Область техники
Изобретение относится к генной инженерии и может быть использовано в биотехнологии, медицине и сельском хозяйстве для создания препаратов генной терапии.
Уровень техники
Генная терапия - это современный медицинский подход, направленный на лечение наследственных и приобретенных заболеваний путем введения нового генетического материала в клетки пациента с целью компенсации или подавления функции мутантного гена и/или исправления генетического дефекта. Конечным продуктом экспрессии гена может являться молекула РНК или белка. Однако осуществление большей части физиологических процессов в организме связано с функциональной активностью белковых молекул, тогда как молекулы РНК являются либо промежуточным продуктом в синтезе белков, либо осуществляют регуляторные функции. Таким образом, целью генной терапии является, в большинстве случаев, введение в организм генов, обеспечивающих транскрипцию и последующую трансляцию белковых молекул, кодируемых этими генами. В рамках описания настоящего изобретения под экспрессией гена подразумевается продукция белковой молекулы, аминокислотная последовательность которой кодируется этим геном. Мутации в генах могут приводить к полной или частичной потере экспрессии белков или же экспрессии вариантов белковых молекул, обладающих нежелательной функциональной активностью. Введение в организм генотерапевтических векторов, кодирующих тот или иной ген, может восстанавливать экспрессию целевых белков. Однако данный подход носит компенсирующий характер и не направлен на исправление генетических дефектов. С открытием систем направленного (таргетного) редактирования нуклеотидных последовательностей стало возможным, вводя в генотерапевтические векторы различные нуклеазы, обладающие специфическими свойствами (например, Cas9), реализовать подход терапевтического геномного редактирования, которое направлено на исправление мутаций в последовательности ДНК, что также является таргетной генной терапией. В этом случае происходит восстановление функции за счет исправление генетических дефектов (Memi F, Ntokou A, Papangeli I, 2018; Hussain W et al., 2019).
Ген Cas9 кодирует белок-нуклеазу. CRISPR/Cas9 система изначально была открыта как компонент иммунной системы бактерий, который позволяет направленно избавляться бактериальным клеткам от нуклеотидных последовательностей бактериофагов (Sapranauskas R, 2011; Mougiakos I, 2017). Поскольку данная система обладает определенной степенью универсальности принципа действия, она нашла широкое применение в биомедицинских и биотехнологических исследованиях. В настоящее время CRISPR/Cas9 система широко используется в рамках научных исследований для редактирования геномов в культурах клеток млекопитающих и лабораторных животных и обладает потенциалом для разработки средств и способов генной терапии. Принцип работы данной системы заключается в том, что эндонуклеаза Cas9 при помощи gRNA, комплементарной определенной последовательности в геноме, вносит разрывы в цепи ДНК, вырезая участок таргетной ДНК. Целостность ДНК в месте разрывов затем восстанавливается с помощью клеточных систем репарации, которые в качестве матрицы для восстановления могут использовать гомологичную цепь ДНК, содержащую корректную последовательность нуклеотидов, или восстанавливать разрывы путем непосредственного соединения соседних нуклеотидов без репарации вырезанного участка ДНК (Salsman J, 2017). Конструирование gRNA таким образом, чтобы эта молекула была комплементарна участку ДНК, в котором содержится та или иная мутация, позволяет с помощью Cas9 направленно вырезать именно этот участок, что и обуславливает потенциал данного механизма действия в коррекции генетического материала, то есть редактировании геномов (Wilson LOW, 2018).
Тем не менее одной из основных проблем использования системы CRISPR/Cas9 для редактирования геномов остается проблема доставки комплекса эндонуклеазы и gRNA в клеточное ядро и в целом проблемы фармакокинетики, которые ограничивают проникающую способность молекул в различные органы и ткани или требуют введения слишком высоких концентраций и использования специальных композиций, обеспечивающих проникновение в клетки (Dowdy SF, 2017). Преодолеть данные ограничения позволяет использование генетических векторов для гетерологичной экспрессии гена Cas9. Наиболее изученными векторами в этой области являются лентивирусные, аденовирусные, аденоассоциированные и прочие вектора вирусного происхождения.
Другой проблемой использования CRISPR/Cas9 системы остается риск неспецифического действия эндонуклеазы. В этом ключе использование векторов, которые не интегрируют в геном и обеспечивают только транзиентную экспрессию генов, является потенциально более безопасным, чем, например, использование лентивирусных и аденоассоциированных векторов (Li L, Hu S, Chen X, 2017). Однако использование любых вирусных векторов для доставки в организм тех или иных последовательностей ограничено тропизмом псевдовирусных частиц к различным тканям, что не всегда позволяет осуществлять эффективное проникновение в целевые клетки и органы (Maginnis MS, 2017). Также потенциал использования любых вирусных векторов ограничен, в том числе, их относительно высокой иммуногенностью, предсуществующим иммунитетом и рисками, связанными с генотерапевтическими векторами вирусного происхождения в целом (Lukashev AN, Zamyatnin AA, 2016).
Таким образом предшествующий уровень техники свидетельствует о том, что существует потребность в разработке эффективных и безопасных генотерапевтических подходов для доставки Cas9 в целевые орагны и ткани.
Генотерапевтические векторы, как известно, разделяют на вирусные, клеточные и ДНК-векторы (Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal Products EMA/CAT/80183/2014). В последнее время в генной терапии всё большее внимание уделяется разработке невирусных систем доставки генетического материала, среди которых лидируют плазмидные векторы. Плазмидные векторы лишены недостатков, присущих клеточным и вирусным векторам. В клетке-мишени они существуют в эписомальной форме, не интегрируют в геном, производство их достаточно дешево, отсутствие иммунного ответа и побочных реакций на введение плазмидного вектора делают их удобным инструментом генной терапии и генетической профилактики (ДНК-вакцины) (Li L, Petrovsky N. // Expert Rev Vaccines. 2016;15(3):313-29).
Тем не менее, ограничениями для использования плазмидных векторов для генной терапии являются: 1) наличие генов устойчивости к антибиотикам для наработки в бактериальных штаммах, 2) наличие различных регуляторных элементов, представленных последовательностями вирусных геномов 3) размер терапевтического плазмидного вектора, определяющий эффективность проникновения вектора в клетку-мишень.
Известно, что Европейское агентство по лекарственным средствам считает необходимым избегать введения маркеров антибиотикорезистентности в разрабатываемые плазмидные векторы для генной терапии (Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011 EMA/CAT/GTWP/44236/2009 Committee for advanced therapies). Данная рекомендация связана, в первую очередь, с потенциальной опасностью проникновения ДНК-вектора или горизонтального переноса генов антибиотикорезистентности в клетки бактерий, представленных в организме в составе нормальной или оппортунистической микрофлоры. Помимо этого, наличие генов антибиотикорезистентности значительно увеличивает размер ДНК-вектора, что приводит к снижению эффективности его проникновения в эукариотические клетки.
Необходимо отметить, что гены антибиотикорезистентности также вносят принципиальный вклад в способ получения ДНК-векторов. В случае наличия генов антибиотикорезистентности штаммы для наработки ДНК-векторов обычно культивируются в среде, содержащей селективный антибиотик, что создает риск наличия следовых количеств антибиотика в недостаточно очищенных препаратах ДНК-векторов. Таким образом, получение ДНК-векторов для генной терапии, в которых отсутствуют гены антибиотикорезистентности, связано с получением штаммов, обладающих такой отличительной особенностью как способность к стабильной амплификации целевых ДНК-векторов в среде без содержания антибиотиков.
Кроме того, Европейское Медицинское Агентство рекомендует избегать наличия в составе терапевтических плазмидных векторов регуляторных элементов для повышения экспрессии целевых генов (промоторов, энхансеров, посттрансляционных регуляторных элементов), являющихся нуклеотидными последовательностями геномов различных вирусов (Draft Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products,http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2015/05/WC500187020.pdf). Данные последовательности, хотя и могут увеличивать уровень экспрессии целевого трансгена, однако создают риск рекомбинации с генетическим материалом вирусов дикого типа и интеграции в геном эукариотической клетки. Более того, целесообразность гиперэкспрессии того или иного гена в целях терапии остается нерешенным вопросом.
Также, существенным моментом является размер терапевтического вектора. Известно, что современные плазмидные векторы зачастую перегружены нефункциональными участками, серьезно увеличивающими размер вектора (Mairhofer J, Grabherr R. // Mol Biotechnol. 2008.39(2):97-104). Например, ген устойчивости к ампициллину в векторах серии pBR322, как правило, состоит из не менее чем 1000 п.н., что составляет более 20% от размера самого вектора. При этом наблюдается обратная зависимость между размером вектора и его способностью проникать в эукариотические клетки – ДНК-векторы с небольшим размером эффективней проникаю в клетки человека и животных. Так, например, в серии экспериментов по трансфекции клеток HeLa ДНК-векторами с размером от 383 до 4548 п.н. было показано, что разница в эффективности проникновения может достигать двух порядков (отличаться в 100 раз) (Hornstein BD et al. // PLoS ONE. 2016;11(12): e0167537.).
Таким образом, при выборе ДНК-вектора в целях безопасности и наибольшей эффективности следует отдавать предпочтение тем конструкциям, в которых не содержатся гены устойчивости к антибиотикам, последовательности вирусного происхождения и размер которых позволяет эффективно проникать в эукариотические клетки. Штамм для получения такого ДНК-вектора в количествах, достаточных для целей генной терапии, должен обеспечивать возможность стабильной амплификации ДНК-вектора с использованием питательных сред, не содержащих антибиотики.
Примером использования рекомбинантных ДНК-векторов для генной терапии является способ получения рекомбинантного вектора для генетической иммунизации по патенту US 9550998 В2. Плазмидный вектор представляет собой суперскученный плазмидный ДНК-вектор и предназначен для экспрессии клонированных генов в клетках животных и человека. Вектор состоит из ориджина репликации, регуляторных элементов, включающих промотор и энхансер цитомегаловируса человека, регуляторные элементы из Т-лимфотропного вируса человека.
Накопление вектора проводят в специальном штамме E. coli без использования антибиотиков за счет антисенс-комплементации гена sacB, введенного в штамм посредством бактериофага. Недостатком данного изобретения является наличие в составе ДНК-вектора регуляторных элементов, представляющих собой последовательности вирусных геномов.
Прототипами настоящего изобретения в части использования генотерапевтических подходов для экспрессии Cas9 в эукариотических клетках являются следующие патенты.
В патенте US8795965B2 описана ДНК молекула, которая кодирует экспрессионную кассету, содержащую последовательность, кодирующую белок Cas9. Недостатком данного изобретения является неопределенность требований к наличию в составе ДНК молекулы регуляторных последовательностей вирусного происхождения, а также неопределенность способов получения данных молекул и их промышленной применимости.
В патенте CN103981216B описан плазмидный вектор, экспрессирующий ген Cas9. Недостатком данного изобретения является использование в составе вектора регуляторных элементов, обеспечивающих экспрессию гена Cas9 в клетках растений, но не млекопитающих, а также наличие в составе вектора генов устойчивости к антибиотикам.
В патенте US9914939B2 описан плазмидный вектор, экспрессирующий ген Cas9. Недостатком данного изобретения является неопределенность требований безопасности и технологичности в отношении используемого плазмидного вектора, в частности наличие/отсутствие в составе вектора последовательностей вирусного происхождения и генов устойчивости к антибиотикам.
Раскрытие изобретения
Задачей изобретения является конструирование генотерапевтического ДНК-вектора для гетерологичной экспрессии гена Cas9 в клетках человека и животных, сочетающего в себе следующие свойства:
I) Эффективность генотерапевтического ДНК-вектора для гетерологичной экспрессии целевого гена в эукариотических клетках.
II) Возможность безопасного применения для реализации различных методов геномного редактирования геномов человека, животного, в том числе при генетической терапии человека и животного за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов.
III) Возможность безопасного применения для реализации различных методов геномного редактирования геномов человека, животного, в том числе при генетической терапии человека и животного за счет отсутствия в составе генотерапевтического ДНК-вектора генов антибиотикорезистентности.
IV) Технологичность получения и возможность наработки генотерапевтического ДНК-вектора в промышленных масштабах.
Пункты II и III предусмотрены в данном техническом решении в соответствии с рекомендациями государственных регуляторов к лекарственным средствам для генной терапии, в частности, Европейского Агентства по лекарственным средствам касательно отказа от введения маркеров антибиотикорезистентности в разрабатываемые плазмидные векторы для генной терапии (Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011, EMA/CAT/GTWP/44236/2009 Committee for advanced therapies) и касательно отказа от введения в разрабатываемые плазмидные векторы для генной терапии элементов вирусных геномов (Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products / 23 March 2015, EMA/CAT/80183/2014, Committee for Advanced Therapies).
Задачей изобретения также является конструирование штаммов, несущих этот генотерапевтический ДНК-вектор, для наработки и производства в промышленных масштабах генотерапевтического ДНК-вектора.
Поставленная задача решается за счет того, что создан генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген Cas9 для гетерологичной экспрессии этого целевого гена в клетках человека и животных при реализации различных методов геномного редактирования, при этом генотерапевтический ДНК-вектор VTvaf17-Cas9 имеет нуклеотидную последовательность SEQ ID №1. При этом созданный генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген Cas9, отличающийся тем, что созданный генотерапевтический ДНК-вектор VTvaf17-Cas9 за счет ограниченного размера векторной части VTvaf17, не превышающей 3200 п.н., обладает способностью эффективно проникать в клетки человека и животных и экспрессировать клонированный в него целевой ген Cas9.
Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген Cas9, отличающийся тем, что в составе генотерапевтического ДНК-вектора отсутствуют нуклеотидные последовательности вирусного происхождения и отсутствуют гены антибиотикорезистентности, обеспечивая возможность его безопасного применения для реализации различных методов геномного редактирования геномов человека, животного, в том числе при генетической терапии человека и животного.
