Настоящее изобретение относится к количественному клеточному способу определения in vitro действия лиганда антитела против CD26, предпочтительно моноклонального антитела против CD26, такого как бегеломаб.
CD26 представляет собой многофункциональный гликопротеин 110 кДа, экспрессирующийся как на клеточной поверхности, так и в растворимой форме. Антиген CD26 экспрессируется различными тканями и органами, такими как легкие, эндотелий, сердце, головной мозг, печень, кишечник, почки, плацента, поджелудочная железа и скелетные мышцы (Abbott С.A. et al., 1994). На клеточном уровне обнаружено, что экспрессия CD26 обладает сильной костимулирующей активностью в популяциях лимфоцитов, в частности, в активированных Т-лимфоцитах, в покоящихся Т-клетках и на В-лимфоцитах (Cordero OJ et al., 2007). Фактически, в специфической подгруппе Т-клеток памяти экспрессия CD26 увеличивается после активации самих Т-клеток (Morimoto С.et al., 1989). Экспрессия CD26 на Т-клетках ассоциирована со способностью этих клеток продуцировать большие количества интерлейкина-2 (IL-2) и сильно пролиферировать в ответ на митогенную стимуляцию. Однако, экспрессия CD26 отрицательно коррелирует с активностью Т-хелперных лимфоцитов (Mattern Т. et al., 1991).
CD26 отличается ферментативной активностью дипептидил-пептидазы IV (DPP-IV). Указанная ферментативная активность специфически стимулирует гидролиз пептидной связи между N-концевой аминокислотой в положении Х-Pro и соседними аминокислотами (Gorrel M.D. et al., 2001). CD26 относится к подгруппе олигопептидаз, которые могут отщеплять N-концевые дипептиды от множества биологически активных субстратов, таких как цитокины, полипептиды, гормоны и хемокины (De Meester I. et al, 1999; Hildebrandt M. et al., 2000).
У людей CD26 также вовлечен в связывание с аденозиндезаминазой (ADA) (Franco R. et al., 1998). Дефицит ADA служит предрасполагающим фактором к иммунодефицитным заболеваниям, не только путем общих механизмов нарушения регуляции иммунной системы, но также путем внутриклеточной аккумуляции токсических метаболитов пуринового метаболизма (Sauer A.V. et al., 2012).
Возможный эффект от связи между CD26 и ADA заключается в факте модуляции локальной внеклеточной концентрации аденозина, которая приводит к отрицательным сигнальным путям в Т-клетках через аденозиновые рецепторы клеточной поверхности. Некоторые моноклональные антитела, специфические в отношении CD26, способны передавать активирующий сигнал в Т-клетки и регулировать иммунные ответы in vitro (Morimoto С.и Schlossman S.F et al, 1998). Таким образом, CD26 ассоциирован с регуляцией воспаления, иммунной, эндокринной и нервной функциями и также патофизиологией СПИД (синдром приобретенного иммунодефицита).
Ввиду своего повсеместного распространения множество патологических состояний коррелируют с нарушенной экспрессией и/или активностью CD26 в отношении тяжести соответствующего патологического состояния. Эти заболевания могут быть разделены на по меньшей мере пять категорий: аутоиммунные и воспалительные заболевания, злокачественные гематологические опухоли, психонейроэндокринные расстройства, инфекционные заболевания и солидные опухоли. Многие исследователи обнаружили, что при различных опосредованных иммунной системой заболеваниях уровни ферментативной активности CD26 в сыворотке крови, по-видимому, изменяются (Klemann С.et al., 2016). В клинических исследованиях обнаружено, что вариации в экспрессии/активности CD26 вовлечены в различные аутоиммунные заболевания, такие как ревматоидный артрит, рассеянный склероз, сахарный диабет I типа и отторжение трансплантата у хозяина (GvHD).
В различных исследованиях, в которых вводили высокоизбирательные ингибиторы CD26, продемонстрировано замедление возникновения диабета, уменьшение инсулита, увеличение количества регуляторных Т-клеток, что свидетельствует о важной роли CD26 в иммунной регуляции.
В нескольких отчетах по пациентам, страдающим ревматоидным артритом, показана корреляция между экспрессией/ферментативной активностью CD26, тяжестью заболевания и лечением. Эти обнаружения могут создать предпосылки для новых терапевтических подходов, имеющих своей целью ингибирование ферментативной активности CD26. Кроме того, обнаружено, что экспрессия CD26 выше на поверхности Т-клеток в крови и спинномозговой жидкости пациентов, страдающих рассеянным склерозом (Ohnuma K. et al., 2011). GvHD является основным осложнением после трансплантации гематопоэтических стволовых клеток (HSCT), представляющей собой важную терапию при многих гематологических заболеваниях (Ferrara J.L.M. et al., 2009). GvFID может быть классифицировано на острое или хроническое на основании времени возникновения и вызываться трансплантацией наивных Т-клеток, полученных из костного мозга донора, повреждая ткань реципиента (Henden A.S., Hill G.R., 2015). GvHD накладывает сильное ограничение на применение трансплантации костного мозга в качестве жизнесохраняющей терапии (Welniak L.A. et al., 2007).
Три состояния как правило необходимы для развития GvHD: (1) костный мозг донора должен содержать иммунокомпетентные Т-клетки, (2) реципиент должен экспрессировать тканевые антигены, которые не представлены у донора, и (3) реципиент не должен обладать способностью запускать эффективный ответ, который мог бы разрушить трансплантированные клетки.
GvHD характеризуется 3 отдельными фазами: 1) на первой фазе ткани повреждаются у хозяина вследствие лучевой терапии и химиотерапии с высвобождением провоспалительных цитокинов, таких как TNF-альфа (фактор некроза опухоли альфа) и IFN-гамма (интерферон гамма); 2) на второй фазе активации аллореактивные Т-клетки донора активируются антигенами, находящимися на клетках, презентирующих антиген реципиента (АРС), 3) наконец возникает клеточная пролиферация с дальнейшей секрецией цитокинов, продуцируемых как цитотоксическими Т-клетками, так и эффекторными Т-клетками (Ferrara J.L.M. et al., 2009).
Патологию, тяжесть и органную специфичность GvHD (aGvHD) определяют по балансу между различными субпопуляциями Т-лимфоцитов: Th1, Th2 и Th17. Фактически показано, что преобладание подтипа Th1 относительно других зависит от цитокинового окружения во время активации Т-клеток донора, последующего взаимодействия с клетками АРС хозяина и их последующей дифференцировки в хелперные Т-клетки.
Основные цитокины Thl представляют собой IFN-γ, IL-2 и TNF альфа. Увеличенные количества цитокинов Th1, таких как TNF и IFN-γ, ассоциированы с более ранним и тяжелым возникновением заболевания вследствие их способности приводить к повышающей регуляции хемокинов и рецепторов, способных стимулировать воспалительную реакцию. Цитокины Th2 представляют собой IL-4, IL-5, IL-10 и IL-13. Обнаружено, что блокирование ответа клетками Th2 ассоциировано с увеличением желудочно-кишечных симптомов и уменьшением уровней поражения печени и кожи. IL-6, который представляет собой провоспалительный цитокин, контролирует баланс между клетками Th17 и регуляторными Т-клетками. Ингибирование IL-6 может представлять собой потенциально эффективную стратегию для уменьшения тяжести aGvHD при НСТ (трансплантации гемопоэтических клеток) путем индукции иммунологической толерантности (Henden A.S., Hill G.R., 2015).
Таким образом, выявление роли CD26 в процессе ответа Т-клеток и изменения уровней цитокинов безусловно может способствовать пониманию возникновения феномена и прогрессирования GvHD (Henden A.S., Hill G.R., 2015; Yi Т. 2009).
На основе этой предпосылки новый терапевтический подход для предотвращения возникновения GvHD основан на применении моноклональных антител против CD26 (Hatano R., 2013; Bacigalupo А., 2016). Антитела против CD26 ранее проходили разработку в доклинических моделях для предотвращения возникновения GvHD в животных моделях заболевания. Хотя роль CD26/DPPIV при GvHD должна быть исследована дополнительно, сообщалось о том, что обработка мышиным антителом против человеческого CD26 (т.е. бегеломабом) эффективна в лечении GvHD у пациентов, страдающих острым стероидорезистентным GvHD (патент США 9376498). Таким образом, клинические данные подтверждают, что инактивация CD26 моноклональным антителом представляет собой подходящий терапевтический подход для эрадикации субпопуляции аутореактивных Т-клеток с последующей регрессией GVHD.
В свете вышеизложенного, очевидна важность рецептора CD26 в качестве молекулярной мишени в новых терапевтических подходах.
Тест на эффективность обеспечивает количественное измерение биологической активности конкретного лекарственного средства и представляет ключевой параметр качества в процессе разработки фармакологической молекулы.
Для разработки теста на эффективность могут быть использованы различные подходы, включая анализы лиганд-рецептор, анализы на животных, клеточные анализы in vitro или другие биохимические анализы (например ферментативные анализы). Тест на эффективность является особенно подходящим в том случае, если он воспроизводит механизм действия конкретного лекарственного средства.
Для биологических продуктов предпочтительно использовать анализ лиганд-рецептор или клеточные эксперименты in vitro. Поскольку последние способны обеспечить прямое измерение аффинности лекарственного средства в отношении своей молекулярной мишени, они также могут быть подходящими для теста на эффективность.
Тем не менее, не всегда возможно использовать эти типы анализов, просто потому что тесты на эффективность должны быть разработаны на основе механизма действия лекарственного средства, но эта информация не всегда доступна, особенно для моноклональных антител. Этот подход особенно затруднен, поскольку биологические лекарства часто обладают множеством механизмов действия in vivo, таких которые особенно сложно воспроизвести в системе in vitro.
Авторы настоящего изобретения обнаружили, что моноклональное антитело против CD26 (бегеломаб) способно вызывать интернализацию рецептора CD26 после специфического связывания при помощи механизма действия, известного как "кэппинг". Явление интернализации приводит к ингибированию высвобождения провоспалительных цитокинов, которые играют ключевую роль в воспалительных процессах, что представляет непосредственное последующее функциональное явление, вызываемое лигандом антитела против CD26. Таким образом, описанный механизм действия подтверждает все применения моноклональных антител при всех аутоиммунных заболеваниях, в которых важно дезактивировать аутореактивные Т-лимфоциты при сохранении их иммунокомпетентности.
