Изобретение относится к области биотехнологии и производству вакцин против возбудителя чумы плотоядных животных, а именно к способу опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени с применением разработанной логарифмической функции зависимости величины цикла количественной оценки и титра инфекционной активности возбудителя чумы плотоядных животных.
Чума плотоядных - высококонтагиозное вирусное заболевание, поражающее многих диких и домашних плотоядных, приводящее к высокой заболеваемости и смертности хозяев [1]. Передача вирусных частиц происходит при контакте или вдыхании загрязненных жидкостей, хотя недавно сообщалось, что блохи являются переносчиками инфекции [3]. Проявление чумы плотоядных варьирует от легких до тяжелых симптомов в зависимости от иммунного статуса хозяина [2].
Возбудитель чумы плотоядных животных относится к семейству Paramyxoviridae, подсемейству Orthoparamyxoviridae, роду Morbillivirus, виду Canine Distemper virus (CDV) [3]. CDV представляет собой оболочечный несегментированный одноцепочечный РНК-содержащий вирус длиной около 15600-15700 н.о. Он содержит геном РНК с отрицательным смыслом, который кодирует шесть белков. Из них белок Н (гемагглютинин) является составной частью оболочечных гликопротеиновых шипов на вирионе и инициирует проникновение вирусной частицы в клетку-хозяина путем прикрепления к клеточным рецепторам [3, 4, 5].
Мономер белка Н представляет собой трансмембранный гликопротеин типа II, состоящий из короткого N-концевого цитоплазматического хвоста, за которым следует трансмембранный домен и большой С-концевой эктодомен. Эктодомен структурирован в виде ножки и шестилопастной (В1-В6) β-пропеллерной складки, окружающей центральную полость [6, 7].
Белок Н играет важную роль в клеточном тропизме. Отсюда следует, что изменчивость антигена может влиять на вирулентность, круг хозяев и эпитопы нейтрализации возбудителя чумы плотоядных.
Анализ базы данных последовательностей генов РНК-вирусов GenBank показал, что, помимо точечных мутаций, важную роль в эволюции несегментированных РНК-вирусов играет рекомбинация. Ген Н имеет самую высокую генетическую изменчивость в геноме чумы плотоядных животных и поэтому обычно используется для молекулярного типирования изолятов/штаммов CDV [8]. Вирус чумы плотоядных имеет географически различные линии происхождения. Штаммы, которые находятся в пределах одной клады и имеют >95% сходства аминокислотной последовательности в белке Н, считаются принадлежащими к одному и тому же генотипу [9].
Система мер для борьбы с чумой плотоядных и его профилактики предусматривает иммунизацию домашних животных [8]. Для этой цели применяют вакцинные препараты. При их изготовлении вируссодержащее сырье исследуют на определение титра инфекционной активности возбудителя чумы плотоядных для оценки его активности в клетках. В 1,0 см3 суспензии вируса определяют количество тканевых цитопатических доз, вызывающих 50%-ное поражение клеток, что фактически отражает концентрацию полных вирусных частиц, содержащих РНК вируса чумы плотоядных.
Традиционно для определения титра инфекционной активности возбудителя чумы плотоядных применяют метод титрования в перевиваемой монослойной культуре клеток почки африканской зеленой мартышки (Vero), с помощью которой вычисляют минимальную дозу вируса, способную вызвать лизис 50% клеток (прототип) [1]. Данный метод имеет некоторые недостатки: 1) длительная процедура титрования, связанная с поражением клеток возбудителем чумы плотоядных; 2) определенная степень субъективности при оценке результатов исследования; 3) высокая стоимость клеточной линии Vero как тест-системы и затраты на ее поддержание; 4) высокая вероятность риска контаминации культуры клеток Vero.
В связи с этим целесообразно провести поиск альтернативного способа опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени.
Техническим результатом может являться предлагаемый метод, как высокочувствительный и специфичный, объективный и позволяющий определять титр инфекционной активности возбудителя чумы плотоядных в вируссодержащих суспензиях в течение 3,5 часов, не требующий использования культуры клеток для анализа, без контаминации исследуемых образцов, исключающий фактор субъективной оценки результатов исследования, поскольку проводится детекция конечных данных с помощью приборного обеспечения.
Задачей настоящего изобретения является разработка высокочувствительного, высокоспецифичного, экспрессного, объективного, не предполагающего применения линий клеток способа опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени с целью устранения вышеуказанных недостатков.
Данная задача решена благодаря созданию опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени. Разработанный способ дает возможность: 1) сократить время проведения анализа вируссодержащих суспензий для определения титра инфекционной активности возбудителя чумы плотоядных животных до 3,5 ч; 2) исключить вероятность контаминации исследуемого образца; 3) увеличить чувствительность и специфичность анализа за счет применения высокоспецифичных оригинальных праймеров и молекулярного зонда, рассчитанных для Н-гена CDV; 4) повысить достоверность проводимого анализа благодаря установлению зависимости между титром инфекционной активности вируса чумы плотоядных (TCDV) и циклом количественной оценки (Cq), представленной в виде логарифмической функции lg TCDV=-0,3002×Cq,+9,4976 с высокой достоверностью аппроксимации (R2=0,9965) и эффективностью амплификации 99,64%. Предложенная модель позволяет опосредованного определять титр инфекционной активности возбудителя чумы плотоядных в сырье для вакцин с помощью разработанного способа.
Сущность изобретения отражена на графических изображениях:
Фиг. 1 - Нуклеотидная последовательность прямого, обратного праймеров и молекулярного зонда при выравнивании с референтным штаммом «Rockborn» возбудителя чумы плотоядных. Примечание: используя выравнивания других изолятов и производственных штаммов получили тот же дизайн олигонуклеотидов; А - дизайн прямого праймера и зонда, Б - дизайн обратного праймера.
