Область техники
Настоящее изобретение относится к области техники генной инженерии и микроорганизмов и, в частности, относится к рекомбинантному штамму для продуцирования (получения) L-глутаминовой кислоты, а также способу его конструирования и применения.
Уровень техники
L-глутаминовая кислота представляет собой важную аминокислоту, которую используют в пищевых продуктах, в клинической медицине и в других областях.
Традиционно L-глутаминовую кислоту получают в основном путем ферментации с использованием продуцирующих L-глутаминовую кислоту бактерий, принадлежащих к роду Brevibactehum, Corynebacterium или Microtatobiotes, или их мутантов.
L-глутаминовая кислота синтезируется путем биосинтеза из α-кетоглутаровой кислоты, промежуточного продукта цикла лимонной кислоты в клетках микроорганизмов. Существует два биосинтетических пути образования L-глутаминовой кислоты из α-кетоглутаровой кислоты через ассимиляцию ионов аммония. Один путь представлен синтезом L-глутаминовой кислоты путем катализа глутаматдегидрогеназой (GDH) в присутствии ионов аммония в высокой концентрации. Другой путь (путь GS/GOGAT) представлен синтезом L-глутаминовой кислоты глутаминсинтетазой и аминотрансферазой глутамина-оксоглутаровой кислоты. Глутаминсинтетаза (GS) катализирует конверсию L-глутаминовой кислоты и ионов аммония в глутамин; аминотрансфераза глутамина-оксоглутаровой кислоты, также известная как «глутаминсинтетаза» (GOGAT), катализирует синтез L-глутаминовой кислоты, в ходе которого две молекулы L-глутаминовой кислоты синтезируются из одной молекулы глутамина, которая была синтезирована GS из одной молекулы α-кетоглутаровой кислоты.
Улучшение получения L-аминокислот путем ферментации может относиться к методикам ферментации, таким как перемешивание и подача кислорода; или к составу питательной среды, например, к концентрации сахара во время ферментации; или к переработке ферментативного бульона в продукт в подходящей форме, например, путем высушивания и грануляции ферментативного бульона, или с помощью ионообменной хроматографии; или может относиться к собственным свойствам соответствующих продуцирующих микроорганизмов.
Способы улучшения свойств указанных продуцирующих микроорганизмов, включают мутагенез, выбор и скрининг мутантов. Штамм, полученный указанным способом, устойчив к антиметаболитам или ауксотрофен по метаболитам, имеющим регуляторное значение, и продуцирует L-аминокислоты, и т.п.
Хотя существует значительное число способов, способных усиливать способность к продуцированию L-глутаминовой кислоты, сохраняется потребность в разработке способов продуцирования L-глутаминовой кислоты для удовлетворения растущего спроса.
Краткое описание изобретения
Задача настоящего изобретения заключается в разработке новой методики для усиления способности бактерии к продуцированию L-глутаминовой кислоты, с обеспечением таким образом способа эффективного продуцирования L-глутаминовой кислоты.
В ходе исследований для решения вышеуказанной задачи авторы настоящего изобретения обнаружили, что ген BBD29_09525 или гомологичный ему ген бактерии может быть модифицирован или улучшен в отношении экспрессии указанного гена для обеспечения усиленной способности бактерии к продуцированию L-глутаминовой кислоты. Настоящее изобретение было выполнено с учетом этих находок.
Согласно настоящему изобретению предложена бактерия для выработки L-глутаминовой кислоты с улучшенной экспрессией полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 3 или гомологичную ей последовательность. Согласно настоящему изобретению также предложен способ получения L-глутаминовой кислоты с использованием микроорганизма.
Согласно первому аспекту настоящего изобретения предложена бактерия для продуцирования L-глутаминовой кислоты, обладающая улучшенной экспрессией полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 3 или гомологичную ей последовательность. В соответствии с настоящим изобретением улучшенная экспрессия представляет собой усиленную экспрессию полинуклеотида, или присутствие точечной мутации в полинуклеотиде, который кодирует аминокислотную последовательность SEQ ID NO: 3 или гомологичную ей последовательность, или присутствие точечной мутации в полинуклеотиде и усиленную экспрессию полинуклеотида, который кодирует аминокислотную последовательность SEQ ID NO: 3 или гомологичную ей последовательность.
Аминокислотная последовательность SEQ ID NO: 3 или гомологичная ей последовательность представляет собой белок, кодируемый геном BBD29_09525 или гомологичным ему геном.
Указанная бактерия имеет усиленную способность к продуцированию L-глутаминовой кислоты по сравнению с немодифицированным штаммом.
Согласно настоящему изобретению термин «бактерия, обладающая способностью к продуцированию L-глутаминовой кислоты» относится к бактерии, обладающей такой способностью к продуцированию и накоплению целевой L-глутаминовой кислоты в культуральной среде и/или в клетке указанной бактерии, что L-глутаминовая кислота может быть собрана при культивировании бактерии в культуральной среде. Бактерия, обладающая способностью к продуцированию L-глутаминовой кислоты, может представлять собой бактерию, способную к накоплению целевой L-глутаминовой кислоты в культуральной среде и/или в клетке указанной бактерии в большем количестве, чем у немодифицированного штамма.
Термин «немодифицированный штамм» относится к контрольному штамму, который не был модифицирован таким образом, чтобы включать некий специфический признак. Таким образом, примеры немодифицированного штамма включают штамм дикого типа и исходный штамм.
Бактерия, обладающая способностью к продуцированию L-глутаминовой кислоты, может представлять собой бактерию, способную к накоплению целевой L-глутаминовой кислоты в культуральной среде в количестве, предпочтительно превышающем 0,5 г/л, и более предпочтительно превышающем 1,0 г/л.
Согласно настоящему изобретению термин «L-глутаминовая кислота» относится к L-глутаминовой кислоте в свободной форме, к ее соли или смеси, если не указано иное.
Указанный полинуклеотид может кодировать аминокислотную последовательность, обладающую приблизительно 90% или более, приблизительно 92% или более, приблизительно 95% или более, приблизительно 97% или более, приблизительно 98% или более, или приблизительно 99% или более гомологией последовательности с аминокислотной последовательностью SEQ ID NO: 3. В настоящем документе термин «гомология» относится к проценту идентичности двух полинуклеотидов или двух полипептидов как модулей. Гомология последовательностей может быть измерена между одним модулем и другим модулем с применением способа, известного в данной области техники. Например, такая гомология последовательностей может быть измерена с помощью алгоритма BLAST.
Экспрессия полинуклеотида может быть усилена: путем замены или мутирования регуляторной последовательности экспрессии, введения мутации в последовательность полинуклеотида, и путем увеличения числа копий полинуклеотида, инсертированных в хромосому или введенных с помощью вектора, или комбинации перечисленного, и т.п.
Регуляторная последовательность экспрессии полинуклеотида может быть модифицирована. Регуляторная последовательность экспрессии контролирует экспрессию полинуклеотида, который функционально с ней связан, и может содержать, например, промоторы, терминаторы, энхансеры, сайленсеры и т.п. Полинуклеотид может иметь измененный инициирующий кодон. Полинуклеотид может быть включен в определенный сайт хромосомы, с увеличением таким образом числа копий. Согласно настоящему документу определенный сайт может включать, например, сайт транспозона или межгенный сайт. Кроме того, полинуклеотид может быть включен в экспрессионный вектор, а указанный экспрессионный вектор введен в клетку-хозяина, с увеличением таким образом числа копий.
Согласно одному из вариантов реализации настоящего изобретения полинуклеотид или имеющий точечную мутацию полинуклеотид включают в определенный сайт хромосомы микроорганизма с увеличением таким образом числа копий.
Согласно одному из вариантов реализации настоящего изобретения полинуклеотид с последовательностью промотора, или имеющий точечную мутацию полинуклеотид с последовательностью промотора встраивают в определенный сайт хромосомы микроорганизма, со сверхэкспрессией таким образом последовательности нуклеиновой кислоты.
Согласно одному из вариантов реализации настоящего изобретения полинуклеотид или имеющий точечную мутацию полинуклеотид включают в экспрессионный вектор, а указанный экспрессионный вектор вводят в клетку-хозяина, с увеличением таким образом числа копий.
Согласно одному из вариантов реализации настоящего изобретения полинуклеотид с последовательностью промотора, или имеющий точечную мутацию полинуклеотид с последовательностью промотора включают в экспрессионный вектор, а указанный экспрессионный вектор вводят в клетку-хозяина, с обеспечением таким образом сверхэкспрессии последовательности аминокислот.
Согласно конкретному варианту реализации настоящего изобретения указанный полинуклеотид может содержать последовательность нуклеотидов SEQ ID NO: 1.
Согласно одному из вариантов реализации настоящего изобретения полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 3, имеет точечную мутацию, такую, что пролин в положении 113 в последовательности аминокислот SEQ ID NO: 3, заменен на другую аминокислоту.
В соответствии с настоящим изобретением предпочтительно пролин в положении 113 заменен на серин.
В соответствии с настоящим изобретением последовательность, представленная в SEQ ID NO: 4, представляет собой последовательность аминокислот после замены пролина на серин в положении 113 в последовательности аминокислот, представленной в SEQ ID NO: 3.
Согласно одному из вариантов реализации настоящего изобретения последовательность полинуклеотида, имеющего точечную мутацию, образуется в результате мутации основания в положении 337 последовательности полинуклеотида, представленной в SEQIDNO: 1.
Согласно настоящему изобретению указанная мутация включает мутацию основания в положении 337 последовательности полинуклеотида, представленной в SEQ ID NO: 1, с заменой цитозина (С) на тимин (Т).
Согласно одному из вариантов реализации настоящего изобретения последовательность полинуклеотида, имеющего точечную мутацию, содержит последовательность полинуклеотида, представленную в SEQ ID NO: 2.
В настоящем документе термин «функционально связанный» относится к функциональной связи между регуляторной последовательностью и последовательностью полинуклеотида, за счет которой регуляторная последовательность контролирует транскрипцию и/или трансляцию указанной последовательности полинуклеотида. Регуляторная последовательность может представлять собой сильный промотор, который может повышать уровень экспрессии полинуклеотида. Регуляторная последовательность может представлять собой промотор, происходящий из микроорганизма, принадлежащего к роду Corynebacterium, или может представлять собой промотор, происходящий из других микроорганизмов. Например, указанный промотор может представлять собой промотор trc, промотор gap, промотор tac, промотор Т7, промотор lac, промотор trp, промотор araBAD или промотор cj7.
Согласно конкретному варианту реализации настоящего изобретения промотор представляет собой промотор полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 3 (ген BBD29_09525).
В настоящем документе термин «вектор» относится к полинуклеотидной конструкции, содержащей регуляторную последовательность гена и последовательность гена, и выполненной с возможностью экспрессии целевого гена в подходящей клетке-хозяине. Термин "вектор" может также относиться к полинуклеотидной конструкции, содержащей последовательность для гомологичной рекомбинации, такой, что при введении этого вектора в клетку-хозяина может быть изменена регуляторная последовательность эндогенного гена в геноме клетки-хозяина, или целевой ген, который может экспрессироваться, может быть инсертирован в определенный сайт генома хозяина. Таким образом, вектор для применения по настоящему изобретению может дополнительно содержать селективный маркер для определения введения вектора в клетку-хозяина или инсерции вектора в хромосому клетки-хозяина. Селективный маркер может включать маркер, обеспечивающий селектируемый фенотип, такой как лекарственная устойчивость, ауксотрофность, устойчивость к цитотоксическим агентам или экспрессия поверхностного белка. На среде, обработанной таким селективным агентом, может быть выбрана трансформированная клетка, поскольку только клетка, которая экспрессирует селективный маркер, может выживать или проявлять иные фенотипические черты. Вектор согласно описанию в настоящем документе хорошо известен специалистам в данной области техники и включает в том числе, но не ограничиваясь перечисленным: плазмиду, бактериофаг (такой как λ бактериофаг или нитевидный бактериофаг М13, и т.п.), космиду или вирусный вектор.
Согласно некоторым конкретным вариантам реализации настоящего изобретения в качестве вектора используют плазмиду pK18mobsacB и плазмиду pXMJ19.
В настоящем документе термин «трансформация» относится к введению полинуклеотида в клетку-хозяина таким образом, чтобы указанный полинуклеотид мог быть репликативно-компетентным в качестве элемента, чужеродного для генома, или за счет встраивания в геном клетки-хозяина. Способ трансформации вектором, применяемый согласно настоящему изобретению, может включать способ введения нуклеиновой кислоты в клетку. Кроме того, в соответствии с описанием близких методик, может быть использован подходящий для клетки-хозяина способ, задействующий электрические импульсы.
Согласно настоящему изобретению микроорганизм может представлять собой дрожжи, бактерию, водоросль или гриб.
В соответствии с настоящим изобретением указанная бактерия может представлять собой микроорганизм, принадлежащий к роду Corynebacterium, такой как Corynebacterium acetoacidophilum, Corynebacterium acetoglutamicum, Corynebacterium callunae, Corynebacterium glutamicum, Brevibacterium flavum, Brevibacterium lactofermentum, Corynebacterium ammoniagenes, Corynebacterium pekinense, Brevibacterium saccharolyticum, Brevibacterium roseum и Brevibacterium thiogenitalis и т.п.
Согласно одному из вариантов реализации настоящего изобретения указанный микроорганизм, принадлежащий к роду Corynebacterium, представляет собой Corynebacterium glutamicum АТСС 13869.
Согласно одному из вариантов реализации настоящего изобретения указанный микроорганизм, принадлежащий к роду Corynebacterium, представляет собой Corynebacterium glutamicum YPGLU001, который обеспечивает высокие уровни продуцирования глутаминовой кислоты, и депонирован как штамм с названием: Corynebacterium glutamicum; латинское название: Corynebacterium glutamicum; штамм №: YPGLU001 в Китайском главном центре коллекций микробиологических культур («China General Microbiological Culture Collection Center»), сокращенно «CGMCC», адрес: No. 3, Yard 1, Beichen West Road, Chaoyang District, Beijing, 23 ноября 2020 г., под номером доступа: CGMCC №21220.
В соответствии с настоящим изобретением указанная бактерия может также иметь дополнительные улучшения, связанные с повышением уровней продуцирования L-глутаминовой кислоты, такие как усиление или снижение активности, или экспрессии генов глутаматдегидрогеназы, глутаминсинтетазы, аминотрансферазы глутамина оксоглутаровой кислоты и т.п., или замена генов чужеродными генами.
Согласно второму аспекту настоящего изобретения предложены последовательность полинуклеотида (полинуклеотидная последовательность), аминокислотная последовательность, кодируемая указанной последовательностью полинуклеотида, рекомбинантный вектор, содержащий указанную последовательность полинуклеотида, и рекомбинантный штамм, содержащий указанную последовательность полинуклеотида.
В соответствии с настоящим изобретением указанная последовательность полинуклеотида включает полинуклеотид, кодирующий полипептид, содержащий аминокислотную последовательность SEQ ID NO: 3, где пролин в положении 113 в последовательности заменен на другую аминокислоту.
В соответствии с настоящим изобретением предпочтительно пролин в положении 113 заменен на серин.
В соответствии с настоящим изобретением, последовательность, представленная в SEQ ID NO: 4, представляет собой последовательность аминокислот после замены пролина на серин в положении 113 в последовательности аминокислот, представленной в SEQ ID NO: 3.
В соответствии с настоящим изобретением предпочтительно указанный полинуклеотид, кодирующий полипептид, содержащий аминокислотную последовательность SEQ ID NO: 3, содержит последовательность полинуклеотида, представленную в SEQ ID NO: 1.
