СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящая заявка подана в электронном виде с помощью EFS-Web и включает поданный в электронном виде список последовательностей в формате.txt. Файл.txt содержит список последовательностей, названный "PC72377-PRV2_Sequence_Listing_ST25_05142019.txt", созданный 14 мая 2019 и имеющий размер 738 КБ. Список последовательностей, содержащийся в этом файле.txt, является частью описания и включен в настоящее описание в качестве ссылки в полном объеме.
Область изобретения
Настоящее изобретение относится к антителам, например, полноразмерным антителам или их антигенсвязывающим фрагментам, специфически связывающимся с GUCY2c (гуанилатциклазой C) и/или CD3 (кластером дифференцировки 3). Настоящее изобретение дополнительно относится к биспецифическим антителам, специфически связывающимся с CD3 и антигеном опухолевых клеток (например, биспецифическим антителам, специфически связывающимся с CD3 и GUCY2c). Настоящее изобретение также относится к родственным молекулам, например, нуклеиновым кислотам, кодирующим такие антитела или биспецифические антитела, композициям и родственным способам, например, способам получения и очистки таких антител и биспецифических антител, и их применению в диагностике и терапии.
УРОВЕНЬ ТЕХНИКИ
Злокачественные новообразования являются основной причиной смерти по всему миру, на долю которой приходится более 7 миллионов случаев смерти каждый год. Смертность от злокачественных новообразований почти всегда связана с распространением первичных опухолей до отдаленных очагов, образующих метастазы, что в конечном итоге приводит к гибели. Это особенно верно в случае злокачественных новообразований желудочно-кишечного тракта, включая аденокарциному пищевода, желудка, толстого кишечника и прямой кишки. Колоректальный рак (CRC) остается четвертым среди самых диагностируемых злокачественных новообразований и второй из основных причин гибели от злокачественных новообразований в США (Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin., 66:7-30, 2016). По всему миру на долю колоректального рака приходится 1,2 миллионов новых случаев и 600000 случаев гибели в год (Brenner H, Kloor M, Pox CP. Colorectal cancer. Lancet, 383:1490-502, 2014).
Гуанилатциклаза C (GUCY2c) (также известная как STAR, рецептор ST, GUC2C, GUCY2C, GC-C и GCC) является трансмембранным рецептором поверхности клетки, участвующим в поддержании жидкости кишечника, гомеостаза электролитов и пролиферации клеток (Carrithers et al., Proc Natl Acad Sci USA 100: 3018-3020, 2003; Mann et al., Biochem Biophys Res Commun 239: 463-466, 1997; Pitari et al., Proc Natl Acad Sci USA 100: 2695-2699, 2003); регистрационный номер GenBank NM.sub.-004963, и регистрационный номер GenPept NP-004954). Эта функция опосредована связыванием гуанилина (Wiegand et al. FEBS Lett. 311:150-154, 1992) и урогуанилина (Hamra et al. Proc Natl Acad Sci USA 9(22):10464-10468, 1993). GUCY2c также является рецептором термостабильного энтеротоксина (ST), являющегося пептидом, продуцируемым E. coli, а также другими организмами-возбудителями инфекций (Rao, M. C. Ciba Found. Symp. 112:74-93, 1985; Knoop F. C. and Owens, M. J. Pharmacol. Toxicol. Methods 28:67-72, 1992). Связывание ST с GUCY2c активирует сигнальный каскад, приводящий к заболеваниям кишечника, например, диареи.
GUCY2c охарактеризована как белок, участвующий в развитии злокачественных новообразований, включая колоректальный рак, рак поджелудочной железы, рак желудка, рак печени и рак пищевода (Carrithers et al., Dis Colon Rectum 39:171-181, 1996; Buc et al. Eur J Cancer 41: 1618-1627, 2005; Carrithers et al., Gastroenterology 107: 1653-1661, 1994; Urbanski et al., Biochem Biophys Acta 1245: 29-36, 1995).
Как белок поверхности клетки, GUCY2c может служить в качестве терапевтической мишени для рецептор-связывающих белков, таких как антитела или лиганды. GUCY2c экспрессируется на апикальной стороне эпителиальных клеток, выстилающих слизистую оболочку тонкого кишечника, толстого кишечника и прямой кишки (Carrithers et al., Dis Colon Rectum 39: 171-181, 1996). Экспрессия GUCY2c сохраняется после злокачественного перерождения эпителиальных клеток кишечника с экспрессией во всех первичных и метастатических колоректальных опухолях (Carrithers et al., 1996; Buc et al.; Carrithers et al., 1994). Экспрессия GUCY2c также определена в клетках пищевода, диагностируемых при пищеводе Барретта, раке пищевода и раке желудка.
Сохраняется потребность в молекулах и/или композициях, которые могут специфически направленно воздействовать и специфически связываться с метастатическими клетками колоректального рака. Существует потребность в улучшенных способах лечения индивидуумов, которые, как предполагают, страдают колоректальным раком, в особенности, индивидуумов, которые, как предполагают, страдают метастазированием клеток колоректального рака.
Разработаны различные стратегии получения биспецифических антител, которые могут рекрутировать T-клетки для опосредования уничтожения опухолевых клеток. Такие биспецифические антитела могут формировать мостик между опухолевой клеткой и эффекторной клеткой иммунной системы человека (NK-клетками, T-клетками, моноцитами, макрофагами или гранулоцитами), таким образом, делая возможным специфическое уничтожение опухолевой клетки. Хотя доказано, что такие биспецифические антитела предпочтительны для терапевтического использования, например, для терапевтических концепций в случае лечения опухолей, сохраняется потребность в биспецифических антителах, которые будут нацелены на GUCY2c-положительные опухолевые клетки, с высокой активностью и специфичностью. Такая молекула будет полезной в качестве терапевтического средства в случае злокачественных новообразований ЖКТ, при лечении злокачественных новообразований и детекции GUCY2c на клетках.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам, включая биспецифические антитела, где антитела специфически связываются с GUCY2c. Изобретение дополнительно относится к биспецифическим антителам, которые могут связываться с GUCY2c и CD3.
В одном из аспектов настоящее изобретение относится к антителу, специфически связывающемуся с гуанилатциклазой C (GUCY2c), где антитело содержит: (a) вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область VH один (CDR1 VH), определяющую комплементарность область VH два (CDR2 VH) и определяющую комплементарность область VH три (CDR3 VH) последовательности VH, приведенной в SEQ ID NO: 11, 19, 26, 33, 41, 48, 52, 57, 60, 62, 64, 65, 67, 69, 71 или 73; и/или (b) вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область VL один (CDR1 VL), определяющую комплементарность область VL два (CDR2 VL) и определяющую комплементарность область VL три (CDR3 VL) последовательности VL, приведенной в SEQ ID NO: 92, 100, 104, 106, 112, 119, 125, 129, 134, 136, 137, 138, 140, 143, 145, 147, 150, 152, 156, 158, 160, 162, 166, 170, 171, 172, 173, 174 или 175.
В одном из вариантов осуществления настоящее изобретение относится к антителу, специфически связывающемуся с GUCY2c, где антитело содержит: (a) область VH, содержащую (i) CDR1 VH, содержащую последовательность SEQ ID NO: 12, 20, 27, 34, 42, 74, 257, 258, 259, 260 или 261; (ii) CDR2 VH, содержащую последовательность SEQ ID NO: 13, 21, 28, 35, 43, 53, 66, 68, 70, 72, 75, 262, 263, 264, 265, 266 или 267; и (iii) CDR3 VH, содержащую последовательность SEQ ID NO: 14, 22, 29, 36 или 44; и/или (b) область VL, содержащую (i) CDR1 VL, содержащую последовательность SEQ ID NO: 93, 101, 105, 107, 113, 120, 148, 153, 163 или 167; (ii) CDR2 VL, содержащую последовательность SEQ ID NO: 78, 94, 102, 108, 114, 141, 144, 146, 149, 151, 157, 159, 161, 164 или 168; и (iii) CDR3 VL, содержащую последовательность SEQ ID NO: 95, 109, 115, 121, 142, 154, 165 или 169.
В одном из вариантов осуществления настоящее изобретение относится к антителу, где: (a) область VH содержит (i) CDR1 VH, содержащую последовательность SEQ ID NO: 74; (ii) CDR2 VH, содержащую последовательность SEQ ID NO: 75; и (iii) CDR3 VH, содержащую последовательность SEQ ID NO: 29; и/или (b) область VL содержит (i) CDR1 VL, содержащую последовательность SEQ ID NO: 148; (ii) CDR2 VL, содержащую последовательность SEQ ID NO: 149; и (iii) CDR3 VL, содержащую последовательность SEQ ID NO: 142.
В другом варианте осуществления настоящее изобретение относится к антителу, где: (a) область VH содержит SEQ ID NO: 11, 19, 26, 33, 41, 48, 52, 57, 60, 62, 64, 65, 67, 69, 71 или 73; и/или (b) область VL содержит SEQ ID NO: 92, 100, 104, 106, 112, 119, 125, 129, 134, 136, 137, 138, 140, 143, 145, 147, 150, 152, 156, 158, 160, 162, 166, 170, 171, 172, 173, 174 или 175.
В еще одном варианте осуществления настоящее изобретение относится к антителу, где область VH содержит последовательность SEQ ID NO: 73; и где область VL содержит последовательность SEQ ID NO: 147.
В одном из вариантов осуществления настоящее изобретение относится к антителу, где область VH содержит последовательность SEQ ID NO: 73 или ее вариант с одной или несколькими консервативными аминокислотными заменами в остатках, не находящихся в области VH; и/или где область VL содержит последовательность SEQ ID NO: 147 или ее вариант с одной или несколькими консервативными аминокислотными заменами в аминокислотах, не находящихся в области VL.
В одном из вариантов осуществления настоящее изобретение относится к антителу, связывающемуся с GUCY2c и содержащему: CDR1 VL против GUCY2c, содержащую аминокислоты, имеющие последовательность RASESV-XL1.30-XL1.30a-YG-XL1.30d-SLLQ; CDR2 VL против GUCY2c, содержащую аминокислоты, имеющие последовательность, приведенную в SEQ ID NO: 149; CDR3 VL против GUCY2c, содержащую аминокислоты, имеющие последовательность, приведенную в SEQ ID NO: 142; CDR1 VH против GUCY2c, содержащую аминокислоты, имеющие последовательность GFTFS-XH1.31-XH1.32-WMH; CDR2 VH против GUCY2c, содержащую аминокислоты, имеющие последовательность EIK- XH2.52A-XH2.53-XH2.54-XH2.55-XH2.56-XH2.57-NVHEKFKD; и CDR3 VH против GUCY2c, содержащую аминокислоты, имеющие последовательность T-XH3.96-XH3.97-XH3.98-XH3.99-XH3.100-G-XH3.100B-WF-XH3.100E-XH3.101-V, где каждый из XL1.30, XL1.30a, XL1.30d, XH1.31, XH1.32, XH2.52A, XH2.53, XH2.54, XH2.55, XH2.56, XH2.57, XH3.96, XH3.97, XH3.98, XH3.99, XH3.100, XH3.100B, XH3.100E и XH3.101 независимо представляет собой аминокислотный остаток из столбцов 4 и 5 таблиц 42A и 42B.
В одном из аспектов XL1.30 является D, N или S; XL1.30a является Y, W или I; XL1.30d является T, S или H; XH1.31 является S, R, W, Y, A, H, P, Y, T, N, K, D, G или V; XH1.32 является Y, R, L, T, K, P, I, N, M, V или S; XH2.52A является P, T или V; XH2.53 является S, A, L или R; XH2.54 является N, T, R, H, K, M, S, A, Y, T или I; XH2.55 является E, R, K, N, Y, G, L, A, M, S, H, D или Q; XH2.56 является L, W, Y, F, V, I, N или H; XH2.57 является T, M, S, L, N, Q или V; XH3.96 является I, F или K; XH3.97 является T, V, L, I, M, F, Y или A; XH3.98 является T, N, R, G, L или I; XH3.99 является T, K, L, W, A, S, M, P, N или R; XH3.100 является E, G, A, H, S, D, T, R, Q, K, Y, L или M; XH3.100B является Y или H; XH3.100E является F или L; и XH3.101 является D, Y, E или S.
В одном из вариантов осуществления настоящее изобретение относится к антителу, где антитело выбрано из группы, состоящей из Fab-фрагмента, Fab'-фрагмента, F(ab’)2-фрагмента, Fd-фрагмента, Fv-фрагмента, одноцепочечного Fv-фрагмента (scFv), стабилизированного дисульфидными связями Fv-фрагмента (dsFv), однодоменного антитела (dAb), моноклонального антитела, химерного антитела, биспецифического антитела, триспецифического антитела, полиспецифического антитела, биспецифического гетеродимерного диатело, биспецифического гетеродимерного IgG, поликлонального антитела, меченого антитела, гуманизированного антитела, антитела человека и их фрагментов.
В другом варианте осуществления антитело по настоящему изобретению дополнительно содержит человеческий или гуманизированный каркас VH и человеческий или гуманизированный каркас VL. В некоторых вариантах осуществления каркас VH содержит последовательность SEQ ID NO: 5, 6, 7, 8, 15, 16, 17, 18, 23, 24, 25, 30, 31, 32, 37, 38, 39, 40, 45, 46, 47, 49, 50, 51, 54, 55, 56, 58, 59, 61 или 63; и/или каркас VL содержит последовательность SEQ ID NO: 80, 81, 82, 83, 86, 87, 88, 89, 96, 97, 98, 99, 103, 110, 111, 116, 117, 118, 122, 123, 124, 126, 127, 128, 130, 131, 132, 133, 135, 139 или 155.
В другом варианте осуществления настоящее изобретение относится к выделенному моноклональному антителу человека, связывающемуся с эпитопом на внеклеточном домене GUCY2c, где эпитоп содержит по меньшей мере один аминокислотный остаток, выбранный из аминокислотных остатков R73, S74, S75, T76, E78, G79, L80, L82, L83, R84 или I86 SEQ ID NO: 406.
В одном из аспектов эпитоп содержит по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять или по меньшей мере десять аминокислотных остатков, выбранных из аминокислотных остатков R73, S74, S75, T76, E78, G79, L80, L82, L83, R84 или I86 SEQ ID NO: 406. В другом аспекте эпитоп содержит аминокислотные остатки R73, S74, S75, T76, E78, G79, L80, L82, L83, R84 или I86 SEQ ID NO: 406. В дополнительном аспекте эпитоп содержит аминокислоты, имеющие последовательность, приведенную в SEQ ID NO: 406. В дополнительном аспекте эпитоп является функциональным эпитопом.
В другом аспекте выделенное антитело человека и контакты аминокислот эпитопа GUCY2c находятся в пределах 3,8 ангстрем, что определяют посредством кристаллографии. В рамках изобретения термин "в пределах 3,8 ангстрем" означает, что контакты составляют 3,8 ангстрем или менее.
В одном из аспектов антитело по настоящему изобретению является биспецифическим антителом. В другом аспекте настоящее изобретение относится к биспецифическому антителу, специфически связывающемуся с GUCY2c и CD3, где биспецифическое антитело содержит первую полипептидную цепь и вторую полипептидную цепь.
В одном из вариантов осуществления (a) первая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 1, содержащий VL антитела против GUCY2c (VL против GUCY2c) и VH антитела против CD3 (VH против CD3), и (ii) первый способствующий гетеродимеризации домен; и (b) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 2, содержащий VL антитела против CD3 (VL против CD3) и VH антитела против GUCY2c (VH против GUCY2c), и (ii) второй способствующий гетеродимеризации домен; где VL против GUCY2c и VH против GUCY2c образуют домен, специфически связывающийся с GUCY2c; и VL против CD3 и VH против CD3 образуют домен, специфически связывающийся с CD3.
В другом варианте осуществления (a) первая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 1, содержащий VL против CD3 и VH против GUCY2c, и (ii) первый способствующий гетеродимеризации домен; и (b) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 2, содержащий VL против GUCY2c и VH против CD3, и (ii) второй способствующий гетеродимеризации домен; где VL против GUCY2c и VH против GUCY2c образуют домен, специфически связывающийся с GUCY2c; и VL против CD3 и VH против CD3 образуют домен, специфически связывающийся с CD3.
В другом варианте осуществления (a) первая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 1, содержащий VH против GUCY2c и VL против CD3, и (ii) первый способствующий гетеродимеризации домен; и (b) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 2, содержащий VH против CD3 и VL против GUCY2c, и (ii) второй способствующий гетеродимеризации домен; где VH против GUCY2c и VL против GUCY2c образуют домен, специфически связывающийся с GUCY2c; и VH против CD3 и VL против CD3 образуют домен, специфически связывающийся с CD3.
В дополнительном варианте осуществления (a) первая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 1, содержащий VH против CD3 и VL против GUCY2c, и (ii) первый способствующий гетеродимеризации домен; и (b) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 2, содержащий VH против GUCY2c и VL против CD3, и (ii) второй способствующий гетеродимеризации домен, где VL против GUCY2c и VH против GUCY2c образуют домен, специфически связывающийся с GUCY2c; и VL против CD3 и VH против CD3 образуют домен, специфически связывающийся с CD3.
В некоторых вариантах осуществления описанного выше каждый из первого способствующего гетеродимеризации домена и второго способствующего гетеродимеризации домена содержит домен CH2 и домен CH3, где аминокислотную последовательность каждого из домена CH2 и/или домена CH3 модифицируют для регуляции гетеродимеризации и/или стабилизации биспецифического антитела.
В некоторых таких вариантах осуществления аминокислотная последовательность домена CH2 и/или домена CH3 содержит по меньшей мере одну аминокислотную модификацию, где: (a) домен CH3 первого способствующего гетеродимеризации домена образует выступ; и (b) домен CH3 второго способствующего гетеродимеризации домена образует впадину.
В другом таком варианте осуществления домен CH3 первого способствующего гетеродимеризации домена содержит мутации Y349C и/или T366W; и домен CH3 второго способствующего гетеродимеризации домена содержит мутации S354C, T366S, L368A и/или Y407V (нумерация в соответствии с индексом EU).
В конкретном варианте осуществления описанного выше первый способствующий гетеродимеризации домен содержит последовательность SEQ ID NO: 188; где второй способствующий гетеродимеризации домен содержит последовательность SEQ ID NO: 189.
В дополнительных вариантах осуществления VL против GUCY2c и VH против CD3 соединяют посредством глицин-серинового линкера; и VL против CD3 и VH против GUCY2c соединяют посредством глицин-серинового линкера. В некоторых таких вариантах осуществления глицин-сериновый линкер является линкером 1, содержащим последовательность SEQ ID NO: 190. В других вариантах осуществления глицин-серин линкер является линкером 2, содержащим последовательность SEQ ID NO: 191.
В дополнительных вариантах осуществления описанного выше домен 1 ковалентно связан с первым способствующим гетеродимеризации доменом посредством цистеинового линкера; и домен 2 ковалентно связан со вторым способствующим гетеродимеризации доменом посредством цистеинового линкера; где цистеиновый линкер содержит по меньшей мере пять аминокислот. В некоторых таких вариантах осуществления цистеиновый линкер является линкером 3, содержащим последовательность SEQ ID NO: 192.
В некоторых вариантах осуществления первая полипептидная цепь ковалентно связана со второй полипептидной цепью посредством по меньшей мере одной дисульфидной связи. В некоторых таких вариантах осуществления по меньшей мере одна дисульфидная связь образуется между линкером 3 первой полипептидной цепи и линкером 3 второй полипептидной цепи. В другом таком варианте осуществления по меньшей мере одна дисульфидная связь образуется между первым способствующим гетеродимеризации доменом и вторым способствующий гетеродимеризации доменом. В конкретных вариантах осуществления, каждая дисульфидная связь образуется посредством связывания двух остатков цистеина.
В дополнительных вариантах осуществления любого из описанных выше биспецифическое антитело по настоящему изобретению содержит (a) CDR1 VH против GUCY2c, содержащую последовательность SEQ ID NO: 12, 20, 27, 34, 42, 74, 257, 258, 259, 260 или 261; (b) CDR2 VH против GUCY2c, содержащую последовательность SEQ ID NO: 13, 21, 28, 35, 43, 53, 66, 68, 70, 72, 75, 262, 263, 264, 265, 266 или 267; (c) CDR3 VH против GUCY2c, содержащую последовательность SEQ ID NO: 14, 22, 29, 36 или 44; (d) CDR1 VL против GUCY2c, содержащую последовательность SEQ ID NO: 93, 101, 105, 107, 113, 120, 148, 153, 163 или 167; (e) CDR2 VL против GUCY2c, содержащую последовательность SEQ ID NO: 78, 94, 102, 108, 114, 141, 144, 146, 149, 151, 157, 159, 161, 164 или 168; и (f) CDR3 VL против GUCY2c, содержащую последовательность SEQ ID NO: 95, 109, 115, 121, 142, 154, 165 или 169.
В конкретном варианте осуществления биспецифическое антитело по настоящему изобретению содержит (a) CDR1 VH против GUCY2c, содержащую последовательность SEQ ID NO: 74 или 259; (b) CDR2 VH против GUCY2c, содержащую последовательность SEQ ID NO: 75 или 267; (c) CDR3 VH против GUCY2c, содержащую последовательность SEQ ID NO: 29; (d) CDR1 VL против GUCY2c, содержащую последовательность SEQ ID NO: 148; (e) CDR2 VL против GUCY2c, содержащую последовательность SEQ ID NO: 149; и (f) CDR3 VL против GUCY2c, содержащую последовательность SEQ ID NO: 142.
В конкретных вариантах осуществления, биспецифическое антитело содержит: (a) область VH против GUCY2c, содержащую последовательность SEQ ID NO: 73; и (b) область VL против GUCY2c, содержащую последовательность, приведенную в SEQ ID NO: 147.
В дополнительных вариантах осуществления настоящее изобретение относится к биспецифическим антителам, содержащим (a) CDR1 VH против CD3, содержащую последовательность SEQ ID NO: 2, 268 или 277; (b) CDR2 VH против CD3, содержащую последовательность SEQ ID NO: 3, 10, 269 или 270; (c) CDR3 VH против CD3, содержащую последовательность SEQ ID NO: 4; (d) CDR1 VL против CD3, содержащую последовательность SEQ ID NO: 77, 85, 91, 278, 279 или 280; (e) CDR2 VL против CD3, содержащую SEQ ID NO: 78, или 281; и (f) CDR3 VL против CD3, содержащую последовательность SEQ ID NO: 79.
В конкретном варианте осуществления настоящее изобретение относится к биспецифическому антителу, содержащему (a) CDR1 VH против CD3, содержащую последовательность SEQ ID NO: 2 или 268; (b) CDR2 VH против CD3, содержащую последовательность SEQ ID NO: 10 или 270; (c) CDR3 VH против CD3, содержащую последовательность SEQ ID NO: 4; (d) CDR1 VL против CD3, содержащую последовательность SEQ ID NO: 91; (e) CDR2 VL против CD3, содержащую последовательность SEQ ID NO: 78; и (f) CDR3 VL против CD3, содержащую последовательность SEQ ID NO: 79.
В одном из вариантов осуществления настоящее изобретение относится к биспецифическому антителу, содержащему: (a) область VH против CD3, содержащую последовательность SEQ ID NO: 1, 9 или 273; и (b) область VL против CD3, содержащую последовательность SEQ ID NO: 76, 84, 90, 274, 275 или 276. В конкретном варианте осуществления биспецифическое антитело по настоящему изобретению содержит: (a) область VH против CD3, содержащую последовательность SEQ ID NO: 9; и (b) область VL против CD3, содержащую последовательность SEQ ID NO: 90.
В одном из вариантов осуществления настоящее изобретение относится к биспецифическому антителу, содержащему: CDR1 VL против GUCY2c, содержащую аминокислоты, имеющие последовательность RASESV-XL1.30-XL1.30a-YG-XL1.30d-SLLQ; CDR2 VL против GUCY2c, содержащую аминокислоты, имеющие последовательность, приведенную в SEQ ID NO: 149; CDR3 VL против GUCY2c, содержащую аминокислоты, имеющие последовательность, приведенную в SEQ ID NO: 142; CDR1 VH против GUCY2c, содержащую аминокислоты, имеющие последовательность GFTFS-XH1.31-XH1.32-WMH; CDR2 VH против GUCY2c, содержащую аминокислоты, имеющие последовательность EIK-XH2.52A-XH2.53-XH2.54-XH2.55-XH2.56-XH2.57-NVHEKFKD; и CDR3 VH против GUCY2c, содержащую аминокислоты, имеющие последовательность T-XH3.96-XH3.97-XH3.98-XH3.99-XH3.100-G-XH3.100B-WF-XH3.100E-XH3.101-V, где каждый из XL1.30, XL1.30a, XL1.30d, XH1.31, XH1.32, XH2.52A, XH2.53, XH2.54, XH2.55, XH2.56, XH2.57, XH3.96, XH3.97, XH3.98, XH3.99, XH3.100, XH3.100B, XH3.100E и XH3.101 независимо представляет собой аминокислотный остаток в соответствии со столбцами 4 и 5 таблиц 42A и 42B.
В одном из аспектов XL1.30 является D, N или S; XL1.30a является Y, W или I; XL1.30d является T, S или H; XH1.31 является S, R, W, Y, A, H, P, Y, T, N, K, D, G или V; XH1.32 является Y, R, L, T, K, P, I, N, M, V или S; XH2.52A является P, T или V; XH2.53 является S, A, L или R; XH2.54 является N, T, R, H, K, M, S, A, Y, T или I; XH2.55 является E, R, K, N, Y, G, L, A, M, S, H, D или Q; XH2.56 является L, W, Y, F, V, I, N или H; XH2.57 является T, M, S, L, N, Q или V; XH3.96 является I, F или K; XH3.97 является T, V, L, I, M, F, Y или A; XH3.98 является T, N, R, G, L или I; XH3.99 является T, K, L, W, A, S, M, P, N или R; XH3.100 является E, G, A, H, S, D, T, R, Q, K, Y, L или M; XH3.100B является Y или H; XH3.100E является F или L; и XH3.101 является D, Y, E или S.
В другом варианте осуществления настоящее изобретение относится к биспецифическому антителу, связывающемуся с эпитопом на внеклеточном домене GUCY2c, где эпитоп содержит по меньшей мере один аминокислотный остаток, выбранный из аминокислотных остатков R73, S74, S75, T76, E78, G79, L80, L82, L83, R84 или I86 SEQ ID NO: 406.
В одном из аспектов эпитоп содержит по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять или по меньшей мере десять аминокислотных остатков, выбранных из аминокислотных остатков R73, S74, S75, T76, E78, G79, L80, L82, L83, R84 или I86 SEQ ID NO: 406. В другом аспекте эпитоп содержит аминокислотные остатки R73, S74, S75, T76, E78, G79, L80, L82, L83, R84 или I86 SEQ ID NO: 406. В дополнительном аспекте эпитоп содержит аминокислоты, имеющие последовательность, приведенную в SEQ ID NO: 406. В дополнительном аспекте эпитоп является функциональным эпитопом.
В другом аспекте выделенное антитело человека и контакты аминокислот эпитопа GUCY2c находятся в пределах 3,8 ангстрем, что определяют посредством кристаллографии.
В одном из аспектов настоящее изобретение относится к биспецифическому антителу, специфически связывающемуся с GUCY2c и конкурирующему за связывание с биспецифическим антителом, представленным в настоящем описании.
В одном из аспектов настоящее изобретение относится к биспецифическому антителу, специфически связывающемуся с GUCY2c и CD3, где биспецифическое антитело содержит первую полипептидную цепь и вторую полипептидную цепь, и где: (a) первая полипептидная цепь содержит следующие области в следующем порядке в направлении от N-конца к C-концу: VL антитела против GUCY2c (VL против GUCY2c) (SEQ ID NO: 147) - линкер 1 (SEQ ID NO: 190) - VH антитела против CD3 (VH против CD3) (SEQ ID NO: 9) - линкер 3 (SEQ ID NO: 192) - первый способствующий гетеродимеризации домен (SEQ ID NO: 188); и (b) вторая полипептидная цепь содержит следующие области в следующем порядке в направлении от N-конца к C-концу: VL антитела против CD3 (VL против CD3) (SEQ ID NO: 90) - линкер 2 (SEQ ID NO: 191) - VH антитела против GUCY2c (VH против GUCY2c) (SEQ ID NO: 73) - линкер 3 (SEQ ID NO: 192) - второй способствующий гетеродимеризации домен (SEQ ID NO: 189); где VL против GUCY2c и VH против GUCY2c образуют домен, специфически связывающийся с GUCY2c; и VL против CD3 и VH против CD3 образуют домен, специфически связывающийся с CD3; где линкер 3 первой полипептидной цепи и линкер 3 второй полипептидной цепи ковалентно связаны друг с другом двумя дисульфидными связями; где каждый из первого способствующего гетеродимеризации домена и второго способствующего гетеродимеризации домена содержит домен CH2 и домен CH3; где домен CH3 первого способствующего гетеродимеризации домена образует выступ, и домен CH3 второго способствующего гетеродимеризации домена образует впадину; где по меньшей мере одна дисульфидная связь образуется между доменом CH3 первого способствующего гетеродимеризации домена и доменом CH3 второго способствующего гетеродимеризации домена.
В некоторых вариантах осуществления настоящее изобретение относится к биспецифическому антителу, дополнительно содержащему человеческий или гуманизированный каркас VH и человеческий или гуманизированный каркас VL. В некоторых таких вариантах осуществления биспецифическое антитело является гуманизированным антителом. В конкретных вариантах осуществления каркас VH содержит последовательность SEQ ID NO: 5, 6, 7, 8, 15, 16, 17, 18, 23, 24, 25, 30, 31, 32, 37, 38, 39, 40, 45, 46, 47, 49, 50, 51, 54, 55, 56, 58, 59, 61 или 63; и/или каркас VL содержит последовательность SEQ ID NO: 80, 81, 82, 83, 86, 87, 88, 89, 96, 97, 98, 99, 103, 110, 111, 116, 117, 118, 122, 123, 124, 126, 127, 128, 130, 131, 132, 133, 135, 139 или 155.
В одном из аспектов биспецифическое антитело по настоящему изобретению специфически связывается с GUCY2c и CD3, где биспецифическое антитело содержит первую полипептидную цепь и вторую полипептидную цепь, где первую полипептидную цепь получают с помощью экспрессирующего вектора с регистрационным номером ATCC PTA-124944 и вторую полипептидную цепь получают с помощью экспрессирующего вектора с регистрационным номером ATCC PTA-124943.
В конкретных вариантах осуществления антитело или биспецифическое антитело по настоящему изобретению (a) связывается с внеклеточным доменом GUCY2c человека; (b) демонстрирует увеличенное время полужизни в сыворотке или опухоли от 30 мин до 100 дней; и/или (c) демонстрирует более низкое значение EC50 от 0,0001 нМ до 100 нМ в присутствии повышенных уровней экспрессии GUCY2c или повышенных уровней плотности рецептора.
Настоящее изобретение дополнительно относится к фармацевтической композиции, содержащей терапевтически эффективное количество антитела или биспецифического антитела, представленного в настоящем описании, и фармацевтически приемлемый носитель.
В другом аспекте настоящее изобретение относится к способу лечения GUCY2c-ассоциированного нарушения у нуждающегося в этом пациента, включающему введение пациенту антитела против GUCY2c по настоящему изобретению. Настоящее изобретение также относится к способу лечения GUCY2c-ассоциированного нарушения у нуждающегося в этом пациента, включающему введение пациенту биспецифического антитела по настоящему изобретению. Настоящее изобретение дополнительно относится к способу лечения GUCY2c-ассоциированного нарушения у нуждающегося в этом пациента, включающему введение пациенту фармацевтической композиции, содержащей антитело против GUCY2c или биспецифическое антитело, представленное в настоящем описании. В некоторых таких вариантах осуществления GUCY2c-ассоциированное нарушение является злокачественным новообразованием. В конкретных вариантах осуществления злокачественное новообразование является злокачественным новообразованием пищеварительной системы, выбранным из группы, состоящей из злокачественного новообразования пищевода, желудка, тонкого кишечника, толстого кишечника, прямой кишки, анального канала, печени, желчного пузыря, аппендикса, желчных протоков и поджелудочной железы.
Настоящее изобретение дополнительно относится к способу лечения GUCY2c-ассоциированного нарушения у нуждающегося в этом пациента, включающему введение пациенту биспецифического антитела, представленного в настоящем описании, или фармацевтической композиции, содержащей биспецифическое антитело, представленное в настоящем описании, где ответ цитолитических T-клеток активирован.
В одном из аспектов настоящее изобретение относится к антителу, биспецифическому антителу или фармацевтической композиции, представленной в настоящем описании, для применения в терапии. Настоящее изобретение дополнительно относится к антителу или биспецифическому антителу, представленному в настоящем описании, для применения в производстве лекарственного средства для применения в терапии. В некоторых вариантах осуществления терапия представляет собой лечение GUCY2c-ассоциированного нарушения. В конкретных вариантах осуществления GUCY2c-ассоциированное нарушение является злокачественным новообразованием. В некоторых вариантах осуществления злокачественное новообразование является злокачественным новообразованием пищеварительной системы, выбранным из группы, состоящей из злокачественного новообразования пищевода, желудка, тонкого кишечника, толстого кишечника, прямой кишки, анального канала, печени, желчного пузыря, аппендикса, желчных протоков и поджелудочной железы. В конкретных вариантах осуществления с помощью терапии активируют ответ цитолитических T-клеток.
В одном из аспектов настоящее изобретение относится к полинуклеотиду, кодирующему антитело или биспецифическое антитело, представленное в настоящем описании. В другом варианте осуществления настоящее изобретение относится к вектору, содержащему полинуклеотиды, представленные в настоящем описании. В еще одном варианте осуществления настоящее изобретение относится к клетке-хозяину, содержащей векторы, представленные в настоящем описании. В некоторых таких вариантах осуществления клетка-хозяин рекомбинантно продуцирует антитело или биспецифическое антитело, представленное в настоящем описании. В конкретных вариантах осуществления клетка-хозяин выбрана из группы, состоящей из линий бактериальных клеток, линий клеток млекопитающего, линий клеток насекомых и линий дрожжевых клеток. В конкретном варианте осуществления линия клеток млекопитающего является линией клеток CHO. В одном из вариантов осуществления антитело или биспецифическое антитело получают с использованием бесклеточной системы синтеза белка in vitro.
В одном из аспектов настоящее изобретение относится к способу получения антитела против GUCY2c или биспецифического антитела, представленного в настоящем описании, включающему культивирование клетки-хозяина в условиях, приводящих к продукции антитела против GUCY2c или биспецифического антитела, представленного в настоящем описании, и очистку антитела или биспецифического антитела от супернатанта культуры.
В другом аспекте настоящее изобретение относится к применению антитела против GUCY2c, биспецифического антитела, фармацевтической композиции, полинуклеотида, вектора или клетки-хозяина, представленных в настоящем описании, в производстве лекарственного средства для лечения GUCY2c-ассоциированного нарушения.
В другом варианте осуществления настоящее изобретение относится к композиции, содержащей биспецифические антитела по изобретению и второе терапевтическое средство.
В другом варианте осуществления настоящее изобретение относится к композиции, содержащей биспецифические антитела по изобретению и противодиарейное средство.
В одном из аспектов противодиарейное средство включает, в качестве неограничивающих примеров, субгаллат висмута, Lactobacillus acidophilus, Saccharomyces boulardii, лоперамид/симетикон, атропин/дифеноксилат, атропин/ дифеноксин, лиофилизат Saccharomyces boulardii, Lactobacillus acidophilus, лоперамид, субсалицилат висмута, Lactobacillus acidophilus/Lactobacillus bulgaricus, Lactobacillus rhamnosus, аттапульгит, крофелемер, фторхинолон, антибиотик или октреотид.
В другом аспекте настоящее изобретение относится к применению композиции, содержащей биспецифическое антитело против GUCY2c и противодиарейное средство, представленное в настоящем описании, в производстве лекарственного средства для лечения GUCY2c-ассоциированного нарушения.
Другие варианты осуществления будут очевидны из следующего подробного описания. Если аспекты или варианты осуществления изобретения описаны в терминах группы Маркуша или другого группирования альтернатив, настоящее изобретение относится не только ко всей группе, указанной в целом, но также и к каждому члену группы по отдельности и всем возможным подгруппам основной группы, а также к основной группе, в которой отсутствует один или более из членов группы. Настоящее изобретение также предусматривает конкретное исключение одного или более из любых членов групп в описываемом в заявке изобретении.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фигурах 1A, 1B, 1C и 1D показаны схемы четырех альтернативных представлений биспецифических антител против CD3-GUCY2c, содержащих первый способствующий гетеродимеризации домен и второй способствующий гетеродимеризации домен, содержащий Fc-цепь, оптимизированную для связывания с помощью "выступа-во-впадину".
На фигуре 2 показано связывание биспецифических антител (A) GUCY2C-0247 и (B) GUCY2C-1608 с опухолевыми клетками T84 с использованием анализ на основе проточной цитометрии.
На фигуре 3 показан анализ на основе проточной цитометрии для определения связывания биспецифических антител (A) GUCY2C-0247 и (B) GUCY2C-1608 с наивными T-клетками человека.
На фигуре 4A показаны GUCY2C-1608 и GUCY2C-0247, рекрутирующие наивные T-клетки человека для индуцирования уничтожения опухолевых клеток T84. На фигуре 4B показана GUCY2C-1608-опосредованная активность цитотоксических T-клеток, наблюдаемая в GUCY2c-экспрессирующих линиях опухолевых клеток, LS1034. На фигуре 4C показана GUCY2C-1608-опосредованная активность цитотоксических T-клеток, наблюдаемая в GUCY2c-экспрессирующей линии опухолевых клеток LS174T. На фигуре 4D показано, что не наблюдали активности при использовании GUCY2C-1608 в GUCY2c-отрицательной линии клеток.
На фигурах 5A-5F показан анализ in vitro высвобождения цитокинов, индуцируемого после GUCY2C-1608-опосредованного рекрутирования наивных T-клеток человека к GUCY2c-экспрессирующим клеткам T84. При анализе на основе Luminex выявляли положительную регуляцию ИФН-гамма человека (фигура 5A), ИЛ-10 (фигура 5B), ИЛ-2 (фигура 5C), ИЛ-4 (фигура 5D), ИЛ-6 (фигура 5E) и ФНО-альфа (фигура 5F).
На фигуре 6 показано дозозависимое ингибирование роста опухоли посредством GUCY2C-0247 в опухолевом ксенотрансплантате линии клеток колоректальной карциномы T84 в модели адоптивного переноса.
На фигуре 7 показано дозозависимое ингибирование роста опухоли посредством GUCY2C-0247 в опухолевом ксенотрансплантате линии клеток колоректальной карциномы HT55 в модели адоптивного переноса.
На фигуре 8 показано дозозависимое ингибирование роста опухоли посредством GUCY2C-1608 в опухолевом ксенотрансплантате линии клеток колоректальной карциномы HT55 в модели адоптивного переноса.
На фигуре 9 показано дозозависимое ингибирование роста опухоли посредством биспецифических антител против GUCY2c, GUCY2C-0247 и GUCY2C-1608, в опухолевом ксенотрансплантате линии клеток колоректального рака LS1034 в модели адоптивного переноса.
На фигуре 10 показано ингибирование роста опухоли посредством биспецифических антител против GUCY2c, GUCY2C-0074, GUCY2C-0098 и GUCY2C-0105, в опухолевом ксенотрансплантате линии клеток колоректального рака LS1034 в модели адоптивного переноса.
На фигуре 11 показано дозозависимое ингибирование роста опухоли посредством GUCY2C-0098 в опухолевом ксенотрансплантате, полученном из пациента с колоректальной карциномой, PDX-CRX-11201 в модели адоптивного переноса.
На фигуре 12 показано дозозависимое ингибирование роста опухоли посредством GUCY2C-1608 в опухолевом ксенотрансплантате, полученном из пациента с колоректальной карциномой, PDX-CRX-11201 в модели адоптивного переноса.
На фигуре 13 показано ингибирование роста опухоли посредством GUCY2C-0240 в опухолевом ксенотрансплантате, полученном из пациента с колоректальной карциномой, PDX-CRX-11201 в модели адоптивного переноса.
На фигуре 14 показано ингибирование роста опухоли посредством GUCY2C-0098 в опухолевом ксенотрансплантате, полученном из пациента с колоректальной карциномой, PDX-CRX-12213 в модели адоптивного переноса.
На фигуре 15 показано ингибирование роста опухоли посредством GUCY2C-0247 в опухолевом ксенотрансплантате, полученном из пациента с колоректальной карциномой, PDX-CRX-24225 в модели адоптивного переноса.
На фигуре 16 показана характеризация способности к индуцированию цГМФ и нейтрализации биспецифического антитела против GUCY2c-CD3 в клетках T84.
На фигуре 17 показан анализ LC/MS GUCY2C-1608 после очистки.
На фигуре 18 показана опосредованная T-клетками цитотоксичность биспецифического антитела против GUCY2C с вариантами против CD3. Биспецифические антитела против GUCY2c с разными вариантами CD3 рекрутируют наивные T-клетки человека для индуцирования уничтожения опухолевых клеток T84.
На фигуре 19 показан анализ DELFIA ELISA конкуренции за пептид ECD GUCY2c.
На фигуре 20 показан конкурентный DELFIA ELISA с использованием антитела MS20.
На фигуре 21 показаны подробности области связывания между пептидом GUCY2c и пептидом scFv GUCY2C-1608 при кристаллографии.
На фигуре 22 показана кристаллическая структура двух антигенсвязывающих участков на биспецифическом антителе против GUCY2c-CD3, где показано, что они разделены приблизительно 70 Å и находятся на противоположных сторонах молекулы, прямо напротив друг друга.
На фигуре 23 показаны результаты комбинированных исследований с использованием антитела против VEGF и акситиниба, демонстрировавших аддитивное ингибирование роста опухоли в комбинации с биспецифическим антителом против GUCY2c-CD3.
На фигуре 24 показаны результаты комбинированных исследований с использованием антитела против PD1, демонстрировавшего аддитивное ингибирование роста опухоли в комбинации с биспецифическим антителом против GUCY2c-CD3.
На фигуре 25 показаны результаты комбинированных исследований с использованием антитела против PD-L1, демонстрировавшего аддитивное ингибирование роста опухоли в комбинации с биспецифическим антителом против GUCY2c-CD3.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Представленное в настоящем описании изобретение относится к антителам, специфически связывающимся с GUCY2c (например, GUCY2c человека, GUCY2c мыши, GUCY2c крысы, GUCY2c яванского макака). Кроме того, представленное в настоящем описании изобретение относится к биспецифическим антителам, специфически связывающимся с CD3 (например, CD3 человека) и опухолевым антигеном (например, GUCY2c). Изобретение также относится к полинуклеотидам, кодирующим эти антитела, композициям, содержащим эти антитела, и способам получения и применения этих антител. Изобретение дополнительно относится к способам лечения состояния, ассоциированного с экспрессией GUCY2c, у индивидуума, такого как злокачественное новообразование, с использованием антител (например, антитела против GUCY2, антитела против CD3 или биспецифического антитела), как представлено в настоящем описании.
Общие способы
В практическом осуществлении настоящего изобретения, если не указано иначе, используют общепринятые способы молекулярной биологии (включая рекомбинантные способы), микробиологии, клеточной биологии, биохимии и иммунологии, известные специалистам в этой области. Такие способы полностью описаны в литературе, такой как, Molecular Cloning: A Laboratory Manual, второе издание (Sambrook et al., 1989) Cold Spring Harbor Press; Oligonucleotide Synthesis (M.J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R.I. Freshney, ed., 1987); Introduction to Cell and Tissue Culture (J.P. Mather and P.E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J.B. Griffiths, and D.G. Newell, eds., 1993-1998) J. Wiley and Sons; Methods in Enzymology (Academic Press, Inc.); Handbook of Experimental Immunology (D.M. Weir and C.C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J.M. Miller and M.P. Calos, eds., 1987); Current Protocols in Molecular Biology (F.M. Ausubel et al., eds., 1987); PCR: The Polymerase Chain Reaction (Mullis et al., eds., 1994); Current Protocols in Immunology (J.E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C.A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: a practical approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal antibodies: a practical approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using antibodies: a laboratory manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999); Antibodies (M. Zanetti and J.D. Capra, eds., Harwood Academic Publishers, 1995). В некоторых случаях, термины с общепринято понятным значением определяют в настоящем описании для ясности и/или для справки, и включение таких определений в настоящее описание не следует обязательно истолковывать как существенное различие по сравнению со значением, как правило, общепринято понятным в этой области.
Настоящее изобретение далее будет подробно описано со ссылкой на следующие определения и примеры. Все патенты и публикации, включая все последовательности, описанные в таких патентах и публикациях, упоминаемых в настоящем описании, специально включены в него в качестве ссылок.
Определения
В основном, если не указано иначе, все специальные термины, обозначения и другие научные термины или терминология, используемые в настоящем описании, должны иметь значение, общепринято понятное специалистам в области, к которой относится настоящее изобретение. Например, термин "и/или", как используют в такой фразе, как "A и/или B", в настоящем описании, предназначен для включения A и B; A или B; A (в отдельности); и B (в отдельности). Аналогично, термин "и/или", как используют в такой фразе, как "A, B и/или C", предназначен для включения каждого из следующих вариантов осуществления: A, B и C; A, B или C; A или C; A или B; B или C; A и C; A и B; B и C; A (в отдельности); B (в отдельности) и C (в отдельности).
В рамках изобретения термины в единственном числе включают соответствующее множественное число, если контекст четко не указывает на иное.
В рамках изобретения числовые диапазоны включают числа, определяющие диапазон.
Термин "приблизительно" и т.п., когда он предшествует списку числовых значений или диапазону, относится к каждому отдельному значению в списке или диапазоне независимо, как если бы каждому отдельному значению в списке или диапазоне непосредственно предшествовал этот термин. Термины означают, что значения, к которым они относятся, являются точными, близкими или схожими с ними. Например, в некоторых вариантах осуществления термин "приблизительно" конкретное значение может означать значение, составляющее 99%, 95% или 90% от того значения. Например, выражение "приблизительно 100" включает 99 и 101 и все значения между ними (например, 99,1, 99,2, 99,3, 99,4, 99,5 и т.д.). В качестве другого примера, если температура составляет 70°C, "приблизительно" 70°C могут составлять 69°C, 66°C или 63°C. Следует понимать, что они являются исключительно примерами.
В рамках изобретения термин "в пределах 3,8 ангстрем" означает, что контакты составляют 3,8 ангстрем или менее, что определяют посредством кристаллографии.
В рамках изобретения нуклеиновые кислоты записывают слева направо в 5'-3' направлении; аминокислотные последовательности записывают слева направо в ориентации от амино- к карбокси-концу, соответственно. Практикующие специалисты могут найти определения и специальные термины в Sambrook et al., 1 989, и Ausubel FM et al., 1993. Следует понимать, что настоящее изобретение не ограничено конкретной описанной методологией, способами и реагентами, т.к. их можно варьировать.
Термины "полипептид", "олигопептид", "пептид" и "белок" используют в настоящем описании взаимозаменяемо для обозначения цепей аминокислот любой длины, предпочтительно, относительно коротких (например, 10-100 аминокислот). Цепь может являться линейной или разветвленной, может содержать модифицированные аминокислоты и/или может прерываться не-аминокислотами. Термины также включают аминокислотную цепь, модифицированную природно или посредством вмешательства; например, посредством образования дисульфидной связи, гликозилирования, липидирования, ацетилирования, фосфорилирования или любой другой манипуляции или модификации, такой как конъюгация с компонентом-меткой. В определение также включены, например, полипептиды, содержащие один или более аналогов аминокислот (включая, например, неприродные аминокислоты и т.д.), а также другие модификации, известные в этой области. Следует понимать, что полипептиды могут находиться в форме отдельных цепей или ассоциированные цепи. Предпочтительно, используют полипептиды млекопитающих (полипептиды, исходно полученные из организма млекопитающего), более предпочтительно - напрямую секретируемые в среду.
"Антитело" является молекулой иммуноглобулина, способной специфически связываться с мишенью, такой как углевод, полинуклеотид, липид, полипептид и т.д., с помощью по меньшей мере одного участка распознавания антигена, находящегося в вариабельной области молекулы иммуноглобулина. В рамках изобретения термин включает поликлональное антитело, моноклональное антитело, химерное антитело, биспецифическое антитело, антитело с двойной специфичностью, бифункциональное антитело, триспецифическое антитело, полиспецифическое антитело, биспецифическое гетеродимерное диатело, биспецифическое гетеродимерное IgG, меченое антитело, гуманизированное антитело, антитело человека и их фрагменты (такие как Fab, Fab’, F(ab’)2, Fv), одноцепочечные (ScFv) и доменные антитела (включая, например, антитела акул и Верблюдовых), слитые белки, содержащие антитело, и любую другую модифицированную конфигурацию молекулы иммуноглобулина, содержащей участок распознавания антигена. Антитело включает антитело любого класса, такого как IgG, IgA или IgM (или их подкласса), и антитело может не принадлежать к какому-либо конкретному классу. В зависимости от аминокислотной последовательности константной области тяжелых цепей антитела иммуноглобулины можно приписывать к разным классам. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них можно дополнительно разделять на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные области тяжелой цепи, соответствующие разным классам иммуноглобулинов, названы альфа, дельта, эпсилон, гамма и мю, соответственно. Структуры субъединиц и трехмерные конфигурации разных классов иммуноглобулинов хорошо известны. Настоящее изобретение также включает "аналоги антител", другие неантительные белковые каркасы, например, слитые белки и/или иммуноконъюгаты, в которых для обеспечения специфического связывания антигена используют CDR. Антитела по изобретению можно получать из любого биологического вида, включая, в качестве неограничивающих примеров, мышь, человека, верблюда, ламу, рыбу, акулу, козу, кролика, курицу и корову.
Термин "антитело" дополнительно включает иммуноглобулиновые молекулы, содержащие четыре полипептидные цепи, две тяжелые цепи (H) и две легкие цепи (L), соединенные друг с другом дисульфидными связями, а также их мультимеры (например, IgM). Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (сокращенно обозначаемую в настоящем описании как VR HC или VH) и константную область тяжелой цепи. Термин "вариабельная область" антитела относится к вариабельной области легкой цепи антитела или вариабельной области тяжелой цепи антитела в отдельности или в комбинации. Константная область тяжелой цепи содержит три домена, CH1, CH2 и CH3. Домены CH1 и CH2 соединены шарнирной областью. Каждая легкая цепь содержит вариабельную область легкой цепи (сокращенно обозначаемую в настоящем описании как VR LC или VL) и константную область легкой цепи. Константная область легкой цепи содержит один домен (CL1). Области VH и VL можно дополнительно разделять на области гипервариабельности, обозначаемые как определяющие комплементарность области (CDR), перемежающие с областями, являющимися более консервативными, обозначаемыми как каркасные области (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. В разных вариантах осуществления изобретения FR антитела против CD3 (или его антигенсвязывающей части) могут являться идентичными последовательностям зародышевой линии человека или природно или искусственно модифицированными. Консенсусную аминокислотную последовательность можно определять посредством параллельного анализа двух или более CDR. CDR в каждой цепи удерживаются в непосредственной близости с помощью FR и вместе с CDR другой цепи участвуют в образовании антигенсвязывающего участка антитела.
В рамках изобретения термин "антигенсвязывающий фрагмент", или "фрагмент антитела", или "антигенсвязывающая часть" относится к одному или более фрагментам антитела, сохраняющим способность специфически связываться с антигеном. Примеры связывающих фрагментов, включенных в термин "антигенсвязывающий фрагмент" антитела, включают (i) вариабельный домен тяжелой цепи антитела (VH), и/или вариабельный домен легкой цепи антитела (VL), или пару VH/VL, полученную из полноразмерных антител или фрагментов антител, таких как домен VH и/или домен VL; (ii) Fab-фрагмент, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (iii) Fab'-фрагмент, по существу, являющийся Fab с частью шарнирной области (например, Fundamental Immunology, Paul ed., 3.sup.rd ed.1993); (iv) F(ab')2-фрагмент, бивалентный фрагмент, содержащий два Fab-фрагмента, соединенных дисульфидным мостиком в шарнирной области; (v) Fd-фрагмент, состоящий из доменов VH и CH1; (vi) Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела; (vii) одноцепочечный F-фрагмента (scFv), отдельная белковая цепь, в которой области VL и VH спариваются с образованием моновалентных молекулы (например, Bird et al. (1988) Science 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883); (viii) стабилизированный дисульфидными связями Fv-фрагмент (dsFv), Fv со сконструированной межмолекулярной дисульфидной связью для стабилизации пары VH-VL; (ix) антитело из отдельного вариабельного домена (sdAb или dAb) (например, Ward et al., (1989) Nature 341:544-546), состоящее из вариабельного домена тяжелой цепи и лишенное легкой цепи; (x) определяющую комплементарность область (CDR) и любые их производные.
В рамках изобретения "антигенсвязывающий фрагмент" антитела может содержать по меньшей мере один вариабельный домен. Вариабельный домен может иметь любой размер или аминокислотную композицию и, как правило, будет содержать по меньшей мере одну CDR, смежную или находящуюся в рамке считывания с одной или более каркасными последовательностями. В рамках изобретения "антигенсвязывающий фрагмент антитела" может содержать гомодимер или гетеродимер (или другой мультимер) любой из конфигураций вариабельной области и константного домена, указанных ниже, в нековалентной связи друг с другом и/или с одной или более мономерными областями VH или VL (например, с помощью дисульфидных связей). Например, вариабельная область может являться димерной и содержать димеры VH-VH, VH-VL или VL-VL. Конфигурации вариабельных и константных доменов, которые можно обнаружить в антигенсвязывающем фрагменте антитела по настоящему изобретению, включают: VH-CH1; VH-CH2; VH-CH3; VH-CH1-CH2; VH-CH1-CH2-CH3; VH-CH2-CH3; VH-VL-CL, VH-VL-CH1, VH-VL-CH2; VH-CL; VL-CH1; VL-CH2; VL-CH3; VL-CH1-CH2; VL-CH1-CH2-CH3; VL-CH2-CH3 и VL-CL. Вариабельные области и константные домены можно связывать друг с другом напрямую или с помощью полной или частичной шарнирной или линкерной области. Шарнирная область может состоять из по меньшей мере 2 (например, 5, 10, 15, 20, 40, 60 или более) аминокислот, что приводит к гибкому или полугибкому связыванию между смежными вариабельными областями и/или константными доменами в одной полипептидной молекуле.
В рамках изобретения "связывающий домен" содержит любую область полипептида (например, антитела), отвечающую за селективное связывание с интересующей молекулой (например, антигеном, лигандом, рецептором, субстратом или ингибитором). Примеры связывающих доменов включают вариабельную область антитела, рецептор-связывающий домен, лиганд-связывающий домен и ферментативный домен.
В рамках изобретения термин "акцепторный каркас человека" является каркасом, содержащим аминокислотную последовательность каркаса вариабельной области легкой цепи (VL) или каркаса вариабельной области тяжелой цепи (VH), полученную из каркаса иммуноглобулина человека или консенсусного каркаса человека, как определено ниже. Акцепторный каркас человека, "полученный из" каркаса иммуноглобулина человека или консенсусного каркаса человека, может содержать ту же аминокислотную последовательность или может содержать модификации аминокислотной последовательности. В некоторых вариантах осуществления количество аминокислотных модификаций составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. В некоторых вариантах осуществления акцепторный каркас VL человека является идентичным по последовательности в отношении последовательности каркаса VL иммуноглобулина человека или консенсусной каркасной последовательности человека.
В рамках изобретения термин "аффинно зрелое" антитело относится к антителу с одной или более модификациями в одной или более вариабельных областях, включающих CDR и FR, по сравнению с родительским антителом, не имеющим таких модификаций, и где такие модификации приводят к улучшению аффинности антитела к антигену.
В рамках изобретения термин "Fc-область", "Fc-домен", "Fc-цепь" или аналогичные термины используют для определения C-концевой области тяжелой цепи IgG. Fc-область IgG содержит два константных домена, CH2 и CH3. Домен CH2 Fc-области IgG человека, как правило, располагается от аминокислоты 231 до аминокислоты 340 в соответствии с системой нумерации индекса EU или от аминокислоты 244 до аминокислоты 360 в соответствии с системой нумерации Kabat. Домен CH3 Fc-области IgG человека, как правило, располагается от аминокислоты 341 до аминокислоты 447 в соответствии с системой нумерации индекса EU или от аминокислоты 361 до аминокислоты 478 в соответствии с системой нумерации Kabat. Домен CH2 Fc-области IgG человека (также обозначаемый как домен "Cγ 2") является уникальным в том, что он не спарен тесно с другим доменом. Вместо этого, две N-связанных разветвленных углеводных цепи расположены между двумя доменами CH2 интактного нативного IgG. Fc-область может содержать нативные последовательности Fc или варианты последовательности Fc. Хотя границы последовательности Fc тяжелой цепи иммуноглобулина могут варьироваться, последовательность Fc тяжелой цепи IgG человека, как правило, определяют как располагающуюся от аминокислотного остатка приблизительно в положении Cys226 или приблизительно положении Pro230 до карбокси-конца последовательности Fc. Если в настоящем описании не указано иначе, нумерацию аминокислотных остатков в Fc-области или константной области осуществляют в соответствии с системой нумерации EU, также обозначаемой как индекс EU, как описано в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
В некоторых вариантах осуществления Fc-цепь начинается в шарнирной области непосредственно выше участка расщепления папаином и заканчивается на C-конце антитела. Таким образом, полная Fc-цепь содержит по меньшей мере шарнирный домен, домен CH2 и домен CH3. В некоторых вариантах осуществления Fc-цепь содержит по меньшей мере один из: шарнирного домена (например, верхней, средней и/или нижней шарнирной области), домена CH2, домена CH3, домена CH4 или его варианта, части или фрагмента. В некоторых вариантах осуществления Fc-домен содержит полную Fc-цепь (т.е. шарнирный домен, домен CH2 и домен CH3). В некоторых вариантах осуществления Fc-цепь содержит шарнирный домен (или его часть), слитый с доменом CH3 (или его частью). В некоторых вариантах осуществления Fc-цепь содержит домен CH2 (или его часть), слитый с доменом CH3 (или его частью). В некоторых вариантах осуществления Fc-цепь состоит из домена CH3 или его частью. В некоторых вариантах осуществления Fc-цепь состоит из шарнирного домена (или его части) и домена CH3 (или его части). В некоторых вариантах осуществления Fc-цепь состоит из домена CH2 (или его части) и домена CH3. В некоторых вариантах осуществления Fc-цепь состоит из шарнирного домена (или его части) и домена CH2 (или его части). В некоторых вариантах осуществления в Fc-цепи отсутствует по меньшей мере часть домена CH2 (например, весь домен CH2 или его часть). Термин "Fc-цепь" в основном, относится к полипептиду, содержащему всю Fc-цепь тяжелой цепи иммуноглобулина или его часть. Это включает, в качестве неограничивающих примеров, полипептиды, содержащие полные домен CH1, шарнирный домен, домены CH2 и/или CH3, а также фрагменты таких пептидов, содержащие только, например, шарнирный домен, домен CH2 и домен CH3. Fc-цепь можно получать из иммуноглобулина любого биологического вида и/или любого подтипа, включая, в качестве неограничивающих примеров, антитело IgG1, IgG2, IgG3, IgG4, IgD, IgA, IgE или IgM человека. Fc-домен включает нативные молекулы Fc и варианты молекул Fc. Как и в случае вариантов Fc и нативных Fc, термин "Fc-цепь" включает молекулы в мономерной или мультимерной форме, отщеплена ли она от целого антитела или получена другими способами. В некоторых вариантах осуществления Fc-цепь содержит карбокси-концевые части обеих тяжелых цепей, удерживаемые вместе посредством дисульфидных связей. В некоторых вариантах осуществления Fc-цепь состоит из домена CH2 и домена CH3.
В этой области термины "Fc-рецептор" и "FcR" используют для описания рецептора, связывающегося с Fc-областью антитела. Предпочтительным FcR является нативная последовательность FcR человека. Кроме того, предпочтительным FcR является FcR, связывающийся с антителом IgG (гамма-рецептор), и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, включая аллельные варианты и альтернативно сплайсированные формы этих рецепторов. Рецепторы FcγRII включают FcγRIIA ("активирующий рецептор") и FcγRIIB ("ингибирующий рецептор"), имеющие схожие аминокислотные последовательности, отличающиеся, главным образом, своими цитоплазматическими доменами. Обзор FcR см. в Ravetch and Kinet, Ann. Rev. Immunol., 9:457-92, 1991; Capel et al., Immunomethods, 4:25-34, 1994; и de Haas et al., J. Lab. Clin. Med., 126:330-41, 1995. Термин "FcR" также включает неонатальный рецептор FcRn, отвечающий за перенос материнских IgG в плод (Guyer et al., J. Immunol., 117:587, 1976; и Kim et al., J. Immunol., 24:249, 1994).
"Нативная последовательность Fc-области" или "Fc-область дикого типа" содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, обнаруживаемой в природе. Термин Fc IgG "дикого типа" человека означает последовательность аминокислот, встречающуюся в природе в популяции людей. Разумеется, как и последовательность Fc может немного варьироваться среди индивидуумов, можно осуществлять одно или более изменений в последовательности дикого типа, и она все равно будет входить в объем изобретения. Например, Fc-область может содержать дополнительные изменения, не относящиеся к настоящему изобретению, такие как мутация в участке гликозилирования, включение неприродной аминокислоты или мутация "выступ-во-впадину".
"Вариант Fc-области" или "вариант Fc-цепи" содержит аминокислотную последовательность, отличающуюся от нативной последовательности Fc-области по меньшей мере одной аминокислотной модификацией, но сохраняющую по меньшей мере одну эффекторную функцию нативной последовательности Fc-области. В некоторых вариантах осуществления вариант Fc-цепи имеет по меньшей мере одну замену аминокислоты по сравнению с нативной последовательностью Fc-цепи или Fc-области родительского полипептида, например, от приблизительно одной до приблизительно десяти замен аминокислот, и предпочтительно - от приблизительно одной до приблизительно пяти замен аминокислот в нативной последовательности Fc-цепи или Fc-цепи родительского полипептида. В настоящем описании вариант Fc-цепи, предпочтительно, будет иметь по меньшей мере приблизительно 80% идентичности последовательности в отношении нативной последовательности Fc-цепи и/или Fc-цепи родительского полипептида, и наиболее предпочтительно - по меньшей мере, приблизительно 90% идентичности последовательности, более предпочтительно - по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99% идентичности последовательности.
В рамках изобретения термин "эффекторные функции" относится к видам биологической активности, приписываемым Fc-цепи (нативной последовательности Fc-цепи или аминокислотной последовательности варианта Fc-цепи) антитела и варьируется в зависимости от изотипа антитела. Примеры эффекторных функций антител включают: связывание C1q и обусловленную комплементом цитотоксичность; связывание Fc-рецептора; антителозависимую клеточную цитотоксичность (ADCC); фагоцитоз; отрицательную регуляцию рецепторов поверхности клетки (например, B-клеточного рецептора) и активацию B-клеток. Для таких эффекторных функций, как правило, необходимо комбинирование Fc-цепи со связывающим доменом (например, вариабельной областью антитела), и их можно оценивать с использованием различных анализов, известных в этой области для оценки таких эффекторных функций антител. Пример измерения эффекторной функции осуществляют с помощью связывания Fcγ3 и/или C1q.
Эффекторные функции антител определяются последовательностями Fc-цепи; эта цепь также является частью, распознаваемой Fc-рецепторами (FcR), обнаруживаемыми на некоторых типах клеток.
В некоторых вариантах осуществления полипептид Fc содержит часть или всю последовательность шарнирной области дикого типа (как правило, на N-конце). В некоторых вариантах осуществления полипептид Fc не содержит функциональную последовательность шарнирной области или последовательность шарнирной области дикого типа.
В рамках изобретения термин "шарнирная область", "шарнирная последовательность" и его варианты включает значение, известное в этой области, проиллюстрированное, например, в Janeway et al., ImmunoBiology: the immune system in health and disease, Elsevier Science Ltd., NY (4th ed., 1999); Bloom et al., Protein Science, 6:407-415, 1997; и Humphreys et al., J. Immunol. Methods, 209:193-202, 1997.
В рамках изобретения термин "иммуноглобулиноподобная шарнирная область", "иммуноглобулиноподобная шарнирная последовательность" или его варианты относится к шарнирной области и шарнирной последовательности иммуноглобулиноподобной или антитело-подобной молекулы (например, иммуноадгезинов). В некоторых вариантах осуществления иммуноглобулиноподобную шарнирную область можно получать из любого подтипа IgG1, IgG2, IgG3 или IgG4 или из IgA, IgE, IgD или IgM, включая их химерные формы, например, химерной шарнирной области IgG1/2.
В некоторых вариантах осуществления шарнирную область можно получать из подтипа IgG1 человека от аминокислоты 216 до аминокислоты 230 в соответствии с системой нумерации индекса EU или от аминокислоты 226 до аминокислоты 243 в соответствии с системой нумерации Kabat. В некоторых вариантах осуществления последовательность может представлять собой EPKSCDKTHTCPPCP (SEQ ID NO: 186). Специалисты в этой области могут отличаться по своему пониманию конкретных аминокислот, соответствующих различным доменам молекулы IgG. Таким образом, N-конец или C-конец доменов, описанных выше, может быть длиннее или короче на 1, 2, 3, 4, 5, 6, 7, 8, 9 или даже 10 аминокислот.
В некоторых вариантах осуществления шарнирная область может являться мутантной на одну или более аминокислот. В некоторых вариантах осуществления шарнирная область может быть укороченной и содержать лишь часть полной шарнирной области. В некоторых вариантах осуществления шарнирная область может содержать только последние 5 аминокислот шарнирной области.
В рамках изобретения термины "связанный", "слитый", "слияние", "ковалентно связанный", "ковалентно соединенный" и "генетически слитый" используют взаимозаменяемо. Эти термины относятся к соединению двух или более элементов или компонентов любыми способами, включая химическую конъюгацию или рекомбинантные способы. В рамках изобретения термин "ковалентно связанный" означает, что указанные фрагменты напрямую ковалентно связывают друг с другом или косвенно ковалентно соединяют друг с другом с помощью промежуточного остатка или остатков, таких как линкерный пептид или фрагмент. В предпочтительном варианте осуществления фрагменты ковалентно слиты. Одним из типов ковалентной связи является пептидная связь. Способы химической конъюгации (например, с использованием гетеробифункциональных перекрестно-сшивающих средств) известны в этой области. Слитые фрагменты также можно подвергать генетическому слиянию. В рамках изобретения термин "генетически слитый" или "генетически связанный" относится к колинейной ковалентной связи или присоединению двух или более белков, полипептидов или их фрагментов через их отдельные пептидные остовы, посредством генетической экспрессии единой полинуклеотидной молекулы, кодирующей эти белки, полипептиды или фрагменты. Такое генетическое слияние приводит к экспрессии единой смежной генетической последовательности. Предпочтительное генетическое слияние осуществляют в рамке считывания, т.е. две или более открытые рамки считывания (ORF) подвергают слиянию с образованием непрерывной более длинной ORF таким образом, чтобы сохранить правильную рамку считывания исходных ORF. Таким образом, получаемый рекомбинантный слитый белок является единым полипептидом, содержащим два или более белковых сегмента, соответствующих полипептидам, кодируемым исходными ORF (при этом сегменты в норме не соединены таким образом в природе). В этом случае, единый полипептид расщепляется при процессинге с образованием димерных молекул, содержащих две полипептидные цепи.
В рамках изобретения термин "модификация" относится к замене, инсерции и/или делеции аминокислоты в полипептидной последовательности, изменению во фрагменте, химически соединенном с белком, или модификации функции белка, например, антитела. Например, модификация может представлять собой измененную функцию антитела или измененную углеводную структуру, присоединенную к белку. В рамках изобретения термин "аминокислотная модификация" относится к мутации (замене), инсерции (добавлению) или делеции одного или более аминокислотных остатков в антителе. Термин "мутация аминокислоты" означает замену по меньшей мере одного существующего аминокислотного остатка другим, отличающимся аминокислотным остатком (например, замену аминокислотного остатка). Термин "делеция аминокислоты" означает удаление по меньшей мере одного аминокислотного остатка в заранее определенном положении в аминокислотной последовательности. Например, мутация L234A означает, что аминокислотный остаток лизин в положении 234 в Fc-области антитела заменен аминокислотным остатком аланином (замена лизина аланином) (нумерация в соответствии с индексом EU). Термин "природный аминокислотный остаток" означает аминокислотный остаток из группы, состоящей из аланина (трехбуквенное обозначение: Ala, однобуквенное обозначение: A), аргинина (Arg, R), аспарагина (Asn, N), аспарагиновой кислоты (Asp, D), цистеина (Cys, C), глутамина (Gln, Q), глутаминовой кислоты (Glu, E), глицина (Gly, G), гистидина (His, H), изолейцина (Ile, I), лейцина (Leu, L), лизина (Lys, K), метионина (Met, M), фенилаланина (Phe, F), пролина (Pro, P), серина (Ser, S), треонина (Thr, T), триптофана (Trp, W), тирозина (Tyr, Y) и валина (Val, V).
В рамках изобретения "консервативная аминокислотная замена" является заменой, при которой аминокислотный остаток заменяют аминокислотным остатком, имеющим схожую боковую цепь. В этой области определены семейства аминокислотных остатков, имеющих схожие боковые цепи. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновую кислоту, глутаминовую кислоту), незаряженными полярными боковыми цепями (например, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, глицин, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин).
В рамках изобретения "заменимый" аминокислотный остаток является остатком, который можно изменять относительно последовательности дикого типа связывающего средства, например, антитела, без устранения или без существенного изменения биологической активности, в то время как изменение "незаменимого" аминокислотного остатка приводит к такому изменению. В антителе незаменимый аминокислотный остаток может являться определяющим специфичность остатком (SDR).
Термин "средство" используют в настоящем описании для обозначения биологической макромолекулы, экстракта из биологических материалов, смеси биологических макромолекул, химического соединения, смеси химических соединений и/или смеси химических соединений и биологических макромолекул. Термин "терапевтическое средство" относится к средству, имеющему биологическую активность.
В рамках изобретения термин "моноклональное антитело" относится к антителу, полученному из популяции, по существу, гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных природных мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифическими и направлены против одного антигенного участка. Кроме того, в отличие от препаратов поликлональных антител, которые, как правило, включают различные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной и той же детерминанты на антигене. Определение "моноклональное" свидетельствует о природе антитела как полученного из, по существу, гомогенной популяции антител, и не следует истолковывать его как требующее получения антитела любым конкретным способом. Например, моноклональные антитела для использования по настоящему изобретению, можно получать гибридомным способом, впервые описанным в Kohler and Milstein, Nature 256:495, 1975, или способами рекомбинантной ДНК, как описано в патенте США № 4816567. Моноклональные антитела также можно выделять из фаговых библиотек, полученных способом, описанным, например, в McCafferty et al., Nature 348:552-554, 1990.
Антитела по настоящему изобретению могут являться "гуманизированными антителами". В рамках изобретения термин "гуманизированное" антитело относится к формам не принадлежащих человеку антител (например, мыши, крысы, кролика, не являющегося человеком примата или другого млекопитающего), являющихся химерными иммуноглобулинами, иммуноглобулиновыми цепями или их фрагментами (такими как Fv, Fab, Fab’, F(ab')2 или другие антигенсвязывающие подпоследовательности антител), содержащими один или более аминокислотных остатков, включенных в них из источника, не принадлежащего человеку. Эти не принадлежащий человеку аминокислотные остатки зачастую обозначают как "импортируемые" остатки, как правило, получаемые из "импортируемого" вариабельного домена. Импортируемый остаток, последовательность или антитело имеет желаемую аффинность и/или специфичность или другую желаемую биологическую активность антитела, представленную в настоящем описании.
Предпочтительно, гуманизированные антитела являются иммуноглобулинами человека (реципиентное антитело), в которых остатки из определяющей комплементарность области (CDR) реципиента заменяют остатками CDR не являющихся человеком видов (донорное антитело), таких как мышь, крыса или кролик, имеющими желаемую специфичность, аффинность и емкость. В некоторых случаях остатки каркасной области Fv (FR) иммуноглобулина человека заменяют соответствующими не принадлежащими человеку остатками. Кроме того, гуманизированное антитело может содержать остатки, не обнаруживаемые ни в реципиентном антителе, ни в импортируемых последовательностях CDR или каркас, но включенные для дополнительного улучшения и оптимизации свойств антител. В основном, гуманизированное антитело будет содержать, по существу, все из по меньшей мере одной и, как правило, двух вариабельных областей, в которых все или, по существу, все из областей CDR соответствуют CDR не принадлежащего человеку иммуноглобулина, и все или, по существу, все из областей FR являются областями из консенсусной последовательности иммуноглобулина человека. Гуманизированное антитело оптимально также будет содержать по меньшей мере часть константной области иммуноглобулина или домен (Fc), как правило, из иммуноглобулина человека. Предпочтительными являются антитела, имеющие Fc-цепи, модифицированные, как описано в WO 99/58572. Другие формы гуманизированных антител имеют одну или более CDR (CDR L1, CDR L2, CDR L3, CDR H1, CDR H2 или CDR H3), измененные относительно исходного антитела, также обозначаемые как одна или более CDR, "полученные из" одной или более CDR из исходного антитела. В рамках изобретения термин "гуманизированный" предназначен для включения деиммунизированных антител.
В рамках изобретения термин "антитело человека" означает антитело, имеющее аминокислотную последовательность, соответствующую последовательности антитела, продуцируемого человеком и/или полученного любым из способов получения антител человека, известных специалистам в этой области или представленных в настоящем описании. Таким образом, в рамках изобретения термин "антитело человека" предназначен для включения антител, имеющих вариабельные и константные области, полученные из последовательностей иммуноглобулинов зародышевой линии человека. Антитела человека по изобретению могут включать аминокислотные остатки, не кодируемые последовательностями иммуноглобулинов зародышевой линии человека (например, мутации, встроенные посредством случайного или сайт-специфического мутагенеза in vitro или соматической мутации in vivo), например, в CDR, и в частности, CDR3. Это определение антитела человека включает антитела, содержащие по меньшей мере один полипептид тяжелой цепи человека или по меньшей мере один полипептид легкой цепи человека. Одним из таких примеров является антитело, содержащее полипептиды легкой цепи мыши и тяжелой цепи человека. Антитела человека можно получать различными способами, известными в этой области. В одном из вариантов осуществления антитело человека выбрано из фаговой библиотеки, где фаговая библиотека экспрессирует антитела человека (Vaughan et al., Nature Biotechnology, 14:309-314, 1996; Sheets et al., Proc. Natl. Acad. Sci. (USA) 95:6157-6162, 1998; Hoogenboom и Winter, J. Mol. Biol., 227:381, 1991; Marks et al., J. Mol. Biol., 222:581, 1991). Антитела человека также можно получать посредством иммунизации животных, в которые локусы иммуноглобулинов человека трансгенно встроены вместо эндогенных локусорв, например, мышей, в которых эндогенные гены иммуноглобулинов частично или полностью инактивированы. Этот подход описан в патентах США №№ 5545807, 5545806, 5569825, 5625126, 5633425 и 5661016. Альтернативно, антитело человека можно получать посредством иммортализации B-лимфоцитов человека, продуцирующих антитело против антигена-мишени (такие B-лимфоциты можно выделять из индивидуума или при клонировании отдельных клеток с использованием кДНК, или их можно иммунизировать in vitro). См., например, Cole et al. Monoclonal antibodies and Cancer Therapy, Alan R. Liss, p. 77, 1985; Boerner et al., J. Immunol., 147(1):86-95, 1991; и патент США № 5750373.
Антитела человека по настоящему изобретению могут существовать по меньшей мере в двух формах, что связано с гетерогенностью шарнирной области. Например, молекула иммуноглобулина содержит стабильную конструкцию из четырех цепей массой приблизительно 150-160 кДа, в которой димеры удерживаются вместе с помощью межцепочечных дисульфидных связей между тяжелыми цепями. Альтернативно, димеры не связаны с помощью межцепочечных дисульфидных связей, и образуется молекула массой приблизительно 75-80 кДа, состоящая из ковалентно соединенных легкой и тяжелой цепей (полу-антитело).
В рамках изобретения термин "рекомбинантное антитело человека" предназначен для включения всех антител человека, полученных, экспрессированных, созданных или выделенных рекомбинантными способами, таких как антитела, экспрессируемые с использованием рекомбинантного экспрессирующего вектора, с помощью которого трансфицируют клетку-хозяина (описано ниже), антитела, выделенные из рекомбинантной комбинаторной библиотеки антител человека (описано ниже), антитела, выделенные из животного (например, мыши), являющегося трансгенным по генам иммуноглобулина человека (см., например, Taylor et al. (1992) Nucl. Acids Res. 20:6287-6295), или антитела, полученные, экспрессированные, созданные или выделенные любыми другими способами, включающими сплайсинг последовательностей генов иммуноглобулина человека с другими последовательностями ДНК. Такие рекомбинантные антитела человека имеют вариабельные и константные области, полученные из последовательностей иммуноглобулинов зародышевой линии человека. Однако в некоторых вариантах осуществления такие рекомбинантные антитела человека подвергают мутагенезу in vitro (или, если используют животное, трансгенное по последовательностям Ig человека, соматическому мутагенезу in vivo), и, таким образом, аминокислотные последовательности областей VH и VL рекомбинантных антител являются последовательностями, которые, хотя и происходят из последовательностей VH и VL зародышевой линии человека и являются родственными им, могут не существовать в природе в репертуаре антител зародышевой линии человека in vivo.
Антитела по изобретению могут являться "химерными" антителами, в которых часть тяжелой и/или легкой цепи является идентичной или гомологичной соответствующим последовательностям в антителах, полученных из конкретных видов или принадлежащих к конкретному классу или подклассу антител, в то время как остальная часть цепей является идентичной или гомологичной соответствующим последовательностям в антителах, полученных из других видов или принадлежащих к другому классу или подклассу антител, а также фрагментами таких антител при условии, что они проявляют желательную биологическая активность (патент США № 4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA 81:6851-6855, 1984). Интересующие химерные антитела в настоящем описании включают приматизированные антитела, содержащие антигенсвязывающие последовательности вариабельной области, полученные из не являющегося человеком примата (например, обезьян Старого Света, человекообразных обезьян и т.д.), и последовательности константной области человека.
"Моновалентное антитело" содержит один антигенсвязывающий участок на молекулу (например, IgG или Fab). В некоторых случаях моновалентное антитело может иметь более одного антигенсвязывающего участка, но связывающие участки предназначены для разных антигенов.
Термин "моноспецифическое антитело" относится к антителу или препарату антител, содержащему два идентичных антигенсвязывающих участка на молекулу (например, IgG) таким образом, что два связывающих участка связываются с идентичным эпитопом на антигене. Таким образом, они конкурируют друг с другом за связывание с одной молекулой антигена. Этот термин включает "моноклональное антитело" или "композицию моноклональных антител". Большинство антител, обнаруживаемых в природе, являются моноспецифическими. В некоторых случаях моноспецифическое антитело может являться моновалентным антителом (например, Fab).
"Бивалентное антитело" содержит два антигенсвязывающих участка на молекулу (например, IgG). В некоторых случаях два связывающих участка имеют одинаковую антигенную специфичность. Однако бивалентные антитела могут являться биспецифическими.
В рамках изобретения "биспецифическое антитело", "антитело с двойной специфичностью", "бифункциональное антитело", "гетеромультимер", "гетеромультимерный комплекс", "биспецифическое гетеродимерное диатело" или "гетеромультимерный полипептид" представляет собой молекулу, содержащую по меньшей мере первый полипептид и второй полипептид, где второй полипептид отличается по аминокислотной последовательности от первого полипептида по меньшей мере на один аминокислотный остаток. В некоторых случаях биспецифическое антитело является искусственным гибридным антителом, имеющих две разные области тяжелой цепи и области легкой цепи. Предпочтительно, биспецифическое антитело имеет специфичность связывания по меньшей мере для двух разных лигандов, антигенов или участков связывания. Таким образом, биспецифические антитела могут связываться одновременно с двумя разными антигенами. Два антигенсвязывающих участка биспецифического антитела связываются с двумя разными эпитопами, которые могут находиться на одной или разных белковых мишенях, например, опухолевой мишени.
В рамках изобретения термин "антиген-мишень", "антиген клетки-мишени", "опухолевый антиген" или "опухолеспецифический антиген" относится к антигенной детерминанте, присутствующей на поверхности клетки-мишени, например, клетки в опухоли, такой как злокачественная клетка или клетка опухолевой стромы.
Термины "мутационная нагрузка" используют в настоящем описании взаимозаменяемо. Мутационная нагрузка опухоли является мерой количества мутаций в опухолевом геноме, определяемой как общее количество мутаций на кодирующую область опухолевого генома. Наблюдают значительную вариабельность мутационной нагрузки среди типов опухолей в диапазоне от всего лишь нескольких мутаций до тысяч мутаций (Alexandrov LB et al., Nature 2013;500(7463):415-421; Lawrence MS et al., Nature 2013;499:214-218; Vogelstein B et al., Science, 2013;339:1546-1558).
В рамках изобретения первая полипептидная цепь и вторая полипептидная цепь "биспецифического антитела" содержат по меньшей мере одну VL антитела и одну VH антитела или ее фрагмент, где оба связывающих домена антитела содержатся в одной полипептидной цепи, и где области VL и VH в каждой полипептидной цепи получены из разных антител.
Биспецифическое антитело, антитело с двойной специфичностью, бифункциональное антитело, гетеромультимер, гетеромультимерный комплекс, биспецифическое гетеродимерное диатело или гетеромультимерный полипептид могут образовывать третичные структуры более высокого порядка, где присутствуют другие полипептиды в дополнение к первому и второму полипептиду. Полипептиды гетеромультимера могут взаимодействовать друг с другом посредством непептидной ковалентной связи (например, дисульфидной связи) и/или нековалентного взаимодействия (например, водородных связей, ионных связей, вандерваальсовых сил и/или гидрофобных взаимодействий).
Биспецифическое антитело, антитело с двойной специфичностью, бифункциональное антитело, гетеромультимер, гетеромультимерный комплекс, биспецифическое гетеродимерное диатело или гетеромультимерный полипептид можно получать посредством конструирования sFv-фрагментов с короткими линкерами (например, приблизительно 3-10 остатков) между областями VH и VL таким образом, что достигают межцепочечного, а не внутрицепочечного спаривания V-областей, что приводит к образованию бивалентного фрагмента, т.е. фрагмента, имеющего два антигенсвязывающих участка. Биспецифические антитела можно получать из полноразмерных антител или фрагментов антител (например, биспецифических антител F(ab′)2). Диатела более подробно описывают, например, в EP404,097; WO93/11161; и Hollinger et al., Proc. Natl. Acad. Sci. USA 90:6444-6448, 1993. Биспецифические антитела являются гетеродимерами двух "перекрестных" sFv-фрагментов, в которых области VH и VL двух антител находятся на разных полипептидных цепях.
В рамках изобретения термин "выделенное антитело" означает антитело, идентифицированное и отделенное и/или выделенное из по меньшей мере одного компонента из своего природного окружения. Например, антитело, отделенное или удаленное от по меньшей мере одного компонента организма или от ткани или клетки, в которой антитело существует или продуцируется в природе, является "выделенным антителом" в соответствии с целями по настоящему изобретению. Выделенное антитело также включает антитело in situ в рекомбинантной клетке. Выделенные антитела являются антителами, подвергнутыми по меньшей мере одной стадии очистки или выделения. В некоторых вариантах осуществления выделенное антитело может, по существу, не содержать другой клеточный материал и/или химические вещества.
В рамках изобретения термин "связанный" или "связь" относится к прямому соединению посредством пептидной связи между первой и второй аминокислотной последовательностью или соединению, включающему третью аминокислотную последовательность, соединенную пептидной связью с первой и второй аминокислотными последовательностями. Например, линкер, соединенной пептидной связью с C-концом одной аминокислотной последовательности и N-концом другой аминокислотной последовательности.
В рамках изобретения термин "линкер" относится к аминокислотной последовательности длиной две или более аминокислот. Линкер может состоять из нейтральных полярных или неполярных аминокислот. Линкер может составлять, например, от 2 до 100 аминокислот в длину, например, от 2 до 50 аминокислот в длину, например, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50 аминокислот в длину. Линкер может являться "расщепляемым", например, посредством саморасщепления или ферментативного или химического расщепления. Участки расщепления в аминокислотных последовательностях и ферменты и химические вещества, расщепляющие по таким участкам, хорошо известны в этой области, а также представлены в настоящем описании.
В рамках изобретения термин "дисульфидная связь" или "цистеин-цистеиновая дисульфидная связь" относится к ковалентному взаимодействию между двумя цистеинами, в которых атомы серы из цистеинов окисляются с образованием дисульфидной связи. Средняя энергия связи в случае дисульфидной связи составляет приблизительно 60 ккал/моль по сравнению с 1-2 ккал/моль в случае водородной связи. В контексте настоящего изобретения цистеины, образующие дисульфидную связь, находятся в каркасных областях одноцепочечного антитела и служат для стабилизации конформации антитела. Остатки цистеина можно встраивать, например, посредством сайт-специфического мутагенезу таким образом, что в молекуле могут образовываться стабилизирующие дисульфидные связи.
Обозначение "выступ-во-впадину" является аналогичным обозначению "выступ и впадина", и их можно использовать взаимозаменяемо.
Термин "выступ" относится по меньшей мере к одной боковой цепи аминокислоты, выступающей с контактной поверхности первого полипептида и, таким образом, располагаемой в компенсаторной впадине в смежной контактной поверхности (т.е. контактной поверхности второго полипептида) для стабилизации гетеродимера, что, таким образом, способствует образованию гетеродимера по сравнению с образованием гомодимера. Выступ может существовать на исходной контактной поверхности, или его можно встраивать синтетически (например, посредством измерения нуклеиновой кислоты, кодирующей контактную поверхность). Как правило, нуклеиновую кислоту, кодирующую контактную поверхность первого полипептида, изменяют так, что она кодирует выступ. Для достижения этого нуклеиновую кислоту, кодирующую по меньшей мере один исходный аминокислотный остаток в контактной поверхности первого полипептида, заменяют нуклеиновой кислотой, кодирующей, по меньшей мере один импортируемый аминокислотный остаток, имеющий больший объем боковой цепи, чем исходный аминокислотный остаток. Верхним пределом количества исходных остатков, подвергаемых замене, является общее количество остатков в контактной поверхности первого полипептида. Некоторые импортируемые остатки для образования выступа, как правило, являются природными аминокислотными остатками и, предпочтительно, выбраны из аргинина (R), фенилаланина (F), тирозина (Y) и триптофана (W).
Выступ "располагается" во впадине, что означает, что пространственное расположение выступа и впадины на контактной поверхности первого полипептида и второго полипептида, соответственно, и размеры выступа и впадины являются такими, что выступ может располагаться во впадине без значительного нарушения нормального соединения первого и второго полипептидов в контактной поверхности. Т.к. выступы, такие как фенилаланин (F), тирозин (Y) и триптофан (W), как правило, не располагаются перпендикулярно от оси контактной поверхности, выравнивание выступа с соответствующей впадиной основано на моделировании пары выступ/впадина на основе трехмерной структуры, например, получаемой посредством рентгеновской кристаллографии или ядерного магнитного резонанса (ЯМР). Этого можно достигать способами, широко распространенными в этой области.
Термин "впадина" относится по меньшей мере к одной боковой цепи аминокислоты, углубленной относительно контактной поверхности второго полипептида и, таким образом, принимающей соответствующий выступ на смежной контактной поверхности первого полипептида. Впадина может существовать на исходной контактной поверхности, или ее можно встраивать синтетически (например, посредством изменения нуклеиновой кислоты, кодирующей контактную поверхность). Как правило, нуклеиновую кислоту, кодирующую контактную поверхность второго полипептида, изменяют так, что она кодирует впадину. Для достижения этого нуклеиновую кислоту, кодирующую по меньшей мере один исходный аминокислотный остаток в контактной поверхности второго полипептида, заменяют ДНК, кодирующей по меньшей мере один "импортируемый" аминокислотный остаток, имеющий меньший объем боковой цепи, чем исходный аминокислотный остаток. Верхним пределом количества исходных остатков, подвергаемых замене, является общее количество остатков в поверхности раздела второго полипептида. Некоторые импортируемые остатки для образования впадины, как правило, являются природными аминокислотными остатками и, предпочтительно, выбраны из аланина (A), серина (S), треонина (T) и валина (V).
В рамках изобретения термины "контактная поверхность", "остаток контактной поверхности", "аминокислота контактной поверхности", "контактный остаток" или "контактная аминокислота", как правило, относятся к любому аминокислотному остатку, присутствующему в домене, который может участвовать в контактах между первым полипептидом и вторым полипептидом.
"Исходный аминокислотный" остаток является остатком, заменяемым "импортируемым аминокислотным" остатком, который может иметь меньший или больший объем боковой цепи, чем исходный остаток. Импортируемый аминокислотный остаток может являться природным или неприродным аминокислотным остатком, но предпочтительно является первым из них. "Природные" аминокислотные остатки являются остатками, кодируемыми генетическим кодом. Термин "неприродный" аминокислотный остаток означает остаток, некодируемый генетическим кодом, но способный ковалентно связываться со смежными аминокислотными остатками полипептидной цепи. Примерами неприродных аминокислотных остатков являются норлейцин, орнитин, норвалин, гомосерин и другие аналоги аминокислотных остатков, такие как описываемые, например, в Ellman et al., Meth. Enzym. 202:301-336 (1991). В некоторых вариантах осуществления способ по настоящему изобретению может включать замену по меньшей мере одного исходного аминокислотного остатка, но можно заменять более одного исходного остатка. Как правило, не более чем общее количество остатков в контактной поверхности первого или второго полипептида может содержать исходные аминокислотные остатки, подвергаемые замене.
В рамках изобретения термин "конкурирует" в отношении антитела означает, что первое антитело или его антигенсвязывающий фрагмент (или часть) связывается с эпитопом, по существу, схоже со связыванием второго антитела или его антигенсвязывающей части, таким образом, что результат связывания первого антитела с его когнатным эпитопом детектируемо снижается в присутствии второго антитела по сравнению со связыванием первого антитела в отсутствие второго антитела. Альтернативный вариант, в котором связывание второго антитела с его эпитопом также детектируемо снижается в присутствии первого антитела, может иметь место, но не является обязательным. Т.е. первое антитело может ингибировать связывание второго антитела с его эпитопом без ингибирования вторым антителом связывания первого антитела с его соответствующим эпитопом. Однако если каждое антитело детектируемо ингибирует связывание другого антитела с его когнатным эпитопом или лигандом с той же, большей или меньшей степенью, указывают, что антитела "перекрестно конкурируют" друг с другом за связывание с их соответствующими эпитопами. Конкурирующие и перекрестно конкурирующие антитела входят в объем настоящего изобретения. Независимо от механизма, посредством которого происходит такая конкуренция или перекрестная конкуренция (например, стерическое препятствие, конформационное изменение или связывание с общим эпитопом или его частью), специалистам в этой области с учетом идей, представленных в настоящем описании, будет понятно, что предусмотрены такие конкурирующие и/или перекрестно конкурирующие антитела, и их можно использовать для способов, представленных в настоящем описании.
В рамках изобретения термин "антителозависимая клеточная цитотоксичность" или "ADCC" относится к клеточно-опосредованной реакции, при которой неспецифические цитотоксические клетки, экспрессирующие Fc-рецепторы (FcR) (например, естественные киллеры (NK), нейтрофилы и макрофаги), распознают связанное антитело на клетке-мишени, а затем вызывают лизис клетки-мишени. Активность ADCC интересующей молекулы можно оценивать с использованием анализа ADCC in vitro, как описано в патентах США №№ 5500362 или 5821337. Эффекторные клетки, которые можно использовать для таких анализов, включают мононуклеарные клетки периферической крови (PBMC) и NK-клетки. Альтернативно или дополнительно, активность ADCC интересующей молекулы можно оценивать in vivo, например, в модели на животных, как описано в Clynes et al., PNAS (USA), 95:652-656, 1998.
В рамках изобретения термин "обусловленная комплементом цитотоксичность" или "CDC" относится к лизису мишени в присутствии комплемента. Путь активации комплемента инициируется связыванием первого компонента системы комплемента (C1q) с молекулой (например, антителом), комплексированной с когнатным антигеном. Для оценки активации комплемента можно осуществлять анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods, 202: 163, 1996.
В рамках изобретения термины "иммуноспецифически связывается", "иммуноспецифически распознает", "специфически связывается", "специфически распознает" и аналогичные термины относятся к молекулам, например, связывающим доменам, специфически связывающимся с антигеном (например, эпитопом или иммунным комплексом) и не связывающимся специфически с другой молекулой. Молекула, специфически связывающаяся с антигеном, может связываться с другими пептидами или полипептидами с более низкой аффинностью, что определяют посредством анализов, известных в этой области, например, иммунологических анализов, BIACORE™ или других анализов. Предпочтительно, молекулы, специфически связывающиеся с антигеном, не реагируют перекрестно с другими белками.
Подходящие "условия умеренной строгости" включают предварительную промывку в растворе 5-кратного SSC, 0,5% SDS, 1,0 мМ ЭДТА (pH 8,0); гибридизацию при 50°C-65°C, 5-кратный SSC, в течение ночи; с последующей двукратной промывкой при 65°C в течение 20 минут каждым из 2-кратного, 0,5-кратного и 0,2-кратного SSC, содержащего 0,1% SDS.
В рамках изобретения "условия высокой строгости" являются следующими: (1) для промывки используют низкую ионную силу и высокую температуру, например, 0,015 M хлорида натрия/0,0015 M цитрата натрия/0,1% додецилсульфата натрия при 50°C; (2) во время гибридизации используют денатурирующее средство, такое как формамид, например, 50% (об./об.) формамида с 0,1% бычьего сывороточного альбумина/0,1% фиколла/0,1% поливинилпирролидона/50 мМ буфером фосфата натрия при pH 6,5 с 750 мМ хлорида натрия, 75 мМ цитрата натрия при 42°C; или (3) используют 50% формамида, 5-кратный SSC (0,75 M NaCl, 0,075 M цитрат натрия), 50 мМ фосфат натрия (pH 6,8), 0,1% пирофосфат натрия, 5-кратный раствор Денхардта, обработанную ультразвуком ДНК молок лососевых (50 мкг/мл), 0,1% SDS и 10% декстран сульфат при 42°C с промывками при 42°C в 0,2-кратном SSC (хлорид натрия/цитрат натрия) и 50% формамиде при 55°C с последующей промывкой высокой строгости, состоящей из 0,1-кратного SSC, содержащего ЭДТА при 55°C. Специалистам в этой области будет понятно, как корректировать температуру, ионную силу и т.д. по мере необходимости для адаптации к таким факторам, как длина зонда и т.п.
Связывающие белки, связывающие домены, CDR или антитела (определяемые в настоящем описании в широком смысле) можно идентифицировать в соответствии с определениями по Kabat, Chothia, совмещением Kabat и Chothia, AbM, Contact, North и/или конформационными определениями или любым способом определения CDR, хорошо известными в этой области. См., например, Kabat et al., 1991, Sequences of Proteins of Immunological Interest, 5th ed. (гипервариабельные области); Chothia et al., Nature 342:877-883, 1989 (структурные петлевые структуры). Идентичность аминокислотных остатков в конкретном антителе, составляющих CDR, можно определять способами, хорошо известными в этой области. Определение CDR по AbM представляет собой компромисс между Kabat и Chothia, и в нем используют программное обеспечение для моделирования антител Oxford Molecular's AbM (Accelrys®). Определение CDR по системе "Contact" основано на наблюдаемых контактах антигена, как описано в MacCallum et al., J. Mol. Biol., 262:732-745, 1996. "Конформационное" определение CDR основано на остатках, вносящих энтальпийный вклад в связывание антигена (см., например, Makabe et al., J. Biol. Chem., 283:1156-1166, 2008). North идентифицировал канонические конформации CDR с использованием другого предпочтительного набора определений CDR (North et al., J. Mol. Biol. 406: 228-256, 2011). В другом подходе, обозначенном в настоящем описании как "конформационное определение" CDR, положения CDR можно идентифицировать как остатки, вносящие энтальпийный вклад в связывание антигена (Makabe et al., J Biol. Chem. 283:1156-1166, 2008). При других определениях границ CDR могут не следовать строго одному из описанных выше подходов, но, несмотря на это, они будут перекрываться с по меньшей мере частью CDR по Kabat, хотя они могут быть короче или длиннее в свете прогнозирования или экспериментальных результатов, касающихся того, что конкретные остатки, или группы остатков или даже целые CDR, не влияют значительно на связывание антигена. В рамках изобретения термин "CDR" может относиться к CDR, определенным с помощью любого подхода, известного в этой области, включая комбинации подходов. В способах, используемых в настоящем описании, можно использовать CDR, определенные с помощью любого из этих подходов. В случае любого указанного варианта осуществления, содержащего более одной CDR, CDR (или другой остаток антитела) можно определять в соответствии с любым из определений по Kabat, Chothia, расширенного определения (комбинации Kabat и Chothia), North, расширенного определения, AbM, Contact и/или конформационных определений.
Остатки в вариабельном домене нумеруют по Kabat, что представляет собой систему нумерации, используемую для вариабельных областей тяжелой цепи или вариабельных областей легкой цепи компиляции антител. См., Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991. При использовании этой системы нумерации конкретная линейная аминокислотная последовательность может содержать меньше аминокислот или дополнительные аминокислоты, соответствующие укорочению или инсерции в FR или CDR вариабельной области. Например, вариабельная область тяжелой цепи может включать инсерцию одной аминокислоты (остаток 52a по Kabat) после остатка 52 H2 и встроенные остатки (например, остатки 82a, 82b и 82c по Kabat) после остатка FR тяжелой цепи 82. Нумерацию остатков по Kabat можно определять для указанного антитела посредством выравнивания областей гомологии последовательности антитела со "стандартной" пронумерованной по Kabat последовательностью. Доступны различные алгоритмы для нумерации по Kabat. Алгоритм, воплощенный в версии 2.3.3 выпуска Abysis (www.abysis.org), использовали в настоящем изобретении для приписывания нумерации Kabat вариабельным областям CDR1 VL, CDR2 VL, CDR3 VL, CDR2 VH и CDR3 VH. Определение AbM использовали для CDR1 VH. Конкретные положения аминокислотных остатков в антителе также можно нумеровать по Kabat.
В рамках изобретения термин "библиотека фагового дисплея" относится к популяции бактериофагов, каждый из которых содержит чужеродную кДНК, рекомбинантно слитую в рамке считывания с поверхностным белком. Фаг экспонирует чужеродный белок, кодируемый кДНК, на своей поверхности. После репликации в бактериальной клетке-хозяине, как правило, E. coli, фаг, содержащий интересующую чужеродную кДНК, подвергают селекции по экспрессии чужеродного белка на поверхности фага.
Термин "эпитоп" относится к части молекулы, которая может распознаваться, контактировать и/или быть связанной антителом в одной или более антигенсвязывающих областей антитела, известных как паратоп. Один антиген может иметь более одного эпитопа. Таким образом, разные антитела могут связываться с разными областями на антигене и могут иметь разные биологические эффекты. Эпитопы зачастую состоят из химически активной поверхностной группировки молекул, таких как аминокислоты или сахарные боковые цепи, и имеют специфические трехмерные структурные характеристики, а также специфические характеристики заряда. В рамках изобретения эпитопы могут являться конформационными или линейными. Конформационный эпитоп образован пространственно близкими аминокислотами из разных сегментов линейной полипептидной цепи. Линейный эпитоп является эпитопом, образованным смежными аминокислотными остатками в полипептидной цепи. В некоторых вариантах осуществления эпитоп может включать фрагменты сахаридов, фосфорильных групп или сульфонильных групп на антигене.
В рамках изобретения термин "антигенный эпитоп" определяют как часть полипептида, с которой антитело может специфически связываться, что определяют любым способом, хорошо известным в этой области, например, с помощью общепринятых иммунологических анализов. "Нелинейный эпитоп" или "конформационный эпитоп" содержит несмежные полипептиды (или аминокислоты) в антигенном белке, с которым связывается антитело, специфическое для эпитопа. После определения желаемого эпитопа на антигене можно получать антитела к этому эпитопу, например, способами, представленными в настоящем описании.
Термины "специфическое связывание" или "специфически связывающийся" при использовании по отношению к взаимодействию антитела и белка или пептида относятся к взаимодействию, зависящему от наличия конкретной структуры (т.е. антигенной детерминанты или эпитопа) на белке; другими словами, антитело распознает и связывается со специфической белковой структурой, а не с белками в целом. Например, если антитело является специфическим для эпитопа "A", наличие белка, содержащего эпитоп A (или свободного, немеченого A), в реакционной смеси, содержащей меченый "A" и антитело, будет снижать количество меченого A, связанного с антителом.
В некоторых вариантах осуществления термин "специфически связывается" означает, например, что антитело связывается с белком с KD приблизительно 0,1 нМ или менее, но более типично - менее приблизительно 1 мкМ. В некоторых вариантах осуществления термин "специфически связывается" означает, что антитело связывается с мишенью иногда с KD по меньшей мере приблизительно 0,1 мкМ или менее, а в других случаях - по меньшей мере, приблизительно 0,01 мкМ или менее, и в других случаях - по меньшей мере приблизительно 1 нМ или менее. Из-за идентичности последовательности между гомологичными белками у разных биологических видов, специфическое связывание может включать антитело, распознающее белок у более одного биологического вида (например, GUCY2c человека и GUCY2c мыши). Аналогично, из-за гомологии в некоторых областях полипептидных последовательностей разных белков, специфическое связывание может включать антитело, распознающее более одного белка. Следует понимать, что в некоторых вариантах осуществления антитело или связывающий фрагмент, специфически связывающийся с первой мишенью, может связываться или не связываться специфически со второй мишенью. В связи с этим, "специфическое связывание" не требует обязательно (хотя может включать) исключительное связывание, т.е. связывание с единственной мишенью. Таким образом, в некоторых вариантах осуществления антитело может специфически связываться с более чем одной мишенью. В некоторых вариантах осуществления множество мишеней может связываться одним и тем же антигенсвязывающим участком на антителе. Например, в некоторых случаях антитело может содержать два идентичных антигенсвязывающих участка, каждый из которых специфически связывается с одним и тем же эпитопом на двух или более белках. В некоторых альтернативных вариантах осуществления антитело может являться мультиспецифическим и содержать по меньшей мере два антигенсвязывающих участка с разными специфичностями. В качестве неограничивающего примера, биспецифическое антитело может содержать один антигенсвязывающий участок, распознающий эпитоп на одном белке (например, CD3 человека), и дополнительно содержать второй, иной антигенсвязывающий участок, распознающий другой эпитоп на втором белке. Как правило, но не обязательно, ссылка на связывание означает специфическое связывание.
Антитело, специфически связывающееся с антигеном, может связываться с другими пептидами или полипептидами с более низкой аффинностью, что определяют с помощью анализов, известных в этой области, например, иммунологических анализов, BIACORE™ или других анализов. Предпочтительно, антитело, специфически связывающееся с антигеном, не реагирует перекрестно с другими белками.
Термины "неспецифическое связывание" или "фоновое связывание" при использовании по отношению к взаимодействию антитела и белка или пептида относятся к взаимодействию, независящему от наличия конкретной структуры (т.е. антитело связывается с белками в целом, а не с конкретной структурой, такой как эпитоп).
В рамках изобретения термин "kon" или "ka" относится к константе скорости ассоциации антитела с антигеном. В частности, константы скорости (kon/ka и koff/kd) и равновесные константы диссоциации измеряют с использованием целого антитела (т.е. бивалентного) и мономерных белков GUCY2c.
В рамках изобретения термин "koff" или "kd" относится к константе скорости диссоциации антитела из комплекса антитело/антиген.
В рамках изобретения термин "KD" относится к равновесной константе диссоциации взаимодействия антитело-антиген.