Создан также способ получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9, который заключается в том, что генотерапевтический ДНК-вектор VTvaf17-Cas9 получают следующим образом: кодирующую часть целевого гена Cas9 клонируют в ДНК-вектор VTvaf17 и получают генотерапевтический ДНК-вектор VTvaf17- Cas9, SEQ ID №1.
Способ применения созданного генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9 для гетерологичной экспрессии этого целевого гена в клетках человека и животных, заключающийся во введении генотерапевтического ДНК-вектора в клетки, органы и ткани человека или животного совместно с молекулами gRNA или генетическими конструкциями, обеспечивающими экспрессию gRNA, и/или во введении в органы и ткани человека или животного аутологичных клеток человека или животного, трансфицированных генотерапевтическим ДНК-вектором совместно с молекулами gRNA или генетическими конструкциями, обеспечивающими экспрессию gRNA или в сочетании обозначенных способов.
Способ получения штамма Escherichia coli SCS110-AF/VTvaf17- Cas9 заключается в электропорации компетентных клеток штамма Escherichia coli SCS110-AF созданным генотерапевтическим ДНК-вектором и последующей селекцией стабильных клонов штамма с использованием селективной среды.
Заявлен штамм Escherichia coli SCS110-AF/VTvaf17-Cas9, несущий генотерапевтический ДНК-вектор для его наработки с возможностью культивирования штамма без использования антибиотиков.
Способ производства в промышленных масштабах генотерапевтического ДНК-вектора заключается в масштабировании бактериальной культуры штамма до количеств, необходимых для наращивания бактериальной биомассы в промышленном ферментере, после чего биомассу используют для выделения фракции, содержащей целевой ДНК-продукт - генотерапевтический ДНК-вектор VTvaf17-Cas9, многостадийно фильтруют и очищают хроматографическими методами.
Изобретение поясняется чертежами, где:
На фиг.1
приведена схема генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9, который представляет собой кольцевую двуцепочечную молекулу ДНК, способную к автономной репликации в клетках бактерии Escherichia coli.
На фиг.1 приведена схема генотерапевтического ДНК-вектора VTvaf17- Cas9.
На схеме отмечены следующие структурные элементы вектора:
EF1a - промоторная область гена человеческого фактора элонгации EF1A с собственным энхансером, содержащимся в первом интроне гена. Служит для обеспечения высокого уровня транскрипции рекомбинантного гена в большинстве тканей человека;
Рамка считывания целевого гена, соответствующая кодирующей части гена Cas9;
hGH-TA – терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
ori – ориджин репликации, служащий для автономной репликации с однонуклеотидной заменой для повышения копийности плазмиды в клетках большинства штаммов Escherichia coli;
RNA-out – регуляторный элемент РНК-out транспозона Tn 10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Eshcerichia coli SCS 110.
Отмечены уникальные сайты рестрикции.
На фиг.2
показаны графики накопления ампликонов кДНК целевого гена, а именно, гена Cas9, в первичной культуре фибробластов кожи человека HDFa (ATCC PCS-201-01) до их трансфекции и через 48 часов после трансфекции этих клеток генотерапевтическим ДНК-вектором VTvaf17- Cas9 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.2 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 – кДНК гена Cas9 в культуре клеток HDFa до трансфекции ДНК-вектором VTvaf17- Cas9;
2 – кДНК гена Cas9 в культуре клеток HDFa после трансфекции ДНК-вектором VTvaf17- Cas9;
3 – кДНК гена B2M в культуре клеток HDFa до трансфекции ДНК-вектором VTvaf17- Cas9;
4 – кДНК гена B2M в культуре клеток HDFa после трансфекции ДНК-вектором VTvaf17- Cas9.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин), приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.3
показаны графики накопления ампликонов кДНК целевого гена, а именно гена Cas9, в первичной культуре кератиноцитов эпидермиса человека HEKa (ATCC PCS-200-011) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором VTvaf17-Cas9 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.3 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена Cas9 в первичной культуре кератиноцитов эпидермиса человека HEKa до трансфекции ДНК-вектором VTvaf17- Cas9;
2 – кДНК гена Cas9 первичной культуре кератиноцитов эпидермиса человека HEKa после трансфекции ДНК-вектором VTvaf17- Cas9;
3 - кДНК гена B2M в первичной культуре кератиноцитов эпидермиса человека HEKa до трансфекции ДНК-вектором VTvaf17- Cas9;
4 - кДНК гена B2M первичной культуре кератиноцитов эпидермиса человека HEKa после трансфекции ДНК-вектором VTvaf17- Cas9.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.4
показаны графики накопления ампликонов кДНК целевого гена, а именно гена Cas9, в первичной культуре клеток меланоцитов эпидермиса человека линии Primary Epidermal Melanocytes; Normal, Human, Adult (HEMa) (ATCC® PCS-200-013™) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором VTvaf17-Cas9 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.4 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена Cas9 в первичной культуре клеток меланоцитов эпидермиса человека HEMa до трансфекции ДНК-вектором VTvaf17-Cas9;
2 – кДНК гена Cas9 в первичной культуре клеток меланоцитов эпидермиса человека HEMa после трансфекции ДНК-вектором VTvaf17-Cas9;
3 - кДНК гена B2M в первичной культуре клеток меланоцитов эпидермиса человека HEMa до трансфекции ДНК-вектором VTvaf17-Cas9;
4 - кДНК гена B2M в первичной культуре клеток меланоцитов эпидермиса человека HEMa после трансфекции ДНК-вектором VTvaf17-Cas9.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.5
показана диаграмма концентрации белка Cas9 в клеточном лизате первичной культуры фибробластов кожи человека HDFa (ATCC PCS-201-01) после трансфекции этих клеток ДНК-вектором VTvaf17-Cas9 с целью оценки функциональной активности, то есть экспрессии на уровне белка, по изменению количества белка Cas9 в лизате клеток.
На фиг.5 отмечены следующие элементы:
культура А – культура клеток фибробластов кожи человека HDFa, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура клеток фибробластов кожи человека HDFa, трансфицированных ДНК-вектором VTvaf17;
культура C - культура клеток фибробластов кожи человека HDFa, трансфицированных ДНК-вектором VTvaf17-Cas9.
На фиг. 6
показана диаграмма концентрации белка Cas9 в лизате клеток первичной культуры кератиноцитов эпидермиса человека HEKa (ATCC PCS-200-01), после трансфекции этих клеток ДНК-вектором VTvaf17-Cas9 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9.
На фиг.6 отмечены следующие элементы:
культура А - первичная культура клеток кератиноцитов эпидермиса человека HEKa, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - первичная культура клеток кератиноцитов эпидермиса человека HEKa, трансфицированных ДНК-вектором VTvaf17;
культура C - первичная культура клеток кератиноцитов эпидермиса человека HEKa, трансфицированных ДНК-вектором VTvaf17-Cas9.
На фиг. 7
показана диаграмма концентрации белка Cas9 в лизате клеток первичной культуры меланоцитов эпидермиса человека линии Primary Epidermal Melanocytes; Normal, Human, Adult (HEMa) (ATCC® PCS-200-013™) после трансфекции этих клеток ДНК-вектором VTvaf17-Cas9 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9.
На фиг.7 отмечены следующие элементы:
культура А - культура клеток меланоцитов эпидермиса человека линии HEMa, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура клеток меланоцитов эпидермиса человека линии HEMa, трансфицированных ДНК-вектором VTvaf17;
культура C - культура клеток меланоцитов эпидермиса человека линии HEMa, трансфицированных ДНК-вектором VTvaf17-Cas9.
На фиг. 8
показана диаграмма концентрации белка Cas9 в лизате клеток яичника сирийского хомячка CHO-K1 (ATCC® CCL-61™) после трансфекции этих клеток генотерапевтическим ДНК-вектором VTvaf17-Cas9 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9.
На фиг.8 отмечены следующие элементы:
культура А - культура клеток яичника сирийского хомячка CHO-K1, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура клеток яичника сирийского хомячка CHO-K1, трансфицированных ДНК-вектором VTvaf17;
культура C - культура клеток яичника сирийского хомячка CHO-K1, трансфицированных ДНК-вектором VTvaf17-Cas9.
На фиг. 9
показана диаграмма концентрации белка Cas9 в лизате культуры мононуклеарных клеток переферической крови человека (PBMC) после трансфекции этих клеток генотерапевтическим ДНК-вектором VTvaf17-Cas9 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9.
На фиг.9 отмечены следующие элементы:
культура А - культура клеток PBMC человека, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура клеток PBMC человека, трансфицированных ДНК-вектором VTvaf17;
культура C - культура клеток PBMC человека, трансфицированных ДНК-вектором VTvaf17-Cas9.
На фиг.10
показана диаграмма концентраций белка Cas9 в биоптатах кожи трех крыс в предварительно эпилированной зоне у крыс линии Wistar после введения в кожу культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9 с целью демонстрации способа применения путем введения аутологичных клеток, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9.
На фиг.10 отмечены следующие элементы:
K1I – биоптат кожи крысы К1 в зоне введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9;
K1II – биоптат кожи крысы в зоне введения аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим ген Cas9;
K1III – биоптат кожи крысы из интактного участка;
К2I – биоптат кожи крысы К2 в зоне введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9;
K2II – биоптат кожи крысы K2 в зоне введения аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим ген Cas9;
K2III – биоптат кожи крысы K2 из интактного участка;
К3I – биоптат кожи крысы К3 в зоне введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9;
K3II – биоптат кожи крысы К3 в зоне введения аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим ген Cas9;
K3III – биоптат кожи крысы К3 из интактного участка.
На фиг. 11
показана диаграмма снижения количества флуоресцирующих клеток 293/GFP Cell Line (Cell Biolabs, Кат. AKR-200) после сочетанной трансфекции этих клеток ДНК-вектором VTvaf17-Cas9, несущим ген Cas9 и GFP-targeting guide RNA for CRISPR (Genaxxon bioscience, Кат. P2008.0010) с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка и способности этого белка осуществлять направленную эндонуклеазную активность с участием специфической gRNA, что приводит к редактированию последовательности ДНК.
На фиг.11 отмечены следующие элементы:
культура А – клетки 293/GFP трансфицированные водой с последующей трансфекцией GFP-targeting guide RNA for CRISPR;
культура В – клетки 293/GFP трансфицированные ДНК-вектором VTvaf17, не содержащим кДНК гена Cas9, с последующей трансфекцией GFP-targeting guide RNA for CRISPR;
культура С – клетки 293/GFP трансфицированные ДНК-вектором VTvaf17-Cas9, несущим ген Cas9, с последующей трансфекцией GFP-targeting guide RNA for CRISPR;
культура D – клетки 293/GFP трансфицированные ДНК-вектором VTvaf17-Cas9, несущим ген Cas9, с последующей трансфекцией водой.
На фиг. 12
Показаны результаты секвенирования ПЦР фрагментов нескольких клонов клеток линии легочной аденокарциномы MCF-7 (ATCC® HTB-22™), в которых произошло редактирование генома после трансфекции VTvaf17-Cas9, несущим ген Cas9, с вектором gRNA_Cloning Vector, несущим последовательности gRNA гена TLR9.
На фигуре изображен участок ДНК, соотвествующей области двухцепочечного разрыва, вносимого sgRNA TLR_1 и TLR_2, где Tlr12 – консенсусная последовательность; WT – последовательность фрагмента гена TLR9 не подвергшегося геномному редактированию или с восстановленной последовательностью; al1, al2, G2 различные аллели у гетерозиготных клонов (на схеме представлены оба аллеля).
Реализация изобретения
На основе ДНК-вектора VTvaf17 размером 3165 п.н. создан генотерапевтический ДНК-вектор, несущий целевой ген Cas9, предназначенный для гетерологичной экспрессии этого целевого гена в клетках человека и животных. При этом способ получения генотерапевтического ДНК-вектора, несущего целевой ген, заключается в том, что в полилинкер генотерапевтического ДНК-вектора VTvaf17 клонируют белок-кодирующую последовательность целевого гена Cas9 (кодирует эндонуклеазу Cas9). Известно, что способность ДНК-векторов проникать в эукариотические клетки обусловлена, главным образом, размером вектора. При этом ДНК-векторы с наименьшим размером обладают более высокой проникающей способностью. Таким образом, предпочтительным является отсутствие в составе вектора элементов, которые не несут функциональной нагрузки, но при этом увеличивают размер ДНК-вектора. Данные особенности ДНК-векторов были учтены при получении генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9, путем отсутствия в составе вектора крупных нефункциональных последовательностей и генов антибиотикорезистентности, что позволило, помимо технологических преимуществ и преимуществ в плане безопасности применения, значительно уменьшить размер полученного генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9. Таким образом, способность проникать в эукариотические клетки полученного генотерапевтического ДНК-вектора обусловлена его небольшими размерами.
Генотерапевтический ДНК-вектор VTvaf17-Cas9 получали следующим образом: кодирующую часть целевого гена Cas9 клонировали в генотерапевтический ДНК-вектор VTvaf17 и получали генотерапевтический ДНК-вектор VTvaf17- Cas9, SEQ ID №1. Кодирующую часть гена Cas9 размером 4221 п.н. получали путем ферментативного синтеза из химически синтезированых олигонуклеотидов. Расщепление продукта синтеза специфическими эндонуклеазами рестрикции проводили с учетом оптимальной процедуры дальнейшего клонирования, причем клонирование в генотерапевтический ДНК-вектор VTvaf17 проводили по сайтам рестрикции BamHI, EcoRI, расположенными в полилинкере вектора VTvaf17. Выбор сайтов рестрикции проводили таким образом, чтобы клонированный фрагмент попадал в рамку считывания экспрессионной кассеты вектора VTvaf17, при этом белок-кодирующая последовательность не содержала сайты рестрикции для выбранных эндонуклеаз. При этом специалистам в данной области техники понятно, что методическая реализация получения генотерапевтического ДНК-вектора VTvaf17-Cas9 может варьировать в рамках выбора известных методов молекулярного клонирования генов, при этом эти способы подпадают под объем настоящего изобретения. Так, например, могут быть использованы различные последовательности олигонуклеотидов для амплификации гена Cas9, различные эндонуклеазы рестрикции или такие лабораторные техники как безлигазное клонирование генов.