Следовательно, в качестве теста на эффективность фармацевтического продукта для задачи определения количества активного соединения в образце функциональным путем как интернализация CD26, так и ингибирование секреции воспалительных цитокинов представляют новый тест, который может быть использован для оценки эффективности какого-либо лиганда антитела против CD26, предпочтительно моноклонального антитела, разрабатываемого для терапевтических и/или диагностических применений.
Описанный подход может быть использован для оценки эффективности любого антитела против CD26 на основе принципиально нового обнаружения механизма действия бегеломаба. Кроме того, использование этого способа обеспечивает количественное измерение как интернализации CD26, так и ингибирования секреции/продукции образующихся цитокинов.
В заключение, эти обнаружения обеспечивают возможность получения чрезвычайно полезного теста на эффективность с точки зрения 1) воспроизводимости, 2) легкости количественного определения активности лиганда антитела против CD26 в конкретном образце и 3) количественного измерения активности лиганда антитела против CD26, независимо от того, является ли оно известным как бегеломаб или вновь идентифицированным антителом.
Таким образом, настоящее изобретение относится к способу определения in vitro эффективности лиганда антитела против CD26, включающему следующие стадии:
а) инкубацию при 37°С популяции человеческих Т-лимфоцитов, экспрессирующих рецептор CD26, в процентной доле более 75% с лигандом антитела против CD26 в концентрации в диапазоне от 0,001 мкг/мл до 150 мкг/мл, предпочтительно от 0,01 мкг/мл до 100 мкг/мл, более предпочтительно от 0,01 мкг/мл до 2 мкг/мл, еще более предпочтительно от 0,01 мкг/мл до 0,5 мкг/мл;
б) инкубацию с антителом против CD26, меченым флуорохромом, которое распознает эпитоп на рецепторе CD26, отличающийся от эпитопа, распознаваемого лигандом антитела против CD26 на стадии а);
в) определение величины MFI (медиана интенсивности флуоресценции) для CD26, измеренной для образца клеток, обработанных лигандом антитела против CD26 (MFIT), и величины MFI для необработанных клеток (MFINT) путем цитофлуориметрического анализа;
г) оценку процентной доли интернализации рецептора CD26 в виде RFI (относительная интенсивность флуоресценции, то есть величина MFI, нормализованная относительно базовой величины), рассчитанной как отношение между величиной MFI для CD26, измеренной для образца клеток, обработанных лигандом антитела против CD26 (MFIT), и величиной MFI для необработанных клеток (MFINT), умноженной на 100, и затем отнятой от 100, в соответствии со следующей формулой:
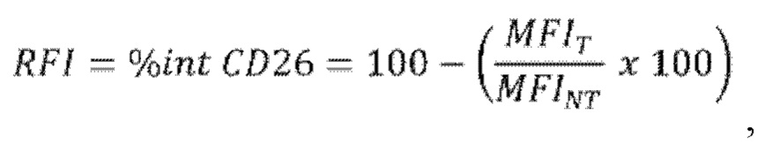
где "%int CD26" или RFI представляет собой процентную долю интернализации CD26, "MFIT" представляет собой величину MFI для клеток, обработанных лигандом CD26 (тестируемые клетки), и "MFINT" представляет собой величину MFI для клеток, не обработанных лигандом CD26 (референсные клетки).
Такая процентная доля (%int CD26 или RFI):
- если составляет менее 20%, то указывает на низкую эффективность лиганда антитела против CD26;
- если находится в диапазоне от 20% до 30%, то указывает на среднюю эффективность лиганда антитела против CD26;
- если составляет выше 30%, то указывает на высокую эффективность лиганда антитела против CD26.
На Фиг. 12, блок А, представлено логическое основание для присвоения критериев "низкая", "средняя" и "высокая" эффективность, соответственно. В частности, обнаружено, что ответ с точки зрения % интернализации CD26 зависит от дозы в соответствии с формой кривой S (сигмоидальная), где при меньшей концентрации лиганда (на этой фигуре обозначен как "RS") она имеет меньшую величину "%int CD26", и наоборот, при большей концентрации лиганда она имеет большую величину "%int CD26" вплоть до максимальной величины (плато) 30%-35%. Если это плато рассматривать как 100% от "%int CD26", тогда возможно нормализовать все величины кривой, как показано на Фиг. 12, блок В.
В этом смысле рассматривается кривая, начинающаяся с 0% до 100% интернализации, и, таким образом, можно определить три различных диапазона эффективности:
1. "низкая", которая относится к лиганду, имеющему процентную долю интернализации в диапазоне от 0% до 50% относительно плато,
2. "средняя", которая относится к лиганду, имеющему процентную долю интернализации в диапазоне от 50% до 90% относительно плато,
3. "высокая", которая относится к лиганду, имеющему процентную долю интернализации выше 90% относительно плато.
Другими словами, эта концепция может быть обобщена в следующей Таблице 1:

В альтернативном воплощении изобретения популяция человеческих лимфоцитов со стадии а) может быть инкубирована при комнатной температуре.
Указанный лиганд антитела против CD26 представляет собой молекулу, способную специфически связываться с рецептором CD26, предпочтительно моноклональное антитело против CD26 или его фрагменты, более предпочтительно, бегеломаб.
В предпочтительном воплощении способа по изобретению концентрация лиганда антитела против CD26 со стадии а) составляет 0,001 мкг/мл, 0,01 мкг/мл, 0,5 мкг/мл или 2 мкг/мл.
В предпочтительном воплощении способа по изобретению антитело против CD26 со стадии б) представляет собой мышиное антитело против человеческого CD26, конъюгированное с флуорохромом АРС (BD Pharmingen; номер по каталогу: 563670, номер клона: М-А261), и его инкубируют в концентрации 2,5 мкг/мл.
В соответствии с еще одним предпочтительным воплощением способа по изобретению флуорохром, используемый на стадии б), представляет собой любой флуорохром, который может быть использован для цитофлуориметрического анализа, выбранный из группы, состоящей из FITC (флуоресцеин изотиоцианат), АРС (аллофикоцианин), РЕ (фикоэритрин), РЕ-Су7, АРС-Н7, PerCP (перидинин-хлорофилл протеин) и РЕ-Су5.5. Указанный флуорохром предпочтительно представляет собой АРС.
Кроме того, в соответствии с предпочтительным воплощением способа по изобретению популяцию CD26+ Т-лимфоцитов со стадии а) отбирают из популяции первичных Т-лимфоцитов и линии опухолевых клеток человеческих Т-лимфоцитов. Предпочтительно, указанная опухолевая клеточная линия человеческих Т-лимфоцитов представляет собой клеточную линию Karpas 299.
В предпочтительном воплощении способа по изобретению цитофлуориметрический анализ со стадии в) осуществляют путем FACS (анализа сортировки клеток с активированной флуоресценцией).
В способе по изобретению также предложена дополнительная стадия верификации эффективности лиганда антитела против CD26, которая предложена для тестирования ингибирования высвобождения цитокинов вышеупомянутым лигандом, где указанные цитокины выбраны из группы, состоящей из IL-8 (интерлейкин-8), IL-1β, IL-6, IL-2, GM-CSF (колониестимулирующий фактор гранулоцитов-макрофагов), IL-6 и TNF-α, осуществляемого на популяции CD26+ человеческих Т-лимфоцитов со стадии а). В предпочтительном воплощении указанные цитокины представляют собой IL-8 и/или IL-1β. Указанная популяция CD26+ человеческих Т-лимфоцитов со стадии а) выбрана из популяции первичных Т-лимфоцитов и опухолевой клеточной линии человеческих Т-лимфоцитов. Предпочтительно, указанная опухолевая клеточная линия человеческих Т-лимфоцитов представляет собой клеточную линию Karpas 299.
В предпочтительном воплощении указанное тестирование ингибирования продукции цитокинов осуществляют путем анализа MesoScale Discovery (MSD).
