Область техники
Настоящее изобретение относится к новому промотору и к способу получения целевого продукта с применением этого промотора.
Предшествующий уровень техники
Успехи в понимании внутриклеточных механизмов, наряду с развитием технологии привели к разработке способов получения различных целевых продуктов посредством регулирования метаболизма клеток. На данный момент чаще всего и непосредственно для получения различных низкомолекулярных или высокомолекулярных соединений и белковых лекарственных средств, таких как аминокислоты, полифенолы, флавоноиды, антитела и натуральные каучуки, используют микроорганизмы, принадлежащие к роду Escherichia или к роду Corynebacterium.
Из множества способов регулирования метаболизма, главным образом были изучены системы экспрессии гена с высокой эффективностью для индукции сверхэкспрессии целевых генов. Например, так как промоторы хорошо известны в качестве наиболее важного фактора, вовлеченного в экспрессию генов, было проведено обширное исследование с использованием нескольких промоторов (Ptac, Ptrc и Plac), происходящих из Е. coli. Кроме того, заявитель настоящего изобретения обнаружил происходящий из Corynebacterium ammoniagenes промотор cj1, демонстрирующий сильную активность, составляющую 296% активности промотора tac из Е. coli, в ходе предыдущего исследования (опубликованный для всеобщего ознакомления корейский патент 10-2006-0068505). Однако все еще существует необходимость разработки способа эффективного продуцирования целевого продукта с высоким выходом.
Описание изобретения
Техническая задача
В результате интенсивного исследования для разработки способов эффективного продуцирования целевого продукта посредством повышения экспрессии целевого гена, авторы настоящего изобретения обнаружили, что новый промотор (cj2.2), полученный посредством модификации последовательности существующего промотора cj1, известного как в высшей степени активный, и установили, что новый промотор имеет активность больше, чем промоторную активность cj 1 примерно в 3 раза или более, и тем самым осуществили настоящее изобретение.
Техническое решение
Задача настоящего изобретения заключается в предложении полинуклеотида, имеющего промоторную активность, включающего полинуклеотидную последовательность SEQ ID NO: 1.
Другая задача настоящего изобретения заключается в предложении вектора, включающего полинуклеотид, имеющий промоторную активность.
Еще одна задача настоящего изобретения заключается в предложении клетки-хозяина, в которую введен вектор.
Еще одна задача настоящего изобретения заключается в предложении способа получения целевого вещества, где способ включает культивирование клетки-хозяина; и извлечение целевого вещества из клетки-хозяина или культуральной среды, в которой культивируют клетку-хозяина.
Полезные эффекты
Так как новый промотор по настоящему изобретению может быть введен в микроорганизм, что приводит к повышению уровня экспрессии и активности гена, связанного с ним, целевой продукт, зависимый от промотора и гена, может эффективно продуцироваться.
Описание графических материалов
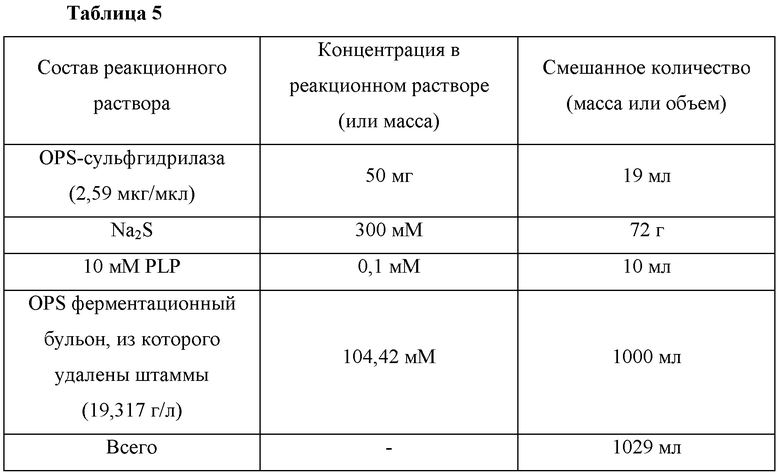
ФИГ. 1 представляет собой диаграмму, иллюстрирующую результаты анализа интенсивности флуоресценции зеленых флуоресцентных белков, экспрессированных в отдельных клонах библиотеки вариантов рекомбинантного вектора, сконструированных согласно одному воплощению настоящего изобретения, где cj 1 представляет собой клон, включающий промотор cj1, и M1 - М20 представляют собой отдельные клоны библиотеки.
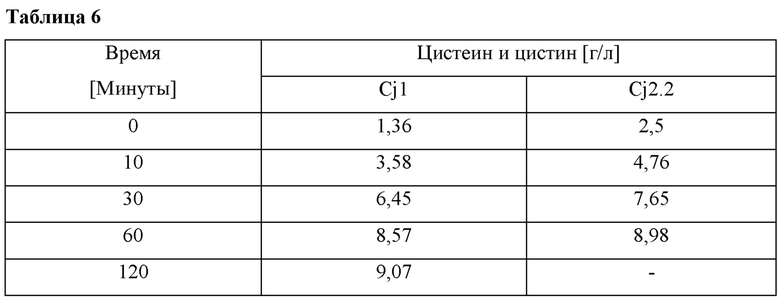
ФИГ. 2 представляет собой карту расщепления вектора, включающего промотор cj2.2 и ген metZ, полученного согласно одному воплощению настоящего изобретения.
