ОБЛАСТЬ ТЕХНИКИ
Данное изобретение относится к микроорганизму, обладающему усиленной способностью продуцировать L-глутамин, и способу получения L-глутамина с его применением.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
L-глутамин представляет собой аминокислоту, широко применяющуюся в различной промышленности, включая фармацевтическую, косметическую и диетические продукты. Он широко применяется в качестве терапевтического средства при нарушениях пищеварения, для улучшения функции печени, улучшения функции головного мозга, улучшения иммунитета, в качестве терапевтического средства при язве желудка, терапевтического средства при алкоголизме, увлажняющего средства в косметических продуктах, пищевой добавки при физических нагрузках и питательной добавки для пациентов.
Соответственно, Corynebacterium glutamicum и Escherichia coli применяются для получения L-глутамина с применением микроорганизмов. В пути биосинтеза L-глутамина α-кетоглутаровая кислота, образующаяся в результате гликолиза и ЦТК (цикла трикарбоновых кислот) применяется в качестве предшественника для образования L-глутамина при помощи глутаматдегидрогеназы, и в итоге L-глутамин образуется в реакции, катализируемой глутаминсинтетазой (Production of glutamate and glutamate-related amino acids: Molecular Mechanism Analysis and Metabolic Engineering, Amino acid Biosynthesis-pathways, regulation and metabolic engineering pp.1-38).
Однако, в соответствии с увеличением потребности в L-глутамине по-прежнему актуальны исследования для эффективного увеличения способности продуцировать L-глутамин.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Техническая задача
В результате попыток увеличить способность продуцировать L-глутамин авторы данного изобретения обнаружили, что L-глутамин с высокой эффективностью продуцируется в микроорганизмах, содержащих фосфоенолпируваткарбоксикиназу и, таким образом, завершили данное изобретение.
Техническое решение
Задачей данного изобретения является предоставление микроорганизма рода Corynebacterium, обладающего ослабленной активностью фосфоенолпируваткарбоксикиназы и способностью продуцировать L-глутамин.
Другой задачей данного изобретения является предоставление способа получения L-глутамина, включающего культивирование в среде микроорганизма рода Corynebacterium, обладающего ослабленной активностью фосфоенолпируваткарбоксикиназы и способностью продуцировать L-глутамин.
Еще одной задачей данного изобретения является предоставление способа получения микроорганизма для получения L-глутамина, включающего ослабление фосфоенолпируваткарбоксикиназы.
Еще одной задачей данного изобретения является предоставление композиции для получения L-глутамина, включающей микроорганизм, в котором ослаблена фосфоенолпируваткарбоксикиназа; среды, в которой культивировали микроорганизм, или их комбинации.
Еще одной задачей данного изобретения является предоставление применения микроорганизма рода Corynebacterium для получения L-глутамина, в котором ослаблена фосфоенолпируваткарбоксикиназа.
Полезные эффекты
Микроорганизм по данному изобретению, содержащий ослабленную фосфоенолпируваткарбоксикиназу, может продуцировать L-глутамин с высокой эффективностью. Полученный L-глутамин можно применять в различных продуктах, включая корма или кормовые добавки для животных, а также продукты питания или пищевые добавки для человека и фармацевтические средства.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Каждое из описаний и воплощений, изложенных в данном изобретении, также может быть применено к другим описаниям и воплощениям. Таким образом, все комбинации различных элементов, изложенных в данном изобретении, входят в объем данного изобретения. Более того, объем данного изобретения не ограничивается конкретным описанием, приведенным ниже.
Кроме того, специалисты в данной области поймут или смогут найти, проведя рутинные эксперименты, множество эквивалентов частных воплощений данного изобретения, описанных в данном документе. Кроме того, такие эквиваленты следует считать входящими в объем данного изобретения. В данном описании дополнительно процитирован ряд публикаций и патентных документов, и приведены ссылки на них. Содержание процитированных публикаций и патентных документов включено во всей полноте путем ссылки для более точного описания уровня техники, к которой относится данное изобретение, и содержания данного изобретения.
В аспекте данного изобретения для решения задач предложен микроорганизм рода Corynebacterium, обладающий ослабленной активностью фосфоенолпируваткарбоксикиназы и способностью продуцировать L-глутамин.
В данном описании термин «фосфоенолпируваткарбоксикиназа (PEPCK; ЕС 4.1.1.32)» относится к ферменту, который задействован в процессе глюконеогенеза и конвертирует оксалоацетат в фосфоенолпируват и диоксид углерода в присутствии ЕТФ (гаммаглутаминтрансфераза). В данном изобретении фосфоенолпируваткарбоксикиназа может использоваться взаимозаменяемо с PEPCK.
Аминокислотная последовательность PEPCK может быть получена из известной базы данных, такой как NCBI Genebank.
Например, PEPCK по данному изобретению может представлять собой белок, происходящий из микроорганизма рода Corynebacterium. Более конкретно, PEPCK по данному изобретению может происходить из Corynebacterium glutamicum, Corynebacterium deserti, Corynebacterium crudilactis, Corynebacterium efficiens, Corynebacterium callunae и тому подобных, без ограничения.
PEPCK по данному изобретению может представлять собой эндогенный белок микроорганизма рода Corynebacterium по данному изобретению.
В данном изобретении PEPCK по данному изобретению может иметь, содержать или состоять из аминокислотной последовательности, приведенной в SEQ ID NO: 1, или может по существу состоять из аминокислотной последовательности.
В данном изобретении аминокислотная последовательность SEQ ID NO: 1 может включать аминокислотную последовательность, гомологичную или идентичную ей по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5%, 99,7% или 99,9%. Кроме того, очевидно, что белок, имеющий аминокислотную последовательность с делецией модификацией, заменой, консервативной заменой или добавлением некоторых последовательностей также входит в объем данного изобретения, при условии, что он имеет аминокислотную последовательность, которая обладает указанной гомологией или идентичностью, и демонстрирует эффективность, соответствующую таковой белка, включающего аминокислотную последовательность SEQ ID NO: 1.
Примеры этого включают случай наличия последовательности вставок или делеций последовательности, мутаций естественного происхождения, молчащих мутаций или консервативных замен на N-конце, С-конце и/или внутри аминокислотной последовательности, которые не изменяют функцию белка по данному изобретению.
«Консервативная замена» относится к замене одной аминокислоты на другую аминокислоту, имеющую схожие структурные и/или химические свойства. Такая аминокислотная замена, как правило, может возникать на основании сходства в полярности, заряде, растворимости, гидрофобности, гидрофильности и/или амфипатических свойств остатков. Как правило, консервативные замены могут оказывать незначительное влияние или не оказывать влияния на активность белка или полипептида.
В данном описании термин «гомология» или «идентичность» относится к степени сходства двух заданных аминокислотных последовательностей или нуклеотидных последовательностей и может выражаться в процентах. Термины гомология и идентичность зачастую используются взаимозаменяемо.
Гомология или идентичность консервативных полинуклеотидов или полипептидов определяется при помощи стандартных алгоритмов выравнивания и при этом могут использоваться значения штрафов за открытие гэпа, установленных в используемой программе по умолчанию. Существенным образом гомологичные или идентичные последовательности обычно способны гибридизоваться в условиях умеренной или высокой жесткости со всей последовательностью или ее частью. Очевидно, что гибридизация также включает гибридизацию с полинуклеотидами, содержащими общие кодоны или кодоны с учетом вырожденности кодонов в полинуклеотидах.
Установить, обладают ли две любые полинуклеотидные или полипептидные последовательности гомологией, сходством или идентичностью, можно, например, при помощи известных компьютерных алгоритмов, таких как программа FASTA с использованием параметров по умолчанию, например, как описано Pearson et al., (1988) [Proc. Natl. Acad. Sci. USA 85]: 2444. В альтернативном варианте гомологию, сходство или идентичность можно устанавливать при помощи алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453), как осуществляется в программе Needleman пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16: 276-277) (версии 5.0.0 или более поздней) (включая пакет программ GCG (Devereux, J., et al., Nucleic Acids Research 12: 387 (1984)), BLASTP, BLASTN, FASTA (Atschul, [S.] [F.,] [ET AL, J MOLEC BIOL 215]: 403 (1990); Guide to Huge Computers, Martin J. Bishop, [ED.,] Academic Press, San Diego, 1994, and [CARILLO ETA/.](1988) SIAM J Applied Math 48: 1073). Например, гомологию, сходство или идентичность можно определять при помощи BLAST базы данных Национального Центра Биотехнологической Информации или ClustalW.
Гомологию, сходство или идентичность полинуклеотидов или полипептидов можно определять путем сравнения информации о последовательности, например, с применением компьютерной программы GAP, такой как у Needleman et al. (1970), J Mol Biol. 48: 443, например, как изложено Smith and Waterman, Adv. Appl. Math (1981) 2: 482. В заключение, программа GAP может определять гомологию, сходство или идентичность по значению, полученному путем деления общего количества символов в более короткой из двух последовательностей на количество одинаково расположенных символов (то есть нуклеотидов или аминокислот). Параметры по умолчанию для программы GAP могут включать (1) бинарную матрицу сравнения (содержащую значение 1 для идентичности и 0 для отсутствия идентичности) и взвешенную матрицу сравнения, предложенную Gribskov et al(1986) Nucl. Acids Res. 14: 6745 (или подстановочную матрицу EDNAFULL (EMBOSS версия NCBI NUC4.4), как изложено Schwartz and Dayhoff, eds., Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp.353-358 (1979); (2) штраф 3,0 для каждого гэпа и дополнительный штраф 0,10 для каждого символа в каждом гэпе (или штраф за открытие гэпа 10, штраф за продолжение гэпа 0,5); и (3) отсутствие штрафа за концевые гэпы.
PEPCK по данному изобретению может кодироваться геном рек. Ген рек по данному изобретению может включать все гены, которые, как известно, кодируют белки, обладающие активностью PEPCK.