Настоящее изобретение далее будет описано для иллюстративных, а не ограничивающих объем изобретения задач в соответствии с предпочтительным воплощением с конкретной ссылкой на графические материалы, на которых:
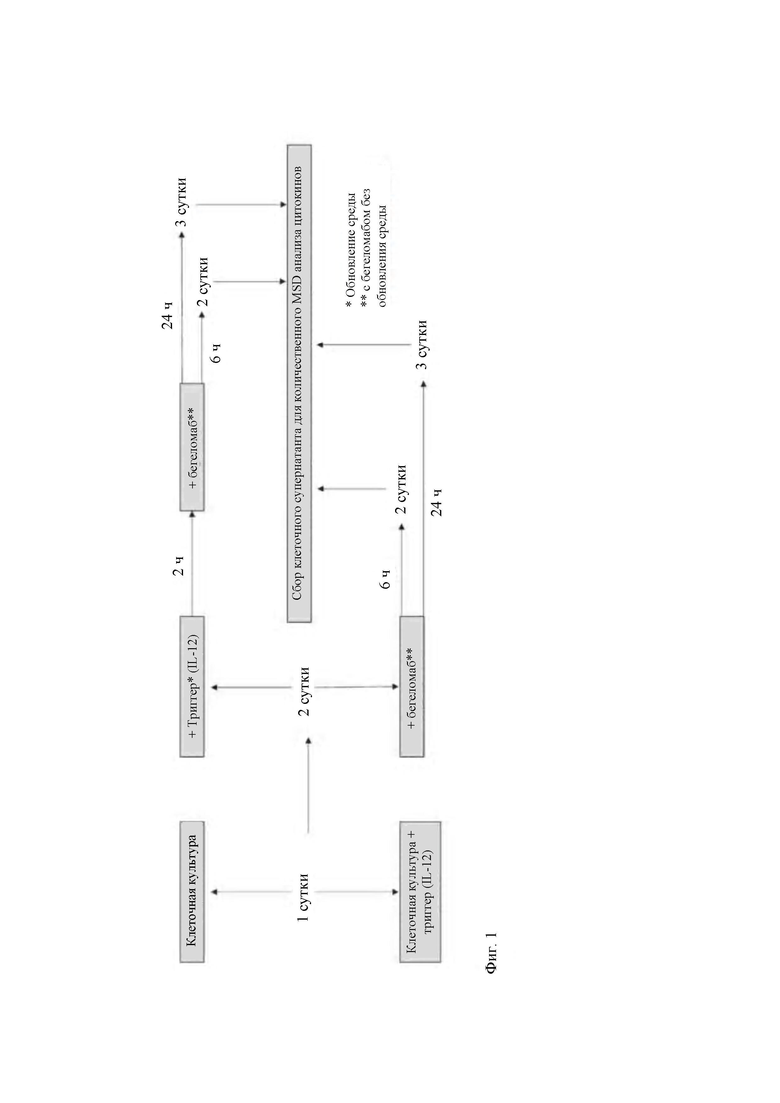
- на Фиг. 1 показана экспериментальная схема клеточной культуры для анализа уровней провоспалительных цитокинов путем MSD-анализа;
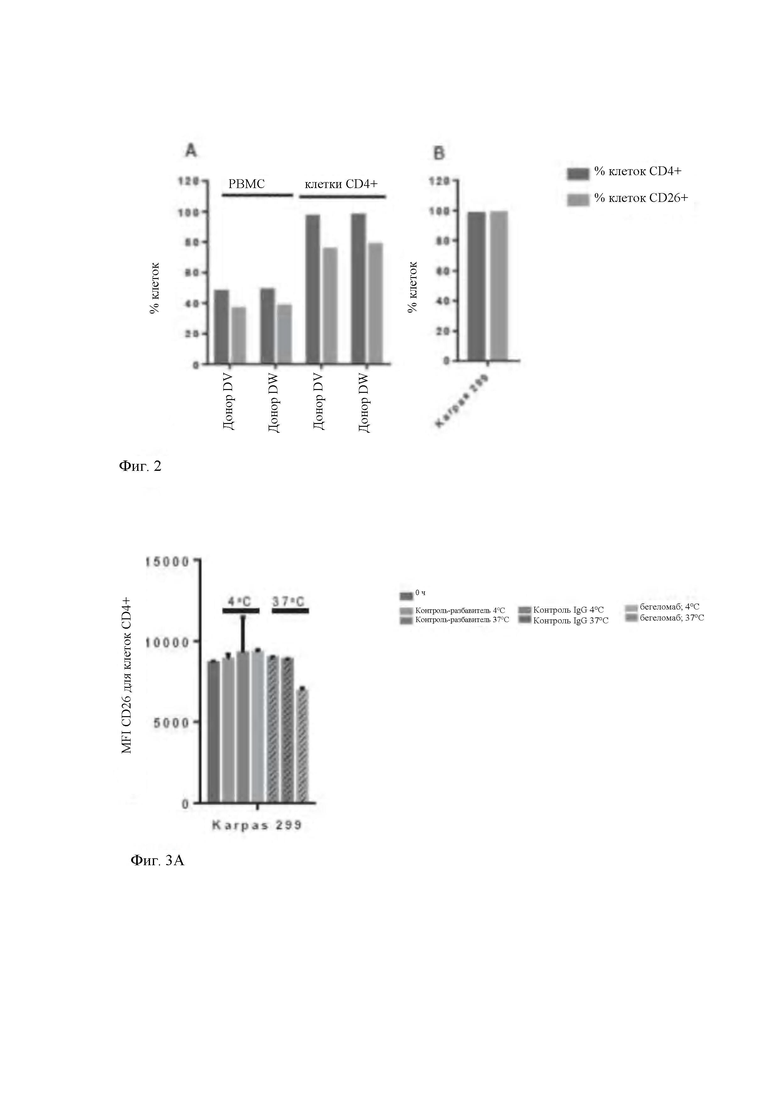
- на Фиг. 2 показан цитофлуориметрический анализ процентной доли клеток CD4+ (темный столбец) и клеток CD26+ (левый столбец) в первичных мононуклеарных клетках периферической крови и очищенных CD4+ Т-клетках (А) и человеческой Т-клеточной линии Karpas 299 (В);
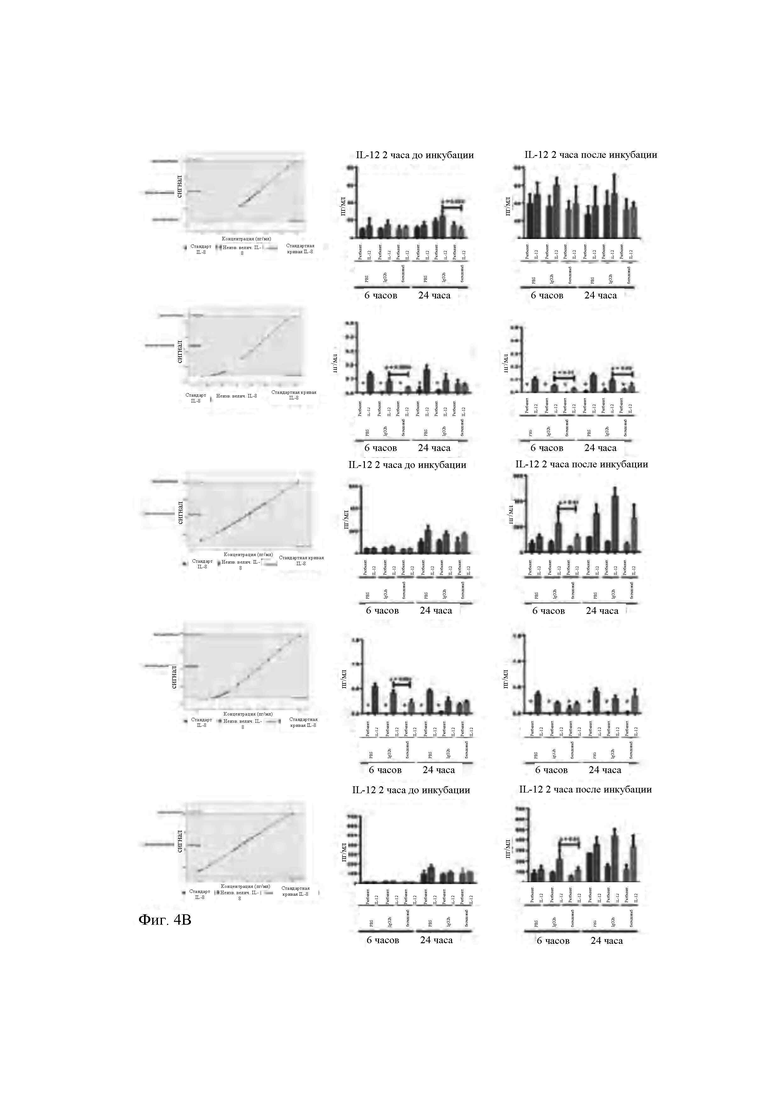
- на Фиг. 3 продемонстрировано действие бегеломаба на интернализацию рецептора CD26 в клетках Karpas 299 после 8 часов инкубации (блок А) и первичных CD3+, CD4+, CD8+ Т-клетках, инкубируемых при 4°С и 37°С (блоки В, С, D) после 6 и 24 часов;
- на Фиг. 4 продемонстрировано ингибирование уровней цитокинов IL-2, IL-8, IL-1β, GM-CSF, IL-6 и TNF-α, вызванное бегеломабом, в Т-клеточной линии Karpas 299 (блоки А) и в первичных Т-клетках (блоки В). На графике слева от оси ординат указан сигнал электрохемилюминесценции, тогда как на оси абсцисс указана соответствующая концентрация;
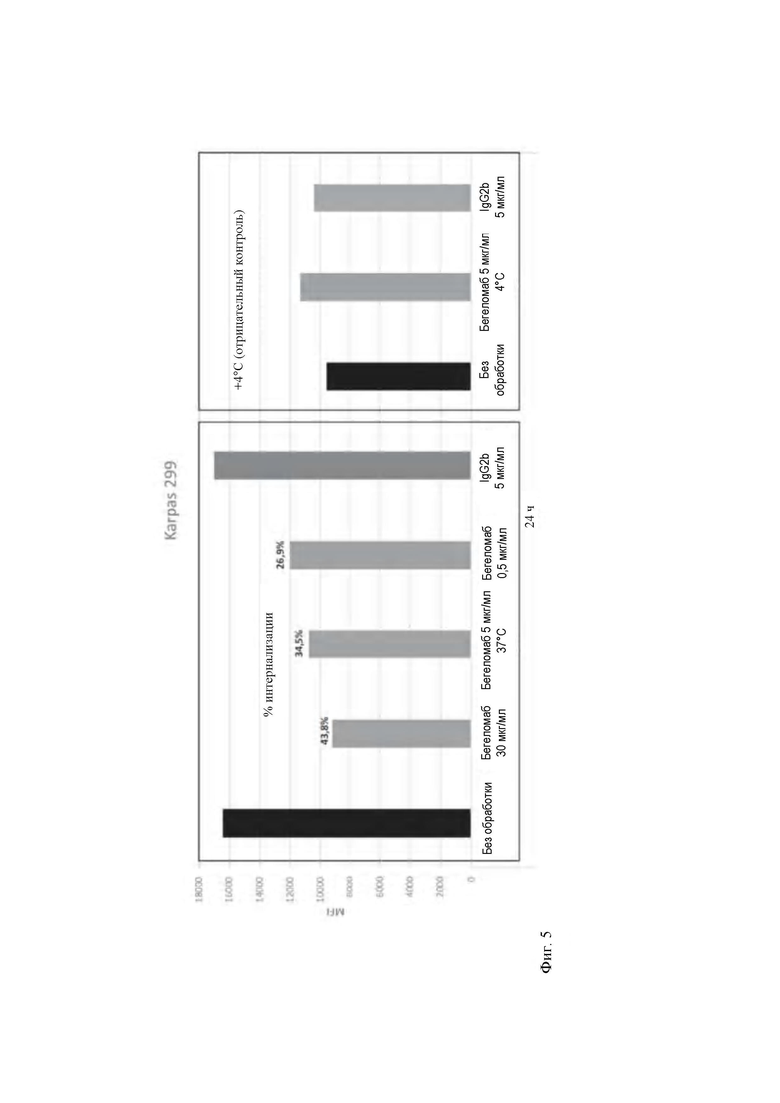
- на Фиг. 5 продемонстрировано уменьшение флуоресценции, ассоциированное с CD26, после 24 часов обработки бегеломабом в клетках Karpas 299;
- на Фиг. 6 продемонстрировано уменьшение флуоресценции, ассоциированное с CD26, после 24 часов в клетках Karpas 299 после обработки, начиная с низкой дозы 0,0005 мкг/мл (3×10-12 М) до более высокой концентрации бегеломаба, равной 150 мкг/мл (1×10-6 М);
- на Фиг. 7 показаны кривые зависимости ответа от дозы для интернализации CD26 (RFI), регистрируемые путем анализа FACS, осуществляемого на образцах с бегеломабом, хранимых при различных температурных условиях (SVI-STB представляет собой образец с сокращенной эффективностью, то есть образец, подвергаемый условиям ускоренного старения при 32,5°С в течение 6 месяцев);
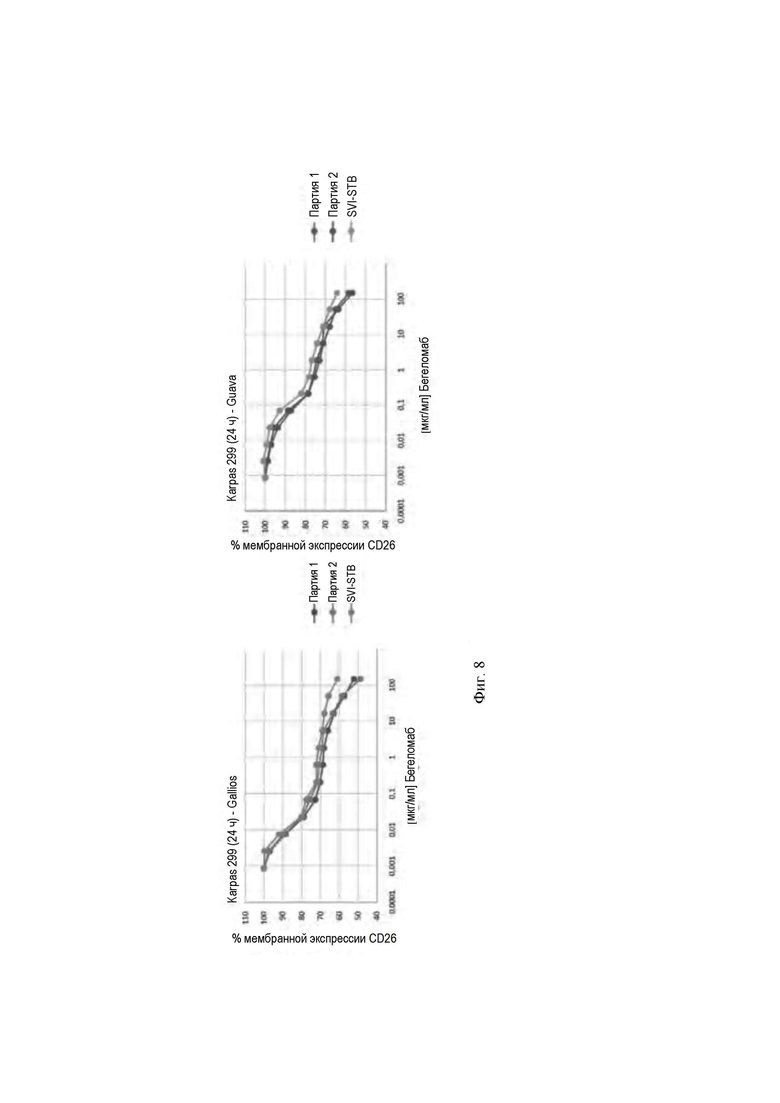
- на Фиг. 8 показаны кривые интернализации CD26 (RFI), регистрируемые путем анализа FACS на различных цитофлуориметрах;
- на Фиг. 9 продемонстрирована относительная эффективность мышиного моноклонального антитела против hCD26 (клон 202.36), наблюдаемая относительно бегеломаба (WS-BEG-013);
- на Фиг. 10 продемонстрирована относительная эффективность мышиного антитела IgG2b изотипа K (клон MG2b-57), наблюдаемая относительно бегеломаба (WS-BEG-013);
- на Фиг. 11 показана величина эффективности для тестируемого образца (TEST), рассчитанная относительно референсного стандарта RS (бегеломаб), для демонстрации того, как относительная активность для любого неизвестного образца, названного TEST, может быть количественно измерена относительно референсного стандарта (RS);
- на Фиг. 12 показаны два иллюстративных графика критериев, применяемых для определения низкой, средней или высокой эффективности.
Для того чтобы лучше проиллюстрировать данное изобретение, приведены следующие примеры, которые следует рассматривать как иллюстративные, а не ограничивающие его.
ПРИМЕР 1: Исследование механизма действия моноклонального антитела бегеломаба
Механизм действия бегеломаба (антитело против CD26) исследовали для разработки теста для оценки эффективности антитела in vitro.
Разработка этого теста, названного как тест на эффективность, весьма важна для измерения биологической активности данного лекарственного средства, поскольку эта активность представляет собой важный компонент контроля качества молекулы во время ранней разработки, последующих стадий разработки и фаз вывода на рынок в объеме контроля качества как разрешение на выпуск партии.
В частности, множество биологических продуктов, таких как моноклональные антитела, реализуют свою функцию путем связывания с клеточной или растворимой мишенью, затем запуская соответствующие клеточные события ниже от молекулярной мишени.
Таким образом, оценивали следующее:
- Экспрессию/сверхэкспрессию CD26 в человеческих Т-клеточных линиях и человеческих первичных Т-клетках.
- Интернализацию CD26 после связывания с бегеломабом.
- Действие бегеломаба на продуцирование и высвобождение воспалительных цитокинов.
МАТЕРИАЛЫ И МЕТОДЫ