Генотерапевтический ДНК-вектор VTvaf17-Cas9 обладает нуклеотидной последовательностью SEQ ID №1. При этом специалистам в данной области техники известно свойство вырожденности генетического кода, из которого следует, что под объем настоящего изобретения также подпадают варианты нуклеотидных последовательностей, отличающихся инсерцией, делецией или заменой нуклеотидов, которые не приводят к изменению полипептидной последовательности, кодируемой целевым геном, и/или не приводят к потере функциональной активности регуляторных элементов вектора VTvaf17. При этом специалистам в данной области техники известно явление генетического полиморфизма, из которого следует, что под объем настоящего изобретения также подпадают варианты нуклеотидных последовательностей гена Cas9, которые при этом кодируют различные варианты аминокислотных последовательностей белка Cas9, не отличающихся от приведенных по своей функциональной активности при физиологических условиях.
Способность проникать в эукариотические клетки и функциональную активность, то есть способность экспрессировать целевой ген, полученного генотерапевтического ДНК-вектора VTvaf17- Cas9 подтверждают путем введения в эукариотические клетки полученного вектора и последующим анализом экспрессии специфической мРНК и/или белкового продукта целевого гена. Наличие специфической мРНК в клетках, в которые был введен генотерапевтический ДНК-вектор VTvaf17- Cas9 свидетельствует как о способности полученного вектора проникать в эукариотические клетки, так и о его способности экспрессировать мРНК целевого гена Cas9. При этом, как известно специалистам в данной области техники, наличие мРНК гена является обязательным условием, но не доказательством трансляции белка, кодируемого целевым геном. Поэтому для подтверждения свойства генотерапевтического ДНК-вектора VTvaf17- Cas9 экспрессировать целевой ген на уровне белка в эукариотических клетках, в которые был введен генотерапевтический ДНК-вектор, проводят анализ концентрации белка, кодируемого целевым геном, с использованием иммунологических методов. Наличие белка Cas9 подтверждает эффективность экспрессии целевого гена в эукариотических клетках с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9. Таким образом, для подтверждения эффективности экспрессии созданного генотерапевтического ДНК-вектора VTvaf17- Cas9, несущего целевой ген, а именно, ген Cas9, использовали следующие методы:
А) ПЦР в реальном времени - изменение накопления ампликонов кДНК целевого гена в лизате клеток человека и животного, после трансфекции различных клеточных линий человека и животного генотерапевтическим ДНК-вектором;
B) Иммуноферментный анализ - изменение количественного уровня целевого белка в лизате клеток человека, животного, после трансфекции различных клеточных линий человека и животных генотерапевтическим ДНК-вектором;
С) Иммуноферментный анализ - изменение количественного уровня целевого белка в супернатанте биоптатов тканей животного, после введения в эти ткани аутологичных клеток этого животного, трансфицированных генотерапевтическим ДНК-вектором;
D) Измерение методом проточной флуорометрии экспрессии гена GFP в клетках, подвергшихся процедуре геномного редактирования, после сочетанной трансфекции этих клеток генотерапевтическим ДНК-вектором VTvaf17- Cas9 и gRNA, что привело к инактивации гена GFP и отсутствию экспрессии флуоресцентного белка в клетках или ее значительному снижению;
Е) Секвенирование участка ДНК клеток человека, животного, подвергшегося процедуре геномного редактирования, после сочетанной трансфекции этих клеток генотерапевтическим ДНК-вектором VTvaf17-Cas9 и вектором, экспрессирующим gRNA, направленным на последовательность гена TLR9.
Для подтверждения реализуемости способа применения созданного генотерапевтического ДНК-вектора VTvaf17- Cas9, несущего целевой ген, а именно, ген Cas9 выполняли:
А) трансфекцию генотерапевтическим ДНК-вектором различных клеточных линий человека, животных;
B) сочетанную трансфекцию генотерапевтическим ДНК-вектором и gRNA различных клеточных линий млекопитающих;
С) сочетанную трансфекцию генотерапевтическим ДНК-вектором и вектором, кодирующим gRNA, различных клеточных линий млекопитающих;
D) введение в ткани животного аутологичных клеток, трансфицированных генотерапевтическими ДНК-вектором;
E) демонстрацию изменения экспрессии редактируемого гена в клеточной линии, подвергшейся процедуре геномного редактирования;
F) демонстрацию изменения последовательности участка редактируемого гена в клеточной линии, подвергшейся геномному редактированию.
Указанные способы применения характеризуются отсутствием потенциальных рисков для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов, и за счет отсутствия в составе генотерапевтического ДНК-вектора генов устойчивости к антибиотикам, что подтверждается отсутствием участков, гомологичных вирусным геномам и генам антибиотикорезистентности в нуклеотидной последовательности генотерапевтического ДНК-вектора VTvaf17- Cas9 (SEQ ID №1).
Как известно специалистам в данной области техники, гены антибиотикорезистентности в составе генотерапевтических ДНК-векторов используются с целью получения этих векторов в препаративных количествах путем наращивания бактериальной биомассы в питательной среде, содержащей селективный антибиотик. В рамках настоящего изобретения в целях возможности безопасного применения генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9, использование селективных питательных сред, содержащих антибиотик, не представляется возможным. В качестве технологического решения для получения генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9, для возможности масштабирования до промышленных масштабов получения генотерапевтического вектора предлагается способ получения штамма для наработки указанных генотерапевтических векторов на основе бактерии Escherichia coli SCS110-AF. Способ получения штамма Escherichia coli SCS110-AF/VTvaf17-Cas9 заключается в получении компетентных клеток штамма Escherichia coli SCS110-AF с введением в эти клетки генотерапевтического ДНК-вектора VTvaf17-Cas9 с помощью методов трансформации (электропорации), общеизвестных специалистам в данной области техники. Полученный штамм Escherichia coli SCS110-AF/VTvaf17-Cas9 используется для наработки генотерапевтического ДНК-вектора VTvaf17-Cas9 с возможностью использования сред без содержания антибиотиков.
Для подтверждения получения штамма Escherichia coli SCS110-AF/VTvaf17-Cas9 проводили трансформацию, селекцию и последующее наращивание с выделением плазмидной ДНК.
Для подтверждения технологичности получения и возможности масштабирования до промышленного производства генотерапевтического ДНК-вектора VTvaf17- Cas9, несущего целевой ген, а именно, ген Cas9, выполняли ферментацию в промышленном масштабе штамма Escherichia coli SCS110-AF/VTvaf17- Cas9, который содержит генотерапевтический ДНК-вектор VTvaf17, несущий целевой ген Cas9.
Способ масштабирования получения бактериальной массы до промышленных масштабов для выделения генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, Cas9, заключается в том, что затравочную культуру штамма Escherichia coli SCS110-AF/VTvaf17- Cas9 инкубируют в объеме питательной среды без содержания антибиотика обеспечивающим подходящую динамику накопления биомассы, по достижению достаточного количества биомассы в логарифмической фазе роста, бактериальную культуру переносят в промышленный ферментер, после чего растят до достижения стационарной фазы роста, затем выделяют фракцию, содержащую целевой ДНК-продукт - генотерапевтический ДНК-вектор VTvaf17- Cas9 многостадийно фильтруют и очищают хроматографическими методами. При этом специалистам в данной области техники понятно, что условия культивирования штаммов, состав питательных сред (за исключением содержания антибиотиков), используемое оборудование, методы очистки ДНК могут варьировать в рамках стандартных операционных процедур в зависимости от отдельно взятой производственной линии, но известные подходы к масштабированию, промышленному получению и очистке ДНК-векторов с использованием штамма Escherichia coli SCS110-AF/VTvaf17-Cas9 подпадают под объем настоящего изобретения.
Описанное раскрытие изобретения подтверждается примерами реализации настоящего изобретения.
Изобретение поясняется следующими примерами.
Пример 1.
Получение генотерапевтического ДНК-вектора VTvaf17- Cas9, несущего целевой ген, а именно, гена Cas9.
Генотерапевтический ДНК-вектор VTvaf17-Cas9 конструировали клонированием кодирующей части гена Cas9 размером 4221 п.н. в ДНК-вектор VTvaf17 размером 3165 п.н. по сайтам рестрикции BamHI и EcoRI. Кодирующую часть гена Cas9 размером 4221 п.н. получали путем ферментативного синтеза из химически синтезированых олигонуклеотидов.
Генотерапевтический ДНК-вектор VTvaf17 конструировали объединением шести фрагментов ДНК, полученных из разных источников:
(а) ориджин репликации получали путем ПЦР-амплификации участка коммерческой плазмиды pBR322 с внесением точечной мутации;
(б) промоторный регион EF1a получали путем ПЦР-амплификации участка геномной ДНК человека;
(в) терминатор транскрипции hGH-TA получали путем ПЦР-амплификации участка геномной ДНК человека;
(г) регуляторный участок транспозона Tn10 РНК-out получали путем синтеза из олигонуклеотидов;
(д) ген устойчивости к канамицину получали путем ПЦР-амплификации участка коммерческой плазмиды pET-28 человека;
(е) полилинкер получали отжигом двух синтетических олигонуклеотидов.
ПЦР-амплификацию проводили с использованием коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США) в соответствии с инструкцией производителя. Фрагменты имеют перекрывающиеся области для возможности их объединения с последующей ПЦР-амплификацией. Объединяли фрагменты (а) и (б) с использованием олигонуклеотидов Ori-F и EF1-R, а также фрагменты (в), (г) и (д) с использованием олигонуклеотидов hGH-F и Kan-R. Затем полученные участки объединяли путем рестрикции с последующим лигированием по сайтам BamHI и NcoI. В результате получали плазмиду, пока еще не содержащую полилинкер. Для его введения проводили расщепление плазмиды по сайтам BamHI и EcoRI, и лигирование с фрагментом (е). Таким образом, получали вектор размером 4182 п.н., несущий ген устойчивости к канамицину, который фланкирован сайтами рестрикции SpeI. Далее этот участок выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор VTvaf17 размером 3165 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков.
Расщепление продукта амплификации кодирующей части гена Cas9 и ДНК-вектора VTvaf17 проводили эндонуклеазами рестрикции BamHI и EcoRI (New England Biolabs, США).
В результате получали ДНК-вектор VTvaf17-Cas9 размером 7351 п.н. с нуклеотидной последовательностью SEQ ID №1 и общей структурой изображенной на фиг.1.
Пример 2.
Подтверждение способности генотерапевтического ДНК-вектора VTvaf17-Cas9, несущего целевой ген, а именно, ген Cas9, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена Cas9, в первичной культуре фибробластов кожи человека HDFa (ATCC PCS-201-01) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17-Cas9, несущим ген Cas9 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Для оценки изменения накопления мРНК целевого гена Cas9, использовалась первичная культура фибробластов кожи человека HDFa. Клеточную культуру HDFa выращивали в стандартных условиях (37 °С, 5% СО2) с использованием питательной среды Fibroblast Growth Kit–Serum-Free (ATCC® PCS-201-040). В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5Ч104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором VTvaf17-Cas9, экспрессирующим ген Cas9 человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) согласно рекомендациям производителя. В пробирке №1 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора ДНК-вектора VTvaf17-Cas9 (концентрация 500 нг/мкл) и 1 мкл реагента Р3000. Аккуратно перемешивали легким встряхиванием. В пробирке №2 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора Lipofectamin 3000. Аккуратно перемешивали легким встряхиванием. Добавляли содержимое пробирки №1 к содержимому пробирки №2, инкубировали 5 мин при комнатной температуре. Полученный раствор по каплям добавляли к клеткам в объеме 40 мкл.
В качестве контроля использовали клетки HDFa, трансфицированные генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена (кДНК гена Cas9 до и после трансфекции генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора VTvaf17 для трансфекции проводили как описано выше.
Суммарную РНК из клеток HDFa выделяли с использованием Trizol Reagent (Invitrogen, США) согласно рекомендациям производителя. В лунку с клетками добавляли 1 мл Trizol Reagent и гомогенизировали с последующим прогреванием в течении 5 мин при 65 °С. Далее образец центрифугировали при 14000g в течении 10 мин и снова прогревали в течении 10 мин при 650С. Далее добавляли 200 мкл хлороформа, плавно перемешивали и центрифугировали при 14000g в течении 10 мин. Затем отбирали водную фазу, добавляли к ней 1/10 объема 3М ацетата натрия, рН 5.2 и равный объем изопропилового спирта. Инкубировали образец при -20 °С в течении 10 мин с последующим центрифугированием при 14000g в течении 10 мин. Осадок промывали 1 мл 70% этилового спирта, высушивали на воздухе и растворяли в 10 мкл воды, свободной от РНКаз. Определение уровня экспрессии мРНК гена Cas9 после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени. Для получения и амплификации кДНК, специфичной для гена Cas9 человека, использовали олигонуклеотиды Cas9_SF и Cas9_SR (перечень последовательностей (1), (2)).
Длина продукта амплификации - 275 п.н.