В рамках изобретения термин "аффинность связывания", как правило, относится к силе суммы всех нековалентных взаимодействий между одним участком связывания молекулы (например, антитела) и его партнера по связыванию (например, антигена). Если не указано иначе, в рамках изобретения термин "аффинность связывания" относится к характерной аффинности связывания, отражающей взаимодействие 1:1 между членами связывающейся пары (например, антителом и антигеном). Аффинность молекулы X к ее партнеру Y, как правило, можно представлять с помощью константы диссоциации (KD). Аффинность молекулы X к ее партнеру Y, как правило, можно представлять с помощью константы диссоциации (Kd). Например, Kd может составлять приблизительно 200 нМ, 150 нМ, 100 нМ, 60 нМ, 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 8 нМ, 6 нМ, 4 нМ, 2 нМ, 1 нМ или менее. Аффинность можно измерять общепринятыми, известными в этой области способами, включая способы, представленные в настоящем описании. Низкоаффинные антитела, как правило, связываются с антигеном медленно и имеют склонность к быстрой диссоциации, в то время как высокоаффинные антитела, как правило, связываются с антигеном быстрее и имеют склонность к тому, чтобы оставаться связанными в течение большего периода времени. В этой области известны различные способы измерения аффинности связывания, любой из которых можно использовать в целях по настоящему изобретению. В частности, термин "аффинность связывания" предназначен для обозначения скорости диссоциации конкретного взаимодействия антиген-антитело. KD представляет собой соотношение скорости диссоциации, также называемой "скоростью обратной реакции (koff)", и скорости ассоциации или "скорости прямой реакции (kon)". Таким образом, KD равна koff/kon, и ее выражают как молярную концентрацию (M). Из этого следует, что, чем меньше KD, тем сильнее аффинность связывания. Таким образом, KD 1 мкМ свидетельствует о слабой аффинности связывания по сравнению с KD 1 нМ. Значения KD антител можно определять способами, хорошо известными в этой области. Одним из способов определения KD антитела является использование поверхностного плазмонного резонанса (SPR), как правило, с использованием биосенсорной системы, такой как система BIACORE™. Кинетический анализ BIACORE™ включает анализ связывания и диссоциации антигена с чипов с иммобилизованными молекулами (например, молекулами, содержащими эпитоп-связывающие домены) на их поверхности. Другим способом определения KD антитела является использование интерферометрия биослоя, как правило, с использованием технологии OCTET® (системы Octet QKe, ForteBio).
Термины "биологически активный", "биологическая активность" и "биологические характеристики" в отношении биспецифического антитела по настоящему изобретению, такого как антитело, фрагмент или его производное, означает обладание способностью связываться с биологической молекулой, если не указано иначе.
В рамках изобретения термины "нуклеиновые кислоты" и "нуклеотидные последовательности" включают молекулы ДНК (например, кДНК или геномную ДНК), молекулы РНК (например, мРНК), комбинации молекул ДНК и РНК или гибридные молекулы ДНК/РНК и аналоги молекул ДНК или РНК. Такие аналоги можно получать с использованием, например, аналогов нуклеотидов, включающих, в качестве неограничивающих примеров, инозин или тритилированные основания. Такие аналоги также могут содержать молекулы ДНК или РНК, содержащие модифицированные остовы, придающие молекулам полезные свойства, такие как, например, резистентность к нуклеазам или повышенную способность проникать через клеточные мембраны. Нуклеиновые кислоты или нуклеотидные последовательности могут являться одноцепочечными, двухцепочечными, могут содержать одноцепочечные и двухцепочечные части, и могут содержать трехцепочечные части, но предпочтительно, является двухцепочечной ДНК.
Настоящее изобретение также относится к полинуклеотидам, кодирующим антитела по изобретению, включая полипептиды и связывающие области антител. Можно получать Полинуклеотиды, кодирующие молекулы по изобретению, и определять нуклеотидную последовательность полинуклеотидов любым известным в этой области способом.
Полинуклеотиды, кодирующие антитела по настоящему изобретению, могут включать следующее: только кодирующую последовательность для варианта, кодирующую последовательность для варианта и дополнительные кодирующие последовательности, такие как функциональный полипептид, или сигнальную или секреторную последовательность или последовательность пробелка; кодирующую последовательность для антитела и некодирующую последовательность, такую как интроны или 5’- и/или 3’-некодирующая последовательность кодирующей последовательности антитела. Термин "полинуклеотид, кодирующий антитело" включает полинуклеотид, включающий дополнительную кодирующую последовательность для варианта, но также полинуклеотид, включающий дополнительную кодирующую и/или некодирующую последовательность. В этой области известно, что полинуклеотидная последовательность, оптимизированная для конкретной клетки-хозяина/экспрессирующей системы, легко можно получать из аминокислотной последовательности желаемого белка (см. GENEART® AG, Regensburg, Germany).
Антитела и их антигенсвязывающие фрагменты по настоящему изобретению могут иметь дополнительные консервативные замены аминокислот или замены заменимых аминокислот, не имеющие значительного эффекта в отношении функций полипептида. Будет ли конкретная замена хорошо переноситься или нет, т.е. не будет ли она негативно влиять на желаемые биологические свойства, такие как связывающая активность, можно определять, как описано в Bowie, JU et al. Science 247: 1306-1310,1990 или Padlan et al. FASEB J. 9: 133-139, 1995.
В рамках изобретения термин "выделенный" относится к материалу, удаленному из исходного окружения (например, природного окружения, если он является природным). Например, природный полинуклеотид или полипептид, присутствующий у живого животного, не является выделенным, но тот же полинуклеотид или полипептид, отделенный от некоторых или всех из сопутствующих материалов в природной системе, является выделенным. Такой полинуклеотид может являться частью вектора, и/или такой полинуклеотид или полипептид может являться частью композиции, например, смеси, раствора или суспензии, содержащей выделенную клетку или культивируемую клетку, содержащую полинуклеотид или полипептид, и все равно будет выделенным в том смысле, что вектор или композиция не является частью своего природного окружения.
В рамках изобретения термин "репликон" относится к любому генетическому элементу, такому как плазмида, хромосома или вирус, функционирующему в качестве автономной единицы репликации полинуклеотидов в клетке.
В рамках изобретения термин "функционально связанный" относится к ситуации, в которой описанные компоненты находятся во взаимосвязи, позволяющей им функционировать заранее определенным образом. Например, контрольную последовательность, "функционально связанную" с кодирующей последовательностью, лигируют таким образом, что достигают экспрессии кодирующей последовательности в условиях, подходящих или совместимых с контрольной последовательностью. В основном, термин "функционально связанный" означает, что соединенные последовательности ДНК являются смежными и, в случае секреторной лидерной последовательности, смежными и в рамке считывания. Однако энхансеры могут не являться смежными. Связывание осуществляют посредством лигирования по удобным участкам рестрикции. Если таких участков нет, используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с общепринятой практикой.
В рамках изобретения термин "вектор" означает конструкцию, с помощью которой можно доставлять и, предпочтительно, экспрессировать один или более интересующих генов или последовательностей в клетке-хозяине. Неограничивающие примеры векторов включают вирусные векторы, экспрессирующие векторы депротеинизированной ДНК или РНК, плазмиду, космиду или фаговые векторы, экспрессирующие векторы ДНК или РНК, ассоциированные с катионными конденсирующими средствами, экспрессирующие векторы ДНК или РНК, инкапсулированные в липосомах, и некоторые эукариотические клетки, такие как клетки-продуценты.
В рамках изобретения термин "последовательность контроля экспрессии" или "контрольная последовательность" относится к полинуклеотидной последовательности, необходимой для осуществления экспрессии кодирующей последовательности, с которой она лигирована. Природа таких контрольных последовательностей отличается в зависимости от организма-хозяина. Например, у прокариот такие контрольные последовательности, как правило, включают промотор, участок связывания рибосомы и терминаторы и, в некоторых случаях, энхансеры. Таким образом, термин "контрольная последовательность" предназначен для включения по меньшей мере всех компонентов, наличие которых является необходимым для экспрессии, а также может включать дополнительные компоненты, наличие которых является благоприятным, например, лидерные последовательности.
"Клетка-хозяин" включает отдельную клетку или культуру клеток, которая может являться или является реципиентом для векторов для включения полинуклеотидных вставок. Клетки-хозяева включают потомство одной клетки-хозяина, и потомство не обязательно должны быть полностью идентичными (по морфологии или по комплементарной геномной ДНК) исходной родительской клетке по причине природной, случайной или преднамеренной мутации. Клетка-хозяин включает клетки, трансфицированные in vivo с использованием полинуклеотидов по настоящему изобретению.
В рамках изобретения "клетки млекопитающих" включают ссылку на клетки, полученные из млекопитающих, включая людей, крыс, мышей, морских свинок, шимпанзе или макак. Клетки можно культивировать in vivo или in vitro.
В рамках изобретения термин "очищенный продукт" относится к препарату продукта, выделенному из клеточных составляющих, с которыми продукт в норме ассоциирован, и/или из других типов клеток, которые могут присутствовать в интересующем образце.
В рамках изобретения термин "по существу, чистый" относится к материалу, являющемуся по меньшей мере на 50% чистым (т.е. свободным от загрязнений), более предпочтительно - по меньшей мере на 90% чистым, более предпочтительно - по меньшей мере 95% чистым, еще более предпочтительно - по меньшей мере 98% чистым, и наиболее предпочтительно - по меньшей мере 99% чистым.
В рамках изобретения термин "злокачественное новообразование" или "злокачественный" относится к физиологическому состоянию у млекопитающих, как правило, отличающемуся нерегулируемым ростом клеток, неоплазией или опухолью, являющейся результатом аномального, неконтролируемого роста клеток. В некоторых аспектах термин "злокачественное новообразование" относится к злокачественной первичной опухоли без метастазирования, оставшейся локализованной. В других аспектах термин "злокачественное новообразование" относится к злокачественной опухоли, инвазирующей и разрушающе соседние структуры тела и распространяющейся в отдаленные очаги. В некоторых аспектах злокачественное новообразование ассоциировано со специфическим антигеном злокачественной опухоли. Неограничивающие примеры злокачественных новообразований включают злокачественное новообразование ротовой полости и глотки, пищеварительной системы (например, пищевода, желудка, тонкого кишечника, толстого кишечника, прямой кишки, анального канала, печени, желчного пузыря, аппендикса, желчных протоков и поджелудочной железы) или эндокринной системы. В некоторых вариантах осуществления злокачественное новообразование пищеварительной системы является раком пищевода, желудка, тонкого кишечника, толстого кишечника, прямой кишки, анального канала, печени, желчного пузыря, аппендикса, желчных протоков и поджелудочной железы.
В рамках изобретения термин "рак пищевода" предназначен для включения общепринятого медицинского определения, определяющего рак пищевода как медицинское состояние, отличающееся злокачественным новообразованием из клеток пищевода. Примеры злокачественного новообразования пищевода включают аденокарциному, плоскоклеточную карциному, хориокарциному, лимфому, саркому и мелкоклеточный рак.
Злокачественное новообразование "желудочно-кишечного тракта" (ЖКТ) представляет собой термин для группы злокачественных новообразований, поражающих пищеварительную систему. В рамках изобретения термин "рак желудка" предназначен для включения общепринятого медицинского определения, определяющего рак желудка как медицинское состояние, отличающееся злокачественным новообразованием из клеток желудка. В частности, рак желудка является заболеванием, при котором злокачественные клетки образуются в выстилке желудка. Рак желудка может развиваться в любой части желудка и может распространяться по всему желудку и в другие органы, в частности, пищевод, легкие и печень.
В рамках изобретения термин "колоректальный рак" предназначен для включения общепринятого медицинского определения, определяющего колоректальный рак как медицинское состояние, отличающееся злокачественным новообразованием из клеток желудочно-кишечного тракта ниже тонкого кишечника (т.е. толстого кишечника, включая слепую кишку, восходящую ободочную кишку, поперечную ободочную кишку, нисходящую ободочную кишку, сигмовидную кишку и прямую кишку). Кроме того, в рамках изобретения термин "колоректальный рак" предназначен для включения медицинских состояний, отличающихся злокачественным новообразованием из клеток двенадцатиперстной кишки и тонкого кишечника (тощей кишки и подвздошной кишки). Определение колоректального рака, используемое в настоящем описании, является более обширным, чем общепринятое медицинское определение, но приведено в таком виде, т.к. клетки двенадцатиперстной кишки и тонкого кишечника также содержат GUCY2c.
В рамках изобретения термин "злокачественная клетка" или "злокачественное новообразование" относится к опухолям или опухолевым клеткам, являющимся инвазивными и/или способными подвергаться метастазированию, т.е. злокачественным клеткам.
В рамках изобретения термин "лечить" или "лечение" означает подход для достижения благоприятных или желаемых клинических результатов. В целях по настоящему изобретению, лечение определяют как введение индивидууму, например, пациенту, молекулы антитела против GUCY2c (например, моноклонального антитела против GUCY2c или биспецифического или полиспецифического антитела против GUCY2c). Такое введение, например, можно осуществлять посредством прямого введения индивидууму или посредством нанесения на выделенную ткань или клетку из индивидуума, которую затем возвращают в организм индивидуума. Молекулу антитела против GUCY2c можно вводить в отдельности или в комбинации с одним или более средствами. Лечение может быть предназначено для излечения, заживления, ослабления, облегчения, изменения, восстановления, улучшения, временного облегчения или влияния на нарушение, симптомы нарушения или предрасположенность к нарушению, например, злокачественному новообразованию. В некоторых вариантах осуществления лечение является полезным при любом одном или более из следующего: (a) лечении, профилактике или улучшении одного или более симптомов состояния, ассоциированного со злокачественными клетками, экспрессирующими GUCY2c, у индивидуума (например, злокачественного новообразования желудочно-кишечного тракта, такого как колоректальный рак (CRC)); (b) ингибировании роста или прогрессирования опухоли у индивидуума, имеющего злокачественную опухоль, экспрессирующую GUCY2c; (c) ингибировании метастазирования злокачественных клеток, экспрессирующих GUCY2c, у индивидуума, имеющего одну или более злокачественных клеток, экспрессирующих GUCY2c; (d) индуцировании регрессирования (например, долговременного регрессирования) опухоли, экспрессирующей GUCY2c; (e) вызывании цитотоксической активности в злокачественных клетках, экспрессирующих GUCY2c; (f) повышении выживаемости без прогрессирования у индивидуума с GUCY2c-ассоциированным нарушением; (g) повышении общей выживаемости индивидуума с GUCY2c-ассоциированным нарушением; (h) снижении использования дополнительных химиотерапевтических или цитотоксических средств у индивидуума с GUCY2c-ассоциированным нарушением; (i) снижении опухолевой нагрузки у индивидуума с GUCY2c-ассоциированным нарушением; или (j) блокировании взаимодействия GUCY2c с другими еще не идентифицированными факторами. Не желая быть связанными какой-либо теорией, полагают, что лечение вызывает ингибирование, абляцию или уничтожение клетки in vitro или in vivo или иное снижение способности клетки, включая аномальную клетку, опосредовать нарушение, например, нарушение, представленное в настоящем описании, такое как злокачественное новообразование.
В рамках изобретения термин "противодиарейное средство" означает лекарственное средство, останавливающее или замедляющее диарею. В дополнение к поддерживающей терапии, которая может включать введение жидкости вместе с прекращением приема лактозы, алкоголя, продуктов с высокой осмолярностью, эти противодиарейные средства можно вводить совместно или в комбинации с биспецифическими антителами против GUCY2c. Противодиарейные средства включают, в качестве неограничивающих примеров, субгаллат висмута, Lactobacillus acidophilus, Saccharomyces boulardii, лоперамид/симетикон, атропин/дифеноксилат, атропин/дифеноксин, лиофилизат Saccharomyces boulardii, Lactobacillus acidophilus, лоперамид, субсалицилат висмута, Lactobacillus acidophilus/Lactobacillus bulgaricus, Lactobacillus rhamnosus, аттапульгит, крофелемер, фторхинолон, антибиотик или октреотид. См. Bensen et al. Recommended Guidelines for the Treatment of Cancer Treatment-Induced Diarrhea. Journal of Clinical Oncology 14: 2918-2926, 2004.
В рамках изобретения термин "индивидуум" предназначен для включения любого животного (например, млекопитающего), включая, в качестве неограничивающих примеров, людей, не являющихся человеком приматов, грызунов и т.п., которое будет реципиентом конкретного лечения. Например, индивидуум может являться пациентом (например, пациентом-человеком или ветеринарным пациентом), имеющим злокачественное новообразование. Как правило, термины "индивидуум" и "пациент" используют в настоящем описании взаимозаменяемо по отношению к человеку.
В рамках изобретения термин "не являющиеся человеком животные" включает всех не являющихся человеком позвоночных, например, не являющихся человеком млекопитающих и немлекопитающих, таких как не являющиеся человеком приматы, овца, собака, корова, курицы, амфибии, пресмыкающиеся, мышь, крыса, кролик или коза и т.д., если не указано иначе.
В рамках изобретения термин "фармацевтически приемлемый" относится к продукту или соединению, одобренному (или одобряемому) надзорным органом федерального правительства или правительства штата или указанному в Фармакопее США или других общепризнанных фармакопей для использования на животных, включая людей.
В рамках изобретения термины "фармацевтически приемлемый эксципиент, носитель или вспомогательное вещество" или "приемлемый фармацевтический носитель" относятся к эксципиенту, носителю или вспомогательному веществу, которое можно вводить индивидууму вместе с по меньшей мере одним антителом по настоящему изобретению и которое не нарушает активность антитела. Эксципиент, носитель или вспомогательное вещество должно являться нетоксичным при введении с антителом в дозах, достаточным для достижения терапевтического эффекта.
В рамках изобретения термин "улучшение" означает уменьшение или улучшение одного или более симптомов по сравнению с отсутствием введения молекулы антитела по изобретению. Термин "улучшение" также включает уменьшение длительности симптома.
В рамках изобретения термин "профилактика" относится к профилактике рецидивирования или дебюта одного или более симптомов нарушения у индивидуума в результате введения профилактического или терапевтического средства.
В рамках изобретения термины "эффективное количество", "терапевтически эффективное количество", "терапевтически достаточное количество" или "эффективная доза" относятся к любому количеству терапевтического средства, являющемуся эффективным или достаточным после введения однократной или многократных доз индивидууму для профилактики, заживления, улучшения или лечения заболевания, нарушения или побочного эффекта, или снижения скорости прогрессирования заболевания или нарушения, или пролонгирования излечения, облегчения или улучшения состояния индивидуума с нарушением, представленным в настоящем описании, за пределами ожидаемого в отсутствие такого лечения. Термин также включает количества, эффективные для усиления нормальной физиологической функции. Эффективное количество можно рассматривать в отношении введения одного или более терапевтических средств, и отдельное средство можно рассматривать для введения в эффективном количестве, если можно достигать или достигают желаемого результата одновременно с одним или более другими средствами.
В рамках изобретения термин "ингибирование роста" опухоли или злокачественного новообразования относится к замедлению, препятствию или прекращению роста и/или метастазирования и не обязательно свидетельствует об общей элиминации роста опухоли.
Активность является мерой активности терапевтического средства, выражаемой в терминах количества, необходимого для достижения эффекта с указанной интенсивностью. Высокоактивное средство вызывает больший ответ при низких концентрациях по сравнению со средством с более низкой активностью, вызывающим меньший ответ при низких концентрациях. Активность является функцией аффинности и эффективности. Термин "эффективность" относится к способности терапевтического средства вызывать биологический ответ после связывания с лигандом-мишенью и количественной величине этого ответа. В рамках изобретения термин "полумаксимальная эффективная концентрация (EC50)" относится к концентрации терапевтического средства, вызывающей ответ, величина которого представляет собой половину между исходным уровнем и максимумом после определенного времени воздействия. Терапевтическое средство может вызывать ингибирование или стимуляцию. Значение EC50 является общеупотребительным, и его используют в настоящем описании в качестве меры активности.
В рамках изобретения термин "комбинированное лечение" или введение "в комбинации с" относится к использованию более одного профилактического и/или терапевтического средства. Использование термина "комбинированное лечение" или "в комбинации" не ограничивает порядок, в котором профилактические и/или терапевтические средства вводят индивидууму с нарушением. Другими словами, комбинированное лечение можно осуществлять посредством раздельного, последовательного или одновременного лечения с использованием терапевтических средств. В случае "последовательного введения" первое вводимое средство может вызывать некоторый физиологический эффект в отношении индивидуума, когда второе средство вводят или оно становится активным в организме индивидуума.
В рамках изобретения термин "одновременное введение" в отношении введения профилактического и/или терапевтического средства относится к введению средств таким образом, что отдельные средства присутствуют в организме индивидуума одновременно. На одновременное введение могут влиять молекулы, составляемые в одной композиции или отдельных композициях, вводимых одновременно или близко по времени. При необходимости, последовательное введение можно осуществлять в любом порядке.
В рамках изобретения термин "GUCY2c" относится к гуанилатциклазе C (GUCY2c) млекопитающего, предпочтительно - белку GUCY2c человека. Термин "GUCY2c" можно использовать взаимозаменяемо с термином "GUCY2C". Нуклеотидная последовательность GUCY2c человека описана как регистрационный номер GenBank: NM.sub.-004963, включенный в настоящее описание в качестве ссылки. Аминокислотная последовательность GUCY2c человека описана как регистрационный номер GenBank NP.sub.-004954, включенный в настоящее описание в качестве ссылки.
Как правило, природный аллельный вариант имеет аминокислотную последовательность, по меньшей мере на 95%, 97% или 99% идентичную белку, представленному под регистрационным номером GenBank NP.sub.-004954. Белок GUCY2c охарактеризован как трансмембранный рецепторный белок поверхности клетки, и считают, что он играет критическую роль в поддержании жидкости кишечника, гомеостазе электролитов и пролиферации клеток.
В рамках изобретения термины "антитело, связывающееся с GUCY2c", "антитело, распознающее GUCY2c", "антитело против GUCY2c", "молекула антитела против GUCY2c" включает молекулу, в которой комбинируют по меньшей мере один связывающий домен антитела (как определено в настоящем описании) с по меньшей мере одним связывающим доменом антитела против GUCY2c (как определено в настоящем описании). Термин "молекула антитела против GUCY2c по настоящему изобретению" относится к антителам и их антигенсвязывающим фрагментам, взаимодействующим или распознающим, например, связывающим (например, связывающим специфически) GUCY2c, например, GUCY2c человека, GUCY2c мыши, GUCY2c крысы, GUCY2c яванского макака.
Термин "кластер дифференцировки 3" или "CD3" относится к мультимерному белковому комплексу, исторически известному как комплекс T3 и состоящему из четырех отдельных полипептидных цепей: эпсилон (ε), гамма (γ), дельта (δ), дзета (ζ), собирающихся и функционирующих как три пары димеров (εγ, εδ, ζζ). Комплекс CD3 служит в качестве T-клеточного корецептора, связанного нековалентно с T-клеточным рецептором (TCR) (Smith-Garvin et al. 2009) и генерирующего активационный сигнал в T-лимфоцитах. Белковый комплекс CD3 является определяющим признаком T-клеточного ростка, таким образом, антитела против CD3 можно эффективно использовать в качестве T-клеточных маркеров (Chetty and Gatter, Journal of Pathology, Vol 172, 4, 303-301 (1994)). Известно, что антитела против CD3 вызывают образование цитотоксических T-клеток посредством активации продукции эндогенных лимфокинов и способны селективно уничтожать опухолевые мишени (Yun et al., Cancer Research, 49:4770-4774, 1989).
Более конкретно, T-клетки экспрессируют комплексы TCR, способные индуцировать антигенспецифические иммунные ответы (Smith-Garvin et al., Annula Review of Immunology, 27: 1, 591-619 (2009)). Антигены являются пептидами, экспрессирующимися опухолевыми клетками и инфицированными вирусами клетками и способными стимулировать иммунные ответы. Внутриклеточно экспрессируемые антигены связаны с молекулами главного комплекса гистосовместимости класса I (MHC класса I) и транспортируются к поверхности, где они презентируются T-клеткам. Если аффинность связывания TCR с MHC класса I в комплексе с антигеном является достаточной, будет инициироваться образование иммунного синапса. Передача сигнала через иммунный синапс опосредован корецепторами CD3, образующими димеры εδ, εγ и ζζ. Эти димеры связываются с TCR и генерируют активационный сигнал в T-лимфоцитах. Этот сигнальный каскад направляет опосредованное T-клетками уничтожение клетки, экспрессирующей антиген. Цитотоксичность опосредована высвобождением и переносом гранзима B и перфорина из T-клетки в клетку-мишень.
В рамках изобретения термины "антитело, связывающееся с CD3", "антитело, распознающее CD3", "антитело против CD3", "молекула антитела против CD3" включает антитела и их антигенсвязывающие фрагменты, специфически распознающие одну субъединицу CD3 (например, эпсилон, дельта, гамма или дзета), а также антитела и их антигенсвязывающие фрагменты, специфически распознающие димерный комплекс из двух субъединиц CD3 (например, димеры гамма/эпсилон, дельта/эпсилон и дзета/дзета CD3). CD3 эпсилон человека приведен под регистрационным номером GenBank NM_000733. Антитела и антигенсвязывающие фрагменты по настоящему изобретению могут связываться с растворимым CD3 и/или CD3, экспрессирующимся на поверхности клетки. Растворимый CD3 включает природные белки CD3, а также рекомбинантные варианты белка CD3, такие как, например, мономерные и димерные конструкции CD3, в которых отсутствует трансмембранный домен или которые иначе не связаны с клеточной мембраной.
Антитела против CD3 могут генерировать активационный сигнал в T-лимфоцитах. Также можно использовать другие активирующие T-клетки лиганды, включая, в качестве неограничивающих примеров CD28, CD134, CD137 и CD27.
Биспецифические антитела против CD3 позволяют обходить потребность в вовлечении MHC-пептид/TCR и вместо этого рекрутируют T-клетки к клеткам-мишеням, экспрессирующим антиген на поверхности клетки. Одно плечо биспецифического антитела связывается с опухолеассоциированным антигеном на поверхности клетки, а другое плечо связывается с белком CD3, являющимся частью комплекса TCR на T-клетках. Совместное вовлечение CD3 на T-клетках и антигена-мишени на опухолевых клетках посредством биспецифического антитела приводит к цитотоксическому ответу. Таким образом, не желая быть связанными какой-либо теорией, полагают, что биспецифические антитела по настоящему изобретению могут позволить T-клетке обходить потребность во взаимодействии TCR и MHC класса I в комплексе с антигеном и вместо этого перенаправляют T-клетки к клеткам-мишеням через прямое совместное вовлечение CD3 (такого как CD3 эпсилон), экспрессирующегося на T-клетке, и GUCY2c, экспрессирующейся на опухоли.
В рамках изобретения термин "биспецифическое антитело против CD3-GUCY2c" относится к молекуле, сконструированной для направления T-клеток индивидуума на уничтожение злокачественных клеток посредством таргетинга опухолевых клеток, экспрессирующих желаемую молекулу. В некоторых вариантах осуществления желаемая молекула является GUCY2c человека. В некоторых вариантах осуществления биспецифическое антитело против CD3-GUCY2c содержит два FV-домена. В некоторых вариантах осуществления биспецифическое антитело против CD3-GUCY2c содержит первый FV-домен против GUCY2c и второй FV-домен против CD3. FV-домены могут являться scFV-доменами.
В рамках изобретения "первый полипептид" является любым полипептидом, подвергаемым связыванию со вторым полипептидом. Первый полипептид и второй полипептид соединены в области контакта. В дополнение к области контакта, первый полипептид может содержать один или более дополнительных доменов, таких как "связывающие домены" (например, вариабельный домен антитела, рецептор-связывающий домен, лиганд-связывающий домен или ферментативный домен) или константные домены антитела (или их части), включая домены CH2, CH1 и CL. Как правило, первый полипептид будет содержать по меньшей мере один домен, полученный из антитела. Этот домен, как правило, является константным доменом, таким как домен CH3 антитела, и может образовывать область контакта первого полипептида. Примеры первых полипептидов включают полипептиды тяжелой цепи антитела, химеры, в которых комбинируют константный домен антитела со связывающим доменом гетерологичного полипептида, рецепторные полипептиды, полипептиды-лиганды и полипептиды вариабельного домена антитела (например, биспецифические антитела).
В дополнение к области контакта, второй полипептид может содержать дополнительные домены, такие как "связывающий домен" (например, вариабельный домен антитела, рецептор-связывающий домен, лиганд-связывающий домен или ферментативный домен), или константные домены антитела (или их части), включая домены CH2, CH1 и CL. Как правило, второй полипептид будет содержать по меньшей мере один домен, полученный из антитела. Этот домен, как правило, является константной областью, такой как домен CH3 антитела, и может образовывать область контакта второго полипептида. Примеры вторых полипептидов включают полипептиды тяжелой цепи антитела, химеры, в которых комбинируют константный домен антитела со связывающим доменом гетерологичного полипептида, и полипептиды вариабельного домена антитела (например, биспецифические антитела).
В рамках изобретения термины "комплекс" или "комплексированный" относятся к связыванию двух или более молекул, взаимодействующих друг с другом с помощью связей и/или сил (например, вандерваальсовых, гидрофобных, гидрофильных сил), не являющихся пептидными связями. В одном из вариантов осуществления комплекс является гетеромультимерным. Следует понимать, что в рамках изобретения термин "белковый комплекс" или "полипептидный комплекс" включает комплексы, включающие небелковое вещество, конъюгированное с белком в белковом комплексе (например, включая, в качестве неограничивающих примеров, химические молекулы, такие как токсин или средство детекции).
Хотя в практическом осуществлении или тестировании настоящего изобретения можно использовать любые материалы и способы, схожие или эквивалентные материалам и способам, представленным в настоящем описании, далее будут описаны предпочтительные материалы и способы.
Материалы и способы
Описаны различные способы получения антител, включающие общепринятый гибридомный способ для получения моноклональных антител, рекомбинантные способы получения антител (включая химерные антитела, например, гуманизированные антитела), получение антител в трансгенных животных и недавно описанную технологию фагового дисплея для получения "полностью человеческих" антител. Эти способы вкратце описаны ниже.
Поликлональные антитела против интересующего антигена, как правило, можно индуцировать у животных посредством многократных подкожных (sc) или интраперитонеальных (ip) инъекций антигена и адъюванта. Антиген (или фрагмент, содержащий целевую аминокислотную последовательность) можно конъюгировать с белком, являющимся иммуногенным для биологического вида, подлежащего иммунизации, например, сывороточный альбумин, бычий тиреоглобулин или ингибитор трипсина сои, с использованием бифункционального или дериватизирующего средства, например малеимидобензоилсульфосукцинимидного сложного эфира (конъюгация через остатки цистеина) или N-гидроксисукцинимида (через остатки лизина). Животных иммунизируют с помощью иммуногенных конъюгатов или производных и через несколько недель животных подвергают бустерной иммунизации посредством подкожной инъекции во множестве участков. Через 7-14 дней у животных забирают кровь и анализируют сыворотку на титр антител. Животных подвергают бустерной иммунизации до достижения плато титра. Предпочтительно, животное подвергают бустерной иммунизации с использованием конъюгата того же антигена, но конъюгированного с другим белком и/или через другой перекрестно сшивающий реагент. Конъюгаты также можно получать в культуре рекомбинантных клеток в виде слитых белков. Кроме того, для усиления иммунного ответа используют средства, вызывающие агрегацию, такие как квасцы.
Моноклональные антитела можно получать из популяции, по существу, гомогенных антител с использованием гибридомного способа, впервые описанного в Kohler & Milstein, Nature 256:495, 1975, или можно получать способами рекомбинантной ДНК (Cabilly et al., патент США № 4816567). В гибридомном способе мышь или другого подходящего животного-хозяина иммунизируют, как описано выше, для индуцирования лимфоцитов, продуцирующих или способных продуцировать антитела, которые будут специфически связываться с белком, используемым для иммунизации. Альтернативно, лимфоциты можно иммунизировать in vitro. Затем лимфоциты подвергают слиянию с миеломными клетками с использованием подходящего средства для слияния, такого как полиэтиленгликоль, для получения гибридомной клетки (Goding, Monoclonal antibodies: Principles and Practice, pp. 59-103 (Academic Press, 1986)). Полученные таким образом гибридомные клетки высевают и выращивают в подходящей среде для культивирования, предпочтительно, содержащей одно или более веществ, ингибирующих рост или выживаемость неслитых, родительских миеломных клеток. Кроме того, гибридомные клетки можно выращивать in vivo в виде асцитных опухолей у животного. Моноклональные антитела, секретируемые субклонами, соответствующим образом отделяют от среды для культивирования, асцитной жидкости или сыворотки общепринятыми способами очистки иммуноглобулинов, такими как, например, с использованием протеин A-сефарозы, хроматография на гидроксиапатите, электрофорез в геле, диализ или аффинная хроматография.
Экспрессию антитела, антигенсвязывающих фрагментов антитела или любой конструкции антитела можно осуществлять в подходящих прокариотических или эукариотических клетках-хозяевах, подобных клеткам CHO, клеткам NSO, клеткам SP2/0, клеткам HEK293, клеткам COS, клеткам PER.C6, дрожжам или клеткам E. coli, и секретируемое антитело выделяют и собирают из клеток (супернатанта культуры клеток, кондиционированного супернатанта культуры клеток, лизата клеток или очищенной совокупности). Общие способы получения антител хорошо известны в этой области и описаны, например, в обзорных статьях Makrides, S.C., Protein Expr. Purif. 17:183-202, 1999; Geisse, S., et al., Protein Expr. Purif. 8:271-282, 1986; Kaufman, R.J., MoI. Biotechnol. 16:151-160, 2000; Werner, R.G., Drug Res. 48:870-880, 1998. В конкретном варианте осуществления культура клеток является культурой клеток млекопитающего, такой как культура клеток китайского хомяка (CHO).
В различных вариантах осуществления выделенные антитела можно подвергать дополнительным стадиям очистки с использованием общепринятых способов хроматографии, известных в этой области. В частности, способы очистки включают, в качестве неограничивающих примеров, аффинную хроматографию (например, аффинную хроматографию с протеином A), ионообменную хроматографию (например, анионообменную хроматографию или катионообменную хроматографию), хроматографию гидрофобных взаимодействий, хроматографию на гидроксиапатите, гель-фильтрацию и/или диализ. Среди них предпочтительным способом очистки является использование хроматографии с протеином A. Матрицей, к которой присоединяют аффинный лиганд, чаще всего является агароза, но доступны и другие матрицы.
В этой области также известны другие способы очистки антител, такие как фракционирование на ионообменной колонке, осаждение этанолом, обращенно-фазовая высокоэффективная хроматография, осаждение этанолом, обращенно-фазовая ВЭЖХ, хроматография на диоксиде кремния, хроматография на гепарин-сефарозе, хроматография на анионообменной или катионообменной смоле (например, на колонке с полиаспарагиновой кислотой), хроматофокусирование, электрофорез в ПААГ с SDS и осаждение сульфатом аммония. Изложенный выше список способов очистки представляет собой исключительно примеры, и не предназначен для ограничения перечисления.
Альтернативно, можно получать трансгенных животных (например, мышей), способных после иммунизации продуцировать полный репертуар антител человека в отсутствие продукции эндогенных иммуноглобулинов. Например, описано, что гомозиготная делеция гена соединительной области тяжелой цепи антитела (J.sub.H) у химерных и мутантных по зародышевой линии мышей приводит к полному ингибированию продукции эндогенных антител. Перенос набора генов иммуноглобулинов зародышевой линии человека таким мутантным по зародышевой мышам будет приводить к продукции антител человека после стимуляции антигеном. См., например, Jakobovits et al., Proc. Natl. Acad. Sci. USA 90:2551-255;1993 и Jakobovits et al., Nature 362:255-258, 1993.
В некоторых вариантах осуществления антитела по настоящему изобретению можно гуманизировать с сохранением высокой аффинности к антигену и других благоприятных биологических свойств. Способы гуманизации не принадлежащих человеку антител хорошо известны в этой области. Гуманизацию, по существу, можно осуществлять способом Winter с соавт. (Jones et al., Nature 321:522-525, 1986; Riechmann et al., Nature 332:323-327, 1988; Verhoeyen et al., Science 239:1534-1536, 1988) посредством замены CDR или последовательностей CDR грызуна соответствующими последовательностями антитела человека. Таким образом, такие гуманизированные антитела являются химерными антителами (Cabilly, выше), где вариабельный домен человека, по существу, меньший, чем интактный, заменяют соответствующей последовательностью не являющихся человеком видов. На практике, гуманизированные антитела, как правило, являются антителами человека, в которых некоторые остатки CDR и, возможно, некоторые остатки FR заменяют остатками из аналогичных участков в антителах грызуна. Важно гуманизировать антитела с сохранением высокой аффинность к антигену и других благоприятных биологических свойств. Для достижения этой цели по предпочтительному способу гуманизированные антитела получают способом анализа родительских последовательностей и различных концептуальных гуманизированных продуктов с использованием трехмерных моделей родительских и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов хорошо известны специалистам в этой области. Доступны компьютерные программы, с помощью которых иллюстрируют и отображают возможные трехмерные конформационные структуры выбранных последовательностей иммуноглобулинов-кандидатов. Изучение таких отображений позволяет анализировать возможную роль остатков в функционировании последовательности иммуноглобулина-кандидата, т.е. анализировать остатки, влияющие на способность иммуноглобулина-кандидата связываться с антигеном. Таким образом, остатки FR можно выбирать и комбинировать из консенсусной и импортируемой последовательности таким образом, что достигают желаемой характеристики антитела, такой как повышенная аффинность к антигенам-мишеням. Более подробно см. WO92/22653, опубликованный 23 декабря 1992 года.
Антитела по настоящему изобретению также можно получать с использованием различных способов фагового дисплея, известных в этой области. В способах фагового дисплея функциональные домены антитела экспонируются на поверхности фаговых частиц, несущих полинуклеотидные последовательности, кодирующие их. В конкретном аспекте такого фага можно использовать для экспонирования антигенсвязывающих доменов, таких как Fab и Fv или стабилизированной дисульфидной связью Fv, экспрессирующихся из репертуара или комбинаторной библиотеки антител (например, человека или мыши). Фаг, экспрессирующий антигенсвязывающий домен, связывающий интересующий антиген, можно подвергать селекции или идентифицировать с использованием антигена, например, с использованием меченого антигена или антигена, связанного или фиксированного на твердой поверхности или частице. Фаг, используемый в этих способах, как правило, является нитевидным фагом, включая fd и M13. Антигенсвязывающие домены экспрессируются в виде рекомбинантно слитого белка с белком гена III или гена VIII фага. Примеры способов фагового дисплея, которые можно использовать для получения иммуноглобулинов или их фрагментов по настоящему изобретению, включают описанные в Brinkmann et al. "Phage display Of Disulfide-Stabilized Fv Fragments," J. Immunol. Methods, 182:41-50, 1995.
Функциональные характеристики множества изотипов IgG и их доменов хорошо известны в этой области. Аминокислотные последовательности IgG1, IgG2, IgG3 и IgG4 известны в этой области. Выбор и/или комбинирование двух или более доменов из конкретных изотипов IgG для использования в способах по изобретению могут быть основаны на любом известном параметре родительских изотипов, включая аффинность к FcγR. Например, области или домены изотипов IgG, проявляющих ограниченное связывание с FcγRIIB или отсутствие связывания, например, IgG2 или IgG4, можно использовать, если биспецифическое антитело желательно конструировать для максимизации связывания с активирующим рецептором и минимизации связывания с ингибиторным рецептором. Аналогично, использование Fc-цепей или доменов из изотипов IgG, о которых известно, что они преференциально связываются с C1q или FcγRIIIA, например, IgG3 можно комбинировать с аминокислотными модификациями Fc, о которых в этой области известно, что они усиливают антителозависимую клеточную цитотоксичность (ADCC) и/или обусловленную комплементом цитотоксичность (CDC), для конструирования биспецифического антитела таким образом, что максимизируют активность эффекторной функции, например, активации комплемента или ADCC. Аналогично, можно получать мутации в Fc-цепях или доменах изотипов IgG, минимизирующие или устраняющие эффекторную функцию Fc-цепи.
Во время процесса поиска, с помощью получения и характеризации антител можно получать информацию о желаемых эпитопах. Затем с помощью этой информации можно осуществлять конкурентный скрининг антител на связывание с тем же эпитопом. Подходом для достижения этого является осуществление исследований конкуренции и перекрестной конкуренции для обнаружения антител, конкурирующих или перекрестно конкурирующих друг с другом, например, антител, конкурирующих за связывание с антигеном. Одним из способов является идентификация эпитопа, с которым связываются антитела, или "эпитопное картирование". Существует множество известных в этой области способов картирования и характеризации локализации эпитопов на белках, включая разрешение кристаллической структуры комплекса антитело-антиген, конкурентные анализы, анализы экспрессии фрагментов генов и анализы на основе синтетических пептидов, как описано, например, в главе 11 Harlow and Lane, Using Antibodies, a Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1999. В дополнительном примере эпитопное картирование можно использовать для определения последовательности, с которой связывается антитело. Эпитопное картирование коммерчески доступно в различных источниках, например, Pepscan Systems (Edelhertweg 15, 8219 PH Lelystad, The Netherlands). В дополнительном примере для идентификации остатков, требующихся, достаточных и/или необходимых для связывания эпитопа можно осуществлять мутагенез антигенсвязывающего домена, эксперименты по замене доменов и мутагенез с аланиновым сканированием. Аффинность связывания и скорость обратной реакции взаимодействия антигенсвязывающего домена можно определять с помощью анализов конкурентного связывания. Одним из примеров анализа конкурентного связывания является радиоиммунологический анализ, включающий инкубацию меченого антигена и детекцию молекулы, связанной с меченым антигеном. Аффинность молекулы по настоящему изобретению к антигену и скорости обратной реакции связывания можно определять из данных насыщения при анализе Скэтчарда.
Аффинности и связывающие свойства антител по настоящему изобретению в отношении антигена исходно можно определять с использованием анализов in vitro (биохимических или иммунологических анализов), известных в этой области для антигенсвязывающего домена, включая, в качестве неограничивающих примеров, твердофазный иммуноферментный анализ (ELISA), поверхностный плазмонный резонанс (SPR), интерферометрию биослоя или анализы иммунопреципитации. Молекулы по изобретению могут иметь связывающие свойства в моделях in vivo (таких как описанные и представленные в настоящем описании), схожие с таковыми в анализах in vitro. Однако настоящее изобретение не исключает молекулы по изобретению, не проявляющие желаемый фенотип в анализах in vitro, но проявляющий желаемый фенотип in vivo.
Необязательным форматом биспецифического антитела является Fv-стратегия на основе ковалентно связанной, биспецифической гетеродимерной структуры диатела, также известной как переориентирующееся антитело с двойной аффинностью (DART®), описанное, например, в патентных публикациях США №№ 2007/0004909, 2009/0060910 и 2010/0174053).
После получения последовательности нуклеиновой кислоты, кодирующей молекулы по изобретению (т.е. связывающие домены), можно получать вектор для получения молекул с помощью технологии рекомбинантных ДНК с использованием способов, хорошо известных в этой области.
Полинуклеотиды, кодирующие связывающие домены антитела (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c) по настоящему изобретению могут включать полинуклеотидную последовательность контроля экспрессии, функционально связанную с последовательностями, кодирующими антитело, включая ассоциированные в природе или гетерологичные промоторные области, известные в этой области. Последовательности контроля экспрессии могут представлять собой эукариотические промоторные системы в векторах, с помощью которых можно трансформировать или трансфицировать эукариотические клетки-хозяева, но также можно использовать контрольные последовательности для прокариотических хозяев. После встраивания вектора в подходящую линию клеток-хозяев, клетку-хозяина выращивают в условиях, подходящих для экспрессии нуклеотидных последовательностей и, при желании, для сбора и очистки антител. Эукариотические линии клеток включают линии клеток CHO, различные линии клеток COS, клетки HeLa, линии миеломных клеток, трансформированные B-клетки или линии эмбриональных клеток почки человека.
В одном из вариантов осуществления ДНК, кодирующую антитела по изобретению, выделяют и секвенируют общепринятыми способами (например, с использованием олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи антител мыши). Гибридомные клетки по изобретению служат предпочтительным источником такой ДНК. После выделения ДНК можно встраивать в экспрессирующие векторы, с помощью которых затем трансфицируют клетки-хозяева, такие как клетки обезьян COS, клетки CHO или миеломные клетки, которые в ином случае не продуцируют иммуноглобулиновый белок, для синтеза моноклональных антител в рекомбинантных клетках-хозяевах. ДНК также можно модифицировать, например, посредством замены кодирующей последовательностью константных доменов тяжелой и легкой цепи человека гомологичных последовательностей мышей, Morrison et al., Proc. Nat. Acad. Sci. 81:6851, 1984. Таким образом, получают химерные антитела, имеющие специфичность связывания моноклонального антитела против антигена по настоящему изобретению.
Как белок поверхности клетки, GUCY2c может служить в качестве терапевтической мишени для рецептор-связывающих белков, таких как антитела или лиганды. В нормальной ткани кишечника GUCY2c экспрессируется на апикальной стороне плотного контакта эпителиальных клеток, образующих барьер между средой просвета и сосудистым компартментом (Almenoff et al., Mol Microbiol 8: 865-873, 1993; и Guarino et al., Dig Dis Sci 32: 1017-1026, 1987). В связи с этим, системное внутривенное введение GUCY2c-связывающего белкового терапевтического средства будет иметь минимальный эффект в отношении кишечных рецепторов GUCY2c, присутствующих на нормальных клетках, при этом делая возможным доступ к неопластическим клеткам желудочно-кишечного тракта, включая злокачественные или метастазирующие клетки рака толстого кишечника, внекишечные или метастазирующие опухоли толстого кишечника, опухоли пищевода, опухоли желудка или аденокарциному в области гастроэзофагеального соединения. Кроме того, GUCY2c интернализуется посредством рецептор-опосредованного эндоцитоза после связывания лиганда (Buc et al. Eur J Cancer 41: 1618-1627, 2005; Urbanski et al., Biochem Biophys Acta 1245: 29-36, 1995).
Можно использовать тканеспецифическую экспрессию и ассоциацию GUCY2c со злокачественным новообразованием, например, желудочно-кишечного происхождения (например, раком толстого кишечника, раком желудка или раком пищевода), для использования GUCY2c в качестве диагностического маркера этого заболевания (Carrithers et al., Dis Colon Rectum 39:171-181, 1996; Buc et al. Eur J Cancer 41: 1618-1627, 2005).
Настоящее изобретение относится к антителу, связывающемуся с GUCY2c (например, GUCY2c человека (например, SEQ ID NO: 224, являющегося производным белка под регистрационным номером: NP_004954,2)), и которое можно использовать в любом одном или более из следующего: (a) лечении, профилактике или улучшении одного или более симптомов состояния, ассоциированного со злокачественными клетками, экспрессирующими GUCY2c, у индивидуума (например, злокачественного новообразования желудочно-кишечного новообразования, такого как колоректальный рак (CRC)); (b) ингибировании роста или прогрессирования опухоли у индивидуума, имеющего злокачественную опухоль, экспрессирующую GUCY2c; (c) ингибировании метастазирования злокачественных клеток, экспрессирующих GUCY2c, у индивидуума, имеющего одну или более злокачественных клеток, экспрессирующих GUCY2c; (d) индуцировании регрессирования (например, долговременного регрессирования) опухоли, экспрессирующей GUCY2c; (e) вызывании цитотоксической активности в отношении злокачественных клеток, экспрессирующих GUCY2c; (f) повышении выживаемости без прогрессирования у индивидуума с GUCY2c-ассоциированным нарушением; (g) повышении общей выживаемости индивидуума с GUCY2c-ассоциированным нарушением; (h) уменьшении использования дополнительных химиотерапевтических или цитотоксических средств у индивидуума с GUCY2c-ассоциированным нарушением; (i) снижении опухолевой нагрузки у индивидуума с GUCY2c-ассоциированным нарушением; или (j) блокировании взаимодействия GUCY2c с другими, еще не идентифицированными факторами.
В одном из аспектов настоящее изобретение относится к антителу, содержащему (a) вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область VH один (CDR1 VH), определяющую комплементарность область VH два (CDR2 VH) и определяющую комплементарность область VH три (CDR3 VH) последовательности VH, приведенной в SEQ ID NO: 11, 19, 26, 33, 41, 48, 52, 57, 60, 62, 64, 65, 67, 69, 71 или 73; и/или (b) вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область VL один (CDR1 VL), определяющую комплементарность область VL два (CDR2 VL) и определяющую комплементарность область VL три (CDR3 VL) последовательности VL, приведенной в SEQ ID NO: 92, 100, 104, 106, 112, 119, 125, 129, 134, 136, 137, 138, 140, 143, 145, 147, 150, 152, 156, 158, 160, 162, 166, 170, 171, 172, 173, 174 или 175.
В другом аспекте настоящее изобретение относится к антителу, имеющему любую из частичных последовательностей тяжелой цепи и/или любую из частичных последовательностей легкой цепи, приведенных в таблице 1. В одном из вариантов осуществления настоящее изобретение относится к антителу, содержащему область VH и/или область VL, где: (a) область VH содержит SEQ ID NO: 11, 19, 26, 33, 41, 48, 52, 57, 60, 62, 64, 65, 67, 69, 71 или 73; и/или (b) область VL содержит SEQ ID NO: 92, 100, 104, 106, 112, 119, 125, 129, 134, 136, 137, 138, 140, 143, 145, 147, 150, 152, 156, 158, 160, 162, 166, 170, 171, 172, 173, 174 или 175.
Таблица 1
(SEQ ID NO: 48)
В таблице 1 подчеркнутые последовательности представляют собой последовательности CDR по Kabat, а выделенные полужирным шрифтом - по Chothia. Определение AbM использовали для CDR1 VH.
Изобретение также относится к частям CDR антител против GUCY2c. Контактные области CDR являются областями антитела, придающими антителу специфичность к антигену. В основном, контактные области CDR включают положения остатков в CDR и зонах Вернье, ограниченных для поддержания правильной петлевой структуры, чтобы антитело связывалось со специфическим антигеном. См., например, Makabe et al., J. Biol. Chem., 283:1156-1166, 2007. Определение областей CDR хорошо известно в этой области. Алгоритм, воплощенный в версии 2.3.3 выпуска Abysis (www.abysis.org), использовали в настоящем изобретении для приписывания нумерации по Kabat вариабельным областям, CDR1 VL, CDR2 VL, CDR3 VL, CDR2 VH и CDR3 VH, за исключением CDR1 VH. Определение AbM использовали для CDR1 VH.
В одном из аспектов настоящее изобретение относится к антителу, имеющему любую из последовательностей CDR VH и/или любую из последовательностей CDR VL, приведенных в таблице 2. В одном из аспектов настоящее изобретение относится к антителу, специфически связывающемуся с гуанилатциклазой C (GUCY2c), где антитело содержит: (a) вариабельную область тяжелой цепи (VH), содержащую (i) определяющую комплементарность область VH один (CDR1 VH), содержащую SEQ ID NO: 12, 20, 27, 34, 42, 74, 257, 258, 259, 260 или 261; (ii) определяющую комплементарность область VH два (CDR2 VH), содержащую SEQ ID NO: 13, 21, 28, 35, 43, 53, 66, 68, 70, 72, 75, 262, 263, 264, 265, 266 или 267; и (iii) определяющую комплементарность область VH три (CDR3 VH), содержащую SEQ ID NO: 14, 22, 29, 36 или 44; и/или (b) вариабельную область легкой цепи (VL), содержащую (i) определяющую комплементарность область VL один (CDR1 VL), содержащую SEQ ID NO: 93, 101, 105, 107, 113, 120, 148, 153, 163 или 167; (ii) определяющую комплементарность область VL два (CDR2 VL), содержащую SEQ ID NO: 78, 94, 102, 108, 114, 141, 144, 146, 149, 151, 157, 159, 161, 164 или 168; и (iii) определяющую комплементарность область VH три (CDR3 VL), содержащую SEQ ID NO: 95, 109, 115, 121, 142, 154, 165 или 169.
В конкретном варианте осуществления настоящее изобретение относится к антителу, где: (a) область VH содержит (i) CDR1 VH, содержащую последовательность SEQ ID NO: 74 или 259; (ii) CDR2 VH, содержащую последовательность SEQ ID NO: 75 или 267; и iii) CDR3 VH, содержащую последовательность SEQ ID NO: 29; и/или (b) область VL содержит (i) CDR1 VL, содержащую последовательность SEQ ID NO: 148; (ii) CDR2 VL, содержащую последовательность SEQ ID NO: 149; и (iii) CDR3 VL, содержащую последовательность SEQ ID NO: 142.
Таблица 2
TYWMQ (SEQ ID NO: 257) (Kabat)
YPGDGM (SEQ ID NO: 262) (Chothia)
KYWMQ (SEQ ID NO: 258) (Kabat)
YPGDGF (SEQ ID NO: 263) (Chothia)
KPSNGL (SEQ ID NO: 264) (Chothia)
DTYIH (SEQ ID NO: 260) (Kabat)
DPANGN (SEQ ID NO: 265) (Chothia)
DYIML (SEQ ID NO: 261) (Kabat)
NPYYGS (SEQ ID NO: 266) (Chothia)
TYWMQ(SEQ ID NO: 257) (Kabat)
YPGDGM (SEQ ID NO: 262) (Chothia)
TYWMQ(SEQ ID NO: 257) (Kabat)
YPGDGM(SEQ ID NO: 262) (Chothia)
DYIML (SEQ ID NO: 261) (Kabat)
NPYYGS(SEQ ID NO: 266) (Chothia)
SYWMH (SEQ ID NO: 259) (Kabat)
KPSNGL(SEQ ID NO: 264) (Chothia)
SYWMH (SEQ ID NO: 259) (Kabat)
KPSNGL(SEQ ID NO: 264) (Chothia)
SYWMH (SEQ ID NO: 259) (Kabat)
KPSNGL (SEQ ID NO: 264) (Chothia)
SYWMH (SEQ ID NO: 259) (Kabat)
KPSNGL(SEQ ID NO: 264) (Chothia)
SYWMH (SEQ ID NO: 259) (Kabat)
KPSNGL (SEQ ID NO: 264) (Chothia)
SYWMH (SEQ ID NO: 259) (Kabat)
KPSNGL (SEQ ID NO: 264) (Chothia)
SYWMH (SEQ ID NO: 259) (Kabat)
KPSNGL (SEQ ID NO: 264) (Chothia)
SYWMH(SEQ ID NO: 259) (Kabat)
KPSNEL (SEQ ID NO: 267) (Chothia)
В одном из аспектов настоящее изобретение относится к антителу, имеющему любую из частичных последовательностей легкой цепи и/или любую из частичных последовательностей тяжелой цепи, приведенных в таблице 3. В одном из вариантов осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, специфически связывающемуся с CD3, где антитело содержит: область VH, содержащую CDR1 VH, CDR2 VH и CDR3 VH последовательности VH, приведенной в SEQ ID NO: 1, 9 или 273; и/или область VL, содержащую CDR1 VL, CDR2 VL и CDR3 VL последовательности VL, приведенной в SEQ ID NO: 76, 84, 90, 274, 275 или 276.
Таблица 3
VH 2B5-1039,
VH 2B5-1040
В таблице 3 подчеркнутые последовательности представляют собой последовательности CDR по Kabat, а выделенные полужирным шрифтом - по Chothia. Определение AbM использовали для CDR1 VH.
В таблице 4 приведены примеры последовательностей CDR для антител против CD3, представленных в настоящем описании.
Таблица 4
DYYMT(SEQ ID NO: 268) (Kabat)
RNRARGYT (SEQ ID NO: 269) (Chothia)
DYYMT(SEQ ID NO: 268) (Kabat)
RNQARGYT (SEQ ID NO: 270) (Chothia)
GFTFSDY (SEQ ID NO: 277) (Chothia)
GFTFSDYYMT (SEQ ID NO: 2) (расширенный)
RNQARGYT (SEQ ID NO: 270) (Chothia)
GFTFSDY (SEQ ID NO: 277) (Chothia)
GFTFSDYYMT (SEQ ID NO: 2) (расширенный)
RNQARGYT (SEQ ID NO: 270) (Chothia)
GFTFSDY (SEQ ID NO: 277) (Chothia)
GFTFSDYYMT
(SEQ ID NO: 2) (расширенный)
RNQARGYT (SEQ ID NO: 270) (Chothia)
(SEQ ID NO: 85)
В одном из вариантов осуществления настоящее изобретение относится к биспецифическому антителу, где: (a) CDR1 VL против GUCY2c содержит последовательность SEQ ID NO: 93, 101, 105, 107, 113, 120, 148, 153, 163 или 167; CDR2 VL против GUCY2c содержит последовательность SEQ ID NO: 78, 94, 102, 108, 114, 141, 144, 146, 149, 151, 157, 159, 161, 164 или 168; и CDR3 VL против GUCY2c содержит последовательность SEQ ID NO: 95, 109, 115, 121, 142, 154, 165 или 169; (b) CDR1 VH против CD3 содержит последовательность SEQ ID NO: 2, 268 или 277; CDR2 VH против CD3 содержит последовательность SEQ ID NO: 3, 10, 269 или 270; и CDR3 VH против CD3 содержит последовательность SEQ ID NO: 4; (c) CDR1 VL против CD3 содержит последовательность SEQ ID NO: 77, 85, 91, 278, 279 или 280; CDR2 VL против CD3 содержит последовательность SEQ ID NO: 78 или 281; и CDR3 VL против CD3 содержит последовательность SEQ ID NO: 79; и (d) CDR1 VH против GUCY2c содержит последовательность SEQ ID NO: 12, 20, 27, 34, 42, 74, 257, 258, 259, 260 или 261; CDR2 VH против GUCY2c содержит последовательность SEQ ID NO: 13, 21, 28, 35, 43, 53, 66, 68, 70, 72, 75, 262, 263, 264, 265, 266 или 267; и CDR3 VH против GUCY2c содержит последовательность SEQ ID NO: 14, 22, 29, 36 или 44.
В некоторых таких вариантах осуществления настоящее изобретение относится к биспецифическому антителу, где: (a) CDR1 VH против GUCY2c содержит последовательность SEQ ID NO: 74 или 259; (b) CDR2 VH против GUCY2c содержит последовательность SEQ ID NO: 75 или 267; (c) CDR3 VH против GUCY2c содержит последовательность SEQ ID NO: 29; (d) CDR1 VL против GUCY2c содержит последовательность SEQ ID NO: 148; (e) CDR2 VL против GUCY2c содержит последовательность SEQ ID NO: 149; и (f) CDR3 VL против GUCY2c содержит последовательность SEQ ID NO: 142.
В дополнительных вариантах осуществления описанного выше изобретение относится к биспецифическому антителу, где: (a) CDR1 VH против CD3 содержит последовательность SEQ ID NO: 2, 268, или 277; (b) CDR2 VH против CD3 содержит последовательность SEQ ID NO: 10 или 270; (c) CDR3 VH против CD3 содержит последовательность SEQ ID NO: 4; (d) CDR1 VL против CD3 содержит последовательность SEQ ID NO: 91, 278, 279 или 280; (e) CDR2 VL против CD3 содержит последовательность SEQ ID NO: 78 или 281; и (f) CDR3 VL против CD3 содержит последовательность SEQ ID NO: 79.
В конкретном варианте осуществления настоящее изобретение относится к полинуклеотиду, содержащему нуклеотидную последовательность, кодирующую вариабельные области тяжелой цепи и/или легкой цепи антитела, как представлено в настоящем описании и в таблице 5, таблице 37A и таблице 37B, включая: CD3-0001, CD3-0004, CD3-0006, GUCY2C-0074, GUCY2C-0077, GUCY2C-0078, GUCY2C-0098, GUCY2C-0104, GUCY2C-0105, GUCY2C-0240, GUCY2C-0315, GUCY2C-0179, GUCY2C-0193, GUCY2C-0210, GUCY2C-0212, GUCY2C-0241, GUCY2C-0247, GUCY2C-1186, GUCY2C-1467, GUCY2C-1478, GUCY2C-1481,GUCY2C-1512, GUCY2C-1518, GUCY2C-1526, GUCY2C-1527, GUCY2C-1538, GUCY2C-1554, GUCY2C-1555, GUCY2C-1556, GUCY2C-1557, GUCY2C-1590, GUCY2C-1591, GUCY2C-1592, GUCY2C-1608, huIGHV3-7, huIGKV1-39, GUCY2C-0405, GUCY2C-0486, GUCY2C-1640, GUCY2C-0250, GUCY2C-1678, GUCY2C-1679 или GUCY2C-1680.
В последовательностях, представленных в настоящем описании, приведены антитела против GUCY2c, антитела против CD3 и биспецифические антитела против CD3-GUCY2c. В некоторых вариантах осуществления эти антитела обозначают по ID их клона (например, GUCY2C-0098). Эти ID клонов можно использовать для обозначения зрелого антитела (например, IgG или биспецифического антитела против CD3). ID клона также может относиться к уникальным областям VH или VL, обнаруживаемым в этом антителе. В случае ID клонов, соответствующих биспецифическим антителам против CD3-GUCY2c (например, GUCY2C-0098), также можно упомянуть уникальную область антитела против CD3 (например, области VL и/или VH), включенную в зрелый белок. В таблице 5 приведен формат антитела и области против CD3 (если она есть), присутствующей в зрелом белке. В таблицах 37A и 37B дополнительно приведены последовательности компонентов этих зрелых антител против GUCY2c, IgG против CD3 и биспецифических антител против CD3-GUCY2c.
Таблица 5
Некоторые из антител против CD3, представленных в настоящем описании, можно обозначать несколькими названиями. Например, CD3-0001 можно обозначать как 2B5v1 или h2B5v1; CD3-0004 можно обозначать как 2B4 или h2B4; и CD3-0006 можно обозначать как 2B5v6 или h2B5v6.
Некоторые из антител против CD3, представленных в настоящем описании, включают, в качестве неограничивающих примеров, варианты CD3-0006 (также обозначаемые как 2B5v6 или h2B5v6). Например, варианты CD3-0006 включают, в качестве неограничивающих примеров, 2B5-1038, 2B5-1039 и 2B5-1040.
Аффинность связывания (KD) антитела, представленного в настоящем описании, к GUCY2c (такой как GUCY2c человека (например, SEQ ID NO: 224, 230 или 232)) или CD3 (SEQ ID NO: 242) может составлять от приблизительно 0,001 нМ до приблизительно 6500 нМ. В некоторых вариантах осуществления аффинность связывания составляет приблизительно 6500 нМ, 6000 нМ, 5500 нМ, 4500 нМ, 4000 нМ, 3500 нМ, 3000 нМ, 2500 нМ, 2000 нМ, 1500 нМ, 1000 нМ, 750 нМ, 500 нМ, 400 нМ, 300 нМ, 250 нМ, 200 нМ, 150 нМ, 100 нМ, 75 нМ, 50 нМ, 45 нМ, 40 нМ, 35 нМ, 30 нМ, 25 нМ, 20 нМ, 19 нМ, 17 нМ, 16 нМ, 15 нМ, 10 нМ, 8 нМ, 7,5 нМ, 7 нМ, 6,5 нМ, 6 нМ, 5,5 нМ, 5 нМ, 4 нМ, 3 нМ, 2 нМ, 1 нМ, 0,5 нМ, 0,3 нМ, 0,1 нМ, 0,01 нМ, 0,002 нМ или 0,001 нМ. В некоторых вариантах осуществления аффинность связывания составляет менее приблизительно 6500 нМ, 6000 нМ, 5500 нМ, 5000 нМ, 4000 нМ, 3000 нМ, 2000 нМ, 1000 нМ, 900 нМ, 800 нМ, 500 нМ, 250 нМ, 200 нМ, 100 нМ, 50 нМ, 30 нМ, 20 нМ, 10 нМ, 7,5 нМ, 7 нМ, 6,5 нМ, 6 нМ, 5 нМ, 4,5 нМ, 4 нМ, 3,5 нМ, 3 нМ, 2,5 нМ, 2 нМ, 1,5 нМ, 1 нМ, 0,5 нМ, 0,1 нМ, 0,05 нМ, 0,01 нМ или менее. В некоторых вариантах осуществления антитело имеет KD от приблизительно 80 до приблизительно 200 пМ, предпочтительно - от приблизительно от 100 до приблизительно 150 пМ.
В некоторых вариантах осуществления настоящее изобретение относится к антителу, связывающемуся с GUCY2c и/или CD3 и конкурирующему с антителом, представленным в настоящем описании, включая: CD3-0001, CD3-0004, CD3-0006, GUCY2C-0074, GUCY2C-0077, GUCY2C-0078, GUCY2C-0098, GUCY2C-0104, GUCY2C-0105, GUCY2C-0240, GUCY2C-0315, GUCY2C-0179, GUCY2C-0193, GUCY2C-0210, GUCY2C-0212, GUCY2C-0241, GUCY2C-0247, GUCY2C-1186, GUCY2C-1467, GUCY2C-1478, GUCY2C-1481, GUCY2C-1512, GUCY2C-1518, GUCY2C-1526, GUCY2C-1527, GUCY2C-1538, GUCY2C-1554, GUCY2C-1555, GUCY2C-1556, GUCY2C-1557, GUCY2C-1590, GUCY2C-1591, GUCY2C-1592, GUCY2C-1608, huIGHV3-7, huIGKV1-39, GUCY2C-0405, GUCY2C-0486, GUCY2C-1640, GUCY2C-0250, GUCY2C-1678, GUCY2C-1679 и GUCY2C-1680 (таблица 5).
В одном из аспектов настоящее изобретение относится к антителу, имеющему любую из последовательностей каркасной области, приведенных в таблице 6. В некоторых вариантах осуществления настоящее изобретение относится к антителу, представленному в настоящем описании, дополнительно содержащему человеческий или гуманизированный каркас VH и человеческий или гуманизированный каркас VL. В некоторых таких вариантах осуществления каркас VH содержит последовательность SEQ ID NO: 5, 6, 7, 8, 15, 16, 17, 18, 23, 24, 25, 30, 31, 32, 37, 38, 39, 40, 45, 46, 47, 49, 50, 51, 54, 55, 56, 58, 59, 61 или 63; и/или каркас VL содержит последовательность SEQ ID NO: 80, 81, 82, 83, 86, 87, 88, 89, 96, 97, 98, 99, 103, 110, 111, 116, 117, 118, 122, 123, 124, 126, 127, 128, 130, 131, 132, 133, 135, 139 или 155.
Таблица 6
В одном из вариантов осуществления настоящее изобретение относится к антителу, где область VH содержит последовательность SEQ ID NO: 73 или ее вариант с одной или несколькими консервативными аминокислотными заменами в остатках, не находящихся в области CDR; и/или где область VL содержит последовательность SEQ ID NO: 147 или ее вариант с одной или несколькими консервативными аминокислотными заменами в аминокислотах, не находящихся в области CDR.
В одном из аспектов настоящее изобретение относится к биспецифическому антителу, специфически связывающемуся с GUCY2c и CD3, где биспецифическое антитело содержит первую полипептидную цепь и вторую полипептидную цепь. В одном из вариантов осуществления первая полипептидная цепь содержит аминокислотную последовательность, приведенную в одной или более из SEQ ID NO: 216 и 220. В другом варианте осуществления вторая полипептидная цепь содержит аминокислотную последовательность, приведенную в одной или более из SEQ ID NO: 216 и 220. В предпочтительном варианте осуществления первая полипептидная цепь содержит аминокислотную последовательность SEQ ID NO: 216; и вторая полипептидная цепь содержит аминокислотную последовательность SEQ ID NO: 220. В некоторых таких вариантах осуществления биспецифическое антитело может специфически связываться с эпитопом GUCY2c и эпитопом CD3 и содержит первую полипептидную цепь и вторую полипептидную цепь, где первая полипептидная цепь содержит последовательность SEQ ID NO: 216, и вторая полипептидная цепь содержит последовательность SEQ ID NO: 220. В одном из вариантов осуществления вторая полипептидная цепь является любым полипептидом, подвергаемым связыванию с первой полипептидной цепью через область контакта.
В одном таком варианте осуществления настоящее изобретение относится к биспецифическому антителу, где: (a) первая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 1, содержащий VL антитела против GUCY2c (VL против GUCY2c) и VH антитела против CD3 (VH против CD3), и (ii) первый способствующий гетеродимеризации домен; и (b) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 2, содержащий VL антитела против CD3 (VL против CD3) и VH антитела против GUCY2c (VH против GUCY2c), и (ii) второй способствующий гетеродимеризации домен; где VL против GUCY2c и VH против GUCY2c образуют домен, специфически связывающийся с GUCY2c; и VL против CD3 и VH против CD3 образуют домен, специфически связывающийся с CD3.
В другом таком варианте осуществления (a) первая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 1, содержащий VL против CD3 и VH против GUCY2c, и (ii) первый способствующий гетеродимеризации домен; и (b) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 2, содержащий VL против GUCY2c и VH против CD3, и (ii) второй способствующий гетеродимеризации домен; где VL против GUCY2c и VH против GUCY2c образуют домен, специфически связывающийся с GUCY2c; и VL против CD3 и VH против CD3 образуют домен, специфически связывающийся с CD3.
В другом таком варианте осуществления (a) первая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 1, содержащий VH против GUCY2c и VL против CD3, и (ii) первый способствующий гетеродимеризации домен; и (b) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 2, содержащий VH против CD3 и VL против GUCY2c, и (ii) второй способствующий гетеродимеризации домен; где VH против GUCY2c и VL против GUCY2c образуют домен, специфически связывающийся с GUCY2c; и VH против CD3 и VL против CD3 образуют домен, специфически связывающийся с CD3.
В дополнительном варианте осуществления (a) первая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 1, содержащий VH против CD3 и VL против GUCY2c, и (ii) первый способствующий гетеродимеризации домен; и (b) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 2, содержащий VH против GUCY2c и VL против CD3, и (ii) второй способствующий гетеродимеризации домен, где VL против GUCY2c и VH против GUCY2c образуют домен, специфически связывающийся с GUCY2c; и VL против CD3 и VH против CD3 образуют домен, специфически связывающийся с CD3.
Изобретение также относится к выделенным полинуклеотидам, кодирующим антитела по изобретению, и векторам и клеткам-хозяевам, содержащим полинуклеотиды. В одном из аспектов полинуклеотид содержит нуклеотидную последовательность, кодирующую вариабельные области тяжелой цепи, вариабельные области легкой цепи, области CDR, каркасные области, Fc-цепи с выступами и впадинами, первые полипептидные цепи и вторые полипептидные цепи, приведенные в таблицах 1-7 и 38. В некоторых вариантах осуществления полинуклеотид содержит последовательность, кодирующую любое из антител CD3-0001, CD3-0004, CD3-0006, GUCY2C-0074, GUCY2C-0077, GUCY2C-0078, GUCY2C-0098, GUCY2C-0104, GUCY2C-0105, GUCY2C-0240, GUCY2C-0315, GUCY2C-0179, GUCY2C-0193, GUCY2C-0210, GUCY2C-0212, GUCY2C-0241, GUCY2C-0247, GUCY2C-1186, GUCY2C-1467, GUCY2C-1478, GUCY2C-1481, GUCY2C-1512, GUCY2C-1518, GUCY2C-1526, GUCY2C-1527, GUCY2C-1538, GUCY2C-1554, GUCY2C-1555, GUCY2C-1556, GUCY2C-1557, GUCY2C-1590, GUCY2C-1591, GUCY2C-1592, GUCY2C-1608, huIGHV3-7, huIGKV1-39, GUCY2C-0405, GUCY2C-0486, GUCY2C-1640, GUCY2C-0250, GUCY2C-1678, GUCY2C-1679 или GUCY2C-1680.
Последовательность, кодирующую интересующее антитело, можно поддерживать в векторе в клетке-хозяине, а затем клетку-хозяина можно выращивать и замораживать для дальнейшего использования. Векторы (включая экспрессирующие векторы) и клетки-хозяева дополнительно представлены в настоящем описании.
Изобретение также относится к слитым белкам, содержащим один или более фрагментов или областей из антител по настоящему изобретению. В одном из вариантов осуществления настоящее изобретение относится к слитому полипептиду, содержащему по меньшей мере 10 смежных аминокислот из вариабельной области легкой цепи, приведенной в SEQ ID NO: 76, 84, 90, 92, 100, 104, 106, 112, 119, 125, 129, 134, 136, 137, 138, 140, 143, 145, 147, 150, 152, 156, 158, 160, 162, 166, 170, 171, 172, 173, 174, 175, 274, 275 или 276; и/или по меньшей мере 10 смежных аминокислот из вариабельной области тяжелой цепи, приведенной в SEQ ID NO: 1, 9, 11, 19, 26, 33, 41, 48, 52, 57, 60, 62, 64, 65, 67, 69, 71, 73 или 273. В других вариантах осуществления настоящее изобретение относится к слитому полипептиду, содержащему по меньшей мере приблизительно 10, по меньшей мере приблизительно 15, по меньшей мере приблизительно 20, по меньшей мере приблизительно 25 или по меньшей мере приблизительно 30 смежных аминокислот из вариабельной области легкой цепи и/или по меньшей мере приблизительно 10, по меньшей мере приблизительно 15, по меньшей мере приблизительно 20, по меньшей мере приблизительно 25 или по меньшей мере приблизительно 30 смежных аминокислот из вариабельной области тяжелой цепи. В другом варианте осуществления слитый полипептид содержит вариабельную область легкой цепи и/или вариабельную область тяжелой цепи, приведенные в любой из пар последовательностей, выбранных из SEQ ID NO: 73 и 147;69 и 147; 60 и 138; 48 и 125; или 26 и 106. В другом варианте осуществления слитый полипептид содержит одну или более CDR. В других вариантах осуществления слитый полипептид содержит CDR H3 (CDR3 VH) и/или CDR L3 (CDR3 VL). В целях по настоящему изобретению слитый белок содержит одно или более антител и другую аминокислотную последовательность, к которой оно не присоединено в нативной молекуле, например, гетерологичную последовательность или гомологичную последовательность из другой области. Неограничивающие примеры гетерологичных последовательностей включают метку, такую как метка FLAG или метка 6His. Метки хорошо известны в этой области. Слитый полипептид можно получать известными в этой области способами, например, синтетически или рекомбинантно. Как правило, слитые белки по настоящему изобретению получают, получая экспрессирующий полинуклеотид, кодирующий их, рекомбинантными способами, представленными в настоящем описании, хотя их также можно получать другими способами, известными в этой области, включая, например, химический синтез.
Изобретение также относится к scFv антител по настоящему изобретению. Одноцепочечные фрагменты вариабельной области получают посредством связывания вариабельных областей легкой и/или тяжелой цепи с использованием короткого соединительного пептида (Bird et al., Science 242:423-426, 1988). Примером соединительного пептида является GGGGSGGGGSGGGGSG (SEQ ID NO: 193), служащего мостиком длиной приблизительно 3,5 нм между карбокси-концом одной вариабельной области и амино-концом другой вариабельной области. Можно конструировать и использовать линкеры с другими последовательностями (Bird et al., Science 242:423-426, 1988). Линкеры должны быть короткими, гибкими полипептидами и, предпочтительно, содержат менее приблизительно 20 аминокислотных остатков. В свою очередь, линкеры можно модифицировать для дополнительных функций, таких как присоединение лекарственных средств или прикрепление к твердым подложкам. Одноцепочечные варианты можно получать рекомбинантно или синтетически. Для синтетического получения scFv можно использовать автоматический синтезатор. Для рекомбинантного получения scFv подходящую плазмиду, содержащую полинуклеотид, кодирующий scFv, можно встраивать в подходящую клетку-хозяина, эукариотическую, такую как дрожжевые клетки, растительные клетки, клетки насекомых или клетки млекопитающих, или прокариотическую, такую как E. coli. Полинуклеотиды, кодирующие интересующие scFv, можно получать общепринятыми способами, такими как лигирование полинуклеотидов. Полученный scFv можно выделять стандартными способами очистки белков, известными в этой области.
Также предусмотрены другие формы одноцепочечных антител, такие как диатела или минитела. Диатела являются бивалентными, биспецифическими антителами, в которых вариабельные области тяжелой цепи (VH) и вариабельные области легкой цепи (VL) экспрессируются на одной полипептидной цепи, но с использованием линкера, слишком короткого для спаривания между двумя доменами на одной цепи, что, таким образом, заставляет домены спариваться с комплементарными доменами другой цепи и приводит к образованию двух антигенсвязывающих участков (см. например, Holliger, P., et al., Proc. Natl. Acad Sci. USA 90:6444-6448, 1993; Poljak, R. J., et al., Structure 2:1121-1123, 1994). Минитело включает области VL и VH нативного антитела, слитые с шарнирной областью и домена CH3 молекулы иммуноглобулина. См., например, патент США № 5837821.
В одном из аспектов настоящее изобретение относится к композициям (таким как фармацевтические композиции), содержащим любые из полинуклеотидов по изобретению. В некоторых вариантах осуществления композиция содержит экспрессирующий вектор, содержащий полинуклеотид, кодирующий любые из антител, представленных в настоящем описании. В других вариантах осуществления композиция содержит любой или оба из полинуклеотидов, приведенных в SEQ ID NO: 246 (кодирующей первую полипептидную цепь GUCY2C-1608) и SEQ ID NO: 247 (кодирующей вторую полипептидную цепь GUCY2C-1608). Экспрессирующие векторы и введение композиций полинуклеотидов представлены в настоящем описании далее.
Полинуклеотиды, комплементарные любым таким последовательностям, также включены в настоящее изобретение. Полинуклеотиды могут являться одноцепочечными (кодирующими или антисмысловыми) или двухцепочечными и могут представлять собой молекулы ДНК (геномной, кДНК или синтетической) или РНК. Молекулы РНК включают зрелую и незрелую мРНК, такую как предшественник мРНК (пре-мРНК) или гетерогенные ядерные мРНК (hnRNA) и зрелые мРНК. Дополнительные кодирующие или некодирующие последовательности, необязательно, могут присутствовать в полинуклеотиде по настоящему изобретению, и полинуклеотид, необязательно, можно связывать с другими молекулами и/или материалами подложек.
Полинуклеотиды могут содержать нативную последовательность (т.е. эндогенную последовательность, кодирующую антитело или его часть) или вариант такой последовательности. Варианты полинуклеотидов содержат одну или более замен, добавлений, делеций и/или инсерций таким образом, что иммунореактивность кодируемого полипептида не снижается относительно нативной иммунореактивной молекулы. Эффект в отношении иммунореактивности кодируемого полипептида, как правило, можно оценивать, как представлено в настоящем описании. Предпочтительно, варианты демонстрируют по меньшей мере приблизительно 70% идентичности, более предпочтительно - по меньшей мере приблизительно 80% идентичности, еще более предпочтительно - по меньшей мере приблизительно 90% идентичности, и наиболее предпочтительно - по меньшей мере приблизительно 95% идентичности в отношении полинуклеотидной последовательности, кодирующей нативное антитело или его часть.
Указывают, что две полинуклеотидные или полипептидные последовательности являются "идентичными", если последовательность нуклеотидов или аминокислот в двух последовательностях является одинаковой при выравнивании для максимального соответствия, как описано ниже. Сравнение двух последовательностей, как правило, осуществляют посредством сравнения последовательностей в окне сравнения для идентификации и сравнения локальных областей сходства последовательностей. В рамках изобретения термин "окно сравнения" относится к сегменту из по меньшей мере приблизительно 20 смежных положений, как правило, от 30 до приблизительно 75 или от 40 до приблизительно 50, в которых последовательность можно сравнивать с референсной последовательностью с таким же количеством смежных положений после оптимального выравнивания двух последовательностей.
Оптимальное выравнивание последовательностей для сравнения можно осуществлять с использованием программы Megalign в пакете биоинформатического программного обеспечения Lasergene (DNASTAR, Inc., Madison, WI) с использованием параметров по умолчанию. Предпочтительно, "процент идентичности последовательности" определяют посредством сравнения двух оптимально выровненных последовательностей в окне сравнения из по меньшей мере 20 положений, где часть полинуклеотидной или полипептидной последовательности в окне сравнения может содержать добавления или делеции (т.е. пропуски) 20 процентов или менее, как правило, от 5 до 15 процентов или от 10 до 12 процентов, по сравнению с референсными последовательностями (несодержащими добавления или делеции) для оптимального выравнивания двух последовательностей. Процент вычисляют посредством определения количества положений, в которых встречаются идентичные основания нуклеиновой кислоты или аминокислотные остатки в обеих последовательностях для получения количества совпавших положений, разделяя количество совпавших положений на общее количество положений в референсной последовательности (т.е. размер окна) и умножая результат на 100 для получения процента идентичности последовательности.
Варианты, дополнительно или альтернативно, могут являться, по существу, гомологичными нативному гену или его части или комплементарной последовательности. Такие варианты полинуклеотидов могут гибридизоваться в умеренно строгих условиях с природной последовательностью ДНК, кодирующей нативное антитело (или комплементарную последовательность).
Специалистам в этой области следует понимать, что в результате вырожденности генетического кода существует множество нуклеотидных последовательностей, кодирующих полипептид, представленный в настоящем описании. Некоторые из этих полинуклеотидов имеют минимальную гомологию по отношению к нуклеотидной последовательности любого нативного гена. Несмотря на это, полинуклеотиды, варьирующиеся из-за различий в использовании кодонов, конкретно включены в настоящее изобретение. Кроме того, аллели генов, содержащие полинуклеотидные последовательности, представленные в настоящем описании, входят в объем настоящего изобретения. Аллели представляю собой эндогенные гены, измененные в результате одной или более мутаций, таких как делеции, добавления и/или замены нуклеотидов. Образующиеся мРНК и белок, необязательно, могут иметь измененную структуру или функцию. Аллели можно идентифицировать стандартными способами (такими как гибридизация, амплификация и/или сравнение последовательностей из баз данных).
В одном из аспектов настоящее изобретение относится к способу получения любых из полинуклеотидов, представленных в настоящем описании. Например, полинуклеотиды по настоящему изобретению можно получать с использованием химического синтеза, рекомбинантных способов или ПЦР. Способы химического синтеза полинуклеотидов хорошо известны в этой области и не требуют подробного обсуждения в настоящем описании. Специалист в этой области может использовать последовательности, представленные в настоящем описании, и коммерческий синтезатор ДНК для получения желаемой последовательности ДНК.
Для получения полинуклеотидов рекомбинантными способами полинуклеотид, содержащий желаемую последовательность, можно встраивать в подходящий вектор и вектор, в свою очередь, можно встраивать в подходящую клетку-хозяина для репликации и амплификации, как представлено в настоящем описании. Полинуклеотиды можно встраивать в клетки-хозяева любыми способами, известными в этой области. Клетки трансформируют посредством встраивания экзогенного полинуклеотида посредством прямого захвата, эндоцитоза, трансфекции, F-скрещивания или электропорации. После встраивания экзогенный полинуклеотид можно поддерживать в клетке в виде неинтегрированного вектора (такого как плазмида) или интегрировать в геном клетки-хозяина. Амплифицированный таким образом полинуклеотид можно выделять из клетки-хозяина способами, хорошо известными в этой области (например, Sambrook et al., 1989).
Альтернативно, ПЦР позволяет воспроизводить последовательности ДНК. Технология ПЦР хорошо известна в этой области и описана в патентах США №№ 4683195, 4800159, 4754065 и 4683202, а также PCR: Polymerase Chain Reaction, Mullis et al. eds., Birkauswer Press, Boston, 1994.
РНК можно получать с использованием выделенной ДНК в подходящем векторе, встраивая его в подходящую клетку-хозяина. Когда клетка делится, и ДНК транскрибируется в РНК, РНК можно выделять способами, хорошо известными специалистам в этой области и описанными, например, в Sambrook et al., 1989, выше.
Подходящие клонирующие векторы можно конструировать стандартными способами или можно выбирать из большого количества клонирующих векторов, доступных в этой области. Хотя выбранный клонирующий вектор может варьироваться в зависимости от используемой клетки-хозяина, клонирующие векторы, которые можно использовать, как правило, будут обладать способностью к саморепликации, могут иметь одну мишень для конкретной эндонуклеазы рестрикции и/или могут нести гены для маркера, который можно использовать в селекции клонов, содержащих вектор. Подходящие примеры включают плазмиды и бактериальные вирусы, например, pUC18, pUC19, Bluescript (например, pBS SK+) и их производные, mp18, mp19, pBR322, pMB9, ColE1, pCR1, RP4, фаговую ДНК и челночные векторы, такие как pSA3 и pAT28. Эти и многие другие клонирующие векторы доступны в коммерческих источниках, таких как BioRad, Strategene и Invitrogen.
Экспрессирующие векторы, как правило, являются реплицируемыми полинуклеотидными конструкциями, содержащими полинуклеотид по изобретению. Следует понимать, что экспрессирующий вектор должен реплицироваться в клетках-хозяевах в виде эписом или интегральной части хромосомной ДНК. Подходящие экспрессирующие векторы включают, в качестве неограничивающих примеров, плазмиды, вирусные векторы, включая аденовирусы, аденоассоциированные вирусы, ретровирусы, космиды, и экспрессирующие векторы, описанные в публикации PCT № WO87/04462. Компоненты вектора, как правило, могут включать в качестве неограничивающих примеров, одно или более из следующего: сигнальную последовательность; участок начала репликации; один или более маркерных генов; подходящие элементы контроля транскрипции (такие как промоторы, энхансеры и терминатор). Для экспрессии (т.е. трансляции), как правило, также необходим один или более элементов контроля трансляции, такие как участки связывания рибосомы, участки инициации трансляции и стоп-кодоны.
Векторы, содержащие интересующие полинуклеотиды, можно встраивать в клетку-хозяина любым из ряда подходящих способов, включая электропорацию, трансфекцию с использованием хлорида кальция, хлорида рубидия, фосфата кальция, DEAE-декстрана или других веществ; бомбардировку микрочастицами; липофекцию и инфекцию (например, если вектор является возбудителем инфекции, таким как вирус осповакцины). Выбор встраиваемых векторов или полинуклеотидов зачастую будет зависеть от признаков клетки-хозяина.
Для выделения генов, кодирующих интересующее антитело, полипептид или белок, можно использовать любые клетки-хозяева, способные гиперэкспрессировать гетерологичную ДНК. Неограничивающие примеры клеток-хозяев млекопитающего включают клетки COS, HeLa и CHO. Также см. публикацию PCT № WO87/04462. Подходящие клетки-хозяева, не принадлежащие млекопитающим, включают прокариот (таких как E. coli или B. subtillis) и дрожжей (таких как S. cerevisae, S. pombe или K. lactis). Предпочтительно, клетки-хозяева экспрессируют кДНК на уровне приблизительно в 5 раз выше, более предпочтительно - в 10 раз выше, даже более предпочтительно - в 20 раз выше, чем у соответствующего эндогенного антитела, или белка GUCY2c, или домена GUCY2c (например, доменов 1-4), что определяют посредством иммунологического анализа или FACS. Можно идентифицировать клетку, гиперэкспрессирующую интересующее антитело или белок.
В одном из аспектов молекула антитела против GUCY2c будет иметь аффинность к GUCY2c, например, что измеряют посредством прямых анализов связывания или конкурентных анализов связывания в диапазоне аффинности от пикомоль до микромоль, предпочтительно - в диапазоне от пикомоль до низких наномолярных значений.
Биспецифические антитела можно получать с использованием антител, представленных в настоящем описании. Способы получения биспецифических антител известны в этой области (см., например, Suresh et al., Methods in Enzymology 121:210, 1986). Общепринято, рекомбинантная продукция биспецифических антител основана на коэкспрессии двух пар тяжелая цепь-легкая цепь иммуноглобулина, при этом две тяжелые цепи имеют разные специфичности (Millstein and Cuello, Nature 305, 537-539, 1983).
В другом варианте осуществления биспецифическое антитело, представленное в настоящем описании, содержит полноразмерное антитело человека, где вариабельная область антитела из гетеродимерного белка может рекрутировать активность иммунной эффекторной клетки человека посредством специфического связывания с эффекторным антигеном (например, антигеном CD3), находящимся на иммунной эффекторной клетке человека, и где вторая вариабельная область антитела из гетеродимерного белка может специфически связываться с антигеном-мишенью. В некоторых вариантах осуществления антитело человека имеет изотип IgG1, IgG2, IgG3 или IgG4. В некоторых вариантах осуществления гетеродимерный белок содержит иммунологически инертную Fc-цепь.
Иммунная эффекторная клетка человека может являться любой из множества иммунных эффекторных клеток, известных в этой области. Например, иммунная эффекторная клетка может являться членом лимфоидного ростка клеток человека, включая, в качестве неограничивающих примеров, T-клетку (например, цитотоксическую T-клетку), B-клетку и естественный киллер (NK). Иммунная эффекторная клетка также, в качестве неограничивающих примеров, может являться членом миелоидного ростка человека, включая, в качестве неограничивающих примеров, моноцит, нейтрофильный гранулоцит и дендритную клетку. Такие иммунные эффекторные клетки могут иметь цитотоксический или апоптотический эффект в отношении клетки-мишени или другой желаемый эффект после активации посредством связывания эффекторного антигена.
Эффекторный антиген является антигеном (например, белком или полипептидом), экспрессирующимся на иммунной эффекторной клетке человека. Неограничивающие примеры эффекторных антигенов, которые могут связываться с гетеродимерным белком (например, гетеродимерным антителом или биспецифическим антителом), включают CD3 человека (или комплекс CD3 (кластер дифференцировки)), CD16, NKG2D, NKp46, CD2, CD28, CD25, CD64 и CD89.
Клетка-мишень может являться клеткой, являющейся нативной или чужеродной для людей. В случае нативной клетки-мишени клетка может трансформироваться в злокачественную клетку или патологически модифицироваться (например, нативная клетка-мишень, инфицированная вирусом, плазмодием или бактерией). В случае чужеродной клетки-мишени клетка является инвазирующим патогеном, таким как бактерия, плазмодий или вирус.
Антиген-мишень экспрессируется на клетке-мишени в состоянии заболевания (например, воспалительного заболевания, пролиферативного заболевания (например, злокачественного новообразования), иммунологического нарушения, неврологического заболевания, нейродегенеративного заболевания, аутоиммунного заболевания, инфекционного заболевания (например, вирусной инфекции или паразитарной инфекции), аллергической реакции, реакции "трансплантат против хозяина" или реакции "хозяин против трансплантата"). Антиген-мишень не является эффекторным антигеном. Неограничивающие примеры антигенов-мишеней включают GUCY2c, BCMA, EpCAM (молекулу адгезии эпителиальных клеток), CCR5 (хемокиновый рецептор типа 5), CD19, HER (рецептор эпидермального фактора роста человека)-2/neu, HER-3, HER-4, EGFR (рецептор эпидермального фактора роста), PSMA, CEA, MUC-1 (муцин), MUC2, MUC3, MUC4, MUC5AC, MUC5B, MUC7, CIhCG, антиген Льюиса Y, CD20, CD33, CD30, ганглиозид GD3, 9-O-ацетил-GD3, GM2, Globo H, фукозил-GM1, поли-SA, GD2, карбоангидразу IX (MN/CA IX), CD44v6, Shh (Sonic Hedgehog), Wue-1, антиген плазматических клеток, (мембраносвязанный) IgE, MCSP (меланомный хондроитинсульфатный протеогликан), CCR8, предшественник ФНО-альфа, STEAP, мезотелин, антиген A33, PSCA (антиген стволовых клеток предстательной железы), Ly-6; десмоглеин 4, неоэпитоп E-кадгерин, фетальный ацетилхолиновый рецептор, CD25, маркер CA19-9, маркер CA-125 и рецептор MIS (ингибирующей мюллеровой субстанции) типа II, sTn (сиалированный антиген Tn; TAG-72), FAP (антиген активации фибробластов), эндосиалин, EGFRvIII, LG, SAS и CD63.
В одном из вариантов осуществления настоящее изобретение относится к биспецифическому антителу, где: (a) CDR1 VL против GUCY2c, CDR2 VL против GUCY2c и CDR3 VL против GUCY2c из VL против GUCY2c содержат последовательность, приведенную в SEQ ID NO: 92, 100, 104, 106, 112, 119, 125, 129, 134, 136, 137, 138, 140, 143, 145, 147, 150, 152, 156, 158, 160, 162, 166, 170, 171, 172, 173, 174 или 175; (b) CDR1 VH против CD3, CDR2 VH против CD3 и CDR3 VH против CD3 из VH против CD3 содержат последовательность, приведенную в SEQ ID NO: 1, 9 или 273; (c) CDR1 VL против CD3, CDR2 VL против CD3 и CDR3 VL против CD3 из VL против CD3 содержат последовательность, приведенную в SEQ ID NO: 76, 84, 90 или 274, 275 или 276; и/или (d) CDR1 VH против GUCY2c, CDR2 VH против GUCY2c и CDR3 VH против GUCY2c из VH против GUCY2c содержат последовательность, приведенную в SEQ ID NO: 11, 19, 26, 33, 41, 48, 52, 57, 60, 62, 64, 65, 67, 69, 71 или 73.
В конкретном варианте осуществления настоящее изобретение относится к биспецифическому антителу, где: (a) область VH против GUCY2c содержит последовательность SEQ ID NO: 73; и (b) область VL против GUCY2c содержит последовательность SEQ ID NO: 147.
В некоторых вариантах осуществления настоящее изобретение относится к биспецифическому антителу, где: (a) CDR1 VL против GUCY2c содержит последовательность SEQ ID NO: 93, 101, 105, 107, 113, 120, 148, 153, 163 или 167; CDR2 VL против GUCY2c содержит последовательность SEQ ID NO: 78, 94, 102, 108, 114, 141, 144, 146, 149, 151, 157, 159, 161, 164 или 168; и CDR3 VL против GUCY2c содержит последовательность SEQ ID NO: 95, 109, 115, 121, 142, 154, 165 или 169; (b) CDR1 VH против CD3 содержит последовательность SEQ ID NO: 2, 268 или 277; CDR2 VH против CD3 содержит последовательность SEQ ID NO: 3, 10, 269 или 270; и CDR3 VH против CD3 содержит последовательность SEQ ID NO: 4; (c) CDR1 VL против CD3 содержит последовательность SEQ ID NO: 77, 85, 91, 278, 279 или 280; CDR2 VL против CD3 содержит последовательность SEQ ID NO: 78 или 281; и CDR3 VL против CD3 содержит последовательность SEQ ID NO: 79; и (d) CDR1 VH против GUCY2c содержит последовательность SEQ ID NO: 12, 20, 27, 34, 42, 74, 257, 258, 259, 260 или 261; CDR2 VH против GUCY2c содержит последовательность SEQ ID NO: 13, 21, 28, 35, 43, 53, 66, 68, 70, 72, 75, 262, 263, 264, 265, 266 или 267; и CDR3 VH против GUCY2c содержит последовательность SEQ ID NO: 14, 22, 29, 36 или 44.
В некоторых вариантах осуществления антитела, которые можно использовать в настоящем изобретении, являются моноклональными антителами, поликлональными антителами, фрагментами антител (например, Fab, Fab’, F(ab’)2, Fv, Fc и т.д.), химерными антителами, биспецифическими антителами, гетероконъюгированными антителами, одноцепочечными антителами (ScFv), их мутантами, слитыми белками, содержащими антительную часть (например, доменное антитело), гуманизированными антителами и любой другой модифицированной конфигурацией молекулы иммуноглобулина, содержащей участок распознавания антигена с необходимой специфичностью, включая варианты гликозилирования антител, варианты аминокислотной последовательности антител и ковалентно модифицированные антитела. Антитела могут являться антителами мыши, крысы, человека или иметь любое другое происхождение (включая химерные или гуманизированные антитела).
В некоторых вариантах осуществления антитело против GUCY2c или CD3, представленное в настоящем описании, является моноклональным антителом. Например, антитело против GUCY2c или CD3 является гуманизированным моноклональным антителом или химерным моноклональным антителом.
Настоящее изобретение относится к биспецифическому антителу, содержащему Fc-цепь или -домен или его часть. В некоторых вариантах осуществления Fc-цепь или ее части содержат один или более константных доменов Fc-цепи IgG1, IgG2, IgG3 или IgG4 (например, домен CH2 или CH3). В другом варианте осуществления настоящее изобретение относится к молекулам, содержащим Fc-цепь или ее часть, где Fc-цепь или ее часть содержит по меньшей мере одну аминокислотную модификацию (например, замену) относительно сравнимой Fc-цепи дикого типа или ее части. Варианты Fc-области хорошо известны в этой области, и их, главным образом, используют для изменения фенотипа антитела, содержащего вариант Fc-области, что оценивают посредством любого анализа связывающей активности или эффекторной функции, хорошо известных в этой области, например, ELISA, анализа SPR или ADCC. Такие варианты Fc-цепи или их части могут повышать период полувыведения из плазмы и стабильность, проявляемые биспецифическим антителом по изобретению, содержащим Fc-цепь или ее часть. В другом варианте осуществления настоящее изобретение относится к применению любого варианта Fc, известного в этой области.
В одном из вариантов осуществления осуществляют одну или более модификаций аминокислот Fc-цепи для снижения аффинности и авидности Fc-цепи и, таким образом, молекулы биспецифического антитела по изобретению к одному или более рецепторам FcγR. В конкретном варианте осуществления настоящее изобретение относится к биспецифическим антителам, содержащим вариант Fc-цепи или его часть, где вариант Fc-цепи содержит по меньшей мере одну аминокислотную модификацию относительно Fc-цепи дикого типа, где вариант Fc-области связывается только с одним FcγR, где FcγR является FcγRIIIA. В другом конкретном варианте осуществления настоящее изобретение относится к биспецифическим антителам, содержащим вариант Fc-цепи или его часть, где вариант Fc-цепи содержит по меньшей мере одну аминокислотную модификацию относительно Fc-цепи дикого типа, где вариант Fc-области связывается только с одним FcγR, где FcγR является FcγRIIA. В другом конкретном варианте осуществления настоящее изобретение относится к биспецифическим антителам, содержащим вариант Fc-цепи или его часть, где вариант Fc-цепи содержит по меньшей мере одну аминокислотную модификацию относительно Fc-цепи дикого типа, где вариант Fc-цепи связывается только с одним FcγR, где FcγR является FcγRIIB. В другом варианте осуществления настоящее изобретение относится к молекулам, содержащим вариант Fc-цепи, где вариант придает или опосредует сниженную активность ADCC (или другую эффекторную функцию) и/или повышенное связывание с FcγRIIB (CD32B) относительно молекулы, несодержащей Fc-цепь или содержащей Fc-цепь дикого типа, что измеряют способами, известными специалистам в этой области и представленным в настоящем описании.
Изобретение также относится к применению Fc-области, содержащей домены или области из двух или более изотипов IgG. Как известно в этой области, аминокислотная модификация Fc-области может значительно влиять на Fc-опосредованную эффекторную функцию и/или связывающую активность. Однако эти изменения функциональных характеристик можно дополнительно улучшать и/или дорабатывать при воплощении в контексте выбранных изотипов IgG. Аналогично, на нативные характеристики изотипа Fc можно воздействовать посредством одной или более аминокислотных модификаций. Множество изотипов IgG (т.е. IgG1, IgG2, IgG3 и IgG4) демонстрируют разные физические и функциональные свойства, включая время полужизни в сыворотке, фиксацию комплемента, аффинности связывания FcγR и активности эффекторных функций (например, ADCC, CDC) из-за различий аминокислотных последовательностей шарнирных областей и/или Fc-областей.
В одном из вариантов осуществления аминокислотную модификацию и Fc-области IgG независимо выбирают на основе их соответствующих отдельных активностей связывающей функции и/или эффекторной функции для конструирования биспецифического антитела с желаемыми характеристиками. В конкретном варианте осуществления аминокислотные модификации и шарнирные области/Fc-области IgG раздельно анализируют на активность связывающей и/или эффекторной функции, как представлено в настоящем описании или известно в этой области в отношении IgG1. В одном из вариантов осуществления аминокислотная модификация и шарнирная область/Fc-область IgG демонстрируют схожую функциональность, например, сниженную активность ADCC (или другую эффекторную функцию) и/или повышенное связывание с FcγRIIB в контексте биспецифического антитела или другой Fc-содержащей молекулы (например, иммуноглобулина). В другом варианте осуществления настоящее изобретение относится к вариантам Fc-области, содержащим комбинации аминокислотных модификаций, известных в этой области, и выбранных областей IgG, демонстрирующие новые свойства, неопределяемые, когда модификации и/или области анализируют независимо, как представлено в настоящем описании.
В некоторых вариантах осуществления каждый из первого способствующего гетеродимеризации домена и второго способствующего гетеродимеризации домена содержит Fc-область, содержащую домен CH2 и домен CH3, где аминокислотная последовательность каждого из домена CH2 и/или домена CH3 содержит по меньшей мере одну аминокислотную модификацию по сравнению с Fc-областью дикого типа, образующую выступ или впадину.
В некоторых вариантах осуществления каждый из первого способствующего гетеродимеризации домена и второго способствующего гетеродимеризации домена содержит домен CH2 и домен CH3, где аминокислотную последовательность каждого из доменов CH2 и/или каждого из доменов CH3 модифицируют для регуляции гетеродимеризации и/или стабилизации биспецифического антитела.
В некоторых таких вариантах осуществления биспецифические антитела по настоящему изобретению содержат первый способствующий гетеродимеризации домен на первой полипептидной цепи и второй способствующий гетеродимеризации домен на второй полипептидной цепи. В совокупности, первый и второй способствующие гетеродимеризации домены регулируют гетеродимеризацию, и/или стабилизируют биспецифическое антитело (например, посредством взаимодействия выступа и впадины на комплементарных способствующих гетеродимеризации доменов), и/или служат для стабилизации биспецифического антитела.
В некоторых вариантах осуществления первый способствующий гетеродимеризации домен и второй способствующий гетеродимеризации домен не являются оба выступами или не являются оба впадинами; и/или первый способствующий гетеродимеризации домен и второй способствующий гетеродимеризации домен образуют Fc-область иммуноглобулина IgG.
В некоторых вариантах осуществления первый способствующий гетеродимеризации домен может содержать Fc-цепь, имеющую домен CH2 и/или CH3, модифицированный так, чтобы он содержал выступ или впадину. В некоторых таких вариантах осуществления аминокислотная последовательность домена CH2 и/или домена CH3 содержит по меньшей мере одну аминокислотную модификацию, где: (a) домен CH3 первого способствующего гетеродимеризации домена образует выступ; и (b) домен CH3 второго способствующего гетеродимеризации домена образует впадину. В другом таком варианте осуществления домен CH3 первого способствующего гетеродимеризации домена содержит мутации Y349C и/или T366W для образования выступа; и домен CH3 второго способствующего гетеродимеризации домена содержит мутации S354C, T366S, L368A, и/или Y407V для образования впадины (нумерация в соответствии с индексом EU). В некоторых конкретных вариантах осуществления мутации приводят к сниженной эффекторной функции.
В некоторых вариантах осуществления первый способствующий гетеродимеризации домен может содержать домен CH2 и/или CH3, модифицированный так, чтобы он содержал выступ, содержащий последовательность SEQ ID NO: 188, если второй способствующий гетеродимеризации домен содержит домен CH2 и/или CH3, модифицированный так, чтобы он содержал впадину. В другом варианте осуществления первый способствующий гетеродимеризации домен может содержать впадину, если второй способствующий гетеродимеризации домен содержит домен CH2 и/или CH3, модифицированный так, чтобы он содержал выступ. В конкретном варианте осуществления описанного выше первый способствующий гетеродимеризации домен содержит последовательность SEQ ID NO: 188 для образования выступа; и где второй способствующий гетеродимеризации домен содержит последовательность SEQ ID NO: 189 для образования впадину. В таблице 7 приведены аминокислотные последовательности Fc-цепей с выступами и впадинами и первых полипептидных цепей и вторых полипептидных цепей биспецифических антител против GUCY2c с различными антителами против GUCY2c и CD3, представленными в настоящем описании (SEQ ID NO: 196, 197, 199, 200, 202, 203, 205, 206, 210, 211, 216, 217, 219, 220, 248, 249, 282, 283, 284, 285, 286 и 287).
