Предпосылки настоящего изобретения
Область техники
Настоящее изобретение относится к микробиологической промышленности, в частности к способу продукции 4-гидрокси-L-изолейцина или его соли.
Описание предшествующего уровня техники
4-Гидрокси-L-изолейцин является аминокислотой, которую можно выделить и очистить из семян пажитника (Trigonella foenum-graecum L. leguminosae). 4-Гидрокси-L-изолейцин проявляет инсулинотропную активность, представляющую большой интерес, поскольку его стимулирующий эффект четко зависит от концентрации глюкозы в среде, как показано, и на выделенной перфузионной поджелудочной железе крыс, и на островках поджелудочной железы человека (Sauvaire, Y. et al. Diabetes, 47: 206-210 (1998)). Такая глюкозная зависимость не подтверждена на сульфонилмочевине (Drucker, D.J., Diabetes, 47: 159-169 (1998)), единственном инсулинотропном препарате, используемом в настоящее время для лечения диабета II типа [или диабета, независимого от инсулина - non-insulin-dependent diabetes (NIDD) mellitus (NIDDM)], и в результате гипогликемия является распространенным нежелательным побочным эффектом при лечении сульфонилмочевиной (Jackson, J. and Bessler, R. Drugs, 22: 211-245; 295-320 (1981); Jennings, A. et al. Diabetes Care, 12: 203-208 (1989)). Повышение глюкозной толерантности также известно (Am. J. Physiol. Endocrinol., Vol.287, E463-E471, 2004). Сообщено об этом повышении активности глюкометаболизма и его потенциальном применении в фармацевтике и здоровом питании (выложенная патентная заявка Японии No. Hei 6-157302, заявка США US 2007-000463A1).
4-Гидрокси-L-изолейцин, обнаруженный только в растениях, благодаря его особому инсулинотропному действию может рассматриваться в качестве нового стимулятора секреции, представляющего потенциальный интерес для лечения диабета II типа, заболевания, характеризующегося недостаточной секрецией инсулина, связанной с устойчивостью к инсулину различной степени (Broca, С. et al. Am. J. Physiol. 277 (Endocrinol. Metab. 40): E617-E623 (1999)).
Метод окисления железа, аскорбиновой кислоты, 2-оксиглутаровой кислоты и изолейцина (зависимого от кислорода) с использованием активности диоксигеназы в экстракте пажитника описан как метод для производства 4-гидрокси-L-изолейцина (Phytochemistry, Vol.44, No. 4, pp.563-566, 1997). Однако этот метод является неудовлетворительным в качестве метода производства 4-гидрокси-L-изолейцина, поскольку фермент ингибируется субстратом при концетрации 20 мМ и выше, фермент не идентифицирован, фермент получают из растительного экстракта и его трудно получить в большом количестве, фермент нестабилен.
Раскрыт эффективный восьмистадийный синтез оптически чистого (2S,3R,4S)-4-гидроксиизолейцина с выходом 39%. Ключевые стадии этого синтеза включают биотрансформацию этил-2-метилацетоацетата в этил- (2S,3S)-2-метил-3-гидрокси-бутаноата с использованием Geotrichum candidum и асимметричный синтез Strecker (Wang, Q. et al., Eur. J. Org. Chem., 834-839 (2002)).
Также раскрыт короткий шестистадийный хемоферментативный синтез (2S,3R,4S)-4-гидроксиизолейцина с общим стереохимическим контролем, последней стадией которого является ферментативное разложение производного N-фенилацетиллактона путем гидролиза с использованием коммерческой пенициллинацилазы G, иммобилизованной на Eupergit C (E-PAC) (Rolland-Fulcrand, V. et al., J. Org. Chem., 873-877 (2004)).
Однако в настоящее время отсутствуют сообщения о продукции (2S,3R,4S)-4-гидроксиизолейцина путем прямого ферментативного гидроксилирования L-изолейцина с использованием бактерии с L-изолейциндиоксигеназной активностью, в частности с использованием бактерии с изначально имеющимися активностями L-изолейциндиоксигеназы и 4-гидрокси-L-изолейциндегидрогеназы, отличающейся тем, что активность 4-гидрокси-L-изолейциндегидрогеназы в указанной бактерии уменьшена или разрушена.
Описание изобретения
Авторы настоящего изобретения предложили новый метод для продукции (2S,3R,4S)-4-гидроксиизолейцина путем прямого ферментативного гидроксилирования L-изолейцина с использованием L-изолейциндиоксигеназы или бактерии с L-изолейциндиоксигеназной активностью. При дальнейшем исследовании обнаружен факт способности некоторых бактерий превращать образованный (2S,3R,4S)-4-гидроксиизолейцин в АМКР.
В результате расширенного исследования, проведенного с учетом вышеупомянутых проблем, авторы настоящего изобретения идентифицировали бактерию с высоким уровнем активности 4-гидрокси-L-изолейциндегидрогеназы, клонировали ген, кодирующий 4-гидрокси-L-изолейциндегидрогеназу, и обнаружили, что для синтеза требуемого (2S,3R,4S)-4-гидрокси-L-изолейцина с использованием бактерии, изначально имеющей L-изолейциндиоксигеназу и 4-гидрокси-L-изолейциндегидрогеназу, необходимо снизить или разрушить в бактерии активность 4-гидрокси-L-изолейциндегидрогеназы для предотвращения превращения требуемого 4-гидрокси-L-изолейцина в АМКР, что привело к выполнению настоящего изобретения.
Целью настоящего изобретения является увеличение продукции (2S,3R,4S)-4-гидрокси-L-изолейцина (используемое обозначение включает в себя как свободную форму, так и его соли, может использоваться аббревиатура "(2S,3R,4S)-4HIL"; в дальнейшем обозначает то же самое), предоставление способа производства (2S,3R,4S)-4-гидрокси-L-изолейцина или его соли путем прямого ферментативного гидроксилирования L-изолейцина с использованием бактерии, изначально имеющей активности L-изолейциндиоксигеназы и 4-гидрокси-L-изолейциндегидрогеназы, отличающейся тем, что активность 4-гидрокси-L-изолейциндегидрогеназы уменьшена или разрушена.
А именно, целью настоящего изобретения является предоставление способа продукции (2S,3R,4S)-4-гидрокси-L-изолейцина или его соли, включающего стадии:
реакции L-изолейцина в водном растворе в присутствии бактерии, изначально имеющей активности L-изолейциндиоксигеназы и 4-гидрокси-L-изолейциндегидрогеназы, и
выделения синтезированного (2S,3R,4S)-4-гидрокси-L-изолейцина, отличающегося тем, что указанная бактерия модифицирована таким образом, что активность 4-гидрокси-L-изолейциндегидрогеназы в указанной бактерии уменьшена или разрушена.
Также целью настоящего изобретения является предоставление вышеописанного способа, отличающегося тем, что гены, кодирующие L-изолейциндиоксигеназу и 4-гидрокси-L-изолейциндегидрогеназу, объединены в составе одного оперона.
Также целью настоящего изобретения является предоставление вышеописанного способа, отличающегося тем, что указанная бактерия модифицирована таким образом, что активность L-изолейциндиоксигеназы в указанной бактерии увеличена.
Также целью настоящего изобретения является предоставление вышеописанного способа, отличающегося тем, что активность 4-гидрокси-L-изолейциндегидрогеназы в указанной бактерии уменьшена или разрушена путем ослабления или подавления экспрессии гена, кодирующего 4-гидрокси-L-изолейциндегидрогеназу.