Согласно одному из вариантов реализации настоящего изобретения указанная последовательность полинуклеотида образуется в результате мутации основания в положении 337 в последовательности полинуклеотида, представленной в SEQ ID NO: 1.
В соответствии с настоящим изобретением мутация относится к изменению основания/нуклеотида в сайте, и способ осуществления мутации может представлять собой по меньшей мере один способ, выбранный из мутагенеза, сайт-направленного ПЦР-мутагенеза и/или гомологичной рекомбинации, и т.п. Согласно настоящему изобретению предпочтительно используют сайт-направленный ПЦР-мутагенез и/или гомологичную рекомбинацию.
Согласно настоящему изобретению мутация включает мутацию основания в положении 337 в последовательности полинуклеотида, представленной в SEQ ID NO: 1, с заменой цитозина (С) на тимин (Т).
Согласно одному из вариантов реализации настоящего изобретения последовательность полинуклеотида содержит последовательность полинуклеотида SEQ ID NO: 2.
В соответствии с настоящим изобретением последовательность аминокислот содержит последовательность аминокислот SEQ ID NO: 4.
В соответствии с настоящим изобретением рекомбинантный вектор конструируют путем введения последовательности полинуклеотида в плазмиду.
Согласно одному из вариантов реализации настоящего изобретения плазмида представляет собой плазмиду pK18mobsacB.
Согласно другому варианту реализации настоящего изобретения плазмида представляет собой плазмиду pXMJ19.
В частности, указанные последовательность полинуклеотида и плазмида могут быть сконструированы в виде рекомбинантного вектора с помощью системы рекомбинации NEBuider.
В соответствии с настоящим изобретением указанный рекомбинантный штамм содержит указанную последовательность полинуклеотида.
Во варианте реализации настоящего изобретения исходный штамм для рекомбинантного штамма представляет собой Corynebacterium glutamicum CGMCC №21220.
В одном из вариантом реализации настоящего изобретения исходный штамм для рекомбинантного штамма представляет собой АТСС 13869.
Согласно третьему аспекту настоящего изобретения также предложен способ конструирования рекомбинантного штамма, продуцирующего L-глутаминовую кислоту.
В соответствии с настоящим изобретением способ конструирования включает следующий этап:
модификацию последовательности полинуклеотида гена BBD29_09525 дикого типа, представленной в SEQ ID NO: 1, у штамма-хозяина, чтобы вызвать мутацию основания в положении 337 с получением рекомбинантного штамма, содержащего кодирующий мутированный BBD29_09525 ген.
В соответствии со способом конструирования согласно настоящему изобретению конструирование включает по меньшей мере что-либо одно из мутагенеза, сайт-направленного ПЦР-мутагенеза и/или гомологичной рекомбинации, и т.п.
В соответствии со способом конструирования согласно настоящему изобретению мутация относится к мутации основания в положении 337 в SEQ ID NO: 1 с заменой цитозина (С) на тимин (Т). В частности, последовательность полинуклеотида, содержащая кодирующий мутированный BBD29_09525 ген, представлена в SEQ ID NO: 2.
Кроме того, способ конструирования включает следующие этапы:
(1) модификацию последовательности нуклеотидов гена BBD29_09525 дикого типа, представленной в SEQ ID NO: 1, чтобы вызвать мутацию основания в положении 337, с получением последовательности полинуклеотида мутированного гена BBD29_09525;
(2) соединение мутированной последовательности полинуклеотида с плазмидой для конструирования рекомбинантного вектора;
(3) введение указанного рекомбинантного вектора в штамм-хозяин с получением рекомбинантного штамма, содержащего мутированный кодирующий BBD29_09525 ген.
В соответствии со способом конструирования согласно настоящему изобретению указанный этап (1) включает конструирование гена BBD29_09525 с точечной мутацией: в соответствии с геномной последовательностью немодифицированного штамма синтез двух пар праймеров P1, Р2 и Р3, Р4 для амплификации фрагмента гена BBD29_09525, и введение точечной мутации в ген BBD29_09525 дикого типа, SEQ ID NO: 1, с применением сайт-направленного ПЦР-мутагенеза с получением последовательности нуклеотидов гена BBD29_09525 с точечной мутацией, SEQ ID NO: 2, обозначенного как BBD29_09525 С337Т.
Согласно одному из вариантов реализации настоящего изобретения геном немодифицированного штамма может происходить из штамма АТСС13869, геномную последовательность которого можно найти на вебсайте NCBI.
Согласно одному из вариантов реализации настоящего изобретения на этапе (1) используют следующие праймеры:
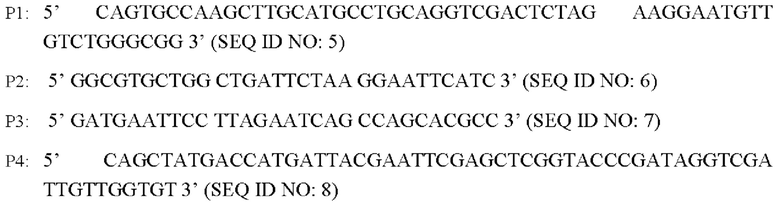
Согласно одному из вариантов реализации настоящего изобретения ПЦР-амплификацию осуществляют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 с при 94°С, ренатурация в течение 30 с при 52°С и удлинение в течение 45 с при 72°С в ходе 30 циклов, и избыточное удлинение в течение 10 мин при 72°С.
Согласно одному из вариантов реализации настоящего изобретения ПЦР-амплификацию с перекрывающимися праймерами осуществляют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 с при 94°С, ренатурация в течение 30 с при 52°С и удлинение в течение 90 с при 72°С в ходе 30 циклов, и избыточное удлинение в течение 10 мин при 72°С.
В соответствии со способом конструирования согласно настоящему изобретению этап
(2) включает конструирование рекомбинантной плазмиды, включающей сборку выделенного и очищенного BBD29_09525 с плазмидой pKlSmobsacB с помощью системы рекомбинации NEBuider с получением рекомбинантной плазмиды.
В соответствии со способом конструирования согласно настоящему изобретению этап
(3) включает конструирование рекомбинантного штамма путем трансформации рекомбинантной плазмидой штамма-хозяина с получением рекомбинантного штамма.
Согласно одному из вариантов реализации настоящего изобретения трансформацию на этапе (3) осуществляют методом электротрансформации.
Согласно одному из вариантов реализации настоящего изобретения штамм-хозяин представляет собой АТСС 13869.
Согласно одному из вариантов реализации настоящего изобретения штамм-хозяин представляет собой Corynebacterium glutamicum CGMCC №21220.
Согласно одному из вариантов реализации настоящего изобретения указанная рекомбинация достигается путем гомологичной рекомбинации.
Согласно четвертому аспекту настоящего изобретения также предложен способ конструирования рекомбинантного штамма, продуцирующего L-глутаминовую кислоту.
В соответствии с настоящим изобретением способ конструирования включает следующие этапы:
амплификация 5'- и 3'-фрагментов плеч гомологии BBD29_09525 и последовательности кодирующей области гена BBD29_09525 с областью его промотора, и введение гена BBD29_09525 или BBD29_09525C337T в геном штамма-хозяина путем гомологичной рекомбинации, что позволит указанному штамму сверхэкспрессировать ген BBD29_09525 или BBD29_09525C337T.
Согласно одному из вариантов реализации настоящего изобретения используют следующие праймеры для амплификации 5'-фрагментов плеч гомологии:

Согласно одному из вариантов реализации настоящего изобретения используют следующие праймеры для амплификации 3'-фрагментов плеча гомологии:
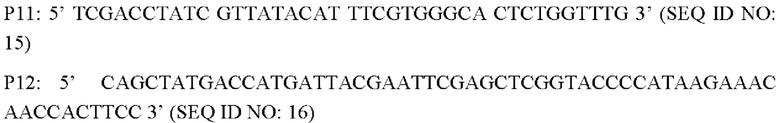
Согласно одному из вариантов реализации настоящего изобретения используют следующие праймеры для амплификации последовательности кодирующей области гена и области его промотора:
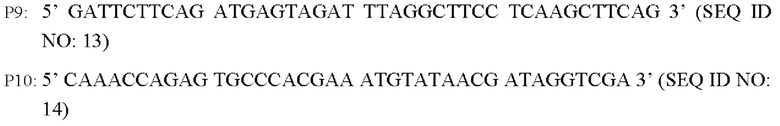
Согласно одному из вариантов реализации настоящего изобретения, опять же с вышеупомянутыми Р7/Р12 в качестве праймеров, осуществляют амплификацию с применением в качестве матриц смеси трех фрагментов, амплифицированного 5'-фрагмента плеча гомологии, 3'-фрагмента плеча гомологии и фрагмента последовательности кодирующей области гена и области его промотора, с получением единого фрагмента плеча гомологии.
Согласно одному из вариантов реализации настоящего изобретения используемая ПЦР-система содержит: 10 × буфер Ex Taq Buffer: 5 мкл, смесь dNTP (2,5 мМ каждый): 4 мкл, Mg2+(25 мМ): 4 мкл, праймеры (10 пМ): каждого по 2 мкл; и Ex Taq (5 Ед/мкл): 0,25 мкл, в общем объеме 50 мкл. ПЦР-амплификацию осуществляют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 с при 94°С, ренатурация в течение 30 с при 52°С, удлинение в течение 120 с при 72°С (30 циклов) и избыточное удлинение в течение 10 мин при 72°С.
Согласно одному из вариантов реализации настоящего изобретения используют систему рекомбинации NEBuider для сборки челночной плазмиды PK18mobsacB с единым фрагментом плеча гомологии для получения интегративной плазмиды.
Согласно одному из вариантов реализации настоящего изобретения штамма-хозяин трансфицируют указанной интегративной плазмидой для введения гена BBD29_09525 или BBD29_09525C337T в геном штамма-хозяина путем гомологичной рекомбинации.
Согласно одному из вариантов реализации настоящего изобретения штамм-хозяин представляет собой Corynebacterium glutamicum CGMCC №21220.
Согласно одному из вариантов реализации настоящего изобретения штамм-хозяин представляет собой АТСС 13869.
Согласно одному из вариантов реализации настоящего изобретения указанный штамм-хозяин представляет собой штамм, несущий последовательность полинуклеотида, представленную в SEQ ID NO: 2.
Согласно пятому аспекту настоящего изобретения также предложен способ конструирования рекомбинантного штамма, продуцирующего L-глутаминовую кислоту.
В соответствии с настоящим изобретением способ конструирования включает следующие этапы:
амплификация последовательности кодирующей области гена BBD29_09525 и области промотора, или последовательности кодирующей области гена BBD29_09525C337T и области промотора, конструирование сверхэкспрессирующего плазмидного вектора и трансформация указанным вектором штамма-хозяина, что позволяет указанному штамму сверхэкспрессировать ген BBD29_09525 или BBD29_09525 С337Т.
Согласно одному из вариантов реализации настоящего изобретения используют следующие праймеры для амплификации последовательности кодирующей области гена и области его промотора:
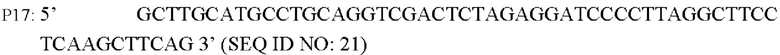
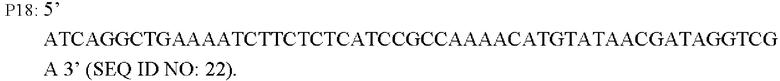
Согласно одному из вариантов реализации настоящего изобретения указанная ПЦР-система содержит: 10 × буфер Ex Taq Buffer: 5 мкл, смесь dNTP (2,5 мМ каждый): 4 мкл, Mg2+(25 мМ): 4 мкл, праймеры (10 пМ): по 2 мкл каждого, Ex Taq (5 Ед/мкл): 0,25 мкл, в общем объеме 50 мкл. ПЦР-амплификацию осуществляют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 с при 94°С, ренатурация в течение 30 с при 52°С, удлинение в течение 120 с при 72°С (30 циклов) и избыточное удлинение в течение 10 мин при 72°С.
Согласно одному из вариантов реализации настоящего изобретения систему рекомбинации NEBuider применяют для сборки челночной плазмиды pXMJ19 с фрагментом BBD29_09525 или BBD29_09525С337Т, имеющим собственные промоторы, с получением сверхэкспрессирующей плазмиды.
Согласно одному из вариантов реализации настоящего изобретения штамм-хозяин представляет собой Corynebacterium glutamicum CGMCC №21220.
Согласно одному из вариантов реализации настоящего изобретения штамм-хозяин представляет собой АТСС 13869.
Согласно одному из вариантов реализации настоящего изобретения указанный штамм-хозяин представляет собой штамм, несущий последовательность полинуклеотида, представленную в SEQ ID NO: 2.
Рекомбинантный штамм, предложенный согласно настоящему изобретению, может индивидуально применяться при ферментации для продуцирования L-глутаминовой кислоты, и может также применяться для продуцирования L-глутаминовой кислоты при гибридной ферментации с другими продуцирующими L-глутаминовую кислоту бактериями.
Согласно другому аспекту настоящего изобретения предложен способ получения L-глутаминовой кислоты, который включает культивирование бактерии; и получение L-глутаминовой кислоты из культуры.
Бактерия может быть культивирована в подходящей культуральной среде в условиях культивирования, известных в данной области техники. Культуральная среда может содержать: источник углерода, источник азота, следовые элементы и комбинацию перечисленного. При культивировании могут корректироваться значения рН культуры. Кроме того, может предотвращаться образование пузырьков при культивировании, например, с применением пеногасителя для предотвращения образования пузырьков. Кроме того, в культуру могут вводиться газы при культивировании. Указанные газы могут включать любые газы, способные поддерживать аэробные условия в культуре. При культивировании температура культуры может составлять от 20°С до 45°С. Вырабатываемая L-глутаминовая кислота может быть выделена из культуры, т.е. культуру обрабатывают серной кислотой или соляной кислотой и т.п., с последующим применением комбинации таких способов, как анионообменная хроматография, конденсация, кристаллизация и изоэлектрическое осаждение.
Согласно настоящему изобретению:
SEQ ID NO: 1: последовательность ORF BBD29_09525 дикого типа


SEQ ID NO: 3: последовательность аминокислот белка, кодируемого BBD29_09525 дикого типа

SEQ ID NO: 4: последовательность аминокислот белка, кодируемого BBD29_09525P113S

BBD29_09525P113S представляет собой BBD29_09525 P113S.
Согласно настоящему изобретению также предложен белок, обозначенный как белок BBD29_09525P113S, отличающийся тем, что указанный белок может представлять собой любое из:
А1) белка, имеющего аминокислотную последовательность SEQ ID NO: 4;
А2) белка, обладающего 80% или более идентичностью белку, указанному в А1), и имеющего ту же функцию, что и белок, указанный в А1), полученного путем осуществления в последовательности аминокислот, представленной в SEQ ID NO: 4, замены, и/или делеции, и/или добавления остатков аминокислот;
A3) слитого белка, имеющего туже функцию, полученного путем присоединения метки к N-концу и/или С-концу А1) или А2).
Согласно настоящему изобретению также предложена молекула нуклеиновой кислоты, обозначенная как BBD29_09525, причем указанная молекула нуклеиновой кислоты BBD29_09525C337T может представлять собой любое из:
B1) молекулы нуклеиновой кислоты, кодирующей белок BBD29_09525P113S;
B2) молекулы ДНК, кодирующая последовательность которой представлена в SEQ ID NO: 2;
B3) молекулы ДНК, нуклеотидная последовательность которой представлена в SEQ ID NO: 2.
Молекула ДНК, представленная в SEQ ID NO: 2, представляет собой ген BBD29-09525C337T согласно настоящему изобретению.