Фиг. 2 - Зависимость титра инфекционной активности возбудителя чумы плотоядных животных (TCDV) и величины цикла количественной оценки (Cq) с помощью метода обратной транскрипции и реакции амплификации в режиме реального времени (n=7, отмечены точки, отображающие средние значения циклов количественной оценки).
Сущность изобретения пояснена следующими перечнями последовательностей штамма «Rockborn» возбудителя чумы плотоядных: SEQ ID NO:1 представляет последовательность нуклеотидов Н-гена кДНК возбудителя чумы плотоядных штамма «Rockborn»;
SEQ ID NO:2 представляет последовательность аминокислот, соответствующих участку Н-гена кДНК возбудителя чумы плотоядных штамма «Rockborn».
Сущность изобретения заключается в подходе по опосредованному определению титра инфекционной активности возбудителя чумы плотоядных животных в сырье для вакцин с помощью метода обратной транскрипции и реакции амплификации в режиме реального времени. Заявляемый способ основан на: 1) элюировании РНК вируса чумы плотоядных животных с применением магнитных коллоидных частиц поверхностно-модифицированного оксида железа (II, III) Fe3O4 с инертной поверхностью за счет обработки 50% раствором полиэтиленимина и диаметром 180-200 нм; 2) проведении обратной транскрипции; 3) амплификации специфического фрагмента Н-гена CDV с применение оригинальных специфических праймеров CDV-F7173, CDV-R7428, а также молекулярного зонда CDV-P7232, меченого флуоресцентным красителем FAM (λmax флуоресценции = 520 нм) и тушителем свечения RTQ-1 (λmax поглощения = 520 нм); 4) проведение реакции амплификации с детекцией ампликонов с помощью флуоресцентного свечения и отображения накопления сигнала в виде логистической кривой; 5) определении титра инфекционной активности CDV с применением логарифмической функции, выраженной в виде уравнения:
lg TCDV = -0,3002 × Cq + 9,4976
с высокой достоверностью аппроксимации (R2=0,9965) и эффективностью амплификации 99,64%.
В настоящее время метод ПНР в режиме реального времени применяют для индикации нуклеиновых кислот и проведения филогенетического анализа различных инфекционных агентов, в том числе возбудителя чумы плотоядных. Для опосредованного определения титра инфекционной активности CDV ранее данный метод с применением разработанной системы оригинальных праймеров и молекулярного зонда не использовался. Таким образом, сведений об аналогах предлагаемого способа опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин авторами не обнаружено.
Разработанный способ опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени по сравнению с прототипом отличается более высокой чувствительностью и специфичностью, объективностью и экспрессностью выполнения анализа и значительным снижением риска контаминации.
В отличие от прототипа разработанный способ включает этапы элюирования РНК CDV животных с применением магнитных коллоидных частиц поверхностно-модифицированного оксида железа (II, III) Fe3O4 с инертной поверхностью и диаметром 180-200 нм; проведения обратной транскрипции; постановки реакции амплификации специфического фрагмента Н-гена CDV с применение оригинальных специфических праймеров и зонда; определения титра инфекционной активности CDV с применением логарифмической функции, выраженной в виде уравнения:
Применение предложенного способа позволит сократить время проведения анализа вируссодержащих суспензий для определения титра инфекционной активности возбудителя чумы плотоядных животных до 3,5 ч; исключить вероятность контаминации и использование клеточных линий; повысить специфичность и чувствительность анализа; увеличить достоверность проводимого анализа. Иными словами, актуально применять предложенный способ для опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени.
Ключевым элементом заявляемого способа является определение значений циклов количественной оценки реакции амплификации нуклеиновой кислоты CDV в ПЦР режиме реального времени и расчет титра инфекционной активности данного вируса с использованием разработанной логарифмической модели зависимости Cq и титра инфекционной активности возбудителя чумы плотоядных.
Сопоставительный анализ с прототипом позволяет сделать вывод, что новизна и изобретательский уровень заявляемого изобретения заключается в применении способа на основе обратной транскрипции, реакции амплификации в режиме реального времени, оригинальных специфичных праймеров и молекулярного зонда, рассчитанных на участок Н-гена CDV, и разработанной логарифмической функции зависимости величины цикла количественной оценки и титра инфекционной активности возбудителя чумы плотоядных для опосредованного определения титра инфекционной активности данного вируса в сырье для вакцин.
Сущность предлагаемого изобретения пояснена на графическом материале - графике зависимости титра инфекционной активности возбудителя чумы плотоядных животных (TCDV) и величины цикла количественной оценки (Cq) с помощью метода обратной транскрипции и реакции амплификации в режиме реального времени (n=7, отмечены точки, отображающие средние значения циклов количественной оценки) (фиг. 2).
С целью определения титра инфекционной активности CDV подготавливают контрольную панель стандартов возбудителя чумы плотоядных штамма «Rockborn», в качестве которых используют лиофильно высушенные суспензии вируса указанного штамма с титрами: 0,0; 1,0; 2,0; 3,0; 4,0; 5,0; 6,0; 7,0; 8,0 lg ТЦД50/см3, которые разводят 1/15 М фосфатно-буферным раствором до требуемого объема и титра инфекционной активности. В качестве отрицательных контролей применяют суспензию клеток Vero, не контаминированную микроорганизмами.
Из всех стандартных положительных образцов и отрицательных контролей, а также тестируемых проб выделяют РНК с помощью коллоидных частиц поверхностно-модифицированного Fe3O4 с инертной поверхностью за счет обработки 50%-ным раствором ПЭИ и диаметром 180-200 нм.
Заблаговременно готовят растворы для элюирования РНК.
Лизирующий раствор. Смешивают гуанидинизотиоцианат (ГТЦ) (47,27 г), тритон Х-100 (2,02 г), поливинилпирролидон (2,00 г), тетрахлоруксусную кислоту (0,73 г) и ацетат аммония (0,40 г), растворяют в деионизированной воде, объем которой доводят до 100 мл. Конечная концентрация ГТЦ составляет 6 М.
