Изобретение относится к лимфокинам, в частности к синергетическому действию фактора некроза опухолей (ФНО) и интерлейкина-4 (ИЛ-4) на ингибирование роста опухолевых клеток и цитотоксичность.
Цитокином называют любую молекулу, которая продуцируется клеткой и поражает ее или другие клетки. Цитокины очень разнообразны и вместе взятые оказывают самое различное действие как на нормальные, так и на патологически измененные клетки. Влияние определенного цитокина на данную клетку определяется как природой цитокина, так и типом клетки-мишени.
Фактор некроза опухолей (ФНО) представляет собой цитокин, который продуцируется в первую очередь активированными моноцитами и весьма плейотропен по своей природе (Agier et al. 1988, Факторы некроза опухолей. Справочник по цитолитическим лимфоцитам и компонентам: Эффекторы иммунной системы, E.R. Podak, ed. , CRC Press Inc. Boca Raton, Fla., p. 105; Goeddel et al., 1987, Факторы некроза опухолей: Структура генов и биологическая активность In: Coldspring Harbor Sumposia on Quantative Biology (1, 597). ФНО ингибирует рост самых разнообразных опухолевых клеток в культуре, но стимулирует рост фибробластов, B-клеток и тимоцитов. Цитотоксическое действие ФНО на опухолевые клетки усиливается интерферонами (Lee et al., 1984, J. Immunol., 133, 103; Sugarman et al., 1985, Science, 230, 1984; Aggarwal et al, 1985, Nature, 318, 665; Vilcek et al., 1986, Взаимодействие между фактором некроза опухолей интерферонами, In: Biology of the Interferon System, Edited by H. Schelllehens and W.E. Stewart, Science Publishers, Amsterdam, p. 249, Fransen et al., 1986, Eur. J Cancer Clin. Oncol., 22, 419-426; Campbell et al., J. Immunol., 141, Schiller et al., 1987 Canc. Res., 47, 2809-2813). Усиление антипролиферативного действия ФНО интерферонами сопровождает восходящую регуляцию рецепторов ФНО (Aggarwal et al., 1985, Nature 318, 665; Fransen et al. , 1986, Eur. J. Cancer. Clin. Oncol., 22, 419-426). Однако восходящая регуляция рецепторов ФНО интерферонами не является первичным механизмом их синергетической цитотоксической реакции (Tsujimoto et al., 1986, J. Immunol. , 137, 2272-2276; Aggrawal and Eessalu, 1987, J. Biol. Chem. 262, 10000-10007).
Синергетические взаимодействия γ- интерферона и ФНО при регуляции роста опухолей описаны в патенте США N 4650674.
Интерлейкин-4 (ИЛ-4) представляет собой иммуномодуляторный цитокин, продуцируемый активированными Т-лимфоцитами. В качестве одного из модуляторов иммунной системы он был впервые описан как ростовой фактор B-клеток благодаря его способности ко-стимулировать пролиферацию B-лимфоцитов (Howard et al., J. Exp. Med. 155, 914, 1982). ИЛ-4 влияет также на другие самые разнообразные клетки, в том числе Т-клетки, тучные клетки, макрофаги и предшественники кровеносных клеток (Paul et al., 1987, Ann. Rev. Immunol, 5, 429; Yokota et al., 1988, Immunol. Rev. 102, 137; Crabstein et al. 1987, J. Exp. Med. 1633, 1405). ИЛ-4 усиливает рост отдельных типов клеток, например, Т-клеток и тучных клеток (Fernander - Botran et al., 1986, Proc. Natl. Acad. Sci. USA, 83, 9689: Spits et al., 1987, J. Immunol., 139, 1142; Zlotnik et al. , 1987, Proc. Natl. Acad. Sci., USA, 84, 3856; Mosmann et al., 1986, Proc. Natl. Acad. Sci, USA, 83, 5654), индуцирует экспрессию антигенов МНС класса II на B-клетках и моноцитах (Tevede et al., 1988, J. Immunol., 140, 1548), повышает содержание антиген-специфичных цитотоксичных Т-клеток и усиливает опосредуемую через макрофаги цитотоксическую активность (Spits et al. , 1988, J. Immunol., 141, 39; Widmer et al., 1987, J. Exp. Med., 166, 1447; Crawford et al., 1987, J. Immunol., 139, 135).
Оказывая нисходящее регулирующее действие на продукцию некоторых цитокинов, в том числе ИЛ-4-1- α , ИЛ- β , γ -ИФН и α- ФНО, ИЛ-4 влияет также на продукцию ИЛ-6 B-клетками по восходящему пути (Essner et al., 1989, J. Immunol. , 142, 3857; Lee et al., 1990, J. of Leu Kocyte Biology, 47, 457-479; Pelman et al. , 1989, J. Exp. Med. 170, 175). Индуцируемая ИЛ-2 активность клеток LAK и NK регулируется ИЛ-4 по нисходящему механизму (Brooks et al., 1988, Clin. Exp. Immunol, 74, 162; Nagler et al., 1988, J. Immunol., 141, 2349). Недавно было установлено, что помимо пролиферативной активности ИЛ-4 обладает способностью ингибировать in vitro рост человеческих лимфоидных образований и опухолей крови (Taylor et al., 1990, Blood, 75, 5).
Изобретение относится к синергетическому действию ИЛ-4 на антипролиферативную активность ФНО. ИЛ-4 усиливает цитотоксическое действие ФНО на самые разнообразные типы клеток, в том числе опухолевые клетки грудной железы, клетки карциномы и лимфомы и многие другие. Синергетическое действие зависит от доз ФНО и ИЛ-4. Степень усиления цитотоксической активности ФНО под воздействием ИЛ-4 сопоставима с ранее описанной для гамма-интерферона.
Изобретение относится также к синергетическому действию интерлейкина-4, продуцируемого T-клетками, на цитотоксическую активность ФНО в опухолевых клетках. В одном из своих воплощений изобретение позволяет получить композицию, в состав которой входят очищенные ФНО и ИЛ-4. Другой аспект изобретения касается получения композиционного материала, имеющего в своем составе высокоочищенные ФНО, ИЛ-4 и гамма-интерферон (γ -ИФН). Еще один аспект изобретения - применение указанной композиции для ингибирования роста опухолевых клеток. Наконец, согласно изобретению эту композицию используют для уничтожения опухолевых клеток.
Антипролиферативная активность ФНО в отношении большого числа различных клеточных линий усиливается ИЛ-4. Сам по себе ИЛ-4 не обладает способностью подавлять рост этих клеточных линий, а в отдельных случаях слабо усиливает его. В количественном отношении степень стимулирующего действия ИЛ-4 на цитотоксическую активность ФНО сопоставима с таковой γ-ИФН. Тем не менее механизм стимулирующего действия ИЛ-4 и γ-ИФН на антипролиферативную или цитотоксическую активность ФНО и лимфотоксина (Лт) отличаются.
Введение животным с опухолью бесклеточной композиции, имеющей в своем составе фактор некроза опухолей и интерлейкин-4, для обработки опухолевых клеток приводит к угнетению роста опухоли. Предпочтительно, чтобы фактор некроза опухолей был человеческого происхождения. В принципе его можно получать из любого хорошо известного источника. В оптимальном случае фактор некроза опухолей получают методами генной инженерии.
ИЛ-4 также должен быть по возможности идентичен человеческому ИЛ-4. Его можно получать из любого известного источника. В оптимальном случае его получают по рекомбинантной технологии.
Изобретение относится также к способу лечения опухолей, согласно которому композицию, содержащую ФНО и ИЛ-4, вводят больному животному в количестве, которое считают эффективным для достижения желаемого результата.
Помимо этого изобретение позволяет получать композицию, в состав которой входят ФНО, ИЛ-4 и γ -ИФН. При введении такой композиции животным с опухолью она вызывает уменьшение скорости роста последней и оказывает избирательное цитотоксическое действие на опухолевые клетки.