Молекула ДНК, представленная в SEQ ID NO: 2 (ген BBD29_09525С337Т), кодирует белок BBD29_09525 P113S, представленный в SEQ ID NO: 4.
Присутствием серина (S) в положении 113 в последовательности аминокислот белка BBD29_09525 P113S (SEQ ID NO: 4) обусловлено мутацией пролина (Р).
Согласно настоящему изобретению также предложен биоматериал, где указанный биоматериал может представлять собой любое из:
B1) экспрессионной кассеты, содержащей молекулу нуклеиновой кислоты BBD29_09525C337T;
B2) рекомбинантного вектора, содержащего молекулу нуклеиновой кислоты BBD29_09525С337Т, или рекомбинантного вектора, содержащего экспрессионную кассету по С1);
B3) рекомбинантного микроорганизма, содержащего молекулу нуклеиновой кислоты BBD29_09525С337Т, или рекомбинантного микроорганизма, содержащего экспрессионную кассету по С1), или рекомбинантного микроорганизма, содержащего рекомбинантный вектор по С2).
Согласно настоящему изобретению также предложено любое из следующих применений любого из Dl) - D8):
F1) применение любого из D1) D8) для регуляции продуцирования L-глутаминовой кислоты у микроорганизма;
F2) применение любого из D1) - D8) для конструирования генетически сконструированной (модифицированной) бактерии для продуцирования L-глутаминовой кислоты;
F3) применение любого из D1) - D8) для продуцирования L-глутаминовой кислоты; где Dl) D8) представляет собой: D1) белок BBD29_09525P113S;
D2) молекулу нуклеиновой кислоты BBD29_095250337; D3) биоматериал;
D4) молекулу ДНК, последовательность нуклеотидов которой представляет собой SEQ ID NO: 1;
D5) молекулу ДНК, обладающую идентичностью 90% или более с молекулой ДНК, представленной в SEQ ID NO: 1, и имеющую ту же функцию, что и указанная молекула ДНК, полученную путем осуществления в последовательности нуклеотидов, представленной в SEQ ID NO: 1, модификации, и/или замены, и/или делеции, и/или добавления одного или нескольких нуклеотидов;
D6) экспрессионную кассету, содержащую молекулу ДНК по D4) или D5);
D7) рекомбинантный вектор, содержащий молекулу ДНК по D4) или D5), или рекомбинантный вектор, содержащий экспрессионную кассету по D6);
D8) рекомбинантный микроорганизм, содержащий молекулу ДНК по D4) или D5), или рекомбинантный микроорганизм, содержащий экспрессионную кассету по D6), или рекомбинантный микроорганизм, содержащий рекомбинантный вектор по D7).
Молекула ДНК, представленная в SEQ ID NO: 1, представляет собой ген BBD29_09525 согласно настоящему изобретению.
Молекула ДНК, представленная в SEQ ID NO: 1 (ген BBD29_09525), кодирует белок, представленный в SEQ ID NO: 3.
В настоящем документе «идентичность» относится к идентичности последовательностей аминокислот или последовательностей нуклеотидов. Международные сайты поиска гомологии в сети Интернет могут применяться для определения идентичности последовательностей аминокислот, например, страница BLAST на главной странице вебсайта NCBI. Например, в Advanced BLAST2.1 может быть использована программа blastp со значением параметра Expect value, установленным на 10, всеми фильтрами в режиме OFF, BLOSUM62 в качестве матрицы и значениями штрафа за введение пропуска, штрафа за удлинение пропуска на один остаток и коэффициента лямбда (Lambda ratio), установленными на 11, 1 и 0,85 (устанавливаемые по умолчанию значения), соответственно, и выполнен поиск идентичности между парой последовательностей аминокислот для расчета, после чего можно получить значение идентичности (%).
Согласно настоящему документу идентичность 80% или более может представлять собой по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность.
Согласно настоящему документу идентичность 90% или более может представлять собой по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность.
Регуляция продуцирования L-глутаминовой кислоты у микроорганизма согласно описанию в настоящем документе может представлять собой повышение или понижение уровня накопления L-глутаминовой кислоты у микроорганизма (т.е. стимуляцию или ингибирование биосинтеза L-глутаминовой кислоты).
Согласно настоящему изобретению также предложен способ повышения продуцирования L-глутаминовой кислоты у микроорганизма, при этом указанный способ включает любое из:
Е1) повышения уровня экспрессии или содержания молекулы нуклеиновой кислоты BBD29_09525C337T в целевом микроорганизме с получением микроорганизма, продуцирующего больше L-глутаминовой кислоты, чем целевой микроорганизм;
Е2) повышения уровня экспрессии или содержания молекулы ДНК по D4) или D5) в целевом микроорганизме с получением микроорганизма, продуцирующего больше L-глутаминовой кислоты, чем целевой микроорганизм;
Е3) осуществления мутации в молекуле ДНК, последовательность нуклеотидов которой представляет собой SEQ ID NO: 1, в целевом микроорганизме с получением микроорганизма, продуцирующего больше L-глутаминовой кислоты, чем целевой микроорганизм.
Согласно описанному выше способу указанная мутация может представлять собой точечную мутацию, т.е. мутацию одного нуклеотида.
В описанном выше способе точечная мутация может представлять собой мутацию остатка пролина в положении 113 в последовательности аминокислот, кодируемой молекулой ДНК, представленной в SEQ ID NO: 1, на остаток другой аминокислоты.
В описанном выше способе точечная мутация может представлять собой мутацию пролина в положении 113 в последовательности аминокислот, кодируемой молекулой ДНК, представленной в SEQ ID NO: 1, на серин, что обеспечивает получение мутированного белка BBD29_09525 P113S, последовательность аминокислот которого представляет собой SEQ ID NO: 4.
Мутация относится к изменению одного или нескольких оснований в гене в результате сайт-направленной мутации, что приводит к изменению аминокислотного состава соответствующего белка, с получением таким образом нового белка или обеспечением новой функции исходного белка, то есть представляет собой сайт-направленную мутацию генов. Методики осуществления сайт-направленных мутаций генов, такие как опосредованный олигонуклеотидными праймерами сайт-направленный мутагенез, ПНР-опосредованный сайт-направленный мутагенез или кассетный мутагенез, хорошо известны специалистам в данной области техники.
Точечная мутация согласно описанию в настоящем документе может представлять собой замену одного основания, инсерцию одного основания или делецию одного основания, и в особенности замену одного основания. Замена одного основания может представлять собой аллельную замену.
Точечная мутация может заключаться в осуществлении модификации нуклеиновой кислоты с заменой цитозина (С) в положении 337 гена BBD29_09525 (SEQ ID NO: 1).
В частности, точечная мутация может заключаться в осуществлении мутации цитозина (С) в положении 337 гена BBD29_09525 (SEQ ID NO: 1) на тимин (Т) с получением молекулы ДНК, представленной в SEQ ID NO: 2.
В настоящем документе рекомбинантный вектор может представлять собой, в частности, рекомбинантный вектор pK18-BBD29_09525C337T, PK18mobsacB-BBD29_09525, PK18mobsacB-BBD29_09525 С337Т, pXMJ19-BBD29_09525 или pXMJ19-BBD29_09525C337T.
Рекомбинантный вектор pK18-BBD29_09525C337T представляет собой рекомбинантный вектор, полученный путем замены фрагмента (фрагмента малого размера) между сайтами распознавания Xbal I WBamH I в векторе pK18mobsacB на фрагмент ДНК, представленный положениями 37 1466 SEQ ID NO: 29 в перечне последовательностей, при сохранении в неизмененном виде других последовательностей вектора pK18mobsacB. Рекомбинантный вектор pK18-BBD29_09525C337T содержит молекулу ДНК, представленную положениями 1-1051 в мутированном гене BBD29_09525 С337Т; представленном в SEQ ID NO: 2.
Рекомбинантный вектор PK18mobsacB-BBD29_09525 применяют для интеграции экзогенного гена BBD29_09525 в хромосому хозяина для сверхэкспрессии гена BBD29_09525 дикого типа у бактерии-продуцента. Рекомбинантный вектор pK18mobsacB-BBD29_09525 представляет собой рекомбинантный вектор, полученный путем замены фрагмента (фрагмента малого размера) между сайтами распознавания Xbal I и/ВатН I в векторе pK18mobsacB на фрагмент ДНК, представленный положениями 37-3407 SEQ ID NO: 30 в перечне последовательностей, при сохранении в неизмененном виде других последовательностей вектора pK18mobsacB.
Рекомбинантный вектор PK18mobsacB-BBD29_09525C337T применяют для интеграции экзогенного гена BBD29_09525 в хромосому хозяина для сверхэкспрессии мутантного типа гена BBD29_09525C337T у бактерии-продуцента. Рекомбинантный вектор pK18mobsacB-BBD29_09525C337T представляет собой рекомбинантный вектор, полученный путем замены фрагмента (фрагмента малого размера) между сайтами распознавания Xbal I и/ВатН I в векторе pK18mobsacB на фрагмент ДНК, представленный положениями 37-3407 SEQ ID NO: 31 в перечне последовательностей, при сохранении в неизмененном виде других последовательностей вектора pK18mobsacB.
Рекомбинантный вектор pXMJ19-BBD29_09525 применяют для экспрессии с помощью плазмиды экзогенного гена BBD29_09525 вне хромосомы для сверхэкспрессии гена BBD29_09525 дикого типа у бактерии-продуцента. Рекомбинантный вектор pXMJ19-BBD29_09525 представляет собой рекомбинантный вектор, полученный путем замены фрагмента (фрагмента малого размера) между сайтами распознавания EcoR I и Kpn I в векторе pXMJ19 на фрагмент ДНК, представленный положениями 37-1927 SEQ ID NO: 32 в перечне последовательностей, при сохранении в неизмененном виде других последовательностей вектора pXMJ19.
Рекомбинантный вектор pXMJ19-BBD29_09525С337Т применяют для экспрессии с помощью плазмиды экзогенного гена BBD29_09525С337Т вне хромосомы для сверхэкспрессии мутантного типа гена BBD29_09525С337Т у бактерии-продуцента. Рекомбинантный вектор pXMJ19-BBD29_09525C337T представляет собой рекомбинантный вектор, полученный путем замены фрагмента (фрагмента малого размера) между сайтами распознавания EcoR I и Kpn I в векторе pXMJ19 на фрагмент ДНК, представленный положениями 37-1927 SEQ ID NO: 33 в перечне последовательностей, при сохранении в неизмененном виде других последовательностей вектора pXMJ19.
Все рекомбинантные векторы из pK18-BBD29_09525С337Т, PK18mobsacB-BBD29_09525, PK18mobsacB-BBD29_09525C337T, pXMJ19-BBD29_09525 и pXMJ19-BBD29_09525C337T включены в заявленный объем настоящего изобретения.
В настоящем документе рекомбинантный микроорганизм может представлять собой, в частности, рекомбинантную бактерию YPG-007, YPG-008, YPG-009, YPG-010 или YPG-011.
Рекомбинантная бактерия YPG-007 представляет собой рекомбинантную бактерию, полученную путем трансформации рекомбинантного вектора pK18-BBD29_09525C337T в Corynebacterium glutamicum CGMCC №21220, и указанная рекомбинантная бактерия YPG-007 содержит мутированный ген BBD29_09525, представленный в SEQ ID NO: 2.
Рекомбинантная бактерия YPG-008 содержит двухкопийный ген BBD29_09525, представленный в SEQ ID NO: 1; рекомбинантная бактерия, содержащая двухкопийный ген BBD29_09525, способна значимо и устойчиво повышать уровень экспрессии гена BBD29_09525. Рекомбинантная бактерия YPG-008 представляет собой сконструированную бактерию, сверхэкспрессирующую ген BBD29_09525 дикого типа из генома.
Рекомбинантная бактерия YPG-009 содержит мутированный ген BBD29_09525C337T, представленный в SEQ ID NO: 2; указанная рекомбинантная бактерия YPG-009 представляет собой сконструированную бактерию, сверхэкспрессирующую мутантный тип гена BBD29_09525С337Т из генома.
Рекомбинантная бактерия YPG-010 содержит двухкопийный ген BBD29_09525, представленный в SEQ ID NO: 1; указанная рекомбинантная бактерия YPG-010 представляет собой сконструированную бактерию, сверхэкспрессирующую ген BBD29_09525 дикого типа на плазмиде, т.е. сверхэкспрессия происходит вне хромосомы с помощью плазмиды pXMJ19-BBD29_09525.
Рекомбинантная бактерия YPG-011 содержит мутированный ген BBD29_09525C337T, представленный в SEQ ID NO: 2; рекомбинантная бактерия YPG-011 представляет собой сконструированную бактерию, сверхэкспрессирующую мутантный тип гена BBD29_09525 С337Т на плазмиде, т.е. сверхэкспрессия происходит вне хромосомы с помощью плазмиды pXMJ19-BBD29_09525C337T.
Все рекомбинантные бактерии из YPG-007, YPG-008, YPG-009, YPG-010 и YPG-011 включены в заявленный объем настоящего изобретения.
Согласно настоящему изобретению также предложен способ конструирования рекомбинантного микроорганизма, при этом указанный способ включает по меньшей мере что-либо одно из:
F1) введения молекулы нуклеиновой кислоты BBD29_09525C337T в целевой микроорганизм с получением рекомбинантного микроорганизма;
F2) введения молекулы ДНК, представленной в SEQ ID NO: 1, в целевой микроорганизм с получением рекомбинантного микроорганизма;
F3) редактирования молекулы ДНК, представленной в SEQ ID NO: 1, с использованием средства редактирования генов (такого как редактирование генов с изменением одного основания) для введения молекулы ДНК, представленной в SEQ ID NO: 2, в целевой микроорганизм.
Указанное введение может быть представлено трансформацией бактерии-хозяина трансформация вектором, несущим молекулу ДНК согласно настоящему изобретению, любыми известными способами трансформации, такими как способ химической трансформации или способ электротрансформации. Введенная молекула ДНК может 5 быть однокопийной или многокопийной. Введение может быть представлено интеграцией экзогенного гена в хромосому хозяина, или экспрессией вне хромосомы с помощью плазмиды.
Согласно настоящему изобретению также предложен способ получения L-глутаминовой кислоты, при этом указанный способ включает продуцирование L-10 глутаминовой кислоты любым из рекомбинантных микроорганизмов согласно описанию в настоящем документе.
Описанный выше способ может представлять собой получение L-глутаминовой кислоты ферментативным способом, а рекомбинантный микроорганизм может относиться к роду Corynebacterium, и в частности, представлять собой Corynebacterium 15 glutamicum и его варианты.
Информация о депонировании: Название штамма: Corynebacterium glutamicum; Латинское название: Corynebacterium glutamicum; штамм №: YPGLU001; Депозитарное учреждение: Китайский главный центр коллекций микробиологических культур, сокращенно CGMCC; адрес: No. 3, Yard 1, Beichen West Road, Chaoyang District, Beijing; 20 Дата депонирования: 23 ноября 2020 г.; номер доступа в депозитарном центре: CGMCC №21220.
Наилучшие способы реализации изобретения
Настоящее изобретение будет подробно описано ниже на конкретных вариантах реализации; указанные примеры приведены исключительно для иллюстрации настоящего изобретения, но не для ограничения объема настоящего изобретения. Примеры, представленные ниже, могут применяться в качестве руководства для дальнейшего улучшения специалистами в данной области техники, и не подразумевают каких-либо ограничений настоящего изобретения.
Если конкретным образом не указано иное, все экспериментальные методы в приведенных ниже примерах представляют собой обычные способы, которые осуществляют в соответствии с методиками или условиями, описанными в литературе, относящейся к данной области техники, или в соответствии с инструкциями по применению продуктов. Если конкретным образом не указано иное, материалы и реагенты, используемые в приведенных ниже примерах, могут быть коммерчески доступными.