Реакцию обратной транскрипции и ПЦР-амплификацию проводили с помощью набора реагентов SYBR GreenQuantitect RT–PCR Kit (Qiagen, США) для ПЦР в режиме реального времени. Реакцию проводили в объеме 20 мкл, содержащих: 25 мкл QuantiTect SYBR Green RT-PCR MasterMix, 2,5 мМ хлорида магния, по 0,5 мкМ каждого праймера, 5 мкл РНК. Реакцию осуществляли на амплификаторе CFX96 (Bio-Rad, США) при следующих условиях: 1 цикл обратной транскрипции при 42°С - 30 минут, денатурация 98°С - 15 мин, затем 40 циклов, включающих денатурацию 94°С - 15 сек, отжиг праймеров 60°C - 30 сек и элонгацию 72°С - 30 сек. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов Cas9 и B2M. В качестве отрицательного контроля использовали деионизированную воду. Количество динамику накопления ампликонов кДНК генов Cas9 и B2M оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 2.
Из фигуры 2 следует, что в результате трансфекции первичной культуры клеток фибробластов человека HDFa генотерапевтическим ДНК-вектором VTvaf17-Cas9, установлено наличие мРНК гена Cas9, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген Cas9 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9 для гетерологичной экспрессии гена Cas9 в эукариотических клетках.
Пример 3.
Подтверждение способности генотерапевтического ДНК-вектора VTvaf17-Cas9, несущего целевой ген, а именно, ген Cas9, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена Cas9, в первичной культуре кератиноцитов эпидермиса человека HEKa (ATCC PCS-200-011) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17-Cas9, несущим ген Cas9 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Первичную культуру клеток кератиноцитов эпидермиса человека HEKa выращивали в среде Keratinocyte Growth Kit (ATCC® PCS-200-040™) в стандартных условиях (37°С, 5% СО2). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5Ч104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором VTvaf17-Cas9, экспрессирующим ген Cas9 человека, проводили как описано в примере 2. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве контроля использовали культуру клеток HEKa, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим целевой ген (кДНК гена Cas9 до и после трансфекции генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили как описано в примере 2.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов Cas9 и B2M. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов Cas9 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 3.
Из фигуры 3 следует, что в результате трансфекции культуры клеток HEKa генотерапевтическим ДНК-вектором VTvaf17-Cas9, установлено наличие мРНК гена Cas9, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген Cas9 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9 для гетерологичной экспрессии гена Cas9 в эукариотических клетках.
Пример 4.
Подтверждение способности генотерапевтического ДНК-вектора VTvaf17-Cas9, несущего целевой ген, а именно, ген Cas9, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена Cas9 в культуре клеток меланоцитов эпидермиса человека линии Primary Epidermal Melanocytes; Normal, Human, Adult (HEMa) (ATCC® PCS-200-013™) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17-Cas9, несущим ген Cas9 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Культуру меланоцитов эпидермиса человека линии Primary Epidermal Melanocytes выращивали в среде, приготовленной с использованием Adult Melanocyte Growth Kit (ATCC® PCS-200-042™) в стандартных условиях (37°С, 5% СО2). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5Ч104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором VTvaf17-Cas9, экспрессирующим ген Cas9 человека, проводили как описано в примере 2. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве контроля использовали культуру клеток меланоцитов эпидермиса человека HEMa, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим целевой ген (кДНК гена Cas9 до и после трансфекции генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили как описано в примере 2.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов Cas9 и B2M. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов Cas9 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 4.
Из фигуры 4 следует, что в результате трансфекции культуры клеток меланоцитов эпидермиса человека линии HEMa генотерапевтическим ДНК-вектором VTvaf17-Cas9 установлено наличие мРНК гена Cas9, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген Cas9 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9 для гетерологичной экспрессии гена Cas9 в эукариотических клетках.
Пример 5.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9, несущего ген Cas9, для гетерологичной экспрессии белка Cas9 в клетках млекопитающих.
Оценивали изменение количества белка Cas9 в лизате клеток первичной культуры фибробластов кожи человека HDFa (ATCC PCS-201-01) после трансфекции этих клеток ДНК-вектором VTvaf17-Cas9, несущим ген Cas9 человека. Клетки выращивали как описано в примере 2.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена Cas9 (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-Cas9, несущий ген Cas9 человека (С). Трансфекцию клеток HDFa проводили как описано в примере 2.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка Cas9 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Cas9 (CRISPR Associated Protein 9) ELISA Kit (Cell Biolabs Inc, Кат. PRB-5079) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка Cas9, входящим в состав набора. Чувствительность метода составляла не менее 1,5 нг/мл, диапазон измерения - от 1.56 нг/мл до 100 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 5.
Из фигуры 5 следует, что в результате трансфекции первичной культуры клеток HDFa человека генотерапевтическим ДНК-вектором VTvaf17-Cas9 установлено наличие белка Cas9 по сравнению с отсутствием такового в контрольных образцах, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген Cas9 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9 для гетерологичной экспрессии Cas9 в эукариотических клетках.
Пример 6.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9, несущего ген Cas9, для гетерологичной экспрессии белка Cas9 в клетках млекопитающих.
Оценивали изменение количества белка Cas9 в лизате клеток первичной культуры кератиноцитов эпидермиса человека HEKa (ATCC PCS-200-011) после трансфекции этих клеток ДНК-вектором VTvaf17-Cas9, несущим ген Cas9 человека. Клетки выращивали как описано в примере 3.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена Cas9 (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-Cas9, несущий ген Cas9 человека (С). Приготовление комплекса ДНК-дендример и трансфекцию клеток проводили согласно методике производителя.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка Cas9 проводили методом твердофазного иммуноферментного анализа с использованием набора Cas9 (CRISPR Associated Protein 9) ELISA Kit (Cell Biolabs Inc, Кат. PRB-5079) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка Cas9, входящим в состав набора. Чувствительность метода составляла не менее 1,5 нг/мл, диапазон измерения - от 1.56 нг/мл до 100 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 6.
Из фигуры 6 следует, что в результате трансфекции первичной культуры клеток кератиноцитов эпидермиса человека HEKa человека генотерапевтическим ДНК-вектором VTvaf17-Cas9 установлено наличие белка Cas9 по сравнению с отсутствием такового в контрольных образцах, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген Cas9 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9 для гетерологичной экспрессии Cas9 в эукариотических клетках.
Пример 7.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9, несущего ген Cas9, для гетерологичной экспрессии белка Cas9 в клетках млекопитающих.
Оценивали изменение количества белка Cas9 в лизате клеток первичной культуры меланоцитов эпидермиса человека Primary Epidermal Melanocytes; Normal, Human, Adult (HEMa) (ATCC® PCS-200-013™) после трансфекции этих клеток ДНК-вектором VTvaf17-Cas9, несущим ген Cas9 человека. Клетки выращивали как описано в примере 4.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена Cas9 (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-Cas9, несущий ген Cas9 человека (С). Приготовление комплекса ДНК-дендример и трансфекцию клеток HEMa проводили согласно методике производителя.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка Cas9 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Cas9 (CRISPR Associated Protein 9) ELISA Kit (Cell Biolabs Inc, Кат. PRB-5079) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка Cas9, входящим в состав набора. Чувствительность метода составляла не менее 1,5 нг/мл, диапазон измерения - от 1.56 нг/мл до 100 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 7.
Из фигуры 7 следует, что в результате трансфекции первичной культуры клеток HEMa человека генотерапевтическим ДНК-вектором VTvaf17-Cas9 установлено наличие белка Cas9 по сравнению с отсутствием такового в контрольных образцах, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген Cas9 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9 для гетерологичной экспрессии Cas9 в эукариотических клетках.
Пример 8.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9, несущего ген Cas9, для гетерологичной экспрессии белка Cas9 в клетках животных.
Оценивали изменение количества белка Cas9 в лизате клеток яичника сирийского хомячка CHO-K1 (ATCC® CCL-61™) после трансфекции этих клеток ДНК-вектором VTvaf17-Cas9, несущим ген Cas9 человека. Клетки выращивали в среде F-12K Medium (Kaighn's Modification of Ham's F-12 Medium) (ATCC® 30-2004™) с добавление 10% сыворотки Fetal Bovine Serum (FBS) (ATCC® 30-2020™) в стандартных условиях.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена Cas9 (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-Cas9, несущий ген Cas9 человека (С). Приготовление комплекса ДНК-дендример и трансфекцию клеток проводили согласно методике производителя.
После трансфекции к 0,5 мл клеточной суспензии добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка Cas9 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Cas9 (CRISPR Associated Protein 9) ELISA Kit (Cell Biolabs Inc, Кат. PRB-5079) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка Cas9, входящим в состав набора. Чувствительность метода составляла не менее 1,5 нг/мл, диапазон измерения - от 1.56 нг/мл до 100 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 8.
Из фигуры 8 следует, что в результате трансфекции культуры клеток CHO-K1 генотерапевтическим ДНК-вектором VTvaf17-Cas9 установлено наличие белка Cas9 по сравнению с отсутствием такового в контрольных образцах, что подтверждает способность вектора проникать в эукариотические клетки животных и экспрессировать ген Cas9 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9 для гетерологичной экспрессии Cas9 в эукариотических клетках животных.
Пример 9.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9, несущего ген Cas9, для гетерологичной экспрессии белка Cas9 в клетках человека.
Оценивали изменение количества белка Cas9 в лизате культуры первичных мононуклеарных клеток переферической крови человека (PBMC) после трансфекции этих клеток ДНК-вектором VTvaf17-Cas9, несущим ген Cas9 человека. Клетки PBMC выделяли из 10 мл венозной крови человека путем разделения фракций в градиенте фиколла 1,119 (ПанЭко, Р051-1) Клетки выращивали в среде RPMI-1640 (ПанЭко, С310п) в стандартных условиях (37С, 5% СО2).
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена Cas9 (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-Cas9, несущий ген Cas9 человека (С). Приготовление комплекса ДНК-дендример и трансфекцию клеток проводили согласно методике производителя.
Через 24 часа после трансфекции 1 мл культуры клеток осаждали центрифугированием, к осадку клеток добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка Cas9 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Cas9 (CRISPR Associated Protein 9) ELISA Kit (Cell Biolabs Inc, Кат. PRB-5079) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка Cas9, входящим в состав набора. Чувствительность метода составляла не менее 1,5 нг/мл, диапазон измерения - от 1.56 нг/мл до 100 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 9.
Из фигуры 9 следует, что в результате трансфекции культуры клеток PBMC генотерапевтическим ДНК-вектором VTvaf17-Cas9 установлено наличие белка Cas9 по сравнению с отсутствием такового в контрольных образцах, что подтверждает способность вектора проникать в клетки человека и экспрессировать ген Cas9 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9 для гетерологичной экспрессии Cas9 в эукариотических клетках млекопитающих.
Пример 10.
Подтверждение эффективности генотерапевтического ДНК-вектора VTvaf17-Cas9, несущего ген Cas9 и реализуемости способа его применения для повышения уровня экспрессии белка Cas9 в тканях животного путем введения аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9.
Для подтверждения эффективности генотерапевтического ДНК-вектора VTvaf17-Cas9, несущего ген Cas9 и реализуемости способа его применения оценивали изменения количества белка Cas9 в коже крыс при введении в кожу крыс культуры аутологичных фибробластов этого животного, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9.
Трем крыса линии Wistar вводили в кожу соответствующую культуру аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9, несущим ген Cas9, и параллельно вводили плацебо, представляющее собой культуру аутологичных фибробластов животного, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим ген Cas9.
Первичную культуру фибробластов крысы выделяли из биоптатов кожи животного. Используя устройство для взятия биопсии кожи Epitheasy 3.5 (Medax SRL, Италия) брали образец биоптата кожи массой около11 мг. Культивирование первичной культуры клеток осуществляли при 37°С в атмосфере, содержащей 5 % СО2 в среде DMEM с 10 % фетальной телячьей сывороткой и ампициллином 100 Ед/мл. Пересев культуры и смену культуральной среды осуществляли каждые 2 дня. Общая продолжительность роста культуры не превышала 25-30 дней. Из культуры клеток отбирали аликвоту, содержащую 5Ч104 клеток. Культуру фибробластов крыс трансфицировали генотерапевтическим ДНК-вектором VTvaf17-Cas9, несущим ген Cas9 или плацебо – вектор VTvaf17, не несущий целевой ген Cas9.
Трансфекцию осуществляли с помощью катионного полимера полиэтиленимина JETPEI (Polyplus Transfection, Франция), согласно инструкции фирмы-производителя. Клетки культивировали 72 часа и затем вводили животным. Введение животному культуры аутогенных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9 и плацебо, представляющее собой культуру аутологичных фибробластов крысы, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, осуществляли туннельным методом в область предварительно эпилированной кожи иглой 27G на глубину около 1 мм. Концентрация модифицированных аутологичных фибробластов в вводимой суспензии составляла примерно 5 млн клеток на 1 мл суспензии, количество вводимых клеток не превышало 10 млн. Очаги введения культуры аутогенных фибробластов располагались на расстоянии 3 - 5 см друг от друга.
Биопсийные образцы отбирали на 4-е сутки после введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9, несущим целевой ген, а именно, ген Cas9 и плацебо. Взятие биопсии осуществляли из участков кожи животных в зоне введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9, несущим целевой ген, а именно, ген Cas9 (I), культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим целевой ген Cas9 (плацебо) (II), а также из участков интактной кожи (III), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка как это описано в примерах 5 - 9.
Диаграммы, полученные в результате анализа, представлены на фиг. 10.
Из фигуры 10 следует, что в коже крыс в области введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9, несущим ген Cas9, установлено наличие белка Cas9 в сравнении с отсутствием такового в контрольных образцах: в области введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим ген Cas9 (плацебо) и в образце из интактной зоны, что свидетельствует об эффективности генотерапевтического ДНК-вектора VTvaf17-Cas9 и подтверждает реализуемость способа его применения для повышения уровня экспрессии Cas9 в тканях млекопитающих, в частности при введении аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-Cas9.
Пример 11.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9, несущего ген Cas9, для редактирования генома в клетках млекопитающих.