В частности, ген рек по данному изобретению может представлять собой рек, происходящий из микроорганизма рода Corynebacterium. Более конкретно, ген рек по данному изобретению может представлять собой pck, происходящий из штамма Corynebacterium glutamicum. Например, ген pck по данному изобретению может представлять собой полинуклеотид, кодирующий WP 011015446.1, происходящий из Corynebacterium glutamicum АТСС13032.
В данном описании термин «полинуклеотид» представляет собой цепь ДНК или РНК определенной длины или более в виде полимера из нуклеотидов, в котором мономеры нуклеотидов соединены в длинную цепь ковалентными связями и, более конкретно, обозначает фрагмент полинуклеотида, кодирующий белок.
Полинуклеотид, кодирующий PEPCK по данному изобретению, может включать нуклеотидную последовательность, кодирующую аминокислотную последовательность, приведенную в SEQ ID NO: 1. В качестве примера данного изобретения полинуклеотид по данному изобретению может иметь или включать нуклеотидную последовательность SEQ ID NO: 2. Кроме того, полинуклеотид по данному изобретению может состоять или по существу состоять из последовательности SEQ ID NO: 2. В частности, PEPCK может кодироваться полинуклеотидом, приведенным в нуклеотидной последовательности SEQ ID NO: 2.
Можно осуществлять различные модификации кодирующей области полинуклеотида по данному изобретению в диапазоне, не изменяющем аминокислотную последовательность PEPCK с учетом вырожденности кодонов или предпочтения кодонов в организмах, которые предназначены для экспрессии PEPCK по данному изобретению. В частности, полинуклеотид по данному изобретению имеет или включает нуклеотидную последовательность, обладающую гомологией или идентичностью 70% или более, 75% или более, 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более или 98% или более с последовательностью SEQ ID NO: 2, или может состоять или по существу состоять из нуклеотидной последовательности, обладающей гомологией или идентичностью 70% или более, 75% или более, 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более или 98% или более с последовательностью SEQ ID NO: 2, без ограничения.
Кроме того, полинуклеотид по данному изобретению может включать в себя любой зонд, который может быть получен из известной генной последовательности, например, любой последовательности, которая может гибридизоваться в жестких условиях с последовательностью, комплементарной всей или части полинуклеотидной последовательности по данному изобретению, без ограничения. «Жесткие условия» относятся к условиям, которые позволяют специфически гибридизоваться полинуклеотидам. Такие условия, в частности, описаны в литературе (см. J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York, 9.50-9.51, 11.7-11.8). Их примеры включают условия, когда полинуклеотиды, обладающие гомологией или идентичностью, гибридизуются, например, полинуклеотиды, обладающие гомологией или идентичностью 70% или более, 75% или более, 76% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более или 99% или более, гибридизуются, а полинуклеотиды, обладающие гомологией или идентичностью, которая ниже указанной, не гибридизуются, или условия, при которых осуществляется отмывка один раз, в частности, от двух до трех раз, при концентрации соли и температуре, соответствующих 60°С, 1×SSC (раствор цитрата и хлорида натрия), 0,1% SDS, в частности, 60°С, 0,1×SSC, 0,1% SDS, более конкретно, 68°С, 0,1×SSC и 0,1% SDS, что представляет собой условие отмывки при стандартной гибридизации по Саузерну.
Для гибридизации необходимо, чтобы две нуклеиновые кислоты имели комплементарные последовательности, хотя возможны несовпадения между основаниями, в зависимости от строгости гибридизации. Термин «комплементарный» используется для описания взаимоотношения между нуклеотидными основаниями, способными гибридизоваться друг с другом. Например, в случае ДНК аденозин комплементарен тимину, а цитозин комплементарен гуанину. Соответственно, полинуклеотид по данному изобретению может также включать по существу схожие последовательности нуклеиновых кислот, а также выделенные фрагменты нуклеиновых кислот, комплементарные полной последовательности.
В частности, полинуклеотид, обладающий гомологией или идентичностью с полинуклеотидом по данному изобретению, можно обнаружить с применением условий гибридизации, включающих стадию гибридизации при значении Tm 55°С, и с применением описанных выше условий. Кроме того, значение Tm может составлять 60°С, 63°С или 65°С, без ограничения, и может быть соответствующим образом скорректировано специалистом в области техники в зависимости от задачи.
Надлежащая строгость для гибридизации полинуклеотидов зависит от длины полинуклеотидов и степени комплементарности, и эти параметры хорошо известны в области техники (например, J. Sambrook et al., выше).
В данном описании термин «микроорганизм (или штамм)» включает как микроорганизмы дикого типа и естественным образом или искусственным образом генетически модифицированные микроорганизмы, представляет собой микроорганизм, у которого определенный механизм ослаблен или усилен вследствие вставки экзогенного гена или усиления или ослабления активности эндогенного гена, и может представлять собой микроорганизм, прошедший генетическую модификацию, чтобы продуцировать желаемый полипептид, белок или продукт.
В данном изобретении термин «ослабление» активности полипептида (включая, например, белки, обозначенные по названию каждого фермента) представляет собой понятие, включающее как активность, пониженную по сравнению с собственной активностью, так и отсутствие активности. Ослабление может использоваться взаимозаменяемо с такими терминами, как инактивация, дефицит, снижение, уменьшение, сокращение и аттенуация.
Ослабление может также включать случай, когда активность самого полипептида снижена или устранена по сравнению с активностью полипептида, исходно присущей микроорганизму, в результате мутации полинуклеотида, кодирующего полипептид, и тому подобного, случая, когда общая степень активности и/или концентрация (уровень экспрессии) полипептида в клетке ниже, чем у нативного штамма вследствие ингибирования экспрессии гена полинуклеотида, кодирующего полипептид, или ингибирования трансляции в полипептид, случая, когда экспрессия полинуклеотида не достигается вовсе и/или случая, когда активность полипептида не проявляется даже несмотря на экспрессию полинуклеотида.
Такое ослабление активности полипептида может осуществляться любым способом, известным в области техники, без ограничения, и может достигаться путем применения различных способов, хорошо известных в области техники ((например, Nakashima N et al., Bacterial cellular engineering by genome editing and gene silencing. Int J Mol Sci. 2014;15(2):2773-2793, Sambrook et al., Molecular Cloning 2012, и тому подобные).
В частности, ослабление активности полипептида в данном изобретении может представлять собой
1) делецию всего гена, кодирующего полипептид, или его части;
2) модификацию области, регулирующей экспрессию (или последовательности, регулирующей экспрессию) для снижения экспрессии гена, кодирующего полипептид;
3) модификацию аминокислотной последовательности, составляющей полипептид (например, делеции/замены/добавления одной или более аминокислот в аминокислотную последовательность), так что активность полипептида устраняется или ослабляется;
4) модификацию нуклеотидной последовательности, кодирующей полипептид, так что активность полипептида устраняется или ослабевает (например, делеции/замены/добавления одного или более нуклеотидов в нуклеотидную последовательность гена, кодирующего полипептид, для кодирования модифицированного полипептида, так что активность полипептида устраняется или ослабляется);
5) модификацию нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-нетранслируемую область транскрипта гена, кодирующего полипептид;
6) внедрение антисмыслового олигонуклеотида (например, антисмысловой РНК), которая комплементарно связывается с транскриптом гена, кодирующего полипептид;
7) добавление последовательности, комплементарной последовательности Шайна-Дальгарно, к переднему концу последовательности Шайна-Дальгарно гена, кодирующего полипептид, с образованием вторичной структуры, к которой рибосома не может присоединиться;
8) добавление промотора, транскрибируемого в противоположном направлении, к 3' концу открытой рамки считывания (ORF) нуклеотидной последовательности, кодирующей полипептид (инженерия на основе обратной транскрипции, RTE); или
9) комбинацию указанного (например, комбинацию двух или более выбранных из (1) - (8)), без конкретного ограничения.
Например,
1) Делеция всего гена, кодирующего полипептид, или его части может представлять собой удаление всего полипептида, кодирующего эндогенный целевой полипептид в хромосоме, замену полинуклеотидом, в котором осуществлена делеция некоторых нуклеотидов или замена маркерным геном.
2) Модификация области, регулирующей экспрессию (или последовательности, регулирующей экспрессию) может представлять собой возникновение мутации в области, контролирующей экспрессию (или последовательности, контролирующей экспрессию) путем делеции, вставки, неконсервативной или консервативной замены или их комбинации, или замену последовательностью, обладающей более слабой активностью. Последовательность, регулирующая экспрессию, включает промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, и последовательность, регулирующую терминацию транскрипции и трансляции, без ограничения.
3) и 4) Модификация аминокислотной последовательности или нуклеотидной последовательности может представлять собой возникновение мутации в аминокислотной последовательности полипептида или полинуклеотидной последовательности, кодирующей полипептид, в результате делеции, вставки, неконсервативной или консервативной замены или их комбинации, так что активность полипептида ослабевает, или замену аминокислотной последовательностью или полинуклеотидной последовательностью, улучшенной для ослабления активности, или аминокислотной последовательностью или полинуклеотидной последовательностью, улучшенной, чтобы не проявлять активность, без ограничения. Например, экспрессия гена может быть ингибирована или ослаблена в результате внедрения мутации в полинуклеотидную последовательность для образования терминирующего кодона, без ограничения.
5) Модификация нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-нетранслируемую область транскрипта гена, кодирующего полипептид, может, например, представлять собой замену нуклеотидной последовательностью, кодирующей другой инициирующий кодон, имеющий более низкую степень экспрессии по сравнению с эндогенным инициирующим кодоном, без ограничения.
6) Внедрение антисмыслового олигонуклеотида (например, антисмысловой РНК), которая комплементарно связывается с транскриптом гена, кодирующего полипептид, можно найти, например, в [Weintraub, Н. et al., Antisense-RNA as a molecular tool for genetic analysis, Reviews - Trends in Genetics, Vol.1(1) 1986].