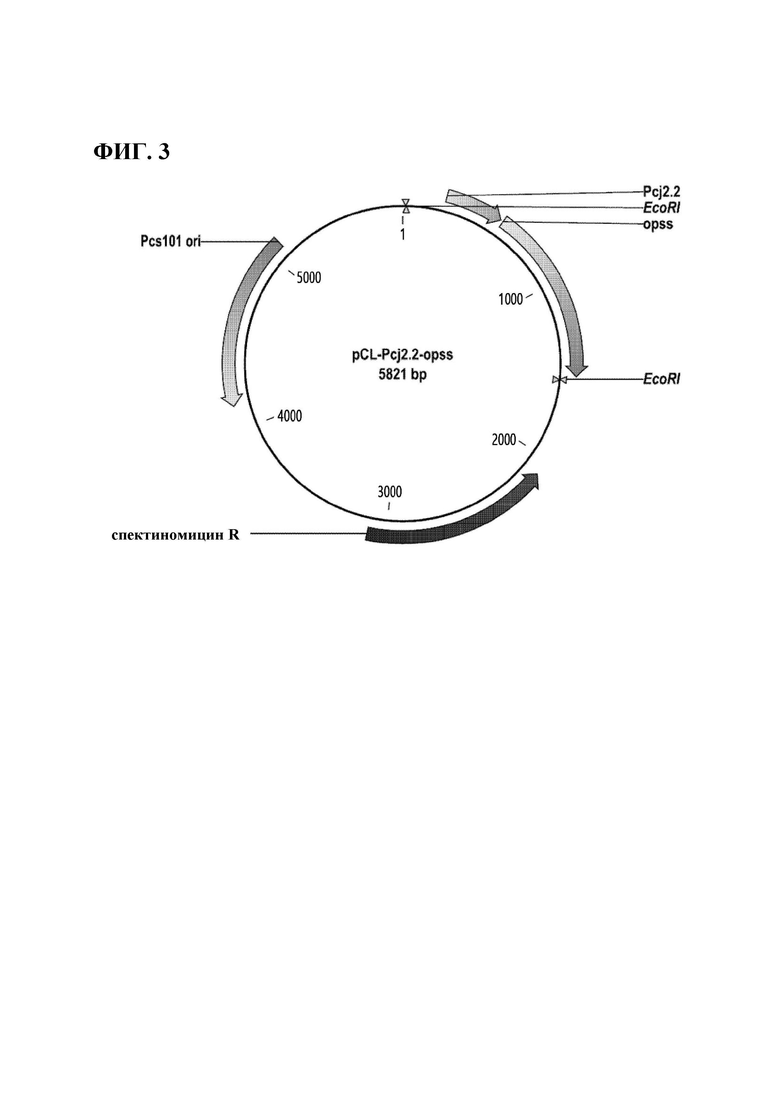
ФИГ. 3 представляет собой карту расщепления вектора, включающего промотор cj2.2 и ген opss, полученного согласно одному воплощению настоящего изобретения.
Лучший вариант осуществления изобретения
Ниже настоящее изобретение будет описано подробно. При этом каждое описание и воплощение, раскрытое в настоящем изобретении, может быть использовано в данном описании изобретения для описания разных описаний и воплощений. Другими словами, все комбинации различных компонентов, раскрытых в настоящем описании изобретения, включены в объем настоящего изобретения. Кроме того, объем настоящего изобретения не следует ограничивать подробным описанием, представленным ниже.
В одном аспекте настоящего изобретения предложен полинуклеотид, имеющий промоторную активность, содержащий полинуклеотидную последовательность SEQ ID NO: 1.
При использовании здесь термин "промотор" относится к нетранслируемой нуклеотидной последовательности, включающей сайт связывания полимеразы, расположенный выше кодирующей области, и имеющей активность инициации транскрипции целевого гена промотора в мРНК, то есть описывает область ДНК, которая управляет инициацией транскрипции конкретного гена, когда с ней связывается полимераза. Промотор может быть расположен в 5'-области сайта инициации транскрипции мРНК.
В настоящем изобретении полинуклеотид, содержащий полинуклеотидную последовательность SEQ ID NO: 1 и имеющий промоторную активность, может использоваться взаимозаменяемо с "полинуклеотидом" или "промотором cj2.2". Везде в описании изобретения можно использовать все описанные выше термины.
Для целей настоящей заявки полинуклеотид может иметь более высокую промоторную активность, чем активность обычных промоторов. Кроме того, полинуклеотид может не только повышать уровень экспрессии целевого гена, функционально связанного с полинуклеотидом в нужной клетке-хозяине, но также повышать уровень экспрессии и активность белка, кодируемого целевым геном, в дополнение к уровню экспрессии целевого гена.
Кроме того, этот полинуклеотид можно использовать в качестве универсального промотора.
При использовании здесь, термин "целевой ген" относится к гену, кодирующему целевой белок, уровень экспрессии которого должен быть повышен, например к гену, вовлеченному в продуцирование аминокислот, без ограничения этим.
Конкретно, ген может представлять собой ген, кодирующий белок, вовлеченный в продуцирование аминокислот, таких как метионин или цистеин, более конкретно ген, кодирующий метионин-превращающий фермент, вовлеченный в продуцирование метионина, или цистеин-превращающий фермент, вовлеченный в продуцирование цистеина, еще более конкретно ген, кодирующий О-ацетилгомосеринсульфгидрилазу, вовлеченную в продуцирование метионина, или О-фосфосеринсульфгидрилазу (OPSS), вовлеченную в продуцирование цистеина, и наиболее конкретно ген metZ или ген Opss, без ограничения ими. Последовательности соответствующих генов легкодоступны специалисту в данной области техники из известной базы данных, такой как GenBank Национального института здоровья (USA).
В настоящем изобретении полинуклеотид может включать нуклеотидную последовательность SEQ ID NO: 1.
Дополнительно, нуклеотидная последовательность по настоящему изобретению может быть модифицирована посредством мутагенеза, хорошо известного в данной области техники, такого как прямая эволюция и сайта-направленный мутагенез.
Таким образом, полинуклеотид может включать полинуклеотид, имеющий нуклеотидную последовательность с гомологией 60% или более, конкретно 70% или более, более конкретно 80% или более, еще более конкретно 83% или более, 84% или более, 88% или более, 90% или более, 93% или более, 95% или более, или 97% или более с нуклеотидной последовательностью SEQ ID NO: 1. Кроме того очевидно, что любой полинуклеотид, имеющий полинуклеотидную последовательность с гомологией, описанной выше, и включающий делецию, модификацию, замену или добавление одной или нескольких последовательностей, входит в объем настоящего изобретения, при условии что этот полинуклеотид сохраняет биологические активности, по существу идентичные или соответствующие активностям нуклеотидной последовательности SEQ ID NO: 1.