Первичные клетки и клеточные линии
Человеческая Т-клеточная линия происходит из Т-клеток лимфомы Karpas 299, приобретенной в SIGMA (Кат. 06072604-1VL). Клетки Karpas 299 выращивали в RPMI1640 с GlutaMAX (Gibco, кат. #72400), дополненной 10% термически инактивированной фетальной бычьей сывороткой (Sigma, кат. #F2442, температурную инактивацию осуществляли в лаборатории), 1 МЕ/мл пенициллина/стрептомицина (Gibco, кат. #15140), 2 мМ L-глутамина (Gibco, кат. #2503) и 50 мкМ β-меркаптоэтанола (Sigma-Aldrich, кат. #М3148). После оттаивания клеткам давали возможность вырасти, и их подвергали криоконсервации.
Клеточную линию анализировали для оценки присутствия микопазмы (MycoAlert™, Lonza, кат. #LT-07) и обнаружили отсутствие в ней последней. Мононуклеарные клетки периферической крови (РВМС) выделяли при помощи разделяющего градиента Ficoll-Hypaque (Lymphoprep, Fresenius) из лейкоцитарной пленки трех здоровых доноров, полученной после информированного согласия в соответствии с протоколом, одобренным Этическим Комитетом Сан Раффаэля (San Raffaele Ethics Committee) (IRB). Для оценки жизнеспособности клеток РВМС окрашивали трипановым синим и подсчитывали в камере Бюркера. РВМС выращивали в полной среде (RPMI 10% FBS плюс 1% PenStrep и глутамин) с низкими дозами IL-2 (50 МЕ/мл) и IL-7 (5 мкг/мл) при 37°С в течение ночи (о/n) до начала эксперимента.
Для определения экспрессии CD26 на клеточной поверхности человеческих первичных CD4+ Т-клеток и Т-клеточных линий приобрели антитело a-CD26-FITC клон BA5b (Biolegend, кат. #302704). Антитело тестировали в диапазоне разведения на первичных Т-клетках трех доноров и на клетках Karpas 299 для определения оптимальной концентрации для цитофлуориметрического анализа. Оптимальную концентрацию используют для определения процентной доли экспрессии CD26 на человеческих первичных CD4+ Т-клетках и Т-клеточных линиях, а также для анализа интернализации CD26.
Экспрессия CD26 на Т-клеточных линиях и человеческих первичных CD4+ Т-клетках Первичные CD4+ клетки двух доноров и клетки Karpas 299 высевали в буфере autoMACS (Miltenyi, кат. #130-091-222) с добавлением 0,2% BSA (бычий сывороточный альбумин) (Miltenyi, кат. #130-091-376) с плотностью 100000 клеток/лунку в 96-луночные планшеты с V-образной формой дна лунки (Costar, кат. #3598). Клетки инкубировали в течение 15 минут в диапазоне концентраций a-CD26-FITC или a-IgG2a-FITC (Biolegend, кат. #400208) от 2,5 мкг/мл до 31,3 нг/мл. Клетки затем промывали и фиксировали в 4% формальдегиде. Процентную долю CD26-положительных клеток определяли посредством цитофлуориметрии с использованием FACSCanto П. IgG-FITC использовали для определения неспецифических связываний.
Количественное определение экспрессии CD26 на человеческих первичных клетках и Т-клеточной линии
Цитофлуориметрический анализ использовали для определения количества CD26, экспрессируемого на клеточной мембране первичных мононуклеарных клеток периферической крови (РВМС) и CD4+ Т-клетках, и также на клеточной линии Karpas 299. CD4+ метку добавляли в качестве Т-клеточного маркера. Первичные CD4+Т-клетки трех доноров и клетки Karpas 299 высевали в буфере MACS с плотностью 100000 клеток/лунку в 96-луночные планшеты с V-образной формой дна лунки. Клетки инкубировали с 10 мкг/мл a-CD26-FITC и 2,5 мкг/мл a-CD4 в течение 15 минут. Добавляли 10 мкг/мл a-IgG2a-FITC и 2,5 мкг/мл a-IgG2b-APC (Biolegend, кат.400612) для того, чтобы обеспечить гейтирование. Клетки затем промывали и фиксировали в 4% формальдегиде. Процентную долю CD4+ и CD26+ клеток определяли путем цитофлуориметрического анализа с использованием FACSCanto П.
Анализ интернализации CD26
Способность бегеломаба вызывать интернализацию рецептора CD26 в Т-клетках (как человеческих первичных Т-клетках, так и в Т-клеточной линии) определяли путем цитофлуориметрического анализа. Анализ осуществляли в двух независимых экспериментах. CD4+ Т-клетки выделяли из трех лейкоцитарных пленок на эксперимент.Затем как РВМС, так и выделенные клетки метили антителом против CD4 и антителом против CD26 для определения чистоты выделенных клеточных популяций, а также процентной доли клеток CD26 до эксперимента. Кроме того, краситель для определения жизнеспособности добавляли для обеспечения того, что эксперимент осуществляется на живых клетках. Клетки анализировали путем цитофлуориметрического анализа.