7) Добавление последовательности, комплементарной последовательности Шайна-Дальгарно, к переднему концу последовательности Шайна-Дальгарно, гена, кодирующего полипептид, с образованием вторичной структуры, к которой рибосома не может присоединиться, может осуществляться для выключения или замедления трансляции мРНК.
Кроме того, 8) добавление промотора, транскрибируемого в противоположном направлении, к 3' концу ORF нуклеотидной последовательности, кодирующей полипептид, может быть для ослабления активности путем создания антисмыслового нуклеотида, комплементарного транскрипту гена, кодирующего полипептид.
Модификация всех полинуклеотидов в микроорганизме по данному изобретению или их части может быть индуцирована путем (а) гомологичной рекомбинации с применением вектора для встраивания в хромосому микроорганизма или редактирования генома с применением полученных с использованием инженерии нуклеаз (например, CRISPR-Cas9) и/или (б) обработки излучением, таким как ультрафиолетовое излучение и радиоактивное излучение, и/или химическими средствами, без ограничения. Способ для модификации всего гена или его части может включать способ с применением технологии рекомбинации ДНК. Например, делеция всего гена или его части может достигаться путем инъекции нуклеотидной последовательности или вектора, содержащего нуклеотидную последовательность, гомологичную целевому гену, в микроорганизм для осуществления гомологичной рекомбинации. Инъецированная нуклеотидная последовательность или вектор может включать доминантный селективный маркер, без ограничения.
Вектор по данному изобретению может включать продукт ДНК, содержащей нуклеотидную последовательность полинуклеотида, кодирующего целевой полипептид, функционально связанную с подходящей областью, регулирующей экспрессию (или последовательностью, регулирующей экспрессию), чтобы целевой полипептид экспрессировался в подходящем хозяине. Область, регулирующая экспрессию, может включать промотор, способный инициировать транскрипцию, любую последовательность оператора для регуляции такой транскрипции, последовательность, кодирующую подходящий сайт связывания рибосомы на мРНК, и последовательность для регуляции терминации транскрипции и трансляции. После трансформации подходящего хозяина вектор может реплицироваться или функционировать независимо от генома хозяина и может интегрироваться в сам геном.
Векторы, использованные в данном изобретении, не ограничены какими-либо конкретными, и можно применять любые векторы, известные в области техники. Примеры обычно используемых векторов включают плазмиды, космиды, вирусы и бактериофаги естественного происхождения или рекомбинантные. Например, в качестве фаговых векторов или космидных векторов могут применяться pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A и Charon21A, а в качестве плазмидных векторов могут применяться векторы на основе pDZ, на основе pBR, на основе pUC, на основе pBluescript II, на основе pGEM, на основе pTZ, на основе pCL и на основе рЕТ. В частности, можно использовать векторы pDZ, pDC, pDCM2, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCClBAC и им подобные.
Например, полинуклеотид, кодирующий целевой полипептид, может быть встроен в хромосому при помощи вектора для встраивания в хромосому клетки. Встраивание полинуклеотида в хромосому может достигаться любым известным в области техники способом, например, посредством гомологичной рекомбинации, без ограничения. Дополнительно можно включать селективный маркер для определения встраивания в хромосому. Селективный маркер используют для отбора клеток, трансформированных вектором, то есть для подтверждения, встроилась ли целевая молекула нуклеиновой кислоты, и могут применяться маркеры, придающие фенотип, позволяющий осуществлять отбор, такой как устойчивость к лекарственным средствам, ауксотрофия, устойчивость к цитотоксическим агентам или экспрессия модифицированных поверхностных полипептидов. В среде, обработанной агентом для селекции, выживают или демонстрируют иные признаки экспрессии, только клетки, экспрессирующие селективный маркер, таким образом, можно осуществлять отбор трансформированных клеток.
В данном описании термин «трансформация» относится к внедрению вектора, который включает в себя полинуклеотид, кодирующий целевой полипептид, в клетку-хозяина или микроорганизм, так чтобы полипептид, кодируемый полинуклеотидом, мог экспрессироваться в клетке-хозяине. Полинуклеотид может быть встроен в хромосому клетки-хозяина и располагаться в хромосоме клетки-хозяина или находиться вне хромосомы, при условии, что после трансформации полинуклеотид может экспрессироваться в клетке-хозяине. Кроме того, полинуклеотид включает ДНК и/или РНК, которые кодируют целевой полипептид. Полинуклеотид может быть внедрен в любой форме, при условии, что он может быть внедрен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть внедрен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генетическую конструкцию, включающую все элементы, необходимые для самостоятельной экспрессии. Как правило, экспрессионная кассета может включать в себя промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, сайт связывания рибосомы и сигнал терминации трансляции. Экспрессионная кассета может быть в форме экспрессирующего вектора, способного к саморепликации. Кроме того, полинуклеотид может быть внедрен в клетку-хозяина как таковой и быть функционально связанным с последовательностью, необходимой для экспрессии в клетке-хозяине, без ограничения.
Кроме того, термин «функционально связанный» означает, что последовательность полинуклеотида функционально связана с последовательностью промотора, который инициирует и опосредует транскрипцию полинуклеотида, кодирующего целевой полипептид по данному изобретению.
Микроорганизм по данному изобретению может представлять собой микроорганизм, обладающий увеличенной способностью продуцировать L-глутамин по сравнению с родительским штаммом или штаммом дикого типа рода Corynebacterium, в котором активность фосфоенолпируваткарбоксикиназы не ослаблена. Иными словами, микроорганизм по данному изобретению может представлять собой микроорганизм, от природы обладающий способностью продуцировать L-глутамин, или микроорганизм, в котором PEPCK или кодирующий ее полинуклеотид ослаблен в родительском штамме, не обладающем способностью продуцировать L-глутамин, без ограничения.
Например, микроорганизм с немодифицированной PEPCK, в котором PEPCK не ослаблена и который представляет собой целевой штамм для сравнения увеличения способности продуцировать L-глутамин, может представлять собой штамм Corynebacterium glutamicum АТСС13032 дикого типа, штамм Corynebacterium glutamicum АТСС13032, обладающий усиленной активностью белка глутаминсинтетазы (GlnA, ЕС 6.3.1.2), микроорганизм, в котором активность соответствующего белка усилена в микроорганизме путем внедрения мутации (D401N) в ген glnA, кодирующий глутаминсинтетазу, штамм Corynebacterium glutamicum КССМ12645Р (WO2021-177731 A1), глутамин-продуцирующий штамм, или штамм Corynebacterium glutamicum KFCC-10680 (патент Кореи 10-0048440), глутамин-продуцирующий штамм, без ограничения.
Например, рекомбинантный штамм, обладающий усиленной продуцирующей способностью, может обладать способностью продуцировать L-глутамин, повышенной приблизительно на 1% или более, в частности, приблизительно на 2% или более, приблизительно на 5% или более, приблизительно на 6% или более, приблизительно на 7% или более, приблизительно на 7,2% или более, приблизительно на 8% или более, приблизительно на 9% или более, или приблизительно 9,2% или более (верхняя граница не ограничена кокой-либо конкретной и может составлять, например, приблизительно 200% или менее, приблизительно 150% или менее, приблизительно 100% или менее, приблизительно 51% или менее, приблизительно 40% или менее, приблизительно 30% или менее, приблизительно 20% или менее или приблизительно 15% или менее) по сравнению со способностью продуцировать L-глутамин родительского штамма до мутации, немодифицированного микроорганизма или микроорганизма с немодифицированной PEPCK. Однако, способность продуцировать L-глутамин не ограничена перечисленной, при условии, что она составляет положительное значение, повышенное по сравнению с продуцирующей способностью родительского штамма до мутации, немодифицированного микроорганизма или микроорганизма с немодифицированной PEPCK. В другом примере микроорганизм, обладающий повышенной способностью продуцировать L-глутамин, может обладать способностью продуцировать L-глутамин, повышенной приблизительно в 1,01 раз или более, приблизительно в 1,02 раз или более, приблизительно в 1,05 раз или более, приблизительно в 1,06 раз или более, приблизительно в 1,07 раз или более, приблизительно в 1,072 раз или более, приблизительно в 1,08 раз или более, приблизительно в 1,09 раз или более, или приблизительно в 1,092 раз или более (верхняя граница не ограничена какой-либо конкретной и может составлять, например, приблизительно 10 раз или менее, приблизительно 5 раз или менее, приблизительно 3 раза или менее, приблизительно 2 раза или менее) по сравнению со способностью продуцировать L-глутамин родительского штамма до мутации, немодифицированного микроорганизма или микроорганизма с немодифицированной PEPCK, однако способность продуцировать L-глутамин не ограничена указанной.
В данном описании термин «немодифицированный микроорганизм» не исключает штаммов, содержащих мутации, которые могут возникать естественным путем в микроорганизмах, и может относиться к штамму дикого типа или к самому нативному штамму или к штамму до изменения его признака вследствие генетической мутации, вызванной естественными или искусственными факторами. Кроме того, в данном описании термин «микроорганизм с немодифицированной PEPCK» может относиться к штамму, в котором описанная здесь PEPCK или кодирующий ее полинуклеотид не ослаблены, или к штамму, в котором PEPCK или кодирующий ее полинуклеотид еще не ослаблены. Микроорганизм с немодифицированной PEPCK по данному изобретению не исключает штаммов, содержащих модификации других белков или генов, нежели модификация PEPCK или кодирующего ее полинуклеотида.
В данном описании термин «немодифицированный микроорганизм» может использоваться взаимозаменяемо со «штаммом до модификации», «микроорганизмом до модификации», «немутированным штаммом», «немодифицированным штаммом», «немутированным микроорганизмом» или «референтным микроорганизмом».