Оценивали снижение флуоресценции клеток за счет нарушения экспрессии гена GFP в клетках 293/GFP Cell Line (Cell Biolabs, Кат. AKR-200) после сочетанной трансфекции этих клеток ДНК-вектором VTvaf17-Cas9, несущим ген Cas9 человека и GFP-targeting guide RNA for CRISPR (Genaxxon bioscience, Кат. P2008.0010). Клетки выращивали в среде DMEM в стандартных условиях.
Для трансфекции клеток ДНК-вектором VTvaf17-Cas9, несущим ген Cas9, использовали Lipofectamine 3000 (ThermoFisher Scientific, США) согласно рекомендациям производителя как это описано в примере 2. Спустя 24 часа после трансфекции ДНК-вектором VTvaf17-Cas9, несущим ген Cas9, клетки трансфицировали GFP-targeting guide RNA for CRISPR. Для трансфекции GFP-targeting guide RNA for CRISPR использовали реагент CRISPRfect E transfection reagent (Genaxxon bioscience, Кат. P2002.0035) согласно инструкции производителя.
В качестве экспериментальных образцов использовали трансфекцию водой с последующей трансфекцией GFP-targeting guide RNA for CRISPR (А), трансфекцию ДНК-вектором VTvaf17, не содержащим кДНК гена Cas9, с последующей трансфекцией GFP-targeting guide RNA for CRISPR (В), трансфекцию ДНК-вектором VTvaf17-Cas9, несущим ген Cas9, с последующей трансфекцией GFP-targeting guide RNA for CRISPR (С), трансфекцию ДНК-вектором VTvaf17-Cas9, несущим ген Cas9, с последующей трансфекцией водой (D).
Спустя 48 часов после второй трансфекции удаляли культуральную среду, клетки ресуспендировали в физиологическом растворе и использовали для оценки количества клеток, экспрессирующих флуоресцентный белок GFP методом проточной флуорометрии на приборе Beckman Coulter's Cytomics FC 500 (Beckman Coulter's, США). Полученные в результате анализа, представлены на фиг. 11.
Из фигуры 11 следует, что в результате трансфекции первичной культуры клеток 293/GFP генотерапевтическим ДНК-вектором VTvaf17-Cas9 с последующей трансфекцией GFP-targeting guide RNA for CRISPR установлено снижение (около 80%) количества клеток, экспрессирующих GFP по сравнению с отсутствием снижения в контрольных образцах, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать функционально активную эндонуклеазу Cas9, которая с помощью GFP-targeting guide RNA осуществляет направленное редактирование гена GFP, что приводит к подавлению экспрессии флуоресцентного белка GFP. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9 для выполнения таргетного редактирования генома эукариотических клеток.
Пример 12.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9, несущего ген Cas9, для редактирования генома в клетках млекопитающих.
Выполняли скрининг клонов клеток линии MCF7 (ATCC® HTB-22™) с целью выявления клонов, в которых произошло направленное редактирование генома, после трансфекции этих клеток смесью ДНК-вектора VTvaf17-Cas9, несущего ген Cas9, с gRNA_Cloning Vector, несущим олигонуклеотиды к участкам экзона гена TLR9.
В качестве sgRNA подобраны специфические олигонуклеотиды, соответствующие 20 нуклеотидным участкам экзона гена TLR9 (Толл-подобный рецептор 9). Подбор sgRNA проводили с помощью сервиса CRISPR DESIGN (http://crispr.mit.edu/). Два синтезированых олигонуклеотида tlrg4f и tlrg4r (список последовательностей (3) и (4)) смешивали в эквимолярных количествах в буфере T4 ДНК лигазы (Thermo Scientific, США), образцы прогревали при 94°С в течение 2 мин, после чего медленно охлаждали до комнатной температуры для образования дуплексов. Далее, олигонуклеотидные дуплексы клонировали в состав вектора gRNA_Cloning Vector (AddGene, #41824). Плазмиды с клонированными олигонуклеотидами наращивали и очищали в препаративных количествах набором Plasmid DNA Purification Kit (Qiagen, США).
Клетки MCF-7 культивировали с использованием среды EMEM Eagle's Minimum Essential Medium (EMEM) (ATCC® 30-2003™) стандартных условиях (37С, 5% CO2). Трансфекцию проводили с использованием набора реагентов Lipofectamine 3000 (Thermoscientific, США) согласно инструкции фирмы производителя. Для трансфекции использовали эквимолярную смесь ДНК-вектора VTvaf17-Cas9, несущего ген Cas9, с gRNA_Cloning Vector, несущим олигонуклеотиды к участкам экзона гена TLR9. В качестве контроля использовали воду или gRNA_Cloning Vector, несущий олигонуклеотиды к участкам экзона гена TLR9, или ДНК-вектор VTvaf17-Cas9, несущий ген Cas9.
Затем клетки с помощью FACS-сортера рассевали в лунки 96-луночного планшента. Для анализа клонов, выросших после сортинга, клетки, растущие в 96-луночных планшетах, промывали PBS, лизировали в 50 мкл реагента ДНК-экспресс (Литех, Россия), после чего проводили пробоподготовку согласно рекомендациям производителя. Полученные образцы использовали в качестве матрицы для амплификации методом ПЦР участка локуса, содержащего участки распознавания sgRNAs в гене TLR9. Полученные ПЦР-фрагменты секвенировали для анализа на генетическом анализаторе ABI Prism 3730xl Genetic Analyser (Applied Biosystems, США). В результате было идентифицировано четыре клона, содержащих изменения нуклеотидных последовательностей гена TLR9, которые явились результатом направленного геномного редактирования. В контрольных образцах не было выявлено клонов, содержащих какие-либо изменения в последовательностях гена TLR9.
Полученные в результате анализа данные, представлены на фиг. 12.
Из фигуры 12 следует, что в результате трансфекции клеточной линии MCF7 смесью ДНК-вектора VTvaf17-Cas9, несущего ген Cas9, с gRNA_Cloning Vector, несущим олигонуклеотиды к участкам экзона гена TLR9, и последующим скринингом клонов клеток было выявлено несколько клонов, в которых произошло направленное редактирование генома, что подтверждает способность вектора VTvaf17-Cas9 проникать в эукариотические клетки и экспрессировать функционально активную эндонуклеазу Cas9, которая с помощью gRNA способна вносить мутацию в целевой ген, например в ген TLR9, что показано путем секвенирования таргетной последовательности. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-Cas9 для выполнения таргетного редактирования генома эукариотических клеток.
Пример 13.
Штамм Escherichia coli SCS110-AF/VTvaf17-Cas9, несущий генотерапевтический ДНК-вектор, и способ его получения.
Конструирование штамма для наработки в промышленных масштабах генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, Cas9, а именно штамма Escherichia coli SCS110-AF/VTvaf17-Cas9, несущего генотерапевтический ДНК-вектор VTvaf17-Cas9 для его наработки с возможностью селекции без использования антибиотиков заключается в получении электрокомпетентных клеток штамма Escherichia coli SCS110-AF с проведением электропорации этих клеток генотерапевтическим ДНК-вектором VTvaf17-Cas9, после чего клетки высеваются на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола. При этом штамм Escherichia coli SCS110-AF для наработки генотерапевтического ДНК-вектора VTvaf17 или генотерапевтических ДНК-векторов на его основе с возможностью положительной селекции без использования антибиотиков получали путем конструирования линейного фрагмента ДНК, содержащего регуляторный элемент RNA-in транспозона Tn10 для селекции без применения антибиотиков размером 64 п.н., ген левансахаразы sacB, продукт которого обеспечивает селекцию на сахарозо-содержащей среде размером 1422 п.н., ген устойчивости к хлорамфениколу catR, необходимый для отбора клонов штамма, в которых прошла гомологичная рекомбинация размером 763 п.н. и две гомологичные последовательности, обеспечивающие процесс гомологичной рекомбинации в области гена recA с одновременной его инактивацией размером 329 п.н. и 233 п.н., после чего проводили трансформацию клеток Escherichia coli путем электропорации и отбирали клоны, выжившие на среде, содержащей 10 мкг/мл хлорамфеникола.
Полученный штамм для наработки был внесен в коллекции Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ с присвоением регистрационных номеров:
штамм Escherichia coli SCS110-AF/VTvaf17-Cas9 – регистрационный № ВКПМ: В- , дата депонирования:
Пример 14.
Способ масштабирования получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9 до промышленного масштаба.
Для подтверждения технологичности получения и возможности производства в промышленных масштабах генотерапевтического ДНК-вектора VTvaf17-Cas9 (SEQ ID №1) проводили масштабную ферментацию штамма Escherichia coli SCS110-AF/VTvaf17-Cas9, который содержит генотерапевтический ДНК-вектор VTvaf17, несущий целевой ген, а именно Cas9. Штамм Escherichia coli SCS110-AF/VTvaf17-Cas9, получали на базе штамма Escherichia coli SCS110-AF (Cell and Gene Therapy LLC, Великобритания) по примеру 13 путем электропорации компетентных клеток этого штамма генотерапевтическим ДНК-вектором VTvaf17-Cas9, несущим целевой ген, а именно, Cas9 с последующим высевом трансформированных клеток на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы и отбором отдельных клонов.
Ферментацию полученного штамма Escherichia coli SCS110-AF/VTvaf17-Cas9, несущего генотерапевтический ДНК-вектор VTvaf17-Cas9 проводили в ферментере объемом 10 л с последующим выделением генотерапевтического ДНК-вектора VTvaf17-Cas9.
Для ферментации штамма Escherichia coli SCS110-AF/VTvaf17-Cas9 готовили среду, содержащую на 10 л: 100 г триптон, 50 г дрожжевой экстракт (Becton Dickinson, США), доводили водой до 8800 мл и автоклавировали при 121 0С 20 мин, затем добавляли 1200 мл 50% (вес/объем) сахарозы. Далее засевали в колбу затравочную культуру штамма Escherichia coli SCS110-AF/VTvaf17-Cas9 в объеме 100 мл. Инкубировали в шейкере-инкубаторе 16 ч при 30°С. Переносили затравочную культуру в ферментер Techfors S (Infors HT, Швейцария), растили до достижения стационарной фазы. Контроль осуществляли измерением оптической плотности культуры при длине волны 600 нм. Клетки осаждали центрифугированием 30 мин при 5000-10000 g. Супернатант удаляли, клеточную массу ресуспендировали в 10% по объему фосфатно-солевого буфера. Повторно центрифугировали 30 мин при 5000-10000g. Супернатант удаляли, к клеточной массе добавляли раствор 20 мМ TrisCl, 1 мМ ЭДТА, 200 г/л сахароза, рН 8,0 в объеме 1000 мл, тщательно перемешивали до образования гомогенной суспензии. Добавляли раствор яичного лизоцима до конечной концентрации 100мкг/мл. Инкубировали 20 мин на льду при бережном перемешивании. Далее, добавляли 2500 мл раствора 0,2 М NaOH, 10 г/л додецилсульфат натрия, инкубировали 10 мин на льду при бережном перемешивании, затем добавляли 3500 мл раствора 3M ацетат натрия, 2М уксусная кислота, рН 5-5,5, инкубировали 10 мин на льду при бережном перемешивании. Полученный образец центрифугировали 20-30 мин при 15000 g или более. Раствор аккуратно декантировали, от остатков осадка избавлялись фильтрацией через грубый фильтр (фильтровальную бумагу). Добавляли РНКазу А (Sigma, США) до конечной концентрации 20 нг/мл, инкубировали ночь 16 ч при комнатной температуре. Раствор центрифугировали 20-30 мин при 15000g, затем фильтровали через мембранный фильтр с порами 0,45 мкм (Millipore, США). Далее проводили ультрафильтрацию через мембрану размером отсечения 100кДа (Millipore, США) и разбавляли до начального объема буфером 25 мМ TrisCl, pH 7.0. Операцию повторяли три – четыре раза. Наносили раствор, на колонку с 250 мл сорбента DEAE sepharose HP (GE, США), уравновешенную раствором 25 мМ TrisCl, pH 7.0. После нанесения образца колонку промывали тремя объемами этого же раствора, а затем проводили элюцию генотерапевтического ДНК-вектора VTvaf17-Cas9 линейным градиентом от раствора 25 мМ TrisCl, pH 7.0 до раствора 25 мМ TrisCl, pH 7.0, 1М NaCl в объеме пяти объемов колонки. Контроль элюции осуществляли по оптической плотности сходящего раствора при 260 нм. Хроматографические фракции, содержащие генотерапевтический ДНК-вектор VTvaf17-Cas9, объединяли и проводили гель-фильтрацию на сорбенте Superdex 200 (GE, США). Колонку уравновешивали фосфатно-солевым буфером. Контроль элюции осуществляли по оптической плотности сходящего раствора при 260 нм, фракции анализировали электрофорезом в агарозном геле. Фракции, содержащие генотерапевтический ДНК-вектор VTvaf17-Cas9 объединяли и хранили при -20°С. Для оценки воспроизводимости техпроцесса обозначенные технологические операции повторяли пятикратно.
Воспроизводимость техпроцесса и количественные характеристики выхода конечного продукта подтверждают технологичность получения и возможность производства в промышленных масштабах генотерапевтического ДНК-вектора VTvaf17-Cas9.
Таким образом, созданный генотерапевтический ДНК-вектор с целевым геном может быть использован для введения в клетки человека, животных и организм млекопитающих, обеспечивая гетерологичную экспрессию эндонуклеазы Cas9, что может использоваться с целью редактирования последовательности генома человека и животных в присутствии gRNA заданной специфичности.