При использовании здесь термин "гомология" относится к степени идентичности с заданной нуклеотидной последовательностью и может быть указана в виде процента. Везде в описании изобретения гомологичная последовательность, имеющая такую же или подобную активность с заданной нуклеотидной последовательностью, обозначается в виде "% гомологии". Гомология с нуклеотидной последовательностью может быть определена с использованием любого алгоритма, известного в данной области техники, например BLAST (Ссылка: Karlin and Altschul, Pro. Natl. Acad. Sci. USA, 90, 5873, 1993) или FASTA, введенного Pearson (Ссылка: Methods Enzymol., 183, 63, 1990). На основе алгоритма BLAST были разработаны программы, известные как BLASTN или BLASTX (Ссылка: http://www.ncbi.nlm.nih.gov).
Термин "жесткие условия" относится к условиям, в которых обеспечивается специфическая гибридизация между полинуклеотид ами. Такие условия подробно раскрыты в известных документах. Например, условия могут включать осуществление гибридизации генов, имеющих высокую гомологию, например гомологию 60% или более, в частности 90% или более, более конкретно 95% или более, еще более конкретно 97% или более, и наиболее конкретно 99% или более, без осуществления гибридизации генов, имеющих гомологию более низкую, чем указанные выше гомологии, или выполнение отмывки один раз, конкретно два или три раза, в обычных условиях отмывки для Саузерн-гибридизации с концентрацией солей и температурой 60°С, 1×SSC и 0,1% SDS, конкретно 60°С, 0,1×SSC и 0,1% SDS, и более конкретно 68°С, 0,1×SSC и 0,1% SDS. Для гибридизации требуется, чтобы два нуклеотида имели комплементарные последовательности, хотя возможно ошибочное спаривание оснований в зависимости от степени жесткости гибридизации. Термин «комплементарный» используют для описания взаимоотношения между нуклеотидными основаниями, способными гибридизоваться друг с другом. Например, что касается ДНК, аденозин комплементарен тимину, и цитозин комплементарен гуанину. Таким образом, настоящее изобретение может включать не только по существу подобную полинуклеотидную последовательность, но также фрагмент полинуклеотида, выделенный, но комплементарный целой последовательности.
В частности, полинуклеотид, имеющий гомологию, может быть обнаружен с использованием описанных выше условий в условиях гибридизации, включающих стадию гибридизации при значении Tm 55°С. Также, значение Tm может представлять собой 60°С, 63°С или 65°С, без ограничения ими, и может подходящим образом регулироваться специалистом в данной области техники согласно цели. Степень жестких условий для гибридизации полинуклеотида может зависеть от длины полинуклеотида и степени комплементарности, и переменные хорошо известны в данной области техники (ссылка на Sambrook et al., supra, 9.50-9.51, 11.7-11.8).
В частности, выражение "включающий нуклеотидную последовательность SEQ ID NO: 1" не исключает добавление, делецию и/или модификацию нуклеотида, которые могут иметь место во время лигирования с целевым геном, с использованием фермента рестрикции или подобного, когда полинуклеотид в качестве промотора лигируют с целевым геном.
Также, полинуклеотид, включающий нуклеотидную последовательность SEQ ID NO: 1 и имеющий промоторную активность, может включать любую нуклеотидную последовательность, гибридизированную с нуклеотидной последовательностью, полностью или частично комплементарной нуклеотидной последовательности SEQ ID NO: 1 в жестких условиях, чтобы иметь активность промотора по настоящему изобретению, без ограничения.
В другом аспекте настоящего изобретения предложен вектор, включающий полинуклеотид, имеющий промоторную активность.
При этом "промотор" является таким, как описано выше.
При использовании здесь, термин "вектор", в виде искусственной молекулы ДНК, включающий генетическое вещество, способное экспрессировать целевой ген в подходящей клетке-хозяине, относится к ДНК-продукту, включающему: подходящую регуляторную последовательность экспрессии гена; и нуклеотидную последовательность целевого гена, функционально связанную с ней.
Конкретно, термин "регуляторная последовательность экспрессии гена" относится к последовательности, включающей полинуклеотид, имеющий промоторную активность и способный экспрессировать целевой ген, функционально связанный с ним.
В частности, регуляторная последовательность экспрессии гена может дополнительно включать последовательность оператора для контроля над транскрипцией, последовательность, кодирующую подходящий сайт связывания с рибосомой мРНК, и ДНК для регуляции окончания транскрипции и трансляции, в дополнение к промотору для инициации транскрипции гена, но без ограничения этим. Кроме того, регуляторная последовательность, подходящая для прокариотов, может включать промотор и сайт связывания с рибосомой, без ограничения ими. Полинуклеотид, имеющий промоторную активность по настоящему изобретению, может являться частью последовательности для регуляции экспрессии гена, как описано выше, согласно цели специалиста в данной области техники.
Кроме того, термин "функционально связанный" означает функционально связанный с нуклеотидной последовательностью целевого гена, так что полинуклеотид, имеющий промоторную активность, инициирует и опосредует транскрипцию целевого гена. Функциональное связывание может быть получено с использованием метода генетической рекомбинации, известного в данной области техники, и сайт-направленное расщепление и лигирование ДНК может быть выполнено с использованием рестрикционного фермента лигазы и подобных, известных в данной области техники, без ограничения ими.
Вектор при использовании здесь может представлять собой любой вектор, экспрессируемый в клетке-хозяине, без ограничения, и клетка-хозяин может быть трансформирована с использованием любого вектора, хорошо известного в данной области техники. Примеры общеизвестных векторов могут включать плазмиду, космиду, вирус и бактериофаг в природной или рекомбинантной форме.