Также целью настоящего изобретения является предоставление вышеописанного способа, отличающегося тем, что указанная бактерия принадлежит к роду Bacillus, Enterobacter или Pseudomonas.
Также целью настоящего изобретения является предоставление вышеописанного способа, отличающегося тем, что указанная бактерия принадлежит к Bacillus thuringiensis, Bacillus weihenstephanensis, Bacillus cereus, Enterobacter agglomerance или Pseudomonas syringae.
Также целью настоящего изобретения является предоставление вышеописанного способа, отличающегося тем, что указанная бактерия - бактериальная культура, клетки, обработанные клетки или клеточный лизат.
Более детально настоящее изобретение описано ниже.
Подробное описание наилучшего способа осуществления изобретения
В настоящем изобретении термин "(2S,3R,4S)-4-гидрокси-L-изолейцин" или "(2S,3R,4S)-4HIL" относится к отдельному химическому соединению или к смеси, содержащей (2S,3R,4S)-4-гидроксиизолейцин. Также термин "2-амино-3-метил-4-кетопентаноат" или "АМКР" относится к отдельному химическому соединению или к смеси, содержащей различные оптические изомеры 2-амино-3-метил-4-кетопентаноата.
Термин "бактерия", как он используется в настоящем описании, включает образующую фермент бактерию, мутант и генетический рекомбинант такой бактерии, в которой существует или увеличена целевая ферментативная активность, и т.п.
"Бактерия, изначально содержащая активности L-изолейциндиоксигеназы и 4-гидрокси-L-изолейциндегидрогеназы" - согласно настоящему изобретению бактерия, на хромосоме которой содержатся гены, кодирующие ферменты с активностями L-изолейциндиоксигеназы и 4-гидрокси-L-изолейциндегидрогеназы. А именно, это бактерия, имеющая гены, кодирущие L-изолейциндиоксигеназу и 4-гидрокси-L-изолейциндегидрогеназу, организованные в оперон, предпочтительно трансляционно-сопряженные. Предпочтительны бактерии, принадлежащие к роду Bacillus, роду Pseudomonas, роду Enterobacter.
Выражение "бактерия, принадлежащая к роду Bacillus" означает, что бактерия классифицирована как принадлежащая к роду Bacillus в соответствии с классификацией, известной специалисту в области микробиологии. Примеры использованной в настоящем изобретении бактерии, принадлежащей к роду Bacillus, включают, но не ограничиваются ими, Bacillus thuringiensis, Bacillus weihenstephanensis, Bacillus cereus. Частные примеры штаммов Bacillus thuringiensis - штамм Bacillus thuringiensis 2-e-2, названный Bacillus thuringiensis AJ110584 и в соответствии с Будапештским Договором депонированный в International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology (Central 6, 1-1-1 Higashi, Tsukuba, Ibaraki 305-8566, Japan) 27 сентября 2006 г. с инвентарным номером PERM BP-10688, штамм Bacillus thuringiensis (serovar israelensis) (ATCC 35646) и штамм Bacillus thuringiensis (serovar israelensis), депонированный во Всероссийской коллекции промышленных микроорганизмов (VKPM) с инвентарным номером VKPM В-197.
Фраза "бактерия, принадлежащая к роду Pseudomonas" означает, что бактерия классифицирована как принадлежащая к роду Pseudomonas в соответствии с классификацией, известной специалисту в области микробиологии. Примеры использованной в настоящем изобретении бактерии, принадлежащей к роду Pseudomonas, включают, но не ограничиваются ими, Pseudomonas syringae.
Фраза "бактерия, принадлежащая к роду Enterobacter" означает, что бактерия классифицирована как принадлежащая к роду Enterobacter в соответствии с классификацией, известной специалисту в области микробиологии. Примеры использованной в настоящем изобретении бактерии, принадлежащей к роду Enterobacter, включают, но не ограничиваются ими, Enterobacter agglomerans. Некоторые виды Enterobacter agglomerans недавно реклассифицированы в Pantoea agglomerans, Pantoea ananatis, Pantoea stewartii и т.п. на основании анализа нуклеотидной последовательности 16S рРНК, и т.д. (Int. J. Syst. Bacteriol., 43, 162-173 (1993)).
L-Изолейциндиоксигеназа из микробных клеток в дальнейшем упоминается в виде аббревиатуры IDO. 4-гидрокси-L-изолейциндегидрогеназа из микробных клеток в дальнейшем упоминается в виде аббревиатуры HIDH.
Активность L-изолейциндиоксигеназы означает активность, катализирующую реакцию гидроксилирования L-изолейцина в присутствии α-кетоглутарата и кислорода, в результате чего образуются 4-HIL, сукцинат и CO2. Активность 4-гидрокси-L-изолейциндегидрогеназы означает активность, катализирующую реакцию образования АМКР из 4-HIL в присутствии NAD+ или NADP. L-Изолейциндиоксигеназную активность IDO настоящего изобретения можно измерить путем анализа образования (2S,3R,4S)-4HIL из L-изолейцина с использованием высокоэффективной жидкостной хроматографии (ВЭЖХ). Также 4-гидрокси -L-изолейциндегидрогеназную активность HIDH настоящего изобретения можно измерить путем анализа образования АМКР из 4-HIL с использованием высокоэффективной жидкостной хроматографии (ВЭЖХ).
Ген, кодирующий HIDH, был идентифицирован при анализе полного генома штамма Bacillus thuringiensis (serovar israelensis) (ATCC 35646), заявлен 17 января 2007 г. в National Center for Biotechnology Information, NIH, Bethesda, MD 20894, USA (Accession No: AAJM00000000.1, GI:74494335), и клонирован из штамма Bacillus thuringiensis (serovar israelensis), полученного из Всероссийской коллекции промышленных микроорганизмов (ВКПМ), хранящегося под инвентарным номером VKPM В-197. Нуклеотидная последовательность из штамма Bacillus thuringiensis (serovar israelensis) ATCC35646, кодирующая HIDH, представлена в SEQ ID NO:1 (см. перечень последовательностей). Аминокислотная последовательность HIDH из штамма Bacillus thuringiensis (serovar israelensis) ATCC35646 представлена в SEQ ID NO: 2.
ДНК, кодирующая HIDH, катализирущую реакцию, в которой 4HIL превращается в АМКР - не только ДНК, показанная в SEQ ID No:1. Это связано с различиями в нуклеотидных последовательностях, наблюдающихся для каждого вида и штамма среди видов, содержащих гены, кодирующие HIDH.
ДНК, кодирующая HIDH, включает не только выделенную ДНК, кодирущую HIDH, но и ДНК, в которой в кодирующую HIDH ДНК, выделенную из хромосомной ДНК HIDH-продуцирующего микроорганизма, искусственно введены мутации, также включена в ДНК настоящего изобретения, при условии, что она кодирует HIDH с активностью, катализирующей указанную реакцию. Методы для искусственного введения мутаций включают обычно используемые методы, такие как метод для введения сайт-специфических мутаций, описанный в Method. in Enzymol., 154 (1987).