Задача, поставленная в данном изобретении, а именно: конструирование генотерапевтического ДНК-вектора для гетерологичной экспрессии гена Cas9 в клетках человека и животных, сочетающего в себе следующие свойства:
I) Эффективность генотерапевтического ДНК-вектора для гетерологичной экспрессии целевого гена в эукариотических клетках;
II) Возможность безопасного применения для реализации различных методов геномного редактирования геномов человека и животного, в том числе при генетической терапии человека и животного за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов;
III) Возможность безопасного применения для реализации различных методов геномного редактирования геномов человека и животного, в том числе при генетической терапии человека и животного за счет отсутствия в составе генотерапевтического ДНК-вектора генов антибиотикорезистентности;
IV) Технологичность получения и возможность наработки генотерапевтического ДНК-вектора в промышленных масштабах,
а также задача по конструированию штамма, несущего этот генотерапевтический ДНК-вектор для производства этого генотерапевтического ДНК-вектора решена, что подтверждается примерами:
для п. I – пример 1, 2, 3, 4, 5; 6; 7; 8; 9; 10; 11;
для п. II и п. III – пример 1, 11, 12;
для п. IV – пример 1, 13, 14.
Промышленная применимость:
Все представленные выше примеры подтверждают промышленную применимость предлагаемого генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9 для гетерологичной экспрессии гена Cas9 в клетках человека и животных, штамма Escherichia coli SCS110-AF/VTvaf17-Cas9, несущего генотерапевтический ДНК-вектор, способа получения генотерапевтического ДНК-вектора, способа производства в промышленных масштабах генотерапевтического ДНК-вектора.
Перечень сокращений
VTvaf17 – вектор генотерапевтический, не содержащий последовательностей вирусных геномов и маркеров антибиотико-резистентности (vector therapeutic virus-antibiotic-free)
gRNA – guided RNA
PBMC – мононуклеарные клетки переферической крови
ДНК - дезоксирибонуклеиновая кислота
кДНК - комплементарная дезоксирибонуклеиновая кислота
РНК - рибонуклеиновая кислота
мРНК - матричная рибонуклеиновая кислота
п.н. – пар нуклеотидов
ПЦР - полимеразная цепная реакция
мл – миллилитр, мкл – микролитр
куб. мм – кубический миллиметр
л - литр
мкг - микрограмм
мг - миллиграмм
г - грамм
мкМ - микромоль
мМ – миллимоль
мин – минута
сек - секунда
об/мин - обороты в минуту
нм - нанометр
см - сантиметр
мВт - милливатт
о.е. ф-относительная единица флуоресценции
РВС – фосфатно-солевой буфер
Список литературы
1. Dowdy SF. Overcoming cellular barriers for RNA therapeutics. Nat Biotechnol. 2017 Mar;35(3):222-229.
2. Draft Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products, http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2015/05/WC500187020.pdf
3. Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal Products EMA/CAT/80183/2014
4. Hornstein BD, Roman D, Arévalo-Soliz LM, Engevik MA, Zechiedrich L. Effects of Circular DNA Length on Transfection Efficiency by Electroporation into HeLa Cells. Ceña V, ed. PLoS ONE. 2016;11(12):e0167537.
5. Hussain W, Mahmood T, Hussain J, Ali N, Shah T, Qayyum S, Khan I. CRISPR/Cas system: A game changing genome editing technology, to treat human genetic diseases. Gene. 2019 Feb 15;685:70-75. doi: 10.1016/j.gene.2018.10.072. Epub 2018 Oct 26. Review. PubMed PMID: 30393194.
6. Li L, Hu S, Chen X. Non-viral delivery systems for CRISPR/Cas9-based genome editing: Challenges and opportunities. Biomaterials. 2018 Jul;171:207-218.
7. Li L, Petrovsky N. Molecular mechanisms for enhanced DNA vaccine immunogenicity. Expert Rev Vaccines. 2016;15(3):313-29
8. Lukashev AN, Zamyatnin AA Jr. Viral Vectors for Gene Therapy: Current State and Clinical Perspectives. Biochemistry (Mosc). 2016 Jul;81(7):700-8.
9. Maginnis MS. Virus-Receptor Interactions: The Key to Cellular Invasion. J Mol Biol. 2018 Aug 17;430(17):2590-2611.
10. Mairhofer J, Grabherr R. Rational vector design for efficient non-viral gene delivery: challenges facing the use of plasmid DNA. Mol Biotechnol. 2008.39(2):97-104
11. Memi F, Ntokou A, Papangeli I. CRISPR/Cas9 gene-editing: Research technologies, clinical applications and ethical considerations. Semin Perinatol. 2018 Dec;42(8):487-500. doi: 10.1053/j.semperi.2018.09.003. Epub 2018 Oct 2. Review. PubMed PMID: 30482590.
12. Mougiakos I, Mohanraju P, Bosma EF, Vrouwe V, Finger Bou M, Naduthodi MIS, Gussak A, Brinkman RBL, van Kranenburg R, van der Oost J. Characterizing a thermostable Cas9 for bacterial genome editing and silencing. Nat Commun. 2017 Nov 21;8(1):1647.
13. Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011 EMA/CAT/GTWP/44236/2009 Committee for advanced therapies
14. Salsman J, Dellaire G. Precision genome editing in the CRISPR era. Biochem Cell Biol. 2017 Apr;95(2):187-201.
15. Sapranauskas R, Gasiunas G, Fremaux C, Barrangou R, Horvath P, Siksnys V. The Streptococcus thermophilus CRISPR/Cas system provides immunity in Escherichia coli. Nucleic Acids Res. 2011 Nov;39(21):9275-82.
16. Wilson LOW, O'Brien AR, Bauer DC. The Current State and Future of CRISPR-Cas9 gRNA Design Tools. Front Pharmacol. 2018 Jul 12;9:749.
17. Молекулярная биология, 2011, том 45, № 1, с. 44-55
Перечень сокращений
VTvaf17 – вектор генотерапевтический, не содержащий последовательностей вирусных геномов и маркеров антибиотико-резистентности (vector therapeutic virus-antibiotic-free)
gRNA – guided RNA
PBMC – мононуклеарные клетки переферической крови
ДНК - дезоксирибонуклеиновая кислота
кДНК - комплементарная дезоксирибонуклеиновая кислота
РНК - рибонуклеиновая кислота
мРНК - матричная рибонуклеиновая кислота
п.н. – пар нуклеотидов
ПЦР - полимеразная цепная реакция
мл – миллилитр, мкл – микролитр
куб. мм – кубический миллиметр
л - литр
мкг - микрограмм
мг - миллиграмм
г - грамм
мкМ - микромоль
мМ – миллимоль
мин – минута
сек - секунда
об/мин - обороты в минуту
нм - нанометр
см - сантиметр
мВт - милливатт
о.е. ф-относительная единица флуоресценции
РВС – фосфатно-солевой буфер.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Селл энд Джин Терапи Лтд (CELL and GENE THERAPY Ltd), Общество с ограниченной ответственностью «Прорывные Инновационные Технологии»,
<120> Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген Cas9 для гетерологичной экспрессии этого целевого гена в клетках человека и животных при реализации различных методов геномного редактирования, способ получения и применения генотерапевтического ДНК-вектора, штамм Escherichia coli SCS110-AF/VTvaf17-Cas9, несущий генотерапевтический ДНК-вектор, способ его получения, способ производства в промышленных масштабах генотерапевтического ДНК-вектора.