Например, pWE15, М13, λLB3, λBL4, λIXII, λASHII, λAPII, λt10, λt11, Charon4A и Charon21A можно использовать в качестве фагового вектора или космидного вектора, и векторы на основе epBR, на основе pUC, на основе pBluescriptII, на основе pGEM, на основе pTZ, на основе pCL и на основе рЕТ можно использовать в качестве плазмидного вектора.
Кроме того, эндогенный промотор хромосомы может быть заменен на полинуклеотид по настоящему изобретению, имеющий промоторную активность, с использованием вектора, применяемый для встраивания в хромосому клетки-хозяина. Например, можно использовать векторы pECCG117, pDZ, pACYC177, pACYC184, pCL, pUC19, pBR322, pMW118, pCC1BAC, pCES208 и pXMJ19, но настоящее изобретение не ограничивается ими.
Кроме того, встраивание полинуклеотида в хромосому может быть выполнено любым способом, известным в данной области техники, например посредством гомологичной рекомбинации.
Так как вектор по настоящему изобретению может быть встроен в хромосому посредством гомологичной рекомбинации, этот вектор может дополнительно включать селективный маркер для идентификации вставки в хромосому. Селективный маркер используют для отбора клеток, трансформированных вектором, то есть для того, чтобы установить, встроен ли в нее полинуклеотид, и может представлять собой любой маркер, который обеспечивает селектируемые фенотипы, такие как устойчивость к лекарственному средству, потребность в питательных веществах, устойчивость к цитотоксическим агентам или экспрессия поверхностных белков. Так как только клетки, экспрессирующие селективный маркер, способны выживать или демонстрировать различные фенотипы в среде, обработанной селективным агентом, посредством этого могут быть отобраны трансформированные клетки.
В другом аспекте настоящего изобретения предлагается клетка-хозяин, в которую введен вектор.
При этом "вектор" является таким, как описано выше.
При использовании здесь термин "клетка-хозяин" относится к трансформанту, трансформированному вектором, содержащим полинуклеотид.
Здесь термин "трансформация" относится к процессу введения вектора, содержащего полинуклеотид, имеющий промоторную активность, в клетку-хозяина так, чтобы целевой ген, регулируемый этим полинуклеотидом, экспрессировался в клетке-хозяине. Конкретно, клетка-хозяин не ограничена до тех пор, пока полинуклеотид вводят в клетку-хозяина и он действует в ней в качестве промотора, и, соответственно, экспрессия целевого гена, регулируемая полинуклеотидом, повышается. Например, клетка-хозяин может представлять собой клетку микроорганизма, растения или животного, конкретно микроорганизм рода Escherichia или рода Corynebacterium, и более конкретно Е. coli в качестве микроорганизма рода Escherichia, без ограничения ими.
Методы трансформации включают любые методы, обеспечивающие ввод вектора в клетку-хозяина, и могут быть выполнены посредством подходящих стандартных техник, хорошо известных в данной области техники, выбранных в зависимости от клетки-хозяина. Например, можно использовать электропорацию, осаждение фосфатом кальция (СаРО4), осаждение хлоридом кальция (CaCl2), микроинъекцию, метод с использованием полиэтиленгликоля (ПЭГ), метод с использованием DEAE-декстрана (диэтиламиноэтил-декстрана), метод с использованием катионных липосом и метод с использованием ацетата лития-ДМСО, но настоящее изобретение не ограничено ими.
В другом аспекте настоящего изобретения предложен способ получения целевого вещества, включающий: культивирование клетки-хозяина; и извлечение целевого вещества из клетки-хозяина или культуральной среды, в которой культивируют клетку-хозяина.
При этом "клетка-хозяин" является такой, как описано выше.
В настоящем изобретении целевое вещество может представлять собой аминокислоту. Конкретно, аминокислота может представлять собой L-аминокислоту, если не указано иное, и может быть выбрана из группы, состоящей из глицина, аланина, валина, лейцина, изолейцина, треонина, серина, цистеина, глутамина, метионина, аспарагиновой кислоты, аспарагина, глутаминовой кислоты, лизина, аргинина, гистидина, фенилаланина, тирозина, триптофана, пролина и любой их комбинации, без ограничения ими. Более конкретно, аминокислота может представлять собой, но без ограничения ими, метионин или цистеин.
При использовании здесь, термин "культивирование" относится к выращиванию клетки-хозяина в искусственно регулируемой подходящей среде. В настоящем изобретении способ получения целевого вещества с использованием клетки-хозяина, включающей полинуклеотид, может быть выполнен любым способом, хорошо известным в данной области техники. Конкретно, культивирование может быть выполнено непрерывно посредством периодического процесса, периодического процесса с подпиткой или повторяющегося периодического процесса с подпиткой, без ограничения ими. Культуральная среда, используемая для культивирования, необходима для удовлетворения потребностей конкретных штаммов в подходящем способе. Примеры источников Сахаров, которые можно использовать в культуральной среде, включают: сахариды и углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза, масла и жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло, жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота, спирты, такие как глицерин и этиловый спирт, и органические кислоты, такие как уксусная кислота. Эти вещества можно использовать отдельно или в комбинации, без ограничения ими.
Примеры доступных источников азота могут включать пептон, дрожжевые экстракты, мясные экстракты, солодовые экстракты, жидкий кукурузный экстракт, соевый шрот и мочевину или неорганические соединения, такие как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота также можно использовать отдельно или в комбинации, без ограничения ими.
Примеры доступных источников фосфора включают дигидрофосфат калия, гидрофосфат калия или их соответствующие соли натрия. Культуральная среда дополнительно может включать соль металла, необходимую для роста, такую как сульфат магния или сульфат железа. Также, в дополнение к указанным выше веществам можно использовать важные для роста вещества, такие как аминокислоты и витамины. Кроме того, в культуральной среде можно использовать необходимые предшественники. Описанные выше вещества можно добавлять к культуральной среде во время процесса культивирования в ходе периодического процесса или непрерывного процесса, подходящего для культур.