ДНК, гибридизующаяся в жестких условиях с ДНК с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности SEQ ID No:1, кодирующая белок, обладающий активностью HIDH, также включена в ДНК настоящего изобретения. Как используется здесь, термин "жесткие условия" означает такие условия, при которых специфические гибриды образуются, тогда как неспецифические гибриды не образуются. Хотя количественное описание этих условий является затруднительным, в качестве примера можно сослаться на условия, при которых молекулы ДНК, имеющие более высокую гомологию, предпочтительно не менее 70%, более предпочтительно не менее 80%, еще более предпочтительно не менее 90% и особенно предпочтительно не менее 95% или более, гибридизуются друг с другом, тогда как молекулы ДНК, имеющие более низкую гомологию, не гибридизуются друг с другом, или такие условия, при которых гибридизация имеет место при обычных условиях отмывки во время проведения гибридизации по Саузерну, которая проводится при концентрации солей, 0.1×SSC, 0.1% SDS при температуре 37°С, предпочтительно 0.1×SSC, 0.1% SDS при температуре 60°С и более предпочтительно 0.1×SSC, 0.1% SDS при температуре 65°С. Длина зонда может быть выбрана соответствующим образом в зависимости от условий гибридизации и обычно варьирует от 100 п.о. до 1 тыс.п.о. Кроме того, определения " HIDH активность" может быть достаточно для активности, которая синтезирует АМКР из 4HIL. Однако в случае, если нуклеотидная последовательность гибридизуется в жестких условиях с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности SEQ ID No:1, она предпочтительно сохраняет HIDH активность величиной 10% или более, предпочтительно 30% или более, более предпочтительно 50% или более и еще более предпочтительно 70% или более, относительно белка с аминокислотной последовательностью SEQ ID No:2 в условиях 37°С и рН 8. Активность HIDH можно измерить путем анализа образования АМКР с использованием высокоэффективной жидкостной хроматографии (ВЭЖХ).
Кроме того, ДНК, кодирующая белок, по существу идентичный HIDH, кодируемой ДНК SEQ ID No:1, также включена в ДНК настоящего изобретения. А именно, следующие ДНК также включены в настоящее изобретение:
(a) ДНК, включающая нуклеотидную последовательность SEQ ID No:1;
(b) ДНК, гибридизующаяся в жестких условиях с ДНК с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности SEQ ID No:1, и кодирующая белок, обладающий HIDH активностью;
(c) ДНК, кодирующая белок, включающий аминокислотную последовательность SEQ ID No:2;
(d) ДНК, кодирующая белок с аминокислотной последовательностью, которая содержит замену, делецию, вставку, добавление или инверсию одной или нескольких аминокислот в последовательности SEQ ID No:2, и обладающий HIDH активностью, и
(e) ДНК, кодирующая белок с аминокислотной последовательнотью, гомологичной по крайней мере на 70%, предпочтительно гомологичной по крайней мере на 80%, более предпочтительно гомологичной по крайней мере на 90% и еще более предпочтительно гомологичной по крайней мере на 95% аминокислотной последовательности SEQ ID No:2, и обладающий HIDH активностью.
Здесь "один или несколько" означает диапазон, вне которого стереоизомерия аминокислотных остатков белка или активность HIDH изменяется незначительно и, в частности, соответствует значениям от 1 до 20, предпочтительно от 1 до 15, более предпочтительно от 1 до 10 и еще более предпочтительно от 1 до 5.
Замена, делеция, вставка, добавление или инверсия одного или нескольких аминокислотных остатков будут представлять собой консервативную(ые) мутацию(и) при условии, что активность фермента при этом сохраняется. Примером консервативной мутации(ий) является(ются) консервативная(ые) замена(ы). Примеры консервативных замен включают замену Ala на Ser или Thr, замену Arg на Gln, His или Lys, замену Asn на Glu, Gln, Lys, His или Asp, замену Asp на Asn, Glu или Gln, замену Cys на Ser или Ala, замену Gln на Asn, Glu, Lys, His, Asp или Arg, замену Glu на Asn, Gln, Lys или Asp, замену Gly на Pro, замену His на Asn, Lys, Gln, Arg или Tyr, замену Ile на Leu, Met, Val или Phe, замену Leu на Ile, Met, Val или Phe, замену Lys на Asn, Glu, Gln, His или Arg, замену Met на Ile, Leu, Val или Phe, замену Phe на Tip, Tyr, Met, Ile или Leu, замену Ser на Thr или Ala, замену Thr на Ser или Ala, замену Trp на Phe или Tyr, замену Tyr на His, Phe или Trp и замену Val на Met, Ile или Leu.
Кроме того, ДНК, гомологичная ДНК SEQ ID No:1, может быть использована в качестве ДНК, кодирующей HIDH. Кодирует ли гомологичная ДНК HIDH, можно подтвердить путем измерения активности в лизате клеток или в лизате клеток микроорганизма, в котором сверхэкспрессируется гомологичная ДНК.
ДНК, гомологичная ДНК SEQ ID No:1, также может быть получена из генома других видов Bacillus, например Bacillus cereus, Bacillus weihenstephanensis. При анализе всех известных бактериальных геномов обнаружено, что несколько микроорганизмов содержат гены с высокой гомологией к гену HIDH Bacillus thuringiensis serovar israelensis (ATCC 35646). Результаты выравнивания аминокислотных последовательностей HIDH Bacillus cereus, Bacillus thuringiensis, Bacillus weihenstephanensis и Pseudomonas syringae показаны на фиг.3. И все эти гены являются вторыми генами в опероне, содержащем ген IDO в качестве первого гена.
Фраза «бактерия модифицирована таким образом, что активность 4-гидрокси-L-изолейциндегидрогеназы в указанной бактерии уменьшена или разрушена» означает, что бактерия модифицирована таким образом, что модифицированная бактерия содержит пониженное количество белка HIDH по сравнению с немодифицированной бактерией, содержит мутантный белок HIDH с пониженной активностью или не способна синтезировать белок HIDH.
Возможный способ уменьшить или разрушить активность HIDH - возможное ослабление или подавление экспрессии гена, кодирующего HIDH. Полное подавление экспрессии гена, кодирующего HIDH, может быть достигнуто путем инактивации гена на хромосоме бактерии.
Фраза "инактивация гена HIDH" означает, что модифицированный ген кодирует полностью нефункциональный белок. Также возможно, что модифицированная область ДНК не способна экспрессировать ген из-зи делеции части гена, сдвига рамки считывания гена, введения миссенс/нонсенс мутации(й) или модификации области, прилегающей к гену, включая последовательности, контролирующие экспрессию гена, такие как промотор, энхансер, аттенюатор, сайт связывания рибосомы и т.д.
Например, для введения мутации путем генной рекомбинации могут использоваться следующие методы. Конструируют мутантный ген и бактерию для ее модификации трансформируют фрагментом ДНК, содержащим мутантный ген. Затем нативный ген на хромосоме заменяют мутантным путем гомологичной рекомбинации и отбирают полученный штамм. Кроме того, введение сайт-специфической мутации путем замены гена с использованием гомологичной рекомбинации изложенным выше способом может быть проведено с использованием плазмиды, лишенной способности реплицироваться в клетке хозяина.
Инактивация гена может быть выполнена традиционными методами, такими как мутагенная обработка с использованием УФ-облучения или обработка нитрозогуанидином (N-метил-1'-нитро-N-нитрозогуанидин), сайт-направленный мутагенез, разрушение гена с использованием гомологичной рекомбинации.
Наличие или отсутствие гена HIDH на хромосоме бактерии можно определить известыми методами, включая ПЦР, блоттинг по Саузерну и т.п. Кроме того, уровень экспрессии гена можно оценить путем измерения количества транскрибируемой с гена мРНК с использованием различных известных методов, включая блоттинг по Нозерну, количественную ПЦР в реальном времени и т.п. Количество кодируемого геном белка можно измерить известными методами, включая SDS-ПААГ с последующим иммуноблоттингом (Вестерн-блоттинг) и т.п.
