Изобретение относится к области биотехнологии, может быть использовано в микробиологической промышленности для получения фермента, используемого в генетической инженерии.
Эндонуклеаза рестрикции (рестриктаза) Есо RV расщепляет двунитевую ДНК по фосфодиэфирной связи в последовательности нуклеотидов ГАТАТЦ и, в отличие от большинства известных рестриктаз, способна расщеплять 5-оксиметилцитозинсодержащие и глюкозилированные ДНК Т-четных бактериофагов.
Цель изобретения обеспечение электрофоретической гомогенности препарата, не содержащего интерферирующих примесей ферментов нуклеинового обмена.
Кроме того, изобретение позволяет удлинить срок хранения без снижения активности целевого продукта и транспортировки в широком диапазоне температур.
На чертеже представлена электрофореграмма очищенного препарата эндонуклеазы рестрикции Eco RV и маркерных белков, где 1 экстракт белков Е. соli; 2 цитохром С; 3 миоглобин; 4 рестриктаза Eco RV; 5 альдолаза; 6 яичный альбумин; 7 бычий сывороточный альбумин.
Изобретение поясняется примерами.
П р и м е р 1. Клетки Е. соli ВКМ В-1450Д, содержащие рекомбинантную плазмидную ДНК с генами рестрикции-модификации Есо RV, выращивают в LB-среде (1% бакто-триптона, 0,5% дрожжевого экстракта, 1% NaCl, 0,5% глюкозы, рН 7,2-7,4) до ОД550 0,8 при 37оС. Культуральную жидкость охлаждают до 4оС во льду и клетки осаждают центрифугированием при 5500 об/мин 30 мин. Полученную биомассу клеток хранят при (-20) (-40оС).
Берут 50 г (сырой вес) биомассы Е. соli ВКМ В-1450Д и суспендируют в 250 мл буферного раствора, содержащего 50 мМ калий фосфатного буфера рН 7,0; 1 мМ ЭДТА, 1 мМ NaN3; 7 мМ β -меркаптоэтанола (буфер А), 400 мМ NaCl и 10% глицерина, на магнитной мешалке при 0оС. После получения гомогенной суспензии добавляют 5 мл одного раствора лизоцима до конечной концентрации 100 мкг/мл и выдерживают при 0оС 15 мин. Полученные сферопласты клеток разрушают ультразвуком. Клеточные обломки убирают центрифугированием при 25000 об/мин в течение 2,5 ч при 4оС.
Надосадочную жидкость переносят в мерный цилиндр (осадки отбрасывают) и разводят в два раза буферным раствором А без хлористого натрия до конечной концентрации NaCl 200 мМ.
Полученный экстракт белков клеток Е. соli ВКМ В-1450Д разделяют путем хроматографии на фосфоцеллюлозе Р11.
Хроматография на фосфоцеллюлозе Р11.
Колонку с фосфоцеллюлозой уравновешивают буферным раствором А, содержащим 200 мМ NaCl и 10% глицерина. Бесклеточный экстракт наносят на колонку с фосфоцеллюлозой со скоростью 40 мл/ч. После нанесения колонку промывают 400 мл буферного раствора, которым была уравновешена колонка с фосфоцеллюлозой. Элюцию белков проводят линейным градиентом концентрации хлористого натрия от 200 до 800 мМ, в буфере А общим объемом 1600 мл. Фракции собирают по 10 мл.
Активность рестриктазы Есо RV определяют в каждой десятой фракции при разведении в 100 раз. Для этого из каждой анализируемой фракции отбирают по 10 мкл, добавляют 990 мкл буферного раствора А и хорошо перемешивают.
Условия определения активности рестриктазы Eco RV следующие: 1,2 мкг ДНК бактериофага Т4 (можно заменить ДНК бактериофагов λ, ⊘ 80 и др. или плазмидной ДНК), 50 мМ трис-HCl рН 7,5, 50 мМ NaCl, 10 мМ MgCl2 в объеме 50 мкл, 1 мкл элюата в разведении 1:100 при 37оС, в течение 15 мин. Продукты реакции анализируют электрофорезом в 0,6-0,8% агарозном геле, трис-ацетатный буфер (трис-боратный буфер). После электрофореза гель окрашивают в водном растворе бромистого этидия (5 мкг/мл) в течение 15 мин, отмывают в дистиллированной воде в две смены и фотографируют в УФ-свете ( λ= 254 нм), используя оранжевый светофильтр и пленку МЗ-2 или микрат 300.
За единицу активности рестриктазы Есо RV принимают минимальное количество фермента, способного расщеплять 1 мкг субстратной ДНК (ДНК фагов Т4, λ и ⊘ 80) за 15 мин при 37оС.
Рестриктаза Есо RV элюируется с фосфоцеллюлозы в интервале концентрации хлористого натрия до 500 до 600 мМ.
Для дальнейшей очистки берут фракции с удельной активностью рестриктазы Есо RV не менее 100 ед/мкл, что составляет 1/5 от максимальной высоты пика удельной активности фермента. Фракции с 80 по 110 объединяют (общий объем 300 мл) и разводят в 2,5 раза буфером А без хлористого натрия (температура 4оС) до конечной концентрации хлористого натрия около 200 мМ и наносят на колонку с голубой сефарозой 6 В.
Хроматография на голубой сефарозе.