При культивировании клетки-хозяина можно регулировать рН, используя надлежащим образом щелочное соединение, такое как гидроксид натрия, гидроксид калия и аммиак, или кислое соединение, такое как фосфорная кислота или серная кислота. Дополнительно, для предотвращения образования пены можно использовать пеногаситель, такой как полигликолевый эфир жирной кислоты. Для поддержания аэробного состояния можно вводить в культуры кислород или кислородсодержащий газ (например воздух).
Температура культур (культуральная среда) обычно может находиться в диапазоне от 20°С до 45°С, конкретно в диапазоне от 25°С до 40°С. Культивирование можно продолжать до получения желательного выхода целевого вещества, конкретно в течение от 10 до 160 часов.
Целевое вещество может быть отделено и извлечено из культур (культуральной среды) посредством любого способа, известного в данной области техники. Отделение может быть выполнено посредством центрифугирования, фильтрации, хроматографии, кристаллизации и тому подобного. Например, супернатант, полученный после удаления биомассы путем центрифугирования культур при низкой скорости, может быть подвергнут ионообменной хроматографии, но настоящее изобретение не ограничивается этим.
Извлечение может дополнительно включать процесс очистки.
Осуществление изобретения
Ниже настоящее изобретение будет описано более подробно со ссылкой на следующие ниже примеры. Однако эти примеры служат только для иллюстративных целей, и настоящее изобретение не следует ограничивать этими примерами.
Пример 1: Конструирование библиотеки вариантов рекомбинантного вектора, включающих последовательность промотора cj1
Для обнаружения промоторов, способных сильно индуцировать экспрессию гена в микробных штаммах, была сконструирована библиотека вариантов на основе промотора cj1, который известен как проявляющий высокую активность (общедоступная корейская патентная публикация No. 10-2006-0068505).
Конкретно, библиотеку вариантов конструировали, используя рекомбинантный вектор pCL-cj1-gfp, включающий последовательность промотора cj1 в качестве матрицы. Библиотеку конструировали, используя набор для ПНР с ошибками (Clontech Diversify® PCR Random Mutagenesis Kit). Полимеразные цепные реакции (ПЦР) проводили, используя праймеры SEQ ID NO: 2 и 3 в условиях, когда могут иметь место вариации. Условия для получения одной вариации на 1000 п. о. являются следующими: после предварительного нагрева при 94°С в течение 30 секунд в присутствии 40 мкМ dGTP (дезоксигуанозина трифосфат) (конечная реакция) без использования MnSO4 (8 мМ), процесс поддерживания при 94°С в течение 30 секунд и затем при 68°С в течение 1 минуты повторяли 25 раз. ПЦР-продукты, полученные посредством этого, подвергали 25 циклам процесса, включающего денатурацию при 95°С в течение 50 секунд с использованием мега-праймера (от 125 до 500 нг), отжиг при 60°С в течение 50 секунд и удлинение при 68°С в течение 12 минут и затем обрабатывали DpnI для конструирования библиотеки путем трансформирования штаммов Е. coli DH5α.
Пример 2: Оценка интенсивности активности зеленого флуоресцентного белка (GFP) рекомбинантного вектора
Для оценки активностей вариантов рекомбинантного вектора из библиотеки, сконструированной в Примере 1 выше, активности зеленых флуоресцентных белков (GFP) сравнивали друг с другом.
Конкретно, библиотеку, состоящую из примерно 10000 или более штаммов, высевали на М9 среду и культивировали при 37°C с 900 об/мин в течение примерно 24 часов. Культивированную Е. coli центрифугировали при 4000 об/мин в течение 10 минут с получением штаммов и суспендировали в фосфатно-солевом буферном растворе (PBS). Затем варианты с высокой интенсивностью флуоресценции зеленого флуоресцентного белка (GFP) отбирали с помощью анализатора флуоресценции.
В первом скрининге были отобраны 20 вариантов, имеющих более высокие активности GFP, чем активность промотора cj1, в примерно 1,1 - примерно 5 раз (ФИГ. 1). Среди них вариант М11 промотора, имеющий самую высокую активность GFP, был назван промотором cj2.2 (SEQ ID NO: 1) и его последовательность была проанализирована.
Пример 3: Получение вектора экспрессии промотора cj2.2
Так как было подтверждено, что промотор cj2.2 имел наибольшую активность среди вариантов рекомбинантного вектора из библиотеки в Примере 2, сайт-направленный мутагенез проводили, используя вектор pCL_Pcj1 для введения промотора cj2.2 в микробные штаммы.
Конкретно, выполняли ПЦР, используя праймеры, представленные SEQ ID NO: 4 и 5. ПЦР включает процесс, включающий денатурацию при 94°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд и удлинение при 68°С в течение 12 минут, и этот процесс повторяли 18 раз.
Продукты, полученные в указанном выше процессе, обрабатывали DpnI и штаммы DH5a трансформировали посредством этого с получением вектора pCL_Pcj2.2, включающего промотор cj2.2.
Пример 4: Измерение активности промотора cj2.2
Пример 4-1: Измерение уровня экспрессии метионин-превращающего фермента (О-ацетилгомосеринсульфгидрилаза)
Для идентификации активности промотора cj2.2 и способности посредством этого индуцировать экспрессию гена, метионин-превращающий фермент (О-ацетилгомосеринсульфгидрилазу) лигировали с промотором cj2.2 и определяли уровень его экспрессии.
