ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] В данной международной заявке на патент заявляется приоритет по предварительной заявке на патент США № 62/067908, поданной 23 октября 2014 года, предварительной заявке на патент США № 62/148656, поданной 16 апреля 2015 года, предварительной заявке на патент США № 62/188353, поданной 2 июля 2015 года, а также предварительной заявке на патент США № 62/210795, поданной 27 августа 2015 года, содержание которых включено в данный документ посредством ссылки в полном объеме.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка подается вместе с Перечнем последовательностей в электронном формате. Перечень последовательностей предоставляется в виде файла под названием "Sequence_Listing_STP25.txt", созданного 30 сентября 2015 года, с последним изменением 22 октября 2015 года, размер которого составляет 83000 байт. Информация в электронном формате Перечня последовательностей включена в данный документ посредством ссылки в полном объеме.
УРОВЕНЬ ТЕХНИКИ
[0003] Использование однодоменных антител (ОдАт) в качестве отдельных антигенсвязывающих белков или в качестве антигенсвязывающего домена в большем белке или полипептиде предоставляет ряд существенных преимуществ по сравнению с использованием обычных антител или фрагментов антител. Преимущества ОдАт включают: только один домен необходим для связывания антигена с высокой аффинностью и с высокой селективностью; ОдАт могут экспрессироваться из одного гена и не требуют посттрансляционной модификации; ОдАт обладают высокой устойчивостью к теплу, рН, протеазам и другим денатурирующим средствам или условиям; получение ОдАт является недорогим; и ОдАт могут достигать мишени и эпитопов, недоступных для обычных антител.
[0004] Существует ряд заболеваний или патологических состояний, таких как рак, которые вызваны аберрантными внутриклеточными или трансмембранными компонентами, такими как нуклеотиды и белки. Устранение аберрантных компонентов может использоваться для профилактики или лечения данных заболеваний или патологических состояний. Существует ряд фармакологических соединений, доступных для лечения, но эти соединения могут быть неэффективными, не иметь возможности быть доставленными или быть токсичными для непораженных заболеванием клеток.
[0005] Другие способы лечения включают использование терапевтических белков или средств, которые содержат экзогенную последовательность-мишень, чтобы терапевтическое средство могло распознаваться рецепторами в клеточной мембране, позволяя терапевтическому средству проходить через клеточную мембрану и проникать в клетку. После того, как терапевтическое средство попадает внутрь клетки, данное терапевтическое средство может взаимодействовать с компонентом-мишенью, для лечения данного заболевания. Тем не менее, использование экзогенной последовательности-мишени может ограничивать тип клеток, который подвергается воздействию терапевтического средства, и увеличивает стоимость получения данного терапевтического средства.
[0006] По указанным выше причинам, существует потребность в композициях и способах лечения или профилактики заболевания, которые не полагаются на экзогенные последовательности-мишени или химические композиции для проникновения в клетку, и которые являются эффективными в направленном воздействии только на пораженные клетки в организме.
[0007] Настоящее изобретение относится к однодоменным антителам (ОдАт), белкам и полипептидам, содержащим данные ОдАт. Данные ОдАт направлены против внутриклеточных компонентов, которые вызывают патологическое состояние или заболевание. Данное изобретение также включает нуклеиновые кислоты, кодирующие ОдАт, белки и полипептиды, а также композиции, содержащие ОдАт. Данное изобретение включает использование композиций, ОдАт, белков или полипептидов для профилактических, терапевтических или диагностических целей. Данное изобретение также включает использование моноклональных антител, направленных на ОдАт по данному изобретению.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0008] Один вариант реализации данного изобретения представляет собой однодоменное антитело (ОдАт), направленное против внутриклеточного компонента. Внутриклеточным компонентом может являться, например, белок, нуклеиновая кислота, липид, углевод, STAT1, STAT2, STAT3, STAT4, STAT5a, STAT5b, STAT6, ФНО-альфа и KRAS.
[0009] В другом варианте реализации настоящее изобретение направлено на ОдАт к STAT3. Необязательно, ОдАт к STAT3 содержит аминокислотную последовательность, приведенную в SEQ ID №: 3 или SEQ ID №: 4.
[0010] В другом варианте реализации данное изобретение направлено на выделенный полипептид, содержащий аминокислотную последовательность, кодирующую ОдАт к STAT, такой как, например, полипептид, приведенный в SEQ ID №: 3 или SEQ ID №: 4.
[0011] В еще одном варианте реализации данное изобретение направлено на клетку- хозяина, и данная клетка-хозяин экспрессирует аминокислотную последовательность ОдАт, такую как, например, аминокислотная последовательность, приведенная в SEQ ID №: 3 или SEQ ID №: 4.
[0012] Один из вариантов реализации данного изобретения представляет собой фармацевтическую композицию, содержащую ОдАт или полипептид, а также фармацевтически приемлемый носитель. Необязательно, ОдАт включает ОдАт к STAT3, содержащее аминокислотную последовательность, приведенную в SEQ ID №: 3 или SEQ ID №: 4, и полипептид включает выделенный полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID №: 3 или SEQ ID №: 4.
[0013] Другим вариантом реализации данного изобретения является способ диагностирования заболевания, опосредованного STAT3 у субъекта, при этом данный способ включает этапы, в которых: а) биологический образец приводят в контакт с ОдАт или полипептидом; б) определяют количество STAT3 в биологическом образце; и в) сравнивают количество, определенное на этапе (б), с эталонным, при этом разница в количестве указывает на наличие расстройства.
[0014] Другим вариантом реализации данного изобретения является способ профилактики или лечения заболевания или расстройства, или профилактики рецидива заболевания, опосредованного STAT3, или для применения в лечении рака или заболеваний, вызванных аномальной клеточной пролиферацией, включающий введение ОдАт к STAT3 или полипептида субъекту, нуждающемуся в этом. Необязательно, ОдАт включает ОдАт к STAT3, содержащее аминокислотную последовательность, приведенную в SEQ ID №: 3 или SEQ ID №: 4, и полипептид включает выделенный полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID №: 3 или SEQ ID №: 4.
[0015] Один вариант реализации данного изобретения представляет собой ОдАт к ФНО-альфа. Необязательно, ОдАт к ФНО-альфа содержит аминокислотную последовательность, приведенную в SEQ ID №: 5, SEQ ID №: 6 или SEQ ID №: 7. Данное изобретение также относится к выделенному полипептиду, содержащему аминокислотную последовательность, приведенную в SEQ ID №: 5, SEQ ID №: 6 или SEQ ID №: 7.
[0016] Другим вариантом реализации данного изобретения является клетка-хозяин, экспрессирующая аминокислотную последовательность, приведенную в SEQ ID №: 5, SEQ ID №: 6 или SEQ ID №: 7.
[0017] В другом варианте реализации данное изобретение также представляет собой фармацевтическую композицию, содержащую ОдАт или полипептид, а также фармацевтически приемлемый носитель. Необязательно, ОдАт включает ОдАт к ФНО- альфа, содержащее аминокислотную последовательность, приведенную в SEQ ID №: 5, SEQ ID №: 6 или SEQ ID №: 7, и полипептид включает выделенный полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID №: 5, SEQ ID №: 6 или SEQ ID №: 7.
[0018] Другим вариантом реализации данного изобретения является способ диагностирования расстройства, опосредованного ФНО-альфа у субъекта, при этом данный способ включает этапы, в которых: а) биологический образец приводят в контакт с ОдАт или полипептидом; б) определяют количество ФНО-альфа в биологическом образце; и в) сравнивают количество, определенное на этапе (б), с эталонным, при этом разница в количестве указывает на наличие заболевания.
[0019] В одном варианте реализации данное изобретение описывает способ профилактики или лечения заболевания или расстройства, или рецидива заболевания или расстройства, опосредованного ФНО-альфа, или для применения в лечении рака или заболеваний, вызванных аномальной клеточной пролиферацией, включающий введение ОдАт к ФНО-альфа или полипептида млекопитающему, нуждающемуся в этом. Необязательно, ОдАт к ФНО-альфа содержит аминокислотную последовательность, приведенную в SEQ ID №: 5, SEQ ID №: 6 или SEQ ID №: 7, и полипептид включает выделенный полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID №: 5, SEQ ID №: 6 или SEQ ID №: 7.
[0020] Один вариант реализации данного изобретения представляет собой ОдАт к KRAS. Необязательно, ОдАт к KRAS содержит аминокислотную последовательность, приведенную в SEQ ID №: 2. В одном аспекте данное изобретение относится к выделенному полипептиду, при этом данный выделенный полипептид содержит аминокислотную последовательность, приведенную в SEQ ID №: 2. В другом аспекте данное изобретение относится к клетке-хозяину, экспрессирующей аминокислотную последовательность, приведенную в SEQ ID №: 2.
[0021] Другой вариант реализации данного изобретения представляет собой фармацевтическую композицию, содержащую ОдАт или полипептид, а также фармацевтически приемлемый носитель. Необязательно, ОдАт включает ОдАт к KRAS, содержащее аминокислотную последовательность, приведенную в SEQ ID №: 2, и полипептид включает выделенный полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID №: 2.
[0022] Дополнительным вариантом реализации данного изобретения является способ диагностирования расстройства, опосредованного KRAS у субъекта, при этом данный способ включает этапы, в которых: а) биологический образец приводят в контакт с ОдАт или полипептидом; б) определяют количество KRAS в указанном биологическом образце; и в) сравнивают количество, определенное на этапе (б), с эталонным, при этом разница в количестве указывает на наличие расстройства. Необязательно, ОдАт включает ОдАт к KRAS, содержащее аминокислотную последовательность, приведенную в SEQ ID №: 2, и полипептид включает выделенный полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID №: 2.
[0023] Данное изобретение также относится к способу лечения заболевания с использованием ОдАт к KRAS, при этом данный способ включает введение эффективного количества ОдАт к KRAS субъекту, нуждающемуся в этом.
[0024] В одном варианте реализации данное изобретение описывает способ профилактики или лечения заболевания или расстройства, или рецидива заболевания или расстройства, опосредованного KRAS, или для применения в лечении рака или заболеваний, вызванных аномальной клеточной пролиферацией, включающий введение ОдАт к KRAS или полипептида млекопитающему, нуждающемуся в этом. Необязательно, ОдАт к KRAS содержит аминокислотную последовательность, приведенную в SEQ ID №: 2, и полипептид включает выделенный полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID №: 2.