Микроорганизм по данному изобретению может представлять собой микроорганизм, содержащий PEPCK, у которой активность ослаблена по сравнению с собственной активностью, или кодирующий ее полинуклеотид; или микроорганизм (например, рекомбинантный микроорганизм), генетически модифицированный, чтобы содержать PEPCK, у которой активность ослаблена по сравнению с собственной активностью, или кодирующий ее полинуклеотид, без ограничения. «Собственная активность» относится к активности определенного полипептида, которой исходно обладал родительский штамм до трансформации, микроорганизм дикого типа или немодифицированный микроорганизм, когда признак изменяется вследствие генетической мутации, вызванной факторами естественного или искусственного происхождения. Данный термин можно использовать взаимозаменяемо с «активностью до модификации». Тот факт, что активность полипептида «ослаблена, инактивирована, дефицитна, уменьшена, снижена, сокращена и аттенуирована» по сравнению с его собственной активностью означает, что активность полипептида снижена по сравнению с активностью определенного полипептида, которой исходно обладал родительский штамм до трансформации или немодифицированный микроорганизм.
В качестве еще одного примера данного изобретения микроорганизм по данному изобретению может представлять собой Corynebacterium glutamicum, Corynebacterium stationis, Corynebacterium crudilactis, Corynebacterium deserti, Corynebacterium efficiens, Corynebacterium callunae, Corynebacterium singulare, Corynebacterium halotolerans, Corynebacterium striatum, Corynebacterium ammoniagenes, Corynebacterium pollutisoli, Corynebacterium imitans, Corynebacterium testudinoris или Corynebacterium flavescens и может, в частности, представлять собой Corynebacterium glutamicum, без ограничения.
В качестве другого примера рекомбинантный микроорганизм по данному изобретению может представлять собой микроорганизм, у которого способность продуцировать L-глутамин усилена путем дополнительного усиления активности некоторых белков пути биосинтеза L-глутамина или путем дополнительного ослабления активности некоторых белков пути распада L-глутамина.
В частности, в данном изобретении микроорганизм рода Corynebacterium может представлять собой микроорганизм (US 7262035 В2), в котором дополнительно осуществлено усиление glnA (например, увеличение числа копий, изменение промотора, дерегулирование активности вследствие аденилирования, ослабление glnE или ослабление активности белка РП), микроорганизм (US 7262035 В2), в котором дополнительно осуществлено усиление глутаматдегидрогеназы (GDH) (например, увеличение числа копий или модификация последовательности, регулирующей экспрессию), или микроорганизм (WO2021-177731 А1), в который дополнительно внедрена мутация, усиливающая glnA, без ограничения.
В данном описании термин «усиление активности» полипептида означает, что активность полипептида увеличена по сравнению с его собственной активностью. Усиление может использоваться взаимозаменяемо с такими терминами, как активация, повышение, сверхэкспрессия и увеличение. При этом, активация, усиление, повышение, сверхэкспрессия и увеличение могут включать как проявление активности, которая исходно отсутствовала, так и проявление активности, улучшенной по сравнению с собственной активностью или активностью до модификации. «Собственная активность» относится к активности определенного полипептида, которой исходно обладал родительский штамм до трансформации или немодифицированный микроорганизм, когда признак изменяется вследствие генетической мутации, вызванной факторами естественного или искусственного происхождения. Данный термин может использоваться взаимозаменяемо с «активностью до модификации». Тот факт, что активность полипептида «усилена», «повышена», «сверхэкспрессирована» или «увеличена» по сравнению с его собственной активностью означает, что активность полипептида улучшена по сравнению с активностью и/или концентрацией (уровнем экспрессии) определенного полипептида, которой исходно обладал родительский штамм до трансформации или немодифицированный микроорганизм.
Усиление может достигаться посредством внедрения чужеродного полипептида или посредством усиления активности и/или концентрации (уровня экспрессии) эндогенного полипептида. Подтвердить, усилилась ли активность белка можно по увеличению степени активности или уровня экспрессии соответствующего полипептида или по увеличению количества продукта, высвобождаемого соответствующим полипептидом.
Для усиления активности полипептида могут применяться различные способы, хорошо известные в области техники, и способ может не ограничиваться каким-либо конкретным, при условии, что активность целевого полипептида может быть усилена по сравнению с таковой микроорганизма до модификации. В частности, усиление может осуществляться посредством генетической инженерии и/или белковой инженерии, которые являются рутинными способами молекулярной биологии и хорошо известны специалистам в области техники, без ограничения (например, Sitnicka et al., Functional Analysis of Genes. Advances in Cell Biology. 2010, Vol.2. 1-16, Sambrook et al., Molecular Cloning 2012).
В частности, усиление активности полипептида по данному изобретению может представлять собой
1) увеличение внутриклеточного числа копий полинуклеотида, кодирующего полипептид;
2) замену области, регулирующей экспрессию гена, на хромосоме, кодирующей полипептид, высокоактивной последовательностью;
3) модификацию нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-нетранслируемую область транскрипта гена, кодирующего полипептид;
4) модификацию аминокислотной последовательности полипептида, так чтобы активность полипептида усиливалась;
5) модификацию полинуклеотидной последовательности, кодирующей полипептид, так чтобы активность полипептида усиливалась (например, модификацию полинуклеотидной последовательности гена полипептида для кодирования модифицированного полипептида, так чтобы активность полипептида усиливалась);
6) внедрение чужеродного полипептида, проявляющего активность полипептида, или чужеродного полинуклеотида, кодирующего его;
7) оптимизацию кодонов полинуклеотида, кодирующего полипептид;
8) анализ третичной структуры полипептида для выбора и модификации или химической модификации экспонированного сайта или
9) комбинацию двух или более выбранных из (1) (8), без конкретного ограничения.
Более конкретно,
1) Увеличение внутриклеточного числа копий полинуклеотида, кодирующего полипептид, может достигаться посредством внедрения в клетку-хозяина вектора, способного реплицироваться и функционировать независимо от хозяина, с которым функционально связан полинуклеотид, кодирующий соответствующий полипептид. В качестве альтернативы увеличение может достигаться путем внедрения одной копии или двух или более копий полинуклеотида, кодирующего соответствующий полипептид, в хромосому клетки-хозяина. Внедрение в хромосому может осуществляться путем внедрения в клетку-хозяина вектора, который способен встраивать полинуклеотид в хромосому клетки-хозяина, без ограничения. Вектор является таким, как описано выше.
2) Замена области, регулирующей экспрессию гена (экспрессию последовательности гена) на хромосоме, кодирующей полипептид, высокоактивной последовательностью может, например, представлять собой возникновение мутации в последовательности в результате делеции, вставки, неконсервативной или консервативной замены или их комбинации, или замену последовательностью, обладающей более высокой активностью, так чтобы активность области, регулирующей экспрессию, дополнительно усилилась. Область, регулирующая экспрессию, может включать промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, и последовательность, регулирующую терминацию транскрипции и трансляции, без конкретного ограничения. Например, замена может осуществляться для замены исходного промотора сильным промотором, без ограничения.
Примеры известных сильных промоторов включают промоторы CJ1-CJ7 (патент US 7662943 В2), промотор lac, промотор trp, промотор trc, промотор tac, промотор PR фага лямбда, промотор PL, промотор tet, промотор gapA, промотор SPL7, промотор SPL13 (sm3) (патент US 10584338 B2), промотор 02 (патент US 10273491 В2), промотор tkt и промотор уссА, без ограничения.
3) Модификация нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-нетранслируемую область транскрипта гена, кодирующего полипептид, может, например, представлять собой замену нуклеотидной последовательностью, кодирующей другой инициирующий кодон, имеющий более высокую степень экспрессии по сравнению с эндогенным инициирующим кодоном, без ограничения.
4) и 5) Модификация аминокислотной последовательности или полинуклеотидной последовательности может представлять собой возникновение мутации в аминокислотной последовательности полипептида или в полинуклеотидной последовательности, кодирующей полипептид, в результате делеции, вставки, неконсервативной или консервативной замены или их комбинации, так что активность полипептида усиливается, или замену аминокислотной последовательностью или полинуклеотидной последовательностью, улучшенной для проявления более сильной активности, или аминокислотной последовательностью или полинуклеотидной последовательностью, улучшенной, чтобы проявлять повышенную активность, без ограничения. В частности, замена может осуществляться путем встраивания полинуклеотида в хромосому посредством гомологичной рекомбинации, без ограничения. Используемый при этом вектор может дополнительно включать селективный маркер для определения встраивания в хромосому.
6) Внедрение чужеродного полинуклеотида, проявляющего активность полипептида, может представлять собой внедрение в клетку-хозяина чужеродного полинуклеотида, кодирующего полипептид, демонстрирующий такую же/схожую активность с таковой полипептида. Чужеродный полинуклеотид не ограничен происхождением или последовательностью, при условии, что он демонстрирует такую же/схожую активность, как и полипептид. Внедрение может осуществляться путем соответствующего выбора и применения специалистом в области техники известного способа трансформации. При экспрессии внедренного полинуклеотида в клетке-хозяине может продуцироваться полипептид, и его активность может быть повышена.
7) Оптимизация кодонов полинуклеотида, кодирующего полипептид, может представлять собой оптимизацию кодонов, так что транскрипция или трансляция эндогенного полинуклеотида в клетке-хозяине повышается, или оптимизацию кодонов чужеродного полинуклеотида, так что в клетке-хозяине достигается оптимизированная транскрипция и трансляция чужеродного полинуклеотида.
8) Анализ третичной структуры полипептида для выбора и модификации или химической модификации экспонированного сайта может осуществляться для определения шаблонного белка-кандидата в соответствии со степенью сходства последовательностей, например, путем сравнения информации о последовательности анализируемого полипептида с базой данных, в которой хранится информация о последовательности известных полипептидов, для подтверждения структур на этом основании, для выбора модифицируемого или химически модифицируемого экспонированного сайта и для модификации или химической модификации экспонированного сайта.