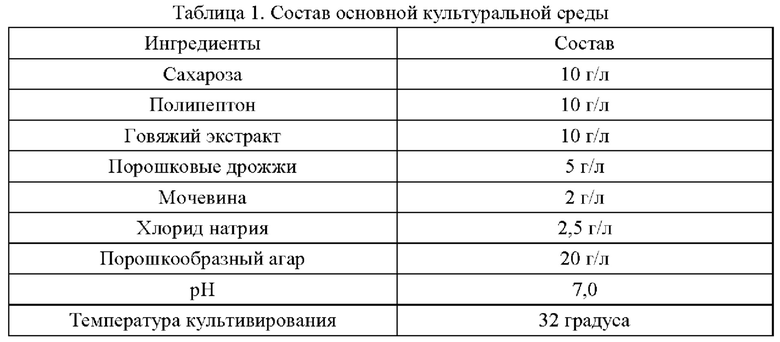
В приведенных ниже примерах состав основной среды, используемой для культивирования штамма, одинаковый, а сахарозу, канамицин или хлорамфеникол добавляют в состав основной среды по мере необходимости. Состав основной среды приведен в табл. 1:
Corynebacterium glutamicum YPGLU001 CGMCC №21220 в примерах ниже был депонирован в Китайском главном центре коллекций микробиологических культур (сокращенно CGMCC, адрес: No. 3, Yard 1, Beichen West Road, Chaoyang District, Beijing, Institute of Microbiology, Chinese Academy of Sciences) 23 ноября 2020 г. под номером доступа CGMCC №21220. Corynebacterium glutamicum YPGLU001 также известен как Corynebacterium glutamicum CGMCC №21220.
Пример 1. Конструирование вектора для трансформации pK18-BBD29_09525, содержащего кодирующую область гена BBD29_09525 с точечной мутацией
В соответствии с последовательностью генома Corynebacterium glutamicum АТСС13869, опубликованной в NCBI, разрабатывали и синтезировали два пары праймеров для амплификации последовательности кодирующей области гена BBD29_09525, и вводили точечную мутацию в штамм Corynebacterium glutamicum CGMCC №21220 путем аллельной замены (подтверждено соответствие кодирующей области гена BBD29_09525 на хромосоме штамма АТСС13869 при секвенировании). Последовательность аминокислот, соответствующая кодируемому белку, представлена в SEQ ID NO: 3, с заменой цитозина (С) в положении 337 последовательности нуклеотидов гена BBD29_09525 на тимин (Т) (SEQ ID NO: 2: BBD29_09525 С337Т), соответственно, с заменой пролина (Р) в положении 113 в последовательности аминокислот, соответствующей кодируемому белку, на серин (S) (SEQ ID NO: 4: BBD29_09525 P113S).
Точечная мутация представляет собой мутацию цитозина (С) в положении 337 в последовательности нуклеотидов гена BBD29_09525 (SEQ ID NO: 1) на тимин (Т), обеспечивающую получение молекулы ДНК, представленной в SEQ ID NO: 2 (мутированный ген BBD29_09525, обозначенный как BBD29_09525C337T).
При этом молекула ДНК, представленная в SEQ ID NO: 1, кодирует белок, имеющий последовательность аминокислот SEQ ID NO: 3 (указанный белок обозначен как белок BBD29_09525).
Молекула ДНК, представленная в SEQ ID NO: 2, кодирует мутантный белок, последовательность аминокислот которого представлена SEQ ID NO: 4 (указанный мутантный белок обозначен как BBD29_09525P113S). Присутствие серина (S) в положении 113 в последовательности аминокислот мутантного белка BBD29_09525P113S (SEQ ID NO: 4) обусловлено мутацией пролина (Р).
Методику ПЦР с перекрывающимися праймерами используют для сайт-направленного мутагенеза генов со следующими разработанными праймерами (синтезированы Invitrogen Corporation, Шанхай). Основание, где происходит мутация, выделено жирным шрифтом:
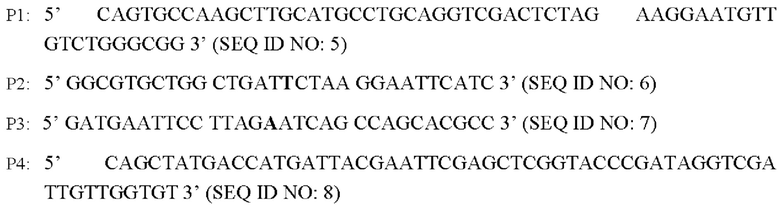
Способ конструирования: используя Corynebacterium glutamicum АТСС13869 в качестве матрицы, осуществляют ПЦР-амплификацию с соответствующими праймерами P1, Р2 и Р3, Р4 с получением двух фрагментов ДНК кодирующей области гена BBD29_09525, каждый из которых содержит мутированное основание и имеет размер 766 и.о. и 768 и.о., соответственно (BBD29_09525 Up и BBD29_09525 Down).
ПЦР-система: 10 × буфер Ex Taq Buffer: 5 мкл, смесь dNTP (каждый 2,5 мМ): 4 мкл, Mg2+(25 мМ): 4 мкл, праймеры (10 пМ): по 2 мкл каждого, Ex Taq (5 Ед/мкл): 0,25 мкл, в общем объеме 50 мкл.
Вышеуказанную ПЦР-амплификацию осуществляют путем предварительной денатурации в течение 5 минут при 94°С, денатурации в течение 30 с при 94°С, ренатурации в течение 30 с при 52°С, удлинения в течение 45 с при 72°С в течение 30 циклов и избыточного удлинения в течение 10 мин при 72°С с получением двух фрагментов ДНК, содержащих кодирующую область гена BBD29_09525, каждый из которых имеет размер 766 п.о. и 768 и.о. (BBD29_09525 Up и BBD29_09525 Down).
Указанные два фрагмента ДНК (BBD29_09525 Up и BBD29_09525 Down) выделяют посредством электрофореза на агарозном геле и очищают, после чего извлекают целевые полосы. Указанные два фрагмента ДНК затем используют в качестве матрицы с Р1 и Р4 в качестве праймеров для ПЦР-амплификации с перекрывающимися праймерами с получением фрагмента длиной 1504 п.о., обозначенного как BBD29_09525 Up-Down (последовательность представлена в SEQ ID NO: 29). В молекуле ДНК, представленной в SEQ ID NO: 29, положения 37-1087 представляют собой фрагмент гена BBD29_09525C337T с сайтом мутации (т.е. положения 1-1051 SEQ ID NO: 2).
ПЦР-система с перекрывающимися праймерами: 10 × буфер Ex Taq: 5 мкл, смесь dNTP (каждый 2,5 мМ): 4 мкл, Mg2+(25 мМ): 4 мкл, праймеры (10 пМ): 2 мкл каждого, Ex Taq (5 Ед/мкл): 0,25 мкл, в общем объеме 50 мкл.
ПЦР-амплификацию с перекрывающимися праймерами осуществляют путем предварительной денатурации в течение 5 минут при 94°С, денатурации в течение 30 с при 94°С, ренатурации в течение 30 с при 52°С, удлинения в течение 90 с при 72°С в течение 30 циклов и избыточного удлинения в течение 10 мин при 72°С.
Фрагмент ДНК BBD29_09525 Up-Down (SEQ ID NO: 29) содержит сайт мутации для введения модификации нуклеиновой кислоты в положении 337 кодирующей области гена BBD29_09525 у Corynebacterium glutamicum CGMCC №21220, в частности, изменения цитозина (С) в положении 337 кодирующей области гена BBD29_09525 у Corynebacterium glutamicum CGMCC №21220 на тимин (Т), в конечном итоге обуславливающего изменение аминокислоты в положении 113 кодируемого белка с пролина (Р) на серин (S).
После расщепления плазмиды pK18mobsacB (приобретена у Addgene Corporation) Xba I /BamH I, BBD29_09525C337T и линеаризированную плазмиду pK18mobsacB выделяют посредством электрофореза на агарозном геле и очищают, с последующей сборкой с применением системы рекомбинации NEBuider, получая вектор рК18-BBD29_09525C337T, плазмиду, содержащую маркер устойчивости к канамицину. При этом вектор pK18-BBD29_09525 отправляют для секвенирования и идентификации в компанию, осуществляющую секвенирование, и содержащий корректную точечную мутацию (С-Т) вектор pK18-BBD29_09525 отправляют на хранение.
В частности, фрагмент ДНК (BBD29_09525 Up-Down) очищают после выделения посредством электрофореза на агарозном геле, с последующим лигированием в плазмиду pK18mobsacB (приобретена у Addgene Corporation, расщеплена Xbal I/BamH I), очищенную после расщепления ферментами (Xbal I/BamH I) в течение 30 минут при 50°С с применением фермента NEBuilder (приобретен у NEB Corporation). Выросший одиночный клон после трансформации продукта лигирования DH5a (приобретен у TAKARA Corporation), подвергают ПЦР-идентификации для обнаружения положительного рекомбинантного вектора pK18-BBD29_09525С337Т, содержащего маркер устойчивости к канамицину (Kanr). Корректно расщепленный рекомбинантный вектор pK18-BBD29-09525C337T отправляют для секвенирования и идентификации в компанию, осуществляющую секвенирование, а содержащий корректную точечную мутацию (С-Т) рекомбинантный вектор pK18-BBD29_09525С337Т отправляют на хранение.
Рекомбинантный вектор pK18-BBD29_09525С337Т представляет собой рекомбинантный вектор, полученный путем замены фрагмента (фрагмента малого размера) между сайтами распознавания Xbal I и/BamH I в векторе pK18mobsacB на фрагмент ДНК, представленный положениями 37-1466 SEQ ID NO: 29 в перечне последовательностей, при сохранении в неизмененном виде других последовательностей вектора pK18mobsacB.
Рекомбинантный вектор pK18-BBD29_09525C337T содержит молекулу ДНК, представленную положениями 1-1051 в мутированном гене BBD29_09525C337T, представленном в SEQ ID NO: 2
Пример 2. Конструирование сконструированного штамма, содержащего BBD29_09525С337Т с точечной мутацией
Способ конструирования: Плазмидой pK18-BBD29_09525С337Т после аллельной замены согласно примеру 1 электротрансформируют Corynebacterium glutamicum CGMCC №21220 для культивирования в культуральной среде. См. в табл. 1 ингредиенты среды и условия культивирования. Каждую из выросших моноколоний идентифицируют с применением праймера Р1 и универсального праймера M13R для определения положительного штамма по полосе, соответствующей размеру приблизительно 1511 п.о., полученной при амплификации. Положительный штамм культивируют на культуральной среде, содержащей 15% сахарозы, и выросшие моноколонии культивируют на культуральных средах с канамицином и без канамицина, соответственно. Штаммы, растущие на культуральной среде без канамицина и не растущие на культуральной среде с канамицином, выбирают для дальнейшей идентификации с помощью ПЦР со следующими праймерами (синтезированы Invitrogen Corporation, Шанхай):
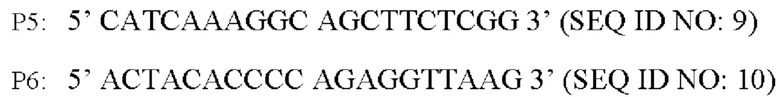
Описанные выше амплифицированные продукты ПЦР подвергают денатурации при высокой температуре и на ледяной бане, после чего проводят SSCP-электрофорез (с амплифицированным фрагментом вектора pK18-BBD29_09525 С337Т в качестве положительного контроля, амплифицированным фрагментом АТСС13869 в качестве отрицательного контроля, и водой в качестве холостого контроля). См. в табл. 2 описание подготовки ПААГ для SSCP-электрофореза и условий электрофореза. Поскольку фрагменты отличаются структурой, их локализация при электрофорезе будет разной. Соответственно, штамм, характеризующийся успешной аллельной заменой, представляет собой такой штамм, локализация фрагмента которого при электрофорезе не совпадает с локализацией отрицательного контрольного фрагмента, и совпадает с локализацией положительного контрольного фрагмента. Праймеры Р5 и Р6 используют также для ПЦР-амплификации целевого фрагмента штамма, характеризующегося успешной аллельной заменой, который затем лигируют в вектор PMD19-T для секвенирования. Используя выравнивание последовательностей для мутированной последовательности оснований, подтверждают успешность аллельной замены у указанного штамма, который обозначен как YPG-007.
Рекомбинантная бактерия YPG-007 содержит мутированный ген BBD29_09525C337T SEQ ID NO: 2.
Пример 3. Конструирование сконструированного штамма со сверхэкспрессируемым геном BBD29_09525 или BBD29_09525C337T в геноме
Для дополнительного изучения и подтверждения того, что сверхэкспрессия гена BBD29_09525 дикого типа или его производного мутантного гена BBD29_09525 у бактерии-продуцента может повышать продуцирование L-глутаминовой кислоты, экзогенный ген интегрируют в хромосому хозяина для конструирования сконструированного штамма с геном BBD29_09525 или геном BBD29_09525, сверхэкспрессируемым из генома.
В соответствии с последовательностью генома Corynebacterium glutamicum АТСС13869, опубликованной в базе NCBI, разрабатывали и синтезировали три пары праймеров для амплификации 5'- и 3'-фрагментов плеч гомологии, а также последовательности кодирующей области гена BBD29_09525 и области промотора для введения гена BBD29_09525 или BBD29_09525С337Т в штамм Corynebacterium glutamicum CGMCC №21220 путем гомологичной рекомбинации.
Разработаны следующие праймеры (синтезированы mvitrogen Corporation, Шанхай):

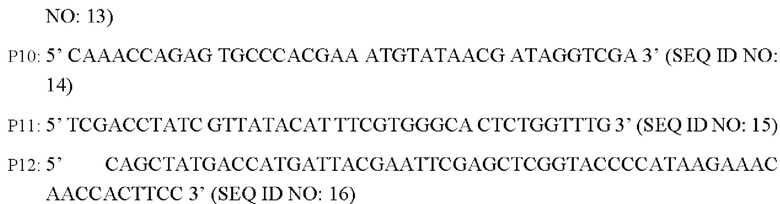
Способ конструирования: Используя Corynebacterium glutamicum АТСС13869 или YPG-007, соответственно, в качестве матрицы, осуществляют ПЦР-амплификацию с праймерами Р7/Р8, Р9/Р10 и Р11/Р22, соответственно, с получением 5'-фрагмента плеча гомологии длиной около 807 п.о., фрагмента кодирующей области гена BBD29_09525 и области промотора длиной 1931 п.о. или фрагмента кодирующей области гена BBD29_09525 С337Т и области промотора длиной около 1931 п.о., и 3'-фрагмента плеча гомологии длиной около 787 п.о. Кроме того, с применением Р7/Р12 в качестве праймеров осуществляют амплификацию со смесью трех описанных выше амплифицированных фрагментов в качестве матрицы с получением единых фрагментов плеч гомологии 1 и 2 (единый фрагмент плеч гомологии 1 длиной 3445 п.о., согласно SEQ ID NO: 30; и единый фрагмент плеч гомологии 2 длиной 3445 п.о., согласно SEQ ID NO: 31). После завершения ПЦР-реакции осуществляют электрофорез амплифицированных продуктов для выделения требуемого фрагмента ДНК длиной приблизительно 3445 п.о. с применением набора с колонкой для выделения ДНК на геле (TIANGEN), который лигируют, используя систему рекомбинации NEBuider, в челночную плазмиду PK18mobsacB, расщепленную Xba I и выделенную с получением интегративной плазмиды (т.е. рекомбинантного вектора) PK18mobsacB-BBD29_09525 или PK18mobsacB-BBD29_09525. Указанная плазмида содержит маркер устойчивости к канамицину, благодаря чему путем скрининга с канамицином может быть получен рекомбинантный штамм, содержащий интегрированную в геном плазмиду.