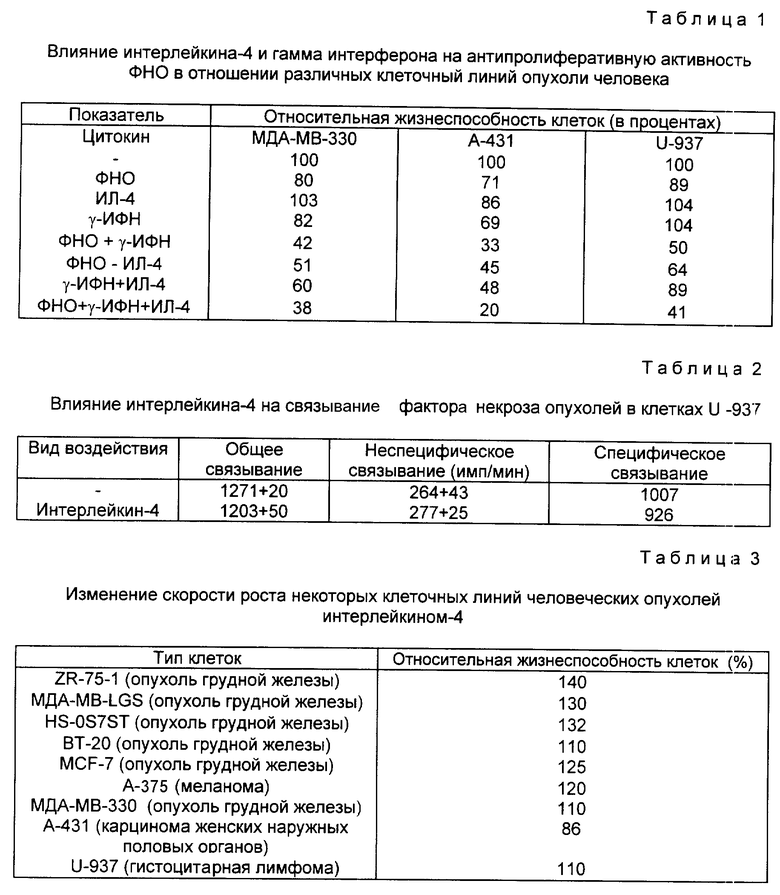
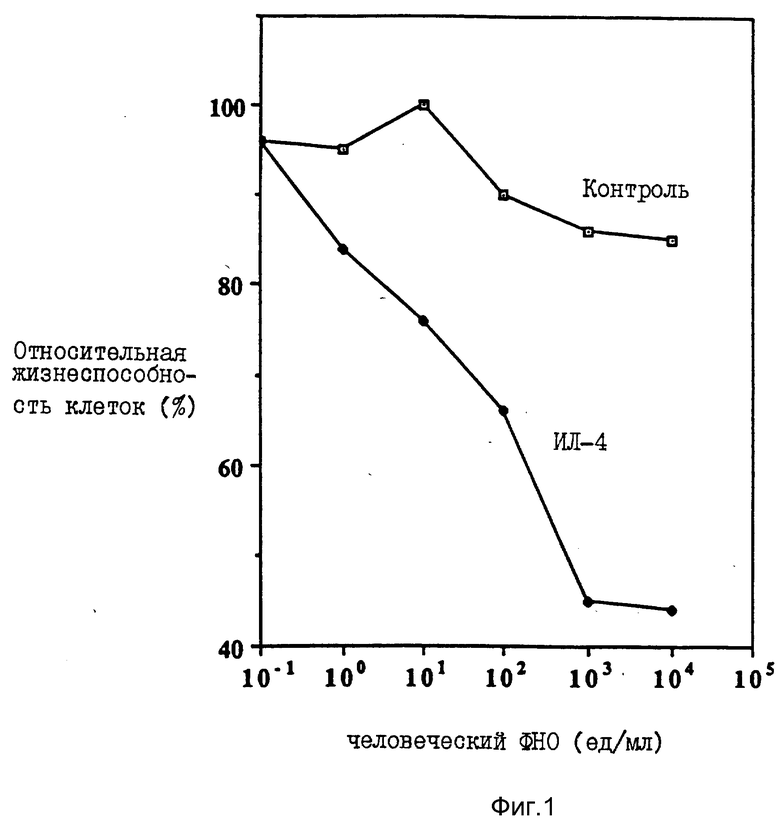
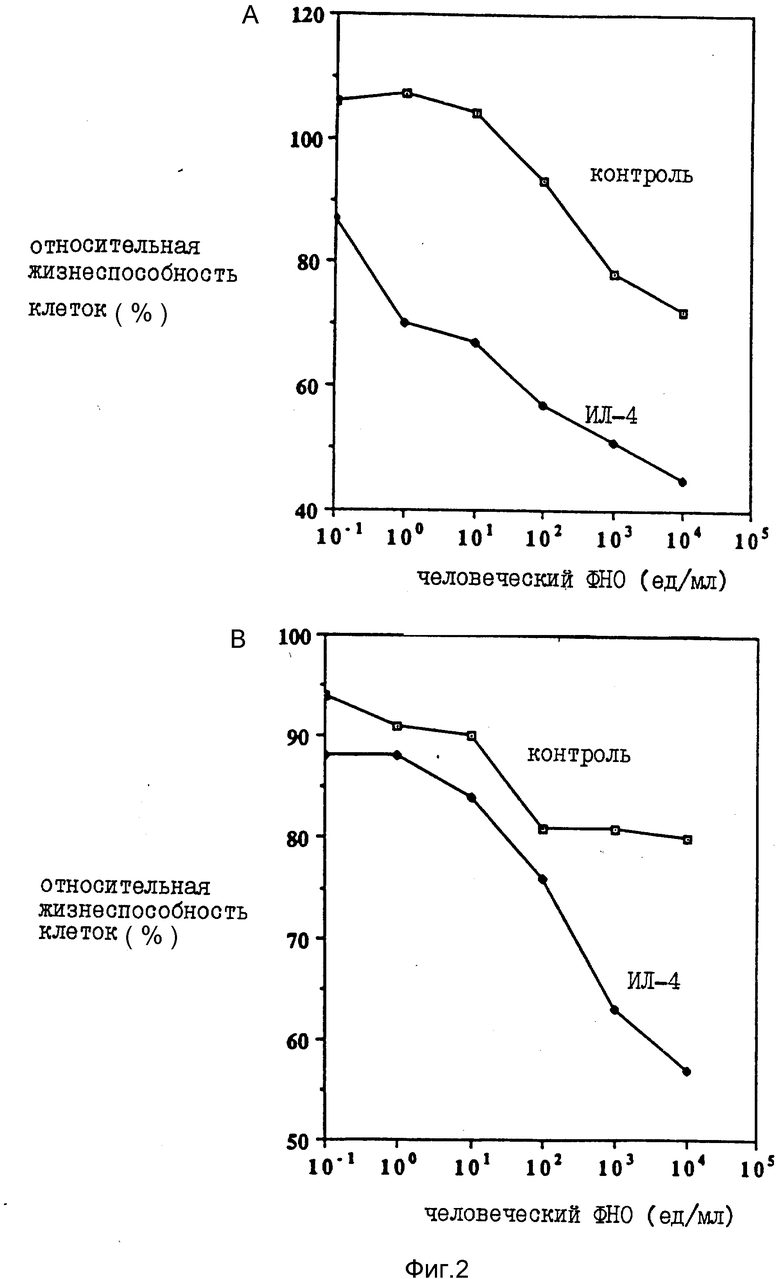
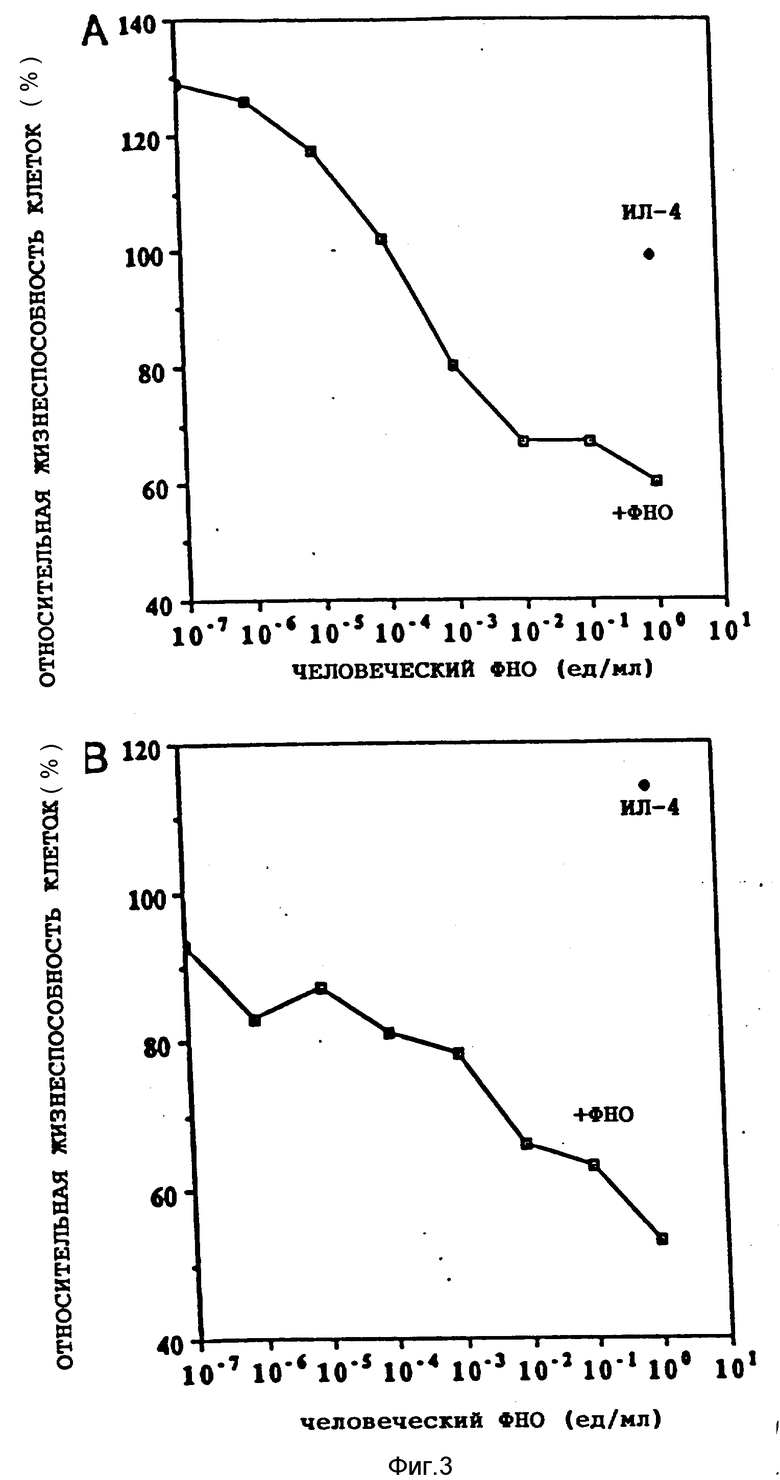
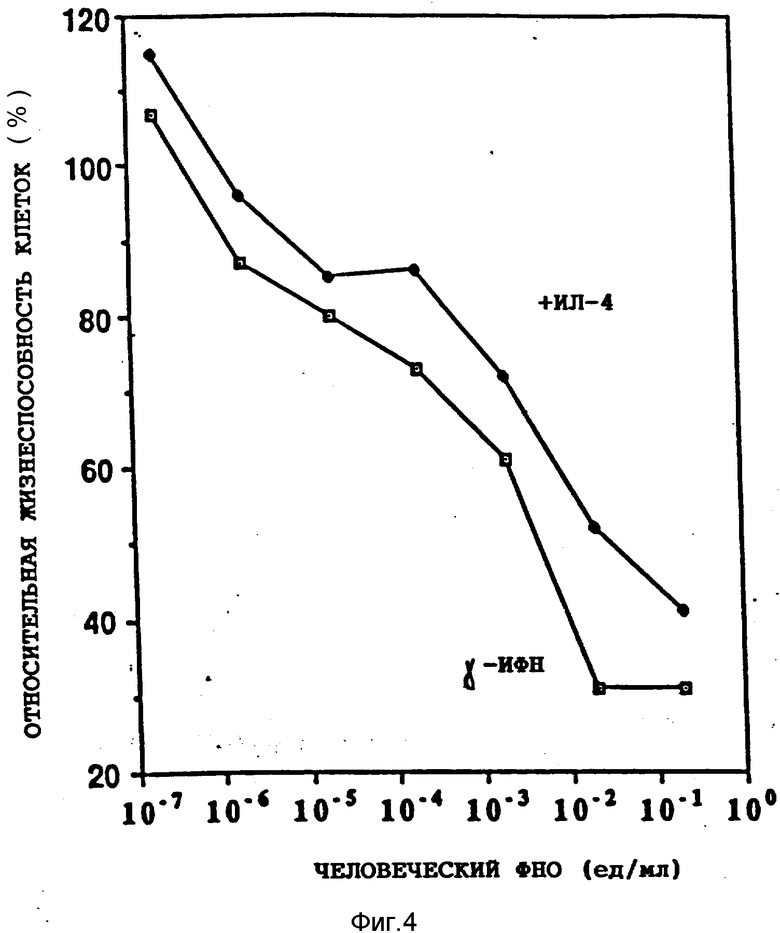
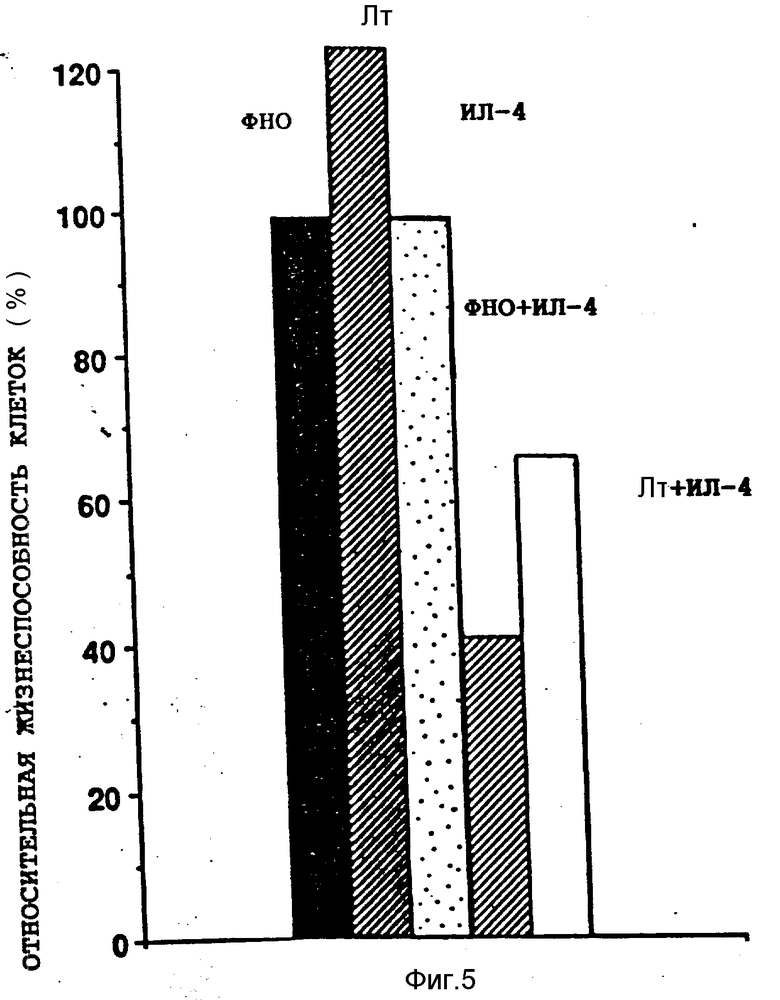
На фиг. 1 иллюстрируется действие разных концентраций ФНО и ИЛ-4 (100 нг/мл) на жизнеспособность клеток опухоли молочной железы женщин (линия МДА-МВ-330); на фиг. 2 - влияние различных концентраций ФНО и ИЛ-4 (100 нг/мл) на жизнеспособность клеточной линии А-431 карциномы человеческого эпителия (A) и клеточной линии U-937 гистиоцитарной лимфомы человека (B); на фиг. 3 - действие различных концентраций ИЛ-4 на цитотоксическую активность ФНО в отношении клеточной линии А-431 (A) и линии МДА-МВ-330 (B); на фиг. 4 - сравнительное действие ИЛ-4 и γ -ИФН на цитотоксическую активность ФНО в отношении клеточной линии МДА-МВ-330; на фиг. 5 - сравнительное действие ИЛ-4 на цитотоксическую активность ФНО и Лт в отношении клеток МДА-МВ-330.
Изобретение касается способа усиления антипролиферативной активности ФНО в опухолевых клетках путем введения в опухоль одновременно ИЛ-4 и ФНО. ИЛ-4 усиливает цитотоксическое действие ФНО на самые разнообразные виды опухолевых