Такое усиление активности полипептида может представлять собой увеличение активности или концентрации (уровня экспрессии) соответствующего полипептида на основании активности или концентрации полипептида, экспрессируемого в штамме дикого типа или штамме микроорганизма до модификации или увеличение количества продукта, образуемого соответствующим полипептидом, без ограничения.
В другом аспекте данного изобретения предложен способ получения L-глутамина, включающий культивирование в среде микроорганизма рода Corynebacterium, обладающего ослабленной активностью фосфоенолпируваткарбоксикиназы и способностью продуцировать L-глутамин.
Способ получения L-глутамина по данному изобретению может включать культивирование в среде микроорганизма, содержащего PEPCK, активность которой ослаблена по сравнению с собственной активностью, или кодирующий ее полинуклеотид; или микроорганизма, генетически модифицированного, чтобы содержать PEPCK, активность которой ослаблена по сравнению с собственной активностью, или кодирующий ее полинуклеотид.
В данном описании термин «культивирование» означает выращивание микроорганизма по данному изобретению в надлежащим образом контролируемых окружающих условиях. Процесс культивирования по данному изобретению можно осуществлять с применением подходящей среды в условиях культивирования, известных в области техники. Специалист в области техники может легко адаптировать такой процесс культивирования в зависимости от выбранного микроорганизма. В частности, культивирование может представлять собой периодическое культивирование, непрерывное культивирование и/или непрерывное культивирование с подпиткой, без ограничения.
В данном описании термин «среда» относится к материалу, в котором питательные вещества, необходимые для культивирования микроорганизма по данному изобретению, смешаны в качестве основных компонентов, и обеспечивающему нутриенты и факторы роста, включая воду, необходимые для выживания и роста. В частности, в качестве среды и других условий культивирования, применяемых для культивирования микроорганизма по данному изобретению, можно использовать любую среду, без конкретного ограничения, при условии, что она представляет собой среду, используемую при стандартном культивировании микроорганизмов. Микроорганизм по данному изобретению можно культивировать в обычной среде, содержащей надлежащие источники углерода, источники азота, источники фосфора, неорганические соединения, аминокислоты и/или витамины, в аэробных условиях при регулировании температуры, рН и тому подобного.
В частности, культуральная среда для микроорганизмов рода Corynebacterium может быть найдена в литературе ["Manual of Methods for General Bacteriology" by the American Society for Bacteriology (Washington D.C., USA, 1981)].
В данном изобретении примеры источников углерода включают углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, сахароза и мальтоза; сахароспирты, такие как маннит и сорбит; органические кислоты, такие как пировиноградная кислота, молочная кислота и лимонная кислота, и аминокислоты, такие как глутаминовая кислота, метионин и лизин. Можно использовать естественные органические источники питательных веществ, такие как гидролизат крахмала, меласса, сырая меласса, рисовые отруби, маниок, выжимки сахарного тростника и жидкий кукурузный экстракт. В частности, можно использовать такие углеводы как глюкоза и стерилизованная предварительно обработанная меласса (то есть меласса, в которой сахара конвертированы в восстанавливающие) и, можно различным образом использовать другие источники углерода в соответствующем количестве, без ограничения. Такие источники углерода можно использовать по-отдельности или в комбинации двух или более из них, без ограничения.
В качестве источников азота можно использовать источники неорганического азота, такие как аммоний, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония и нитрат аммония; и источники органического азота, такие как аминокислоты, такие как глутаминовая кислота, метионин и глутамин, пептон, NZ амин, мясной экстракт, дрожжевой экстракт, мальтозный экстракт, кукурузный экстракт, гидролизат казеина, рыба или продукты ее переработки, обезжиренный соевый жмых или продукты его переработки. Такие источники азота можно использовать по отдельности или в комбинации двух или более из них, без ограничения.
Примеры источников фосфора могут включать калия дигидрофосфат и калия гидрофосфат или натрия дигидрофосфат и натрия гидрофосфат. В дополнение к перечисленному в качестве неорганических соединений можно использовать хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и тому подобные, а также аминокислоты, витамины, соответствующие предшественники и/или тому подобное. Эти компоненты или предшественники можно добавлять в среду либо порциями, либо непрерывно, однако способ добавления компонентов или предшественников этим не ограничивается.
Для корректировки рН среды в процессе культивирования микроорганизма по данному изобретению в среду можно соответствующим образом добавлять такие соединения, как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота. Для подавления образования пены в процессе культивирования можно добавлять пеногасители, такие как сложный полигликоле вый эфир жирной кислоты. Для поддержания аэробного состояния среды в среду можно вводить кислород или кислородсодержащий газ, либо для поддержания анаэробных или микроаэробных условий газ можно не вводить или можно вводить азот, водород или углекислый газ, однако способ регулирования атмосферы этим не ограничивается.
Для культуры по данному изобретению температуру культивирования можно поддерживать от 20°С до 45°С, в частности, от 25°С до 40°С, и время культивирования может составлять приблизительно от 10 до 160 часов, однако условия культивирования этим не ограничиваются.
L-глутамин, продуцируемый культурой по данному изобретению, может секретироваться в среду или оставаться в клетках.
Способ получения L-глутамина по данному изобретению может дополнительно включать получение микроорганизма по данному изобретению, получение среды для культивирования микроорганизма или их комбинацию (в любом порядке), например, перед стадией культивирования.
Способ получения L-глутамина по данному изобретению может дополнительно включать выделение L-глутамина из культуральной среды (среды после проведения культивирования) или культивированного микроорганизма. После стадии культивирования может быть дополнительно включена стадия выделения.
Выделение может осуществляться для сбора необходимого L-глутамина с применением подходящего способа, известного в области техники, в зависимости от способа культивирования микроорганизма по данному изобретению, например, периодического культивирования, непрерывного культивирования или культивирования с подпиткой. Например, может применяться центрифугирование, фильтрация, обработка преципитирующим агентом для кристаллизованных белков (высаливание), экстракция, обработка ультразвуком, ультрафильтрация, диализ, различные виды хроматографии, такие как хроматография молекулярных сит (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография и аффинная хроматография, ВЭЖХ (высокоэффективная жидкостная хроматография) или их комбинация. Необходимый L-глутамин можно выделять из среды или микроорганизма подходящими способами, известными в области техники.
Способ получения L-глутамина по данному изобретению может дополнительно включать стадию очистки. Очистку можно осуществлять подходящими способами, известными в области техники. Например, когда способ получения L-глутамина по данному изобретению включает как стадию выделения, так и стадию очистки, стадию выделения и стадию очистки можно осуществлять непрерывно или с перерывами, независимо от порядка, или можно осуществлять одновременно или путем объединения в одну стадию, однако способ осуществления стадии выделения и стадии очистки этим не ограничивается.
В способе по данному изобретению PEPCK, полинуклеотид, вектор и микроорганизм и тому подобное являются такими, как описано в других аспектах.
В еще одном аспекте данного изобретения предложена композиция для получения L-глутамина, включающая микроорганизм, в котором ослаблена фосфоенолпируваткарбоксикиназа; среда, в которой культивировали микроорганизм, или их комбинация.
Композиция по данному изобретению может дополнительно содержать любой подходящий эксципиент, обычно применяющийся в композициях для получения L-глутамина, и такой эксципиент может, например, представлять собой консерванты, смачивающие вещества, диспергирующие вещества, суспендирующие вещества, буферы, стабилизаторы или вещества, регулирующие тоничность, без ограничения.
В еще одном аспекте данного изобретения предложен способ получения микроорганизма для продуцирования L-глутамина, включающий ослабление PEPCK.
В еще одном аспекте данного изобретения предложено применение микроорганизма рода Corynebacterium, в котором ослаблена фосфоенолпируваткарбоксикиназа, для получения L-глутамина.
PEPCK, ослабление, микроорганизм рода Corynebacterium и тому подобное являются такими, как описано в других аспектах.
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Далее данное изобретение будет описано более подробно с отсылкой к примерам. При этом, приведенные ниже Примеры являются лишь предпочтительными воплощениями для иллюстрации данного изобретения и, следовательно, не предназначены для ограничения объема данного изобретения. При этом, технические признаки, которые не описаны в данном документе, могут быть в достаточной степени поняты и легко реализованы специалистами в области техники, к которой относится данное изобретение, или в схожих областях техники.
Пример 1. Конструирование вектора для ослабления гена рек
Для ослабления гена рек, кодирующего фосфоенолпируваткарбоксикиназу, конструировали вектор для делеции гена.
В частности, для конструирования штамма, в котором осуществлена делеция части гена рек, осуществляли ПЦР с использованием хромосомы Corynebacterium glutamicum АТСС13032 в качестве матрицы и праймеров с SEQ ID NO: 3 и SEQ ID NO: 4, SEQ ID NO: 1 и SEQ ID NO: 2, соответственно. Последовательности использованных при этом праймеров приведены в Таблице 1 ниже.

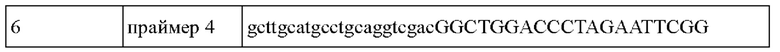
В качестве полимеразы для реакции ПЦР использовали ДНК-полимеразу высокой точности PfuUltra™ (Stratagene). Условия ПЦР представляли собой денатурацию при 95°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и реакцию полимеризации при 72°С в течение 1 мин, повторявшиеся 28 раз. В результате получали 5' концевой фрагмент ДНК 929 пн, несущий инициирующий кодон гена рек, и 3' концевой фрагмент ДНК 1000 пн, несущий терминирующий кодон гена рек, соответственно. Два амплифицированных фрагмента ДНК очищали с применением набора для очистки продуктов ПЦР (QUIAGEN) и использовали в качестве встраиваемых фрагментов ДНК для конструирования плазмиды. При этом осуществляли клонирование, выбирая молярные концентрации (М) вектора pDZ (патент Кореи 2009-0094433), обработанного рестрикционным ферментом Sail и подвергнутого тепловой обработке при 65°С в течение 20 минут, и встраиваемого фрагмента ДНК, амплифицированного посредством ПЦР, в соотношении 1:2 и используя набор для клонирования Infusion (TaKaRa) согласно прилагающейся инструкции для конструирования вектора pDZ-APEPCK для делеции гена рек.