Конкретно, ген metZ (корейская патентная публикация №10-1250651), кодирующий О-ацетилгомосеринсульфгидрилазу, клонировали в pCL_Pcj2.2 вектор, полученный в Примере 3. ПЦР выполняли, используя хромосому Rhodobacters phaeroides в качестве матрицы и праймеры, представленные SEQ ID NO: 6 и 7, и в повторяющемся режиме выполняли 30 циклов процесса, включающего денатурацию при 94°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд и удлинение при 72°С в течение 2 минут. Фрагменты ДНК, полученные из ПЦР, расщепляли с помощью BamHI/SalI и клонировали в вектор pCL_Pcj2.2, расщепленный посредством такого же фермента рестрикции. Вектор, полученный, как описано выше, был назван pCL-Pcj2.2-metZ, и схематическое изображение вектора показано на ФИГ. 2.
Штаммы Е. coli K12 трансформировали с помощью вектора pCL-Pcj2.2-metZ и культивировали на среде LB для чашек с добавлением 50 мкг/л спектиномицина для отбора колоний. Отобранные колонии высевали на 3 мл среды LB с добавлением 50 мкг/л спектиномицина, и культивировали при 37°С, 200 об/мин в течение 16 часов. Сразу же после пересева полученных продуктов в 25 мл новой жидкой среды LB (колба 250 мл) и культивирования продуктов до получения OD600 от 0,5 до 0,6 (в течение 2-3 часов) при тех же условиях культивирования, продукты культивировали в 500 мл среды LB (1 л сосуд) с добавлением 4% глюкозы. После завершения культивирования отбирали 1 мл культуральной среды и удаляли супернатант посредством центрифугирования с получением клеток. После промывания клеток 0,1 М калий-фосфатным буфером (рН 7,5) клетки суспендировали с помощью 1 мл калий-фосфатного буфера и лизировали с помощью ультразвукового диспергатора.
Затем измеряли активность метионин-превращающего фермента (О-ацетилгомосеринсульфгидрилазы), содержащегося в лизатах. Для измерения активности фермента 5 мкл лизатов добавляли к смешанному раствору субстрата, где были смешаны 1 мл О-ацетилгомосерина (80 г/л), 0,01 мл метилмеркаптана натрия (15%, масс./об.), 0,01 мл О-ацетилгомосеринсульфгидрилазы и 0,1 мМ пиридоксаль-5'-фосфата, и смесь перемешивали при 800 об/мин и 33°С для выполнения ферментативных реакций. После завершения реакций анализировали концентрации метионина и О-ацетилгомосерина посредством ВЭЖХ и вычисляли активность белка обычным способом, используя результаты анализа ВЭЖХ. Результаты показаны в Таблице 1 ниже.
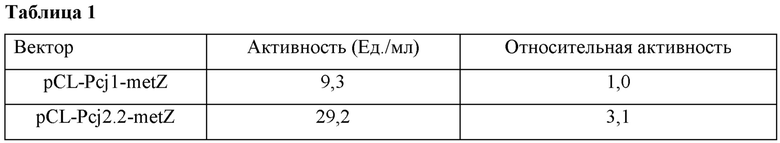
Как показано в Таблице 1, было подтверждено, что промотор cj2.2 по настоящему изобретению повышал активность О-ацетилгомосеринсульфгидрилазы до активности примерно в 3,1 раза большей, чем активность обычного промотора cj1, как известно, сильно индуцирующего экспрессию гена в микробных штаммах.
Таким образом, было подтверждено, что промотор cj2.2 имеет более сильную активность, чем обычные промоторы, и, соответственно, активность целевого фермента, кодируемого геном, связанным с ним, может быть дополнительно повышена.
Пример 4-2: Измерение уровня экспрессии цистеин-превращающего фермента (О-фосфосеринсульфгидрилазы)
Для определения активности промотора cj2.2 и, тем самым, способности индуцировать экспрессию гена, цистеин-превращающий фермент (О-фосфосеринсульфгидрилазу) лигировали с промотором cj2.2 и определяли уровень его экспрессии.
Конкретно, О-фосфосеринсульфгидрилазу (ниже упоминаемую как "OPSS", корейская патентная публикация №10-1208267) клонировали в вектор, полученный в Примере 3. ПЦР выполняли, используя хромосому Mycobacterium smegmatics в качестве матрицы, и праймеры, представленные SEQ ID NO: 8 и 9, и вектор получали способом, аналогичным способу из Примера 4-1. Полученный вектор был назван pCL-Pcj2.2-opss, и схематическое изображение этого вектора показано на ФИГ. 3.
Штаммы Е. coli K12 трансформировали с помощью вектора pCL-Pcj2.2-opss и культивировали на среде для чашек LB с добавлением 50 мкг/л спектиномицина для селекции колоний. Отобранными колониями засевали 5 мл среды LB с добавлением 50 мкг/л спектиномицина, и культивировали при 37°С, 200 об/мин в течение 16 часов. Сразу же после пересева полученных продуктов в 25 мл новой жидкой среды LB (250 мл колба) и культивирования продуктов до получения OD600 от 0,5 до 0,6 (в течение 2-3 часов) в таких же условиях культивирования, продукты культивировали в 500 мл среды LB (1 л сосуд) с добавлением 4% глюкозы, до истощения глюкозы. Клетки лизировали путем добавления 2% ксилена (об./об.) к культуральной среде и лизаты использовали для оценки активности.
Затем активность фермента OPSS, полученного с использованием вектора pCL-Pcj2.2-opss, оценивали согласно известному способу в условиях, раскрытых в документах, представленных в Mino K and Ishikawa K, FEBS letters, 551: 133-138, 2003; BurnsKE et al, J. Am.Chem. Soc., 127: 11602-11603, 2005; и Westrop GD et al, J. Biol.Chem., 281: 25062-25075, 2006). Субстрат добавляли в миллилитрах, и условия для измерения активности фермента показаны в Таблице 2 ниже.
Смесь, показанную в Таблице 2, за исключением фермента, предварительно инкубировали при 37°С в течение 5 минут, и добавляли к ней 50 мг фермента OPSS с последующим взаимодействием при 37°С. Затем добавляли 0,1 мл 33,2% ТСА (трихлоруксусная кислота) в реакционный раствор для прекращения реакции и реакционный раствор разбавляли 0,1 н. HCl с последующим исследованием посредством ЖХ (жидкостной хроматографии) для анализа цистеина и цистина. Измеренные активности показаны в Таблице 3 ниже.