Методы приготовления плазмидной ДНК, рестрикции и лидирования ДНК, трансформации, выбора нуклеотидов в качестве праймера и т.п. могут быть обычными методами, известными специалисту в этой области. Эти методы описаны, например, в Sambrook, J., Fritsch, E.F., and Maniatis, Т., "Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989).
Для эффективной продукции 4-HIL необходимо увеличить активность белка IDO, катализирующего реакцию образования 4-HIL из L-изолейцина.
Нуклеотидная последовательность, кодирующая IDO, и аминокислотная последовательность IDO штамма Bacillus thuringiensis (serovar israelensis) VKPM В-197 представлены в SEQ ID NO:3 и 4 соответственно.
ДНК, кодирующая IDO, катализирующую реакцию, в которой (2S,3R,4S)-4HIL образуется из L-изолейцина- не только ДНК, показанная в SEQ ID No: 3. Это связано с различиями в нуклеотидных последовательностях, наблюдающихся для каждого вида и штамма среди видов Bacillus, которые образуют IDO, катализирующую реакцию образования (2S,3R,4S)-4HIL из L-изолейцина.
ДНК настоящего изобретения включает не только выделенные ДНК, кодирующие IDO, но ДНК, в которых в ДНК, кодирующей IDO, выделенной из хромосомной ДНК IDO-продуцирующих микроорганизмов, искусственно введены мутации, также включены в ДНК настоящего изобретения при условии, что ДНК кодирует IDO с активностью, катализирующей указанную реакцию. Методы для искусственного введения мутаций включают обычно используемые методы, такие как метод для введения сайт-специфических мутаций, описанный в Method. in Enzymol., 154 (1987).
ДНК, гибридизующаяся в жестких условиях с ДНК с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности SEQ ID No:3, кодирующая белок, обладающий активностью IDO, также включена в ДНК настоящего изобретения. Как используется здесь, термин "жесткие условия" означает такие условия, при которых специфические гибриды образуются, тогда как неспецифические гибриды не образуются. Хотя количественное описание этих условий является затруднительным, в качестве примера можно сослаться на условия, при которых молекулы ДНК, имеющие более высокую гомологию, предпочтительно не менее 70%, более предпочтительно не менее 80%, еще более предпочтительно не менее 90% и особенно предпочтительно не менее 95% или более, гибридизуются друг с другом, тогда как молекулы ДНК, имеющие более низкую гомологию, не гибридизуются друг с другом, или такие условия, при которых гибридизация имеет место при обычных условиях отмывки во время проведения гибридизации по Саузерну, которая проводится при концентрации солей, 0,1×SSC, 0,1% SDS при температуре 37°С, предпочтительно 0,1×SSC, 0,1% SDS при температуре 60°С и более предпочтительно 0,1×SSC, 0,1% SDS при температуре 65°С. Длина зонда может быть выбрана соответствующим образом, в зависимости от условий гибридизации и обычно варьирует от 100 п.о. до 1 тыс.п.о. Кроме того, определения "L-изолейциндиоксигеназная активность" может быть достаточно для активности, которая синтезирует (2S,3R,4S)-4HIL из L-изолейцина. Однако в случае, если нуклеотидная последовательность гибридизуется в жестких условиях с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности SEQ ID No:3, она предпочтительно сохраняет L-изолейциндиоксигеназную активность величиной 10% или более, предпочтительно 30% или более, более предпочтительно 50% или более и еще более предпочтительно 70% или более, относительно белка с аминокислотной последовательностью SEQ ID No:4 в условиях 37°С и рН 8.
L-Изолейциндиоксигеназная активность IDO может быть определена путем анализа образования (2S,3R,4S)-4HIL из L-изолейцина с использованием высокоэффективной жидкостной хроматографии (ВЭЖХ) (см. пример 2 и фиг.3).
Кроме того, ДНК, кодирующая белок, по существу идентичный IDO, кодируемой ДНК SEQ ID No:3, также включена в ДНК настоящего изобретения. А именно, следующие ДНК также включены в настоящее изобретение:
(a) ДНК, включающая нуклеотидную последовательность SEQ ID No:3;
(b) ДНК, гибридизующаяся в жестких условиях с ДНК с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности SEQ ID No:3 и кодирующая белок, обладающий L-изолейциндиоксигеназной активностью;
(c) ДНК, кодирующая белок, включающий аминокислотную последовательность SEQ ID No:4;
(d) ДНК, кодирующая белок с аминокислотной последовательностью, которая содержит замену, делецию, вставку, добавление или инверсию одной или нескольких аминокислот в последовательности SEQ ID No:4, и обладающий L-изолейциндиоксигеназной активностью, и
(e) ДНК, кодирующая белок с аминокислотной последовательностью, гомологичной по крайней мере на 70%, предпочтительно гомологичной по крайней мере на 80%, более предпочтительно гомологичной по крайней мере на 90% и еще более предпочтительно гомологичной по крайней мере на 95% аминокислотной последовательности SEQ ID No:4 и обладающий L-изолейциндиоксигеназной активностью.
Здесь "один или несколько" означает диапазон, вне которого стереоизомерия аминокислотных остатков белка или активность L-изолейциндиоксигеназы изменяется незначительно и, в частности, соответствует значениям от 1 до 20, предпочтительно от 1 до 15, более предпочтительно от 1 до 10 и еще более предпочтительно от 1 до 5.
Замена, делеция, вставка, добавление или инверсия одного или нескольких аминокислотных остатков будут представлять собой консервативную(ые) мутацию(и) при условии, что активность фермента при этом сохраняется. Примером консервативной мутации(ий) является(ются) консервативная(ые) замена(ы). Примеры консервативных замен включают замену Ala на Ser или Thr, замену Arg на Gln, His или Lys, замену Asn на Glu, Gln, Lys, His или Asp, замену Asp на Asn, Glu или Gln, замену Cys на Ser или Ala, замену Gln на Asn, Glu, Lys, His, Asp или Arg, замену Glu на Asn, Gln, Lys или Asp, замену Gly на Pro, замену His на Asn, Lys, Gln, Arg или Tyr, замену Ile на Leu, Met, Val или Phe, замену Leu на Ile, Met, Val или Phe, замену Lys на Asn, Glu, Gln, His или Arg, замену Met на Ile, Leu, Val или Phe, замену Phe на Trp, Tyr, Met, Ile или Leu, замену Ser на Thr или Ala, замену Thr на Ser или Ala, замену Trp на Phe или Tyr, замену Tyr на His, Phe или Trp и замену Val на Met, Ile или Leu.
ДНК, гомологичная ДНК SEQ ID NO: 3, также может быть получена из генома других видов Bacillus, например Bacillus cereus, Bacillus weihenstephanensis.
Кроме того, кодирующие IDO гомологичные ДНК из других бактерий могут быть получены клонированием на основе гомологии со следующими генами (см. таблицу 1): Bacillus, Escherichia, Corynebacterium, Arthrobacter, Aspergillus, Pseudomonas, Granulibacter, Methylobacillus, Granulibacter, Acidiphilium, Agrobacterium, Gluconobacter, Caulobacter, Stigmatella, Myxococcus, Polaromonas, Caulobacter, Polaromonas, Sphingomonas, Acidovorax, Mycobacterium, Azotobacter, Vibrio, Polynucleobacter, Streptomyces и т.п.