Пример 2: Конструирование штамма рода Corynebacterium на основе дикого типа, обладающего способностью продуцировать L-глутамин
Штамм, продуцирующий L-глутамин, получали из Corynebacterium glutamicum АТСС13032 дикого типа. В частности, для увеличения активности глутаминсинтетазы, последнего фермента пути биосинтеза, конструировали штамм (регистрационный номер КССМ12645Р), в который был внедрен мутантный glnA (D401N), ген, кодирующий глутаминсинтетазу (SEQ ID NO: 11).
В частности, для конструирования штамма, в который была внедрена мутация glnA (D401N), проводили ПЦР с использованием хромосомы Corynebacterium glutamicum АТСС13032 в качестве матрицы и праймеров с SEQ ID NO: 7 и SEQ ID NO: 8 или SEQ ID NO: 9 и SEQ ID NO: 10, соответственно. Последовательности использованных при этом праймеров приведены в Таблице 2 ниже.


В качестве полимеразы для реакции ПЦР использовали ДНК-полимеразу высокой точности PfuUltra™. Условия ПЦР представляли собой денатурацию при 95°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и реакцию полимеризации при 72°С в течение 1 мин, повторявшиеся 28 раз. В результате получали 5' концевой фрагмент ДНК 929 пн и 3' концевой фрагмент ДНК, несущий ген glnA с мутацией D401N, соответственно. Два амплифицированных фрагмента ДНК очищали с применением набора для очистки продукта ПЦР и использовали в качестве встраиваемых фрагментов ДНК для конструирования плазмиды. При этом осуществляли клонирование, выбирая молярные концентрации (М) вектора pDZ (патент Кореи 2009-0094433), обработанного рестрикционным ферментом spel и подвергнутого тепловой обработке при 65°С в течение 20 минут, и встраиваемого фрагмента ДНК, амплифицированного посредством ПЦР, в соотношении 1:2 и используя набор для клонирования Infusion согласно прилагающейся инструкции с конструированием pDZ-glnA(D401N) для встраивания мутантного гена glnA(D401N).
Затем Corynebacterium glutamicum АТСС13032 трансформировали сконструированным вектором путем электропорации и применения электрического импульса (Van der Rest et al., Appl. Microbial. Biotechnol. 52:541-545, 1999) с получением штамма, содержащего в хромосоме мутацию glnA (D401N) в результате гомологичной рекомбинации с хромосомой. Штамм обозначали Corynebacterium glutamicum СА11-4021 и затем депонировали в Корейском центре культур микроорганизмов (КССМ), международном депозитарии, в соответствии с Будапештским соглашением, 19 декабря 2019 с присвоением ему регистрационного номера КССМ12645Р.
Пример 3. Конструирование и оценка продуцирующего L-глутамин штамма, имеющего ослабленный ген рек
Ген pck в Corynebacterium glutamicum СА11-4021, продуцирующем L-глутамин штамме, сконструированном в Примере 2, ослабляли. Более конкретно, Corynebacterium glutamicum СА11-4021 трансформировали вектором, сконструированным в Примере 1, путем электропорации и осуществляли второй кроссовер для получения продуцирующего L-глутамин штамма, в котором была осуществлена делеция гена рек в хромосоме, и обозначали Corynebacterium glutamicum СА11-4023.
Способность продуцировать глутамин измеряли путем культивирования Corynebacterium glutamicum СА11-4023 следующим образом.
Во-первых, каждый штамм инокулировали в 25 мл среды для посева, находящейся в колбе с угловыми перегородками объемом 250 мл, и культивировали при встряхивании со скоростью 200 об/мин при температуре 30°С в течение 20 часов. Затем 1 мл раствора посевной культуры инокулировали в 24 мл среды для продуцирования, находящейся в колбе с угловыми перегородками объемом 250 мл, и культивировали при встряхивании со скоростью 200 об/мин при температуре 32°С в течение 48 часов. Составы среды для посева и среды для продуцирования, соответственно, показаны ниже. По окончании культивирования измеряли концентрацию L-глутамина при помощи ВЭЖХ (высокоэффективная жидкостная хроматография) (Waters 2478). Результаты измерения способности продуцировать глутамин и скорости потребления Сахаров показаны в Таблице 3 ниже.
Среда для посева (рН 7,0)
Глюкоза 20 г, пептон 10 г, дрожжевой экстракт 5 г, мочевина 1,5 г, KH2PO4 4 г, K2HPO4 8 г, MgSO4 × 7Н2О 0,5 г, биотин 100 мкг, тиамин-HCl 1000 мкг, кальция пантотенат 2000 мкг и никотинамид 2000 мкг (из расчета на 1 л дистиллированной воды).
Среда для продуцирования глутамина (рН 8,0)
Сахар-сырец 60 г, (NH4)2SO4 45 г, соевый белок 0,48 г, СаСО3 50 г, MgSO4 × 7Н2О 0,4 г, KH2PO4 1 г, тиамина гидрохлорид 0,2 мг, биотин 0,3 мг, никотинамид 60 мг, FeSO4 × 7Н2О 10 мг и MnSO4 × Н2О 10 мг (из расчета на 1 л дистиллированной воды).


В результате подтверждали, что способность продуцировать глутамин у СА11-4023, в котором был ослаблен ген рек, улучшалась на 7,2% по сравнению с таковой родительского штамма СА11-4021.
Штамм СА11-4023 депонировали в Корейском центре культур микроорганизмов (КССМ), международном депозитарии, в соответствии с Будапештским соглашением, 22 декабря 2020 с присвоением ему регистрационного номера КССМ 12916Р.
Пример 4. Конструирование и оценка штамма, продуцирующего L-глутамин в высоких концентрациях, имеющего ослабленный ген рек
Ген рек в штамме Corynebacterium glutamicum KFCC-10680 (патент Кореи 10-0048440), известном штамме, продуцирующем глутамин, ослабляли таким же образом, как в Примере 3. Более конкретно, Corynebacterium glutamicum KFCC-10680 трансформировали вектором pDZ-ДРЕРСК, сконструированным в Примере 1, путем электропорации и осуществляли второй кроссовер для получения штамма, в котором была осуществлена делеция гена рек в хромосоме, обозначали Corynebacterium glutamicum KFCC-10680Apck и культивировали следующим образом для измерения способности продуцировать глутамин.
Во-первых, каждый штамм инокулировали в 25 мл среды для посева, находящейся в колбе с угловыми перегородками объемом 250 мл, и культивировали при встряхивании со скоростью 200 об/мин при температуре 30°С в течение 20 часов. Затем 1 мл раствора посевной культуры инокулировали в 24 мл среды для продуцирования, находящейся в колбе с угловыми перегородками объемом 250 мл, и культивировали при встряхивании со скоростью 200 об/мин при температуре 32°С в течение 48 часов. Составы среды для посева и среды для продуцирования, соответственно, показаны ниже. По окончании культивирования измеряли концентрацию L-глутамина при помощи ВЭЖХ (Waters 2478). Результаты измерения способности продуцировать глутамин и скорости потребления Сахаров показаны в Таблице 4 ниже.
Среда для посева (рН 7,0)
Глюкоза 20 г, пептон 10 г, дрожжевой экстракт 5 г, мочевина 1,5 г, KH2PO4 4 г, K2HPO4 8 г, MgSO4 × 7H2O 0,5 г, биотин 100 мкг, тиамин-HCl 1000 мкг, кальция пантотенат 2000 мкг и никотинамид 2000 мкг (из расчета на 1 л дистиллированной воды).
Среда для продуцирования глутамина (рН 8,0)
Сахар-сырец 60 г, (NH4)2SO4 45 г, соевый белок 0,48 г, СаСО3 50 г, MgSO4 × 7Н2О 0,4 г, KH2PO4 1 г, тиамина гидрохлорид 0,2 мг, биотин 0,3 мг, никотинамид 60 мг, FeSO4 × 7Н2О 10 мг и MnSO4 × Н2О 10 мг (из расчета на 1 л дистиллированной воды).
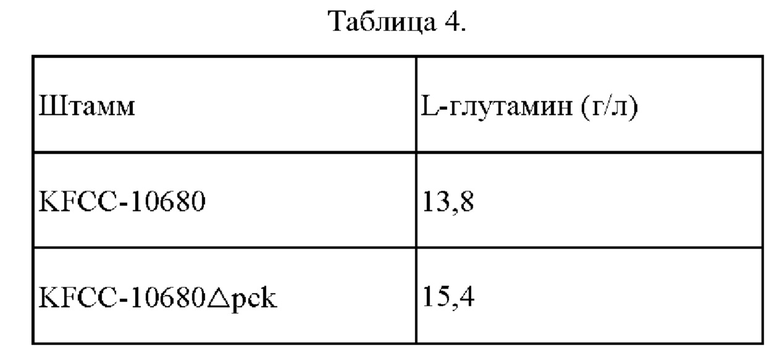
В результате подтверждали, что способность продуцировать L-глутамин у KFCC-10680Δpck, в котором была осуществлена делеция гена рек, улучшалась на 9,2% по сравнению с таковой родительского штамма KFCC-10680.