<160> 1
<170> BiSSAP 1.3.6
<210> 1
<211> 7351
<212> DNA
<213> Homo Sapiens
<400> 1
cgtgaggctc cggtgcccgt cagtgggcag agcgcacatc gcccacagtc cccgagaagt 60
tggggggagg ggtcggcaat tgaaccggtg cctagagaaa gtggcgcggg gtaaactggg 120
aaagtgatgt cgtgtactgg ctccgccttt ttcccgaggg tgggggagaa ccgtatataa 180
gtgcagtagt cgccgtgaac gttctttttc gcaacgggtt tgccgccaga acacaggtaa 240
gtgccgtgtg tggttcccgc gggcctggcc tctttacggg ttatggccct tgcgtgcctt 300
gaattacttc cacgcccctg gctgcagtac gtgattcttg atcccgagct tcgggttgga 360
agtgggtggg agagttcgag gccttgcgct taaggagccc cttcgcctcg tgcttgagtt 420
gaggcctggc ttgggcgctg gggccgccgc gtgcgaatct ggtggcacct tcgcgcctgt 480
ctcgctgctt tcgataagtc tctagccatt taaaattttt gatgacctgc tgcgacgctt 540
tttttctggc aagatagtct tgtaaatgcg ggccaagatc tgcacactgg tatttcggtt 600
tttggggccg cgggcggcga cggggcccgt gcgtcccagc gcacatgttc ggcgaggcgg 660
ggcctgcgag cgcggccacc gagaatcgga cgggggtagt ctcaagctgg ccggcctgct 720
ctggtgcctg gcctcgcgcc gccgtgtatc gccccgccct gggcggcaag gctggcccgg 780
tcggcaccag ttgcgtgagc ggaaagatgg ccgcttcccg gccctgctgc agggagctca 840
aaatggagga cgcggcgctc gggagagcgg gcgggtgagt cacccacaca aaggaaaagg 900
gcctttccgt cctcagccgt cgcttcatgt gactccacgg agtaccgggc gccgtccagg 960
cacctcgatt agttctcgag cttttggagt acgtcgtctt taggttgggg ggaggggttt 1020
tatgcgatgg agtttcccca cactgagtgg gtggagactg aagttaggcc agcttggcac 1080
ttgatgtaat tctccttgga atttgccctt tttgagtttg gatcttggtt cattctcaag 1140
cctcagacag tggttcaaag tttttttctt ccatttcagg tgtcgtgaaa actaccccta 1200
aaagccaagg atccgccacc atggccccaa agaagaagcg gaaggtcggt atccacggag 1260
tcccagcagc cgacaagaag tacagcatcg gcctggacat cggcaccaac tctgtgggct 1320
gggccgtgat caccgacgag tacaaggtgc ccagcaagaa attcaaggtg ctgggcaaca 1380
ccgaccggca cagcatcaag aagaacctga tcggagccct gctgttcgac agcggcgaaa 1440
cagccgaggc cacccggctg aagagaaccg ccagaagaag atacaccaga cggaagaacc 1500
ggatctgcta tctgcaagag atcttcagca acgagatggc caaggtggac gacagcttct 1560
tccacagact ggaagagtcc ttcctggtgg aagaggataa gaagcacgag cggcacccca 1620
tcttcggcaa catcgtggac gaggtggcct accacgagaa gtaccccacc atctaccacc 1680
tgagaaagaa actggtggac agcaccgaca aggccgacct gcggctgatc tatctggccc 1740
tggcccacat gatcaagttc cggggccact tcctgatcga gggcgacctg aaccccgaca 1800
acagcgacgt ggacaagctg ttcatccagc tggtgcagac ctacaaccag ctgttcgagg 1860
aaaaccccat caacgccagc ggcgtggacg ccaaggccat cctgtctgcc agactgagca 1920
agagcagacg gctggaaaat ctgatcgccc agctgcccgg cgagaagaag aatggcctgt 1980
tcggaaacct gattgccctg agcctgggcc tgacccccaa cttcaagagc aacttcgacc 2040
tggccgagga tgccaaactg cagctgagca aggacaccta cgacgacgac ctggacaacc 2100
tgctggccca gatcggcgac cagtacgccg acctgtttct ggccgccaag aacctgtccg 2160
acgccatcct gctgagcgac atcctgagag tgaacaccga gatcaccaag gcccccctga 2220
gcgcctctat gatcaagaga tacgacgagc accaccagga cctgaccctg ctgaaagctc 2280
tcgtgcggca gcagctgcct gagaagtaca aagagatttt cttcgaccag agcaagaacg 2340
gctacgccgg ctacattgac ggcggagcca gccaggaaga gttctacaag ttcatcaagc 2400
ccatcctgga aaagatggac ggcaccgagg aactgctcgt gaagctgaac agagaggacc 2460
tgctgcggaa gcagcggacc ttcgacaacg gcagcatccc ccaccagatc cacctgggag 2520
agctgcacgc cattctgcgg cggcaggaag atttttaccc attcctgaag gacaaccggg 2580
aaaagatcga gaagatcctg accttccgca tcccctacta cgtgggccct ctggccaggg 2640
gaaacagcag attcgcctgg atgaccagaa agagcgagga aaccatcacc ccctggaact 2700
tcgaggaagt ggtggacaag ggcgcttccg cccagagctt catcgagcgg atgaccaact 2760
tcgataagaa cctgcccaac gagaaggtgc tgcccaagca cagcctgctg tacgagtact 2820
tcaccgtgta taacgagctg accaaagtga aatacgtgac cgagggaatg agaaagcccg 2880
ccttcctgag cggcgagcag aaaaaggcca tcgtggacct gctgttcaag accaaccgga 2940
aagtgaccgt gaagcagctg aaagaggact acttcaagaa aatcgagtgc ttcgactccg 3000
tggaaatctc cggcgtggaa gatcggttca acgcctccct gggcacatac cacgatctgc 3060
tgaaaattat caaggacaag gacttcctgg acaatgagga aaacgaggac attctggaag 3120
atatcgtgct gaccctgaca ctgtttgagg acagagagat gatcgaggaa cggctgaaaa 3180
cctatgccca cctgttcgac gacaaagtga tgaagcagct gaagcggcgg agatacaccg 3240
gctggggcag gctgagccgg aagctgatca acggcatccg ggacaagcag tccggcaaga 3300
caatcctgga tttcctgaag tccgacggct tcgccaacag aaacttcatg cagctgatcc 3360
acgacgacag cctgaccttt aaagaggaca tccagaaagc ccaggtgtcc ggccagggcg 3420
atagcctgca cgagcacatt gccaatctgg ccggcagccc cgccattaag aagggcatcc 3480
tgcagacagt gaaggtggtg gacgagctcg tgaaagtgat gggccggcac aagcccgaga 3540
acatcgtgat cgaaatggcc agagagaacc agaccaccca gaagggacag aagaacagcc 3600
gcgagagaat gaagcggatc gaagagggca tcaaagagct gggcagccag atcctgaaag 3660
aacaccccgt ggaaaacacc cagctgcaga acgagaagct gtacctgtac tacctgcaga 3720
atgggcggga tatgtacgtg gaccaggaac tggacatcaa ccggctgtcc gactacgatg 3780
tggaccatat cgtgcctcag agctttctga aggacgactc catcgacaac aaggtgctga 3840
ccagaagcga caagaaccgg ggcaagagcg acaacgtgcc ctccgaagag gtcgtgaaga 3900
agatgaagaa ctactggcgg cagctgctga acgccaagct gattacccag agaaagttcg 3960
acaatctgac caaggccgag agaggcggcc tgagcgaact ggataaggcc ggcttcatca 4020
agagacagct ggtggaaacc cggcagatca caaagcacgt ggcacagatc ctggactccc 4080
ggatgaacac taagtacgac gagaatgaca agctgatccg ggaagtgaaa gtgatcaccc 4140
tgaagtccaa gctggtgtcc gatttccgga aggatttcca gttttacaaa gtgcgcgaga 4200
tcaacaacta ccaccacgcc cacgacgcct acctgaacgc cgtcgtggga accgccctga 4260
tcaaaaagta ccctaagctg gaaagcgagt tcgtgtacgg cgactacaag gtgtacgacg 4320
tgcggaagat gatcgccaag agcgagcagg aaatcggcaa ggctaccgcc aagtacttct 4380
tctacagcaa catcatgaac tttttcaaga ccgagattac cctggccaac ggcgagatcc 4440
ggaagcggcc tctgatcgag acaaacggcg aaaccgggga gatcgtgtgg gataagggcc 4500
gggattttgc caccgtgcgg aaagtgctga gcatgcccca agtgaatatc gtgaaaaaga 4560
ccgaggtgca gacaggcggc ttcagcaaag agtctatcct gcccaagagg aacagcgata 4620
agctgatcgc cagaaagaag gactgggacc ctaagaagta cggcggcttc gacagcccca 4680
ccgtggccta ttctgtgctg gtggtggcca aagtggaaaa gggcaagtcc aagaaactga 4740
agagtgtgaa agagctgctg gggatcacca tcatggaaag aagcagcttc gagaagaatc 4800
ccatcgactt tctggaagcc aagggctaca aagaagtgaa aaaggacctg atcatcaagc 4860
tgcctaagta ctccctgttc gagctggaaa acggccggaa gagaatgctg gcctctgccg 4920
gcgaactgca gaagggaaac gaactggccc tgccctccaa atatgtgaac ttcctgtacc 4980
tggccagcca ctatgagaag ctgaagggct cccccgagga taatgagcag aaacagctgt 5040
ttgtggaaca gcacaagcac tacctggacg agatcatcga gcagatcagc gagttctcca 5100
agagagtgat cctggccgac gctaatctgg acaaagtgct gtccgcctac aacaagcacc 5160
gggataagcc catcagagag caggccgaga atatcatcca cctgtttacc ctgaccaatc 5220
tgggagcccc tgccgccttc aagtactttg acaccaccat cgaccggaag aggtacacca 5280
gcaccaaaga ggtgctggac gccaccctga tccaccagag catcaccggc ctgtacgaga 5340
cacggatcga cctgtctcag ctgggaggcg acaaaaggcc ggcggccacg aaaaaggccg 5400
gccaggcaaa aaagaaaaag taagaattcc ctgtgacccc tccccagtgc ctctcctggc 5460
cctggaagtt gccactccag tgcccaccag ccttgtccta ataaaattaa gttgcatcat 5520
tttgtctgac taggtgtcct tctataatat tatggggtgg aggggggtgg tatggagcaa 5580