Таблица 7
(SEQ ID NO: 196)
(SEQ ID NO: 197)
(SEQ ID NO: 199)
(SEQ ID NO: 200)
(SEQ ID NO: 202)
(SEQ ID NO: 203)
(SEQ ID NO: 205)
(SEQ ID NO: 206)
(SEQ ID NO: 210)
(SEQ ID NO: 211)
GUCY2C-1608_выступ
(SEQ ID NO: 216)
(SEQ ID NO: 217)
(SEQ ID NO: 220)
(SEQ ID NO: 248)
(SEQ ID NO: 249)
(SEQ ID NO: 282)
(SEQ ID NO: 283)
(SEQ ID NO: 284)
(SEQ ID NO: 285)
(SEQ ID NO: 286)
(SEQ ID NO: 287)
Одним из способов определения аффинности связывания антител против GUCY2c или CD3 является измерение аффинности связывания монофункциональных Fab-фрагментов антитела. Для получения монофункциональных Fab-фрагментов антитело (например, IgG) можно расщеплять папаином или экспрессировать рекомбинантно. Аффинность Fab-фрагмента антитела против GUCY2c можно определять посредством поверхностного плазмонного резонанса (с использованием системы поверхностного плазмонного резонанса (SPR) BIACORE™ 3000™, BIACORE™, INC, Piscataway NJ) с использованием сенсорных чипов с предварительно иммобилизованным стрептавидином (SA) или с использованием антитела против Fc мыши или антитела против Fc человека и подвижного буфера HBS-EP (0,01 M HEPES, pH 7,4, 0,15 M NaCl, 3 мМ ЭДТА, 0,005% об./об. поверхностно-активного вещества P20). Биотинилированную или Fc-слитую GUCY2c человека можно разводить в буфере HBS-EP до концентрации менее 0,5 мкг/мл и впрыскивать в отдельные каналы чипа с использованием переменного времени контакта для получения двух диапазонов плотности антигена, 50-200 единиц ответа (RU) для подробных исследований кинетики или 800-1000 RU для скрининговых анализов. Исследования регенерации показали, что с помощью 25 мМ NaOH в 25% об./об. этаноле эффективно удаляют связанный Fab при сохранении активности GUCY2c на чипе при более чем 200 инъекциях. Как правило, серийные разведения (охватывающие концентрации, составляющие 0,1-10-кратную KD) образцов очищенных Fab впрыскивают в течение 1 мин при 100 мкл/минуту и допускают время диссоциации до 2 часов. Концентрации белков Fab определяют посредством ELISA и/или электрофореза в ПААГ с SDS с использованием Fab в известной концентрации (что определяют посредством анализа аминокислот) в качестве стандарта. Кинетические данные о скорости ассоциации (kon) и скорости диссоциации (koff) получают одновременно посредством глобальной аппроксимации по модели связывания Ленгмюра 1:1 (Karlsson, R. Roos, H. Fagerstam, L. Petersson, B., Methods Enzymology 6. 99-110, 1994) с использованием программного обеспечения BIAevaluation. Значения равновесной константы диссоциации (KD) вычисляют как koff/kon. Этот способ подходит для использования в определении аффинности связывания антитела с любой GUCY2c, включая GUCY2c человека, GUCY2c другого млекопитающего (такого как GUCY2c мыши, GUCY2c крысы или GUCY2c примата), а также различными формами GUCY2c (например, гликозилированной GUCY2c). Аффинность связывания антитела, как правило, измеряют при 25°C, но ее также можно измерять при 37°C.
Антитела, представленные в настоящем описании, можно получать любым известным в этой области способом. Для получения линий гибридомных клеток путь и схема иммунизации животных-хозяев, как правило, соответствуют установленным и общепринятым способам стимуляции и продукции антител, как дополнительно представлено в настоящем описании. Общие способы получения антител человека и мыши известны в этой области и/или представлены в настоящем описании.
Предполагают, что на любое млекопитающее, включая людей, или продуцирующие антитела клетки из них можно воздействовать так, чтобы они служили основой для получения линий клеток млекопитающего, включая линии клеток человека и линии гибридомные клеток. Как правило, животного-хозяина инокулируют интраперитонеально, внутримышечно, перорально, подкожно, внутриподошвенно и/или интрадермально с использованием количества иммуногена, представленное в настоящем описании.
Гибридомы можно получать из лимфоцитов и иммортализованных миеломных клеток с использованием общего способа гибридизации соматических клеток из Kohler, B. and Milstein, C., Nature 256:495-497, 1975 или его модифицированной версии из Buck, D. W., et al., In Vitro, 18:377-381, 1982. В гибридизации можно использовать доступные миеломные линии, включая в качестве неограничивающих примеров X63-Ag8.653 и клетки из Salk Institute, Cell Distribution Center, San Diego, Calif., USA. Как правило, способ включает слияние миеломных клеток и лимфоидных клеток с использованием фузогена, такого как полиэтиленгликоль, или электрическими способами, хорошо известными специалистам в этой области. После слияния клетки отделяют от среды для слияния и выращивают в селективной среде для выращивания, такой как гипоксантин-аминоптерин-тимидиновая (HAT) среда, для устранения негибридизовавшихся родительских клеток. Любые из сред, представленных в настоящем описании, дополненных или недополненных сывороткой, можно использовать для культивирования гибридом, секретирующих моноклональные антитела. В качестве другой альтернативы способу слияния клеток, для получения моноклональных антител по настоящему изобретению можно использовать EBV-иммортализованные B-клетки. Гибридомы, при желании, выращивают и субклонируют и супернатанты анализируют на антииммуногенную активность общепринятыми способами иммунологических анализов (например, посредством радиоиммунологического анализа, ферментативного иммунологического анализа или флуоресцентного иммунологического анализа).
Гибридомы, которые можно использовать в качестве источника антител, включают все производные, клетки-потомки родительских гибридом, продуцирующих моноклональные антитела, специфические в отношении GUCY2c, CD3 или их частей.
Гибридомы, продуцирующие такие антитела, можно выращивать in vitro или in vivo известными способами. Моноклональные антитела можно выделять из сред для культивирования или физиологических жидкостей общепринятыми способами очистки иммуноглобулинов, при желании, такими как осаждение сульфатом аммония, электрофорез в геле, диализ, хроматография и ультрафильтрации. Нежелательную активность, при ее наличии, можно устранять, например, пропуская препарат через адсорбенты, полученные из иммуногена, прикрепленного к твердой фазе, и осуществляя элюцию или высвобождая желаемые антитела из иммуногена. Популяцию антител (например, моноклональных антител) можно получать посредством иммунизации животного-хозяина с использованием GUCY2c или CD3 человека или их фрагмента, содержащего целевую аминокислотную последовательность, конъюгированную с белком, являющимся иммуногенным для биологического вида, подлежащего иммунизации, например, гемоцианином морского блюдца, сывороточным альбумином, бычьим тиреоглобулином или ингибитором трипсина сои с использованием бифункционального или дериватизирующего средства, например, малеимидобензоилсульфосукцинимидного сложного эфира (конъюгация через остатки цистеина), N-гидроксисукцинимида (через остатки лизина), глутаральдегида, янтарного ангидрида, SOCl2, или R1N=C=NR, где R и R1 являются разными алкильными группами.
При желании, интересующее антитело (моноклональное или поликлональное) можно секвенировать, а затем полинуклеотидную последовательность можно клонировать в вектор для экспрессии или размножения. Последовательность, кодирующую интересующее антитело, можно поддерживать в векторе в клетке-хозяине, а затем клетку-хозяина можно выращивать и замораживать для дальнейшего использования. Получение рекомбинантных моноклональных антител в культуре клеток можно осуществлять посредством клонирования генов антител из B-клеток способами, известными в этой области. См., например, Tiller et al., J. Immunol. Methods, 3291122008; патент США № 7314622.
Альтернативно, полинуклеотидную последовательность можно использовать для генетического воздействия для гуманизации антитела или для улучшения аффинности или других характеристик антител. Например, константную область можно конструировать так, чтобы она как можно больше напоминала константные области человека во избежание иммунного ответа, если антитело используют в клинических испытаниях и лечении людей. Желательным может являться генетическое воздействие на последовательность антитела для достижения более высокой аффинности к GUCY2c или CD3 и большей эффективности в ингибировании GUCY2c.
Существуют четыре общие стадии гуманизации моноклонального антитела. Они представляют собой: (1) определение нуклеотидной и прогнозируемой аминокислотной последовательности начальных вариабельных областей легких и тяжелых цепей антитела, (2) дизайн гуманизированного антитела, т.е. принятие решения о том, какую каркасную область антитела использовать во время гуманизации, (3) конкретные способы гуманизации и (4) трансфекцию и экспрессию гуманизированного антитела. См., например, патенты США №№ 4816567, 5807715, 5866692, 6331415, 5530101, 5693761, 5693762, 5585089 и 6180370.
Описан ряд молекул гуманизированных антител, содержащих антигенсвязывающий участок, полученный из не принадлежащего человеку иммуноглобулина, включая химерные антитела, имеющие V-области грызуна или модифицированные V-области грызуна и соответствующие CDR, слитые с константными областями человека. См., например, Winter et al. Nature 349:293-299, 1991, Lobuglio et al. Proc. Nat. Acad. Sci. USA 86:4220-4224, 1989, Shaw et al. J Immunol. 138:4534-4538, 1987, и Brown et al. Cancer Res. 47:3577-3583, 1987. В других источниках описывают CDR грызуна, пересаженные на поддерживающую каркасную область (FR) человека перед слиянием с подходящей константной областью антитела человека. См., например, Riechmann et al. Nature 332:323-327, 1988, Verhoeyen et al. Science 239:1534-1536, 1988, и Jones et al. Nature 321:522-525, 1986. В другом источнике описывают CDR грызуна, поддерживаемые рекомбинантно сконструированными каркасными областями грызуна. См., например, европейскую патентную публикацию № EP0519596. Эти "гуманизированные" молекулы конструируют для минимизации нежелательного иммунного ответа на молекулы антител грызуна против человека, что ограничивает длительность и эффективность терапевтического использования этих веществ на реципиентах-людях. Например, константную область антитела можно конструировать таким образом, что она является иммунологически инертной (например, не запускает опосредованный комплементом лизис). См., например, патентную заявку PCT № PCT/GB99/01441; патентную заявку Великобритании № 9809951.8. Другие способы гуманизации антител, которые также можно использовать, описаны в Daugherty et al., Nucl. Acids Res. 19:2471-2476, 1991, в патентах США №№ 6180377, 6054297, 5997867, 5866692, 6210671 и 6350861 и публикации PCT № WO01/27160.
Общие принципы, касающиеся гуманизированных антител, описанных выше, также можно использовать для кастомизации антител для использования, например, на собаках, кошках, приматах, лошадях и крупном рогатом скоте. Кроме того, можно комбинировать один или более аспектов гуманизации антитела, представленного в настоящем описании, например, пересадку CDR, мутации каркаса и мутации CDR.
В одном из вариантов полностью человеческие антитела можно получать с использованием коммерчески доступных мышей, сконструированных для экспрессии конкретных белков иммуноглобулина человека. Трансгенные животные, полученные для достижения более желательного (например, полностью человеческие антитела) или более устойчивого иммунного ответа, также можно использовать для получения гуманизированных антител или антител человека. Примерами такой технологии являются Xenomouse™ от Abgenix, Inc. (Fremont, CA) и HuMAb-Mouse® и TC Mouse™ от Medarex, Inc. (Princeton, NJ).
Альтернативно, антитела можно получать рекомбинантно и экспрессировать любым известным в этой области способом. В качестве другой альтернативы, антитела можно получать рекомбинантно с помощью технологии фагового дисплея. См., например, патенты США №№ 5565332, 5580717, 5733743 и 6265150 и Winter et al., Annu. Rev. Immunol. 12:433-455, 1994. Альтернативно, можно использовать технологию фагового дисплея (McCafferty et al., Nature 348:552-553, 1990) для получения антител человека и фрагментов антител in vitro из репертуара генов вариабельных областей (V) иммуноглобулина из иммунизированных доноров. В этом способе гены V-домена антитела клонируют в рамке считывания в ген основного или минорного белка оболочки нитевидного бактериофага, такого как M13 или fd, и экспонируют как функциональные фрагменты антител на поверхности фаговой частицы. Т.к. частица нитевидного фага содержит одноцепочечную копию ДНК генома фага, селекция на основе функциональных свойств антитела также приводит к селекции гена, кодирующего антитело, проявляющего эти свойства. Таким образом, фаг имитирует некоторые из свойств B-клетки. Фаговый дисплей можно осуществлять в различных форматах; обзор см., например, Johnson, Kevin S. and Chiswell, David J., Current Opinion in Structural Biology 3:564-571, 1993. Для фагового дисплея можно использовать несколько источников сегментов V-генов. Clackson et al., Nature 352:624-628, 1991 выделяли разнообразный набор антител против оксазолона из небольшой рандомной комбинаторной библиотеки V-генов, полученных из селезенок иммунизированных мышей. Можно конструировать репертуар V-генов неиммунизированных людей-доноров и можно выделять антитела против разнообразного набора антигенов (включая аутоантигены), по существу, способами, описанными в Mark et al., J. Mol. Biol. 222:581-597, 1991, или Griffith et al., EMBO J. 12:725-734, 1993. При природном иммунном ответе гены антител накапливают мутации в большом количестве (соматическая супермутация). Некоторые вносимые изменения будут придавать более высокую аффинность, и B-клетки, экспонирующие высокоаффинный поверхностный иммуноглобулин, преференциально делятся и дифференцируются во время последующей стимуляции антигеном. Этот природный процесс можно имитировать способом, известным как "перестановка цепей" (Marks et al., Bio/Technol. 10:779-783, 1992). В этом способе аффинность "первичных" антител человека, полученных посредством фагового дисплея, можно улучшать, последовательно заменяя гены V-области тяжелой и легкой цепи репертуаром природных вариантов (репертуаром) генов V-доменов, полученным из неиммунизированных доноров. Этот способ делает возможным получение антител и фрагментов антител с аффинностями в диапазоне пМ-нМ. Стратегия получения очень большого репертуара фаговых антител (также известная как "библиотеки всеобщей матери") описана в Waterhouse et al., Nucl. Acids Res. 21:2265-2266, 1993. Перестановку генов также можно использовать для получения антител человека из антител грызунов, где антитело человека обладает аффинностями и специфичностями, схожими с исходным антителом грызуна. В этом способе, также обозначаемом как "импринтинг эпитопов", ген V-домена тяжелой или легкой цепи антител грызуна, полученный способом фагового дисплея, заменяют репертуаром генов V-доменов человека, получая химеры грызун-человек. Селекция с использованием антигена приводит к выделению вариабельных областей человека, способных восстанавливать функциональный антигенсвязывающий участок, т.е. эпитоп регулирует (импринтирует) выбор партнера. Когда способ повторяют для замены оставшегося V-домена грызуна, получают антитело человека (см. публикацию PCT № WO93/06213). В отличие от общепринятой гуманизации антител грызунов посредством пересадки CDR, этот способ обеспечивает получение полностью человеческих антител, не имеющих остатки каркаса или CDR грызуна.
Антитела можно получать рекомбинантно, сначала выделяя антитела и продуцирующие антитела клетки из животных-хозяев, получая последовательность гена и используя последовательность гена для рекомбинантной экспрессии антитела в клетках-хозяевах (например, клетках CHO). Другим способом, который можно использовать, является экспрессия последовательности антитела в растениях (например, табаке) или трансгенном молоке. Описаны способы рекомбинантной экспрессии антител в растениях или молоке. См., например, Peeters, et al. Vaccine 19:2756, 2001; Lonberg, N. and D. Huszar Int. Rev. Immunol 13:65, 1995; и Pollock, et al., J Immunol Methods 231:147, 1999. В этой области известны способы получения производных антител, например, гуманизированных, одноцепочечных и т.д.
Для выделения антител, специфических в отношении GUCY2c, CD3 или интересующих антигенов, также можно использовать иммунологические анализы и способы сортировки на основе проточной цитометрии, такие как активируемая флуоресценцией сортировка клеток (FACS).
Антитела, представленные в настоящем описании, можно связывать с множеством разных твердых подложек или носителей. Такие подложки могут являться активными и/или инертными. Хорошо известные подложки включают полипропилен, полистирол, полиэтилен, декстран, нейлон, амилазы, стекло, природные и модифицированные целлюлозы, полиакриламиды, агарозы и магнетит. В целях по изобретению подложка может являться растворимой или нерастворимой. Специалистам в этой области будут известны другие подходящие подложки для связывания антител, или они смогут определять их с использованием рутинного экспериментирования. В некоторых вариантах осуществления подложка содержит фрагмент, нацеленный на миокард.
ДНК, кодирующую моноклональные антитела, легко выделяют и секвенируют общепринятыми способами (например, с использованием олигонуклеотидных зондов, которые могут специфически связываться с генами, кодирующими тяжелые и легкие цепи моноклональных антител). Гибридомные клетки служат в качестве предпочтительного источника такой ДНК. После выделения ДНК можно помещать в экспрессирующие векторы (такие как экспрессирующие векторы, описанные в публикации PCT № WO87/04462), с помощью которых затем трансфицируют клетки-хозяева, такие как клетки E. coli, клетки обезьян COS, клетки CHO или миеломные клетки, которые в ином случае не продуцируют иммуноглобулиновый белок, для достижения синтеза моноклональных антител в рекомбинантных клетках-хозяевах. См., например, публикацию PCT № WO87/04462. ДНК также можно модифицировать, например, посредством замены кодирующей последовательностью константных областей тяжелой и легкой цепи человека гомологичных последовательностей мыши, Morrison et al., Proc. Nat. Acad. Sci. 81:6851, 1984, или посредством ковалентного соединения с кодирующей последовательностью иммуноглобулина всей или части кодирующей последовательности неиммуноглобулинового полипептида. Таким образом, получают "химерные" или "гибридные" антитела, имеющие специфичность связывания моноклонального антитела по настоящему изобретению.
Антитела против GUCY2c, CD3 или других антигенов, представленные в настоящем описании, можно идентифицировать или охарактеризовывать известными в этой области способами, при этом определяют и/или измеряют снижение уровней экспрессии GUCY2c, CD3 или другого антигена. В некоторых вариантах осуществления антитело против GUCY2c идентифицируют посредством инкубации средства-кандидата с GUCY2c и мониторинга связывания и/или сопутствующего снижения уровней экспрессии GUCY2c. Анализ связывания можно осуществлять с использованием очищенных полипептидов GUCY2c или клеток, от природы экспрессирующих или трансфицированных для экспрессии полипептидов GUCY2c. В одном из вариантов осуществления анализ связывания является анализом конкурентного связывания, где оценивают способность антитела-кандидата конкурировать с известным антителом против GUCY2c за связывание с GUCY2c. Анализ можно осуществлять в различных форматах, включая формат ELISA.
После исходной идентификации активность антитела-кандидата против GUCY2c, CD3 или другого антигена можно дополнительно подтверждать и уточнять посредством биологических анализов, известных для тестирования целевых видов биологической активности. Альтернативно, биологические анализы можно использовать для прямого скрининга кандидатов. Некоторые из способов идентификации и характеризации антител подробно описаны в примерах.
Антитела против GUCY2c, CD3 или другого антигена можно охарактеризовывать способами, хорошо известными в этой области. Например, одним из способов является идентификация эпитопа, с которым оно связывается, или эпитопное картирование. Существует множество известных в этой области способов картирования и характеризации локализации эпитопов на белках, включая разрешение кристаллической структуры комплекса антитело-антиген, конкурентные анализы, анализы экспрессии фрагментов генов и синтетические анализы на основе пептидов, как описано, например, в главе 11 Harlow and Lane, Using Antibodies, a Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1999. В дополнительном примере эпитопное картирование можно использовать для определения последовательности, с которой связывается антитело. Эпитопное картирование является коммерчески доступным в различных источниках, например, Pepscan Systems (Edelhertweg 15, 8219 PH Lelystad, The Netherlands). Эпитоп может являться линейным эпитопом, т.е. содержаться в одном фрагменте аминокислот, или конформационным эпитопом, образованным при трехмерном взаимодействии аминокислот, которые могут не содержаться в одном фрагменте. Пептиды различной длины (например, по меньшей мере 4-6 аминокислот) можно выделять или синтезировать (например, рекомбинантно) и использовать для анализов связывания с антителом против GUCY2c, CD3 или другим антигеном. В другом примере эпитоп, с которым связывается антитело против GUCY2c, CD3 или другого антигена, можно определять посредством системного скрининга с использованием перекрывающихся пептидов, полученных из последовательности GUCY2c, CD3 или другого антигена, и определения связывания с антителом против GUCY2c, CD3 или другого антигена. В соответствии с анализами экспрессии фрагментов генов, открытую рамку считывания, кодирующую GUCY2c, CD3 или другой антиген, фрагментируют случайным образом или с помощью специфических генетических конструкций, и определяют реактивность экспрессируемых фрагментов GUCY2c, CD3 или другого антигена и антитела, подлежащего тестированию. Фрагменты генов можно получать, например, посредством ПЦР, а затем транскрибировать и транслировать в белок in vitro в присутствии радиоактивных аминокислот. Затем определяют связывание антитела с радиоактивно мечеными фрагментами GUCY2c, CD3 или другого антигена с помощью иммунопреципитации и электрофореза в геле. Некоторые эпитопы также можно идентифицировать с использованием больших библиотек случайных пептидных последовательностей, экспонируемых на поверхности фаговых частиц (фаговые библиотеки). Альтернативно, определенную библиотеку перекрывающихся пептидных фрагментов можно тестировать на связывание с тестовым антителом в простых анализах связывания. В дополнительном примере для идентификации остатков, требующихся, достаточных и/или необходимых для связывания эпитопа, можно осуществлять мутагенез антигенсвязывающего домена, эксперименты по перестановке доменов и мутагенез с аланиновым сканированием. Например, эксперименты по перестановке доменов можно осуществлять с использованием мутантной GUCY2c, CD3 или другого антигена, где различные фрагменты GUCY2c, CD3 или другого антигенного белка заменяют (перестанавливают) последовательностями из GUCY2c другого биологического вида (например, мыши) или близкородственного, но антигенно отличающегося отдельного белка (например, Trop-1). Оценивая связывание антитела с мутантной GUCY2c, CD3 или другим антигеном, можно оценивать важность конкретного фрагмента GUCY2c, CD3 или другого антигена для связывания антитела.
Другим способом, который можно использовать для характеризации антитела против GUCY2c, CD3 или другого антигена, является использование конкурентных анализов с другими антителами, которые, как известно, связываются с тем же антигеном, т.е. различными фрагментами на GUCY2c, CD3 или другом антигене, для определения того, связывается ли антитело против GUCY2c, CD3 или другого антигена с тем же эпитопом, что и другие антитела, соответственно. Конкурентные анализы хорошо известны специалистам в этой области.
Для регуляции экспрессии антитела против GUCY2c, CD3 или другого антигена можно использовать экспрессирующий вектор. Специалисту в этой области известно введение экспрессирующих векторов для достижения экспрессии экзогенного белка in vivo. См., например, патенты США №№ 6436908, 6413942 и 6376471. Введение экспрессирующих векторов включает локальное или системное введение, включая инъекцию, пероральное введение, биобаллистическую пушку или катетерное введение и местное введение. В другом варианте осуществления экспрессирующий вектор вводят напрямую в симпатический ствол или ганглий или коронарную артерию, предсердие, желудочек или перикард.
Также можно использовать направленную доставку терапевтических композиций, содержащих экспрессирующий вектор, или субгеномных полинуклеотидов. Рецептор-опосредованные способы доставки ДНК описаны, например, в Findeis et al., Trends Biotechnol., 11:202, 1993; Chiou et al., Gene Therapeutics: Methods And Applications Of Direct Gene Transfer, J.A. Wolff, ed., 1994; Wu et al., J. Biol. Chem., 263:621, 1988; Wu et al., J. Biol. Chem., 269:542, 1994; Zenke et al., Proc. Natl. Acad. Sci. USA, 87:3655, 1990; и Wu et al., J. Biol. Chem., 266:338, 1991. Терапевтические композиции, содержащие полинуклеотид, вводят в диапазоне от приблизительно 100 нг до приблизительно 200 мг ДНК для локального введения в способе генной терапии. В способе генной терапии также можно использовать диапазоны концентраций от приблизительно 500 нг до приблизительно 50 мг, от приблизительно 1 мкг до приблизительно 2 мг, от приблизительно 5 мкг до приблизительно 500 мкг и от приблизительно 20 мкг до приблизительно 100 мкг ДНК. Терапевтические полинуклеотиды и полипептиды можно доставлять с использованием носителей для доставки генов. Носитель для доставки генов может иметь вирусное или невирусное происхождение (см., в целом, Jolly, Cancer Gene Therapy,1:51, 1994; Kimura, Human Gene Therapy, 5:845, 1994; Connelly, Human Gene Therapy, 1:185, 1995; и Kaplitt, Nature Genetics, 6:148, 1994). Экспрессию таких кодирующих последовательностей можно индуцировать с использованием эндогенных промоторов млекопитающих или гетерологичных промоторов. Экспрессия кодирующей последовательности может являться конститутивной или регулируемой.
Вирусные векторы для доставки желаемого полинуклеотида и экспрессии в желаемой клетке хорошо известны в этой области. Неограничивающие примеры носителей на основе вирусов включают рекомбинантные ретровирусы (см., например, публикации PCT №№ WO90/07936; WO94/03622; WO93/25698; WO93/25234; WO93/11230; WO93/10218; WO91/02805; патенты США №№ 5219740 и 4777127; патент Великобритании № 2200651; и патент EP № EP0 345 242), векторы на основе альфавирусов (например, векторы вируса Синдбис, вируса леса Семлики (ATCC VR-67; ATCC VR-1247), вируса лихорадки реки Росс (ATCC VR-373; ATCC VR-1246) и вируса венесуэльского энцефалита лошадей (ATCC VR-923; ATCC VR-1250; ATCC VR 1249; ATCC VR-532)) и векторы на основе аденоассоциированного вируса (AAV) (см., например, публикации PCT №№ WO94/12649, WO93/03769; WO93/19191; WO94/28938; WO95/11984 и WO95/00655). Также можно использовать введение ДНК, связанной с убитым аденовирусом, как описано в Curiel, Hum. Gene Ther., 3:147, 1992.
Также можно использовать невирусные носители для доставки и способы, включая, в качестве неограничивающих примеров, конденсированную с поликатионами ДНК, связанную или несвязанную с убитым аденовирусом в отдельности (см., например, Curiel, Hum. Gene Ther., 3:147, 1992); связанную с лигандом ДНК (см., например, Wu, J. Biol. Chem., 264:16985, 1989); эукариотическая клетки-носители для доставки (см., например, патент США № 5814482; публикации PCT №№ WO95/07994; WO96/17072; WO95/30763 и WO97/42338) и нейтрализацию заряда ядра или слияние с клеточными мембранами. Также можно использовать депротеинизированную ДНК. Примеры способов встраивания депротеинизированной ДНК описаны в публикации PCT № WO90/11092 и патенте США № 5580859. Липосомы, которые могут действовать в качестве носителей для доставки генов, описаны в патенте США № 5422120; публикациях PCT №№ WO95/13796; WO94/23697; WO91/14445; и европейской публикации № EP 0524968. Дополнительные подходы описаны в Philip, Mol. Cell Biol., 14:2411, 1994 и Woffendin, Proc. Natl. Acad. Sci., 91:1581, 1994.
В некоторых вариантах осуществления настоящее изобретение относится к композициям, включая фармацевтические композиции, содержащие антитела по изобретению, представленные в настоящем описании или полученные способами, представленными в настоящем описании, и имеющие характеристики, представленные в настоящем описании. В рамках изобретения фармацевтические композиции могут содержать одно или более антител, связывающихся с GUCY2c, одно или более биспецифических антител, связывающихся с CD3 и опухолевым антигеном GUCY2c, и/или один или более полинуклеотидов, содержащих последовательности, кодирующие одно или более из этих антител. Эти композиции могут дополнительно содержать подходящие эксципиенты, такие как фармацевтически приемлемые эксципиенты, включая буферы, хорошо известные в этой области.
Изобретение также относится к способам получения любых из таких антител. Антитела по настоящему изобретению можно получать известными в этой области способами. Полипептиды можно получать посредством протеолитической или другой деградации антител, рекомбинантными способами (т.е. в виде единых или слитых полипептидов), как описано выше, или посредством химического синтеза. Полипептиды антител, в особенности, более короткие полипептиды до приблизительно 50 аминокислот, удобно получать посредством химического синтеза. Способы химического синтеза известны в этой области и коммерчески доступны. Например, антитело можно получать с помощью автоматизированного полипептидного синтезатора с использованием твердофазного способа. Также см., патенты США №№ 5807715, 4816567 и 6331415.
Гетероконъюгированные антитела, содержащие два ковалентно соединенных антитела, также входят в объем изобретения. Такие антитела используют для таргетинга клеток иммунной системы к нежелательным клеткам (патент США № 4676980) и для лечения инфекции ВИЧ (публикации PCT №№ WO91/00360 и WO92/200373; и европейская патентная публикация № EP03089). Гетероконъюгированные антитела можно получать любыми удобными способами перекрестной сшивки. Подходящие сшивающие средства и способы хорошо известны в этой области и описаны в патенте США № 4676980.
Химерные или гибридные антитела также можно получать in vitro известными способами синтетической химии белков, включая способы, включающие сшивающие средства. Например, иммунотоксины можно конструировать с использованием реакции дисульфидного обмена или посредством образования тиоэфирной связи. Примеры подходящих реагентов для этой цели включают иминотиолат и метил-4-меркаптобутиримидат.
В рекомбинантных гуманизированных антителах Fcγ-часть можно модифицировать во избежание взаимодействия с Fcγ-рецептором, и комплементом и иммунной системой. Способы получения таких антител описаны в публикации PCT № WO99/58572. Например, константную область можно конструировать так, чтобы она больше напоминала константные области человека во избежание иммунного ответа, если антитело используют в клинических испытаниях и лечении людей. См., например, патенты США №№ 5997867 и 5866692.
Антитела против GUCY2c и биспецифические антитела против CD3-GUCY2c, представленные в настоящем описании, могут содержать одну или более замен, инсерций и/или делеций аминокислот в каркасных областях и/или областях CDR вариабельных доменов тяжелых и легких цепей по сравнению с соответствующими последовательностями зародышевой линии, из которых получают антитела. Такие мутации можно устанавливать посредством сравнения аминокислотных последовательностей, представленных в настоящем описании, с последовательностями зародышевой линии, доступными, например, в общедоступных базах данных о последовательностях антител. Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам, полученным из любых аминокислотных последовательностей, представленных в настоящем описании, где одна или более аминокислот в одной или более каркасных областей и/или областей CDR подвергнуты мутации в соответствующие остатки последовательности зародышевой линии, из которой получали антитело, или в соответствующие остатки другой последовательности зародышевой линии человека, или в консервативную аминокислотную замену соответствующих остатков зародышевой линии (такие изменения в последовательности в совокупности в настоящем описании обозначают как "мутации зародышевой линии"). Специалист в этой области, начиная с последовательностей вариабельной области тяжелой и легкой цепи, представленных в настоящем описании, легко может получать многочисленные антитела и антигенсвязывающие фрагменты, содержащие одну или более отдельных мутаций зародышевой линии или их комбинаций. В некоторых вариантах осуществления все из остатков каркаса и/или CDR в доменах VH и/или VL подвергнуты обратной мутации в остатки, обнаруживаемые в исходной последовательности зародышевой линии, из которой получали антитело. В других вариантах осуществления только некоторые остатки подвергают обратной мутации в исходную последовательность зародышевой линии, например, только мутантные остатки, обнаруживаемые в первых 8 аминокислотах FR1 или в последних 8 аминокислотах FR4, или только мутантные остатки, обнаруживаемые в CDR1, CDR2 или CDR3. В других вариантах осуществления один или более остатков из каркаса и/или CDR подвергают мутации в соответствующие остатки другой последовательности зародышевой линии (т.е. последовательности зародышевой линии, отличающейся от последовательности зародышевой линии, из которой антитело получали исходно).
Кроме того, антитела по настоящему изобретению могут содержать любую комбинацию двух или более мутаций зародышевой линии в каркасных областях и/или областях CDR, например, где некоторые отдельные остатки подвергнуты мутации в соответствующий остаток конкретной последовательности зародышевой линии, в то время как некоторые другие остатки, отличающиеся от исходной последовательности зародышевой линии, сохраняют или подвергают мутации в соответствующий остаток другой последовательности зародышевой линии. После получения антитела и антигенсвязывающие фрагменты, содержащие одну или более мутаций зародышевой линии, легко можно тестировать на одно или более желаемых свойств, таких как улучшенная специфичность связывания, повышенная аффинность связывания, улучшенные или повышенные антагонистические или агонистические биологические свойства (соответственно), сниженная иммуногенность и т.д. Антитела и антигенсвязывающие фрагменты, полученные этим общим способом, включены в настоящем изобретении.
Настоящее изобретение также относится к антителам против GUCY2c и биспецифическим антителам против CD3-GUCY2c, содержащим варианты любых аминокислотных последовательностей HC VR, LC VR и/или CDR, представленных в настоящем описании, имеющих одну или более консервативных замен. Например, настоящее изобретение относится к антителам против GUCY2c и биспецифическим антителам против CD3-GUCY2c, имеющим аминокислотные последовательности HC VR, LC VR и/или CDR с, например, 10 или менее, 8 или менее, 6 или менее, 4 или менее и т.д. консервативных аминокислотных замен относительно любой из аминокислотных последовательностей HC VR, LC VR и/или CDR, как представлено в настоящем описании.
Таким образом, настоящее изобретение относится к модификациям вариантов антител и полипептидов по изобретению, как представлено в настоящем описании, включая функционально эквивалентные антитела, не влияющим значимо на их свойства, и вариантам, имеющим повышенную или сниженную активность и/или аффинность. Например, аминокислотную последовательность можно подвергать мутации для получения антитела с желаемой аффинностью связывания с GUCY2c и/или CD3. Модификация полипептидов является общепринятой практикой в этой области и не требует подробного обсуждения в настоящем описании. Примеры модифицированных полипептидов включают полипептиды с консервативными заменами аминокислотных остатков, одной или более делециями или добавлениями аминокислот, которые не изменяют значительно, неблагоприятно функциональную активность или которые приводят к созреванию (повышают) аффинность полипептида к его лиганду, или использование химических аналогов.
Инсерции аминокислотной последовательности включают, амино- и/или карбокси-концевые слияния с диапазоном длин от одного остатка до полипептидов, содержащих сотню или более остатков, а также инсерции отдельных или многочисленных аминокислотных остатков в последовательности. Примеры концевых инсерций включают антитело с N-концевым метионильным остатком или антитело, слитое с эпитопной меткой. Другие инсерционные варианты молекулы антитела включают слияние фермента или полипептида, повышающего время полужизни антитела в кровотоке, с N- или C-концом антитела.
Варианты с заменой включают удаление по меньшей мере одного аминокислотного остатка в молекуле антитела и встраивание другого остатка вместо него. Участки, представляющие наибольший интерес для замещающего мутагенеза, включают гипервариабельные области, но также предусмотрены изменения FR. Консервативные замены приведены в таблице 8 под заголовком "консервативные замены". Если такие замены приводят к изменению биологической активности, то можно встраивать более существенные изменения, обозначенные в таблице 8 как "примеры замен" или описанные ниже по отношению к классам аминокислот, и подвергать скринингу полученные продукты.
Таблица 8
Значительные модификации биологических свойств антитела осуществляют посредством выбора замен, значительно отличающихся по своему эффекты в отношении (a) структуры остова полипептида в области замены, например, в виде конформации листа или спирали, (b) заряда или гидрофобности молекулы в целевом участке или (c) объема боковой цепи. Природные аминокислотные остатки разделяют на группы с учетом общих свойств боковой цепи:
(1) Неполярные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) Полярные незаряженные: Cys, Ser, Thr, Asn, Gln;
(3) Кислые (отрицательно заряженные): Asp, Glu;
(4) Основные (положительно заряженные): Lys, Arg;
(5) Остатки, влияющие на ориентацию цепи: Gly, Pro; и
(6) Ароматические: Trp, Tyr, Phe, His.
Неконсервативные замены осуществляют посредством замены члена одного из этих классов членом другого класса.
Любой остаток цистеина, не участвующий в поддержании правильной конформации антитела, также можно заменять, как правило, серином, для улучшения окислительной стабильности молекулы и предотвращения аномальной перекрестной сшивки. И наоборот, цистеиновые связи можно добавлять в антитело для улучшения его стабильности, в частности, если антитело является фрагментом антитела, таким как Fv-фрагмент.
Модификации аминокислот могут находиться в диапазоне от изменения или модификации одной или более аминокислот для завершения реконструкции области, такой как вариабельная область. Изменения вариабельной области могут приводить к изменению аффинности и/или специфичности связывания. В некоторых вариантах осуществления в домене CDR осуществляют от не более одной до пяти консервативных аминокислотных замен. В других вариантах осуществления в домене CDR осуществляют от не более одной до трех консервативных аминокислотных замен. В других вариантах осуществления домен CDR является CDR3 VH и/или CDR3 VL.
Модификации также включают гликозилированные и негликозилированные полипептиды, а также полипептиды с другими посттрансляционными модификациями, такими как, например, гликозилирование разными сахарами, ацетилирование и фосфорилирование. Антитела являются гликозилированными в консервативных положениях в их константных областях (Jefferis and Lund, Chem. Immunol. 65:111-128, 1997; Wright and Morrison, TibTECH 15:26-32, 1997). Олигосахаридные боковые цепи иммуноглобулинов влияют на функцию белка (Boyd et al., Mol. Immunol. 32:1311-1318, 1996; Wittwe and Howard, Biochem. 29:4175-4180, 1990) и внутримолекулярное взаимодействие между частями гликопротеина, которые могут влиять на конформацию и представлять собой трехмерную поверхность гликопротеина (Jefferis and Lund, выше; Wyss и Wagner, Current Opin. Biotech. 7:409-416, 1996). Олигосахариды также могут служить для таргетинга указанного гликопротеина к некоторым молекулам с учетом специфических распознающих структур. Также сообщают, что гликозилирование антител влияет на антителозависимую клеточную цитотоксичность (ADCC). В частности, сообщают, что клетки CHO с тетрациклин-регулируемой экспрессией β(1,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII), гликозилтрансферазы, катализирующей образование GlcNAc в точках ветвления, имеют улучшенную активность ADCC (Umana et al., Mature Biotech. 17:176-180, 1999).
Гликозилирование антител, как правило, является N-связанным или O-связанным. Термин "N-связанный" относится к присоединению молекулы углевода к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-X-серин, аспарагин-X-треонин и аспарагин-X-цистеин, где X является любой аминокислотой, за исключением пролина, являются распознаваемыми последовательностями для ферментативного присоединения молекулы углевода к боковой цепи аспарагина. Таким образом, наличие этих трипептидных последовательностей в полипептиде приводит к образованию потенциального участка гликозилирования. Термин "O-связанное" гликозилирование относится к присоединению одного из сахаров N-ацетилгалактозамина, галактозы или ксилозы к гидроксиаминокислоте, чаще всего, серину или треонину, хотя также можно использовать 5-гидроксипролин или 5-гидроксилизин.
Добавление участков гликозилирования в антитело удобно осуществлять посредством изменения аминокислотной последовательности таким образом, что она содержит одну или более из описанных выше трипептидных последовательностей (в случае N-связанных участков гликозилирования). Изменение также можно осуществлять посредством добавления или замены одним или более остатками серина или треонина в последовательности исходного антитела (в случае O-связанных участков гликозилирования).
Профиль гликозилирования антител также можно изменять без изменения основополагающей нуклеотидной последовательности. Гликозилирование, главным образом, зависит от клетки-хозяина, используемой для экспрессии антитела. Т.к. тип клеток, используемый для экспрессии рекомбинантных гликопротеинов, например, антител, в качестве потенциальных терапевтических средств, редко представляет собой нативную клетку, можно ожидать вариаций профиля гликозилирования антител (см., например, Hse et al., J. Biol. Chem. 272:9062-9070, 1997).
В дополнение к выбору клеток-хозяев, факторы, влияющие на гликозилирование во время рекомбинантной продукции антител, включают режим выращивания, состав средств, плотность культуры, оксигенацию, pH, схемы очистки т.п. Предложены различные способы изменения профиля гликозилирования, достигаемого в конкретном организме-хозяине, включая встраивание или гиперэкспрессию некоторых ферментов, вовлеченных в продукцию олигосахаридов (патенты США №№ 5047335, 5510261 и 5278299). Гликозилирование или некоторые типы гликозилирования можно ферментативно удалять из гликопротеина, например, с использованием эндогликозидаз H (Endo H), N-гликозидазы F, эндогликозидазы F1, эндогликозидазы F2, эндогликозидазы F3. Кроме того, рекомбинантную клетку-хозяина можно генетически конструировать так, чтобы она была дефектной по процессингу некоторых типов полисахаридов. Эти и схожие способы хорошо известны в этой области.
Другие способы модификации включают использование способов связывания, известных в этой области, включая в качестве неограничивающих примеров, ферментативные способы, окислительную замену и хелатирование. Модификации можно использовать, например, для присоединения меток для иммунологических анализов. Модифицированные полипептиды получают способами, общепринятыми в этой области, и их можно подвергать скринингу с использованием стандартных способов, известных в этой области, некоторые из которых описаны ниже и в примерах.
В некоторых вариантах осуществления изобретения антитело содержит модифицированную константную область, такую как константная область, имеющая повышенную аффинность к Fc-рецептору гамма человека, является иммунологически инертным или частично инертным, например, не запускает комплемент-опосредованный лизис, не стимулирует антителозависимую клеточную цитотоксичность (ADCC) или не активирует макрофаги; или имеет сниженные активности (по сравнению с немодифицированным антителом) в любом одном или более из следующего: запуске комплемент-опосредованного лизиса, стимуляции антителозависимой клеточной цитотоксичности (ADCC) или активации микроглии. Для достижения оптимального уровня и/или комбинации эффекторных функций можно использовать различные модификации константной области. См., например, Morgan et al., Immunology 86:319-324, 1995; Lund et al., J. Immunology 157:4963-9 157:4963-4969, 1996; Idusogie et al., J. Immunology 164:4178-4184, 2000; Tao et al., J. Immunology 143: 2595-2601, 1989; и Jefferis et al., Immunological Reviews 163:59-76, 1998. В некоторых вариантах осуществления константная область модифицирована, как описано в Eur. J. Immunol., 29:2613-2624, 1999; заявке PCT № PCT/GB99/01441; и/или заявке Великобритании № 9809951.8. В других вариантах осуществления константная область является агликозилированной по N-связанному гликозилированию. В некоторых вариантах осуществления константная область является агликозилированной по N-связанному гликозилированию в результате мутации гликозилированного аминокислотного остатка или фланкирующих остатков, являющихся частью последовательности распознавания N-гликозилирования в константной области. Например, N-участок гликозилирования N297 можно подвергать мутации в A, Q, K или H. См., Tao et al., J. Immunology 143: 2595-2601, 1989; и Jefferis et al., Immunological Reviews 163:59-76, 1998. В некоторых вариантах осуществления константная область является агликозилированной по N-связанному гликозилированию. Константная область может являться агликозилированной по N-связанному гликозилированию ферментативно (например, посредством удаления углевода с помощью фермента ПНГазы) или посредством экспрессии в клетке-хозяине с недостаточностью гликозилирования.
Другие модификации антител включают антитела, модифицированные, как описано в публикации PCT № WO99/58572. Эти антитела содержат, в дополнение к связывающему домену, направленному против молекулы-мишени, эффекторный домен, имеющий аминокислотную последовательность, по существу, гомологичную всей или части константной области тяжелой цепи иммуноглобулина человека. Эти антитела могут связываться с молекулой-мишенью без запуска значительного комплемент-зависимого лизиса или клеточно-опосредованного разрешения мишени. В некоторых вариантах осуществления эффекторный домен может специфически связываться с FcRn и/или FcγRIIb. Они, как правило, основаны на химерных доменах, полученных из двух или более доменов CH2 тяжелой цепи иммуноглобулина человека. Антитела, модифицированные таким образом, особенно пригодны для использования в хронической терапии антителами, для избегания воспалительных и других нежелательных реакций на общепринятую терапию антителами.
Настоящее изобретение включает аффинно зрелые варианты осуществления. Например, аффинно зрелые антитела можно получать известными в этой области способами (Marks et al., Bio/Technology, 10:779-783, 1992; Barbas et al., Proc Nat. Acad. Sci, USA 91:3809-3813, 1994; Schier et al., Gene, 169:147-155, 1995; Yelton et al., J. Immunol., 155:1994-2004, 1995; Jackson et al., J. Immunol., 154(7):3310-9, 1995, Hawkins et al., J. Mol. Biol., 226:889-896, 1992; и публикация PCT № WO04/058184).
Следующие способы можно использовать для коррекции аффинности антитела и для характеризации CDR. Один из способов характеризации CDR антитела и/или изменения (такого как улучшение) аффинности связывания полипептида, такого как антитело, называется "сканирующий мутагенез библиотеки". Как правило, сканирующий мутагенез библиотеки осуществляют следующим образом. Одно или более положений аминокислот в CDR заменяют двумя или более (например, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20) аминокислотами с использованием способов, известных в этой области. Таким образом получают небольшие библиотеки клонов (в некоторых вариантах осуществления одну для каждого положения аминокислоты, подвергаемого анализу), каждая из которых имеет сложность два или более члена (если две или более аминокислот заменяют в каждом положении). Как правило, библиотека также включает клон, содержащий нативную (незамещенную) аминокислоту. Небольшое количество клонов, например, приблизительно 20-80 клонов (в зависимости от сложности библиотеки), из каждой библиотеки подвергают скринингу на аффинность связывания с полипептидом мишенью (или другой мишенью связывания) и идентифицируют кандидатов с повышенным, тем же, сниженным или отсутствующим связыванием. Способы определения аффинности связывания хорошо известны в этой области. Аффинность связывания можно определять с использованием анализа поверхностного плазмонного резонанса BIACORE™, с помощью которого определяют различия аффинности связывания приблизительно в 2 раза или более. BIACORE™ особенно подходит, если исходное антитело уже связывается с относительно высокой аффинностью, например, KD приблизительно 10 нМ или менее. Скрининг с использованием анализа поверхностного плазмонного резонанса BIACORE™ описан в примерах в настоящем описании.
Аффинность связывания можно определять с использованием биосенсора Kinexa, сцинтилляционного анализа сближения, ELISA, иммунологического анализа ORIGEN (IGEN), гашения флуоресценции, переноса флуоресценции и/или дрожжевого дисплея. Аффинность связывания также можно подвергать скринингу с использованием подходящего биологического анализа.
В некоторых вариантах осуществления каждое положение аминокислоты в CDR заменяют (в некоторых вариантах осуществления одно за один раз) всеми 20 природными аминокислотами с использованием известных в этой области способов мутагенеза (некоторые из которых представлены в настоящем описании). Это приводит к получению небольших библиотек клонов (в некоторых вариантах осуществления одной библиотеки на каждое положение аминокислоты, подвергаемое анализу), каждая из которых имеет сложность 20 членов (если все 20 аминокислот заменяют в каждом положении).
В некоторых вариантах осуществления библиотека, подлежащая скринингу, содержит замены в двух или более положениях, которые могут находиться в одной и той же CDR или двух или более CDR. Таким образом, библиотека может содержать замены в двух или более положениях в одной CDR. Библиотека может содержать замену в двух или более положениях в двух или более CDR. Библиотека может содержать замену в 3, 4, 5 или более положениях, где указанные положения, обнаруживаемые в двух, трех, четырех, пяти или шести CDR. Замену можно получать с использованием кодонов с низкой избыточностью. См., например, таблицу 2 из Balint et al., Gene 137(1):109-18, 1993.
Кандидатов с улучшенным связыванием можно секвенировать, таким образом, идентифицируя мутанта с заменой в CDR, приводящей к улучшенной аффинности (также обозначаемой как "улучшенная" замена). Кандидатов, которые связываются, также можно секвенировать, таким образом, идентифицируя замену CDR, сохраняющую связывание.
Можно осуществлять множество раундов скрининга. Например, кандидаты (каждый из которых содержит замену аминокислоты в одном или более положениях одной или более CDR) с улучшенным связыванием также подходят для дизайна второй библиотеки, содержащей по меньшей мере исходную и замещенную аминокислоту в каждом улучшенном положении CDR (т.е. положении аминокислоты в CDR, в котором мутант с заменой демонстрирует улучшенное связывание). Получение и скрининг или селекцию этой библиотеки дополнительно описывают ниже.
Сканирующий мутагенез библиотеки также представляет собой способ характеризации CDR, поскольку частота клонов с улучшенным связыванием, тем же связыванием, сниженным связыванием или отсутствием связывания также дает информацию, касающуюся важности каждого положения аминокислоты для стабильности комплекса антитело-антиген. Например, если положение CDR сохраняет связывание при изменении во все 20 аминокислот, это положение идентифицируют как положение, которое вряд ли требуется для связывания антигена. И наоборот, если положение в CDR сохраняет связывание лишь при небольшой доле замен, это положение идентифицируют как положение, важное для функции CDR. Таким образом, способами сканирующего мутагенеза библиотеки получают информацию, касающуюся положений в CDR, которые можно изменять во множество разных аминокислот (включая все 20 аминокислот), и положений в CDR, которые нельзя изменять или которые можно изменять в несколько аминокислот.
Кандидатов с улучшенной аффинностью можно комбинировать во второй библиотеке, включающей улучшенную аминокислоту, исходную аминокислоту в этом положении, и которая может дополнительно включать дополнительные замены в этом положении, в зависимости от сложности библиотеки, желаемой или допускаемой с использованием желаемого способа скрининга или селекции. Кроме того, при желании, смежное положение аминокислоты можно рандомизировать в по меньшей мере две или более аминокислот. Рандомизация смежных аминокислот может делать возможной дополнительную конформационную гибкость в мутантной CDR, что, в свою очередь, может делать возможным или облегчать встраивание большего количества улучшающих мутаций. Библиотека также может содержать замену в положениях, не демонстрирующих улучшенную аффинность в первом раунде скрининга.
Вторую библиотеку подвергают скринингу или селекции по членам библиотеки с улучшенной и/или измененной аффинностью связывания с использованием любого известного в этой области способа, включая скрининг с использованием анализа поверхностного плазмонного резонанса BIACORE™, и селекцию с использованием любого известного в этой области способа селекции, включая фаговый дисплей, дрожжевой дисплей и рибосомный дисплей.
Настоящее изобретение также относится к композициям, содержащим антитела, конъюгированные (например, связанные) со средством, облегчающим связывание с твердой подложкой (такой как биотин или авидин). Для простоты, как правило, можно ссылаться на антитела с пониманием того, что эти способы используют по отношению к любому из вариантов осуществления антител против GUCY2c, представленных в настоящем описании. Термин "конъюгация", как правило, относится к связыванию этих компонентов, как представлено в настоящем описании. Связывания (как правило, представляющего собой фиксацию этих компонентов вблизи друг друга по меньшей мере для введения) можно достигать рядом способов. Например, прямая реакция между средством и антителом возможна, если каждый из компонентов включает заместитель, способный реагировать с другим заместителем. Например, нуклеофильная группа, такая как аминогруппа или сульфгидрильная группа, на одном компоненте может реагировать с карбонил-содержащей группой, такой как ангидрид или галогенангидрид, или с алкильной группой, содержащей хорошую уходящую группу (например, галид) на другом.
В некоторых вариантах осуществления в качестве первой стадии выбирают первый и второй полипептид (и любые дополнительные полипептиды, образующие биспецифическое антитело). Как правило, нуклеиновую кислоту, кодирующую эти полипептиды, необходимо выделять таким образом, что ее можно изменять так, чтобы она кодировала выступ, или впадину, или и то, и другое, как определено в настоящем описании. Однако мутации можно вносить синтетическими способами, например, с использованием пептидного синтезатора. Кроме того, в случае, если замещенный остаток является неприродным остатком, доступен способ по Noren et al., выше, для получения полипептидов, имеющих такие замены. Кроме того, часть биспецифического антитела соответствующим образом получают рекомбинантно в культуре клеток, а другие части молекулы получают указанными выше способами.
Способы выделения антител и получения биспецифических антител являются следующими. Однако следует понимать, что биспецифические антитела можно получать из других полипептидов, или они могут включать другие полипептиды, с использованием способов, известных в этой области. Например, нуклеиновую кислоту, кодирующую интересующий полипептид (например, лиганд, рецептор или фермент), можно выделять из библиотеки кДНК, полученной из ткани, которая, как считают, содержит мРНК полипептида и экспрессирует ее на детектируемом уровне. Библиотеки подвергают скринингу с использованием зондов (таких как антитела или олигонуклеотиды из приблизительно 20-80 оснований), сконструированных для идентификации интересующего гена или белка, кодируемого им. Скрининг кДНК или геномной библиотеки с использованием выбранного зонда можно осуществлять стандартными способами, как описано в главах 10-12 Sambrook et al., Molecular Cloning: A Laboratory Manual (New York: Cold Spring Harbor Laboratory Press, 1989).
Как указано выше, термин "биспецифическое антитело" относится к комплексу двух или более полипептидных цепей, каждая из которых содержит по меньшей мере одну область VL антитела и одну область VH антитела или ее фрагмент, где области VL и VH в каждой полипептидной цепи получают из разных антител. В конкретных аспектах биспецифическое антитело включает димеры или тетрамеры полипептидных цепей, содержащие области VL и VH. Отдельные полипептидные цепи, содержащие мультимерные белки, можно ковалентно соединять по меньшей мере с одним другим пептидом мультимера посредством межцепочечных дисульфидных связей.
Биспецифические антитела по настоящему изобретению одновременно направленно воздействуют на T-клетки (CD3) и опухолевые клетки (GUCY2c) и успешно направляют и активируют T-клеточную цитотоксичность в отношении опухолевых клеток, экспрессирующих GUCY2c.
Каждая полипептидная цепь биспецифического антитела содержит область VL и область VH, которые можно ковалентно связывать с помощью линкера, где линкер является глицин-сериновым линкером, содержащим остатки глицина и серина таким образом, что связывающие домены антитела ограничены в отношении самосборки. В конкретном варианте осуществления глицин-сериновый линкер является линкером 1, содержащим последовательность SEQ ID NO: 190. Кроме того, каждая полипептидная цепь содержит домен гетеродимеризации, способствующий гетеродимеризации и/или стабилизации множества полипептидных цепей и снижающий вероятность гомодимеризации разных полипептидных цепей. Домен гетеродимеризации может находиться на N-конце полипептидной цепи или C-конце. Домен гетеродимеризации может содержать цистеиновый линкер (линкер 2), имеющий длину 1, 2, 3, 4, 5, 6 или более аминокислотных остатков. Взаимодействие двух полипептидных цепей может приводить к образованию двух пар VL/VH, образующих два эпитоп-связывающих домена, т.е. бивалентной молекулы. Ни одна из областей VH или VL не ограничена в каком-либо положении в полипептидной цепи, т.е. не ограничена амино-концом или карбокси-концом, области также не ограничены по своим положениям относительно друг друга, т.е. область VL может являться N-концевой относительно области VH и наоборот. Единственным ограничением является то, чтобы комплементарная полипептидная цепь была доступна для образования функционального биспецифического антитела. Если области VL и VH получают из антител, специфических для разных антигенов, для образования функционального биспецифического антитела необходимо взаимодействие двух разных полипептидных цепей, т.е. образование гетеродимера. В отличие от этого, если две разные полипептидные цепи свободны в своем взаимодействии, например, в рекомбинантной системе экспрессии, одна из которых содержит VLA и VHB (A является первым эпитопом, и B является вторым эпитопом), а другая содержит VLB и VHA, два разных участка связывания могут образовывать: VLA-VHA и VLB-VHB. Во всех парах полипептидных цепей биспецифического антитела возможно смещение или неправильное связывание двух цепей, например, взаимодействие областей VL-VL или VH-VH. Однако очистку функциональных биспецифических антител легко осуществлять с учетом иммуноспецифичности правильно димеризованного участка связывания с использованием любого способа на основе аффинности, известного в этой области или представленного в настоящем описании, например, аффинной хроматографии.
В одном из вариантов осуществления полипептидные цепи биспецифического антитела могут содержать различные линкеры и пептиды. Линкеры и пептиды могут содержать 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или более аминокислот. В некоторых вариантах осуществления VL против GUCY2c и VH против CD3 соединяют с помощью линкера 1 или линкера 2; и VL против CD3 и VH против GUCY2c соединяют с помощью линкера 1 или линкера 2. В некоторых таких вариантах осуществления линкер 1 содержит последовательность SEQ ID NO: 190. В других вариантах осуществления линкер 2 содержит последовательность SEQ ID NO: 191.
В некоторых вариантах осуществления домен 1 ковалентно связывают с первым способствующим гетеродимеризации доменом посредством цистеинового линкера и домен 2 ковалентно связывают со вторым способствующий гетеродимеризации доменом посредством цистеинового линкера. Каждый из цистеиновых линкеров включает по меньшей мере один остаток цистеина, делающий возможным образование внутримолекулярной дисульфидной связи. В дополнительных вариантах осуществления описанного выше цистеиновый линкер содержит по меньшей мере пять аминокислот. В некоторых таких вариантах осуществления цистеиновый линкер является линкером 3, содержащим последовательность SEQ ID NO: 192.
В некоторых вариантах осуществления первую полипептидную цепь ковалентно связывают со второй полипептидной цепью посредством по меньшей мере одной дисульфидной связи. В некоторых таких вариантах осуществления по меньшей мере одна дисульфидная связь образуется между линкером 3 первой полипептидной цепи и линкером 3 второй полипептидной цепи. В другом таком варианте осуществления по меньшей мере одна дисульфидная связь образуется между первым способствующим гетеродимеризации доменом и вторым способствующим гетеродимеризации доменом. В конкретных вариантах осуществления каждая дисульфидная связь образуется при связывании двух остатков цистеина.
Биспецифические антитела по изобретению одновременно могут связываться с двумя отдельными и отличающимися эпитопами. В некоторых вариантах осуществления по меньшей мере один участок связывания эпитопа является специфическим в отношении детерминанты CD3, экспрессирующейся на иммунной эффекторной клетке, например, экспрессирующейся на T-лимфоцитах. В одном из вариантов осуществления молекула биспецифического антитела связывается с детерминантой эффекторной клетки, а также активирует эффекторную клетку.
В конкретных вариантах осуществления биспецифическое антитело по настоящему изобретению (a) связывается с внеклеточным доменом GUCY2c человека, (b) демонстрирует повышенное время полужизни в сыворотке и опухоли от 30 мин до 100 дней; и/или (c) демонстрирует более низкое значение EC50 от 0,0001 нМ до 100 нМ в присутствии повышенных уровней экспрессии GUCY2c или повышенных уровней плотности рецепторов.
В одном из вариантов осуществления эпитоп-связывающий домен может связываться с опухолеспецифическим антигеном GUCY2c, ассоциированным с раком молочной железы, яичников, щитовидной железы, предстательной железы, шейки матки, легких (включая, в качестве неограничивающих примеров, немелкоклеточный рак легких и мелкоклеточный рак легких), мочевого пузыря, эндометрия, головы и шеи, яичек и глиобластомой. В некоторых вариантах осуществления злокачественное новообразование является злокачественным новообразованием пищеварительной системы, выбранным из группы, состоящей из злокачественного новообразования пищевода, желудка, тонкого кишечника, толстого кишечника, прямой кишки, анального канала, печени, желчного пузыря, аппендикса, желчных протоков и поджелудочной железы. В конкретных вариантах осуществления терапия активирует ответ цитолитических T-клеток.
Биспецифические антитела по настоящему изобретению содержат антигенсвязывающие домены, как правило, полученные из иммуноглобулинов или антител. Антитела, из которых получают связывающие домены, используемые в способах по изобретению, можно получать из любого животного, включая птиц и млекопитающих (например, человека, не являющегося человеком примата, мышь, осла, овцу, кролика, козу, морскую свинку, верблюда, лошадь или курицу). Предпочтительно, антитела являются человеческими или гуманизированными моноклональными антителами.
Биспецифические антитела по настоящему изобретению можно охарактеризовывать различными способами. В частности, молекулы по изобретению можно анализировать на способность иммуноспецифически связываться с антигеном. Молекулы, идентифицированные как иммуноспецифически связывающиеся с антигеном, затем можно анализировать на их специфичность и аффинность к антигену.
Биспецифическое антитело по настоящему изобретению можно получать различными способами, хорошо известными в этой области, включая синтез белков de novo и рекомбинантную экспрессию нуклеиновых кислот, кодирующих связывающие белки. Желаемые последовательности нуклеиновой кислоты можно получать рекомбинантными способами (например, посредством ПЦР-мутагенеза ранее полученного варианта желаемого полинуклеотида) или посредством твердофазного синтеза ДНК. Как правило, используют способы рекомбинантной экспрессии. В одном из аспектов настоящее изобретение относится к полинуклеотиду, содержащему последовательность, кодирующую VH и/или VL против CD3; в другом аспекте изобретение относится к полинуклеотиду, содержащему последовательность, кодирующую VH и/или VL против GUCY2c. Из-за вырожденности генетического кода различные последовательности нуклеиновой кислоты кодируют каждую аминокислотную последовательность иммуноглобулина, и настоящее изобретение относится ко всем нуклеиновым кислотам, кодирующим связывающие белки, представленные в настоящем описании.
Биспецифическое антитело по настоящему изобретению может содержать два scFV-домена, связывающихся с антигеном-мишенью и CD3, слитые с Fc-доменом IgG. В одном из вариантов осуществления изобретения биспецифическое антитело содержит два scFV-домена, связывающихся с GUCY2c и CD3 эпсилон, слитые с Fc-доменом IgG1. В частности, на фигурах 1A, 1B, 1C и 1D показаны четыре альтернативные схемы биспецифических антител против CD3-GUCY2c. Биспецифическое антитело содержит первый способствующий гетеродимеризации и второй способствующий гетеродимеризации домен. Каждый первый способствующий гетеродимеризации и второй способствующий гетеродимеризации домен может содержать домен CH2 и/или домен CH3 Fc-области иммуноглобулина, где домен CH2 и/или домен CH3 изменяют так, чтобы он содержал выступ или впадину. Модификации Fc-части биспецифического антитела описаны ниже.
В аспекте настоящего изобретения биспецифическое антитело, как показано на фигурах 1A, 1B, 1C и 1D, содержит первую полипептидную цепь и вторую полипептидную цепь.
В одном из аспектов настоящее изобретение относится к биспецифическому антителу, представленному в настоящем описании, где: (a) первая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 1,содержащий VL антитела против GUCY2c (VL против GUCY2c) и VH антитела против CD3 (VH против CD3), и (ii) первый способствующий гетеродимеризации домен; и (b) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 2, содержащий VL антитела против CD3 (VL против CD3) и VH антитела против GUCY2c (VH против GUCY2c), и (ii) второй способствующий гетеродимеризации домен.
В другом аспекте изобретение относится к биспецифическому антителу, представленному в настоящем описании, где: (a) первая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 1, содержащий VL против CD3 и VH против GUCY2c, и (ii) первый способствующий гетеродимеризации домен; и (b) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 2, содержащий VL против GUCY2c и VH против CD3, и (ii) второй способствующий гетеродимеризации домен.
В еще одном аспекте изобретение относится к биспецифическому антителу, представленному в настоящем описании, где: (a) первая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 1, содержащий VH против GUCY2c и VL против CD3, и (ii) первый способствующий гетеродимеризации домен; и (b) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 2, содержащий VH против CD3 и VL против GUCY2c, и (ii) второй способствующий гетеродимеризации домен.
В другом аспекте изобретение относится к биспецифическому антителу, представленному в настоящем описании, где: (a) первая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 1, содержащий VH против CD3 и VL против GUCY2c, и (ii) первый способствующий гетеродимеризации домен; и (b) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: (i) домен 2, содержащий VH против GUCY2c и VL против CD3 и (ii) второй способствующий гетеродимеризации домен.
В некоторых вариантах осуществления описанного выше VL против GUCY2c и VH против GUCY2c образуют домен, специфически связывающийся с GUCY2c; и VL против CD3 и VH против CD3 образуют домен, специфически связывающийся с CD3.
В дополнительных вариантах осуществления любого из описанных выше биспецифическое антитело по настоящему изобретению содержит (a) CDR1 VH против GUCY2c, содержащую последовательность SEQ ID NO: 12, 20, 27, 34, 42, 74, 257, 258, 259, 260,или 261; (b) CDR2 VH против GUCY2c, содержащую последовательность SEQ ID NO: 13, 21, 28, 35, 43, 53, 66, 68, 70, 72, 75, 262, 263, 264, 265, 266 или 267; (c) CDR3 VH против GUCY2c, содержащую последовательность SEQ ID NO: 14, 22, 29, 36 или 44; (d) CDR1 VL против GUCY2c, содержащую последовательность SEQ ID NO: 93, 101, 105, 107, 113, 120, 148, 153, 163 или 167; (e) CDR2 VL против GUCY2c, содержащую последовательность SEQ ID NO: 78, 94, 102, 108, 114, 141, 144, 146, 149, 151, 157, 159, 161, 164 или 168; и (f) CDR3 VL против GUCY2c, содержащую последовательность SEQ ID NO: 95, 109, 115, 121, 142, 154, 165 или 169.
В конкретном варианте осуществления биспецифическое антитело по настоящему изобретению содержит (a) CDR1 VH против GUCY2c, содержащую последовательность SEQ ID NO: 74; (b) CDR2 VH против GUCY2c, содержащую последовательность SEQ ID NO: 75; (c) CDR3 VH против GUCY2c, содержащую последовательность SEQ ID NO: 29; (d) CDR1 VL против GUCY2c, содержащую последовательность SEQ ID NO: 148; (e) CDR2 VL против GUCY2c, содержащую последовательность SEQ ID NO: 149; и (f) CDR3 VL против GUCY2c, содержащую последовательность SEQ ID NO: 142.
В конкретных вариантах осуществления биспецифическое антитело содержит: (a) область VH против GUCY2c, содержащую последовательность SEQ ID NO: 73; и (b) область VL против GUCY2c, содержащую последовательность, приведенную в SEQ ID NO: 147.
В дополнительных вариантах осуществления настоящее изобретение относится к биспецифическим антителам, содержащим (a) CDR1 VH против CD3, содержащую последовательность SEQ ID NO: 2; (b) CDR2 VH против CD3, содержащую последовательность SEQ ID NO: 3 или 10; (c) CDR3 VH против CD3, содержащую последовательность SEQ ID NO: 4; (d) CDR1 VL против CD3, содержащую последовательность SEQ ID NO: 77, 85, или 91; (e) CDR2 VL против CD3, содержащую SEQ ID NO: 78; и (f) CDR3 VL против CD3, содержащую последовательность SEQ ID NO: 79.
В конкретном варианте осуществления настоящее изобретение относится к биспецифическому антителу, содержащему (a) CDR1 VH против CD3, содержащую последовательность SEQ ID NO: 2; (b) CDR2 VH против CD3, содержащую последовательность SEQ ID NO: 10; (c) CDR3 VH против CD3, содержащую последовательность SEQ ID NO: 4; (d) CDR1 VL против CD3, содержащую последовательность SEQ ID NO: 91; (e) CDR2 VL против CD3, содержащую последовательность SEQ ID NO: 78; и (f) CDR3 VL против CD3, содержащую последовательность SEQ ID NO: 79.
В одном из вариантов осуществления настоящее изобретение относится к биспецифическому антителу, содержащему: (a) область VH против CD3, содержащую последовательность SEQ ID NO: 1, 9 или 273; и (b) область VL против CD3, содержащую последовательность SEQ ID NO: 76, 84, 90, 274, 275 или 276. В конкретном варианте осуществления биспецифическое антитело по настоящему изобретению содержит: (a) область VH против CD3, содержащую последовательность SEQ ID NO: 9; и (b) область VL против CD3, содержащую последовательность SEQ ID NO: 90.
В другом аспекте по изобретению линкер 3 может содержать укороченную нижнюю шарнирную область IgG1 человека, имеющую последовательность GCPPCP (SEQ ID NO: 192), с по меньшей мере одним остатком глицина, предшествующим нижней шарнирной области.
В одном из аспектов первый способствующий гетеродимеризации домен может содержать Fc-цепь, содержащую домен CH2 и/или CH3, измененную так, чтобы содержать Fc-цепь с выступом, содержащую последовательность SEQ ID NO: 188, 196, 199, 202, 205, 210, 216, или 248, 282, 284 или 286; или Fc-цепь с впадиной, содержащую последовательность SEQ ID NO: 189, 197, 200, 203, 206, 211, 217, 220, 249, 283, 285 или 287. В другом аспекте второй способствующий гетеродимеризации домен может содержать Fc-цепь, содержащую домен CH2 и/или CH3, измененную так, чтобы содержать Fc-цепь с выступом, содержащую последовательность SEQ ID NO: 188, 196, 199, 202, 205, 210, 216 или 248, 282, 284 или 286, если такая первая гетеродимерная область содержит Fc-цепь с впадиной, или Fc-цепь с впадиной, содержащую последовательность SEQ ID NO: 189, 197, 200, 203, 206, 211, 217, 220, 249, 283, 285 или 287, если такая первая гетеродимерная область содержит Fc-цепь с выступом (таблица 7).
В таблице 9 приведены последовательности нуклеиновых кислот первой полипептидной цепи и второй полипептидной цепи биспецифических антител с различными антителами против GUCY2c м CD3, представленными в настоящем описании. В SEQ ID NO: 246 и 247 приведены соответствующие нуклеотидные последовательности по изобретению.
Таблица 9
Эффекторные нуль-мутации в CH2-CH3 IgG1 человека
Fc-цепь IgG1 человека модифицировали для встраивания мутаций L234A, L235A и G237A (SEQ ID NO: 187, нумерация в соответствии с индексом EU) с использованием стандартного, направляемого праймерами ПЦР-мутагенеза для снижения эффекторной функции по причине связывания с FcγRIII, получая нуль-фенотип эффекторной функции (Canfield et al., J. Exp. Med (1991) 173:1483-1491; Shields et al., J. Biol. Chem.(2001) 276:6591-604).
Мутации "выступы-во-впадины" в CH2-CH3 IgG1 человека
"Выступы-во-впадины" представляют собой эффективную стратегию дизайна, известную в этой области для конструирования гомодимеров тяжелой цепи антитела для гетеродимеризации. В этом подходе вариант с "выступом" получали посредством замены небольшой аминокислоты более крупной аминокислотой в одной цепи Fc-цепи IgG1, например, Y349C и T366W, (нумерация в соответствии с индексом EU). "Выступ" конструировали для вставки во "впадину" в домене CH3 комплементарной цепи Fc-цепи, полученной посредством замены крупного остатка меньшим остатком, например, S354C, T366S, L368A и Y407V (нумерация в соответствии с индексом EU).
В некоторых вариантах осуществления встраивали комплементарные мутации для достижения гетеродимеризации получаемых Fc-цепей, таким образом, что каждая Fc-цепь будет нести набор мутаций, Y349C и T366W в случае Fc-цепи с выступом (SEQ ID NO: 188), или S354C, T366S, L368A и Y407V в случае Fc-цепи с впадиной (SEQ ID NO: 189), как показано в таблице 10. При совместной трансфекции в подходящую ДНК млекопитающего-хозяина, кодирующую аминокислотные последовательности например SEQ ID NO: 246 и 247, получают Fc-домен, т.е. преимущественно биспецифическое антитело содержит одну Fc-цепь с выступом, связанную с одной Fc-цепью с впадиной.
Таблица 10
Биспецифические антитела против CD3-GUCY2c по настоящему изобретению являются стабильными против агрегации в исследованиях термической стабильности и мощными слитыми белками биспецифическое антитело-Fc против CD3 и GUCY2c человека. Fc-домен с "выступом-во-впадину" делает возможной улучшенную экспрессию в клетках CHO и улучшенную очистку, приводящую к высокой чистоте желаемого гетеродимера. Мутации, сконструированные в Fc-домене, устраняют связывание FcγR, таким образом, потенциально позволяя избегать ADCC-опосредованного истощения T-клеток. Кроме того, включение Fc-домена в биспецифическое антитело повышает стабильность молекул, анализируемую посредством дифференциальной сканирующей калориметрии (DSC).
Биспецифическое антитело можно конструировать так, чтобы оно содержало по меньшей мере один остаток цистеина, который может взаимодействовать с соответствующим остатком цистеина на другой полипептидной цепи по изобретению с образованием межцепочечной дисульфидной связи. Межцепочечные дисульфидные связи могут служить для стабилизации биспецифического антитела, улучшая экспрессию и выделение в рекомбинантных системах, что приводит к стабильному и единообразному составу, а также улучшая стабильность выделенного и/или очищенного продукта in vivo. Остаток или остатки цистеина можно встраивать в виде отдельной аминокислоты или как часть более крупной аминокислотной последовательности, например, шарнирной области, в любой части полипептидной цепи. В конкретном аспекте по меньшей мере один остаток цистеина конструируют так, чтобы он находился на C-конце полипептидной цепи.
В одном из аспектов настоящее изобретение относится к биспецифическому антителу, специфически связывающемуся с GUCY2c и конкурирующему за связывание с биспецифическим антителом, представленным в настоящем описании. В некоторых таких вариантах осуществления биспецифическое антитело специфически связывается с эпитопом GUCY2c.
В одном из аспектов настоящее изобретение относится к биспецифическому антителу, специфически связывающемуся с GUCY2c и CD3, где биспецифическое антитело содержит первую полипептидную цепь и вторую полипептидную цепь, и где: (a) первая полипептидная цепь содержит следующие области в следующем порядке в направлении от N-конца к C-концу: VL антитела против GUCY2c (VL против GUCY2c) (SEQ ID NO: 147) - линкер 1 (SEQ ID NO: 190) - VH антитела против CD3 (VH против CD3) (SEQ ID NO: 9) - линкер 3 (SEQ ID NO: 192) - первый способствующий гетеродимеризации домен (SEQ ID NO: 188); и (b) вторая полипептидная цепь содержит следующие области в следующем порядке в направлении от N-конца к C-концу: VL антитела против CD3 (VL против CD3) (SEQ ID NO: 90) - линкер 2 (SEQ ID NO: 191) - VH антитела против GUCY2c (VH против GUCY2c) (SEQ ID NO: 73) - линкер 3 (SEQ ID NO: 192) - второй способствующий гетеродимеризации домен (SEQ ID NO: 189).
В другом аспекте изобретение относится к биспецифическому антителу, специфически связывающемуся с GUCY2c и CD3, где биспецифическое антитело содержит первую полипептидную цепь и вторую полипептидную цепь, и где: (a) первая полипептидная цепь содержит следующие области в следующем порядке в направлении от N-конца к C-концу: VL антитела против GUCY2c (VL против GUCY2c) (SEQ ID NO: 147) - линкер 1 (SEQ ID NO: 190) - VH антитела против CD3 (VH против CD3) (SEQ ID NO: 9) - линкер 3 (SEQ ID NO: 192) - первый способствующий гетеродимеризации домен (SEQ ID NO: 188); и (b) вторая полипептидная цепь содержит следующие области в следующем порядке в направлении от N-конца к C-концу: VL антитела против CD3 (VL против CD3) (SEQ ID NO: 90) - линкер 2 (SEQ ID NO: 191) -VH антитела против GUCY2c (VH против GUCY2c) (SEQ ID NO: 73) - линкер 3 (SEQ ID NO: 192) -второй способствующий гетеродимеризации домен (SEQ ID NO: 189); где VL против GUCY2c и VH против GUCY2c образуют домен, специфически связывающийся с GUCY2c; и VL против CD3 и VH против CD3 образуют домен, специфически связывающийся с CD3; где линкер 3 первой полипептидной цепи и линкер 3 второй полипептидной цепи ковалентно связаны друг с другом двумя дисульфидными связями; где каждый из первого способствующего гетеродимеризации домена и второго способствующего гетеродимеризации домена содержит домен CH2 и домен CH3; где домен CH3 первого способствующего гетеродимеризации домена образует выступ, и домен CH3 второго способствующего гетеродимеризации домена образует впадину; где по меньшей мере одна дисульфидная связь образуется между доменом CH3 первого способствующего гетеродимеризации домена и доменом CH3 второго способствующего гетеродимеризации домена.
В некоторых таких вариантах осуществления настоящее изобретение относится к биспецифическому антителу, специфически связывающемуся с эпитопом GUCY2c и эпитопом CD3.
В некоторых вариантах осуществления настоящее изобретение относится к биспецифическому антителу, дополнительно содержащему человеческий или гуманизированный каркас VH и человеческий или гуманизированный каркас VL. В некоторых таких вариантах осуществления биспецифическое антитело является гуманизированным антителом. В конкретных вариантах осуществления каркас VH содержит последовательность SEQ ID NO: 5, 6, 7, 8, 15, 16, 17, 18, 23, 24, 25, 30, 31, 32, 37, 38, 39, 40, 45, 46, 47, 49, 50, 51, 54, 55, 56, 58, 59, 61 или 63; и/или каркас VL содержит последовательность SEQ ID NO: 80, 81, 82, 83, 86, 87, 88, 89, 96, 97, 98, 99, 103, 110, 111, 116, 117, 118, 122, 123, 124, 126, 127, 128, 130, 131, 132, 133, 135, 139 или 155.
В одном из аспектов биспецифическое антитело по настоящему изобретению специфически связывается с GUCY2c и CD3, где биспецифическое антитело содержит первую полипептидную цепь и вторую полипептидную цепь; где первую полипептидную цепь получают с помощью экспрессирующего вектора с регистрационным номером ATCC PTA-124944; и вторую полипептидную цепь получают с помощью экспрессирующего вектора с регистрационным номером ATCC PTA-124943.
В одном из аспектов настоящее изобретение относится к биспецифическому антителу, способному специфически связываться с GUCY2c и CD3, содержащему первую полипептидную цепь и вторую полипептидную цепь, где первая полипептидная цепь содержит последовательность SEQ ID NO: 216, и вторая полипептидная цепь содержит последовательность SEQ ID NO: 220.
В некоторых таких вариантах осуществления настоящее изобретение относится к биспецифическому антителу, где первая и вторая полипептидные цепи ковалентно связаны друг с другом по меньшей мере одной дисульфидной связью.
Настоящее изобретение дополнительно относится к фармацевтической композиции, содержащей терапевтически эффективное количество антитела или биспецифического антитела, представленного в настоящем описании, и фармацевтически приемлемый носитель.
Настоящее изобретение относится к способам и композициям для лечения, профилактики или контроля злокачественного новообразования у индивидуума, включающим введение индивидууму терапевтически эффективного количества антитела или биспецифических антител, сконструированных по изобретению, где молекула дополнительно связывается с антигеном злокачественной опухоли. Молекулы по изобретению могут особенно подходить для профилактики, ингибирования, уменьшения роста и/или регрессирования первичных опухолей и метастазирования злокачественных клеток. Хотя они и не ограничены каким-либо конкретным механизмом действия, молекулы по изобретению могут опосредовать эффекторную функцию, что может приводить к клиренсу опухоли, уменьшению опухоли или их комбинации.
Антитела (например, против GUCY2c или биспецифические антитела против CD3-GUCY2c) по настоящему изобретению можно использовать по-разному, включая, в качестве неограничивающих примеров, способы терапевтического лечения и способы диагностики.
В одном из аспектов настоящее изобретение относится к способу лечения состояния, ассоциированного с экспрессией GUCY2c, у индивидуума. В некоторых вариантах осуществления способ лечения состояния, ассоциированного с экспрессией GUCY2c у индивидуума, включает введение нуждающемуся в этом индивидууму эффективного количества композиции (например, фармацевтической композиции), содержащей антитела против GUCY2c или биспецифические антитела против GUCY2c, представленные в настоящем описании. Состояния, ассоциированные с экспрессией GUCY2c, включают, в качестве неограничивающих примеров, аномальную экспрессию GUCY2c, измененную или аномальную экспрессию GUCY2c, злокачественные клетки, экспрессирующие GUCY2c, и пролиферативное нарушение (например, злокачественное новообразование).
Таким образом, изобретение относится к способам лечения злокачественного новообразования, отличающегося антигеном GUCY2c, посредством конструирования биспецифического антитела для иммуноспецифического распознавания антигена GUCY2c и антигена CD3 на T-клетках. Биспецифические антитела, сконструированные по изобретению, можно использовать для профилактики или лечения злокачественного новообразования, т.к. они обладают цитотоксической активностью благодаря индуцируемым антителом против CD3, активированным киллерным T-клеткам.
В конкретном варианте осуществления настоящее изобретение относится к способу лечения GUCY2c-ассоциированного нарушения у нуждающегося в этом пациента, включающему введение пациенту антитела против GUCY2c, представленного в настоящем описании. Настоящее изобретение также относится к способу лечения GUCY2c-ассоциированного нарушения у нуждающегося в этом пациента, включающему введение пациенту биспецифического антитела по настоящему изобретению.
Настоящее изобретение дополнительно относится к способу лечения GUCY2c-ассоциированного нарушения у нуждающегося в этом пациента, включающему введение пациенту фармацевтической композиции, содержащей антитело против GUCY2c или биспецифическое антитело, представленное в настоящем описании. В некоторых таких вариантах осуществления GUCY2c-ассоциированное нарушение является злокачественным новообразованием. В конкретных вариантах осуществления злокачественное новообразование является раком молочной железы, яичников, щитовидной железы, предстательной железы, шейки матки, легких (включая, в качестве неограничивающих примеров, немелкоклеточный рак легких и мелкоклеточный рак легких), мочевого пузыря, эндометрия, головы и шеи, яичка или глиобластому. В некоторых вариантах осуществления злокачественное новообразование является злокачественным новообразованием пищеварительной системы, выбранным из группы, состоящей из злокачественного новообразования пищевода, желудка, тонкого кишечника, толстого кишечника, прямой кишки, анального канала, печени, желчного пузыря, аппендикса, желчных протоков и поджелудочной железы. В конкретных вариантах осуществления с помощью терапии активируют ответ цитолитических T-клеток.
В одном из аспектов настоящее изобретение относится к антителу, биспецифическому антителу или фармацевтической композиции, представленным в настоящем описании, для использования в терапии. В конкретном варианте осуществления изобретение также относится к антителу против GUCY2c или биспецифическому антителу против CD3-GUCY2c для использования в способе лечения злокачественного новообразования, определенном в настоящем описании.
Настоящее изобретение дополнительно относится к антителу или биспецифическому антителу, представленному в настоящем описании, для использования в производстве лекарственного средства для использования в терапии. В некоторых вариантах осуществления терапия представляет собой лечения GUCY2c-ассоциированного нарушения. В конкретных вариантах осуществления GUCY2c-ассоциированное нарушение является злокачественным новообразованием. В некоторых вариантах осуществления злокачественное новообразование выбрано из группы, состоящей из рака молочной железы, яичников, щитовидной железы, предстательной железы, шейки матки, легких (включая, в качестве неограничивающих примеров, немелкоклеточный рак легких и мелкоклеточный рак легких), мочевого пузыря, эндометрия, головы и шеи, яичка или глиобластомы. В некоторых вариантах осуществления злокачественное новообразование является злокачественным новообразованием пищеварительной системы, выбранным из группы, состоящей из злокачественного новообразования пищевода, желудка, тонкого кишечника, толстого кишечника, прямой кишки, анального канала, печени, желчного пузыря, аппендикса, желчных протоков и поджелудочной железы. В конкретных вариантах осуществления с помощью терапии активируют ответ цитолитических T-клеток.
В одном из аспектов настоящее изобретение относится к полинуклеотиду, кодирующему антитело или биспецифическое антитело, представленное в настоящем описании. В другом варианте осуществления настоящее изобретение относится к вектору, содержащему полинуклеотиды, представленные в настоящем описании. В еще одном варианте осуществления настоящее изобретение относится к клетке-хозяину, трансформированной или трансфицированной с использованием векторов, представленных в настоящем описании. В некоторых таких вариантах осуществления клетка-хозяин рекомбинантно продуцирует антитело или биспецифическое антитело, представленное в настоящем описании. В конкретных вариантах осуществления, клетка-хозяин выбрана из группы, состоящей из линий бактериальных клеток, линий клеток млекопитающих, линий клеток насекомых и линий дрожжевых клеток. В конкретном варианте осуществления линия клеток млекопитающих является линией клеток CHO. В одном из вариантов осуществления антитело или биспецифическое антитело получают с использованием бесклеточной система синтеза белка in vitro.
В одном из аспектов настоящее изобретение относится к способу лечения злокачественного новообразования у нуждающегося в этом индивидуума, включающему a) получение биспецифического антитела, представленного в настоящем описании, и b) введение указанного биспецифического антитела указанному пациенту. В некоторых вариантах осуществления настоящее изобретение относится к способу лечения состояния, ассоциированного со злокачественными клетками, экспрессирующими опухолевый антиген, у индивидуума, включающему a) получение антитела, представленного в настоящем описании, и b) введение нуждающемуся в этом индивидууму эффективного количества фармацевтической композиции, содержащей антитело, представленное в настоящем описании. Неограничивающие примеры злокачественного новообразования, подлежащего лечению, включают рак пищевода, рак желудка, рак тонкого кишечника, рак толстого кишечника, рак прямой кишки, рак анального канала, рак печени, рак желчного пузыря, рак аппендикса, рак желчных протоков и рак поджелудочной железы. Таким образом, композиции и способы, представленные в настоящем описании, можно использовать для лечения, визуализации, детекции и вакцинации против указанных выше первичных и метастатических злокачественных новообразований.
В некоторых вариантах осуществления настоящее изобретение относится к способу ингибирования роста или прогрессирования опухоли у индивидуума, имеющего злокачественные клетки, экспрессирующие GUCY2c, включающему введение нуждающемуся в этом индивидууму эффективного количества композиции, содержащей антитела против GUCY2c или биспецифические антитела против CD3-GUCY2c, представленные в настоящем описании. В других вариантах осуществления настоящее изобретение относится к способу ингибирования метастазирования клеток, экспрессирующих GUCY2c, у индивидуума, включающему введение нуждающемуся в этом индивидууму эффективного количества композиции, содержащей антитела против GUCY2c или биспецифические антитела против CD3-GUCY2c, как представлено в настоящем описании. В других вариантах осуществления настоящее изобретение относится к способу индуцирования регрессирования опухоли в злокачественных клетках у индивидуума, включающему введение нуждающемуся в этом индивидууму эффективного количества композиции, содержащей антитела против GUCY2c или биспецифические антитела против CD3-GUCY2c, представленные в настоящем описании.
В конкретном аспекте антитела против GUCY2c или биспецифическое антитело против CD3-GUCY2c по изобретению ингибируют или снижают рост злокачественных клеток по меньшей мере на 99%, по меньшей мере на 95%, по меньшей мере на 90%, по меньшей мере на 85%, по меньшей мере на 80%, по меньшей мере на 75%, по меньшей мере на 70%, по меньшей мере на 60%, по меньшей мере на 50%, по меньшей мере на 45%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 35%, по меньшей мере на 30%, по меньшей мере на 25%, по меньшей мере на 20% или по меньшей мере на 10% относительно роста злокачественных клеток в отсутствие антитела или биспецифического антитела по изобретению.
В конкретном аспекте антитела против GUCY2c или биспецифические антитела против CD3-GUCY2c уничтожают клетки или ингибируют или снижают рост злокачественных клеток по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 100% лучше, чем в отсутствие антитела или биспецифических антител по изобретению.
В одном из вариантов осуществления настоящее изобретение относится к способу лечения аутоиммунного нарушения у индивидуума, включающему введение нуждающемуся в этом индивидууму эффективного количества композиции, содержащей антитела против GUCY2c или биспецифические антитела против CD3-GUCY2c, представленные в настоящем описании.
В рамках изобретения аутоиммунные нарушения включают, в качестве неограничивающих примеров, воспалительное заболевание кишечника, системную красную волчанку, диабет (типа I), целиакию, агаммаглобулинемию, аутоиммунную энтеропатию, аутоиммунный гепатит, болезнь Крона, пемфигоид желудочно-кишечного тракта, полимиозит, поперечный миелит, язвенный колит и васкулит.
В одном из аспектов настоящее изобретение относится к эффективному количеству композиции (например, фармацевтической композиции), содержащей антитела (например, антитела против GUCY2c или биспецифические антитела против CD3-GUCY2c), представленные в настоящем описании, для лечения состояния (например, злокачественного новообразования), ассоциированного с экспрессией GUCY2c, у нуждающегося в этом индивидуума. В одном из вариантов осуществления настоящее изобретение относится к эффективному количеству композиции (например, фармацевтической композиции), содержащей антитела (например, антитела против GUCY2c или биспецифические антитела против CD3-GUCY2c), как представлено в настоящем описании, для ингибирования роста или прогрессирования опухоли у индивидуума, имеющие злокачественные клетки, экспрессирующие GUCY2c. В некоторых вариантах осуществления настоящее изобретение относится к эффективному количеству композиции (например, фармацевтической композиции), содержащей антитела (например, антитела против GUCY2c или биспецифические антитела против CD3-GUCY2c), как представлено в настоящем описании, для ингибирования метастазирования злокачественных клеток, экспрессирующих GUCY2c, у нуждающегося в этом индивидуума. В некоторых вариантах осуществления настоящее изобретение относится к эффективному количеству композиции (например, фармацевтической композиции), содержащей антитела (например, антитела против GUCY2c или биспецифические антитела против CD3-GUCY2c), представленные в настоящем описании.
В другом аспекте изобретение относится к антителам (например, антителам GUCY2c или биспецифическим антителам против CD3-GUCY2c), представленным в настоящем описании, для использования в лечении состояния (например, злокачественного новообразования), ассоциированного с экспрессией GUCY2c, у нуждающегося в этом индивидуума. В некоторых вариантах осуществления настоящее изобретение относится к антителам (например, антителам против GUCY2c или биспецифическим антителам против CD3-GUCY2c), представленным в настоящем описании, для ингибирования роста или прогрессирование опухоли у индивидуума, имеющего злокачественные клетки, экспрессирующие GUCY2c. В некоторых вариантах осуществления настоящее изобретение относится к антителам (например, антителам против GUCY2c или биспецифическим антителам против CD3-GUCY2c), представленным в настоящем описании, для ингибирования метастазирования злокачественных клеток, экспрессирующих GUCY2c, у нуждающегося в этом индивидуума. В некоторых вариантах осуществления настоящее изобретение относится к антителам (например, антителам GUCY2c или биспецифическим антителам против CD3-GUCY2c), представленным в настоящем описании, для индуцирования регрессирования опухоли у индивидуума, имеющего злокачественные клетки, экспрессирующие GUCY2c.
В другом аспекте изобретение относится к применению антител (например, антител против GUCY2c или биспецифических антител против CD3-GUCY2c), представленных в настоящем описании, в производстве лекарственного средства для лечения состояния (например, злокачественного новообразования), ассоциированного с экспрессией GUCY2c. В некоторых вариантах осуществления настоящее изобретение относится к применению антител (например, антител против GUCY2c или биспецифических антител против CD3-GUCY2c), представленных в настоящем описании, в производстве лекарственного средства для ингибирования роста или прогрессирования опухоли. В некоторых вариантах осуществления настоящее изобретение относится к применению антител (например, антител против GUCY2c или биспецифических антител против CD3-GUCY2c), представленных в настоящем описании, в производстве лекарственного средства для ингибирования метастазирования злокачественных клеток, экспрессирующих GUCY2c. В некоторых вариантах осуществления настоящее изобретение относится к применению антител (например, антител против GUCY2c или биспецифических антител против CD3-GUCY2c), представленных в настоящем описании, в производстве лекарственного средства для индуцирования регрессирования опухоли.
В другом аспекте настоящее изобретение относится к способу детекции, диагностики и/или мониторинга состояния, ассоциированного с экспрессией GUCY2c. Например, антитела (например, антитела против GUCY2c или биспецифические антитела против CD3-GUCY2c), представленные в настоящем описании, можно метить детектируемым фрагментом, таким как визуализирующее средство и фермент-субстратная метка. Антитела, представленные в настоящем описании, также можно использовать для диагностических анализов in vivo, таких как визуализация in vivo (например, ПЭТ или SPECT), или в качестве реагента для окрашивания.
Таким образом, в дополнение к перенаправлению T-клеток к опухолеспецифическим антигенам, биспецифические антитела против CD3-GUCY2c по настоящему изобретению можно использовать для доставки других диагностических или терапевтических средств к клеткам, экспрессирующим опухолевый антиген на своей поверхности. Таким образом, биспецифическое антитело можно присоединять прямо или косвенно, например, через линкер, с лекарственным средством таким образом, что его будут доставлять непосредственно в клетки, несущие опухолевый антиген. Терапевтические средства включают, в качестве неограничивающих примеров, нуклеиновые кислоты, белки, пептиды, аминокислоты или производные, гликопротеины, радиоактивные изотопы, липиды, углеводы или рекомбинантные вирусы. Диагностические или терапевтические средства на основе нуклеиновых кислот включают, в качестве неограничивающих примеров, антисмысловые нуклеиновые кислоты, дериватизированные олигонуклеотиды для ковалентной перекрестной сшивки с одиночной или дуплексной ДНК и триплекс-образующие олигонуклеотиды.
Альтернативно, диагностические или терапевтические средства, связанные с биспецифическими антителами по настоящему изобретению, могут представлять собой систему инкапсуляции, такую как липосома или мицелла, содержащая терапевтическое средство, такое как лекарственное средство, нуклеиновая кислота (например, антисмысловая нуклеиновая кислота) или другое терапевтическое средство, предпочтительно, защищенное от прямого воздействия кровотока. Способы получения липосом, присоединенных к антителам, хорошо известны специалистам в этой области. См., например, патент США № 4957735; и Connor, et al., Pharm. Ther. 28:341-365 (1985).
В некоторых вариантах осуществления способы, представленные в настоящем описании, дополнительно включают стадию лечения индивидуума с использованием дополнительной формы терапии. В некоторых вариантах осуществления дополнительная форма терапии является дополнительной противоопухолевой терапией, включая, в качестве неограничивающих примеров, химиотерапию, лучевую терапию, хирургическое вмешательство, гормональную терапию и/или дополнительную иммунотерапию.
Настоящее изобретение дополнительно включает введение молекулы по изобретению в комбинации с другими терапевтическими средствами, известными специалистам в этой области для лечения или профилактики злокачественного новообразования, включая, в качестве неограничивающих примеров, стандартные и экспериментальные химиотерапевтические средства, биологические терапевтические средства, иммунотерапевтические средства, лучевую терапию или хирургическое вмешательство. В некоторых аспектах молекулы по изобретению можно вводить в комбинации с терапевтически или профилактически эффективным количеством одного или более средств, терапевтических антител или других средств, известных специалистам в этой области для лечения и/или профилактики злокачественного новообразования.
Таким образом, способы лечения злокачественного новообразования включают введение нуждающемуся в этом индивидууму эффективного количества антитела или биспецифического антитела по настоящему изобретению в комбинации с химиотерапевтическим средством. Такое комбинированное лечение можно вводить раздельно, последовательно, или одновременно. Подходящие химиотерапевтические средства включают, в качестве неограничивающих примеров, по меньшей мере одно дополнительное средство, такое как бевацизумаб, цетуксимаб, сиролимус, панитумумаб, 5-фторурацил(5-FU), капецитабин, тивозаниб, иринотекан, оксалиплатин, цисплатин, трифлуридин, типирацил, лейковорин, гемцитабин и/или эрлотиниба гидрохлорид.
Дозируемые количества и частоты введения, представленные в настоящем описании, включены в термины "терапевтически эффективный" и "профилактически эффективный". Доза и частота дополнительно варьируются в зависимости от факторов, специфических для каждого пациента, в зависимости от специфических вводимых терапевтических или профилактических средств, тяжести и типа злокачественных новообразований, пути введение, а также возраста, массы тела, ответа и медицинского анамнеза пациента. Специалист в этой области может выбирать подходящие схемы лечения, рассматривая такие факторы и следуя, например, дозам, описанным в литературе и рекомендуемым в Настольном справочнике врача (56-ое издание, 2002 год).
Антитела или биспецифические антитела по настоящему изобретению могут находиться в фармацевтической композиции для введения, составляемой так, чтобы подходить для выбранного способа введения, и содержащей фармацевтически приемлемый дилюент или эксципиенты, такие как буферы, поверхностно-активные вещества, консерванты, солюбилизаторы, изотонические средства, стабилизирующие средства, носители и т.п. В Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton Pa., 18th ed., 1995 представлено руководство по способам составления, как правило, известным практикующим специалистам.
Эти фармацевтические композиции можно вводить любыми способами, известными в этой области, с помощью которых, как правило, достигают назначенной цели лечения злокачественного новообразования. Путь введения может являться парентеральным, определенным в настоящем описании как относящийся к способам введения, включающим, в качестве неограничивающих примеров, чреспищеводную, внутриопухолевую, трансколоноскопическую, чрескожную, внутривенную, внутримышечную, интраперитонеальную, подкожную и внутрисуставную инъекцию и инфузию. Способ введения и дозирования молекул по изобретению (например, антител против GUCY2c, биспецифического антитела против CD3-GUCY2c, фармацевтической композиции) зависит от типа заболевания, подлежащего лечению, при необходимости, его стадии, антигена, подлежащего контролю, типа сопутствующего лечения, если оно есть, частоты лечения, природы желаемого эффекта, а также массы тела, возраста, состояния здоровья, питания и пола пациента. Таким образом, конкретный используемый режим дозирования может варьироваться в широких пределах и, таким образом, может отклоняться от режимов дозирования, приведенных в настоящем описании.
Для введения можно использовать различные составы антител по настоящему изобретению (например, антител против GUCY2c или биспецифических антител против CD3-GUCY2c). В некоторых вариантах осуществления антитела (например, антитела против GUCY2c или биспецифическое антитело против CD3-GUCY2c) можно вводить в отдельности. В некоторых вариантах осуществления антитело (например, антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c) и фармацевтически приемлемый эксципиент могут находиться в различных составах. Фармацевтически приемлемые эксципиенты известны в этой области и являются относительно инертными веществами, облегчающими введение фармакологически эффективного вещества. Например, эксципиент может придавать форму или консистенцию или действовать в качестве дилюента. Подходящие эксципиенты включают, в качестве неограничивающих примеров, стабилизирующие средства, увлажнители и эмульгаторы, соли для варьирования осмолярности, инкапсулирующие средства, буферы и усилители проникновения через кожу. Эксципиенты, а также составы для парентерального и непарентерального введения лекарственного средства, приведены в Remington, The Science and Practice of Pharmacy 21st Ed. Mack Publishing, 2005.
В некоторых вариантах осуществления эти средства составляют для введения посредством инъекции (например, интраперитонеально, внутривенно, подкожно, внутримышечно и т.д.). Таким образом, эти средства можно комбинировать с фармацевтически приемлемыми носителями, такими как физиологический раствор, раствор Рингера, раствор декстрозы и т.п. Конкретный режим дозирования, т.е. доза, время и повторение, будут зависеть от конкретного индивидуума и медицинского анамнеза индивидуума.
Антитела (например, антитела против GUCY2c или биспецифические антитела против CD3-GUCY2c), представленные в настоящем описании, можно вводить любым подходящим способом, включая инъекцию (например, интраперитонеальную, внутривенную, подкожную, внутримышечную и т.д.). Антитело, например, моноклональное антитело или биспецифическое антитело, также вводят посредством ингаляции, как представлено в настоящем описании. Как правило, в случае введения антитела по настоящему изобретению (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c) доза зависит от организма, подвергаемого лечению и конкретного способа введения. В одном из вариантов осуществления диапазон доз антитела по настоящему изобретению будет составлять от приблизительно 0,001 мкг/кг массы тела до приблизительно 20000 мкг/кг массы тела. Термин "масса тела" используют, когда пациента подвергают лечению. Если обрабатывают выделенные клетки, термин "масса тела" в рамках изобретения относится к "общей массе клеток". Термин "общая масса тела" можно использовать в отношении обработки выделенной клетки и лечения пациента. Все концентрации и уровни обработки, выраженные в настоящей заявке как "масса тела" или просто "кг", также считают охватывающими аналогичные концентрации "общей массы клеток" и "общей массы тела". Однако специалистам в этой области будет понятна полезность различных диапазонов доз, например, от 0,01 мкг/кг массы тела до 20000 мкг/кг массы тела, от 0,02 мкг/кг массы тела до 15000 мкг/кг массы тела, от 0,03 мкг/кг массы тела до 10000 мкг/кг массы тела, от 0,04 мкг/кг массы тела до 5000 мкг/кг массы тела, от 0,05 мкг/кг массы тела до 2500 мкг/кг массы тела, от 0,06 мкг/кг массы тела до 1000 мкг/кг массы тела, от 0,07 мкг/кг массы тела до 500 мкг/кг массы тела, от 0,08 мкг/кг массы тела до 400 мкг/кг массы тела, от 0,09 мкг/кг массы тела до 200 мкг/кг массы тела или от 0,1 мкг/кг массы тела до 100 мкг/кг массы тела. Кроме того, специалистам будет понятно, что можно использовать различные уровни доз, например, 0,0001 мкг/кг, 0,0002 мкг/кг, 0,0003 мкг/кг, 0,0004 мкг/кг, 0,005 мкг/кг, 0,0007 мкг/кг, 0,001 мкг/кг, 0,1 мкг/кг, 1,0 мкг/кг, 1,5 мкг/кг, 2,0 мкг/кг, 5,0 мкг/кг, 10,0 мкг/кг, 15,0 мкг/кг, 30,0 мкг/кг, 50 мкг/кг, 75 мкг/кг, 80 мкг/кг, 90 мкг/кг, 100 мкг/кг, 120 мкг/кг, 140 мкг/кг, 150 мкг/кг, 160 мкг/кг, 180 мкг/кг, 200 мкг/кг, 225 мкг/кг, 250 мкг/кг, 275 мкг/кг, 300 мкг/кг, 325 мкг/кг, 350 мкг/кг, 375 мкг/кг, 400 мкг/кг, 450 мкг/кг, 500 мкг/кг, 550 мкг/кг, 600 мкг/кг, 700 мкг/кг, 750 мкг/кг, 800 мкг/кг, 900 мкг/кг, 1 мкг/кг, 5 мкг/кг, 10 мкг/кг, 12 мкг/кг, 15 мг/кг, 20 мг/кг и/или 30 мг/кг. Все эти дозы являются примерами, и ожидают, что в настоящем изобретении можно использовать любую дозу между этими значениями. Для антитела по настоящему изобретению можно использовать любые из указанных выше диапазонов доз или уровней доз. В случае повторных введений в течение нескольких дней или более, в зависимости от состояния, лечение продолжают до возникновения желаемой супрессии симптомов или до достижения достаточных терапевтических уровней, например, для ингибирования или задержки роста/прогрессирования опухоли или метастазирования злокачественных клеток.
Также можно использовать другие режимы дозирования в зависимости от профиля фармакокинетического распада, который хочет достигнуть практикующий специалист. В одном из вариантов осуществления антитело по настоящему изобретению вводят в усиленной начальной дозе, с последующей более высокой и/или непрерывной, по существу постоянной дозой. В некоторых вариантах осуществления предусмотрено введение от одного до четырех раз в неделю. В других вариантах осуществления предусмотрено введение раз в месяц, или раз в два месяца, или раз в три месяца. Прогресс этой терапии легко подвергать мониторингу с помощью общепринятых способов и анализов. Схему дозирования (включающую антитело (например, антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c)) можно варьировать с течением времени.
В целях по настоящему изобретению, подходящая доза антитела (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c) будет зависеть от антитела (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c) или его используемых композиций, типа и тяжести симптомов, подвергаемых лечению, если средство вводят в терапевтических целях, предшествующей терапии, клинического анамнеза пациента и ответа на средство, скорости выведения вводимого средства у пациента и решения лечащего врача. Как правило, клиницист будет вводить антитело (например, антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c) до достижения дозы, позволяющей достигать желаемого результата. Дозу и/или частоту можно варьировать в течение лечения. При определении дозы, как правило, будут учитывать эмпирические факторы, такие как время полужизни. Например, антитела, совместимые с иммунной системой человека, такие как гуманизированные антитела или полностью человеческие антитела, можно использовать для пролонгирования времени полужизни антитела и для предотвращения атаки антитела иммунной системой организма-хозяина. Частоту введения можно определять и корректировать в течение терапии, и она, как правило, но не обязательно, основана на лечении, и/или супрессии, и/или улучшении, и/или задержке симптомов, например, ингибировании или задержке роста опухоли и т.д. Альтернативно, подходящими могут являться составы антител с замедленным высвобождением (например, антител GUCY2c или биспецифического антитела против CD3-GUCY2c). В этой области известны различные составы и устройства для достижения замедленного высвобождения.
В одном из вариантов осуществления дозы антитела (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c) можно определять эмпирически у индивидуумов, подвергнутых одному или более введениям антитела (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c). Индивидуумам вводят увеличивающиеся дозы антитела (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c). Для оценки эффективности можно отслеживать показатель заболевания.
В некоторых вариантах осуществления введение антитела (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c) приводит по меньшей мере к одному эффекту, выбранному из группы, состоящей из ингибирования роста опухоли, регрессирования опухоли, снижения размера опухоли, снижения количества опухолевых клеток, задержки роста опухоли, абскопального эффекта, ингибирования метастазирования опухоли, уменьшения метастатических очагов с течением времени, снижения использования химиотерапевтических или цитотоксических средств, уменьшения опухолевой массы, повышения выживаемости без прогрессирования, повышения общей выживаемости, полного ответа, частичного ответа и стабильного заболевания.