На основании приведенного описания специалист в области техники поймет, что возможны и другие частные воплощения изобретения, не изменяющие сути изобретения или его существенных признаков. Таким образом, следует понимать, что приведенное выше воплощение не ограничивает объем изобретения, а является иллюстративным во всех аспектах. Следует понимать, что объем изобретения ограничен прилагаемой формулой изобретения, а не приведенным выше подробным описанием изобретения, и поэтому все изменения или модификации, вытекающие из значений и объема формулы изобретения, и эквивалентные им понятия следует считать входящими в объем данного изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> МИКРООРГАНИЗМ, ОБЛАДАЮЩИЙ УСИЛЕННОЙ СПОСОБНОСТЬЮ ПРОДУЦИРОВАТЬ
L-ГЛУТАМИН, И СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНА С ЕГО ПРИМЕНЕНИЕМ
<130> OPA21345
<150> KR 10-2020-0156903
<151> 2020-11-20
<160> 12
<170> KoPatentIn 3.0
<210> 1
<211> 610
<212> PRT
<213> Unknown
<220>
<223> Аминокислотная последовательность фосфоенолпируваткарбоксикиназы
<400> 1
Met Thr Thr Ala Ala Ile Arg Gly Leu Gln Gly Glu Ala Pro Thr Lys
1 5 10 15
Asn Lys Glu Leu Leu Asn Trp Ile Ala Asp Ala Val Glu Leu Phe Gln
20 25 30
Pro Glu Ala Val Val Phe Val Asp Gly Ser Gln Ala Glu Trp Asp Arg
35 40 45
Met Ala Glu Asp Leu Val Glu Ala Gly Thr Leu Ile Lys Leu Asn Glu
50 55 60
Glu Lys Arg Pro Asn Ser Tyr Leu Ala Arg Ser Asn Pro Ser Asp Val
65 70 75 80
Ala Arg Val Glu Ser Arg Thr Phe Ile Cys Ser Glu Lys Glu Glu Asp
85 90 95
Ala Gly Pro Thr Asn Asn Trp Ala Pro Pro Gln Ala Met Lys Asp Glu
100 105 110
Met Ser Lys His Tyr Ala Gly Ser Met Lys Gly Arg Thr Met Tyr Val
115 120 125
Val Pro Phe Cys Met Gly Pro Ile Ser Asp Pro Asp Pro Lys Leu Gly
130 135 140
Val Gln Leu Thr Asp Ser Glu Tyr Val Val Met Ser Met Arg Ile Met
145 150 155 160
Thr Arg Met Gly Ile Glu Ala Leu Asp Lys Ile Gly Ala Asn Gly Ser
165 170 175
Phe Val Arg Cys Leu His Ser Val Gly Ala Pro Leu Glu Pro Gly Gln
180 185 190
Glu Asp Val Ala Trp Pro Cys Asn Asp Thr Lys Tyr Ile Thr Gln Phe
195 200 205
Pro Glu Thr Lys Glu Ile Trp Ser Tyr Gly Ser Gly Tyr Gly Gly Asn
210 215 220
Ala Ile Leu Ala Lys Lys Cys Tyr Ala Leu Arg Ile Ala Ser Val Met
225 230 235 240
Ala Arg Glu Glu Gly Trp Met Ala Glu His Met Leu Ile Leu Lys Leu
245 250 255
Ile Asn Pro Glu Gly Lys Ala Tyr His Ile Ala Ala Ala Phe Pro Ser
260 265 270
Ala Cys Gly Lys Thr Asn Leu Ala Met Ile Thr Pro Thr Ile Pro Gly
275 280 285
Trp Thr Ala Gln Val Val Gly Asp Asp Ile Ala Trp Leu Lys Leu Arg
290 295 300
Glu Asp Gly Leu Tyr Ala Val Asn Pro Glu Asn Gly Phe Phe Gly Val
305 310 315 320
Ala Pro Gly Thr Asn Tyr Ala Ser Asn Pro Ile Ala Met Lys Thr Met
325 330 335
Glu Pro Gly Asn Thr Leu Phe Thr Asn Val Ala Leu Thr Asp Asp Gly
340 345 350
Asp Ile Trp Trp Glu Gly Met Asp Gly Asp Ala Pro Ala His Leu Ile
355 360 365
Asp Trp Met Gly Asn Asp Trp Thr Pro Glu Ser Asp Glu Asn Ala Ala
370 375 380
His Pro Asn Ser Arg Tyr Cys Val Ala Ile Asp Gln Ser Pro Ala Ala
385 390 395 400
Ala Pro Glu Phe Asn Asp Trp Glu Gly Val Lys Ile Asp Ala Ile Leu
405 410 415
Phe Gly Gly Arg Arg Ala Asp Thr Val Pro Leu Val Thr Gln Thr Tyr
420 425 430
Asp Trp Glu His Gly Thr Met Val Gly Ala Leu Leu Ala Ser Gly Gln
435 440 445
Thr Ala Ala Ser Ala Glu Ala Lys Val Gly Thr Leu Arg His Asp Pro
450 455 460
Met Ala Met Leu Pro Phe Ile Gly Tyr Asn Ala Gly Glu Tyr Leu Gln
465 470 475 480
Asn Trp Ile Asp Met Gly Asn Lys Gly Gly Asp Lys Met Pro Ser Ile
485 490 495
Phe Leu Val Asn Trp Phe Arg Arg Gly Glu Asp Gly Arg Phe Leu Trp
500 505 510
Pro Gly Phe Gly Asp Asn Ser Arg Val Leu Lys Trp Val Ile Asp Arg
515 520 525
Ile Glu Gly His Val Gly Ala Asp Glu Thr Val Val Gly His Thr Ala
530 535 540
Lys Ala Glu Asp Leu Asp Leu Asp Gly Leu Asp Thr Pro Ile Glu Asp
545 550 555 560
Val Lys Glu Ala Leu Thr Ala Pro Ala Glu Gln Trp Ala Asn Asp Val
565 570 575
Glu Asp Asn Ala Glu Tyr Leu Thr Phe Leu Gly Pro Arg Val Pro Ala
580 585 590
Glu Val His Ser Gln Phe Asp Ala Leu Lys Ala Arg Ile Ser Ala Ala
595 600 605
His Ala
610
<210> 2
<211> 1833
<212> DNA
<213> Unknown
<220>
<223> Нуклеотидная последовательность фосфоенолпируваткарбоксикиназы
<400> 2
atgactactg ctgcaatcag gggccttcag ggcgaggcgc cgaccaagaa taaggaactg 60
ctgaactgga tcgcagacgc cgtcgagctc ttccagcctg aggctgttgt gttcgttgat 120
ggatcccagg ctgagtggga tcgcatggcg gaggatcttg ttgaagccgg taccctcatc 180
aagctcaacg aggaaaagcg tccgaacagc tacctagctc gttccaaccc atctgacgtt 240
gcgcgcgttg agtcccgcac cttcatctgc tccgagaagg aagaagatgc tggcccaacc 300
aacaactggg ctccaccaca ggcaatgaag gacgaaatgt ccaagcatta cgctggttcc 360
atgaaggggc gcaccatgta cgtcgtgcct ttctgcatgg gtccaatcag cgatccggac 420
cctaagcttg gtgtgcagct cactgactcc gagtacgttg tcatgtccat gcgcatcatg 480
acccgcatgg gtattgaagc gctggacaag atcggcgcga acggcagctt cgtcaggtgc 540
ctccactccg ttggtgctcc tttggagcca ggccaggaag acgttgcatg gccttgcaac 600
gacaccaagt acatcaccca gttcccagag accaaggaaa tttggtccta cggttccggc 660
tacggcggaa acgcaatcct ggcaaagaag tgctacgcac tgcgtatcgc atctgtcatg 720
gctcgcgaag aaggatggat ggctgagcac atgctcatcc tgaagctgat caacccagag 780
ggcaaggcgt accacatcgc agcagcattc ccatctgctt gtggcaagac caacctcgcc 840
atgatcactc caaccatccc aggctggacc gctcaggttg ttggcgacga catcgcttgg 900
ctgaagctgc gcgaggacgg cctctacgca gttaacccag aaaatggttt cttcggtgtt 960
gctccaggca ccaactacgc atccaaccca atcgcgatga agaccatgga accaggcaac 1020
accctgttca ccaacgtggc actcaccgac gacggcgaca tctggtggga aggcatggac 1080
ggcgacgccc cagctcacct cattgactgg atgggcaacg actggacccc agagtccgac 1140
gaaaacgctg ctcaccctaa ctcccgttac tgcgtagcaa tcgaccagtc cccagcagca 1200
gcacctgagt tcaacgactg ggaaggcgtc aagatcgacg caatcctctt cggtggacgt 1260
cgcgcagaca ccgtcccact ggttacccag acctacgact gggagcacgg caccatggtt 1320
ggtgcactgc tcgcatccgg tcagaccgca gcttccgcag aagcaaaggt cggcacactc 1380
cgccacgacc caatggcaat gctcccattc attggctaca acgctggtga atacctgcag 1440
aactggattg acatgggtaa caagggtggc gacaagatgc catccatctt cctggtcaac 1500
tggttccgcc gtggcgaaga tggacgcttc ctgtggcctg gcttcggcga caactctcgc 1560
gttctgaagt gggtcatcga ccgcatcgaa ggccacgttg gcgcagacga gaccgttgtt 1620
ggacacaccg ctaaggccga agacctcgac ctcgacggcc tcgacacccc aattgaggat 1680
gtcaaggaag cactgaccgc tcctgcagag cagtgggcaa acgacgttga agacaacgcc 1740
gagtacctca ctttcctcgg accacgtgtt cctgcagagg ttcacagcca gttcgatgct 1800
ctgaaggccc gcatttcagc agctcacgct taa 1833
<210> 3
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 1
<400> 3
ggggatcctc tagagtcgac ctgcgacgac tggaaaacca tgg 43
<210> 4
<211> 47
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 2
<400> 4
gcagttctta agcgtgaact actagttaaa actttaggtg agacaac 47
<210> 5
<211> 47
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 3
<400> 5
gttgtctcac ctaaagtttt aactagtagt tcacgcttaa gaactgc 47
<210> 6
<211> 42
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 4
<400> 6
gcttgcatgc ctgcaggtcg acggctggac cctagaattc gg 42
<210> 7
<211> 42
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 5