Колонку (2,5 х 40 см) с голубой сефарозой промывают 400 мл 1 М NaCl и уравновешивают буфером А, содержащим 200 м М NaCl. Объединенные фракции после хроматографии на фосфоцеллюлозе, содержащие рестриктазу Есо RV, наносят на колонку с голубой сефарозой со скоростью 50 мл/ч и промывают 400 мл буферного раствора, которым уравновешена колонка. Рестриктазу элюируют линейным градиентом концентрации хлористого натрия от 200 до 1000 мМ в буфере А. Фракции собирают по 10 мл. Активность рестриктазы Eco RV определяют в каждой десятой фракции в разведении 1:50. Фермент элюируется с колонки в интервале концентрации хлористого натрия от 400 до 600 мМ. Фракции (с 40 по 70), содержащие рестриктазу Есо RV с удельной активностью 50 ед/мкл и более, объединяют и очищают на гидроксилапатите (ГАП).
Хроматография на гидроксилапатите (ГАП).
Колонку с гидроксилапатитом (1,5 х 10 см) промывают 500 мМ калийфосфатным буфером, рН 7,0, и уравновешивают буферным раствором, содержащим 25 мМ калийфосфатного буфера рН 7,0, 250 мМ NaCl, 1 мМ ЭДТА, 1 мМ NaN3, 7 мМ β-меркаптоэтанола. Объединенные фракции после хроматографии на голубой сефарозе разводят в 2 раза раствором, содержащим 1 мМ ЭДТА, 1 мМ NaN3, 7 мМ -меркаптоэтанола до конечной концентрации калий-фосфатного буфера 25 мМ и хлористого натрия около 250 мМ. После нанесения колонку с ГАП промывают буферным раствором, которым была уравновешена колонка. Рестриктазу Eco RV элюируют с ГАП линейным градиентом концентрации калий-фосфатного буфера от 250 до 500 мМ рН 7,0 общим объемом 250 мл. Фракции собирают по 5 мл. Фермент элюируется с ГАП в интервале концентраций калий-фосфатного буфера рН 7,0 от 175 до 300 мМ.
Функции (с 15 по 25), содержащие рестриктазу Есо RV с удельной активностью 50 ед/мкл и более, объединяют (общий объем 50 мл) и диализуют против 500 мл буфера А, содержащего 200 мМ NaCl, три смены через 6 ч при 4оС.
Стабилизация рестриктазы Есо RV лиофилизацией.
К полученному препарату фермента (50 мл) добавляют 10 мг бычьего сывороточного альбумина до конечной концентрации 200 мкг/мл, растворяют и препарат переносят в толстостенную емкость, замораживают раствор фермента на внутренних стенках емкости, используя сухой лед (-70оС). После чего препарат рестриктазы лиофилизуют. После лиофилизации препарат фермента собирают со стенок стеклянной палочкой в виде хлопьев (порошка) белого цвета. Лиофилизированный препарат рестриктазы Есо RV хранится без доступа воздуха при температуре от (-70) (+22)оС в течение 6 мес.
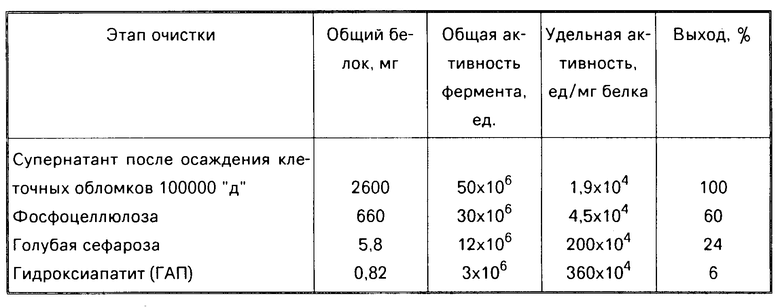
Степень очистки и выход рестриктазы Есо RV на этапах очистки при использовании биомассы с удельной активностью 1 х 106 ед/г сырых клеток Е. соli ВКМ В-1450Д представлены в таблице.
В случае использования для выделения фермента биомассы клеток с удельной активностью 5х106 и более ед/г сырых клеток, получают препарат фермента с удельной активностью 15х106 ед/мг белка и более с выходом более 10%
П р и м е р 2. Способ осуществляют как в примере 1, используя для выделения рестриктазы Есо RV биомассу Е. соli ВКМR-4D активностью 10х106 ед/г сырых клеток Получают фермент-рестриктазу Есо RV с удельной активностью 20х106 ед/мг белка.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТОВ СИСТЕМЫ РЕСТРИКЦИИ-МОДИФИКАЦИИ | 1984 |
|
SU1262952A1 |
| СПОСОБ ПОЛУЧЕНИЯ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ BSE 21 I | 1999 |
|
RU2184777C2 |
| Способ выделения эндонуклеазы рестрикции Ара 1 | 1989 |
|
SU1655986A1 |
| СПОСОБ ПОЛУЧЕНИЯ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ HINF I | 1997 |
|
RU2136749C1 |
| Способ выделения рестриктаз II класса | 1989 |
|
SU1698289A1 |
| Способ получения рестриктазы С @ I | 1988 |
|
SU1546485A1 |
| Способ конструирования плазмидной ДНК,штамм @ @ -продуцент эндонуклеазы рестрикции @ и способ получения эндонуклеазы рестрикции @ | 1981 |
|
SU1040791A1 |
| Способ получения сайт-специфической эндонуклеазы @ @ | 1983 |
|
SU1120019A1 |
| Способ получения эндонуклеазы-рестриктазы, расщепляющей последовательность нуклеотидов 5 @ =G @ CGC=3 @ | 1988 |
|
SU1576565A1 |
| Способ получения рестриктазы, способной узнавать и расщеплять последовательность нуклеотидов GTCGAC | 1989 |
|
SU1752769A1 |

| Способ конструирования плазмидной ДНК,штамм @ @ -продуцент эндонуклеазы рестрикции @ и способ получения эндонуклеазы рестрикции @ | 1981 |
|
SU1040791A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