клеток, неограничивающий перечень которых включает клетки опухоли грудных желез (линия МДА-МВ-330), клетки карциномы женских наружных половых органов (линия А-431), клетки гистиоцитарной лимфомы человека (линия U-937). Синергетический эффект зависит от дозы ФНО и ИЛ-4. Сам по себе ИЛ-4 обладает слабо выраженным действием на клетки указанных типов, а в некоторых случаях даже проявляет выраженную пролиферативную активность. Степень стимулирующего действия ИЛ-4 на цитотоксичность ФНО сопоставима с таковой γ-ИФН.
Изобретение позволяет получать синергетическую композицию, имеющую в своем составе ФНО и цитокин Т-клеток: ИЛ-4. Эта композиция оказывает более выраженное цитотоксическое действие на опухолевые клетки нежели один ФНО. Изобретение позволяет также получать композицию, содержащую высокоочищенные ФНО и ИЛ-4. Эти цитокины могут иметь своими источниками любые материалы, включая природные и рекомбинантные белки. Согласно другому аспекту изобретения получают композиционный материал, содержащий высокоочищенные ФНО, ИЛ-4 и гамма-интерферон ( γ -ИНФ). Кроме того, изобретение относится к применению такой композиции для подавления роста опухолевых клеток. Наконец, ее можно использовать для уничтожения таких клеток.