Введение антитела (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c) в соответствии со способом по настоящему изобретению может являться непрерывным или дробным, в зависимости, например, от физиологического состояния реципиента, является ли цель введения терапевтической или профилактической, и других факторов, известных специалистам в этой области. Введение антитела (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c) может являться, по существу, непрерывным в течение заранее выбранного периода времени или может представлять собой серию дробных доз.
В некоторых вариантах осуществления могут присутствовать несколько антител (например, антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c). Могут присутствовать по меньшей мере одно, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять или более разных антител (например, антител против GUCY2c или биспецифических антител против CD3-GUCY2c). Как правило, антитела (например, антитела против GUCY2c или биспецифические антитела против CD3-GUCY2c) могут иметь комплементарные активности, не влияющие друг на друга негативно. Например, можно использовать одно или более из следующих антител: первое антитело против GUCY2c или CD3, направленное против одного эпитопа на GUCY2c или CD3, и второе антитело против GUCY2c или CD3, направленное против другого эпитопа на GUCY2c или CD3.
Терапевтические составы антитела (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c), используемые по настоящему изобретению, получают для хранения посредством смешивания антитела, имеющего желаемую степень чистоты, с необязательными фармацевтически приемлемыми носителями, эксципиентами или стабилизаторами (Remington, The Science and Practice of Pharmacy 21st Ed. Mack Publishing, 2005), в форме лиофилизированных составов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в используемых дозах и концентрациях, и могут содержать буферы, такие как фосфат, цитрат и другие органические кислоты; соли, такие как хлорид натрия; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол и m-крезол); низкомолекулярные (менее приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие средства, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексные соединения с металлами (например, комплексы Zn-белок); и/или неионные поверхностно-активные средства, такие как TWEENTM, PLURONICTM или полиэтиленгликоль (PEG).
Липосомы, содержащие антитело (например, антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c), получают известными в этой области способами, как описано в Epstein, et al., Proc. Natl. Acad. Sci. USA 82:3688, 1985; Hwang, et al., Proc. Natl Acad. Sci. USA 77:4030, 1980; и патентах США №№ 4485045 и 4544545. Липосомы с повышенным временем циркуляции описаны в патенте США № 5013556. Особенно подходящие липосомы можно получать способом обращенно-фазового выпаривания с использованием липидной композиции, содержащей фосфатидилхолин, холестерин и PEG-дериватизированный фосфатидилэтаноламин (PEG-PE). Липосомы экструдируют через фильтры с определенным размером пор для получения липосом с желаемым диаметром.
Активные ингредиенты также можно заключать в микрокапсулы, полученные, например, способами коацервации или посредством интерфазной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и микрокапсулы из полиметилметакрилата, соответственно, в коллоидных системах доставки лекарственного средства (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсиях. Такие способы описаны в Remington, The Science and Practice of Pharmacy 21st Ed. Mack Publishing, 2005.
Можно получать препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих антитело, где матрицы находятся в форме профилированных изделий, например, пленок или микрокапсул. Примеры матриц с замедленным высвобождением включают полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат) или поливиниловый спирт), полилактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и 7-этил-L-глутамата, недеградируемый этилен-винилацетат, деградируемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOTTM (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и лейпролид ацетат), ацетат избутират сахарозы и поли-D-(-)-3-гидроксимасляная кислота.
Составы, предназначенные для использования для введения in vivo, должны быть стерильными. Этого легко достигать, например, посредством фильтрации через мембраны для стерилизации фильтрацией. Композиции терапевтического антитела (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c), как правило, помещают в контейнер, имеющий стерильное входное отверстие, например, мешок для внутривенного раствора или флакон с пробкой, прокалываемой иглой для подкожных инъекций.
Композиции по настоящему изобретению могут находиться в стандартных лекарственных формах, таких как таблетки, пилюли, капсулы, порошки, гранулы, растворы или суспензии или суппозитории, для перорального, парентерального или ректального введения или введения посредством ингаляции или инсуффляции.
Для получения твердых композиций, таких как таблетки, основной активный ингредиент смешивают с фармацевтическим носителем, например, общепринятыми ингредиентами для таблетирования, таких как кукурузный крахмал, лактоза, сахароза, сорбит, тальк, стеариновая кислота, стеарат магния, дифосфат кальция или камеди, и другими фармацевтическими дилюентами, например, водой, для получения твердой композиции до придания ей лекарственной формы, содержащей однородную смесь соединения по настоящему изобретению или его нетоксичную фармацевтически приемлемую соль. Упоминание этих композиций до придания им лекарственной формы как однородных означает, что активный ингредиент равномерно диспергирован по всей композиции таким образом, что композицию легко можно разделять на в равной степени эффективные стандартные лекарственные формы, такие как таблетки, пилюли и капсулы. Затем эту твердую композицию до придания ей лекарственной формы разделяют на стандартные лекарственные формы описанного выше типа, содержащие от 0,1 до приблизительно 500 мг активного ингредиента по настоящему изобретению. Таблетки или пилюли новой композиции можно покрывать или иначе экстемпорально получать для получения лекарственной формы, обеспечивающей преимущество пролонгированного действия. Например, таблетка или пилюля может содержать внутренний компонент дозы и внешний компонент дозы, последний из которых находится в виде оболочки вокруг первого из них. Два компонента можно разделять кишечнорастворимым слоем, предназначенным для устойчивости к распаду в желудке и позволяющим внутреннему компоненту попадать в двенадцатиперстную кишку или иметь замедленное высвобождение. Для таких кишечнорастворимых слоев или покрытий можно использовать различные материалы, включающие ряд полимерных кислот и смесей полимерных кислот с такими материалами, как шеллак, цетиловый спирт и ацетат целлюлозы.
Подходящие поверхностно-активные средства включают, в частности, неионные средства, такие как полиоксиэтиленсорбитаны (например, TweenTM 20, 40, 60, 80 или 85) и другие сорбитаны (например, SpanTM 20, 40, 60, 80 или 85). Композиции с поверхностно-активным средством, как правило, будут содержать от 0,05 до 5% поверхностно-активного средства и могут содержать от 0,1 до 2,5%. Следует понимать, что, при необходимости, можно добавлять другие ингредиенты, например, маннит или другие фармацевтически приемлемые носители.
Подходящие эмульсии можно получать с использованием коммерчески доступных жировых эмульсий, таких как IntralipidTM, LiposynTM, InfonutrolTM, LipofundinTM и LipiphysanTM. Активный ингредиент можно растворять в заранее смешанной композиции эмульсии или, альтернативно, его можно растворять в масле (например, соевом масле, сафлоровом масле, хлопковом масле, сезамовом масле, кукурузном масле или миндальном масле) и эмульсии, полученной после смешивания с фосфолипидом (например, фосфолипидами яйца, фосфолипидами сои или лецитином сои) и водой. Следует понимать, что можно добавлять другие ингредиенты, например, глицерин или глюкозу, для корректировки тоничности эмульсии. Подходящие эмульсии, как правило, будут содержать до 20% масла, например, от 5 до 20%. Жировая эмульсия может содержать жировые капли от 0,1 до 1,0 мкм, в частности, от 0,1 до 0,5 мкм, и иметь pH в диапазоне от 5,5 до 8,0.
Композиции эмульсий могут являться композициями, полученными посредством смешивания антитела (например, антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c) с IntralipidTM или его компонентами (соевым маслом, фосфолипидами яйца, глицерином и водой).
Композиции для ингаляции или инсуффляции включают растворы и суспензии в фармацевтически приемлемых водных или органических растворителях или их смесях и порошки. Жидкие или твердые композиции могут содержать подходящие фармацевтически приемлемые эксципиенты, как описано выше. В некоторых вариантах осуществления композиции вводят пероральным или назальным респираторным путем для локального или системного эффекта. Композиции в предпочтительно стерильных фармацевтически приемлемых растворителях можно небулизировать с помощью газов. Небулизируемые растворы можно вдыхать непосредственно из небулизирующего устройства, или небулизирующее устройство можно прикреплять к маске для лица, тенту или дыхательному аппарату с положительным перемежающимся давлением. Композиции в виде раствора, суспензии или порошка можно вводить, предпочтительно, перорально или назально с помощью устройства, посредством которого состав вводят соответствующим образом.
Композиции в объеме изобретения включают все композиции, где антитело (антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c) присутствует в количестве, являющемся эффективным для достижения желаемого медицинского эффекта при лечении злокачественного новообразования. Хотя индивидуальные потребности могут варьироваться от одного пациента к другому, определение оптимальных диапазонов эффективных количеств всех компонентов входит в объем навыков специалиста в этой области.
Примеры таких композиций, а также способов составления, также описаны выше и ниже. В некоторых вариантах осуществления композиция содержит одно или более антител (например, антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c). В некоторых вариантах осуществления антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c является антителом человека, гуманизированным антителом или химерным антителом. В некоторых вариантах осуществления антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c содержит константную область, способную запускать желаемый иммунный ответ, такой как антитело-опосредованный лизис или ADCC. В других вариантах осуществления антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c содержит константную область, не запускающую нежелательный иммунный ответ, такой как антитело-опосредованный лизис или ADCC.
Следует понимать, что композиции могут содержать несколько антител (например, антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c), например, смесь антител против GUCY2c или биспецифических антител против CD3-GUCY2c, распознающих разные эпитопы GUCY2c или CD3 и GUCY2c. Другие примеры композиций содержат несколько антител против GUCY2c или биспецифических антител против CD3-GUCY2c, распознающих одни и те же эпитопы, или разные типы антител против GUCY2c биспецифических антител против CD3-GUCY2c, или антитела, связывающиеся с разными эпитопами GUCY2c (например, GUCY2cA человека) или CD3 и GUCY2c (CD3 и GUCY2c человека).
В некоторых вариантах осуществления антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c можно вводить в комбинации с введением одного или более дополнительных терапевтических средств. Они включают, в качестве неограничивающих примеров, введение биотерапевтического средства и/или химиотерапевтического средства, такого как, в качестве неограничивающих примеров, вакцина, терапию CAR-T-клетками, лучевую терапию, цитокиновую терапию, биспецифическое антитело против CD3, ингибитор других иммуносупрессорных путей, ингибитор ангиогенеза, активатор T-клеток, ингибитор метаболического пути, ингибитор mTOR, ингибитор пути аденозина, ингибитор тирозинкиназ включая, в качестве неограничивающих примеров, Inlyta, ингибиторы ALK и сунитиниб, ингибитор BRAF, эпигенетический модификатор, ингибитор IDO1, ингибитор JAK, ингибитор STAT, ингибитор циклин-зависимой киназы, биотерапевтическое средство (включая, в качестве неограничивающих примеров, антитела против VEGF, VEGFR, EGFR, Her2/neu, других рецепторов факторов роста, CD40, CD-40L, CTLA-4, OX-40, 4-1BB, TIGIT и ICOS), иммуногенное средство (например, аттенуированные злокачественные клетки, опухолевые антигены, антигенпрезентирующие клетки, такие как дендритные клетки с введенным опухолевым антигеном или нуклеиновыми кислотми, иммуностимулирующие цитокины (например, ИЛ-2, ИФНα2, ГМ-КСФ) и клетки, трансфицированные с использованием генов, кодирующих иммуностимулирующие цитокины, такие как, в качестве неограничивающих примеров, ГМ-КСФ).
Примеры биотерапевтических средств включают терапевтические антитела, иммуномодулирующие средства и терапевтические иммунные клетки.
Терапевтические антитела могут иметь специфичность в отношении различных антигенов. Например, терапевтические антитела могут быть направлены против опухолеассоциированного антигена, таким образом, что связывание антитела с антигеном способствует гибели клетки, экспрессирующей антиген. В другом примере терапевтические антитела могут быть направлены против антигена (например, PD-1) на иммунной клетке, таким образом, что связывание антитела предотвращает отрицательную регуляцию активности клетки, экспрессирующей антиген (и, таким образом, способствует активности клетки, экспрессирующей антиген). В некоторых случаях терапевтическое антитело может функционировать посредством множества разных механизмов (например, оно может i) способствовать гибели клетки, экспрессирующей антиген, а также ii) предотвращать индуцирование антигеном отрицательной регуляции активности иммунных клеток, контактирующих с клеткой, экспрессирующей антиген).
Терапевтические антитела могут быть направлены против, например, антигенов, указанных ниже. Для некоторых антигенов примеры антител против антигена также включены ниже (в прямых/круглых скобках после антигена). Следующие антигены также можно обозначать в настоящем описании как "антигены-мишени" или т.п. Антигены-мишени для терапевтических антител в настоящем описании включают, например: 4-1BB (например, утомилумаб); 5T4; A33; альфа-фолатный рецептор 1 (например, мирветуксимаб соравтанзин); Alk-1; BCMA [например, PF-06863135 (см. патент США № 9969809)]; BTN1A1 (например, см. WO2018222689); CA-125 (например, абаговомаб); карбоангидразу IX; CCR2; CCR4 (например, могамулизумаб); CCR5 (например, леронлимаб); CCR8; CD3 [например, блинатумомаб (биспецифическое антитело против CD3/CD19), PF-06671008 (биспецифическое антитело против CD3/P-кадгерина), PF-06863135 (биспецифическое антитело против CD3/BCMA), CD19 (например, блинатумомаб, MOR208); CD20 (например ибритумомаб тиуксетан, обинутузумаб, офатумумаб, ритуксимаб, ублитуксимаб); CD22 (инотузумаб озогамицин, моксетумомаб пасудотокс); CD25; CD28; CD30 (например, брентуксимаб ведотин); CD33 (например, гемтузумаб озогамицин); CD38 (например, даратумумаб, исатуксимаб), CD40; CD-40L; CD44v6; CD47; CD52 (например, алемтузумаб); CD63; CD79 (например, полатузумаб ведотин); CD80; CD123; CD276/B7-H3 (например, омбуртамаб); CDH17; CEA; ClhCG; CTLA-4 (например, ипилимумаб, тремелимумаб), CXCR4; десмоглеин 4; DLL3 (например, ровалпитузумаб тесирин); DLL4; E-кадгерин; EDA; EDB; EFNA4; EGFR (например, цетуксимаб, депатуксизумаб мафодотин, нецитумумаб, панитумумаб); EGFRvIII; эндосиалин; EpCAM (например, опортузумаб монатокс); FAP; фетальный ацетилхолиновый рецептор; FLT3 (например, см. WO2018/220584); GD2 (например, динутуксимаб, 3F8); GD3; GITR; GloboH; GM1; GM2; GUCY2C (например, PF-07062119); HER2/neu [например, маргетуксимаб, пертузумаб, трастузумаб; ado-трастузумаб эмтанзин, трастузумаб дуокармазин, PF-06804103 (см. US8828401)]; HER3; HER4; ICOS; IL-10; ITG-AvB6; LAG-3 (например, релатлимаб); антиген Льюиса Y; LG; Ly-6; M-CSF [например PD-0360324 (см. патент США № 7326414)]; MCSP; мезотелин; MUC1; MUC2; MUC3; MUC4; MUC5AC; MUC5B; MUC7; MUC16; Notch1; Notch3; нектин-4 (например, энфортумаб ведотин); OX40 [например, PF-04518600 (см. патент США № 7960515)]; P-кадгерин [например PF-06671008 (см. WO2016/001810)]; PCDHB2; PD-1 [например, BCD-100, камрелизумаб, цемиплимаб, генолимзумаб (CBT-501), MEDI0680, ниволумаб, пембролизумаб, RN888 (см. WO2016/092419), синтилимаб, спартализумаб, STI-A1110, тислелизумаб, TSR-042]; PD-L1 (например, атезолизумаб, дурвалумаб, BMS-936559 (MDX-1105) или LY3300054); PDGFRA (например, оларатумаб); антиген плазматических клеток; поли-SA; PSCA; PSMA; PTK7 [например, PF-06647020 (см. патент США № 9409995)]; Ror1; SAS; SCRx6; SLAMF7 (например, элотузумаб); SHH; SIRPa (например, ED9, Effi-DEM); STEAP; TGF-бета; TIGIT; TIM-3; TMPRSS3; предшественник ФНО-альфа; TROP-2 (например, сацитузумаб говитекан); TSPAN8; VEGF (например, бевацизумаб, бролуцизумаб); VEGFR1 (например, ранибизумаб); VEGFR2 (например, рамуцирумаб, ранибизумаб); Wue-1.
Терапевтические антитела могут иметь любой подходящий формат. Например, терапевтические антитела могут иметь любой формат, описанный где-либо в настоящем описании. В некоторых вариантах осуществления терапевтическое антитело может являться "голым" антителом. В некоторых вариантах осуществления терапевтическое антитело можно связывать с лекарственным средством/средством (также известным как "конъюгат антитело-лекарственное средство" (ADC)). В некоторых вариантах осуществления терапевтическое антитело против конкретного антигена можно включать в мультиспецифическое антитело (например, биспецифическое антитело).
В некоторых вариантах осуществления антитело против антигена можно конъюгировать с лекарственным средством/средством. Связанные молекулы антитело-лекарственное средство также обозначают как "конъюгаты антитело-лекарственное средство" (ADC). Лекарственные средства/средства можно соединять с антителом прямо или косвенно через линкер. Чаще всего, токсические лекарственные средства соединяют с антителом таким образом, что связывание ADC с соответствующим антигеном способствует уничтожению клеток, экспрессирующих антиген. Например, ADC, которые соединяют с токсическими лекарственными средствами, особенно подходят для таргетинга опухолеассоциированных антигенов для стимуляции уничтожения опухолевых клеток, экспрессирующих опухолеассоциированные антигены. В других вариантах осуществления средства, которые можно соединять с антителом, могут являться, например, иммуномодулирующим средством (например, для модуляции активности иммунных клеток вблизи ADC), визуализирующим средством (например, для облегчения визуализации ADC у индивидуума или в биологическом образце индивидуума) или средством для повышения времени полужизни в сыворотке или биологической активности антитела.
Способы конъюгации цитотоксического средства или других терапевтических средств с антителами описаны в различных публикациях. Например, химическую модификацию можно осуществлять в антителах через амины боковой цепи лизина или сульфгидрильные группы цистеина, активированные посредством восстановления межцепочечных дисульфидных связей, для того, чтобы произошла реакция конъюгации. См., например, Tanaka et al., FEBS Letters 579:2092-2096, 2005, и Gentle et al., Bioconjugate Chem. 15:658-663, 2004. Также описаны реакционноспособные остатки цистеина, сконструированные в конкретных участках антител для специфической конъюгации лекарственного средства с определенной стехиометрией. См., например, Junutula et al., Nature Biotechnology, 26:925-932, 2008. Конъюгация с использованием метки, содержащей глутамин-донор ацила, или эндогенного глутамина, сделанного реакционноспособным (т.е. со способностью образовывать ковалентную связь в качестве донора ацила) посредством конструирования полипептида в присутствии трансглутаминазы и амина (например, цитотоксического средства, содержащего или прикрепленного к реакционноспособному амину), также описана в международных заявках WO2012/059882 и WO2015015448. В некоторых вариантах осуществления ADC может иметь любой из признаков или характеристик ADC, представленных WO2016166629, таким образом, включенной в качестве ссылки для всех целей.
Лекарственные средства/средства, которые можно соединять с антителом в формате ADC, могут включать, например, цитотоксические средства, иммуномодулирующие средства, визуализирующие средства, терапевтические белки, биополимеры или олигонуклеотиды.
Примеры цитотоксических средств, которые можно включать в ADC, включают антрациклин, ауристатин, доластатин, комбретастатин, дуокармицин, димер пирролобензодиазепина, димер индолино-бензодиазепинп, ендиин, гелданамицин, майтанзин, пуромицин, таксан, алкалоид барвинка, камптотецин, тубулизин, гемиастерлин, сплайсостатин, пладиенолид и их стереоизомеры, изостереомеры, аналоги или производные.
Примеры иммуномодулирующих средств, которые можно включать в ADC, включают ганцикловир, этанерцепт, такролимус, сиролимус, воклоспорин, циклоспорин, рапамицин, циклофосфамид, азатиоприн, микофенолата мофетил, метотрексат, глюкокортикоид и его аналоги, цитокины, факторы роста стволовых клеток, лимфотоксины, фактор некроза опухоли (ФНО), гемопоэтические факторы, интерлейкины (например, интерлейкин-1 (ИЛ-1), ИЛ-2, ИЛ-3, ИЛ-6, ИЛ-10, ИЛ-12, ИЛ-18 и ИЛ-21), колониестимулирующие факторы (например, гранулоцитарный колониестимулирующий фактор (Г-КСФ) и гранулоцитарно-макрофагальный колониестимулирующий фактор (ГМ-КСФ)), интерфероны (например, интерферон-альфа, -бета и гамма.), фактор роста стволовых клеток, обозначенный как "фактор S 1", эритропоэтин и тромбопоэтин или их комбинацию.
Примеры визуализирующих средств, которые можно включать в ADC, включают флуоресцеин, родамин, лантаноид-активированные фосфоры и их производные или радиоактивный изотоп, связанный с хелатором. Неограничивающие примеры флуорофоров включают флуоресцеинизотиоцианат (FITC) (например, 5-FITC), флуоресцеин амидит (FAM) (например, 5-FAM), эозин, карбоксифлуоресцеин, эритрозин, Alexa FluorTM (например, Alexa 350, 405, 430, 488, 500, 514, 532, 546, 555, 568, 594, 610, 633, 647, 660, 680, 700 или 750), карбокситетраметилродамин (TAMRA) (например, 5,-TAMRA), тетраметилродамин (TMR) и сульфородамин (SR) (например, SR101). Неограничивающие примеры хелаторов включают 1,4,7,10-тетраазациклододекан-N, N',N'',N'''-тетрауксуснкю кислоту (DOTA), 1,4,7-триазациклононан-1,4,7-триуксусную кислоту (NOTA), 1,4,7-триазациклононан, 1-глутаровая кислота-4,7-уксусную кислоту (дефероксамин), диэтилентриаминпентауксусную кислоту (DTPA) и 1,2-бис(o-аминофенокси)этан-N, N,N',N'-тетрауксусную кислоту (BAPTA).
Примеры терапевтических белков, которые можно включать в ADC, включают токсин, гормон, фермент и фактор роста.
Примеры биосовместимых полимеров, которые можно включать в ADC, включают водорастворимые полимеры, такие как полиэтиленгликоль (PEG) или его производные и цвиттер-ион-содержащие биосовместимые полимеры (например, фосфорилхолин-содержащий полимер).
Примеры биосовместимых полимеров, которые можно включать в ADC, включают антисмысловые олигонуклеотиды.
В некоторых вариантах осуществления антитело против антигена, представленное в настоящем описании, можно включать в молекулу биспецифического антитела. Биспецифические антитела являются моноклональными антителами, имеющими специфичность связывания в отношении по меньшей мере двух разных антигенов.
Иммуномодулирующие средства включают различные типы молекул, которые могут стимулировать иммунный ответ у индивидуума, такие как агонисты паттерн-распознающего рецептора (PRR), иммуностимулирующие цитокины и противоопухолевые вакцины.
Паттерн-распознающие рецепторы (PRR) являются рецепторами, экспрессирующимися клетками иммунной системы и распознающими различные молекулы, ассоциированные с патогенами и/или повреждением или гибелью клеток. PRR участвуют во врожденном иммунном ответе и адаптивном иммунном ответе. Агонисты PRR можно использовать для стимуляции иммунного ответа в индивидуума. Существует множество классов молекул PRR, включая Толл-подобные рецепторы (TLR), RIG-I-подобные рецепторы (RLR), рецепторы, подобные нуклеотид-связывающему домену олигомеризации (NOD) (NLRs), лектиновые рецепторы C-типа (CLR) и белок-стимулятор интерфероновых генов (STING).
Термины "TLR" и "Толл-подобный рецептор" относятся к любому Толл-подобному рецептору. Толл-подобные рецепторы являются рецепторами, участвующими в активации иммунных ответов. TLR распознают, например, патоген-ассоциированные молекулярные паттерны (PAMP), экспрессирующиеся в микроорганизмах, а также эндогенные молекулярные паттерны, ассоциированные с повреждением (DAMP), высвобождающиеся из умерших или умирающих клеток.
Молекулы, активирующие TLR (и, таким образом, активирующие иммунные ответы), обозначают в настоящем описании как "агонисты TLR". Агонисты TLR могут включать, например, низкомолекулярные соединения (например, органическую молекулу, имеющую молекулярную массу до приблизительно 1000 Да), а также крупные молекулы (например, олигонуклеотиды и белки). Некоторые агонисты TLR являются специфическими для одного типа TLR (например, TLR3 или TLR9), в то время как некоторые агонисты TLR активируют два или более типа TLR (например, и TLR7, и TLR8).
Примеры агонистов TLR, представленные в настоящем описании, включают агонисты TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8 и TLR9.
Примеры низкомолекулярных агонистов TLR включают описанные, например, в патентах США №№ 4689338; 4929624; 5266575; 5268376; 5346905; 5352784; 5389640; 5446153; 5482936; 5756747; 6110929; 6194425; 6331539; 6376669; 6451810; 6525064; 6541485; 6545016; 6545017; 6573273; 6656938; 6660735; 6660747; 6664260; 6664264; 6664265; 6667312; 6670372; 6677347; 6677348; 6677349; 6683088; 6756382; 6797718; 6818650 и 77091214; патентных публикациях США №№ 2004/0091491, 2004/0176367 и 2006/0100229; и международных патентных публикациях №№ WO 2005/18551, WO 2005/18556, WO 2005/20999, WO 2005/032484, WO 2005/048933, WO 2005/048945, WO 2005/051317, WO 2005/051324, WO 2005/066169, WO 2005/066170, WO 2005/066172, WO 2005/076783, WO 2005/079195, WO 2005/094531, WO 2005/123079, WO 2005/123080, WO 2006/009826, WO 2006/009832, WO 2006/026760, WO 2006/028451, WO 2006/028545, WO 2006/028962, WO 2006/029115, WO 2006/038923, WO 2006/065280, WO 2006/074003, WO 2006/083440, WO 2006/086449, WO 2006/091394, WO 2006/086633, WO 2006/086634, WO 2006/091567, WO 2006/091568, WO 2006/091647, WO 2006/093514 и WO 2006/098852.
Дополнительные примеры низкомолекулярных агонистов TLR включают некоторые производные пурина (такие как описываемые в патентах США №№ 6376501 и 6028076), некоторые производные амида имидазохинолина (такие как описываемые в патенте США № 6069149), некоторые производные имидазопиридина (такие как описываемые в патенте США № 6518265), некоторые производные бензимидазола (такие как описываемые в патенте США № 6387938), некоторые производные 4-аминопиримидина, слитые с пятичленным азот-содержащим гетероциклическим ядром (такие как производные аденина, описанные в патентах США №№ 6376501, 6028076 и 6329381 и в WO 02/08905) и некоторые производные 3-.бета.-D-рибофуранозилтиазоло[4,5-d]пиримидина (такие как описываемые в патентной публикации США № 2003/0199461) и некоторые низкомолекулярные иммунопотенциирующие соединения, такие как описываемые, например, в патентной публикации США № 2005/0136065.
Примеры крупных агонистов TLR включают олигонуклеотидные последовательности. Некоторые агонисты TLR-олигонуклеотидные последовательности содержат динуклеотиды цитозин-гуанин (CpG) и описаны, например, в патентах США №№ 6194388, 6207646, 6239116, 6339068 и 6406705. Некоторые CpG-содержащие олигонуклеотиды могут включать синтетические иммуномодулирующие структурные мотивы, такие как описываемые, например, в патентах США №№ 6426334 и 6476000. В других агонистах TLR-нуклеотидных последовательностях отсутствуют последовательности CpG, и они описаны, например, в международной патентной публикации № WO 00/75304. Другие агонисты TLR-нуклеотидные последовательности включают гуанозин- и уридин-богатую одноцепочечную РНК (ssRNA), такую как описанная, например, в Heil et al, Science, vol. 303, pp. 1526-1529, Mar. 5, 2004.
Другие агонисты TLR включают биологические молекулы, такие как аминоалкилглюкозаминидфосфаты (AGP), и описаны, например, в патентах США №№ 6113918, 6303347, 6525028 и 6649172.
Агонисты TLR также включают инактивированные патогены или их фракции, которые могут активировать множество разных типов рецепторов TLR. Примеры агонистов TLR, полученных из патогенов, включают BCG, экстракт Mycobacterium obuense, талимоген лагерпарепвек (T-Vec) (полученный из HSV-1) и Pexa-Vec (полученный из вируса осповакцины).
В некоторых вариантах осуществления агонист TLR может являться агонистическим антителом, специфически связывающимся с TLR.
Ниже приведено краткое описание различных TLR, а также агонистов TLR. Перечисление агонистов TLR ниже для конкретного TLR не следует истолковывать как указание того, что упомянутый агонист TLR обязательно активирует только этот TLR (например, некоторые молекулы могут активировать множество типов TLR или даже множество классов PRR). Например, некоторые молекулы, приведенные ниже в качестве примеров агонистов TLR4, также могут являться агонистами TLR5.
Термины "TLR1" и "Толл-подобный рецептор 1" относятся к любой форме рецептора TLR1, а также вариантам, изоформам и видовым гомологам, сохраняющим по меньшей мере часть активности TLR1. Если не указано иначе, например, со ссылкой на TLR1 человека, TLR1 включает нативную последовательность TLR1 всех видов млекопитающих, например человека, обезьяны и мыши. Один из примеров TLR1 человека приведен под регистрационным номером UniProt Q15399.
В рамках изобретения термин "агонист TLR1" означает любую молекулу, которая после связывания с TLR1 (1) стимулирует или активирует TLR1, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие TLR1 или (3) повышает, способствует или индуцирует экспрессию TLR1. Агонисты TLR1, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению, включают, например, бактериальные липопротеины и их производные, связывающиеся с TLR1.
Примеры агонистов TLR1, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают, например, бактериальные липопротеины и их производные, такие как SPM-105 (полученные из автоклавированных микобактерий), OM-174 (производное липида A), OmpS1 (порин из Salmonella typhi), OmpS1 (порин из Salmonella typhi), OspA (из Borrelia burgdorferi), MALP-2 (макрофаг-активирующий липопептид-2kD микоплазмы), STF (растворимый туберкулезный фактор), CU-T12-9, дипровоцим и липопептиды, полученные из компонентов клеточной стенки, такие как PAM2CSK4, PAM3CSK4 и PAM3Cys.
TLR1 может образовывать гетеродимер с TLR2 и, таким образом, многие агонисты TLR1 также являются агонистами TLR2.
Термины "TLR2" и "Толл-подобный рецептор 2" относятся к любой форме рецептора TLR2, а также вариантам, изоформам и видовым гомологам, сохраняющим по меньшей мере часть активности TLR2. Если не указано иначе, например, со ссылкой на TLR2 человека, TLR2 включает нативную последовательность TLR2 всех видов млекопитающих, например, человека, обезьяны и мыши. Один из примеров TLR2 человека приведен под регистрационным номером UniProt O60603.
Термин "агонист TLR2" в рамках изобретения означает любую молекулу, которая после связывания TLR2 (1) стимулирует или активирует TLR2, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие TLR2 или (3) повышает, способствует или индуцирует экспрессию TLR2. Агонисты TLR2, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению, включают, например, бактериальные липопротеины и их производные, которые связываются с TLR2.
Примеры агонистов TLR2, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают, например, бактериальные липопротеины (например, диацилированные липопротеины) и их производные, такие как SPM-105 (полученный их автоклавированных микобактерий), OM-174 (производное липида A), OmpS1 (порин из Salmonella typhi), OmpS1 (порин из Salmonella typhi), OspA (из Borrelia burgdorferi), MALP-2 (макрофаг-активирующий липопептид-2kD микоплазмы), STF (растворимый туберкулезный фактор), CU-T12-9, дипровоцим, ампливант и липопептиды, полученные из компонентов клеточной стенки, такие как PAM2CSK4, PAM3CSK4 и PAM3Cys.
TLR2 может образовывать гетеродимер с TLR1 или TLR6, и, таким образом, многие агонисты TLR2 также являются агонистами TLR1 или TLR6.
Термины "TLR3" и "Толл-подобный рецептор 3" относятся к любой форме рецептора TLR3, а также вариантам, изоформам и видовым гомологам, сохраняющим по меньшей мере часть активности TLR3. Если не указано иначе, например, со ссылкой на TLR3 человека, TLR3 включает нативную последовательность TLR3 всех видов млекопитающих, например, человека, обезьяны и мыши. Один из примеров TLR3 человека приведен под регистрационным номером UniProt O15455.
В рамках изобретения термин "агонист TLR3" означает любую молекулу, которая после связывания с TLR3 (1) стимулирует или активирует TLR3, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие TLR3 или (3) повышает, способствует или индуцирует экспрессию TLR3. Агонисты TLR3, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению включают, например, лиганды нуклеиновой кислоты, связывающиеся с TLR3.
Примеры агонистов TLR3, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают лиганды TLR3, такие как синтетическая дцРНК, полиинозин-полицитидиловая кислота ["поли(I:C)"] (доступна, например, в InvivoGen в высокомолекулярных (HMW) и низкомолекулярных (LMW) препаратах), полиаденил-полиуридиловая кислота ["поли(A:U)"] (доступная, например, в InvivoGen), поли-ICLC (см. Levy et al., Journal of Infectious Diseases, vol. 132, no. 4, pp. 434-439, 1975), амплиген (см. Jasani et al., Vaccine, vol. 27, no. 25-26, pp. 3401-3404, 2009), хилтонол, ринтатолимод и RGC100 (см. Naumann et al., Clinical and Developmental Immunology, vol. 2013, article ID 283649).
Термины "TLR4" и "Толл-подобный рецептор 4" относятся к любой форме рецептора TLR4, а также вариантам, изоформам и видовым гомологам, сохраняющим по меньшей мере часть активности TLR4. Если не указано иначе, например, со ссылкой на TLR4 человека, TLR4 включает нативную последовательность TLR4 всех видов млекопитающих, например, человека, обезьяны и мыши. Один из примеров TLR4 человека приведен под регистрационным номером UniProt O00206.
в рамках изобретения термин "агонист TLR4" означает любую молекулу, которая после связывания с TLR4 (1) стимулирует или активирует TLR4, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие TLR4 или (3) повышает, способствует или индуцирует экспрессию TLR4. Агонисты TLR4, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению, включают, например, бактериальные липополисахариды (LPS) и их производные, связывающиеся с TLR4.
Примеры агонистов TLR4, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают, например, бактериальные липополисахариды (LPS) и их производные, такие как B:0111 (Sigma), монофосфорил-липид A (MPLA), 3DMPL (3-O-деацилированный MPL), GLA-AQ, G100, AS15, ASO2, GSK1572932A (GlaxoSmithKline, UK).
Термины "TLR5" и "Толл-подобный рецептор 5" относятся к любой форме рецептора TLR5, а также вариантам, изоформам и видовым гомологам, сохраняющим по меньшей мере часть активности TLR5. Если не указано иначе, например, со ссылкой на TLR5 человека, TLR5 включает нативную последовательность TLR5 всех видов млекопитающих, например, человека, обезьяны и мыши. Один из примеров TLR5 человека приведен под регистрационным номером UniProt O60602.
В рамках изобретения термин "агонист TLR5" означает любую молекулу, которая после связывания с TLR5 (1) стимулирует или активирует TLR5, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие TLR5 или (3) повышает, способствует или индуцирует экспрессию TLR5. Агонисты TLR5, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению, включают, например, бактериальные флагеллины и их производные, связывающиеся с TLR5.
Примеры агонистов TLR5, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают, например, бактериальный флагеллин, очищенный из B. subtilis, флагеллин, очищенный из P. aeruginosa, флагеллин, очищенный из S. typhimurium, и рекомбинантный флагеллин (все из которых доступны в InvivoGen), энтолимод (CBLB502; фармакологически оптимизированное производное флагеллина).
Термины "TLR6" и "Толл-подобный рецептор 6" относятся к любой форме рецептора TLR6, а также вариантам, изоформам и видовым гомологам, сохраняющим по меньшей мере часть активности TLR6. Если не указано иначе, например, со ссылкой на TLR6 человека, TLR6 включает нативную последовательность TLR6 всех видов млекопитающих, например, человека, обезьяны и мыши. Один из примеров TLR6 человека приведен под регистрационным номером UniProt Q9Y2C9.
В рамках изобретения термин "агонист TLR6" означает любую молекулу, которая после связывания с TLR6 (1) стимулирует или активирует TLR6, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие TLR6 или (3) повышает, способствует или индуцирует экспрессию TLR6. Агонисты TLR6, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению, включают, например, бактериальные липопептиды и их производные, связывающиеся с TLR6.
Примеры агонистов TLR6, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают, например, многие из агонистов TLR2, представленных выше, т.к. TLR2 и TLR6 могут образовывать гетеродимер. TLR6 также может образовывать гетеродимер с TLR4, и агонисты TLR6 могут включать различные агонисты TLR4, представленные выше.
Термины "TLR7" и "Толл-подобный рецептор 7" относятся к любой форме рецептора TLR7, а также вариантам, изоформам и видовым гомологам, сохраняющим по меньшей мере часть активности TLR7. Если не указано иначе, например, со ссылкой на TLR7 человека, TLR7 включает нативную последовательность TLR7 всех видов млекопитающих, например, человека, обезьяны и мыши. Один из примеров TLR7 человека приведен под регистрационным номером UniProt Q9NYK1.
В рамках изобретения термин "агонист TLR7" означает любую молекулу, которая после связывания с TLR7 (1) стимулирует или активирует TLR7, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие TLR7 или (3) повышает, способствует или индуцирует экспрессию TLR7. Агонисты TLR7, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению включают, например, лиганды нуклеиновой кислоты, связывающиеся с TLR7.
Примеры агонистов TLR7, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают рекомбинантную одноцепочечную ("ss")РНК, имидазохинолиновые соединения, такие как имиквимод (R837), гардиквимод и резиквимод (R848); локсорибин (7-аллил-7,8-дигидро-8-оксо-гуанозин) и родственные соединения; 7-тиа-8-оксогуанозин, 7-деазагуанозин и родственные гуанозиновые аналоги; ANA975 (Anadys Pharmaceuticals) и родственные соединения; SM-360320 (Sumimoto); 3M-01, 3M-03, 3M-852 и 3M-S-34240 (3M Pharmaceuticals); GSK2245035 (GlaxoSmithKline; молекула 8-оксоаденина), AZD8848 (AstraZeneca; молекула 8-оксоаденин), MEDI9197 (Medimmune; ранее известный как 3M-052), ssRNA40 и аналоги аденозина, такие как UC-1V150 (Jin et al., Bioorganic Medicinal Chem Lett (2006) 16:4559-4563, соединение 4). Многие агонисты TLR7 также являются агонистами TLR8.
Термины "TLR8" и "Толл-подобный рецептор 8" относятся к любой форме рецептора TLR8, а также вариантам, изоформам и видовым гомологам, сохраняющим по меньшей мере часть активности TLR8. Если не указано иначе, например, со ссылкой на TLR8 человека, TLR8 включает нативную последовательность TLR8 всех видов млекопитающих, например, человека, обезьяны и мыши. Одним из примеров TLR8 человека приведен под регистрационным номером UniProt Q9NR97.
В рамках изобретения термин "агонист TLR8" означает любую молекулу, которая после связывания с TLR8 (1) стимулирует или активирует TLR8, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие TLR8 или (3) повышает, способствует или индуцирует экспрессию TLR8. Агонисты TLR8, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению, включают, например, лиганды нуклеиновой кислоты, связывающиеся с TLR8.
Примеры агонистов TLR8, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают рекомбинантную одноцепочечную оцРНК, имиквимод (R837), гардиквимод, резиквимод (R848), 3M-01, 3M-03, 3M-852 и 3M-S-34240 (3M Pharmaceuticals); GSK2245035 (GlaxoSmithKline; молекула 8-оксоадениан), AZD8848 (AstraZeneca; молекула 8-оксоаденина), MEDI9197 (Medimmune; ранее известный как 3M-052), поли-G10, мотолимод и различные агонисты TLR7, представленные выше (как описано выше, многие агонисты TLR7 также являются агонистами TLR8).
Термины "TLR9" и "Толл-подобный рецептор 9" относятся к любой форме рецептора TLR9, а также вариантам, изоформам и видовым гомологам, сохраняющим по меньшей мере часть активности TLR9. Если не указано иначе, например, со ссылкой на человек TLR9, TLR9 включает нативную последовательность TLR9 всех видов млекопитающих, например, человека, обезьяны и мыши. Одним из примеров TLR9 человека приведен под регистрационным номером UniProt Q9NR96.
В рамках изобретения термин "агонист TLR9" означает любую молекулу, которая после связывания с TLR9 (1) стимулирует или активирует TLR9, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие TLR9 или (3) повышает, способствует или индуцирует экспрессию TLR9. Агонисты TLR9, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению, включают, например, лиганды нуклеиновой кислоты, связывающиеся с TLR9.
Примеры агонистов TLR9, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают неметилированную CpG-содержащую ДНК, иммуностимулирующие олигодезоксинуклеотиды (ODN), такие как CpG-содержащие ODN, такие как CpG24555, CpG10103, CpG7909 (PF-3512676/агатолимод), CpG1018, AZD1419, ODN2216, MGN1703, SD-101, 1018ISS и CMP-001. Агонисты TLR9 также включают нуклеотидные последовательности, содержащие синтетический цитозин-фосфат-2'-дезокси-7-деазагуанозиновый динуклеотид (CpR) (Hybridon, Inc.), dSLIM-30L1 и комплексы иммуноглобулин-ДНК. Примеры агонистов TLR9 описаны в WO2003/015711, WO2004/016805, WO2009/022215, PCT/US95/01570, PCT/US97/19791 и патентах США №№ 8552165, 6194388 и 6239116, каждый из которых включен, таким образом, в настоящее описание в качестве ссылки для всех целей.
RLR включают различные цитозольные PRR, определяющие, например, дцРНК. Примеры RLR включают, например, индуцируемый ретиноевой кислотой ген I (RIG-I), ген 5, ассоциированный с дифференцировкой меланомы (MDA-5) и белок Laboratory of Genetics and Physiology 2 (LGP2).
В рамках изобретения термин "агонист RLR" означает любую молекулу, которая после связывания с RLR (1) стимулирует или активирует RLR, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие RLR или (3) повышает, способствует или индуцирует экспрессию RLR. Агонисты RLR, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению, включают, например, нуклеиновые кислоты и их производные, связывающиеся с RLR, и агонистические моноклональные антитела (mAb), специфически связывающееся с RLR.
Примеры агонистов RLR, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают, например, короткую двухцепочечную РНК с некэпированным 5’-трифосфатом (агонист RIG-I); поли I:C (агонист MDA-5) и BO-112 (агонист MDA-A).
NLR включают различные PRR, определяющие, например, молекулы молекулярных паттернов, ассоциированных с повреждением (DAMP). NLR включают подсемейства NLRA-A, NLRB-B, NLRC-C и NLRP-P. Примеры NLR включают, например, NOD1, NOD2, NAIP, NLRC4 и NLRP3.
В рамках изобретения термин "агонист NLR" означает любую молекулу, которая после связывания с NLR (1) стимулирует или активирует NLR, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие NLR или (3) повышает, способствует или индуцирует экспрессию NLR. Агонисты NLR, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению, включают, например, DAMP и их производные, связывающиеся с NLR, и агонистические моноклональные антитела (mAb), специфически связывающееся с NLR.
Примеры агонистов NLR, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают, например, липосомальный мурамилтрипептид/мифамуртид (агонист NOD2).
CLR включают различные PRR, определяющие, например, углеводы и гликопротеины. CLR включают трансмембранные CLR и секретируемые CLR. Примеры CLR включают, например, DEC-205/CD205, макрофагальный рецептор маннозы (MMR), дектин-1, дектин-2, Mincle, DC-SIGN, DNGR-1 и маннозо-связывающий лектин (MBL).
В рамках изобретения термин "агонист CLR" означает любую молекулу, которая после связывания с CLR (1) стимулирует или активирует CLR, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие CLR или (3) повышает, способствует или индуцирует экспрессию CLR. Агонисты CLR, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению, включают, например, углеводы и их производные, связывающиеся с CLR и агонистические моноклональные антитела (mAb), специфически связывающееся с CLR.
Примеры агонистов CLR, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают, например, MD-фракцию (очищенный растворимый бета-глюкановый экстракт из Grifola frondosa) и Imprime PGG (бета 1,3/1,6-глюкановый PAMP, полученный из дрожжей).
Белок STING функционирует как цитозольный сенсор ДНК и адаптерный белок в передаче сигнала интерферона типа 1. Термины "STING" и "стимулятор генов интерферонов" относятся к любой форме белка STING, а также вариантам, изоформам и видовым гомологам, сохраняющим по меньшей мере часть активности STING. Если не указано иначе, например, со ссылкой на STING человека, STING включает нативную последовательность STING всех видов млекопитающих, например, человека, обезьяны и мыши. Один из примеров TLR9 человека приведен под регистрационным номером UniProt Q86WV6. STING также известен как TMEM173.
В рамках изобретения "термин STING" означает любую молекулу, которая после связывания с TLR9 (1) стимулирует или активирует STING, (2) повышает, способствует, индуцирует или пролонгирует активность, функцию или присутствие STING или (3) повышает, способствует или индуцирует экспрессию STING. Агонисты STING, которые можно использовать в любом из способов лечения, лекарственных средств и применении по настоящему изобретению, включают, например, лиганды нуклеиновой кислоты, связывающиеся с STING.
Примеры агонистов STING, которые можно использовать в способах лечения, лекарственных средствах и применении по настоящему изобретению, включают различные иммуностимулирующие нуклеиновые кислоты, такие как синтетическая двухцепочечная ДНК, циклическая ди-ГМФ, циклическая ГМФ-АМФ (цГАМФ), синтетические циклические динуклеотиды (CDN), такие как MK-1454 и ADU-S100 (MIW815), и низкомолекулярные соединения, такие как P0-424.
Другие PRR включают, например, ДНК-зависимый активатор ИФН-регуляторных факторов (DAI) и белок Absent in Melanoma 2 (AIM2).
Иммуностимулирующие цитокины включают различные сигнальные белки, стимулирующие иммунный ответ, такие как интерфероны, интерлейкины и гемопоэтические факторы роста.
Примеры иммуностимулирующих цитокинов включают ГМ-КСФ, Г-КСФ, ИФН-альфа, ИФН-гамма; ИЛ-2 (например, денилейкин-дифтитокс), ИЛ-6, ИЛ-7, ИЛ-11, ИЛ-12, ИЛ-15, ИЛ-18, ИЛ-21 и ФНО-альфа.
Иммуностимулирующие цитокины могут иметь любой подходящий формат. В некоторых вариантах осуществления иммуностимулирующий цитокин может являться рекомбинантной версией цитокина дикого типа. В некоторых вариантах осуществления иммуностимулирующий цитокин может являться мутеином, имеющим одно или более изменений аминокислот по сравнению с соответствующим цитокином дикого типа. В некоторых вариантах осуществления иммуностимулирующий цитокин можно включать в химерный белок, содержащий цитокин и по меньшей мере один другой функциональный белок (например, антитело). В некоторых вариантах осуществления иммуностимулирующий цитокин можно ковалентно связывать с лекарственным средством/средством (например, любым лекарственным средством/средством, описанным где-либо в настоящем описании, в качестве возможного компонента ADC).
Противоопухолевые вакцины включают различные композиции, содержащие опухолеассоциированные антигены (или которые можно использовать для получения опухолеассоциированного антигена у индивидуума), и, таким образом, их можно использовать для провоцирования иммунного ответа у индивидуума, которые будут направлены на опухолевые клетки, содержащие опухолеассоциированный антиген.
Примеры материалов, которые можно включать в противоопухолевую вакцину, включают, аттенуированные злокачественные клетки, опухолевые антигены, антигенпрезентирующие клетки, такие как дендритные клетки, в которые добавлен опухолевый антиген или нуклеиновые кислоты, кодирующие опухолеассоциированный антигены. В некоторых вариантах осуществления противоопухолевую вакцину можно получать с использованием собственных злокачественных клеток пациента. В некоторых вариантах осуществления противоопухолевую вакцину можно получать с использованием биологического материала не из собственных злокачественных клеток пациента.
Противоопухолевые вакцины включают, например, сипулеуцел-T и талимоген лагерпарепвек (T-VEC).
Терапия иммунными клетками включает лечение пациента иммунными клетками, способными к таргетингу злокачественных клеток. Терапия иммунными клетками включает, например, инфильтрирующие опухоль лимфоциты (TIL) и T-клетки с химерными антигенными рецепторами (CAR-T-клетки).
Примеры химиотерапевтическими средствами включают алкилирующие средства, такие как тиотепа и циклофосфамид; алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метилмеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилоломеламин; ацетогенины (в частности, буллатацин и буллатацинон); камптотецин (включая синтетический аналог топотекан); бриостатин; каллистатин; CC-1065 (включая его синтетические аналоги адозелесин, карзелесин и бизелесин); криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги, KW-2189 и CBI-TMI); элеутеробин; панкратистатин; саркодиктин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, холофосфамид, эстрамустин, ифосфамид, мехлоретамин, гидрохлорид оксида мехлоретамина, мелфалан, новэмбихин, фенестерин, преднимустин, трофосфамид, урамустин; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин, ранимустин; антибиотики, такие как ендииновые антибиотики (например, калихимицин, в частности, калихимицин гамма 1I и калихимицин фи I1, см., например, Agnew, Chem. Intl. Ed. Engl., 33:183-186 (1994); динемицин, включая динемицин A; бисфосфонаты, такие как клодронат; эсперамицин; а также неокарциностатин хромофор и родственные хромопротеиновые хромофоры ендииновых антибиотиков), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, каминомицин, карцинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин (включая морфолино-доксорубицин, цианоморфолино-доксорубицин, 2-пирролино-доксорубицин и дезоксидоксорубицин), пегилированный липосомальный доксорубицин, эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин C, микофеноловую кислоту, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, пропионат дромостанолона, эпитиостанол, мепитиостан, тестолактон; антиадреналовые средства, такие как аминоглутемид, митотан, трилостан; пополнитель фолиевой кислоты, такой как фролиновая кислота; ацеглатон; альдофосфамид гликозид; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатрексат; дефофамин; демеколцин; диазиквон; эфлорнитин; ацетат эллиптиния; эпотилон; этоглюцид; нитрат галлия; гидроксимочевину; лентинан; лонидамин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопидамол; нитракрин; пентостатин; фенамет; пирарубицин; лозоксантрон; подофиллиновую кислоту; 2-этилгидразид; прокарбазин; разоксан; ризоксин; сизофуран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2′,2″-трихлортриэтиламин; трихотецены (в частности, токсин T-2, верракурин A, роридин A и ангуидин); уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гaцитозин; арабинозид ("Ara-C"); циклофосфамид; тиотепа; таксоиды, например, паклитаксел и доксетаксел; хлорамбуцил; гемцитабин; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митоксантрон; винкристин; винорелбин; новантрон; тенипозид; эдатрексат; дауномицин; аминоптерин; кселода; ибандронат; CPT-11; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин; и фармацевтически приемлемые соли, кислоты или производные любых из указанных выше соединений. Также включены противогормональные средства, регулирующие или ингибирующие действие гормонов на опухоли, такие как антиэстрогены и селективные модуляторы эстрогеновых рецепторов (SERM), включая, например, тамоксифен, ралоксифен, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоусифен, LY117018, онапристон и торемифен (фарестон); ингибиторы ароматазы, ингибирующие фермент ароматазу, регулирующую продукцию эстрогена в надпочечников, такие как, например, 4(5)-имидазолы, аминоглутетимид, мегестрол ацетат, экземестан, форместан, фадрозол, ворозол, летрозол и анастрозол; и антиандрогены, такие как флутамид, нилутамид, бикалутамид, лейпролид и гозерелин; ингибиторы KRAS; ингибиторы MCT4; ингибиторы MAT2a; ингибиторы тирозинкиназ, такие как сунитиниб, акситиниб; ингибиторы alk/c-Met/ROS, такие как кризотиниб, лорлатиниб; ингибиторы mTOR, такие как темсиролимус, гедатолисиб; ингибиторы src/abl, такие как босутиниб; ингибиторы циклин-зависимых киназ (CDK), такие как палбоциклиб, PF-06873600; ингибиторы erb, такие как дакомитиниб; ингибиторы PARP, такие как талазопариб; ингибиторы SMO, такие как гласдегиб, PF-5274857; ингибиторы EGFR T790M, такие как PF-06747775; ингибиторы EZH2, такие как PF-06821497; ингибиторы PRMT5, такие как PF-06939999; ингибиторы TGFRβr1, такие как PF-06952229; и фармацевтически приемлемые соли, кислоты или производные любого из указанных выше соединений. В конкретных вариантах осуществления такое дополнительное терапевтическое средство является бевацизумабом, цетуксимабом, сиролимусом, панитумумабом, 5-фторурацилом (5-FU), капецитабином, тивозанибом, иринотеканом, оксалиплатином, цисплатином, трифлуридином, типирацилом, лейковорином, гемцитабином, регорафинибом или гидрохлоридом эрлотиниба.
В некоторых вариантах осуществления антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c используют в комбинации с одним или более другими терапевтическими средствами, направленно воздействующими на модулятор иммунных контрольных точек или костимуляторное средство, такими как, в качестве неограничивающих примеров, средство, направленно воздействующее на CTLA-4, LAG-3, B7-H3, B7-H4, B7-DC (PD-L2), B7-H5, B7-H6, B7-H8, B7-H2, B7-1, B7-2, ICOS, ICOS-L, TIGIT, CD2, CD47, CD80, CD86, CD48, CD58, CD226, CD155, CD112, LAIR1, 2B4, BTLA, CD160, TIM1, TIM-3, TIM4, VISTA (PD-H1), OX40, OX40L, GITR, GITRL, CD70, CD27, 4-1BB, 4-BBL, DR3, TL1A, CD40, CD40L, CD30, CD30L, LIGHT, HVEM, SLAM (SLAMF1, CD150), SLAMF2 (CD48), SLAMF3 (CD229), SLAMF4 (2B4, CD244), SLAMF5 (CD84), SLAMF6 (NTB-A), SLAMCF7 (CS1), SLAMF8 (BLAME), SLAMF9 (CD2F), CD28, CEACAM1 (CD66a), CEACAM3, CEACAM4, CEACAM5, CEACAM6, CEACAM7, CEACAM8, CEACAM1-3AS, CEACAM3C2, CEACAM1-15, PSG1-11, CEACAM1-4C1, CEACAM1-4S, CEACAM1-4L, IDO, TDO, CCR2, путь CD39-CD73-аденозин (A2AR), BTK, TIK, CXCR2, CXCR4, CCR4, CCR8, CCR5, CSF-1 или модулятор врожденного иммунного ответа.
В некоторых вариантах осуществления антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c используют в комбинации, например, с антагонистическим антителом против CTLA-4, таким как, например, ипилимумаб; антагонистическим антителом против LAG-3, таким как BMS-986016 и IMP701; антагонистическим антителом против TIM-3; антагонистическим антителом против B7-H3, таким как, например, MGA271; антагонистическим антителом против VISTA; антагонистическим антителом против TIGIT; антитело; антителом против CD80; антителом против CD86; антагонистическим антителом против B7-H4; агонистическим антителом против ICOS; агонистическим антителом против CD28; модулятором врожденного иммунного ответа (например, TLR, KIR, NKG2A) и ингибитором IDO.
В некоторых вариантах осуществления антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c используют в комбинации с агонистом OX40, таким как, например, агонистическое антитело против OX-40. В некоторых вариантах осуществления антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c используют в комбинации с агонистом GITR, таким как, например, агонистическое антитело против GITR, такое как, в качестве неограничивающих примеров, TRX518. В некоторых вариантах осуществления антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c используют в комбинации с ингибитором IDO. В некоторых вариантах осуществления антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c используют в комбинации с цитокиновой терапией, такой как, в качестве неограничивающих примеров, ИЛ-15, КСФ-1, М-КСФ-1 и т.д.
В некоторых вариантах осуществления антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c используют в комбинации с одним или более другими терапевтическими антителами, такими как, в качестве неограничивающих примеров, антитело против CD19, CD22, CD40, CD52 или CCR4.
В некоторых вариантах осуществления композиция антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c содержит по меньшей мере одно дополнительное средство, такое как бевацизумаб, цетуксимаб, сиролимус, панитумумаб, 5-фторурацил(5-FU), капецитабин, тивозаниб, иринотекан, оксалиплатин, цисплатин, трифлуридин, типирацил, лейковорин, гемцитабин и эрлотиниба гидрохлорид.
В некоторых вариантах осуществления антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c можно вводить совместно или вводить последовательно до или после другого средства с интервалами в диапазоне от минут до недель. В вариантах осуществления, если другие средства, и/или белки, или полинуклеотиды вводят раздельно, как правило, будут обеспечивать то, что между каждым введением не проходил значительный период времени, таким образом, что средство и композиция по настоящему изобретению все равно смогут вызывать преимущественно комбинированный эффект в отношении индивидуума. В таких случаях предусмотрено, что можно вводить оба средства в пределах приблизительно 12-24 ч. друг относительно друга и, более предпочтительно - в пределах приблизительно 6-12 ч. друг относительно друга. Однако в некоторых случаях желательным может являться значительное увеличение периода времени для введения, где между соответствующими введениями проходит от нескольких дней (2, 3, 4, 5, 6 или 7) до нескольких недель (1, 2, 3, 4, 5, 6, 7 или 8).
В некоторых вариантах осуществления композицию антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c комбинируют со схемой лечения, дополнительно включающей общепринятую терапию, выбранную из группы, состоящей из: хирургического вмешательства, лучевой терапии, химиотерапии, таргетированной терапии, иммунотерапии, гормональной терапии, ингибирования ангиогенеза и паллиативной терапии.
Композиция, используемая в настоящем изобретении, может дополнительно содержать фармацевтически приемлемые носители, эксципиенты или стабилизаторы (Remington: The Science and practice of Pharmacy 21st Ed., 2005, Lippincott Williams and Wilkins, Ed. K. E. Hoover) в форме лиофилизированных составов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в дозах и концентрациях и могут содержать буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и m-крезол); низкомолекулярные (менее приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстраны; хелатирующие средства, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексные соединения с металлами (например, комплексы Zn-белок); и/или неионные поверхностно-активные средства, такие как TWEEN™, PLURONIC™ или полиэтиленгликоль (PEG). Фармацевтически приемлемые эксципиенты дополнительно приведены в настоящем описании.
В одном из аспектов настоящее изобретение относится к способу получения антитела против GUCY2c или биспецифического антитела, как представлено в настоящем описании, включающему культивирование клетки-хозяина в условиях, приводящих к продукции антитела против GUCY2c или биспецифического антитела, представленного в настоящем описании, и очистку антитела или биспецифического антитела из культуры.
В другом аспекте настоящее изобретение относится к применению антитела против GUCY2c, биспецифического антитела, фармацевтической композиции, полинуклеотида, супернатанта, вектора или клетки-хозяина, представленных в настоящем описании, в производстве лекарственного средства для детекции, диагностики и/или лечения GUCY2c-ассоциированного нарушения.
Дополнительный аспект изобретения относится к набору, содержащему антитело против GUCY2c или биспецифическое антитело против CD3-GUCY2c, как представлено в настоящем описании выше, и инструкции по использованию в соответствии с любыми из способов по изобретению, представленных в настоящем описании. Как правило, эти инструкции включают описание введения антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c для описанного выше терапевтического лечения. Этот набор содержит любую фармацевтическую композицию, представленную в настоящем описании. Фармацевтические композиции и другие реагенты могут находиться в наборах в любой удобной форме, такой как, например, раствор или порошок.
В другом аспекте набор дополнительно содержит одно или более других профилактических или терапевтических средств, которые можно использовать в лечении злокачественного новообразования, в одном или более контейнерах. В одном из вариантов осуществления другое профилактическое или терапевтическое средство является химиотерапевтическим средством. В других аспектах профилактическое или терапевтическое средство является биологическим или гормональным терапевтическим средством.
Некоторые аспекты фармацевтических композиций, профилактических или терапевтических средств по изобретению, предпочтительно, тестируют in vitro, в системе культуры клеток и в модели на животных организм, такой как система модели на грызунах, на желаемую терапевтическую активность перед использованием на людях.
Токсичность и эффективность профилактических и/или терапевтических способов по настоящему изобретению можно определять стандартными фармацевтическими способами в культурах клеток или на экспериментальных животных, например, для определения LD50 (дозы, летальной для 50% популяции) и ED50 (дозы, терапевтически эффективной для 50% популяции). Соотношение дозы между токсическими и терапевтическими эффектами представляет собой терапевтический индекс, и его можно выражать как соотношение LD50/ED50. Профилактические и/или терапевтические средства, демонстрирующие большие терапевтические индексы, являются предпочтительными.
Кроме того, для оценки профилактической и/или терапевтической полезности терапевтических средств или комбинированной терапии, представленных в настоящем описании, для лечения или профилактики злокачественного новообразования можно использовать любые анализы, известные специалистам в этой области.
Инструкции, касающиеся использования антитела против GUCY2c или биспецифического антитела против CD3-GUCY2c, представленного в настоящем описании, как правило, включают информацию о дозе, режиме дозирования и пути введения для предполагаемого лечения. Контейнеры могут представлять собой однократные дозы, многодозовые упаковки или субоднократные дозы. Инструкции, поставляемые в наборах по изобретению, как правило, являются письменными инструкциями на ярлыке или вкладыше в упаковку (например, бумажном листе, включенном в набор), но машиночитаемые инструкции (например, инструкции на магнитном или оптическом устройстве хранения данных) также являются приемлемыми.
Наборы по настоящему изобретению находятся в подходящей упаковке. Подходящая упаковка включает, в качестве неограничивающих примеров, флаконы, ампулы, пробирки, бутыли, банки, гибкую упаковку (например, запаянные майларовые или пластиковые пакеты) и т.п. для каждой фармацевтической композиции и других включенных реагентов, например, буферов, сбалансированных солевых растворов и т.д., для использования в введении фармацевтических композиций индивидуумам. Также предусмотрены упаковки для использования в комбинации с конкретным устройством, таким как ингалятор, устройство для назального введения (например, атомайзер) или инфузионное устройство, такое как минипомпа. Набор может иметь стерильное входное отверстие (например, контейнер может являться мешком для внутривенного раствора или флаконом, имеющим пробку, прокалываемую иглой для подкожных инъекций). Контейнер также может иметь стерильное входное отверстие (например, контейнер может являться мешком для внутривенного раствора или флаконом, имеющим пробку, прокалываемую иглой для подкожных инъекций). По меньшей мере одно активное средство в композиции является антителом против GUCY2c или биспецифическим антителом против CD3-GUCY2c. Контейнер может дополнительно содержать второе фармацевтически активное средство.
Как правило, набор содержит контейнер и ярлык или вкладыши в упаковку на контейнере или прикрепленным к контейнеру.
Депонирование биологического материала
Типичные материалы по настоящему изобретению депонированы в American Type Culture Collection, 10801 University Boulevard, Manassas, Va. 20110-2209, USA, на 13 февраля 2018 года. Вектор цепи A GUCY2C-1608 (VH-впадина SEQ ID NO: 220), имеющий регистрационный номер ATCC PTA-124943, содержит вставку ДНК (SEQ ID NO: 247), кодирующую цепь VH-впадина биспецифического антитела GUCY2C-1608, и вектор цепи B GUCY2C-1608 (VL-выступ SEQ ID NO: 216), имеющий регистрационный номер ATCC PTA-124944, содержит вставку ДНК (SEQ ID NO: 246), кодирующую цепь VL-выступ биспецифического антитела GUCY2C-1608.
Депонирование осуществляли в соответствии с положениями Будапештского договора о международном признании депонирования микроорганизмов для целей патентной процедуры и нормативных актов в рамках него (Будапештский договор). Это гарантирует поддержание жизнеспособной культуры депозита в течение 30 лет с даты депонирования. Депозит будет сделан доступным ATCC в рамках Будапештского договора и является объектом соглашения между Pfizer Inc. и ATCC, обеспечивающей постоянный и неограниченный доступ потомства культуры депозита для публики после выдачи пертинентного патента США или выкладки любой патентной заявки США или иностранной патентной заявки, в зависимости от того что произойдет первым и обеспечивающей доступность потомства кому-либо, определенному Ведомством по патентам и товарным знакам США как имеющему право на это в соответствии с разделом 122 35 U.S.C. и правилами Ведомства, соответствующими ему (включая раздел 1.14 37 C.F.R. с конкретной ссылкой на 886 OG 638).
Правоприобретатель по настоящей заявке согласен на то, что, если материалы депозита погибнут, или будут утрачены, или будут разрушены при культивировании в подходящих условиях, материалы незамедлительно будут заменены по уведомлении такими же материалами. Доступность депонируемого материала не следует истолковывать как лицензию на практическое осуществление настоящего изобретения в нарушение прав, предоставленных властью любого правительства в соответствии с патентным правом.
Следующие примеры конкретных аспектов осуществления настоящего изобретения представлены исключительно в иллюстративных целях и не предназначены для ограничения объема настоящего изобретения каким-либо образом.
Примеры
Пример 1: Иммунизация и гибридомное получение антител мыши против GUCY2c
Рекомбинантный иммуноген-белок GUCY2c человека или клетки, гиперэкспрессирующие GUCY2c человека на своей поверхности (SEQ ID NO: 224), инъецировали мышам Balb/c для получения гибридом. Самок мышей Balb/c возрастом восемь недель иммунизировали растворимым белком huGUCY2c-mIgG2a или линией клеток 300.19, экспрессирующей GUCY2c человека (SEQ ID 224/225). В случае мышей, иммунизированных рекомбинантным белком huGUCY2c, животным вводили дозы по протоколу RIMMS (Kilpatrick, et al. Hybridoma. 1997. 16:381-389). В кратком изложении, мышей иммунизировали шесть раз рекомбинантным Fc-слитым белком GUCY2c человека-IgG2a мыши (SEQ ID NO: 230) в течение двух недель посредством подкожной инъекции. Используемыми адъювантами являлись полный и неполный адъюванты Фрейнда. В случае иммунизации на основе клеток мышей Balb/c иммунизировали с помощью 5×106 клеток 300.19, гиперэкспрессирующих GUCY2c человека, дважды в неделю в течение одного месяца без адъюванта посредством i.p. инъекции. Для определения титров антитела мыши против GUCY2c получали образцы крови иммунизированных животных и исследовали GUCY2c-специфические титры посредством твердофазного иммуноферментного анализа (ELISA) с использованием рекомбинантного GUCY2c mIgG2a-Fc человека (SEQ ID NO: 230) и GUCY2c-mIgG2a-Fc яванского макака (SEQ ID NO: 234). Животных с наиболее высокими титрами GUCY2c выбирали для получения гибридомы. В случае мышей, иммунизированных растворимым рекомбинантным белком, для слияния использовали лимфоциты LN и спленоциты, в то время как в случае мышей, иммунизированных 300.19/hGUCY2c, для слияния использовали только спленоциты. Для получения гибридомы спленоциты и/или лимфоциты лимфоузлов подвергали слиянию с миеломой P3X в соотношении 1:1 с использованием PEG. Слитые клетки подвергали селекции в присутствии HAT в течение семи дней, после чего гибридомы поддерживали в HT-содержащих средах перед скринингом. Для идентификации специфических в отношении антитела против huGUCY2c гибридом, супернатанты гибридом подвергали скринингу на реактивность IgG против huGUCY2c посредством ELISA. Затем потенциальные гибридомы против GUCY2c объединяли и анализировали на реактивность в отношении GUCY2c человека и яванского макака на поверхности трансфицированных линий клеток 300.19 и первичных линий опухолевых клеток, экспрессирующих GUCY2c. В кратком изложении, линии клеток 300.19, экспрессирующих различные белки GUCY2c (SEQ ID NO: 224, 226 и 228), высевали в количестве 4×104 клеток/лунку в стерильные 96-луночные планшеты для культивирования тканей (Becton Dickinson) в день 1 и инкубировали при 37°C/5% CO2 в течение 1-2 дней до достижения конфлюэнтного монослоя. Клетки 4 раза промывали PBS+Ca2+ и Mg2+ и блокировали в течение 1 часа при комнатной температуре с помощью 3% молока/1% BSA/PBS+Ca2+ и Mg2+. Серийно разведенные образцы (1:3 в блокирующем буфере) наносили на планшет. Планшеты инкубировали при комнатной температуре в течение 2 часов перед промывкой PBS+Ca2+ и Mg2+. Затем добавляли HRP-конъюгированное вторичное антитело, антитело козы против Fc IgG мыши (Thermo Scientific), разведенное (1:4.000) в блокирующем буфере, и инкубировали с клетками в течение 1 часа. Планшеты промывали PBS+Ca2+ и Mg2+ перед проявлением с помощью раствора субстрата TMB в течение 10 минут, а затем добавляли 0,18M⋅H2SO4 для остановки реакции. Измеряли поглощение при OD 450 нМ, строили график данных и анализировали с помощью программного обеспечения Microsoft Excel и Graphpad-Prism. Идентифицировали гибридомы, специфически связывающиеся с GUCY2c человека и яванского макака.
Пример 2: Связывание гибридомных антител против GUCY2c в анализах проточной цитометрии и иммуногистохимии
Гибридомные антитела против GUCY2c подвергали скринингу посредством проточной цитометрии на связывание на поверхности клетки с GUCY2c человека на опухолевых клетках T84 и HT55. Клетки HT29 использовали в качестве отрицательных контролей. Клетки подвергали диссоциации с помощью буфера для диссоциации клеток (Sigma) и блокировали на льду с помощью PBS+3% BSA. Клетки окрашивали на льду с использованием первичного антитела в течение 1 часа, а затем промывали холодным PBS. Добавляли 10 мкг/мл вторичного антитела (PE-конъюгированного вторичного антитела против мыши, Jackson Immunoresearch) на льду в течение часа. Клетки снова промывали холодным PBS, ресуспендировали в PBS, окрашивали DAPI для различения живых/погибших клеток и анализировали на связывание GUCY2c с помощью BD LSRII Fortessa. Идентифицировали несколько клонов, демонстрирующих специфическое связывание с GUCY2c-экспрессирующими линиями клеток (T84 и HT55) и отсутствие связывания с GUCY2c-отрицательными клетками (HT29).
Гибридомы из мышей BALB/c, иммунизированных против GUCY2c, тестировали на иммунореактивность в фиксированных формалином погруженных в парафин клеточных осадках. Клеточные осадки, полученные при этом скрининге, состояли из родительских клеток 300.19, клеток 300.19, гиперэкспрессирующих GUCY2c мыши, яванского макака или человека, одной линии клеток колоректального рака человека, экспрессирующих высокие эндогенные уровни GUCY2c (T84), и одной линии клеток колоректального рака человека, отрицательных по экспрессии GUCY2c (HT29). Линии клеток фиксировали в течение 24 ч. в 10% нейтральном забуференном формалине (Thermo Scientific) и центрифугировали при 300×g в течение 4 минут (мин) для осаждения. Формалин удаляли и клеточные осадки осторожно ресуспендировали предварительно нагретым до 50ºC Histogel (Thermo Scientific). Клеточные осадки, заключенные в Histogel, охлаждали при 4°C в течение 1-2 ч. перед обработкой в течение ночи в автоматизированном гистопроцессоре VIP (Ткань-Tek). Обработанные клеточные осадки заключали в парафин. Нарезали срезы микропанелей клеток по пять мкм, переносили на водяную баню и помещали на предметные стекла Superfrost Excell (Fisher). Стеклам позволяли сохнуть в течение ночи. После депарафинизации и регидратации тканевых срезов осуществляли термически-индуцируемое демаскирование эпитопов с помощью варочного автоклава Retriever 2100 (Electron Microscopy Sciences) в буфере Borg Decloaker, pH 9,5 (Biocare Medical), или цитратном буфере, pH 6,0 (Thermo Scientific), с последующим охлаждением до комнатной температуры (RT). Активность эндогенной пероксидазы инактивировали с помощью Peroxidazed 1 (Biocare Medical) в течение 10 мин. Неспецифические белковые взаимодействия блокировали в течение 10 мин с помощью Background Punisher (Biocare Medical). Каждую гибридому инкубировали без разведения в течение 1 ч. в условиях термически-индуцируемого демаскирования эпитопов. Срезы промывали в TBS и определяли связывание гибридомы с помощью Envision+ Mouse HRP (DAKO) в течение 30 мин. Препараты промывали в TBS и определяли иммунореактивность с использованием набора Betazoid DAB Chromogen Kit (Biocare Medical) в течение 5 мин с последующим промыванием дистиллированной водой. Иммуноокрашенные среды быстро докрашивали гематоксилина CAT (Biocare Medical), промывали водопроводной водой, дегидратировали батареей спиртов, очищали в ксилоле и накрывали покровным стеклом с помощью гистологической среды Permount (FisherChemicals). Препараты оценивал ветеринарный патолог для анализа иммунореактивности. При этом скрининге идентифицировали клоны с иммунореактивностью в отношении GUCY2c человека, яванского макака и мыши.
Результаты, касающиеся связывания гибридомных антител против GUCY2c по данным проточной цитометрии и иммуногистохимии (IHC), приведены в таблице 11.
Таблица 11
Пример 3: Количественный анализ уровней экспрессии GUCY2c на линиях клеток
Плотность рецепторов GUCY2c на линиях клеток измеряли с использованием клона GUCY2C-9H3, конъюгированного в соотношении 1:1 с фикоэритрином (Thermo Scientific). 1×105 клеток окрашивали с помощью 10 мкг/мл PE-меченого mAb против GUCY2c для насыщающего связывания. Клетки промывали и ресуспендировали в буфере для FACS с DAPI и анализировали с использованием BD LSRII Fortessa с программным обеспечением FACS Diva. Меченые QuantiBRITE PE бусы (BD) использовали в тех же условиях определения нагрузки PE для вычисления количества PE-меченых антител на клетку (ABC) с учетом геометрического среднего интенсивности флуоресценции PE. Измерения плотности рецептора GUCY2c в линиях опухолевых клеток человека приведены в таблице 12. Линии клеток с плотностями рецептора GUCY2c охарактеризовывали в этом анализе и использовали в последующих анализах (например, анализах цитотоксичности) для оценки функциональной активности биспецифических антител против CD3-GUCY2c.
Таблица 12
Пример 4: Клонирование и секвенирование гибридомных антител против GUCY2c
Супернатанты гибридом с подтвержденной связывающей активностью на поверхности клетки субклонировали в формат IgG1 мыши. Выделяли РНК и получали последовательности ДНК вариабельной (V) области из экспрессирующихся антител посредством клонирования с помощью полимеразной цепной реакции с обратной транскрипцией (RT-ПЦР), как описано ниже.
От одного до пяти миллионов субклонированных гибридомных клеток гомогенизировали для выделения тотальной РНК с помощью набора QIAGEN RNAeasy Mini. Затем первую цепь кДНК получали с использованием набора SuperScript III RT (Invitrogen). Затем получали двухцепочечную кДНК для вариабельных областей IgG против GUCY2c и амплифицировали посредством ПЦР с использованием праймеров для константных областей тяжелой цепи (IgG1, IgG2a, IgG2b) и легкой цепи (каппа или лямбда) IgG мыши, как описано ниже. Условия циклов ПЦР являлись следующими: 1 цикл при 95°C в течение 1 мин; 25 циклов при 95°C в течение 1 мин, 63°C в течение 1 мин и 72°C в течение 1 мин. Получаемые продукты RT-ПЦР клонировали в клонирующий вектор TOPO-Blunt (Invitrogen) и секвенировали.
Остатки в вариабельном домене нумеровали по Kabat, что представляет собой систему нумерации, используемую для вариабельных доменов тяжелой цепи или вариабельных доменов легких цепей компиляции антител (Kabat et al., 1991, Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD). При использовании этой системы нумерации точная линейная аминокислотная последовательность может содержать меньше или больше аминокислот, что соответствует укорочению или инсерции в FR или CDR вариабельного домена. Например, вариабельный домен тяжелой цепи может включать инсерцию одной аминокислоты (остатка 52a по Kabat) после остатка 52 H2 и встроенные остатки (например, остатки 82a, 82b и 82c по Kabat) после остатка 82 каркаса (FR) тяжелой цепи. Нумерацию остатков по Kabat можно определять для указанного антитела посредством выравнивания в областях гомологии последовательности антитела со "стандартной", пронумерованной по Kabat последовательностью. Доступны различные алгоритмы приписывания нумерации по Kabat. В настоящем описании для приписывания нумерации по Kabat CDR1 VL, CDR2 VL, CDR3 VL, CDR2 VH и CDR3 VH вариабельных областей используют алгоритм, воплощенный в версии 2.3.3 выпуска Abysis (www.abysis.org). Определение AbM используют для CDR1 VH. Остатки FR являются остатками вариабельного домена антитела, иными, чем остатки CDR. Каркас домена VH или VL содержит четыре подобласти каркаса, FR1, FR2, FR3 и FR4, перемежающиеся с CDR в следующем порядке: FR1 - CDR1 - FR2 - CDR2 - FR3 - CDR3 - FR4.
Химерные T-клеточные биспецифические белки с V-областями мыши и модифицированные константные области IgG человека (hIgG1) получали следующим образом: кДНК V-области из родительских векторов мыши TOPO субклонировали в экспрессирующие векторы млекопитающих, состоящие из первого и второго способствующего гетеродимеризации домена, каждый из которых содержит цистеиновый линкер (линкер 3), такой как GCPPCP (SEQ ID NO: 192), и Fc-цепь с "выступом" (SEQ ID NO: 188) или Fc-цепь с "впадиной" (SEQ ID NO: 189). В этом случае, вариабельную область тяжелой цепи (VH) антитела против GUCY2c мыши (SEQ ID NO: 11, 19, 26, 33 и 41) подвергали слиянию в рамке считывания с вариабельной областью легкой цепи (VL) антитела против CD3 человека (SEQ ID NO: 84), разделяя линкером GGGSGGGG (SEQ ID NO: 190), а затем другим линкером GCPPCP (SEQ ID NO: 192), соединяя с Fc-цепью с "впадиной" (SEQ ID NO: 189). Области VL против GUCY2c мыши (SEQ ID NO: 92, 100, 104, 106, 112 и 119) подвергали слиянию в рамке считывания с линкером GGGSGGGG (SEQ ID NO: 190), областью VH против CD3 человека (SEQ ID NO: 1), с последующим линкером GCPPCP (SEQ ID NO: 192) и Fc-цепь с "выступом" (SEQ ID NO: 188). Фрагменты амплифицировали посредством стандартной ПЦР с использованием праймеров, включающих липкие концы по 15-25 нуклеотидов, имеющих гомологию с принимающим вектором, для клонирования с изотермальной сборкой (ITA). Фрагменты рекомбинировали в указанные выше линеаризованные векторы с использованием набора для клонирования Infusion ITA (Clontech).
Затем продукты лигирования трансформировали в компетентные E. coli DH5α (Life Technologies) и выращивали в течение ночи при 37°C на чашках с агарозой, содержащей 100 мкг/мл карбенициллина. Колонии подсчитывали, отбирали и выращивали в течение ночи в бульоне YT с 100 мкг/мл карбенициллина и выделяли ДНК стандартными способами. Затем все конструкции ДНК секвенировали по обеим цепям перед экспрессией в клетках млекопитающего с использованием общепринятых способов секвенирования.
Пример 5: Экспрессия и очистка химерных биспецифических антител против CD3-GUCY2c
С помощью комплементарных пар конструкций (по 12,5 мкг каждой цепи) котрансфицировали 25 мл культуры в логарифмической фазе роста, содержащих 1 миллион клеток/мл клеток HEK 293 с использованием набора для трансфекции ExpiFectamine™ 293 (Life Technologies). Через 24 часа после трансфекции добавляли энхансер трансфекции ExpiFectamine и клеткам позволяли расти еще в течение 4-5 дней перед сбором. Затем собирали истощенные культуры, центрифугировали для удаления клеточного детрита, а затем пропускали через фильтр 20 мкм.
Затем очищенные кондиционированные среды, содержащие биспецифические антитела против CD3-GUCY2c, очищали с использованием аффинной хроматографии с протеином A. Образцы вводили в микроколонки 0,45 мл (Repligen), предварительно заполненные смолой MabSelect SuRe™ с протеином A (GE Healthcare) с использованием жидкостного манипулятора (Tecan). Связанный белок промывали PBS, pH 7,2, затем элюировали 20 мМ лимонной кислотой, 150 мМ хлоридом натрия, pH 3,5, и нейтрализовали 2 M Трис, pH 8,0. Затем образцы обессаливали в PBS, pH 7,2 с использованием капельных колонок G25 Sephadex (GE Healthcare) по инструкциям производителя. Очищенные белки анализировали на чистоту с использованием эксклюзионной хроматографии аналитического масштаба с помощью колонки Mab HTP (Tosoh Bioscience) и Aglient 1200 ВЭЖХ по инструкциям производителя. Концентрации определяли посредством измерения поглощения при OD280 нм с использованием микроспектрофотометра (Trinean).
Пример 6: Анализ связывания рекомбинантного белка
Белки подвергали скринингу на связывание с рекомбинантными белками человека, яванского макака и мыши с помощью ELISA. Очищенными ECD GUCY2c человека, яванского макака, мыши или белками CD3 эпсилон-дельта человека покрывали 384-луночные планшеты для ELISA Nunc-Maxisorb в количестве 1 мкг/мл в 20 мкл в PBS-CMF (без кальция и магния) при 4°C. Планшеты 3 раза промывали PBS+0,05% TWEEN20 и блокировали PBS+3% молока в течение 1 ч., встряхивали при RT. Блокирующий раствор удаляли, добавляли образцы, серийно разведенные в блокирующем растворе, и инкубировали в течение 1 ч. со встряхиванием при RT. Планшеты промывали 3 раза, как описано выше, и добавляли вторичное антитело (антитело козы против Fc IgG человека-HRP - Southern Biotech L2047-05, 1:5000; антитело козы против Fc IgG мыши-HRP - Thermo Scientific, разведенное 1:4000; или антитело мыши против пента-HIS-HRP, Qiagen, 1:1000), с последующим встряхиванием в течение 1 ч. при RT. Планшеты промывали, как описано выше, и проявляли сигнал с использованием TMB, реакцию останавливали с помощью H2SO4 и считывали поглощение при 450 нм с помощью спектрофотометра для чтения планшетов Envision (Perkin Elmer). Также периодически использовали альтернативный способ детекции вместо TMB/H2SO4. После добавления и первичной промывки добавляли 20 мкл на лунку конъюгата антитела против IgG человека с европием (Perkin Elmer EU-N1), разведенного 1:1000 в аналитическом буфере DELFIA (Perkin Elmer 1244-330) и инкубировали при RT в течение 1 ч. Затем планшеты промывали 3 раза 1-кратным промывочным раствором DELFIA (Perkin Elmer 1244-114), добавляли 20 мкл на лунку усиливающего раствора (Perkin Elmer 1244-105) и инкубировали в течение 30 минут. Затем измеряли флуоресценцию с временным разрешением при 320 и 615 нм с использованием спектрофотометра для чтения планшетов EnVision® (многоканального спектрофотометра для чтения планшетов EnVision®, Perkin Elmer) по инструкциям производителя.
Для тестирования биспецифического связывания с GUCY2c и CD3 осуществляли ELISA в сэндвич-формате. CD3 эпсилон-дельта человека (SEQ ID NO: 242) покрывали 384-луночные планшеты для ELISA Nunc-Maxisorb в количестве 1 мкг/мл в 20 мкл PBS-CMF (без кальция и магния) при 4°C. Планшеты промывали 3 раза PBS+0,05% TWEEN20 и блокировали PBS+3% молока в течение 1 ч., встряхивая при RT. Удаляли блокирующий раствор, добавляли образцы, серийно разведенные в блокирующем растворе, и инкубировали в течение 1 ч. со встряхиванием при RT. Планшеты промывали 3 раза, как описано выше, и добавляли очищенный слитый белок ECD GUCY2c человека, яванского макака или мыши-Fc IgG2a мыши в планшет в количестве 1 мкг/мл и инкубировали при RT, встряхивая в течение 1 ч. Планшеты промывали 3 раза, как описано выше, и добавляли вторичное антитело (антитело козы против Fc IgG мыши-HRP - Thermo Scientific, разведенное 1:4000) с последующим встряхиванием в течение 1 ч. при RT. Планшеты промывали, как описано выше, и проявляли сигнал с использованием TMB, реакцию останавливали с помощью H2SO4 и считывали поглощение при 450 нм с помощью спектрофотометра для чтения планшетов EnVision® (Perkin Elmer). Также периодически использовали альтернативный способ детекции вместо TMB/H2SO4. После добавления и промывки очищенного слитого белка ECD GUCY2c человека, яванского макака или мыши-Fc IgG2a мыши, добавляли 20 мкл на лунку конъюгата IgG против мыши с европием (Perkin Elmer EU-N1), разведенного 1:1000 в аналитическом буфере DELFIA (Perkin Elmer 1244-330), и инкубировали при RT в течение 1 ч. Планшеты промывали 3 раза промывочным буфером DELFIA и инкубировали в течение 30 минут с 20 мкл усиливающего раствора (Perkin Elmer 1244-105). Затем измеряли флуоресценцию с разрешением по времени при 3240 и 615 нм с использованием спектрофотометра для чтения планшетов EnVision® (Perkin Elmer) по инструкциям производителя. Химерные биспецифические антитела против CD3-GUCY2c подвергали скринингу на связывающую активность против рекомбинантной GUCY2c человека и CD3 эпсилон человека с помощью ELISA. Лучшие 6 клонов демонстрировали двойную связывающую активность в нижнем или субнаномолярном диапазоне (таблица 13).
Таблица 13
Пример 7: Опосредованная биспецифическим антителом T-клеточная активность
PBMC человека выделяли из крови здорового донора с использованием Histopaque-177 (Sigma). Наивные T-клетки выделяли из PBMC с использованием набора для обогащения T-клеток от Stem Cell Technologies (отрицательная селекция T-клеток). GUCY2c-экспрессирующие опухолевые клетки человека T84, трансфицированные с использованием экспрессирующей конструкции люциферазы, ресуспендировали в среде R10 (RPMI, 10% FBS, 1% пенициллин/стрептомицин, 3 мл 45% глюкозы). T-клетки также ресуспендировали в средах R10 и добавляли к клеткам T84 при соотношении эффектора и мишени (соотношении E:T) 10:1 или 5:1. Клетки обрабатывали серийными разведениями биспецифических антител против GUCY2c или биспецифическим антителом против CD3 отрицательного контроля и инкубировали при 37°C в течение 48 часов с последующим измерением сигнала люциферазы с использованием реагента Neolite (Perkin Elmer) с использованием устройства Victor (Perkin Elmer). Значения EC50 вычисляли с помощью Graphpad PRISM с использованием четырех-параметрического нелинейного регрессионного анализа. Мишене-отрицательные клетки HCT116 или HT29 не демонстрировали какой-либо опосредованной биспецифическим антителом против GUCY2c цитотоксичности. Результаты анализов уничтожения клеток с использованием химерных биспецифических антител против CD3-GUCY2c (соотношение E:T=10:1) приведены в таблице 14. Все биспецифические антитела против CD3-GUCY2c демонстрировали специфическое и мощное T-клеточно-зависимое уничтожение GUCY2c-положительных клеток.
Таблица 14
Результаты анализов уничтожения клеток с использованием гуманизированных биспецифических антител против CD3-GUCY2c (соотношение E:T=5:1) приведены в таблице 15. Все биспецифические антитела демонстрировали T-клеточно-зависимую цитотоксичность в отношении GUCY2c-положительных клеток.
Таблица 15
Пример 8: Биннинг эпитопов с помощью Octet
Биспецифические белки GUCY2C-0074, -0077, -0104, -0105 и -0098 оценивали на конкурентное и неконкурентное связывание с GUCY2c человека (SEQ ID NO: 224) с использованием анализа тандемного биннинга с помощью OctetRED 384 (ForteBio). Анализ Octet осуществляли при комнатной температуре. Биотинилированную GUCY2c человека разводили до 10 мкг/мл фосфатно-солевом буфере (PBS) без кальция и магния и фиксировали на биосенсорах со стрептавидином (SA) (18-5020, ForteBio) по инструкциям производителя. Биспецифические белки разбавляли до 300 нМ в PBS. Покрытые GUCY2c биосенсоры погружали в первый биспецифический белок на 300 секунд, затем погружали во второй биспецифический белок на 300 секунд. Каждое из биспецифических антител тестировали таким попарно комбинаторным способом. Биспецифические антитела, конкурирующие за одну и ту же область связывания на GUCY2c человека, группировали в одном бине. Посредством биннинга эпитопов химерных биспецифических антител против CD3-GUCY2c с помощью Octet определяли уникальные группы эпитопов, распознаваемые связывающими доменами против GUCY2c (таблица 16). Символом "+" указаны конкурирующие эпитопы; символом "+/-" указаны частично перекрывающиеся эпитопы; символом "-" указаны неконкурирующие эпитопы; и "NT" указано, что сравнительный анализ не проводили.
Таблица 16
Пример 9: Гуманизация связывающих доменом антитела мыши против GUCY2c
Гуманизацию вариабельных областей антитела мыши против GUCY2c осуществляли с использованием стратегии пересадки CDR. Комплементарную ДНК (кДНК), содержащую акцепторный каркас человека, VH3-7 (SEQ ID NO: 176) в случае тяжелой цепи и VK1-39 (SEQ ID NO: 180) в случае легкой цепи, с донорными последовательностями определяющих комплементарность областей (CDR) против GUCY2c синтезировали в векторах, содержащих константную область IgG1-3m человека или каппа-цепи человека в случае легкой цепи. Вектор тяжелой цепи состоял из области CH1 (SEQ ID NO: 185), шарнирной области (SEQ ID NO: 186) и области CH2-CH3, включающей мутации, влияющие на эффекторную функцию, L234A, L235A и G237A (SEQ ID NO: 187; нумерация в соответствии с индексом EU). Вектор легкой цепи состоял из домена CL каппа (SEQ ID NO: 184). Встраивали Обратные мутации в последовательность мыши в каркасных областях областей VH и VL и оценивали их на полное восстановление связывающей активности. Выравнивание областей VH и VL акцепторного каркаса человека, родительских клонов мыши и полностью активных гуманизированных вариантов приведено в таблицах 17A, 17B, 18A и 18B ниже для клона GUCY2C-0098.
Обратные мутации в последовательность мыши в каркасных областях областей VH и VL встраивали и оценивали на полное восстановление связывающей активности. Выравнивание областей VH и VL акцепторного каркаса человека, родительских клонов мыши и полностью активного гуманизированного варианта GUCY2C-0241 приведено в таблицах 17A и 17B ниже для клона GUCY2C-0098.
Таблица 17A
(SEQ ID NO: 5)
SYWMS (SEQ ID NO: 271) Kabat
(SEQ ID NO: 30)
SYWMH (SEQ ID NO: 259) (Kabat)
(SEQ ID NO: 5)
SYWMH (SEQ ID NO: 259) (Kabat)
Таблица 17B
KQDGSE (SEQ ID NO: 272) Chothia
KPSNGL (SEQ ID NO: 264) (Chothia)
(SEQ ID NO: 29)
KPSNGL (SEQ ID NO: 264) (Chothia)
(SEQ ID NO: 29)
Выравнивание области VL акцепторного каркаса человека, родительского клона мыши и полностью гуманизированного варианта приведено в таблицах 18A и 18B для клона GUCY2C-0098. Подчеркнуты CDR при нумерации по Kabat. Полужирным шрифтом выделены исходные, не принадлежащие человеку аминокислоты зародышевой линии, присутствующие в полностью гуманизированном клоне.
Таблица 18A
(SEQ ID NO: 80)
(SEQ ID NO: 181)
(SEQ ID NO: 81)
(SEQ ID NO: 96)
(SEQ ID NO: 107)
(SEQ ID NO: 110)
(SEQ ID NO: 126)
(SEQ ID NO: 107)
(SEQ ID NO: 81)
Таблица 18B
(SEQ ID NO: 182)
(SEQ ID NO: 82)
(SEQ ID NO: 183)
(SEQ ID NO: 108)
(SEQ ID NO: 111)
(SEQ ID NO: 109)
(SEQ ID NO: 108)
(SEQ ID NO: 82)
В таблице 19 показано, что гуманизированные GUCY2c-связывающие домены как в IgG1 человека сохраняли связывание с рекомбинантным белком GUCY2c по результатам ELISA.
Таблица 19
В таблице 20 показано, что гуманизированные GUCY2c-связывающие домены как в биспецифических антителах против CD3 сохраняли одновременное связывание с рекомбинантными белками GUCY2c человека и CD3 человека. GUCY2C-0240 является гуманизированной версией биспецифического антитела против CD3 IgG GUCY2C-0166. GUCY2C-0247 и GUCY2C-0250 являются гуманизированными биспецифическими антителами против CD3 GUCY2C-0241 с разными вариабельными областями против CD3.
Таблица 20
Пример 10: Дальнейшая гуманизация и оптимизация GUCY2C-связывающих доменов с использованием фагового дисплея
В некоторых вариантах осуществления замена является заменой зародышевой линии человека, в которой остаток CDR заменяют соответствующим остатком зародышевой линии человека для повышения содержания аминокислот человека и снижения потенциальной иммуногенности антитела. Например, если используют каркас VH3-7 человека зародышевой линии и пример антитела GUCY2C-0241, выравнивание CDR1 VH антитела GUCY2C-0241 (SEQ ID NO: 27) и VH3-7 зародышевой линии человека приведено в таблице 21 (с использованием системы нумерации Kabat с определением AbM CDR1 VH):
Таблица 21
В случае положений 26, 28, 29, 31, 32, 33 и 34 остаток зародышевой линии человека и соответствующий остаток GUCY2C-0241 являются одинаковыми, и замена зародышевой линии невозможна. В случае положений 27, 30 и 35 (выделены полужирным шрифтом и подчеркнуты) остаток зародышевой линии человека и соответствующий остаток GUCY2C-0241 отличаются. Остатки GUCY2C-0241 в этих положениях можно заменять соответствующим остатком VH3-7 зародышевой линии человека для дополнительного повышения содержания остатков человека.
Антитело его или антигенсвязывающий фрагмент может содержать каркас VH, содержащий последовательность каркаса VH зародышевой линии человека. Последовательность каркаса VH можно получать из VH3 зародышевой линии человека, VH1 зародышевой линии, VH5 зародышевой линии или VH4 зародышевой линии. Предпочтительными каркасами тяжелой цепи зародышевой линии человека являются каркасы, полученные из VH1, VH3 или VH5 зародышевой линии. Например, можно использовать каркасы VH из следующих зародышевых линий: IGHV3-23, IGHV3-7 или IGHV1-69 (названия зародышевых линий основаны на определении зародышевой линии IMGT). Предпочтительными каркасами легкой цепи зародышевой линии человека являются каркасы, полученные из Vκ или Vλ зародышевой линии. Например, можно использовать каркасы VL из следующих зародышевых линий: IGKV1-39 или IGKV3-20 (названия зародышевых линий основаны на определении зародышевой линии IMGT). Альтернативно или дополнительно, каркасная последовательность может являться консенсусной каркасной последовательностью зародышевой линии человека, такой как каркас консенсусной последовательности Vλ1 человека, консенсусной последовательности Vκ1, консенсусной последовательности Vκ2, консенсусной последовательности Vκ3, консенсусной последовательности зародышевой линии VH3, консенсусной последовательности зародышевой линии VH1, консенсусной последовательности зародышевой линии VH5 или консенсусной последовательности зародышевой линии VH4. Последовательности каркасов зародышевой линии человека доступны в различных общедоступных базах данных, таких как V-base, IMGT, NCBI или Abysis.
Антитело его или антигенсвязывающий фрагмент может содержать каркас VL, содержащий последовательность каркаса VL зародышевой линии человека. Каркас VL может содержать одну или более замен, добавлений или делеций аминокислот при сохранении функционального и структурного сходства с зародышевой линией, из которой его получают. В некоторых аспектах каркас VL является по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентичным каркасной последовательности VL зародышевой линии человека. В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит каркас VL, содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 замен, добавлений или делеций аминокислот относительно последовательности каркаса VL зародышевой линии человека.
Каркас VL зародышевой линии человек может являться каркасом DPK9 (название IMGT: IGKV1-39). Каркас VL зародышевой линии человека может являться каркасом DPK12 (название IMGT: IGKV2D-29). Каркас VL зародышевой линии человека может являться каркасом DPK18 (название IMGT: IGKV2-30). Каркас VL зародышевой линии человека может являться каркасом DPK24 (название IMGT: IGKV4-1). Каркас VL зародышевой линии человека может являться каркасом HK102_V1 (название IMGT: IGKV1-5). Каркас VL зародышевой линии человека может являться каркасом DPK1 (название IMGT: IGKV1-33). Каркас VL зародышевой линии человека может являться каркасом DPK8 (название IMGT: IGKV1-9). Каркас VL зародышевой линии человека может являться каркасом DPK3 (название IMGT: IGKV1-6).Каркас VL зародышевой линии человека может являться каркасом DPK21 (название IMGT: IGKV3-15). Каркас VL зародышевой линии человека может являться каркасом Vg_38K (название IMGT: IGKV3-11). Каркас VL зародышевой линии человека может являться каркасом DPK22 (название IMGT: IGKV3-20). Каркас VL зародышевой линии человека может являться каркасом DPK15 (название IMGT: IGKV2-28). Каркас VL зародышевой линии человека может являться каркасом DPL16 (название IMGT: IGLV3-19). Каркас VL зародышевой линии человека может являться каркасом DPL8 (название IMGT: IGLV1-40). Каркас VL зародышевой линии человека может являться каркасом V1-22 (название IMGT: IGLV6-57). Каркас VL зародышевой линии человека может являться консенсусной последовательностью каркаса Vλ человека. Каркас VL зародышевой линии человека может являться консенсусной последовательностью каркаса Vλ1 человека. Каркас VL зародышевой линии человека может являться консенсусной последовательностью каркаса Vλ3 человека. Каркас VL зародышевой линии человека может являться консенсусной последовательностью каркаса Vκ человека. Каркас VL зародышевой линии человека может являться консенсусной последовательностью каркаса Vκ1 человека. Каркас VL зародышевой линии человека может являться консенсусной последовательностью каркаса Vκ2 человека. Каркас VL зародышевой линии человека может являться консенсусной последовательностью каркаса Vκ3 человека.
Определение паратопов антител против GUCY2c посредством расширенной бинарной замены
Описан способ определения критических остатков CDR, с помощью которого выбирают функциональные варианты антител из библиотеки, содержащей остаток зародышевой линии человека или соответствующий остаток грызуна в каждом положении CDR, за исключением CDR-H3 (Townsend et al., 2015, Proc. Natl. Acad. Sci. USA. 112(50):15354-15359). Конструировали фаговую библиотеку scFv, в которой все положения CDR, за исключением CDR3 VH GUCY2C-0241, отличающейся от зародышевой линии человека (VH3-7/VK1-39), рандомизировали так, чтобы приблизительно 50% клонов кодировали аминокислоту GUCY2C-0241 мыши и приблизительно 50% кодировали аминокислоту зародышевой линии человека в этом положении (таблицы 17A, 17B, 18A и 18B). Библиотеки получали и подвергали 3-4 раундам селекции по GUCY2c человека (SEQ ID NO: 232), как описано ниже. Клоны, сохранявшие связывание с GUCY2c, выделяли и определяли их последовательности.
В этом эксперименте положения, в которых последовательность человека наблюдали у менее чем приблизительно 20% связывающих клонов, определяли как необходимые остатки мыши. Остатки мыши преимущественно сохранялись в 25 положениях (остатки тяжелой цепи H35, E50, P52a, S53, N54, G55, L56, T57, N58, I60, E61, K62, F63 и N65; остатки легкой цепи E27, V29, D30, L32, M33, Q34, N53, E55, T91, R92 и K93 в соответствии с системой нумерации Kabat, и определением AbM использовали для CDR1 VH; систему нумерации Chothia использовали для CDR1 VL), что свидетельствует о том, что замена этих остатков остатками зародышевой линии человека является крайне неблагоприятной. Остатки мыши и человека обнаруживали со схожей частотой (определяемой как содержание остатков человека более 20%) в остатках тяжелой цепи 27 (F/Y) и 30 (S/T) и остатках легкой цепи 54 (L/V) и 94 (A/V; в соответствии с системой нумерации Kabat; определение AbM использовали для CDR1 VH), что свидетельствует о том, что последовательность мыши в этих положениях не является критической.
Кроме того, в стратегию расширенной бинарной замены включали остатки каркаса (FW), ранее требовавшие обратной мутации в аминокислоты мыши (положения тяжелой цепи I48 и G49 в соответствии с нумерацией по Kabat). После селекции и скрининга фага аминокислотную последовательность человека для положения каркаса тяжелой цепи 48 определяли в приблизительно 10% случаев, что позволяет предполагать, что обратная мутация является предпочтительной. Положение тяжелой цепи 49 содержало аминокислотный остаток мыши в 100% случаев, что позволяет предполагать, что эта обратная мутация является необходимой. Частота аминокислотной последовательности мыши или человека в положениях CDR или FW после расширенной бинарной замены приведена в таблице 22. В таблице 22 показана частота включения аминокислот человека в положениях CDR GUCY2C-0098, тестируемых посредством расширенной бинарной замены (с использованием системы нумерации Kabat, и для CDR1 VH использовали определение AbM; для CDR1 VL использовали систему нумерации Chothia).
Таблица 22
Дальнейшая оптимизация связывающих доменов после GUCY2c
Варианты антител против GUCY2c с улучшенной стабильностью выделяли следующими способами. Области VH и VL гуманизированного клона GUCY2C-0241 (SEQ ID NO: 60 и 137) синтезировали в виде одноцепочечного вариабельного фрагмента (scFv) в фаговом экспрессирующем векторе, содержащем C-концевые метки His6 и c-myc (Blue Heron). Встраивали случайные мутации в домены CDR VH и VL посредством "мягкой" рандомизации NNK (способами, ранее описанными в патенте США № 9884921). Олигонуклеотиды, сконструированные для селективной мутации одного остатка CDR за один раз, синтезировал поставщик (IDT), а затем их использовали для внесения изменений в матрицу нуклеиновой кислоты ранее описанными способами (Townsend et al., 2015, Proc. Natl. Acad. Sci. USA. 112(50):15354-15359). В дизайн олигонуклеотидов также включают мутацию, сконструированную для устранения потенциального участка посттрансляционной модификации (G55E тяжелой цепи), в рамках которой удаляют потенциальный участок дезамидирования аспарагина NG (Chelius et al., Anal. Chem. 77(18):6004-6011, 2005). Мутантные клоны выбирали и отправляли на секвенирование для оценки частоты мутации. Затем эти фаговые экспрессирующие векторы, содержащие подвергнутые мутагенезу варианты CDR VH и VL GUCY2C-0241 (SEQ ID NO: 60 и 137), объединяли в небольшие подбиблиотеки для фагового дисплея.
Рекомбинантные белки GUCY2c человека, яванского макака и мыши использовали на стадиях селекции и последующего скрининга (SEQ ID NO: 230, 232, 234, 236, 238 и 240). Для селекции, включающей биотинилированные белки, аликвоты описанных выше совокупностей фагов и магнитные частицы с стрептавидином (Dynabeads M-280 с стрептавидином) блокировали раздельно в 3% молоке/PBS в течение 1 часа при комнатной температуре, 60°C или 65°C в ротационном смесителе (20 об/мин). После инкубации при разных температурах блокированные совокупности фагов центрифугировали при 4000 об/мин в течение 10 минут, супернатанты переносили в новые пробирки и смешивали с 6% блокирующим буфером. Блокированных фагов инкубировали с избыточными молярными концентрациями реагентов для деселекции (т.е. стрептавидиновых частиц, несодержащих мишень, содержащих двухцепочечную ДНК, инсулин и/или реагенты экстракта мембран), инкубировали при комнатной температуре в течение 1 часа на роторном шейкере (20 об/мин), смешивали с блокированными магнитными частицами с стрептавидином и инкубировали еще в течение часа. Подвергнутую деселекции библиотеку собирали посредством осаждения частиц с использованием магнитного сепаратора. Используя устройство для магнитной очистки Kingfisher (ThermoFisher, Springfield NJ, USA) биотинилированный антиген для селекции (в различных концентрациях, указанных в таблице 23) инкубировали с подвергнутой деселекции фаговой библиотеки в 96-луночном планшете с глубокими лунками в течение 1 часа при комнатной температуре с периодическим смешиванием. Частицы разделяли с использованием магнитного сепаратора на устройстве Kingfisher и промывали 4 раза PBS/0,1% Tween20 и 3 раза PBS в отдельных 96-луночных планшетах с глубокими лунками. Связанного фага элюировали посредством инкубации с 100 мМ триэтиламина (TEA) в течение 8 минут в новом 96-луночном планшете с последующим отделением от магнитных частиц. Элюированного фага нейтрализовали с помощью 2 M Трис-HCl, pH 7,5. Пример условий селекции фага для оптимизации связывающего домена антитела против GUCY2c приведены в таблице 23. В таблице 23 также показаны условия селекции, используемые для фагового дисплея. "Dslxn" означает используемые условия деселекции. SA=стрептавидин; D=ДНК; I=инсулин; M=белки мембранного экстракта. "Therm" соответствует термической инкубации, осуществляемой перед инкубацией с мишенью.