Клетки CD4+ оценили как имеющие хорошее качество, поскольку обнаруженная чистота популяции CD4+ составляла >95%; процентная доля клеток CD26+ в популяции CD4+ составляла >75%. Т-клеточную линию высевали в течение всей ночи (о/п) в количестве 100000 клеток/лунку в 96-луночных плоскодонных планшетах (Corning Costar, кат. #3598), покрытых 2 мкг/мл антитела против CD3/антитела против CD28 (eBioscience, кат. #16-0037-85 и 16-0289-85, соответственно), и инкубировали их в течение 2 часов при 37°С.После инкубации в течение ночи среду меняли и клетки инкубировали с 0,03 мг/мл (2×10-7 М) бегеломаба, IgG2b в качестве контроля (Sigma-Aldrich, кат. #SAB4700729 или Biolegend, кат. #401202) или PBS (Gibco, кат. #10010) в качестве контрольного разбавителя в общем объеме 100 мкл. Каждое условие тестировали в трех параллелях (три лунки с клетками/условие). Обработку осуществляли при 4°С и при 37°С в течение 8 часов. Клетки затем метили и процентную долю клеток CD26+ в клетках CD4+ определяли путем цитофлуориметрического анализа. Дополнительно, группу клеток метили и анализировали до начала обработки для определения процентной доли клеток CD4+CD26+ в нулевой момент времени. В день проведения эксперимента РВМС выделяли путем центрифугирования при 1500 об/мин в течение 5 минут и инкубировали с бегеломабом в различных концентрациях в течение 6 и 24 часов, соответственно. После инкубации с бегеломабом РВМС промывали и метили следующими моноклональными антителами: человеческие антитела против CD3, CD4, CD8 и CD26, конъюгированные с FITC-, РЕ-, АРС, АРС-Н7- (Biolegend). Ранее было показано, что моноклональное антитело против CD26 может связываться с эпитопом, отличающимся от эпитопа для бегеломаба. Антитело, конъюгированное с флуорофором, против изотипа, соответствующего изотипу бегеломаба (IgG2b), всегда использовали в качестве отрицательного контроля. Образцы анализировали с использованием проточного цитофлуориметра FACS Canto II (BD Biosciences). Все данные анализировали с использованием программного обеспечения Flow Jo (Tree Star Inc.) и выражали в виде относительной интенсивности флуоресценции (RFI) клеток CD4+ или CD8+. Эту величину рассчитывают путем деления средней интенсивности флуоресценции образца, меченого антителом против CD26, на соответствующий изотопический контроль.
9-плексный анализ MSD для провоспалителъных цитокинов Для оценки действия бегеломаба при активации Т-клеток оценивали продукцию провоспалительных цитокинов.
Для этой задачи использовали анализ MesoScale Discovery (MSD), а именно 9-плексный анализ человеческих провоспалительных цитокинов (кат. #15007 В-2). Как первичные Т-клетки, так и Т-клеточную линию Karpas 299, помещали в количестве 100000 клеток/лунку в 96-лучночные плоскодонные планшеты, покрытые антителом против CD3/антителом против CD28, как описано выше (раздел 4.1.1.2), и инкубировали в течение ночи.
Клетки хранили в среде RPMI 1640 с GlutaMAX и дополненной 10% HI-FBS, 1% пенициллином/стрептомицином, 2 мМ L-глутамином и 50 мкМ Р-меркаптоэтанолом. Клетки обрабатывали IL-12 и бегеломабом, как описано на Фиг. 1. Кратко, клетки предварительно инкубировали в течение ночи с 20 нг/мл IL-12 (R&D systems, кат. #219-IL-0059) и контрольным разбавителем (0,1% бычий сывороточный альбумин (BSA, Sigma-Aldrich, кат. #А2153-1 кг)-PBS) или инкубировали в течение ночи в контрольном разбавителе без IL-12. На следующие сутки клетки без предварительной инкубации в течение ночи с IL-12 инкубировали в течение 2 часов с 20 нг/мл IL-12. Затем все клетки инкубировали с 0,03 мг/мл (2×10-7 М) IgG2b (Biolegend), бегеломабом или контрольным разбавителем PBS в течение 6 часов или 24 часов. Во всех экспериментах супернатант собирали после определенного периода времени и замораживали при -80°С до новых анализов.
Для тестирования эффекта "покрывания" антителом против CD3 и антителом против CD28 в отношении секреции цитокинов клетки тестировали на покрытых и не покрытых планшетах и предварительно инкубировали в течение ночи с IL-12 и затем обрабатывали в течение 24 часов бегеломабом/IgG2b/PBS.
Анализ MSD цитокинов обнаруживает уровни интерлейкина 2 (IL-2), IL-8, IL-1β, колониестимулирующего фактора гранулоцитов-макрофагов (GM-CSF), интерферона гамма (IFN-γ), IL-6, IL-10, фактора некроза опухоли альфа (TNF-α) и IL-12p70. Анализ MSD цитокинов проводили в соответствии с указаниями производителя, планшеты фактически блокировали в течение ночи при 4°С вместо 1 часа при комнатной температуре для предотвращения возможности неспецифических связываний.
Стандартную кривую строили с использованием среды, адаптированной во время клеточной фазы эксперимента. Программное обеспечение MSD Discovery Workbench 4.0.12 использовали для построения стандартных кривых и для расчета концентрации цитокинов (пг/мл). Величины, полученные для образцов, накладывали на стандартную кривую для определения того, оказываются ли они в пределах ее диапазона. Графики для всех цитокинов строили с использованием GraphPad Prism 6. Исходно, для осуществления статистического анализа ингибирование бегеломабом, вдвое превышающее ингибирование для контроля IgG2b, произвольно установили в качестве порогового уровня.
Однако, поскольку различные условия демонстрировали 1,9-кратное ингибирование, эту пороговую величину сдвинули до 1,9. Для оценки статистически значимых различий в условиях обработки использовали двусторонний тест ANOVA с последующим тестом Тьюки в Graph Pad Prism 6.0, с получением р-величины, скорректированной на множественных тестах.
РЕЗУЛЬТАТЫ
Экспрессия/сверхэкспрессия CD26 в человеческих Т-клеточных линиях и в человеческих первичных Т-клетках
На Фиг. 2 представлена процентная доля клеток CD4+ и CD26+ в популяции мононуклеарных первичных клеток, полученных из периферической крови, в очищенных CD4+ Т-клетках и в линии Karpas 299. Результаты, полученные путем цитофлуориметрического анализа, демонстрируют, что процентная доля клеток CD4+ и CD26+ в первичных мононуклеарных клетках составляет приблизительно более 45% и более 35%, соответственно, после выделения CD4+ Т-клеток эти процентные доли составляют более 95% и более 75%, соответственно (Фиг. 2, блок А). Почти все клетки Karpas 299 экспрессируют CD4 (более 95%), а также высокие уровни CD26 (более 95%) (Фиг. 2, блок В). В завершение, процентная доля CD26-положительньгх клеток в выделенной популяции первичных CD4+ Т-клеток и в клеточной линии Karpas 299 приблизительно составляет 75% и 95%, соответственно. Вышеприведенные процентные доли также достаточны для измерения изменений уровней CD26 на последующих экспериментальных стадиях.
Интернализация CD26
Цитофлуориметрический анализ использовали для определения индуцированной бегеломабом интернализации CD26 в первичных Т-клетках и в Т-клеточных линиях.
Такой же анализ использовали также для идентификации дозозависимой взаимосвязи между уменьшением присутствия CD26 и концентрациями бегеломаба. В Т-клеточной линии Karpas 299 определяли процентную долю клеток CD26+, а также среднюю интенсивность флуоресценции (MFI) CD26+ в популяции CD4+. В Т-клеточной линии Karpas 299 обнаружено уменьшение CD26 MFI в популяции CD4+ только при 37°С после 8 часов (ингибирование на 7% после 4 часов - данные не показаны на фигуре - и 22,3% после 8 часов, соответственно) (Фиг. 3, блок А). Это уменьшение является статистически значимым при MFI для CD26 в популяции CD4+, измеренной после 8 часов инкубации с бегеломабом при 37 °С относительно инкубации, осуществляемой при 4°С, т.е. когда все биологические процессы ингибируются низкой температурой. Полученные данные свидетельствуют о том, что количество рецепторов CD26 на одну клетку уменьшается в клетках Karpas 299 при 37°С после связывания с бегеломабом. Поскольку такое же биологическое явление не наблюдается при 4°С, можно сделать вывод, что интернализация рецептора CD26 после связывания с бегеломабом представляет механизм действия антитела в Т-клеточной линии Karpas 299. Для обеспечения того, что интернализация рецептора CD26 в клетках Karpas 299, вызванная связыванием с бегеломабом, может быть использована в качестве теста для оценки эффективности выпускаемых партий для внедрения в клиническую практику, осуществляли эксперимент зависимости ответа от дозы. В вышеупомянутом эксперименте клетки Karpas 299 обрабатывали увеличивающимися концентрациями бегеломаба при 37 градусах, и в частности, 0,5 мкг/мл (3×10-9 М), 5 мкг/мл (3×10-8 М) и 30 мкг/мл (2×10-7 М) в течение 24 часов (Фиг. 5). Полученные результаты представляют огромный интерес, поскольку обнаружено, что увеличивающиеся концентрации антитела против CD26 способны вызывать прогрессирующее увеличение интернализации CD26, экспрессируемого на мембране. В частности, дозы бегеломаба 0,5 мкг/мл, 5 мкг/мл и 30 мкг/мл, 3×10-9 М, 3×10-8 М и 2×10-7 М, соответственно, соответствуют процентным долям интернализации 26,9%, 34,5% и 43,8%. Этот зависимый от дозы эффект представляет собой необходимое условие для разработки теста на эффективность.
Для оценки действия бегеломаба на первичные Т-клетки мононуклеарные клетки, полученные из периферической крови, инкубировали с бегеломабом при трех различных условиях (6 мкг/мл; 30 мкг/мл и 150 мкг/мл, 4×10-8 М, 2×10-7 М и 1×10-6 М, соответственно) в течение 6 и 24 часов при 37°С.После инкубации клетки немедленно метили для оценки экспрессии в них CD26. В качестве контроля мононуклеарные клетки инкубировали с самой высокой концентрацией бегеломаба (150 мкг/мл, т.е. 1×10-6 М) при 4°С.После 6 часов обнаружено ощутимое, хотя не значимое, уменьшение уровней интернализации CD26 (RFI) по сравнению с базовыми уровнями CD26 в CD3+, CD4+ и CD8+ Т-клетках (Фиг. 3, блок В). Тем не менее, статистически значимые результаты (*, Р<0,05) получены после 24 часов, в частности, для CD8+ Т-клеток (Фиг. 3, блок С). Как ранее обнаружено для клеточной линии Karpas 299, действие понижающей регуляции CD26 уменьшается тогда, когда мононуклеарные клетки инкубируют с бегеломабом при 4°С (Фиг. 3, блок D), свидетельствуя о явлении активного блокирования интернализации комплекса антитело-СХ)26. Кратко, после 24 часов инкубации с бегеломабом экспрессия CD26 уменьшается до 43,8% в клетках Karpas 299 при обработке концентрациями 30 мкг/мл (2×10-7 М) бегеломаба, и это явление зависит от дозы антитела против CD26 (Фиг. 5). Как ожидалось, при 4°С не было обнаружено никаких изменений экспрессии CD26 на поверхности анализируемых клеток. Полученные результаты ясно указывают на то, что рецептор CD26 интернализуется в ответ на связывание с бегеломабом с последующим устранением критической активации сигнала для субпопуляции CD26+ Т-клеток. Высвобождение провоспалительных цитокинов
Учитывая, что интернализация CD26 может обладать различными последующими воздействиями на Т-клеточную активацию, уровни некоторых провоспалительных цитокинов оценивали после обработки бегеломабом в первичных Т-клетках и клетках Karpas 299.