Понятие "индивидуум" в данном контексте означает любое животное, предпочтительно млекопитающее, еще более предпочтительно грызуна, кошку, собаку, корову или человека.
Термин "фактор некроза опухолей" употребляется для обозначения полипептида отличающегося от лимфотоксина, который характеризуется наличием преобладающей цитотоксической активности и аминокислотной последовательности, функционально гомологичной зрелому фактору некроза опухолей; а также фрагмента такого полипептида и производного самого полипептида или его фрагмента. Подробная характеристика фактора некроза опухолей, который может быть использован для реализации данного изобретения, приведена в патенте США N 4650674.
Термином "преобладающая цитотоксическая активность" обозначают усиленные разрушения или подавление роста опухолевых клеток по сравнению с нормальными клетками в одних и тех же условиях. О характере преобладающей цитотоксической активности судят по воздействию полипептида на опухолевые клетки in vivo или in vitro в сопоставлении с его действием на нормальные клетки или ткани. Диагностическим признаком в условиях in vitro обычно служит клеточный лизис, тогда как в опытах in vivo о проявлениях преобладающей цитотоксической активности судят по наличию опухолевого некроза. В то же время цитотоксическая активность может проявляться в форме цитостаза или антипролиферативной активности. Системы тестирования такой активности общеизвестны. Можно использовать, в частности, описанную ниже методику оценки клеточного лизиса для определения специфической активности фактора некроза опухолей. Другой пример методики биотестирования приведен в работе B. Aggrawal с соавторами из сборника "Гормоны тимуса и лимфокины" (Thymic Hormones and Lymphokines, 1983, под редакцией A. Goldstein (Spring Symposium on Health Seiences, George Washington). Упоминаемые в этой работе клетки линии A549 могут быть получены из Американского банка опухолевых культур, где они зарегистрированы под кодовым номером CCZ185.
Специфическую активность ФНО чаще выражают в терминах его литического действия нежели цитостаза. За единицу ФНО принято такое его количество, которое необходимо для лизиса 50% клеток-мишеней. Это не исключает возможности применения других способов тестирования специфической активности ФНО, в частности, методов, основанных на оценке скорости роста клеток.
Нативный фактор некроза опухолей из нормальных биологических источников имеет молекулярную массу порядка 17000 при расчете по данным электрофореза в полиакриламидном геле с додецилсульфатом натрия (ДДС-ПААГ). Его электрическая точка равна примерно 5,3. Его изоэлектрическая точка равна примерно 5,3. Обработка ФНО трипсином вызывает множественный гидролиз его молекулы. Нативный ФНО был получен путем очистки с помощью обращенно-фазовой ВЭЖХ.
В общем случае полипептид, идентифицируемый как фактор некроза опухолей, имеет в своем составе области, гомологичные белку с активностью фактора некроза опухолей, описанному в патенте США N 4650674, или его фрагментам. Гомологичная область охватывает непрерывную последовательность аминокислотных остатков в положениях 20-100, особенно ее участки между остатками 35-66 и 110-133.
Наиболее достоверным способом подтверждения идентичности того или иного полипептида фактору некроза опухолей является выявление способности антисыворотки, вызывающей нейтрализацию цитолитической активности полной молекулы ФНО, в той же мере нейтрализовать цитолитическую активность данного полипептида. Следует, однако, признать, что иммунологическая идентичность не всегда совпадает с цитотоксической. Антитела, нейтрализующие фактор некроза опухолей, необязательно связывают идентифицируемый белок, поскольку они не обладают специфическим сродством к тем участкам фактора некроза, которые отвечают за его цитотоксическую активность. Напротив, антитела могут связываться с биологически инертными участками ФНО и вызывать его нейтрализацию вследствие стерического блокирования. Это не исключает идентичности белка фактору некроза опухолей по признакам гомологичности и биологической активности.
Следует отметить, что такие признаки ФНО, как молекулярный вес, изоэлектрическая точка и т.п., свойственные нативному фактору некроза опухолей или фактору дикого типа, выделенному из периферических лимфоцитов или культур известных клеточных линий, характеризуют только нативные формы ФНО. Однако понятие "фактор некроза опухолей" согласно вышеприведенному определению включает и другие формы, которые могут не иметь всех признаков нативного фактора. Под указанное определение подпадает не только нативный фактор некроза опухолей, но и родственные ему полипептиды с цитотоксической активностью. Так, например, производные ФНО (мутанты, полученные путем вставок или делеций, и слитые пептиды) по молекулярной массе отличаются от нативного человеческого фактора некроза опухолей (слитые белки, имеющие в своем составе полную молекулу фактора некроза опухолей; предшественники ФНО и мутанты со вставками имеют более значительную молекулярную массу, чем зрелый нативный фактор некроза опухолей, тогда как у мутантов с делециями она меньше). Точно так же фактор некроза опухолей может быть преобразован методами генной инженерии с целью уменьшения или элиминирования чувствительности к гидролитическому воздействию трипсином или другими протеазами. Наконец, пост-трансляционный процессинг предшественника человеческого ФНО в клеточных линиях млекопитающих, отличных от приматов, может сопровождаться микрогетерогенностью его N-концевого участка с заменой N-концевого аминокислотного остатка - валина.
Следует также иметь в виду, что, говоря о "способности" вызывать цитотоксический эффект или некроз опухолей in vivo, имеют в виду, что понятие "фактор некроза опухолей" включает полипептиды, которые могут быть преобразованы путем ферментативного гидролиза из неактивного состояния (как у зимогена) в полипептидный фрагмент, обладающий желаемой биологической активностью. В типичном случае неактивными предшественниками служат слитые пептиды, в составе которых полная молекула ФНО по C-концевому остатку связана пептидной связью с человеческим белком или его фрагментом. Состав аминокислотных остатков в месте пептидной связи или на примыкающих участках подбирают таким образом, чтобы иметь возможность освободить фактор некроза опухолей посредством протеолитического гидролиза в условиях in vivo либо на одном из этапов производственного регламента. Полученный таким образом фактор некроза опухолей обладает цитотоксической активностью согласно определению.
Хотя термином "фактор некроза опухолей" обычно обозначают человеческий ФНО, он в равной степени пригоден для ФНО из других источников: мышей, свиней, лошадей или крупного рогатого скота, если соответствующие полипептиды имеют признаки гомологичности и цитотоксической активности согласно определению. ФНО не обладает видовой специфичностью, иными словами, человеческий ФНО активен в отношении опухолей мышей и других животных. По этой причине ФНО одного вида животных можно использовать для терапии опухолей у другого вида.
Фактор некроза опухолей может существовать в форме многомерных молекул. Производные ФНО подпадают под определение фактора некроза опухолей. Участки молекулы фактора некроза опухолей между аминокислотными остатками в положениях 35-66 и 110-133 характеризуются высокой степенью гомологичности (50%) лимфотоксину. Гидрофобные C-концевые участки, соответствующие участку 150-157 фактора некроза опухолей, также весьма консервативны в обеих молекулах. Поскольку оба пептида проявляют цитотоксическую активность и вызывают некроз опухолей in vivo, полагают, что именно эти участки обуславливают сходство биологического действия фактора некроза опухолей и лимфотоксина. В связи с этим при практической реализации изобретения получаемая композиция может вместо ФНО содержать лимфотоксин.
Термин "интерлейкин-4" или "ИЛ-4" в данном контексте используется для обозначения полипептида, который первоначально был описан как фактор, продуцируемый клетками лимфомы EZ-4- после их активации ТРА и индуцирующий пролиферацию мышиных клеток, активированных субмитогенными дозами антииммуноглобулина. ИЛ-4 называли также ростовым фактором B-клеток (РФВК), фактором, стимулирующим B-клетки (ФСВК), ростовым фактором II Т-клеток (РФТК-II), и ростовом фактором II тучных клеток (РФТК-II) (Howard et al., 1983, J. Exp. Med. , 155, 194,923; Yokota et al., 1988, Jmmunol. Rev., 102,137). ИЛ-4, образующийся в многочисленных популяциях Т-клеток/лимфоцитов, определяют с помощью иммунологических методов (различные варианты иммуноферментативного анализа) или посредством нортен-блот анализа (Paliard et al., 1988). В типичном случае активность ИЛ-4 может быть количественно охарактеризована с помощью биологической методики, которая основана на оценке усиления роста Т-клеток, как описано, например, в работе Spits с соавт. (J. Immunol., 1987, 139,1142-1147). Применяются, однако, и иные способы определения активности или идентификации ИЛ-4, Так, например, другой распространенный метод биотестирования состоит в активации периферических моноядерных клеток крови обработкой фитогемагглютинином на протяжении 72 ч при 37oC с последующим отмыванием и наслаиванием клеток (5000/0,1 мл/ячейка) на плату с 96 ячейками. Тестируемые пробы материала, содержащего ИЛ-4, серийно разводят и инкубируют с активированными клетками при 37oC в течение 72 ч. В инкубационную систему вводят меченый H3 тимидин и по включению последнего в клетки судят об активности ИЛ-4. Одна единица ИЛ-4 соответствует количеству цитокина, которое вызывает 50% пролиферации.