[0025] В одном варианте реализации данное изобретение описывает способ введения ОдАт по данному изобретению, при этом данный способ включает внутривенное введение, внутримышечное введение, пероральное введение, ректальное введение, внутриглазное введение, энтеральное введение, парентеральное введение, подкожное введение, чрескожное введение, введение в виде глазных капель, введение в виде назального спрея, введение посредством ингаляции или небулайзера, местное введение и введение в виде имплантируемого лекарственного средства.
[0026] В другом варианте реализации данное изобретение описывает способ лечения заболевания, профилактики заболевания или профилактики рецидива заболевания с использованием ОдАт по данному изобретению в комбинации с одним или более соединениями. Необязательно, одно или более соединений представляют собой ингибитор транскрипции.
[0027] В другом варианте реализации данное изобретение описывает способ измерения уровней ОдАт, при этом данный способ включает этапы, в которых: а) создают моноклональное антитело мыши, направленное против одного или более доменов ОдАт; б) проводят иммунологический анализ для определения количества ОдАт у субъекта; и в) выполняют количественный анализ ОдАт у субъекта.
ГРАФИЧЕСКИЕ МАТЕРИАЛЫ
[0028] Эти и другие признаки, аспекты и преимущества настоящего изобретения станут более понятными с учетом следующего описания, прилагаемой формулы изобретения и прилагаемых графических материалов, в которых:
На Фиг. 1 проиллюстрирована схематическая карта экспрессионного вектора pTT21-stt VHH13 VHH13-ОдАт к STAT3;
На Фиг. 2 проиллюстрирована схематическая карта экспрессионного вектора pTT21-stt VHH14 VHH14-ОдАт к STAT3;
На Фиг. 3 проиллюстрированы результаты анализа иммунопреципитации с использованием полученного в бактерии VHH13 (SEQ ID №: 3) к STAT3 и полученного в бактерии VHH14 (SEQ ID №: 4) к STAT3;
На Фиг. 4 проиллюстрированы результаты анализа иммунопреципитации с использованием полученного в бактерии VHH13 (SEQ ID №: 3) к STAT3;
На Фиг. 5 проиллюстрировано ингибирование роста полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в ксенотрансплантатной модели MDA-MB-231 при дозе, составляющей 0,5 мг/кг/день;
На Фиг. 6 проиллюстрировано ингибирование роста полученного в бактерии VHH13-ОдАт (SEQ ID № 3) к STAT3 в ксенотрансплантатной модели MDA-MB-231 при дозах, находящихся в пределах от 1 мг/кг дважды в день до 2 мг/кг дважды в день или 2 мг/кг/день;
На Фиг. 7 проиллюстрировано ингибирование роста полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в ксенотрансплантатной модели MDA-MB-231 при дозе, составляющей 5 мг/кг/дважды в день;
На Фиг. 8 проиллюстрировано ингибирование роста полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в ксенотрансплантатной модели DU145 при дозе, составляющей 5 мг/кг/дважды в день;
На Фиг. 9 проиллюстрировано ингибирование роста полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в ксенотрансплантатной модели PANC-1 при дозе, составляющей 5 мг/кг/дважды в день;
На Фиг. 10 проиллюстрировано ингибирование роста полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в ксенотрансплантатной модели MCF-7 при дозе, составляющей 1 мг/кг/трижды в день;
На Фиг. 11 проиллюстрировано ингибирование роста полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в ксенотрансплантатной модели BT-474 при дозе, составляющей 1 мг/кг/трижды в день;
На Фиг. 12 проиллюстрирована цитотоксичность ФНО-альфа в клетках U937;
На Фиг. 13 проиллюстрирована цитотоксичность стауроспорина в клетках U937; и
На Фиг. 14 проиллюстрировано ингибирование цитотоксичности ФНО-альфа с помощью ОдАт к ФНО-альфа.
ОПИСАНИЕ
[0029] При использовании по тексту данного документа, следующие термины и их варианты имеют значения, указанные ниже, если контекстом, в котором используется этот термин, явно не подразумевается иное значение.
[0030] Термины в единственном числе и аналогичные референты, используемые в данном документе, следует толковать как охватывающие как единственное, так и множественное число, если их использование в контексте не указывает на иное.
[0031] Термин "антигенная детерминанта" означает эпитоп на антигене, распознаваемый антигенсвязывающей молекулой (такой как ОдАт или полипептид по данному изобретению), и более конкретно - антигенсвязывающим участком антигенсвязывающей молекулы. Термины "антигенная детерминанта" и "эпитоп" также могут использоваться взаимозаменяемо. Считается, что аминокислотная последовательность, которая может связываться с конкретной антигенной детерминантой, эпитопом, антигеном или белком, обладает аффинностью и/или специфичностью к ним, "против" или "направлена против" антигенной детерминанты, эпитопа, антигена или белка.
[0032] При использовании по тексту данного документа, термин "включать" и варианты данного термина, такие как "включающий" и "включает", не предназначены для исключения других добавок, компонентов, целых чисел или этапов.
[0033] Предполагается, что ОдАт, полипептиды и белки, описанные в данном документе, могут содержать так называемые "консервативные" аминокислотные замены, которые могут быть в целом описаны как аминокислотные замены, в которых аминокислотный остаток заменен другим аминокислотным остатком с аналогичной химической структурой, и которые имеют незначительное или, по сути, не имеют никакого влияния на функцию, активность или другие биологические свойства полипептида. Консервативные аминокислотные замены хорошо известны в данной области техники. Консервативные замены представляют собой замены, в которых одна аминокислота в следующих группах (а)-(д) заменена другой аминокислотой в пределах той же группы: (а) малые алифатические, неполярные или слегка полярные остатки: Ala, Ser, Thr, Pro и Gly; (б) полярные, отрицательно заряженные остатки и их (незаряженные) амиды: Asp, Asn, Glu и Gln; (в) полярные, положительно заряженные остатки: His, Arg и Lys; (г) большие алифатические, неполярные остатки: Met, Leu, Ile, Val и Cys; и (д) ароматические остатки: Phe, Tyr и Trp. Другие консервативные замены включают: Ala на Gly или на Ser; Arg на Lys; Asn на Gln или на His; Asp на Glu; Cys на Ser; Gln на Asn; Glu на Asp; Gly на Ala или на Pro; His на Asn или на Gln; Ile на Leu или на Val; Leu на Ile или на Val; Lys на Arg, на Gln или на Glu; Met на Leu, на Tyr или на Ile; Phe на Met, на Leu или на Tyr; Ser на Thr; Thr на Ser; Trp на Tyr; Tyr на Trp; и/или Phe на Val, на Ile или на Leu.
[0034] Термин "домен", при использовании по тексту данного документа, как правило, означает глобулярную область цепи антитела и, в частности, глобулярную область тяжелой цепи антитела, или полипептид, который, по сути, состоит из такой глобулярной области.
[0035] Аминокислотная последовательность и структура ОдАт, как правило, состоит из четырех каркасных областей или "FR", которые называют "каркасной областью 1" или "FR1"; "каркасной областью 2" или "FR2"; "каркасной областью 3" или "FR3"; и "каркасной областью 4" или "FR4", соответственно. Каркасные области прерываются тремя определяющими комплементарность областями или "CDR", которые называют "определяющей комплементарность областью 1" или "CDR1"; "определяющей комплементарность областью 2" или "CDR2"; и "определяющей комплементарность областью 3" или "CDR3", соответственно.
[0036] При использовании по тексту данного документа, термин "гуманизированное ОдАт" означает ОдАт, в котором один или более аминокислотных остатков в аминокислотной последовательности природной последовательности VHH были заменены одним или более аминокислотными остатками, которые встречаются в соответствующем положении в домене VH из обычного 4-цепочечного антитела человека. Это может быть осуществлено с помощью способов, которые хорошо известны в данной области техники. Например, FR ОдАт могут быть заменены вариабельными FR человека.
[0037] При использовании по тексту данного документа, "выделенная" нуклеиновая кислота или аминокислота была отделена по меньшей мере от одного другого компонента, с которым она обычно связана, такого как ее источник или среда, другая нуклеиновая кислота, другой белок/полипептид, другой биологический компонент или макромолекула, или контаминант, примесь или второстепенный компонент.
[0038] Термин "млекопитающее" означает особь, принадлежащую к классу млекопитающих, и включает, не ограничиваясь ими, людей, домашних и сельскохозяйственных животных, а также зоопарковых, спортивных и домашних животных, таких как коровы, лошади, овцы, собаки и кошки.
[0039] При использовании по тексту данного документа, предполагается, что термин "фармацевтически приемлемый носитель" включает любые растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические и замедляющие абсорбцию средства, и т.п., совместимые с фармацевтическим введением. Подходящие носители описаны в последнем издании Remington's Pharmaceutical Sciences, стандартном справочнике в данной области. Предпочтительные примеры таких носителей или разбавителей включают, не ограничиваясь ими, воду, солевой раствор, растворы Рингера, раствор декстрозы, ФСБ (фосфатно-солевой буферный раствор) и 5% сывороточный альбумин человека. Также могут использоваться липосомы, катионные липиды и неводные носители, такие как нелетучие масла. Применение таких сред и средств для фармацевтически активных веществ хорошо известно в данной области техники. Их использование в композиции по данному изобретению предполагается за исключением тех случаев, когда обычная среда или средство являются несовместимыми с терапевтическим средством, описанным выше.
[0040] Термин "количественный иммунологический анализ" означает любые средства измерения количества антигена, присутствующего в образце, с использованием антитела. Методы выполнения количественных иммунологических анализов включают, не ограничиваясь ими, твердофазный иммуноферментный анализ (ELISA), анализ посредством специфического мечения анализируемого вещества с повторным захватом (SALRA), жидкостную хроматографию, масс-спектрометрию, сортировку клеток с активированной флуоресценцией и тому подобное.
[0041] Термин "раствор" означает композицию, содержащую растворитель и растворенное вещество, и включает истинные растворы и суспензии. Примеры растворов включают твердые, жидкие или газообразные вещества, растворенные в жидкости, и твердые частицы или мицеллы, взвешенные в жидкости.