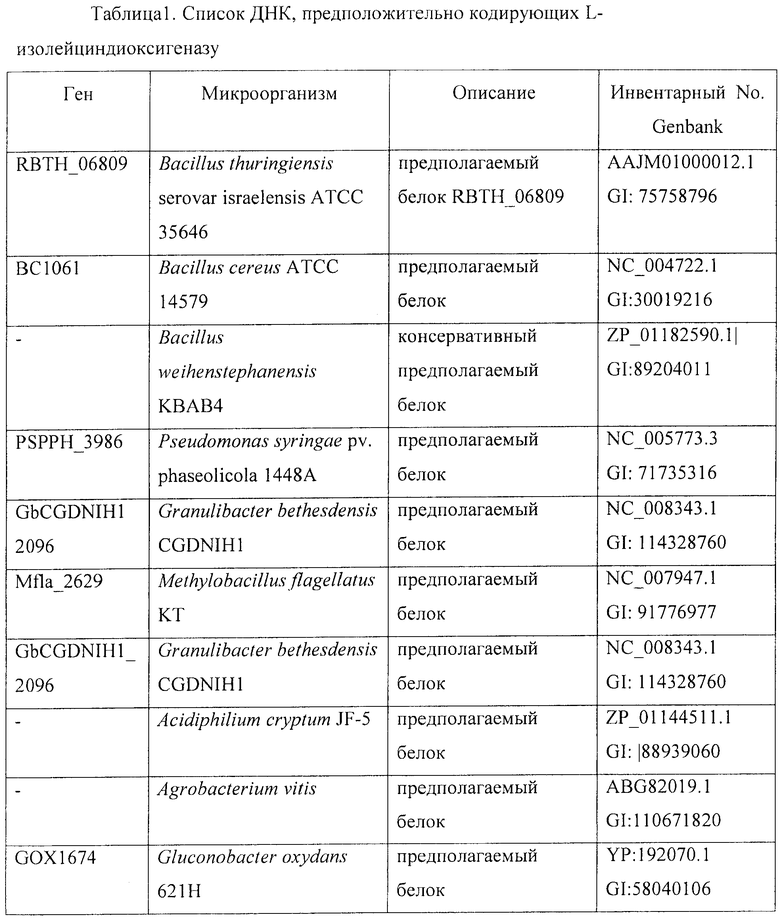

Фраза "бактерия, модифицированная таким образом, что в ней увеличена активность L-изолейциндиоксигеназы" означает, что указанная бактерия модифицирована таким образом, что эта бактерия содержит повышенное количество белка IDO в сравнении с немодифицированной бактерией. Одним из путей увеличения количества белка IDO является повышение экспрессии гена, кодирующего белок IDO.
Фраза "усиление экспрессии гена" означает, что экспрессия гена выше, чем в немодифицированном штамме, например в штамме дикого типа. Примеры таких модификаций включают увеличение числа копий экспрессируемого(ых) гена(ов) на клетку, увеличение уровня экспрессии гена(ов) и т.д. Количество числа копий экспрессируемого гена измеряют, например, путем рестрикции хромосомной ДНК с последующим блоттингом по Саузерну с использованием зонда, приготовленного на основе последовательности гена, флуоресцентной гибридизации in situ (FISH), и т.п. Уровень экспресии гена можно измерить различными известными методами, включая блоттинг по Нозерну, количественную ОТ-ПЦР в реальном времени и т.п. Количество белка, кодируемого геном, можно определить известными методами, включая электрофорез в SDS-ПААГ с последующим иммуноблоттингом (Вестерн-блоттинг), и т.п.
"Трансформация бактерии ДНК, кодирующей белок" означает введение ДНК в бактерию, например, традиционными методами. В результате трансформации этой ДНК увеличится экспрессия гена, кодирующего белок настоящего изобретения, и увеличится активность белка в бактериальных клетках. Методы трансформации включают любые известные упомянутые ранее методы.
Методы усиления экспрессии гена включают увеличение числа копий гена. Введение гена в вектор, способный функционировать в бактерии настоящего изобретения, увеличивает число копий гена. Для этого предпочтительно использовать многокопийные векторы.
Усиление экспрессии гена может также быть достигнуто путем введения множества копий гена в бактериальную хромосому с использованием, например, гомологичной рекомбинации.
Увеличение числа копий гена также может быть достигнуто путем введения множества копий гена в хромосомную ДНК бактерии. Для введения множества копий гена в бактериальную хромосому выполняют гомологичную рекомбинацию с использованием последовательности, существующей во множестве копий, в качестве мишеней в хромосомной ДНК. Последовательности с множеством копий в хромосомной ДНК включают, но не ограничиваются ими, повторяющуюся ДНК, или инвертированные повторы на концах транспозонных элементов.
Усиление экспрессии гена также может быть достигнуто путем помещения ДНК настоящего изобретения под контроль сильного промотора. Использование сильного промотора можно комбинировать с увеличением копий гена.
Кроме того, также возможно ввести нуклеотидную замену в области промотора гена на бактериальной хромосоме, ведущую к усилению функции промотора. Изменение последовательности, контролирующей экспрессию, можно осуществить, например, тем же способом, что и замещение гена с использованием плазмиды, экспрессия которой чувствительна к температуре, как раскрыто в заявке РСТ WO 00/18935 и выложенной патентной заявке Японии No. 1-215280.
Способы приготовления плазмидной ДНК включают, но не ограничиваются ими, рестрикцию и лигирование ДНК, трансформацию, выбор олигонуклеотида для использования в качестве праймера и т.п. или другие методы, известные специалисту в данной области. Эти методы описаны, например, в "Molecular Cloning A Laboratory Manual, Third Edition", Cold Spring Harbor Laboratory Press (2001).
Способ продукции (2S,3R,4S)-4-гидрокси-L-изолейцина настоящего изобретения включает одну стадию прямого ферментативного гидроксилирования L-изолейцина для получения (2S,3R,4S)-4HIL, представленную следующей реакцией:
L-изолейцин+α-кетоглутарат+О2→4HIL+сукцинат+СО2
при этом реакцию осуществляют в присутствии L-изолейцина в качестве молекулы- акцептора одного атома кислорода, α-кетоглутарата в качестве молекулы-акцептора еще одного атома кислорода, одной молекулы кислорода в качестве донора двух атомов кислорода и бактерии, содержащей IDO, катализирующую реакцию.
Для осуществления реакции гидроксилирования с использованием IDO реакционная смесь, содержащая L-изолейцин, α-кетоглутарат и бактерию, содержащую IDO, катализирующую реакцию, поддерживают при подходящей температуре от 20 до 50°С в покое, при встряхивании или помешивании в течение от 30 минут до 5 дней при рН 5-12.
Образующийся в реакционной смеси (2S,3R,4S)-4HIL можно выделить или очистить в соответствии с известными методами.
Примеры методов выделения и очистки могут включать метод, в котором (2S,3R,4S)-4HIL контактирует с ионообменной смолой для адсорбции основных аминокислот с последующей элюцией и кристаллизацией, и метод, в котором продукт, полученный после элюции, обесцвечивают и фильтруют через активированный уголь с последующей кристаллизацией для получения (2S,3R,4S)-4HIL.