ggggcaagtt gggaagacaa cctgtagggc ctgcggggtc tattgggaac caagctggag 5640
tgcagtggca caatcttggc tcactgcaat ctccgcctcc tgggttcaag cgattctcct 5700
gcctcagcct cccgagttgt tgggattcca ggcatgcatg accaggctca gctaattttt 5760
gtttttttgg tagagacggg gtttcaccat attggccagg ctggtctcca actcctaatc 5820
tcaggtgatc tacccacctt ggcctcccaa attgctggga ttacaggcgt gaaccactgc 5880
tcccttccct gtccttacgc gtagaattgg taaagagagt cgtgtaaaat atcgagttcg 5940
cacatcttgt tgtctgatta ttgatttttg gcgaaaccat ttgatcatat gacaagatgt 6000
gtatctacct taacttaatg attttgataa aaatcattaa ctagtccatg gctgcctcgc 6060
gcgtttcggt gatgacggtg aaaacctctg acacatgcag ctcccggaga cggtcacagc 6120
ttgtctgtaa gcggatgccg ggagcagaca agcccgtcag ggcgcgtcag cgggtgttgg 6180
cgggtgtcgg ggcgcagcca tgacccagtc acgtagcgat agcggagtgt atactggctt 6240
aactatgcgg catcagagca gattgtactg agagtgcacc atatgcggtg tgaaataccg 6300
cacagatgcg taaggagaaa ataccgcatc aggcgctctt ccgcttcctc gctcactgac 6360
tcgctgcgct cggtcgttcg gctgcggcga gcggtatcag ctcactcaaa ggcggtaata 6420
cggttatcca cagaatcagg ggataacgca ggaaagaaca tgtgagcaaa aggccagcaa 6480
aaggccagga accgtaaaaa ggccgcgttg ctggcgtttt tccataggct ccgcccccct 6540
gacgagcatc acaaaaatcg acgctcaagt cagaggtggc gaaacccgac aggactataa 6600
agataccagg cgtttccccc tggaagctcc ctcgtgcgct ctcctgttcc gaccctgccg 6660
cttaccggat acctgtccgc ctttctccct tcgggaagcg tggcgctttc tcatagctca 6720
cgctgtaggt atctcagttc ggtgtaggtc gttcgctcca agctgggctg tgtgcacgaa 6780
ccccccgttc agcccgaccg ctgcgcctta tccggtaact atcgtcttga gtccaacccg 6840
gtaagacacg acttatcgcc actggcagca gccactggta acaggattag cagagcgagg 6900
tatgtaggcg gtgctacaga gttcttgaag tggtggccta actacggcta cactagaaga 6960
acagtatttg gtatctgcgc tctgctgaag ccagttacct tcggaaaaag agttggtagc 7020
tcttgatccg gcaaacaaac caccgctggt agcggtggtt tttttgtttg caagcagcag 7080
attacgcgca gaaaaaaagg atctcaagaa gatcctttga tcttttctac ggggtctgac 7140
gctcagtgga acgaaaactc acgttaaggg attttggtca tgagattatc aaaaaggatc 7200
ttcacctaga tccttttaaa ttaaaaatga agttttaaat caatctaaag tatatatgag 7260
taaacttggt ctgacagtta ccaatgctta atcagtgagg cacctatctc agcgatctgt 7320
ctatttcgtt catccatagt tgcctgactc c 7351
<---
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ОЛИГОНУКЛЕОТИДОВ
(1) Cas9_SF CATCGAGCAGATCAGCGAGT
(2) Cas9_FR CGATCCGTGTCTCGTACAGG
(3) tlrg4f CACCGACTGGGTGTACAACGAGCTT
(4) tlrg4r AAACAAGCTCGTTGTACACCCAGTC
<---

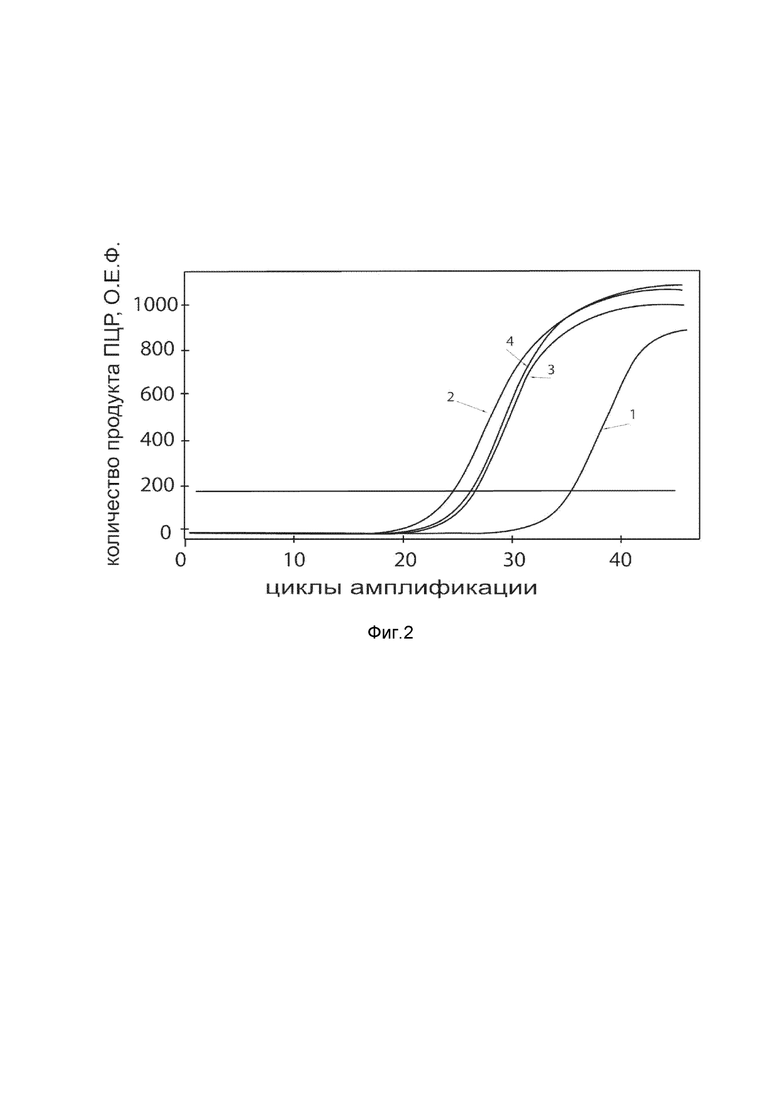


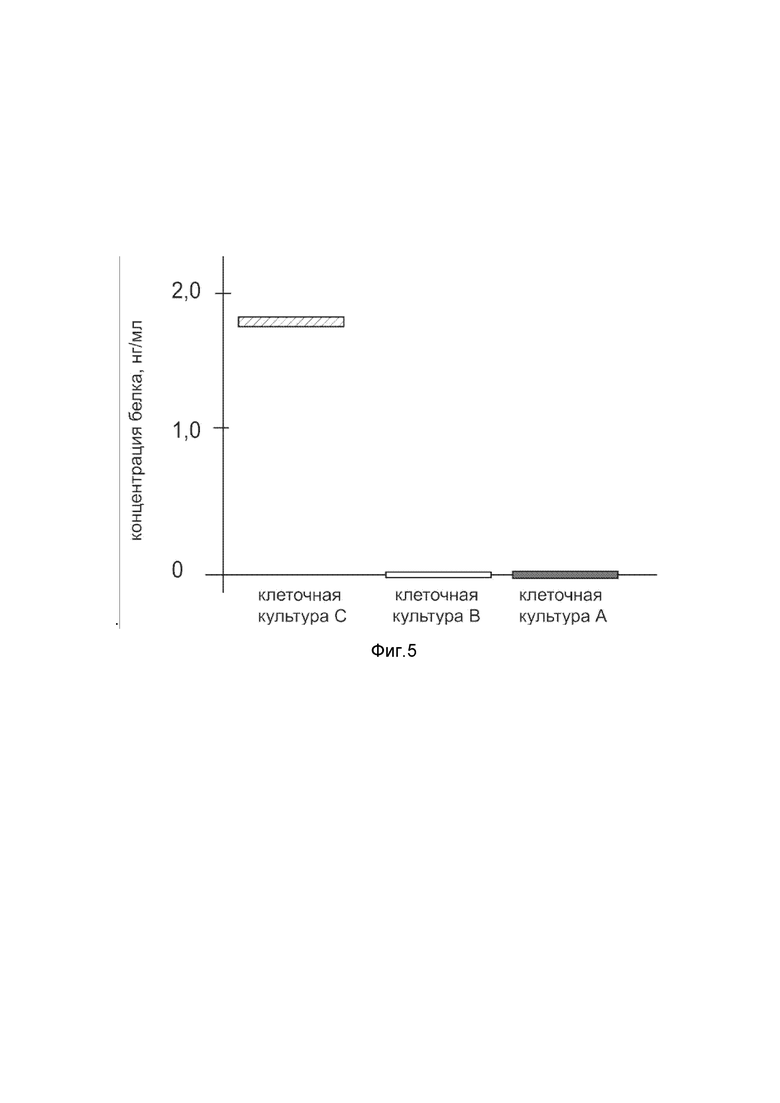
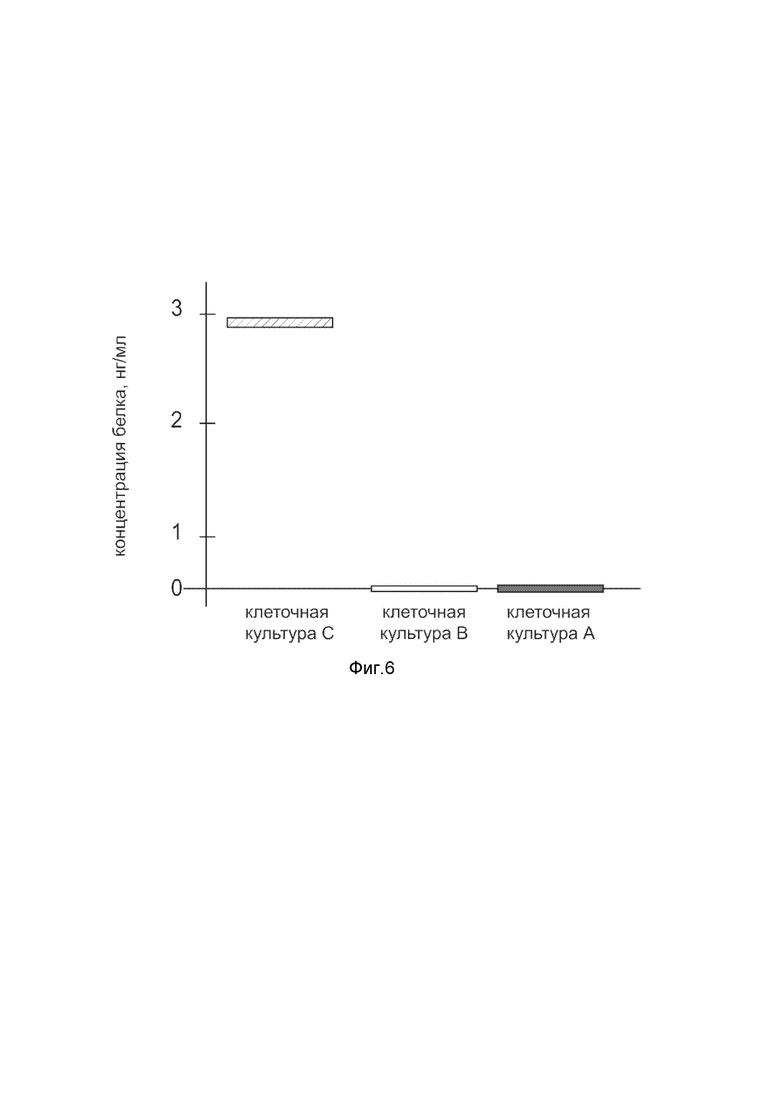
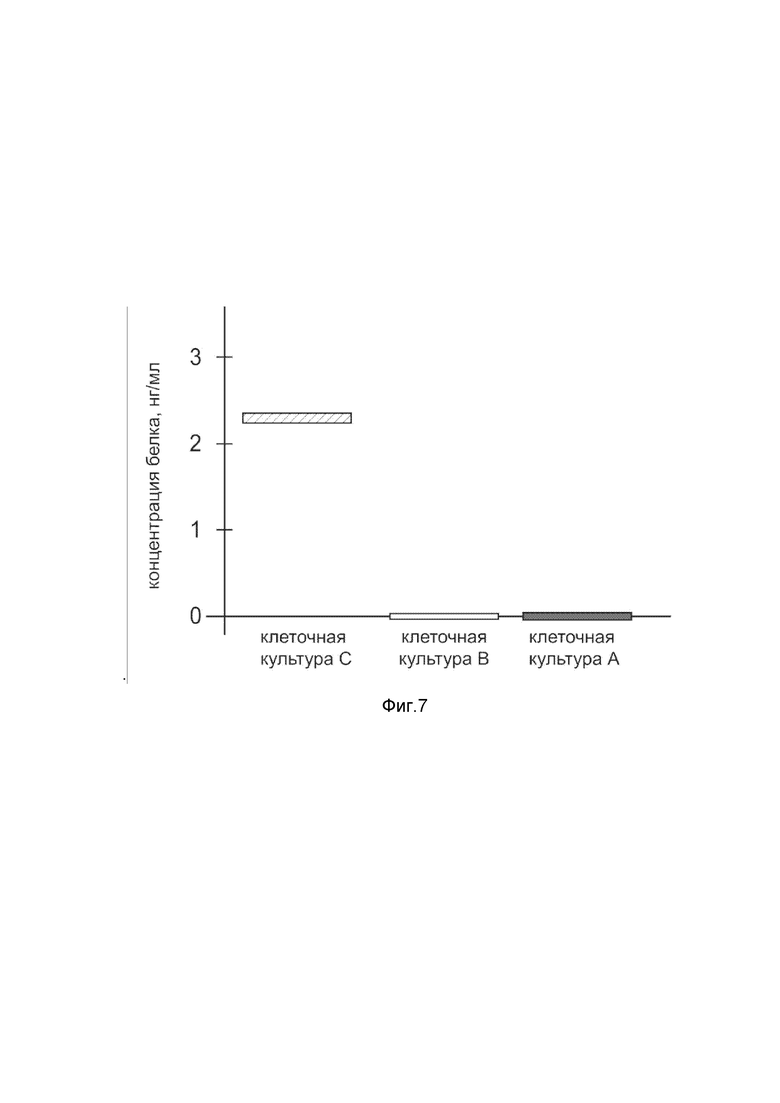
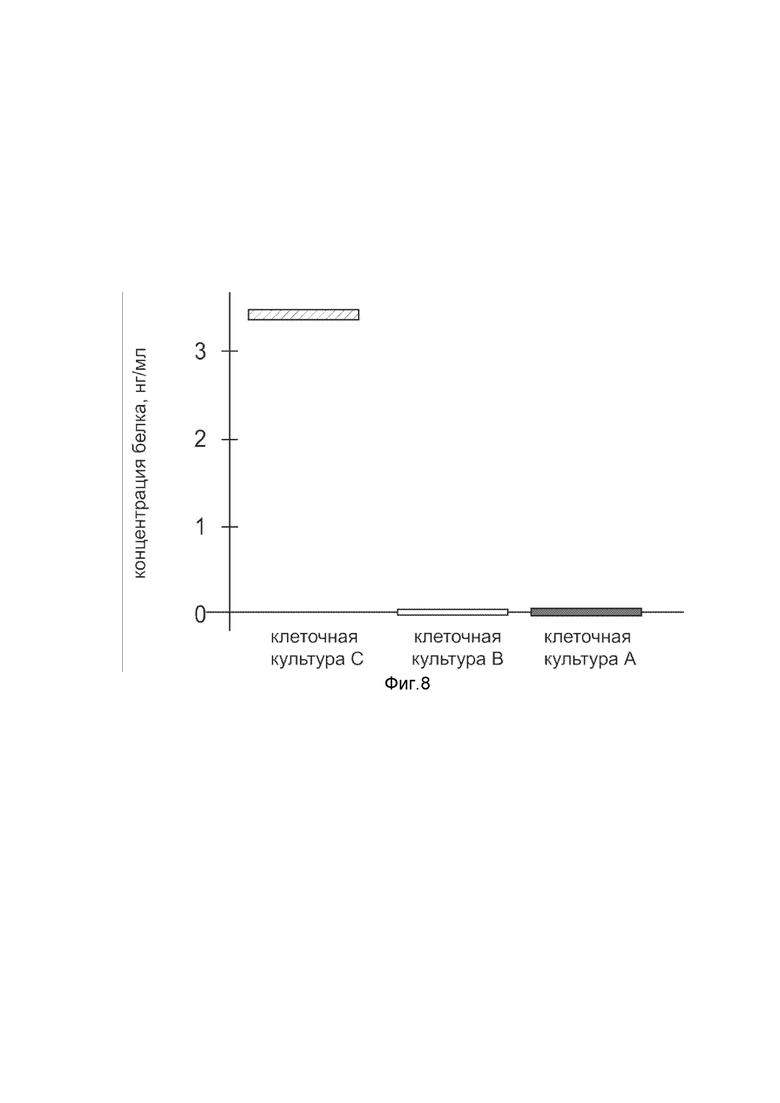
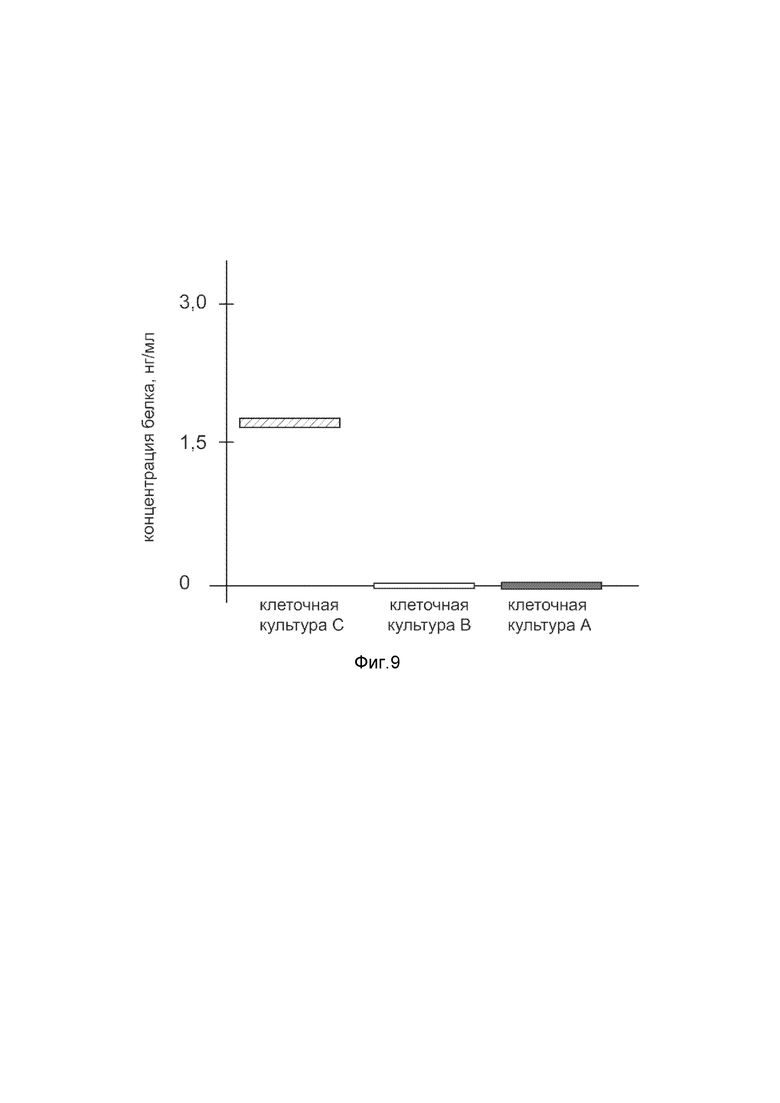



Изобретение относится к генной инженерии и может быть использовано в биотехнологии, медицине и сельском хозяйстве для создания препаратов генной терапии. Предложен генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген Cas9 для гетерологичной экспрессии этого целевого гена в клетках человека и животных при реализации различных методов геномного редактирования. При этом генотерапевтический ДНК-вектор VTvaf17-Cas9 имеет нуклеотидную последовательность SEQ ID №1. Предложены также способ получения указанного вектора, применение вектора, штамм Escherichia coli, несущий указанный вектор, а также способ производства в промышленных масштабах указанного вектора. Изобретение может быть использовано для безопасного применения для генетической терапии человека и животных. 6 н. и 2 з.п. ф-лы, 12 ил., 14 пр.
1. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген Cas9, для гетерологичной экспрессии этого целевого гена в клетках человека и животных при реализации методов геномного редактирования, при этом генотерапевтический ДНК-вектор VTvaf17-Cas9 имеет нуклеотидную последовательность SEQ ID №1.
2. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген Cas9 по п. 1, отличающийся тем, что созданный генотерапевтический ДНК-вектор VTvaf17-Cas9 по п. 1 за счет ограниченного размера векторной части VTvaf17, не превышающей 3200 п.н., обладает способностью эффективно проникать в клетки человека и животных и экспрессировать клонированный в него целевой ген Cas9.
3. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген Cas9 по п. 1, отличающийся тем, что в составе генотерапевтического ДНК-вектора отсутствуют нуклеотидные последовательности вирусного происхождения и отсутствуют гены антибиотикорезистентности, обеспечивая возможность его безопасного применения для реализации различных методов геномного редактирования геномов человека и животного, в том числе при генетической терапии человека и животного.
4. Способ получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9 по п. 1, заключающийся в том, что генотерапевтический ДНК-вектор VTvaf17-Cas9 по п. 1 получают следующим образом: кодирующую часть целевого гена Cas9 клонируют в ДНК-вектор VTvaf17 и получают генотерапевтический ДНК-вектор VTvaf17-Cas9.
5. Способ применения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген Cas9 по п. 1, для гетерологичной экспрессии этого целевого гена в клетках человека и животных, заключающийся во введении генотерапевтического ДНК-вектора по п. 1 в клетки, органы и ткани человека или животного совместно с gRNA и/или во введении в органы и ткани человека или животного аутологичных клеток человека или животного, трансфицированных генотерапевтическим ДНК-вектором по п. 1 совместно с gRNA или в сочетании обозначенных способов.
6. Способ получения штамма Escherichia coli SCS110-AF/VTvaf17-Cas9, который заключается в электропорации компетентных клеток штамма Escherichia coli SCS110-AF генотерапевтическим ДНК-вектором по п. 1 и последующей селекцией стабильных клонов штамма с использованием селективной среды.
7. Штамм, полученный по п. 6, Escherichia coli SCS110-AF/VTvaf17-Cas9, несущий генотерапевтический ДНК-вектор по п. 1 для гетерологичной экспрессии целевого гена в клетках человека и животных.
8. Способ производства в промышленных масштабах генотерапевтического ДНК-вектора по п. 1, заключающийся в масштабировании бактериальной культуры штамма по п. 7 до количеств, необходимых для наращивания бактериальной биомассы в промышленном ферментере, после чего биомассу используют для выделения фракции, содержащей целевой ДНК-продукт - генотерапевтический ДНК-вектор VTvaf17-Cas9 по п. 1, многостадийно фильтруют и очищают хроматографическими методами.
| CN 104480144 A, 01.04.2015 | |||
| CN 103981216 B, 22.06.2016 | |||
| LIU C | |||
| ET AL | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| J Control Release | |||
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| СМИРНОВ А.В | |||
| И ДР | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Вавиловский журнал генетики и селекции | |||
| Токарный резец | 1924 |
|
SU2016A1 |