[0042] Термин "специфичность" включает ряд различных видов антигенов или антигенных детерминант, с которыми конкретная антигенсвязывающая молекула или антигенсвязывающая белковая молекула могут связываться. Специфичность антигенсвязывающего белка можно определить на основе аффинности и/или авидности. Аффинность, выраженная константой равновесия при диссоциации антигена и антигенсвязывающего белка (KD), является показателем силы связывания между антигенной детерминантой и антигенсвязывающим участком на антигенсвязывающем белке: чем меньше значение KD, тем сильнее сила связывания между антигенной детерминантой и антигенсвязывающей молекулой (в альтернативном варианте, аффинность также может быть выражена в виде аффинной константы (KA), которая представляет собой 1/KD). Как будет понятно специалистам в данной области техники, аффинность можно определить в зависимости от конкретного рассматриваемого антигена. Авидность является показателем силы связывания между антигенсвязывающей молекулой и антигеном. Авидность относится как к аффинности между антигенной детерминантой и ее антигенсвязывающим участком на антигенсвязывающей молекуле, так и к количеству соответствующих участков связывания, присутствующих на антигенсвязывающей молекуле. Специфическое связывание антигенсвязывающего белка с антигеном или антигенной детерминантой можно определить с помощью любых известных методов, таких как, например, анализ Скэтчарда и/или анализы конкурентного связывания, такие как радиоиммунологические анализы (РИА), иммуноферментные анализы (ИФА) и конкурентные сэндвич-анализы.
[0043] При использовании по тексту данного документа, термин "рекомбинантный" относится к применению методов генной инженерии (например, клонирования и амплификации), применяемых для получения ОдАт по данному изобретению.
[0044] Термин "однодоменное антитело", "ОдАт" или "VHH" может, как правило, означать полипептид или белок, содержащий аминокислотную последовательность, которая состоит из четырех каркасных областей, прерываемых тремя определяющими комплементарность областями. Это представлено в виде FR1-CDR1-FR2-CDR2-FR3- CDR3-FR4. ОдАт по данному изобретению также включает полипептид или белок, который содержит аминокислотную последовательность ОдАт. Как правило, ОдАт продуцируются у представителей семейства верблюдовых, таких как ламы, но также могут быть синтезированы с помощью методов, которые хорошо известны в данной области техники. При использовании по тексту данного документа, вариабельные домены, присутствующие в природных антителах, содержащих тяжелые цепи, также называются "доменами VHH", чтобы их можно было отличить от вариабельных доменов тяжелой цепи, которые присутствуют в обычных 4-цепочечных антителах, называемых "доменами VH", и от вариабельных доменов легкой цепи, которые присутствуют в обычных 4-цепочечных антителах, называемых "доменами VL". Термины "VHH" и "ОдАт" используются в данном документе взаимозаменяемо. Нумерация аминокислотных остатков ОдАт или полипептида осуществляется в соответствии с общей нумерацией для доменов VH, изложенной Kabat et al. ("Sequence of proteins of immunological interest", Министерство здравоохранения США, NIH Бетесда, штат Мэриленд, публикация № 91). В соответствии с этой нумерацией, FR1 ОдАт содержит аминокислотные остатки в положениях 1-30, CDR1 ОдАт содержит аминокислотные остатки в положениях 31-36, FR2 ОдАт содержит аминокислоты в положениях 36-49, CDR2 ОдАт содержит аминокислотные остатки в положениях 50-65, FR3 ОдАт содержит аминокислотные остатки в положениях 66-94, CDR3 ОдАт содержит аминокислотные остатки в положениях 95-102 и FR4 ОдАт содержит аминокислотные остатки в положениях 103-113.
[0045] Термин "синтетический" означает продуцирование посредством химического или ферментативного синтеза in vitro.
[0046] Термин "мишень", при использовании по тексту данного документа, означает любой компонент, антиген или фрагмент, который распознается ОдАт. Термин "внутриклеточная мишень" означает любой компонент, антиген или фрагмент, присутствующий внутри клетки. "Трансмембранная мишень" представляет собой компонент, антиген или фрагмент, который расположен внутри клеточной мембраны. "Внеклеточная мишень" означает компонент, антиген или фрагмент, который находится вне клетки.
[0047] Термин "терапевтическая композиция", при использовании по тексту данного документа, означает вещество, которое предполагает наличие терапевтического эффекта, такое как фармацевтические композиции, генетические материалы, биопрепараты и другие вещества. Генетические материалы включают вещества, предполагающие наличие прямого или косвенного генетического терапевтического эффекта, такие как генетические векторы, генетические регуляторные элементы, генетические структурные элементы, ДНК, РНК и тому подобное. Биопрепараты включают вещества, которые представляют собой живую материю, или полученные из живой материи, предполагающей наличие терапевтического эффекта.
[0048] При использовании по тексту данного документа, словосочетания "терапевтически эффективное количество" и "профилактически эффективное количество" означают количество, которое обеспечивает терапевтический эффект в лечении, профилактике или ведении заболевания или выраженного симптома заболевания. Терапевтически эффективное количество может лечить заболевание или патологическое состояние, симптом заболевания или предрасположенность к заболеванию, с целью вылечить, исцелить, ослабить, облегчить, изменить, устранить, улучшить, нормализовать или повлиять на заболевание, симптомы заболевания или предрасположенность к заболеванию. Конкретное количество, которое является терапевтически эффективным, может быть легко определено обычным практикующим врачом, и может варьироваться в зависимости от факторов, известных в данной области техники, таких как, например, вид заболевания, анамнез и возраст пациента, стадия заболевания и введение других терапевтических средств.
[0049] Настоящее изобретение относится к однодоменным антителам (ОдАт), которые направлены против внутриклеточных компонентов, а также к белкам и полипептидам, содержащим ОдАт и нуклеотиды, кодирующие данные белки и полипептиды. Данное изобретение также может относиться к ОдАт, которые направлены против межклеточных, трансклеточных и внеклеточных мишеней или антигенов. Данное изобретение также включает нуклеиновые кислоты, кодирующие ОдАт, белки и полипептиды, а также композиции, содержащие ОдАт. Данное изобретение включает использование композиций, ОдАт, белков или полипептидов для профилактических, терапевтических или диагностических целей.
[0050] ОдАт имеют ряд уникальных структурных характеристик и функциональных свойств, которые делают ОдАт весьма целесообразными для использования в качестве функциональных антигенсвязывающих доменов или белков. ОдАт функционально связываются с антигеном при отсутствии вариабельного домена легкой цепи, и могут выполнять функцию отдельной, относительно малой, функциональной антигенсвязывающей структурной единицы, домена или белка. Это отличает ОдАт от доменов обычных антител, которые сами по себе не выполняют функцию антигенсвязывающего белка или домена, но должны объединяться с обычными фрагментами антител, такими как фрагменты Fab или фрагмент ScFv, чтобы связывать антиген.
[0051] ОдАт могут быть получены с помощью способов, которые хорошо известны в данной области техники. Например, один из способов получения ОдАт включает (а) иммунизацию верблюдовых одним или более антигенами, (б) выделение периферических лимфоцитов из иммунизированных верблюдовых, получение тотальной РНК и синтез соответствующих кДНК, (в) создание библиотеки фрагментов кДНК, кодирующих домены VHH, (г) транскрибирование кодирующих домен VHH кДНК, полученных на этапе (в), с мРНК с помощью ПЦР, преобразование мРНК в формат рибосомного дисплея и выбор домена VHH с помощью рибосомного дисплея, и (д) экспрессирование домена VHH в подходящий вектор и, необязательно, очистка экспрессированного домена VHH.
[0052] Другой способ получения ОдАт по данному изобретению осуществляется посредством приготовления нуклеиновой кислоты, кодирующей ОдАт, с помощью методов синтеза нуклеиновых кислот, с последующей экспрессией нуклеиновой кислоты in vivo или in vitro. Кроме того, ОдАт, полипептиды и белки по данному изобретению могут быть получены с помощью методов синтеза или полусинтеза для получения белков, полипептидов или других аминокислотных последовательностей.
[0053] ОдАт по данному изобретению, как правило, будут связываться со всеми природными или синтетическими аналогами, вариантами, мутантами, аллелями, частями и фрагментами мишени или по меньшей мере с теми аналогами, вариантами, мутантами, аллелями, частями и фрагментами мишени, которые содержат одну или более антигенных детерминант или эпитопов, которые по сути являются такими же, как антигенная детерминанта или эпитоп, с которыми ОдАт по данному изобретению связываются в мишени дикого типа. ОдАт по данному изобретению могут связываться с такими аналогами, вариантами, мутантами, аллелями, частями и фрагментами с аффинностью и/или специфичностью, которая является такой же, или которая выше или ниже, чем аффинность и специфичность, с которой ОдАт по данному изобретению связываются с мишенью дикого типа. Также в пределах объема данного изобретения предполагается, что ОдАт по данному изобретению связываются с некоторыми аналогами, вариантами, мутантами, аллелями, частями и фрагментами мишени, но не связываются с другими. Кроме того, ОдАт по данному изобретению может быть гуманизированным, и может быть одновалентным или поливалентным, и/или мультиспецифическим. Кроме того, ОдАт по данному изобретению могут связываться с фосфорилированной формой белка-мишени, а также с нефосфорилированной формой белка-мишени. ОдАт могут быть связаны с другими молекулами, такими как альбумин или другие макромолекулы.
[0054] Кроме того, в пределы объема данного изобретения входит то, что ОдАт являются поливалентными, то есть ОдАт может иметь два или более белков или полипептидов, которые направлены против двух или более различных эпитопов мишени. В таком поливалентном ОдАт белок или полипептид могут быть направлены, например, против таких же эпитопов, по сути, эквивалентных эпитопов или разных эпитопов. Разные эпитопы могут располагаться на одной мишени или они могут находиться на двух или более разных мишенях.
[0055] Также предполагается, что последовательность одного или более ОдАт по данному изобретению может быть соединена или связана с одной или более линкерными последовательностями. Линкер может представлять собой, например, последовательность белка, содержащую комбинацию серинов, глицинов и аланинов.
[0056] Кроме того, в пределы объема данного изобретения входит использование частей, фрагментов, аналогов, мутантов, вариантов, аллелей и/или производных ОдАт по данному изобретению, при условии, что они являются пригодными для описанных применений.
[0057] Поскольку ОдАт по данному изобретению главным образом предназначены для терапевтического и/или диагностического применения, они направлены против мишеней млекопитающих, предпочтительно человека. Тем не менее, возможно, что ОдАт, описанные в данном документе, вступают в перекрестную реакцию с мишенями других видов, например, с мишенями из одного или более других видов приматов или других животных (например, мыши, крысы, кролика, свиньи или собаки) и, в частности, в моделях на животных для заболеваний и расстройств, обусловленных заболеванием, связанным с данными мишенями.
[0058] В другом своем аспекте данное изобретение относится к нуклеиновой кислоте, которая кодирует ОдАт по данному изобретению. Такая нуклеиновая кислота может находиться, например, в виде генетической конструкции.