Раствор для отмывания 1 представляет собой 80% раствор этилового спирта.
Раствор для отмывания 2 представляет собой 80% изопропанол.
Раствор для элюирования РНК представляет собой буферный раствор ТЕ, состоящий из трис(оксиметил)аминометан (0,01 М) и этилендиамин-тетраацетата (0,001 М), свободного от РНКаз и катионов Mg2+ (pH 7,20-7,30).
К 100 мкл суспензии CDV добавляют 500 мкл полученного лизирующего раствора, инкубируют содержимое в процессе перемешивания в течение 5 мин., затем продлевают инкубирование в открытом сухом термостате при температуре 60±2°С. К полученному лизату добавляют 10 мкл коллоидных частиц поверхностно-модифицированного Fe3O4 с диаметром 180-200 нм. Содержимое пробирок тщательно перемешивают на вортексе, и помещают в магнитный штатив до полного осветления раствора. Не вынимая пробирки из магнитного штатива, удаляют супернатант, не захватывая магнитные частицы. Иными словами, оставляют магнитные частицы, которые сенсибилизированы РНК и балластными компонентами. На следующем этапе проводят отмывание магнитных частиц от балластных составляющих. Для этого в каждую пробирку добавляют по 500 мкл раствора для отмывания 1, тщательно перемешивают на вортексе, и снова помещают в магнитный штатив до полного осветления раствора с последующим удалением супернатанта. Для очистки от гидрофобных примесей в каждую пробирку добавляют по 400 мл раствора для отмывки 2, и повторяют процедуру с удалением жидкости в магнитном штативе. Пробирки с открытыми крышками оставляют в магнитном поле до полного высыхания магнитных частиц. В пробирки добавляют по 50 мкл раствора для элюирования (буфер ТЕ) и прогревают содержимое при температуре 60±2°С в течение 5 мин, периодически встряхивая их на вортексе. После процесса десорбции пробирки помещают в магнитный штатив и после полного осветления раствора, не задевая частицы, отбирают элюаты РНК. Полученный экстракт суммарной РНК хранят при температуре -20±2°С или сразу используют в дальнейшей работе.
На следующем этапе проводят реакцию амплификации в режиме реального времени для исследования контрольных образцов и проб. Для постановки реакции готовят реакционную смесь, рецептура которой представлена в таблиде 1. Дизайн праймеров и молекулярного зонда отражены в таблице 2.
Расчет дизайна праймеров и зонда проводили на основании нуклеотидных последовательностей Н-гена CDV производственного штамма «Rockborn» и других изолятов, опубликованных в базах данных GenBank [10].
В качестве гомологичных участку Н-гена возбудителя чумы плотоядных олигонуклеотидов используют:
CDV-F7173-праймер  (в позиции №2 внесена замена R, которая предполагает альтернативное применение двух вариантов нуклеотидов: G/A - purine),
(в позиции №2 внесена замена R, которая предполагает альтернативное применение двух вариантов нуклеотидов: G/A - purine),
CDV-R7428-праймер  и CDV-P7232-зонд
и CDV-P7232-зонд  (фиг. 1) в концентрации 10 пМ на реакцию. Для элонгации применяют дезоксирибонуклеозидтрифосфаты с их концентрацией в реакционной смеси по 0,2 мМ. В качестве основы используют буферный раствор (5х), содержание которого составляет 20% от общего объема реакционной смеси. В смесь также добавляют хлорида магния до концентрации 2,5 мМ. В качестве катализаторов обратной транскрипции и ГТЦР в режиме реального времени применяют следующие ферменты: MMLV-обратная транскриптаза (1 ед.) и Tag ДНК-зависимую ДНК-полимеразу (1 ед.). Элюаты РНК CDV каждого образца добавляют к реакционной смеси по 5 мкл. Итоговый объем смеси для проведения одной реакции составляет 25 мкл.
(фиг. 1) в концентрации 10 пМ на реакцию. Для элонгации применяют дезоксирибонуклеозидтрифосфаты с их концентрацией в реакционной смеси по 0,2 мМ. В качестве основы используют буферный раствор (5х), содержание которого составляет 20% от общего объема реакционной смеси. В смесь также добавляют хлорида магния до концентрации 2,5 мМ. В качестве катализаторов обратной транскрипции и ГТЦР в режиме реального времени применяют следующие ферменты: MMLV-обратная транскриптаза (1 ед.) и Tag ДНК-зависимую ДНК-полимеразу (1 ед.). Элюаты РНК CDV каждого образца добавляют к реакционной смеси по 5 мкл. Итоговый объем смеси для проведения одной реакции составляет 25 мкл.
