd того же самого связывающего белка для человеческого TGFβ3, согласно измерениям по методу поверхностного плазмонного резонанса.
В дополнительном варианте осуществления настоящее изобретение направлено на выделенный связывающий белок, который связывается с TGFβ1, где связывающий белок содержит первую полипептидную цепь и вторую полипептидную цепь, при этом каждая из первой и второй полипептидных цепей имеет следующую формулу:
(VD1)-(линкер 1)n-(VD2)-(линкер 2)m-(шарнирная область)p-(Fc-область),
где VD1 содержит первый вариабельный домен, выбранный из группы, состоящей из VL-домена, выделенного из антитела, способного связываться с TGFβ1, и VH-домена, выделенного из антитела, способного связываться с TGFβ1, и VD2 содержит второй вариабельный домен, выбранный из группы, состоящей из VL-домена, выделенного из антитела, способного связываться с TGFβ1, и VH-домена, выделенного из антитела, способного связываться с TGFβ1; и где n равняется 0 или 1, m равняется 0 или 1, и p равняется 0 или 1.
В одном варианте осуществления настоящее изобретение направлено на выделенный TGFβ1-связывающий димер scFv-Fc, который избирательно связывается с TGFβ1. Димер scFv-Fc может содержать два полипептидных мономера, каждый из которых имеет следующую формулу в направлении от N-конца к C-концу: (VH-домен)-(линкер)-(VL-домен)-(шарнирная область)-(Fc-область). В другом варианте осуществления раскрыт выделенный связывающий белок, связывающийся с TGFβ1, который содержит первую полипептидную цель и вторую полипептидную цепь. Как первая, так и вторая полипептидные цепи могут иметь следующую формулу в направлении от N-конца к C-концу: (VH-домен)-(линкер 1)n-(VL-домен)-(линкер 2)m-(шарнирная область)p-(Fc-область). P может равняться 0 или 1, n может равняться 0 или 1, и m может равняться 0 или 1. В одном аспекте первая и вторая полипептидные цепи могут быть идентичными и могут образовывать димер.
В другом варианте осуществления раскрытый TGFβ1-связывающий белок может содержать полипептидную цепь, имеющую следующую формулу в направлении от N-конца к C-концу: (VH-домен)-(линкер 1)n-(VL-домен)-(линкер 2)m-(шарнирная область)p-(Fc-область), где p может равняться 0 или 1, n может равняться 0 или 1, и m может равняться 0 или 1.
VH-домен раскрытого связывающего белка может содержать область 1, определяющую комплементарность, вариабельного домена тяжелой цепи (HCDR1), область 2, определяющую комплементарность, вариабельного домена тяжелой цепи (HCDR2) и область 3, определяющую комплементарность, вариабельного домена тяжелой цепи (HCDR3). В одном аспекте HCDR1 может иметь аминокислотную последовательность SEQ ID NO: 22, HCDR2 может иметь аминокислотную последовательность SEQ ID NO: 23, и HCDR3 может иметь аминокислотную последовательность SEQ ID NO: 24, SEQ ID NO: 25, SEQ ID NO: 26 или SEQ ID NO: 30.
Каркасные области VH-домена могут быть выбраны из последовательности вариабельного домена тяжелой цепи зародышевого типа. VH-домен может быть выбран, например, из последовательностей человеческого VH-домена, указанных под SEQ ID NO: 1 или SEQ ID NO: 2, или его варианта, имеющего модификации до четырех аминокислот включительно.
VL-домен раскрытого связывающего белка может содержать область 1, определяющую комплементарность, вариабельного домена легкой цепи (LCDR1), область 2, определяющую комплементарность, вариабельного домена легкой цепи (LCDR2) и область 3, определяющую комплементарность, вариабельного домена легкой цепи (LCDR3). В одном аспекте LCDR1 может иметь аминокислотную последовательность SEQ ID NO: 27, LCDR2 может иметь аминокислотную последовательность SEQ ID NO: 28, и LCDR3 может иметь аминокислотную последовательность SEQ ID NO: 29.
Каркасные области VL-домена могут быть выбраны из последовательности вариабельного домена лямбда- или каппа-цепи зародышевого типа. VL-домен может быть выбран, например, из последовательностей человеческого Vκ-домена, указанных под SEQ ID NO: 5 или SEQ ID NO: 6, или его варианта, имеющего модификации до четырех аминокислот включительно. В одном варианте осуществления каждый полипептид димера может содержать VH-домен, указанный под SEQ ID NO: 1, и Vκ-домен, указанный под SEQ ID NO: 5, которые представляют собой соответственно VH- и VL-домены, присутствующие в метелимумабе.
В одном варианте осуществления вариабельные домены в scFv-компоненте могут быть связаны с помощью гибкого линкера, имеющего длину приблизительно 15 аминокислот. "Приблизительно" в данном контексте означает, что длина линкера может варьироваться в пределах до плюс-минус четырех дополнительных аминокислот включительно. В целях оптимальной гибкости линкер состоит преимущественно из глициновых и сериновых остатков. Например, линкер может представлять собой линкер типа [G4S]3. Линкер может иметь аминокислотную последовательность SGGGSGGGGSGGGGS (SEQ ID NO: 3), аминокислотную последовательность GGGGSGGGGSGGGGS (SEQ ID NO: 4) или их вариант, имеющий до четырех аминокислотных модификаций включительно. Для целей данного раскрытия "имеющий до x аминокислотных модификаций включительно" означает, что специалист в данной области может изменить аминокислоты в количестве до x включительно на другие аминокислоты без значительного изменения структуры и функции полипептида.
В другом варианте осуществления p равняется 1, и scFv-компонент соединен с Fc-областью с помощью шарнирной области. Шарнирная область может содержать аминокислотные последовательности, полученные из шарнирной области человеческого IgG1 или IgG4. Например, шарнирная область может содержать аминокислотную последовательность PKSCDKTHTCPPCPAPELLGGP (SEQ ID NO: 7) или ее вариант, имеющий до четырех аминокислотных модификаций включительно. В одном варианте осуществления длина шарнирной области может варьироваться от 3 до 15 аминокислот. Если шарнирная область получена из человеческого IgG1, то она может содержать аминокислотную последовательность CPPCP (SEQ ID NO: 21). Кроме того, вариант шарнирной области под SEQ ID NO: 7, который также представляет собой шарнирную область человеческого IgG1, может содержать аминокислотную последовательность CPPCP (SEQ ID NO: 21).
В другом варианте осуществления m равняется 1, и линкер 2 присутствует между scFv-компонентом и шарнирной областью. В одном аспекте линкер 2 может содержать аминокислотную последовательность GGSG (SEQ ID NO: 20) или ее вариант, имеющий до 2 аминокислотных модификаций включительно.
Fc-область может содержать два или три константных домена, например, CH2-домен и CH3-домен. Fc-область можно получить, например, из человеческого IgG1, человеческого IgG4 или варианта человеческого IgG1 или IgG4, имеющего до десяти аминокислотных модификаций включительно. В одном варианте осуществления каждой полипептид димера имеет последовательность, указанную под SEQ ID NO: 9. Структура димера scFv-Fc под SEQ ID NO: 9 показана на фиг. 2. Димер scFv-Fc может связываться с TGFβ1 избирательно. Димер scFv-Fc может характеризоваться кажущейся константой диссоциации, составляющей менее 1 нМ или даже менее 0,1 нМ. Кажущуюся константу диссоциации можно измерить, например, путем применения биологического анализа с использованием A549 или по методу поверхностного плазмонного резонанса.
В другом варианте осуществления раскрыт выделенный полинуклеотид, который может содержать нуклеотидную последовательность, кодирующую димер scFv-Fc. Выделенный полинуклеотид может представлять собой кДНК, рекомбинантную ДНК или синтетическую ДНК. Клетка-хозяин может содержать выделенную нуклеиновую кислоту. Клетка-хозяин может представлять собой человеческую клетку, такую как человеческая эмбриональная клетка почки 293 (HEK293) и линии клеток, происходящие из нее, или она может представлять собой клетку яичника китайского хомячка (CHO). Способ получения димера scFv-Fc может предусматривать культивирование клетки-хозяина в условиях, подходящих для выработки димера scFv-Fc. Димер scFv-Fc можно очищать. Степень чистоты может составлять 90%, 95%, 99%, 99,5% или более.
Димер scFv-Fc по настоящему изобретению может быть элементом композиции. Композиция может представлять собой фармацевтическую композицию. Фармацевтическая композиция может содержать терапевтически эффективное количество димера scFv-Fc. Композиция может дополнительно содержать один или несколько биологически активных компонентов, наполнителей или разбавителей.
Способ лечения заболевания или состояния, непосредственно или опосредованно обусловленного активностью TGFβ1, у человека может включать введение фармацевтической композиции, содержащей терапевтически эффективное количество димера scFv-Fc. Заболевание или состояние может быть выбрано из группы, состоящей из фиброзного заболевания, рака, иммуноопосредованного заболевания, например, диффузного кожного системного склероза, заболевания, характеризующегося нарушением ремоделирования костной ткани, заболевания почек и/или их комбинаций. Димер scFv-Fc можно применять в производстве лекарственного препарата для лечения заболевания или нарушения, выбранного из группы, состоящей из фиброзного заболевания, рака, иммуноопосредованного заболевания, например, диффузного кожного системного склероза, заболевания, характеризующегося нарушением ремоделирования костной ткани, заболевания почек и/или их комбинаций. Лечение заболевания или нарушения может включать нейтрализацию TGFβ1 или ингибирование передачи сигналов, опосредованной TGFβ1. Лечение заболевания или нарушения может включать ингибирование выработки фибронектина, выработки фактора роста эндотелия сосудов (VEGF), пролиферации эпителиальных клеток, пролиферации эндотелиальных клеток, пролиферации гладкомышечных клеток или подавления иммунитета, опосредованных TGFβ1. Лечение заболевания или нарушения может включать повышение активности естественных клеток-киллеров.
КРАТКОЕ ОПИСАНИЕ НЕКОТОРЫХ ПРОЕКЦИЙ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Графические материалы, представленные в данном документе, приведены с целью иллюстрации и не должны использоваться для ограничения объема настоящего изобретения.
На фиг. 1 представлены общие структуры различных форматов.
На фиг. 2 представлены результаты анализа связывания с TGFβ1 на Biacore, которые показывают утрату аффинности при превращении scFv(CAT191) в молекулу IgG4 полной длины (CAT192).
На фиг. 3 показаны результаты биологического анализа с использованием клеток A549, в котором сравниваются ингибиторные эффекты различных конструкций антител в отношении выработки IL-11, стимулируемой TGFβ1: scFv-диатело с линкером из 5 аминокислот (SEQ ID NO: 14); CAT191(scFv) (SEQ ID NO: 12); CAT191(scFv-Fc) (SEQ ID NO: 9) и CAT192(IgG4) (легкая цепь под SEQ ID NO: 10 и тяжелая цепь под SEQ ID NO: 11).
На фиг. 4 представлены результаты фармакокинетических тестов для определения периода полувыведения CAT191 (scFv-Fc) после внутривенного (IV) введения.
На фиг. 5 представлены результаты фармакокинетических тестов для определения периода полувыведения CAT191 (scFv-Fc) после внутрибрюшинного (IP) введения.
На фиг. 6 показаны результаты изучения TGFβ1-специфичного связывания CAT191(scFv-Fc), полученного из клеток CHO.
На фиг. 7 показаны результаты клеточного анализа активности CAT191(scFv-Fc), полученного из клеток CHO.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Раскрытые димеры scFv-Fc связываются с TGFβ1 и нейтрализуют его избирательно и с высокой аффинностью и авидностью. ScFv-области могут состоять из таких же VH- и VL-доменов, как и у метелимумаба. Димеры scFv-Fc преимущественно характеризуются более высокой эффективностью нейтрализации TGFβ1, чем в случае применения вариабельных доменов в других форматах. Ввиду своего относительно малого размера и удлиненного периода полувыведения из сыворотки крови димеры scFv-Fc по настоящему изобретению являются превосходными кандидатами для терапевтических путей применения.
Как используется в данном документе, "первый элемент и/или второй элемент" означает определенное раскрытие первого или второго элемента по отдельности или первого и второго элементов в комбинации. Формы существительного единственного числа включают ссылку на формы множественного числа, если контекст четко не определяет иное.
"Выделенный" полинуклеотид (или нуклеиновая кислота) или белок удален от его природной формы и/или изменен по сравнению с ней с помощью технологий генной инженерии. "Очищенные" нуклеиновая кислота или белок могут быть фактически чистыми, например, по меньшей мере на 90% чистыми, или находиться в однородной форме.
"Избирательное связывание" или "избирательно связывающийся" с человеческим TGFβ1 означает, что связывающий белок (например, димер scFv-Fc) способен связываться с человеческим TGFβ1 с более высокой аффинностью, чем при связывании с человеческим TGFβ2 или человеческим TGFβ3, например, с константой диссоциации для человеческого TGFβ1, по меньшей мере на 50% более низкой, чем его константа диссоциации для человеческого TGFβ2 или человеческого TGFβ3, согласно измерениям по методу поверхностного плазмонного резонанса.
Димеры scFv-Fc
В одном варианте осуществления вариабельные домены димера scFv-Fc по настоящему изобретению содержат области, определяющие комплементарность (CDR), из CDR, раскрытых в патенте США № 6492497 (например, SEQ ID NO: 11-19 патента США № 6492497), включенном в данный документ посредством ссылки. CDR-области перечислены ниже:
HCDR1 SYGMH SEQ ID No. 22
HCDR2 VISYDGSIKYYADSVKG SEQ ID No. 23
HCDR3 TGEYSGYDTSGVEL SEQ ID No. 24
TGEYSGYDTDPQYS SEQ ID No. 25
TGFYSGYDTPASPD SEQ ID No. 26
LCDR1 RASQGIGDDLG SEQ ID No. 27
LCDR2 GTSTLQS SEQ ID No. 28
LCDR3 LQDSNYPLT SEQ ID No. 29
Неожиданно был выявлен консенсусный связывающий мотив HCDR3, имеющий следующую последовательность:
HCDR3 TGX1YSGYDTX2X3X4X5X6 SEQ ID No. 30
При этом: X1 может представлять собой любую аминокислоту (предпочтительно E или F) или отсутствовать,
X2 может представлять собой любую аминокислоту (предпочтительно S, D или P) или отсутствовать,
X3 может представлять собой любую аминокислоту (предпочтительно G, P или A) или отсутствовать,
X4 может представлять собой любую аминокислоту (предпочтительно V, Q или S) или отсутствовать,
X5 может представлять собой любую аминокислоту (предпочтительно E, Y или P) или отсутствовать,
X6 может представлять собой любую аминокислоту (предпочтительно L, S или D) или отсутствовать.