Рекомбинантный вектор pK18mobsacB-BBD29_09525 представляет собой рекомбинантный вектор, полученный путем замены фрагмента (фрагмента малого размера) между сайтами распознавания Xbal I и/BamH I в векторе pK18mobsacB на фрагмент ДНК, представленный положениями 37-3407 SEQ ID NO: 30 в перечне последовательностей, при сохранении в неизмененном виде других последовательностей вектора pK18mobsacB.
Рекомбинантный вектор pK18mobsacB-BBD29_09525С337Т представляет собой рекомбинантный вектор, полученный путем замены фрагмента (фрагмента малого размера) между сайтами распознавания Xbal I и/BamH I в векторе pK18mobsacB на фрагмент ДНК, представленный положениями 37-3407 SEQ ID NO: 31 в перечне последовательностей, при сохранении в неизмененном виде других последовательностей вектора pK18mobsacB. ПЦР-система: 10 × буфер Ex Taq Buffer: 5 мкл, смесь dNTP (каждый 2,5 мМ): 4 мкл, Mg2+(25 мМ): 4 мкл, праймеры (10 пМ): по 2 мкл каждого, Ex Taq (5 Ед/мкл): 0,25 мкл, в общем объеме 50 мкл.
ПЦР-амплификацию осуществляют путем предварительной денатурации в течение 5 минут при 94°С, денатурации в течение 30 с при 94°С, ренатурации в течение 30 с при 52°С, удлинения в течение 120 с при 72°С (30 циклов) и избыточного удлинения в течение 10 мин при 72°С.
Каждую из этих двух интегративных плазмид (PK18mobsacB-BBD29_09525 и PK18mobsacB-BBD29_09525C337T) вводят в штамм Corynebacterium glutamicum CGMCC №21220 путем электротрансформации, и осуществляют ПЦР-идентификацию выросших моноколоний с праймерами Р13/Р14 для обнаружения положительного штамма, содержащего фрагмент длиной приблизительно 1821 п.о., полученный при ПЦР-амплификации, тогда как штамм, не содержащий каких-либо амплифицированных этим способом фрагментов, представляет собой исходный штамм. При скрининге с 15% сахарозы положительный штамм культивируют на культуральных средах с канамицином и без канамицина, соответственно, и штаммы, растущие на культуральной среде без канамицина и не растущие на культуральной среде с канамицином подвергают дальнейшей ПЦР с праймерами Р15/Р16 для идентификации. Штамм, из которого амплифицирован фрагмент длиной приблизительно 1769 п.о., представляет собой штамм с геном BBD29_09525 или BBD29_09525С337Т, интегрированным в геном Corynebacterium glutamicum CGMCC №21220, обозначенный как YPG-008 (без точечной мутации) и YPG-009 (с точечной мутацией).
Рекомбинантная бактерия YPG-008 содержит двухкопийный ген BBD29_09525 согласно SEQ ID NO: 1; указанная рекомбинантная бактерия, содержащая двухкопийный ген BBD29_09525, способна значимо и устойчиво повышать уровень экспрессии гена BBD29_09525. Рекомбинантная бактерия YPG-008 представляет собой сконструированную бактерию, сверхэкспрессирующую ген BBD29_09525 дикого типа из генома.
Рекомбинантная бактерия YPG-009 содержит мутированный ген BBD29_09525C337T согласно SEQ ID NO: 2; рекомбинантная бактерия YPG-009 представляет собой сконструированную бактерию, сверхэкспрессирующую мутантный ген BBD29_09525C337T из генома.
Праймеры для ПЦР-идентификации представлены ниже:

Пример 4. Конструирование модифицированного штамма, сверхэкспрессирующего ген BBD29_09525 или BBD29_09525C337T с плазмиды
В соответствии с последовательностью генома Corynebacterium glutamicum АТСС13869, опубликованной в базе NCBI, разрабатывали и синтезировали пару праймеров для амплификации последовательности кодирующей области гена BBD29_09525 и области промотора. Разработаны следующие праймеры (синтезированы hwitrogen Corporation, Шанхай):
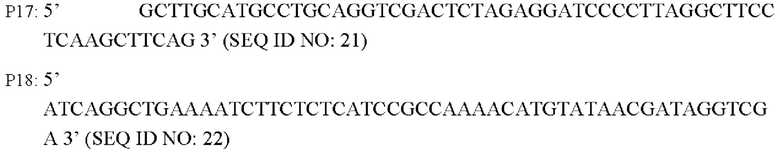
Способ конструирования: Используя YPG-008 или YPG-007, соответственно, в качестве матрицы, осуществляют ПЦР-амплификацию с праймерами Р17/р18 с получением фрагмента гена BBD29_09525 и его промотора длиной 1961 п.о. (SEQ ID NO: 32) или фрагмента гена BBD29_09525 С337Ти его промотора длиной 1961 п.о. (SEQ ID NO: 33). Осуществляют электрофорез амплифицированных продуктов для выделения требуемого фрагмента ДНК в 1961 и.о. с применением набора с колонкой для выделения ДНК на геле, который лигируют с использованием системы рекомбинации NEBuider в челночную плазмиду pXMJ19, расщепленную EcoR I/Kpn I, и выделяют, получая сверхэкспрессирующую плазмиду (т.е. рекомбинантный вектор) pXMJ19-BBD29_09525 или pXMJ19-BBD29_09525. Указанная плазмида содержит маркер устойчивости к хлорамфениколу, и трансформация штамма плазмидой может быть достигнута при скрининге с хлорамфениколом.
Рекомбинантный вектор pXMJ19-BBD29_09525 представляет собой рекомбинантный вектор, полученный путем замены фрагмента (фрагмента малого размера) между сайтами распознавания EcoR I и Kpn I в векторе pXMJ19 на фрагмент ДНК, представленный положениями 37-1927 SEQ ID NO: 32 в перечне последовательностей, при сохранении в неизмененном виде других последовательностей вектора pXMJ19.
Рекомбинантный вектор pXMJ19-BBD29_09525C337T представляет собой рекомбинантный вектор, полученный путем замены фрагмента (фрагмента малого размера) между сайтами распознавания EcoR I и Kpn I в векторе pXMJ19 на фрагмент ДНК, представленный положениями 37-1927 SEQ ID NO: 33 в перечне последовательностей, при сохранении в неизмененном виде других последовательностей вектора pXMJ19.
ПЦР-система: 10 × буфер Ex Taq Buffer: 5 мкл, смесь dNTP (каждый 2,5 мМ): 4 мкл, Mg2+(25 мМ): 4 мкл, праймеры (10 пМ): по 2 мкл каждого, Ex Taq (5 Ед/мкл): 0,25 мкл, в общем объеме 50 мкл.
ПЦР-амплификацию осуществляют путем предварительной денатурации в течение 5 минут при 94°С, денатурации в течение 30 с при 94°С, ренатурации в течение 30 с при 52°С и удлинения в течение 120 с при 72°С (30 циклов) и избыточного удлинения в течение 10 мин при 72°С.
Каждую из двух плазмид (pXMJ19-BBD29_09525 и pXMJ19-BBD29_09525 С337Т) вводят в штамм Corynebacterium glutamicum CGMCC №21220 путем электротрансформации, и осуществляют ПЦР-идентификацию выросших моноколоний с праймерами M13R (-48) и р18 для обнаружения трансформированного штамма, содержащего фрагмент длиной приблизительно 2000 п.о., полученный при ПЦР-амплификации, обозначенного как YPG-010 (без точечной мутации) и YPG-011 (с точечной мутацией).
Рекомбинантная бактерия YPG-010 содержит двухкопийный ген BBD29_09525 согласно SEQ ID NO: 1; рекомбинантная бактерия YPG-010 представляет собой сконструированную бактерию, сверхэкспрессирующую ген BBD29_09525 дикого типа на плазмиде, т.е. сверхэкспрессия происходит вне хромосомы с плазмиды pXMJ19-BBD29_09525.
Рекомбинантная бактерия YPG-011 содержит мутированный ген BBD29_09525C337T согласно SEQ ID NO: 2; рекомбинантная бактерия YPG-011 представляет собой сконструированную бактерию, сверхэкспрессирующую мутантный ген BBD29_09525С337Т на плазмиде, т.е. сверхэкспрессия происходит вне хромосомы с плазмиды pXMJ19-BBD29_09525C337T.
Пример 5. Конструирование сконструированного штамма с делетированным в геноме геном BBD29_09525.
В соответствии с последовательностью генома Corynebacterium glutamicum АТСС13869, опубликованной в базе NCBI, синтезируют две пары праймеров для амплификации фрагментов на обоих концах кодирующей области гена BBD29_09525, в виде 5'- и 3'-фрагментов плеч гомологии. Разработаны следующие праймеры (синтезированы Invitrogen Corporation, Шанхай):

Способ конструирования: Используя в качестве матрицы Corynebacterium glutamicum АТСС13869, осуществляют ПЦР-амплификацию с праймерами Р19/Р20 и Р21/Р22, соответственно, с получением 5'-фрагмента плеча гомологии длиной 804 п.о. и 3'-фрагмента плеча гомологии длиной 807 п.о. Кроме того, Р19/Р22 используют в качестве праймеров для ПЦР с перекрывающимися праймерами с получением единого фрагмента плеча гомологии длиной 1571 п.о. После завершения ПЦР-реакции проводят электрофорез амплифицированных продуктов для выделения требуемого фрагмента ДНК длиной 1571 п.о. с применением набора с колонкой для выделения ДНК на геле, который лигируют с помощью системы рекомбинации NEBuider в челночную плазмиду, плазмиду pkl8mobsacB, расщепленную Xba I и выделенную с получением нокаутной плазмиды. Указанная плазмида содержит маркер устойчивости к канамицину.
Указанной нокаутной плазмидой электротрансформируют штамм Corynebacterium glutamicum CGMCC №21220 и осуществляют ПЦР-идентификацию выросших моноколоний, в каждом случае со следующими праймерами (синтезированы Invitrogen Corporation, Шанхай):
Штамм с полосами, соответствующими 1497 п.о. и 3000 п.о., амплифицированными в ходе описанной выше ПЦР-амплификации, представляет собой положительный штамм, тогда как штамм с единственной амплифицированной полосой, соответствующей 3000 п.о., представляет собой исходный штамм. При скрининге на культуральной среде с 15% сахарозы положительный штамм культивируют на культуральных средах с канамицином и без канамицина, соответственно, и выбирают штаммы, растущие на культуральной среде без канамицина и не растущие на культуральной среде с канамицином, для дальнейшей ПЦР-идентификации с праймерами Р23/Р24. Штамм с амплифицированной полосой, соответствующей 1497 п.о., представляет собой генетически сконструированный штамм с нокаутом кодирующей области гена BBD29_09525, обозначенный как YPG-012 (нокаутирован ген BBD29_09525 в геноме Corynebacterium glutamicum CGMCC №21220).
Пример 6. Эксперименты с ферментацией для получения L-глутаминовой кислоты
Эксперименты с ферментацией осуществляют на штаммах (YPG-007, YPG-008, YPG-009, YPG-010, YPG-011 и YPG-012), сконструированных согласно приведенным выше примерам 2-5, и исходном штамме Corynebacterium glutamicum CGMCC №21220, в ферментативном реакторе типа BLBIO-5GC-4-H (приобретен у Shanghai Bailun Biological Technology Co., Ltd.); культуральная среда представлена в табл. 3, процесс управления представлен в табл. 4. Для каждого штамма эксперименты проводят в трех повторностях. Результаты приведены в табл. 5.

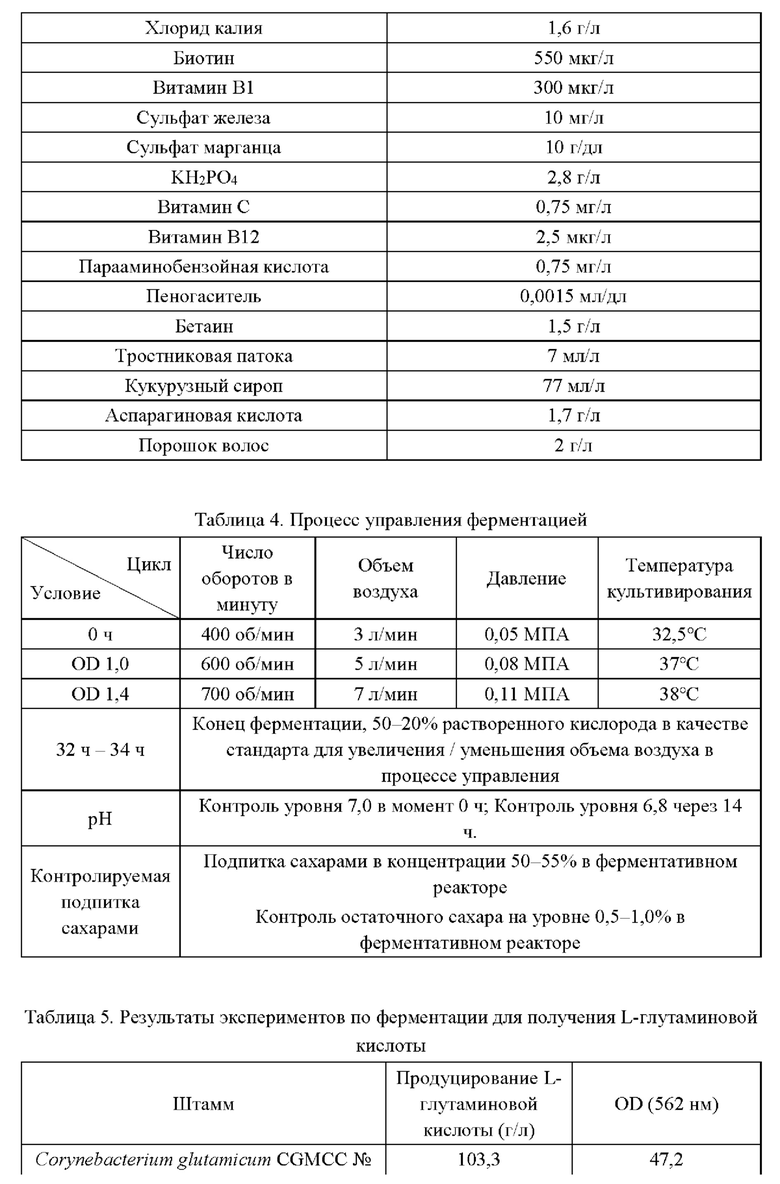
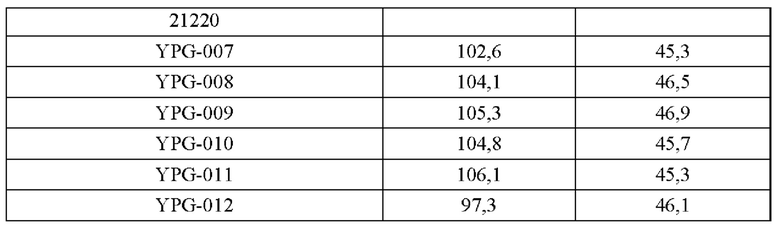
Результаты из табл. 5 демонстрируют, что сверхэкспрессия гена BBD29_09525, или точечная мутация в кодирующей области гена BBD29_09525, BBD29_09525С337Т, и/или сверхэкспрессия гена BBD29_09525 или мутантного гена BBD29_09525С337Т у Corynebacterium glutamicum способствуют повышению продуцирования L-глутаминовой кислоты, тогда как ослабление экспрессии или нокаут гена BBD29_09525 неблагоприятны для накопления L-глутаминовой кислоты.