[0059] В другом аспекте данное изобретение относится к хозяину или клетке- хозяину, которая экспрессирует или может экспрессировать ОдАт по данному изобретению и/или которая содержит нуклеиновую кислоту, кодирующую ОдАт по данному изобретению. Последовательности ОдАт могут использоваться для инсерции в геном любого организма с созданием генетически модифицированного организма (ГМО). Примеры включают, не ограничиваясь ими, растения, бактерии, вирусы и животных.
[0060] Данное изобретение дополнительно относится к способам получения или создания ОдАт, нуклеиновых кислот, кодирующих ОдАт, клеток-хозяев, экспрессирующих или способных экспрессировать такие ОдАт, продуктов и композиций, содержащих ОдАт по данному изобретению.
[0061] Кроме того, данное изобретение относится к способам применения и использованию ОдАт, нуклеиновых кислот, кодирующих ОдАт, клеток-хозяев, продуктов и композиций, описанных в данном документе. Такой продукт или композиция может, например, представлять собой фармацевтическую композицию для лечения или профилактики заболевания, или представлять собой продукт или композицию, предназначенную для диагностики. ОдАт могут использоваться в различных анализах, например, анализах ELISA и анализах масс-спектрометрии для измерения уровней ОдАт в сыворотке крови и тканях.
[0062] В другом аспекте нуклеиновая кислота, кодирующая одно или более ОдАт по данному изобретению, может быть вставлена в геном организма для лечения или профилактики заболеваний.
[0063] Данное изобретение, как правило, относится к ОдАт, а также к белкам или полипептидам, содержащим или, по сути, состоящим из одного или более таких ОдАт, которые могут использоваться для профилактических, терапевтических и/или диагностических целей.
[0064] Способы и композиции, описанные в данном изобретении, могут применяться для лечения заболевания, описанного в данном документе, и могут применяться в любой дозировке и/или препарате, описанных в данном документе или известных по иным причинам, а также с любым способом введения, описанным в данном документе или по иным причинам известным специалисту в данной области техники.
[0065] ОдАт по данному изобретению, в частности, VHH к STAT3, VHH к KRAS и VHH к ФНО-альфа по данному изобретению, могут использоваться для лечения и профилактики злокачественных заболеваний, в том числе, не ограничиваясь ими: множественной миеломы, лейкозов (HTLV-1-зависимого, эритролейкоза, острого миелогенного лейкоза (ОМЛ), хронического миелогенного лейкоза (ХМЛ) и лейкоза из больших гранулярных лимфоцитов (ЛБГЛ)), лимфом (ВЭБ-ассоциированной/Беркитта, фунгоидного микоза, Т-клеточной лимфомы кожи, неходжкинской лимфомы (НХЛ), анапластической крупноклеточной лимфомы (АККЛ)), рака молочной железы, трижды негативного рака молочной железы, рака головы и шеи, меланомы, рака яичников, рака легкого, рака поджелудочной железы, рака предстательной железы, сарком, остеосаркомы, саркомы Капоши, саркомы Юинга, гепатоцеллюлярного рака, глиомы, нейробластомы, астроцитомы, рака ободочной и прямой кишки, опухолей Вильмса, рака почки, рака мочевого пузыря, рака эндометрия, рака шейки матки, рака пищевода, плоскоклеточного рака кожи, базальноклеточного рака и любых метастатических раковых заболеваний. ОдАт могут использоваться у больных раком, чтобы помочь предотвратить или уменьшить потерю веса или кахексию, обусловленные раком.
[0066] ОдАт, в частности, ОдАт к STAT3 и к ФНО-альфа по данному изобретению, также могут использоваться для лечения и профилактики заболеваний, таких как, не ограничиваясь ими: аутоиммунные заболевания (например, ревматоидный артрит, язвенный колит, болезнь Крона, бактериальный колит, астма, склеродермия, волчанка, энцефаломиелит, артериит, васкулит, гломерулонефрит, увеит, увеоретинит, рассеянный склероз), поликистоз почек, дерматологические заболевания (например, псориаз, очаговая алопеция, атопический дерматит, келоидные/гипертрофические рубцы, липома, болезнь Педжета и актинический кератоз), гнойный гидраденит, трансплантация (например, паренхиматозных органов, костного мозга, руки, лица, конечностей, любой части тела), мышечная дистрофия и мышечная атрофия, обусловленные раковыми заболеваниями и старением, эндометриоз, макулярная дегенерация, дегенерация сетчатки, инсульт, эпилепсия, черепно-мозговые травмы и повреждения спинного мозга, гипертензия, гипертрофия сердца, болезнь Альцгеймера, гипертензия легочной артерии, сахарный диабет 2 типа и анкилозирующий спондилит. Кроме того, ОдАт могут оказывать воздействие на орфанные заболевания. Примеры таких редких орфанных заболеваний включают, не ограничиваясь ими, трижды негативные раковые заболевания молочной железы, раковые заболевания поджелудочной железы, ОМЛ (острый миелоидный лейкоз), раковые заболевания головы и шеи, множественную миелому и химиоустойчивые раковые заболевания.
[0067] Вирусные инфекции можно лечить посредством направленного воздействия на внутриклеточные вирусные белки в инфицированных клетках. Вирусные белки, такие как обратная транскриптаза ВИЧ, могут блокировать жизненный цикл вируса. ОдАт по данному изобретению может также оказывать направленное воздействие на внутриклеточные вирусные белки, такие как VP24 вируса Эбола, и тем самым блокирует способность вируса Эбола останавливать противовирусный иммунный ответ хозяина. ОдАт по данному изобретению могут использоваться для воздействия на заболевания, при которых наблюдается сверхэкспрессия внутриклеточной молекулы. С помощью ОдАт можно лечить болезнь Хантингтона.
[0068] ОдАт по данному изобретению можно использовать с одним или более соединениями. Например, ОдАт по данному изобретению можно использовать с ингибиторами JAK/STAT, такими как, например, куркумин, ресвератрол, кукурбитацин A, B, E, I, Q, флавопиридол, дезокситетрангомицин, производные циклопентенона, N- ацил-гомосерин лактон, производные индирубина, меизоиндиго, тирфостин, содержащие платину соединения (например, IS3-295), пептидомиметики, антисмысловые олигонуклеотиды, S3I-201, производные фосфотирозин трипептида, ингибиторы протеазы ВИЧ (например, нелфинавир, индинавир, саквинавир и ритонавир), JSI-124, XpYL, Ac-pYLPQTV-NH2, ISS 610, CJ-1383, пириметамин, метформин, атипримод, S3I- M2001, STX-0119; производное N-[2-(1,3,4-оксадиазолил)]-4 хинолинкарбоксамида, S3I-1757, LY5; 5,8-диоксо-6(пиридин-3-иламино)-5,8-дигидро-нафталин-1-сульфонамид, витацинстин, статтик, STA-21, LLL-3, LLL12, XZH-5, SF-1066, SF-1087, 17o, криптотаншинон, FLL32, FLL62, C188-9, BP-1108 и BP-1075, галиеллалактон, JQ1, 5, 15 DPP, WP1066, никлозамид, SD1008, нифуроксазид, криптотаншинон, BBI хинон и руксолитиниба фосфат. Одно или более соединений могут увеличивать терапевтический эффект и увеличивать эффективность ОдАт по данному изобретению. Кроме того, эффективность ОдАт можно увеличить посредством объединения его с пептидами, пептидомиметиками и другими лекарственными средствами, такими как, например, не ограничиваясь ими, циметидин, аторвастатин, целекоксиб, метформин и циметидин. Кроме того, что касается лучевой терапии, ОдАт к STAT3 могут превращать радиорезистентные раковые заболевания в радиочувствительные раковые заболевания.
[0069] Также предполагается, что одно или более ОдАт по данному изобретению можно объединять или ОдАт по данному изобретению можно объединять с другими ОдАт.
[0070] Предполагается, что некоторые ОдАт по данному изобретению могут проходить через клеточную мембрану и проникать в клетку без дополнительного оказания направленного воздействия последовательностей белка на ОдАт и без экзогенных соединений, которые направляют ОдАт связываться с рецепторами клеточной поверхности и проходить через клеточную мембрану.
[0071] После прохождения через клеточную мембрану эти ОдАт могут оказывать направленное воздействие на трансмембранные или внутриклеточные молекулы или антигены. Эти внутриклеточные или трансмембранные мишени могут представлять собой, например, белки, углеводы, липиды, нуклеиновые кислоты, мутантные белки, вирусные белки и прионы. Мишени ОдАт могут выполнять функцию ферментов, структурных белков клетки, внутриклеточных участков молекул клеточных мембран, молекул в мембранах органелл, любого типа молекулы РНК, любых областей ДНК или хромосомы, метилированных или неметилированных нуклеиновых кислот, частично собранных молекул в рамках механизма синтеза клетки, молекул вторичных посредников и молекул в механизмах клеточной сигнализации. Мишени могут включать все молекулы в цитоплазме, ядре, органеллах и клеточной мембране. Направленное воздействие на молекулы, предназначенные для секреции или размещения в клеточной мембране, может быть осуществлено в цитоплазме до выхода из клетки.
[0072] Мишени ОдАт могут быть у людей, животных, растений, грибов, паразитов, простейших, бактерий, вирусов, прионов, прокариотических клеток и эукариотических клеток. Некоторыми примерами меж- и внутриклеточных сигнальных молекул и белковых групп, которые могут быть мишенями ОдАт по данному изобретению, являются: продукты онкогена, гормоны, цитокины, факторы роста, нейромедиаторы, киназы (в том числе тирозинкиназа, серинкиназа и треонинкиназа), фосфатазы, убиквитин, циклические нуклеотиды, циклазы (аденилат и гуанилат), G-белки, фосфодиэстеразы, суперсемейство ГТФаз, иммуноглобулины (антитела, фрагменты Fab, связующие вещества, ОдАт), суперсемейство иммуноглобулинов, инозитолфосфатные липиды, стероидные рецепторы, кальмодулин, группа CD (например, CD4, CD8, CD28 и т.д.), факторы транскрипции, ФНО-бета, ФНО-альфа и бета, суперсемейство лигандов ФНО, сигнальные молекулы рецепторов Notch, сигнальные молекулы рецепторов Hedgehog, сигнальные молекулы рецепторов Wnt, сигнальные молекулы толл-подобных рецепторов, каспазы, актин, миозин, миостатин, 12-липоксигеназа, 15-липоксигеназа, суперсемейство липоксигеназ, обратная транскриптаза, вирусы и их белки, амилоидные белки, коллаген, сопряженный с G-белком рецептор, мутантные нормальные белки, прионы, Ras, Raf, Myc, Src, BCR/ABL, MEK, Erk, Mos, Tpl2, MLK3, TAK, DLK, MKK, p38, MAPK, MEKK, ASK, SAPK, JNK, BMK, MAP, JAK, PI3K, циклооксигеназа, STAT1, STAT2, STAT3, STAT4, STAT5a, STAT5b, STAT6, Myc, p53, BRAF, NRAS, KRAS, HRAS и хемокины.