Воспалительные цитокины могут быть классифицированы на две группы: группа 1) вовлеченная в острое воспаление; группа 2) ответственная за хроническое воспаление. Цитокины, такие как IL-1, TNF-α, IL-6, IL-11, IL-8 и другие хемокины, G-CSF и GM-CSF относятся к первой группе. Вторая группа может быть дополнительно разделена на цитокины, которые опосредуют гуморальный ответ, такие как IL-4, IL-5, IL-6, IL-7 и IL-13, и цитокины, которые опосредуют клеточный ответ, такие как IL-1, IL-2, IL-3, IL-4, IL-7, IL-9, IL-10, IL-12, интерфероны, трансформирующий фактор роста Р и фактор некроза опухоли α и β (Shaikh PZ, 2011).
Провоспалительные цитокины представляют собой молекулы, вовлеченные в ряд аутоиммунных заболеваний, таких как ревматоидный артрит, рассеянный склероз, GvHD. В патогенезе этих заболеваний CD26 играет ключевую роль, особенно в воспалительном процессе. По этой причине оценивали ингибирование некоторых провоспалительных цитокинов бегеломабом. Более конкретно, анализировали уровни следующих цитокинов: интерлейкин-2 (IL-2), IL-8, IL-1β, колониестимулирующий фактор гранулоцитов-макрофагов (GM-CSF), интерферон гамма (IFN-γ), IL-6, IL-10, фактор некроза опухоли альфа (TNF-α) и IL-12p70. Серии разведений образцов супернатанта осуществляли в качестве предварительного теста в данном анализе, и результаты показали, что неразбавленные образцы находятся в диапазоне стандартной кривой для всех цитокинов, за исключением супернатантов клеток IL-12p70, на которые воздействовали IL-12 в качестве сигнала (не показано). По этой причине 25 мкл неразведенного супернатанта стандарта тестировали в анализе MSD. В обоих экспериментах стандартные кривые для всех цитокинов во всех анализируемых планшетах демонстрировали хорошо калиброванные кривые.
Анализируемые образцы наносили на стандартные графики для определения того, попадают ли они в диапазон обнаружения. Уровни всех цитокинов, за исключением IL-12p70, измеряли и анализировали, хотя изменения уровней цитокинов обнаруживали между первичными Т-клеточными донорами и Т-клеточной линией Karpas 299 с уровнями ниже уровня обнаружения в некоторых условиях. Уровни IL-1β и IL-6 были очень низкими во всех анализируемых клеточных популяциях в обоих экспериментах. Обработка клеток IL-12 очевидно вызывает продуцирование IL-1β, IFN-γ, IL-6 и IL-10 при множестве состояний, как ранее сообщалось в литературе (Vacaflores A. et al., 2016). Ингибирование продукции цитокинов после обработки бегеломабом обнаруживали таким образом для IL-2, IL-8, IL-1β, колониестимулирующого фактора гранулоцитов-макрофагов (GM-CSF), IL-6 и фактора некроза опухоли альфа (TNF-α) (Фиг. 4).
Значительное уменьшение уровней IL-8 и IL-1β после инкубации с бегеломабом обнаружено в тех же самых условиях, как в клетках Karpas 299, так и в первичных Т-клетках.
В следующей таблице 2 предложен обзор ингибирующего действия бегеломаба на секрецию различных цитокинов в первичных Т-клетках и клетках Karpas 299, демонстрируя, в частности, ингибирование уровней провоспалительных цитокинов, вызванное бегеломабом после инкубации клеток при 37°С в течение 6 или 24 часов.

Кроме того, для тестирования эффекта "покрывания" в отношении секреции цитокинов клетки тестировали на планшетах, покрытых и не покрытых антителом против CD3/антителом против CD28, и обрабатывали IL-12, бегеломабом или соответствующим контрольным разбавителем, как описано в разделе "Материалы и методы". В общем, обнаружено, что уровни цитокинов не обнаруживаются или оказываются меньше в не покрытых планшетах по сравнению с покрытыми планшетами. Последнее свидетельствует о том, что покрывание важно для разработки аналитического окна для исследования действия бегеломаба при определении уровней цитокинов. Следовательно, эффект бегеломаба в отношении измерения уровней цитокинов с использованием не покрытых планшетов далее не анализировали. На основе этого эксперимента полученные результаты показали, что бегеломаб способен значительно уменьшать продукцию специфических цитокинов, таких как IL-8, IL-1β и IL-6, как в первичных Т-клетках, так и в клетках Karpas 299, особенно подчеркивая механизм действия антитела против CD26 (бегеломаб) при лечении аутоиммунных заболеваний.
ПРИМЕР 2: Клеточный анализ для определения эффективности лигандов антитела против CD26
Задача настоящей экспериментальной части заключается в том, чтобы разработать клеточный анализ активности лигандов антитела против CD26, начиная с известного моноклонального антитела против CD26 бегеломаба.
Бегеломаб представляет собой моноклональное антитело против CD26, продуцируемое клетками гибридомы, клиническая мишень которых заключается в ингибировании активации Т-лимфоцитов донора в аллогенных трансплантатах.
Задача анализа заключается в том, чтобы измерить вариацию в уровнях CD26, присутствующих на клеточной мембране человеческих Т-лимфоцитов (Karpas 299, CD4-положительные), после обработки последних in vitro увеличивающимися концентрациями бегеломаба.
CD26 представляет собой мембранный гликопротеин, вовлеченный в активацию Т-лимфоцитов путем дипептидил-пептидазной активности. Предварительные данные демонстрируют способность бегеломаба вызывать интернализацию CD26 на поверхности человеческих Т-лимфоцитов и клеток Karpas 299 после 24-часовой обработки.
В частности, клетки инкубируют при 37°С в течение 24 часов увеличивающимися концентрациями бегеломаба и затем окрашивают двумя антителами, способными соответственно связывать рецептор CD4 и CD26 и конъюгированными с флуорофорами, которые излучают при различных длинах волн. Во время анализа FACS для каждой тестированной концентрации бегеломаба интенсивность флуоресценции измеряют пропорционально количеству CD26, присутствующих на клеточной поверхности каждой единичной клетки. Этот параметр затем количественно определяют при помощи прибора путем расчета медианы для 20000 единичных событий регистрации флуоресцентного сигнала, выбранных на популяции CD4-положительных клеток.
Протокол окрашивания клеточных образцов и анализ FACS После окончания заранее установленного периода инкубации с бегеломабом клеточные образцы обрабатывают в соответствии со следующим протоколом окрашивания антителами, конъюгированными с флуорофорами, и затем анализируют посредством FACS для экспрессии поверхностного CD26:
- перенос содержимого каждой лунки в пробирки Eppendorf объемом 1,5 мл;
- центрифугирование при 10000 об/мин в течение 5 минут;
- промывание осадка 1 мл охлажденного DPBS;
- центрифугирование при 10000 об/мин в течение 5 минут;
- сушка осадка и окрашивание следующими антителами в соответствии с протоколом инкубации для анализа FACS, как описано ниже в таблице 3.

- перемешивание образца на вортексе и инкубация при к.т.в течение 30 минут в темноте;
- добавление 1 мл DPBS и центрифугирование при 10000 об/мин в течение 5 минут;
- удаление супернатанта и промывание осадка 1 мл охлажденного DPBS 1X;
- центрифугирование при 10000 об/мин в течение 5 минут;
- ресуспендирование клеточного осадка в концентрации 0,7-1Е6 клеток/мл в DPBS-5% FBS и хранение образцов в темноте при +4 °С до визуализации посредством FACS. Если визуализация не может быть осуществлена в течение суток, клетки фиксируют путем добавления параформальдегида до конечной концентрации 0,36%.
Во время анализа FACS для каждой тестируемой концентрации бегеломаба интенсивность флуоресценции измеряют относительно количества CD26, присутствующих на клеточной поверхности каждой единичной CD4-положительной клетки. Этот параметр затем количественно определяют при помощи прибора в качестве средней величины, получаемой путем регистрации флуоресценции для 20000 событий, выбранных среди популяции CD4-положительных клеток. Для регистрации двух флуоресценций, ассоциированных с FITC и АРС, нет необходимости осуществлять компенсирующую активность, поскольку два флуорофора излучают при различных длинах волн.
Данные затем обрабатывают с использованием программы BD FACS Diva, которая дает возможность рассчитать уровень экспрессии CD26 как среднюю величину для всех регистрируемых событий (выбранных в популяции CD4-положительньгх клеток).
РЕЗУЛЬТАТЫ
Биологическую активность бегеломаба измеряли в виде способности вызывать интернализацию CD26, представленного на поверхности клеточной мембраны клеток Karpas 299.
Количество интернализованного CD26 после обработки увеличивающимися концентрациями бегеломаба измеряли в виде уменьшения средней величины флуоресценции, излучаемой специфическим антителом (АРС мышиное антитело против человеческого CD26, клон М-А261 RUO BD Pharmigen code 563670), связанным с CD26, экспрессированным на поверхности клеточной мембраны.
Задача заключается в том, чтобы построить кривые зависимости ответа от концентрации для каждой партии продукта для сравнения с кривой зависимости ответа от концентрации, получаемой путем обработки клеток рабочим стандартом (WS) при помощи сравнительного теста для оценки эффективности лота относительно WS, при котором параллельность кривых является необходимым условием приемлемости анализа (тест параллельности, осуществляемый при помощи программного обеспечения PLA.3 Analysis).
Как видно из графика на Фиг. 5, после 24 часов обработки бегеломабом происходит уменьшение флуоресценции, ассоциированное с CD26, на приблизительно 40% для клеток Karpas 299.
Кроме того, этот эффект в клетках Karpas 299 оказывается зависит от концентрации, представляя собой условие, рассматриваемое как важное для разработки клеточного анализа.