Настоящее изобретение было подробно описано выше. Специалисты в данной области техники смогут широко реализовать настоящее изобретение на практике, используя эквивалентные параметры, концентрации и условия без отступления от существа и объема настоящего изобретения, не прибегая к избыточному экспериментированию. Хотя были предложены конкретные примеры реализации настоящего изобретения, следует понимать, что возможны его дальнейшие улучшения. Таким образом, в соответствии с принципами настоящего изобретения, настоящим документом охвачены все варианты, применения или улучшения настоящего изобретения, в том числе изменения, вносимые с использованием обычных методик, известных в данной области техники, выходящие за пределы заявленного в настоящем документе объема изобретения. Некоторые существенные признаки могут быть использованы в соответствии с прилагаемой формулой изобретения.
Промышленное применение
Согласно настоящему изобретению было обнаружено, что через снижение экспрессии или нокаут гена BBD29_09525 кодируемый этим геном продукт влияет на способность к продуцированию L-глутаминовой кислоты, и что рекомбинантный штамм, полученный путем введения точечной мутации в кодирующую последовательность, или путем увеличения числа копий, или сверхэкспрессии указанного гена, способствует продуцированию глутаминовой кислоты в более высокой концентрации по сравнению с немодифицированным штаммом.
В частности, согласно настоящему изобретению сначала конструируют генетически сконструированную бактерию YPG-007 с точечной мутацией (С-Т) путем введения точечной мутации в кодирующую область гена BBD29_09525 (SEQ ID NO: 1) Corynebacterium glutamicum CGMCC №21220 путем аллельной замены. Для дополнительного изучения и подтверждения того, что сверхэкспрессия гена BBD29_09525 дикого типа или мутантного гена BBD29_09525 С337Т у бактерии-продуцента может повышать продуцирование L-глутаминовой кислоты, экзогенный ген интегрируют в хромосому хозяина или экспрессируют вне хромосомы плазмидой, соответственно, конструируя таким образом сконструированные бактерии YPG-008, YPG-009, YPG-010 и YPG-011, сверхэкспрессирующие ген BBD29_09525 или ген BBD29_09525С337Т из генома и плазмиды. Эксперименты предполагают, что ген BBD29_09525 и его варианты вовлечены в биосинтез L-глутаминовой кислоты. Используя сверхэкспрессию или нокаут гена BBD29 11265, или сайт-направленную мутацию (такую как точечная мутация) в указанном гене, можно регулировать уровень накопления L-глутаминовой кислоты у микроорганизма. Точечная мутация в кодирующей области гена BBD29_09525 или сверхэкспрессия гена BBD29_09525 или его мутантного гена BBD29_09525С337Т у бактерии-продуцента способствует повышению продуцирования и увеличению показателя конверсии L-глутаминовой кислоты, тогда как нокаут или ослабление экспрессии гена BBD29_09525 неблагоприятны для накопления L-глутаминовой кислоты. Ген BBD29_09525 и его вариант (такой как ген BBD29_09525 С337Т) могут применяться для конструирования генетически сконструированного штамма для продуцирования L-глутаминовой кислоты, чтобы способствовать повышению продуцирования L-глутаминовой кислоты, и для получения высокопродуктивного и высококачественного штамма для промышленного производства, полезного для широкого диапазона вариантов применения и имеющего высокую экономическую ценность для промышленного производства L-глутаминовой кислоты.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> INNER MONGOLIA EPPEN BIOTECHCO., LTD / ИННЕР МОНГОЛИА ЭППЕН БИОТЕХ
КО., ЛТД
<120> Recombinant Strain with Modified Gene BBD29_09525 for Producing
L-Glutamic Acid, and Method for Constructing the Same and Use
Thereof / РЕКОМБИНАНТНЫЙ ШТАММ С МОДИФИЦИРОВАННЫМ ГЕНОМ BBD29_09525
ДЛЯ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ, А ТАКЖЕ СПОСОБ ЕГО
КОНСТРУИРОВАНИЯ И ЕГО ПРИМЕНЕНИЕ
<130> GNPDH21085
<150> CN202011631266.8
<151> 2020-12-30
<160> 33
<170> PatentIn, версия 3.5
<210> 1
<211> 1503
<212> ДНК
<213> Corynebacterium glutamicum
<220>
<221> CDS/Кодирующая последовательность
<222> (1)..(1503)
<400> 1
atgtcagatt ccccgaagaa cgcaccgagg attaccgatg aggcagatgt agttctcatt 60
ggtgccggta tcatgagctc cacgctgggt gcaatgctgc gtcagctgga gccaagctgg 120
actcagatcg tcttcgagcg tttggatgga ccggcacaag agtcgtcctc cccgtggaac 180
aatgcaggaa ccggccactc tgctctatgc gagctgaact acaccccaga ggttaagggc 240
aaggttgaaa ttgccaaggc tgtaggaatc aacgagaagt tccaggtttc ccgtcagttc 300
tggtctcacc tcgttgaaga gggcgtgctg gctgatccta aggaattcat caaccctgtt 360
cctcacgtat ctttcggcca gggcgcagac caggttgcat acatcaaggc tcgctacgaa 420
gctttgaagg atcacccact cttccagggc atgacctacg ctgacgatga agctaccttc 480
accgagaagc tgcctttgat ggcaaagggc cgtgacttct ctgatccagt agcaatctct 540
tggatcgatg aaggcaccga catcaactac ggtgctcaga ccaagcagta cctggatgca 600
tctgaagttg aaggcactga aatccgctat ggccacgaag tcaagagcat caaggctgat 660
ggcgcaaagt ggatcgtgac cgtcaagaac gtacacactg gcgacaccaa gaccatcaag 720
gcaaacttcg tgttcgtcgg cgcaggcggg tacgcactgg atctgcttcg cagcgcaggc 780
atcccacagg tcaagggctt cgctggattc ccagtatccg gcctgtggct tcgttgcacc 840
aacgaggaac tgatcgagca gcacgcagcc aaggtatatg gcaaggcatc tgttggcgct 900
cctccaatgt ctgttcctca ccttgacacc cgcgttatcg agggtgaaaa gggtctgctc 960
tttggacctt acggtggctg gacccctaag ttcttgaagg aaggctccta cctggacctg 1020
ttcaagtcca tccgcccaga caacattcct tcctaccttg gcgttgctgc tcaggaattt 1080
gatctgacca agtaccttgt cactgaagtt ctcaaggacc aggacaagcg tatggatgct 1140
cttcgcgagt acatgccaga ggcacaaaac ggcgattggg agaccatcgt tgccggacag 1200
cgtgttcagg ttattaagcc tgcaggattc cctaagttcg gttccctgga attcggcacc 1260
accttgatca acaactccga aggcaccatc gccggattgc tcggtgcttc ccctggagca 1320
tccatcgcac cttccgcaat gatcgagctg cttgagcgtt gcttcggtga ccgcatgatc 1380
gagtggggcg acaagctgaa ggacatgatc ccttcctacg gcaagaagct tgcttccgag 1440
ccagcactgt ttgagcagca gtgggcacgc acccagaaga ccctgaagct tgaggaagcc 1500
taa 1503
<210> 2
<211> 1503
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<220>
<221> CDS/Кодирующая последовательность
<222> (1)..(1503)
<400> 2
atgtcagatt ccccgaagaa cgcaccgagg attaccgatg aggcagatgt agttctcatt 60
ggtgccggta tcatgagctc cacgctgggt gcaatgctgc gtcagctgga gccaagctgg 120
actcagatcg tcttcgagcg tttggatgga ccggcacaag agtcgtcctc cccgtggaac 180
aatgcaggaa ccggccactc tgctctatgc gagctgaact acaccccaga ggttaagggc 240
aaggttgaaa ttgccaaggc tgtaggaatc aacgagaagt tccaggtttc ccgtcagttc 300
tggtctcacc tcgttgaaga gggcgtgctg gctgattcta aggaattcat caaccctgtt 360
cctcacgtat ctttcggcca gggcgcagac caggttgcat acatcaaggc tcgctacgaa 420
gctttgaagg atcacccact cttccagggc atgacctacg ctgacgatga agctaccttc 480
accgagaagc tgcctttgat ggcaaagggc cgtgacttct ctgatccagt agcaatctct 540
tggatcgatg aaggcaccga catcaactac ggtgctcaga ccaagcagta cctggatgca 600
tctgaagttg aaggcactga aatccgctat ggccacgaag tcaagagcat caaggctgat 660
ggcgcaaagt ggatcgtgac cgtcaagaac gtacacactg gcgacaccaa gaccatcaag 720
gcaaacttcg tgttcgtcgg cgcaggcggg tacgcactgg atctgcttcg cagcgcaggc 780
atcccacagg tcaagggctt cgctggattc ccagtatccg gcctgtggct tcgttgcacc 840
aacgaggaac tgatcgagca gcacgcagcc aaggtatatg gcaaggcatc tgttggcgct 900
cctccaatgt ctgttcctca ccttgacacc cgcgttatcg agggtgaaaa gggtctgctc 960
tttggacctt acggtggctg gacccctaag ttcttgaagg aaggctccta cctggacctg 1020
ttcaagtcca tccgcccaga caacattcct tcctaccttg gcgttgctgc tcaggaattt 1080
gatctgacca agtaccttgt cactgaagtt ctcaaggacc aggacaagcg tatggatgct 1140
cttcgcgagt acatgccaga ggcacaaaac ggcgattggg agaccatcgt tgccggacag 1200
cgtgttcagg ttattaagcc tgcaggattc cctaagttcg gttccctgga attcggcacc 1260
accttgatca acaactccga aggcaccatc gccggattgc tcggtgcttc ccctggagca 1320
tccatcgcac cttccgcaat gatcgagctg cttgagcgtt gcttcggtga ccgcatgatc 1380
gagtggggcg acaagctgaa ggacatgatc ccttcctacg gcaagaagct tgcttccgag 1440
ccagcactgt ttgagcagca gtgggcacgc acccagaaga ccctgaagct tgaggaagcc 1500
taa 1503
<210> 3
<211> 500
<212> БЕЛОК
<213> Corynebacterium glutamicum
<400> 3
Met Ser Asp Ser Pro Lys Asn Ala Pro Arg Ile Thr Asp Glu Ala Asp
1 5 10 15
Val Val Leu Ile Gly Ala Gly Ile Met Ser Ser Thr Leu Gly Ala Met
20 25 30
Leu Arg Gln Leu Glu Pro Ser Trp Thr Gln Ile Val Phe Glu Arg Leu
35 40 45
Asp Gly Pro Ala Gln Glu Ser Ser Ser Pro Trp Asn Asn Ala Gly Thr
50 55 60
Gly His Ser Ala Leu Cys Glu Leu Asn Tyr Thr Pro Glu Val Lys Gly
65 70 75 80
Lys Val Glu Ile Ala Lys Ala Val Gly Ile Asn Glu Lys Phe Gln Val
85 90 95
Ser Arg Gln Phe Trp Ser His Leu Val Glu Glu Gly Val Leu Ala Asp
100 105 110
Pro Lys Glu Phe Ile Asn Pro Val Pro His Val Ser Phe Gly Gln Gly
115 120 125
Ala Asp Gln Val Ala Tyr Ile Lys Ala Arg Tyr Glu Ala Leu Lys Asp
130 135 140
His Pro Leu Phe Gln Gly Met Thr Tyr Ala Asp Asp Glu Ala Thr Phe
145 150 155 160
Thr Glu Lys Leu Pro Leu Met Ala Lys Gly Arg Asp Phe Ser Asp Pro
165 170 175
Val Ala Ile Ser Trp Ile Asp Glu Gly Thr Asp Ile Asn Tyr Gly Ala
180 185 190
Gln Thr Lys Gln Tyr Leu Asp Ala Ser Glu Val Glu Gly Thr Glu Ile
195 200 205
Arg Tyr Gly His Glu Val Lys Ser Ile Lys Ala Asp Gly Ala Lys Trp
210 215 220
Ile Val Thr Val Lys Asn Val His Thr Gly Asp Thr Lys Thr Ile Lys
225 230 235 240
Ala Asn Phe Val Phe Val Gly Ala Gly Gly Tyr Ala Leu Asp Leu Leu
245 250 255
Arg Ser Ala Gly Ile Pro Gln Val Lys Gly Phe Ala Gly Phe Pro Val
260 265 270
Ser Gly Leu Trp Leu Arg Cys Thr Asn Glu Glu Leu Ile Glu Gln His
275 280 285
Ala Ala Lys Val Tyr Gly Lys Ala Ser Val Gly Ala Pro Pro Met Ser
290 295 300
Val Pro His Leu Asp Thr Arg Val Ile Glu Gly Glu Lys Gly Leu Leu
305 310 315 320
Phe Gly Pro Tyr Gly Gly Trp Thr Pro Lys Phe Leu Lys Glu Gly Ser
325 330 335
Tyr Leu Asp Leu Phe Lys Ser Ile Arg Pro Asp Asn Ile Pro Ser Tyr
340 345 350
Leu Gly Val Ala Ala Gln Glu Phe Asp Leu Thr Lys Tyr Leu Val Thr
355 360 365
Glu Val Leu Lys Asp Gln Asp Lys Arg Met Asp Ala Leu Arg Glu Tyr
370 375 380
Met Pro Glu Ala Gln Asn Gly Asp Trp Glu Thr Ile Val Ala Gly Gln
385 390 395 400
Arg Val Gln Val Ile Lys Pro Ala Gly Phe Pro Lys Phe Gly Ser Leu
405 410 415
Glu Phe Gly Thr Thr Leu Ile Asn Asn Ser Glu Gly Thr Ile Ala Gly
420 425 430
Leu Leu Gly Ala Ser Pro Gly Ala Ser Ile Ala Pro Ser Ala Met Ile
435 440 445
Glu Leu Leu Glu Arg Cys Phe Gly Asp Arg Met Ile Glu Trp Gly Asp
450 455 460
Lys Leu Lys Asp Met Ile Pro Ser Tyr Gly Lys Lys Leu Ala Ser Glu
465 470 475 480
Pro Ala Leu Phe Glu Gln Gln Trp Ala Arg Thr Gln Lys Thr Leu Lys
485 490 495
Leu Glu Glu Ala
500
<210> 4
<211> 500
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 4
Met Ser Asp Ser Pro Lys Asn Ala Pro Arg Ile Thr Asp Glu Ala Asp
1 5 10 15
Val Val Leu Ile Gly Ala Gly Ile Met Ser Ser Thr Leu Gly Ala Met
20 25 30
Leu Arg Gln Leu Glu Pro Ser Trp Thr Gln Ile Val Phe Glu Arg Leu
35 40 45
Asp Gly Pro Ala Gln Glu Ser Ser Ser Pro Trp Asn Asn Ala Gly Thr
50 55 60
Gly His Ser Ala Leu Cys Glu Leu Asn Tyr Thr Pro Glu Val Lys Gly
65 70 75 80
Lys Val Glu Ile Ala Lys Ala Val Gly Ile Asn Glu Lys Phe Gln Val
85 90 95
Ser Arg Gln Phe Trp Ser His Leu Val Glu Glu Gly Val Leu Ala Asp
100 105 110
Ser Lys Glu Phe Ile Asn Pro Val Pro His Val Ser Phe Gly Gln Gly
115 120 125
Ala Asp Gln Val Ala Tyr Ile Lys Ala Arg Tyr Glu Ala Leu Lys Asp
130 135 140
His Pro Leu Phe Gln Gly Met Thr Tyr Ala Asp Asp Glu Ala Thr Phe