[0073] KRAS представляет собой гомолог онкогена Ras Кирстена из генов семейства Ras млекопитающих. KRAS кодирует белок, который является представителем небольшого суперсемейства ГТФ. Данный белок причастен к различным злокачественным опухолям, в том числе к аденокарциноме легкого, муцинозной аденоме, протоковой карциноме поджелудочной железы и карциноме толстой и прямой кишки. В нормальных условиях, представители семейства Ras влияют на клеточный рост и события дифференцировки в системе сигнализации на основе компартментализации внутриклеточной мембраны. Тем не менее, онкогенный Ras может дерегулировать процессы, которые регулируют пролиферацию клеток и апоптоз.
[0074] ОдАт к KRAS были разработаны для направленного воздействия на KRAS дикого типа и мутантный KRAS (G12D), чтобы блокировать его роль в злокачественных клетках, таких как, например, клетки, связанные с раком ободочной и прямой кишки, раком поджелудочной железы, раком желчных протоков, раком легких, лейкозами и другими метастатическими злокачественными опухолями. Не будучи связанными конкретным механизмом, полагают, что ОдАт к KRAS связывает KRAS и блокирует нисходящие сигналы KRAS в злокачественных клетках. Кроме того, ОдАт к KRAS может успешно лечить злокачественные опухоли, которые являются устойчивыми к анти-EGFR биопрепаратам (например, цетуксимабу и панитумумабу).
[0075] С помощью методов, которые хорошо известны в данной области, рекомбинантный мутантный белок KRAS (G12D) человека использовали для создания ОдАт, которые направлены против или могут связываться с эпитопом KRAS или мутантного KRAS (G12D), или других мутантов KRAS. Кроме того, ОдАт могут создаваться для других мутантов KRAS. Для создания ОдАт к KRAS, рекомбинантный полноразмерный KRAS (ID гена: 3845) человека экспрессировали в Escherichia coli.
[0076] Получали и скринировали несколько ОдАт. Последовательность ДНК одного ОдАт к KRAS (G12D), именуемая KRAS_13 (SEQ ID №: 1), показана ниже:
5'Gaggtgcagctggtggagtctgggggaggctcggtgcagactggagggtctctgagactctcctgtgcagtttctggaaatatcggc agcagctactgcatgggctggttccgccaggctccagggaagaagcgcgaggcggtcgcacgtattgtacgtgatggtgccactggcta cgcagactacgtgaagggccgattcaccatctcccgagacagcgccaagaacactctgtatctgcaaatgaacaggctgatacctgagg acactgccatctactactgtgcggcagacctgcccccaggttgtttgactcaggcgatttggaattttggttatcggggccagggaaccctg gtcaccgtctcctca-3'
[0077] Аминокислотная последовательность ОдАт (SEQ ID № 2) к KRAS (G12D), KRAS_13, показана ниже, с подчеркнутыми CDR:
EVQLVESGGGSVQTGGSLRLSCAVSGNIGSSYCMGWFRQAPGKKREAVA
RIVRDGATGYADYVKGRFTISRDSAKNTLYLQMNRLIPEDTAIYYCAA
DLPPGCLTQAIWNFGYRGQGTLVTVSS
[0078] Кроме того, настоящее изобретение включает одно или более мышиных моноклональных антител, которые направлены против одного или более доменов ОдАт к KRAS по данному изобретению. Мышиное моноклональное антитело можно создать с помощью методов, которые известны специалистам в данной области техники, например, мышиное моноклональное антитело можно получить с помощью мышиной гибридомы. Мышиное моноклональное антитело можно использовать в диагностических анализах, например, антитело можно использовать в иммунологическом анализе, таком как ELISA или масс-спектрометрический анализ, с целью измерения количества ОдАт к KRAS в сыворотке крови пациента. Цитотоксичность ОдАт KRAS (G12D) испытывали на клетках рака поджелудочной железы человека PANC-1, как описано ниже.
[0079] STAT3 является представителем семейства белков-переносчиков сигнала и активаторов транскрипции (STAT), которые осуществляют как сигнальную трансдукцию, так и активацию транскрипционных функций. STAT3 широко экспрессируется и активируется через фосфорилирование по тирозину и/или серину в качестве связывающего ДНК белка в ответ на различные цитокины и факторы роста, такие как ЭФР, ИЛ-6, ТФР, ИЛ-2 и Г-КСФ. Фосфопротеин STAT3 образует гомодимеры и гетеродимеры с другими представителями семейства STAT и транслоцируется в ядро с целью модуляции транскрипции различных генов, и в результате играет ключевую роль во многих клеточных процессах, таких как клеточный рост, апоптоз, ангиогенез, ускользание от иммунологического надзора и выживаемость.
[0080] ОдАт к STAT3 можно давать пациентам и другим организмам для лечения заболеваний, вызванных фосфорилированным и нефосфорилированным STAT3, а также для профилактики развития заболевания или рецидива заболевания. Например, пациенты, перенесшие трансплантацию органов и трансплантацию костного мозга, подвержены более высокому риску в отношении SCCA и BCCA кожи из-за иммуносупрессивных препаратов, которые они принимают. Введение ОдАт к STAT3 может уменьшать или устранять этот риск. На пациентов, получавших лечение от злокачественной опухоли, которые подвержены риску рецидива, лечение ОдАт к STAT3 окажет благоприятное действие. На основании семейного анамнеза и HLA-типа, некоторые индивидуумы будут подвергаться повышенному риску в отношении некоторых видов аутоиммунных заболеваний и могут получить пользу от лечения с использованием ОдАт со снижением риска развития этого аутоиммунного заболевания. Риск рака молочной железы можно уменьшить за счет ведения анти-STAT3 препаратов, таких как GLG-302, как показано в недавнем исследовании NCI.
[0081] Помимо ингибирования STAT3, ОдАт к STAT3 также может ингибировать STAT1, STAT2, STAT4, STAT5a, STAT5b и STAT6 благодаря высокой степени гомологии между этими молекулами.
[0082] Рекомбинантный белок STAT3 человека использовали для получения ОдАт к STAT, которые были направлены против или могли связываться с эпитопом STAT3. Для создания ОдАт к STAT3, рекомбинантный полноразмерный STAT3 (ID гена: 6774) человека экспрессировали бакуловирусом в клетках насекомых Sf9. ОдАт к STAT клонировали в векторы, которые могут экспрессироваться как в клетках бактерий, так и млекопитающих, как показано на Фиг. 1 и 2.
[0083] ОдАт к STAT3 по данному изобретению может использоваться для направленного воздействия на STAT3 и все другие молекулы STAT внутри клетки с целью ингибирования роста клеток, такого как, например, подавление роста раковых клеток. Кроме того, ОдАт к STAT3 может ингибировать рост клеток в других пролиферативных заболеваниях, таких как псориаз и макулярная дегенерация, через ФРЭС.
[0084] Не ограничиваясь конкретным механизмом действия, полагают, что ОдАт к STAT3 может устранять индуцированную раком супрессию иммунного ответа за счет снижения уровней STAT3 в антигенпрезентирующих клетках, таких, как, например, дендритные клетки хозяина. Ингибирование STAT3 способствует противораковому ответу врожденной и приобретенной иммунной системой пациента (т.е. дендритными клетками, макрофагами, нейтрофилами, Т-клетками, NK-клетками и В-клетками).
[0085] С помощью методов, которые хорошо известны в данной области техники, несколько ОдАт к STAT получали и подвергали скринингу на способность подавлять рост раковых клеток и индуцировать апоптоз в клеточных линиях рака, как описано ниже. Испытывали цитотоксичность и антипролиферативные активности ОдАт к STAT3. Кроме того, испытывали переносимость ОдАт к STAT3 in vitro и in vivo. Получение мышиных моноклональных антител, направленных против одного или более доменов ОдАт к STAT, описано ниже.
[0086] Аминокислотная последовательность одного ОдАт к STAT3, именуемого VHH13 (SEQ ID № 3), показана ниже:
HVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSGISTGGLI
TYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSRFDCYRGSWFNRYM
YNSWGQGTQVTVSS
Три CDR подчеркнуты.