VH-домен содержит HCDR1 под SEQ ID NO: 22, HCDR2 под SEQ ID NO: 23 и одну из HCDR3, выбранных из группы, состоящей из SEQ ID NO: 24, SEQ ID NO: 25, SEQ ID NO: 26 и SEQ ID NO: 30. Последовательности CDR могут быть отделены в любом месте каркасными областями в количестве от одной до четырех в следующем порядке от N-конца: FW1 - CDR1 - FW2 - CDR2 - FW3 - CDR3 - FW4. Каркасные области VH-домена могут быть выбраны из последовательности вариабельного домена тяжелой цепи зародышевого типа. В одном варианте осуществления последовательности FW-области могут быть выбраны из одной и той же последовательности человеческого вариабельного домена тяжелой цепи зародышевого типа. VL-домен содержит LCDR1 под SEQ ID NO: 7, LCDR2 под SEQ ID NO: 28 и LCDR3 под SEQ ID NO: 29. Каркасные области VL-домена могут быть выбраны из последовательности вариабельного домена лямбда- или каппа-цепи зародышевого типа, например, из одной и той же последовательности человеческого вариабельного домена лямбда- или каппа-цепи зародышевого типа. В настоящее время из уровня техники известно приблизительно 40 последовательностей вариабельных доменов тяжелых цепей зародышевого типа, а также приблизительно 40 последовательностей вариабельных доменов каппа-цепей зародышевого типа и приблизительно 30 последовательностей вариабельных доменов лямбда-цепей зародышевого типа, например, VH3, Vκ1, VH 1-69 и VH 1-e.
В другом варианте осуществления составные VH- или VL-домены можно получить путем применения последовательностей CDR, раскрытых в данном документе. Например, кристаллические структуры VH- или VL-доменов можно применять в качестве ориентира для получения составного домена с применением последовательностей CDR из одного антитела и с применением FW-областей зародышевого типа из другого антитела. Более подробную информацию можно найти в публикации заявки на патент США № 20020099179; а также в Homes and Foote, J Immunol. 1997 Mar 1;158(5):2192-201, обе из которых настоящим включены в данное раскрытие посредством ссылки.
Димеры scFv-Fc по настоящему изобретению могут состоять из таких же VH- и VL-доменов, как и у метелимумаба, имеющих последовательности, указанные под SEQ ID NO: 1 и SEQ ID NO: 5 соответственно. VH-домен может быть замещен VH-доменом, имеющим последовательности, указанные под SEQ ID NO: 2; VL-домен может быть замещен VL-доменом, имеющим последовательности, указанные под SEQ ID NO: 6. Эти VH- и VL-домены раскрыты в патенте США № 6492497 (например, SEQ ID NO: 4, 6, 8 и 10), включенном в данный документ посредством ссылки.
"Вариабельный домен" (VD) относится к гипервариабельному связывающему домену иммуноглобулина или лигандсвязывающему домену рецептора, участвующему в связывании с антигеном/лигандом, как известно специалистам в данной области. Вариабельные домены обычно называют по их расположению в иммуноглобулине или происхождению; например, вариабельные домены легкой цепи иммуноглобулина (VL), вариабельные домены тяжелой цепи иммуноглобулина (VH), вариабельные домены тяжелой цепи верблюжьего иммуноглобулина (VHH).
"Вариант" вариабельного домена содержит аминокислотные добавления, замены и/или делеции по сравнению с эталонной последовательностью. "Вариант" VH- или VL-доменов может иметь до четырех таких аминокислотных модификаций включительно. Например, один из двух доменов может содержать аминокислотную замену, в то время как другой домен является немодифицированным, или оба из доменов могут содержать аминокислотные замены. Модификации, в ходе которых добавляются или удаляются аминокислотные остатки, можно выполнить на N-конце или C-конце VH- или VL-домена. Например, можно удалить N-концевой остаток VH-домена.
Например, можно производить до четырех аминокислотных замен включительно для деиммунизации димера scFv-Fc. Деиммунизацию можно осуществлять, например, в соответствии со способом Harding et al. (2010) mAbs 2: 256-265.
Например, остатки каркасной области VH- и/или VL-доменов можно заменять для повышения стабильности димеров scFv-Fc и/или уменьшения их склонности к агрегации. Недостаточная стабильность может влиять на способность экспрессируемых димеров scFv-Fc к правильному сворачиванию при рекомбинантной экспрессии, приводя к тому, что часть экспрессируемых антител является нефункциональной. Антитела с низкой стабильностью также могут быть предрасположены к образованию потенциально иммуногенных агрегатов или могут иметь ухудшенную авидность или сохраняемость. В частности, полипептиды scFv могут демонстрировать проблемы со стабильностью, растворимостью, экспрессией, агрегацией, продуктами распада и возможностью производства в целом в системах экспрессии как у бактерий, так и у млекопитающих. Аминокислотные замены в каркасной области, которые, как предполагается, повышают стабильность и/или снижают склонность к агрегации VH- и/или VL-домена, например, в полипептиде scFv, раскрыты, например, в WO 2007/109254. Замены в соответствующих остатках в VH- и VL-доменах по настоящему изобретению, как предполагается, аналогичным образом повышают стабильность димеров scFv-Fc и/или снижают их склонность к агрегации.
Замены, которые могут быть допустимыми, как предполагается, включают в себя те замены, в ходе которых аминокислота из SEQ ID NO: 1, 2, 5 или 6 будет замещена соответствующей аминокислотой, которая встречается в другой последовательности человеческого VH- или VL-домена зародышевого типа. Замена аминокислоты каркасной области на аминокислоту, встречающуюся в любой из этих последовательностей зародышевого типа, может быть допустимой. Например, остаток VH-домена под SEQ ID NO: 1 можно заменить аминокислотой, присутствующей в соответствующем положении в любой последовательности VH зародышевого типа, например, последовательности зародышевого типа из DP-10 (VH 1-69) или DP-88 (VH 1-e). Соответствующие положения в этом случае определяются путем выравнивания последовательностей между различными последовательностями зародышевого типа с помощью методик выравнивания, хорошо известных из уровня техники, например, ClustalW.
Дополнительные замены, которые, как предполагается, являются допустимыми, представляют собой замены, производимые в отношении аминокислоты, большая часть боковой цепи которой подвергается воздействию растворителя, что определяется в анализе трех сокристаллических структур. Площадь поверхности остатка, доступную для растворителя, можно оценить с использованием методик, хорошо известных из уровня техники. Кроме того, предполагается, что замены аминокислот, погруженных вглубь вариабельных доменов, будут более допустимыми, если боковая цепь аминокислоты не создает стерического несоответствия с прилегающими остатками. По этой причине аминокислоты, погруженные вглубь, как правило, заменяют аминокислотами с боковыми цепями аналогичного или меньшего размера. Например, замена погруженного вглубь остатка Ile на Leu, Val, Ala или Gly, как предполагается, является допустимой. Возможное стерическое несоответствие, создаваемое заменой, можно предсказать путем анализа трех сокристаллических структур. Дополнительные замены, которые, как предполагается, являются допустимыми, представляют собой замены, при которых сохраняются существующие электростатические взаимодействия в вариабельных доменах, например, диполь-дипольные взаимодействия, взаимодействия индуцированных диполей, водородные связи или ионные связи.
Дополнительные аминокислотные замены вариабельных доменов включают в себя замены, которые, как предполагается, придают новые полезные свойства антителам или их антигенсвязывающим фрагментам. Например, предполагаемые сайты N-гликозилирования в VH- и/или VL-доменах можно удалить для предупреждения или снижения образования N-гликоформ. Аминоконцевой остаток можно заменить остатком Gln для того, чтобы вызвать пироглутамилирование, которое может уменьшить число вариантов, отличающихся зарядом. Аминокислотные замены можно применять для снижения изоэлектрической точки, в результате чего может уменьшаться, например, скорость выведения полипептидных антител IgG.
Поверхностные остатки вариабельных доменов можно заменять, например, остатками Cys или Lys, которые затем можно ковалентно модифицировать и соединить с молекулами, придающими полезные характеристики антителам или их антигенсвязывающим фрагментам, например, выявляемой меткой, токсином, нацеливающим фрагментом или белком. Например, остаток Cys можно соединить с цитотоксическим лекарственным средством с образованием конъюгата с лекарственным средством. Остатки Cys также можно соединить с молекулами, которые увеличивают период полувыведения из сыворотки крови, например, полиэтиленгликолем (PEG) или сывороточным альбумином. Такие аминокислотные модификации рассматриваются, например, в Beck et al. (2010) Nature 10: 345-52.
Выявляемые метки включают в себя радиоактивные метки, такие как 131I или 99Tc, которые можно присоединить к антителам или их антигенсвязывающим фрагментам с помощью способов, известных из уровня техники. Метки также включают в себя ферментные метки, такие как пероксидаза хрена. Метки дополнительно включают в себя химические фрагменты, такие как биотин, которые можно выявить посредством связывания со специфическим когнатным выявляемым фрагментом, например, меченым авидином. Можно присоединить другие фрагменты, которые облегчают очистку. Например, антитела или их антигенсвязывающие фрагменты можно метить His с помощью хорошо известных способов рекомбинантной модификации и экспрессии.
VH- и VL-домены димеров scFv-Fc связаны друг с другом с помощью линкера, называемого в данном документе линкером 1. Линкеры, подходящие для получения scFv-фрагмента, хорошо известны из уровня техники. См., например, Bird et al. (1988) Science, 242: 423-426; Huston et al. (1988) Proc. Nat'l Acad. Sci. USA 85: 5879-5883. Это можно выполнить, например, путем внутрирамочного слияния кодирующих нуклеиновых кислот и экспрессии белка слияния в подходящей клетке-хозяине. Подходящие линкеры включают в себя линкеры типа [G4S]3. Линкеры типа [G4S]3 состоят из повторяющихся звеньев из глициновых и сериновых остатков. Например, такие линкеры могут иметь последовательность SGGGSGGGGSGGGGS (SEQ ID NO: 3) или GGGGSGGGGSGGGGS (SEQ ID NO: 4) или их вариант, имеющий до четырех аминокислотных модификаций включительно. Модификации могут включать в себя делеции или вставки, которые изменяют длину линкера, или аминокислотные замены, предпочтительно с Gly на Ser или наоборот. Линкеры типа [G4S]3 широко применяли для связывания вариабельных доменов в структуре scFv, поскольку данные линкеры являются гипоаллергенными и вызывают минимальные конформационные нарушения в вариабельных доменах. См., например, Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85: 5879-83.
В димерах scFv-Fc между VL-доменом и шарнирной областью необязательно вставлена короткая линкерная последовательность, называемая в данном документе линкером 2. Эта линкерная последовательность повышает гибкость scFv-компонента относительно Fc-компонента. В одном варианте осуществления линкер 2 имеет последовательность GGSG (SEQ ID NO: 20). Подходящие модификации линкера GGSG включают в себя изменение его длины на одну-четыре аминокислоты или замену одной-двух аминокислот, предпочтительно с Gly на Ser или наоборот.
Шарнирная область представляет собой гибкий домен, который соединяет scFv-часть с Fc-областью. Гибкость шарнирной области в молекулах IgG и IgA способствует тому, что Fab-фрагменты принимают взаимное расположение с широким диапазоном углов между ними, что позволяет связываться с эпитопами, разделенными различными расстояниями. Подходящая шарнирная область включает в себя, например, шарнирную область человеческого IgG1, имеющую аминокислотную последовательность PKSCDKTHTCPPCPAPELLGGP (SEQ ID NO: 7). Эта последовательность соответствует части верхней шарнирной области, средней шарнирной области и N-концевой части CH2-домена человеческого IgG1, как раскрыто, например, на фиг. 4B патента США № 8048421. Шарнирная область человеческого IgG1 содержит два остатка Cys, которые могут образовывать дисульфидные связи с остатками Cys шарнирной области в соответствующем мономере. Часть шарнирной области человеческого IgG1, которая образует дисульфидные связи, содержит аминокислотную последовательность CPPCP (SEQ ID NO: 21). Варианты шарнирной области человеческого IgG1 могут содержать эту последовательность.
ScFv-компонент подвергнут внутрирамочному слиянию с Fc-областью, которая образует Fc-компонент димера. Подходящие Fc-области содержат две или три константные области. Fc-области включают в себя области из человеческого IgG1, указанные под SEQ ID NO: 8, или IgG4, указанные в CH2- и CH3-доменах под SEQ ID NO: 11. Fc-область антитела опосредует его период полувыведения из сыворотки крови и эффекторные функции, такие как комплементзависимая цитотоксичность (CDC), антителозависимая клеточная цитотоксичность (ADCC) и антителозависимый клеточный фагоцитоз (ADCP).
Модификации можно производить в шарнирной области и Fc-области для улучшения различных свойств димеров scFv-Fc. В одном варианте осуществления помимо модификаций шарнирной области можно модифицировать одну, две, три, четыре, пять или до десяти аминокислот встречающейся в природе человеческой Fc-области включительно. Например, Fc-область можно модифицировать для увеличения периода полувыведения из сыворотки крови димера scFv-Fc. Период полувыведения IgG зависит от его pH-зависимого связывания с рецептором FcRn. FcRn, который экспрессируется на поверхности эндотелиальных клеток, связывается с IgG pH-зависимым образом и защищает его от разрушения. Например, было показано, что мутации, расположенные на границе между CH2- и CH3-доменами, увеличивают аффинность связывания с FcRn и период полувыведения IgG1 in vivo. Такие модификации рассматриваются, например, в Strohl WR., 2009. Optimization of Fc-mediated effector functions of monoclonal antibodies. Curr Opin Biotechnol. 20(6):685-91; и Vaccaro C. et al., 2005. Engineering the Fc region of immunoglobulin G to modulate in vivo antibody levels. Nat Biotechnol. 23(10):1283-8.