Таблица 23
Элюированного фага использовали для инфицирования 10 мл культуры E. coli ER2738, выращенной до средне-логарифмической фазы (соответствующей OD600~0,5). Бактерии инфицировали фагом в течение 1 часа при 37°C со встряхиванием при 150 об/мин, концентрировали после стадии центрифугирования и высевали на чашки с 2-кратным YT-агаром для биологических анализов, содержащих 2% глюкозу и 100 мкг/мл ампициллина (2-кратный YTAG). Для определения титров фага различные разведения культуры E. coli, инфицированных исходным или выходным фагом, также высевали на 2-кратный агар YTAG. После выращивания в течение ночи при 30°C добавляли 10 мл 2-кратной среды YTAG в каждый планшет для биологического анализа и клетки ресуспендировали, соскребая бактериальный "газон". К этой суспензии клеток добавляли глицерин для достижения конечной концентрации 17% и хранили в аликвотах при -80°C до дальнейшего использования. Для "спасения" фага для следующего раунда селекции 100 мкл этой суспензии клеток использовали для инокуляции 20 мл 2-кратной среды YTAG, выращивали при 37°C (300 об/мин) до OD600 0,3-0,5. Затем клетки суперинфицировали с помощью 3,3 мкл хелперного фага MK13K07 и инкубировали при 37°C (150 об/мин) в течение 1 часа. Затем клетки центрифугировали и ресуспендировали осадок в содержащей канамицин/несодержащей глюкозу среде (2-кратной YT с 50 мкг/мл канамицина и 100 мкг/мл ампициллина). Эту культуру выращивали в течение ночи при 30°C (300 об/мин). Фаг собирали в супернатанте после центрифугирования, и он был готов для использования в следующем раунде селекции, как описано выше.
Получение необработанного периплазматического материала для применения в ELISA
ScFv можно экспрессировать на поверхности фаговой частицы или в растворе в бактериальном периплазматическом пространстве в зависимости от используемых условий выращивания. Для индуцирования высвобождения scFv в периплазму на 96-луночные планшеты с глубокими лунками, содержащие 2-кратные среды YT с 0,1% глюкозой/100 мкг/мл ампициллина, инокулировали материал из размороженных глицериновых стоков (один клон на лунку) с использованием устройства-сборщика колоний QPix (Molecular Devices) и выращивали при 37°C (999 об/мин) в течение ~4 часов. Культуры индуцировали с использованием IPTG в конечной концентрации 0,02 мМ и выращивали в течение ночи при 30°C (999 об/мин). Содержимое бактериальной периплазмы (PeriPreps) высвобождали с помощью осмотического шока. В кратком изложении, планшеты центрифугировали и осадки ресуспендировали в 150 мкл периплазматического буфера TES (50 мМ Трис, 1 мМ ЭДТА, 20% сахарозы, pH7,4) с последующим добавлением 150 мкл 1:5 TES:воды и инкубировали на льду в течение 30 минут. Планшеты центрифугировали в течение 20 минут при 4000 об/мин и собирали scFv-содержащий супернатант.
Конкурентный ELISA
Планшеты для ELISA (NUNC-Maxisorb 460372) покрывали 1 мкг/мл интересующего антигена в буфере PBS в течение ночи при 4°C. Покрывающий раствор выбрасывали, а затем планшеты блокировали при комнатной температуре в течение 2 часов с помощью 70 мкл PBS-3% BSA на лунку. Блокирующий раствор выбрасывали и добавляли 20 мкл первичного антитела, представлявшего собой смесь неизвестного образца в виде PeriPreps или очищенного белка (в заранее определенном разведении, как правило, 1:5) и чистого родительского антитела в желаемой концентрации (как правило, EC50-EC80). Планшеты инкубировали в течение 2 часов при комнатной температуре с медленным встряхиванием. Планшеты промывали 5 раз 100 мкл на лунку промывочного буфера (Perkin Elmer 1244-114) и инкубировали в течение 1 часа с 20 мкл конъюгата вторичного антитела с европием, разведенного 1:1000 в аналитическом буфере Delfia (Perkin Elmer 1244-111) в случае стрептавидин-Eu или антитела против IgG человека, разведенного 1:500, в случае антитела против IgG мыши. Планшеты промывали 5 раз, как описано выше, и инкубировали в течение 30 минут с 20 мкл усиливающего раствора (Perkin Elmer 1244-105). Затем измеряли флуоресценцию с разрешением по времени при 320 и 615 нм с использованием спектрофотометра для чтения планшетов EnVision® (EnVision® многоканального спектрофотометра для чтения планшетов, Perkin Elmer) по инструкциям производителя.
Преобразование ScFv в scFv-Fc, IgG и биспецифическое антитело
ScFv, демонстрирующее желаемые свойства, выбирали для субклонирования в scFv-Fc, где Fc-домен IgG1 человека присоединяли к 3'-концу scFv. В кратком изложении, фрагменты амплифицировали посредством стандартной полимеразной цепной реакции (ПЦР) с праймерами BssHII в случае 5’-амплификации и BclI в случае 3’-амплификации. Фрагменты разрезали соответствующими ферментами рестрикции по инструкциям производителя (New England Biolabs). Ампликоны scFv-Fc против GUCY2c выделяли из геля (набор для очистки из геля Qiagen) и лигировали в предварительно расщепленный экспрессирующий вектор млекопитающего, являющийся собственностью Pfizer, содержащий Fc-домен IgG1 человека. С помощью конструкций ScFv-Fc трансформировали E. coli и секвенировали, как описано выше. Затем экспрессирующие векторы с проверенной последовательностью использовали для транзиторной экспрессии в клетках млекопитающих HEK 293, как описано выше. Образцы очищали с использованием аффинной хроматографии с протеином A с последующей заменой буфера на PBS, как описано выше. Затем очищенные образцы подвергали скринингу на связывающую активность в отношении рекомбинантного белка GUCY2c и стабильность. Примером последовательности scFv-Fc является GUCY2C-0405 (SEQ ID NO: 213). Этот формат состоит из scFv в ориентации VH-VL, соединенных линкером 4, являющимся глицин-сериновым линкером (SEQ ID NO: 193). VH-VL соединяют с модифицированным Fc-доменом IgG1 человека (SEQ ID NO: 187) с использованием двух линкерных последовательностей, линкера 5 (SEQ ID NO: 194) и линкера 6 (SEQ ID NO: 195).
Оптимизированные scFv также субклонировали в IgG1 человека и T-клеточные биспецифические форматы, как описано выше. Праймеры, специфические для генов вариабельных областей, конструировали для клонирования ITA, как описано ранее.
Пример 11: Поверхностный плазмонный резонанс оптимизированных антител против GUCY2c
Оптимизированные антитела против GUCY2c оценивали на связывание посредством поверхностного плазмонного резонанса с использованием устройства BIACORE™ 8K (GE Healthcare). Используя сенсорный чип CM5, сначала наносили антитело против Fc человека (GE BR-1008-39) по инструкциям производителя. IgG человека против GUCY2c поливали чип в течение 30 сек при 0,5 мкг/мл, 10 мкл/мин в забуференном физиологическом растворе Хэнкса (HBS) -EP+ pH=7,4. Затем белку GUCY2c человека (SEQ ID NO: 224) в концентрации 450 нМ, 150 нМ и 50 нМ позволяли ассоциироваться, поливая им чип в течение 54 сек при 50 мкл/мин, а затем диссоциировать в течение 300 сек. Чип регенерировали после каждого прогона с помощью 3-кратного 3M MgCl2 в течение 30 сек при 5 BIACORE™. Аффинность связывания (KD, нМ) оптимизированных связывающих доменов против GUCY2c в формате IgG1 человека с рекомбинантным белком GUCY2c человека приведены в таблице 24. Несколько оптимизированных связывающих доменов против GUCY2c, таких как GUCY2C-0486 (SEQ ID NO: 214 и 215), демонстрировали улучшенную аффинность связывания по сравнению с родительским, неоптимизированным связывающим доменом (GUCY2C-0241).
Таблица 24
Пример 12: Анализ связывания способом термической проверки
Для оценки улучшенной термической стабильности оптимизированных вариантов, периплазматические препараты оптимизированных фаговых клонов (описанных выше) или очищенные белки оценивали на связывающую активность после термической проверки при повышенных температурах в устройстве для ПЦР. 384-луночные белые планшеты для ELISA Greiner bio-one (781074) покрывали 1 мкг/мл целевого белка покрывающего буфера (25 мМ Na2CO3, 75 мМ NaHCO3, pH 9,6) O/N при 4°C. Планшеты промывали 3 раза PBS+0,05% TWEEN20, а затем блокировали в течение 1 ч. при встряхивании при RT с блокирующим буфером (PBS+3% молока). Отдельно подготавливали планшеты для разведения периплазматических препаратов или очищенного белка при связывании 1:10 периплазматических препаратов или 1 мкг/мл (EC80) рекомбинантного белка при комнатной температуре в блокирующем буфере и 100 мкл переносили в 96-луночный планшет для ПЦР. Затем планшет для ПЦР, содержащий 100 мкл разведенного образца, нагревали при желаемых температурах (как правило, от 55°C до 75°C) в течение 30 минут. Планшетам позволяли охлаждаться в течение 15 минут при комнатной температуре, а затем центрифугировали при 4000 об/мин в течение 10 минут. Блокированные планшеты для ELISA опустошали и добавляли 20 мкл термически проверенного белкового супернатанта на лунку, затем инкубировали в течение 1 ч., встряхивали при RT. Планшеты промывали 3 раза, как описано выше, и добавляли 20 мкл реагента белка против 6xHIS-европий (Perkin Elmer EU-N1) или реагента белка конъюгата антитела против IgG человека и европия (Perkin Elmer EU-N1), разведенного 1:1000 в аналитическом буфере DELFIA (Perkin Elmer 1244-330), на лунку и инкубировали при RT в течение 1 ч. со встряхиванием. Планшеты промывали 3 раза 1-кратным промывочным раствором DELFIA (Perkin Elmer 1244-114), в каждую лунку добавляли 20 мкл усиливающего раствора (Perkin Elmer 1244-105) и инкубировали в течение 30 минут. Планшеты считывали на спектрофотометре для чтения планшетов Envision для определения флуоресценции с разрешением по времени сначала при OD320, затем при OD615 по инструкциям производителя. Температуру, при которой сохранялось 50% активности, регистрировали как T50. При термическом анализе связывания на стабильность оптимизированных GUCY2c-связывающих доменов как scFv-Fc наблюдали улучшенную термическую стабильность после инкубации при повышенных температурах по сравнению с неоптимизированным родительским клоном GUCY2C-0241 (обозначенным как GUCY2C-0295 в формате scFv-Fc) (таблицы 25A и 25B).
В таблицах 25A и 25B показана флуоресценция с разрешением по времени (TRF) для оптимизированных GUCY2c-связывающих доменов (в виде белков scFv-Fc) при повышенной температуре. Значение T50 представляет собой температуру, при которой клон сохраняет 50% связывающей активности.
Таблица 25A
Таблица 25B
Аминокислотные последовательности клонов антитела против GUCY2c, демонстрирующие улучшенную термическую стабильность, сравнивали с родительским клоном GUCY2C-0241 (VH SEQ ID NO: 60 и VL SEQ ID NO: 137). Изменения содержания аминокислот в CDR VH, приводящие к улучшенной стабильности, приведены в таблице 26A. Изменения в положениях 59, 60, 61 и 62, по-видимому, оказывают наибольшее влияние на стабильность, коррелируя с наибольшими изменениями стабильности (в соответствии с системой нумерации Kabat с использованием определения AbM для CDR1 VH).
Таблица 26A
Изменения содержания аминокислот в CDR VL, приводящие к улучшенной стабильности, приведены в таблице 26B (использовали систему нумерации Chothia для CDR1 VL и систему нумерации Kabat для CDR2 VL и CDR3). Изменения в положениях 53, 54, 55 в CDR2 VL и положениях 93 и 94 в CDR3 VL, по-видимому, оказывают наибольшее влияние на стабильность, коррелируя с наибольшими изменениями стабильности.
Таблица 26B
Пример 13: Связывание рекомбинантного белка оптимизированными биспецифическими антителами против CD3-GUCY2c
Оптимизированные связывающие домены против GUCY2c оценивали в биспецифическом формате на одновременное связывание рекомбинантного белка с GUCY2c человека (SEQ ID NO: 224) и CD3 человека (SEQ ID NO: 242) при комнатной температуре и 60°C способом DELFIA, описанным выше. Оптимизированные биспецифические антитела также оценивали на прямое связывание с GUCY2c человека (SEQ ID NO: 232) и GUCY2c яванского макака (SEQ ID NO: 236) при комнатной температуре и 60°C способом DELFIA, описанным выше. Двойное связывание оптимизированных биспецифических антител против GUCY2c с GUCY2c человека и CD3 человека приведены в таблице 27. Оптимизированные биспецифические антитела против CD3-GUCY2c, такие как GUCY2C-1478, демонстрировали сильное связывание при комнатной температуре и 60°C.
Таблица 27
Результаты анализа прямого связывания оптимизированных биспецифических антител против GUCY2c с GUCY2c человека и яванского макака приведены в таблице 28. Оптимизированные биспецифические антитела против CD3-GUCY2C, такие как GUCY2C-1478, демонстрировали сильное связывание при комнатной температуре с GUCY2c человека и яванского макака, но при этом наблюдали некоторую потерю сигнала связывания после инкубации при 60°C.
Таблица 28
Пример 14: Связывание клеток биспецифическими антителами против CD3-GUCY2c по результатам проточной цитометрии
Оптимизированные биспецифические антитела против CD3 титровали на связывание поверхности клетки с GUCY2c на клетках T84 и с CD3 человека на наивных T-клетках человека, выделенных из свежих мононуклеарных клеток периферической крови человека (PBMC), описанными выше способами проточной цитометрии. Гуманизированные и оптимизированные клоны против GUCY2c GUCY2C-0247 и GUCY2C-1608 демонстрировали аффинность связывания с клетками T84, экспрессирующими GUCY2c человека, в нижнем и субнаномолярном диапазоне (фигуры 2A и 2B). Гуманизированные и оптимизированные биспецифические антитела против GUCY2c также оценивали на связывание с CD3 на поверхности клетки посредством проточной цитометрии (фигуры 3A и 3B) и наблюдали аффинность связывания с наивными T-клетками человека в нижнем или среднем наномолярном диапазоне.
Пример 15: Уменьшение риска иммуногенности биспецифических антител против GUCY2c
Если некоторые терапевтические белки вводят пациентам, нежелательные иммунные ответы, такие как образование антител против лекарственных средств (ADA), влияют на эффективность лекарственного средства и вызывает проблемы в отношении безопасности для пациента (Yin L., et al., Cell Immunol. Jun; 295(2):118-26, 2015). ADA можно классифицировать на две группы: нейтрализующие ADA (NAb) или ненейтрализующие ADA (не-NAb) в зависимости от того, ингибируют ли они фармакологическую активность терапевтического белка (Yin L., et al., Cell Immunol. Jun; 295(2):118-26, 2015). Существует два возможных механизма, посредством которых NAb и не-NAb могут вносить вклад в снижение эффективности лекарственного средства. Во-первых, NAb напрямую блокируют связывание терапевтического белка с молекулой-мишенью, таким образом, снижая ее терапевтическую эффективность (Yin L., et al., Cell Immunol. Jun; 295(2):118-26, 2015). Во-вторых, NAb и не-NAb могут вносить вклад в повышенный клиренс, влияя на фармакокинетику (PK) TPP, таким образом, препятствуя эффективности лекарственного средства (Yin L., et al., Cell Immunol. Jun; 295(2):118-26, 2015). Иммуногенность в отношении терапевтического белка может генерироваться в рамках T-клеточно-зависимого и T-клеточно-независимого путей. В T-клеточно-зависимом пути T-клетки активируются посредством распознавания образующихся из терапевтического белка антигенных пептидов, презентируемых молекулами главного комплекса гистосовместимости класса II (MHC II) в антигенпрезентирующих клетках. Затем активированные T-клетки стимулируют B-клетки так, что они образуют ADA, специфические в отношении терапевтического белка (Yin L., et al., Cell Immunol. Jun; 295(2):118-26, 2015). Для минимизации вероятности иммуногенности, возникающей в результате T-клеточно-зависимых ответов, предпринимались попытки снизить антигенный потенциал последовательностей биспецифических антител против CD3-GUCY2c способами in silico, описанными ниже.
Последовательности анализировали двумя способами (подробно описанными ниже) для идентификации потенциальных T-клеточных эпитопов. Любую последовательность, отмеченную по правилам, представленным в настоящем описании для любого способа, считали эпитопом. В настоящем описании исследуют последовательности, главным образом, на уровне аминокислотных 9-меров.
Способ 1: Последовательности подвергают анализу EpiMatrix в пакете программ ISPRI (ISPRI v 1.8.0, EpiVax Inc., Providence, RI (2017); Schafer JRA, Jesdale BM, George JA, Kouttab NM, De Groot AS. Prediction of well-conserved HIV-1 ligands using a matrix-based algorithm, EpiMatrix. Vaccine 16(19), 1880-84, 1998). Необработанные результаты представляют собой ранжирование вероятности связывания каждого 9-мерного аминокислотного фрагмента относительно 8 разных типов HLA. Таким образом, существует 8 предсказаний ("наблюдений") для каждого 9-мера. 9-меры получают, начиная с каждого индивидуального положения линейной нумерации последовательности (таким образом, один и тот же 9-мер может встречаться более одного раза в одной и той же последовательности). Если любые 4 наблюдения свидетельствуют о том, что 9-мер входит в лучшие 5% связывающих средств (что означает, что согласно предсказанию он будет в лучших 5% связывающих средств для по меньшей мере 4 типов HLA), 9-мер считают прогнозируемым эпитопом ("эпитоп"). Альтернативно, если любое 1 из 8 предсказаний свидетельствует о том, что 9- мер входит в лучший 1% связывающих средств, 9-мер также считают прогнозируемым эпитопом.
Способ 2: Последовательности подвергали анализу с использованием консенсусного способа связывания MHC-II (Wang P, Sidney J, Kim Y, Sette A, Lund O, Nielsen M, Peters B. Peptide binding predictions for HLA DR, DP и DQ molecules. BMC Bioinformatics 11:568, 2010; Wang P, Sidney J, Dow C, Mothé B, Sette A, Peters B. A systematic assessment of MHC class II peptide binding predictions and evaluation of a консенсусн approach. PLoS Comput Biol. 4(4),e1000048, 2008in IEDB (Vita R, Overton JA, Greenbaum JA, Ponomarenko J, Clark JD, Cantrell JR, Wheeler DK, Gabbard JL, Hix D, Sette A, Peters B. The immune epitope database (IEDB) 3,0. Nucleic Acids Res. Jan 28 (43), D405-12, 2015; IEDB MHC-II Binding Predictions, http://www.iedb.org).
Выходные данные программного обеспечения распределяют результаты по 15-мерам. Получают консенсусные баллы и ранжирование процентилей для каждой комбинации 15-мера и типа HLA. Но индивидуальные баллы, из которых получают консенсусные результаты для каждого 15-мера, представляют собой ранжирование некоторых 9-меров, обнаруживаемых в 15-мере: в каждом способе, используемом для получения консенсусных результатов, отчет предоставляется в виде ранга процентили для 9-мера в 15-мере, и консенсусные баллы, принятые за значение для общего 15-мера, представляет собой предсказание для 9-мера, имеющего медианные баллы. Авторы настоящего изобретения классифицировали 9-мер как эпитоп, если (a) его выбирают как консенсусный типичный результат для 15-мера, и (b) он имеет ранг процентили в лучших 10% связывающих средств для рассматриваемого типа HLA, и если критерии (a) и (b) совпадают для трех или более отдельных типов HLA для одного и того же 9-мера (т.е. три наблюдения). Рассматриваемые типы HLA представляли собой DRB1*01, 1*03, 1*04, 1*07, 1*08, 1*11, 1*13 и 1*15, являющиеся теми же типами HLA в стандартном отчете ISPRI/EpiMatrix. Таким образом, хотя первичными выходными данными способа является ранжирование 15-мер, авторы настоящего изобретения по-новому истолковывают данные для получения списка прогнозируемых 9-мерных эпитопов для простоты сравнения со способа 1.
Авторы настоящего изобретения классифицировали каждый эпитоп как эпитоп зародышевой линии или не-зародышевой линии. В случае антител авторы настоящего изобретения дополнительно классифицировали каждый эпитоп с учетом его локализации в антителе (CDR или не-CDR). Авторы настоящего изобретения фильтровали последовательности V-доменов человека, полученные из IMGT (www.imgt.org), для удаления зародышевых линий, аннотированных как псевдогены или открытые рамки считывания (ORF). Любой прогнозируемый 9-мерный эпитоп, обнаруживаемый в остальных последовательностях, считали эпитопом зародышевой линии. Эпитопы, обнаруживаемые в областях J или C (включая IgG1, IgG2, IgG3 и IgG4) или соединениях между этими областями, также классифицировали как эпитопы зародышевой линии. Иными словами, эпитоп классифицируют как эпитоп не-зародышевой линии. Определения CDR основаны на системе нумерации Kabat (Kabat EA, Wu TT, Perry HM, Gottesman KS, Foeller C. Sequences of Proteins of Immunological Interest, US Department of Health and Human Services, NIH Publication No. 91-3242, 1991), где CDR определяют как включающие следующие остатки: CDR1 VH (H26-H35, включая инсерции, такие как H35A, до H36, но не включая его), CDR2 VH (H50-H65 включительно), CDR3 VH (H95-H102 включительно), CDR1 VL (L24-L34 включительно), CDR2 VL (L54-L56, включительно), CDR3 VL (L89-L97, включительно). Прогнозируемый 9-мерный эпитоп является эпитопом CDR, если любая из этих аминокислот является частью области CDR. Необходимо отметить, что выбранное авторами настоящего изобретения исходное положение (H26) для CDR1 VH отличается от описанного в некоторых других публикациях при использовании аннотации по Kabat.
В случае отдельной цепи или пары доменов VH и VL антитела общие баллы можно вычислять, суммируя каждый из 9-меров следующим образом. Исследуют все отдельные комбинации 9-мера и типа HLA ("наблюдения"), независимо от того, является ли 9-мер эпитопом. Если конкретное наблюдение свидетельствует о том, что пептид входит в лучшие 5% связывающих средств для казанного типа HLA, Z-баллы EpiMatrix для этого наблюдения добавляют к промежуточной сумме, ассоциированной со всей последовательностью белка. Также регистрируют общее количество исследуемых наблюдений. Единственным исключением является то, что все наблюдения, касающиеся 9-меров, идентифицированных посредством ISPRI как "T-регитопы", считают имеющими баллы EpiMatrix, равные нулю. В промежуточной сумме исходные баллы 0,05*2,2248 вычитают из каждого наблюдения (включая T-регитопы). Конечные баллы вычисляют следующим образом:
Скорректированные баллы T-reg = (промежуточная сумма)*1000/(количество наблюдений)
Более низкие баллы свидетельствуют о более низком прогнозируемом иммуногенном потенциале.
Баллы включают только прогнозирование с помощью ISPRI/EpiMatrix и не включают информацию, полученную с помощью IEDB. Таким образом, любые сильные HLA-связывающие средства, прогнозируемые с помощью IEDB, но не ISPRI, не вносят вклад в баллы. Теоретически, последовательности могут содержать множество IEDB-прогнозируемых HLA-связывающих средств и все равно иметь благоприятные скорректированные баллы T-reg, если с помощью EpiMatrix также не прогнозируют те же последовательности как вероятные связывающие средства.
Выравнивание неоптимизированных и оптимизированных областей VH и VL против GUCY2c с зародышевыми линиями. Остатки, выделенные полужирным шрифтом, представляют собой горячие точки T-клеточного эпитопа, определяемые описанными выше способами in silico. В некоторых вариантах осуществления замена является заменой зародышевой линии человека, при которой остаток CDR заменяют соответствующим остатком зародышевой линии человека для повышения содержания аминокислот человека и снижения потенциальной иммуногенности антитела (как описано выше). В других вариантах осуществления замена представляет собой другую аминокислоту, снижающую потенциальную иммуногенность антитела и не мешающую связывающим свойствам антитела.
Например, если каркас VH3-7 зародышевой линии человека используют с примером антитела GUCY2C-0241, то выравнивание CDR2 VH GUCY2C-0241 (SEQ ID NO: 28) и VH3-7 зародышевой линии человека (SEQ ID NO: 178) является таким, как представлено в таблице 29 ниже (с использованием системы нумерации Kabat):
Таблица 29
В случае положений 51, 52, 59 и 64 остаток зародышевой линии человека и соответствующий остаток GUCY2C-0241 являются одинаковыми, и замена зародышевой линии невозможна. В случае положений 50, 52A, 53, 54, 55, 56, 57, 58, 60, 61, 62, 63 и 65 (выделены полужирным шрифтом), остаток зародышевой линии человека и соответствующий остаток GUCY2C-0241 отличаются. Если анализу T-клеточных эпитопов in silico подвергают последовательность GUCY2C-0241, идентифицируют потенциальные пептидные последовательности (подчеркнуты). Затем их дополнительно анализируют на содержимое непринадлежащей человеку зародышевой линии с баллами T-клеточных эпитопов >10 и остатки непринадлежащей человеку зародышевой линии используют для мутагенеза (показано выше полужирным шрифтом и подчеркнуто). Используя мутагенез с "мягкой" рандомизацией (способами, ранее описанными в патенте США № 9884921), эти остатки подвергали мутагенезу и тестировали на сохранение связывающей активности и сохранение стабильности.
Пример 16: Активность опосредованного T-клетками уничтожения клеток оптимизированными биспецифическими антителами против CD3-GUCY2c
Оптимизированные биспецифические антитела против CD3-GUCY2c оценивали на опосредованную T-клетками цитотоксичность в отношении опухолевых клеток, как описано ранее. Оптимизированные биспецифические антитела против GUCY2c демонстрировали улучшенное эффективное опосредованное T-клетками уничтожение опухолевых клеток T84. Результаты анализов уничтожения клеток с использованием гуманизированного биспецифического антитела против CD3-GUCY2c (соотношение E:T=5:1) приведены в таблице 30 и на фигурах 4A-4D.
Таблица 30
Пример 17: Анализ дифференциальной сканирующей калориметрии (DSC)
Белки разводили в фосфатно-солевом буфере (PBS) до 0,6 мг/мл в объеме 400 мкл. PBS использовали в качестве буфера пустой пробы в референсной ячейке. PBS содержал 137 мМ NaCl, 2,7 мМ KCl, 8,1 мМ Na2HPO4 и 1,47 мМ KH2PO4, pH 7,2. Образцы распределяли в лоток для образцов MicroCal VP-Capillary DSC с помощью автосемплера (Malvern Instruments Ltd, Malvern, UK). Образцы уравновешивали в течение 5 минут при 10°C, а затем доводили до 110°C со скоростью 100°C в час. Выбирали период фильтрации 16 секунд. Необработанные данные подвергали поправке на фон и нормализовали концентрацию белка. Для аппроксимации данных к модели MN2-State с соответствующим количеством переходов использовали программное обеспечение Origin 7.0 (OriginLab Corporation, Northampton, MA). Результаты дифференциальной сканирующей калориметрии оптимизированных связывающих доменов против GUCY2c в формате scFv-Fc приведены в таблице 31. Оптимизированные клоны демонстрируют улучшенную температуру разворачивания (Tm1) по сравнению с родительским, неоптимизированным клоном GUCY2C-0247 (обозначенным как GUCY2C-0295 в формате scFv-Fc).
Таблица 31
Пример 18: Анализ поверхностного плазмонного резонанса (SPR) биспецифических антител против CD3-GUCY2c
Аффинности связывания биспецифических антител против CD3-GUCY2c с GUCY2c человека (SEQ ID NO: 224), GUCY2c яванского макака (SEQ ID NO: 236) и GUCY2C мыши (SEQ ID NO: 240) определяли с использованием устройства BIACORE™ T200 (GE Healthcare) при 25°C и 37°C со скоростью сбора 10 Гц. GUCY2c человека, GUCY2c яванского макака и GUCY2c мыши напрямую иммобилизовали на поверхности трех разных проточных кювет сенсорного чипа CM5 (BR100530, GE Healthcare) с использованием набора для присоединения по аминогруппе (BR100050, GE Healthcare) по инструкциям производителя. Конечные уровни иммобилизации GUCY2c человека, яванского макака и мыши составляли 142 резонансных единиц (RU), 61 RU и 201 RU соответственно. Проточную кювету 1 использовали в качестве референсной проточной кюветы. Серию трехкратных разведений биспецифических антител против CD3-GUCY2c с концентрациями в диапазоне от 450 нМ до 5,56 нМ впрыскивали на поверхность сенсора в течение 52 секунд при скорости потока 50 мкл/мин. Осуществляли мониторинг диссоциации в течение 400 секунд и регенерировали поверхность 10 мМ глицином, pH 2,1. Подвижный буфер и буфер для образцов представляли собой 10 мМ HEPES, pH 7,4, 0,15 M NaCl, 3 мМ ЭДТА, 0,05% P-20 (HBS-EP+). Данные получали в двух параллелях. Аффинности связывания и константы скорости определяли посредством аппроксимации данных получаемой сенсограммы к модели 1:1 Ленгмюра с помощью аналитического программного обеспечения BIACORE™ T200 версии 3.0 (GE Healthcare). Аффинности связывания биспецифических антител против GUCY2c с CD3 человека (SEQ ID NO: 242) и CD3 яванского макака (SEQ ID NO: 244) определяли с использованием устройства BIACORE™ T200 (GE Healthcare) при 25°C и 37°C со скоростью сбора 10 Гц. CD3 человека и яванского макака захватывали на поверхности трех разных проточных кювет сенсорного чипа CM5 (BR100050, GE Healthcare) с использованием набора для захвата по His-группе (28995056, GE Healthcare) по инструкциям производителя. Уровни захвата CD3 человека (SEQ ID NO: 242) составляли 8 резонансных единиц (RU) и 16 RU на проточных кюветах 2 и 3, соответственно, при этом 10 RU CD3 яванского макака захватывали на проточной кювете 4. Проточную кювету 1 использовали в качестве референса. Серию трехкратных разведений биспецифического белка против GUCy2c с концентрациями в диапазоне от 100 нМ до 3,7 нМ впрыскивали на сенсорную поверхность в течение 55 секунд при скорости потока 50 мкл/мин. Осуществляли мониторинг диссоциации в течение 300 секунд и поверхность регенерировали 10 мМ глицином, pH 1,5. Подвижный буфер и буфер для образцов представляли собой 10 мМ HEPES, pH 7,4, 0,15 M NaCl, 3 мМ ЭДТА, 0,05% P-20 (HBS-EP+). Данные получали в двух параллелях. Аффинности связывания и константы скорости определяли посредством аппроксимации получаемых данных сенсограммы к модели Ленгмюра 1:1 с помощью аналитического программного обеспечения BIACORE™ T200 версии 3.0 (GE Healthcare). Результаты анализа связывания BIACORE™ биспецифических антител против CD3-GUCY2c с GUCY2c человека, яванского макака и мыши в разных условиях, включая ориентацию анализа и температуру, приведены в таблицах 32A-32J. Гуманизированные и оптимизированные GUCY2C-связывающие домены демонстрировали сохраненное или улучшенное связывание с GUCY2C и CD3 в качестве биспецифических антител. В некоторых случаях наблюдали снижение аффинности связывания при повышенной температуре (37°C).
В таблице 32A показана кинетика связывания биспецифических антител против CD3-GUCY2c с GUCY2c человека. Используемым способом захвата являлся захват с помощью антитела против IgG человека при 25°C.
Таблица 32A
В таблице 32B показана кинетика связывания биспецифических антител против CD3-GUCY2c с GUCY2c человека. Используемым способом захвата являлась прямая иммобилизация при 25°C.
Таблица 32B
В таблице 32C показана кинетика связывания биспецифических антител против CD3-GUCY2c с GUCY2c человека. Используемым способом захвата являлась прямая иммобилизация при 37°C.
Таблица 32C
В таблице 32D показана кинетика связывания биспецифических антител против CD3-GUCY2c с GUCY2c яванского макака. Используемым способом захвата являлся захват с помощью антитела против IgG человека при 25°C.
Таблица 32D
В таблице 32E показана кинетика связывания биспецифических антител против CD3-GUCY2c с GUCY2c яванского макака. Используемым способом захвата являлась прямая иммобилизация при 25°C.
Таблица 32E
Таблица 32F показана кинетика связывания биспецифических антител против CD3-GUCY2c с GUCY2c яванского макака. Используемым способом захвата являлась прямая иммобилизация при 37°C.
Таблица 32F
В таблице 32H показана кинетика связывания биспецифических антител против CD3-GUCY2c с GUCY2c мыши. Используемым способом захвата являлся захват с помощью антитела против IgG человека при 25°C.
Таблица 32H
В таблице 32I показана кинетика связывания биспецифических антител против CD3-GUCY2c с GUCY2c мыши. Используемым способом захвата являлась прямая иммобилизация при 25°C.
Таблица 32I
В таблице 32J показана кинетика связывания биспецифических антител против CD3-GUCY2c с GUCY2c мыши. Используемым способом захвата являлась прямая иммобилизация при 37°C.
Таблица 32J
В таблицах 33A-33F показан анализ связывания BIACORE™ биспецифических антител против CD3-GUCY2c с CD3 человека и яванского макака в разных условиях, включая ориентацию анализа и температуру.
В таблице 33A показана кинетика связывания биспецифических антител против CD3-GUCY2c с белком CD3 человека. Используемым способом захвата являлся захват с помощью антитела против IgG человека при 25°C.
Таблица 33A
В таблице 33B показана кинетика связывания биспецифических антител против CD3-GUCY2c с CD3 человека. Используемым способом захвата являлся захват с помощью антитела против His при 25°C.
Таблица 33B
В таблице 33C показана кинетика связывания биспецифических антител против CD3-GUCY2c с CD3 человека. Используемым способом являлся захват с помощью антитела против His при 25°C.
Таблица 33C
В таблице 33D показана кинетика связывания биспецифических антител против CD3-GUCY2c с CD3 яванского макака. Используемым способом являлся захват с помощью антитела против His при 25°C.
Таблица 33D
В таблице 33E показана кинетика связывания биспецифических антител против CD3-GUCY2c с CD3 яванского макака. Используемым способом являлся захват с помощью антитела против His при 25°C.
Таблица 33E
В таблице 33F показана кинетика связывания биспецифических антител против CD3-GUCY2c с CD3 яванского макака. Используемым способом являлся захват с помощью антитела против His при 25°C.
Таблица 33F
Пример 19: Измерения высвобождения цитокинов in vitro в случае биспецифических антител против CD3-GUCY2c
Клетки T84 обрабатывали T-клетками (соотношение E:T 5:1) и GUCY2C-1608 в разных дозах. Супернатанты собирали в разные моменты времени (24 ч., 48 ч., 96 ч.), анализировали с использованием мультиплексного анализа Lumninex по инструкциям производителя и считывали с помощью Luminex 200 с использованием программного обеспечения xPONENT. Измеряемыми цитокинами являлись ИФН-гамма, ИЛ-10, ИЛ-2, ИЛ-4, ИЛ-6, ФНО-альфа человека. На фигурах 5A-5F проиллюстрированы профили высвобождения цитокинов в случае GUCY2C-1608. Эти цитокины подвергались положительной регуляции при обработке GUCY2C-1608 клетками T84 в присутствии T-клеток, что подтверждает механизм опосредованной T-клетками активности биспецифического антитела против CD3-GUCY2c в GUCY2c-экспрессирующих клетках.
Пример 20: Оценка in vivo активности, опосредованной биспецифическим антителом против CD3-GUCY2c
PBMC человека размораживали в средах (X-VIVO 15 (Lonza). Добавляли 5% сывороточного альбумина человека (Gemini #100-318), 1% пенициллина/стрептомицина, 0,01 мМ 2-меркаптоэтанола на приблизительно 5 миллионов клеток на мл. Клетки центрифугировали и ресуспендировали в буфере Robosep (Stem Cell Technologies) в концентрации 50 миллионов клеток на мл. T-клетки выделяли с использованием набора для обогащения T-клеток человека EasySep (Stem Cell Technologies). T-клетки активировали и выращивали с использованием набора для активации/экспансии T-клеток человека (Miltenyi). В день 2 T-клетки переносили в устройство для культивирования клеток G-Rex для экспансии, в среды добавляли ИЛ-2 (Stem Cell Technologies) и восполняли через 2 дня. T-клетки собирали через 1 неделю после активации/экспансии. Во время сбора частицы удаляли с помощью магнита и клетки ресуспендировали в DPBS в количестве 1×107 клеток/мл для инокуляции in vivo.
Для исследований ксенотрансплантатов мышам NSG подкожно в бок инокулировали линии клеток колоректальной опухоли (T84, H55, LS1034) или полученные из пациента фрагменты ксенотрансплантата (PDX-CRX-11201, PDX-CRX-12213, PDX-CRX-24225).
Опухоли измеряли с использованием цифрового штангенциркуля и вычисляли объемы с помощью модифицированной формулы эллипсоида 1/2 × длина × ширина 2. Мышей рандомизировали и стадировали по размеру опухоли 150-200 мм3. Начальную дозу биспецифического антитела против GUCY2c, биспецифического антитела отрицательного контроля или носителя вводили животным в день 0 и 2 миллиона культивированных T-клеток инокулировали на следующий день. Мышам вводили дозы в виде болюсных инъекций 0,2 мл еженедельно до 5 раз, и все введения соединений и T-клеток осуществляли внутривенно через боковую хвостовую вену каждого животного. Опухоли измеряли дважды в неделю вместе с непрерывным мониторингом признаков реакции "трансплантат-против-хозяина" (фигуры 6-15). Все биспецифические антитела против CD3-GUCY2c демонстрировали дозозависимую опосредованную T-клетками противоопухолевую активность в моделях линии клеток ксенотрансплантата и полученного из пациента ксенотрансплантата.
Пример 21: Анализ циклического гуанозинмонофосфата (цГМФ) для характеризации биспецифических антител против GUCY2c-CD3 в отношении потенциальной активности модуляции пути GUCY2c
Для тестирования продукции циклического гуанозиномонофосфата (цГМФ) клетками T84 в ответ на стимуляцию пути GUCY2c 2×106 клеток T84 высевали в 24-луночный планшет на ночь. Клетки промывали DMEM, содержащей 50 мМ HEPES, и обрабатывали DMEM, содержащей 50 мМ HEPES и 1 мМ 3-изобутил-1-метилксантин (IBMX), в течение 15 минут. Затем клетки обрабатывали GUCY2C-1608 или биспецифическим антителом отрицательного контроля в концентрациях в диапазоне от 0,1 до 99 мкМ. В качестве положительного контроля добавляли агониста GUCY2c, энтеротоксин STp (BACHEM) (от 0,02 до 51 мкМ) в течение 45 минут. Твердые частицы удаляли посредством центрифугирования и использовали 200 мкл супернатанта для анализа продукции цГМФ с использованием набора cGMP Parameter Assay Kit (R&D Systems) по инструкциям производителя. Для оценки потенциальной нейтрализующей активности GUCY2C-1608 добавляли 0,2 мкМ STp в том же диапазоне, что и GUCY2C-1608 или биспецифическое антитело отрицательного контроля, как описано выше, на 45 минут, и супернатанты аналогично анализировали на уровни цГМФ.
Активность пути GUCY2c оценивали в клетках T84 посредством измерения продукции цГМФ. Хотя агонист пути GUCY2c бактериальный энтеротоксин STp повышал цГМФ дозозависимым образом, продукция цГМФ не повышалась при добавлении увеличивающихся концентраций GUCY2C-1608. Кроме того, GUCY2C-1608 не влияло на продукцию цГМФ, индуцируемую 0,2 мкМ STp (фигура 16). Эти результаты свидетельствуют о том, что GUCY2C-1608 не является агонистом пути GUCY2c и не нейтрализует функцию пути GUCY2c в присутствии лиганда.
Пример 22: ELISA полиспецифичности в отношении ДНК и инсулина
Предполагают, что неспецифическое связывание антител с иными молекулами, чем мишени, является механизмом быстрого клиренса in vivo (Hötzel et al., 2010, MAbs 4(6):753-760). Доказательства такого полиреактивного неспецифического связывания можно получать посредством измерения связывания препаратов мембран (Xu et al., Protein Eng. Des. Select. 26(10):663-70, 2013), бакуловирусных частиц (Hötzel et al., mAbs 4, 753-760, (2012)) или отрицательно заряженных субстратов, таких как ДНК, инсулин и гепарин (Tiller et al., J. Immunol. Methods 329(1-2):112-124, 2008). В ELISA на ДНК и инсулин использовали способ низкой строгости, исходно разработанный для детекции низкоаффинных аутоантител от пациентов с системной красной волчанкой (Tiller et al., J. Immunol. Methods 329, 112, 2008; патент США № 7314622).
384-луночные планшеты для ELISA (Nunc Maxisorp) покрывали в течение ночи при 4°C ДНК (10 мкг/мл) и инсулином (5 мкг/мл) в PBS, pH 7,5. ELISA, адаптированный из анализов, описанных в Tiller et al., J. Immunol. Methods 329, 112, 2008; патенте США № 7314622, осуществляли с помощью автоматизированного устройства для манипуляций с жидкостями PerkinElmer Janus. Лунки промывали водой, блокировали 50 мкл буфера ELISA на полиреактивность (PEB; PBS, содержащий 0,5% Tween-20, 1 мМ ЭДТА) в течение 1 часа при комнатной температуре и три раза промывали водой. Серийно разведенные mAb в 25 мкл добавляли в лунки в четырех параллелях и инкубировали в течение 1 ч. при комнатной температуре. Планшеты 3 раза промывали водой и в каждую лунку добавляли 25 мкл 10 нг/мл антитела козы против IgG человека (Fcγ-фрагмент-специфического), конъюгированного с пероксидазой хрена (Jackson ImmunoResearch). Планшеты инкубировали в течение 1 ч. при комнатной температуре, 3 раза промывали 80 мкл воды и в каждую лунку добавляли 25 мкл субстрата TMB (Sigma Aldrich). Реакции останавливали через 6 минут 50 секунд посредством добавления 25 мкл 0,18 M ортофосфорной кислоты в каждую лунку и считывали поглощение при 450 нм. Баллы связывания с ДНК и инсулином вычисляли как соотношение сигнала ELISA антитела в количестве 10 мкг/мл и сигнала в лунке, содержащей буфер. В таблице 34 показан анализ полиспецифичности биспецифических антител против CD3-GUCY2c с использованием ELISA связывания ДНК и инсулина. Оптимизированные биспецифические антитела демонстрировали низкий потенциал неспецифического связывания.
Таблица 34
Пример 23: Спектроскопический анализ самовзаимодействия наночастиц с аффинным захватом (AC-SINS)
Антитело и антитело-подобные белки обладают потенциалом к взаимодействию с самими собой, особенно при повышенных концентрациях. Это самовзаимодействие может приводить к проблемам с вязкостью, ассоциированными с составом, во время разработки лекарственных средств, а также повышенному риску клиренса (Avery et al., mAbs, 2018, Vol. 0, N0. 0, 1-12). С помощью спектроскопического анализа самовзаимодействия наночастиц с аффинным захватом измеряют самовзаимодействие, и его используют для облегчения прогнозирования высокой вязкости и потенциально плохих фармакокинетических свойств.
Анализ AC-SINS стандартизировали в 384-луночном формате с помощью автоматизированного устройства для манипуляций с жидкостями Perkin-Elmer Janus. 20 нм наночастиц золота (Ted Pella, Inc., #15705) покрывали смесью 80% антитела козы против Fc человека (Jackson ImmunoResearch Laboratories, Inc. # 109-005-098) и 20% неспецифических поликлональных антител козы (Jackson ImmunoResearch Laboratories, Inc. # 005-000-003), подвергнутых замене буфера на 20 мМ ацетат натрия, pH 4,3, и разводили до 0,4 мг/мл. После инкубации в течение одного часа при комнатной температуре незанятые участки на наночастицах золота блокировали тиолированным полиэтиленгликолем (2 кДа). Затем покрытые наночастицы концентрировали в 10 раз с использованием шприцевого фильтра и добавляли 10 мкл к 100 мкл mAb при 0,05 мг/мл в PBS, pH 7,2. Покрытые наночастицы инкубировали с интересующим антителом в течение 2 ч. в 96-луночном полипропиленовом планшете, а затем переносили в 384-луночный полистироловый планшет и считывали с помощью спектрофотометра Tecan M1000. Поглощение считывали при 450-650 с приращениями 2 нм, и для идентификации максимального поглощения, сглаживания данных и аппроксимации данных с использованием полинома второго порядка использовали Microsoft Excel macro. Сглаженное максимальное поглощение средней пустой пробы (только буфер PBS) вычитали из сглаженного максимального поглощения образца антитела для определения баллов AC-SINS антитела. В таблице 35 приведен анализ неспецифичности AC-SINS для оптимизированных биспецифических антител против CD3-GUCY2c, показавший низкий потенциал самовзаимодействия и неспецифичности.
Таблица 35
Пример 24: Получение стабильной линии клеток, экспрессия и очистка биспецифического антитела GUCY2c-1608
Две цепи, кодирующие биспецифическое антитело GUCY2C-1608, клонировали в экспрессирующий вектор млекопитающего pRY19-GA-Q, содержащий двойной промотор и участок рекомбинации для интеграции по одному участку (SSI) (Zhang, L. et al. Biotechnology Progress., 31: 1645-1656, 2015) в геном клетки CHO. SSI основана на инсерции интересующего гена с использованием основанной на рекомбиназе замене кассеты в хромосомном участке, известном высокой стабильной экспрессией генов. Конструкции GUCY2C-1608-впадина и GUCY2C-1608-выступ клонировали в экспрессирующий вектор в кассете один и два, соответственно. С помощью полученной плазмиды GUCY2C-1608 трансфицировали 10E9 клеток CHO-K1 SV посредством электропорации с использованием вектора pRY19, содержащего интересующий ген, с последующим положительным и отрицательным селекционным давлением с использованием гигромицина-B и ганцикловира. Эти популяции CHO подвергали 3-недельной фазе восстановления. Когда наблюдаемая жизнеспособность превышала 90%, получали банки клеток для последующего использования. Затем полученные популяции CHO подвергали 12-дневной экспрессии на платформе для культивирования с подпиткой в средах CD-CHO (ThermoFisher Scientific, Waltham, MA). В конце 12-дневного периода клетки центрифугировали при 3000×g в течение 10 минут с последующей стерильной фильтрацией кондиционированных сред с использованием фильтра-пробки для бутыли PES 0,2 мкм (Corning, Oneonta, NY). Кроме того, стабильные популяции CHO масштабировали для контролируемого качающегося биореактора объемом 10 л (Finesse, Santa Clara, CA). Популяции CHO выращивали в собственных средах Pfizer в рамках 12-дневного культивирования с подпиткой, включавшего химически определенную подпитку для поддержания жизнеспособности культуры. После завершения культуру подвергали глубинной фильтрации с использованием фильтровального тракта, состоящего из фильтра 20" 5 мкм Pall Profile II с последующим фильтром 10" 0,2 мкм Pall Supor (Pall Corporation, Port Washington, NY). Титры оценивали с использованием аффинной хроматографии с протеином A. Затем осуществляли очистку белка способами, известными в этой области, включая хроматографию с протеином A, хроматографию гидрофобных взаимодействий и ионообменную хроматографию. Титр экспрессии белка GUCY2C-1608 из стабильно трансфицированной популяции клеток CHO приведен в таблице 36. Титры экспрессии для популяций клона GUCY2C-1608 свидетельствовали об уровнях экспрессии от 0,48 до 0,84 г/литр.
Таблица 36
Пример 25: Масс-спектрометрический анализ биспецифического антитела GUCY2C-1608
Анализ LC/MS осуществляли для подтверждения образования правильного спаренного очищенного биспецифического антитела GUCY2C-1608. Молекулярные массы биспецифической молекулы определяют по ее уникальным аминокислотным последовательностям, и при определении точной молекулярной массы получают доказательства наличия правильно спаренных биспецифических молекул против GUCY2c. В кратком изложении, образец биспецифического антитела против CD3-GUCY2c анализировали посредством анализа LC/MS с помощью устройства для ВЭЖХ Waters Acquity H-класса, сопряженного с масс-спектрометром Bruker maXis II Q-ToF. Аналиты нагружали на колонку SMT SAM C2 (2,1 мм × 100 мм, поддерживаемую при 80°C) и элюировали с использованием третичного градиента буфера B (80% 1-пропанола, 20% ацетонитрила с 0,05% трифторуксусной кислоты) и буфера C (100% ацетонитрила с 0,05% трифторуксусной кислоты) при скорости потока 300 мкл/мин (5 мин линейного повышения до 19% B и 4% C, 20 мин линейного повышения до 22% B и 16% C, 10 мин линейного повышения до 0% B и 98% C). Масс-спектрометрическую детекцию осуществляли в позитивном режиме с капиллярным напряжением 3,5 кВ. Анализ данных осуществляли с помощью деконволюции MaxEnt 1 в программе Compass DataAnalysis v4.2. На фигуре 17 показана высокая чистота при прогнозируемой молекулярной массе конечного очищенного биспецифического антитела GUCY2C-1608.
Пример 26: Последовательности
В таблицах 37A и 37B приведено обобщение SEQ ID NO антител или фрагментов антител, представленных в настоящем описании. В таблице 37A используют систему нумерации ABM для последовательностей CDR1 VH. В таблице 37A используют систему нумерации Kabat для всех других последовательностей, включая CDR2 VH.
Таблица 37A
Таблица 37B
В таблице 38 приведены последовательности, представленные в настоящем описании.
Таблица 38
2B5-1038 VH,
2B5-1039 VH,
2B5-1040 VH,
CDR1 (расширенный/ABM)
2B5-1038 VH,
2B5-1039 VH,
2B5-1040 VH,
CDR3
2B5-1038 VH,
2B5-1039 VH,
2B5-1040 VH,
CDR2 (Kabat)
GUCY2C-1608_VH
GUCY2C-1608_выступ
Регистрационный номер GenBank NP_004954.2
Регистрационный номер GenBank NM_004963.3
Регистрационный номер GenBank NP_001120790.1
Регистрационный номер GenBank NM_001127318.1
2B5-1038 VH,
2B5-1039 VH,
2B5-1040 VH,
CDR1 (Kabat)
2B5-1038 VH,
2B5-1039 VH,
2B5-1040 VH,
CDR2 (Chothia)
2B5-1039 VH,
2B5-1040 VH
VH 2B5-1039,
VH 2B5-1040,
CDR1 (Chothia)
VL 2B5-1039,
VL 2B5-1040,
CDR2
Легкая цепь IgG кролика версии клона 9H3
Тяжелая цепь IgG кролика версии клона 9H3
Легкая цепь IgG1 мыши клона 9H3
Тяжелая цепь IgG1 мыши клона 9H3
Легкая цепь_
Химерное 9H3 IgG1 человека 3m в pTT5
Тяжелая цепь
Химерное 9H3 IgG1 человека 3m в pTT5
Последовательности антител по настоящему изобретению пронумерованы с использованием Abysis AbM. С помощью системы нумерации Abysis AbM приписывают нумерацию по Kabat и определяют начало и конец каждой CDR по правилам Kabat, за исключением CDR H1. CDR H1 определяют по AbM.
Пример 27: Три варианта антитела против CD3 демонстрировали улучшенную аффинность и цитотоксичность в биспецифических антителах против GUCY2C-CD3
Три варианта антитела против CD3, 2B5-1038 (SEQ ID NO: 273 и 274), 2B5-1039 (SEQ ID NO: 273 и 275), и 2B5-1040 (SEQ ID NO: 273 и 276), полученные из 2B5v6, переформатировали в молекулы биспецифическое диатело-Fc, спаренные с последовательностью противоопухолевого антитела против GUCY2c в одной из конфигураций, показанных на фигуре 1, с использованием стандартных способов клонирования, экспрессии и очистки, представленных в настоящем описании выше. Получаемые биспецифические антитела, приведенные в таблице 39, тестировали на цитотоксичность in vitro с использованием анализа перенаправления T-клеток, аффинность связывания с использованием анализа поверхностного плазмонного резонанса (SPR) и неспецифичность с использованием AC-SIN. Риск иммуногенности оценивали с помощью Epivax Tool для прогнозирования in silico потенциальных T-клеточных эпитопов. Интактные данные масс-спектроскопии свидетельствуют о том, что все биспецифические антитела имеют правильное спаривание и на 100% являются гетеродимерами. В таблице 39 показано, что все три биспецифические антитела (1) являются приблизительно в 2 раза более активными в анализе цитотоксичности in vitro по сравнению с контрольным биспецифическим GUCY2C-1608, спаренным с антителом против CD3 2B5v6 (фигура 18); (2) имеют аффинность связывания с рекомбинантным CD3 человек по результатам SPR, соответствующую активности цитотоксичности; (3) имеют баллы иммуногенности, являющуюся улучшенными по сравнению с контролем; и (4) баллы AC-SIN оставались теми же, что и у контрольного биспецифического антитела GUCY2C-1608.
Таблица 39
Пример 28:Созревание аффинности связывающего домена антитела против GUCY2c
Для повышения аффинности связывающего домена антитела против GUCY2c к GUCY2c использовали подход фагового дисплея. Конструировали четыре библиотеки фагового дисплея и получали их способами, описанными выше. Библиотеки получали посредством встраивания мутаций в CDR H1, H2 и H3 и CDR L1 антитела GUCY2C-1608 (SEQ ID NO: 221). Фаговые библиотеки подвергали "спасению", как описано выше, и селекции с использованием стратегии конкурентной селекции, показанной в таблице 40. Для раунда 1 фаг инкубировали с стрептавидиновыми частицами, комплексированными с биотинилированной GUCY2c человека (5 нМ; SEQ ID NO: 232). После 3 промывок в реакционную смесь добавляли 100-кратный избыток небиотинилированного антигена (500 нМ) и смесь вращали в течение ночи при 4°C. Частицы промывали на следующий день для удаления фага, диссоциировавшего от частиц, и оставшегося связанного фага элюировали с использованием TEA. Для раунда 2 селекцию повторяли, как описано выше, против 0,5 нМ антигена без какой-либо конкуренции. Отбирали колонии и получали периплазматические препараты, как описано выше. Клоны подвергали скринингу на улучшенную активность связывания с использованием конкурентного ELISA, описанного выше. Клоны, демонстрировавшие улучшенное связывание по сравнению с родительскими вариантами, выбирали для дальнейшего анализа. После скрининга осуществляли секвенирование ДНК, как описано выше, лучших клонов для идентификации мутаций, вносящих вклад в улучшенную активность связывания GUCY2c.
После исходной библиотеки для селекции посредством созревания аффинности области CDR этих лучших вариантов комбинировали в новую библиотеку и осуществляли дополнительные селекции фагового дисплея, приведенные в таблице 41. Раунд 1 селекции осуществляли, как описано выше для исходной библиотеки для селекции посредством созревания аффинности, за исключением того, что использовали 0,5 нМ биотинилированного антигена. Для каждой библиотеки было две группы селекции раунда 2, в которых фага перед селекцией стимулировали или не стимулировали с помощью инкубации при 65°C. После селекции фага отбирали колонии и получали периплазматические препараты, как описано выше. Клоны подвергали скринингу посредством конкурентного ELISA, описанного выше. После скрининга осуществляли секвенирование ДНК лучших клонов для идентификации мутаций, вносящих вклад в улучшенную активность связывания GUCY2c. В таблицах 40 и 41 показана стратегия селекции фагового дисплея, используемая для библиотек созревания аффинности (C=библиотека; R=раунд; Dslxn=условия деселекции; Therm=термические условия; SA=стрептавидин; D=ДНК; I=инсулин; M=мембранный экстракт).
Таблица 40
Таблица 41
Аминокислотные последовательности клонов антитела против GUCY2c, демонстрировавших улучшенную аффинность при конкурентном ELISA PeriPrep, сравнивали с родительским клоном GUCY2C-1608 (VH SEQ ID NO: 73 и VL SEQ ID NO: 147). Изменения содержания аминокислот в CDR VH, приводившие к улучшенной стабильности, приведены в таблице 42A. Изменения в положениях 54, 55, 56, 97 и 99, по-видимому, имеют наибольшее влияние на аффинность, что коррелирует с наибольшими изменениями флуоресценции с разрешением по времени (в соответствии с системой нумерации Kabat, с использованием определения AbM для CDR1 VH).
Таблица 42A
(H1)
(H2)
(H3)
Изменения содержания аминокислот в CDR VL, приводящие к улучшенной аффинности при конкурентном ELISA PeriPrep, показаны в таблице 42B (систему нумерации Chothia использовали для CDR1 VL). Изменения в положениях 30, 30a и 30d в CDR1 VL, по-видимому, имеют наибольшее влияние на аффинность, что коррелирует с наибольшими изменениями флуоресценции с разрешением по времени.
Таблица 42B
(L1)
После скрининга в формате scFv в виде периплазматических препаратов последовательности VH и VL лучших клонов клонировали в полный формат IgG1/каппа человека. Гены VH и VL амплифицировали посредством ПЦР из фагового вектора и субклонировали в векторы IgG1 человека и каппа человека способами молекулярной биологии, описанными выше. Затем версию IgG1 человека этих лучших аффинно зрелых клонов транзиторно экспрессировали и очищали способами, также описанными выше. Затем очищенные белки анализировали посредством поверхностного плазмонного резонанса, как описано выше, против рекомбинантного белка GUCY2c человека (SEQ ID NO: 232). В таблице 43 показаны скорость прямой реакции (ka), скорость обратной реакции (kd) и равновесная константа (KD) для аффинно зрелых клонов по сравнению с родительским GUCY2C-1608 в форме IgG (GUCY2c-1640; SEQ ID NO: 214 и 223). Несколько вариантов демонстрировали аффинность, улучшенную в 3,4 раза по сравнению с родительским антителом. В таблице 44 приведены полные аминокислотные последовательности VH и VL этих аффинно зрелых версий GUCY2C-1608.
Таблица 43. Поверхностный плазмонный резонанс аффинно зрелых IgG против GUCY2c относительно белка внеклеточного домена (ECD) GUCY2c человека.
Таблица 44. Аминокислотные последовательности аффинно зрелых областей VH и VL
Пример 29: Определение эпитопов посредством пептидного картирования внеклеточного домена GUCY2C
Пептидное картирование осуществляли для определения эпитопа на внеклеточном домене (ECD) GUCY2c, с которым связывается основное антитело против GUCY2c. Для этого подхода получали 20-мерные пептиды с перекрыванием 15 аминокислот, охватывающие весь внеклеточный домен в трансмембранном домене GUCY2c, состоящем из аминокислот 1-440 (таблица 45). Пептиды синтезировали и предоставляли в виде панелей в лиофилизированном формате (New England Peptide, Gardner MA). Панели переразводили ацетонитрилом до 100 мкг/мл, а затем дополнительно разбавляли до 20 мкг/мл буфером карбоната/бикарбоната натрия. Затем очищенные антитела или биспецифические антитела против GUCY2c-CD3 тестировали с помощью Delfia ELISA на связывание с различными пептидами с использованием описанных выше способов Delfia ELISA, при этом разведенные пептиды использовали в качестве антигенов-мишеней. GUCY2C-1608 демонстрировало сильное связывание с пептидами 19 и 20 (SEQ ID NO: 334 и 335), что измеряли по флуоресценции с разрешением по времени (TRF). Данные о связывании по результатам TRF для GUCY2C-1608 относительно всех пептидов приведены в таблице 45. Последовательность NSGDCRSSTCEGLDLLRKISNAQRM (SEQ ID NO: 405) охватывается этими пептидами. Область перекрывания пептидов представляет собой RSSTCEGLDLLRKIS (SEQ ID NO: 406), указывая на эпитоп в области.
Таблица 45
Затем определяли конкуренцию пептида 19 GUCY2c (SEQ ID NO: 334) с полным внеклеточным доменом в конкурентном ELISA DELFIA способами, описанными выше, и показано, что он эффективно конкурирует с полным внеклеточным доменом (фигура 19). Это позволяет предполагать, что пептид 19 (SEQ ID NO 334) содержит специфический эпитоп, с которым связывается GUCY2C-1608, на полном внеклеточном домене GUCY2c.
GUCY2C-1608 связывается с иным эпитопом ECD GUCY2c, чем Антитело MS20
Связывание GUCY2C-1608 с внеклеточным доменом GUCY2c также сравнивали с известными с антителами против GUCY2c. Антитело MS20, описанное в US2013/315923, не имеет описанного эпитопа. Для сравнения этого антитела с GUCY2C-1608 осуществляли конкурентный ELISA Delfia с использованием модифицированного способа конкурентного ELISA Delfia, описанного выше. В этом случае белок GUCY2c человека (SEQ ID NO: 232) наносили в виде покрытия на планшет, как описано выше. GUCY2C-1640 (IgG-версию GUCY2C-1608; SEQ ID NO: 214 и 223) или антитело MS20 серийно разводили, начиная с 20 мкг/мл, затем комбинировали с химерной IgG-версией кролика GUCY2C-0098 (SEQ ID NO: 440 и 441, состоящих из SEQ ID NO: 26 и 106, слитых с константным доменом кролика) в количестве 250 нг/мл (значение EC50, определенное отдельно). Планшеты промывали, как описано выше, и добавляли вторичное антитело против кролика с европием (Perkin Elmer), разведенное 1:1000, для детекции. Сигнал TRF определяли, как описано ранее. Снижение сигнала TRF свидетельствует о конкурентном связывании, т.к. химерная версия кролика GUCY2C-0098 вытесняется из связывания с GUCY2c человека. Как и ожидали, GUCY2C-1640 вытесняет химерную версию кролика GUCY2C-0098, в то время как MS20 не вытесняет (фигура 20). Таким образом, GUCY2C-1608 связывается с иным эпитопом ECD GUCY2c, чем антитело MS20.
Пример 30: GUCY2c-1608 связывается с иным эпитопом ECD GUCY2c, чем антитело 5F9
Описано, что антитела, описанные в US2011/0110936, связываются с разными последовательностями эпитопов на ECD GUCY2c из пептида 19 (SEQ ID NO 334), как представлено в настоящем описании ниже.
В лунку 96-луночного планшета добавляли 3×106 клеток, экспрессирующих GUCY2c человека, крысы и курицы (CID1814, 1815 и 1818, соответственно), точечные мутанты CID1941-1950, "патч"-мутанты CID1868-1873 (патч-мутанты состоят из 2-5 аминокислотных остатков GUCY2c человека, замененных аминокислотными остатками из эквивалентных положений в GUCY2c курицы), или химеры GUCY2c человека-курицы CID1874-1881 (>5 аминокислот GUCY2c человека, замененных остатками GUCY2c курицы), или подтверждающие обратные химеры CID1939 и 1940 (>5 аминокислот GUCY2c курицы, замененных остатками GUCY2c человека), или отрицательный контроль CID1431 (конструкция отрицательного контроля, несодержащая последовательность GUCY2s, но имеющая те же эпитопные метки, что и другие конструкции, экспрессирующиеся на поверхности дрожжей). Клетки инкубировали с антителом против V5 (Abcam 27671), моновалентными или бивалентными антителами против GUCY2c, антителом GUCY2C-1608 или антителом 5F9 (антителом Millennium, описанным в US2011/0110936) в течение 1 часа при комнатной температуре. Затем клетки промывали три раза буфером PBS+0,5% BSA. Затем клетки инкубировали с PE-конъюгированным антителом против IgG мыши, или PE-конъюгированным антителом против IgG человека, или PE-конъюгированным антителом против HA (Miltenyi Biotec 130-092-257) в течение 30 минут при 4°C в темноте. Клетки промывали три раза буфером PBS+0,5% BSA, а затем анализировали с помощью проточного цитометра.
MFI (медиану интенсивности флуоресценции) антитела против GUCY2c, связывающегося с клетками, нормализовали по экспрессии на поверхности клетки, измеряемой по MFI антител против меток. Все клетки экспрессировали V5 и HA на поверхности клеток для нормализации экспрессии. В таблице 46 показано нормализованное относительное связывание антител против GUCY2c с каждой конструкцией GUCY2c. Химеры (за исключением CID1939 и CID1940), точечные мутанты и патч-мутанты, не связывающиеся с Ab против GUCY2c, демонстрируют, что остатки GUCY2c, измененные в этих конструкциях, важны для связывания с антителом. Восстановление связывания GUCY2C-1608, но не антитела 5F9 с обратными химерами CID1939 и CID1940 подтверждает, что антитело GUCY2C- 1608 связывается с остатками 68-87 на GUCY2c (SEQ ID NO: 224), и свидетельствует о том, что участок связывания антитела 5F9 не перекрывается с участком связывания антитела GUCY2C-1608.
Таблица 46
Пример 31: Кристаллизация и определение структуры комплекса scFv против GUCY2C-1608 и GUCY2c-пептида
Для исследований кристаллизации комплекс scFv GUCY2C-1608 (SEQ ID NO 409) и пептида ECD GUCY2c № 19 68NSGDCRSSTCEGLDLLRKIS87 (SEQ ID NO 334) получали в молярном соотношении 1:1,2 и концентрировали до 8,8 мг/мл в белковом растворе, содержащем TBS, при pH 7,5. Кристаллы получали способом висячей капли посредством диффузии в парах в условиях, содержащих 20% PEG 3350, 200 мМ сульфата лития, бис-Трис, pH 5,5. Стержневые кристаллы имели симметрию, соответствующую моноклинной пространственной группе=P212121 с параметрами ячейки a=70,68 Å; b=80,57 Å; c=90,7 Å и с двумя копиями пептидных комплексов scFv GUCY2C-1608 в кристаллографической асимметричной единице. Кристаллы криоконсервировали с использованием резервуарного раствора, содержащего 20% этиленгликоля, и быстро замораживали в жидком азоте. Набор данных для разрешения 1,6 Å получали для одного замороженного кристалла с помощью пучка синхротронного излучения IMCA 17-ID в Argonne National Laboratory (APS). Данные обрабатывали и масштабировали с использованием autoPROC, и конечный набор данных являлся на 71,4% полным.
Структуру разрешали посредством молекулярной замены с использованием PHASER, начиная с одноцепочечного Fv-фрагмента, полученного из структуры биспецифического антитела против GUCY2c и CD3 (фигура 22). Разрешения достигали посредством поиска двух копий scFv GUCY2C-1608. На полученных картах электронной плотности, вычисленных с использованием двух копий scFv GUCY2C-1608 в качестве модели, недвусмысленно видны дополнительные электронные плотности для двух пептидов, каждый из которых связан с соответствующей копией scFv GUCY2C-1608.
С помощью нескольких повторяющихся раундов ручной корректировки и перестройки модели с использованием COOT и кристаллографической детализации с использованием autoBUSTER получали конечную модель GUCY2C-1608 scFv+пептид с кристаллографическими показателями Rрабоч. 19,7% и Rсвободн. 21,7%, где Rрабоч.= ||Fнабл.|-|Fвыч.||/|Fнабл.|, и Rсвободн эквивалентно Rрабоч., но вычислено для случайно выбранных 5% отражений, опущенных во время детализации.
A. Описание структуры scFv GUCY2C-1608-пептид
GUCY2c-пептид длиной 17 остатков принимает, главным образом, α-спиральную конформацию после связывания с областями CDR scFv GUCY2C-1608 (фигура 21). α-спираль прижата дисульфидной SS-связью к N-концу пептида. Общая площадь поверхности связывания, погруженная относительно области взаимодействия, составляет ~1,320 Å2, что является неожиданно большим для фрагмента длиной всего лишь 17 аминокислотных остатков. Поверхность связывания является, главным образом, гидрофобной с некоторыми специфическими полярными контактами в центре и контактами водородной связи на периферии (фигура 21).
Связывающие контакты эпитопов, находящиеся в пределах 3,8 Å от scFv GUCY2C-1608, определены подчеркнутыми остатками в пептидной последовательности 68NSGDCRSSTCEGLDLLRKIS87 и приведены в таблице 47. Аминокислоты, составляющие >75% площади связывающей поверхности, выделены полужирным шрифтом. Связывающий паратоп scFv GUCY2C-1608 определен 5 областями CDR: CDR-H1, CDR-H2, CDR-H3, CDR-L1 и CDR-L3. Аминокислоты scFv GUCY2C-1608 (паратоп) и GUCY2c-пептида (эпитоп), вносящие вклад в контакт в пределах 3,8 Å, приведены в таблице 48.
Таблица 47. Остатки эпитопа GUCY2c
Таблица 48. Аминокислоты, определяющие паратоп (цепи H+L) и эпитоп (цепь C)
B. Кристаллизация и определение структуры биспецифических антител против GUCY2c и CD3
Для исследований кристаллизации получали модифицированную версию GUCY2C-1608 с меткой His6 или меткой FLAG на C-конце каждой цепи субъединицы VL-VH для облегчения очистки. Биспецифическое антитело против GUCY2c и CD3 для кристаллографии получали с использованием последовательностей VH GUCY2C-1637 (SEQ ID NO: 407) и VL GUCY2C-1637 (SEQ ID NO: 408). С помощью конструкций транзиторно трансфицировали клетки FreeStyle™ 293 HEK, как описано выше. После сбора кондиционированных сред 2,5 мл смолы Ni-NTA, заранее уравновешенной TBS, позволяли связываться с белком в течение 1 часа при 4°C с помощью орбитального миксера. Затем смолу собирали и помещали в колонку Applied Biosystems (Life Technologies, Grand Island, NY, USA). Сначала смолу промывали до исходного уровня буфером A (50 мМ фосфата натрия, 300 мМ хлорида натрия, pH 8,0), затем пятью CV буфера A, дополненного 20 мМ имидазола, и, наконец, элюировали буфером A+250 мМ имидазола. Фракции с наибольшей чистотой объединяли и диализовали в PBS с использованием кассеты 30 мл отсечкой по молекулярной массе 10 кДа в течение 2 часов при 4°C. Затем диализованный белок дополнительно очищали посредством хроматографии с использованием метки FLAG. Сорок мл смолы M2 против метки FLAG (Sigma, St. Louis, MO, USA) заранее уравновешивали TBS и позволяли связываться в 1,4 л кондиционированной среды в течение ночи при 4°C. Затем смолу собирали и упаковывали в колонку для хроматографии на M2 против FLAG, промывали до исходного уровня с использованием TBS (20 CV) и сначала элюировали 0,1 M буфером для пептида FLAG, а в конце элюировали с использованием 0,1 M глицина, pH 3,0. Элюированный белок незамедлительно нейтрализовали с помощью 10% 1,0 M Трис, pH 8,0. Затем фракции с наибольшей чистотой объединяли и дополнительно очищали посредством эксклюзионной хроматографии с использованием колонки Superdex200 (GE Healthcare, Piscataway, NJ, USA) и хранили в TBS.
Для исследований кристаллизации очищенную с помощью His/FLAG версию GUCY2C-1608 концентрировали до 12 мг/мл в растворе белка, содержащем TBS, pH 7,5. Кристаллы получали способом висячей капли посредством диффузии в парах в условиях, включающих 1 М малоната натрия, pH 6, 2% полимера этиленимина. Кристаллы имели симметрию, соответствующую гексагональной пространственной группе=P3221 с параметрами ячейки a=b=119,55 Å и c=121,85 Å и с одной копией диатела в кристаллографической асимметричной единице. Кристаллы криоконсервировали с использованием резервуарного раствора, содержащего 2,8 M малоната натрия, pH 6, и быстро замораживали в жидком азоте. Набор данных для разрешения 2,03 Å получали для одного замороженного кристалла с помощью пучка синхротронного излучения IMCA 17-ID в Argonne National Laboratory (APS). Данные обрабатывали и масштабировали с использованием autoPROC, и конечный набор данных являлся на 49,3% полным.
Структуру разрешали посредством молекулярной замены с использованием PHASER, начиная с моделей одноцепочечного Fv-фрагмента, полученных с помощью записи в Brookhaven PDB 1moe. Разрешения достигали посредством отдельного поиска для каждой из четырех субъединиц молекулы диатело. С помощью нескольких повторяющихся раундов ручной корректировки и перестройки модели с использованием COOT и кристаллографической детализации с использованием autoBUSTER получали конечную модель диатела с кристаллографическими показателями Rрабоч. 19,1% и Rсвободн. 20,8%, где Rрабоч.= ||Fнабл.|-|Fвыч.||/|Fнабл.|, и Rсвободн. эквивалентно Rрабоч., но вычислено для случайно выбранных 5% отражений, опущенных во время детализации.
C. Описание структуры диатела против GUCY2C-CD3
Два антигенсвязывающих участка на биспецифическом антителе против GUCY2c и CD3 разделены приблизительно 70 Å и находятся на противоположных сторонах молекулы, непосредственно друг напротив друга (фигура 22). Области CDR против CD3 и CDR против GUCY2c расположены на отдалении от области контакта субъединиц.
Пример 32: Оценка in vivo комбинации биспецифического антитела против GUCY2c-CD3 с mAb против VEGF-A и акситинибом
PBMC человека размораживали в средах (X-VIVO 15 (Lonza). Добавляли 5% сывороточного альбумина человека (Gemini #100-318), 1% пенициллина/стрептомицина, 0,01 мМ 2-меркаптоэтанола на приблизительно 5 миллионов клеток на мл. Клетки центрифугировали и ресуспендировали в буфере Robosep (Stem Cell Technologies) в концентрации 50 миллионов клеток на мл. T-клетки выделяли с использованием набора для обогащения T-клеток человека EasySep (Stem Cell Technologies). T-клетки активировали и выращивали с использованием набора для активации/экспансии T-клеток человека (Miltenyi). В день 2 T-клетки переносили в устройство для культивирования клеток G-Rex для экспансии и в среду добавляли ИЛ-2 (Stem Cell Technologies) и восполняли через 2 дня. T-клетки собирали через 1 неделю после активации/экспансии. Во время сбора частицы удаляли с помощью магнита и клетки ресуспендировали в DPBS в количестве 1×107 клеток/мл для инокуляции in vivo.
Мышам NSG подкожно в бок инокулировали полученные из пациента фрагменты колоректального ксенотрансплантата PDX-CRX-11201. Опухоли измеряли с использованием цифрового штангенциркуля и вычисляли объемы с помощью модифицированной формулы эллипсоида ½ × длина × ширина2. Мышей рандомизировали и стадировали по размеру опухоли 150-200 мм3. Начальную дозу биспецифического антитела против GUCY2c (GUCY2C-1608), биспецифического антитела отрицательного контроля или носителя вводили животным в день 0 и 2 миллиона культивированных T-клеток инокулировали на следующий день. Мышам вводили дозы в виде болюсных инъекций 0,2 мл еженедельно до 3 раз, и все введения соединений и T-клеток осуществляли внутривенно через хвостовую вену каждого животного. Комбинированные средства вводили, начиная со дня 0. mAb против VEGF (G6-31) вводили каждые 3 дня до 4 раз внутривенно через хвостовую вену. Акситиниб вводили дважды в сутки в течение до 14 дней с помощью желудочного зонда. Опухоли измеряли дважды в неделю вместе с непрерывным мониторингом признаков реакции "трансплантат-против-хозяина".
Биспецифическое антитело против GUCY2c и CD3, вводимое в количестве 0,05 мг/кг, демонстрировало частичный противоопухолевый ответ в модели адоптивного переноса PDX-11201. mAb против VEGF и акситиниб также демонстрировали частичный противоопухолевый ответ в этой модели при введении в отдельности в количестве 5 мг/кг и 15 мг/кг, соответственно. Комбинация биспецифического антитела против GUCY2c и CD3 с mAb против VEGF или акситинибом приводила к аддитивным противоопухолевым ответам, демонстрируя полное ингибирование опухоли.
GUCY2C-1608 демонстрировало повышенное ингибирование роста опухоли при комбинировании с mAb против VEGF, а также акситинибом в модели полученного из пациента ксенотрансплантата колоректальной карциномы PDX-CRX-11201 с адоптивным переносом T-клеток.
Пример 33: Оценка in vivo комбинации биспецифического антитела против GUCY2c и CD3 с антителом против PD1
PBMC человека размораживали в средах (X-VIVO 15 (Lonza). Добавляли 5% сывороточного альбумина человека (Gemini #100-318), 1% пенициллина/стрептомицина, 0,01 мМ 2-меркаптоэтанола на приблизительно 5 миллионов клеток на мл. Клетки центрифугировали и ресуспендировали в буфере Robosep (Stem Cell Technologies) в концентрации 50 миллионов клеток на мл. T-клетки выделяли с использованием набора для обогащения T-клеток человека EasySep (Stem Cell Technologies). T-клетки активировали и выращивали с использованием набора для активации/экспансии T-клеток человека (Miltenyi). В день 2 T-клетки переносили в устройство для культивирования клеток G-Rex для экспансии, в среды добавляли ИЛ-2 (Stem Cell Technologies) и восполняли через 2 дня. T-клетки собирали через 1 неделю после активации/экспансии. Во время сбора частицы удаляли с помощью магнита и клетки ресуспендировали в DPBS в количестве 1×107 клеток/мл для инокуляции in vivo.
Мышам NSG подкожно в бок инокулировали клетки линии колоректального рака LS1034. Опухоли измеряли с использованием цифрового штангенциркуля и вычисляли объемы с помощью модифицированной формулы эллипсоида ½ × длина × ширина2. Мышей рандомизировали и стадировали по размеру опухоли 150-200 мм3. Начальную дозу биспецифического антитела против GUCY2c (GUCY2C-1608) в количестве 0,03 мг/кг, биспецифического антитела отрицательного контроля в количестве 0,1 мг/кг, антитела против PD1 (биоаналог пембролизумаба D265A, мутант по эффекторной функции Fc hIgG1a) в количестве 5 мг/кг или носителя вводили животным в день 0 и 2 миллиона культивированных T-клеток инокулировали на следующий день. Мышам вводили дозы в виде болюсных инъекций 0,2 мл еженедельно до 3 раз, и все введения соединений и T-клеток осуществляли внутривенно через боковую хвостовую вену каждого животного. Опухоли измеряли дважды в неделю вместе с непрерывным мониторингом признаков реакции "трансплантат-против-хозяина".
Биспецифическое антитело против GUCY2c и CD3, вводимое в количестве 0,03 мг/кг, демонстрировало частичный противоопухолевый ответ в модели адоптивного переноса LS1034. Антитело против PD-1 не демонстрировало противоопухолевый ответ в этой модели при введении в количестве 10 мг/кг. Комбинирование биспецифического антитела против GUCY2c и CD3 с антителом против PD-1 приводило к аддитивному противоопухолевому ответу (фигура 24).
GUCY2C-1608 демонстрировало повышенное ингибирование роста опухоли при комбинировании с mAb против PD1 в модели полученного из линии клеток колоректальной карциномы ксенотрансплантата LS1034 с адоптивным переносом T-клеток.
Пример 34: Оценка in vivo комбинации биспецифического антитела против GUCY2c и CD3 с антителом против PDL1
PBMC человека размораживали в средах (X-VIVO 15 (Lonza). Добавляли 5% сывороточного альбумина человека (Gemini #100-318), 1% пенициллина/стрептомицина, 0,01 мМ 2-меркаптоэтанола на приблизительно 5 миллионов клеток на мл. Клетки центрифугировали и ресуспендировали в буфере Robosep (Stem Cell Technologies) в концентрации 50 миллионов клеток на мл. T-клетки выделяли с использованием набора для обогащения T-клеток человека EasySep (Stem Cell Technologies). T-клетки активировали и выращивали с использованием набора для активации/экспансии T-клеток человека (Miltenyi). В день 2 T-клетки переносили в устройство для культивирования клеток G-Rex для экспансии, в среды добавляли ИЛ-2 (Stem Cell Technologies) и восполняли через 2 дня. T-клетки собирали через 1 неделю после активации/экспансии. Во время сбора частицы удаляли с помощью магнита и клетки ресуспендировали в DPBS в количестве 1×107 клеток/мл для инокуляции in vivo.
Мышам NSG подкожно в бок инокулировали клетки линии колоректального рака LS1034. Опухоли измеряли с использованием цифрового штангенциркуля и вычисляли объемы с помощью модифицированной формулы эллипсоида ½ × длина × ширина2. Мышей рандомизировали и стадировали по размеру опухоли 150-200 мм3. Начальную дозу биспецифического антитела против GUCY2c (GUCY2C-1608) в количестве 0,03 мг/кг, или биспецифического антитела отрицательного контроля в количестве 0,1 мг/кг, или носителя вводили животным в день 0 и 2 миллиона культивированных T-клеток инокулировали на следующий день. Мышам вводили дозы в виде болюсных инъекций 0,2 мл еженедельно до 3 раз, и все введения соединений и T-клеток осуществляли внутривенно через боковую хвостовую вену каждого животного. Антитело против PDL1 вводили внутривенно, начиная со дня 0, каждые 3 дня до 6 раз. Опухоли измеряли дважды в неделю вместе с непрерывным мониторингом признаков реакции "трансплантат-против-хозяина".
Биспецифическое антитело против GUCY2c и CD3, вводимое в количестве 0,03 мг/кг, демонстрировало частичный противоопухолевый ответ в модели адоптивного переноса LS1034. Антитело против PD-L1 не демонстрировало противоопухолевый ответ в этой модели при введении в количестве 10 мг/кг. Комбинирование биспецифического антитела против GUCY2c и CD3 с антителом против PD-L1 приводило к аддитивному противоопухолевому ответу (фигура 25).
GUCY2C-1608 демонстрировало повышенное ингибирование роста опухоли при комбинировании с mAb против PD-L1 в модели полученного из линии клеток колоректальной карциномы ксенотрансплантата LS1034 с адоптивным переносом T-клеток.
Пример 35: Исследование фазы 1 с повышением дозы и изучением ранее установленной максимальной переносимой дозы в расширенной популяции для оценки безопасности, переносимости, фармакокинетики, фармакодинамики и противоопухолевой активности биспецифического антитела GUCY2c-1608 на пациентах с опухолями желудочно-кишечного тракта на поздней стадии
В этом примере описано открытое, многоцентровое, многодозовое исследование фазы 1 безопасности, PK и PD биспецифического антитела GUCY2c-1608. Исследование включает две части, повышения дозы в качестве единственного средства и подбора дозы в комбинации (с антителом против PD-1 или антителом против VEGF) (части 1A и 1B, соответственно) с последующим исследованием ранее установленной максимальной переносимой дозы в расширенной популяции (часть 2). Последовательным когортам пациентов с опухолями желудочно-кишечного тракта на поздней стадии/метастазирующими, включая колоректальную аденокарциному, аденокарциному желудка и пищевода, неявлявшихся кандидатами для схем лечения, о которых известно, что они приносят клиническую пользу, будут вводить увеличивающиеся дозы биспецифического антитела GUCY2c-1608 в части 1A исследования. В части 1B будут проводить подбор доз биспецифического антитела GUCY2c-1608 в комбинации (с антителом против PD-1 или антителом против VEGF) на пациентах с колоректальной аденокарциномой, аденокарциномами желудка и пищевода на поздней стадии/метастазирующими. В части 2 (фазе исследования ранее установленной максимальной переносимой дозы в расширенной популяции) будут оценивать рекомендуемую дозу фазы 2 (RP2D), выбранную в части 1 (фазе повышения дозы), в виде монотерапии или в комбинации (с антителом против PD-1 или антителом против VEGF) на пациентах с колоректальными аденокарциномами, ранее подвергавшимися лечению. Когорты для оценки биомаркеров, для которых необходимы парные биоптаты опухоли этих пациентов, можно использовать в частях 1A и 1B при приближении к максимальной переносимой дозе (MTD) или в случае RP2D/MTD и в подгруппах групп исследования ранее установленной максимальной переносимой дозы в расширенной популяции в части 2.
Увеличивающиеся дозы GUCY2c-1608 будут вводить в виде еженедельных подкожных (SC) инъекций в 21-дневных циклах. Предполагаемая начальная доза SC составляет 0,6 мкг/кг.
После определения GUCY2c-1608 RP2D/MTD в виде монотерапии будут собирать дозовые когорты части 1B для исследования безопасности, переносимости и предварительной противоопухолевой активности при этом уровне доз в комбинации (с антителом против PD-1 или антителом против VEGF). В фазу исследования ранее установленной максимальной переносимой дозы в расширенной популяции (часть 2) будут оценивать RP2D GUCY2c-1608 в виде монотерапии и комбинированного средства (с антителом против PD-1 или антителом против VEGF) на пациентах с колоректальными аденокарциномами на поздней стадии/метастазирующими.
Также будет иметь место по меньшей мере 72-часовой интервал между первой дозой, вводимой каждому из исходных пациентов (т.е. пациентов, включенных в исходную оценку дозолимитирующей токсичности (DLT)), включенных в исследование на новом уровне дозы. Всех пациентов будут наблюдать стационарно в течение по меньшей мере 48 часов или более после первой дозы SC цикла 1 дня 1 (C1D1) части 1.
Лечение с использованием GUCY2c-1608 будут продолжать до прогрессирования заболевания, отказа пациента или возникновения неприемлемой токсичности, в зависимости от того, что произойдет первым, если исследователь и медицинский наблюдатель не согласятся на лечение в условиях прогрессирования заболевания с учетом индивидуальной оценки пользы/риска.
Альтернативная схема дозирования с введением доз IV
Можно оценивать внутривенное (IV) введение, учитывая доступные клинические данные (включая безопасность/переносимость, PK, PD и др.), полученные с помощью когорт повышения дозы с введением SC.
В этом исследовании можно проводить оценку усиленной начальной дозы, чтобы сделать возможным последующий более высокий уровень дозы.
Критерии для повышения дозы
Для когорт введения уровни начальной дозы приведены в таблице 49; промежуточные дозы можно оценивать с учетом клинических результатов. В случае когорт введения SC во время использования начальных уровней повышения доз максимальные повышения дозы будут составлять до 250%, но их будут корректировать до не более 100% после наблюдения DLT.
Таблица 49
* Промежуточные дозы можно оценивать с учетом клинических результатов
Повышение дозы будут прекращать при достижении критериев прекращения, что определяет специалист в этой области.
Часть 1A исследования предназначена для определения MTD для схемы введения доз SC, однако, если схему дозирования переключают на IV, то будут определять MTD для введения доз IV. Если для IV необходимо включение усиленной начальной дозы, будут определять MTD для IV с использованием схемы введения усиленной начальной дозы.
Часть 1B будет предназначена для определения MTD GUCY2c-1608 в виде комбинированного средства (с антителом против PD-1 или антителом против VEGF).
RP2D является дозой, выбранной для дальнейшего исследования с учетом результатов фазы 1 исследования.
Выбор пациентов
Для повышения дозы (часть 1A и 1B): гистологическая или цитологическая диагностика колоректальной аденокарциномы или аденокарциномы желудка или пищевода на поздней стадии/метастазирующей, являющейся резистентной к стандартной терапии, или для которой стандартная терапия недоступна.
Для исследования ранее установленной максимальной переносимой дозы в расширенной популяции (часть 2): гистологическая или цитологическая диагностика колоректальной аденокарциномы, ранее подвергавшейся лечению, с по меньшей мере одним измеримым очагом, ранее не облученным, что определяют по RECIST версии 1.1, являющейся резистентной к стандартной терапии, или для которой стандартная терапия недоступна.
Лечение синдрома высвобождения цитокинов
Лечение синдрома высвобождения цитокинов (CRS) и пересмотренная оценочная система CRS адаптированы из D.W.Lee, et al: Current Concepts in the Diagnosis and Management of Cytokine Release Syndrome. Blood 124 (2014) 188-195.
Введение антитела против ИЛ-6: Рассматривают введение 4 мг/кг в течение 1 часа и введение второй дозы в случае отсутствия клинического улучшения в течение от 24 до 48 часов.
Описание каждого патента, патентной заявки и публикации, процитированных в настоящем описании, включено, таким образом, в настоящее описание в качестве ссылки в полном объеме.
Эквивалент
Хотя настоящее изобретение описано со ссылкой на конкретные варианты осуществления, очевидно, что специалисты в этой области могу разрабатывать другие варианты осуществления и варианты настоящего изобретения без отклонения от сущности и объема изобретения. Объект изобретения включает все эти другие варианты осуществления и варианты, включая комбинации и подкомбинации различных элементов, признаков, функций и/или свойств, представленных в настоящем описании. В следующей формуле изобретения конкретно указаны некоторые комбинации и покомбинации, рассматриваемые в качестве новых и неочевидных. Изобретение, воплощенное в других комбинациях и подкомбинациях признаков, функций, элементов и/или свойств, может быть заявлено в настоящей заявке, в заявках, по которым испрашивается приоритет по настоящей заявке, или в связанных заявках. Такие пункты формулы изобретения, направленные на другое изобретение или то же изобретение и имеющие более широкий, более узкий, равный или иной объем по сравнению с исходной формулой изобретения, также считают включенными в объем настоящего изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ПФАЙЗЕР ИНК.