[0087] Аминокислотная последовательность второго ОдАт к STAT3, именуемого VHH14 (SEQ ID № 4), показана ниже:
QVQLVESGGGSVQAGGSLRLSCVASTYTGCMGWFRQAPGKEREGVAALSSRGFAGH
YTDSVKGRFSISRDYVKNAVYLQMNTVKPEDAAMYYCAAREGWECGETWLDRTAG
GHTYWGQGTLVTVSS
[0088] И снова, три CDR подчеркнуты. Белковые последовательности других ОдАт к STAT3, которые были получены, являются такими, как показано ниже:
STAT3_10 (SEQ ID № 5):
(1) DVQLVESGGGSVQAGGSLRLSCVASTYTGCMGWFRQAPGKEREGVAA
(48) LSSRGFAGHYTDSVKGRFSISRDYVKNAVYLQMNTVKPEDAAMYYCAARE
(98) GWECGETWLDRTAGGHTYWGQGTQVTVSS STAT3_34 (SEQ ID № 6):
(1) DVQLVESGGGSVQAGGSLRLSCVASTYTGCMGWFRQAPGKEREGVAA
(48) LSSRGFAGHYTDSVKGRFSISRDYVKNAVYLQMNTVKPEDAAMYYCAARE
(98) GWECGETWLDRTAGGHTYWGQGTQVTVSS
STAT3_19 (SEQ ID № 7):
(1) HVQLVESGGGSVQAGGSLRLSCVASTYTGCMGWFRQAPGKEREGVAA
(48) LSSRGFAGHYTDSVKGRFSISRDYVKNAVYLQMNTVKPEDAAMYYCAARE
(98) GWECGETWLDRTAGGHTYWGQGTQVTVSS
STAT3_14 (SEQ ID № 8):
(1) QVQLVESGGGSVQAGGSLRLSCVASTYTGCMGWFRQAPGKEREGVAA
(48) LSSRGFAGHYTDSVKGRFSISRDYVKNAVYLQMNTVKPEDAAMYYCAARE
(98) GWECGETWLDRTAGGHTYWGQGTLVTVSS
STAT3_35 (SEQ ID № 9):
(1) QVQLVESGGGSVQAGGSLRLSCVASTYTGCMGWFRQAPGKEREGVAA
(48) LSSRGFAGHYTDSVKGRFSISRDYVKNAVYLQMNTVKPEDAAMYYCAARE
(98) GWECGETWLDRTAGGHTYWGQGTLVTVSS
STAT3_9 (SEQ ID № 10):
(1) QVQLVESGGGSVQAGGSLRLSCVASTYTGCMGWFRQAPGKEREGVAA
(48) LSSRGFAGHYTDSVKGRFSISRDYVKNAVYLQMNTVKPEDAAMYYCAARE
(98) GWECGETWLDRTAGGHTYWGQGTLVTVSS
STAT3_30 (SEQ ID № 11):
(1) QVQLVESGGGSVQAGGSLRLSCVASTYTGCMGWFRQAPGKEREGVAA
(48) LSSRGFAGHYTDSVKGRFSISRDYVKNAVYLQMNTVKPEDAAMYYCAARE
(98) GWECGETWLDRTAGGHTYWGQGTLVTVSS
STAT3_23 (SEQ ID № 12):
(1) QVQLVESGGGSVQAGGSLRLSCVASTYTGCMGWFRQAPGKEREGVAA
(48) LSSRGFAGHYTDSVKGRFSISRDYVKNAVYLQMNTVKPEDAAMYYCAARE
(98) GWECGETWLDRTAGSHTYWGQGTLVTVSS
STAT3_24 (SEQ ID № 13):
(1) EVQLVESGGGSVQAGGSLRLSCVASTYTGCMGWFRQAPGKEREGVAA
(48) LSSRGFAGHYTDSVKGRFSISRDYVKNAVYLQMNTVKPEDAAMYYCAARE
(98) GWECGETWLDRTAGGHTYWGQGTLVTVSS
STAT3_36 (SEQ ID № 14):
(1) DVQLVESGGGSVQAGDSLRLSCVASTYTGCMGWFRQAPGKEREGVAA
(48) LSSRGFAGHYTDSVKGRFSISRDYVKNAVYLQMNTVKPEDAAMYYCAARE
(98) GWECGETWLDRTAGGHTYWGQGTLVTVSS
STAT3_12 (SEQ ID № 15):
(1) QVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTLVTVSS
STAT3_16 (SEQ ID № 16):
(1) QVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTNNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTLVTVSS
STAT3_11 (SEQ ID № 17):
(1) EVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTLVTVSS
STAT3_20 (SEQ ID № 18):
(1) DVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTLVTVSS
STAT3_2 (SEQ ID № 19):
(1) DVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTLVTVSS
STAT3_15 (SEQ ID № 20):
(1) DVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTLVTVSS
STAT3_6 (SEQ ID № 21):
(1) HVQLVESEGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTLVTVSS
STAT3_33 (SEQ ID № 22):
(1) QVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTQVTVSS
STAT3_17 (SEQ ID № 23):
(1) QVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTQVTVSS
STAT3_25 (SEQ ID № 24):
(1) EVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMSSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTQVTVSS
STAT3_32 (SEQ ID № 25):
(1) DVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTQVTVSS
STAT3_13 (SEQ ID № 26):
(1) HVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTQVTVSS
STAT3_39 (SEQ ID № 27):
(1) HVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTQVTVSS
STAT3_4 (SEQ ID № 28):
(1) HVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTQVTVSS
STAT3_29 (SEQ ID № 29):
(1) HVQLVESGGGSVQAGGSLRLSCAASGANGGRSCMGWFRQVPGKEREGVSG
(51) ISTGGLITYYADSVKGRFTISQDNTKNTLYLQMNSLKPEDTAMYYCATSR
(101) FDCYRGSWFNRYMYNSWGQGTQVTVSS
[0089] Соответствующие последовательности ДНК ОдАт к STAT3 являются такими, как показано ниже:
Stat3_VHH-10 (SEQ ID № 30):
5'-gatgtgcagctggtggagtctgggggaggctcggtgcaggctggaggctctctgagactctcctgtgtagcctctacatacaccggc tgcatgggctggttccgccaggctcctggaaaggagcgcgagggagtcgcagctcttagtagccgtggttttgccgggcactataccgactccgtg aagggccgattctccatctcccgagactacgtcaagaatgcggtgtatctgcaaatgaacactgtgaaacctgaggacgctgccatgtactactgtgc agcacgggagggatgggagtgcggtgagacctggttggaccggaccgccgggggccatacctactggggccaggggacccaggtcaccgtct cctca-3'
Stat3_VHH-14 (SEQ ID № 31):
5'-caggtgcagctggtggagtctgggggaggctcggtgcaggctggaggctctctgagactctcctgtgtagcctctacatacaccggc tgcatgggctggttccgccaggctcctggaaaggagcgcgagggagtcgcagctcttagtagccgtggttttgccgggcactataccgactccgtg aagggccgattctccatctcccgagactacgtcaagaatgcggtgtatctgcaaatgaacactgtgaaacctgaggacgctgccatgtactactgtgc agcacgggagggatgggagtgcggtgagacctggttggaccggaccgccgggggccatacctactggggccaggggaccctggtcaccgtctc ctca-3'
Stat3_VHH-12 (SEQ ID № 32):
5'-caggtgcagctggtggagtctgggggaggctcggtgcaggctggagggtctctgagactctcctgtgcagcctctggagccaatgg tggtcggagctgcatgggctggttccgccaggttccagggaaggagcgcgagggggtttctggtatttcaaccggtggtcttattacatactatgccg actccgtgaagggccgattcaccatctcccaagacaacaccaagaacacgctgtatctgcaaatgaacagcctgaaacctgaggacactgccatgt actactgtgcgacgagtcggtttgactgctatagaggctcttggttcaaccgatatatgtataacagttggggccaggggaccctggtcaccgtctcct ca-3'
Stat3_VHH-13 (SEQ ID № 33):
5'-catgtgcagctggtggagtctgggggaggctcggtgcaggctggagggtctctgagactctcctgtgcagcctctggagccaacgg tggtcggagctgcatgggctggttccgccaggttccagggaaggagcgcgagggggtttctggtatttcaaccggtggtcttattacatactatgccg actccgtgaagggccgattcaccatctcccaagacaacaccaagaacacgctgtatctgcaaatgaacagcctgaaacctgaggacactgccatgt actactgtgcgacgagtcggtttgactgctatagaggctcttggttcaaccgatatatgtataacagttggggccaggggacccaggtcactgtctcct ca-3'
Stat3_VHH-20 (SEQ ID № 34):
5'-gatgtgcagctggtggagtctgggggaggctcggtgcaggctggagggtctctgagactctcctgtgcagcctctggagccaatgg tggtcggagctgcatgggctggttccgccaggttccagggaaggagcgcgagggggtttctggtatttcaaccggtggtcttattacatactatgccg actccgtgaagggccgattcaccatctcccaagacaacaccaagaacacgctgtatctgcaaatgaacagcctgaaacctgaggacactgccatgt actactgtgcgacgagtcggtttgactgctatagaggctcttggttcaaccgatatatgtataacagttggggccaggggaccctggtcaccgtctcct ca-3'
Stat3_VHH-23 (SEQ ID № 35):
5'-caggtgcagctggtggagtctgggggaggctcggtgcaggctggaggctctctgagactctcctgtgtagcctctacatacaccggc tgcatgggctggttccgccaggctcctggaaaggagcgcgagggagtcgcagctcttagcagccgtggttttgccgggcactataccgactccgtg aagggccgattctccatctcccgagactacgtcaagaatgcggtgtatctgcaaatgaacactgtgaaacctgaggacgctgccatgtactactgtgc agcacgggagggatgggagtgcggtgagacctggttggaccggaccgccgggagccatacctactggggccaggggaccctggtcaccgtctc ctca-3'
Stat3_VHH-24 (SEQ ID № 36):
5'-gaggtgcagctggtggagtctgggggaggctcggtgcaggctggaggctctctgagactctcctgtgtagcctctacatacaccgg ctgcatgggctggttccgccaggctcctggaaaggagcgcgagggagtcgcagctcttagtagccgtggttttgccgggcactataccgactccgt gaagggccgattctccatctcccgagactacgtcaagaatgcggtgtatctgcaaatgaacactgtgaaacctgaggacgctgccatgtactactgtg cagcacgggagggatgggagtgcggtgagacctggttggaccgaaccgccgggggccatacctactggggccaggggaccctggtcaccgtct cctca-3'
Stat3_VHH-25 (SEQ ID № 37):
5'-gaggtgcagctggtggagtctgggggaggctcggtgcaggctggagggtctctgagactctcctgtgcagcctctggagccaatg gtggtcggagctgcatgggctggttccgccaggttccagggaaggagcgcgagggggtttctggtatttcaaccggtggtcttattacatactatgcc gactccgtgaagggtcgattcaccatctcccaagacaacaccaagaacacgctgtatctgcaaatgagcagcctgaaacctgaggacactgccatg tactactgtgcgacgagtcggtttgactgctatagaggctcttggttcaaccgatatatgtataacagttggggccaggggacccaggtcaccgtctcc tca-3'
Stat3_VHH-19 (SEQ ID № 38):