Другие модификации шарнирной области и/или Fc-области могут усиливать или ослаблять эффекторные функции. Четыре изотипа человеческого IgG связываются с активирующими Fcγ-рецепторами (FcγRI, FcγRIIa, FcγRIIIa), ингибирующим FcγRIIb-рецептором и первым компонентом системы комплемента (C1q) с различными значениями аффинности, что обуславливает осуществление различных эффекторных функций. Например, связывание IgG с FcγR или C1q зависит от остатков, расположенных в шарнирной области и CH2-домене IgG. Одна или несколько аминокислотных замен этих остатков могут влиять на эффекторную функцию путем модулирования взаимодействия IgG с FcγR или C1q. Известны другие замены, которые влияют на эффекторную функцию. Эти модификации рассматриваются, например, в Strohl (2009) "Optimization of Fc-mediated effector functions of monoclonal antibodies," Curr. Opin. Biotechnol. 20:685-91.
Иллюстративные модификации шарнирной области и/или Fc-области обобщены в таблице 1.
Таблица 1. Иллюстративные модификации шарнирной области и Fc-области
1. Hinton et al. (2004) J. Biol. Chem. 279(8):6213-16.
2. Vaccaro et al. (2005) Nature Biotechnol. 23(10):1283-88.
3. Armour et al. (1999) Eur. J. Immunol. 29(8):2613-24.
4. Shields et al. (2001) J. Biol. Chem. 276(9):6591-604.
5. Idusogie et al. (2000) J. Immunol. 164(8):4178-84.
6. Idusogie et al. (2001) J. Immunol. 166(4):2571-75.
7. Lazar et al. (2006) Proc. Nat'l Acad. Sci. USA 103(11): 4005-10.
8. Ryan et al. (2007) Mol. Cancer Ther. 6: 3009-18.
9. Datta-Mannan et al. (2007) Drug Metab. Dispos. 35: 86-94.
10. Steurer et al. (1995) J. Immunol. 155(3):1165-74.
11. Richards et al. (2008) Mol. Cancer Ther. 7(8):2517-27.
12. Labrijn et al. (2009) Nature Biotechnol. 27(8):767-71.
Кроме того, рекомбинантные аминокислотные модификации можно применять для снижения структурной однородности экспрессируемых полипептидов. Иллюстративным примером является Peters et al. (2012) J. Biol. Chem. 287(29): 24525-33, в котором раскрыты замены Cys на Ser в шарнирной области IgG4, уменьшающие гетерогенность дисульфидных связей и повышающие термостабильность Fab-домена. Аналогично, Zhang et al. (2010) Anal. Chem. 82: 1090-99 раскрывают конструирование шарнирной области IgG2 с целью ограничения перестановки дисульфидных связей и образования структурных изомеров в терапевтических путях применения. Аминокислотные модификации CH3-домена также можно применять для удаления карбоксиконцевых остатков Lys с целью снижения числа вариантов, отличающихся зарядом. Аминокислотные модификации также можно применять для улучшения фармакологической функции рекомбинантных антител или их антигенсвязывающих фрагментов. Например, аминокислотные модификации можно применять для повышения активации системы комплемента, усиления антителозависимой клеточной цитотоксичности (ADCC) посредством повышения связывания с FcγRIIIA или снижения связывания с FcγRIIIB и/или увеличения периода полувыведения из сыворотки крови посредством повышения связывания с FcRn. Такие аминокислотные модификации рассматриваются, например, в Beck et al. (2010) Nature 10: 345-52.
Нуклеиновые кислоты и способы получения димеров scFv-Fc
В дополнительном аспекте настоящего изобретения предлагаются нуклеиновые кислоты, кодирующие димеры scFv-Fc. Выделенная нуклеиновая кислота может представлять собой, например, синтетическую ДНК, мРНК, не встречающуюся в природе, или кДНК. Примеры включают нуклеиновые кислоты, кодирующие VH- и VL-домены, указанные под SEQ ID NO: 3, 5, 7 и 9 в патенте США № 6492497. Дополнительные нуклеиновые кислоты включают в себя последовательность, указанную под SEQ ID NO: 13 в настоящем изобретении, которая кодирует диатело с линкером из 5 аминокислот, указанное под SEQ ID NO: 14, и последовательность, указанную под SEQ ID NO: 15, которая кодирует димер пептида с лейциновыми застежками, имеющий аминокислотную последовательность, указанную под SEQ ID NO: 16. Дополнительные нуклеиновые кислоты включают в себя последовательность, указанную под SEQ ID NO: 17, которая кодирует CAT191(scFv-Fc), имеющий аминокислотную последовательность, указанную под SEQ ID NO: 9. Нуклеиновую кислоту можно вставить в плазмиду, вектор или кассету транскрипции или экспрессии. Можно получить нуклеиновые кислоты, кодирующие димеры scFv-Fc, и экспрессируемые антитела можно протестировать с помощью традиционных методик, хорошо известных из уровня техники, таких как раскрытые в Borsi et al. (2002) Int. J. Cancer 102: 75-85.
Рекомбинантная клетка-хозяин может содержать одну или несколько вышеуказанных конструкций. Способы получения димеров scFv-Fc включают обеспечение экспрессии кодирующей нуклеиновой кислоты в клетке-хозяине в условиях, подходящих для выработки димеров scFv-Fc, и извлечение антител. Способ извлечения антител может включать выделение и/или очистку антител. Способ получения может включать составление антител в виде композиции, содержащей по меньшей мере один дополнительный компонент, такой как фармацевтически приемлемый наполнитель.
Предполагается, что термин "рекомбинантная клетка-хозяин" (или просто "клетка-хозяин"), используемый в данном документе, относится к клетке, в которую была введена экзогенная ДНК. Следует понимать, что такие термины предназначены для обозначения не только конкретной рассматриваемой клетки, но и потомства такой клетки. Поскольку в последующих поколениях могут иметь место определенные модификации вследствие мутации либо влияния окружающей среды, такое потомство в действительности может не быть идентичным родительской клетке, но по-прежнему включаться в объем термина "клетка-хозяин", используемого в данном документе. Клетки-хозяева предпочтительно включают в себя прокариотические и эукариотические клетки, выбранные из любого из царств живых организмов. Предпочтительные эукариотические клетки включают в себя клетки простейших, грибов, растений и животных. Клетки-хозяева наиболее предпочтительно включают в себя без ограничений линию прокариотических клеток E. coli; линии клеток млекопитающих CHO, HEK 293 и COS; линию клеток насекомых Sf9 и клетку гриба Saccharomyces cerevisiae.
Можно выбрать или сконструировать подходящие векторы, содержащие нуклеиновую кислоту, кодирующую димеры scFv-Fc, и содержащие соответствующие регуляторные последовательности, в том числе промоторные последовательности, терминаторные последовательности, последовательности полиаденилирования, энхансерные последовательности, маркерные гены и другие последовательности, при необходимости. Векторы могут представлять собой, например, плазмиды, фаги, фагмиды, аденовирусы, AAV, лентивирусы. Из уровня техники хорошо известны методики и протоколы манипуляции с нуклеиновой кислотой, например, при получении конструкций нуклеиновых кислот, мутагенезе, секвенировании, введении ДНК в клетки и экспрессии генов.
Предполагается, что термин "вектор", используемый в данном документе, относится к молекуле нуклеиновой кислоты, способной к транспорту другой нуклеиновой кислоты, с которой она была связана. Одним типом вектора является "плазмида", которая относится к кольцевой двухнитевой петле ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, где в вирусный геном могут быть лигированы дополнительные сегменты ДНК. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую их вводят (например, бактериальные векторы, имеющие бактериальную точку начала репликации, и эписомные векторы для млекопитающих). Другие векторы (например, неэписомные векторы для млекопитающих) могут интегрироваться в геном клетки-хозяина после введения в клетку-хозяина и, таким образом, реплицируются вместе с геномом хозяина. Кроме того, определенные векторы способны управлять экспрессией генов, с которыми они функционально связаны. Такие векторы называются в данном документе "рекомбинантными векторами экспрессии" (или просто "векторами экспрессии"). Как правило, векторы экспрессии, обладающие полезностью в технологиях рекомбинантных ДНК, часто имеют форму плазмид. В настоящем описании термины "плазмида" и "вектор" можно использовать взаимозаменяемо, поскольку плазмида является наиболее широко применяемой формой вектора. Однако, подразумевается, что настоящее изобретение включает и другие такие формы векторов экспрессии, такие как вирусные векторы (например, ретровирусы, дефектные по репликации, аденовирусы и аденоассоциированные вирусы), выполняющие эквивалентные функции.
Введение таких нуклеиновых кислот в клетку-хозяина можно выполнять с помощью методик, хорошо известных из уровня техники. Для эукариотических клеток подходящие методики могут включать в себя, например, трансфекцию с использованием фосфата кальция, DEAE-декстрановый метод, электропорацию, трансфекцию, опосредованную липосомами, и трансдукцию с использованием ретровирусов или других вирусов. Для бактериальных клеток подходящие методики могут включать в себя трансформацию с использованием хлорида кальция, электропорацию и трансфекцию с использованием бактериофага. После введения можно обуславливать или обеспечивать экспрессию нуклеиновой кислоты, например, путем культивирования клеток-хозяев в условиях, подходящих для экспрессии гена. В одном варианте осуществления нуклеиновая кислота по настоящему изобретению интегрируется в геном, например, хромосому, клетки-хозяина. Интеграции можно содействовать путем включения последовательностей, которые содействуют рекомбинации с геномом, в соответствии со стандартными методиками.
Хорошо известны системы клонирования и экспрессии полипептида в ряде различных клеток-хозяев. Подходящие клетки-хозяева включают в себя клетки бактерий, клетки млекопитающих, клетки растений, клетки насекомых, грибов, дрожжей и трансгенных растений и животных. Линии клеток млекопитающих, доступные в данной области техники для экспрессии гетерологичного полипептида, включают в себя клетки яичника китайского хомячка (CHO), клетки HeLa, клетки почки новорожденного хомяка, клетки меланомы мыши, клетки меланомы крысы, человеческие эмбриональные клетки почек, например, клетки HEK293, человеческие эмбриональные клетки сетчатки и многие другие. Экспрессия антител и фрагментов антител в прокариотических клетках, таких как клетки E. coli, надежно отработана в данной области техники. Для обзора см., например, Plückthun Bio/Technology 9: 545-551 (1991). Экспрессия в культивируемых эукариотических клетках также доступна специалистам в данной области, как рассматривается, например, в Andersen et al. (2002) Curr. Opin. Biotechnol. 13: 117-23.
Димеры scFv-Fc могут быть гликозилированными, естественным путем либо вследствие выбора хозяина для экспрессии, например, клеток CHO, HEK293 или NSO (ECACC 85110503), или они могут быть негликозилированными, например, в случае выработки в результате экспрессии в прокариотической клетке. Гликозилирование можно также целенаправленно изменить, например, посредством ингибирования фукозилирования, с целью повышения активности ADCC получаемого в результате димера scFv-Fc.
Способы применения антител или их антигенсвязывающих фрагментов
Димеры scFv-Fc можно применять в способе лечения или диагностики организма человека или животного, таком как способ лечения (который может включать профилактическое лечение) заболевания или нарушения у пациента-человека, который включает введение эффективного количества для лечения пациента. Состояния, подвергаемые лечению, включают в себя любые состояния, в которых играет роль TGFβ1, например, фиброзное заболевание, рак, иммуноопосредованное заболевание и заживление ран, например, диффузный системный склероз, заболевание, характеризующееся нарушением ремоделирования костной ткани, заболевание почек и/или их комбинации.
Было показано, что антитела, специфичные к человеческому TGFβ1, являются эффективными в животных моделях для лечения опосредованных TGFβ1 гломерулонефрита (Border et al. (1990) Nature 346: 371-374), рубцевания нервной ткани (Logan et al. (1994) Eur. J. Neurosci. 6: 355-363), рубцевания кожи (Shah et al. (1992) Lancet 339: 213-214; Shah et al. (1994) J. Cell Science 107: 1137-1157; Shah et al. (1995) J. Cell Science 108: 985-1002) и легочного фиброза (Giri et al. (1993) Thorax 48: 959-966). Кроме того, было показано, что антитела к TGFβ1, 2 и 3 являются эффективными в моделях фиброза легких, радиационно-индуцированного фиброза (патент США № 5616561), миелофиброза, ожогов, контрактуры Дюпюитрена, язв желудка и ревматоидного артрита (Wahl et al. (1993) Exp. Medicine 177: 225-230).
Димеры scFv-Fc применимы для лечения заболевания и состояния, непосредственно или опосредованно обусловленного активностью TGFβ1. Димеры scFv-Fc могут избирательно ингибировать активность человеческой изоформы TGFβ1 in vitro или in vivo. Виды активности изоформ TGFβ1 включают без ограничений передачу сигнала, опосредованную TGFβ, отложение внеклеточного матрикса (ECM), ингибирование пролиферации эпителиальных и эндотелиальных клеток, содействие пролиферации гладкомышечных клеток, индукцию экспрессии коллагена III типа, индукцию экспрессии TGF-β, фибронектина, VEGF и IL-11, связывание с пептидом, ассоциированным с латентностью, подавление иммунитета, индуцированное опухолью, содействие ангиогенезу, активацию миофибробластов, содействие метастазированию и ингибирование активности NK-клеток. Например, димеры scFv-Fc применимы для лечения фокально-сегментарного гломерулосклероза (FSGS), фиброза печени (HF), острого инфаркта миокарда (AMI), идиопатического легочного фиброза (IPF), склеродермии (SSc) и синдрома Марфана.