Способ очистки природного ИЛ-4 из содержащей сыворотку культуральной среды описан Lee с соавт. (J. Biol. Chem. 1988, 263, 10817-10823). После катионно-обменной хроматографии на S-сефарозе содержащие ИЛ-4 фракции элюировали в градиенте NaCl (0 - 0,5 M) в 50 мМ HEPES pH 7,0 и подвергали гель-фильтрации на сефадексе g - 100. Очистку фракций с ИЛ-4 продолжали с помощью обращенно-фазовой ВЭЖХ в градиенте 20-70% (объем/объем) ацетонитрила в 0,1% (объем/объем) трифторуксусной кислоты.
Эта методика позволяет обнаруживать до 60% присутствующего в пробах ИЛ-4 и обеспечивает его 3000-кратную очистку (специфическая активность 2,6 • 107 ед/мг). Очистка ИЛ-4 из бессывороточных супернатантов культуральной среды требует всего одной стадии хроматографирования на сефарозе-СМ и обеспечивает обнаружение 40% ИЛ-4 при его 100-кратной очистке (специфическая активность 3 • 107 ед/мг).
Для практической реализации изобретения пригоден любой полипептид, обладающий ИЛ-4 активностью и имеющий в своем составе аминокислотную последовательность, гомологичную последовательности зрелого ИЛ-4, а также фрагмент такого полипептида и производное самого полипептида или его фрагмента.
Об активности ИЛ-4 судят по его воздействию на клеточную пролиферацию, хотя для целей изобретения его используют для усиления цитотоксичности ФНО. Это, однако, не исключает возможность использования других способов оценки специфической активности ИЛ-4, в частности, методом, основанным на иных биологических эффектах в клетках - мишенях.
Физико-химические свойства ИЛ-4 изучены Yokota et al., Jmmunol. Rev., 1988, 102; 137 и Leе et al., (J. Biol, Chem., 1988, 263, 10817-10823). Нативный ИЛ-4 из биологических источников имеет молекулярную массу порядка 20000-22000 Да (по данным гель-фильтрации) и изоэлектрическую точку 6,7 при удалении сахарной группы и 10,53 при наличии последней. Такой ИЛ-4 чувствителен к гидролизу трипсином или химотрипсином. Аминокислотная последовательность ИЛ-4 состоит из 129 остатков с расчетной молекулярной массой 15000 Да. Расчетная молекулярная масса совпадает с ее экспериментальной оценкой методом ДДС-электрофореза в ДДС-ПААГ (1500, 18000 и 19000). Расхождение между теоретической и экспериментальной оценкой обусловлено наличием углеводного компонента.
Расчетная молекулярная масса рекомбинантного ИЛ-4 составляет приблизительно 22000 Да, по данным гель-фильтрации, и 15000 Да при оценке с помощью электрофореза в ДДС-ПААГ. Изоэлектрическая точка равна 10,53. Препарат гидролизуется под воздействием трипсина или химотрипсина.
Он сохраняет стабильность на протяжении 3 месяцев при 4oC, более одного месяца при комнатной температуре и в течение примерно 10 мин при 56oC. Кроме того, ИЛ-4 стабилен в относительно широком диапазоне значений pH (от 2 до 10). Он не обладает видовой специфичностью постольку, поскольку человеческий ИЛ-4 действует на мышиные клетки. Вместе с тем ИЛ-4 мышей действует только на клетки этого вида.
Степень гомологичности аминокислотной последовательности, которая служит признаком принадлежности того или иного соединения к ИЛ-4, согласно определению последнего может отличаться в зависимости от того, на каком участке молекулы она оценивается: внутри или вне области, отвечающей за цитотоксическую активность данного полипептида. Ключевые домены, определяющие цитотоксическую активность, должны иметь высокую степень гомологичности, тогда как области, не имеющие существенного значения для сохранения информации ИЛ-4 или его связывания с рецепторами, могут обладать слабой гомологичностью, что не мешает относить идентифицируемый полипептид к классу ИЛ-4. В то же время ключевые домены могут обладать выраженной цитотоксической активностью и сохранять гомологию согласно определению после замещения аминокислотных остатков, имеющих функционально сходные боковые цепи. Термин "функционально сходные" относится к доминантным признакам боковых цепей, таким как основность, нейтральность или кислотность, либо наличие и отсутствие стерической размерности.
Одним из наиболее существенных показателей при установлении идентичности того или иного полипептида ИЛ-4 служит способность антисыворотки, нейтрализующей биологическую активность ИЛ-4, в той же мере подавлять биологическую активность идентифицируемого соединения, вместе с тем доказать, что иммунологическая идентичность и цитотоксическая идентичность необязательно совпадают. Антитела, нейтрализующие ИЛ-4, не всегда связывают другой белок, потому что не обладают специфическим сродством к тем участкам молекулы, которые отвечают за проявление цитотоксической активности. Вместе с тем такие антитела могут связываться с биологически инертными участками и ингибировать биологическую активность посредством простой стерической блокады. Таким образом, блок с мутацией в инертной зоне теряет способность взаимодействия с антителами, что не мешает ему соответствовать определению ИЛ-4 по признакам высокой гомологичности и биологической активности.
Следует отметить, что такие признаки ИЛ-4, как молекулярная масса, изоэлектрическая точка и т.п., свойственные нативному ИЛ-4 или ИЛ-4 дикого типа, выделенному из периферических лимфоцитов или культур известных клеточных линий, характеризуют только нативные формы ИЛ-4. Однако понятие "ИЛ-4" согласно вышеприведенному определению включает и другие формы, которые могут не иметь всех признаков нативного ИЛ-4. Под указанное определение подпадает не только нативный ИЛ-4, но и родственные ему полипептиды. Так, например, производные ИЛ-4 (мутанты, полученные путем вставок и делеций, или слитые пептиды) по молекулярной массе отличаются от нативного ИЛ-4 (слитые белки, имеющие в своем составе полную молекулу ИЛ-4, и мутанты со вставками имеют большую молекулярную массу, чем нативный ИЛ-4, тогда как у мутантов с делециями она меньше). Точно так же ИЛ-4 может быть преобразован методами генной инженерии с целью уменьшения или элиминирования чувствительности к гидролитическому действию трипсина и других протеаз.
Композицию согласно изобретению можно вводить вместе с одним или несколькими полипептидами в неактивном состоянии, которые поддаются преобразованию путем ферментативного гидролиза из неактивной формы (как у зимогена) в полипептидный фрагмент, обладающий желаемой биологической активностью. В типичном случае неактивными предшественниками служат слитые пептиды, в составе которых один или несколько ИЛ-4, ФНО или γ -ИФН связаны по C-концам с человеческим белком или его фрагментом, либо прямо или косвенно с любым другим полипептидом, пригодным для реализации данного изобретения. Состав аминокислотных остатков в месте такой связи и на примыкающих участках подбирают таким образом, чтобы иметь возможность освободить ИЛ-4, ФНО или γ -ИФН протеолитическим гидролизом in vivo либо на одном из этапов производственного регламента in vitro. Получаемая таким образом композиция обладает цитотоксической активностью согласно определению.
Хотя термин "ИЛ-4" обычно обозначают человеческий ИЛ-4, он в равной степени пригоден для ИЛ-4 из других источников: мышей, свиней, лошадей и крупного рогатого скота, если соответствующие полипептиды имеют признаки гомологичности и цитотоксической активности согласно определению.
Для целей изобретения пригодны также многомерные формы ИЛ-4.
Производные ИЛ-4 тоже подпадают под определение ИЛ-4. К числу таких производных относятся мутанты с измененной аминокислотной последовательностью, гликозилированные формы, ковалентно связанные или агрегированные конъюганты с другими химическими компонентами. Ковалентные конъюганты получают, связывая известными способами требуемые соединения с определенными группировками боковых цепей аминокислотных остатков ИЛ-4 или с его N и C-концевыми участками.
Как и в случае ФНО, к мутантным производным ИЛ-4 принадлежат варианты ИЛ-4 или его фрагментов с направленными, т.е. сайт-специфичными мутациями, которые делают эти соединения пригодными для целей изобретения. Мутантами ИЛ-4 считают полипептиды, которые по признаку гомологичности подпадают под вышеприведенное определение ИЛ-4, но имеют измененную делецией, заменой или вставкой аминокислотную последовательность, что отличает ее от последовательности ИЛ-4. Способы направленных мутаций посредством замены нужных оснований в соответствующих ДНК хорошо известны (например, мутагенез с использованием праймера М 13).