5'-catgtgcagctggtggagtctggggggggctcggtgcaggctggaggctctctgagactctcctgtgtagcctctacatacaccggc tgcatgggctggttccgccaggctcctggaaaggagcgcgagggagtcgcagctcttagtagccgtggttttgccgggcactataccgactccgtg aagggccgattctccatctcccgagactacgtcaagaatgcggtgtatctgcaaatgaacactgtgaaacctgaggacgctgccatgtactactgtgc agcacgggagggatgggagtgcggtgagacctggttggaccggaccgccgggggccatacctactggggccaggggacccaggtcaccgtct cctca-3'
Stat3_VHH-32 (SEQ ID № 39):
5'-gatgtgcagctggtggagtctgggggaggctcggtgcaggctggagggtctctgagactctcctgtgcagcctctggagccaatg gtggtcggagctgcatgggctggttccgccaggttccagggaaggagcgcgagggggtttctggtatttcaaccggtggtcttattacatactatgcc gactccgtgaagggccgattcaccatctcccaagacaacaccaagaacacgctgtatctgcaaatgaacagcctgaaacctgaggacactgccatg tactactgtgcgacgagtcggtttgactgctatagaggctcttggttcaaccgatatatgtataacagttggggccaggggacccaggtcaccgtctcc tca-3'
Stat3_VHH-33 (SEQ ID № 40):
5'-caggtgcagctggtggagtctgggggaggctcggtgcaggctggagggtctctgagactctcctgtgcagcctctggagccaatg gtggtcggagctgcatgggctggttccgccaggttccagggaaggagcgcgagggggtttctggtatttcaaccggtggtcttattacatactatgcc gactccgtgaagggccgattcaccatctcccaagacaacaccaagaacacgctgtatctgcaaatgaacagcctgaaacctgaggacactgccatg tactactgtgcgacgagtcggtttgactgctatagaggctcttggttcaaccgatatatgtataacagttggggccaggggacccaggtcaccgtctcc tca-3'
Stat3_VHH-36 (SEQ ID № 41):
5'-gatgtgcagctggtggagtctgggggaggctcggtgcaggctggagactctctgagactctcctgtgtagcctctacatacaccggc tgcatgggctggttccgccaggctcctggaaaggagcgcgagggagtcgcagctcttagtagccgtggttttgccgggcactataccgactccgtg aagggccgattctccatctcccgagactacgtcaagaatgcggtgtatctgcaaatgaacactgtgaaacctgaggacgctgccatgtactactgtgc agcacgggagggatgggagtgcggtgagacctggttggaccggaccgccgggggccatacctactggggccaggggaccctggtcactgtctc ctca-3'
Stat3_VHH-11 (SEQ ID № 42):
5'-gtgcagctggtggagtctgggggaggctcggtgcaggctggagggtctctgagactctcctgtgcagcctctggagccaatggtg gtcggagctgcatgggctggttccgccaggttccagggaaggagcgtgagggggtttctggtatttcaaccggtggtcttattacatactatgccgac tccgtgaagggccgattcaccatctcccaagacaacaccaagaacacgctgtatctgcaaatgaacagcctgaaacctgaggacactgccatgtact actgtgcgacgagtcggtttgactgctatagaggctcttggttcaaccgatatatgtataacagttggggccaggggaccctggtcactgtctcctca-3'
Stat3_VHH-6 (SEQ ID № 43):
5'-gtgcagctggtggagtctgagggaggctcggtgcaggctggagggtctctgagactctcctgtgcagcctctggagccaatggtg gtcggagctgcatgggctggttccgccaggttccagggaaggagcgcgagggggtttctggtatttcaaccggtggtcttattacatactatgccgac tccgtgaagggccgattcaccatctcccaagacaacaccaagaacacgctgtatctgcaaatgaacagcctgaaacctgaggacactgccatgtact actgtgcgacgagtcggtttgactgctatagaggctcttggttcaaccgatatatgtataacagttggggccaggggaccctggtcaccgtctcctca-3'
Stat3_VHH-1 (SEQ ID № 44):
5'-gtgcagctggtggagtctgggggaggctcggtgcaggctggagggtctctgagactctcctgtgcagcctctggagccaatggtg gtcggagctgcatgggctggttccgccaggttccagggaaggagcgcgagggggtttctggtatttcaaccggtggtcttattacatactatgccgac tccgtgaagggccgattcaccatctcccaagacaacaccaataacacgctgtatctgcaaatgaacagcctgaaacctgaggacactgccatgtact actgtgcgacgagtcggtttgactgctatagaggctcttggttcaaccgatatatgtataacagttggggccaggggaccctggtcactgtctcctca-3'
[0090] Кроме того, настоящее изобретение включает одно или более мышиных моноклональных антител, которые направлены против одного или более доменов ОдАт к STAT3 по данному изобретению. Мышиное моноклональное антитело можно создать с помощью методов, которые известны специалистам в данной области техники, например, мышиное моноклональное антитело можно получить с помощью мышиной гибридомы. Мышиное моноклональное антитело можно использовать в диагностических анализах, например, антитело можно использовать в иммунологическом анализе, таком как ELISA, с целью измерения количества ОдАт к STAT3 в сыворотке крови пациента. Следует понимать, что данный способ не ограничивается ОдАт к STAT3, и может использоваться для получения мышиного антитела, направленного на любое из ОдАт по данному изобретению.
[0091] Ген ФНО-альфа кодирует многофункциональный провоспалительный цитокин, который принадлежит суперсемейству фактора некроза опухоли (ФНО). Этот цитокин в основном секретируется макрофагами. Данный цитокин участвует в регуляции широкого спектра биологических процессов, в том числе регуляции роста, дифференцировке, воспалении, вирусной репликации, онкогенезе, а также аутоиммунных заболеваниях; и в вирусных, бактериальных, грибковых и паразитарных инфекциях. К тому же было установлено, что индуцируя геморрагический некроз опухолей, ФНО участвует в онкогенезе, опухолевых метастазах, вирусной репликации, септическом шоке, лихорадке, воспалении, кахексии и аутоиммунных заболеваниях, включая болезнь Крона и ревматоидный артрит, а также реакцию "трансплантат против хозяина".
[0092] Настоящее изобретение относится к ОдАт, белкам и полипептидам, которые направлены против ФНО-альфа, в частности, против ФНО-альфа человека внутри клетки или клеточной мембраны так, чтобы предотвратить секрецию ФНО-альфа клетками.
[0093] Предполагается, что ОдАт к ФНО-альфа и полипептиды по данному изобретению можно использовать для профилактики и/или лечения заболеваний и расстройств, связанных с ФНО-альфа и/или опосредованных им, таких как воспаление, ревматоидный артрит, болезнь Крона, язвенный колит, синдром воспаленного кишечника, рассеянный склероз, болезнь Аддисона, аутоиммунный гепатит, аутоиммунный паротит, сахарный диабет 1 типа, эпидидимит, гломерулонефрит, болезнь Грейвса, синдром Гийена-Барре, болезнь Хашимото, гемолитическая анемия, системная красная волчанка, мужское бесплодие, рассеянный склероз, тяжелая миастения, пузырчатка, псориаз, ревматическая лихорадка, ревматоидный артрит, саркоидоз, склеродермия, синдром Шегрена, спондилоартропатии, тиреоидит, васкулит и потеря веса из-за рака и кахексии.
[0094] ФНО-альфа существует в различных формах; есть мономерные и мультимерные формы, в том числе тримерная форма. В пределы объема данного изобретения входит то, что ОдАт, белки и полипептиды по данному изобретению связываются с ФНО-альфа в его различных формах, т.е. мономерной форме или мультимерных формах. Таким образом, если ОдАт, белки и полипептиды по данному изобретению направлены на ФНО-альфа, следует понимать, что это также включает ОдАт, белки и полипептиды, направленные против ФНО-альфа в его тримерной форме.
[0095] Известно, что сигнальная трансдукция с помощью ФНО включает сшивание рецепторов ФНО тримером молекул ФНО, который содержит три участка связывания рецептора (см., например, Peppel et al., J. Exp. Med., 174 (1991), 1483-1489).
[0096] Рекомбинантный белок ФНО-альфа человека использовали для создания ОдАт, которые направлены против или могут связываться с эпитопом ФНО-альфа. Для создания ОдАт к ФНО-альфа, рекомбинантный полноразмерный ФНО-альфа (ID гена: 7124) человека экспрессировали в Escherichia coli и использовали в качестве антигена-мишени.
[0097] Было получено тридцать пять ОдАт к белку ФНО-альфа. Эти антитела к ФНО-альфа разделяли на три группы, основываясь на гомологии последовательностей.
[0098] Аминокислотная последовательность первого ОдАт к ФНО-альфа, именуемого VHH66-ОдАт (SEQ ID № 45) к ФНО-альфа, показана ниже:
HVQLVESGGGSVEAGGSLRLSCAASGFRYAAYCMGWFRQADGKEREGVATIDIDGLT THADSVKGRFTISRDNAKNTLSLQMNDLKPEDTAMYYCAADRDRCGSIWTYAYKYRG-QGTLVTVSS
Три CDR подчеркнуты.
[0099] Аминокислотная последовательность второго ОдАт к ФНО-альфа, именуемого VHH69-ОдАт (SEQ ID № 46) к ФНО-альфа, показана ниже:
EVQLVESGGGSVLAGGSLRLSCVASGFTSRYNYMAWFRQAPGKEREGVATIGTASGS ADYYGSVKDRFTISQDNAKNTVYLQMNSLKPEDTAMYYCAARTYGTISLTPSDYRYWGQGT LVTVSS
Три CDR подчеркнуты.