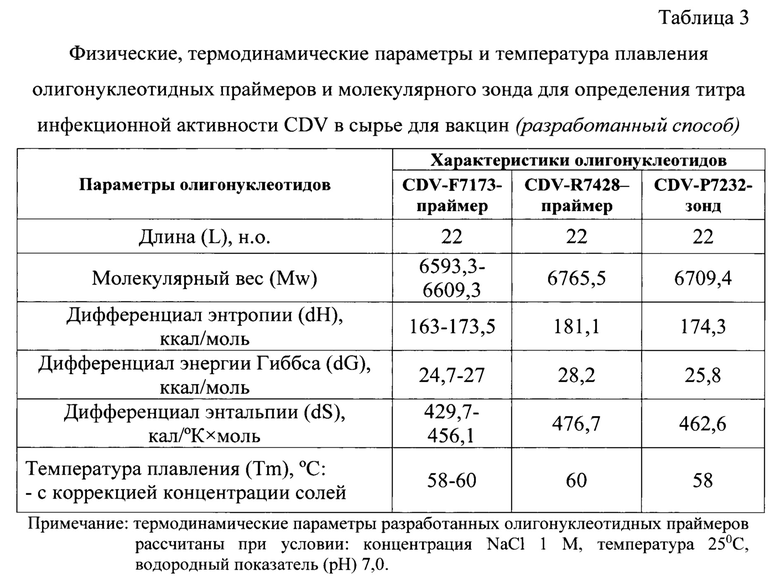
Олигонуклеотиды для кДНК-матрицы подбирали в соответствии с рядом общих правил, которые отражены в работах В. Deiman и R. Sooknanan [11]. Длины CDV-F7173, CDV-R7428 и CDV-P7232 составляют по 22 н.о. каждый, что соответствует требованиям. Молекулярный вес олигонуклеотидов прямого, обратного праймеров и зонда равен 6593,3-6609,3; 6765,5; 6709,4, соответственно. Праймеры и зонд очищены в полиакриламидном геле и с помощью высокоэффективной жидкостной хроматографии, соответственно. Нуклеотидная последовательность зонда не комплементарна олигонуклеотидным праймерам. Отсутствуют 4 и более подряд одинаковых нуклеотидов в цепи праймеров и зонда. Флуорофор FAM присоединен к 5'-концу, а гаситель флуоресценции RTQ1 - к 3'-концу. Данные условия соответствуют требованиям, предъявляемым к олигонуклеотидным праймерам и молекулярному зонду, которые участвуют в ПНР в режиме реального времени. В качестве флуоресцентного красителя был выбран FAM с длиной волны максимальной флуоресценцией 520 нм. Для тушения свечения использовали гаситель флуоресценции RTQ1 с длиной волны максимального поглощения при 520 нм и возможном диапазоне гашения 470-570 нм. Иными словами, была выбрана подходящая пара «флуорофор-гаситель».
При анализе нуклеотидных последовательностей олигонуклеотидов установили, что для праймеров и зонда не характерно образование «шпилек», а также не выявлено 3'-комплементарности и сайтов, отжигающих сами на себя. Расчет вероятности образования «шпилек» и димеров олигонуклеотидов проводили при условии, что минимальное количество пар оснований, необходимых для димеризации, - 5, а для образования «шпилек», - 4.
Проведено определение температур плавления (Tm) для олигонуклеотидных праймеров и зонда. Точное определение температуры плавления играет важную роль в молекулярно-биологических исследованиях, в том числе при подборе олигонуклеотидных праймеров и зонда для ГТЦР в режиме реального времени кДНК CDV. Для оценки температуры плавления олигонуклеотидов применяли метод, учитывающий концентрации ионов K+ и диметилсульфооксида (DMSO) с применением формулы [11].
Физические, термодинамические константы и расчет температур плавления разработанных олигонуклеотидных праймеров и молекулярного зонда представлены в таблице 3. Из нее следует, что энтропия, энергия Гиббса и энтальпия для прямого праймера CDV-F7173 составили 163-173,5 ккал/моль, 24,7-27 ккал/моль, 429,7-456,1 кал/(°K×моль), соответственно. Энтропия, энергия Гиббса и энтальпия для CDV-R7428 составили 181,1 ккал/моль, 28,2 ккал/моль, 476,7 кал/(°K×моль), соответственно. Энтропия, энергия Гиббса и энтальпия для CDV-P7232 - 174,3 ккал/моль, 25,8 ккал/моль, 462,6 кал/(°K×моль), соответственно. Данные значения необходимы для расчета температур плавления представленных олигонуклеотидов. Tm при использовании алгоритма ближайших соседей для прямого, обратного праймеров и зонда составили 58-60, 60, 58°С, соответственно.
Экспериментально было выявлено, что оптимальная температура отжига рассматриваемых олигонуклеотидов составляет 60°С.Для проведения ПЦР в режиме реального времени было решено проводить гибридизацию праймеров и зонда с участком Н-гена CDV при температуре 60°С.
Последовательности разработанных праймеров и молекулярного зонда проверили на наличие нежелательных совпадений с другими последовательностями нуклеиновых кислот с использованием Банка данных последовательности нуклеиновых кислот возбудителя чумы плотоядных. Последовательности праймеров и зонда также проанализировали на наличие внутренних вторичных структур с помощью программы сворачивания нуклеиновых кислот с помощью программы Mfold. Выявлено, что для разработанных олигонуклеотидов нежелательных совпадений с другими последовательностями нуклеиновых кислот, а также наличия внутренних вторичных структур не обнаружено.
Постановку реакции осуществляли в детектирующем термоциклере любой марки при температурных и временных параметрах, сведения о которых представлены в таблице 4.
Обратную транскрипцию проводят при температуре 42°С в течение 30 мин. Перед проведением ПЦР в режиме реального времени осуществляют предварительный прогрев смеси при температуре 95°С в течение 5 мин. для активации фермента Taq ДНК-полимеразы и инактивации MMLV-ревертазы.
ПЦР в режиме реального времени включает в себя 3 подэтапа: денатурацию, отжиг олигонуклеотидов, элонгацию. Денатурацию осуществляют при температуре 95°С в течение 20 с, отжиг олигонуклеотидов и элонгацию совмещают для сокращения времени проведения анализа и проводят при температуре 60°С в течение 40 с (таблица 4).
Результаты реакции амплификации в режиме реального времени анализируют, оценивая и сравнивая графики накопления флуоресцентного сигнала по значениям циклов количественной оценки Cq, определенных с помощью пересечения пороговой линии и логарифмическим отображением функции  Учет результатов в реакции происходит на каждом цикле. Флуориметр определяет уровень флуоресценции и строит кинетическую кривую в координатах: уровень флуоресценции - цикл реакции амплификации. В случае присутствия в исследуемой пробе специфической кДНК-матрицы кинетическая кривая имеет экспоненциальную зависимость (график представлен в виде логистической кривой). Положительными считаются пробы, которым соответствуют сигмоиды, полученные при анализе флуоресценции красителя FAM, входящего в состав молекулярного зонда. Пробы считаются отрицательными, если при их анализе отсутствует логистическая кривая в виде графика, приближенного к экспоненте.