В способе согласно настоящему изобретению бактерия - бактериальная культура, клетки, обработанные клетки или лизат клеток.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
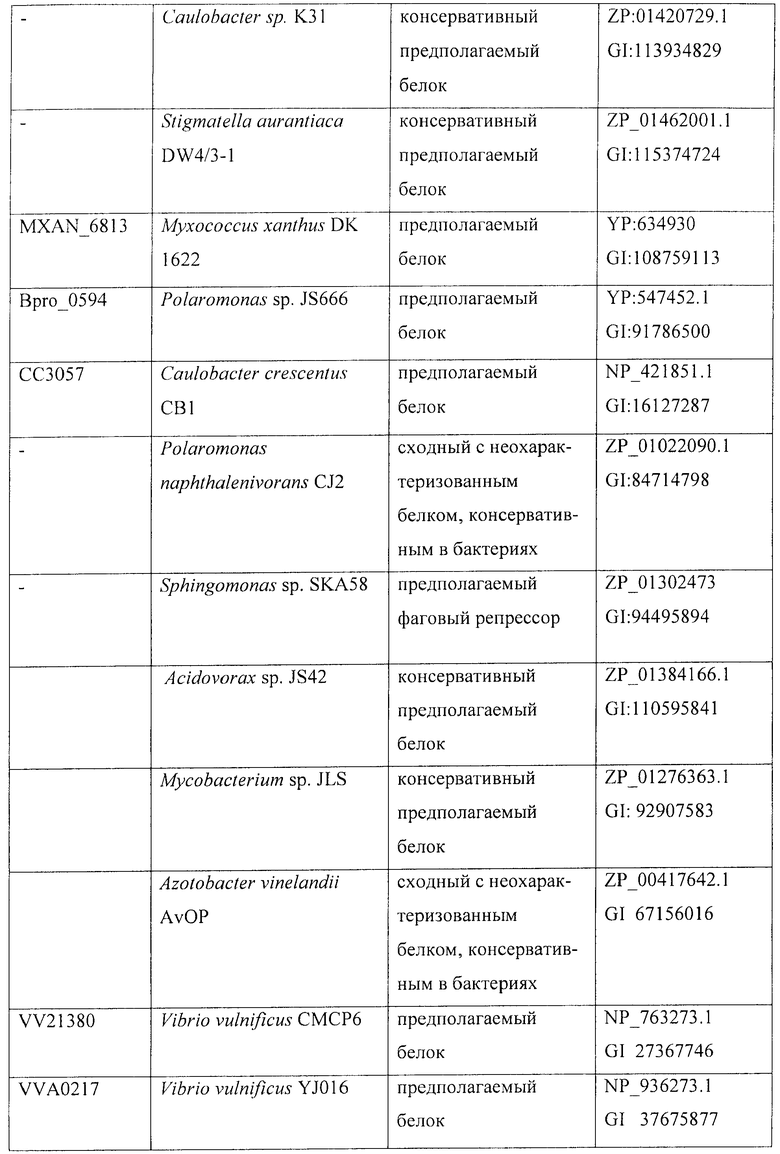
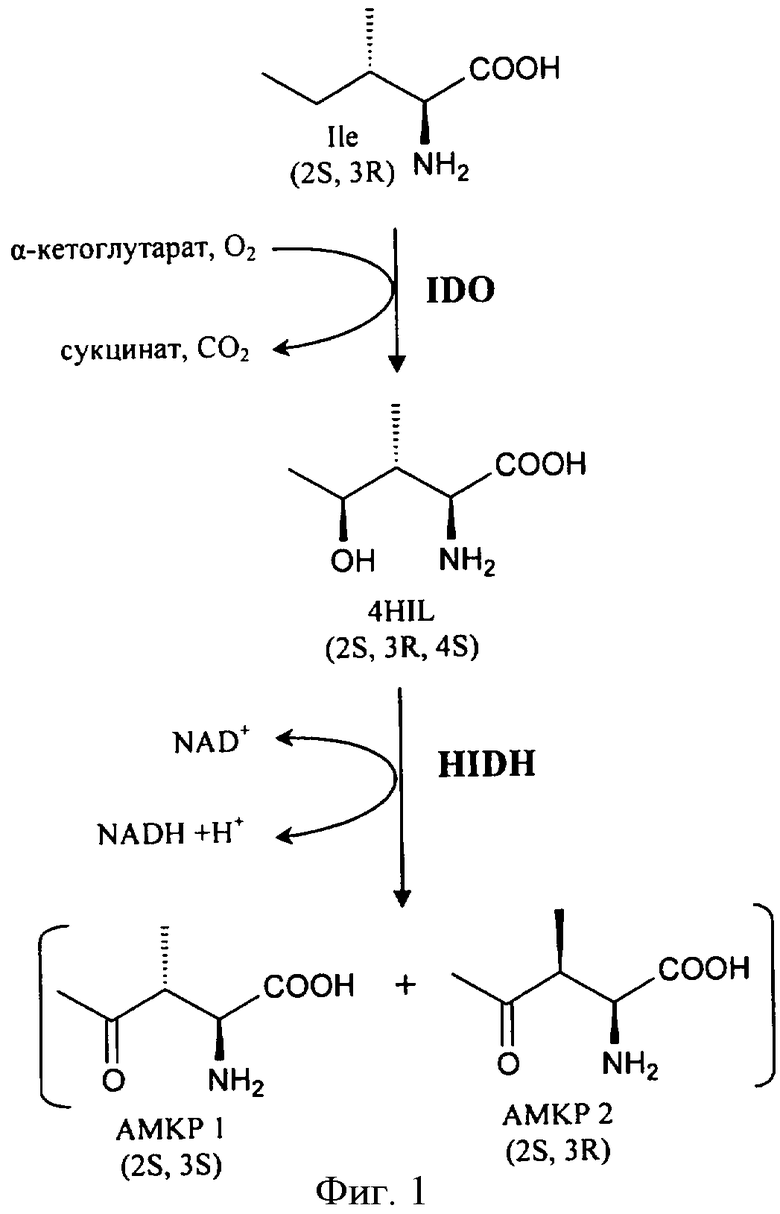
На фиг.1 показан предполагаемый путь синтеза АМКР в Bacillus fhuringiensis. На фиг.2 показаны: А - структура оперона, содержащего гены, кодирующие IDO и HIDH Bacillus thuringiensis serovar israelensis (ATCC 35646), трансляционно сопряженные с использованием "ATGA"; В - структура сконструированной рекомбинантной плазмиды, содержащей IDO-HIDH оперон.