Говоря о выраженной гомогенности ИЛ-4 или фактора некроза опухолей, имеют в виду такие ИЛ-4 и фактор некроза, в которых практически отсутствуют другие белки из того же источника, из которого были выделены нативные соединения. Это означает, что гомогенные ИЛ-4 и/или фактор некроза опухолей практически не содержит белков плазмы крови, таких как альбумин, фибриноген, сериновые протеазы, α-глобулин, цитотоксичные полипептиды, иные, чем фактор некроза опухолей (лимфотоксин или интерфероны), или другие белки клеток или организма, послужившие синтетическим источником полипептида (в том числе цельных клеток или твердых клеточных остатков). В то же время достаточно чистые препараты могут иметь в своем составе нижеописанные стабилизаторы и недействующие компоненты; заданные количества белков клеток или организма, служивших синтетическим источником; белков из иных источников нежели клетки или организм, из которых получены ИЛ-4 или фактор некроза опухолей; синтетические полипептиды, например, поли-Z-лизин. Рекомбинантные ИЛ-4 и фактор некроза опухолей, предназначенные для экспрессии в аллогенных (т.е. бактериальных) клетках-хозяевах, должны быть, разумеется, совершенно свободны от белков источника гена.
Для целей изобретения ИЛ-4 и фактор некроза опухолей предпочтительно синтезируют в культурах рекомбинантных организмов, используя общепринятые методы.
Композицию согласно изобретению для последующего введения получают посредством смешивания ИЛ-4 и фактора некроза опухолей требуемой степени чистоты с физиологически приемлемыми носителями, т.е. с носителями, которые нетоксичны для реципиентов в используемых дозах или концентрациях. Обычно при этом используют буферные растворы, антиоксиданты (например, аскорбиновую кислоту), низкомолекулярные полипептиды (менее 10 аминокислотных остатков), белки, аминокислоты, углеводы (в том числе глюкозу или декстрины), хелирующие агенты типа ЭДТА и другие стабилизаторы или нейтральные компоненты. Состав носителя должен обеспечивать стабилизацию композиции. Композиция, предназначенная для терапевтического использования, подлежит стерилизации, которую на практике осуществляют посредством фильтрации через стерильные мембраны. Как правило, композицию хранят в форме лиофилизированного препарата.
Поскольку интерфероны являются синергистами ИЛ-4 и фактора некроза опухолей, α, β и γ -интерфероны предпочтительно используют в сочетании с композициями фактор некроза опухолей ИЛ-4 или с композициями, содержащими ИЛ-4, ФНО и лимфотоксин. ФНО должен иметь специфическую активность 100 ед/нг, а ИЛ-4 и γ -ИФН активность порядка 10 ед/нг. Типичные композиции содержат ИЛ-4 и/или ИФН и фактор некроза опухолей и/или лимфотоксин при соотношении единиц активности в диапазоне от 0,1:1 до 200:1, обычно 10:1. Фактор некроза опухолей в таких композициях может быть частично или полностью заменен лимфотоксином. Степень замены, разумеется, зависит от задач и условий терапии.
Композиции ИЛ-4/ФНО предназначены для введения животным с опухолями. Введение осуществляют любым из общепринятых способов, т.е. внутривенно, внутрибрюшинно, внутримышечно, посредством инфузии в пораженные участки ткани или инъекции стерильных растворов, а также с использованием систем дозированного введения (см. ниже). ИЛ-4/ФНО можно вводить в пораженную ткань, т.е. путем непосредственно инъекции в солидные опухоли. При наличии диссеминированных опухолей, как, например, при лейкемии, предпочтительно внутривенное введение композиции или введение в лимфатическую систему. Животным с опухолями органов брюшной полости (например, в случае рака яичника) желательно вводить композицию посредством внутрибрюшинной инфузии, используя для этой цели оборудование для перитонеального диализа и совместимые растворители. Обычно композицию ИЛ-4/ФНО вводят путем непрерывной инфузии, хотя допускается и разовое введение.
Предпочтительно вводить ИЛ-4/ФНО с помощью имплантированных дозаторов. Примерами материалов, пригодных для таких систем введения белков с молекулярной массой димеров и тримеров фактора некроза опухолей, могут служить сополимеры L-глютаминовой кислоты и гамма-этил-L-глютамина (U. Sidman et al., Biopolymers, 1983, 22/1, 547-556), поли-(2-окси-этил-метакрилат) (R. Sanger et al. , Biomed. Matter Res., 1981, 15, 167-277 и R. Sanger, Chem. Tech., 1982, 12, 98-105) или этилен-винилацетат (R. Sanger et al., там же). Такие материалы имплантируют на место хирургически удаленных опухолей. По альтернативной методике композицию ИЛ-4/ФНО вводят в составе капсул из полупроницаемых мембран или липосом непосредственно в опухоль. Такой способ введения особенно ценен при неоперабельных опухолях, например, опухолях мозга.
Способ лечения опухолей согласно изобретению посредством введения их носителям ФНО и ИЛ-4 предполагает как одновременное, так и последовательное введение этих полипептидов. Известно, что эффект одновременного введения можно воспроизвести поочередным введением сначала одного, а потом другого полипептида. Относительные сроки введения каждого из препаратов известны из предшествующего опыта и зависят исключительно от скорости разложения in vivo первого полипептида.
Доза вводимой композиции определяется, в частности, способом ее введения, природой опухоли и состоянием индивидуума. При введении ИЛ-4/ФНО непосредственно в пораженную ткань доза композиции в пересчете на единицу веса меньше, чем при внутривенной инфузии. В то же время некоторые формы солидных опухолей более устойчивы к действию фактора некроза по сравнению с другими типами (например, при лейкемии). В соответствии с этим приходится подбирать дозировки и изменять пути введения композиции, чтобы обеспечить максимальное проявление ее цитотоксической активности по отношению к данному виду опухоли. Для этой цели используют исследование биопсийного материала от носителей опухолей или такие диагностические методы, как тестирование с применением карциноэмбрионального антигена для выявления повышенного уровня токсичности в рекомбинантном материале. В большинстве случае доза фактора некроза опухолей для мышей при внутривенной инъекции составляет приблизительно 120 мкг/кг веса тела/день, обеспечивая достаточно высокую терапевтическую эффективность in vivo в отсутствии токсичности. Доза ИЛ-4 до 120 мкг/кг веса тела/день при внутривенном введении мышам также не вызывает токсических эффектов и дает хороший терапевтический эффект in vivo.
Фактор некроза опухолей не обладает видовой специфичностью, с точки зрения цитотоксической активности, поэтому для лечения опухолей человека можно вместо человеческого ФНО использовать ФНО свиней или крупного рогатого скота. Тем не менее при введении фактора некроза опухолей тому или иному виду животных же6лательно использовать в качестве его источника тот же самый вид во избежание образования аутоантител.
Все ссылки на литературные источники в тексте описания следует рассматривать лишь как источники информации.
Более исчерпывающее представление о сути описанного в предшествующем разделе изобретения можно получить из нижеследующих примеров. Эти примеры служат для иллюстрации отдельных аспектов изобретения и не являются ограничивающими, за исключением специально оговоренных случаев.
Пример 1. Процедуры культивирования клеток.
Все линии культивировали в условиях, исключающих слияние клеток. Последние наслаивали на среду ДМЭМ, обогащенную глютамином (2 мМ), пенициллином (100 ед/мл), стрептомицином (100 мкг/мл) и плодной телячьей сывороткой (10%) из расчета 0,3-1,3•106 клеток/25 мм чашка Петри. Инкубацию проводили в термостате с контролируемой влажностью в атмосфере 5% CO2.
Пример 2. Оценка цитотоксичности.
Для оценки цитотоксичности 5•103 клеток в 0,1 мл среды помещали в ячейки платы на 96 ячеек. После инкубации при 37oC в течение ночи при повышенной влажности в атмосфере 5% CO2 среду удаляли, а в ячейки добавляли серийные разделения тестируемых проб в 0,1 мл среды. Плату оставляли при 37oC на 72 ч, затем среду снова удаляли и оценивали жизнеспособность клеток, используя окрашивание по Sugarman et al., Science, 1985, 230, 943). Жизнеспособность оценивали в процентах по результатам определения оптической плотности в присутствии тестируемого материала и без него. Разность от деления первой полученной величины на вторую умножали на 100.
Пример 3. Радиорецепторный анализ.