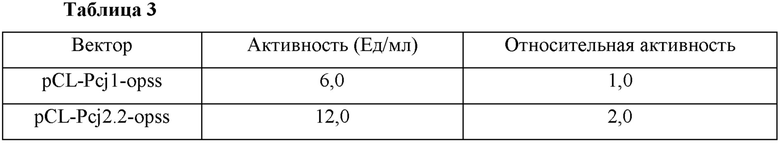
Как показано в Таблице 3, было подтверждено, что промотор cj2.2 по настоящему изобретению повышал активность О-фосфосеринсульфгидрилазы примерно в два раза больше, чем обычный промотор cj1, как известно, сильно индуцирующий экспрессию гена в микробных штаммах.
Таким образом, было подтверждено, что промотор cj2.2 имеет более сильную активность, чем обычные промоторы, и, следовательно, активность целевого фермента, кодируемого геном, связанным с ним, может быть дополнительно повышена.
Пример 5: Получение целевого продукта с использованием фермента, полученного при помощи промотора cj2.2
Пример 5-1: Получение метионина
Так как было подтверждено, что активность метионин-превращающего фермента (О-ацетилгомосеринсульфгидрилазы) была повышена в результате использования промотора cj2.2 согласно Примеру 4-1, активность промотора cj2.2 и эффективность продуцирования метионина в реакторной системе определяли по продуцированию целевого продукта метионина с использованием фермента.
Между тем О-ацетилгомосеринсульфгидрилаза превращает О-ацетилгомосерин в метионин.
Конкретно, в качестве субстрата ферментативной реакции использовали О-ацетилгомосерин, полученный посредством ферментирования микробного штамма, раскрытого в корейском патенте №10-1250651. После удаления клеток из культивируемого ферментационного бульона с использованием центрифуги, 500 мл культурального раствора, имеющего концентрацию примерно 70 г/л, использовали в качестве субстрата. В качестве фермента О-ацетилгомосеринсульфгидрилазы использовали 50 мл лизатов (ферментный раствор), полученных в Примере 4-1. В ферментативной реакции использовали 50 мл метилмеркаптана натрия и добавляли к нему пиридоксаль-5'-фосфат (Sigma, USA) в концентрации 0,1 мМ. Реакцию выполняли при рН 7,0, 33°С при перемешивании 700 об/мин. Реакцию проводили в течение 3 часов, с добавлением в нее метилмеркаптана натрия. Концентрации О-ацетилгомосерина и L-метионина анализировали в зависимости от времени посредством ВЭЖХ, и результаты показаны в Таблице 4 ниже.
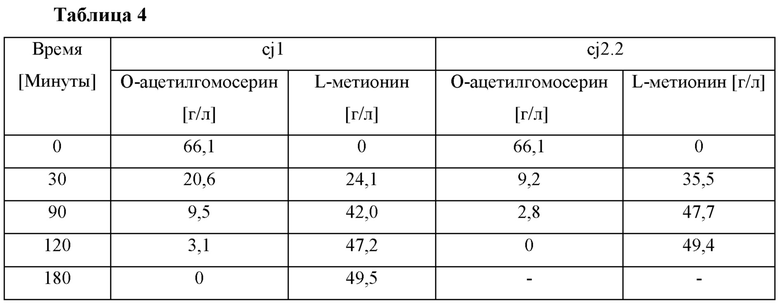
Как показано в Таблице 4, было подтверждено, что скорость превращения из О-ацетилгомосерина в L-метионин при использовании промотора cj2.2 по настоящему изобретению была более высокой, чем при использовании обычного промотора cj1, как известно, сильно индуцирующего экспрессию гена в микробных штаммах, и, таким образом, посредством промотора cj2.2 L-метионин получали быстрее.
Это обусловлено тем, что активность целевого продукта, кодируемого геном, то есть метионин-превращающего фермента (О-ацетилгомосеринсульфгидрилазы), была повышена, когда ген экспрессировался с использованием промотора cj2.2 по настоящему изобретению.
Пример 5-2: Получение цистеина
Так как было подтверждено, что активность цистеин-превращающего фермента (О-фосфосеринсульфгидрилазы) была повышена при использовании промотора cj2.2 согласно Примеру 4-2, активность промотора cj2.2 и эффективность продуцирования цистеина в реакторной системе определяли по продуцированию целевого продукта цистеина с использованием фермента
При этом О-фосфосеринсульфгидрилаза превращает OPS в цистеин (или цистин, который представляет собой оксид цистеина).
Конкретно, концентрацию цистеина в реакционном растворе для превращения количественно определяли методом Гайтона и анализа посредством ЖХ. Кроме того, цистин, полученный в результате окисления цистеина, количественно определяли посредством ЖХ-анализа. Сначала, до реакции превращения в масштабе 1 л сосуда, ферментационный бульон OPS, полученный путем ферментации в масштабе 1 л сосуда, центрифугировали (при 10000 об/мин в течение 10 минут при 4°С) с получением супернатанта и супернатант подвергали мембранной фильтрации (0,45 мкМ) для удаления штаммов. К ферментационному бульону OPS (19,317 г/л) добавляли 72 г Na2S и осадки, полученные из него, удаляли, используя фильтровальную бумагу Whatman (6 мкм). Затем добавляли в него 10 мМ пиридоксаль-5'-фосфата (PLP) и смесь предварительно инкубировали при 37°С, 200 об/мин в течение 5 минут и, наконец, добавляли в него 50 мг OPS-сульфгидрилазы для выполнения реакции превращения в масштабе 1 л сосуда. Условия измерения активности фермента такие, как показано в Таблице 5 ниже.