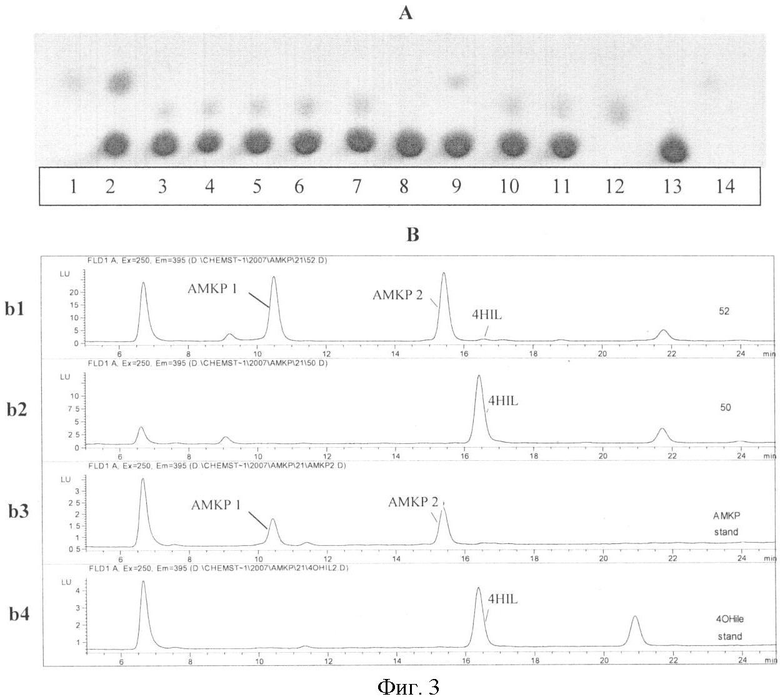
На фиг.3 показана биоконверсия изолейцина в АМКР с использованием штама Е.coli, обладающего активностями IDO и HIDH, А - ТСХ анализ продукции АМКР. Дорожки: 1, 14 - стандартный 4HIL; 12 - стандартный АМКР; 13 - стандартный Ile; 2 - культуральная жидкость штамма TG1[pMW119-IDO (2-е-2)]; 3-7 - культуральная жидкость штаммов TG1[pMW119-IDO-HIDH (2-е-2)] (пять независимых клонов); 8 - культуральная жидкость штамма TG1 [pMW119]; 9 - культуральная жидкость штамма TG1 [pMW119-IDO(VKPM В-197)]; 10-11 - культуральная жидкость штаммов TG1 [pMW119-IDO-HIDH (VKPM В-197)] (два независимых клона). В - ВЭЖХ анализ продукции АМКР. b1 - культуральная жидкость штамма TG1 [pMW119-IDO-HIDH (2-е-2)]; b2 - культуральная жидкость штамма TG1 [pMW119-IDO (2-е-2)]; b3 - стандартный АМКР; b4 - стандартный 4HIL. На фиг.4 показан результат выравнивания аминокислотных последовательностей HIDH Bacillus thuringiensis serovar israelensis ATCC 35646 (ZP_00738909). короткоцепочечной дегидрогеназы/редуктазы SDR Bacillus weihenstephanensis KBAB4 (ZP_01182591), 3-оксоацил-[ацил-переносящий белок]-редуктазы Bacillus cereus ATCC 14579 (NP_830848), гипотетического белка GOS_9257808 [метагеном морского вида] (ЕВН46681) и 3-оксоацил-[ацил-переносящий белок]-редуктазы Pseudomonas syringae pv. phaseolicola 1448A (YP_276118).
Примеры
Настоящее изобретение будет более подробно описано ниже со ссылкой на следующие не ограничивающие настоящее изобретение примеры.
Пример 1. Анализ генома Bacillus thuringiensis и клонирование оперона IDO-HIDH Bacillus thurinsiensis.
Анализ генома Bacillus thuringiensis показал, что трансляция гена IDO (RBTH_06809) сопряжена со следующим соседним геном RBTH_06810 (фиг. 2, А). Обычно такая "синхронизация " используется для эквимолярного синтеза ферментов, катализирующих последовательные реакции. Кроме того, RBTH_06810 кодирует белок, гомологичный 3-оксоацил-[ацилпереносящий белок] редуктазе (дегидрогеназе). Поэтому предположили, что этот ген кодирует HIDH.
Для подтверждения этой гипотезы клонировали оперон IDO-HIDH штамма Bacillus thuringiensis 2-e-2 (PERM ВР-10688) и Bacillus thuringiensis (serovar israelensis) (VKPM B-197). Для этого фрагмент ДНК длиной 1.5 т.п.н., содержащий гены IDO-HIDH фланкированные сайтами рестриктаз BamHl и Sad, амплифицировали в ПЦР с использованием праймеров SVS170 (SEQ ID NO: 5) и SVS171 (SEQ ID NO: 6) и хромосомной ДНК штаммов 2-e-2 или VKPM B-197 в качестве матрицы. Полученные фрагменты обрабатывали рестриктазами BamHI и SacI и лигировали в вектор pMW119, предварительно обработанный теми же рестриктазами. Полученные плазмиды вводили в штамм Е. coli TG1. В результате сконструировали штаммы- продуценты IDO/HIDH: TG1[pMW119-IDO-HIDH (2-e-2)] и TG1[pMW119-IDO-HIDH (VKPM B-197)] (фиг.2, В). Также фрагменты хромосомной ДНК длиной 0,8 т.п.н., содержащие ген IDO, амплифицировали в ПЦР с использованием праймеров SVS170 (SEQ ID No: 5) и хромосомной ДНК штаммов 2-e-2 или VKPM B-197 в качестве матрицы. Полученные ПЦР-фрагменты обрабатывали эндонуклеазами BamHl и SacI и затем лигировали в вектор pMW119, предварительно обработанный теми же рестриктазами. Полученные плазмиды вводили в штамм Е. coli TG1. В результате сконструировали штаммы - продуценты IDO: TG1 [pMW119-IDO (2-e-2)] и TG1 [pMW119-IDO (VKPM B-197)].
Сконструированные рекомбинантные штаммы Е. coli - продуценты IDO/HIDH использовали для превращения изолейцина в АМКР (фиг.3, таблица 2). Клетки штаммов инокулировали в 2 мл среды MI50 и культивировали при 32°С в течение 72 ч в пробирках с энергичным встряхиванием. После культивирования определяли концентрации АМКР и 4HIL в культуральной жидкости с использованием ТСХ и ВЭЖХ. Для ТСХ использовали пластины размером 10×15 см, покрытые 0,11-мм слоем силикагеля Сорбфил, не содержащего флуоресцентный индикатор (Акционерное общество сорбполимер, Краснодар, Россия). Пластинки Сорбфил экспонировали в подвижной фазе следующего состава: пропан-2-ол : этилацетат : 25% водного аммиака : вода = 40:40:7:16 (v/v). Раствор (2%) нингидрина в ацетоне использовали для визуализации.
Состав использованной среды MI50 (г/л):
СаСО3 стерилизовали сухим жаром при 180°С в течение 2 часов. Доводили рН до 7,0 с использованием NaOH.
Как видно из полученных данных, экспрессия только IDO вела к синтезу 4HIL, тогда как экспрессия IDO-HIDH вела к почти полной трансформации 4HIL в АМКР. Таким образом, было установлено, что ген RBTH_06810 кодирует HIDH.
Пимер 2. Измерение с использованием ВЭЖХ накопленных 4-HIL и АМКР
ВЭЖХ анализ: использовали хроматограф высокого давления (Waters, USA) со спектрофлуорометром серии 1100 (Agilent, USA). Выбранные диапазоны длин волн: длина волны возбуждения 250 нм, область длин волн эмиссии 320-560 нм. Разделение с помощью метода accq-tag осуществляли на колонке Nova-PakTM С 18 150×3,9 мм, 4 мкм (Waters, США) при +40°С. Объем пробы составлял 5 мкл. Образование производных аминокислот и их разделение осуществляли в соответствии с рекомендациями производителя Waters (Liu, H. et al., J. Chromatogr. A, 828, 383-395 (1998); Waters accq-tag chemistry package. Instruction manual. Millipore Corporation, pp.1-9 (1993)). Для получения производных аминокислот с 6-аминохинолил-N-гидроксисукцинимидилкарбаматом использовали набор Accq-FluorTM (Waters, США). Анализ с помощью метода accq-tag осуществляли с использованием концентрированного элюента Accq-tag Eluent A (Waters, США). Все растворы готовили с использованием воды Milli-Q, стандартные растворы хранили при +4°С.
Пример 3. Поиск гомологов HIDH Bacillus thuringiensis serovar israelensis (ATCC 35646).
В результате анализа всех известных бактериальных геномов обнаружили, что несколько микроорганизмов содержат гены с высокой гомологией с геном HIDH Bacillus thuringiensis serovar israelensis (ATCC 35646). Результат выравнивания аминокислотных последовательностей HIDH Bacillus cereus, Bacillus thuringiensis, Bacillus weihenstephanensis и Pseudomonas syringae показан на Фиг.3. Предположили, что все эти белки могут проявлять активность HIDH.