Учет результатов в реакции происходит на каждом цикле. Флуориметр определяет уровень флуоресценции и строит кинетическую кривую в координатах: уровень флуоресценции - цикл реакции амплификации. В случае присутствия в исследуемой пробе специфической кДНК-матрицы кинетическая кривая имеет экспоненциальную зависимость (график представлен в виде логистической кривой). Положительными считаются пробы, которым соответствуют сигмоиды, полученные при анализе флуоресценции красителя FAM, входящего в состав молекулярного зонда. Пробы считаются отрицательными, если при их анализе отсутствует логистическая кривая в виде графика, приближенного к экспоненте.
Устанавливают зависимость между циклом количественной оценки и титром инфекционной активности CDV в сырье для вакцин в процессе построения логарифмической функции. Оценивают величину эффективности реакции амплификации (Е) по формуле:
где, k - угловой коэффициент в зависимости Ct=-k×lg TFCV+b, а также достоверность аппроксимации (R2). На основе разработанной модели рассчитывают значение титра инфекционной активности CDV в сырье для вакцин.
Сущность предлагаемого изобретения пояснена примерами его использования, которые не ограничивают объем изобретения.
Пример 1. Выявление зависимости титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцины и цикла количественной оценки ПЦР в виде логарифмической функции.
Для выявления зависимости титра инфекционной активности CDV в сырье для вакцин и цикла количественной оценки ПЦР подготавливали контрольную панель стандартов вируса чумы плотоядных штамма «Rockborn», в качестве которых использовали лиофилизированные суспензии вируса указанного штамма с титрами: 1,0; 2,0; 3,0; 4,0; 5,0; 6,0; 7,0; 8,0 lg ТЦД50/см3, которые растворяли в 1/15 М фосфатном буферном растворе до требуемого объема. В качестве отрицательных контролей применяли суспензию клеток Vero, не контаминированную микроорганизмами.
Из всех стандартных положительных образцов и отрицательных контролей, а также тестируемых проб выделяли РНК в соответствии с методикой, представленной выше.
Проводили обратную транскрипцию и ПЦР в режиме реального времени для исследования стандартов, как описано выше. Результаты реакции анализировали, оценивая и сравнивая графики накопления флуоресцентного сигнала по значениям циклов количественной оценки Cq, определенных с помощью пересечения пороговой линии и логарифмическим отображением функции  Устанавливали зависимость между циклом количественной оценки и титром инфекционной активности CDV в сырье для вакцин в процессе построения логарифмической функции. Полученные данные отражены на фиг. 2 и выражены в виде логарифмической функции:
Устанавливали зависимость между циклом количественной оценки и титром инфекционной активности CDV в сырье для вакцин в процессе построения логарифмической функции. Полученные данные отражены на фиг. 2 и выражены в виде логарифмической функции:
с высокой достоверностью аппроксимации (R2=0,9965) и эффективностью амплификации 99,64%. Предложенная модель позволяет опосредованного определять титр инфекционной активности вируса чумы плотоядных в сырье для вакцин с помощью разработанного способа.
Пример 2. Применение способа опосредованного определения титра инфекционной активности CDV в сырье для вакцин с использованием метода обратной транскрипции и реакции амплификации в режиме реального времени.
В исследовании использовали 6 суспензий культурального CDV с титрами инфекционной активности 6,50; 6,75; 7,00; 7,25; 7,50; 7,75 lg ТЦД50/см3, соответственно (пробы №1-6). В качестве положительного контроля применяли суспензию культурального CDV с титром инфекционной активности 7,00 lg ТЦД50/см3. В качестве отрицательных контролей применяли суспензию клеток Vero, не зараженную микроорганизмами. Испытуемые пробы и контрольные образцы исследовали в 7 повторностях. Этап элюирования РНК, и постановку обратной транскрипции и ПНР в режиме реального времени проводили, как описано выше.
Средние значения пороговых циклов амплификации для проб №1-6 составляли 10,01±0,01, 9,14±0,02, 8,33±0,01, 7,50±0,01, 6,67±0,01, 5,80±0,01, соответственно. Пользуясь разработанной логарифмической функцией:
с высокой достоверностью аппроксимации (R2=0,9965) и эффективностью амплификации 99,64%, рассчитали средние значения титра инфекционной активности CDV для проб №1-6, которые составили 6,49; 6,75; 6,99; 7,24; 7,50; 7,76 lg ТЦД50/см3, соответственно. Для положительного контроля значение порогового цикла амплификации составило 8,32±0,01, что соответствовало титру инфекционной активности CDV, равному 7,00 lg ТЦД50/см3. Для отрицательных контролей экспоненциальные графики не были сформированы, что означало отсутствие CDV в данных образцах. Полученные результаты свидетельствовали о высокой степени точности разработанного способа опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцины с помощью метода обратной транскрипции и реакции амплификации в режиме реального времени.
Иными словами, разработанный способ позволяет рассчитывать титр инфекционной активности CDV в сырье для вакцин методом ПЦР в режиме реального времени.
Пример 3. Выявление степени достоверности определения титра инфекционной активности CDV в сырье для вакцин с применением разработанного способа.
Для анализа использовали 280 суспензий культурального CDV с титрами инфекционной активности от 1,00 до 8,00 lg ТЦД50/см3. В качестве положительного контроля применяли суспензию культурального CDV с титром инфекционной активности вируса 7,00 lg ТЦД50/см3. В качестве отрицательных контролей применяли суспензию клеток Vero, не зараженную микроорганизмами. Испытуемые пробы и контрольные образцы исследовали в трех повторностях. Этапы экстрагирования нуклеиновых кислот и постановку обратной транскрипции и ГЩР в режиме реального времени проводили, как отражено выше. Результаты анализа представлены в таблице 5.