Для анализа рецепторного связывания использовали модификацию метода, предложенную Aggarwal et al. , Nature, 1985, 318, 665). 0,25 • 106 клеток инкубировали в полипропиленовых пробирках (12 • 75 мм) в 0,2 мл свежеприготовленной и охлажденной на льду среды (ДМЭМ с 10%-ной плодной телячьей сывороткой), содержащей I125 ФНО (0,2 • 106 имп/мин), в присутствии (неспецифическое связывание) или в отсутствии (общее связывание) 100-кратного избыточного количества немеченного ФНО. После инкубации при 4oC на протяжении 60 мин среду удаляли, клетки трижды отмывали посредством центрифугирования, после чего определяли связанную с ним радиоактивность. Все измерения проводили трижды.
Пример 4. Усиление антипролиферативной активности ФНО интеролейкином-4.
Стимулирующее действие интерлейкина-4 на антипролиферативную активность ФНО регистрировали в различных типах опухолевых клеток. Действие ФНО в различных концентрациях в присутствии и без ИЛ-4 при постоянной концентрации последнего 100 нг/мл на клеточную линию МДА-МВ-330 из опухоли женской грудной железы показано на фиг. 1. 5 • 103 клеток инкубировали в 0,1 мл среды, содержавшей указанные количества цитокинов, в течение 72 ч. Жизнеспособность клеток оценивали, как описано в примере 1. Все измерения повторяли трижды.
В отсутствии ИЛ-4 ФНО даже при концентрации 10000 ед/мл оказывал очень слабое действие на излучение клетки. Однако при добавлении ИЛ-4 в инкубационную систему рост клеток подавлялся пропорционально концентрациям ФНО. Как следует из фиг. 1, при концентрации ФНО 10000 ед/мл ингибирование роста не превышало 10%. После добавления 100 нг/мл ИЛ-4 ингибирование роста опухолевых клеток достигло 60%. Усиление цитотоксической активности ФНО под воздействием ИЛ-4 наблюдали также в клеточных линиях из опухоли женских наружных половых органов (А-431) и гистоцитарной лимфы (U-937). На фиг. 2 показано действие различных концентраций ФНО при постоянной концентрации ИЛ-4 (100 нг/мл) на относительную жизнеспособность в процентах клеток эпителиальной карциномы человека (линии А-431, фиг. 2А) и клеток гистоцитарной лимфомы человека (U-936, фиг. 2А). 0,5 • 103 клеток каждого типа инкубировали в 0,1 мл среды, содержащей указанные количества цитокинов, после чего оценивали жизнеспособность клеток, как указано в примере 1. Продолжительность инкубации во всех случаях составляла 72 ч.
Как видно из фиг. 2, ИЛ-4 значительно изменял модулирующее действие ФНО на клетки линии А-431 при концентрации ФНО всего 0,01 ед/мл (фиг. 2А).
Еще более выраженным действием на активность ФНО интерлейкин-4 обладал в клетках линии U-937 при концентрации ФНО 1000 ед/мл (фиг. 2B).
Было изучено также влияние ИЛ-4 на действие ФНО при переменных концентрациях первого из цитокинов. При инкубации 5 • 103 клеток в 0,1 мл среды при указанных концентрациях цитокинов на протяжении 72 ч с последующей оценкой жизнеспособности клеток по описанной в примере 1 методике все измерения проводили в трехкратной повторности. На фиг. 3 показано действие различных концентраций ИЛ-4 на цитотоксическую активность ФНО в концентрации 1000 ед/мл в отношении клеточной линии А-431 (фиг. 3А) и линии МДА-МВ-330 (фиг. 3B). Показано также действие одного ИЛ-4 в концентрации 1 мкг/мл. В присутствии ФНО и ИЛ-4 наблюдали пропорциональное концентрациям последнего ингибирование клеточного роста (фиг. 3А). Один ИЛ-4 практически не влиял на него даже в концентрации 1 мкг/мл. Аналогичное, зависимое от дозы ИЛ-4 ингибирование наблюдали в культуре клеток опухоли женской грудной железы (фиг. 3B).
Пример 5. Сопоставление стимулирующего действия ИЛ-4 и γ -ИФН на цитотоксическую активность ФНО.
Ранее было показано, что гамма-интерферон также усиливает цитотоксическую активность ФНО в самых разнообразных клеточных линиях из опухолей (патент США N 4650674, Lee et al. 1984, 133, 1003: J. Immunol. Sugarman et al., 1985, Science, 230, 943; Aggrawal et al., 1985, Nature, 318, 665; Vilcek et al., 1986, Interaction Between Tumor Necrosis Factor and Interferons (Взаимодействие между фактором некроза опухолей и интерферонами), In: The Biology of Interferon System, Edited by H. Schellekenes and W.E. Stewart, Elsevier Science Publishers, Amsterdam, page 249; Fransen et. al., 1986 Eur. J. Cancer Clin. Oncol., 22, 419-426; Campbell et al., (1988, J, Immunol., 141, 2325 - 2329; Schiller et al., 1987. Cancer Res, 47, 2809-2813; Tsujimoto et al., 1986. J.Immunol., 137, 2272-2276).
Фиг. 4 иллюстрирует сравнительное действие ИЛ-4 и γ -ИНФ на цитотоксическую активность ФНО в отношении клеточной линии МДА-МВ-330. 5•103 клеток этой линии инкубировали в 0,1 мл среды, содержащей различные концентрации ФНО при постоянных концентрациях либо ИЛ-4, либо ИФН (по 100 нг/мл каждого) в течение 72 ч. После окончания инкубации оценивали жизнеспособность клеток, как описано в примере 1. Все измерения производили в трехкратной последовательности.
Для того чтобы выяснить, действуют ли ИЛ-4 и γ -ИФН на цитотоксическую активность ФНО посредством сходных или различных механизмов, изучали их влияния на антипролиферативную эффективность ФНО. Приведенные на фиг.4 результаты показывают, что в клетках опухоли грудной железы женщин при равных концентрациях ИЛ-4 и γ -ИФН (100 нг/мл) они в равной степени усиливают ингибирующее действие ФНО на клеточный рост. Последний уменьшился в обоих случаях на 60-70%. Аналогичные результаты были получены в экспериментах на клеточных линиях А-431 и U-937 (табл.1).
5•103 клеток инкубировали в 0,1 мл среды, содержащей либо ФНО (200 нг/мл), либо ИЛ-4 (100 нг/мл), либо γ -ИФН (100 нг/мл), либо указанные выше сочетания цитокинов. При экспериментах с клетками А-431 концентрации всех цитокинов оставались такими же, за исключением концентрации γ -ИФН, которая составляла 1 нг/мл. После 72-ч инкубации жизнеспособность клеток оценивали посредством окрашивания, как описано в примере 1.
ИЛ-4 несколько усиливает антипролиферативное действие γ-ИФН, хотя и в меньшей степени, чем действие ФНО (табл.1). Кроме того, при использовании комбинации всех 3-х цитокинов был получен максимальный эффект. Так, например, при тестировании на клеточной линии А-431 ФНО ингибировал рост клеток только на 30%. Ингибирование возросло до 55% в присутствии ФНО вместе с ИЛ-4 и до 80% после введения в инкубационную систему всех трех цитокинов (т.е. γ -ИФН, ИЛ-4 и ФНО; табл.1).
Полученные результаты показывают, что механизм стимулирующего действия ИЛ-4 на цитотоксическую активность ФНО отличается от механизма действия γ-ИФН. Для того чтобы подтвердить это различие, сопоставляли действие обоих цитокинов на индукцию рецепторов ФНО. Известно, что γ-ИФН обладает способностью индуцировать эти рецепторы. Приведенные в табл.2 результаты совершенно четко показывают, что преинкубация клеток U-937 с ИЛ-4 на протяжении ночи не сопровождалась усилением специфического связывания ФНО. Иными словами, биосинтез новых рецепторов под воздействием ИЛ-4 отсутствовал. Это подтверждает вывод о том, что механизмы потенциирующего действия ИЛ-4 и γ-ИФН на цитотоксическую активность ФНО отличаются.
Клетки U-937 инкубировали с ИЛ-4 в концентрации 1 мкг/мл при 37oC в течение 24 ч, после чего отмывали от среды. Затем 250000 полученных клеток переносили в инкубационную систему, содержащую меченый I125 ФНО (0,2 •106 имп/мин) в присутствии 100 нМ немеченого ФНО или без него. Инкубацию продолжали при 4oC в 0,2 мл среды на протяжении 1 ч. После этого клетки снова отмывали и с помощью методики, описанной в примере 2, определяли количество связанной ими радиоактивности. Все измерения проводили в трехкратной последовательности.
ИЛ-4 сам по себе также вызывал пролиферацию некоторых типов клеток. Как следует из табл. 3, он усиливал рост клеточной линии ZR-75-1 из опухоли грудных желез. В меньшей степени усиление клеточного роста имело место и в других типах клеток. В то же время ИЛ-4 несколько усиливал рост клеток А-431 (табл.3). Рост-стимулирующее действие ИЛ-4 оказалось зависимым от дозы.