Пример 4. Конструирование штамма с инактивированным геном HIDH
1. Делеция гена HIDH
Штамм с делецией гена HIDH может быть сконструирован с использованием следующей техники. Фрагмент ДНК, содержащий маркер для селекции, можно получить в ПЦР с использованием соответствующих праймеров. Каждый праймер должен содержать область, комплементарную области, локализованной на 5'- конце или 3'- конце гена HIDH. Делецию гена HIDH можно осуществить путем замены нативного гена HIDH полученным фрагментом ДНК с использованием гомологичной рекомбинации.
2. Подтверждение делеции гена HIDH с использованием ПЦР
Мутанты с делецией гена HIDH, содержащие ген маркера для селекции, можно проверить с использованием ПЦР. В ПЦР для подтверждения следует использовать локус-специфические праймеры. Делеция гена HIDH может быть подтверждена при различии в длине продуктов ПЦР, полученных при использовании в качестве матрицы хромосомных ДНК родительских и мутантных штаммов.
Хотя указанное изобретение описано в деталях со ссылкой на наилучший способ осуществления изобретения, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.
Каждому из упомянутых выше документов соответствует ссылка, и все цитируемые документы являются частью описания настоящего изобретения.







| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПРОДУКЦИИ 2-АМИНО-3-МЕТИЛ-4-КЕТОПЕНТАНОАТА | 2007 |
|
RU2392323C2 |
| СПОСОБ КОНСТРУИРОВАНИЯ БАКТЕРИИ-ПРОДУЦЕНТА (2S,3R,4S)-4-ГИДРОКСИ-L-ИЗОЛЕЙЦИНА, БАКТЕРИЯ-ПРОДУЦЕНТ (2S,3R,4S)-4-ГИДРОКСИ-L-ИЗОЛЕЙЦИНА И СПОСОБ ПРОДУКЦИИ (2S,3R,4S)-ГИДРОКСИ-L-ИЗОЛЕЙЦИНА ИЛИ ЕГО СОЛИ | 2007 |
|
RU2395580C2 |
| БАКТЕРИЯ - ПРОДУЦЕНТ ПРОДУКТА РЕАКЦИИ, КАТАЛИЗИРУЕМОЙ БЕЛКОМ, ОБЛАДАЮЩИМ АКТИВНОСТЬЮ 2-ОКСОГЛУТАРАТ-ЗАВИСИМОГО ФЕРМЕНТА, И СПОСОБ ПРОДУКЦИИ УКАЗАННОГО ПРОДУКТА | 2008 |
|
RU2444568C2 |
| СПОСОБ ПРОДУКЦИИ 4-ГИДРОКСИ-L-ИЗОЛЕЙЦИНА | 2007 |
|
RU2402608C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГИДРОКСИЛИРОВАННОГО L-ЛЕЙЦИНА И БАКТЕРИЯ, ТРАНСФОРМИРОВАННАЯ ДНК, КОДИРУЮЩЕЙ ДИОКСИГЕНАЗУ | 2011 |
|
RU2468085C1 |
| НОВАЯ АЛЬДОЛАЗА, ДНК, КОДИРУЮЩАЯ АЛЬДОЛАЗУ, КЛЕТКИ, ТРАНСФОРМИРОВАННЫЕ ДНК, СПОСОБ ПОЛУЧЕНИЯ АЛЬДОЛАЗЫ И СПОСОБ ПОЛУЧЕНИЯ 4-ГИДРОКСИ-L-ИЗОЛЕЙЦИНА (ВАРИАНТЫ) | 2006 |
|
RU2338784C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ МЕТОДОМ ФЕРМЕНТАЦИИ, ШТАММ БАКТЕРИИ ESCHERICHIA COLI - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 1999 |
|
RU2207376C2 |
| СПОСОБ ПРОИЗВОДСТВА 4-ГИДРОКСИ-L-ИЗОЛЕЙЦИНА ИЛИ ЕГО СОЛИ | 2006 |
|
RU2355767C2 |
| МАЛАЯ СУБЪЕДИНИЦА ИЗОЗИМА III И ИЗОЗИМ III СИНТЕТАЗЫ АЦЕТОГИДРОКСИКИСЛОТ ИЗ ESCHERICHIA COLI, ФРАГМЕНТ ДНК (ВАРИАНТЫ), ШТАММ БАКТЕРИИ ESCHERICHIA COLI - ПРОДУЦЕНТ L-ВАЛИНА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ L-ВАЛИНА | 2000 |
|
RU2209246C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2007 |
|
RU2364628C2 |

Изобретение относится к биотехнологии и представляет собой способ продукции (2S,3R,4S)-4-гидрокси-L-изолейцина или его соли, включающий стадии реакции L-изолейцина в водном растворе в присутствии бактерии, изначально обладающей активностями L-изолейциндиоксигеназы и 4-гидрокси-L-изолейциндегидрогеназы, и выделения синтезированного (2S,3R,4S)-4-гидрокси-L-изолейцина. При этом указанная бактерия модифицирована таким образом, что активность 4-гидрокси-L-изолейциндегидрогеназы в указанной бактерии разрушена. Изобретение позволяет упростить способ получения (2S,3R,4S)-4-гидрокси-L-изолейцина. 5 з.п. ф-лы, 4 ил., 2 табл.
1. Способ продукции (2S,3R,4S)-4-гидрокси-L-изолейцина или его соли, включающий стадии:
реакции L-изолейцина в водном растворе в присутствии бактерии, изначально обладающей активностями L-изолейциндиоксигеназы и 4-гидрокси-L-изолейциндегидрогеназы, и
выделения синтезированного (2S,3R,4S)-4-гидрокси-L-изолейцина, отличающийся тем, что указанная бактерия модифицирована таким образом, что активность 4-гидрокси-L-изолейциндегидрогеназы в указанной бактерии разрушена.
2. Способ по п.1, отличающийся тем, что гены, кодирующие L- изолейциндиоксигеназу и 4-гидрокси-L-изолейциндегидрогеназу, составляют оперон.
3. Способ по п.1, отличающийся тем, что указанная бактерия модифицирована таким образом, что активность L-изолейциндиоксигеназы в указанной бактерии увеличена.
4. Способ по п.1, отличающийся тем, что активность 4-гидрокси-L-изолейциндегидрогеназы в указанной бактерии разрушена путем подавления экспрессии гена, кодирующего 4-гидрокси-L-изолейциндегидрогеназу.
5. Способ по п.1, отличающийся тем, что указанная бактерия принадлежит к Bacillus thuringiensis, Bacillus weihenstephanensis, Bacillus cereus, Enterobacter agglomerance или Pseudomonas syringae.
6. Способ по п.1, отличающийся тем, что указанной бактерией являются бактериальная культура, клетки, обработанные клетки или клеточный лизат.
| HAEFELE С et.al | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Электрическое сопротивление для нагревательных приборов и нагревательный элемент для этих приборов | 1922 |
|
SU1997A1 |
| Устройство для разметки | 1984 |
|
SU1268397A1 |
| US 20070043240 A1, 22.02.2007 | |||
| US 20030219880 A1, 27.11.2003 | |||
| ЛЕНИНДЖЕР А | |||
| Основы биохимии, том 2 | |||
| Приспособление для установки двигателя в топках с получающими возвратно-поступательное перемещение колосниками | 1917 |
|
SU1985A1 |