Затем 100 мкл реакционного раствора отбирали и смешивали с 100 мкл 33,2% ТС А каждые 0, 10, 30, 60 и 120 минут для прекращения реакции, и реакционный раствор разбавляли 0,1 н. HCl с последующим анализом посредством ЖХ для анализа цистеина и цистина. Также, цистеин и цистин определяли количественно, используя метод Гайтона. Результаты анализа показаны в Таблице 6 ниже.
Как показано в Таблице 6, было подтверждено, что цистеин и цистин продуцировались быстрее при использовании промотора cj2.2 по настоящему изобретению, чем при использовании обычного промотора cj1, как известно, сильно индуцирующего экспрессию гена в микробных штаммах.
Это обусловлено тем, что активность целевого продукта, кодируемого геном, то есть цистеин-превращающего фермента (О-фосфосеринсульфгидрилазы), была повышена, когда ген экспрессировался с использованием промотора cj2.2 по настоящему изобретению.
Указанное выше описание настоящего изобретения представлено для иллюстрации, и специалисту в данной области техники понятно, что могут быть произведены различные изменения и модификации без изменения технической концепции и существенных признаков настоящего изобретения. Поэтому очевидно, что описанные выше воплощения являются иллюстративными во всех аспектах и не ограничивают настоящее изобретение. Различные воплощения, раскрытые в данном описании изобретения, не предназначены для ограничения, с истинным объемом и сущностью, указанными следующей формулой изобретения. Настоящее изобретение подлежит ограничению только терминами прилагаемой формулы изобретения, вместе с полным объемом эквивалентов, на которые такая формула изобретения имеет право.
--->
Перечень последовательностей
<110> CJ CheilJedang Corporation
<120> Novel promoter and uses thereof
<130> OPA19010
<150> KR 10-2018-0032253
<151> 2018-03-20
<160> 9
<170> KoPatentIn 3.0
<210> 1
<211> 307
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор cj2.2
<400> 1
caccgcgggc ttattccatt acatggaatg accaggaatg gcagggaatg cgacgaaatt 60
gactgtgtcg ggagcttctg atccgatgct gccaaccagg agagaaaata atgacatgtg 120
caggcacgct ggtgagctgg agatttatga tctcaagtac cttttttctt gcactcgagg 180
gggctgagtg ccagaatggt tgctgacacc aggttgaggt tggtacacac tcaccaatcc 240
tgccgtcgcg ggcgcctgcg tggaacataa accttgagtg aaaccaaatc taggagatta 300
agatatc 307
<210> 2
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер для F_ПЦР с ошибками
<400> 2
ttgcatgcct gcacaccgcg ggctta 26
<210> 3
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер для R_ПЦР с ошибками
<400> 3
agtgaattcg agctcggtac ccgggg 26
<210> 4
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> cj2.2 сайт-направленный F_праймер
,
<400> 4
ataaaccttg agtgaaacca aatctaggag attaa 35
<210> 5
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> cj2.2 сайт-направленный R_праймер
<400> 5
ttaatctcct agatttggtt tcactcaagg tttat 35
<210> 6
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> metZ F_праймер
<400> 6
aattgtcgac atgggtaacg cgtttcgtga ag 32
<210> 7
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> metZ R_праймер
<400> 7
aattggatcc tcagatcacc gcgagcgc 28
<210> 8
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> OPSS F_праймер
<400> 8
aattgtcgac atgacgcgct acgactcc 28
<210> 9
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> OPSS R_праймер
<400> 9
aattggatcc ttattccagc gcgtcctc 28
<---
Группа изобретений относится к полинуклеотиду, имеющему промоторную активность, а также его использованию для получения целевого вещества. Предложен полинуклеотид, имеющий промоторную активность и содержащий полинуклеотидную последовательность SEQ ID NO: 1. Также предложены экспрессионный вектор, содержащий указанный полинуклеотид, клетка-хозяин, которая содержит указанный полинуклеотид или в которую введен экспрессионный вектор. Также предложен способ получения целевого вещества, включающий культивирование клетки-хозяина, содержащей указанный полинуклеотид, функционально связанный с геном, кодирующим целевой белок, и извлечение целевого вещества из клетки-хозяина или из культуральной среды, в которой культивируют клетку-хозяина. Группа изобретений обеспечивает повышение уровня экспрессии целевого гена, функционально связанного с полинуклеотидом в нужной клетке-хозяине, а также уровня экспрессии и активности белка, кодируемого целевым геном. 4 н. и 2 з.п. ф-лы, 3 ил., 6 табл., 5 пр.
1. Полинуклеотид, имеющий промоторную активность, содержащий полинуклеотидную последовательность SEQ ID NO: 1.
2. Экспрессионный вектор, содержащий полинуклеотид по п. 1, имеющий промоторную активность.
3. Экспрессионный вектор по п. 2, содержащий ген, кодирующий целевой белок.
4. Клетка-хозяин, которая содержит полинуклеотид по п. 1 или в которую введен экспрессионный вектор по п. 2, для получения целевого продукта.
5. Клетка-хозяин по п. 4, которая представляет собой бактериальную клетку, принадлежащую роду Corynebacterium или роду Escherichia.
6. Способ получения целевого вещества, включающий:
культивирование клетки-хозяина, содержащей полинуклеотид по п.1, имеющий промоторную активность, функционально связанный с геном, кодирующим целевой белок, и извлечение целевого вещества из клетки-хозяина или из культуральной среды, в которой культивируют клетку-хозяина.
| KR 1020060068505 A, 21.06.2006 | |||
| ЭКЗОТЕРМИЧЕСКИЙ СТЕРЖЕНЬ-РЕЗАК | 1997 |
|
RU2169065C2 |
| KR 100725336 B1, 07.06.2007 | |||
| EUN PAIK J., RAK LEE B | |||
| Isolation of transcription initiation signals from Corynebacterium ammoniagenes and comparison of their gene expression levels in C | |||
| ammoniagenes and Escherichia coli | |||
| Видоизменение пишущей машины для тюркско-арабского шрифта | 1923 |
|
SU25A1 |