Димеры scFv-Fc применимы для лечения заболеваний и состояний, включающих без ограничений фиброзные заболевания (такие как гломерулонефрит, рубцевание нервной ткани, рубцевание кожи, легочный фиброз, фиброз легких, радиационно-индуцированный фиброз, фиброз печени, миелофиброз), ожоги, иммуноопосредованные заболевания, воспалительные заболевания (в том числе ревматоидный артрит), отторжение трансплантата, рак, контрактуру Дюпюитрена и язвы желудка. Они также применимы для лечения, предупреждения и снижения риска возникновения форм почечной недостаточности, в том числе без ограничений: диабетической нефропатии (при диабете I типа и II типа), радиационно-индуцированной нефропатии, обструктивной нефропатии, диффузного системного склероза, легочного склероза, отторжения трансплантата, наследственного заболевания почек (например, поликистозного заболевания почек, спонгиозной почки, подковообразной почки), гломерулонефрита, нефросклероза, нефрокальциноза, системной красной волчанки, синдрома Шегрена, болезни Бергера, системной или гломерулярной гипертензии, тубулоинтерстициальной нефропатии, почечного тубулярного ацидоза, туберкулеза почки и инфаркта почки. В частности, они применимы в комбинации с антагонистами ренин-ангиотензин-альдостероновой системы, в том числе без ограничений: ингибиторами ренина, ингибиторами ангиотензинпревращающего фермента (ACE), антагонистами рецепторов Ang II (также известными как "блокаторы рецепторов Ang II") и антагонистами альдостерона. Способы применения димеров scFv-Fc в комбинации с такими антагонистами указаны, например, в WO 2004/098637.
Димеры scFv-Fc также применимы для лечения заболеваний и состояний, ассоциированных с отложением ECM, в том числе системного склероза, послеоперационных спаек, келоидного и гипертрофического рубцевания, пролиферативной витреоретинопатии, хирургического дренирования глаукомы, повреждения роговицы, катаракты, болезни Пейрони, острого респираторного дистресс-синдрома взрослых, цирроза печени, рубцевания после инфаркта миокарда, рестеноза после ангиопластики, рубцевания после субарахноидального кровоизлияния, рассеянного склероза, фиброза после ламинэктомии, фиброза после восстановления сухожилий и других органов, рубцевания в результате удаления татуировок, билиарного цирроза (в том числе склерозирующего холангита), перикардита, плеврита, трахеостомии, проникающего повреждения центральной нервной системы, эозинофильного миалгического синдрома, рестеноза сосудов, веноокклюзионной болезни, панкреатита и псориатической артропатии.
Димеры scFv-Fc дополнительно являются применимыми для содействия реэпителизации при таких заболеваниях и состояниях, как венозные язвы, трофические язвы (пролежни), диабетические язвы, участки после трансплантации, донорские участки после трансплантации, ссадины и ожоги, заболевания бронхиального эпителия, такие как астма, ARDS, заболевания кишечного эпителия, такие как мукозит, ассоциированный с лечением цитотоксическими препаратами, язвы пищевода (рефлюксная болезнь), язвы желудка, повреждения тонкой кишки и толстой кишки (воспалительное заболевание кишечника).
Димеры scFv-Fc также можно применять для содействия пролиферации эндотелиальных клеток, например, при стабилизации атеросклеротических бляшек, содействии заживлению сосудистых анастомозов, или для ингибирования пролиферации гладкомышечных клеток, как, например, при заболевании артерий, рестенозе и астме.
Димеры scFv-Fc применимы для усиления иммунного ответа на инфекции, опосредованные макрофагами. Они также применимы для уменьшения подавления иммунитета, вызываемого, например, опухолями, СПИДом или гранулематозными заболеваниями. Димеры scFv-Fc применимы для лечения гиперпролиферативных заболеваний, таких как виды рака, в том числе без ограничений рак молочной железы, предстательной железы, яичника, желудка, почки, поджелудочной железы, ободочной и прямой кишки, кожи, легкого, шейки матки и мочевого пузыря, глиома, мезотелиома, а также различные виды лейкемии и саркомы, такие как саркома Капоши, и применимы для лечения или предупреждения рецидивирования или метастазирования таких опухолей. Димеры scFv-Fc по настоящему изобретению также применимы для ингибирования метастазирования, опосредованного циклоспорином.
В контексте противоопухолевой терапии "лечение" включает любое медицинское вмешательство, приводящее к замедлению роста опухоли или уменьшению метастазирования опухоли, а также частичной ремиссии рака с целью увеличения ожидаемой продолжительности жизни пациента.
Способы лечения включают введение димера scFv-Fc или фармацевтических композиций, содержащих димер scFv-Fc. Димеры scFv-Fc можно применять в производстве лекарственного препарата для введения. Например, способ получения лекарственного препарата или фармацевтической композиции включает составление димера scFv-Fc с фармацевтически приемлемым наполнителем. Композицию можно вводить отдельно или в комбинации с другими средствами лечения одновременно либо последовательно в зависимости от состояния, подлежащего лечению.
Введение выполняют предпочтительно в "терапевтически эффективном количестве", достаточном для того, чтобы оказать благоприятный эффект на пациента. Таким благоприятным эффектом может быть по меньшей мере облегчение по меньшей мере одного симптома конкретного заболевания или состояния. Фактическое вводимое количество, а также скорость и временная динамика введения будут зависеть от природы и тяжести заболевания или состояния, подвергаемого лечению. Назначение лечения, например, решения относительно дозы и т. д., можно определить на основании доклинических и клинических исследований, план которых находится в пределах компетенции специалиста в данной области.
Точная доза будет зависеть от ряда факторов, в том числе того, предназначается ли димер scFv-Fc для диагностики или для лечения, размера и расположения участка, подлежащего обработке, и природы любой выявляемой метки или другой молекулы, присоединенной к димеру scFv-Fc. Обычная доза димера scFv-Fc, например, может находиться в диапазоне от 100 мкг до 1 грамма для путей системного применения и от 1 мкг до 1 мг для путей местного применения. Разовую дозу для лечения взрослого пациента можно пропорционально скорректировать для детей и младенцев. Лечение можно повторять ежедневно, два раза в неделю, один раз в неделю, один раз в месяц или через другие промежутки по усмотрению врача. Лечение может быть периодическим, и период между введениями составляет приблизительно две недели или более, предпочтительно приблизительно три недели или более, более предпочтительно приблизительно четыре недели или более, или приблизительно один месяц.
Предполагается, что уровни доз, составляющие приблизительно 0,1, 0,3, 1, 3, 10 или 15 мг на кг веса тела пациента, являются полезными и безопасными. Например, у крыс и мышей эффективная доза в острой фазе составляла 0,5-5 мг/кг. Таким образом, в случае длительного введения доз людям можно вводить 0,3-10 мг/кг на основании ожидаемого периода полувыведения, составляющего 21 день. Дозы могут быть достаточными для обеспечения эффективности и при этом достаточно низкими для облегчения оптимального введения. Например, доза, составляющая менее 50 мг, облегчает подкожное введение. Внутривенное введение можно применять в качестве пути доставки в случае тяжелых заболеваний, где могут быть необходимыми высокие дозы и длительные промежутки между введениями доз. Подкожное введение может усиливать потенциальный иммунный ответ на препарат. При локальном введении в случае локализованного заболевания можно снизить количество вводимого препарата и повысить концентрацию в месте действия, что может улучшить безопасность.
Димеры scFv-Fc по настоящему изобретению можно вводить путем инъекции, например, подкожно, внутривенно, внутриполостным путем (например, после резекции опухоли), внутриочаговым путем, внутрибрюшинно или внутримышечно. Димеры scFv-Fc также можно доставлять путем ингаляции, или местно (например, внутриглазным, интраназальным, ректальным путем, в раны, на кожу), или перорально.
Димер scFv-Fc обычно будут вводить в форме фармацевтической композиции, которая может содержать по меньшей мере один компонент в дополнение к димеру scFv-Fc. Таким образом, фармацевтические композиции могут содержать фармацевтически приемлемый наполнитель, носитель, буфер, стабилизатор или другие материалы, хорошо известные специалистам в данной области. Такие материалы должны быть нетоксичными и не должны оказывать мешающее влияние на эффективность активного ингредиента. Такие материалы могут включать в себя, например, любые возможные растворители, диспергирующие среды, покрытия, антибактериальные и противогрибковые средства, изотонические средства и средства, замедляющие всасывание. Некоторыми примерами фармацевтически приемлемых носителей являются вода, солевой раствор, фосфатно-солевой буферный раствор, декстроза, глицерин, этанол и т. п., а также их комбинации. Во многих случаях будет предпочтительным включать в композицию изотонические средства, например, сахара, многоатомные спирты, такие как маннит, сорбит, или хлорид натрия. Дополнительными примерами фармацевтически приемлемых веществ являются смачивающие средства или вспомогательные вещества, такие как эмульгирующие средства, консерванты или буферы, которые повышают сохраняемость или эффективность.
Точная природа носителя или другого материала будет зависеть от пути введения. В случае внутривенной инъекции или инъекции в участок поражения активный ингредиент будет находиться в форме парентерально приемлемого водного раствора, который является апирогенным и характеризуется подходящими pK, изотоничностью и стабильностью. Специалисты в данной области вполне в состоянии получить подходящие растворы с применением, например, изотонических основ, таких как хлорид натрия для инъекций, раствор Рингера для инъекций и лактатный раствор Рингера для инъекций. Можно включать консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки.
Димер scFv-Fc можно составлять в жидких, мягких или твердых формах, таких как жидкие растворы (например, растворы для инъекций и инфузий), дисперсии или суспензии, порошки, липосомы и суппозитории. Предпочтительная форма зависит от предполагаемого способа введения, терапевтического применения, физико-химических свойств молекулы и пути доставки. Составы могут содержать наполнители или комбинации наполнителей, например: сахара, аминокислоты и поверхностно-активные вещества. Жидкие составы могут предусматривать широкий диапазон концентраций димеров scFv-Fc и pH. Твердые составы можно получать, например, путем лиофилизации, высушивания распылением или высушивания с использованием сверхкритической флюидной технологии.
Терапевтические композиции можно составлять в виде раствора, микроэмульсии, дисперсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства. Стерильные растворы для инъекций можно получить путем включения димера scFv-Fc в соответствующий растворитель с одним из ингредиентов, перечисленных выше, или их комбинацией с последующей стерилизацией фильтрованием. Как правило, дисперсии получают путем включения активного соединения в стерильную основу, которая содержит основную диспергирующую среду и другие ингредиенты из перечисленных выше. В случае стерильных порошков для получения стерильных растворов для инъекций предпочтительными способами получения являются вакуумная сушка и сублимационная сушка, в результате которых получают порошкообразный активный ингредиент вместе с любым дополнительным желательным ингредиентом из его предварительно стерилизованного фильтрованием раствора. Надлежащую текучесть раствора можно поддерживать, например, путем применения покрытия, такого как лецитин, путем сохранения размера частиц в дисперсии или путем применения поверхностно-активных веществ. Длительного всасывания композиций для инъекций можно достичь путем включения в композицию средства, которое замедляет всасывание, например, моностеаратных солей и желатина.
В определенных вариантах осуществления активное соединение можно получить с носителем, который будет защищать димер scFv-Fc от быстрого высвобождения, как, например, в составе с контролируемым высвобождением, в том числе в имплантатах, трансдермальных пластырях и микроинкапсулированных системах доставки. Можно применять биоразлагаемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, сложные полиортоэфиры и полимолочная кислота. Многие способы получения таких составов запатентованы или общеизвестны для специалистов в данной области.
Способ применения димера scFv-Fc может включать обуславливание или обеспечивание связывания с TGFβ. Такое связывание может иметь место in vivo, например, после введения димера scFv-Fc пациенту, или оно может иметь место in vitro, например, в ELISA, вестерн-блоттинге, иммуноцитохимических анализах, иммунопреципитационных анализах, аффинной хроматографии или клеточных анализах, или в терапевтических способах ex vivo, например, в способах, в которых клетки или жидкости организма приводят в контакт с димером scFv-Fc ex vivo и затем вводят пациенту.
Предлагается набор, содержащий димер scFv-Fc. Димер scFv-Fc можно пометить с целью обеспечения его реактивности в образце, подлежащем определению. Наборы можно использовать, например, в диагностическом анализе. Набор может содержать инструкции по применению компонентов. В набор могут быть включены вспомогательные материалы для содействия в осуществлении или обеспечения осуществления такого способа.
Реактивность димера scFv-Fc в образце можно определять с помощью любых соответствующих средств, например, радиоиммунологического анализа (RIA). Можно смешивать радиоактивно меченный антиген с немеченым антигеном (тестовым образцом) и обеспечивать его связывание с димером scFv-Fc. Связанный антиген физически отделяют от несвязанного антигена, и определяют количество радиоактивного антигена, связанного с димером scFv-Fc. Также можно применять анализ конкурентного связывания с нерадиоактивным антигеном, используя антиген или аналог, связанный с репортерной молекулой. Репортерная молекула может представлять собой флуорохром, фосфор или краситель. Подходящие флуорохромы включают в себя флуоресцеин, родамин, фикоэритрин и техасский красный. Подходящие хромогенные красители включают в себя диаминобензидин.
Другие репортеры включают в себя макромолекулярные коллоидные частицы или материал в виде частиц, как, например, латексные гранулы, которые являются окрашенными, магнитными или парамагнитными, и биологически или химически активные средства, которые могут непосредственно или опосредованно обуславливать визуальное наблюдение выявляемых сигналов, их выявление электронными средствами или регистрацию иным способом. Эти молекулы могут представлять собой ферменты, катализирующие реакции, в ходе которых, например, проявляются или изменяются цвета или которые обуславливают изменения электрических свойств. Они могут быть возбудимыми на молекулярном уровне, так что электронные переходы между энергетическими состояниями приводят к характерным спектральным поглощениям или испусканиям. Они могут включать в себя химические объекты, применяемые совместно с биосенсорами. Можно использовать системы выявления на основе биотина/авидина или биотина/стрептавидина и щелочной фосфатазы. Сигналы, генерируемые конъюгатами антитело-репортер, можно применять для получения измеримых количественно абсолютных или относительных данных о соответствующем связывании антитела в образцах.