Chang, Chew
Guntas, Gurkan
Katragadda, Madan
Mathur, Divya
Root, Adam
Mosyak, Lidia
LaVallie, Edward
<120> АНТИТЕЛА ПРОТИВ GUCY2C И ИХ ПРИМЕНЕНИЕ
<130> PC72377
<160> 445
<170> PatentIn version 3.5
<210> 1
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 1
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Asn Arg Ala Arg Gly Tyr Thr Ser Asp His Asn Pro
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr Val Leu Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 2
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 2
Gly Phe Thr Phe Ser Asp Tyr Tyr Met Thr
1 5 10
<210> 3
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 3
Phe Ile Arg Asn Arg Ala Arg Gly Tyr Thr Ser Asp His Asn Pro Ser
1 5 10 15
Val Lys Gly
<210> 4
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 4
Asp Arg Pro Ser Tyr Tyr Val Leu Asp Tyr
1 5 10
<210> 5
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 5
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 6
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 6
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala
1 5 10
<210> 7
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 7
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 8
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 8
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
1 5 10
<210> 9
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 9
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Asn Gln Ala Arg Gly Tyr Thr Ser Asp His Asn Pro
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr Val Leu Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 10
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 10
Phe Ile Arg Asn Gln Ala Arg Gly Tyr Thr Ser Asp His Asn Pro Ser
1 5 10 15
Val Lys Gly
<210> 11
<211> 114
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 11
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Asn Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Trp Met Gln Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Tyr Pro Gly Asp Gly Met Thr Thr Tyr Thr Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Val Arg Lys Gly Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val
100 105 110
Ser Ser
<210> 12
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 12
Gly Tyr Thr Phe Thr Thr Tyr Trp Met Gln
1 5 10
<210> 13
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 13
Ala Ile Tyr Pro Gly Asp Gly Met Thr Thr Tyr Thr Gln Lys Phe Lys
1 5 10 15
Asp
<210> 14
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 14
Lys Gly Met Asp Tyr
1 5
<210> 15
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 15
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Asn Leu Ser Cys Lys Ala Ser
20 25
<210> 16
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 16
Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly
1 5 10
<210> 17
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 17
Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln
1 5 10 15
Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val Tyr Tyr Cys Val Arg
20 25 30
<210> 18
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 18
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser
1 5 10
<210> 19
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 19
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Lys Tyr
20 25 30
Trp Met Gln Trp Ile Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Tyr Pro Gly Asp Gly Phe Thr Thr Tyr Thr Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asn Tyr Gly Arg Thr Tyr Gly Gly Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 20
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 20
Gly Tyr Thr Phe Thr Lys Tyr Trp Met Gln
1 5 10
<210> 21
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 21
Ala Ile Tyr Pro Gly Asp Gly Phe Thr Thr Tyr Thr Gln Lys Phe Lys
1 5 10 15
Gly
<210> 22
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 22
Arg Asn Tyr Gly Arg Thr Tyr Gly Gly Asp Tyr
1 5 10
<210> 23
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 23
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser
20 25
<210> 24
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 24
Trp Ile Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly
1 5 10
<210> 25
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 25
Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr Met Gln
1 5 10 15
Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 26
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 26
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Ile Glu Lys Phe
50 55 60
Lys Asn Lys Ala Thr Leu Thr Val Asp Lys Ser Ala Thr Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ala Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 27
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 27
Gly Tyr Thr Phe Thr Ser Tyr Trp Met His
1 5 10
<210> 28
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 28
Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Ile Glu Lys Phe Lys
1 5 10 15
Asn
<210> 29
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 29
Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val
1 5 10
<210> 30
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 30
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser
20 25
<210> 31
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 31
Lys Ala Thr Leu Thr Val Asp Lys Ser Ala Thr Thr Ala Tyr Met Gln
1 5 10 15
Leu Ser Ser Leu Thr Ala Glu Asp Ser Ala Val Tyr Tyr Cys Thr Arg
20 25 30
<210> 32
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 32
Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser
1 5 10
<210> 33
<211> 115
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 33
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Thr Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Ala Asn Tyr Asp Pro Lys Phe
50 55 60
Gln Gly Lys Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Phe Tyr Cys
85 90 95
Ser Ser Leu Gly Thr Gly Thr Tyr Trp Gly Gln Gly Thr Thr Leu Thr
100 105 110
Val Ser Ser
115
<210> 34
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 34
Gly Phe Asn Ile Lys Asp Thr Tyr Ile His
1 5 10
<210> 35
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 35
Arg Ile Asp Pro Ala Asn Gly Asn Ala Asn Tyr Asp Pro Lys Phe Gln
1 5 10 15
Gly
<210> 36
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 36
Leu Gly Thr Gly Thr Tyr
1 5
<210> 37
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 37
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Thr Ala Ser
20 25
<210> 38
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 38
Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile Gly
1 5 10
<210> 39
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 39
Lys Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn Thr Ala Tyr Leu Gln
1 5 10 15
Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Phe Tyr Cys Ser Ser
20 25 30
<210> 40
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 40
Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
1 5 10
<210> 41
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 41
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Asp Tyr
20 25 30
Ile Met Leu Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asn Ser Asn Pro Tyr Tyr Gly Ser Thr Ser Tyr Asn Leu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met His Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Tyr Tyr Gly Ser Ser Pro Tyr Trp Tyr Phe Asp Val
100 105 110
Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 42
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 42
Gly Tyr Ser Phe Thr Asp Tyr Ile Met Leu
1 5 10
<210> 43
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 43
Asn Ser Asn Pro Tyr Tyr Gly Ser Thr Ser Tyr Asn Leu Lys Phe Lys
1 5 10 15
Gly
<210> 44
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 44
Ser Gly Tyr Tyr Gly Ser Ser Pro Tyr Trp Tyr Phe Asp Val
1 5 10
<210> 45
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 45
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser
20 25
<210> 46
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 46
Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile Gly
1 5 10
<210> 47
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 47
Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met His
1 5 10 15
Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 48
<211> 114
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 48
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Trp Met Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Tyr Pro Gly Asp Gly Met Thr Thr Tyr Thr Gln Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Ala Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Arg Lys Gly Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser
<210> 49
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 49
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
1 5 10
<210> 50
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 50
Arg Phe Thr Ile Ser Ala Asp Lys Ala Lys Asn Ser Ala Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg
20 25 30
<210> 51
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 51
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 52
<211> 114
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 52
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Trp Met Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Tyr Pro Gly Asp Gly Met Thr Thr Tyr Thr Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Arg Lys Gly Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser
<210> 53
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 53
Ala Ile Tyr Pro Gly Asp Gly Met Thr Thr Tyr Thr Gln Lys Phe Gln
1 5 10 15
Gly
<210> 54
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 54
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser
20 25
<210> 55
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 55
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile Gly
1 5 10
<210> 56
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 56
Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr Met Glu
1 5 10 15
Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg
20 25 30
<210> 57
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 57
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Asp Tyr
20 25 30
Ile Met Leu Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Asn Ser Asn Pro Tyr Tyr Gly Ser Thr Ser Tyr Asn Leu Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Tyr Tyr Gly Ser Ser Pro Tyr Trp Tyr Phe Asp Val
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 58
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 58
Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 59
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 59
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
1 5 10
<210> 60
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 60
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Ile Glu Lys Phe
50 55 60
Lys Asn Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 61
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 61
Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Thr Arg
20 25 30
<210> 62
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 62
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Ile Glu Lys Phe
50 55 60
Lys Asn Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 63
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 63
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Ala
1 5 10
<210> 64
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 64
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Ile Glu Lys Phe
50 55 60
Lys Asn Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 65
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 65
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Ile His Pro Lys Phe
50 55 60
Lys Asn Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 66
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 66
Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Ile His Pro Lys Phe Lys
1 5 10 15
Asn
<210> 67
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 67
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Asn Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 68
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 68
Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Asn Glu Lys Phe Lys
1 5 10 15
Asn
<210> 69
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 69
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asn Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 70
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 70
Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Val His Glu Lys Phe Lys
1 5 10 15
Asn
<210> 71
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 71
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Ala Glu Gln Phe
50 55 60
Lys Asn Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 72
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 72
Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Ala Glu Gln Phe Lys
1 5 10 15
Asn
<210> 73
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 73
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Glu Leu Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 74
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 74
Gly Phe Thr Phe Ser Ser Tyr Trp Met His
1 5 10
<210> 75
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 75
Glu Ile Lys Pro Ser Asn Glu Leu Thr Asn Val His Glu Lys Phe Lys
1 5 10 15
Asp
<210> 76
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 76
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Thr Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 77
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 77
Thr Ser Ser Gln Ser Leu Phe Asn Val Arg Ser Arg Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 78
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 78
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 79
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 79
Lys Gln Ser Tyr Asp Leu Phe Thr
1 5
<210> 80
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 80
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys
20
<210> 81
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 81
Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 82
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 82
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 83
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 83
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 84
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 84
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 85
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 85
Lys Ser Ser Gln Ser Leu Phe Asn Val Arg Ser Arg Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 86
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 86
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys
20
<210> 87
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 87
Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Ser
1 5 10 15
<210> 88
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 88
Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys
20 25 30
<210> 89
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 89
Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
1 5 10
<210> 90
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 90
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Thr Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 91
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 91
Thr Ser Ser Gln Ser Leu Phe Asn Val Arg Ser Gln Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 92
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 92
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Pro Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Leu Glu Glu Asp Asp Thr Ala Met Phe Phe Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 93
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 93
Arg Ala Ser Glu Ser Val Asp Asn Tyr Gly Ile Ser Phe Met Asn
1 5 10 15
<210> 94
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 94
Ala Ala Ser Asn Pro Gly Ser
1 5
<210> 95
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 95
Gln Gln Ser Lys Glu Val Pro Tyr Thr
1 5
<210> 96
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 96
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys
20
<210> 97
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 97
Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 98
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 98
Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser
1 5 10 15
Leu Asn Ile His Pro Leu Glu Glu Asp Asp Thr Ala Met Phe Phe Cys
20 25 30
<210> 99
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 99
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
1 5 10
<210> 100
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 100
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Phe
20 25 30
Asp Ile Ser Phe Met Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Met Glu Glu Asp Asp Thr Ala Met Tyr Phe Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 101
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 101
Arg Ala Ser Glu Ser Val Asp Asn Phe Asp Ile Ser Phe Met Asn
1 5 10 15
<210> 102
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 102
Ala Ala Ser Asn Gln Gly Ser
1 5
<210> 103
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 103
Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser
1 5 10 15
Leu Asn Ile His Pro Met Glu Glu Asp Asp Thr Ala Met Tyr Phe Cys
20 25 30
<210> 104
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 104
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Gly Glu Ser Val Asp Asn Phe
20 25 30
Asp Ile Ser Phe Met Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Met Glu Glu Asp Asp Thr Ala Met Tyr Phe Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 105
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 105
Arg Ala Gly Glu Ser Val Asp Asn Phe Asp Ile Ser Phe Met Asn
1 5 10 15
<210> 106
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 106
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Asp Asp Ile Ala Met Tyr Phe Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 107
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 107
Arg Ala Ser Glu Ser Val Asp Tyr Tyr Gly Thr Ser Leu Met Gln
1 5 10 15
<210> 108
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 108
Ala Ala Ser Asn Val Glu Ser
1 5
<210> 109
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 109
Gln Gln Thr Arg Lys Val Tyr Thr
1 5
<210> 110
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 110
Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 111
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 111
Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser
1 5 10 15
Leu Asn Ile His Pro Val Glu Glu Asp Asp Ile Ala Met Tyr Phe Cys
20 25 30
<210> 112
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 112
Asp Ile Val Met Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Lys Gly Val Thr Thr Ser
20 25 30
Gly Tyr Ser Tyr Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Ala Ala Thr Tyr Tyr Cys Gln His Ser Arg
85 90 95
Glu Phe Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105 110
<210> 113
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 113
Arg Ala Ser Lys Gly Val Thr Thr Ser Gly Tyr Ser Tyr Met His
1 5 10 15
<210> 114
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 114
Leu Ala Ser Asn Leu Glu Ser
1 5
<210> 115
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 115
Gln His Ser Arg Glu Phe Pro Leu Thr
1 5
<210> 116
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 116
Asp Ile Val Met Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys
20
<210> 117
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 117
Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Asn Ile His Pro Val Glu Glu Glu Asp Ala Ala Thr Tyr Tyr Cys
20 25 30
<210> 118
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 118
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
1 5 10
<210> 119
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 119
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala Val Ser Val Gly
1 5 10 15
Glu Lys Val Thr Val Ser Cys Lys Ser Ser Gln Ser Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Arg Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser
65 70 75 80
Ile Ser Ser Val Lys Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 120
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 120
Lys Ser Ser Gln Ser Leu Leu Tyr Ser Ser Asn Gln Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 121
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 121
Gln Gln Tyr Tyr Ser Tyr Pro Thr
1 5
<210> 122
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 122
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala Val Ser Val Gly
1 5 10 15
Glu Lys Val Thr Val Ser Cys
20
<210> 123
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 123
Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 124
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 124
Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Ser Ile Ser Ser Val Lys Ala Glu Asp Leu Ala Val Tyr Tyr Cys
20 25 30
<210> 125
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 125
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met Asn Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Pro Gly Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 126
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 126
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys
20
<210> 127
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 127
Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 128
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 128
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
1 5 10
<210> 129
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 129
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Ala Ala Ser Asn Pro Gly Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 130
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 130
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys
20
<210> 131
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 131
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr
1 5 10 15
<210> 132
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 132
Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys
20 25 30
<210> 133
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 133
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 134
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 134
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ser Gln Ser Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 135
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 135
Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 136
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 136
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 137
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 137
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 138
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 138
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 139
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 139
Trp Tyr Gln Gln Lys Pro Gly Lys Pro Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 140
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 140
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Arg Tyr Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 141
<211> 7
<212> БЕЛОК
<213> Artificial Sequene
<220>
<223> Синтетическая пептидная последовательность
<400> 141
Ala Ala Ser Lys Arg Tyr Ser
1 5
<210> 142
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 142
Gln Gln Thr Arg Lys Ala Tyr Thr
1 5
<210> 143
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 143
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Trp Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 144
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 144
Ala Ala Ser Lys Leu Trp Ser
1 5
<210> 145
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 145
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Val Ala Pro Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 146
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 146
Ala Ala Ser Lys Val Ala Pro
1 5
<210> 147
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 147
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 148
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 148
Arg Ala Ser Glu Ser Val Asp Tyr Tyr Gly Ser Ser Leu Leu Gln
1 5 10 15
<210> 149
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 149
Ala Ala Ser Lys Leu Ala Ser
1 5
<210> 150
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 150
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Ile Ala Pro Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 151
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 151
Ala Ala Ser Asn Ile Ala Pro
1 5
<210> 152
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 152
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Arg Tyr Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Glu Tyr Thr Phe Gly Gln Gly Thr Lys Leu Lys Ile Lys
100 105 110
<210> 153
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 153
Arg Ala Ser Glu Ser Val Asp Tyr Tyr Gly Ser Ser Leu Met Gln
1 5 10 15
<210> 154
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 154
Gln Gln Thr Arg Lys Glu Tyr Thr
1 5
<210> 155
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 155
Phe Gly Gln Gly Thr Lys Leu Lys Ile Lys
1 5 10
<210> 156
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 156
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Ala Pro Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 157
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 157
Ala Ala Ser Asn Val Ala Pro
1 5
<210> 158
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 158
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser His Arg Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 159
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 159
Ala Ala Ser His Arg Ala Ser
1 5
<210> 160
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 160
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Glu Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 161
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 161
Ala Ala Ser Asn Val Ala Ser
1 5
<210> 162
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 162
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly His Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Arg Tyr Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Ser Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 163
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 163
Arg Ala Ser Glu Ser Val Asp Tyr Tyr Gly His Ser Leu Met Gln
1 5 10 15
<210> 164
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 164
Ala Ala Ser Asn Arg Tyr Ser
1 5
<210> 165
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 165
Gln Gln Thr Arg Lys Ala Tyr Ser
1 5
<210> 166
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 166
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Asp Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Glu Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 167
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 167
Arg Ala Ser Gln Ser Val Asp Tyr Tyr Gly Ser Ser Leu Leu Gln
1 5 10 15
<210> 168
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 168
Asp Ala Ser Lys Leu Ala Ser
1 5
<210> 169
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 169
Gln Gln Glu Arg Lys Ala Tyr Thr
1 5
<210> 170
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 170
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Glu Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 171
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 171
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Asp Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 172
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 172
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 173
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 173
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Asp Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Glu Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 174
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 174
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Glu Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 175
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 175
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Asp Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 176
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 176
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val
100 105
<210> 177
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 177
Gly Phe Thr Phe Ser Ser Tyr Trp Met Ser
1 5 10
<210> 178
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 178
Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val Lys
1 5 10 15
Gly
<210> 179
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 179
Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val
1 5
<210> 180
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 180
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 181
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 181
Arg Ala Ser Gln Ser Ile Ser Ser Tyr Leu Asn
1 5 10
<210> 182
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 182
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 183
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 183
Gln Gln Ser Tyr Ser Thr Pro Tyr Thr
1 5
<210> 184
<211> 107
<212> БЕЛОК
<213> Homo sapiens
<400> 184
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 185
<211> 98
<212> БЕЛОК
<213> Homo sapiens
<400> 185
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 186
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 186
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 187
<211> 217
<212> БЕЛОК
<213> Homo sapiens
<400> 187
Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 188
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 188
Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 189
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 189
Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 190
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 190
Gly Gly Gly Ser Gly Gly Gly Gly
1 5
<210> 191
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 191
Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5
<210> 192
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 192
Gly Cys Pro Pro Cys Pro
1 5
<210> 193
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 193
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
<210> 194
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 194
Arg Thr Ser Asp Gln
1 5
<210> 195
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 195
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 196
<211> 462
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 196
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Pro Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Leu Glu Glu Asp Asp Thr Ala Met Phe Phe Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly
115 120 125
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
130 135 140
Gly Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro
145 150 155 160
Gly Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Arg Ala Arg Gly
165 170 175
Tyr Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser
180 185 190
Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr
210 215 220
Tyr Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
225 230 235 240
Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
275 280 285
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro
355 360 365
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 197
<211> 456
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 197
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Gln Gln Ser Gly
115 120 125
Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Asn Leu Ser Cys Lys Ala
130 135 140
Ser Gly Tyr Thr Phe Thr Thr Tyr Trp Met Gln Trp Val Lys Gln Arg
145 150 155 160
Pro Gly Gln Gly Leu Glu Trp Ile Gly Ala Ile Tyr Pro Gly Asp Gly
165 170 175
Met Thr Thr Tyr Thr Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Ala
180 185 190
Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Ala Ser
195 200 205
Glu Asp Ser Ala Val Tyr Tyr Cys Val Arg Lys Gly Met Asp Tyr Trp
210 215 220
Gly Gln Gly Thr Ser Val Thr Val Ser Ser Gly Cys Pro Pro Cys Pro
225 230 235 240
Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys
245 250 255
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
260 265 270
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
275 280 285
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
290 295 300
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
305 310 315 320
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
325 330 335
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
340 345 350
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met
355 360 365
Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro
370 375 380
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
385 390 395 400
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
405 410 415
Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
420 425 430
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
435 440 445
Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 198
<211> 918
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 198
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Pro Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Leu Glu Glu Asp Asp Thr Ala Met Phe Phe Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly
115 120 125
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
130 135 140
Gly Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro
145 150 155 160
Gly Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Arg Ala Arg Gly
165 170 175
Tyr Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser
180 185 190
Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr
210 215 220
Tyr Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
225 230 235 240
Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
275 280 285
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro
355 360 365
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Asp Ile
450 455 460
Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly Glu Arg
465 470 475 480
Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Asn Val Arg Ser
485 490 495
Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
500 505 510
Lys Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln Ser Tyr
545 550 555 560
Asp Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Gly Gly
565 570 575
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Gln Gln Ser Gly Ala Glu
580 585 590
Leu Ala Arg Pro Gly Ala Ser Val Asn Leu Ser Cys Lys Ala Ser Gly
595 600 605
Tyr Thr Phe Thr Thr Tyr Trp Met Gln Trp Val Lys Gln Arg Pro Gly
610 615 620
Gln Gly Leu Glu Trp Ile Gly Ala Ile Tyr Pro Gly Asp Gly Met Thr
625 630 635 640
Thr Tyr Thr Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Ala Asp Lys
645 650 655
Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Ala Ser Glu Asp
660 665 670
Ser Ala Val Tyr Tyr Cys Val Arg Lys Gly Met Asp Tyr Trp Gly Gln
675 680 685
Gly Thr Ser Val Thr Val Ser Ser Gly Cys Pro Pro Cys Pro Ala Pro
690 695 700
Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
705 710 715 720
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
725 730 735
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
740 745 750
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
755 760 765
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
770 775 780
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
785 790 795 800
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
805 810 815
Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys
820 825 830
Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp
835 840 845
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
850 855 860
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser
865 870 875 880
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
885 890 895
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
900 905 910
Leu Ser Leu Ser Pro Gly
915
<210> 199
<211> 461
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 199
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Asp Asp Ile Ala Met Tyr Phe Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Arg Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 200
<211> 464
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 200
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Gln Val Gln Leu Gln Gln Pro Gly
115 120 125
Ala Glu Leu Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala
130 135 140
Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Met His Trp Val Lys Gln Arg
145 150 155 160
Pro Gly Gln Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn Gly
165 170 175
Leu Thr Asn Tyr Ile Glu Lys Phe Lys Asn Lys Ala Thr Leu Thr Val
180 185 190
Asp Lys Ser Ala Thr Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ala
195 200 205
Glu Asp Ser Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr Glu
210 215 220
Gly Tyr Trp Phe Phe Asp Val Trp Gly Ala Gly Thr Thr Val Thr Val
225 230 235 240
Ser Ser Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
275 280 285
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Ser
370 375 380
Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 201
<211> 925
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 201
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Asp Asp Ile Ala Met Tyr Phe Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Arg Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Asp Ile Val
450 455 460
Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly Glu Arg Ala
465 470 475 480
Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Asn Val Arg Ser Arg
485 490 495
Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
500 505 510
Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
515 520 525
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
530 535 540
Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln Ser Tyr Asp
545 550 555 560
Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu
580 585 590
Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr
595 600 605
Thr Phe Thr Ser Tyr Trp Met His Trp Val Lys Gln Arg Pro Gly Gln
610 615 620
Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn
625 630 635 640
Tyr Ile Glu Lys Phe Lys Asn Lys Ala Thr Leu Thr Val Asp Lys Ser
645 650 655
Ala Thr Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ala Glu Asp Ser
660 665 670
Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp
675 680 685
Phe Phe Asp Val Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser Gly
690 695 700
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
705 710 715 720
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
725 730 735
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
740 745 750
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
755 760 765
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
770 775 780
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
785 790 795 800
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
805 810 815
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
820 825 830
Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val
835 840 845
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
850 855 860
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
865 870 875 880
Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
885 890 895
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
900 905 910
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
915 920 925
<210> 202
<211> 463
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 202
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala Val Ser Val Gly
1 5 10 15
Glu Lys Val Thr Val Ser Cys Lys Ser Ser Gln Ser Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Arg Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser
65 70 75 80
Ile Ser Ser Val Lys Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Arg Ala Arg
165 170 175
Gly Tyr Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser
210 215 220
Tyr Tyr Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser
225 230 235 240
Ser Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser
245 250 255
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
260 265 270
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
275 280 285
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
290 295 300
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
305 310 315 320
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
325 330 335
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
340 345 350
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
355 360 365
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys
370 375 380
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
385 390 395 400
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
405 410 415
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
420 425 430
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
435 440 445
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 203
<211> 465
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 203
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Gln Gln Ser Gly
115 120 125
Pro Glu Leu Val Lys Pro Gly Ala Ser Val Lys Ile Ser Cys Lys Ala
130 135 140
Ser Gly Tyr Ser Phe Thr Asp Tyr Ile Met Leu Trp Val Lys Gln Ser
145 150 155 160
His Gly Lys Ser Leu Glu Trp Ile Gly Asn Ser Asn Pro Tyr Tyr Gly
165 170 175
Ser Thr Ser Tyr Asn Leu Lys Phe Lys Gly Lys Ala Thr Leu Thr Val
180 185 190
Asp Lys Ser Ser Ser Thr Ala Tyr Met His Leu Asn Ser Leu Thr Ser
195 200 205
Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Ser
210 215 220
Ser Pro Tyr Trp Tyr Phe Asp Val Trp Gly Ala Gly Thr Thr Val Thr
225 230 235 240
Val Ser Ser Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly
465
<210> 204
<211> 928
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 204
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala Val Ser Val Gly
1 5 10 15
Glu Lys Val Thr Val Ser Cys Lys Ser Ser Gln Ser Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Arg Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser
65 70 75 80
Ile Ser Ser Val Lys Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Arg Ala Arg
165 170 175
Gly Tyr Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser
210 215 220
Tyr Tyr Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser
225 230 235 240
Ser Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser
245 250 255
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
260 265 270
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
275 280 285
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
290 295 300
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
305 310 315 320
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
325 330 335
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
340 345 350
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
355 360 365
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys
370 375 380
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
385 390 395 400
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
405 410 415
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
420 425 430
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
435 440 445
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Asp
450 455 460
Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly Glu
465 470 475 480
Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Asn Val Arg
485 490 495
Ser Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro
500 505 510
Pro Lys Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val Pro
515 520 525
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
530 535 540
Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln Ser
545 550 555 560
Tyr Asp Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Gly
565 570 575
Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Gln Gln Ser Gly Pro
580 585 590
Glu Leu Val Lys Pro Gly Ala Ser Val Lys Ile Ser Cys Lys Ala Ser
595 600 605
Gly Tyr Ser Phe Thr Asp Tyr Ile Met Leu Trp Val Lys Gln Ser His
610 615 620
Gly Lys Ser Leu Glu Trp Ile Gly Asn Ser Asn Pro Tyr Tyr Gly Ser
625 630 635 640
Thr Ser Tyr Asn Leu Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp
645 650 655
Lys Ser Ser Ser Thr Ala Tyr Met His Leu Asn Ser Leu Thr Ser Glu
660 665 670
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Ser Ser
675 680 685
Pro Tyr Trp Tyr Phe Asp Val Trp Gly Ala Gly Thr Thr Val Thr Val
690 695 700
Ser Ser Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
705 710 715 720
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
725 730 735
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
740 745 750
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
755 760 765
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
770 775 780
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
785 790 795 800
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
805 810 815
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
820 825 830
Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Ser
835 840 845
Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
850 855 860
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
865 870 875 880
Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys
885 890 895
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
900 905 910
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
915 920 925
<210> 205
<211> 462
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 205
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met Asn Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Pro Gly Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly
115 120 125
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
130 135 140
Gly Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro
145 150 155 160
Gly Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Arg Ala Arg Gly
165 170 175
Tyr Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser
180 185 190
Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr
210 215 220
Tyr Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
225 230 235 240
Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
275 280 285
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro
355 360 365
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 206
<211> 456
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 206
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Tyr Thr Phe Thr Thr Tyr Trp Met Gln Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Ile Gly Ala Ile Tyr Pro Gly Asp Gly
165 170 175
Met Thr Thr Tyr Thr Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Ala
180 185 190
Asp Lys Ala Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg Lys Gly Met Asp Tyr Trp
210 215 220
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Cys Pro Pro Cys Pro
225 230 235 240
Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys
245 250 255
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
260 265 270
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
275 280 285
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
290 295 300
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
305 310 315 320
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
325 330 335
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
340 345 350
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met
355 360 365
Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro
370 375 380
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
385 390 395 400
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
405 410 415
Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
420 425 430
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
435 440 445
Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 207
<211> 918
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 207
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met Asn Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Pro Gly Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly
115 120 125
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
130 135 140
Gly Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro
145 150 155 160
Gly Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Arg Ala Arg Gly
165 170 175
Tyr Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser
180 185 190
Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr
210 215 220
Tyr Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
225 230 235 240
Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
275 280 285
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro
355 360 365
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Asp Ile
450 455 460
Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly Glu Arg
465 470 475 480
Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Asn Val Arg Ser
485 490 495
Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
500 505 510
Lys Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln Ser Tyr
545 550 555 560
Asp Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Gly Gly
565 570 575
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
580 585 590
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
595 600 605
Tyr Thr Phe Thr Thr Tyr Trp Met Gln Trp Val Arg Gln Ala Pro Gly
610 615 620
Lys Gly Leu Glu Trp Ile Gly Ala Ile Tyr Pro Gly Asp Gly Met Thr
625 630 635 640
Thr Tyr Thr Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Ala Asp Lys
645 650 655
Ala Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
660 665 670
Thr Ala Val Tyr Tyr Cys Val Arg Lys Gly Met Asp Tyr Trp Gly Gln
675 680 685
Gly Thr Leu Val Thr Val Ser Ser Gly Cys Pro Pro Cys Pro Ala Pro
690 695 700
Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
705 710 715 720
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
725 730 735
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
740 745 750
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
755 760 765
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
770 775 780
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
785 790 795 800
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
805 810 815
Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys
820 825 830
Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp
835 840 845
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
850 855 860
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser
865 870 875 880
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
885 890 895
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
900 905 910
Leu Ser Leu Ser Pro Gly
915
<210> 208
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 208
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 209
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 209
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Ile Glu Lys Phe
50 55 60
Lys Asn Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 210
<211> 461
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 210
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Arg Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 211
<211> 464
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 211
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Met His Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn Gly
165 170 175
Leu Thr Asn Tyr Ile Glu Lys Phe Lys Asn Arg Phe Thr Ile Ser Val
180 185 190
Asp Lys Ala Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr Glu
210 215 220
Gly Tyr Trp Phe Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val
225 230 235 240
Ser Ser Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
275 280 285
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Ser
370 375 380
Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 212
<211> 925
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 212
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Arg Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Asp Ile Val
450 455 460
Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly Glu Arg Ala
465 470 475 480
Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Asn Val Arg Ser Arg
485 490 495
Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
500 505 510
Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
515 520 525
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
530 535 540
Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Lys Gln Ser Tyr Asp
545 550 555 560
Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
580 585 590
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr
595 600 605
Thr Phe Thr Ser Tyr Trp Met His Trp Val Arg Gln Ala Pro Gly Lys
610 615 620
Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn
625 630 635 640
Tyr Ile Glu Lys Phe Lys Asn Arg Phe Thr Ile Ser Val Asp Lys Ala
645 650 655
Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
660 665 670
Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp
675 680 685
Phe Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly
690 695 700
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
705 710 715 720
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
725 730 735
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
740 745 750
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
755 760 765
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
770 775 780
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
785 790 795 800
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
805 810 815
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
820 825 830
Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val
835 840 845
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
850 855 860
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
865 870 875 880
Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
885 890 895
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
900 905 910
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
915 920 925
<210> 213
<211> 483
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 213
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asn Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Asp Ile Gln Leu Thr Gln
130 135 140
Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr
145 150 155 160
Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr Gly Ser Ser Leu Leu Gln
165 170 175
Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala
180 185 190
Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly
195 200 205
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp
210 215 220
Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg Lys Ala Tyr Thr Phe Gly
225 230 235 240
Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Ser Asp Gln Glu Pro Lys
245 250 255
Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
260 265 270
Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
275 280 285
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
290 295 300
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
305 310 315 320
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
325 330 335
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
340 345 350
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
355 360 365
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
370 375 380
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
385 390 395 400
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
405 410 415
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
420 425 430
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
435 440 445
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
450 455 460
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
465 470 475 480
Leu Ser Pro
<210> 214
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 214
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 215
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 215
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asn Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 216
<211> 461
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 216
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Gln Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 217
<211> 465
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 217
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Thr Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn
165 170 175
Gly Leu Thr Asn Val His Glu Lys Phe Lys Asn Arg Phe Thr Ile Ser
180 185 190
Val Asp Lys Ala Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr
210 215 220
Glu Gly Tyr Trp Phe Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr
225 230 235 240
Val Ser Ser Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly
465
<210> 218
<211> 926
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 218
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Gln Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Asp Ile Gln
450 455 460
Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val
465 470 475 480
Thr Ile Thr Cys Thr Ser Ser Gln Ser Leu Phe Asn Val Arg Ser Gln
485 490 495
Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
500 505 510
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Ser Arg
515 520 525
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
530 535 540
Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln Ser Tyr Asp
545 550 555 560
Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly
565 570 575
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
580 585 590
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
595 600 605
Tyr Thr Phe Thr Ser Tyr Trp Met His Trp Val Arg Gln Ala Pro Gly
610 615 620
Lys Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr
625 630 635 640
Asn Val His Glu Lys Phe Lys Asn Arg Phe Thr Ile Ser Val Asp Lys
645 650 655
Ala Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
660 665 670
Thr Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr
675 680 685
Trp Phe Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
690 695 700
Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val
705 710 715 720
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
725 730 735
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
740 745 750
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
755 760 765
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
770 775 780
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
785 790 795 800
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
805 810 815
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
820 825 830
Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala
835 840 845
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
850 855 860
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
865 870 875 880
Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg
885 890 895
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
900 905 910
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
915 920 925
<210> 219
<211> 10
<212> БЕЛОК
<213> Homo sapiens
<400> 219
Glu Pro Lys Ser Ser Asp Lys Thr His Thr
1 5 10
<210> 220
<211> 465
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 220
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Thr Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Ser Ser Tyr Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn
165 170 175
Glu Leu Thr Asn Val His Glu Lys Phe Lys Asp Arg Phe Thr Ile Ser
180 185 190
Val Asp Lys Ala Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr
210 215 220
Glu Gly Tyr Trp Phe Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr
225 230 235 240
Val Ser Ser Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly
465
<210> 221
<211> 926
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 221
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Gln Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Asp Ile Gln
450 455 460
Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val
465 470 475 480
Thr Ile Thr Cys Thr Ser Ser Gln Ser Leu Phe Asn Val Arg Ser Gln
485 490 495
Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
500 505 510
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Ser Arg
515 520 525
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
530 535 540
Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln Ser Tyr Asp
545 550 555 560
Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly
565 570 575
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
580 585 590
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
595 600 605
Phe Thr Phe Ser Ser Tyr Trp Met His Trp Val Arg Gln Ala Pro Gly
610 615 620
Lys Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn Glu Leu Thr
625 630 635 640
Asn Val His Glu Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys
645 650 655
Ala Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
660 665 670
Thr Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr
675 680 685
Trp Phe Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
690 695 700
Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val
705 710 715 720
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
725 730 735
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
740 745 750
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
755 760 765
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
770 775 780
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
785 790 795 800
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
805 810 815
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
820 825 830
Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala
835 840 845
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
850 855 860
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
865 870 875 880
Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg
885 890 895
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
900 905 910
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
915 920 925
<210> 222
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 222
Cys Pro Pro Cys Pro
1 5
<210> 223
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 223
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Glu Leu Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 224
<211> 1080
<212> БЕЛОК
<213> Homo sapiens
<400> 224
Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser Val
1 5 10 15
Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu Glu
20 25 30
Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln Asn
35 40 45
Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp Gly
50 55 60
Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly Leu
65 70 75 80
Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val Leu
85 90 95
Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp Thr
100 105 110
Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser Cys
115 120 125
Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys Leu
130 135 140
Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe Lys
145 150 155 160
Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu Thr
165 170 175
Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser Tyr
180 185 190
Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys Glu
195 200 205
Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile Ile
210 215 220
Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg Ala
225 230 235 240
Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp Gln
245 250 255
Tyr Leu Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val Leu
260 265 270
Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe Ser
275 280 285
Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu Asn
290 295 300
Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn Gly
305 310 315 320
Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu Thr
325 330 335
Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp Val
340 345 350
Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys Tyr
355 360 365
Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro Val
370 375 380
Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn Asp
385 390 395 400
Ile Thr Gly Arg Gly Pro Gln Ile Leu Met Ile Ala Val Phe Thr Leu
405 410 415
Thr Gly Ala Val Val Leu Leu Leu Leu Val Ala Leu Leu Met Leu Arg
420 425 430
Lys Tyr Arg Lys Asp Tyr Glu Leu Arg Gln Lys Lys Trp Ser His Ile
435 440 445
Pro Pro Glu Asn Ile Phe Pro Leu Glu Thr Asn Glu Thr Asn His Val
450 455 460
Ser Leu Lys Ile Asp Asp Asp Lys Arg Arg Asp Thr Ile Gln Arg Leu
465 470 475 480
Arg Gln Cys Lys Tyr Asp Lys Lys Arg Val Ile Leu Lys Asp Leu Lys
485 490 495
His Asn Asp Gly Asn Phe Thr Glu Lys Gln Lys Ile Glu Leu Asn Lys
500 505 510
Leu Leu Gln Ile Asp Tyr Tyr Asn Leu Thr Lys Phe Tyr Gly Thr Val
515 520 525
Lys Leu Asp Thr Met Ile Phe Gly Val Ile Glu Tyr Cys Glu Arg Gly
530 535 540
Ser Leu Arg Glu Val Leu Asn Asp Thr Ile Ser Tyr Pro Asp Gly Thr
545 550 555 560
Phe Met Asp Trp Glu Phe Lys Ile Ser Val Leu Tyr Asp Ile Ala Lys
565 570 575
Gly Met Ser Tyr Leu His Ser Ser Lys Thr Glu Val His Gly Arg Leu
580 585 590
Lys Ser Thr Asn Cys Val Val Asp Ser Arg Met Val Val Lys Ile Thr
595 600 605
Asp Phe Gly Cys Asn Ser Ile Leu Pro Pro Lys Lys Asp Leu Trp Thr
610 615 620
Ala Pro Glu His Leu Arg Gln Ala Asn Ile Ser Gln Lys Gly Asp Val
625 630 635 640
Tyr Ser Tyr Gly Ile Ile Ala Gln Glu Ile Ile Leu Arg Lys Glu Thr
645 650 655
Phe Tyr Thr Leu Ser Cys Arg Asp Arg Asn Glu Lys Ile Phe Arg Val
660 665 670
Glu Asn Ser Asn Gly Met Lys Pro Phe Arg Pro Asp Leu Phe Leu Glu
675 680 685
Thr Ala Glu Glu Lys Glu Leu Glu Val Tyr Leu Leu Val Lys Asn Cys
690 695 700
Trp Glu Glu Asp Pro Glu Lys Arg Pro Asp Phe Lys Lys Ile Glu Thr
705 710 715 720
Thr Leu Ala Lys Ile Phe Gly Leu Phe His Asp Gln Lys Asn Glu Ser
725 730 735
Tyr Met Asp Thr Leu Ile Arg Arg Leu Gln Leu Tyr Ser Arg Asn Leu
740 745 750
Glu His Leu Val Glu Glu Arg Thr Gln Leu Tyr Lys Ala Glu Arg Asp
755 760 765
Arg Ala Asp Arg Leu Asn Phe Met Leu Leu Pro Arg Leu Val Val Lys
770 775 780
Ser Leu Lys Glu Lys Gly Phe Val Glu Pro Glu Leu Tyr Glu Glu Val
785 790 795 800
Thr Ile Tyr Phe Ser Asp Ile Val Gly Phe Thr Thr Ile Cys Lys Tyr
805 810 815
Ser Thr Pro Met Glu Val Val Asp Met Leu Asn Asp Ile Tyr Lys Ser
820 825 830
Phe Asp His Ile Val Asp His His Asp Val Tyr Lys Val Glu Thr Ile
835 840 845
Gly Asp Ala Tyr Met Val Ala Ser Gly Leu Pro Lys Arg Asn Gly Asn
850 855 860
Arg His Ala Ile Asp Ile Ala Lys Met Ala Leu Glu Ile Leu Ser Phe
865 870 875 880
Met Gly Thr Phe Glu Leu Glu His Leu Pro Gly Leu Pro Ile Trp Ile
885 890 895
Arg Ile Gly Val His Ser Gly Pro Cys Ala Ala Gly Val Val Gly Ile
900 905 910
Lys Met Pro Arg Tyr Cys Leu Phe Gly Asp Thr Val Asn Thr Ala Ser
915 920 925
Arg Met Glu Ser Thr Gly Leu Pro Leu Arg Ile His Val Ser Gly Ser
930 935 940
Thr Ile Ala Ile Leu Lys Arg Thr Glu Cys Gln Phe Leu Tyr Glu Val
945 950 955 960
Arg Gly Glu Thr Tyr Leu Lys Gly Arg Gly Asn Glu Thr Thr Tyr Trp
965 970 975
Leu Thr Gly Met Lys Asp Gln Lys Phe Asn Leu Pro Thr Pro Pro Thr
980 985 990
Val Glu Asn Gln Gln Arg Leu Gln Ala Glu Phe Ser Asp Met Ile Ala
995 1000 1005
Asn Ser Leu Gln Lys Arg Gln Ala Ala Gly Ile Arg Ser Gln Lys
1010 1015 1020
Pro Arg Arg Val Ala Ser Tyr Lys Lys Gly Thr Leu Glu Tyr Leu
1025 1030 1035
Gln Leu Asn Thr Thr Asp Lys Glu Ser Thr Tyr Phe Thr Arg Thr
1040 1045 1050
Arg Pro Leu Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Ala Ala
1055 1060 1065
Asn Asp Ile Leu Asp Tyr Lys Asp Asp Asp Asp Lys
1070 1075 1080
<210> 225
<211> 3240
<212> ДНК
<213> Homo sapiens
<400> 225
tcccaggtga gtcagaactg ccacaatggc agctatgaaa tcagcgtcct gatgatgggc 60
aactcagcct ttgcagagcc cctgaaaaac ttggaagatg cggtgaatga ggggctggaa 120
atagtgagag gacgtctgca aaatgctggc ctaaatgtga ctgtgaacgc tactttcatg 180
tattcggatg gtctgattca taactcaggc gactgccgga gtagcacctg tgaaggcctc 240
gacctactca ggaaaatttc aaatgcacaa cggatgggct gtgtcctcat agggccctca 300
tgtacatact ccaccttcca gatgtacctt gacacagaat tgagctaccc catgatctca 360
gctggaagtt ttggattgtc atgtgactat aaagaaacct taaccaggct gatgtctcca 420
gctagaaagt tgatgtactt cttggttaac ttttggaaaa ccaacgatct gcccttcaaa 480
acttattcct ggagcacttc gtatgtttac aagaatggta cagaaactga ggactgtttc 540
tggtacctta atgctctgga ggctagcgtt tcctatttct cccacgaact cggctttaag 600
gtggtgttaa gacaagataa ggagtttcag gatatcttaa tggaccacaa caggaaaagc 660
aatgtgatta ttatgtgtgg tggtccagag ttcctctaca agctgaaggg tgaccgagca 720
gtggctgaag acattgtcat tattctagtg gatcttttca atgaccagta cttggaggac 780
aatgtcacag cccctgacta tatgaaaaat gtccttgttc tgacgctgtc tcctgggaat 840
tcccttctaa atagctcttt ctccaggaat ctatcaccaa caaaacgaga ctttgctctt 900
gcctatttga atggaatcct gctctttgga catatgctga agatatttct tgaaaatgga 960
gaaaatatta ccacccccaa atttgctcat gctttcagga atctcacttt tgaagggtat 1020
gacggtccag tgaccttgga tgactggggg gatgttgaca gtaccatggt gcttctgtat 1080
acctctgtgg acaccaagaa atacaaggtt cttttgacct atgataccca cgtaaataag 1140
acctatcctg tggatatgag ccccacattc acttggaaga actctaaact tcctaatgat 1200
attacaggcc ggggccctca gatcctgatg attgcagtct tcaccctcac tggagctgtg 1260
gtgctgctcc tgctcgtcgc tctcctgatg ctcagaaaat atagaaaaga ttatgaactt 1320
cgtcagaaaa aatggtccca cattcctcct gaaaatatct ttcctctgga gaccaatgag 1380
accaatcatg ttagcctcaa gatcgatgat gacaaaagac gagatacaat ccagagacta 1440
cgacagtgca aatacgacaa aaagcgagtg attctcaaag atctcaagca caatgatggt 1500
aatttcactg aaaaacagaa gatagaattg aacaagttgc ttcagattga ctattacaac 1560
ctgaccaagt tctacggcac agtgaaactt gataccatga tcttcggggt gatagaatac 1620
tgtgagagag gatccctccg ggaagtttta aatgacacaa tttcctaccc tgatggcaca 1680
ttcatggatt gggagtttaa gatctctgtc ttgtatgaca ttgctaaggg aatgtcatat 1740
ctgcactcca gtaagacaga agtccatggt cgtctgaaat ctaccaactg cgtagtggac 1800
agtagaatgg tggtgaagat cactgatttt ggctgcaatt ccattttacc tccaaaaaag 1860
gacctgtgga cagctccaga gcacctccgc caagccaaca tctctcagaa aggagatgtg 1920
tacagctatg ggatcatcgc acaggagatc attctgcgga aagaaacctt ctacactttg 1980
agctgtcggg accggaatga gaagattttc agagtggaaa attccaatgg aatgaaaccc 2040
ttccgcccag atttattctt ggaaacagca gaggaaaaag agctagaagt gtacctactt 2100
gtaaaaaact gttgggagga agatccagaa aagagaccag atttcaaaaa aattgagact 2160
acacttgcca agatatttgg actttttcat gaccaaaaaa atgaaagcta tatggatacc 2220
ttgatccgac gtctacagct atattctcga aacctggaac atctggtaga ggaaaggaca 2280
cagctgtaca aggcagagag ggacagggct gacagactta actttatgtt gcttccaagg 2340
ctagtggtaa agtctctgaa ggagaaaggc tttgtggagc cggaactata tgaggaagtt 2400
acaatctact tcagtgacat tgtaggtttc actactatct gcaaatacag cacccccatg 2460
gaagtggtgg acatgcttaa tgacatctat aagagttttg accacattgt tgatcatcat 2520
gatgtctaca aggtggaaac catcggtgat gcgtacatgg tggctagtgg tttgcctaag 2580
agaaatggca atcggcatgc aatagacatt gccaagatgg ccttggaaat cctcagcttc 2640
atggggacct ttgagctgga gcatcttcct ggcctcccaa tatggattcg cattggagtt 2700
cactctggtc cctgtgctgc tggagttgtg ggaatcaaga tgcctcgtta ttgtctattt 2760
ggagatacgg tcaacacagc ctctaggatg gaatccactg gcctcccttt gagaattcac 2820
gtgagtggct ccaccatagc catcctgaag agaactgagt gccagttcct ttatgaagtg 2880
agaggagaaa catacttaaa gggaagagga aatgagacta cctactggct gactgggatg 2940
aaggaccaga aattcaacct gccaacccct cctactgtgg agaatcaaca gcgtttgcaa 3000
gcagaatttt cagacatgat tgccaactct ttacagaaaa gacaggcagc agggataaga 3060
agccaaaaac ccagacgggt agccagctat aaaaaaggca ctctggaata cttgcagctg 3120
aataccacag acaaggagag cacctatttt acgcgtacgc ggccgctcga gcagaaactc 3180
atctcagaag aggatctggc agcaaatgat atcctggatt acaaggatga cgacgataag 3240
<210> 226
<211> 1080
<212> БЕЛОК
<213> Macaca fascicularis
<400> 226
Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser Val
1 5 10 15
Leu Met Met Asp Asn Ser Ala Phe Ala Glu Pro Leu Glu Asn Val Glu
20 25 30
Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln Asn
35 40 45
Ala Gly Leu Asn Val Thr Val Asn Ala Ser Phe Met Tyr Ser Asp Gly
50 55 60
Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly Leu
65 70 75 80
Asp Leu Leu Arg Lys Ile Ser Asn Ala Lys Arg Met Gly Cys Val Leu
85 90 95
Met Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp Thr
100 105 110
Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser Cys
115 120 125
Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys Leu
130 135 140
Thr Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe Lys
145 150 155 160
Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu Ser
165 170 175
Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser Tyr
180 185 190
Phe Ser His Glu Leu Ser Phe Lys Leu Val Leu Arg Gln Asp Lys Glu
195 200 205
Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile Val
210 215 220
Met Cys Gly Asp Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg Ala
225 230 235 240
Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp Gln
245 250 255
Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val Leu
260 265 270
Val Leu Thr Gln Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe Ser
275 280 285
Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu Asn
290 295 300
Gly Ile Leu Leu Phe Gly His Met Leu Lys Thr Phe Leu Glu Asn Gly
305 310 315 320
Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu Thr
325 330 335
Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp Val
340 345 350
Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys Tyr
355 360 365
Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Gln Thr Asn Pro Val
370 375 380
Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn Asp
385 390 395 400
Ile Thr Asp Arg Gly Pro Gln Ile Leu Met Ile Ala Val Phe Thr Leu
405 410 415
Thr Gly Ala Val Val Leu Leu Leu Leu Val Ala Leu Leu Met Leu Arg
420 425 430
Lys Tyr Lys Lys Asp Tyr Glu Leu Arg Gln Lys Lys Trp Ser His Ile
435 440 445
Pro Pro Glu Asn Ile Phe Pro Leu Glu Thr Asn Glu Thr Asn His Val
450 455 460
Ser Leu Lys Ile Asp Asp Asp Lys Arg Arg Asp Thr Ile Gln Arg Leu
465 470 475 480
Arg Gln Cys Lys Tyr Asp Lys Lys Arg Val Ile Leu Lys Asp Leu Lys
485 490 495
His Asn Asp Gly Asn Phe Thr Glu Lys Gln Lys Ile Glu Leu Asn Lys
500 505 510
Leu Leu Gln Ile Asp Tyr Tyr Asn Leu Thr Lys Phe Tyr Gly Thr Val
515 520 525
Lys Leu Asp Thr Met Ile Phe Gly Val Ile Glu Tyr Cys Glu Arg Gly
530 535 540
Ser Leu Arg Glu Val Leu Asn Asp Thr Ile Ser Tyr Pro Asp Gly Thr
545 550 555 560
Phe Met Asp Trp Glu Phe Lys Ile Ser Val Leu Tyr Asp Ile Ala Lys
565 570 575
Gly Met Ser Tyr Leu His Ser Ser Lys Thr Glu Val His Gly Arg Leu
580 585 590
Lys Ser Thr Asn Cys Val Val Asp Ser Arg Met Val Val Lys Ile Thr
595 600 605
Asp Phe Gly Cys Asn Ser Ile Leu Pro Pro Lys Lys Asp Leu Trp Thr
610 615 620
Ala Pro Glu His Leu Arg Gln Ala Asn Val Ser Gln Lys Gly Asp Val
625 630 635 640
Tyr Ser Tyr Gly Ile Ile Ala Gln Glu Ile Ile Leu Arg Lys Glu Thr
645 650 655
Phe Tyr Thr Ser Ser Cys Arg Asp Arg Asn Glu Lys Ile Phe Arg Val
660 665 670
Glu Asn Ser Asn Gly Met Lys Pro Phe Arg Pro Asp Leu Phe Leu Glu
675 680 685
Thr Ala Glu Glu Lys Glu Leu Glu Val Tyr Leu Leu Val Lys Ser Cys
690 695 700
Trp Glu Glu Asp Pro Glu Lys Arg Pro Asp Phe Lys Lys Ile Glu Thr
705 710 715 720
Thr Leu Ala Lys Ile Phe Gly Leu Phe His Asp Gln Lys Asn Glu Ser
725 730 735
Tyr Met Asp Thr Leu Ile Arg Arg Leu Gln Leu Tyr Ser Arg Asn Leu
740 745 750
Glu His Leu Val Glu Glu Arg Thr Gln Leu Tyr Lys Ala Glu Arg Asp
755 760 765
Arg Ala Asp Arg Leu Asn Phe Met Leu Leu Pro Arg Leu Val Val Lys
770 775 780
Ser Leu Lys Glu Lys Gly Phe Val Glu Pro Glu Leu Tyr Glu Glu Val
785 790 795 800
Thr Ile Tyr Phe Ser Asp Ile Val Gly Phe Thr Thr Ile Cys Lys Tyr
805 810 815
Ser Thr Pro Met Glu Val Val Asp Met Leu Asn Asp Ile Tyr Lys Ser
820 825 830
Phe Asp His Ile Val Asp His His Asp Val Tyr Lys Val Glu Thr Ile
835 840 845
Gly Asp Ala Tyr Met Val Ala Ser Gly Leu Pro Lys Arg Asn Gly Asn
850 855 860
Arg His Ala Ile Asp Ile Ala Lys Met Ala Leu Glu Ile Leu Ser Phe
865 870 875 880
Met Gly Thr Phe Glu Leu Glu His Leu Pro Gly Leu Pro Ile Trp Ile
885 890 895
Arg Ile Gly Val His Ser Gly Pro Cys Ala Ala Gly Val Val Gly Ile
900 905 910
Lys Met Pro Arg Tyr Cys Leu Phe Gly Asp Thr Val Asn Thr Ala Ser
915 920 925
Arg Met Glu Ser Thr Gly Leu Pro Leu Arg Ile His Val Ser Gly Ser
930 935 940
Thr Ile Ala Ile Leu Lys Arg Thr Glu Cys Gln Phe Leu Tyr Glu Val
945 950 955 960
Arg Gly Glu Thr Tyr Leu Lys Gly Arg Gly Asn Glu Thr Thr Tyr Trp
965 970 975
Leu Thr Gly Met Lys Asp Gln Lys Phe Asn Leu Pro Thr Pro Pro Thr
980 985 990
Val Glu Asn Gln Gln Arg Leu Gln Ala Glu Phe Ser Asp Met Ile Ala
995 1000 1005
Asn Ser Leu Gln Lys Arg Gln Ala Ala Gly Ile Arg Ser Gln Lys
1010 1015 1020
Pro Arg Arg Val Ala Ser Tyr Lys Lys Gly Thr Leu Glu Tyr Leu
1025 1030 1035
Gln Leu Asn Thr Thr Asp Lys Glu Ser Thr Tyr Phe Thr Arg Thr
1040 1045 1050
Arg Pro Leu Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Ala Ala
1055 1060 1065
Asn Asp Ile Leu Asp Tyr Lys Asp Asp Asp Asp Lys
1070 1075 1080
<210> 227
<211> 3240
<212> ДНК
<213> Macaca fascicularis
<400> 227
tcacaggtga gtcagaactg ccacaatggc agctatgaaa tcagcgtcct gatgatggac 60
aactcagcct ttgcagagcc cctggaaaac gtggaagatg cggtgaatga ggggctggaa 120
atagtgagag gacgtctgca aaacgctggc ctaaatgtga ctgtgaatgc ttctttcatg 180
tattcggatg gtctgattca taactccggc gactgccgga gcagcacctg tgaaggcctt 240
gacctactca ggaaaatttc aaatgcaaaa cggatgggct gtgtcctcat ggggccctca 300
tgtacatact ccaccttcca gatgtacctt gacacagaat tgagctaccc catgatctca 360
gctggaagtt ttggattgtc atgtgactat aaagaaacct taaccaggct gatgtctcca 420
gctagaaagt tgacatactt cttggttaac ttttggaaaa ccaatgatct acccttcaaa 480
acttattcct ggagcacttc gtatgtttac aagaatggta cggagtccga ggactgtttc 540
tggtacctta acgctctgga ggccagtgtt tcctatttct cccacgaact cagttttaag 600
ttggtgttaa gacaagataa ggagtttcag gatatcttaa tggaccacaa caggaaaagc 660
aatgtgattg ttatgtgtgg tgatccagag ttcctctaca agttgaaggg tgaccgagca 720
gtggctgaag acattgtcat tattctagtg gatcttttca atgaccagta ctttgaggac 780
aatgtcacag cccctgacta tatgaaaaat gtccttgttc tgacgcagtc tcctggcaat 840
tccctcctaa atagctcttt ctccaggaat ctatccccaa caaaacgaga ctttgctctt 900
gcctatttga atggaatcct gctctttgga catatgctaa agacatttct tgaaaatgga 960
gaaaatatta ccacccccaa atttgctcat gctttcagga atctcacttt tgaagggtat 1020
gacggtccag tgaccttgga tgactggggg gatgtggaca gtaccatggt gcttctgtat 1080
acgtctgtgg acaccaagaa atacaaggtt cttttgacct atgataccca cgtaaatcag 1140
accaaccctg tggatatgag ccccacattc acttggaaga actctaaact tcctaatgat 1200
attacagacc ggggccctca gatcctgatg attgcagtct tcaccctcac cggagctgtg 1260
gtgctgctcc tgcttgtcgc tctcctgatg ctcagaaaat ataaaaaaga ttatgaactt 1320
cgtcagaaaa aatggtccca cattcctcct gaaaatatct ttcctctgga gaccaatgag 1380
accaatcacg ttagcctgaa gatcgatgat gacaaaagac gagatacaat ccagagacta 1440
cgacagtgca aatacgacaa aaagcgagtg attctcaaag atctcaagca caatgatggt 1500
aatttcactg aaaaacagaa gatagaattg aacaagttgc ttcagattga ctattacaac 1560
ctgaccaagt tctatggcac cgtgaaactt gataccatga tcttcggggt gatagaatac 1620
tgtgagagag gatccctccg ggaagtttta aatgacacaa tttcctaccc tgatggcaca 1680
ttcatggatt gggagtttaa gatctctgtc ctgtatgaca ttgctaaggg aatgtcatat 1740
ctgcactcca gtaagacaga agtccatggt cgtctgaaat ctaccaactg cgtagtggac 1800
agtagaatgg tggtgaagat cactgatttt ggctgcaatt ccattttacc tccaaaaaaa 1860
gacctgtgga cagctccaga gcacctccgc caagccaacg tctctcagaa aggagatgtg 1920
tacagctacg ggatcatcgc acaggagatc atcctgcgga aagaaacctt ctacacttcg 1980
agctgtcgag accggaacga gaagattttc agagtggaaa attccaatgg aatgaaaccc 2040
ttccgtccag atttattctt ggaaacggca gaggaaaaag agctagaagt gtacctactt 2100
gtaaaaagct gttgggaaga agatccagaa aagagaccag atttcaaaaa aattgagact 2160
acacttgcca agatatttgg actttttcat gaccaaaaaa atgaaagcta tatggatacc 2220
ttgatccgac gtctacagct atattctcga aacctggaac atctggtaga ggaaaggaca 2280
cagctataca aggcagagag ggacagggct gacagactta actttatgtt gcttccaagg 2340
ctagtggtaa agtctctgaa ggagaaaggc tttgtagagc cggaactata tgaggaagtt 2400
acaatctact tcagtgacat tgtaggtttc actactatct gcaaatacag cacccccatg 2460
gaagtggtgg acatgcttaa tgacatctat aagagttttg accacattgt tgatcatcat 2520
gatgtctaca aggtggaaac cattggtgat gcctacatgg tggctagtgg tttgcctaag 2580
agaaatggca atcggcatgc aatagacatt gccaagatgg ccttggaaat cctcagcttc 2640
atggggacct ttgagctgga gcatcttcct ggcctcccaa tatggattcg cattggcgtt 2700
cactctggtc cctgcgctgc tggagttgtg ggaatcaaga tgcctcgtta ttgtctattt 2760
ggagatacag tcaacacagc ctctaggatg gaatccactg gcctcccttt gaggattcat 2820
gtgagtggct ccaccatagc cattctgaag agaactgagt gccagttcct gtatgaagtg 2880
agaggagaaa cgtacttaaa gggaagagga aatgagacta cctactggct gaccgggatg 2940
aaggaccaga aattcaacct gccaacccct cctactgtgg agaatcaaca gcgtttgcaa 3000
gcagaatttt cagacatgat tgccaactct ttacagaaaa gacaggcggc agggataaga 3060
agccaaaaac ccagacgagt agccagctat aaaaaaggca ctctggaata cttgcaactg 3120
aataccacgg acaaggagag cacctatttt acgcgtactc ggccgctcga gcagaaactc 3180
atctcagaag aggatctggc agcaaatgat atcctggatt acaaggatga cgacgataag 3240
<210> 228
<211> 1083
<212> БЕЛОК
<213> Mus musculus
<400> 228
Val Phe Trp Ala Ser Gln Val Arg Gln Asn Cys Arg Asn Gly Ser Tyr
1 5 10 15
Glu Ile Ser Val Leu Met Met Asp Asn Ser Ala Tyr Lys Glu Pro Met
20 25 30
Gln Asn Leu Arg Glu Ala Val Glu Glu Gly Leu Asp Ile Val Arg Lys
35 40 45
Arg Leu Arg Glu Ala Asp Leu Asn Val Thr Val Asn Ala Thr Phe Ile
50 55 60
Tyr Ser Asp Gly Leu Ile His Lys Ser Gly Asp Cys Arg Ser Ser Thr
65 70 75 80
Cys Glu Gly Leu Asp Leu Leu Arg Glu Ile Thr Arg Asp His Lys Met
85 90 95