145 150 155 160
Thr Glu Lys Leu Pro Leu Met Ala Lys Gly Arg Asp Phe Ser Asp Pro
165 170 175
Val Ala Ile Ser Trp Ile Asp Glu Gly Thr Asp Ile Asn Tyr Gly Ala
180 185 190
Gln Thr Lys Gln Tyr Leu Asp Ala Ser Glu Val Glu Gly Thr Glu Ile
195 200 205
Arg Tyr Gly His Glu Val Lys Ser Ile Lys Ala Asp Gly Ala Lys Trp
210 215 220
Ile Val Thr Val Lys Asn Val His Thr Gly Asp Thr Lys Thr Ile Lys
225 230 235 240
Ala Asn Phe Val Phe Val Gly Ala Gly Gly Tyr Ala Leu Asp Leu Leu
245 250 255
Arg Ser Ala Gly Ile Pro Gln Val Lys Gly Phe Ala Gly Phe Pro Val
260 265 270
Ser Gly Leu Trp Leu Arg Cys Thr Asn Glu Glu Leu Ile Glu Gln His
275 280 285
Ala Ala Lys Val Tyr Gly Lys Ala Ser Val Gly Ala Pro Pro Met Ser
290 295 300
Val Pro His Leu Asp Thr Arg Val Ile Glu Gly Glu Lys Gly Leu Leu
305 310 315 320
Phe Gly Pro Tyr Gly Gly Trp Thr Pro Lys Phe Leu Lys Glu Gly Ser
325 330 335
Tyr Leu Asp Leu Phe Lys Ser Ile Arg Pro Asp Asn Ile Pro Ser Tyr
340 345 350
Leu Gly Val Ala Ala Gln Glu Phe Asp Leu Thr Lys Tyr Leu Val Thr
355 360 365
Glu Val Leu Lys Asp Gln Asp Lys Arg Met Asp Ala Leu Arg Glu Tyr
370 375 380
Met Pro Glu Ala Gln Asn Gly Asp Trp Glu Thr Ile Val Ala Gly Gln
385 390 395 400
Arg Val Gln Val Ile Lys Pro Ala Gly Phe Pro Lys Phe Gly Ser Leu
405 410 415
Glu Phe Gly Thr Thr Leu Ile Asn Asn Ser Glu Gly Thr Ile Ala Gly
420 425 430
Leu Leu Gly Ala Ser Pro Gly Ala Ser Ile Ala Pro Ser Ala Met Ile
435 440 445
Glu Leu Leu Glu Arg Cys Phe Gly Asp Arg Met Ile Glu Trp Gly Asp
450 455 460
Lys Leu Lys Asp Met Ile Pro Ser Tyr Gly Lys Lys Leu Ala Ser Glu
465 470 475 480
Pro Ala Leu Phe Glu Gln Gln Trp Ala Arg Thr Gln Lys Thr Leu Lys
485 490 495
Leu Glu Glu Ala
500
<210> 5
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 5
cagtgccaag cttgcatgcc tgcaggtcga ctctagaagg aatgttgtct gggcgg 56
<210> 6
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 6
ggcgtgctgg ctgattctaa ggaattcatc 30
<210> 7
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 7
gatgaattcc ttagaatcag ccagcacgcc 30
<210> 8
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 8
cagctatgac catgattacg aattcgagct cggtacccga taggtcgatt gttggtgt 58
<210> 9
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 9
catcaaaggc agcttctcgg 20
<210> 10
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 10
actacacccc agaggttaag 20
<210> 11
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 11
cagtgccaag cttgcatgcc tgcaggtcga ctctaggacc cgcttgccat acgaag 56
<210> 12
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 12
ctgaagcttg aggaagccta aatctactca tctgaagaat c 41
<210> 13
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 13
gattcttcag atgagtagat ttaggcttcc tcaagcttca g 41
<210> 14
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 14
caaaccagag tgcccacgaa atgtataacg ataggtcga 39
<210> 15
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 15
tcgacctatc gttatacatt tcgtgggcac tctggtttg 39
<210> 16
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 16
cagctatgac catgattacg aattcgagct cggtacccca taagaaacaa ccacttcc 58
<210> 17
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 17
gtccaaggtg acggccgcac 20
<210> 18
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 18
tgacttctct gatccagtag 20
<210> 19
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 19
gtactgcttg gtctgagcac 20
<210> 20
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 20
atattcggcc cagcagcagc 20
<210> 21
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 21
gcttgcatgc ctgcaggtcg actctagagg atccccttag gcttcctcaa gcttcag 57
<210> 22
<211> 53
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 22
atcaggctga aaatcttctc tcatccgcca aaacatgtat aacgataggt cga 53
<210> 23
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 23
cagtgccaag cttgcatgcc tgcaggtcga ctctaggcct gactgatttt gggctg 56
<210> 24
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 24
gagataaaag gaagttgaac atcttctaac tgctttcttt 40
<210> 25
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 25
aaagaaagca gttagaagat gttcaacttc cttttatctc 40
<210> 26
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 26
cagctatgac catgattacg aattcgagct cggtacccag atgacctcgg cagcagct 58
<210> 27
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 27
gcctgactga ttttgggctg 20
<210> 28
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 28
agatgacctc ggcagcagct 20
<210> 29
<211> 1504
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 29
cagtgccaag cttgcatgcc tgcaggtcga ctctagaagg aatgttgtct gggcggatgg 60
acttgaacag gtccaggtag gagccttcct tcaagaactt aggggtccag ccaccgtaag 120
gtccaaagag cagacccttt tcaccctcga taacgcgggt gtcaaggtga ggaacagaca 180
ttggaggagc gccaacagat gccttgccat ataccttggc tgcgtgctgc tcgatcagtt 240
cctcgttggt gcaacgaagc cacaggccgg atactgggaa tccagcgaag cccttgacct 300
gtgggatgcc tgcgctgcga agcagatcca gtgcgtaccc gcctgcgccg acgaacacga 360
agtttgcctt gatggtcttg gtgtcgccag tgtgtacgtt cttgacggtc acgatccact 420
ttgcgccatc agccttgatg ctcttgactt cgtggccata gcggatttca gtgccttcaa 480
cttcagatgc atccaggtac tgcttggtct gagcaccgta gttgatgtcg gtgccttcat 540
cgatccaaga gattgctact ggatcagaga agtcacggcc ctttgccatc aaaggcagct 600
tctcggtgaa ggtagcttca tcgtcagcgt aggtcatgcc ctggaagagt gggtgatcct 660
tcaaagcttc gtagcgagcc ttgatgtatg caacctggtc tgcgccctgg ccgaaagata 720
cgtgaggaac agggttgatg aattccttag aatcagccag cacgccctct tcaacgaggt 780
gagaccagaa ctgacgggaa acctggaact tctcgttgat tcctacagcc ttggcaattt 840
caaccttgcc cttaacctct ggggtgtagt tcagctcgca tagagcagag tggccggttc 900
ctgcattgtt ccacggggag gacgactctt gtgccggtcc atccaaacgc tcgaagacga 960
tctgagtcca gcttggctcc agctgacgca gcattgcacc cagcgtggag ctcatgatac 1020
cggcaccaat gagaactaca tctgcctcat cggtaatcct cggtgcgttc ttcggggaat 1080
ctgacatgtt caacttcctt ttatctccgc gagttctctg ccgaacacca acacaaggcg 1140
tcatctcata aatgatcgcc agcacccgaa tgcaagtgtt ggccatattt ttgactgacg 1200
cgtggggcgt tacccttctt gtttcttaca agaaagcgtg ggtacacatt gtccgtaaat 1260
atagagtgta cgcctgccac ctcgagctga ataaacaagc acgggatatt cccctttctc 1320
atgcgaattc ctcccccgtg ctaccccctc ccccaatttg caaagttagt aaatgtgaaa 1380
ttcgcaaaca tgagtagtat cgcggaagtt tcaccacgtg aacctttcag tcggctcgcg 1440
cacaccacac caacaatcga cctatcgggt accgagctcg aattcgtaat catggtcata 1500
gctg 1504
<210> 30
<211> 3445
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 30
cagtgccaag cttgcatgcc tgcaggtcga ctctaggacc cgcttgccat acgaagtcct 60
ggagaagatc tccacccgca tcaccaacga agttccagac gtaaaccgcg tggttttgga 120
cgtaacctcc aagccaccag gaaccatcga atgggagtag gccttaaatg agccttcgtt 180
aagcggcaat caccttatcg gtgattgccg ctttcccatt tctccgggtt ttctggaact 240
ttttgggcgt atgctgggaa tgatcttatt attttgattt cagaaagcag gagagaccag 300
atgagcgaaa tccttgaaac ctactgggca ccccacttcg gaaacaccga tgaagccgca 360
gcactcgttt catacttggc acaagcttcc ggtgatccta ttgaggttca caccctgttc 420
ggggatttag gtttagacgg actctctgga aactacaccg acactgagat cgacggctac 480
ggcgacgcat tcctgctggt tgcagcacta gcagtgttga tggctgaaaa caaagcatcc 540
ggcggcgtga atctgggtga agttggggga gctgataaat cgatccggct gcatgttgaa 600
tccaaggaaa acacccagat caacaccgca ttgaagtact ttgcgctttc cccagaagac 660
cacgcagcgg cagatcgctt cgatgaggat gacctgtctg agcttgccaa cttgagtgaa 720
gagctgcgcg gacagctgga ctaattgctg cccgtttaag gagtccgatt cttcagatga 780
gtagatttag gcttcctcaa gcttcagggt cttctgggtg cgtgcccact gctgctcaaa 840
cagtgctggc tcggaagcaa gcttcttgcc gtaggaaggg atcatgtcct tcagcttgtc 900
gccccactcg atcatgcggt caccgaagca acgctcaagc agctcgatca ttgcggaagg 960
tgcgatggat gctccagggg aagcaccgag caatccggcg atggtgcctt cggagttgtt 1020
gatcaaggtg gtgccgaatt ccagggaacc gaacttaggg aatcctgcag gcttaataac 1080
ctgaacacgc tgtccggcaa cgatggtctc ccaatcgccg ttttgtgcct ctggcatgta 1140
ctcgcgaaga gcatccatac gcttgtcctg gtccttgaga acttcagtga caaggtactt 1200
ggtcagatca aattcctgag cagcaacgcc aaggtaggaa ggaatgttgt ctgggcggat 1260
ggacttgaac aggtccaggt aggagccttc cttcaagaac ttaggggtcc agccaccgta 1320
aggtccaaag agcagaccct tttcaccctc gataacgcgg gtgtcaaggt gaggaacaga 1380
cattggagga gcgccaacag atgccttgcc atataccttg gctgcgtgct gctcgatcag 1440
ttcctcgttg gtgcaacgaa gccacaggcc ggatactggg aatccagcga agcccttgac 1500
ctgtgggatg cctgcgctgc gaagcagatc cagtgcgtac ccgcctgcgc cgacgaacac 1560
gaagtttgcc ttgatggtct tggtgtcgcc agtgtgtacg ttcttgacgg tcacgatcca 1620
ctttgcgcca tcagccttga tgctcttgac ttcgtggcca tagcggattt cagtgccttc 1680
aacttcagat gcatccaggt actgcttggt ctgagcaccg tagttgatgt cggtgccttc 1740
atcgatccaa gagattgcta ctggatcaga gaagtcacgg ccctttgcca tcaaaggcag 1800
cttctcggtg aaggtagctt catcgtcagc gtaggtcatg ccctggaaga gtgggtgatc 1860
cttcaaagct tcgtagcgag ccttgatgta tgcaacctgg tctgcgccct ggccgaaaga 1920
tacgtgagga acagggttga tgaattcctt aggatcagcc agcacgccct cttcaacgag 1980
gtgagaccag aactgacggg aaacctggaa cttctcgttg attcctacag ccttggcaat 2040
ttcaaccttg cccttaacct ctggggtgta gttcagctcg catagagcag agtggccggt 2100
tcctgcattg ttccacgggg aggacgactc ttgtgccggt ccatccaaac gctcgaagac 2160
gatctgagtc cagcttggct ccagctgacg cagcattgca cccagcgtgg agctcatgat 2220
accggcacca atgagaacta catctgcctc atcggtaatc ctcggtgcgt tcttcgggga 2280
atctgacatg ttcaacttcc ttttatctcc gcgagttctc tgccgaacac caacacaagg 2340
cgtcatctca taaatgatcg ccagcacccg aatgcaagtg ttggccatat ttttgactga 2400
cgcgtggggc gttacccttc ttgtttctta caagaaagcg tgggtacaca ttgtccgtaa 2460
atatagagtg tacgcctgcc acctcgagct gaataaacaa gcacgggata ttcccctttc 2520
tcatgcgaat tcctcccccg tgctaccccc tcccccaatt tgcaaagtta gtaaatgtga 2580
aattcgcaaa catgagtagt atcgcggaag tttcaccacg tgaacctttc agtcggctcg 2640
cgcacaccac accaacaatc gacctatcgt tatacatttc gtgggcactc tggtttggtt 2700
accaggatgg gttagtcatt ctgatcagcg aattccacgt tcacatcgcc aattccagag 2760
ttcacaacca gattcagcat tggaccttct agatcagcat tgtgggcggt gagatctcca 2820
acatcacagc gcgctgtgcc cacaccggcg gtacaactta ggctcacggg cacatcatcg 2880
ggcagggtga ccatgacttc gccgatccct gaggtgattt ggatgttttg ttcctgatcc 2940
aattgggtga ggtggctgaa atcgaggttc atttcaccca cgccagaggt gtagctgctg 3000
aggagttcat cgttggtggg gatgagattg acatcgccga ttccagggtc gtcttcaaag 3060
tagatgggat cgatatttga aataaacagg cctgcgaggg cgctcatgac aactccggta 3120
ccaactacac cgccgacaat ccatggccac acatggcgct ttttctgagg cttttgtgga 3180
gggacttgta catcccaggt gttgtattgg ttttgggcaa gtggatccca atgaggcgct 3240
tcgggggttt gttgcgcgaa gggtgcatag tagccctcaa cgggggtgat agtgcttaga 3300
tctggttggg gttgtgggta gagatcttcg tttttcatgg tggcatcctc agaaacagtg 3360
aattcagtgg tgagtagtcc gcggggtgga agtggttgtt tcttatgggg taccgagctc 3420
gaattcgtaa tcatggtcat agctg 3445
<210> 31
<211> 3445
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 31
cagtgccaag cttgcatgcc tgcaggtcga ctctaggacc cgcttgccat acgaagtcct 60
ggagaagatc tccacccgca tcaccaacga agttccagac gtaaaccgcg tggttttgga 120
cgtaacctcc aagccaccag gaaccatcga atgggagtag gccttaaatg agccttcgtt 180
aagcggcaat caccttatcg gtgattgccg ctttcccatt tctccgggtt ttctggaact 240
ttttgggcgt atgctgggaa tgatcttatt attttgattt cagaaagcag gagagaccag 300
atgagcgaaa tccttgaaac ctactgggca ccccacttcg gaaacaccga tgaagccgca 360
gcactcgttt catacttggc acaagcttcc ggtgatccta ttgaggttca caccctgttc 420
ggggatttag gtttagacgg actctctgga aactacaccg acactgagat cgacggctac 480
ggcgacgcat tcctgctggt tgcagcacta gcagtgttga tggctgaaaa caaagcatcc 540