В настоящем изобретении также предлагается применение димера scFv-Fc для измерения уровней антигенов в конкурентном анализе. Димер scFv-Fc можно связать с репортерной молекулой таким образом, что при связывании, например, происходит физическое или оптическое изменение. Репортерная молекула может непосредственно или опосредованно генерировать выявляемые и предпочтительно измеримые сигналы. Репортерные молекулы можно связать непосредственно или опосредованно, ковалентно, например, посредством пептидной связи, или нековалентно. Димер scFv-Fc и белковый репортер можно связать пептидной связью и экспрессировать рекомбинантным путем в виде белка слияния.
Дополнительные аспекты и варианты осуществления настоящего изобретения будут очевидны специалистам в данной области в свете настоящего раскрытия, включающего следующее пояснение на экспериментальных примерах.
Примеры
Пример 1. Аффинность и активность scFv и антитела IgG4
CAT192(IgG4) (метелимумаб) представляет собой человеческое моноклональное антитело IgG4, которое избирательно нейтрализует TGF-β1. TGFβ1 (20-600 RU) иммобилизовали на чипе CM5 на Biacore с помощью химического вещества на основе NHS/EDC. На поверхность вводили различные количества CAT192(IgG4) для отслеживания связывания с TGFβ1, определяемого по методу поверхностного плазмонного резонанса. Данные анализировали в модели связывания 1:1 с целью определения констант связывания. Было обнаружено, что CAT192(IgG4) связывается с TGFβ1 с относительно низкой аффинностью, определенной по методу поверхностного плазмонного резонанса, по сравнению со связыванием исходного scFv CAT191, как показано на фиг. 2. CAT192 (IgG4) также демонстрировал относительно низкую эффективность (IC50=~ 10 нМ) в анализе активности с использованием клеток A549, в котором измеряли ингибирование выработки IL-11, стимулируемой TGFβ1. Иллюстративные результаты анализа с использованием клеток A549 показаны на фиг. 3. Анализ с использованием клеток A549 выполняли в соответствии с процедурой, раскрытой в Rapoza et al. (2006) "Development of an in vitro potency assay for therapeutic TGFβ antagonists: the A549 cell bioassay," J. Immunol. Methods 316: 18-26. Хотя кажущаяся константа диссоциации, составляющая ~ 10 нМ, демонстрировала специфичное связывание с TGFβ1, более высокая относительная активность была бы благоприятной для терапевтических путей применения CAT192 (IgG4).
Пример 2. Модифицированные антитела IgG1
Аффинность CAT192 (IgG4) можно незначительно усилить путем применения определенных денатурирующих условий, что свидетельствует о том, что сворачивание антитела могло вызывать утрату аффинности в ходе превращения scFv в IgG4. Было высказано предположение, что сворачивание IgG4 является уникальным (Aalberse and Schuurman "IgG4 breaking the rules", Immunology 105:9-19). Обмен Fab-фрагментами в IgG4 и взаимодействие Fab с CH2-доменом Fc, возможно, могут объяснять эту утрату аффинности CAT192 (IgG4). Таким образом, CAT192 подвергали перестройке с получением варианта IgG1 путем замещения Fc IgG4 (CH1-, CH2- и CH3-доменов) консенсусной последовательностью IgG1. ДНК, кодирующую CAT192 (IgG1), синтезировали из GeneArt и субклонировали в вектор экспрессии pCEP4(-E+I)Dest.
CAT192 (IgG1) получали в результате трансфекции HEK293 и очищали с помощью колонки с белком A. Перестройка CAT192 из IgG4 в IgG1, однако, не повышала его аффинность. Fab-фрагменты, образуемые из IgG1 и IgG4, также не повышали его аффинности. Было сделано заключение, что высокая аффинность CAT191(scFv) (SEQ ID NO: 12) утрачивалась в ходе превращения в формат антитела полной длины, будь то IgG1 или IgG4. Это было неожиданным, поскольку из scFv-компонентов, полученных из библиотеки, часто конструировали IgG в формате полной длины для терапевтических исследований.
Пример 3. Различные конструкции димеров
Путем применения поверхностного плазмонного резонанса было обнаружено, что CAT191 (scFv) (SEQ ID NO: 12) связывается с TGFβ1 с высокой аффинностью, однако CAT191 (scFv) не обладал авидностью, необходимой для эффективной нейтрализации TGFβ1. Соответственно, тестировали различные другие форматы с применением scFv-компонента в качестве основного структурного элемента. Общие форматы фрагментов антител, в том числе тестируемые форматы, представлены на фиг. 1.
Тестируемые форматы включали диатело, димер пептида (например, димер пептида с лейциновыми застежками) и димер scFv-Fc. В диателе scFv CAT191 линкер типа (Gly4Ser)3 был замещен коротким линкером из 5 аминокислот (GSSGG) (SEQ ID NO: 19) с целью создания нековалентного двухвалентного связывающего средства (димерного диатела). Каждый мономер имел последовательность, указанную под SEQ ID NO: 14. Каждый мономер димера пептида с лейциновыми застежками имел последовательность, указанную под SEQ ID NO: 16. Наконец, каждый мономер димера scFv-Fc имел последовательность, указанную под SEQ ID NO: 9. Диатело и димер пептида экспрессировали в E. coli, а scFv-Fc экспрессировали в клетках HEK293.
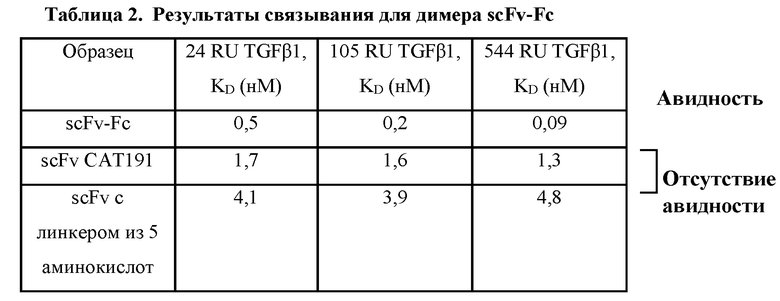
Димер пептида с лейциновыми застежками было трудно экспрессировать, а частично очищенный димер демонстрировал лишь среднюю аффинность, измеренную по методу поверхностного плазмонного резонанса. Диатело (scFv с линкером из 5 аминокислот) демонстрировало лишь среднюю аффинность, но не авидность. В отличие от этого, было обнаружено, что димер scFv-Fc, полученный в результате транзиентной трансфекции HEK293, специфично связывается с TGFβ1 с высокой аффинностью и авидностью. Результаты связывания, выраженные в виде кажущихся констант диссоциации, полученных по методу поверхностного плазмонного резонанса, обобщены ниже в таблице 2.
Активность нейтрализации TGFβ1 в различных форматах также сравнивали в биологическом анализе с использованием клеток A549. На фиг. 3 показаны результаты биологического анализа с использованием клеток A549 для диатела ("scFv-диатело с линкером из 5 аминокислот"), CAT191(scFv) ("scFv"), димера scFv-Fc ("CAT191 (scFv-Fc)") и CAT192(IgG4) ("CAT192"). Как видно на фиг. 3, димер scFv-Fc демонстрировал кажущуюся константу диссоциации в данном анализе, более чем на четыре порядка величины более низкую, чем у CAT192 (~ 10-3 нМ по сравнению с ~ 101 нМ).
Пример 4. Клон scFv-Fc
CAT191 (scFv-Fc) клонировали и получали в большем масштабе в клетках CHO. Последовательность, кодирующую scFv-Fc CAT191, амплифицировали путем ПЦР из вектора экспрессии на основе pCEP4 с помощью набора геноспецифичных прямых и обратных праймеров. В качестве части ПЦР-амплификации в последовательность, кодирующую scFv-Fc CAT191, вводили следующие изменения: 1) добавление сайтов для эндонуклеазы на 5'- и 3'-концах, 2) добавление консенсусной последовательности Козак непосредственно выше стартового кодона, 3) изменение стоп-кодона "TAG" на "TAA" и 4) мутацию 4 тимидиновых нуклеотидов выше стоп-кодона по типу замены на гуанозин с удалением тем самым эндогенного донорного сайта сплайсинга. Мутация донорного сайта сплайсинга не приводила к изменению аминокислот.
Амплифицированную путем ПЦР последовательность, кодирующую CAT191, субклонировали в челночный вектор с целью облегчения проверки последовательности и молекулярного клонирования. После проверки последовательности последовательность, кодирующую CAT191, клонировали в векторы экспрессии pGZ600 и pGZ620 от Genzyme. В обоих векторах применяли промотор гена β-актина хомяка для управления экспрессией трансгена CAT191. Они также содержали селектируемый маркер DHFR, который управлялся отдельным промотором (SV40), с целью обеспечения отбора в клетках CHO. Линию клеток-хозяев CHO-8D6 трансфицировали плазмидой экспрессии pGZ600-CAT191 либо pGZ620-CAT191. После короткого периода восстановления трансфицированные клетки помещали в среду для роста с дефицитом нуклеотидов для отбора с целью получения пулов стабильных трансфектантов. После извлечения в пулов в результате отбора проводили второй цикл отбора в присутствии 20 нМ метотрексата. Пулы CHO, отобранные таким образом, увеличивали в масштабе, и кондиционированные среды применяли для очистки с помощью колонки с белком A.
Характеристики белка, вырабатываемого клетками CHO, получали с помощью SDS-PAGE, связывания на Biacore, SEC-HPLC и анализа активности с использованием клеток A549. Результаты подтверждали, что димер scFv-Fc характеризовался более высокой аффинностью и активностью и специфично нейтрализовал TGFβ1. Результат сравнения активности был благоприятным для панспецифичного антитела GC1008 (фиг. 6 и фиг. 7).
Пример 5. Период полувыведения из кровотока
Период полувыведения CAT191 (scFv-Fc) из кровотока тестировали в мышиной модели с применением плана исследования, представленного в таблице 3.
Таблица 3. Период полувыведения димера scFv-Fc из кровотока
Кровь забирали из ретроорбитального сплетения в определенные моменты времени после внутрибрюшинного (IP) или внутривенного (IV) введения. Примерно 60 мкл цельной крови собирали в гематокритные капилляры и обрабатывали для получения сыворотки крови. До проведения анализа все образцы хранили при -80°C. Концентрацию CAT191(scFv-Fc) определяли с помощью ELISA. Результаты этого фармакокинетического исследования представлены на фиг. 4 и фиг. 5. Результаты свидетельствовали о том, что период полувыведения из кровотока составлял 1,5-2,0 дня, являясь намного более длительным, чем для обычной молекулы scFv, для которой он составляет несколько часов.
Пример 6. Стабильность димеров scFv-Fc
Стабильность CAT191(scFv-Fc), хранящегося при -80°C, отслеживали в течение года с помощью SEC-HPLC, связывания с TGFβ1 на Biacore и анализа активности с использованием A549. Во время периода тестирования изменение агрегации, аффинности или активности не наблюдалось. Материал, хранящийся при температуре 4°C, проявлял незначительное, но устойчивое увеличение агрегации в течение 1 года. Уникальная комбинация меньшего размера, высокой избирательности, активности в отношении TGFβ1 и длительного периода полувыведения in vivo сделала CAT191(scFv-Fc) превосходным кандидатом для терапевтических путей применения.