Gly Cys Ala Leu Met Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met
100 105 110
Tyr Leu Asp Thr Glu Leu Asn Tyr Pro Met Ile Ser Ala Gly Ser Tyr
115 120 125
Gly Leu Ser Cys Asp Tyr Lys Glu Thr Leu Thr Arg Ile Leu Pro Pro
130 135 140
Ala Arg Lys Leu Met Tyr Phe Leu Val Asp Phe Trp Lys Val Asn Asn
145 150 155 160
Ala Ser Phe Lys Pro Phe Ser Trp Asn Ser Ser Tyr Val Tyr Lys Asn
165 170 175
Gly Ser Glu Pro Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala
180 185 190
Gly Val Ser Tyr Phe Ser Glu Val Leu Asn Phe Lys Asp Val Leu Arg
195 200 205
Arg Ser Glu Gln Phe Gln Glu Ile Leu Thr Gly His Asn Arg Lys Ser
210 215 220
Asn Val Ile Val Met Cys Gly Thr Pro Glu Ser Phe Tyr Asp Val Lys
225 230 235 240
Gly Asp Leu Gln Val Ala Glu Asp Thr Val Val Ile Leu Val Asp Leu
245 250 255
Phe Ser Asn His Tyr Phe Glu Glu Asn Thr Thr Ala Pro Glu Tyr Met
260 265 270
Asp Asn Val Leu Val Leu Thr Leu Pro Ser Glu Gln Ser Thr Ser Asn
275 280 285
Thr Ser Val Ala Glu Arg Phe Ser Ser Gly Arg Ser Asp Phe Ser Leu
290 295 300
Ala Tyr Leu Glu Gly Thr Leu Leu Phe Gly His Met Leu Gln Thr Phe
305 310 315 320
Leu Glu Asn Gly Glu Asn Val Thr Gly Pro Lys Phe Ala Arg Ala Phe
325 330 335
Arg Asn Leu Thr Phe Gln Gly Phe Ala Gly Pro Val Thr Leu Asp Asp
340 345 350
Ser Gly Asp Ile Asp Asn Ile Met Ser Leu Leu Tyr Val Ser Leu Asp
355 360 365
Thr Arg Lys Tyr Lys Val Leu Met Lys Tyr Asp Thr His Lys Asn Lys
370 375 380
Thr Ile Pro Val Ala Glu Asn Pro Asn Phe Ile Trp Lys Asn His Lys
385 390 395 400
Leu Pro Asn Asp Val Pro Gly Leu Gly Pro Gln Ile Leu Met Ile Ala
405 410 415
Val Phe Thr Leu Thr Gly Ile Leu Val Val Leu Leu Leu Ile Ala Leu
420 425 430
Leu Val Leu Arg Lys Tyr Arg Arg Asp His Ala Leu Arg Gln Lys Lys
435 440 445
Trp Ser His Ile Pro Ser Glu Asn Ile Phe Pro Leu Glu Thr Asn Glu
450 455 460
Thr Asn His Ile Ser Leu Lys Ile Asp Asp Asp Arg Arg Arg Asp Thr
465 470 475 480
Ile Gln Arg Val Arg Gln Cys Lys Tyr Asp Lys Lys Lys Val Ile Leu
485 490 495
Lys Asp Leu Lys His Ser Asp Gly Asn Phe Ser Glu Lys Gln Lys Ile
500 505 510
Asp Leu Asn Lys Leu Leu Gln Ser Asp Tyr Tyr Asn Leu Thr Lys Phe
515 520 525
Tyr Gly Thr Val Lys Leu Asp Thr Arg Ile Phe Gly Val Val Glu Tyr
530 535 540
Cys Glu Arg Gly Ser Leu Arg Glu Val Leu Asn Asp Thr Ile Ser Tyr
545 550 555 560
Pro Asp Gly Thr Phe Met Asp Trp Glu Phe Lys Ile Ser Val Leu Asn
565 570 575
Asp Ile Ala Lys Gly Met Ser Tyr Leu His Ser Ser Lys Ile Glu Val
580 585 590
His Gly Arg Leu Lys Ser Thr Asn Cys Val Val Asp Ser Arg Met Val
595 600 605
Val Lys Ile Thr Asp Phe Gly Cys Asn Ser Ile Leu Pro Pro Lys Lys
610 615 620
Asp Leu Trp Thr Ala Pro Glu His Leu Arg Gln Ala Thr Ile Ser Gln
625 630 635 640
Lys Gly Asp Val Tyr Ser Phe Ala Ile Ile Ala Gln Glu Ile Ile Leu
645 650 655
Arg Lys Glu Thr Phe Tyr Thr Leu Ser Cys Arg Asp His Asn Glu Lys
660 665 670
Ile Phe Arg Val Glu Asn Ser Tyr Gly Lys Pro Phe Arg Pro Asp Leu
675 680 685
Phe Leu Glu Thr Ala Asp Glu Lys Glu Leu Glu Val Tyr Leu Leu Val
690 695 700
Lys Ser Cys Trp Glu Glu Asp Pro Glu Lys Arg Pro Asp Phe Lys Lys
705 710 715 720
Ile Glu Ser Thr Leu Ala Lys Ile Phe Gly Leu Phe His Asp Gln Lys
725 730 735
Asn Glu Ser Tyr Met Asp Thr Leu Ile Arg Arg Leu Gln Leu Tyr Ser
740 745 750
Arg Asn Leu Glu His Leu Val Glu Glu Arg Thr Gln Leu Tyr Lys Ala
755 760 765
Glu Arg Asp Arg Ala Asp His Leu Asn Phe Met Leu Leu Pro Arg Leu
770 775 780
Val Val Lys Ser Leu Lys Glu Lys Gly Ile Val Glu Pro Glu Leu Tyr
785 790 795 800
Glu Glu Val Thr Ile Tyr Phe Ser Asp Ile Val Gly Phe Thr Thr Ile
805 810 815
Cys Lys Tyr Ser Thr Pro Met Glu Val Val Asp Met Leu Asn Asp Ile
820 825 830
Tyr Lys Ser Phe Asp Gln Ile Val Asp His His Asp Val Tyr Lys Val
835 840 845
Glu Thr Ile Gly Asp Ala Tyr Val Val Ala Ser Gly Leu Pro Met Arg
850 855 860
Asn Gly Asn Arg His Ala Val Asp Ile Ser Lys Met Ala Leu Asp Ile
865 870 875 880
Leu Ser Phe Ile Gly Thr Phe Glu Leu Glu His Leu Pro Gly Leu Pro
885 890 895
Val Trp Ile Arg Ile Gly Val His Ser Gly Pro Cys Ala Ala Gly Val
900 905 910
Val Gly Ile Lys Met Pro Arg Tyr Cys Leu Phe Gly Asp Thr Val Asn
915 920 925
Thr Ala Ser Arg Met Glu Ser Thr Gly Leu Pro Leu Arg Ile His Met
930 935 940
Ser Ser Ser Thr Ile Thr Ile Leu Lys Arg Thr Asp Cys Gln Phe Leu
945 950 955 960
Tyr Glu Val Arg Gly Glu Thr Tyr Leu Lys Gly Arg Gly Thr Glu Thr
965 970 975
Thr Tyr Trp Leu Thr Gly Met Lys Asp Gln Glu Tyr Asn Leu Pro Ser
980 985 990
Pro Pro Thr Val Glu Asn Gln Gln Arg Leu Gln Thr Glu Phe Ser Asp
995 1000 1005
Met Ile Val Ser Ala Leu Gln Lys Arg Gln Ala Ser Gly Lys Lys
1010 1015 1020
Ser Arg Arg Pro Thr Arg Val Ala Ser Tyr Lys Lys Gly Phe Leu
1025 1030 1035
Glu Tyr Met Gln Leu Asn Asn Ser Asp His Asp Ser Thr Tyr Phe
1040 1045 1050
Thr Arg Thr Arg Pro Leu Glu Gln Lys Leu Ile Ser Glu Glu Asp
1055 1060 1065
Leu Ala Ala Asn Asp Ile Leu Asp Tyr Lys Asp Asp Asp Asp Lys
1070 1075 1080
<210> 229
<211> 3249
<212> ДНК
<213> Mus musculus
<400> 229
gtgttctggg cctctcaggt gaggcagaac tgccgcaatg gcagctacga gatcagcgtc 60
ctgatgatgg acaactcagc ctacaaagaa cctatgcaaa acctgaggga ggctgtggag 120
gaaggactgg acatagtgcg aaagcgcctg cgtgaagccg acctaaatgt gactgtgaac 180
gcgactttca tctactccga cggtctgatt cataagtcag gtgactgccg gagcagcacc 240
tgtgaaggcc ttgacctact cagggagatt acaagagatc ataagatggg ctgcgccctc 300
atggggccct cgtgcacgta ttccaccttc cagatgtacc tcgacacaga gttgaactat 360
cccatgattt ccgctggaag ttatggattg tcctgtgact ataaggaaac cctaaccagg 420
atcctgcctc cagccaggaa gctgatgtac ttcttggtcg atttctggaa agtcaacaat 480
gcatctttca aacccttttc ctggaactct tcgtatgttt acaagaatgg atcggaacct 540
gaagattgtt tctggtacct caatgctctg gaggctgggg tgtcctattt ttctgaggtg 600
ctcaacttca aggatgtact gagacgcagc gaacagttcc aggaaatctt aacaggccat 660
aacagaaaga gcaatgtgat tgttatgtgt ggcacgccag aaagcttcta tgatgtgaaa 720
ggtgacctcc aagtggctga agatactgtt gtcatcctgg tagatctgtt cagtaaccat 780
tactttgagg agaacaccac agctcctgag tatatggaca atgtcctcgt cctgacgctg 840
ccgtctgaac agtccacctc aaacacctct gtcgccgaga ggttttcatc ggggagaagt 900
gacttttctc tcgcttactt ggagggaacc ttgctatttg gacacatgct gcagacgttt 960
cttgaaaatg gagaaaatgt cacgggtccc aagtttgctc gtgcattcag gaatctcact 1020
tttcaaggct ttgcaggacc tgtgactctg gatgacagtg gggacattga caacattatg 1080
tcccttctgt atgtgtctct ggataccagg aaatacaagg ttcttatgaa gtatgacacc 1140
cacaaaaaca aaactattcc ggtggctgag aaccccaact tcatctggaa gaaccacaag 1200
ctccccaatg acgttcctgg gctgggccct caaatcctga tgattgccgt cttcacgctc 1260
acggggatcc tggtagttct gctgctgatt gccctcctcg tgctgagaaa atacagaaga 1320
gatcatgcac ttcgacagaa gaaatggtcc cacattcctt ctgaaaacat ctttcctctg 1380
gagaccaacg agaccaacca catcagcctg aagattgacg atgacaggag acgagacaca 1440
atccagagag tgcgacagtg caaatacgac aagaagaaag tgattctgaa agacctcaag 1500
cacagcgacg ggaacttcag tgagaagcag aagatagacc tgaacaagtt gctgcagtct 1560
gactactaca acctgactaa gttctacggc accgtgaagc tggacaccag gatctttggg 1620
gtggttgagt actgcgagag gggatccctc cgggaagtgt taaacgacac aatttcctac 1680
cctgacggca cgttcatgga ttgggagttt aagatctctg tcttaaatga catcgctaag 1740
gggatgtcct acctgcactc cagtaagatt gaagtccacg ggcgtctcaa atccaccaac 1800
tgcgtggtgg acagccgcat ggtggtgaag atcaccgact ttgggtgcaa ttccatcctg 1860
cctccaaaaa aagacctgtg gacggccccg gagcacctgc gccaggccac catctctcag 1920
aaaggagacg tgtacagctt cgccatcatt gcccaggaga tcatcctccg taaggagact 1980
ttttacacgc tgagctgtcg ggatcacaat gagaagattt tcagagtgga aaattcatac 2040
gggaaacctt tccgcccaga cctcttcctg gagactgcag atgagaagga gctggaggtc 2100
tatctacttg tcaaaagctg ttgggaggag gatccagaaa agaggccaga tttcaagaaa 2160
atcgagagca cactggccaa gatatttggc cttttccatg accagaaaaa cgagtcttac 2220
atggacacct tgatccgacg tctccagctg tactctcgaa acctggaaca tctggtggag 2280
gaaaggactc agctgtacaa ggcggagagg gacagggctg accaccttaa cttcatgctc 2340
ctcccacggc tggtggtaaa gtcactgaag gagaaaggca tcgtggagcc agagctgtac 2400
gaagaagtca caatctactt cagtgacatt gtgggcttca ccaccatctg caagtatagc 2460
acgcccatgg aggtggtgga catgctcaac gacatctaca agagctttga ccagattgtg 2520
gaccaccatg acgtctacaa ggtagaaacc atcggtgacg cctacgtggt ggccagcggt 2580
ctgcctatga gaaacggcaa ccgacacgcg gtagacattt ccaagatggc cttggacatc 2640
ctcagcttca tagggacctt tgagttggag catctccctg gcctccccgt gtggatccgc 2700
attggagttc attctgggcc ctgcgctgct ggtgttgtgg ggatcaagat gcctcgctat 2760
tgcctgtttg gagacactgt caacactgcc tccaggatgg aatccaccgg cctccccttg 2820
aggattcaca tgagcagctc caccataacc atcctgaaga gaacggattg ccagttcctg 2880
tatgaagtga ggggagaaac ctacttaaag ggaagaggga cagagaccac atactggctg 2940
actgggatga aggaccaaga atacaacctg ccatccccac cgacagtgga gaaccaacag 3000
cgtctgcaga ctgagttctc agacatgatc gttagcgcct tacagaaaag acaggcctcg 3060
ggcaagaaga gccggaggcc cactcgggtg gccagctaca agaaaggctt tctggaatac 3120
atgcagctga acaattcaga ccacgatagc acctatttta cgcgtacgcg gccgctcgag 3180
cagaaactca tctcagaaga ggatctggca gcaaatgata tcctggatta caaggatgac 3240
gacgataag 3249
<210> 230
<211> 672
<212> БЕЛОК
<213> Homo sapiens
<400> 230
Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser Val
1 5 10 15
Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu Glu
20 25 30
Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln Asn
35 40 45
Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp Gly
50 55 60
Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly Leu
65 70 75 80
Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val Leu
85 90 95
Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp Thr
100 105 110
Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser Cys
115 120 125
Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys Leu
130 135 140
Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe Lys
145 150 155 160
Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu Thr
165 170 175
Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser Tyr
180 185 190
Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys Glu
195 200 205
Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile Ile
210 215 220
Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg Ala
225 230 235 240
Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp Gln
245 250 255
Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val Leu
260 265 270
Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe Ser
275 280 285
Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu Asn
290 295 300
Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn Gly
305 310 315 320
Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu Thr
325 330 335
Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp Val
340 345 350
Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys Tyr
355 360 365
Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro Val
370 375 380
Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn Asp
385 390 395 400
Ile Thr Gly Arg Gly Pro Gln Gly Gly Gly Gly Ser Glu Asn Leu Tyr
405 410 415
Phe Gln Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Pro Arg Gly
420 425 430
Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys Pro Ala Pro Asn Leu
435 440 445
Glu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Ile Lys Asp Val
450 455 460
Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys Val Val Val Asp Val
465 470 475 480
Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp Phe Val Asn Asn Val
485 490 495
Glu Val His Thr Ala Gln Thr Gln Thr His Arg Glu Asp Tyr Asn Ser
500 505 510
Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln His Gln Asp Trp Met
515 520 525
Ser Gly Lys Ala Phe Ala Cys Ala Val Asn Asn Lys Asp Leu Pro Ala
530 535 540
Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly Ser Val Arg Ala Pro
545 550 555 560
Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu Met Thr Lys Lys Gln
565 570 575
Val Thr Leu Thr Cys Met Val Thr Asp Phe Met Pro Glu Asp Ile Tyr
580 585 590
Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu Asn Tyr Lys Asn Thr
595 600 605
Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe Met Tyr Ser Lys Leu
610 615 620
Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn Ser Tyr Ser Cys Ser
625 630 635 640
Val Val His Glu Gly Leu His Asn His His Thr Thr Lys Ser Phe Ser
645 650 655
Arg Thr Pro Gly Gly Gly Pro Pro Asp Tyr Lys Asp Asp Asp Asp Lys
660 665 670
<210> 231
<211> 2076
<212> ДНК
<213> Homo sapiens
<400> 231
atggagacag acacactgct cctgtgggtg ctgcttctgt gggtgccagg ttccactgga 60
tcacaggtga gtcagaactg ccacaatggc agctatgaaa tcagcgtcct gatgatgggc 120
aactcagcct ttgcagagcc cctgaaaaac ttggaagatg cggtgaatga ggggctggaa 180
atagtgagag gacgtctgca aaatgctggc ctaaatgtga ctgtgaacgc tactttcatg 240
tattcggatg gtctgattca taactcaggc gactgccgga gtagcacctg tgaaggcctc 300
gacctactca ggaaaatttc aaatgcacaa cggatgggct gtgtcctcat agggccctca 360
tgtacatact ccaccttcca gatgtacctt gacacagaat tgagctaccc catgatctca 420
gctggaagtt ttggattgtc atgtgactat aaagaaacct taaccaggct gatgtctcca 480
gctagaaagt tgatgtactt cttggttaac ttttggaaaa ccaacgatct gcccttcaaa 540
acttattcct ggagcacttc gtatgtttac aagaatggta cagaaactga ggactgtttc 600
tggtacctta atgctctgga ggctagcgtt tcctatttct cccacgaact cggctttaag 660
gtggtgttaa gacaagataa ggagtttcag gatatcttaa tggaccacaa caggaaaagc 720
aatgtgatta ttatgtgtgg tggtccagag ttcctctaca agctgaaggg tgaccgagca 780
gtggctgaag acattgtcat tattctagtg gatcttttca atgaccagta ctttgaggac 840
aatgtcacag cccctgacta tatgaaaaat gtccttgttc tgacgctgtc tcctgggaat 900
tcccttctaa atagctcttt ctccaggaat ctatcaccaa caaaacgaga ctttgctctt 960
gcctatttga atggaatcct gctctttgga catatgctga agatatttct tgaaaatgga 1020
gaaaatatta ccacccccaa atttgctcat gctttcagga atctcacttt tgaagggtat 1080
gacggtccag tgaccttgga tgactggggg gatgttgaca gtaccatggt gcttctgtat 1140
acctctgtgg acaccaagaa atacaaggtt cttttgacct atgataccca cgtaaataag 1200
acctatcctg tggatatgag ccccacattc acttggaaga actctaaact tcctaatgat 1260
attacaggcc ggggccctca gggaggcgga ggttccgaga atttatactt ccagggcgga 1320
ggcggttccg gcggcggagg aagcgagccc cgcggaccga caatcaagcc ctgtcctcca 1380
tgcaaatgcc cagcacctaa cctcgagggt ggaccatccg tcttcatctt ccctccaaag 1440
atcaaggatg tactcatgat ctccctgagc cccatagtca catgtgtggt ggtggatgtg 1500
agcgaggatg acccagatgt ccagatcagc tggtttgtga acaacgtgga agtacacaca 1560
gctcagacac aaacccatag agaggattac aacagtactc tccgggtggt cagtgccctc 1620
cccatccagc accaggactg gatgagtggc aaggctttcg catgcgccgt caacaacaaa 1680
gacctcccag cgcccatcga gagaaccatc tcaaaaccca aagggtcagt aagagctcca 1740
caggtatatg tcttgcctcc accagaagaa gagatgacta agaaacaggt cactctgacc 1800
tgcatggtca cagacttcat gcctgaagac atttacgtgg agtggaccaa caacgggaaa 1860
acagagctaa actacaagaa cactgaacca gtcctggact ctgatggttc ttacttcatg 1920
tacagcaagc tgagagtgga aaagaagaac tgggtggaaa gaaatagcta ctcctgttca 1980
gtggtccacg agggtctgca caatcaccac acgactaaga gcttctcccg gactccgggt 2040
ggcgggccac ccgactacaa ggacgacgat gacaaa 2076
<210> 232
<211> 418
<212> БЕЛОК
<213> Homo sapiens
<400> 232
Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser Val
1 5 10 15
Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu Glu
20 25 30
Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln Asn
35 40 45
Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp Gly
50 55 60
Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly Leu
65 70 75 80
Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val Leu
85 90 95
Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp Thr
100 105 110
Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser Cys
115 120 125
Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys Leu
130 135 140
Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe Lys
145 150 155 160
Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu Thr
165 170 175
Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser Tyr
180 185 190
Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys Glu
195 200 205
Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile Ile
210 215 220
Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg Ala
225 230 235 240
Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp Gln
245 250 255
Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val Leu
260 265 270
Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe Ser
275 280 285
Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu Asn
290 295 300
Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn Gly
305 310 315 320
Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu Thr
325 330 335
Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp Val
340 345 350
Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys Tyr
355 360 365
Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro Val
370 375 380
Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn Asp
385 390 395 400
Ile Thr Gly Arg Gly Pro Gln Gly Gly Gly Gly Ser Glu Asn Leu Tyr
405 410 415
Phe Gln
<210> 233
<211> 1254
<212> ДНК
<213> Homo sapiens
<400> 233
tcacaggtga gtcagaactg ccacaatggc agctatgaaa tcagcgtcct gatgatgggc 60
aactcagcct ttgcagagcc cctgaaaaac ttggaagatg cggtgaatga ggggctggaa 120
atagtgagag gacgtctgca aaatgctggc ctaaatgtga ctgtgaacgc tactttcatg 180
tattcggatg gtctgattca taactcaggc gactgccgga gtagcacctg tgaaggcctc 240
gacctactca ggaaaatttc aaatgcacaa cggatgggct gtgtcctcat agggccctca 300
tgtacatact ccaccttcca gatgtacctt gacacagaat tgagctaccc catgatctca 360
gctggaagtt ttggattgtc atgtgactat aaagaaacct taaccaggct gatgtctcca 420
gctagaaagt tgatgtactt cttggttaac ttttggaaaa ccaacgatct gcccttcaaa 480
acttattcct ggagcacttc gtatgtttac aagaatggta cagaaactga ggactgtttc 540
tggtacctta atgctctgga ggctagcgtt tcctatttct cccacgaact cggctttaag 600
gtggtgttaa gacaagataa ggagtttcag gatatcttaa tggaccacaa caggaaaagc 660
aatgtgatta ttatgtgtgg tggtccagag ttcctctaca agctgaaggg tgaccgagca 720
gtggctgaag acattgtcat tattctagtg gatcttttca atgaccagta ctttgaggac 780
aatgtcacag cccctgacta tatgaaaaat gtccttgttc tgacgctgtc tcctgggaat 840
tcccttctaa atagctcttt ctccaggaat ctatcaccaa caaaacgaga ctttgctctt 900
gcctatttga atggaatcct gctctttgga catatgctga agatatttct tgaaaatgga 960
gaaaatatta ccacccccaa atttgctcat gctttcagga atctcacttt tgaagggtat 1020
gacggtccag tgaccttgga tgactggggg gatgttgaca gtaccatggt gcttctgtat 1080
acctctgtgg acaccaagaa atacaaggtt cttttgacct atgataccca cgtaaataag 1140
acctatcctg tggatatgag ccccacattc acttggaaga actctaaact tcctaatgat 1200
attacaggcc ggggccctca gggaggcgga ggttccgaga atttatactt ccag 1254
<210> 234
<211> 672
<212> БЕЛОК
<213> Macaca fascicularis
<400> 234
Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser Val
1 5 10 15
Leu Met Met Asp Asn Ser Ala Phe Ala Glu Pro Leu Glu Asn Val Glu
20 25 30
Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln Asn
35 40 45
Ala Gly Leu Asn Val Thr Val Asn Ala Ser Phe Met Tyr Ser Asp Gly
50 55 60
Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly Leu
65 70 75 80
Asp Leu Leu Arg Lys Ile Ser Asn Ala Lys Arg Met Gly Cys Val Leu
85 90 95
Met Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp Thr
100 105 110
Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser Cys
115 120 125
Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys Leu
130 135 140
Thr Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe Lys
145 150 155 160
Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu Ser
165 170 175
Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser Tyr
180 185 190
Phe Ser His Glu Leu Ser Phe Lys Leu Val Leu Arg Gln Asp Lys Glu
195 200 205
Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile Val
210 215 220
Met Cys Gly Asp Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg Ala
225 230 235 240
Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp Gln
245 250 255
Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val Leu
260 265 270
Val Leu Thr Gln Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe Ser
275 280 285
Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu Asn
290 295 300
Gly Ile Leu Leu Phe Gly His Met Leu Lys Thr Phe Leu Glu Asn Gly
305 310 315 320
Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu Thr
325 330 335
Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp Val
340 345 350
Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys Tyr
355 360 365
Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Gln Thr Asn Pro Val
370 375 380
Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn Asp
385 390 395 400
Ile Thr Asp Arg Gly Pro Gln Gly Gly Gly Gly Ser Glu Asn Leu Tyr
405 410 415
Phe Gln Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Pro Arg Gly
420 425 430
Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys Pro Ala Pro Asn Leu
435 440 445
Glu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Ile Lys Asp Val
450 455 460
Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys Val Val Val Asp Val
465 470 475 480
Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp Phe Val Asn Asn Val
485 490 495
Glu Val His Thr Ala Gln Thr Gln Thr His Arg Glu Asp Tyr Asn Ser
500 505 510
Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln His Gln Asp Trp Met
515 520 525
Ser Gly Lys Ala Phe Ala Cys Ala Val Asn Asn Lys Asp Leu Pro Ala
530 535 540
Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly Ser Val Arg Ala Pro
545 550 555 560
Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu Met Thr Lys Lys Gln
565 570 575
Val Thr Leu Thr Cys Met Val Thr Asp Phe Met Pro Glu Asp Ile Tyr
580 585 590
Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu Asn Tyr Lys Asn Thr
595 600 605
Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe Met Tyr Ser Lys Leu
610 615 620
Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn Ser Tyr Ser Cys Ser
625 630 635 640
Val Val His Glu Gly Leu His Asn His His Thr Thr Lys Ser Phe Ser
645 650 655
Arg Thr Pro Gly Gly Gly Pro Pro Asp Tyr Lys Asp Asp Asp Asp Lys
660 665 670
<210> 235
<211> 2016
<212> ДНК
<213> Macaca fascicularis
<400> 235
tcacaggtga gtcagaactg ccacaatggc agctatgaaa tcagcgtcct gatgatggac 60
aactcagcct ttgcagagcc cctggaaaac gtggaagatg cggtgaatga ggggctggaa 120
atagtgagag gacgtctgca aaacgctggc ctaaatgtga ctgtgaatgc ttctttcatg 180
tattcggatg gtctgattca taactccggc gactgccgga gcagcacctg tgaaggcctt 240
gacctactca ggaaaatttc aaatgcaaaa cggatgggct gtgtcctcat ggggccctca 300
tgtacatact ccaccttcca gatgtacctt gacacagaat tgagctaccc catgatctca 360
gctggaagtt ttggattgtc atgtgactat aaagaaacct taaccaggct gatgtctcca 420
gctagaaagt tgacatactt cttggttaac ttttggaaaa ccaatgatct acccttcaaa 480
acttattcct ggagcacttc gtatgtttac aagaatggta cggagtccga ggactgtttc 540
tggtacctta acgctctgga ggccagtgtt tcctatttct cccacgaact cagttttaag 600
ttggtgttaa gacaagataa ggagtttcag gatatcttaa tggaccacaa caggaaaagc 660
aatgtgattg ttatgtgtgg tgatccagag ttcctctaca agttgaaggg tgaccgagca 720
gtggctgaag acattgtcat tattctagtg gatcttttca atgaccagta ctttgaggac 780
aatgtcacag cccctgacta tatgaaaaat gtccttgttc tgacgcagtc tcctgggaat 840
tccctcctaa atagctcttt ctccaggaat ctatccccaa caaaacgaga ctttgctctt 900
gcctatttga atggaatcct gctctttgga catatgctaa agacatttct tgaaaatgga 960
gaaaatatta ccacccccaa atttgctcat gctttcagga atctcacttt tgaagggtat 1020
gacggtccag tgaccttgga tgactggggg gatgtggaca gtaccatggt gcttctgtat 1080
acgtctgtgg acaccaagaa atacaaggtt cttttgacct atgataccca cgtaaatcag 1140
accaaccctg tggatatgag ccccacattc acttggaaga actctaaact tcctaatgat 1200
attacagacc ggggccctca gggaggcgga ggttccgaga atttatactt ccagggcgga 1260
ggcggttccg gcggcggagg aagcgagccc cgcggaccga caatcaagcc ctgtcctcca 1320
tgcaaatgcc cagcacctaa cctcgagggt ggaccatccg tcttcatctt ccctccaaag 1380
atcaaggatg tactcatgat ctccctgagc cccatagtca catgtgtggt ggtggatgtg 1440
agcgaggatg acccagatgt ccagatcagc tggtttgtga acaacgtgga agtacacaca 1500
gctcagacac aaacccatag agaggattac aacagtactc tccgggtggt cagtgccctc 1560
cccatccagc accaggactg gatgagtggc aaggctttcg catgcgccgt caacaacaaa 1620
gacctcccag cgcccatcga gagaaccatc tcaaaaccca aagggtcagt aagagctcca 1680
caggtatatg tcttgcctcc accagaagaa gagatgacta agaaacaggt cactctgacc 1740
tgcatggtca cagacttcat gcctgaagac atttacgtgg agtggaccaa caacgggaaa 1800
acagagctaa actacaagaa cactgaacca gtcctggact ctgatggttc ttacttcatg 1860
tacagcaagc tgagagtgga aaagaagaac tgggtggaaa gaaatagcta ctcctgttca 1920
gtggtccacg agggtctgca caatcaccac acgactaaga gcttctcccg gactccgggt 1980
ggcgggccac ccgactacaa ggacgacgat gacaaa 2016
<210> 236
<211> 418
<212> БЕЛОК
<213> Macaca fascicularis
<400> 236
Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser Val
1 5 10 15
Leu Met Met Asp Asn Ser Ala Phe Ala Glu Pro Leu Glu Asn Val Glu
20 25 30
Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln Asn
35 40 45
Ala Gly Leu Asn Val Thr Val Asn Ala Ser Phe Met Tyr Ser Asp Gly
50 55 60
Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly Leu
65 70 75 80
Asp Leu Leu Arg Lys Ile Ser Asn Ala Lys Arg Met Gly Cys Val Leu
85 90 95
Met Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp Thr
100 105 110
Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser Cys
115 120 125
Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys Leu
130 135 140
Thr Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe Lys
145 150 155 160
Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu Ser
165 170 175
Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser Tyr
180 185 190
Phe Ser His Glu Leu Ser Phe Lys Leu Val Leu Arg Gln Asp Lys Glu
195 200 205
Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile Val
210 215 220
Met Cys Gly Asp Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg Ala
225 230 235 240
Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp Gln
245 250 255
Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val Leu
260 265 270
Val Leu Thr Gln Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe Ser
275 280 285
Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu Asn
290 295 300
Gly Ile Leu Leu Phe Gly His Met Leu Lys Thr Phe Leu Glu Asn Gly
305 310 315 320
Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu Thr
325 330 335
Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp Val
340 345 350
Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys Tyr
355 360 365
Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Gln Thr Asn Pro Val
370 375 380
Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn Asp
385 390 395 400
Ile Thr Asp Arg Gly Pro Gln Gly Gly Gly Gly Ser Glu Asn Leu Tyr
405 410 415
Phe Gln
<210> 237
<211> 1254
<212> ДНК
<213> Macaca fascicularis
<400> 237
tcacaggtga gtcagaactg ccacaatggc agctatgaaa tcagcgtcct gatgatggac 60
aactcagcct ttgcagagcc cctggaaaac gtggaagatg cggtgaatga ggggctggaa 120
atagtgagag gacgtctgca aaacgctggc ctaaatgtga ctgtgaatgc ttctttcatg 180
tattcggatg gtctgattca taactccggc gactgccgga gcagcacctg tgaaggcctt 240
gacctactca ggaaaatttc aaatgcaaaa cggatgggct gtgtcctcat ggggccctca 300
tgtacatact ccaccttcca gatgtacctt gacacagaat tgagctaccc catgatctca 360
gctggaagtt ttggattgtc atgtgactat aaagaaacct taaccaggct gatgtctcca 420
gctagaaagt tgacatactt cttggttaac ttttggaaaa ccaatgatct acccttcaaa 480
acttattcct ggagcacttc gtatgtttac aagaatggta cggagtccga ggactgtttc 540
tggtacctta acgctctgga ggccagtgtt tcctatttct cccacgaact cagttttaag 600
ttggtgttaa gacaagataa ggagtttcag gatatcttaa tggaccacaa caggaaaagc 660
aatgtgattg ttatgtgtgg tgatccagag ttcctctaca agttgaaggg tgaccgagca 720
gtggctgaag acattgtcat tattctagtg gatcttttca atgaccagta ctttgaggac 780
aatgtcacag cccctgacta tatgaaaaat gtccttgttc tgacgcagtc tcctgggaat 840
tccctcctaa atagctcttt ctccaggaat ctatccccaa caaaacgaga ctttgctctt 900
gcctatttga atggaatcct gctctttgga catatgctaa agacatttct tgaaaatgga 960
gaaaatatta ccacccccaa atttgctcat gctttcagga atctcacttt tgaagggtat 1020
gacggtccag tgaccttgga tgactggggg gatgtggaca gtaccatggt gcttctgtat 1080
acgtctgtgg acaccaagaa atacaaggtt cttttgacct atgataccca cgtaaatcag 1140
accaaccctg tggatatgag ccccacattc acttggaaga actctaaact tcctaatgat 1200
attacagacc ggggccctca gggaggcgga ggttccgaga atttatactt ccag 1254
<210> 238
<211> 673
<212> БЕЛОК
<213> Mus musculus
<400> 238
Val Phe Trp Ala Ser Gln Val Arg Gln Asn Cys Arg Asn Gly Ser Tyr
1 5 10 15
Glu Ile Ser Val Leu Met Met Asp Asn Ser Ala Tyr Lys Glu Pro Met
20 25 30
Gln Asn Leu Arg Glu Ala Val Glu Glu Gly Leu Asp Ile Val Arg Lys
35 40 45
Arg Leu Arg Glu Ala Asp Leu Asn Val Thr Val Asn Ala Thr Phe Ile
50 55 60
Tyr Ser Asp Gly Leu Ile His Lys Ser Gly Asp Cys Arg Ser Ser Thr
65 70 75 80
Cys Glu Gly Leu Asp Leu Leu Arg Glu Ile Thr Arg Asp His Lys Met
85 90 95
Gly Cys Ala Leu Met Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met
100 105 110
Tyr Leu Asp Thr Glu Leu Asn Tyr Pro Met Ile Ser Ala Gly Ser Tyr
115 120 125
Gly Leu Ser Cys Asp Tyr Lys Glu Thr Leu Thr Arg Ile Leu Pro Pro
130 135 140
Ala Arg Lys Leu Met Tyr Phe Leu Val Asp Phe Trp Lys Val Asn Asn
145 150 155 160
Ala Ser Phe Lys Pro Phe Ser Trp Asn Ser Ser Tyr Val Tyr Lys Asn
165 170 175
Gly Ser Glu Pro Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala
180 185 190
Gly Val Ser Tyr Phe Ser Glu Val Leu Asn Phe Lys Asp Val Leu Arg
195 200 205
Arg Ser Glu Gln Phe Gln Glu Ile Leu Thr Gly His Asn Arg Lys Ser
210 215 220
Asn Val Ile Val Met Cys Gly Thr Pro Glu Ser Phe Tyr Asp Val Lys
225 230 235 240
Gly Asp Leu Gln Val Ala Glu Asp Thr Val Val Ile Leu Val Asp Leu
245 250 255
Phe Ser Asn His Tyr Phe Glu Glu Asn Thr Thr Ala Pro Glu Tyr Met
260 265 270
Asp Asn Val Leu Val Leu Thr Leu Pro Ser Glu Gln Ser Thr Ser Asn
275 280 285
Thr Ser Val Ala Glu Arg Phe Ser Ser Gly Arg Ser Asp Phe Ser Leu
290 295 300
Ala Tyr Leu Glu Gly Thr Leu Leu Phe Gly His Met Leu Gln Thr Phe
305 310 315 320
Leu Glu Asn Gly Glu Asn Val Thr Gly Pro Lys Phe Ala Arg Ala Phe
325 330 335
Arg Asn Leu Thr Phe Gln Gly Phe Ala Gly Pro Val Thr Leu Asp Asp
340 345 350
Ser Gly Asp Ile Asp Asn Ile Met Ser Leu Leu Tyr Val Ser Leu Asp
355 360 365
Thr Arg Lys Tyr Lys Val Leu Met Lys Tyr Asp Thr His Lys Asn Lys
370 375 380
Thr Ile Pro Val Ala Glu Asn Pro Asn Phe Ile Trp Lys Asn His Lys
385 390 395 400
Leu Pro Asn Asp Val Pro Gly Leu Gly Pro Gln Ile Leu Met Gly Gly
405 410 415
Gly Gly Ser Glu Asn Leu Tyr Phe Gln Gly Gly Gly Gly Ser Gly Gly
420 425 430
Gly Gly Ser Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys
435 440 445
Lys Cys Pro Ala Pro Asn Leu Glu Gly Gly Pro Ser Val Phe Ile Phe
450 455 460
Pro Pro Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val
465 470 475 480
Thr Cys Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile
485 490 495
Ser Trp Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr
500 505 510
His Arg Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro
515 520 525
Ile Gln His Gln Asp Trp Met Ser Gly Lys Ala Phe Ala Cys Ala Val
530 535 540
Asn Asn Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro
545 550 555 560
Lys Gly Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu
565 570 575
Glu Glu Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp
580 585 590
Phe Met Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr
595 600 605
Glu Leu Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser
610 615 620
Tyr Phe Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu
625 630 635 640
Arg Asn Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His
645 650 655
His Thr Thr Lys Ser Phe Ser Arg Thr Pro Gly His His His His His
660 665 670
His
<210> 239
<211> 2019
<212> ДНК
<213> Mus musculus
<400> 239
gtgttctggg cctctcaggt gaggcagaac tgccgcaatg gcagctacga gatcagcgtc 60
ctgatgatgg acaactcagc ctacaaagaa cctatgcaaa acctgaggga ggctgtggag 120
gaaggactgg acatagtgcg aaagcgcctg cgtgaagccg acctaaatgt gactgtgaac 180
gcgactttca tctactccga cggtctgatt cataagtcag gtgactgccg gagcagcacc 240
tgtgaaggcc ttgacctact cagggagatt acaagagatc ataagatggg ctgcgccctc 300
atggggccct cgtgcacgta ttccaccttc cagatgtacc tcgacacaga gttgaactat 360
cccatgattt ccgctggaag ttatggattg tcctgtgact ataaggaaac cctaaccagg 420
attctgcctc cagccaggaa gctgatgtac ttcttggtcg atttctggaa agtcaacaat 480
gcatctttca aacccttttc ctggaactct tcgtatgttt acaagaatgg atcggaacct 540
gaagattgtt tctggtacct caatgctctg gaggctgggg tgtcctattt ttctgaggtg 600
ctcaacttca aggatgtact gagacgcagc gaacagttcc aggaaatctt aacaggccat 660
aacagaaaga gcaatgtgat tgttatgtgt ggcacgccag aaagcttcta tgatgtgaaa 720
ggtgacctcc aagtggctga agatactgtt gtcatcctgg tagatctgtt cagtaaccat 780
tactttgagg agaacaccac agctcctgag tatatggaca atgtcctcgt cctgacgctg 840
ccgtctgaac agtccacctc aaacacctct gtcgccgaga ggttttcatc ggggagaagt 900
gacttttctc tcgcttactt ggagggaacc ttgctatttg gacacatgct gcagacgttt 960
cttgaaaatg gagaaaatgt cacgggtccc aagtttgctc gtgcattcag gaatctcact 1020
tttcaaggct ttgcaggacc tgtgactctg gatgacagtg gggacattga caacattatg 1080
tcccttctgt atgtgtctct ggataccagg aaatacaagg ttcttatgaa gtatgacacc 1140
cacaaaaaca aaactattcc ggtggctgag aaccccaact tcatctggaa gaaccacaag 1200
ctccccaatg acgttcctgg gctgggccct caaatcctga tgggaggcgg aggttccgag 1260
aatttatact tccagggcgg aggcggttcc ggcggcggag gaagcgagcc ccgcggaccg 1320
acaatcaagc cctgtcctcc atgcaaatgc ccagcaccta acctcgaggg tggaccatcc 1380
gtcttcatct tccctccaaa gatcaaggat gtactcatga tctccctgag ccccatagtc 1440
acatgtgtgg tggtggatgt gagcgaggat gacccagatg tccagatcag ctggtttgtg 1500
aacaacgtgg aagtacacac agctcagaca caaacccata gagaggatta caacagtact 1560
ctccgggtgg tcagtgccct ccccatccag caccaggact ggatgagtgg caaggctttc 1620
gcatgcgccg tcaacaacaa agacctccca gcgcccatcg agagaaccat ctcaaaaccc 1680
aaagggtcag taagagctcc acaggtatat gtcttgcctc caccagaaga agagatgact 1740
aagaaacagg tcactctgac ctgcatggtc acagacttca tgcctgaaga catttacgtg 1800
gagtggacca acaacgggaa aacagagcta aactacaaga acactgaacc agtcctggac 1860
tctgatggtt cttacttcat gtacagcaag ctgagagtgg aaaagaagaa ctgggtggaa 1920
agaaatagct actcctgttc agtggtccac gagggtctgc acaatcacca cacgactaag 1980
agcttctccc ggactccggg tcaccatcac catcaccat 2019
<210> 240
<211> 425
<212> БЕЛОК
<213> Mus musculus
<400> 240
Val Phe Trp Ala Ser Gln Val Arg Gln Asn Cys Arg Asn Gly Ser Tyr
1 5 10 15
Glu Ile Ser Val Leu Met Met Asp Asn Ser Ala Tyr Lys Glu Pro Met
20 25 30
Gln Asn Leu Arg Glu Ala Val Glu Glu Gly Leu Asp Ile Val Arg Lys
35 40 45
Arg Leu Arg Glu Ala Asp Leu Asn Val Thr Val Asn Ala Thr Phe Ile
50 55 60
Tyr Ser Asp Gly Leu Ile His Lys Ser Gly Asp Cys Arg Ser Ser Thr
65 70 75 80
Cys Glu Gly Leu Asp Leu Leu Arg Glu Ile Thr Arg Asp His Lys Met
85 90 95
Gly Cys Ala Leu Met Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met
100 105 110
Tyr Leu Asp Thr Glu Leu Asn Tyr Pro Met Ile Ser Ala Gly Ser Tyr
115 120 125
Gly Leu Ser Cys Asp Tyr Lys Glu Thr Leu Thr Arg Ile Leu Pro Pro
130 135 140
Ala Arg Lys Leu Met Tyr Phe Leu Val Asp Phe Trp Lys Val Asn Asn
145 150 155 160
Ala Ser Phe Lys Pro Phe Ser Trp Asn Ser Ser Tyr Val Tyr Lys Asn
165 170 175
Gly Ser Glu Pro Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala
180 185 190
Gly Val Ser Tyr Phe Ser Glu Val Leu Asn Phe Lys Asp Val Leu Arg
195 200 205
Arg Ser Glu Gln Phe Gln Glu Ile Leu Thr Gly His Asn Arg Lys Ser
210 215 220
Asn Val Ile Val Met Cys Gly Thr Pro Glu Ser Phe Tyr Asp Val Lys
225 230 235 240
Gly Asp Leu Gln Val Ala Glu Asp Thr Val Val Ile Leu Val Asp Leu
245 250 255
Phe Ser Asn His Tyr Phe Glu Glu Asn Thr Thr Ala Pro Glu Tyr Met
260 265 270
Asp Asn Val Leu Val Leu Thr Leu Pro Ser Glu Gln Ser Thr Ser Asn
275 280 285
Thr Ser Val Ala Glu Arg Phe Ser Ser Gly Arg Ser Asp Phe Ser Leu
290 295 300
Ala Tyr Leu Glu Gly Thr Leu Leu Phe Gly His Met Leu Gln Thr Phe
305 310 315 320
Leu Glu Asn Gly Glu Asn Val Thr Gly Pro Lys Phe Ala Arg Ala Phe
325 330 335
Arg Asn Leu Thr Phe Gln Gly Phe Ala Gly Pro Val Thr Leu Asp Asp
340 345 350
Ser Gly Asp Ile Asp Asn Ile Met Ser Leu Leu Tyr Val Ser Leu Asp
355 360 365
Thr Arg Lys Tyr Lys Val Leu Met Lys Tyr Asp Thr His Lys Asn Lys
370 375 380
Thr Ile Pro Val Ala Glu Asn Pro Asn Phe Ile Trp Lys Asn His Lys
385 390 395 400
Leu Pro Asn Asp Val Pro Gly Leu Gly Pro Gln Ile Leu Met Gly Gly
405 410 415
Gly Gly Ser Glu Asn Leu Tyr Phe Gln
420 425
<210> 241
<211> 1275
<212> ДНК
<213> Mus musculus
<400> 241
gtgttctggg cctctcaggt gaggcagaac tgccgcaatg gcagctacga gatcagcgtc 60
ctgatgatgg acaactcagc ctacaaagaa cctatgcaaa acctgaggga ggctgtggag 120
gaaggactgg acatagtgcg aaagcgcctg cgtgaagccg acctaaatgt gactgtgaac 180
gcgactttca tctactccga cggtctgatt cataagtcag gtgactgccg gagcagcacc 240
tgtgaaggcc ttgacctact cagggagatt acaagagatc ataagatggg ctgcgccctc 300
atggggccct cgtgcacgta ttccaccttc cagatgtacc tcgacacaga gttgaactat 360
cccatgattt ccgctggaag ttatggattg tcctgtgact ataaggaaac cctaaccagg 420
attctgcctc cagccaggaa gctgatgtac ttcttggtcg atttctggaa agtcaacaat 480
gcatctttca aacccttttc ctggaactct tcgtatgttt acaagaatgg atcggaacct 540
gaagattgtt tctggtacct caatgctctg gaggctgggg tgtcctattt ttctgaggtg 600
ctcaacttca aggatgtact gagacgcagc gaacagttcc aggaaatctt aacaggccat 660
aacagaaaga gcaatgtgat tgttatgtgt ggcacgccag aaagcttcta tgatgtgaaa 720
ggtgacctcc aagtggctga agatactgtt gtcatcctgg tagatctgtt cagtaaccat 780
tactttgagg agaacaccac agctcctgag tatatggaca atgtcctcgt cctgacgctg 840
ccgtctgaac agtccacctc aaacacctct gtcgccgaga ggttttcatc ggggagaagt 900
gacttttctc tcgcttactt ggagggaacc ttgctatttg gacacatgct gcagacgttt 960
cttgaaaatg gagaaaatgt cacgggtccc aagtttgctc gtgcattcag gaatctcact 1020
tttcaaggct ttgcaggacc tgtgactctg gatgacagtg gggacattga caacattatg 1080
tcccttctgt atgtgtctct ggataccagg aaatacaagg ttcttatgaa gtatgacacc 1140
cacaaaaaca aaactattcc ggtggctgag aaccccaact tcatctggaa gaaccacaag 1200
ctccccaatg acgttcctgg gctgggccct caaatcctga tgggaggcgg aggttccgag 1260
aatttatact tccag 1275
<210> 242
<211> 214
<212> БЕЛОК
<213> Homo sapiens
<400> 242
Met Gln Ser Gly Thr His Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Val Gly Val Trp Gly Gln Asp Gly Asn Glu Glu Met Gly Gly Ile Thr
20 25 30
Gln Thr Pro Tyr Lys Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr
35 40 45
Cys Pro Gln Tyr Pro Gly Ser Glu Ile Leu Trp Gln His Asn Asp Lys
50 55 60
Asn Ile Gly Gly Asp Glu Asp Asp Lys Asn Ile Gly Ser Asp Glu Asp
65 70 75 80
His Leu Ser Leu Lys Glu Phe Ser Glu Leu Glu Gln Ser Gly Tyr Tyr
85 90 95
Val Cys Tyr Pro Arg Gly Ser Lys Pro Glu Asp Ala Asn Phe Tyr Leu
100 105 110
Tyr Leu Arg Ala Arg Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Pro Ile Glu Glu Leu Glu Asp Arg Val Phe Val Asn
130 135 140
Cys Asn Thr Ser Ile Thr Trp Val Glu Gly Thr Val Gly Thr Leu Leu
145 150 155 160
Ser Asp Ile Thr Arg Leu Asp Leu Gly Lys Arg Ile Leu Asp Pro Arg
165 170 175
Gly Ile Tyr Arg Cys Asn Gly Thr Asp Ile Tyr Lys Asp Lys Glu Ser
180 185 190
Thr Val Gln Val His Tyr Arg Met Cys Gln Ser Cys Val Glu Leu Asp
195 200 205
His His His His His His
210
<210> 243
<211> 642
<212> ДНК
<213> Homo sapiens
<400> 243
atgcaatccg gtacgcactg gagagtcttg ggtctgtgcc ttttgtctgt tggcgtatgg 60
gggcaagacg ggaacgaaga aatgggaggc attacacaaa caccatacaa ggtatcaatt 120
agcggcacga cggttatact gacatgtcca caatatccag gcagcgaaat tctgtggcag 180
cacaatgaca agaatattgg gggagatgaa gacgacaaaa atatcggtag cgacgaggac 240
catctgtctc tgaaggaatt ttcagaactt gaacaatctg gctattatgt gtgctacccg 300
cgaggcagca aaccggaaga tgcaaacttt tacctttatc tgagagcaag gggcggcgga 360
ggctctgggg gcggaggcag cggcggagga ggatcaccaa tcgaggaatt ggaagatagg 420
gtattcgtaa attgtaacac cagcattaca tgggtggaag ggaccgttgg aactctcctg 480
agcgatatca cacgactgga tcttggtaaa cgaatcctgg acccacgcgg aatctataga 540
tgtaacggaa ctgatattta caaagacaaa gaatctactg tgcaagttca ctaccgaatg 600
tgtcaatcat gcgttgaact cgatcaccac caccatcatc ac 642
<210> 244
<211> 205
<212> БЕЛОК
<213> Macaca fascicularis
<400> 244
Met Gln Ser Gly Thr His Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Val Gly Val Trp Gly Gln Asp Gly Asn Glu Glu Met Gly Ser Ile Thr
20 25 30
Gln Thr Pro Tyr Gln Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr
35 40 45
Cys Ser Gln His Leu Gly Ser Glu Ala Gln Trp Gln His Asn Gly Lys
50 55 60
Asn Lys Gly Asp Ser Gly Asp Gln Leu Phe Leu Pro Glu Phe Ser Glu
65 70 75 80
Met Glu Gln Ser Gly Tyr Tyr Val Cys Tyr Pro Arg Gly Ser Asn Pro
85 90 95
Glu Asp Ala Ser His His Leu Tyr Leu Lys Ala Arg Gly Gly Gly Gly
100 105 110
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Pro Val Glu Glu Leu
115 120 125
Glu Asp Arg Val Phe Val Lys Cys Asn Thr Ser Val Thr Trp Val Glu
130 135 140
Gly Thr Val Gly Thr Leu Leu Thr Asn Asn Thr Arg Leu Asp Leu Gly
145 150 155 160
Lys Arg Ile Leu Asp Pro Arg Gly Ile Tyr Arg Cys Asn Gly Thr Asp
165 170 175
Ile Tyr Lys Asp Lys Glu Ser Ala Val Gln Val His Tyr Arg Met Cys
180 185 190
Gln Asn Cys Val Glu Leu Asp His His His His His His
195 200 205
<210> 245
<211> 615
<212> ДНК
<213> Macaca fascicularis
<400> 245
atgcaaagcg gaactcattg gcgcgtcctg ggactctgtc tgctctccgt gggagtctgg 60
ggacaagatg gaaacgaaga gatgggaagc attacacaaa caccatacca agtctccatt 120
agcggcacta ccgtcattct gacatgttcc caacatctgg gcagcgaagc tcaatggcag 180
cacaatggaa agaataaggg cgatagcgga gaccaactgt ttctgccaga atttagcgaa 240
atggagcaat ccggctatta cgtgtgctac ccacgcggca gcaaccctga agatgctagc 300
catcacctct atctgaaggc tcgcggaggc ggaggcagcg gcggcggagg atccggcgga 360
ggcggaagcc cagtcgagga actggaagat cgcgtcttcg tgaagtgtaa caccagcgtc 420
acatgggtgg aaggcaccgt cggaactctc ctgactaaca acacacgcct ggatctcgga 480
aaacgcatcc tggacccacg cggaatctat agatgtaacg gaactgatat ttacaaagac 540
aaagaatccg ctgtgcaagt ccactaccgc atgtgtcaaa actgtgtcga actggatcat 600
caccatcacc atcac 615
<210> 246
<211> 1386
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 246
gacatccagc tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgca gagccagtga aagtgttgat tattatggca gtagtttatt gcagtggtat 120
cagcagaaac cagggaaagc ccctaagctc ctgatctatg ctgcatccaa actagcttct 180
ggggtcccat caaggttcag tggcagtgga tctgggacag atttcactct caccatcagc 240
agtctgcaac ctgaagattt tgcaacttac tactgtcagc aaactcggaa agcgtatacg 300
tttggccagg ggaccaagct ggagatcaaa ggtggaggta gcgggggcgg cggggaagtt 360
caactcgttg agtctggcgg gggattggtt caacccggtg gaagccttag attgtcatgt 420
gccgcctccg gctttacatt tagcgactat tacatgacct gggtgagaca agctccaggc 480
aaaggacttg aatgggtggc ctttatcaga aatcaggccc gcggctacac aagcgaccat 540
aatccctccg tgaaaggaag atttaccatc agccgggaca atgctaaaaa ttcactttac 600
cttcaaatga actctcttag agccgaggac accgccgtat actactgcgc aagagataga 660
ccaagttatt acgtcctgga ttactggggc cagggaacaa ccgtcaccgt gtcttctgga 720
tgcccaccgt gcccagcacc tgaagccgct ggggcaccgt cagtcttcct cttcccccca 780
aaacccaagg acaccctcat gatctcccgg acccctgagg tcacatgcgt ggtggtggac 840
gtgagccacg aagaccctga ggtcaagttc aactggtacg tggacggcgt ggaggtgcat 900
aatgccaaga caaagccgcg ggaggagcag tacaacagca cgtaccgtgt ggtcagcgtc 960
ctcaccgtcc tgcaccagga ctggctgaat ggcaaggagt acaagtgcaa ggtctccaac 1020
aaagccctcc cagcccccat cgagaaaacc atctccaaag ccaaagggca gccccgagaa 1080
ccacaggtgt gcaccctgcc cccatcccgg gaggagatga ccaagaacca ggtcagcctg 1140
tggtgcctgg tcaaaggctt ctatcccagc gacatcgccg tggagtggga gagcaatggg 1200
cagccggaga acaactacaa gaccacgcct cccgtgctgg actccgacgg ctccttcttc 1260
ctctatagca agctcaccgt ggacaagagc aggtggcagc aggggaacgt cttctcatgc 1320
tccgtgatgc atgaggctct gcacaaccac tacacgcaga agagcctctc cctgtccccc 1380
ggaaag 1386
<210> 247
<211> 1398
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 247
gacatccaga tgacccagtc cccctcttct ctgtctgcct ctgtgggcga cagagtgacc 60
atcacctgca caagctcaca gtcactgttt aatgtccgca gccagaaaaa ctatcttgcg 120
tggtatcagc agaagcctgg caaggctccc aagctgctga tctactgggc cagtacacga 180
gaatccggcg tgccttccag attctccggc tctggctctg gcaccgattt caccctgacc 240
atctcctccc tccagcctga ggatttcgcc acctactact gcaaacagtc ttacgacctt 300
ttcacttttg gcggcggaac aaaggtggag atcaagggcg gaggtggatc tggcggcgga 360
ggcgaggtgc agctggtgga gtctggggga ggcttggtcc agcctggggg gtccctgaga 420
ctctcctgtg cagcctctgg cttcaccttc agcagctact ggatgcactg ggtccgccag 480
gctccaggga aggggctgga gtggattgga gagattaaac ctagcaacga acttactaac 540
gtccatgaaa agttcaagga ccgattcacc atctccgtgg acaaggccaa gaactcagcc 600
tatctgcaaa tgaacagcct gagagccgag gacacggctg tgtattactg tacaagaacg 660
attacgacga cggagggata ctggttcttc gatgtctggg gccaagggac actggtcacc 720
gtctcttcag gatgtccacc gtgcccagca cctgaagccg ctggggcacc gtcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1080
cagccccgag aaccacaggt gtacaccctg cccccatgcc gggaggagat gaccaagaac 1140
caggtcagcc tgtcctgcgc ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1260
ggctccttct tcctcgttag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1380
tccctgtccc ccggaaag 1398
<210> 248
<211> 461
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 248
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Arg Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 249
<211> 464
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 249
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Thr Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Met His Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn Gly
165 170 175
Leu Thr Asn Tyr Ile Glu Lys Phe Lys Asn Arg Phe Thr Ile Ser Val
180 185 190
Asp Lys Ala Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr Glu
210 215 220
Gly Tyr Trp Phe Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val
225 230 235 240
Ser Ser Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
275 280 285
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Ser
370 375 380
Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 250
<211> 925
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 250
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Arg Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Asp Ile Gln
450 455 460
Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val
465 470 475 480
Thr Ile Thr Cys Thr Ser Ser Gln Ser Leu Phe Asn Val Arg Ser Arg
485 490 495
Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
500 505 510
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Ser Arg
515 520 525
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
530 535 540
Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln Ser Tyr Asp
545 550 555 560
Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
580 585 590
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr
595 600 605
Thr Phe Thr Ser Tyr Trp Met His Trp Val Arg Gln Ala Pro Gly Lys
610 615 620
Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn
625 630 635 640
Tyr Ile Glu Lys Phe Lys Asn Arg Phe Thr Ile Ser Val Asp Lys Ala
645 650 655
Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
660 665 670
Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp
675 680 685
Phe Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly
690 695 700
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
705 710 715 720
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
725 730 735
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
740 745 750
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
755 760 765
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
770 775 780
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
785 790 795 800
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
805 810 815
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
820 825 830
Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val
835 840 845
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
850 855 860
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
865 870 875 880
Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
885 890 895
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
900 905 910
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
915 920 925
<210> 251
<211> 1073
<212> БЕЛОК
<213> Homo sapiens
<400> 251
Met Lys Thr Leu Leu Leu Asp Leu Ala Leu Trp Ser Leu Leu Phe Gln
1 5 10 15
Pro Gly Trp Leu Ser Phe Ser Ser Gln Val Ser Gln Asn Cys His Asn
20 25 30
Gly Ser Tyr Glu Ile Ser Val Leu Met Met Gly Asn Ser Ala Phe Ala
35 40 45
Glu Pro Leu Lys Asn Leu Glu Asp Ala Val Asn Glu Gly Leu Glu Ile
50 55 60
Val Arg Gly Arg Leu Gln Asn Ala Gly Leu Asn Val Thr Val Asn Ala
65 70 75 80
Thr Phe Met Tyr Ser Asp Gly Leu Ile His Asn Ser Gly Asp Cys Arg
85 90 95
Ser Ser Thr Cys Glu Gly Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala
100 105 110
Gln Arg Met Gly Cys Val Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr
115 120 125
Phe Gln Met Tyr Leu Asp Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala
130 135 140
Gly Ser Phe Gly Leu Ser Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu
145 150 155 160
Met Ser Pro Ala Arg Lys Leu Met Tyr Phe Leu Val Asn Phe Trp Lys
165 170 175
Thr Asn Asp Leu Pro Phe Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val
180 185 190
Tyr Lys Asn Gly Thr Glu Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala
195 200 205
Leu Glu Ala Ser Val Ser Tyr Phe Ser His Glu Leu Gly Phe Lys Val
210 215 220
Val Leu Arg Gln Asp Lys Glu Phe Gln Asp Ile Leu Met Asp His Asn
225 230 235 240
Arg Lys Ser Asn Val Ile Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr
245 250 255
Lys Leu Lys Gly Asp Arg Ala Val Ala Glu Asp Ile Val Ile Ile Leu
260 265 270
Val Asp Leu Phe Asn Asp Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro
275 280 285
Asp Tyr Met Lys Asn Val Leu Val Leu Thr Leu Ser Pro Gly Asn Ser
290 295 300
Leu Leu Asn Ser Ser Phe Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp
305 310 315 320
Phe Ala Leu Ala Tyr Leu Asn Gly Ile Leu Leu Phe Gly His Met Leu
325 330 335
Lys Ile Phe Leu Glu Asn Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala
340 345 350
His Ala Phe Arg Asn Leu Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr
355 360 365
Leu Asp Asp Trp Gly Asp Val Asp Ser Thr Met Val Leu Leu Tyr Thr
370 375 380
Ser Val Asp Thr Lys Lys Tyr Lys Val Leu Leu Thr Tyr Asp Thr His
385 390 395 400
Val Asn Lys Thr Tyr Pro Val Asp Met Ser Pro Thr Phe Thr Trp Lys
405 410 415
Asn Ser Lys Leu Pro Asn Asp Ile Thr Gly Arg Gly Pro Gln Ile Leu
420 425 430
Met Ile Ala Val Phe Thr Leu Thr Gly Ala Val Val Leu Leu Leu Leu
435 440 445
Val Ala Leu Leu Met Leu Arg Lys Tyr Arg Lys Asp Tyr Glu Leu Arg
450 455 460
Gln Lys Lys Trp Ser His Ile Pro Pro Glu Asn Ile Phe Pro Leu Glu
465 470 475 480
Thr Asn Glu Thr Asn His Val Ser Leu Lys Ile Asp Asp Asp Lys Arg
485 490 495
Arg Asp Thr Ile Gln Arg Leu Arg Gln Cys Lys Tyr Asp Lys Lys Arg
500 505 510
Val Ile Leu Lys Asp Leu Lys His Asn Asp Gly Asn Phe Thr Glu Lys
515 520 525
Gln Lys Ile Glu Leu Asn Lys Leu Leu Gln Ile Asp Tyr Tyr Asn Leu
530 535 540
Thr Lys Phe Tyr Gly Thr Val Lys Leu Asp Thr Met Ile Phe Gly Val
545 550 555 560
Ile Glu Tyr Cys Glu Arg Gly Ser Leu Arg Glu Val Leu Asn Asp Thr
565 570 575
Ile Ser Tyr Pro Asp Gly Thr Phe Met Asp Trp Glu Phe Lys Ile Ser
580 585 590
Val Leu Tyr Asp Ile Ala Lys Gly Met Ser Tyr Leu His Ser Ser Lys
595 600 605
Thr Glu Val His Gly Arg Leu Lys Ser Thr Asn Cys Val Val Asp Ser
610 615 620
Arg Met Val Val Lys Ile Thr Asp Phe Gly Cys Asn Ser Ile Leu Pro
625 630 635 640
Pro Lys Lys Asp Leu Trp Thr Ala Pro Glu His Leu Arg Gln Ala Asn
645 650 655
Ile Ser Gln Lys Gly Asp Val Tyr Ser Tyr Gly Ile Ile Ala Gln Glu
660 665 670
Ile Ile Leu Arg Lys Glu Thr Phe Tyr Thr Leu Ser Cys Arg Asp Arg
675 680 685
Asn Glu Lys Ile Phe Arg Val Glu Asn Ser Asn Gly Met Lys Pro Phe
690 695 700
Arg Pro Asp Leu Phe Leu Glu Thr Ala Glu Glu Lys Glu Leu Glu Val
705 710 715 720
Tyr Leu Leu Val Lys Asn Cys Trp Glu Glu Asp Pro Glu Lys Arg Pro
725 730 735
Asp Phe Lys Lys Ile Glu Thr Thr Leu Ala Lys Ile Phe Gly Leu Phe
740 745 750
His Asp Gln Lys Asn Glu Ser Tyr Met Asp Thr Leu Ile Arg Arg Leu
755 760 765
Gln Leu Tyr Ser Arg Asn Leu Glu His Leu Val Glu Glu Arg Thr Gln
770 775 780
Leu Tyr Lys Ala Glu Arg Asp Arg Ala Asp Arg Leu Asn Phe Met Leu
785 790 795 800
Leu Pro Arg Leu Val Val Lys Ser Leu Lys Glu Lys Gly Phe Val Glu
805 810 815
Pro Glu Leu Tyr Glu Glu Val Thr Ile Tyr Phe Ser Asp Ile Val Gly
820 825 830
Phe Thr Thr Ile Cys Lys Tyr Ser Thr Pro Met Glu Val Val Asp Met
835 840 845
Leu Asn Asp Ile Tyr Lys Ser Phe Asp His Ile Val Asp His His Asp
850 855 860
Val Tyr Lys Val Glu Thr Ile Gly Asp Ala Tyr Met Val Ala Ser Gly
865 870 875 880
Leu Pro Lys Arg Asn Gly Asn Arg His Ala Ile Asp Ile Ala Lys Met
885 890 895
Ala Leu Glu Ile Leu Ser Phe Met Gly Thr Phe Glu Leu Glu His Leu
900 905 910
Pro Gly Leu Pro Ile Trp Ile Arg Ile Gly Val His Ser Gly Pro Cys
915 920 925
Ala Ala Gly Val Val Gly Ile Lys Met Pro Arg Tyr Cys Leu Phe Gly
930 935 940
Asp Thr Val Asn Thr Ala Ser Arg Met Glu Ser Thr Gly Leu Pro Leu
945 950 955 960
Arg Ile His Val Ser Gly Ser Thr Ile Ala Ile Leu Lys Arg Thr Glu
965 970 975
Cys Gln Phe Leu Tyr Glu Val Arg Gly Glu Thr Tyr Leu Lys Gly Arg
980 985 990
Gly Asn Glu Thr Thr Tyr Trp Leu Thr Gly Met Lys Asp Gln Lys Phe
995 1000 1005
Asn Leu Pro Thr Pro Pro Thr Val Glu Asn Gln Gln Arg Leu Gln
1010 1015 1020
Ala Glu Phe Ser Asp Met Ile Ala Asn Ser Leu Gln Lys Arg Gln
1025 1030 1035
Ala Ala Gly Ile Arg Ser Gln Lys Pro Arg Arg Val Ala Ser Tyr
1040 1045 1050
Lys Lys Gly Thr Leu Glu Tyr Leu Gln Leu Asn Thr Thr Asp Lys
1055 1060 1065
Glu Ser Thr Tyr Phe
1070
<210> 252
<211> 3850
<212> ДНК
<213> Homo sapians
<400> 252
gaccagagag aagcgtgggg aagagtgggc tgagggactc cactagaggc tgtccatctg 60
gattccctgc ctccctagga gcccaacaga gcaaagcaag tgggcacaag gagtatggtt 120
ctaacgtgat tggggtcatg aagacgttgc tgttggactt ggctttgtgg tcactgctct 180
tccagcccgg gtggctgtcc tttagttccc aggtgagtca gaactgccac aatggcagct 240
atgaaatcag cgtcctgatg atgggcaact cagcctttgc agagcccctg aaaaacttgg 300
aagatgcggt gaatgagggg ctggaaatag tgagaggacg tctgcaaaat gctggcctaa 360
atgtgactgt gaacgctact ttcatgtatt cggatggtct gattcataac tcaggcgact 420
gccggagtag cacctgtgaa ggcctcgacc tactcaggaa aatttcaaat gcacaacgga 480
tgggctgtgt cctcataggg ccctcatgta catactccac cttccagatg taccttgaca 540
cagaattgag ctaccccatg atctcagctg gaagttttgg attgtcatgt gactataaag 600
aaaccttaac caggctgatg tctccagcta gaaagttgat gtacttcttg gttaactttt 660
ggaaaaccaa cgatctgccc ttcaaaactt attcctggag cacttcgtat gtttacaaga 720
atggtacaga aactgaggac tgtttctggt accttaatgc tctggaggct agcgtttcct 780
atttctccca cgaactcggc tttaaggtgg tgttaagaca agataaggag tttcaggata 840
tcttaatgga ccacaacagg aaaagcaatg tgattattat gtgtggtggt ccagagttcc 900
tctacaagct gaagggtgac cgagcagtgg ctgaagacat tgtcattatt ctagtggatc 960
ttttcaatga ccagtacttt gaggacaatg tcacagcccc tgactatatg aaaaatgtcc 1020
ttgttctgac gctgtctcct gggaattccc ttctaaatag ctctttctcc aggaatctat 1080
caccaacaaa acgagacttt gctcttgcct atttgaatgg aatcctgctc tttggacata 1140
tgctgaagat atttcttgaa aatggagaaa atattaccac ccccaaattt gctcatgctt 1200
tcaggaatct cacttttgaa gggtatgacg gtccagtgac cttggatgac tggggggatg 1260
ttgacagtac catggtgctt ctgtatacct ctgtggacac caagaaatac aaggttcttt 1320
tgacctatga tacccacgta aataagacct atcctgtgga tatgagcccc acattcactt 1380
ggaagaactc taaacttcct aatgatatta caggccgggg ccctcagatc ctgatgattg 1440
cagtcttcac cctcactgga gctgtggtgc tgctcctgct cgtcgctctc ctgatgctca 1500
gaaaatatag aaaagattat gaacttcgtc agaaaaaatg gtcccacatt cctcctgaaa 1560
atatctttcc tctggagacc aatgagacca atcatgttag cctcaagatc gatgatgaca 1620
aaagacgaga tacaatccag agactacgac agtgcaaata cgacaaaaag cgagtgattc 1680
tcaaagatct caagcacaat gatggtaatt tcactgaaaa acagaagata gaattgaaca 1740
agttgcttca gattgactat tacaacctga ccaagttcta cggcacagtg aaacttgata 1800
ccatgatctt cggggtgata gaatactgtg agagaggatc cctccgggaa gttttaaatg 1860
acacaatttc ctaccctgat ggcacattca tggattggga gtttaagatc tctgtcttgt 1920
atgacattgc taagggaatg tcatatctgc actccagtaa gacagaagtc catggtcgtc 1980
tgaaatctac caactgcgta gtggacagta gaatggtggt gaagatcact gattttggct 2040
gcaattccat tttacctcca aaaaaggacc tgtggacagc tccagagcac ctccgccaag 2100
ccaacatctc tcagaaagga gatgtgtaca gctatgggat catcgcacag gagatcatcc 2160
tgcggaaaga aaccttctac actttgagct gtcgggaccg gaatgagaag attttcagag 2220
tggaaaattc caatggaatg aaacccttcc gcccagattt attcttggaa acagcagagg 2280
aaaaagagct agaagtgtac ctacttgtaa aaaactgttg ggaggaagat ccagaaaaga 2340
gaccagattt caaaaaaatt gagactacac ttgccaagat atttggactt tttcatgacc 2400
aaaaaaatga aagctatatg gataccttga tccgacgtct acagctatat tctcgaaacc 2460
tggaacatct ggtagaggaa aggacacagc tgtacaaggc agagagggac agggctgaca 2520
gacttaactt tatgttgctt ccaaggctag tggtaaagtc tctgaaggag aaaggctttg 2580
tggagccgga actatatgag gaagttacaa tctacttcag tgacattgta ggtttcacta 2640
ctatctgcaa atacagcacc cccatggaag tggtggacat gcttaatgac atctataaga 2700
gttttgacca cattgttgat catcatgatg tctacaaggt ggaaaccatc ggtgatgcgt 2760
acatggtggc tagtggtttg cctaagagaa atggcaatcg gcatgcaata gacattgcca 2820
agatggcctt ggaaatcctc agcttcatgg ggacctttga gctggagcat cttcctggcc 2880
tcccaatatg gattcgcatt ggagttcact ctggtccctg tgctgctgga gttgtgggaa 2940
tcaagatgcc tcgttattgt ctatttggag atacggtcaa cacagcctct aggatggaat 3000
ccactggcct ccctttgaga attcacgtga gtggctccac catagccatc ctgaagagaa 3060
ctgagtgcca gttcctttat gaagtgagag gagaaacata cttaaaggga agaggaaatg 3120
agactaccta ctggctgact gggatgaagg accagaaatt caacctgcca acccctccta 3180
ctgtggagaa tcaacagcgt ttgcaagcag aattttcaga catgattgcc aactctttac 3240
agaaaagaca ggcagcaggg ataagaagcc aaaaacccag acgggtagcc agctataaaa 3300
aaggcactct ggaatacttg cagctgaata ccacagacaa ggagagcacc tatttttaaa 3360
cctaaatgag gtataaggac tcacacaaat taaaatacag ctgcactgag gcagcgacct 3420
caagtgtcct gaaagcttac attttcctga gacctcaatg aagcagaaat gtacttaggc 3480
ttggctgccc tgtctggaac atggactttc ttgcatgaat cagatgtgtg ttctcagtga 3540
aataactacc ttccactctg gaaccttatt ccagcagttg ttccagggag cttctacctg 3600
gaaaagaaaa gaaatgaata gactatctag aacttgagaa gattttattc ttatttcatt 3660
tattttttgt ttgtttattt ttatcgtttt tgtttactgg ctttccttct gtattcataa 3720
gattttttaa attgtcataa ttatatttta aatacccatc ttcattaaag tatatttaac 3780
tcataatttt tgcagaaaat atgctatata ttaggcaaga ataaaagcta aaggtttccc 3840
aaaaaaaaaa 3850
<210> 253
<211> 1050
<212> БЕЛОК
<213> Macaca fascicularis
<400> 253
Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser Val
1 5 10 15
Leu Met Met Asp Asn Ser Ala Phe Ala Glu Pro Leu Glu Asn Val Glu
20 25 30
Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln Asn
35 40 45
Ala Gly Leu Asn Val Thr Val Asn Ala Ser Phe Met Tyr Ser Asp Gly
50 55 60
Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly Leu
65 70 75 80
Asp Leu Leu Arg Lys Ile Ser Asn Ala Lys Arg Met Gly Cys Val Leu
85 90 95
Met Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp Thr
100 105 110
Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser Cys
115 120 125
Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys Leu
130 135 140
Thr Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe Lys
145 150 155 160
Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu Ser
165 170 175
Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser Tyr
180 185 190
Phe Ser His Glu Leu Ser Phe Lys Leu Val Leu Arg Gln Asp Lys Glu
195 200 205
Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile Val
210 215 220
Met Cys Gly Asp Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg Ala
225 230 235 240
Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp Gln
245 250 255
Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val Leu
260 265 270
Val Leu Thr Gln Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe Ser
275 280 285
Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu Asn
290 295 300
Gly Ile Leu Leu Phe Gly His Met Leu Lys Thr Phe Leu Glu Asn Gly
305 310 315 320
Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu Thr
325 330 335
Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp Val
340 345 350
Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys Tyr
355 360 365
Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Gln Thr Asn Pro Val
370 375 380
Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn Asp
385 390 395 400
Ile Thr Asp Arg Gly Pro Gln Ile Leu Met Ile Ala Val Phe Thr Leu
405 410 415
Thr Gly Ala Val Val Leu Leu Leu Leu Val Ala Leu Leu Met Leu Arg
420 425 430
Lys Tyr Lys Lys Asp Tyr Glu Leu Arg Gln Lys Lys Trp Ser His Ile
435 440 445
Pro Pro Glu Asn Ile Phe Pro Leu Glu Thr Asn Glu Thr Asn His Val
450 455 460
Ser Leu Lys Ile Asp Asp Asp Lys Arg Arg Asp Thr Ile Gln Arg Leu
465 470 475 480
Arg Gln Cys Lys Tyr Asp Lys Lys Arg Val Ile Leu Lys Asp Leu Lys
485 490 495
His Asn Asp Gly Asn Phe Thr Glu Lys Gln Lys Ile Glu Leu Asn Lys
500 505 510
Leu Leu Gln Ile Asp Tyr Tyr Asn Leu Thr Lys Phe Tyr Gly Thr Val
515 520 525
Lys Leu Asp Thr Met Ile Phe Gly Val Ile Glu Tyr Cys Glu Arg Gly
530 535 540
Ser Leu Arg Glu Val Leu Asn Asp Thr Ile Ser Tyr Pro Asp Gly Thr
545 550 555 560
Phe Met Asp Trp Glu Phe Lys Ile Ser Val Leu Tyr Asp Ile Ala Lys
565 570 575
Gly Met Ser Tyr Leu His Ser Ser Lys Thr Glu Val His Gly Arg Leu
580 585 590
Lys Ser Thr Asn Cys Val Val Asp Ser Arg Met Val Val Lys Ile Thr
595 600 605
Asp Phe Gly Cys Asn Ser Ile Leu Pro Pro Lys Lys Asp Leu Trp Thr
610 615 620
Ala Pro Glu His Leu Arg Gln Ala Asn Val Ser Gln Lys Gly Asp Val
625 630 635 640
Tyr Ser Tyr Gly Ile Ile Ala Gln Glu Ile Ile Leu Arg Lys Glu Thr
645 650 655
Phe Tyr Thr Ser Ser Cys Arg Asp Arg Asn Glu Lys Ile Phe Arg Val
660 665 670
Glu Asn Ser Asn Gly Met Lys Pro Phe Arg Pro Asp Leu Phe Leu Glu
675 680 685
Thr Ala Glu Glu Lys Glu Leu Glu Val Tyr Leu Leu Val Lys Ser Cys
690 695 700
Trp Glu Glu Asp Pro Glu Lys Arg Pro Asp Phe Lys Lys Ile Glu Thr
705 710 715 720
Thr Leu Ala Lys Ile Phe Gly Leu Phe His Asp Gln Lys Asn Glu Ser
725 730 735
Tyr Met Asp Thr Leu Ile Arg Arg Leu Gln Leu Tyr Ser Arg Asn Leu
740 745 750
Glu His Leu Val Glu Glu Arg Thr Gln Leu Tyr Lys Ala Glu Arg Asp
755 760 765
Arg Ala Asp Arg Leu Asn Phe Met Leu Leu Pro Arg Leu Val Val Lys
770 775 780
Ser Leu Lys Glu Lys Gly Phe Val Glu Pro Glu Leu Tyr Glu Glu Val
785 790 795 800
Thr Ile Tyr Phe Ser Asp Ile Val Gly Phe Thr Thr Ile Cys Lys Tyr
805 810 815
Ser Thr Pro Met Glu Val Val Asp Met Leu Asn Asp Ile Tyr Lys Ser
820 825 830
Phe Asp His Ile Val Asp His His Asp Val Tyr Lys Val Glu Thr Ile
835 840 845
Gly Asp Ala Tyr Met Val Ala Ser Gly Leu Pro Lys Arg Asn Gly Asn
850 855 860
Arg His Ala Ile Asp Ile Ala Lys Met Ala Leu Glu Ile Leu Ser Phe
865 870 875 880
Met Gly Thr Phe Glu Leu Glu His Leu Pro Gly Leu Pro Ile Trp Ile
885 890 895
Arg Ile Gly Val His Ser Gly Pro Cys Ala Ala Gly Val Val Gly Ile
900 905 910
Lys Met Pro Arg Tyr Cys Leu Phe Gly Asp Thr Val Asn Thr Ala Ser
915 920 925
Arg Met Glu Ser Thr Gly Leu Pro Leu Arg Ile His Val Ser Gly Ser
930 935 940
Thr Ile Ala Ile Leu Lys Arg Thr Glu Cys Gln Phe Leu Tyr Glu Val
945 950 955 960
Arg Gly Glu Thr Tyr Leu Lys Gly Arg Gly Asn Glu Thr Thr Tyr Trp
965 970 975
Leu Thr Gly Met Lys Asp Gln Lys Phe Asn Leu Pro Thr Pro Pro Thr
980 985 990
Val Glu Asn Gln Gln Arg Leu Gln Ala Glu Phe Ser Asp Met Ile Ala
995 1000 1005
Asn Ser Leu Gln Lys Arg Gln Ala Ala Gly Ile Arg Ser Gln Lys
1010 1015 1020
Pro Arg Arg Val Ala Ser Tyr Lys Lys Gly Thr Leu Glu Tyr Leu
1025 1030 1035
Gln Leu Asn Thr Thr Asp Lys Glu Ser Thr Tyr Phe
1040 1045 1050
<210> 254
<211> 3150
<212> ДНК
<213> Macaca fascicularis
<400> 254
tcacaggtga gtcagaactg ccacaatggc agctatgaaa tcagcgtcct gatgatggac 60
aactcagcct ttgcagagcc cctggaaaac gtggaagatg cggtgaatga ggggctggaa 120
atagtgagag gacgtctgca aaacgctggc ctaaatgtga ctgtgaatgc ttctttcatg 180
tattcggatg gtctgattca taactccggc gactgccgga gcagcacctg tgaaggcctt 240
gacctactca ggaaaatttc aaatgcaaaa cggatgggct gtgtcctcat ggggccctca 300
tgtacatact ccaccttcca gatgtacctt gacacagaat tgagctaccc catgatctca 360
gctggaagtt ttggattgtc atgtgactat aaagaaacct taaccaggct gatgtctcca 420
gctagaaagt tgacatactt cttggttaac ttttggaaaa ccaatgatct acccttcaaa 480
acttattcct ggagcacttc gtatgtttac aagaatggta cggagtccga ggactgtttc 540
tggtacctta acgctctgga ggccagtgtt tcctatttct cccacgaact cagttttaag 600
ttggtgttaa gacaagataa ggagtttcag gatatcttaa tggaccacaa caggaaaagc 660
aatgtgattg ttatgtgtgg tgatccagag ttcctctaca agttgaaggg tgaccgagca 720
gtggctgaag acattgtcat tattctagtg gatcttttca atgaccagta ctttgaggac 780
aatgtcacag cccctgacta tatgaaaaat gtccttgttc tgacgcagtc tcctggcaat 840
tccctcctaa atagctcttt ctccaggaat ctatccccaa caaaacgaga ctttgctctt 900
gcctatttga atggaatcct gctctttgga catatgctaa agacatttct tgaaaatgga 960
gaaaatatta ccacccccaa atttgctcat gctttcagga atctcacttt tgaagggtat 1020
gacggtccag tgaccttgga tgactggggg gatgtggaca gtaccatggt gcttctgtat 1080
acgtctgtgg acaccaagaa atacaaggtt cttttgacct atgataccca cgtaaatcag 1140
accaaccctg tggatatgag ccccacattc acttggaaga actctaaact tcctaatgat 1200
attacagacc ggggccctca gatcctgatg attgcagtct tcaccctcac cggagctgtg 1260
gtgctgctcc tgcttgtcgc tctcctgatg ctcagaaaat ataaaaaaga ttatgaactt 1320
cgtcagaaaa aatggtccca cattcctcct gaaaatatct ttcctctgga gaccaatgag 1380
accaatcacg ttagcctgaa gatcgatgat gacaaaagac gagatacaat ccagagacta 1440
cgacagtgca aatacgacaa aaagcgagtg attctcaaag atctcaagca caatgatggt 1500
aatttcactg aaaaacagaa gatagaattg aacaagttgc ttcagattga ctattacaac 1560
ctgaccaagt tctatggcac cgtgaaactt gataccatga tcttcggggt gatagaatac 1620
tgtgagagag gatccctccg ggaagtttta aatgacacaa tttcctaccc tgatggcaca 1680
ttcatggatt gggagtttaa gatctctgtc ctgtatgaca ttgctaaggg aatgtcatat 1740
ctgcactcca gtaagacaga agtccatggt cgtctgaaat ctaccaactg cgtagtggac 1800
agtagaatgg tggtgaagat cactgatttt ggctgcaatt ccattttacc tccaaaaaaa 1860
gacctgtgga cagctccaga gcacctccgc caagccaacg tctctcagaa aggagatgtg 1920
tacagctacg ggatcatcgc acaggagatc atcctgcgga aagaaacctt ctacacttcg 1980
agctgtcgag accggaacga gaagattttc agagtggaaa attccaatgg aatgaaaccc 2040
ttccgtccag atttattctt ggaaacggca gaggaaaaag agctagaagt gtacctactt 2100
gtaaaaagct gttgggaaga agatccagaa aagagaccag atttcaaaaa aattgagact 2160
acacttgcca agatatttgg actttttcat gaccaaaaaa atgaaagcta tatggatacc 2220
ttgatccgac gtctacagct atattctcga aacctggaac atctggtaga ggaaaggaca 2280
cagctataca aggcagagag ggacagggct gacagactta actttatgtt gcttccaagg 2340
ctagtggtaa agtctctgaa ggagaaaggc tttgtagagc cggaactata tgaggaagtt 2400
acaatctact tcagtgacat tgtaggtttc actactatct gcaaatacag cacccccatg 2460
gaagtggtgg acatgcttaa tgacatctat aagagttttg accacattgt tgatcatcat 2520
gatgtctaca aggtggaaac cattggtgat gcctacatgg tggctagtgg tttgcctaag 2580
agaaatggca atcggcatgc aatagacatt gccaagatgg ccttggaaat cctcagcttc 2640
atggggacct ttgagctgga gcatcttcct ggcctcccaa tatggattcg cattggcgtt 2700
cactctggtc cctgcgctgc tggagttgtg ggaatcaaga tgcctcgtta ttgtctattt 2760
ggagatacag tcaacacagc ctctaggatg gaatccactg gcctcccttt gaggattcat 2820
gtgagtggct ccaccatagc cattctgaag agaactgagt gccagttcct gtatgaagtg 2880
agaggagaaa cgtacttaaa gggaagagga aatgagacta cctactggct gaccgggatg 2940
aaggaccaga aattcaacct gccaacccct cctactgtgg agaatcaaca gcgtttgcaa 3000
gcagaatttt cagacatgat tgccaactct ttacagaaaa gacaggcggc agggataaga 3060
agccaaaaac ccagacgagt agccagctat aaaaaaggca ctctggaata cttgcaactg 3120
aataccacgg acaaggagag cacctatttt 3150
<210> 255
<211> 1072
<212> БЕЛОК
<213> Mus musculus
<400> 255
Met Thr Ser Leu Leu Gly Leu Ala Val Arg Leu Leu Leu Phe Gln Pro
1 5 10 15
Ala Leu Met Val Phe Trp Ala Ser Gln Val Arg Gln Asn Cys Arg Asn
20 25 30
Gly Ser Tyr Glu Ile Ser Val Leu Met Met Asp Asn Ser Ala Tyr Lys
35 40 45
Glu Pro Met Gln Asn Leu Arg Glu Ala Val Glu Glu Gly Leu Asp Ile
50 55 60
Val Arg Lys Arg Leu Arg Glu Ala Asp Leu Asn Val Thr Val Asn Ala
65 70 75 80
Thr Phe Ile Tyr Ser Asp Gly Leu Ile His Lys Ser Gly Asp Cys Arg
85 90 95
Ser Ser Thr Cys Glu Gly Leu Asp Leu Leu Arg Glu Ile Thr Arg Asp
100 105 110
His Lys Met Gly Cys Ala Leu Met Gly Pro Ser Cys Thr Tyr Ser Thr
115 120 125
Phe Gln Met Tyr Leu Asp Thr Glu Leu Asn Tyr Pro Met Ile Ser Ala
130 135 140
Gly Ser Tyr Gly Leu Ser Cys Asp Tyr Lys Glu Thr Leu Thr Arg Ile
145 150 155 160
Leu Pro Pro Ala Arg Lys Leu Met Tyr Phe Leu Val Asp Phe Trp Lys
165 170 175
Val Asn Asn Ala Ser Phe Lys Pro Phe Ser Trp Asn Ser Ser Tyr Val
180 185 190
Tyr Lys Asn Gly Ser Glu Pro Glu Asp Cys Phe Trp Tyr Leu Asn Ala
195 200 205
Leu Glu Ala Gly Val Ser Tyr Phe Ser Glu Val Leu Asn Phe Lys Asp
210 215 220
Val Leu Arg Arg Ser Glu Gln Phe Gln Glu Ile Leu Thr Gly His Asn
225 230 235 240
Arg Lys Ser Asn Val Ile Val Met Cys Gly Thr Pro Glu Ser Phe Tyr
245 250 255
Asp Val Lys Gly Asp Leu Gln Val Ala Glu Asp Thr Val Val Ile Leu
260 265 270
Val Asp Leu Phe Ser Asn His Tyr Phe Glu Glu Asn Thr Thr Ala Pro
275 280 285
Glu Tyr Met Asp Asn Val Leu Val Leu Thr Leu Pro Ser Glu Gln Ser
290 295 300
Thr Ser Asn Thr Ser Val Ala Glu Arg Phe Ser Ser Gly Arg Ser Asp
305 310 315 320
Phe Ser Leu Ala Tyr Leu Glu Gly Thr Leu Leu Phe Gly His Met Leu
325 330 335
Gln Thr Phe Leu Glu Asn Gly Glu Asn Val Thr Gly Pro Lys Phe Ala
340 345 350
Arg Ala Phe Arg Asn Leu Thr Phe Gln Gly Phe Ala Gly Pro Val Thr
355 360 365
Leu Asp Asp Ser Gly Asp Ile Asp Asn Ile Met Ser Leu Leu Tyr Val
370 375 380
Ser Leu Asp Thr Arg Lys Tyr Lys Val Leu Met Lys Tyr Asp Thr His
385 390 395 400
Lys Asn Lys Thr Ile Pro Val Ala Glu Asn Pro Asn Phe Ile Trp Lys
405 410 415
Asn His Lys Leu Pro Asn Asp Val Pro Gly Leu Gly Pro Gln Ile Leu
420 425 430
Met Ile Ala Val Phe Thr Leu Thr Gly Ile Leu Val Val Leu Leu Leu
435 440 445
Ile Ala Leu Leu Val Leu Arg Lys Tyr Arg Arg Asp His Ala Leu Arg
450 455 460
Gln Lys Lys Trp Ser His Ile Pro Ser Glu Asn Ile Phe Pro Leu Glu
465 470 475 480
Thr Asn Glu Thr Asn His Ile Ser Leu Lys Ile Asp Asp Asp Arg Arg
485 490 495
Arg Asp Thr Ile Gln Arg Val Arg Gln Cys Lys Tyr Asp Lys Lys Lys
500 505 510
Val Ile Leu Lys Asp Leu Lys His Ser Asp Gly Asn Phe Ser Glu Lys
515 520 525
Gln Lys Ile Asp Leu Asn Lys Leu Leu Gln Ser Asp Tyr Tyr Asn Leu
530 535 540
Thr Lys Phe Tyr Gly Thr Val Lys Leu Asp Thr Arg Ile Phe Gly Val
545 550 555 560
Val Glu Tyr Cys Glu Arg Gly Ser Leu Arg Glu Val Leu Asn Asp Thr
565 570 575
Ile Ser Tyr Pro Asp Gly Thr Phe Met Asp Trp Glu Phe Lys Ile Ser
580 585 590
Val Leu Asn Asp Ile Ala Lys Gly Met Ser Tyr Leu His Ser Ser Lys
595 600 605
Ile Glu Val His Gly Arg Leu Lys Ser Thr Asn Cys Val Val Asp Ser
610 615 620
Arg Met Val Val Lys Ile Thr Asp Phe Gly Cys Asn Ser Ile Leu Pro
625 630 635 640
Pro Lys Lys Asp Leu Trp Thr Ala Pro Glu His Leu Arg Gln Ala Thr
645 650 655
Ile Ser Gln Lys Gly Asp Val Tyr Ser Phe Ala Ile Ile Ala Gln Glu
660 665 670
Ile Ile Leu Arg Lys Glu Thr Phe Tyr Thr Leu Ser Cys Arg Asp His
675 680 685
Asn Glu Lys Ile Phe Arg Val Glu Asn Ser Tyr Gly Lys Pro Phe Arg
690 695 700
Pro Asp Leu Phe Leu Glu Thr Ala Asp Glu Lys Glu Leu Glu Val Tyr
705 710 715 720
Leu Leu Val Lys Ser Cys Trp Glu Glu Asp Pro Glu Lys Arg Pro Asp
725 730 735
Phe Lys Lys Ile Glu Ser Thr Leu Ala Lys Ile Phe Gly Leu Phe His
740 745 750
Asp Gln Lys Asn Glu Ser Tyr Met Asp Thr Leu Ile Arg Arg Leu Gln
755 760 765
Leu Tyr Ser Arg Asn Leu Glu His Leu Val Glu Glu Arg Thr Gln Leu
770 775 780
Tyr Lys Ala Glu Arg Asp Arg Ala Asp His Leu Asn Phe Met Leu Leu
785 790 795 800
Pro Arg Leu Val Val Lys Ser Leu Lys Glu Lys Gly Ile Val Glu Pro
805 810 815
Glu Leu Tyr Glu Glu Val Thr Ile Tyr Phe Ser Asp Ile Val Gly Phe
820 825 830
Thr Thr Ile Cys Lys Tyr Ser Thr Pro Met Glu Val Val Asp Met Leu
835 840 845
Asn Asp Ile Tyr Lys Ser Phe Asp Gln Ile Val Asp His His Asp Val
850 855 860
Tyr Lys Val Glu Thr Ile Gly Asp Ala Tyr Val Val Ala Ser Gly Leu
865 870 875 880
Pro Met Arg Asn Gly Asn Arg His Ala Val Asp Ile Ser Lys Met Ala
885 890 895
Leu Asp Ile Leu Ser Phe Ile Gly Thr Phe Glu Leu Glu His Leu Pro
900 905 910
Gly Leu Pro Val Trp Ile Arg Ile Gly Val His Ser Gly Pro Cys Ala
915 920 925
Ala Gly Val Val Gly Ile Lys Met Pro Arg Tyr Cys Leu Phe Gly Asp
930 935 940
Thr Val Asn Thr Ala Ser Arg Met Glu Ser Thr Gly Leu Pro Leu Arg
945 950 955 960
Ile His Met Ser Ser Ser Thr Ile Thr Ile Leu Lys Arg Thr Asp Cys
965 970 975
Gln Phe Leu Tyr Glu Val Arg Gly Glu Thr Tyr Leu Lys Gly Arg Gly
980 985 990
Thr Glu Thr Thr Tyr Trp Leu Thr Gly Met Lys Asp Gln Glu Tyr Asn
995 1000 1005
Leu Pro Ser Pro Pro Thr Val Glu Asn Gln Gln Arg Leu Gln Thr
1010 1015 1020
Glu Phe Ser Asp Met Ile Val Ser Ala Leu Gln Lys Arg Gln Ala
1025 1030 1035
Ser Gly Lys Lys Ser Arg Arg Pro Thr Arg Val Ala Ser Tyr Lys
1040 1045 1050
Lys Gly Phe Leu Glu Tyr Met Gln Leu Asn Asn Ser Asp His Asp
1055 1060 1065
Ser Thr Tyr Phe
1070
<210> 256
<211> 3951
<212> ДНК
<213> Mus musculus
<400> 256
gaccagtgtg gcaagaccag aaaggtgcgt ggggaagaga agaccaaggg actctgctag 60
cgactctcca gaggggctcc ctgtgtctct aaaagcgagc aatccagggg ggcatggtgc 120
tacggtgagc caggtcatga cgtcactgct gggcttggct gtgcggttac tgctcttcca 180
gcccgcgctg atggtgttct gggcctctca ggtgaggcag aactgccgca atggcagcta 240
cgagatcagc gtcctgatga tggacaactc agcctacaaa gaacctatgc aaaacctgag 300
ggaggctgtg gaggaaggac tggacatagt gcgaaagcgc ctgcgtgaag ccgacctaaa 360
tgtgactgtg aacgcgactt tcatctactc cgacggtctg attcataagt caggtgactg 420
ccggagcagc acctgtgaag gccttgacct actcagggag attacaagag atcataagat 480
gggctgcgcc ctcatggggc cctcgtgcac gtattccacc ttccagatgt acctcgacac 540
agagttgaac tatcccatga tttccgctgg aagttatgga ttgtcctgtg actataagga 600
aaccctaacc aggatcctgc ctccagccag gaagctgatg tacttcttgg tcgatttctg 660
gaaagtcaac aatgcatctt tcaaaccctt ttcctggaac tcttcgtatg tttacaagaa 720
tggatcggaa cctgaagatt gtttctggta cctcaatgct ctggaggctg gggtgtccta 780
tttttctgag gtgctcaact tcaaggatgt actgagacgc agcgaacagt tccaggaaat 840
cttaacaggc cataacagaa agagcaatgt gattgttatg tgtggcacgc cagaaagctt 900
ctatgatgtg aaaggtgacc tccaagtggc tgaagatact gttgtcatcc tggtagatct 960
gttcagtaac cattactttg aggagaacac cacagctcct gagtatatgg acaatgtcct 1020
cgtcctgacg ctgccgtctg aacagtccac ctcaaacacc tctgtcgccg agaggttttc 1080
atcggggaga agtgactttt ctctcgctta cttggaggga accttgctat ttggacacat 1140
gctgcagacg tttcttgaaa atggagaaaa tgtcacgggt cccaagtttg ctcgtgcatt 1200
caggaatctc acttttcaag gctttgcagg acctgtgact ctggatgaca gtggggacat 1260
tgacaacatt atgtcccttc tgtatgtgtc tctggatacc aggaaataca aggttcttat 1320
gaagtatgac acccacaaaa acaaaactat tccggtggct gagaacccca acttcatctg 1380
gaagaaccac aagctcccca atgacgttcc tgggctgggc cctcaaatcc tgatgattgc 1440
cgtcttcacg ctcacgggga tcctggtagt tctgctgctg attgccctcc tcgtgctgag 1500
aaaatacaga agagatcatg cacttcgaca gaagaaatgg tcccacattc cttctgaaaa 1560
catctttcct ctggagacca acgagaccaa ccacatcagc ctgaagattg acgatgacag 1620
gagacgagac acaatccaga gagtgcgaca gtgcaaatac gacaagaaga aagtgattct 1680
gaaagacctc aagcacagcg acgggaactt cagtgagaag cagaagatag acctgaacaa 1740
gttgctgcag tctgactact acaacctgac taagttctac ggcaccgtga agctggacac 1800
caggatcttt ggggtggttg agtactgcga gaggggatcc ctccgggaag tgttaaacga 1860
cacaatttcc taccctgacg gcacgttcat ggattgggag tttaagatct ctgtcttaaa 1920
tgacatcgct aaggggatgt cctacctgca ctccagtaag attgaagtcc acgggcgtct 1980
caaatccacc aactgcgtgg tggacagccg catggtggtg aagatcaccg actttgggtg 2040
caattccatc ctgcctccaa aaaaagacct gtggacggcc ccggagcacc tgcgccaggc 2100
caccatctct cagaaaggag acgtgtacag cttcgccatc attgcccagg agatcatcct 2160
ccgtaaggag actttttaca cgctgagctg tcgggatcac aatgagaaga ttttcagagt 2220
ggaaaattca tacgggaaac ctttccgccc agacctcttc ctggagactg cagatgagaa 2280
ggagctggag gtctatctac ttgtcaaaag ctgttgggag gaggatccag aaaagaggcc 2340
agatttcaag aaaatcgaga gcacactggc caagatattt ggccttttcc atgaccagaa 2400
aaacgagtct tacatggaca ccttgatccg acgtctccag ctgtactctc gaaacctgga 2460
acatctggtg gaggaaagga ctcagctgta caaggcggag agggacaggg ctgaccacct 2520
taacttcatg ctcctcccac ggctggtggt aaagtcactg aaggagaaag gcatcgtgga 2580
gccagagctg tacgaagaag tcacaatcta cttcagtgac attgtgggct tcaccaccat 2640
ctgcaagtat agcacgccca tggaggtggt ggacatgctc aacgacatct acaagagctt 2700
tgaccagatt gtggaccacc atgacgtcta caaggtagaa accatcggtg acgcctacgt 2760
ggtggccagc ggtctgccta tgagaaacgg caaccgacac gcggtagaca tttccaagat 2820
ggccttggac atcctcagct tcatagggac ctttgagttg gagcatctcc ctggcctccc 2880
cgtgtggatc cgcattggag ttcattctgg gccctgcgct gctggtgttg tggggatcaa 2940
gatgcctcgc tattgcctgt ttggagacac tgtcaacact gcctccagga tggaatccac 3000
cggcctcccc ttgaggattc acatgagcag ctccaccata accatcctga agagaacgga 3060
ttgccagttc ctgtatgaag tgaggggaga aacctactta aagggaagag ggacagagac 3120
cacatactgg ctgactggga tgaaggacca agaatacaac ctgccatccc caccgacagt 3180
ggagaaccaa cagcgtctgc agactgagtt ctcagacatg atcgttagcg ccttacagaa 3240
aagacaggcc tcgggcaaga agagccggag gcccactcgg gtggccagct acaagaaagg 3300
ctttctggaa tacatgcagc tgaacaattc agaccacgat agcacctatt tttagaccaa 3360
gtgaggtctg agaactgaca gtagcaacct cctatatcat gaatctgtat tttccagaga 3420
cctcaacaac atagacaagc acttagcctc agtgccctga ctggaacgta gaaccaaccc 3480
ctcaagtcat gtgggtctga ttttgggttg gttggttggt tggttggttg gttttggttt 3540
tgttgagaca gagtttcgtg tatcccaagc tggtctcaaa ctcactgtgt agcagcagat 3600
gactttggac ttctaagatc atccgtgtgt gttcctgaca gtgtgatgag tgtatacgtc 3660
agagccctgt cccacagttc tccatggagc atcaacctga atggaggagc gaggaggggg 3720
aatgtcctgt gctgtacaga acttggggtt ttgttctaat tttatctctg gtttggtttt 3780
gttttctggc tccttccctc tctatgtgtg agagaagttt ttaaattgtc tgaattgtat 3840
gctaagtagc ttatctacaa gaaagtgtgt ttaactagtg atttttgcag aaaccatgct 3900
ggatattagg taaaaaataa aagtgtttag agtctaaaaa aaaaaaaaaa a 3951
<210> 257
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 257
Thr Tyr Trp Met Gln
1 5
<210> 258
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 258
Lys Tyr Trp Met Gln
1 5
<210> 259
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 259
Ser Tyr Trp Met His
1 5
<210> 260
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 260
Asp Thr Tyr Ile His
1 5
<210> 261
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 261
Asp Tyr Ile Met Leu
1 5
<210> 262
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 262
Tyr Pro Gly Asp Gly Met
1 5
<210> 263
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 263
Tyr Pro Gly Asp Gly Phe
1 5
<210> 264
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 264
Lys Pro Ser Asn Gly Leu
1 5
<210> 265
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 265
Asp Pro Ala Asn Gly Asn
1 5
<210> 266
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 266
Asn Pro Tyr Tyr Gly Ser
1 5
<210> 267
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 267
Lys Pro Ser Asn Glu Leu
1 5
<210> 268
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 268
Asp Tyr Tyr Met Thr
1 5
<210> 269
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 269
Arg Asn Arg Ala Arg Gly Tyr Thr
1 5
<210> 270
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 270
Arg Asn Gln Ala Arg Gly Tyr Thr
1 5
<210> 271
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 271
Ser Tyr Trp Met Ser
1 5
<210> 272
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 272
Lys Gln Asp Gly Ser Glu
1 5
<210> 273
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 273
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Asn Gln Ala Arg Gly Tyr Thr Ser Asp His Asn Pro
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr Val Leu Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 274
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 274
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Thr Ser Asp Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Gly Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Asp Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 275
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 275
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Thr Ser Ser Glu Ser Leu Phe Asn Val
20 25 30
Arg Ser Gly Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Asp Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 276
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 276
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Thr Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Gly Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Asp Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 277
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 277
Gly Phe Thr Phe Ser Asp Tyr
1 5
<210> 278
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 278
Thr Ser Asp Gln Ser Leu Phe Asn Val Arg Ser Gly Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 279
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 279
Thr Ser Ser Glu Ser Leu Phe Asn Val Arg Ser Gly Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 280
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 280
Thr Ser Ser Gln Ser Leu Phe Asn Val Arg Ser Gly Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 281
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 281
Trp Ala Ser Asp Arg Glu Ser
1 5
<210> 282
<211> 461
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 282
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Gln Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 283
<211> 465
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 283
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Thr Ser Asp Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Gly Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Asp Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Ser Ser Tyr Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn
165 170 175
Glu Leu Thr Asn Val His Glu Lys Phe Lys Asp Arg Phe Thr Ile Ser
180 185 190
Val Asp Lys Ala Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr
210 215 220
Glu Gly Tyr Trp Phe Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr
225 230 235 240
Val Ser Ser Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly
465
<210> 284
<211> 461
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 284
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Gln Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 285
<211> 465
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 285
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Thr Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Gly Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Asp Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Ser Ser Tyr Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn
165 170 175
Glu Leu Thr Asn Val His Glu Lys Phe Lys Asp Arg Phe Thr Ile Ser
180 185 190
Val Asp Lys Ala Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr
210 215 220
Glu Gly Tyr Trp Phe Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr
225 230 235 240
Val Ser Ser Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly
465
<210> 286
<211> 461
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 286
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Gln Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
<210> 287
<211> 465
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 287
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Thr Ser Ser Glu Ser Leu Phe Asn Val
20 25 30
Arg Ser Gly Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Asp Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Ser Ser Tyr Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn
165 170 175