Интерпретацию результатов проводили, пользуясь разработанной логарифмической функцией:
(R2=0,9965, Е=99,64%) с получением значений TCDV для каждой из 280 проб. Для положительного контроля значение порогового цикла амплификации составило 8,32±0,01, что соответствовало титру инфекционной активности CDV, равному 7,00 lg ТЦД50/см3. Для отрицательных контролей экспоненциальные графики не были сформированы, что означало отсутствие CDV в данных образцах.
Анализируемые пробы и контроли также тестировали в ПЦР в режиме реального времени. Выявили, что данные, полученные с помощью разработанного способа, коррелировали с со значениями стандартов на 99,75-100,00% для 8,0-6,0 lg ТЦДзо/см3 (n=70), на 98,33-99,85% для 5,9-4,0 lg ТЦД50/см3 (n=70), на 96,33-99,56% для 3,9-1,5 lg ТЦД50/см3 (n=70), на 96,08-99,54% для 1,4-1,0 lg ТЦД50/см3 (n=70). Полученные результаты свидетельствовали о высокой степени точности разработанного способа опосредованного определения титра инфекционной активности CDV в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени.
Иными словами, разработанный способ позволяет с высокой степенью достоверности оценивать титр инфекционной активности CDV в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени.
Пример 4. Определение аналитической чувствительности способа опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени.
При определении аналитической чувствительности способа опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени подготавливали серию стандартов CDV разных штаммов с титрами инфекционной активности, равными 1,00-8,00 lg ТЦД50/см3 с шагом 0,1 lg ТЦД50/см3. Контрольные образцы тестировали в 4 повторностях. Этапы элюирования нуклеиновой кислоты и постановку обратной транскрипции и ПЦР в режиме реального времени, как описано выше. Выявлено, что с достоверностью 96,08-100,00% разработанным способом определены титры инфекционной активности возбудителя чумы плотоядных со значениями от 1,0 до 8,0 lg ТЦД50/см3. При изготовлении вакцин применяют вируссодержащее сырье только с титрами инфекционной активности вируса>6,00 lg ТЦД50/см3. Для данных образцов достоверность определения титра инфекционной активности возбудителя чумы плотоядных составляла 99,75-100,00%.
Следовательно, аналитическая чувствительность тест-системы, разработанной для способа опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени составляет не менее 1,0 lg ТЦД50/см3 с достоверностью результатов исследования не менее 96,08%.
Пример 5. Исследование специфичности способа опосредованного определения титра инфекционной акщивности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени.
При анализе специфичности способа опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени, исследовали суспензии CDV, а также штамма «РВ-97» вируса бешенства, штамма «Грей» возбудителя парвовирусного энтерита, штамма «Рич» возбудителя коронавирусного энтерита собак, штамма «ВГНКИ» аденовируса 1 серотипа. Количество инфекционных доз вирусов в суспензиях составлял не менее 6,0 lg ТЦД50/см3. Исследования проводили в 5 повторностях.
Этап элюирования РНК, постановку обратной транскрипции, ПЦР в режиме реального времени проводили, как описано выше. Для проб, содержащих другие вирусы, не наблюдалось формирования графиков экспоненты, и они не выходили за пороговый уровень флуоресцентного сигнала (0,01 у.е.) (таблица 6). Таким образом, разработанный способ является специфичным по отношению к возбудителю чумы плотоядных и может быть использован для его количественного определения.
Основными преимуществами предлагаемого изобретения является возможность исключить вероятность контаминации в ходе реакции и субъективность при учете результатов анализа; снизить время проведения анализа вируссодержащего сырья для вакцин до 3,5 ч; увеличить специфичность и чувствительность анализа по определению титра инфекционной активности возбудителя чумы плотоядных. В предлагаемом изобретении между титром инфекционной активности вирусом чумы плотоядных (TCDV) и циклом количественной оценки (Cq) установлена зависимость, отраженная в виде логарифмической функции:
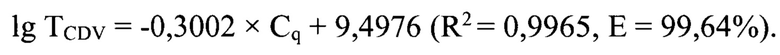
Разработанная математическая модель дает возможность определять титр инфекционной активности CDV в сырье при производстве вакцин.
Предлагаемое изобретение позволяет быстро и с высокой степенью достоверности рассчитывать титр инфекционной активности CDV в сырье для вакцин на основе разработанного способа с применением оригинальных специфических олигонуклеотидных праймеров и зонда для участка Н-гена CDV и разработанной логарифмической модели.
Источники информации, принятые во внимание при составлении описания изобретения к заявке на выдачу патента РФ на изобретение «Способ опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени»:
1. Elia G, et al. Virological and serological findings in dogs with naturally occurring distemper. J. Virol. Methods. 2015;213:123-130. doi: 10.1016/j.j viromet.2014.12.004.
2. Beineke A, Puff C, Seehusen F, Baumgartner W. Pathogenesis and immunopathology of systemic and nervous canine distemper. Vet. Immunol. Immunopathol. 2009;127:1-18. doi: 10.1016/j.vetimm.2008.09.023.
3. Rendon-Marin S, Budaszewski RF, Canal CW, Ruiz-Saenz J. Tropism and molecular pathogenesis of canine distemper virus. Virol. J. 2019; 16:30. doi: 10.1186/sl2985-019-1136-6.
4. Han G-Z, Liu X-P, Li S-S. Cross-species recombination in the haemagglutinin gene of canine distemper virus. Virus Res. 2008;136:198-201. doi: 10.1016/j.virusres.2008.04.022.
5. Lempp C, et al. New aspects of the pathogenesis of canine distemper leukoencephalitis. Viruses. 2014;6:2571-2601. doi: 10.3390/v6072571.
6. Anis E, Holford AL, Galyon GD, Wilkes RP. Antigenic analysis of genetic variants of canine distemper virus. Vet. Microbiol. 2018;219:154-160. doi: 10.1016/j.vetmic.2018.03.014.
7. Carina R, et al. Virus isolation and full-length genome sequencing of a representative canine distemper virus wild type strain of the South America 2 clade. J. Virol. Methods. 2020;279:113857.