Все документы, цитируемые во всем данном раскрытии, в том числе без ограничения научные публикации, патенты и публикации заявок на патенты, настоящим включены в данное раскрытие посредством ссылки так, как если бы в данном документе было воспроизведено полное их содержание.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1: клон SL15 VH-домена человеческого IgG1 (SQN4 US6492497)
EVQLVESGGGVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKELEWVAVISYDGSIKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARTGEYSGYDTDPQYSWGQGTTVTVSS
SEQ ID NO: 2: клон JT182 VH-домена человеческого IgG1 (SQN10 US6492497)
QVQLVESGGGVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKELEWVAVISYDGSIKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARTGEYSGYDTPASPDWGQGTTVTVSS
SEQ ID NO: 3: синтетический линкер
SGGGSGGGGSGGGGS
SEQ ID NO: 4: синтетический линкер
GGGGSGGGGSGGGGS
SEQ ID NO: 5: клон SL15A Vκ-домена человеческого IgG1: (SQN6 US6492497)
EIVLTQSPSSLSASVGDRVTITCRASQGIGDDLGWYQQKPGKAPILLIYGTSTLQSGVPSRFSGSGSGTDFTLTINSLQPEDFATYYCLQDSNYPLTFGGGTRLEIK
SEQ ID NO: 6: клон SL15S Vκ-домена человеческого IgG1: (SQN8 US6492497)
EIVLTQSPSSLSASVGDRVTITCRSSQGIGDDLGWYQQKPGKAPILLIYGTSTLQSGVPSRFSGSGSGTDFTLTINSLQPEDFATYYCLQDSNYPLTFGGGTRLEIK
SEQ ID NO: 7: шарнирная область человеческого IgG1
PKSCDKTHTCPPCPAPELLGGP
SEQ ID NO: 8: Fc-область человеческого IgG1
SVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
SEQ ID NO: 9: CAT191(scFv-Fc)
EVQLVESGGGVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKELEWVAVISYDGSIKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARTGEYSGYDTDPQYSWGQGTTVTVSSSGGGSGGGGSGGGGSEIVLTQSPSSLSASVGDRVTITCRSSQGIGDDLGWYQQKPGKAPILLIYGTSTLQSGVPSRFSGSGSGTDFTLTINSLQPEDFATYYCLQDSNYPLTFGGGTRLEIKGGSGPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
SEQ ID NO: 10: легкая цепь CAT192 (IgG4)
EWLTQSPSSLSASVGDRVTITCRASQGIGDDLGWYQQKPGKAPILLIYGTSTLQSGVPSRFSGSGSGTDFTLTINSLQPEDFATYYCLQDSNYPLTFGGGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO: 11: тяжелая цепь CAT192 (IgG4)
EVQLVESGGGVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKELEWVAVISYDGSIKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARTGEYSGYDTDPQYSWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK
SEQ ID NO: 12: CAT191(scFv)
EVQLVESGGGVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKELEWVAVISYDGSIKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARTGEYSGYDTDPQYSWGQGTTVTVSSSGGGSGGGGSGGGGSEIVLTQSPSSLSASVGDRVTITCRSSQGIGDDLGWYQQKPGKAPILLIYGTSTLQSGVPSRFSGSGSGTDFTLTINSLQPEDFATYYCLQDSNYPLTFGGGTRLEIK
SEQ ID NO: 13: нуклеиновая кислота, кодирующая диатело с линкером из 5 аминокислот
atgaccatgattacgccaagctttggagccttttttttggagattttcaacgtgaaaaaattattattcgcaattcctttagttgttcctttctatgcggcccagccggccatggccgaggtgcagctggtggagtctgggggaggcgtggtccagcctgggaggtccctgagactctcctgtgcagcctctggattcaccttcagtagctatggcatgcactgggtccgccaggctccaggcaaggagctggagtgggtggcagttatatcatatgatggaagtattaaatactatgcagactccgtgaagggccgattcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagctgaggacacggctgtgtattactgtgcgcgaactggtgaatatagtggctacgatacggacccccagtactcctgggggcaagggaccacggtcaccgtctcctcaggttcctctggcggtgaaattgtgctgactcagtctccatcctccctgtctgcatctgtaggagacagagtcaccatcacttgccggtcaagtcagggcattggagatgatttgggctggtatcagcagaagccagggaaagcccctatcctcctgatctatggtacatccactttacaaagtggggtcccgtcaaggttcagcggcagtggatctggcacagatttcactctcaccatcaacagcctgcagcctgaagattttgcaacttattactgtctacaagattccaattacccgctcactttcggcggagggacacgactggagattaaacgtgcggccgcacatcatcatcaccatcacggggccgcagaacaaaaactcatctcagaagaggatctgaatggggccgcatagtagctcgagatcaaacgggctagccagccagaactcgccccggaagaccccgaggatgtcgagcaccaccaccaccac
SEQ ID NO: 14: диатело с линкером из 5 аминокислот
EVQLVESGGGVVQPGRSLRLSCAASGFTESSYGMHWVRQAPGKELEWVAVISYDGSIKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARTGEYSGYDTDPQYSWGQGTTVTVSSGSSGGEIVLTQSPSSLSASVGDRVTITCRSSQGIGDDLGWYQQKPGKAPILLIYGTSTLQSGVPSRFSGSGSGTDFTLTINSLQPEDFATYYCLQDSNYPLTFGGGTRLEIKRAAAHHHHHHGAAEQKLISEEDLNGAA
SEQ ID NO: 15: нуклеиновая кислота, кодирующая димер пептида с лейциновыми застежками
gaggtgcagctggtggagtctgggggaggcgtggtccagcctgggaggtccctgagactctcctgtgcagcctctggattcaccttcagtagctatggcatgcactgggtccgccaggctccaggcaaggagctggagtgggtggcagttatatcatatgatggaagtattaaatactatgcagactccgtgaagggccgattcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagctgaggacacggctgtgtattactgtgcgcgaactggtgaatatagtggctacgatacggacccccagtactcctgggggcaagggaccacggtcaccgtctcctcaagtggaggcggttcaggcggaggtggcagcggcggtggcggatcggaaattgtgctgactcagtctccatcctccctgtctgcatctgtaggagacagagtcaccatcacttgccggtcaagtcagggcattggagatgatttgggctggtatcagcagaagccagggaaagcccctatcctcctgatctatggtacatccactttacaaagtggggtcccgtcaaggttcagcggcagtggatctggcacagatttcactctcaccatcaacagcctgcagcctgaagattttgcaacttattactgtctacaagattccaattacccgctcactttcggcggagggacacgactggagattaaacgtgcggccgcacatcatcatcaccatcacggggccgcagaacaaaaactcatctcagaagaggatctgaatggggccgcacccaagcccagtacccccccaggttcttcaggcgaactggaagaactgctgaaacatctgaaagaactgctgaaaggcccgcgtaaaggcgaactggaagaactgctgaaacatctgaaagaactgctgaaaggcggtgcgccgggcggtcatcatcatcaccatcat
SEQ ID NO: 16: димер пептида с лейциновыми застежками
EVQLVESGGGVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKELEWVAVISYDGSIKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARTGEYSGYDTDPQYSWGQGTTVTVSSSGGGSGGGGSGGGGSEIVLTQSPSSLSASVGDRVTITCRSSQGIGDDLGWYQQKPGKAPILLIYGTSTLQSGVPSRFSGSGSGTDFTLTINSLQPEDFATYYCLQDSNYPLTFGGGTRLEIKRAAAHHHHHHGAAEQKLISEEDLNGAAPKPSTPPGSSGELEELLKHLKELLKGPRKGELEELLKHLKELLKGGAPGGHHHHHH
SEQ ID NO: 17: нуклеиновая кислота, кодирующая CAT191(scFv-Fc)
gaggtgcagctggtggagtctgggggaggcgtggtccagcctgggaggtccctgagactctcctgtgcagcctctggattcaccttcagtagctatggcatgcactgggtccgccaggctccaggcaaggagctggagtgggtggcagttatatcatatgatggaagtattaaatactatgcagactccgtgaagggccgattcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagctgaggacacggctgtgtattactgtgcgcgaactggtgaatatagtggctacgatacggacccccagtactcctgggggcaagggaccacggtcaccgtctcctcaagtggaggcggttcaggcggaggtggcagcggcggtggcggatcggaaattgtgctgactcagtctccatcctccctgtctgcatctgtaggagacagagtcaccatcacttgccggtcaagtcagggcattggagatgatttgggctggtatcagcagaagccagggaaagcccctatcctcctgatctatggtacatccactttacaaagtggggtcccgtcaaggttcagcggcagtggatctggcacagatttcactctcaccatcaacagcctgcagcctgaagattttgcaacttattactgtctacaagattccaattacccgctcactttcggcggagggacacgactggagattaaaggtggcagcggacctaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgagctgaccaagaaccaggtcagcctgacgtgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcagatggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatagtag
SEQ ID NO: 18: человеческий TGFβ1
ALDTNYCFSSTEKNCCVRQLYIDFRKDLGWKWIHEPKGYHANFCLGPCPYIWSLDTQYSKVLALYNQHNPGASAAPCCVPQALEPLPIVYYVGRKPKVEQLSNMIVRSCKCS
SEQ ID No. 19
GSSGG
SEQ ID No. 20
GGSG
SEQ ID No. 21
CPPCP
SEQ ID No. 22
SYGMH
SEQ ID No. 23
VISYDGSIKYYADSVKG
SEQ ID No. 24
TGEYSGYDTSGVEL
SEQ ID No. 25
TGEYSGYDTDPQYS
SEQ ID No. 26
TGFYSGYDTPASPD
SEQ ID No. 27
RASQGIGDDLG
SEQ ID No. 28
GTSTLQS
SEQ ID No. 29
LQDSNYPLT
SEQ ID No. 30
TGX1YSGYDTX2X3X4X5X6
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Pan, Clark
Qiu, Huawei
Bird, Julie
<120> ДИМЕРЫ SCFV-FC, КОТОРЫЕ СВЯЗЫВАЮТСЯ С ТРАНСФОРМИРУЮЩИМ ФАКТОРОМ РОСТА β1 С ВЫСОКОЙ АФФИННОСТЬЮ, АВИДНОСТЬЮ И СПЕЦИФИЧНОСТЬЮ
<130> 578962
<150> 62/128133
<151> 2015-03-04
<160> 30
<170> PatentIn версия 3.5
<210> 1
<211> 123
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Glu Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Ile Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Glu Tyr Ser Gly Tyr Asp Thr Asp Pro Gln Tyr Ser
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 2
<211> 123
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Glu Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Ile Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Glu Tyr Ser Gly Tyr Asp Thr Pro Ala Ser Pro Asp
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 3
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 3
Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 4
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> синтетический
<400> 4
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 5
<211> 107
<212> БЕЛОК
<213> Homo sapiens
<400> 5
Glu Ile Val Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Gly Asp Asp
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Ile Leu Leu Ile
35 40 45
Tyr Gly Thr Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Ser Asn Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 6
<211> 107
<212> БЕЛОК
<213> Homo sapiens
<400> 6
Glu Ile Val Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Gly Ile Gly Asp Asp
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Ile Leu Leu Ile
35 40 45
Tyr Gly Thr Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Ser Asn Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 7
<211> 22
<212> БЕЛОК
<213> Homo sapiens
<400> 7
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
1 5 10 15
Glu Leu Leu Gly Gly Pro
20
<210> 8
<211> 209
<212> БЕЛОК
<213> Homo sapiens
<400> 8
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
1 5 10 15
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
20 25 30
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
35 40 45
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
50 55 60
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
65 70 75 80
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
85 90 95
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
100 105 110
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
115 120 125