Glu Leu Thr Asn Val His Glu Lys Phe Lys Asp Arg Phe Thr Ile Ser
180 185 190
Val Asp Lys Ala Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr
210 215 220
Glu Gly Tyr Trp Phe Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr
225 230 235 240
Val Ser Ser Gly Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly
465
<210> 288
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 288
gacatccaga tgacccagtc cccctcttct ctgtctgcct ctgtgggcga cagagtgacc 60
atcacctgca caagcgacca gtcactgttt aatgtccgca gcggcaaaaa ctatcttgcg 120
tggtatcagc agaagcctgg caaggctccc aagctgctga tctactgggc cagtgaccga 180
gaatccggcg tgccttccag attctccggc tctggctctg gcaccgattt caccctgacc 240
atctcctccc tccagcctga ggatttcgcc acctactact gcaaacagtc ttacgacctt 300
ttcacttttg gcggcggaac aaaggtggag atcaag 336
<210> 289
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 289
gaagtgcagc ttgttgaatc tggcggcggt ttggttcagc ccggtggatc actgcgactc 60
agttgcgcag ctagcggctt caccttttct gattactaca tgacatgggt acgacaggcg 120
ccaggcaagg gtttggaatg ggtagcattc atacgcaatc aggcacgcgg gtacacttca 180
gaccacaatc cctcagtaaa aggaagattt accatctcaa gagacaatgc caaaaattca 240
ctctacctgc aaatgaactc acttcgcgcc gaggataccg ccgtgtatta ctgtgccaga 300
gacagaccat cttattacgt gctggactat tggggacagg gcactacagt caccgtcagc 360
tct 363
<210> 290
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 290
gacatccaga tgacccagtc cccctcttct ctgtctgcct ctgtgggcga cagagtgacc 60
atcacctgca caagctcaga gtcactgttt aatgtccgca gcggcaaaaa ctatcttgcg 120
tggtatcagc agaagcctgg caaggctccc aagctgctga tctactgggc cagtgaccga 180
gaatccggcg tgccttccag attctccggc tctggctctg gcaccgattt caccctgacc 240
atctcctccc tccagcctga ggatttcgcc acctactact gcaaacagtc ttacgacctt 300
ttcacttttg gcggcggaac aaaggtggag atcaag 336
<210> 291
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 291
gaagtgcagc ttgttgaatc tggcggcggt ttggttcagc ccggtggatc actgcgactc 60
agttgcgcag ctagcggctt caccttttct gattactaca tgacatgggt acgacaggcg 120
ccaggcaagg gtttggaatg ggtagcattc atacgcaatc aggcacgcgg gtacacttca 180
gaccacaatc cctcagtaaa aggaagattt accatctcaa gagacaatgc caaaaattca 240
ctctacctgc aaatgaactc acttcgcgcc gaggataccg ccgtgtatta ctgtgccaga 300
gacagaccat cttattacgt gctggactat tggggacagg gcactacagt caccgtcagc 360
tct 363
<210> 292
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 292
gacatccaga tgacccagtc cccctcttct ctgtctgcct ctgtgggcga cagagtgacc 60
atcacctgca caagctcaca gtcactgttt aatgtccgca gcggcaaaaa ctatcttgcg 120
tggtatcagc agaagcctgg caaggctccc aagctgctga tctactgggc cagtgaccga 180
gaatccggcg tgccttccag attctccggc tctggctctg gcaccgattt caccctgacc 240
atctcctccc tccagcctga ggatttcgcc acctactact gcaaacagtc ttacgacctt 300
ttcacttttg gcggcggaac aaaggtggag atcaag 336
<210> 293
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 293
gaagtgcagc ttgttgaatc tggcggcggt ttggttcagc ccggtggatc actgcgactc 60
agttgcgcag ctagcggctt caccttttct gattactaca tgacatgggt acgacaggcg 120
ccaggcaagg gtttggaatg ggtagcattc atacgcaatc aggcacgcgg gtacacttca 180
gaccacaatc cctcagtaaa aggaagattt accatctcaa gagacaatgc caaaaattca 240
ctctacctgc aaatgaactc acttcgcgcc gaggataccg ccgtgtatta ctgtgccaga 300
gacagaccat cttattacgt gctggactat tggggacagg gcactacagt caccgtcagc 360
tct 363
<210> 294
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 294
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Arg
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Lys Leu Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Leu Ser Asp Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 295
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 295
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly His Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 296
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 296
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Glu Leu Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Pro Leu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 297
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 297
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 298
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 298
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Glu Leu Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Pro Leu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 299
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 299
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 300
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 300
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Arg Trp Asn Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 301
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 301
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 302
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 302
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Arg Gly Phe Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 303
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 303
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 304
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 304
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Thr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Glu Leu Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 305
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 305
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 306
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 306
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Pro
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Thr Gly Trp Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Thr Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 307
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 307
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Lys His Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 308
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 308
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Arg Gly Trp Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 309
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 309
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Arg His Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 310
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 310
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Thr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Arg Gly Phe Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Gln Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 311
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 311
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asn Trp Tyr
20 25 30
Gly Ser Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 312
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 312
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Thr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Thr Lys Tyr Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Ile Phe Thr Arg Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 313
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 313
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asn Trp Tyr
20 25 30
Gly Ser Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 314
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 314
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Thr Lys Tyr Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Phe Thr Asn Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 315
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 315
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Ser Ile Tyr
20 25 30
Gly Ser Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 316
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 316
Met Lys Thr Leu Leu Leu Asp Leu Ala Leu Trp Ser Leu Leu Phe Gln
1 5 10 15
Pro Gly Trp Leu
20
<210> 317
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 317
Leu Asp Leu Ala Leu Trp Ser Leu Leu Phe Gln Pro Gly Trp Leu Ser
1 5 10 15
Phe Ser Ser Gln
20
<210> 318
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 318
Trp Ser Leu Leu Phe Gln Pro Gly Trp Leu Ser Phe Ser Ser Gln Val
1 5 10 15
Ser Gln Asn Cys
20
<210> 319
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 319
Gln Pro Gly Trp Leu Ser Phe Ser Ser Gln Val Ser Gln Asn Cys His
1 5 10 15
Asn Gly Ser Tyr
20
<210> 320
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 320
Ser Phe Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu
1 5 10 15
Ile Ser Val Leu
20
<210> 321
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 321
Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser Val Leu Met
1 5 10 15
Met Gly Asn Ser
20
<210> 322
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 322
His Asn Gly Ser Tyr Glu Ile Ser Val Leu Met Met Gly Asn Ser Ala
1 5 10 15
Phe Ala Glu Pro
20
<210> 323
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 323
Glu Ile Ser Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu
1 5 10 15
Lys Asn Leu Glu
20
<210> 324
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 324
Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu Glu Asp
1 5 10 15
Ala Val Asn Glu
20
<210> 325
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 325
Ala Phe Ala Glu Pro Leu Lys Asn Leu Glu Asp Ala Val Asn Glu Gly
1 5 10 15
Leu Glu Ile Val
20
<210> 326
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 326
Leu Lys Asn Leu Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg
1 5 10 15
Gly Arg Leu Gln
20
<210> 327
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 327
Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln Asn
1 5 10 15
Ala Gly Leu Asn
20
<210> 328
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 328
Gly Leu Glu Ile Val Arg Gly Arg Leu Gln Asn Ala Gly Leu Asn Val
1 5 10 15
Thr Val Asn Ala
20
<210> 329
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 329
Arg Gly Arg Leu Gln Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr
1 5 10 15
Phe Met Tyr Ser
20
<210> 330
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 330
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
1 5 10 15
Gly Leu Ile His
20
<210> 331
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 331
Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp Gly Leu Ile His Asn
1 5 10 15
Ser Gly Asp Cys
20
<210> 332
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 332
Thr Phe Met Tyr Ser Asp Gly Leu Ile His Asn Ser Gly Asp Cys Arg
1 5 10 15
Ser Ser Thr Cys
20
<210> 333
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 333
Asp Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu
1 5 10 15
Gly Leu Asp Leu
20
<210> 334
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 334
Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly Leu Asp Leu Leu
1 5 10 15
Arg Lys Ile Ser
20
<210> 335
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 335
Arg Ser Ser Thr Cys Glu Gly Leu Asp Leu Leu Arg Lys Ile Ser Asn
1 5 10 15
Ala Gln Arg Met
20
<210> 336
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 336
Glu Gly Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly
1 5 10 15
Cys Val Leu Ile
20
<210> 337
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 337
Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val Leu Ile Gly
1 5 10 15
Pro Ser Cys Thr
20
<210> 338
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 338
Asn Ala Gln Arg Met Gly Cys Val Leu Ile Gly Pro Ser Cys Thr Tyr
1 5 10 15
Ser Thr Phe Gln
20
<210> 339
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 339
Gly Cys Val Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met
1 5 10 15
Tyr Leu Asp Thr
20
<210> 340
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 340
Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp Thr Glu
1 5 10 15
Leu Ser Tyr Pro
20
<210> 341
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 341
Tyr Ser Thr Phe Gln Met Tyr Leu Asp Thr Glu Leu Ser Tyr Pro Met
1 5 10 15
Ile Ser Ala Gly
20
<210> 342
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 342
Met Tyr Leu Asp Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser
1 5 10 15
Phe Gly Leu Ser
20
<210> 343
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 343
Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser Cys
1 5 10 15
Asp Tyr Lys Glu
20
<210> 344
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 344
Met Ile Ser Ala Gly Ser Phe Gly Leu Ser Cys Asp Tyr Lys Glu Thr
1 5 10 15
Leu Thr Arg Leu
20
<210> 345
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 345
Ser Phe Gly Leu Ser Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met
1 5 10 15
Ser Pro Ala Arg
20
<210> 346
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 346
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
1 5 10 15
Leu Met Tyr Phe
20
<210> 347
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 347
Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys Leu Met Tyr Phe Leu
1 5 10 15
Val Asn Phe Trp
20
<210> 348
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 348
Met Ser Pro Ala Arg Lys Leu Met Tyr Phe Leu Val Asn Phe Trp Lys
1 5 10 15
Thr Asn Asp Leu
20
<210> 349
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 349
Lys Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro
1 5 10 15
Phe Lys Thr Tyr
20
<210> 350
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 350
Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe Lys Thr Tyr Ser
1 5 10 15
Trp Ser Thr Ser
20
<210> 351
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 351
Lys Thr Asn Asp Leu Pro Phe Lys Thr Tyr Ser Trp Ser Thr Ser Tyr
1 5 10 15
Val Tyr Lys Asn
20
<210> 352
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 352
Pro Phe Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly
1 5 10 15
Thr Glu Thr Glu
20
<210> 353
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 353
Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu Thr Glu Asp
1 5 10 15
Cys Phe Trp Tyr
20
<210> 354
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 354
Tyr Val Tyr Lys Asn Gly Thr Glu Thr Glu Asp Cys Phe Trp Tyr Leu
1 5 10 15
Asn Ala Leu Glu
20
<210> 355
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 355
Gly Thr Glu Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala
1 5 10 15
Ser Val Ser Tyr
20
<210> 356
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 356
Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser Tyr Phe
1 5 10 15
Ser His Glu Leu
20
<210> 357
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 357
Leu Asn Ala Leu Glu Ala Ser Val Ser Tyr Phe Ser His Glu Leu Gly
1 5 10 15
Phe Lys Val Val
20
<210> 358
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 358
Ala Ser Val Ser Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu
1 5 10 15
Arg Gln Asp Lys
20
<210> 359
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 359
Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys Glu
1 5 10 15
Phe Gln Asp Ile
20
<210> 360
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 360
Gly Phe Lys Val Val Leu Arg Gln Asp Lys Glu Phe Gln Asp Ile Leu
1 5 10 15
Met Asp His Asn
20
<210> 361
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 361
Leu Arg Gln Asp Lys Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg
1 5 10 15
Lys Ser Asn Val
20
<210> 362
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 362
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
1 5 10 15
Ile Met Cys Gly
20
<210> 363
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 363
Leu Met Asp His Asn Arg Lys Ser Asn Val Ile Ile Met Cys Gly Gly
1 5 10 15
Pro Glu Phe Leu
20
<210> 364
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 364
Arg Lys Ser Asn Val Ile Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr
1 5 10 15
Lys Leu Lys Gly
20
<210> 365
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 365
Ile Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp
1 5 10 15
Arg Ala Val Ala
20
<210> 366
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 366
Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg Ala Val Ala Glu
1 5 10 15
Asp Ile Val Ile
20
<210> 367
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 367
Tyr Lys Leu Lys Gly Asp Arg Ala Val Ala Glu Asp Ile Val Ile Ile
1 5 10 15
Leu Val Asp Leu
20
<210> 368
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 368
Asp Arg Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe
1 5 10 15
Asn Asp Gln Tyr
20
<210> 369
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 369
Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp Gln Tyr Leu
1 5 10 15
Glu Asp Asn Val
20
<210> 370
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 370
Ile Leu Val Asp Leu Phe Asn Asp Gln Tyr Leu Glu Asp Asn Val Thr
1 5 10 15
Ala Pro Asp Tyr
20
<210> 371
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 371
Phe Asn Asp Gln Tyr Leu Glu Asp Asn Val Thr Ala Pro Asp Tyr Met
1 5 10 15
Lys Asn Val Leu
20
<210> 372
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 372
Leu Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val Leu Val
1 5 10 15
Leu Thr Leu Ser
20
<210> 373
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 373
Thr Ala Pro Asp Tyr Met Lys Asn Val Leu Val Leu Thr Leu Ser Pro
1 5 10 15
Gly Asn Ser Leu
20
<210> 374
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 374
Met Lys Asn Val Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu
1 5 10 15
Asn Ser Ser Phe
20
<210> 375
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 375
Met Lys Asn Val Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu
1 5 10 15
Asn Ser Ser Phe
20
<210> 376
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 376
Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe Ser
1 5 10 15
Arg Asn Leu Ser
20
<210> 377
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 377
Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe Ser Arg Asn Leu Ser Pro
1 5 10 15
Thr Lys Arg Asp
20
<210> 378
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 378
Leu Asn Ser Ser Phe Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe
1 5 10 15
Ala Leu Ala Tyr
20
<210> 379
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 379
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
1 5 10 15
Asn Gly Ile Leu
20
<210> 380
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 380
Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu Asn Gly Ile Leu Leu
1 5 10 15
Phe Gly His Met
20
<210> 381
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 381
Phe Ala Leu Ala Tyr Leu Asn Gly Ile Leu Leu Phe Gly His Met Leu
1 5 10 15
Lys Ile Phe Leu
20
<210> 382
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 382
Leu Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu
1 5 10 15
Asn Gly Glu Asn
20
<210> 383
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 383
Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn Gly Glu Asn Ile
1 5 10 15
Thr Thr Pro Lys
20
<210> 384
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 384
Leu Lys Ile Phe Leu Glu Asn Gly Glu Asn Ile Thr Thr Pro Lys Phe
1 5 10 15
Ala His Ala Phe
20
<210> 385
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 385
Glu Asn Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg
1 5 10 15
Asn Leu Thr Phe
20
<210> 386
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 386
Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu Thr Phe Glu
1 5 10 15
Gly Tyr Asp Gly
20
<210> 387
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 387
Phe Ala His Ala Phe Arg Asn Leu Thr Phe Glu Gly Tyr Asp Gly Pro
1 5 10 15
Val Thr Leu Asp
20
<210> 388
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 388
Arg Asn Leu Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp
1 5 10 15
Trp Gly Asp Val
20
<210> 389
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 389
Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp Val Asp
1 5 10 15
Ser Thr Met Val
20
<210> 390
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 390
Pro Val Thr Leu Asp Asp Trp Gly Asp Val Asp Ser Thr Met Val Leu
1 5 10 15
Leu Tyr Thr Ser
20
<210> 391
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 391
Asp Trp Gly Asp Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val
1 5 10 15
Asp Thr Lys Lys
20
<210> 392
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 392
Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys Tyr
1 5 10 15
Lys Val Leu Leu
20
<210> 393
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 393
Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys Tyr Lys Val Leu Leu Thr
1 5 10 15
Tyr Asp Thr His
20
<210> 394
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 394
Val Asp Thr Lys Lys Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val
1 5 10 15
Asn Lys Thr Tyr
20
<210> 395
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 395
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
1 5 10 15
Val Asp Met Ser
20
<210> 396
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 396
Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro Val Asp Met Ser Pro
1 5 10 15
Thr Phe Thr Trp
20
<210> 397
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 397
Val Asn Lys Thr Tyr Pro Val Asp Met Ser Pro Thr Phe Thr Trp Lys
1 5 10 15
Asn Ser Lys Leu
20
<210> 398
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 398
Pro Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro
1 5 10 15
Asn Asp Ile Thr
20
<210> 399
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 399
Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn Asp Ile Thr Gly
1 5 10 15
Arg Gly Pro Gln
20
<210> 400
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 400
Lys Asn Ser Lys Leu Pro Asn Asp Ile Thr Gly Arg Gly Pro Gln Ile
1 5 10 15
Leu Met Ile Ala
20
<210> 401
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 401
Pro Asn Asp Ile Thr Gly Arg Gly Pro Gln Ile Leu Met Ile Ala Val
1 5 10 15
Phe Thr Leu Thr
20
<210> 402
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 402
Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val Leu Ile Gly
1 5 10 15
Pro Ser Cys Thr
20
<210> 403
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 403
Asn Ala Gln Arg Met Gly Cys Val Leu Ile Gly Pro Ser Cys Thr Tyr
1 5 10 15
Ser Thr Phe Gln
20
<210> 404
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 404
Gly Cys Val Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met
1 5 10 15
Tyr Leu Asp Thr
20
<210> 405
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 405
Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly Leu Asp Leu Leu
1 5 10 15
Arg Lys Ile Ser Asn Ala Gln Arg Met
20 25
<210> 406
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 406
Arg Ser Ser Thr Cys Glu Gly Leu Asp Leu Leu Arg Lys Ile Ser
1 5 10 15
<210> 407
<211> 254
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 407
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Thr Ser Ser Gln Ser Leu Phe Asn Val
20 25 30
Arg Ser Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asp Leu Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Ser Ser Tyr Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly Glu Ile Lys Pro Ser Asn
165 170 175
Glu Leu Thr Asn Val His Glu Lys Phe Lys Asp Arg Phe Thr Ile Ser
180 185 190
Val Asp Lys Ala Lys Asn Ser Ala Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Thr Arg Thr Ile Thr Thr Thr
210 215 220
Glu Gly Tyr Trp Phe Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr
225 230 235 240
Val Ser Ser Gly Gly Cys Gly Gly His His His His His His
245 250
<210> 408
<211> 252
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 408
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Ser Ser Leu Leu Gln Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Ala Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg
85 90 95
Lys Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Asp Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Phe Ile Arg Asn Gln Ala Arg Gly Tyr
165 170 175
Thr Ser Asp His Asn Pro Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Pro Ser Tyr Tyr
210 215 220
Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
225 230 235 240
Gly Cys Gly Gly Asp Tyr Lys Asp Asp Asp Asp Lys
245 250
<210> 409
<211> 257
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 409
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Glu Leu Thr Asn Val His Glu Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Asp Ile Gln Leu Thr Gln
130 135 140
Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr
145 150 155 160
Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr Gly Ser Ser Leu Leu Gln
165 170 175
Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala
180 185 190
Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly
195 200 205
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp
210 215 220
Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Arg Lys Ala Tyr Thr Phe Gly
225 230 235 240
Gln Gly Thr Lys Leu Glu Ile Lys Thr Gly Ser Glu Asn Leu Tyr Phe
245 250 255
Gln
<210> 410
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 410
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 411
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 411
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Arg Gln Lys Cys His Asn Gly Thr Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Asp Asn Ser Ala Tyr Lys Glu Pro Leu Gln Asn Leu
65 70 75 80
Arg Asp Ala Val Glu Glu Gly Leu Asp Ile Val Arg Lys Arg Leu Arg
85 90 95
Glu Ala Glu Leu Asn Val Thr Val Asn Ala Thr Phe Ile Tyr Ser Asp
100 105 110
Gly Leu Ile His Lys Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Glu Ile Thr Arg Asp Arg Lys Met Gly Cys Val
130 135 140
Leu Met Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Asn Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Ile Leu Pro Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asp Phe Trp Lys Val Asn Asn Ala Pro Phe
195 200 205
Lys Thr Phe Ser Trp Asn Ser Ser Tyr Val Tyr Lys Asn Gly Ser Glu
210 215 220
Pro Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Gly Val Ser
225 230 235 240
Tyr Phe Ser Glu Val Leu Ser Phe Lys Asp Val Leu Arg Arg Ser Glu
245 250 255
Gln Phe Gln Glu Ile Leu Met Gly Arg Asn Arg Lys Ser Asn Val Ile
260 265 270
Val Met Cys Gly Thr Pro Glu Thr Phe Tyr Asn Val Lys Gly Asp Leu
275 280 285
Lys Val Ala Asp Asp Thr Val Val Ile Leu Val Asp Leu Phe Ser Asn
290 295 300
His Tyr Phe Glu Asp Asp Thr Arg Ala Pro Glu Tyr Met Asp Asn Val
305 310 315 320
Leu Val Leu Thr Leu Pro Pro Glu Lys Phe Ile Ala Asn Ala Ser Val
325 330 335
Ser Gly Arg Phe Pro Ser Glu Arg Ser Asp Phe Ser Leu Ala Tyr Leu
340 345 350
Glu Gly Thr Leu Leu Phe Gly His Met Leu Gln Thr Phe Leu Glu Asn
355 360 365
Gly Glu Ser Val Thr Thr Pro Lys Phe Ala Arg Ala Phe Arg Asn Leu
370 375 380
Thr Phe Gln Gly Leu Glu Gly Pro Val Thr Leu Asp Asp Ser Gly Asp
385 390 395 400
Ile Asp Asn Ile Met Cys Leu Leu Tyr Val Ser Leu Asp Thr Arg Lys
405 410 415
Tyr Lys Val Leu Met Ala Tyr Asp Thr His Lys Asn Gln Thr Ile Pro
420 425 430
Val Ala Thr Ser Pro Asn Phe Ile Trp Lys Asn His Arg Leu Pro Asn
435 440 445
Asp Val Pro Gly Leu Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 412
<211> 1028
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 412
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Ser Arg Cys Asn Asn Asn Asn Tyr Met Ile Asn Val Met Leu
50 55 60
Met Pro Asp Ser Asp Phe Pro Ser Thr Ser Glu Asn Leu Thr Ser Ala
65 70 75 80
Val Glu Glu Ala Leu Ser Thr Ile Gln Asn Glu Leu Glu Thr Glu Gly
85 90 95
Val Lys Val Thr Val Asn Ala Ser Phe His His Phe Arg Ser Ser Leu
100 105 110
Tyr Val Ser Gln Gly Cys Arg Thr Ser Thr Cys Glu Gly Val Glu Leu
115 120 125
Ile Lys Gln Ile Phe Glu Asn Gly Thr Leu Gly Cys Ala Val Ile Gly
130 135 140
Pro Ala Cys Thr Tyr Ala Thr Tyr Gln Met Val Ser Val Glu Thr Ile
145 150 155 160
Pro Ser Leu Pro Leu Ile Ser Val Gly Ser Phe Gly Leu Ser Cys Asp
165 170 175
Tyr Lys Glu Asn Leu Thr Arg Leu Leu Thr Pro Ala Arg Lys Val Asn
180 185 190
Asp Phe Phe Tyr Tyr Phe Trp Asn Glu Ile Gln Gln Pro Phe Lys Thr
195 200 205
Ser Thr Trp Glu Ser Val Tyr Leu Tyr Lys Lys Thr Asp Asn Ser Glu
210 215 220
Gln Cys Leu Trp Tyr Met Asn Ala Leu Asp Ala Gly Val Thr Gln Phe
225 230 235 240
Ser Glu Lys Leu Lys Phe Lys Asp Ile Val Arg Thr Gln Asp Gln Phe
245 250 255
Arg Arg Leu Val Lys Asn Pro Lys Arg Lys Ser Asn Val Ile Ile Met
260 265 270
Cys Gly Thr Pro Ala Asp Ile Arg Gln Asp Leu Gly Thr Glu Thr Val
275 280 285
Asp Lys Asp Ile Val Ile Ile Leu Ile Asp Leu Phe Lys Asn Thr Tyr
290 295 300
Phe Arg Asn Thr Thr Ser Ala Arg Tyr Met Gln Asn Val Leu Val Leu
305 310 315 320
Thr Leu Pro Pro Ala Asn Asn Asn Phe Ser Thr Arg Thr Thr Asp Thr
325 330 335
Ser Leu Leu Glu Asp Asp Phe Val Ile Gly Tyr Tyr Asn Ala Val Leu
340 345 350
Leu Phe Gly His Ile Leu Lys Lys Phe Ile Phe Ser Gln Ser Pro Val
355 360 365
Leu Pro Thr Ser Phe Ile Asn Glu Phe Arg Asn Ile Thr Phe Glu Gly
370 375 380
Ala Gln Gly Pro Val Thr Leu Asp Glu Phe Gly Asp Ile Asp Asn Asn
385 390 395 400
Leu Thr Leu Leu Tyr Thr Thr Gln Ser Ala Ser Asp Pro Gln Tyr Arg
405 410 415
Val Leu Met Tyr Phe Asn Thr Gln Glu Asn Asp Thr Tyr Val Val Ser
420 425 430
Thr Ser Pro Asp Phe Ile Trp Lys Ser His Arg Leu Pro Ser Asp Ile
435 440 445
Pro Ser Thr Gly Pro His Arg Thr Ser Asp Gln Ala Ser Gly Ala His
450 455 460
His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp Tyr Ala
465 470 475 480
Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser Thr Pro
485 490 495
Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His Ser Thr
500 505 510
Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser His Ser
515 520 525
Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr Ser His
530 535 540
Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser Thr Ser
545 550 555 560
Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu Thr Ser
565 570 575
Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr Glu Thr
580 585 590
Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val Thr Glu
595 600 605
Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr Ser Ser
610 615 620
Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile Met Ser
625 630 635 640
Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val Tyr Thr
645 650 655
Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser Leu Ile
660 665 670
Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr Ser Ser
675 680 685
Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr Thr Pro
690 695 700
Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser Ser Thr
705 710 715 720
Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser Ser Val
725 730 735
Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr Pro Ser
740 745 750
Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser Thr Pro
755 760 765
Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His Ile Thr
770 775 780
Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser His Ser
785 790 795 800
Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr Ser Glu
805 810 815
Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val Ser Thr
820 825 830
Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr Ala Val
835 840 845
Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile Pro Thr
850 855 860
Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser Thr Ser
865 870 875 880
Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile Ser Thr
885 890 895
Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile Gln Ser
900 905 910
Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met Ser Thr
915 920 925
Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser Ser Phe
930 935 940
Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr Pro Pro
945 950 955 960
Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro Thr Thr
965 970 975
Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His Lys Leu
980 985 990
Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala Ala Asp
995 1000 1005
Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro Leu Val
1010 1015 1020
Leu Leu Ser Leu Leu
1025
<210> 413
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 413
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Asn
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Phe Asp
100 105 110
Gly Leu Leu His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Val Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 414
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 414
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asp Ser Asp Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Gly Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 415
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 415
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Val Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Asn Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 416
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 416
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Thr Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Ser Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 417
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 417
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Arg Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Ala Phe Leu Tyr Lys Leu Lys Gly Thr Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Arg Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 418
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 418
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Glu Phe Arg Asn Ile
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 419
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 419
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Thr Ser Thr Cys Glu Gly
115 120 125
Val Glu Leu Ile Lys Gln Ile Phe Glu Asn Gly Thr Leu Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 420
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 420
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Asn Glu Ile Gln Gln Pro Phe
195 200 205
Lys Thr Ser Thr Trp Glu Ser Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 421
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 421
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Lys Thr Asp Asn
210 215 220
Ser Glu Gln Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 422
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 422
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Lys Phe Lys Asp Ile Val Arg Thr Gln Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 423
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 423
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Asp
245 250 255
Gln Phe Arg Arg Leu Val Lys Asn Pro Lys Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 424
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 424
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Thr Glu
275 280 285
Thr Val Asp Lys Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 425
<211> 1029
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 425
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Arg Asn Thr Thr Ser Ala Arg Tyr Met Gln Asn Val Leu
305 310 315 320
Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe Ser
325 330 335
Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu Asn
340 345 350
Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn Gly
355 360 365
Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu Thr
370 375 380
Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp Val
385 390 395 400
Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys Tyr
405 410 415
Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro Val
420 425 430
Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn Asp
435 440 445
Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly Ala
450 455 460
His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp Tyr
465 470 475 480
Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser Thr
485 490 495
Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His Ser
500 505 510
Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser His
515 520 525
Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr Ser
530 535 540
His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser Thr
545 550 555 560
Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu Thr
565 570 575
Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr Glu
580 585 590
Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val Thr
595 600 605
Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr Ser
610 615 620
Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile Met
625 630 635 640
Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val Tyr
645 650 655
Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser Leu
660 665 670
Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr Ser
675 680 685
Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr Thr
690 695 700
Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser Ser
705 710 715 720
Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser Ser
725 730 735
Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr Pro
740 745 750
Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser Thr
755 760 765
Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His Ile
770 775 780
Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser His
785 790 795 800
Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr Ser
805 810 815
Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val Ser
820 825 830
Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr Ala
835 840 845
Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile Pro
850 855 860
Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser Thr
865 870 875 880
Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile Ser
885 890 895
Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile Gln
900 905 910
Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met Ser
915 920 925
Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser Ser
930 935 940
Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr Pro
945 950 955 960
Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro Thr
965 970 975
Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His Lys
980 985 990
Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala Ala
995 1000 1005
Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro Leu
1010 1015 1020
Val Leu Leu Ser Leu Leu
1025
<210> 426
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 426
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Ala Ser Asp Pro Gln
405 410 415
Tyr Arg Val Leu Met Tyr Phe Asn Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 427
<211> 1028
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 427
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Ser Arg Cys Asn Asn Asn Asn Tyr Met Ile Asn Val Met Leu
50 55 60
Met Pro Asp Ser Asp Phe Pro Ser Thr Ser Glu Asn Leu Thr Ser Ala
65 70 75 80
Val Glu Glu Ala Leu Ser Thr Ile Gln Asn Glu Leu Glu Thr Glu Gly
85 90 95
Val Lys Val Thr Val Asn Ala Ser Phe His His Phe Arg Ser Ser Leu
100 105 110
Tyr Val Ser Gln Gly Cys Arg Ser Ser Thr Cys Glu Gly Leu Asp Leu
115 120 125
Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Ala Val Ile Gly
130 135 140
Pro Ala Cys Thr Tyr Ala Thr Tyr Gln Met Val Ser Val Glu Thr Ile
145 150 155 160
Pro Ser Leu Pro Leu Ile Ser Val Gly Ser Phe Gly Leu Ser Cys Asp
165 170 175
Tyr Lys Glu Asn Leu Thr Arg Leu Leu Thr Pro Ala Arg Lys Val Asn
180 185 190
Asp Phe Phe Tyr Tyr Phe Trp Asn Glu Ile Gln Gln Pro Phe Lys Thr
195 200 205
Ser Thr Trp Glu Ser Val Tyr Leu Tyr Lys Lys Thr Asp Asn Ser Glu
210 215 220
Gln Cys Leu Trp Tyr Met Asn Ala Leu Asp Ala Gly Val Thr Gln Phe
225 230 235 240
Ser Glu Lys Leu Lys Phe Lys Asp Ile Val Arg Thr Gln Asp Gln Phe
245 250 255
Arg Arg Leu Val Lys Asn Pro Lys Arg Lys Ser Asn Val Ile Ile Met
260 265 270
Cys Gly Thr Pro Ala Asp Ile Arg Gln Asp Leu Gly Thr Glu Thr Val
275 280 285
Asp Lys Asp Ile Val Ile Ile Leu Ile Asp Leu Phe Lys Asn Thr Tyr
290 295 300
Phe Arg Asn Thr Thr Ser Ala Arg Tyr Met Gln Asn Val Leu Val Leu
305 310 315 320
Thr Leu Pro Pro Ala Asn Asn Asn Phe Ser Thr Arg Thr Thr Asp Thr
325 330 335
Ser Leu Leu Glu Asp Asp Phe Val Ile Gly Tyr Tyr Asn Ala Val Leu
340 345 350
Leu Phe Gly His Ile Leu Lys Lys Phe Ile Phe Ser Gln Ser Pro Val
355 360 365
Leu Pro Thr Ser Phe Ile Asn Glu Phe Arg Asn Ile Thr Phe Glu Gly
370 375 380
Ala Gln Gly Pro Val Thr Leu Asp Glu Phe Gly Asp Ile Asp Asn Asn
385 390 395 400
Leu Thr Leu Leu Tyr Thr Thr Gln Ser Ala Ser Asp Pro Gln Tyr Arg
405 410 415
Val Leu Met Tyr Phe Asn Thr Gln Glu Asn Asp Thr Tyr Val Val Ser
420 425 430
Thr Ser Pro Asp Phe Ile Trp Lys Ser His Arg Leu Pro Ser Asp Ile
435 440 445
Pro Ser Thr Gly Pro His Arg Thr Ser Asp Gln Ala Ser Gly Ala His
450 455 460
His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp Tyr Ala
465 470 475 480
Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser Thr Pro
485 490 495
Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His Ser Thr
500 505 510
Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser His Ser
515 520 525
Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr Ser His
530 535 540
Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser Thr Ser
545 550 555 560
Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu Thr Ser
565 570 575
Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr Glu Thr
580 585 590
Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val Thr Glu
595 600 605
Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr Ser Ser
610 615 620
Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile Met Ser
625 630 635 640
Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val Tyr Thr
645 650 655
Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser Leu Ile
660 665 670
Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr Ser Ser
675 680 685
Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr Thr Pro
690 695 700
Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser Ser Thr
705 710 715 720
Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser Ser Val
725 730 735
Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr Pro Ser
740 745 750
Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser Thr Pro
755 760 765
Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His Ile Thr
770 775 780
Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser His Ser
785 790 795 800
Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr Ser Glu
805 810 815
Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val Ser Thr
820 825 830
Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr Ala Val
835 840 845
Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile Pro Thr
850 855 860
Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser Thr Ser
865 870 875 880
Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile Ser Thr
885 890 895
Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile Gln Ser
900 905 910
Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met Ser Thr
915 920 925
Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser Ser Phe
930 935 940
Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr Pro Pro
945 950 955 960
Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro Thr Thr
965 970 975
Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His Lys Leu
980 985 990
Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala Ala Asp
995 1000 1005
Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro Leu Val
1010 1015 1020
Leu Leu Ser Leu Leu
1025
<210> 428
<211> 1028
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 428
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Ser Arg Cys Asn Asn Asn Asn Tyr Met Ile Asn Val Met Leu
50 55 60
Met Pro Asp Ser Asp Phe Pro Ser Thr Ser Glu Asn Leu Thr Ser Ala
65 70 75 80
Val Glu Glu Ala Leu Ser Thr Ile Gln Asn Glu Leu Glu Thr Glu Gly
85 90 95
Val Lys Val Thr Val Asn Ala Ser Phe His His Phe Arg Ser Ser Leu
100 105 110
Tyr Val Ser Gln Gly Cys Arg Ser Ser Thr Cys Glu Gly Leu Asp Leu
115 120 125
Leu Arg Lys Ile Ser Glu Asn Gly Thr Leu Gly Cys Ala Val Ile Gly
130 135 140
Pro Ala Cys Thr Tyr Ala Thr Tyr Gln Met Val Ser Val Glu Thr Ile
145 150 155 160
Pro Ser Leu Pro Leu Ile Ser Val Gly Ser Phe Gly Leu Ser Cys Asp
165 170 175
Tyr Lys Glu Asn Leu Thr Arg Leu Leu Thr Pro Ala Arg Lys Val Asn
180 185 190
Asp Phe Phe Tyr Tyr Phe Trp Asn Glu Ile Gln Gln Pro Phe Lys Thr
195 200 205
Ser Thr Trp Glu Ser Val Tyr Leu Tyr Lys Lys Thr Asp Asn Ser Glu
210 215 220
Gln Cys Leu Trp Tyr Met Asn Ala Leu Asp Ala Gly Val Thr Gln Phe
225 230 235 240
Ser Glu Lys Leu Lys Phe Lys Asp Ile Val Arg Thr Gln Asp Gln Phe
245 250 255
Arg Arg Leu Val Lys Asn Pro Lys Arg Lys Ser Asn Val Ile Ile Met
260 265 270
Cys Gly Thr Pro Ala Asp Ile Arg Gln Asp Leu Gly Thr Glu Thr Val
275 280 285
Asp Lys Asp Ile Val Ile Ile Leu Ile Asp Leu Phe Lys Asn Thr Tyr
290 295 300
Phe Arg Asn Thr Thr Ser Ala Arg Tyr Met Gln Asn Val Leu Val Leu
305 310 315 320
Thr Leu Pro Pro Ala Asn Asn Asn Phe Ser Thr Arg Thr Thr Asp Thr
325 330 335
Ser Leu Leu Glu Asp Asp Phe Val Ile Gly Tyr Tyr Asn Ala Val Leu
340 345 350
Leu Phe Gly His Ile Leu Lys Lys Phe Ile Phe Ser Gln Ser Pro Val
355 360 365
Leu Pro Thr Ser Phe Ile Asn Glu Phe Arg Asn Ile Thr Phe Glu Gly
370 375 380
Ala Gln Gly Pro Val Thr Leu Asp Glu Phe Gly Asp Ile Asp Asn Asn
385 390 395 400
Leu Thr Leu Leu Tyr Thr Thr Gln Ser Ala Ser Asp Pro Gln Tyr Arg
405 410 415
Val Leu Met Tyr Phe Asn Thr Gln Glu Asn Asp Thr Tyr Val Val Ser
420 425 430
Thr Ser Pro Asp Phe Ile Trp Lys Ser His Arg Leu Pro Ser Asp Ile
435 440 445
Pro Ser Thr Gly Pro His Arg Thr Ser Asp Gln Ala Ser Gly Ala His
450 455 460
His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp Tyr Ala
465 470 475 480
Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser Thr Pro
485 490 495
Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His Ser Thr
500 505 510
Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser His Ser
515 520 525
Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr Ser His
530 535 540
Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser Thr Ser
545 550 555 560
Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu Thr Ser
565 570 575
Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr Glu Thr
580 585 590
Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val Thr Glu
595 600 605
Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr Ser Ser
610 615 620
Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile Met Ser
625 630 635 640
Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val Tyr Thr
645 650 655
Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser Leu Ile
660 665 670
Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr Ser Ser
675 680 685
Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr Thr Pro
690 695 700
Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser Ser Thr
705 710 715 720
Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser Ser Val
725 730 735
Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr Pro Ser
740 745 750
Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser Thr Pro
755 760 765
Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His Ile Thr
770 775 780
Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser His Ser
785 790 795 800
Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr Ser Glu
805 810 815
Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val Ser Thr
820 825 830
Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr Ala Val
835 840 845
Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile Pro Thr
850 855 860
Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser Thr Ser
865 870 875 880
Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile Ser Thr
885 890 895
Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile Gln Ser
900 905 910
Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met Ser Thr
915 920 925
Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser Ser Phe
930 935 940
Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr Pro Pro
945 950 955 960
Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro Thr Thr
965 970 975
Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His Lys Leu
980 985 990
Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala Ala Asp
995 1000 1005
Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro Leu Val
1010 1015 1020
Leu Leu Ser Leu Leu
1025
<210> 429
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 429
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Asn
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 430
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 430
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Phe Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 431
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 431
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Leu His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 432
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 432
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Thr Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 433
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 433
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Val Asp Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 434
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 434
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Glu Leu Leu Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 435
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 435
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Ile Arg Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 436
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 436
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Lys Lys Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 437
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 437
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Gln Ile Ser Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 438
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 438
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Ser Gln Val Ser Gln Asn Cys His Asn Gly Ser Tyr Glu Ile Ser
50 55 60
Val Leu Met Met Gly Asn Ser Ala Phe Ala Glu Pro Leu Lys Asn Leu
65 70 75 80
Glu Asp Ala Val Asn Glu Gly Leu Glu Ile Val Arg Gly Arg Leu Gln
85 90 95
Asn Ala Gly Leu Asn Val Thr Val Asn Ala Thr Phe Met Tyr Ser Asp
100 105 110
Gly Leu Ile His Asn Ser Gly Asp Cys Arg Ser Ser Thr Cys Glu Gly
115 120 125
Leu Asp Leu Leu Arg Lys Ile Phe Asn Ala Gln Arg Met Gly Cys Val
130 135 140
Leu Ile Gly Pro Ser Cys Thr Tyr Ser Thr Phe Gln Met Tyr Leu Asp
145 150 155 160
Thr Glu Leu Ser Tyr Pro Met Ile Ser Ala Gly Ser Phe Gly Leu Ser
165 170 175
Cys Asp Tyr Lys Glu Thr Leu Thr Arg Leu Met Ser Pro Ala Arg Lys
180 185 190
Leu Met Tyr Phe Leu Val Asn Phe Trp Lys Thr Asn Asp Leu Pro Phe
195 200 205
Lys Thr Tyr Ser Trp Ser Thr Ser Tyr Val Tyr Lys Asn Gly Thr Glu
210 215 220
Thr Glu Asp Cys Phe Trp Tyr Leu Asn Ala Leu Glu Ala Ser Val Ser
225 230 235 240
Tyr Phe Ser His Glu Leu Gly Phe Lys Val Val Leu Arg Gln Asp Lys
245 250 255
Glu Phe Gln Asp Ile Leu Met Asp His Asn Arg Lys Ser Asn Val Ile
260 265 270
Ile Met Cys Gly Gly Pro Glu Phe Leu Tyr Lys Leu Lys Gly Asp Arg
275 280 285
Ala Val Ala Glu Asp Ile Val Ile Ile Leu Val Asp Leu Phe Asn Asp
290 295 300
Gln Tyr Phe Glu Asp Asn Val Thr Ala Pro Asp Tyr Met Lys Asn Val
305 310 315 320
Leu Val Leu Thr Leu Ser Pro Gly Asn Ser Leu Leu Asn Ser Ser Phe
325 330 335
Ser Arg Asn Leu Ser Pro Thr Lys Arg Asp Phe Ala Leu Ala Tyr Leu
340 345 350
Asn Gly Ile Leu Leu Phe Gly His Met Leu Lys Ile Phe Leu Glu Asn
355 360 365
Gly Glu Asn Ile Thr Thr Pro Lys Phe Ala His Ala Phe Arg Asn Leu
370 375 380
Thr Phe Glu Gly Tyr Asp Gly Pro Val Thr Leu Asp Asp Trp Gly Asp
385 390 395 400
Val Asp Ser Thr Met Val Leu Leu Tyr Thr Ser Val Asp Thr Lys Lys
405 410 415
Tyr Lys Val Leu Leu Thr Tyr Asp Thr His Val Asn Lys Thr Tyr Pro
420 425 430
Val Asp Met Ser Pro Thr Phe Thr Trp Lys Asn Ser Lys Leu Pro Asn
435 440 445
Asp Ile Thr Gly Arg Gly Pro Gln Arg Thr Ser Asp Gln Ala Ser Gly
450 455 460
Ala His His His His His His Gly Ala Tyr Pro Tyr Asp Val Pro Asp
465 470 475 480
Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly Gly Thr Ser
485 490 495
Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr Pro Ser His
500 505 510
Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu Thr Pro Ser
515 520 525
His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser Glu Thr Thr
530 535 540
Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr Thr Glu Ser
545 550 555 560
Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr Ile Thr Glu
565 570 575
Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile Ser Ser Thr
580 585 590
Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser Ile Thr Val
595 600 605
Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser Thr Leu Thr
610 615 620
Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr Ser Ser Ile
625 630 635 640
Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu Thr Ser Val
645 650 655
Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr Ser Thr Ser
660 665 670
Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro Ser Phe Thr
675 680 685
Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala Ser Ser Tyr
690 695 700
Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn Thr Thr Ser
705 710 715 720
Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser Ser Ser Ser
725 730 735
Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser Tyr Ser Thr
740 745 750
Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro Leu His Ser
755 760 765
Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr Thr Ser His
770 775 780
Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu Thr Tyr Ser
785 790 795 800
His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr Glu Ser Thr
805 810 815
Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr Ser Thr Val
820 825 830
Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr Ser Glu Thr
835 840 845
Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser Thr Asp Ile
850 855 860
Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val Gly Thr Ser
865 870 875 880
Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro Thr Asp Ile
885 890 895
Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser Pro Ser Ile
900 905 910
Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser Pro Thr Met
915 920 925
Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr Pro Ile Ser
930 935 940
Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro Thr Gln Thr
945 950 955 960
Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser Pro Ala Pro
965 970 975
Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser Thr Leu His
980 985 990
Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr Ile Gly Ala
995 1000 1005
Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu Ala Pro
1010 1015 1020
Leu Val Leu Leu Ser Leu Leu
1025 1030
<210> 439
<211> 617
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 439
Met Gln Leu Leu Arg Cys Phe Ser Ile Phe Ser Val Ile Ala Ser Val
1 5 10 15
Leu Ala Gln Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp
20 25 30
Ser Thr Gly Gly Gly Ser Gly Gly Gly Ala Gln Ala Ala Gly Ala His
35 40 45
Ser Arg Thr Ser Asp Gln Ala Ser Gly Ala Gly Ala Tyr Pro Tyr Asp
50 55 60
Val Pro Asp Tyr Ala Gly His Gly Thr Ser Gly Gly Gly Gly Ser Gly
65 70 75 80
Gly Thr Ser Thr Pro Ser Tyr Thr Thr Ser Ile Ile Ser Thr Glu Thr
85 90 95
Pro Ser His Ser Thr Pro Ser Ser Ser Thr Ser Ile Thr Thr Thr Glu
100 105 110
Thr Pro Ser His Ser Thr Pro Ser Tyr Thr Ser Ser Val Ser Thr Ser
115 120 125
Glu Thr Thr Ser His Ser Thr Pro Ser Glu Thr Ser Ser Ser Arg Thr
130 135 140
Thr Glu Ser Thr Ser Tyr Ser Ser Pro Ser Ser Thr Ser Ser Asn Thr
145 150 155 160
Ile Thr Glu Thr Ser Ser His Ser Thr Pro Ser Thr Ala Thr Ser Ile
165 170 175
Ser Ser Thr Glu Thr Pro Ser Ser Ser Ile Pro Ser Val Ser Ser Ser
180 185 190
Ile Thr Val Thr Glu Ser Ser Ser His Ser Thr Pro Gly Ala Thr Ser
195 200 205
Thr Leu Thr Ser Ser Glu Thr Ser Thr Trp Ser Thr Pro Ser Ser Thr
210 215 220
Ser Ser Ile Met Ser Ser Ser Tyr Thr Ser Ala Asp Thr Pro Ser Glu
225 230 235 240
Thr Ser Val Tyr Thr Ser Ser Glu Thr Pro Ser Ser Ser Ser Pro Thr
245 250 255
Ser Thr Ser Leu Ile Ser Ser Ser Lys Ser Thr Ser Thr Ser Thr Pro
260 265 270
Ser Phe Thr Ser Ser Ile Thr Ser Thr Glu Thr Ser Ser Tyr Ser Ala
275 280 285
Ser Ser Tyr Thr Pro Ser Val Ser Ser Thr Ala Ser Ser Ser Lys Asn
290 295 300
Thr Thr Ser Ser Thr Ala Ser Ile Ser Ser Ala Glu Thr Val Ser Ser
305 310 315 320
Ser Ser Ser Ser Val Ser Ser Thr Ile Pro Ser Ser Gln Ser Thr Ser
325 330 335
Tyr Ser Thr Pro Ser Phe Ser Ser Ser Ala Thr Ser Ser Val Thr Pro
340 345 350
Leu His Ser Thr Pro Ser Leu Pro Ser Trp Val Thr Thr Ser Lys Thr
355 360 365
Thr Ser His Ile Thr Pro Gly Leu Thr Ser Ser Met Ser Ser Ser Glu
370 375 380
Thr Tyr Ser His Ser Thr Pro Gly Phe Thr Ser Ser Ile Thr Ser Thr
385 390 395 400
Glu Ser Thr Ser Glu Ser Thr Pro Ser Leu Ser Ser Ser Thr Ile Tyr
405 410 415
Ser Thr Val Ser Thr Ser Thr Thr Ala Ile Thr Ser His Phe Thr Thr
420 425 430
Ser Glu Thr Ala Val Thr Pro Thr Pro Val Thr Pro Ser Ser Leu Ser
435 440 445
Thr Asp Ile Pro Thr Thr Ser Leu Arg Thr Leu Thr Pro Ser Ser Val
450 455 460
Gly Thr Ser Thr Ser Leu Thr Thr Thr Thr Asp Phe Pro Ser Ile Pro
465 470 475 480
Thr Asp Ile Ser Thr Leu Pro Thr Arg Thr His Ile Ile Ser Ser Ser
485 490 495
Pro Ser Ile Gln Ser Thr Glu Thr Ser Ser Leu Val Gly Thr Thr Ser
500 505 510
Pro Thr Met Ser Thr Val Arg Met Thr Leu Arg Ile Thr Glu Asn Thr
515 520 525
Pro Ile Ser Ser Phe Ser Thr Ser Ile Val Val Ile Pro Glu Thr Pro
530 535 540
Thr Gln Thr Pro Pro Val Leu Thr Ser Ala Thr Gly Thr Gln Thr Ser
545 550 555 560
Pro Ala Pro Thr Thr Val Thr Phe Gly Ser Thr Asp Ser Ser Thr Ser
565 570 575
Thr Leu His Lys Leu Gln Gly Asp Ser Gly Glu Thr Ile Gly Lys Tyr
580 585 590
Ile Gly Ala Ala Asp Ser Leu Gly Gly Ser Val Leu Leu Leu Ala Leu
595 600 605
Ala Pro Leu Val Leu Leu Ser Leu Leu
610 615
<210> 440
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 440
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Asp Asp Ile Ala Met Tyr Phe Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Gly Asp
100 105 110
Pro Val Ala Pro Thr Val Leu Ile Phe Pro Pro Ala Ala Asp Gln Val
115 120 125
Ala Thr Gly Thr Val Thr Ile Val Cys Val Ala Asn Lys Tyr Phe Pro
130 135 140
Asp Val Thr Val Thr Trp Glu Val Asp Gly Thr Thr Gln Thr Thr Gly
145 150 155 160
Ile Glu Asn Ser Lys Thr Pro Gln Asn Ser Ala Asp Cys Thr Tyr Asn
165 170 175
Leu Ser Ser Thr Leu Thr Leu Thr Ser Thr Gln Tyr Asn Ser His Lys
180 185 190
Glu Tyr Thr Cys Arg Val Thr Gln Gly Thr Thr Ser Val Val Gln Ser
195 200 205
Phe Asn Arg Gly Asp Cys
210
<210> 441
<211> 445
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 441
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Ile Glu Lys Phe
50 55 60
Lys Asn Lys Ala Thr Leu Thr Val Asp Lys Ser Ala Thr Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ala Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser Gly Gln Pro Lys Ala Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Cys Gly Asp Thr Pro Ser Ser Thr
130 135 140
Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Leu Pro Glu Pro Val Thr
145 150 155 160
Val Thr Trp Asn Ser Gly Thr Leu Thr Asn Gly Val Arg Thr Phe Pro
165 170 175
Ser Val Arg Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Ser
180 185 190
Val Thr Ser Ser Ser Gln Pro Val Thr Cys Asn Val Ala His Pro Ala
195 200 205
Thr Asn Thr Lys Val Asp Lys Thr Val Ala Pro Ser Thr Cys Ser Lys
210 215 220
Pro Thr Cys Pro Pro Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Ile
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Asp Asp Pro Glu Val Gln
260 265 270
Phe Thr Trp Tyr Ile Asn Asn Glu Gln Val Arg Thr Ala Arg Pro Pro
275 280 285
Leu Arg Glu Gln Gln Phe Asn Ser Thr Ile Arg Val Val Ser Thr Leu
290 295 300
Pro Ile Ala His Gln Asp Trp Leu Arg Gly Lys Glu Phe Lys Cys Lys
305 310 315 320
Val His Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Arg Gly Gln Pro Leu Glu Pro Lys Val Tyr Thr Met Gly Pro Pro
340 345 350
Arg Glu Glu Leu Ser Ser Arg Ser Val Ser Leu Thr Cys Met Ile Asn
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Lys Asn Gly Lys
370 375 380
Ala Glu Asp Asn Tyr Lys Thr Thr Pro Ala Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Pro Thr Ser Glu Trp Gln
405 410 415
Arg Gly Asp Val Phe Thr Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Ile Ser Arg Ser Pro Gly Lys
435 440 445
<210> 442
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 442
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Asp Asp Ile Ala Met Tyr Phe Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr
100 105 110
Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu
115 120 125
Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro
130 135 140
Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn
145 150 155 160
Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His
180 185 190
Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile
195 200 205
Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 443
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 443
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Ile Glu Lys Phe
50 55 60
Lys Asn Lys Ala Thr Leu Thr Val Asp Lys Ser Ala Thr Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ala Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro
115 120 125
Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met
130 135 140
Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val
180 185 190
Pro Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His
195 200 205
Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys
210 215 220
Gly Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro
245 250 255
Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val
260 265 270
Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr
275 280 285
Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu
290 295 300
Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys
305 310 315 320
Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro
340 345 350
Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile
355 360 365
Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly
370 375 380
Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp
385 390 395 400
Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp
405 410 415
Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His
420 425 430
Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 444
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 444
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Tyr Tyr
20 25 30
Gly Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Asp Asp Ile Ala Met Tyr Phe Cys Gln Gln Thr Arg
85 90 95
Lys Val Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 445
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая пептидная последовательность
<400> 445
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Lys Pro Ser Asn Gly Leu Thr Asn Tyr Ile Glu Lys Phe
50 55 60
Lys Asn Lys Ala Thr Leu Thr Val Asp Lys Ser Ala Thr Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ala Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Thr Ile Thr Thr Thr Glu Gly Tyr Trp Phe Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО К CD3 И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2802272C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2839153C2 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ПОЛИПЕПТИД, ВСТРОЕННЫЙ В УЧАСТОК КАРКАСНОЙ ОБЛАСТИ 3 | 2019 |
|
RU2796254C2 |
| СКОНСТРУИРОВАННЫЕ PH-ЗАВИСИМЫЕ АНТИТЕЛА К CD3, А ТАКЖЕ СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2020 |
|
RU2832079C2 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| ВАРИАНТНЫЕ СВЯЗЫВАЮЩИЕ CD3 ДОМЕНЫ И ИХ ПРИМЕНЕНИЕ В КОМБИНИРОВАННОЙ ТЕРАПИИ ПРИ ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2810222C2 |
| УЛУЧШЕННЫЕ АНТИ-FLT3 АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2019 |
|
RU2830982C2 |
| ТЕХНОЛОГИИ МУЛЬТИМЕРИЗАЦИИ | 2014 |
|
RU2714733C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2021 |
|
RU2840407C1 |
| CD3-СВЯЗЫВАЮЩИЙ ДОМЕН | 2015 |
|
RU2742691C2 |
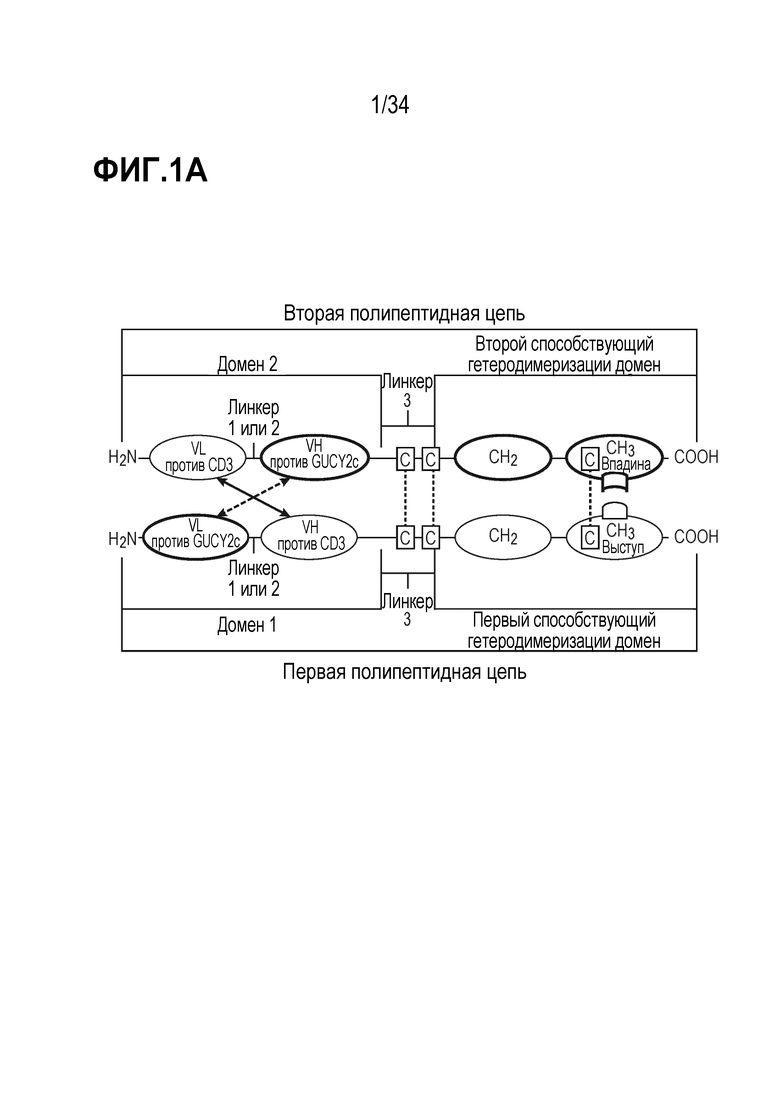
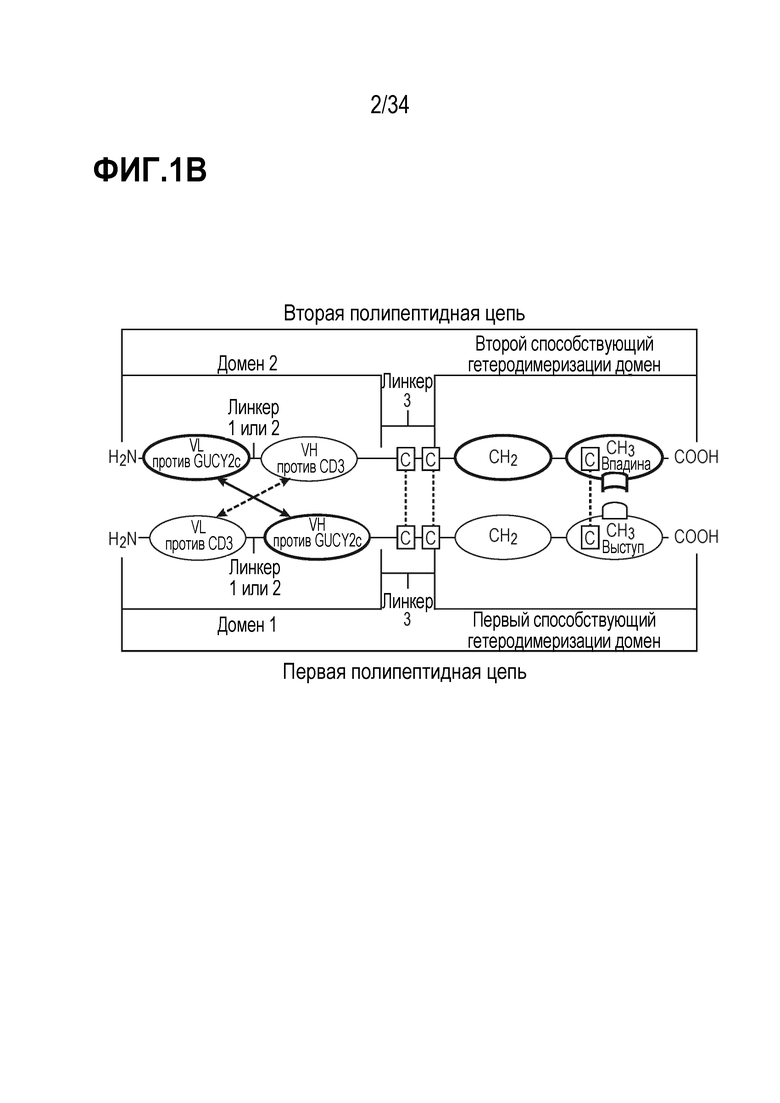
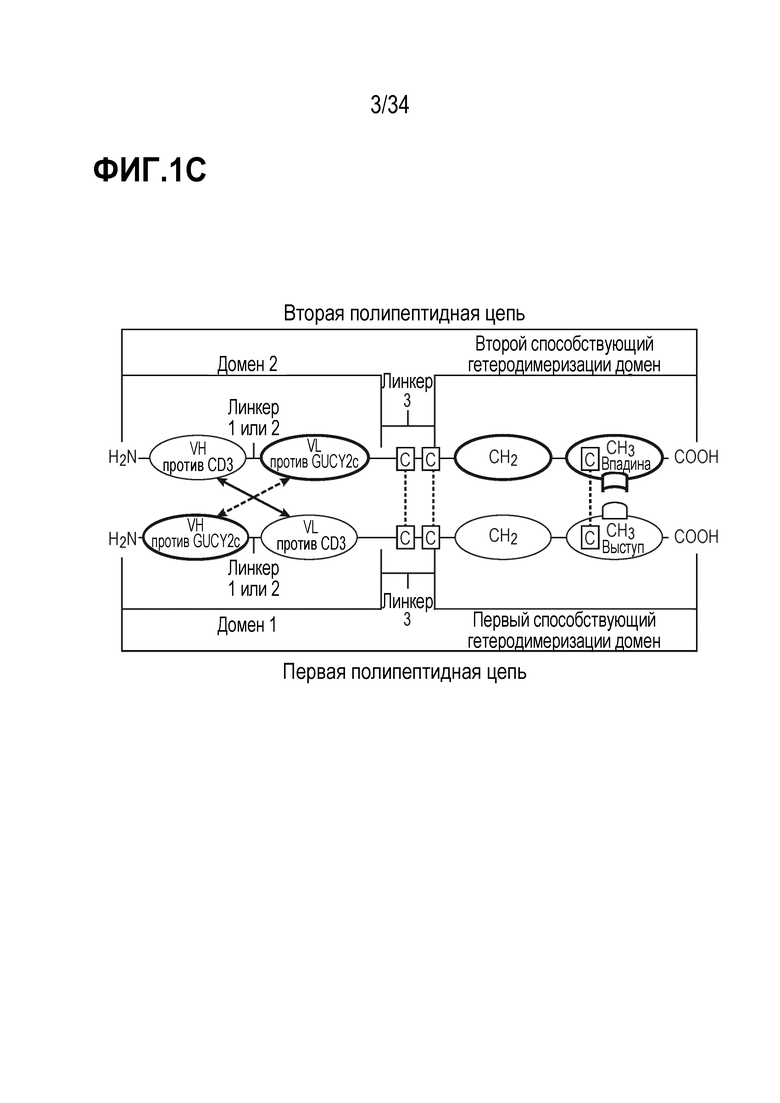
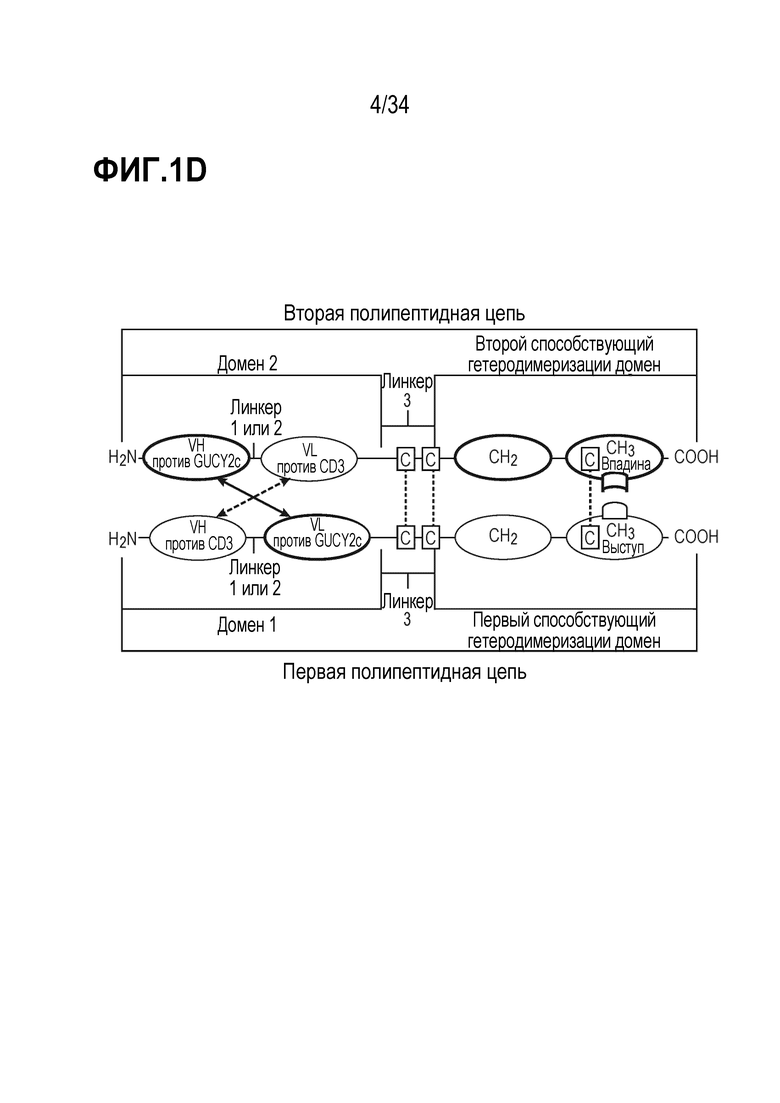
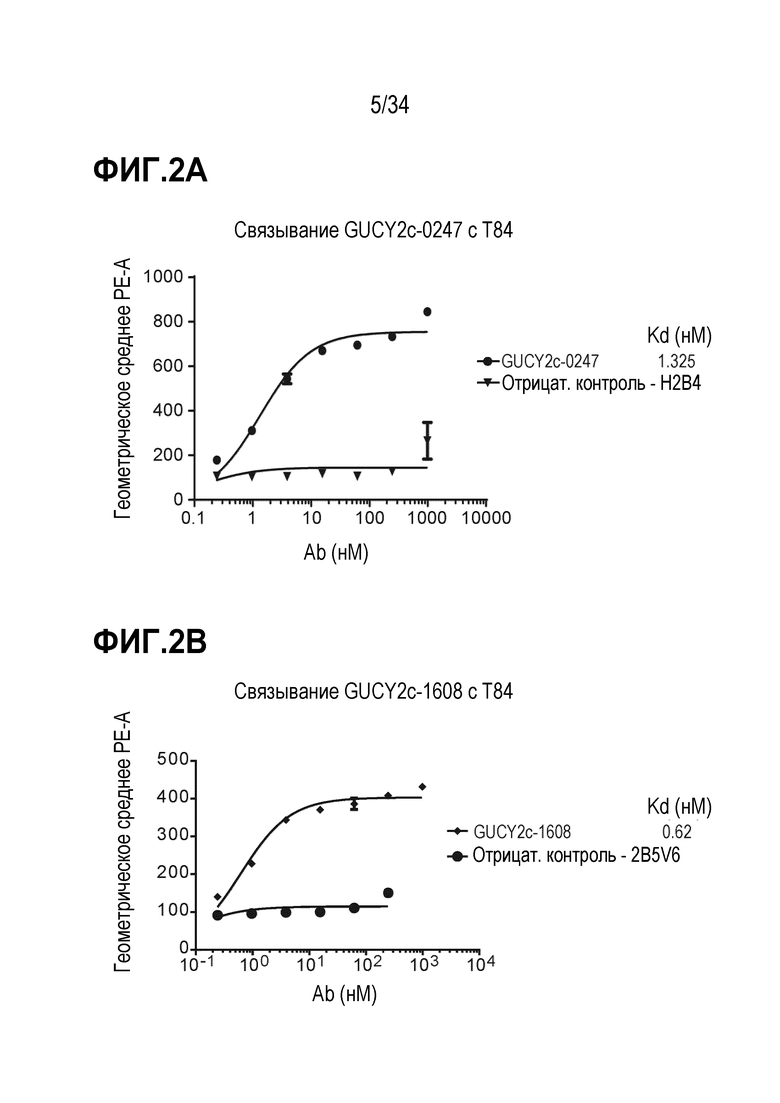
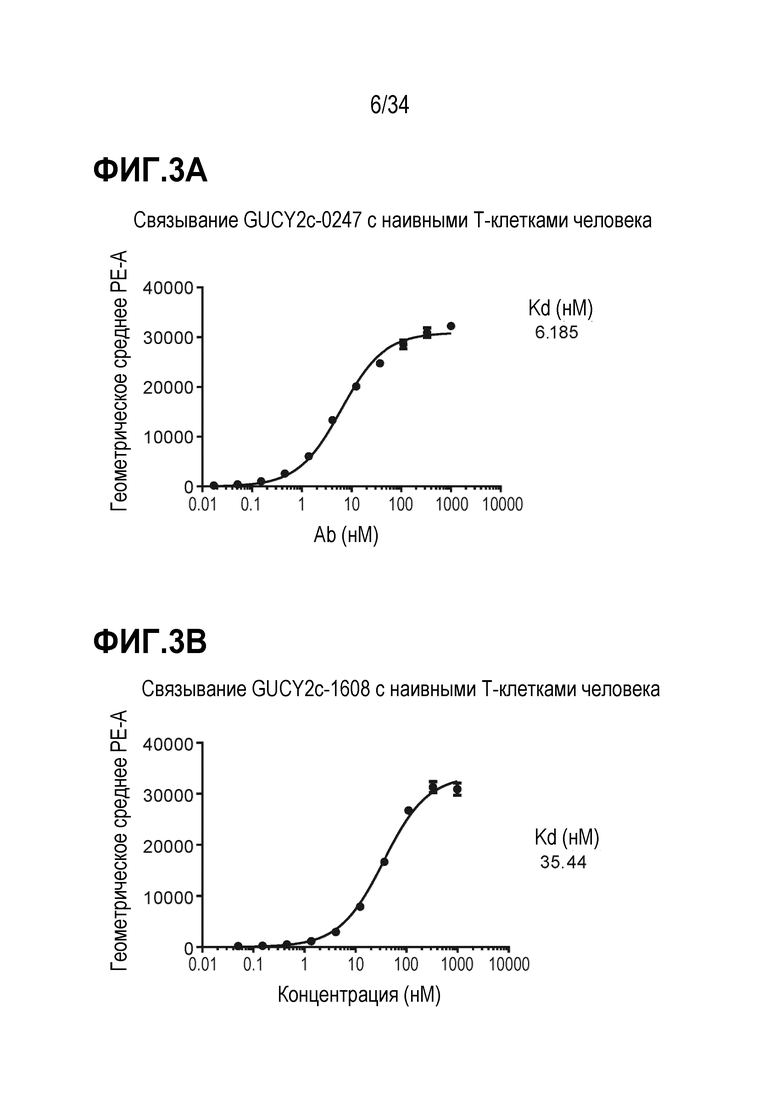
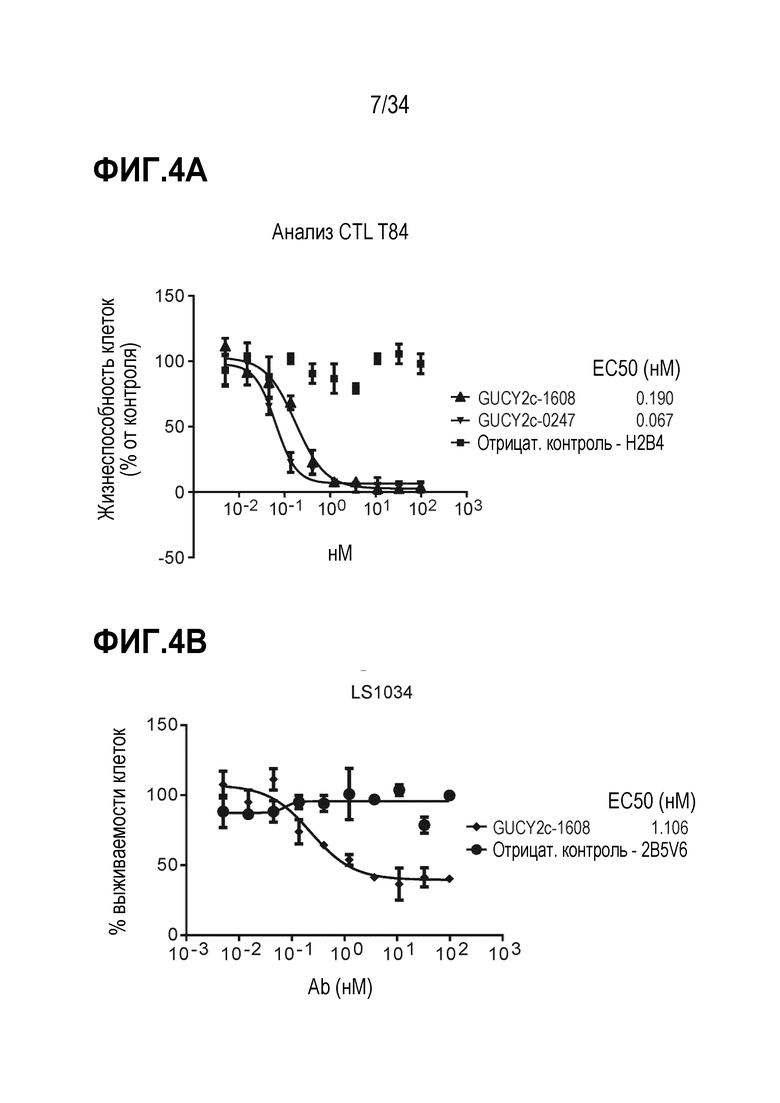
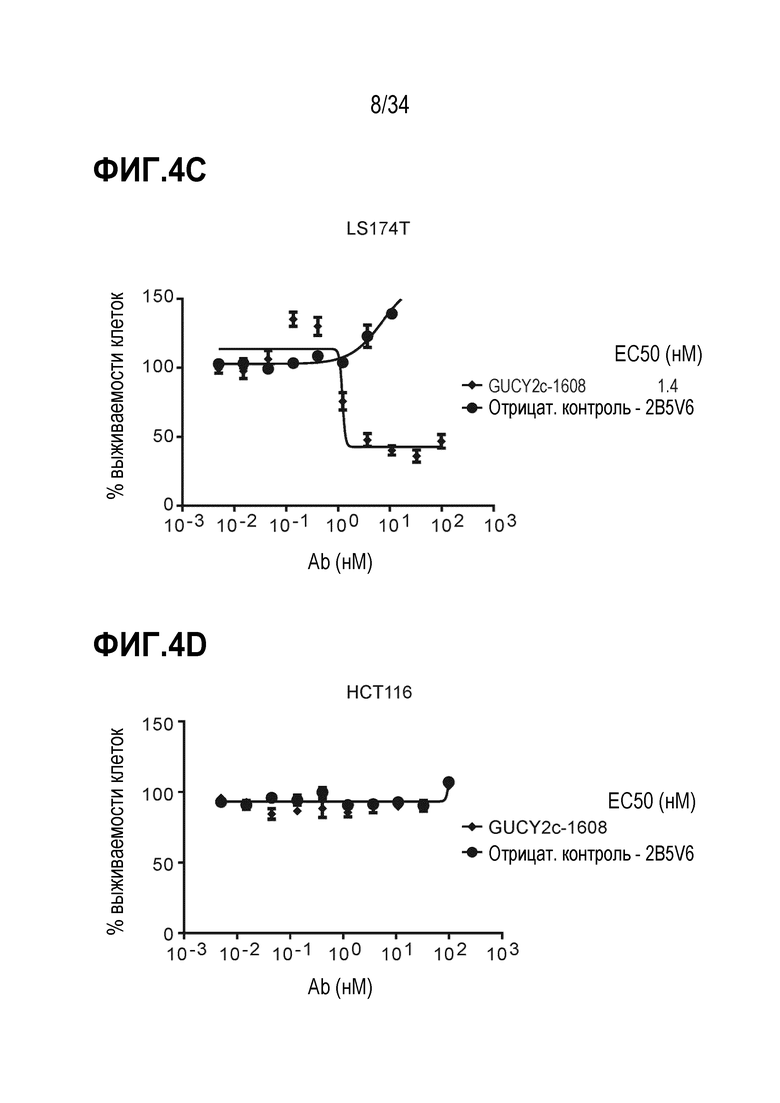
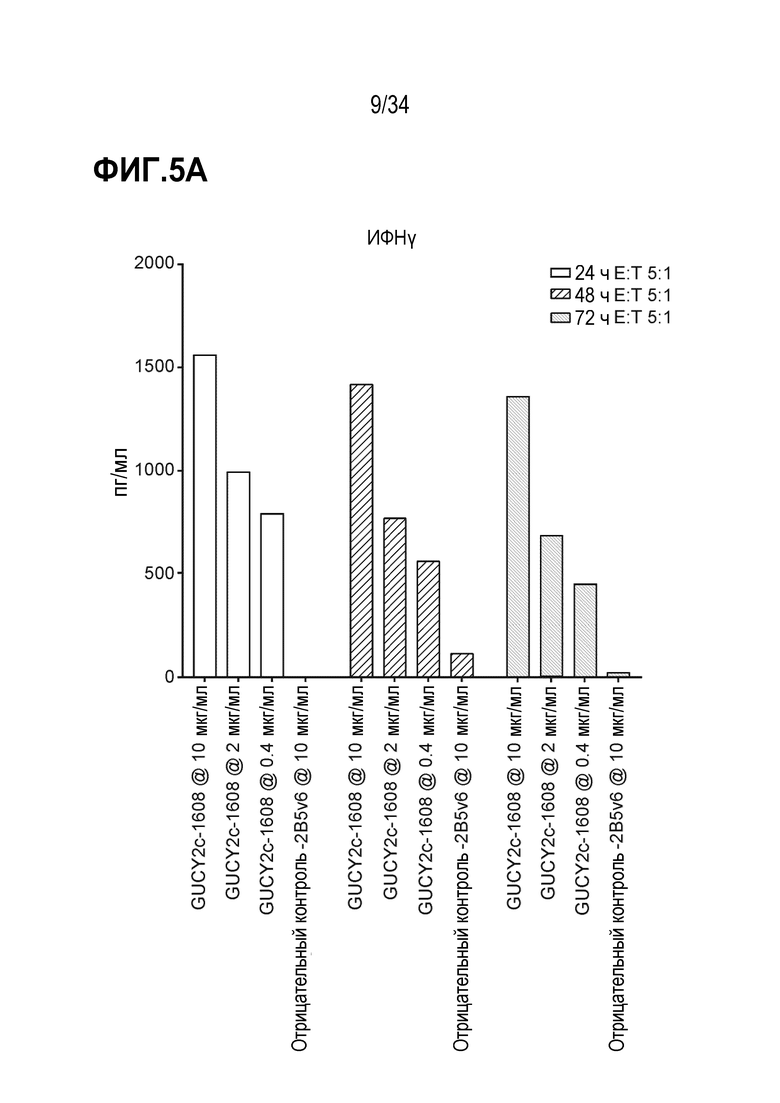
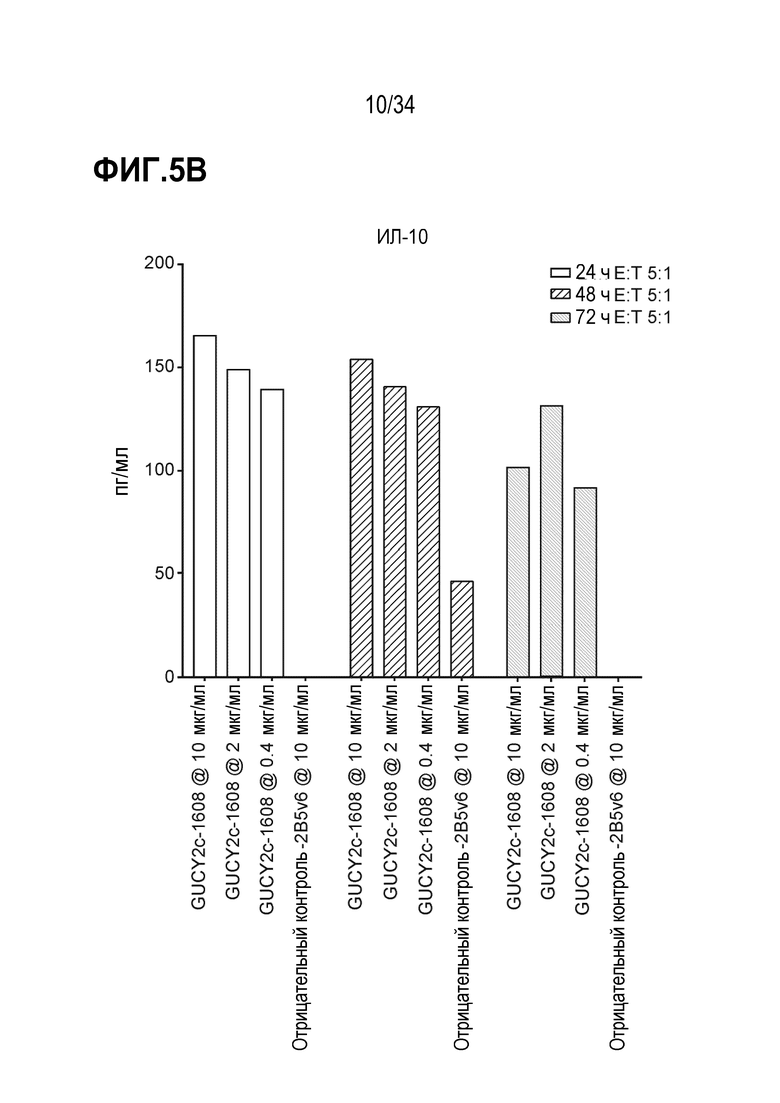
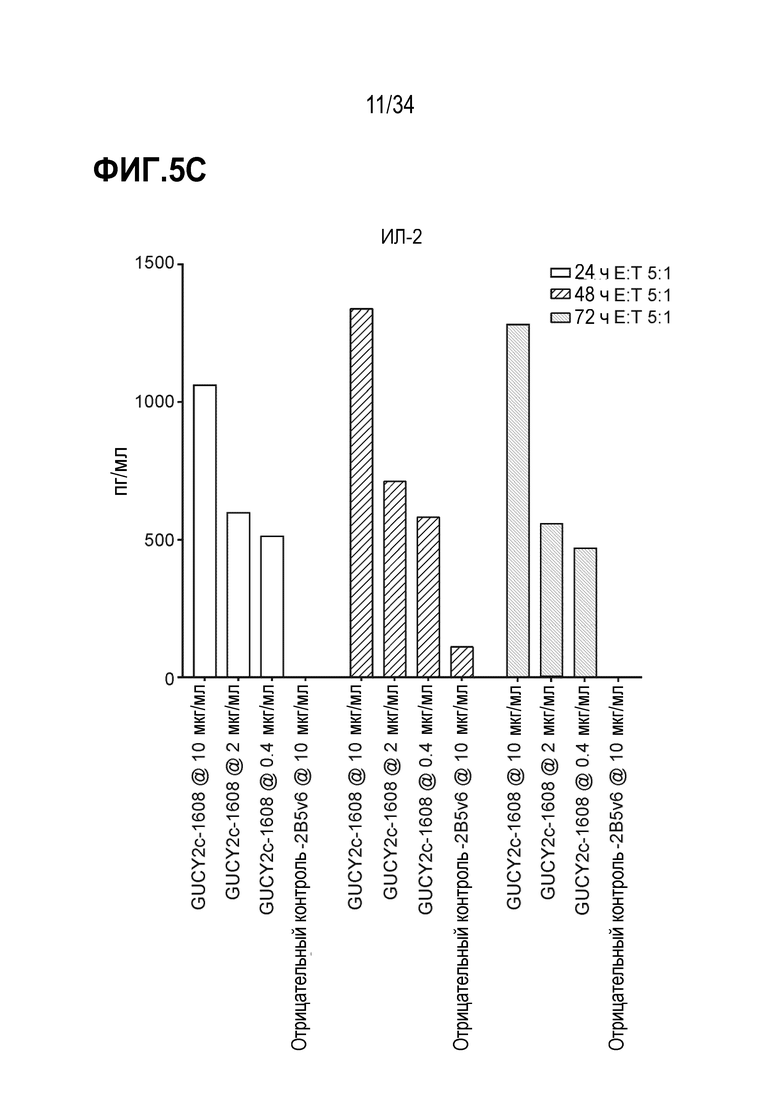
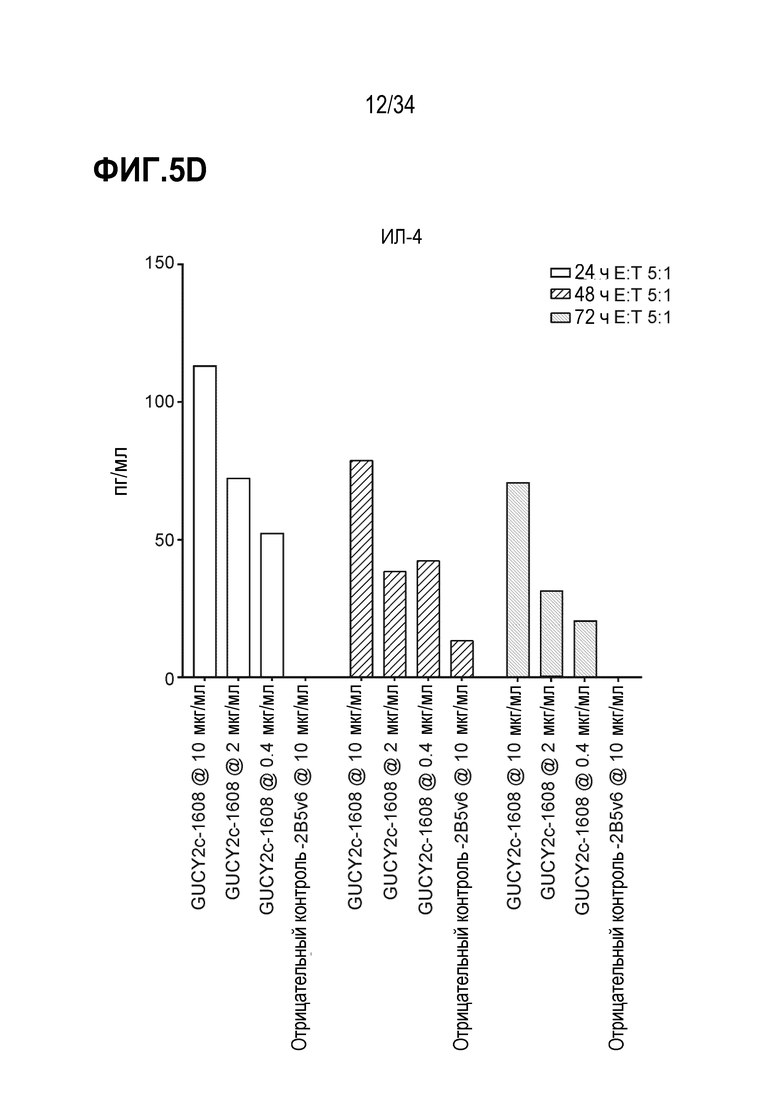
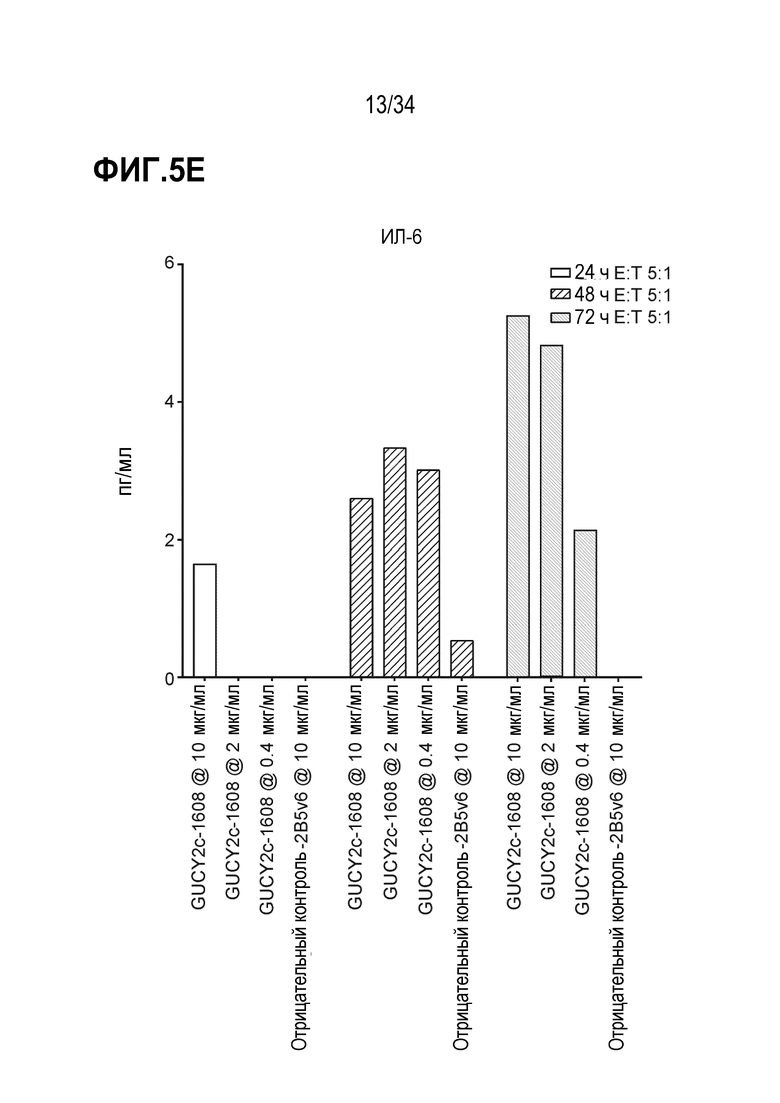
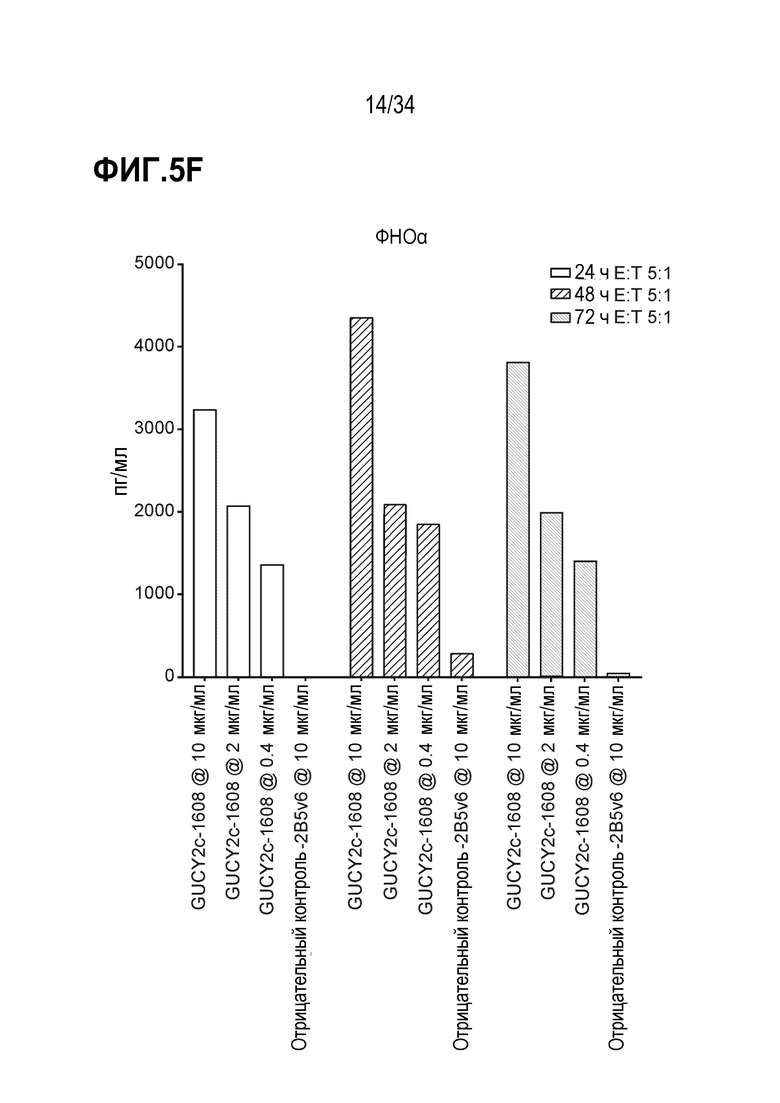
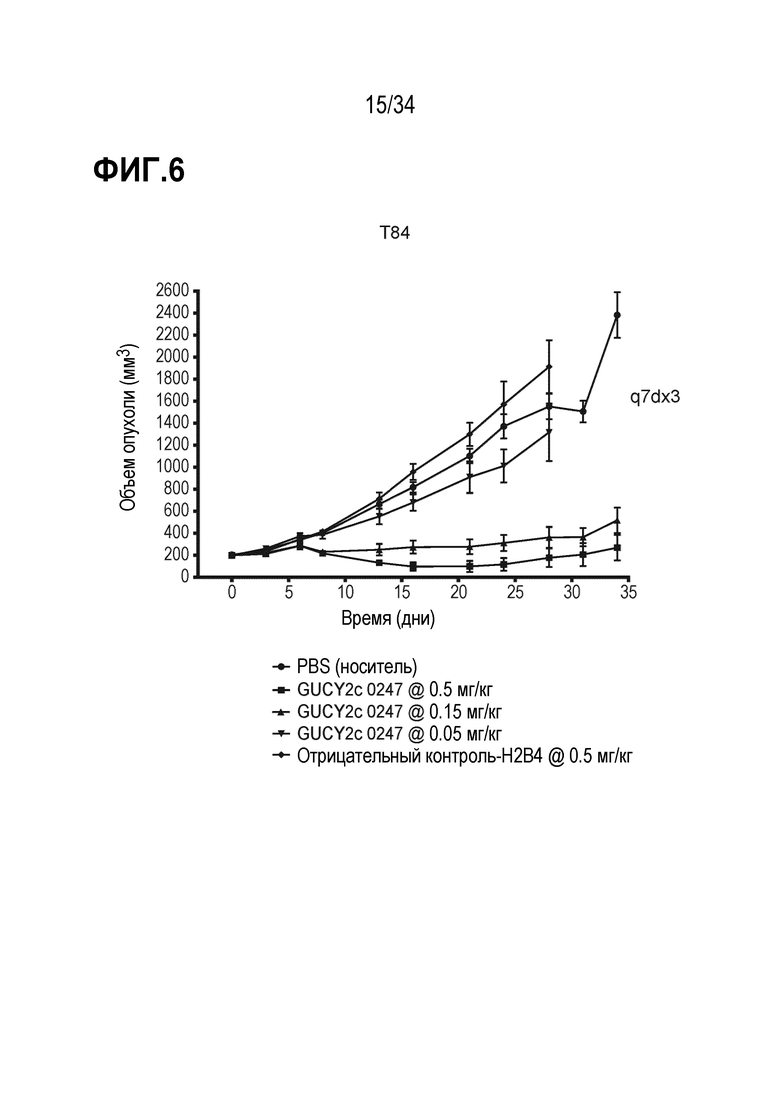
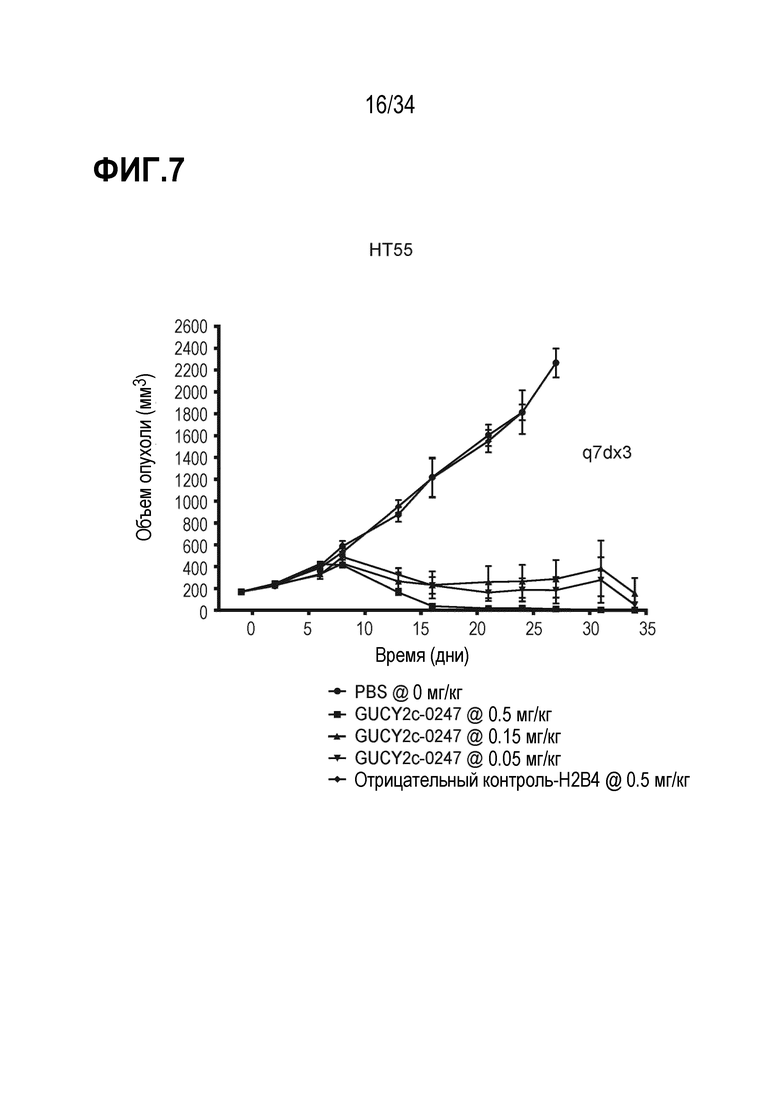
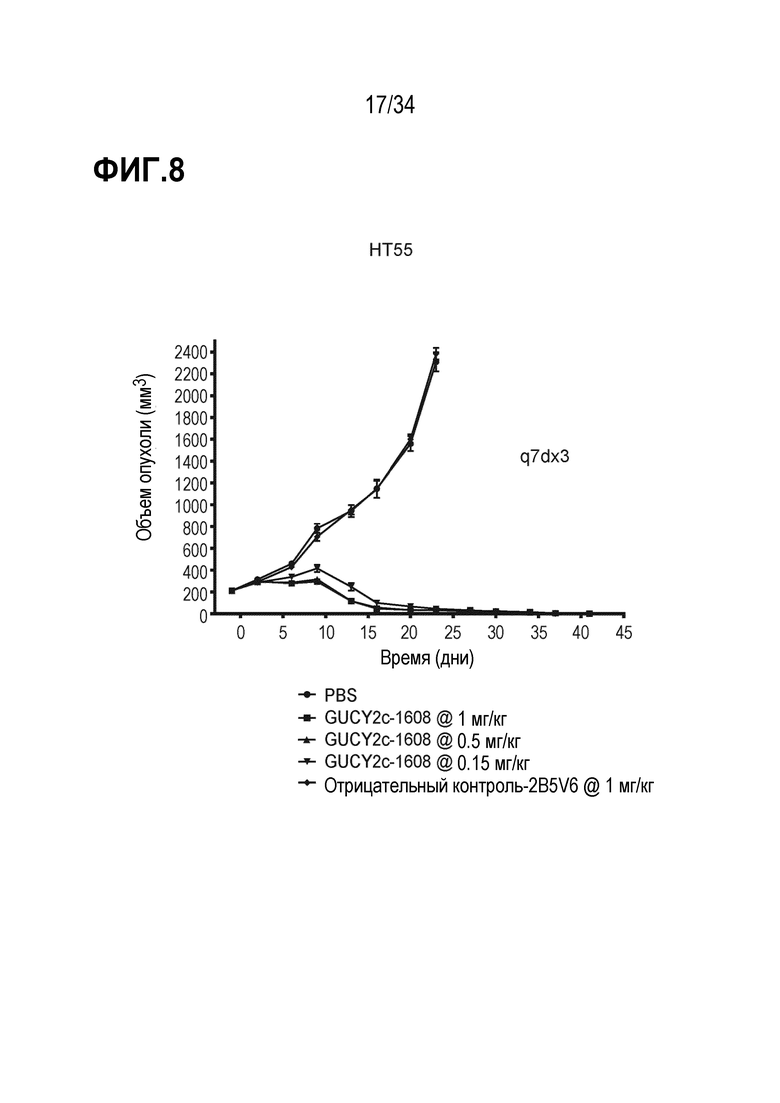
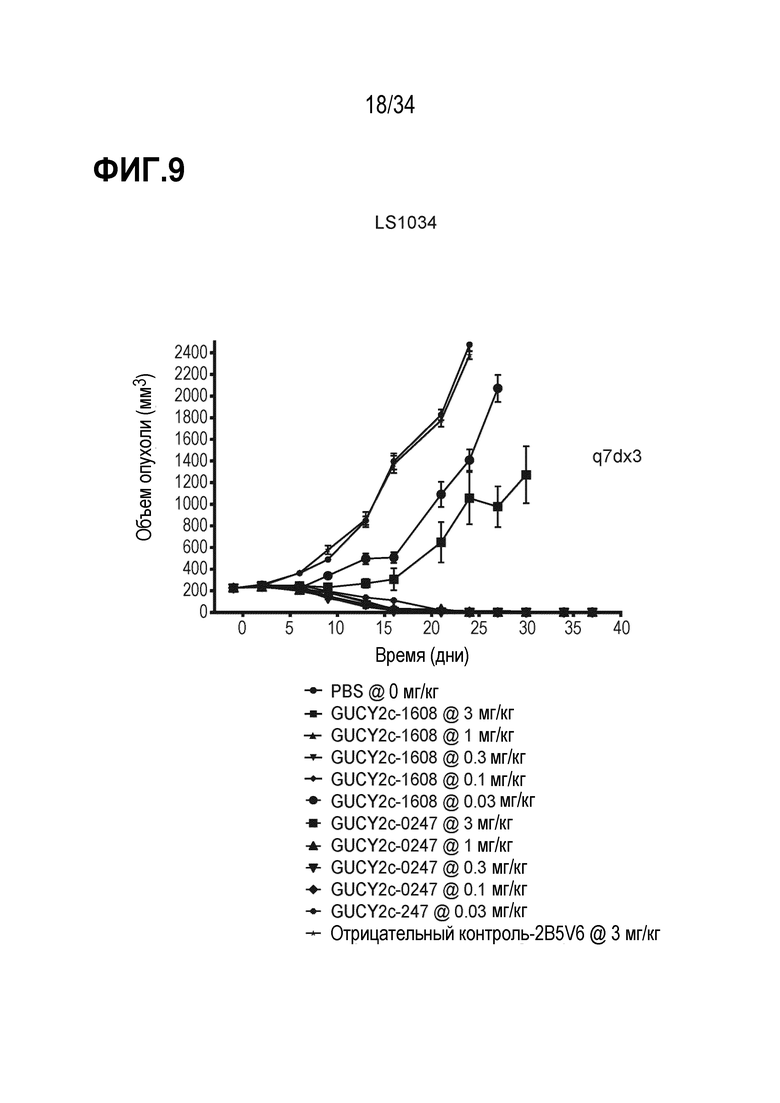
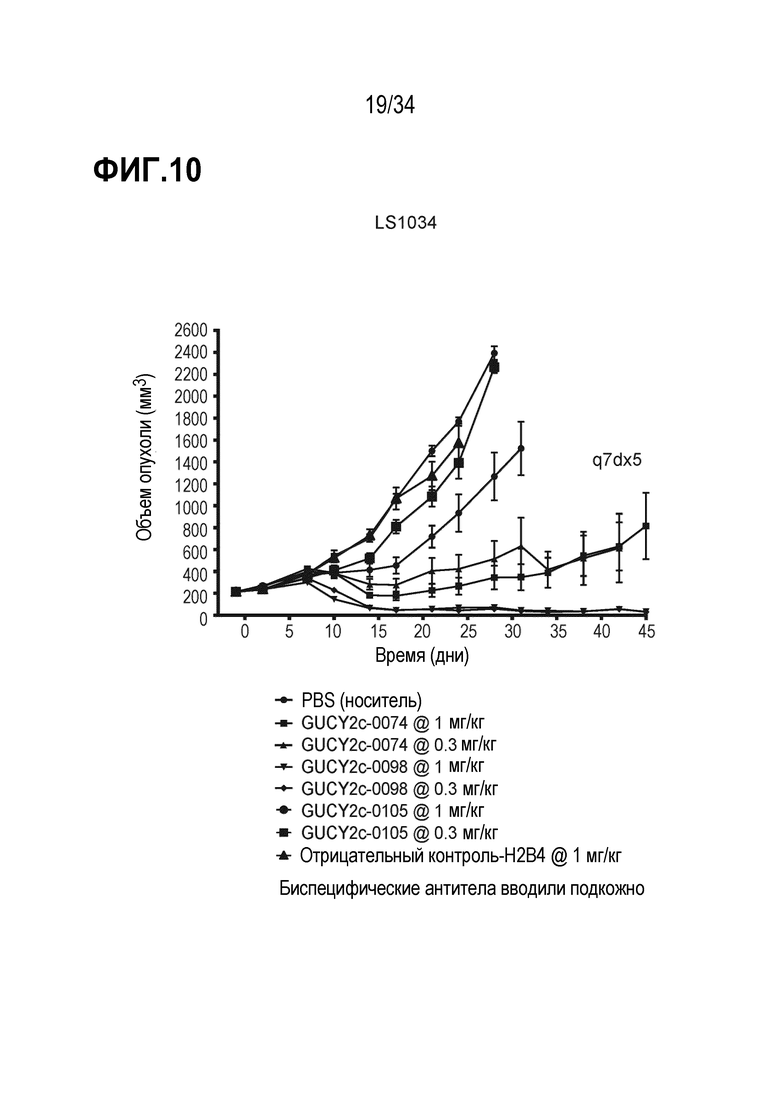
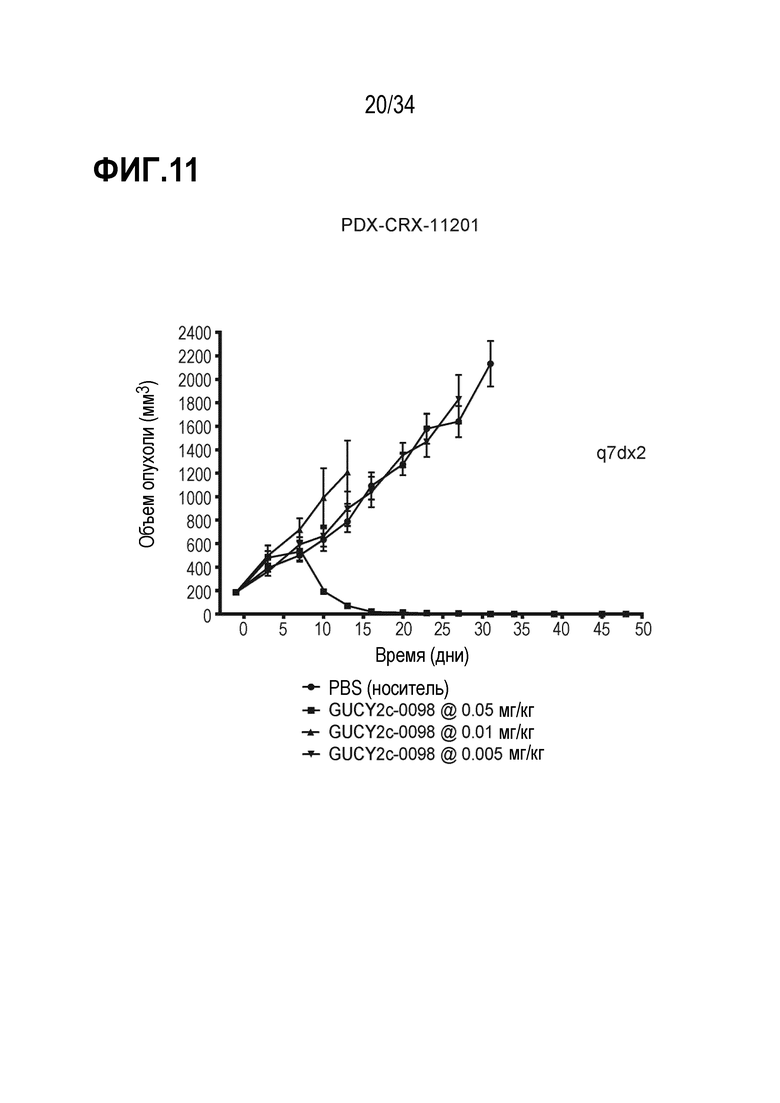
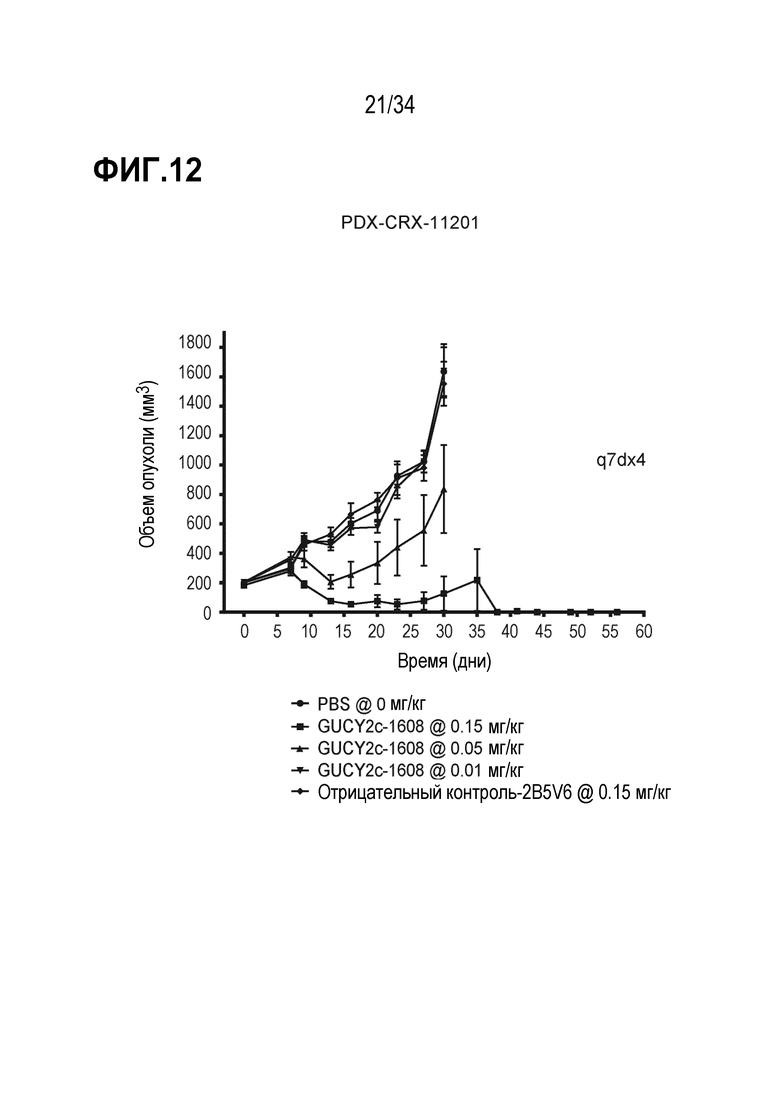
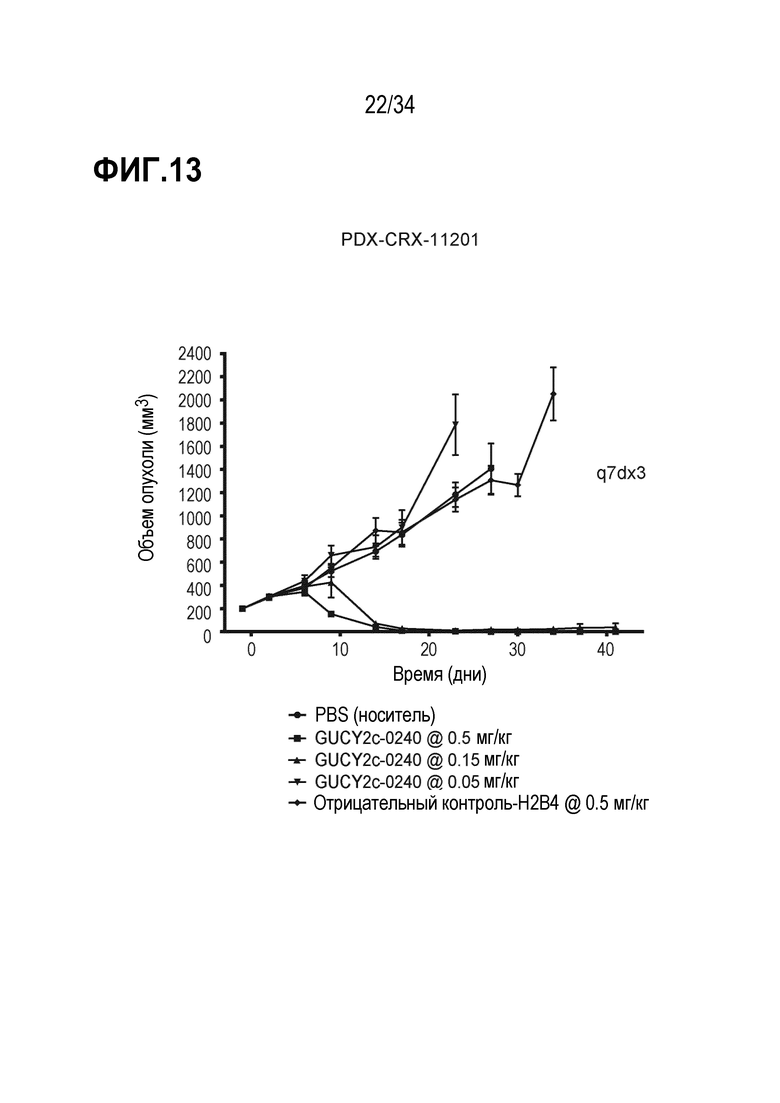
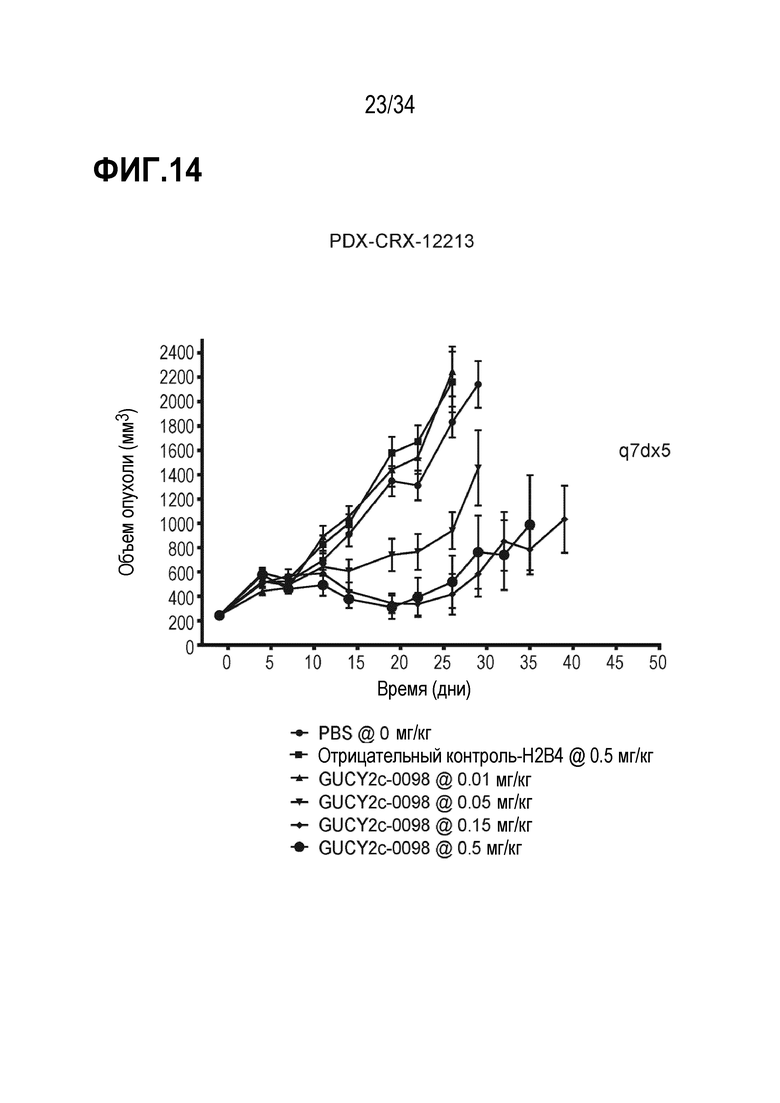
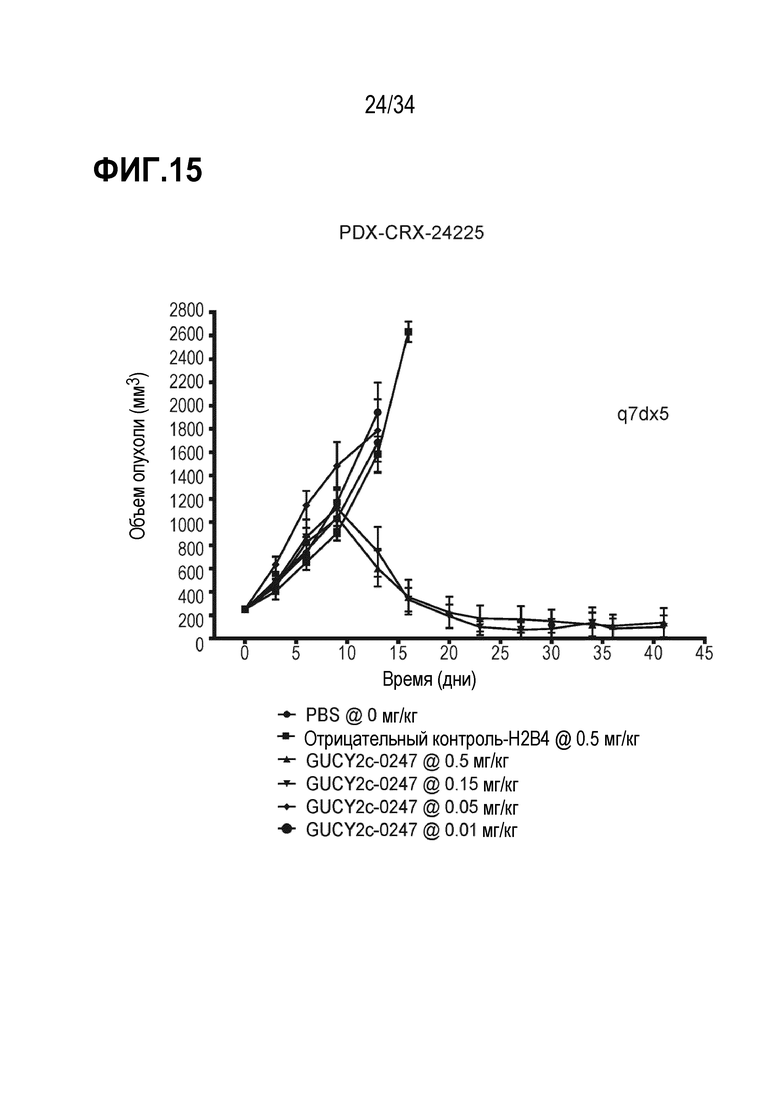
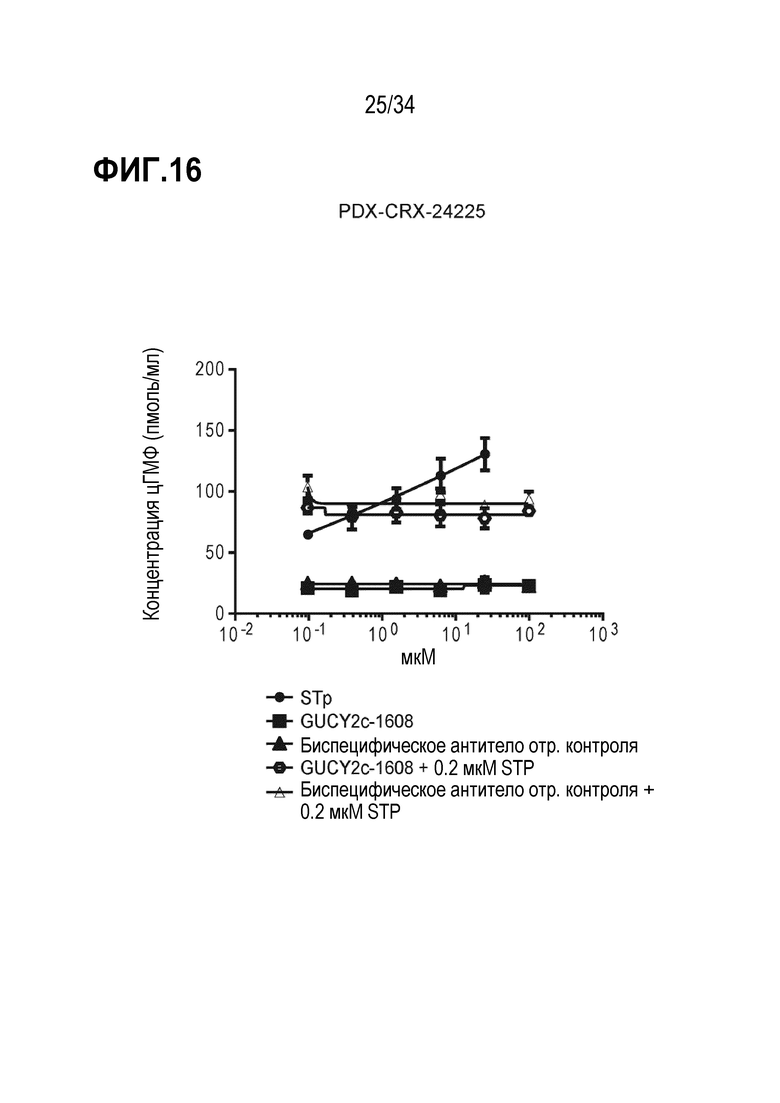
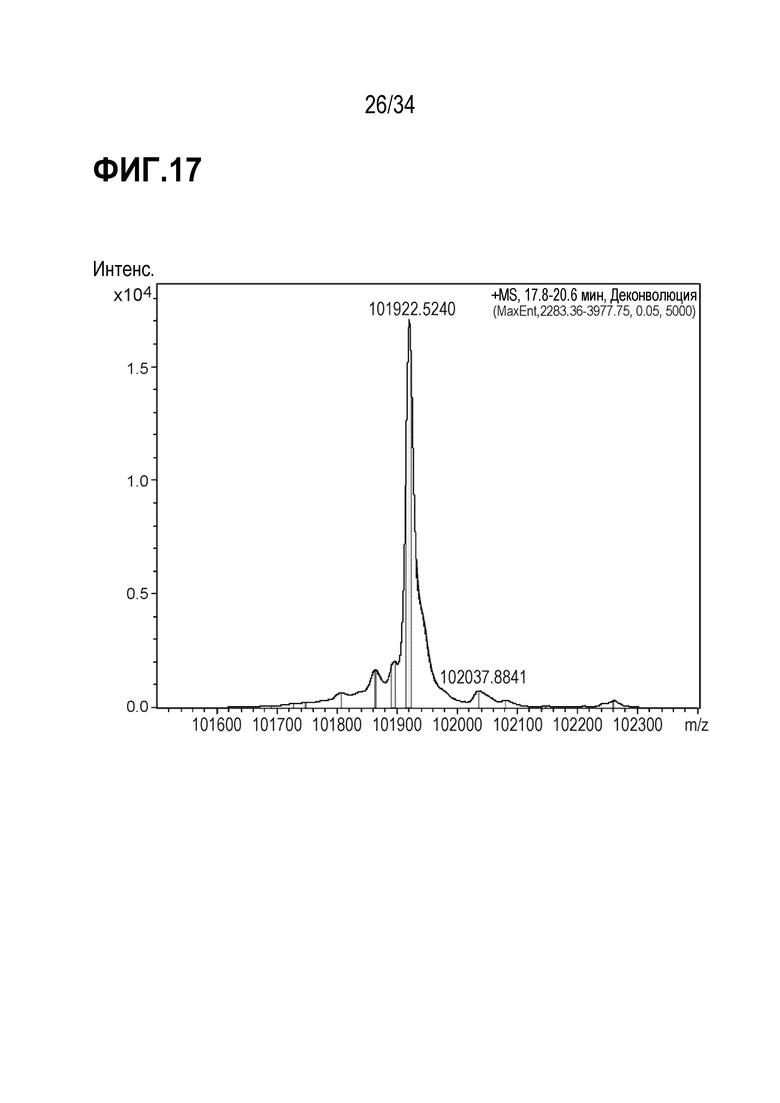
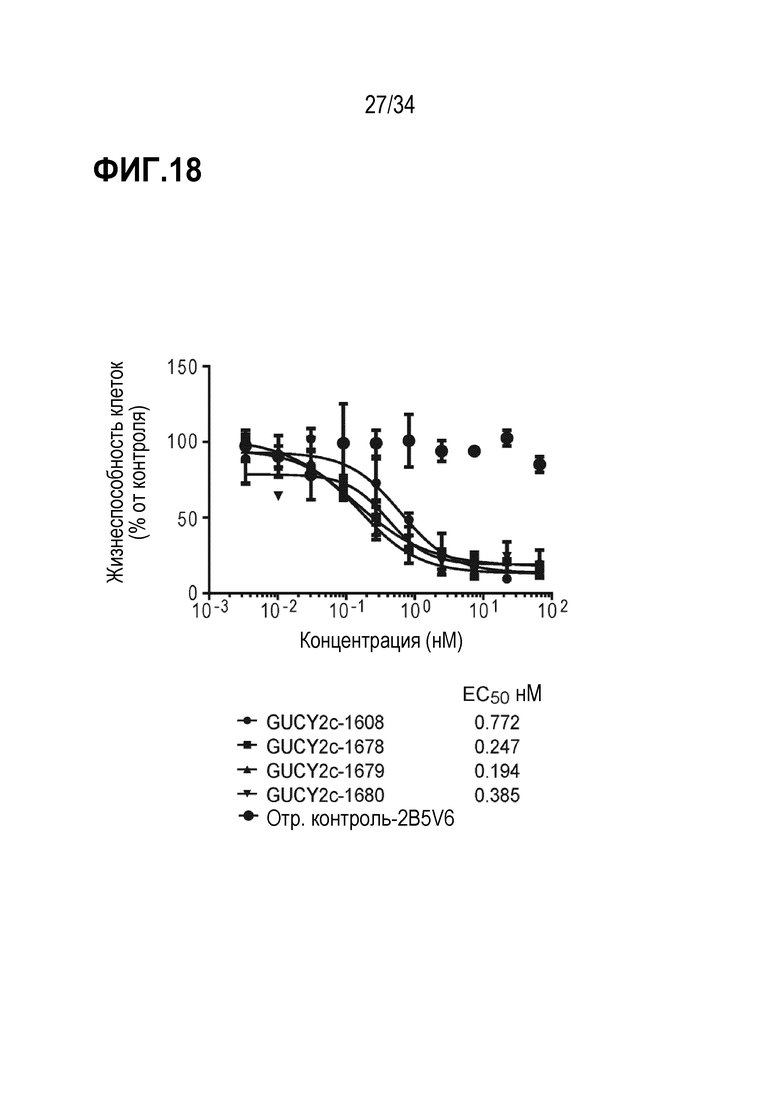
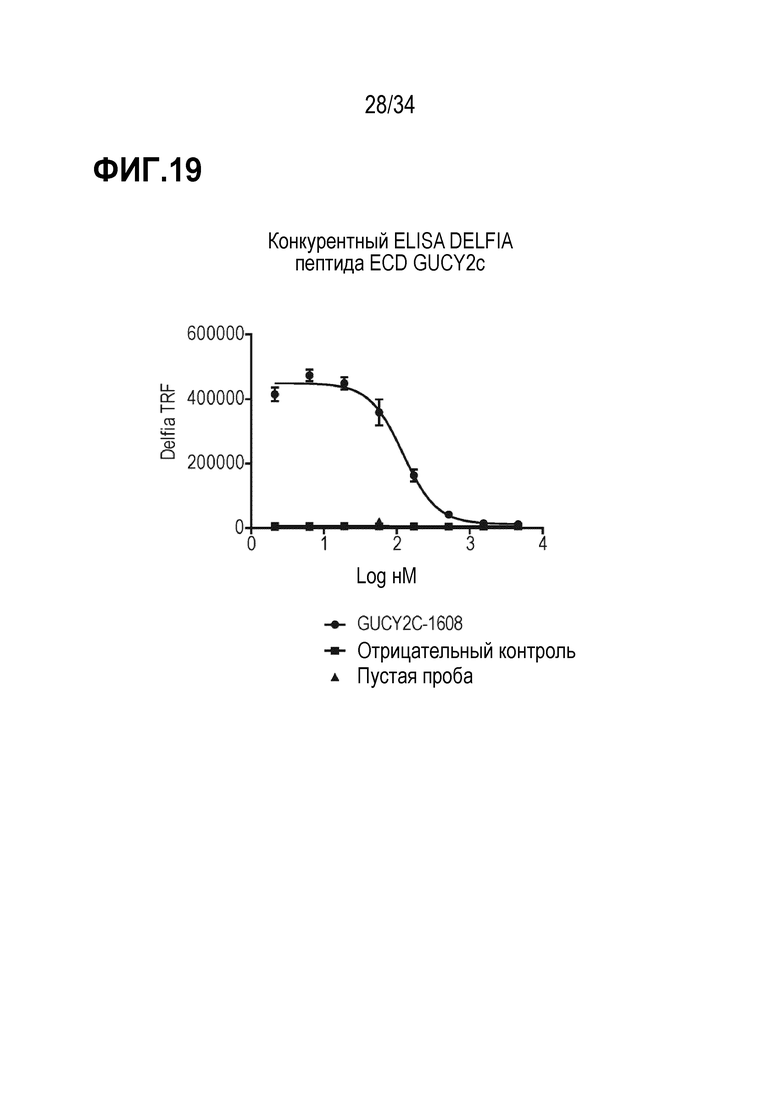
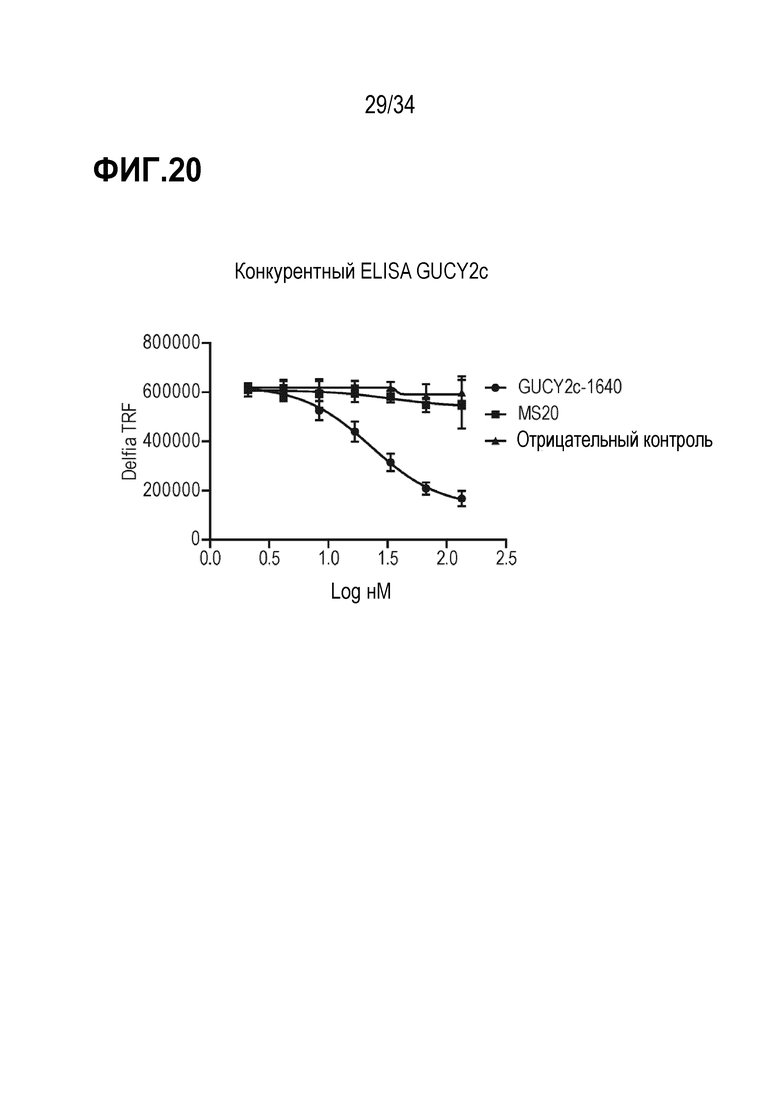
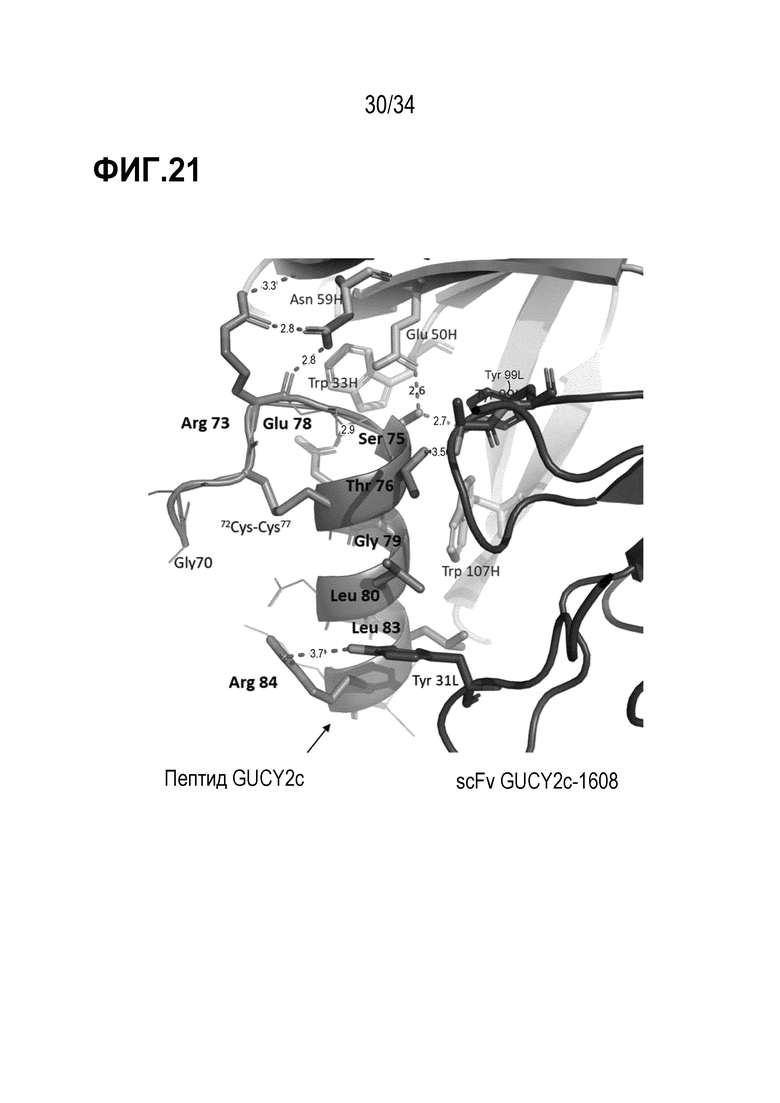
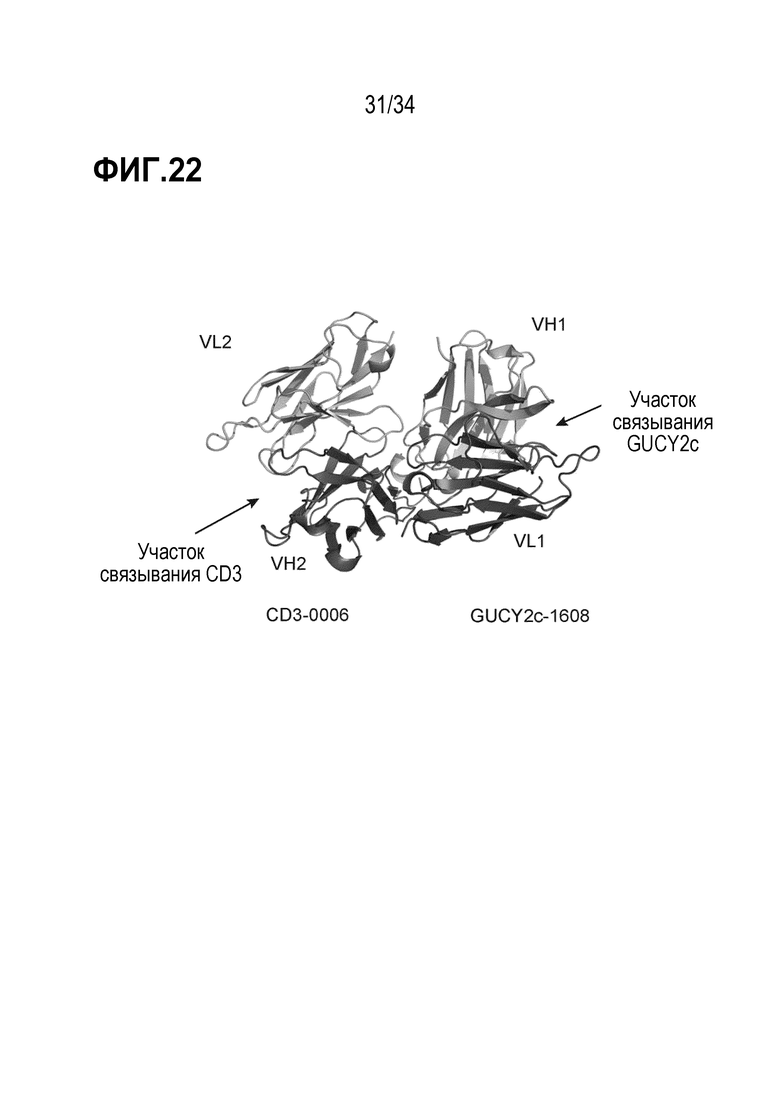
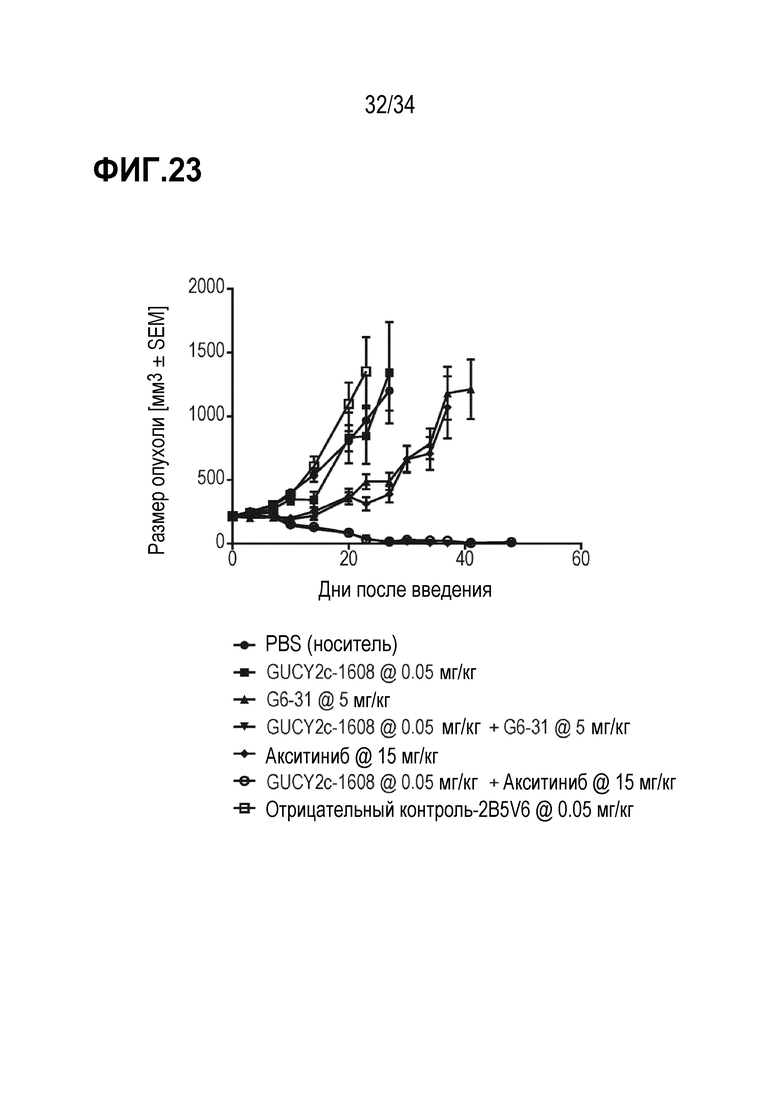
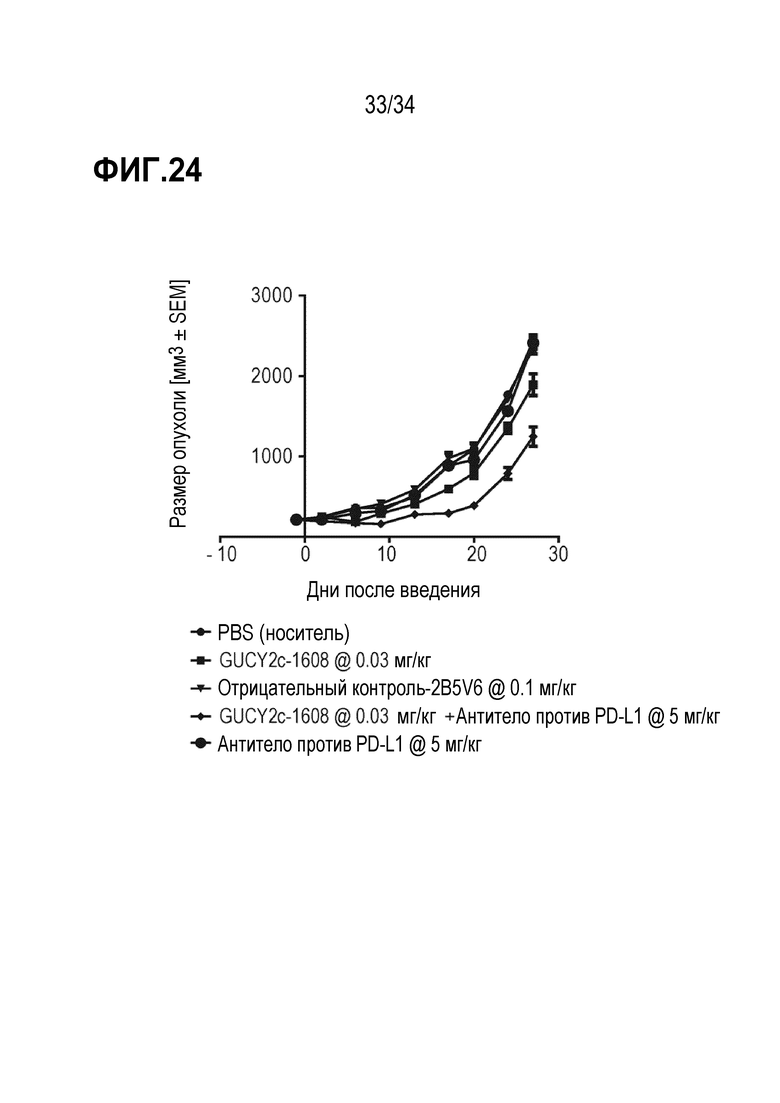
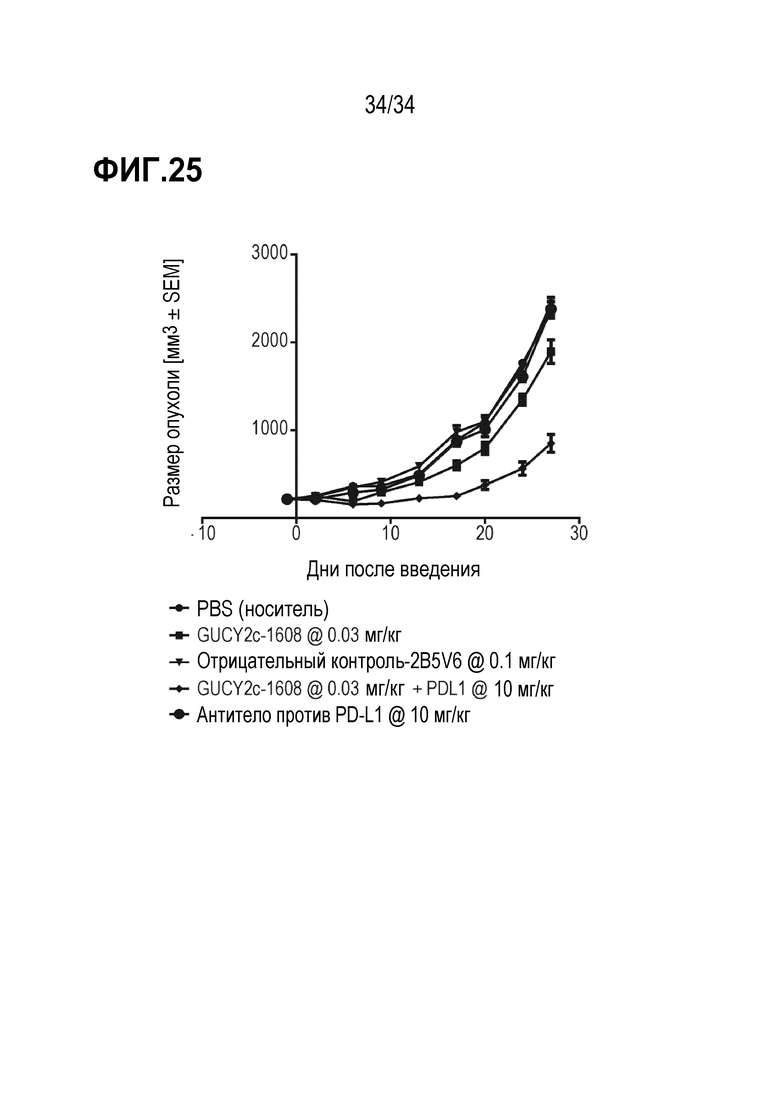
Группа изобретений относится к биотехнологии. Представлены: антитело, которое специфически связывается с гуанилатциклазой C (GUCY2c), биспецифические антитела, которые специфически связываются с GUCY2c и CD3, способы лечения заболеваний, связанных с GUCY2c, у пациента, молекулы нуклеиновых кислот, векторы экспрессии, содержащие полинуклеотид, клетки-хозяина для получения антител и композиции для лечения заболеваний, связанных с GUCY2c. Также раскрыты способы получения антитела и биспецифического антитела. Изобретение позволяет получить новые антитела с высокой аффинностью к GUCY2 и CD3 . 18 н. и 18 з.п. ф-лы, 38 ил., 49 табл., 35 пр.
1. Антитело, которое специфически связывается с гуанилатциклазой C (GUCY2c), где антитело содержит:
a. вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область VH один (CDR1 VH), определяющую комплементарность область VH два (CDR2 VH), и определяющую комплементарность область VH три (CDR3 VH) последовательности VH с SEQ ID NO:73; и
b. вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область VL один (CDR1 VL), определяющую комплементарность область VL два (CDR2 VL), и определяющую комплементарность область VL три (CDR3 VL) последовательности VL с SEQ ID NO:147.
2. Антитело по п.1, где:
a. область VH содержит
(i) CDR1 VH, содержащую последовательность SEQ ID NO:74 или 259;
(ii) CDR2 VH, содержащую последовательность SEQ ID NO:75 или 267; и
(iii) CDR3 VH, содержащую последовательность SEQ ID NO:29; и
b. область VL содержит
(i) CDR1 VL, содержащую последовательность SEQ ID NO:148;
(ii) CDR2 VL, содержащую последовательность SEQ ID NO:149; и
(iii) CDR3 VL, содержащую последовательность SEQ ID NO:142.
3. Антитело по п.1, где:
а. область VH содержит последовательность SEQ ID NO:73; и
b. область VL содержит последовательность SEQ ID NO:147.
4. Антитело по п.1, где
область VH содержит последовательность SEQ ID NO:73; и
область VL содержит последовательность SEQ ID NO:147.
5. Антитело по п.1, где
область VH содержит последовательность SEQ ID NO:73 или ее вариант с одной или несколькими консервативными аминокислотными заменами в остатках, которые не находятся в области CDR; и/или
область VL содержит последовательность SEQ ID NO:147 или ее вариант с одной или несколькими консервативными аминокислотными заменами в аминокислотах, которые не находятся в области CDR.
6. Антитело по п.1, где антитело выбрано из группы, состоящей из Fab-фрагмента, Fab'-фрагмента, F(ab')2-фрагмента, Fd-фрагмента, Fv-фрагмента, одноцепочечного Fv-фрагмента (scFv), стабилизированного дисульфидными связями Fv-фрагмента (dsFv), однодоменного фрагмента антитела (dAb), моноклонального антитела, химерного антитела, поликлонального антитела, гуманизированного антитела, антитела человека и их фрагментов.
7. Антитело по п.1, содержащее
каркас VH человека или гуманизированный каркас VH и
каркас VL человека или гуманизированный каркас VL.
8. Антитело по п.7, где
каркас VH содержит последовательность SEQ ID NO: 5, 6, 7, 8, 15, 16, 17, 18, 23, 24, 25, 30, 31, 32, 37, 38, 39, 40, 45, 46, 47, 49, 50, 51, 54, 55, 56, 58, 59, 61 или 63; и/или
каркас VL содержит последовательность SEQ ID NO: 80, 81, 82, 83, 86, 87, 88, 89, 96, 97, 98, 99, 103, 110, 111, 116, 117, 118, 122, 123, 124, 126, 127, 128, 130, 131, 132, 133, 135, 139 или 155.
9. Выделенное моноклональное антитело человека по любому из пп.1-8, которое связывается с эпитопом на внеклеточном домене GUCY2c,
где эпитоп содержит по меньшей мере один аминокислотный остаток, выбранный из аминокислотных остатков R73, S74, S75, T76, E78, G79, L80, L82, L83, R84 или I86 последовательности SEQ ID NO:406.
10. Биспецифическое антитело, которое специфически связывается с GUCY2c и CD3, где биспецифическое антитело содержит первую полипептидную цепь и вторую полипептидную цепь, где:
a. (i) первая полипептидная цепь содержит в направлении от N-конца к C-концу: домен 1, содержащий область VL антитела против GUCY2c (VL GUCY2c) и область VH антитела против CD3 (VH CD3), и первый способствующий гетеродимеризации домен; и
(ii) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: домен 2, содержащий область VL антитела против CD3 (VL CD3) и область VH антитела против GUCY2c (VH GUCY2c), и второй способствующий гетеродимеризации домен;
b. (i) первая полипептидная цепь содержит в направлении от N-конца к C-концу: домен 1, содержащий VL CD3 и VH GUCY2c, и первый способствующий гетеродимеризации домен; и
(ii) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: домен 2, содержащий VL GUCY2c и VH CD3, и второй способствующий гетеродимеризации домен;
с. (i) первая полипептидная цепь содержит в направлении от N-конца к C-концу: домен 1, содержащий VH GUCY2c и VL CD3, и первый способствующий гетеродимеризации домен; и
(ii) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: домен 2, содержащий VH CD3 и VL GUCY2c, и второй способствующий гетеродимеризации домен; или
d. (i) первая полипептидная цепь содержит в направлении от N-конца к C-концу: домен 1, содержащий VH CD3 и VL GUCY2c, и первый способствующий гетеродимеризации домен; и
(ii) вторая полипептидная цепь содержит в направлении от N-конца к C-концу: домен 2, содержащий VH GUCY2c и VL CD3, и второй способствующий гетеродимеризации домен, и
где VL GUCY2c и VH GUCY2c образуют домен, специфически связывающийся с GUCY2c; и VL CD3 и VH CD3 образуют домен, специфически связывающийся с CD3, и где
VH GUCY2c содержит:
(i) CDR1 VH GUCY2c содержит последовательность SEQ ID NO:74 или 259;
(ii) CDR2 VH GUCY2c содержит последовательность SEQ ID NO:75 или 267;
(iii) CDR3 VH GUCY2c содержит последовательность SEQ ID NO:29;
VL GUCY2c содержит:
(i) CDR1 VL GUCY2c содержит последовательность SEQ ID NO:148;
(ii) CDR2 VL GUCY2c содержит последовательность SEQ ID NO:149; и
(iii) CDR3 VL GUCY2c содержит последовательность SEQ ID NO:142;
VH CD3 содержит
(i) CDR1 VH CD3 содержит последовательность SEQ ID NO:2 или 268;
(ii) CDR2 VH CD3 содержит последовательность SEQ ID NO:10 или 270; и
(iii) CDR3 VH CD3 содержит последовательность SEQ ID NO:4; и
VL CD3 содержит
(i) CDR1 VL CD3 содержит последовательность SEQ ID NO:91;
(ii) CDR2 VL CD3 содержит последовательность SEQ ID NO:78; и
(iii) CDR3 VL CD3 содержит последовательность SEQ ID NO:79.
11. Биспецифическое антитело по п.10, где каждый из первого способствующего гетеродимеризации домена и второго способствующего гетеродимеризации домена содержит Fc-область, содержащую домен CH2 и домен CH3,
где аминокислотная последовательность каждого из домена CH2 и/или CH3 содержит по меньшей мере одну аминокислотную модификацию по сравнению с Fc-областью дикого типа, приводящую к образованию выступа или впадины.
12. Биспецифическое антитело по п.11, где и первый способствующий гетеродимеризации домен, и второй способствующий гетеродимеризации домен не являются выступами или впадинами; и/или
где первый способствующий гетеродимеризации домен и второй способствующий гетеродимеризации домен образуют Fc-область иммуноглобулина IgG.
13. Биспецифическое антитело по п.10, где
VL GUCY2c и VH CD3 соединены глицин-сериновым линкером; и
VL CD3 и VH GUCY2c соединены глицин-сериновым линкером.
14. Биспецифическое антитело по п.10, где
домен 1 ковалентно связан с первым способствующим гетеродимеризации доменом посредством цистеинового линкера; и
домен 2 ковалентно связан со вторым способствующий гетеродимеризации доменом посредством цистеинового линкера;
где цистеиновый линкер содержит по меньшей мере пять аминокислот.
15. Биспецифическое антитело по п.10, где:
(i) первая полипептидная цепь содержит в направлении от N-конца к С-концу: домен 1, содержащий область VL антитела GUCY2c (VL GUCY2c) и область VH антитела против CD3 (VH CD3), и первый способствующий гетеродимеризации домен; и
(ii) вторая полипептидная цепь содержит в направлении от N-конца к С-концу: домен 2, содержащий область VL антитела против CD3 (VL CD3) и область VH антитела GUCY2c (VH GUCY2c), и второй способствующий гетеродимеризации домен; и где
VH GUCY2c содержит
(i) CDR1 VH GUCY2c, содержащую аминокислотную последовательность SEQ ID NO:259,
(ii) CDR2 VH GUCY2c, содержащую аминокислотную последовательность SEQ ID NO:75, и
(iii) CDR3 VH GUCY2c, содержащую аминокислотную последовательность SEQ ID NO:29;
VL GUCY2c содержит
(i) CDR1 VL GUCY2c, содержащую аминокислотную последовательность SEQ ID NO:148,
(ii) CDR2 VL GUCY2c, содержащую аминокислотную последовательность SEQ ID NO:149, и
(iii) CDR3 VL GUCY2c, содержащую аминокислотную последовательность SEQ ID NO:142;
VH CD3 содержит
(i) CDR1 VH CD3, содержащую аминокислотную последовательность SEQ ID NO:268,
(ii) CDR2 VH CD3, содержащую аминокислотную последовательность SEQ ID NO:10, и
(iii) CDR3 VH CD3, содержащую аминокислотную последовательность SEQ ID NO:4; и
VL CD3 содержит
(i) CDR1 VL CD3, содержащую аминокислотную последовательность SEQ ID NO:91,
(ii) CDR2 VL CD3, содержащую аминокислотную последовательность SEQ ID NO:78, и
(iii) CDR3 VL CD3, содержащую аминокислотную последовательность SEQ ID NO:79.
16. Биспецифическое антитело по п.10, где
(i) первая полипептидная цепь содержит в направлении от N-конца к С-концу: домен 1, содержащий область VL антитела GUCY2c (VL GUCY2c) и область VH антитела против CD3 (VH CD3), и первый способствующий гетеродимеризации домен; и
(ii) вторая полипептидная цепь содержит в направлении от N-конца к С-концу: домен 2, содержащий область VL антитела против CD3 (VL CD3) и область VH антитела GUCY2c (VH GUCY2c), и второй способствующий гетеродимеризации домен; и где
VH GUCY2c содержит
(i) CDR1 VH GUCY2c, содержащий аминокислотную последовательность SEQ ID NO:74,
(ii) CDR2 VH GUCY2c, содержащий аминокислотную последовательность SEQ ID NO:75, и
(iii) CDR3 VH GUCY2c, содержащий аминокислотную последовательность SEQ ID NO:29;
VL GUCY2c содержит
(i) CDR1 VL GUCY2c, содержащую аминокислотную последовательность SEQ ID NO:148,
(ii) CDR2 VL GUCY2c, содержащую аминокислотную последовательность SEQ ID NO:149, и
(iii) CDR3 VL GUCY2c, содержащий аминокислотную последовательность SEQ ID NO:142;
VH CD3 содержит
(i) CDR1 VH CD3, содержащую аминокислотную последовательность SEQ ID NO:2,
(ii) CDR2 VH CD3, содержащую аминокислотную последовательность SEQ ID NO:10, и
(iii) CDR3 VH CD3, содержащую аминокислотную последовательность SEQ ID NO:4; и
VL CD3 содержит
(i) CDR1 VL CD3, содержащую аминокислотную последовательность SEQ ID NO:91,
(ii) CDR2 VL CD3, содержащую аминокислотную последовательность SEQ ID NO:78, и
(iii) CDR3 VL CD3, содержащую аминокислотную последовательность SEQ ID NO:79.
17. Биспецифическое антитело по п.10, где:
a. VH GUCY2c содержит аминокислотную последовательность SEQ ID NO:73; и
b. VL GUCY2c содержит аминокислотную последовательность SEQ ID NO:147.
18. Биспецифическое антитело по п.10, где:
a. VH CD3 содержит аминокислотную последовательность SEQ ID NO:9; и
b. VL CD3 содержит аминокислотную последовательность SEQ ID NO:90.
с. VH GUCY2c содержит аминокислотную последовательность SEQ ID NO:73; и
d. VL GUCY2c содержит аминокислотную последовательность SEQ ID NO:147.
19. Биспецифическое антитело, специфически связывающееся с GUCY2c и CD3, где биспецифическое антитело содержит первую полипептидную цепь и вторую полипептидную цепь, и где:
a. первая полипептидная цепь содержит следующие области в следующем порядке в направлении от N-конца к C-концу:
VL антитела против GUCY2c (VL GUCY2c) - Линкер 1 - VH антитела против CD3 (VH CD3) - Линкер 3 - первый способствующий гетеродимеризации домен,
где VL GUCY2c содержит аминокислотную последовательность SEQ ID NO:147,
Линкер 1 содержит аминокислотную последовательность SEQ ID NO:190,
VH CD3 содержит аминокислотную последовательность SEQ ID NO:9,
Линкер 3 содержит аминокислотную последовательность SEQ ID NO:192, и
первый способствующий гетеродимеризации домен содержит аминокислотную последовательность SEQ ID NO:188; и
b. вторая полипептидная цепь содержит следующие области в следующем порядке в направлении от N-конца к C-концу:
VL антитела против CD3 (VL CD3) - Линкер 2 - VH антитела против GUCY2c (VH GUCY2c) - Линкер 3 - второй способствующий гетеродимеризации домен,
где VL CD3 содержит аминокислотную последовательность SEQ ID NO:90,
Линкер 2 содержит аминокислотную последовательность SEQ ID NO:191,
VH GUCY2c содержит аминокислотную последовательность SEQ ID NO:73,
Линкер 3 содержит аминокислотную последовательность SEQ ID NO:192 и
второй способствующий гетеродимеризации домен содержит аминокислотную последовательность SEQ ID NO:189.
20. Биспецифическое антитело по п.19, где
VL GUCY2c и VH GUCY2c образуют домен, специфически связывающийся с GUCY2c; и
VL CD3 и VH CD3 образуют домен, специфически связывающийся с CD3;
где Линкер 3 первой полипептидной цепи и Линкер 3 второй полипептидной цепи ковалентно связаны друг с другом двумя дисульфидными связями; где каждый из первого способствующего гетеродимеризации домена и второго способствующего гетеродимеризации домена содержит домен CH2 и домен CH3;
где домен CH3 первого способствующего гетеродимеризации домена образует выступ, и домен CH3 второго способствующего гетеродимеризации домена образует впадину; и
где по меньшей мере одна дисульфидная связь образуется между доменом CH3 первого способствующего гетеродимеризации домена и доменом CH3 второго способствующего гетеродимеризации домена.
21. Биспецифическое антитело по п.10, где биспецифическое антитело является гуманизированным антителом.
22. Биспецифическое антитело, которое специфически связывается с GUCY2c и с CD3, содержащее первую полипептидную цепь и вторую полипептидную цепь,
где первая полипептидная цепь содержит аминокислотную последовательность SEQ ID NO:216, и
вторая полипептидная цепь содержит аминокислотную последовательность SEQ ID NO:220.
23. Способ лечения заболевания, связанного с GUCY2c, у пациента, включающий введение пациенту антитела по п.1, биспецифического антитела по п.10 или фармацевтической композиции, содержащей такое антитело или биспецифическое антитело.
24. Молекула нуклеиновой кислоты, кодирующая антитело по п.1.
25. Молекула нуклеиновой кислоты, кодирующая биспецифическое антитело по п.10.
26. Вектор экспрессии, содержащий полинуклеотид по п.24.
27. Вектор экспрессии, содержащий полинуклеотид по п.25.
28. Клетка-хозяин для получения антитела по п.1, содержащая вектор по п.26.
29. Клетка-хозяин для получения биспецифического антитела по п.10, содержащая вектор по п.27.
30. Способ получения антитела, включающий культивирование клетки-хозяина по п.28 в условиях, приводящих к продукции антитела по п.1, и очистку антитела из супернатанта культуры.
31. Способ получения биспецифического антитела, включающий культивирование клетки-хозяина по п.29 в условиях, приводящих к продукции биспецифического антитела по п.10, и очистку биспецифического антитела из супернатанта культуры.
32. Композиция для лечения заболевания, связанного с GUCY2c, содержащая эффективное количество биспецифического антитела по п.10 и второе терапевтическое средство.
33. Способ лечения заболевания, связанного с GUCY2c, у пациента, включающий введение проведение пациенту комбинированной терапии, которая включает композицию по п.32.
34. Композиция для лечения заболевания, связанного с GUCY2c, содержащая эффективное количество биспецифического антитело по п.10 и противодиарейное средство.
35. Способ лечения заболевания, связанного с GUCY2c у индивидуума, включающий введение индивидууму комбинированного терапевтического средства, содержащего композицию по п.34.
36. Фармацевтическая композиция, для лечения заболевания, связанного с GUCY2c, содержащая эффективное количество биспецифического антитела по п.10 и фармацевтически приемлемый носитель.
| EVA DAHLEN et al.: "Bispecific antibodies in cancer immunotherapy", THERAPEUTIC ADVANCES IN VACCINES AND IMMUNOTHERAPY BISPECIFIC ANTIBODIES IN CANCER IMMUNOTHERAPY, vol | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| WO 2013055809 A1, 18.0.2013 | |||
| US 2017233472 A1, 17.08.2017 | |||
| US 2013315923 Al, 28.11.2013 | |||
| RU 2012143519 А,10.05.2014. | |||