<400> 7
ggggatcctc tagagtcgac gtggcgtttg aaaccccgga ag 42
<210> 8
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 6
<400> 8
catcgagcca cacgctccag tgaacaagga cctctacgaa c 41
<210> 9
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 7
<400> 9
gttcgtagag gtccttgttc actggagcgt gtggctcgat g 41
<210> 10
<211> 44
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 8
<400> 10
gcttgcatgc ctgcaggtcg acttagcagt cgaagtacaa ttcg 44
<210> 11
<211> 477
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность glnA(D401N)
<400> 11
Val Ala Phe Glu Thr Pro Glu Glu Ile Val Lys Phe Ile Lys Asp Glu
1 5 10 15
Asn Val Glu Phe Val Asp Val Arg Phe Thr Asp Leu Pro Gly Thr Glu
20 25 30
Gln His Phe Ser Ile Pro Ala Ala Ser Phe Asp Ala Asp Thr Ile Glu
35 40 45
Glu Gly Leu Ala Phe Asp Gly Ser Ser Ile Arg Gly Phe Thr Thr Ile
50 55 60
Asp Glu Ser Asp Met Asn Leu Leu Pro Asp Leu Gly Thr Ala Thr Leu
65 70 75 80
Asp Pro Phe Arg Lys Ala Lys Thr Leu Asn Val Lys Phe Phe Val His
85 90 95
Asp Pro Phe Thr Arg Glu Ala Phe Ser Arg Asp Pro Arg Asn Val Ala
100 105 110
Arg Lys Ala Glu Gln Tyr Leu Ala Ser Thr Gly Ile Ala Asp Thr Cys
115 120 125
Asn Phe Gly Ala Glu Ala Glu Phe Tyr Leu Phe Asp Ser Val Arg Tyr
130 135 140
Ser Thr Glu Met Asn Ser Gly Phe Tyr Glu Val Asp Thr Glu Glu Gly
145 150 155 160
Trp Trp Asn Arg Gly Lys Glu Thr Asn Leu Asp Gly Thr Pro Asn Leu
165 170 175
Gly Ala Lys Asn Arg Val Lys Gly Gly Tyr Phe Pro Val Ala Pro Tyr
180 185 190
Asp Gln Thr Val Asp Val Arg Asp Asp Met Val Arg Asn Leu Ala Ala
195 200 205
Ser Gly Phe Ala Leu Glu Arg Phe His His Glu Val Gly Gly Gly Gln
210 215 220
Gln Glu Ile Asn Tyr Arg Phe Asn Thr Met Leu His Ala Ala Asp Asp
225 230 235 240
Ile Gln Thr Phe Lys Tyr Ile Ile Lys Asn Thr Ala Arg Leu His Gly
245 250 255
Lys Ala Ala Thr Phe Met Pro Lys Pro Leu Ala Gly Asp Asn Gly Ser
260 265 270
Gly Met His Ala His Gln Ser Leu Trp Lys Asp Gly Lys Pro Leu Phe
275 280 285
His Asp Glu Ser Gly Tyr Ala Gly Leu Ser Asp Ile Ala Arg Tyr Tyr
290 295 300
Ile Gly Gly Ile Leu His His Ala Gly Ala Val Leu Ala Phe Thr Asn
305 310 315 320
Ala Thr Leu Asn Ser Tyr His Arg Leu Val Pro Gly Phe Glu Ala Pro
325 330 335
Ile Asn Leu Val Tyr Ser Gln Arg Asn Arg Ser Ala Ala Val Arg Ile
340 345 350
Pro Ile Thr Gly Ser Asn Pro Lys Ala Lys Arg Ile Glu Phe Arg Ala
355 360 365
Pro Asp Pro Ser Gly Asn Pro Tyr Leu Gly Phe Ala Ala Met Met Met
370 375 380
Ala Gly Leu Asp Gly Ile Lys Asn Arg Ile Glu Pro His Ala Pro Val
385 390 395 400
Asn Lys Asp Leu Tyr Glu Leu Pro Pro Glu Glu Ala Ala Ser Ile Pro
405 410 415
Gln Ala Pro Thr Ser Leu Glu Ala Ser Leu Lys Ala Leu Gln Glu Asp
420 425 430
Thr Asp Phe Leu Thr Glu Ser Asp Val Phe Thr Glu Asp Leu Ile Glu
435 440 445
Ala Tyr Ile Gln Tyr Lys Tyr Asp Asn Glu Ile Ser Pro Val Arg Leu
450 455 460
Arg Pro Thr Pro Gln Glu Phe Glu Leu Tyr Phe Asp Cys
465 470 475
<210> 12
<211> 1434
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность glnA(D401N)
<400> 12
gtggcgtttg aaaccccgga agaaattgtc aagttcatca aggatgaaaa cgtcgagttc 60
gttgacgttc gattcaccga ccttcccggc accgagcagc acttcagcat cccagctgcc 120
agcttcgatg cagatacaat cgaagaaggt ctcgcattcg acggatcctc gatccgtggc 180
ttcaccacga tcgacgaatc tgacatgaat ctcctgccag acctcggaac ggccaccctt 240
gatccattcc gcaaggcaaa gaccctgaac gttaagttct tcgttcacga tcctttcacc 300
cgcgaggcat tctcccgcga cccacgcaac gtggcacgca aggcagagca gtacctggca 360
tccaccggca ttgcagacac ctgcaacttc ggcgccgagg ctgagttcta cctcttcgac 420
tccgttcgct actccaccga gatgaactcc ggcttctacg aagtagatac cgaagaaggc 480
tggtggaacc gtggcaagga aaccaacctc gacggcaccc caaacctggg cgcaaagaac 540
cgcgtcaagg gtggctactt cccagtagca ccatacgacc aaaccgttga cgtgcgcgat 600
gacatggttc gcaacctcgc agcttccggc ttcgctcttg agcgtttcca ccacgaagtc 660
ggtggcggac agcaggaaat caactaccgc ttcaacacca tgctccacgc ggcagatgat 720
atccagacct tcaagtacat catcaagaac accgctcgcc tccacggcaa ggctgcaacc 780
ttcatgccta agccactggc tggcgacaac ggttccggca tgcacgctca ccagtccctc 840
tggaaggacg gcaagccact cttccacgat gagtccggct acgcaggcct gtccgacatc 900
gcccgctact acatcggcgg catcctgcac cacgcaggcg ctgttctggc gttcaccaac 960
gcaaccctga actcctacca ccgtctggtt ccaggcttcg aggctccaat caacctggtg 1020
tactcacagc gcaaccgttc cgctgctgtc cgtatcccaa tcaccggatc caacccgaag 1080
gcaaagcgca tcgaattccg cgctccagac ccatcaggca acccatacct gggctttgca 1140
gcgatgatga tggccggcct cgacggcatc aagaaccgca tcgagccaca cgctccagtg 1200
aacaaggacc tctacgaact accaccagag gaagctgcat ccattccaca ggcaccaacc 1260
tccctggaag catccctgaa ggcactgcag gaagacaccg acttcctcac cgagtctgac 1320
gtcttcaccg aggatctcat cgaggcgtac atccagtaca agtacgacaa cgagatctcc 1380
ccagttcgcc tgcgcccaac cccgcaggaa ttcgaattgt acttcgactg ctaa 1434
<---
Изобретение относится к биотехнологии. Предложен способ получения L-глутамина, включающий культивирование микроорганизма Corynebacterium glutamicum, обладающего ослабленной активностью фосфоенолпируваткарбоксикиназы вследствие делеции гена, кодирующего фосфоенолпируваткарбоксикиназу, в среде. Также предложено применение указанного микроорганизма Corynebacterium glutamicum для получения L-глутамина. Изобретение обеспечивает продуцирование L-глутамина с высоким выходом. 2 н. и 5 з.п. ф-лы, 4 табл., 4 пр.
1. Способ получения L-глутамина, включающий культивирование микроорганизма Corynebacterium glutamicum, обладающего ослабленной активностью фосфоенолпируваткарбоксикиназы вследствие делеции гена, кодирующего фосфоенолпируваткарбоксикиназу, в среде.
2. Способ получения L-глутамина по п. 1, дополнительно включающий выделение L-глутамина из среды или микроорганизма после стадии культивирования.
3. Способ получения L-глутамина по п. 1, где фосфоенолпируваткарбоксикиназа представляет собой эндогенный белок микроорганизма Corynebacterium glutamicum.
4. Способ получения L-глутамина по п. 1, где фосфоенолпируваткарбоксикиназа состоит из аминокислотной последовательности SEQ ID NO: 1 или аминокислотной последовательности, на 90% или более идентичной SEQ ID NO: 1.
5. Применение микроорганизма Corynebacterium glutamicum для получения L-глутамина, где микроорганизм обладает ослабленной фосфоенолпируваткарбоксикиназой вследствие делеции гена, кодирующего фосфоенолпируваткарбоксикиназу.
6. Применение по п. 5, где фосфоенолпируваткарбоксикиназа представляет собой эндогенный белок микроорганизма Corynebacterium glutamicum.
7. Применение по п. 5, где фосфоенолпируваткарбоксикиназа состоит из аминокислотной последовательности SEQ ID NO: 1 или аминокислотной последовательности, на 90% или более идентичной SEQ ID NO: 1.
| US 20020065403 A1, 30.05.2002 | |||
| Мутантный штамм Corynebacterium glutamicum, продуцирующий L-глутамин (варианты), и способ получения L-глутамина | 2015 |
|
RU2665830C1 |
| МУТАНТНАЯ ГЛУТАМИНСИНТЕТАЗА, ФРАГМЕНТ ДНК, ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ L-ГЛУТАМИНА И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2001 |
|
RU2230114C2 |
| WO 2016032158 A1, 03.03.2016. | |||