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
130 135 140
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
145 150 155 160
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
165 170 175
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
180 185 190
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
195 200 205
Lys
<210> 9
<211> 480
<212> БЕЛОК
<213> Homo sapiens
<400> 9
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Glu Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Ile Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Glu Tyr Ser Gly Tyr Asp Thr Asp Pro Gln Tyr Ser
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ser Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln
130 135 140
Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr
145 150 155 160
Cys Arg Ser Ser Gln Gly Ile Gly Asp Asp Leu Gly Trp Tyr Gln Gln
165 170 175
Lys Pro Gly Lys Ala Pro Ile Leu Leu Ile Tyr Gly Thr Ser Thr Leu
180 185 190
Gln Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
195 200 205
Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr
210 215 220
Tyr Cys Leu Gln Asp Ser Asn Tyr Pro Leu Thr Phe Gly Gly Gly Thr
225 230 235 240
Arg Leu Glu Ile Lys Gly Gly Ser Gly Pro Lys Ser Cys Asp Lys Thr
245 250 255
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
260 265 270
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
275 280 285
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
290 295 300
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
305 310 315 320
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
325 330 335
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
340 345 350
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
355 360 365
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
370 375 380
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
385 390 395 400
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
405 410 415
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
420 425 430
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
435 440 445
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
450 455 460
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
465 470 475 480
<210> 10
<211> 213
<212> БЕЛОК
<213> Homo sapiens
<400> 10
Glu Trp Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp
1 5 10 15
Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Gly Asp Asp Leu
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Ile Leu Leu Ile Tyr
35 40 45
Gly Thr Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Ser Asn Tyr Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 11
<211> 450
<212> БЕЛОК
<213> Homo sapiens
<400> 11
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Glu Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Ile Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Glu Tyr Ser Gly Tyr Asp Thr Asp Pro Gln Tyr Ser
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
195 200 205
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
210 215 220
Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
260 265 270
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
435 440 445
Gly Lys
450
<210> 12
<211> 245
<212> БЕЛОК
<213> Homo sapiens
<400> 12
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Glu Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Ile Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Glu Tyr Ser Gly Tyr Asp Thr Asp Pro Gln Tyr Ser
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ser Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln
130 135 140
Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr
145 150 155 160
Cys Arg Ser Ser Gln Gly Ile Gly Asp Asp Leu Gly Trp Tyr Gln Gln
165 170 175
Lys Pro Gly Lys Ala Pro Ile Leu Leu Ile Tyr Gly Thr Ser Thr Leu
180 185 190
Gln Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
195 200 205
Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr
210 215 220
Tyr Cys Leu Gln Asp Ser Asn Tyr Pro Leu Thr Phe Gly Gly Gly Thr
225 230 235 240
Arg Leu Glu Ile Lys
245
<210> 13
<211> 984
<212> ДНК
<213> Homo sapiens
<400> 13
atgaccatga ttacgccaag ctttggagcc ttttttttgg agattttcaa cgtgaaaaaa 60
ttattattcg caattccttt agttgttcct ttctatgcgg cccagccggc catggccgag 120
gtgcagctgg tggagtctgg gggaggcgtg gtccagcctg ggaggtccct gagactctcc 180
tgtgcagcct ctggattcac cttcagtagc tatggcatgc actgggtccg ccaggctcca 240
ggcaaggagc tggagtgggt ggcagttata tcatatgatg gaagtattaa atactatgca 300
gactccgtga agggccgatt caccatctcc agagacaatt ccaagaacac gctgtatctg 360
caaatgaaca gcctgagagc tgaggacacg gctgtgtatt actgtgcgcg aactggtgaa 420
tatagtggct acgatacgga cccccagtac tcctgggggc aagggaccac ggtcaccgtc 480
tcctcaggtt cctctggcgg tgaaattgtg ctgactcagt ctccatcctc cctgtctgca 540
tctgtaggag acagagtcac catcacttgc cggtcaagtc agggcattgg agatgatttg 600
ggctggtatc agcagaagcc agggaaagcc cctatcctcc tgatctatgg tacatccact 660
ttacaaagtg gggtcccgtc aaggttcagc ggcagtggat ctggcacaga tttcactctc 720
accatcaaca gcctgcagcc tgaagatttt gcaacttatt actgtctaca agattccaat 780
tacccgctca ctttcggcgg agggacacga ctggagatta aacgtgcggc cgcacatcat 840
catcaccatc acggggccgc agaacaaaaa ctcatctcag aagaggatct gaatggggcc 900
gcatagtagc tcgagatcaa acgggctagc cagccagaac tcgccccgga agaccccgag 960
gatgtcgagc accaccacca ccac 984
<210> 14
<211> 262
<212> БЕЛОК
<213> Homo sapiens
<400> 14
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Glu Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Glu Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Ile Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Glu Tyr Ser Gly Tyr Asp Thr Asp Pro Gln Tyr Ser
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Ser Ser Gly Gly
115 120 125
Glu Ile Val Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
130 135 140
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Gly Ile Gly Asp Asp
145 150 155 160
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Ile Leu Leu Ile
165 170 175
Tyr Gly Thr Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
180 185 190
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
195 200 205
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Ser Asn Tyr Pro Leu
210 215 220
Thr Phe Gly Gly Gly Thr Arg Leu Glu Ile Lys Arg Ala Ala Ala His
225 230 235 240
His His His His His Gly Ala Ala Glu Gln Lys Leu Ile Ser Glu Glu
245 250 255
Asp Leu Asn Gly Ala Ala
260
<210> 15
<211> 984
<212> ДНК
<213> Homo sapiens
<400> 15
gaggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatggca tgcactgggt ccgccaggct 120
ccaggcaagg agctggagtg ggtggcagtt atatcatatg atggaagtat taaatactat 180
gcagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctgtgt attactgtgc gcgaactggt 300
gaatatagtg gctacgatac ggacccccag tactcctggg ggcaagggac cacggtcacc 360
gtctcctcaa gtggaggcgg ttcaggcgga ggtggcagcg gcggtggcgg atcggaaatt 420
gtgctgactc agtctccatc ctccctgtct gcatctgtag gagacagagt caccatcact 480
tgccggtcaa gtcagggcat tggagatgat ttgggctggt atcagcagaa gccagggaaa 540
gcccctatcc tcctgatcta tggtacatcc actttacaaa gtggggtccc gtcaaggttc 600
agcggcagtg gatctggcac agatttcact ctcaccatca acagcctgca gcctgaagat 660
tttgcaactt attactgtct acaagattcc aattacccgc tcactttcgg cggagggaca 720
cgactggaga ttaaacgtgc ggccgcacat catcatcacc atcacggggc cgcagaacaa 780
aaactcatct cagaagagga tctgaatggg gccgcaccca agcccagtac ccccccaggt 840
tcttcaggcg aactggaaga actgctgaaa catctgaaag aactgctgaa aggcccgcgt 900
aaaggcgaac tggaagaact gctgaaacat ctgaaagaac tgctgaaagg cggtgcgccg 960
ggcggtcatc atcatcacca tcat 984
<210> 16
<211> 328
<212> БЕЛОК
<213> Homo sapiens
<400> 16
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Glu Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Ile Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Glu Tyr Ser Gly Tyr Asp Thr Asp Pro Gln Tyr Ser
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ser Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln
130 135 140
Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr
145 150 155 160
Cys Arg Ser Ser Gln Gly Ile Gly Asp Asp Leu Gly Trp Tyr Gln Gln
165 170 175
Lys Pro Gly Lys Ala Pro Ile Leu Leu Ile Tyr Gly Thr Ser Thr Leu
180 185 190
Gln Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
195 200 205
Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr
210 215 220
Tyr Cys Leu Gln Asp Ser Asn Tyr Pro Leu Thr Phe Gly Gly Gly Thr
225 230 235 240
Arg Leu Glu Ile Lys Arg Ala Ala Ala His His His His His His Gly
245 250 255
Ala Ala Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Asn Gly Ala Ala
260 265 270
Pro Lys Pro Ser Thr Pro Pro Gly Ser Ser Gly Glu Leu Glu Glu Leu
275 280 285
Leu Lys His Leu Lys Glu Leu Leu Lys Gly Pro Arg Lys Gly Glu Leu
290 295 300
Glu Glu Leu Leu Lys His Leu Lys Glu Leu Leu Lys Gly Gly Ala Pro
305 310 315 320
Gly Gly His His His His His His
325
<210> 17
<211> 1446
<212> ДНК
<213> Homo sapiens
<400> 17
gaggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatggca tgcactgggt ccgccaggct 120
ccaggcaagg agctggagtg ggtggcagtt atatcatatg atggaagtat taaatactat 180
gcagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctgtgt attactgtgc gcgaactggt 300
gaatatagtg gctacgatac ggacccccag tactcctggg ggcaagggac cacggtcacc 360
gtctcctcaa gtggaggcgg ttcaggcgga ggtggcagcg gcggtggcgg atcggaaatt 420
gtgctgactc agtctccatc ctccctgtct gcatctgtag gagacagagt caccatcact 480
tgccggtcaa gtcagggcat tggagatgat ttgggctggt atcagcagaa gccagggaaa 540
gcccctatcc tcctgatcta tggtacatcc actttacaaa gtggggtccc gtcaaggttc 600
agcggcagtg gatctggcac agatttcact ctcaccatca acagcctgca gcctgaagat 660
tttgcaactt attactgtct acaagattcc aattacccgc tcactttcgg cggagggaca 720