Этот обнаруженный эффект является специфическим для бегеломаба, поскольку пул IgG2b (класс IgG, к которому относится бегеломаб) не вызывает интернализацию CD26 при любых тестируемых условиях.
Для достижения оптимальных условий роста для оценки биологического действия было принято решение повторить инкубацию с бегеломабом в течение 24 часов и использовать CD3+ Т-лимфоциты после стимуляции с использованием Dynabeads Human T-Activator CD3/CD28 (Gibco kit, Life Technologies, код 11131D).
% интернализации CD26
Как видно из графика на Фиг. 6, после 24 часов обработки бегеломабом обнаруживается уменьшение флуоресценции, ассоциированное с CD26, на приблизительно 48% для клеток Karpas 299 после обработки самой высокой концентрацией бегеломаба (эквивалентной 150 мкг/мл или 1×10-6 М).
Удлинение кривой зависимости ответа от концентрации обеспечивает возможность достижения более низкого предела ответа, который равен 0,01 мкг/мл (7×l0-11 М).
Отсутствие эффекта пула IgG2b (используемого в качестве отрицательного контроля) было подтверждено.
Последующие анализы разрабатывали для уменьшения сложности кривой (для верификации того, имеет ли кривая монофазную или многофазную картину).
Оценка эффективности в различных образцах бегеломаба
Затем осуществляли оценку эффективности для трех образцов бегеломаба, один из которых хранили в условиях ускоренного старения при 32,5°С в течение 6 месяцев (препарат SVI-STB).
Исходя из анализа FACS, отсутствуют существенные различия между двумя независимыми препаратами партии 1 и партии 2 с точки зрения кинетики интернализации CD26.
Препарат SVI-STB, образец бегеломаба, подвергнутый деградации путем инкубации при 32,5°С в течение 6 месяцев, наоборот, по-видимому менее эффективен чем другие в индуцировании интернализации CD26 (Фиг. 7). Этот результат подтверждает возможность данного способа улавливать измененные образцы бегеломаба.
Как видно из графика на Фиг. 7, восемь выбранных концентраций бегеломаба демонстрируют высокую вариабельность в центральной области кривой образца бегеломаба, хранимого при 4°С, и образца бегеломаба, который претерпевает температурный сдвиг до 32,5°С в течение продолжительных периодов времени.
Два образца бегеломаба, оба из которых хранили при оптимальных условиях при 4°С, демонстрировали отсутствие вариабельности в центральной области кривой.
Оценка эффективности с использованием различных проточных цитофлуориметров
Для подтверждения того, что результаты, полученные для интернализации CD26 после инкубации с антителом против С26, также могут быть воспроизведены путем изменения технологии визуализации, авторы изобретения осуществили эксперименты по интернализации с использованием двух цитофлуориметров различных производителей.
Полученные результаты (Фиг. 8) ясно демонстрируют, что технология, относящаяся к различным цитофлуориметрам, не изменяет результаты кривых интернализации.
В частности, в обоих экспериментах образец бегеломаба, который хранили при неоптимальных условиях (SVI-STB), легко может быть идентифицирован, демонстрируя высокую вариабельность в центральной части кривой по сравнению с кривой двух других образцов бегеломаба, хранимых при оптимальных условиях.
ВЫВОДЫ
Проведенные экспериментальные тесты продемонстрировали путем использования клеточной линии Karpas 299, что клеточный анализ, основанный на индуцированной бегеломабом интернализации CD26 мембраной, обладает всеми характеристиками для того, чтобы адаптировать функционал тестирования выпускаемой партии лекарственного средства для клинического применения.
В частности, тест, основанный на интернализации CD26 после инкубации с бегеломабом, обеспечивает возможность идентификации специфическим и воспроизводимым образом любых возможных отклонений в характеристиках лекарственного средства, производимого для клинического применения.
ПРИМЕР 3: Валидация анализа на эффективность при использовании других антител против CD26
Анализ на эффективность тестировали с использованием двух других антител против CD26:
- мышиное mAb против hCD26 (клон 202.36) Thermo Fisher кат.: МА1-35147; Lot: 311030F;
- IgG2b, мышиный изотип K (клон MG2b-57) BioLegend кат.: М1395 в качестве отрицательного контроля.
Для обнаружения CD26 в мембране использовали мышиное антитело против человеческого CD26, конъюгированное с АРС (клон М-А261) BD Pharmingen. Кат.: 563670; Lotto: 8116645.
На Фиг. 9 и 10 показана относительная эффективность, обнаруженная относительно бегеломаба (WS-BEG-013). Исходя из полученных результатов, клон антитела 202.36 демонстрирует относительную эффективность приблизительно 0,01, тогда как клон антитела MG2b-57 демонстрирует полное отсутствие активности. Как можно наблюдать, тест на эффективность в соответствии с настоящим изобретением также может быть применим в отношении других лигандов CD26.
Кроме того, на Фиг. 9 при сравнении интернализации CD26 при концентрации, близкой к достижению асимптоты для референсного соединения WS-BEG-013 (2 мкг/мл), максимальная интернализация CD26 более 30% наблюдается только для WS-BEG-013 (32,5%), тогда как для специфически тестируемого антитела клон 202.36 составляет гораздо ниже 20% (11,5%), подтверждая в соответствии с обозначенными критериями, что оно представляет собой антитело с низкой эффективностью, т.е. менее 20% (то есть менее 50% плато в %).
На Фиг. 11 проиллюстрированы величины эффективности для тестируемого образца (TEST), рассчитанные относительно референсного стандарта (RS), для демонстрации того, как относительная активность может быть количественно измерена относительно бегеломаба. В этом случае референсный стандарт (RS) представляет собой бегеломаб в концентрации 1,8 мг/мл, тогда как тестируемый образец (TEST) представляет собой бегеломаб в концентрации 1,26 мг/мл, то есть 70% относительно концентрации RS, при ожидаемой относительной эффективности RS, равной отношению концентраций TEST/RS (например, 1,26/1,8 = 0,7, то есть 70%). На Фиг. 11 продемонстрировано то, как при отношении концентраций 70% обнаруживается относительная эффективность 70% как ожидалось (расчет, осуществляемый в соответствии с руководствами Фармакопеи США USP 1034 "Analysis of Biological Assays" mediante software PLA 3.0).
На графике на Фиг. 11 величина MFI 170 представляет максимальную величину экспрессии мембранного CD26 (максимальная горизонтальная асимптота), тогда как величина 110 представляет величину MFI, при которой достигается максимальная интернализация CD26, и поэтому величина, соответствующая минимальной мембранной экспрессии (минимальная горизонтальная асимптота). В этом диапазоне, определяемом в виде абсолютной величины для каждого анализа, оценки эффективности осуществляют путем измерения относительного расстояния кривой для тестируемого соединения от референсной кривой для бегеломаба.
Гистограммы, продемонстрированные на графике, демонстрируют процентную долю интернализации CD26 относительно его максимальной мембранной экспрессии, представленной в виде максимальной измеренной величины MFI (170). Эта процентная доля при сравнении с тестируемым соединением связана с относительной эффективностью:
- при величине MFI 130 RS интернализует 23,1% CD26, присутствующего в мембране, относительно тестируемого образца (TEST), который интернализует только 18,5%) (отношение тестируемый образец/референсный стандарт = 0,80);
- при величине MFI ПО (минимальное присутствие мембранного CD26) RS интернализует 12,1% CD26, присутствующего в мембране, относительно тестируемого образца, который интернализует только 9,9% (отношение тестируемый образец/референсный стандарт = 0,82).
Можно отметить, что величина эффективности не просто представляет собой отношение процентных долей интернализации в один момент времени, а рассчитывается как относительная величина между двумя ЕС50 (средние эффективные концентрации) 0,04342 мкг/мл для референсного стандарта и 0,06644 мкг/мл для тестируемого образца, соответственно.
Эффективности, рассчитываемые в интересуемом примере, фактически могут быть оценены как ЕС50 RS/EC50 Test, с получением величины, равной 0,65, т.е. способной подтверждать, что эффективности тестируемого образца составляют приблизительно 70% относительно эффективности RS.
Кроме того, путем сравнения интернализации CD26 в концентрации, близкой к достижению минимальной горизонтальной асимптоты для RS (2 мкг/мл), максимальная интернализация CD26 более 30% обнаруживается как для RS (36%), так и для тестируемого образца (35,9%), подтверждая в соответствии с обозначенными критериями, что в обоих случаях имеется высокая степень эффективности, т.е. более 30%.
СПИСОК ЛИТЕРАТУРЫ
- Abbott С.A., Baker Е., Sutherland G.R., McCaughan G.W. Immunogenetics 1994 40: 331-338.
- Cordero OJ, Yang CP, Bell EB. Immunobiology 2007, 212:85-94.
- Morimoto C, Torimoto Y., Levinson G., Rudd C.E., Schrieber M., Dang N.H., LetvinN.L. J. Immunol. 1989, 143: 3430-3439.
- Mattern Т., Scholz W., Feller A.C, Flad H.-D., Ulme, A.J. Scand. J. Immunol. 1991, 33:737-48.
- Gorrel M.D., Gysbers V., Mccaughan G. W. Scand. J. Immunol. 2001, 54: 249-264.
- De Meester I., Korom S., Van Damme J., Scharpe' S. Immunol. Today 1999, 20:367-375.
- Hildebrandt M., Reutter W., Arck P., Rose M., Klapp B.F. Clin. Sci. 2000, 99:93-104.
- Franco R., Valenzuela A., Lluis C, Blanco J Immunol. Rev. 1998, 161:27-42.
- Sauer A.V., Brigida I., Carriglio N., Aiuti A. Front Immunol. 2012, 3: 265.
- Morimoto C, Schlossman S.F. Immunol. Rev. 1998, 161:55-70.
- Klemann C, Wagner L., Stephan M., von Horsten S. Clin Exp Immunol. 2016, 185(1):1-21.
- Ohnuma K., Hosono O., Dang NH., Morimoto C. Adv Clin Chem. 2011, 53:51-84.
- Ferrara JLM, Levine JE, Reddy P and Holler E. Lancet 2009, 373:1550-1561.
- Welniak LA, Blazar BR and Murphy WJ. Annu Rev Immunol 2007, 25:139-170.
- Henden A.S., Hill GR. J Immunol 2015; 194:4604-4612.
- Yi Т., Chen Y., Wang L.,Du G., Huang D., Zhao D., Johnston H., Young J., Todorov I., Umetsu D.T., Chen L., Iwakura Y., Kandeel F., Forman S., Zeng D. Blood. 2009, 114(14):3101-12.