ggcggcgtga atctgggtga agttggggga gctgataaat cgatccggct gcatgttgaa 600
tccaaggaaa acacccagat caacaccgca ttgaagtact ttgcgctttc cccagaagac 660
cacgcagcgg cagatcgctt cgatgaggat gacctgtctg agcttgccaa cttgagtgaa 720
gagctgcgcg gacagctgga ctaattgctg cccgtttaag gagtccgatt cttcagatga 780
gtagatttag gcttcctcaa gcttcagggt cttctgggtg cgtgcccact gctgctcaaa 840
cagtgctggc tcggaagcaa gcttcttgcc gtaggaaggg atcatgtcct tcagcttgtc 900
gccccactcg atcatgcggt caccgaagca acgctcaagc agctcgatca ttgcggaagg 960
tgcgatggat gctccagggg aagcaccgag caatccggcg atggtgcctt cggagttgtt 1020
gatcaaggtg gtgccgaatt ccagggaacc gaacttaggg aatcctgcag gcttaataac 1080
ctgaacacgc tgtccggcaa cgatggtctc ccaatcgccg ttttgtgcct ctggcatgta 1140
ctcgcgaaga gcatccatac gcttgtcctg gtccttgaga acttcagtga caaggtactt 1200
ggtcagatca aattcctgag cagcaacgcc aaggtaggaa ggaatgttgt ctgggcggat 1260
ggacttgaac aggtccaggt aggagccttc cttcaagaac ttaggggtcc agccaccgta 1320
aggtccaaag agcagaccct tttcaccctc gataacgcgg gtgtcaaggt gaggaacaga 1380
cattggagga gcgccaacag atgccttgcc atataccttg gctgcgtgct gctcgatcag 1440
ttcctcgttg gtgcaacgaa gccacaggcc ggatactggg aatccagcga agcccttgac 1500
ctgtgggatg cctgcgctgc gaagcagatc cagtgcgtac ccgcctgcgc cgacgaacac 1560
gaagtttgcc ttgatggtct tggtgtcgcc agtgtgtacg ttcttgacgg tcacgatcca 1620
ctttgcgcca tcagccttga tgctcttgac ttcgtggcca tagcggattt cagtgccttc 1680
aacttcagat gcatccaggt actgcttggt ctgagcaccg tagttgatgt cggtgccttc 1740
atcgatccaa gagattgcta ctggatcaga gaagtcacgg ccctttgcca tcaaaggcag 1800
cttctcggtg aaggtagctt catcgtcagc gtaggtcatg ccctggaaga gtgggtgatc 1860
cttcaaagct tcgtagcgag ccttgatgta tgcaacctgg tctgcgccct ggccgaaaga 1920
tacgtgagga acagggttga tgaattcctt agaatcagcc agcacgccct cttcaacgag 1980
gtgagaccag aactgacggg aaacctggaa cttctcgttg attcctacag ccttggcaat 2040
ttcaaccttg cccttaacct ctggggtgta gttcagctcg catagagcag agtggccggt 2100
tcctgcattg ttccacgggg aggacgactc ttgtgccggt ccatccaaac gctcgaagac 2160
gatctgagtc cagcttggct ccagctgacg cagcattgca cccagcgtgg agctcatgat 2220
accggcacca atgagaacta catctgcctc atcggtaatc ctcggtgcgt tcttcgggga 2280
atctgacatg ttcaacttcc ttttatctcc gcgagttctc tgccgaacac caacacaagg 2340
cgtcatctca taaatgatcg ccagcacccg aatgcaagtg ttggccatat ttttgactga 2400
cgcgtggggc gttacccttc ttgtttctta caagaaagcg tgggtacaca ttgtccgtaa 2460
atatagagtg tacgcctgcc acctcgagct gaataaacaa gcacgggata ttcccctttc 2520
tcatgcgaat tcctcccccg tgctaccccc tcccccaatt tgcaaagtta gtaaatgtga 2580
aattcgcaaa catgagtagt atcgcggaag tttcaccacg tgaacctttc agtcggctcg 2640
cgcacaccac accaacaatc gacctatcgt tatacatttc gtgggcactc tggtttggtt 2700
accaggatgg gttagtcatt ctgatcagcg aattccacgt tcacatcgcc aattccagag 2760
ttcacaacca gattcagcat tggaccttct agatcagcat tgtgggcggt gagatctcca 2820
acatcacagc gcgctgtgcc cacaccggcg gtacaactta ggctcacggg cacatcatcg 2880
ggcagggtga ccatgacttc gccgatccct gaggtgattt ggatgttttg ttcctgatcc 2940
aattgggtga ggtggctgaa atcgaggttc atttcaccca cgccagaggt gtagctgctg 3000
aggagttcat cgttggtggg gatgagattg acatcgccga ttccagggtc gtcttcaaag 3060
tagatgggat cgatatttga aataaacagg cctgcgaggg cgctcatgac aactccggta 3120
ccaactacac cgccgacaat ccatggccac acatggcgct ttttctgagg cttttgtgga 3180
gggacttgta catcccaggt gttgtattgg ttttgggcaa gtggatccca atgaggcgct 3240
tcgggggttt gttgcgcgaa gggtgcatag tagccctcaa cgggggtgat agtgcttaga 3300
tctggttggg gttgtgggta gagatcttcg tttttcatgg tggcatcctc agaaacagtg 3360
aattcagtgg tgagtagtcc gcggggtgga agtggttgtt tcttatgggg taccgagctc 3420
gaattcgtaa tcatggtcat agctg 3445
<210> 32
<211> 1961
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 32
gcttgcatgc ctgcaggtcg actctagagg atccccttag gcttcctcaa gcttcagggt 60
cttctgggtg cgtgcccact gctgctcaaa cagtgctggc tcggaagcaa gcttcttgcc 120
gtaggaaggg atcatgtcct tcagcttgtc gccccactcg atcatgcggt caccgaagca 180
acgctcaagc agctcgatca ttgcggaagg tgcgatggat gctccagggg aagcaccgag 240
caatccggcg atggtgcctt cggagttgtt gatcaaggtg gtgccgaatt ccagggaacc 300
gaacttaggg aatcctgcag gcttaataac ctgaacacgc tgtccggcaa cgatggtctc 360
ccaatcgccg ttttgtgcct ctggcatgta ctcgcgaaga gcatccatac gcttgtcctg 420
gtccttgaga acttcagtga caaggtactt ggtcagatca aattcctgag cagcaacgcc 480
aaggtaggaa ggaatgttgt ctgggcggat ggacttgaac aggtccaggt aggagccttc 540
cttcaagaac ttaggggtcc agccaccgta aggtccaaag agcagaccct tttcaccctc 600
gataacgcgg gtgtcaaggt gaggaacaga cattggagga gcgccaacag atgccttgcc 660
atataccttg gctgcgtgct gctcgatcag ttcctcgttg gtgcaacgaa gccacaggcc 720
ggatactggg aatccagcga agcccttgac ctgtgggatg cctgcgctgc gaagcagatc 780
cagtgcgtac ccgcctgcgc cgacgaacac gaagtttgcc ttgatggtct tggtgtcgcc 840
agtgtgtacg ttcttgacgg tcacgatcca ctttgcgcca tcagccttga tgctcttgac 900
ttcgtggcca tagcggattt cagtgccttc aacttcagat gcatccaggt actgcttggt 960
ctgagcaccg tagttgatgt cggtgccttc atcgatccaa gagattgcta ctggatcaga 1020
gaagtcacgg ccctttgcca tcaaaggcag cttctcggtg aaggtagctt catcgtcagc 1080
gtaggtcatg ccctggaaga gtgggtgatc cttcaaagct tcgtagcgag ccttgatgta 1140
tgcaacctgg tctgcgccct ggccgaaaga tacgtgagga acagggttga tgaattcctt 1200
aggatcagcc agcacgccct cttcaacgag gtgagaccag aactgacggg aaacctggaa 1260
cttctcgttg attcctacag ccttggcaat ttcaaccttg cccttaacct ctggggtgta 1320
gttcagctcg catagagcag agtggccggt tcctgcattg ttccacgggg aggacgactc 1380
ttgtgccggt ccatccaaac gctcgaagac gatctgagtc cagcttggct ccagctgacg 1440
cagcattgca cccagcgtgg agctcatgat accggcacca atgagaacta catctgcctc 1500
atcggtaatc ctcggtgcgt tcttcgggga atctgacatg ttcaacttcc ttttatctcc 1560
gcgagttctc tgccgaacac caacacaagg cgtcatctca taaatgatcg ccagcacccg 1620
aatgcaagtg ttggccatat ttttgactga cgcgtggggc gttacccttc ttgtttctta 1680
caagaaagcg tgggtacaca ttgtccgtaa atatagagtg tacgcctgcc acctcgagct 1740
gaataaacaa gcacgggata ttcccctttc tcatgcgaat tcctcccccg tgctaccccc 1800
tcccccaatt tgcaaagtta gtaaatgtga aattcgcaaa catgagtagt atcgcggaag 1860
tttcaccacg tgaacctttc agtcggctcg cgcacaccac accaacaatc gacctatcgt 1920
tatacatgtt ttggcggatg agagaagatt ttcagcctga t 1961
<210> 33
<211> 1961
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 33
gcttgcatgc ctgcaggtcg actctagagg atccccttag gcttcctcaa gcttcagggt 60
cttctgggtg cgtgcccact gctgctcaaa cagtgctggc tcggaagcaa gcttcttgcc 120
gtaggaaggg atcatgtcct tcagcttgtc gccccactcg atcatgcggt caccgaagca 180
acgctcaagc agctcgatca ttgcggaagg tgcgatggat gctccagggg aagcaccgag 240
caatccggcg atggtgcctt cggagttgtt gatcaaggtg gtgccgaatt ccagggaacc 300
gaacttaggg aatcctgcag gcttaataac ctgaacacgc tgtccggcaa cgatggtctc 360
ccaatcgccg ttttgtgcct ctggcatgta ctcgcgaaga gcatccatac gcttgtcctg 420
gtccttgaga acttcagtga caaggtactt ggtcagatca aattcctgag cagcaacgcc 480
aaggtaggaa ggaatgttgt ctgggcggat ggacttgaac aggtccaggt aggagccttc 540
cttcaagaac ttaggggtcc agccaccgta aggtccaaag agcagaccct tttcaccctc 600
gataacgcgg gtgtcaaggt gaggaacaga cattggagga gcgccaacag atgccttgcc 660
atataccttg gctgcgtgct gctcgatcag ttcctcgttg gtgcaacgaa gccacaggcc 720
ggatactggg aatccagcga agcccttgac ctgtgggatg cctgcgctgc gaagcagatc 780
cagtgcgtac ccgcctgcgc cgacgaacac gaagtttgcc ttgatggtct tggtgtcgcc 840
agtgtgtacg ttcttgacgg tcacgatcca ctttgcgcca tcagccttga tgctcttgac 900
ttcgtggcca tagcggattt cagtgccttc aacttcagat gcatccaggt actgcttggt 960
ctgagcaccg tagttgatgt cggtgccttc atcgatccaa gagattgcta ctggatcaga 1020
gaagtcacgg ccctttgcca tcaaaggcag cttctcggtg aaggtagctt catcgtcagc 1080
gtaggtcatg ccctggaaga gtgggtgatc cttcaaagct tcgtagcgag ccttgatgta 1140
tgcaacctgg tctgcgccct ggccgaaaga tacgtgagga acagggttga tgaattcctt 1200
agaatcagcc agcacgccct cttcaacgag gtgagaccag aactgacggg aaacctggaa 1260
cttctcgttg attcctacag ccttggcaat ttcaaccttg cccttaacct ctggggtgta 1320
gttcagctcg catagagcag agtggccggt tcctgcattg ttccacgggg aggacgactc 1380
ttgtgccggt ccatccaaac gctcgaagac gatctgagtc cagcttggct ccagctgacg 1440
cagcattgca cccagcgtgg agctcatgat accggcacca atgagaacta catctgcctc 1500
atcggtaatc ctcggtgcgt tcttcgggga atctgacatg ttcaacttcc ttttatctcc 1560
gcgagttctc tgccgaacac caacacaagg cgtcatctca taaatgatcg ccagcacccg 1620
aatgcaagtg ttggccatat ttttgactga cgcgtggggc gttacccttc ttgtttctta 1680
caagaaagcg tgggtacaca ttgtccgtaa atatagagtg tacgcctgcc acctcgagct 1740
gaataaacaa gcacgggata ttcccctttc tcatgcgaat tcctcccccg tgctaccccc 1800
tcccccaatt tgcaaagtta gtaaatgtga aattcgcaaa catgagtagt atcgcggaag 1860
tttcaccacg tgaacctttc agtcggctcg cgcacaccac accaacaatc gacctatcgt 1920
tatacatgtt ttggcggatg agagaagatt ttcagcctga t 1961
<---
Изобретение относится к области биотехнологии, в частности к белку для применения в продуцировании L-глутаминовой кислоты. Указанный белок характеризуется тем, что содержит аминокислотную последовательность SEQ ID NO: 4. Кроме того, настоящее изобретение также обеспечивает молекулу нуклеиновой кислоты, кодирующую указанный белок, рекомбинантный микроорганизм, содержащий такую молекулу, и способ его конструирования, применение нуклеиновой кислоты для продуцирования L-глутаминовой кислоты, а также способ повышения продуцирования L-глутаминовой кислоты и способ ее получения. Настоящее изобретение обеспечивает усиление способности бактерии к продуцированию L-глутаминовой кислоты. 7 н.п. ф-лы, 5 табл., 6 пр.
1. Белок для применения в продуцировании L-глутаминовой кислоты, характеризующийся тем, что он содержит аминокислотную последовательность SEQ ID NO: 4.
2. Молекула нуклеиновой кислоты для применения в продуцировании L-глутаминовой кислоты, характеризующаяся тем, что указанная молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, кодирующую белок по п. 1.
3. Рекомбинантный микроорганизм для продуцирования L-глутаминовой кислоты, содержащий молекулу нуклеиновой кислоты по п. 2, экспрессионную кассету, содержащую молекулу нуклеиновой кислоты по п. 2, или рекомбинантный вектор, содержащий молекулу нуклеиновой кислоты по п. 2.
4. Применение молекулы нуклеиновой кислоты по п. 2 для конструирования генетически модифицированной бактерии для продуцирования L-глутаминовой кислоты.
5. Способ повышения продуцирования L-глутаминовой кислоты в микроорганизме, характеризующийся тем, что он включает:
повышение уровня экспрессии или содержания молекулы нуклеиновой кислоты по п. 2 в целевом микроорганизме с получением микроорганизма, продуцирующего больше L-глутаминовой кислоты, чем целевой микроорганизм.
6. Способ конструирования рекомбинантного микроорганизма по п. 3, характеризующийся тем, что указанный способ включает:
введение молекулы нуклеиновой кислоты по п. 2 в целевой микроорганизм с получением рекомбинантного микроорганизма.
7. Способ получения L-глутаминовой кислоты, характеризующийся тем, что указанный способ включает продуцирование L-глутаминовой кислоты с использованием рекомбинантного микроорганизма по п. 3.
| US 2007141680 A1, 21.06.2007 | |||
| WO 2020135658 A1, 02.07.2020 | |||
| SHYAMKUMAR, R., MOORTHY et al., Production of L-glutamic Acid with Corynebacterium glutamicum (NCIM 2168) and Pseudomonas reptilivora (NCIM 2598): A Study on Immobilization and Reusability, Avicenna journal of medical biotechnology, 2014, 6(3), 163-168 | |||
| ШТАММ ENTEROBACTER AGGLOMERANS, СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ МИКРООРГАНИЗМА ДЛЯ ВЫРАБОТКИ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 2000 |
|
RU2261272C2 |