cgactggaga ttaaaggtgg cagcggacct aaatcttgtg acaaaactca cacatgccca 780
ccgtgcccag cacctgaact cctgggggga ccgtcagtct tcctcttccc cccaaaaccc 840
aaggacaccc tcatgatctc ccggacccct gaggtcacat gcgtggtggt ggacgtgagc 900
cacgaagacc ctgaggtcaa gttcaactgg tacgtggacg gcgtggaggt gcataatgcc 960
aagacaaagc cgcgggagga gcagtacaac agcacgtacc gtgtggtcag cgtcctcacc 1020
gtcctgcacc aggactggct gaatggcaag gagtacaagt gcaaggtctc caacaaagcc 1080
ctcccagccc ccatcgagaa aaccatctcc aaagccaaag ggcagccccg agaaccacag 1140
gtgtacaccc tgcccccatc ccgggatgag ctgaccaaga accaggtcag cctgacgtgc 1200
ctggtcaaag gcttctatcc cagcgacatc gccgtggagt gggagagcaa tgggcagccg 1260
gagaacaact acaagaccac gcctcccgtg ctggactccg acggctcctt cttcctctac 1320
agcaagctca ccgtggacaa gagcagatgg cagcagggga acgtcttctc atgctccgtg 1380
atgcatgagg ctctgcacaa ccactacacg cagaagagcc tctccctgtc tccgggtaaa 1440
tagtag 1446
<210> 18
<211> 112
<212> БЕЛОК
<213> Homo sapiens
<400> 18
Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser Thr Glu Lys Asn Cys Cys
1 5 10 15
Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys Asp Leu Gly Trp Lys Trp
20 25 30
Ile His Glu Pro Lys Gly Tyr His Ala Asn Phe Cys Leu Gly Pro Cys
35 40 45
Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr Ser Lys Val Leu Ala Leu
50 55 60
Tyr Asn Gln His Asn Pro Gly Ala Ser Ala Ala Pro Cys Cys Val Pro
65 70 75 80
Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr Tyr Val Gly Arg Lys Pro
85 90 95
Lys Val Glu Gln Leu Ser Asn Met Ile Val Arg Ser Cys Lys Cys Ser
100 105 110
<210> 19
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 19
Gly Ser Ser Gly Gly
1 5
<210> 20
<211> 4
<212> БЕЛОК
<213> Homo sapiens
<400> 20
Gly Gly Ser Gly
1
<210> 21
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 21
Cys Pro Pro Cys Pro
1 5
<210> 22
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 22
Ser Tyr Gly Met His
1 5
<210> 23
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 23
Val Ile Ser Tyr Asp Gly Ser Ile Lys Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 24
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 24
Thr Gly Glu Tyr Ser Gly Tyr Asp Thr Ser Gly Val Glu Leu
1 5 10
<210> 25
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 25
Thr Gly Glu Tyr Ser Gly Tyr Asp Thr Asp Pro Gln Tyr Ser
1 5 10
<210> 26
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 26
Thr Gly Phe Tyr Ser Gly Tyr Asp Thr Pro Ala Ser Pro Asp
1 5 10
<210> 27
<211> 11
<212> БЕЛОК
<213> Homo sapiens
<400> 27
Arg Ala Ser Gln Gly Ile Gly Asp Asp Leu Gly
1 5 10
<210> 28
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 28
Gly Thr Ser Thr Leu Gln Ser
1 5
<210> 29
<211> 9
<212> БЕЛОК
<213> Homo sapiens
<400> 29
Leu Gln Asp Ser Asn Tyr Pro Leu Thr
1 5
<210> 30
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> другой_признак
<222> (3)..(3)
<223> Xaa может представлять собой любую втречающуюся в природе аминокислоту
<220>
<221> другой_признак
<222> (10)..(14)
<223> Xaa может представлять собой любую втречающуюся в природе аминокислоту
<400> 30
Thr Gly Xaa Tyr Ser Gly Tyr Asp Thr Xaa Xaa Xaa Xaa Xaa
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЕ АНТИТЕЛА IGG, КОТОРЫЕ СВЯЗЫВАЮТСЯ С ТРАНСФОРМИРУЮЩИМ ФАКТОРОМ РОСТА БЕТА-1 С ВЫСОКОЙ АФФИННОСТЬЮ, АВИДНОСТЬЮ И СПЕЦИФИЧНОСТЬЮ | 2016 |
|
RU2728858C2 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ ДЛЯ ЛЕЧЕНИЯ ВИРУСНОЙ ИНФЕКЦИИ ГЕПАТИТА В И СВЯЗАННЫХ СОСТОЯНИЙ | 2016 |
|
RU2733496C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2839153C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ, СВЯЗЫВАЮЩИЕСЯ С 5T4 И 4-1BB, И ОТНОСЯЩИЕСЯ К НИМ КОМПОЗИЦИИ И СПОСОБЫ | 2018 |
|
RU2804458C2 |
| Белки, связывающие NKG2D, CD16 и опухолеассоциированный антиген | 2018 |
|
RU2816716C2 |
| СЛИТЫЙ БЕЛОК GDF15 ДЛИТЕЛЬНОГО ДЕЙСТВИЯ И СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2020 |
|
RU2836280C1 |
| УЛУЧШЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ РЕЦЕПТОРЫ | 2018 |
|
RU2825816C2 |
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2766199C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
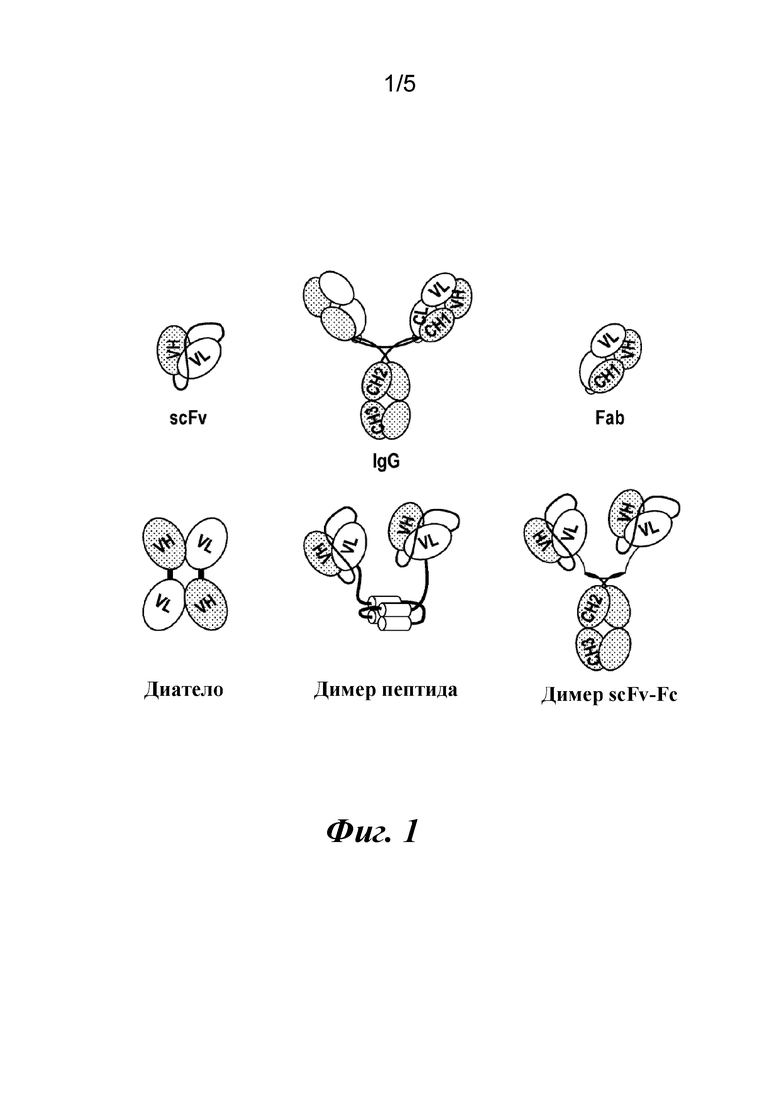
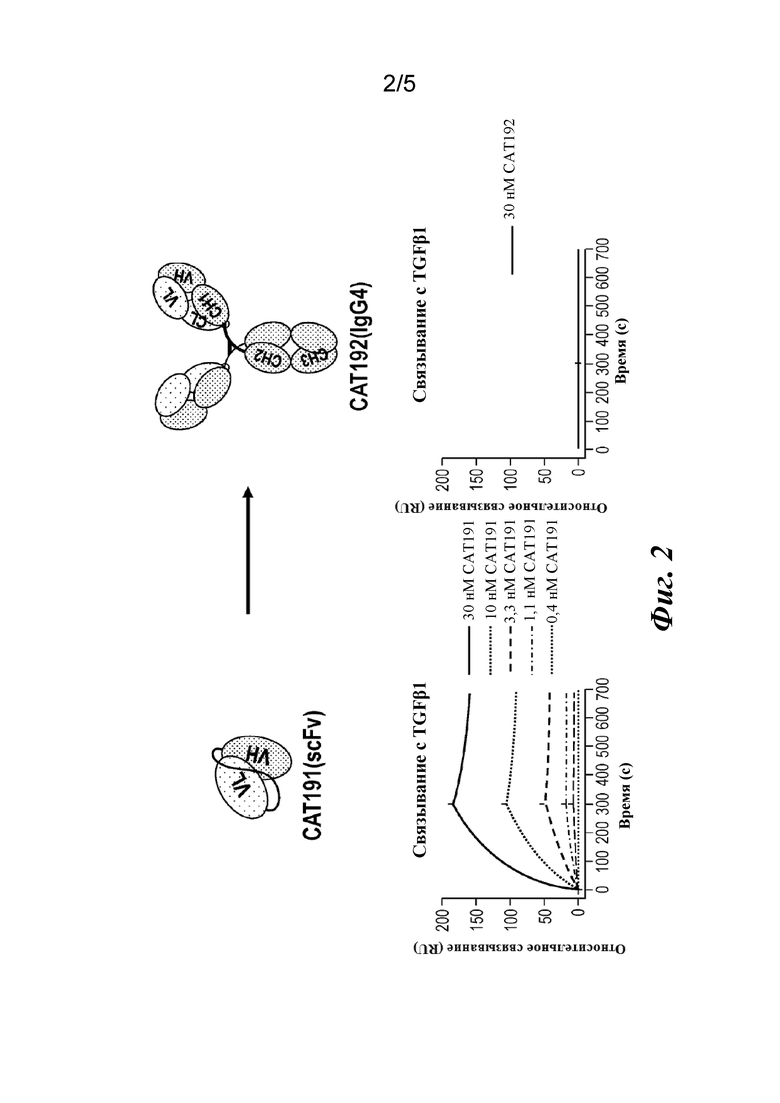
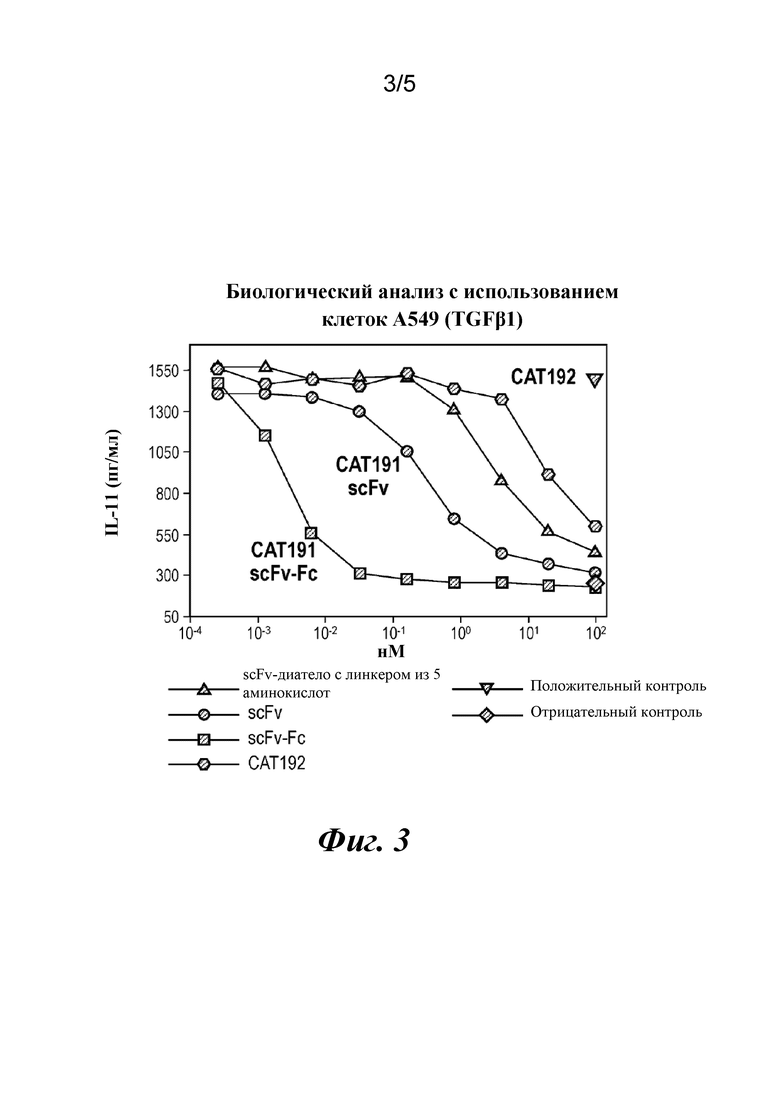
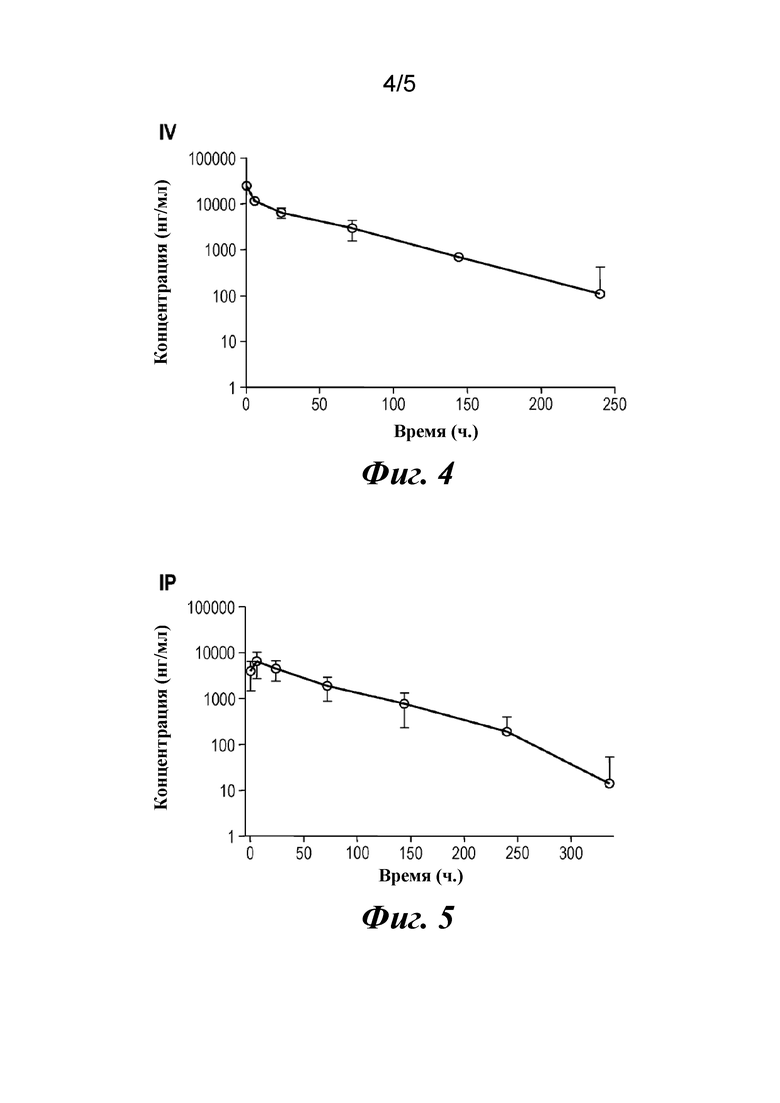
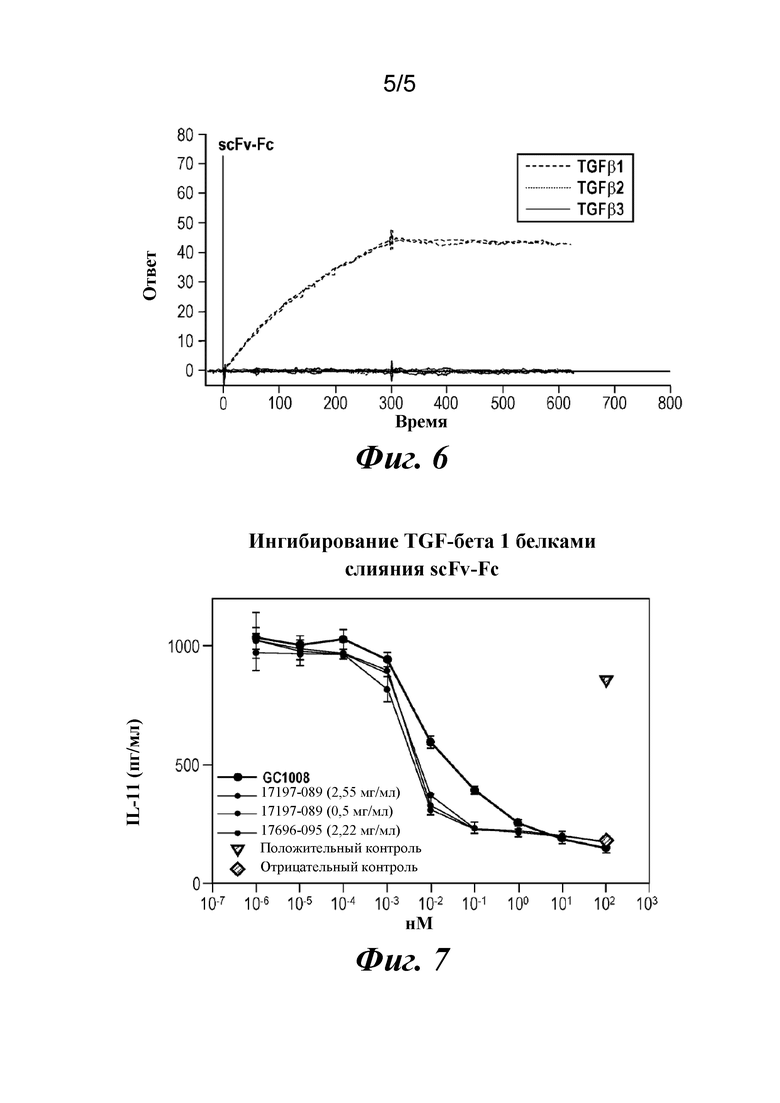
Изобретение относится к области биотехнологии. Предложен выделенный связывающий белок, который избирательно связывается с TGFβ1. Указанный связывающий белок является димером scFv-Fc. Также предложено применение указанного связывающего белка в составе фармацевтической композиции для лечения заболеваний человека, таких как фиброзное заболевание, рак, диффузный кожный системный склероз, заболевание, характеризующееся нарушением ремоделирования костной ткани, или заболевание почек. Изобретение обеспечивает избирательное связывание и нейтрализацию активности TGFβ1 с высокой аффинностью и авидностью. Димеры scFv-Fc являются кандидатами для терапевтических путей применения ввиду комбинации их меньшего размера, высокой избирательности, активности в отношении TGFβ1 и длительного периода полувыведения in vivo. 3 н. и 9 з.п. ф-лы, 7 ил., 3 табл., 6 пр.
1. Выделенный связывающий белок, который избирательно связывается с TGFβ1, где указанный связывающий белок является димером, образованным полипептидной цепью, имеющей формулу в направлении от N-конца к C-концу:
(VH-домен)-(линкер 1)-(VL-домен)-(линкер 2)-(шарнирная область)-(Fc-область),
где
VH-домен содержит область, определяющую комплементарность, тяжелой цепи (HCDR) 1, имеющую аминокислотную последовательность SEQ ID NO: 22, HCDR2, имеющую аминокислотную последовательность SEQ ID NO: 23, и HCDR3, имеющую аминокислотную последовательность SEQ ID NO: 24, 25 или 26; и
VL-домен содержит область, определяющую комплементарность, легкой цепи (LCDR) 1, имеющую аминокислотную последовательность SEQ ID NO: 27 или аминокислотную последовательность SEQ ID NO: 27 c заменой A2S, LCDR2, имеющую аминокислотную последовательность SEQ ID NO: 28, и LCDR3, имеющую аминокислотную последовательность SEQ ID NO: 29;
линкер 1 представляет собой линкер типа [G4S]3;
линкер 2 представлен SEQ ID NO: 20 или ее вариантом, где вариант отличается по длине от SEQ ID NO: 20 на одну-четыре аминокислоты или отличается от SEQ ID NO: 20 наличием до двух аминокислотных замен глицина на серин или серина на глицин;
шарнирная область включает аминокислотную последовательность шарнирной области человеческого IgG1 или IgG4 или аминокислотную последовательность SEQ ID NO: 7 или 21.
2. Выделенный связывающий белок по п. 1, где
VH-домен содержит последовательность человеческого VH-домена, представленную SEQ ID NO: 1 или 2, или ее вариант, имеющий до четырех аминокислотных модификаций включительно; и
VL-домен содержит последовательность человеческого Vκ-домена, представленную SEQ ID NO: 5 или 6, или ее вариант, имеющий до четырех аминокислотных модификаций включительно.
3. Выделенный связывающий белок по любому из пп. 1 или 2, где VH- и VL-домены содержат аминокислотные последовательности, представленные SEQ ID NO: 1 и 5, соответственно.
4. Выделенный связывающий белок по п. 1, где полипептидная цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 9.
5. Выделенный связывающий белок по любому из пп. 1-4, где линкер 1 содержит аминокислотную последовательность SGGGSGGGGSGGGGS (SEQ ID NO: 3) или GGGGSGGGGSGGGGS (SEQ ID NO: 4).
6. Выделенный связывающий белок по любому из пп. 1-5, где линкер 2 представлен SEQ ID NO: 20.
7. Выделенный связывающий белок по любому из пп. 1-6, где Fc-область получена из человеческого IgG1, человеческого IgG4 или варианта человеческого IgG1 или IgG4, в котором модифицированы до десяти аминокислот включительно.
8. Выделенный связывающий белок по любому из пп. 1-7, где выделенный связывающий белок обладает по меньшей мере одной из следующих характеристик:
a) имеет IC50 в отношении человеческого TGFβ1, составляющую менее 1 нМ, в биологическом анализе с использованием клеток A549;
b) характеризуется Kd для человеческого TGFβ1, по меньшей мере на приблизительно 50% более низкой, чем Kd для человеческого TGFβ2, согласно измерениям по методу поверхностного плазмонного резонанса; и
c) характеризуется Kd для человеческого TGFβ1, по меньшей мере на приблизительно 50% более низкой, чем Kd для человеческого TGFβ3, согласно измерениям по методу поверхностного плазмонного резонанса.
9. Фармацевтическая композиция, содержащая терапевтически эффективное количество выделенного связывающего белка по любому из пп. 1-8 и фармацевтически приемлемый наполнитель, для лечения заболевания у человека, нуждающегося в ингибировании активности TGFβ1.
10. Применение выделенного связывающего белка по любому из пп. 1-8 для лечения заболевания у человека, нуждающегося в ингибировании активности TGFβ1.
11. Применение по п. 10, где заболевание выбрано из группы, состоящей из фиброзного заболевания, рака и иммуноопосредованного заболевания.
12. Применение по п. 10, где заболевание представляет собой диффузный кожный системный склероз, заболевание, характеризующееся нарушением ремоделирования костной ткани, или заболевание почек.
| WO 2006116002 A2, 02.11.2006 | |||
| POWERS D.B | |||
| et al., Expression of single-chain Fv-Fc fusions in Pichia pastoris, Journal of Immunological Methods, 2001, Vol.251, Issues 1-2, pp.123-135 | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| ПЕПТИДЫ, ИНГИБИРУЮЩИЕ ТРАНСФОРМИРУЮЩИЙ ФАКТОР РОСТА TGFБЕТА1 | 1999 |
|
RU2232771C2 |