- Hatano R., Ohnuma K., Yamamoto J., Dang N.H., Yamada Т., Morimoto C. Br J Haematol. 2013, 162(2):263-77.
- Bacigalupo A., Deeg J., Caballero D., Gualandi F., Raiola A.M., Varaldo R., Di Grazia C, Van Lint M.T. Abstract 671. ASH Meeting December 2016.
- Brevetto US 9,376,498.
- Shaikh P. Z. Int. J. of Pharm. & Life Sci. (IJPLS) 2011, 2(11): 1247-1263.
- Vacaflores, A., Chapman, N. M., Harty, J. Т., Richer, M. J., & Houtman, J. C. PLoS One. 2016, 9;11(6):e0157175.
- Seidel U.J.E., SchlegelP., Lang P. Front Immunol. 2013, 4: 76.
| название | год | авторы | номер документа |
|---|---|---|---|
| РАЗМНОЖЕНИЕ И ПРИМЕНЕНИЕ НЕГЕМОПОЭТИЧЕСКИХ ТКАНЕРЕЗИДЕНТНЫХ γδ Т-КЛЕТОК | 2016 |
|
RU2784566C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРЕДОТВРАЩЕНИЯ ИЛИ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, ОПОСРЕДОВАННЫХ РЕГУЛЯТОРНЫМИ Т-КЛЕТКАМИ | 2017 |
|
RU2727900C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ T-КЛЕТОК ДЛЯ T-КЛЕТОЧНОЙ ТЕРАПИИ | 2020 |
|
RU2834232C1 |
| FC-ОПТИМИЗИРОВАННЫЕ АНТИТЕЛА К CD25 ДЛЯ ИСТОЩЕНИЯ ОПУХОЛЕСПЕЦИФИЧЕСКИХ КЛЕТОК | 2018 |
|
RU2839380C1 |
| ПЕПТИДЫ СО СПОСОБНОСТЬЮ СВЯЗЫВАТЬСЯ СО СКУРФИНОМ И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2502741C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОПУЛЯЦИИ CD4+CD25+Foxp3+ Т-ЛИМФОЦИТОВ ЧЕЛОВЕКА ex vivo, СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ | 2008 |
|
RU2391401C2 |
| СПОСОБ ОБОГАЩЕНИЯ РЕГУЛЯТОРНЫХ CD4CD25FOXP3T-КЛЕТОК ЧЕЛОВЕКА ex vivo | 2010 |
|
RU2437933C1 |
| ТЕРАПЕВТИЧЕСКИЕ АГЕНТЫ И АУТОИММУННЫЕ ЗАБОЛЕВАНИЯ | 1996 |
|
RU2203088C2 |
| ЛИГАНД GITR И СВЯЗАННЫЕ С ЛИГАНДОМ GITR МОЛЕКУЛЫ И АНТИТЕЛА И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2369636C2 |
| СПОСОБ ПОДАВЛЕНИЯ ВИЧ-ИНФИЦИРОВАННЫХ КЛЕТОК МЛЕКОПИТАЮЩЕГО И БЕЛКОВЫЙ РЕКОМБИНАНТНЫЙ РЕЦЕПТОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 1995 |
|
RU2165703C2 |








Настоящее изобретение относится к количественному клеточному способу определения in vitro действия лиганда антитела против CD26, предпочтительно моноклонального антитела против CD26, такого как бегеломаб. Способ определения in vitro эффективности лиганда антитела против CD26, включает следующие стадии: а) инкубацию при 37°С или при комнатной температуре популяции человеческих Т-лимфоцитов, экспрессирующих рецептор CD26, в процентной доле более 75% с лигандом антитела против CD26 в концентрации в диапазоне от 0,001 мкг/мл до 150 мкг/мл; б) инкубацию с антителом против антитела CD26, меченным флуорохромом, которое распознает эпитоп CD26, отличающийся от эпитопа, распознаваемого лигандом антитела против CD26, используемого на стадии а); в) определение величины MFI (медиана интенсивности флуоресценции) для CD26, измеренной для образца клеток, обработанных лигандом антитела против CD26 (MFIT), и величины MFI для необработанных клеток (MFINT) путем цитофлуориметрического анализа; г) оценку процентной доли интернализации рецептора CD26 (%int CD26) или RFI (относительная интенсивность флуоресценции), рассчитываемой в соответствии со следующей формулой: 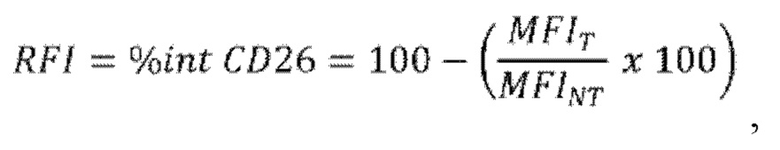 где: если величина %int CD26 составляет менее 20%, это указывает на низкую эффективность лиганда антитела против CD26; если находится в диапазоне от 20% до 30%, это указывает на среднюю эффективность лиганда антитела против CD26; если составляет более 30%, это указывает на высокую эффективность лиганда антитела против CD26. Техническим результатом является возможность количественного измерения как интернализации CD26, так и ингибирования секреции/продукции образующихся цитокинов. 10 з.п. ф-лы, 14 ил., 3 табл.
где: если величина %int CD26 составляет менее 20%, это указывает на низкую эффективность лиганда антитела против CD26; если находится в диапазоне от 20% до 30%, это указывает на среднюю эффективность лиганда антитела против CD26; если составляет более 30%, это указывает на высокую эффективность лиганда антитела против CD26. Техническим результатом является возможность количественного измерения как интернализации CD26, так и ингибирования секреции/продукции образующихся цитокинов. 10 з.п. ф-лы, 14 ил., 3 табл.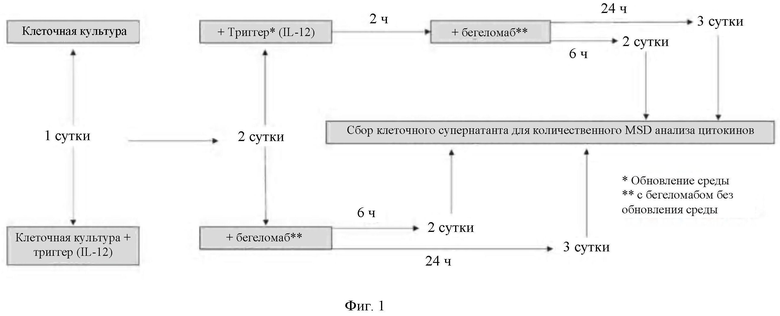
1. Способ определения in vitro эффективности лиганда антитела против CD26, включающий следующие стадии:
а) инкубацию при 37°С или при комнатной температуре популяции человеческих Т-лимфоцитов, экспрессирующих рецептор CD26, в процентной доле более 75% с лигандом антитела против CD26 в концентрации в диапазоне от 0,001 мкг/мл до 150 мкг/мл;
б) инкубацию с антителом против антитела CD26, меченным флуорохромом, которое распознает эпитоп CD26, отличающийся от эпитопа, распознаваемого лигандом антитела против CD26, используемого на стадии а);
в) определение величины MFI (медиана интенсивности флуоресценции) для CD26, измеренной для образца клеток, обработанных лигандом антитела против CD26 (MFIT), и величины MFI для необработанных клеток (MFINT) путем цитофлуориметрического анализа;
г) оценку процентной доли интернализации рецептора CD26 (%int CD26) или RFI (относительная интенсивность флуоресценции), рассчитываемой в соответствии со следующей формулой:

где, если величина %int CD26:
- составляет менее 20%, это указывает на низкую эффективность лиганда антитела против CD26;
- находится в диапазоне от 20% до 30%, это указывает на среднюю эффективность лиганда антитела против CD26;
- составляет более 30%, это указывает на высокую эффективность лиганда антитела против CD26.
2. Способ по п. 1, где указанная концентрация лиганда антитела против CD26 со стадии а) находится в диапазоне от 0,01 мкг/мл до 100 мкг/мл, более предпочтительно от 0,01 мкг/мл до 2 мкг/мл, еще более предпочтительно от 0,01 мкг/мл до 0,5 мкг/мл.
3. Способ по любому из пп. 1, 2, где лиганд антитела против CD26 со стадии а) представляет собой антитело или его фрагменты.
4. Способ по п. 3, где указанное антитело против CD26 представляет собой моноклональное антитело бегеломаб.
5. Способ по любому из пп. 1-4, где указанный флуорохром со стадии б) выбран из группы, состоящей из FITC (флуоресцеин изотиоцианат), АРС (аллофикоцианин), РЕ (фикоэритрин), РЕ-Су7, АРС-Н7, PerCP (перидинин-хлорофилл протеин) и РЕ-Су5.5, предпочтительно АРС.
6. Способ по любому из пп. 1-5, где популяция человеческих CD26+ Т-лимфоцитов со стадии а) выбрана из популяции первичных Т-лимфоцитов и опухолевой клеточной линии человеческих Т-лимфоцитов.
7. Способ по п. 6, где указанная опухолевая клеточная линия человеческих Т-лимфоцитов представляет собой клеточную линию Karpas 299.
8. Способ по любому из пп. 1-7, где цитофлуориметрический анализ со стадии в) осуществляют путем FACS-анализа (анализа сортировки клеток с активированной флуоресценцией).
9. Способ по любому из пп. 1-8, дополнительно включающий стадию анализа ингибирования продукции воспалительных цитокинов, выбранных из группы, состоящей из IL-8 (интерлейкин-8), IL-1β, IL-6, IL-2, GM-CSF (колониестимулирующий фактор гранулоцитов-макрофагов), IL-6 и TNF-α (фактор некроза опухоли альфа), осуществляемого на популяции человеческих CD26+ Т-лимфоцитов со стадии а).
10. Способ по п. 9, где указанное ингибирование продукции цитокинов оценивают путем анализа MesoScale Discovery.
11. Способ по п. 9 или 10, где указанная популяция человеческих CD26+ Т-лимфоцитов представляет собой клеточную линию Karpas 299.
| WO 2002092127 A1, 21.11.2002 | |||
| YAMADA, K., HAYASHI, M., MADOKORO, H., NISHIDA, H., DU, W., OHNUMA, K., YAMADA, T | |||
| Прибор для получения стереоскопических впечатлений от двух изображений различного масштаба | 1917 |
|
SU26A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Прибор для получения стереоскопических впечатлений от двух изображений различного масштаба | 1917 |
|
SU26A1 |