5•103 клеток каждого типа инкубировали с интерлейкином-4 в концентрации 1 мкг/мл на протяжении 72 ч, после чего определяли относительную жизнеспособность клеток окрашиванием, как описано в примере 1. Все измерения производили в трехкратной повторности.
Пример 6. Усиление антипролиферативного действия лимфотоксина интерлейкина-4.
Интерлейкин-4 усиливал антипролиферативное действие лимфотоксина (Лт) на опухолевые клетки. Антипролиферативное действие Лт в сочетании с ИЛ-4 при их концентрациях соответственно 100 ед/мл и 1 мкг/мл сопоставляли с аналогичным действием ФНО (1000 ед/мл) в присутствии или в отсутствии ИЛ-4 (1 мкг/мл). С этой целью клетки линии МДА-МВ-330 из опухоли грудной железы женщин в количестве 5 • 103 инкубировали в 0,1 мл среды, содержащей указанные количества цитокинов, на протяжении 72 ч. О жизнеспособности клеток судили с помощью описанной в примере 1 методики. Все измерения производили в трехкратной последовательности. Полученные результаты испытаний представлены на фиг. 5.
Сам по себе ФНО практически не влиял на состояние клеток, его ингибирующее действие не превышало 5%. В то же время Лт вызывал выраженный пролиферативный эффект. Жизнеспособность клеток в присутствии одного ИЛ-4 составляла 95-98%. При добавлении ИЛ-4 в инкубационную систему, содержащую ФНО или Лт, наблюдали значительное ингибирование клеточного роста. Как видно из фиг. 5, рост клеток задерживался почти на 60% в присутствии ИЛ-4 вместе с ФНО и на 40% в присутствии ИЛ-4 в комбинации с Лт.
Механизм, посредством которого γ -ИФН изменяет реакцию клеток на ФНО, остается невыясненным. Известно, что восходящая регуляция рецепторов ФНО сопровождается восходящей регуляцией цитотоксического действия ФНО гамма-интерфероном (Aggarwal et al. , Nature, 1985, 318, 665; Vilcek et al., The Biology of Interferon System, 1986, с.249). Однако исследования показали, что ИЛ-4 не регулирует рецепторы ФНО по восходящему пути в пораженных им клетках. И хотя эти наблюдения свидетельствуют о разных механизмах стимулирующего действия ИЛ-4 и γ -ИФН на активность ФНО, имеются сообщения, показывающие, что восходящая регуляция рецепторов ФНО γ -интерфероном недостаточна для проявления их синергетического действия (Tsujimoto et al., J. Immunol. , 1986, 137, 2272-2276; Aggarwal et al., J. Biol. Chem., 1987, 262, 10000-10007). По нашим наблюдениям γ-ИФН одновременно усиливает действие ИЛ-4 и ФНО. Возможно потому, что активность обоих этих агентов регулируется посредством разных механизмов. Известно также, что ИЛ-4 усиливает цитотоксическую активность ФНО (Ruggiero and Baglioni, J. Immunol., 1987, 138, 0661-0663; Holtmann and Wallach. J. Immunol., 1987, 139, 1161-1167). Итак, механизм, через который опосредуется стимулирующее действие ИЛ-4 на активность ФНО, неизвестен. В отличие от γ-ИФН ИЛ-1 оказывает нисходящее регуляторное действие на рецепторы ФНО. Помимо цитокинов известны и другие соединения, которые усиливают цитотоксическую активность ФНО, например, ингибиторы белкового синтеза. Аналогичный эффект дает повышение температуры Watanabe et al. , Cancer Res., 1988, 48, 650-653; Niits et al., Cancer Res., 1988, 48, 654-657). Исследования с ингибиторами белкового синтеза показали, что ФНО индуцирует биосинтез некоторых белков, которые защищают клетку от действия этого фактора (Himero et al., Cancer Res., 1990, 50, 4941-4945; Watanabe et al., Immunoparmacol, Immunotoxicol., 1988, 10, 479-499). Известно также, что антипролиферативное действие ФНО подавляется эпидермальным ростовым фактором и трансформирующим ростовым фактором (Sugarman et al., Cancer Res., 1987, 47, и 780-786) в клетках разного типа. Показано, в частности, что эти факторы продуцируются различными опухолями. Нe исключено поэтому, что функция ИЛ-4 опосредуется через ингибирование активности этих факторов. С другой стороны, функция ИЛ-4 может быть связана с нисходящей регуляцией экспрессии онкогенов c-foc и c-мус, которые, как известно, принимают участие в пролиферации клеток.
Имеются сообщения о клонировании кДНК рецепторов ФНО, γ -ИФН и ИЛ-4 (Idzerda et al., J. Exp. Med., 1990, 171, 861-873; Shall et al., Cell 1990, 61, 361-370; Smith et al. , Science, 1990, 248, 1019-1023, Anget et al., Cell, 1988, 55, 273-280). Однако структура этих рецепторов такова, что сами по себе не могут обеспечить трансдукцию сигнала. Описана нисходящая модуляция ИЛ-4 продукции ФНО макрофагами (Essneret et al., J. Immunol., 1989, 142, 3857; Hamilton, Proc. Natl. Acad. Sci. USA, 1989, 86, 3803), однако известна также способность ИЛ-4 усиливать опосредуемую через макрофаги цитотоксичность (Somers and Erikson, Cell Immunol., 1989 , 122, 178-187). Последний эффект, как было показано, обусловлен преимущественно ФНО (Feinman et al., 1987, J. Immunol., 138, 635-640). Непонятно поэтому, почему усиление цитотоксичности макрофагов под воздействием ИЛ-4 сопровождается ингибированием продукции ФНО. Возможно, что повышение цитотоксичности макрофагов для опухолевых клеток под воздействием ИЛ-4 опосредуется через синергетическое взаимодействие между ФНО и ИЛ-4, описанное выше. Установлено, что опухолевые клетки, продуцирующие ИЛ-4, обладают противоопухолевой активностью in vivo по отношению к опухолям другого типа (Tepper et al., Cell, 1989, 57, 503-512). Поскольку сам по себе ИЛ-4 не оказывает непосредственного антипролиферативного действия на опухолевые клетки в условиях культуры, механизм его функционирования в качестве противоопухолевого средства in vivo остается неясным. Возможно, что в этой ситуации его противоопухолевая активность объясняется активацией макрофагов и усилением противоопухолевой активности фактора некроза опухолей.
Из приведенного описания следует, что изобретение полностью отвечает указанным целям и обладает всеми заявленными преимуществами. Описанные компоненты, методы и процедуры обеспечивают предпочтительные пути реализации изобретения, они иллюстрируют различные его аспекты, но не ограничивают область применения изобретения. Характер изобретения допускает различные модификации и иные способы применения, которые отражены в прилагаемой формуле.
Описан состав материала, содержащего фактор некроза опухолей (ФНО) и интерлейкин-4 (ИЛ-4). Эта композиция ингибировала рост раковых клеток молочной железы женщин, клеток карциномы женских наружных половых органов и клеток гистиоцитарной лимфомы человека пропорционально ее дозам. При раздельном воздействии ФНО или ИЛ-4 на раковые клетки женской грудной железы характер их роста существенно не изменялся. В то же время при сочетанном воздействии двух этих цитокинов они пропорционально дозам угнетали рост указанных клеток. Кроме того, гамма-интерферон (γ- ИФН) еще более усиливал антипролиферативную активность композиции, содержащей ФНО и ИЛ-4. Аналогичной активностью обладала композиция, содержащая ИЛ-4 и лимфотоксин. 3 с. и 3 з.п. ф-лы, 3 табл., 5 ил.
Фактор некроза опухоли - 1
Интерлейкин-4 - 0,1 - 200
2. Композиция по п.1, отличающаяся тем, что используют человеческий фактор некроза опухолей.
Фактор некроза опухоли - 1
Интерферон - 0,1 - 200
Интерлейкин-4 - 0,1 - 200
5. Бесклеточная композиция с синергидным эффектом для воздействия на опухолевые клетки, содержащая два активных компонента, один из которых - интерлейкин-4, отличающаяся тем, что вторым компонентом является лимфотоксин при следующем соотношении компонентов в единицах активности:
Интерлейкин-4 - 0,1 - 200
Лимфотоксин - 1
6. Композиция по п.5, отличающаяся тем, что используют человеческий интерлейкин-4.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| US, патент, 4863727, А 61 К 37/02, 1989 | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Forni G | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| "International Journal of Cancer" | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |
| Способ крашения тканей | 1922 |
|
SU62A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| SU , патент, 1405691, А3, А 61 К 38/21, 1988 | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| SU, авторское свидетельство, 1454410, AI, А 61 К 38/21, 1989. | |||