8. Freitas LA, Leme RA, Saporiti V, Alfieri AA, Alfieri AF. Molecular analysis of the full-length F gene of Brazilian strains of canine distemper virus shows lineage co-circulation and variability between field and vaccine strains. Virus. Res. 2019;264:8-15. doi: 10.1016/j.virusres.2019.02.009.
9. Sarute N, et al. Molecular typing of canine distemper virus strains reveals the presence of a new genetic variant in South America. Virus Gene. 2014;48:474-478. doi: 10.1007/s1 1262-014-1054-z.
10. GenBank. [Электронный ресурс] / URL: http://www.ncbi.nlm.nih.gov. (Дата обращения: 07.04.2023).
11. Sooknanan R., van Gemen В., Malek L. Nucleis acid sequence-based amplification // Molecular methods for virus detection-London: Academic press, 1995. - P. 261-285.



--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="CDV titre.xml"
softwareName="WIPO Sequence" softwareVersion="2.1.2"
productionDate="2023-05-31">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>0</ApplicationNumberText>
<FilingDate>2023-05-31</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>507</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>0</ApplicationNumberText>
<FilingDate>2023-05-31</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">ФГБУ "Федеральный центр охраны
здоровья животных" (ФГБУ "ВНИИЗЖ")</ApplicantName>
<ApplicantNameLatin>FGBI "ARRIAH"</ApplicantNameLatin>
<InventorName languageCode="ru">Доронин Максим
Игоревич</InventorName>
<InventorNameLatin>Doronin Maksim Igorevich </InventorNameLatin>
<InventionTitle languageCode="ru">Способ опосредованного определения
титра инфекционной активности возбудителя чумы плотоядных в сырье для
вакцин методом обратной транскрипции и реакции амплификации в режиме
реального времени</InventionTitle>
<SequenceTotalQuantity>2</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>607</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..607</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q3">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>CDV</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MLSYQDKVGAFYKDNARANSTKLSLVTEEHGGRRPPYLLFVLLILLVGI
LALLAITGVRFHQVSTSNMEFSRLLKEDMEKSEAVHHQVIDVLTPLFKIIGDEIGLRLPQKLNEIKQFIL
QKTNFFNPNREFDFRDLHWCINPPSKVKVNFTNYCESIGIRKAIASAANPILLSALSGGRSDIFPPHRCS
GATTSVGKVFPLSVSLSMSLISRTSEIINMLTAISDGVYGKTYLLVPDDIEREFDTQEIRVFEIGFIKRW
LNDMPLLQTTNYMVLPENSKAKVCTIAVGELTLASLCVEESTVLLYHDSSGSQDGILVVTLGIFWATPMD
HIEEVIPVAHPSMEKIHITNHRGFIKDSIATWMVPALASEKQEEQKGCLESACQRKTYPMCNQTSWEPFG
GRQLPSYGRLTLPLDASVDLQLNISFTYGPVILNGDGMDYYESPLLNSGWLTIPPKNGTISGLINKAGRG
DQFTVLPHVLTFAPRESSGNCYLPIQTSQIIDRDVLIESNIVVLPTQSFRYVIATYDISRSDHAIVYYVY
DPIRTISYTHPFRLTTKGRPDFLRIECFVWDNNLWCHQFYRFEADIANSTTSVENLVRIRFSCNRSKP</
INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>1821</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1821</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>genomic DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>CDV</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atgctctcctaccaagacaaggtgggtgccttctacaaggataatgcaa
gagccaattcaaccaagctgtccttagtgacagaagaacatgggggcaggagaccaccttatttgttgtt
tgtccttctcatcttattggttggaatcctggccttgcttgctatcactggagttcgatttcaccaagta
tcaactagcaatatggaatttagcagattgctgaaagaggatatggagaaatcagaggccgtacatcacc
aagtcatagatgtcttgacaccgctcttcaagattattggagatgagattgggttacggttgccacaaaa
gctaaacgagatcaaacaatttatccttcaaaagacaaatttcttcaatccgaacagagaattcgacttc
cgcgatctccactggtgcattaacccgcctagtaaggtcaaggtgaattttactaattactgtgagtcaa
ttgggatcagaaaagctattgcatcggcagcaaatcctatccttttatcagccctatctgggggcagaag
tgacatattcccaccacacagatgcagtggagctactacttcagtaggcaaagttttccccctatcagtc
tcattatccatgtctttgatctcaagaacctcagagataatcaatatgctgaccgctatctcagacggcg
tgtatggcaaaacttacttgctagtgcctgatgatatagaaagagagttcgacactcaagagattcgagt
ctttgaaatagggttcatcaaaaggtggctgaatgacatgccattactccaaacaaccaactatatggta
ctcccggagaattccaaagccaaggtatgtactatagcagtgggtgagttgacactggcttccttgtgtg
tagaagagagcactgtattattatatcatgacagcagtggttcacaagatggtattctagtagtgacact
ggggatattttgggcaacacctatggatcacattgaggaagtgatacctgtcgctcacccatcaatggag
aaaatacatataacaaaccaccggggttttataaaagattcaattgcaacctggatggtgcctgccctgg
cctctgagaaacaagaagaacaaaaaggttgtctggagtcagcttgtcaaagaaaaacctaccccatgtg
caaccaaacgtcatgggaacccttcggaggaagacagttgccatcttatgggcggttgacattacctcta
gatgcaagtgttgaccttcaacttaacatatcgttcacatacggtccggttatactgaatggagatggta
tggattattatgaaagcccacttttgaactccggatggcttaccattcctcccaaaaacggaacaatctc
tggattgataaacaaagcaggtagaggagaccagttcactgtactcccccatgtgttaacatttgcgccc
agggaatcaagtggaaattgttatttacctattcaaacatctcaaattatagatagagatgtcctcattg
agtccaatatagtggtgttgcctacacagagttttagatatgtcatagcaacgtatgacatatcacgaag
tgatcatgcgattgtttattatgtttatgacccaatccggacgatttcttatacgcacccatttagacta
actaccaagggtagacctgatttcctaaggattgaatgttttgtgtgggataacaatttgtggtgtcacc
aattttacagattcgaggctgacatcgccaactctacaaccagtgttgagaatttagtccgtataagatt
ctcatgtaaccgttcaaaacct</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ определения титра инфекционной активности возбудителя чумы плотоядных животных в сырье для изготовления культуральных инактивированных вакцин | 2024 |
|
RU2831062C1 |
| Способ опосредованного определения концентрации вирионов возбудителя чумы плотоядных животных в сырье для вакцин с помощью двойного математического дифференциала для точки crossing point сигмоиды при амплификации участка гена CDVgp1 | 2024 |
|
RU2840386C1 |
| Способ опосредованного определения концентрации рибонуклеопротеина возбудителя чумы плотоядных в сырье для вакцин по данным цикла количественной оценки при амплификации N-гена | 2023 |
|
RU2829837C1 |
| Способ дифференциации штамма "Рокборн" вируса чумы плотоядных от других штаммов ВЧП | 2024 |
|
RU2840521C1 |
| Способ опосредованного определения титра инфекционной активности вакцинного штамма "РВ-97" вируса бешенства в сырье для вакцины на основе анализа кривых плавления ампликонов высокого разрешения с применением интеркалирующего красителя третьего поколения EvaGreen | 2023 |
|
RU2830983C1 |
| Способ опосредованного определения титра инфекционной активности калицивируса кошек в сырье для вакцины методом ПЦР в режиме реального времени | 2023 |
|
RU2809221C1 |
| Способ опосредованного определения титра инфекционной активности мастаденовируса собак в сырье для вакцин методом ПЦР в режиме реального времени | 2023 |
|
RU2808585C1 |
| Способ опосредованного определения титра инфекционной активности вируса инфекционного гепатита собак генотипа CAV-1 в сырье для культуральных вакцин с помощью полимеразной цепной реакции в режиме реального времени по циклу количественной оценки Cq | 2023 |
|
RU2812441C1 |
| Способ опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в неинактивированном сырье для культуральных вакцин с помощью реакции амплификации целевого участка UL26.5-гена вирусной ДНК | 2024 |
|
RU2834248C1 |
| Способ опосредованного определения титра инфекционной активности калицивируса кошек в неинактивированном сырье для культуральных вакцин с помощью количественного учета реакции амплификации участка ORF1-гена вирусной РНК | 2023 |
|
RU2821027C1 |

Изобретение относится к области биотехнологии. Описан способ опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени с применением разработанной логарифмической функции зависимости величины цикла количественной оценки и титра инфекционной активности возбудителя чумы плотоядных животных. Основными преимуществами предлагаемого изобретения является возможность исключить вероятность контаминации в ходе реакции и субъективность при учете результатов анализа; снизить время проведения анализа вируссодержащего сырья для вакцин до 3,5 ч; увеличить специфичность и чувствительность анализа по определению титра инфекционной активности возбудителя чумы плотоядных. В предлагаемом изобретении между титром инфекционной активности вируса чумы плотоядных и циклом количественной оценки установлена зависимость, отраженная в виде логарифмической функции, которая дает возможность определять титр инфекционной активности CDV в сырье для производства вакцин. Предлагаемое изобретение позволяет быстро и с высокой степенью достоверности рассчитывать титр инфекционной активности CDV в сырье для вакцин на основе разработанного способа с применением оригинальных специфических олигонуклеотидных праймеров и зонда для участка Н-гена CDV и разработанной логарифмической модели. 1 з.п. ф-лы, 2 ил., 6 табл., 5 пр.
1. Способ опосредованного определения титра инфекционной активности возбудителя чумы плотоядных в сырье для вакцин методом обратной транскрипции и реакции амплификации в режиме реального времени, включающий следующие стадии:
- выделение РНК возбудителя чумы плотоядных с помощью коллоидных частиц поверхностно-модифицированного Fe3O4 с инертной поверхностью за счет обработки 50%-ным раствором ПЭИ и диаметром 180-200 нм;
- проведение обратной транскрипции и реакции амплификации с детекцией флуоресцентного сигнала логистических кривых исследуемых образцов с использованием разработанных оригинальных олигонуклеотидов:
CDV-F7173-праймер с дизайном 5'-TRCTGTTTGTCCTTCTCATCCT-3',
CDV-R7428-праймер с дизайном 5'- TCCCTGTTCGGATTGAAGAAGT-3',
CDV-P7232-зонд с дизайном 5'-FAM-GAGTTCGATTTCACCAAGTATC-RTQ1-3', которые рассчитаны на участок Н-гена возбудителя чумы плотоядных;
- расчет цикла количественной оценки по данным реакции амплификации в режиме реального времени;
- определение титра инфекционной активности культурального вируса чумы плотоядных в сырье для вакцины на основе разработанной логарифмической регрессионной функции: lg TCDV=-0,3002×Cq+9,4976 с достоверностью аппроксимации 0,9965 и эффективностью амплификации 99,64%.
2. Способ по п. 1, отличающийся тем, что время проведения анализа сокращается до 3,5 ч, аналитическая чувствительность составляет не менее 1,0 lg ТЦД50/см3.
| СТРУННЫЙ ОСЦИЛЛЯТОР | 1928 |
|
SU18355A1 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИГЕНА ВИРУСА ЧУМЫ ПЛОТОЯДНЫХ ДЛЯ ПРИГОТОВЛЕНИЯ ЭРИТРОЦИТАРНОГО ДИАГНОСТИКУМА | 2022 |
|
RU2787396C1 |
| Льнотеребильная машина | 1931 |
|
SU27240A1 |
| А.Л | |||
| Елаков | |||
| Разработка метода контроля антигенной активности вакцин против чумы плотоядных, Автореферат на соискание ученой степени кандидата наук, Москва 2000. | |||