[0100] Аминокислотная последовательность третьего ОдАт к ФНО-альфа, именуемого VHH62-ОдАт (SEQ ID № 47) к ФНО-альфа, показана ниже:
QVQLVESGGGPVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCSWAQG TQGTLVTVSS
[0101] Три CDR подчеркнуты. Другие ОдАт к ФНО-альфа, которые были обнаружены, включают приведенные ниже последовательности, и снова CDR подчеркнуты:
ФНО_2 (SEQ ID № 48):
QVQLVESGGGSVEAGRSLRLSCAASGFRYAAYCMGWFRQADGKEREGVATIDIDGQT THADSVKGRFTISRDNAKNTLSLQMNDLKPEDTAMYYCAADRDRCGSIWTYAYKYRGQGTQ VTVSS
ФНО_46 (SEQ ID № 49):
QVQLVESGGGSVEAGGSLRLSCAASGFRYAAYCMGWFRQADGKEREGVATIDIDGQT THADSVKGRFTISRDNVKNTLSLQMNDLKPEDTAMYYCAADRDRCGSIWTYAYKYRGQGTQ VTVSS
ФНО_71 (SEQ ID № 50):
QVQLVESGGGSVEAGGSLRLSCAASGFRYAAYCMGWFRQADGKEREGVATIDIDGLT THADSVKGRFTISRDNAKNTLSLQMNDLKPEDTAMYYCAADRDRCGSIWTYAYKYRGQGTQ VTVSS
ФНО_21 (SEQ ID № 51):
QVQLVESGGGSVEAGGSLRLSCAASGFRYAAYCMGWFRQADGKEREGVATIDIDGQT THADSVKGRFTISRDNAKNTLSLQMNDLKPEDTAMYYCAADRDRCGSIWTYAYKYRGQGTQ VTVSS
ФНО_38 (SEQ ID № 52):
EVQLVESGGGSVEAGGSLRLSCAASGFRYAAYCMGWFRQADGKEREGVATIDIDGQT THADSVKGRFTISRDNAKNTLSLQMNDLKPEDTAMYYCAADRDRCGSIWTYAYKYRGQGTQ VTVSS
ФНО_18 (SEQ ID № 53):
EVQLVESGGGSVEAGGSLRLSCAASGFRYAAYCMGWFRQADGKEREGVATIDIDGLT THADSVKGRFTISRDNAKNTLSLQMNDLKPEDTAMYYCAADRDRCGSIWTYAYKYRGQGTL VTVSS
ФНО_37 (SEQ ID № 54):
DVQLVESGGGSVEAGGSLRLSCAASGFRYAAYCMGWFRQADGKEREGVATIDIDGQT THADSVKGRFTISRDNAKNTLSLQMNDLKPEDTAMYYCAADRDRCGSIWTYAYKYRGQGTL VTVSS
ФНО_66 (SEQ ID № 55):
HVQLVESGGGSVEAGGSLRLSCAASGFRYAAYCMGWFRQADGKEREGVATIDIDGLT THADSVKGRFTISRDNAKNTLSLQMNDLKPEDTAMYYCAADRDRCGSIWTYAYKYRGQGTL VTVSS
ФНО_68 (SEQ ID № 56):
HVQLVESGGGSVEAGGSLRLSCAASGFRYAAYCMGWFRQADGKEREGVATIDIDGLA THADSVKGRFTISRDNAKNTLSLQMNDLKPEDTAMYYCAADRDRCGSIWTYAYKYRGQGTL VTVSS ФНО_78 (SEQ ID № 57):
HVQLVESGGGSVEAGGSLRLSCAASGFRYAAYCMGWFRQADRKEREGVATIDIDGQT THADSVKGRFTISRDNAKNTLSLQMNDLKPEDTAMYYCAADRDRCGSIWTYAYKYRGQGTQ VTVSS
ФНО_67 (SEQ ID № 58):
HVQLVESGGGSVQAGGSLRLSCAASGFRYAAYCMGWFRQADGKVREGVATIDIDGQ TTHADSVKGRFTISRDNAKNTLSLQMNDLKPEDTAMYYCAADRDRCGSIWTYAYKYRGQGT LVTVSS
ФНО_6 (SEQ ID № 59):
QVQLVESGGGSVQAGGSLRLSCAASGFIDSFGVMAWFRQAPGKEREGVAAVYRRAG DTYYADSVKGRFTISRDNAKNTVYLQMNSLKPEDSAMYYCAARTYGSVSSWTGYKYWGQG TQVTVSS
ФНО_7 (SEQ ID № 60):
DVQLVESGGGSVQAGGSLRLSCAASGFIDSFGVMAWFRQTPGKEREGVAAVYRRAG DTYYADSVKGRFTISRDNAKNTVYLQMNSLKPEDSAMYYCAARTYGSVSSWTGYKYWGQG TQVTVSS
ФНО_13 (SEQ ID № 61):
DVQLVESGGGSVQVGGSLTLSCAVSGYTDSYGVMAWFRQAPGKEREGVASIYRNSGI TYYPDSVKGRFTISRDNAKNTVLLQMNSLKPEDSATYYCAVRSFGSVSTWAGYVYWGQGTQ VTVSS
ФНО_60 (SEQ ID № 62):
DVQLVESGGGSVQAGGSLRLSCAASGFIDSFGVMAWFRQAPGKEREGVAAVYRRAG DTYYADSVKGRFTISRDNAKNTVYLQMNSLKPEDSAMYYCAARTYGSVSSWTGYKYWGRG TQVTVSS
ФНО_73 (SEQ ID № 63):
DVQLVESGGGSVRAGGSLRLSCTASGDTSKSDCMAWFRQAPGKERERVGAIYTRNGY THYADSVNGRFTISQDNAKNALYLQMSGLKPEDTAMYYCAARFRIYGQCVEDDDIDYWGQG TLVTVSS
ФНО_69 (SEQ ID № 64):
EVQLVESGGGSVLAGGSLRLSCVASGFTSRYNYMAWFRQAPGKEREGVATIGTASGS ADYYGSVKDRFTISQDNAKNTVYLQMNSLKPEDTAMYYCAARTYGTISLTPSDYRYWGQGT LVTVSS
ФНО_76 (SEQ ID № 65):
QVQVVEYGGGSVQAGETVRLSCTASGFTFAEADMGWYRQAPGHEWELVSNITTEGIT SEASSSYADSVRGRFTIFDNAKNMVYLQMNSLKHEDTAVYYCAPDPYAYSTYREYCTWAQG TQGTLVTVSS
ФНО_62 (SEQ ID № 66):
QVQLVESGGGPVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCSWAQG TQGTLVTVSS
ФНО_43 (SEQ ID № 67):
QVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCTWAQG TQGTLVTVSS
ФНО_15 (SEQ ID № 68):
QVQPVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCTWAQG AQGTLVTVSS
ФНО_11 (SEQ ID № 69):
QVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCSWAQG TQGTQVTVSS
ФНО_17 (SEQ ID № 70):
QVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCTWAQG TQGTQVTVSS
ФНО_63 (SEQ ID № 71):
QVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCTWAQG TQGTLVTVSS
ФНО_20 (SEQ ID № 72):
HVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCTWAQG TQGTQVTVSS
ФНО_58 (SEQ ID № 73):
EVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCTWAQG TQGALVTVSS
ФНО_27 (SEQ ID № 74):
EVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCTWAQG TQGTLVTVSS
ФНО_28 (SEQ ID № 75):
EVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCSWAQG TQGTQVTVSS
ФНО_4 (SEQ ID № 76):
EVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCTWAQG TQGTQVTVSS
ФНО_14 (SEQ ID № 77):
DVQLVESRGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCTWAQG TQGTLVTVSS
ФНО_3 (SEQ ID № 78):
DVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHVCELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCSWAQG TQGTQVTVSS
ФНО_1 (SEQ ID № 79):
DVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGLECELVSTITTEGITS EASSYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSEYCTWAQGT QGTLVTVSS
ФНО_45 (SEQ ID № 80):
DVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS EASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCTWAQG TQGTLVTVSS
ФНО_22 (SEQ ID № 81):
DVQLVESGGGSVQAGETLRLSCTASGFTFAEADMGWYRQAPGHECELVSTITTEGITS VASSYYADSVRGRFTISRDNAKNMVYLQMNSLKPEDTAVYYCAPDPYAYSTYSDYCTWAQG TQGTQVTVSS
[0102] Испытывали ингибирование роста нескольких ОдАт к ФНО-альфа in vitro, как описано ниже. Кроме того, настоящее изобретение включает одно или более мышиных моноклональных антител, которые направлены против одного или более доменов ОдАт к ФНО-альфа по данному изобретению. Мышиное моноклональное антитело можно создать с помощью методов, которые известны специалистам в данной области техники, как описано ниже. Мышиное моноклональное антитело можно использовать в диагностических анализах, например, антитело можно использовать в иммунологическом анализе, таком как ELISA, с целью измерения количества ОдАт к ФНО-альфа в сыворотке крови пациента.
[0103] Белки RAF представляют собой семейство серин/треонин-специфических киназ, которые служат в качестве центрального промежуточного звена при передаче внеклеточных сигналов на митоген-активируемый протеинкиназный путь, который регулирует клеточный рост, дифференцировку и выживаемость. BRAF является представителем семейства RAF, который активируется представителями семейства Ras при индуцированной фактором роста стимуляции. Активный Ras может индуцировать гетеродимеризацию cRaf и BRAF, и это может объяснить наблюдаемый кооперативный эффект cRaf и BRaf в клетках, отвечающих на сигналы фактора роста. Активирующие мутации в гене BRAF присутствуют в большом проценте злокачественных меланом человека и пропорционально в раковых заболеваниях толстой кишки. Подавляющее большинство этих мутаций приводит к замене валина на глутаминовую кислоту в остатке 599 в сегменте активации BRAF.
[0104] ОдАт к BRAF были разработаны для направленного воздействия на BRAF дикого типа и мутантный BRAF, чтобы блокировать его роль в злокачественных клетках, таких как, например, клетки, связанные с раком толстой кишки и другими злокачественными опухолями.
[0105] С помощью методов, которые хорошо известны в данной области, рекомбинантный белок BRAF человека использовали для создания ОдАт, которые направлены против или могут связываться с эпитопом BRAF.
[0106] Кроме того, настоящее изобретение включает одно или более мышиных моноклональных антител, которые направлены против одного или более доменов ОдАт к BRAF по данному изобретению. Мышиное моноклональное антитело можно создать с помощью методов, которые известны специалистам в данной области техники. Мышиное моноклональное антитело можно использовать в диагностических анализах, например, антитело можно использовать в иммунологическом анализе, таком как ELISA, с целью измерения количества ОдАт к BRAF в сыворотке крови пациента.
ПРИМЕРЫ
ПРИМЕР 1: VHH13-ОДАТ (SEQ ID № 3) К STAT3 СВЯЗЫВАЕТ STAT3
[0107] В этом примере аффинность двух мишеней VHH к STAT3 измеряли с помощью безмаркерного анализа связывания на основе Octet. В данном анализе в качестве зондов-антигенов использовали VHH13-ОдАт (SEQ ID №: 3) к STAT3, к KRAS (отрицательный контроль) и GST-STAT3 (16 кДа одновалентный антиген, Creative BioMart № STAT3-1476H). Белок GST-STAT3 в концентрации 20 мкг/мл иммобилизовали в ФСБ с использованием аминопропилсилановых (APS) биосенсоров Dip and Read, специально предназначенных для гидрофобного белка. Затем зонды погружали в лунки с белком GST-STAT3, VHH13-ОдАт (SEQ ID №: 3) к STAT3 или к KRAS в указанной концентрации. Измеряли скорость ассоциации (скорость образования) антигена. Сенсоры гасили 1% БСА в воде. Зонды погружали в буфер для анализа (ФСБ) и измеряли скорость диссоциации (скорость распада).
[0108] Аффинность, представленную константой равновесия для диссоциации антигена и антигенсвязывающего белка (KD), определяли исходя из полученной константы аффинности (KA), и KD с помощью метода глобальной аппроксимации 1:1 с использованием программного обеспечения Fortebio, как показано ниже в Таблице 1. Аффинность определяли посредством усреднения значений KD для кривых со значениями R2 > 0,95. Результат для 250 нМ VHH13 к STAT3 был опущен, так как он представляет собой резко отклоняющееся значение. Было установлено, что аффинность VHH13-ОдАт (SEQ ID № 3) к STAT3 составляла 1,16×10-7. Аффинность VHH к KRAS не была определена.
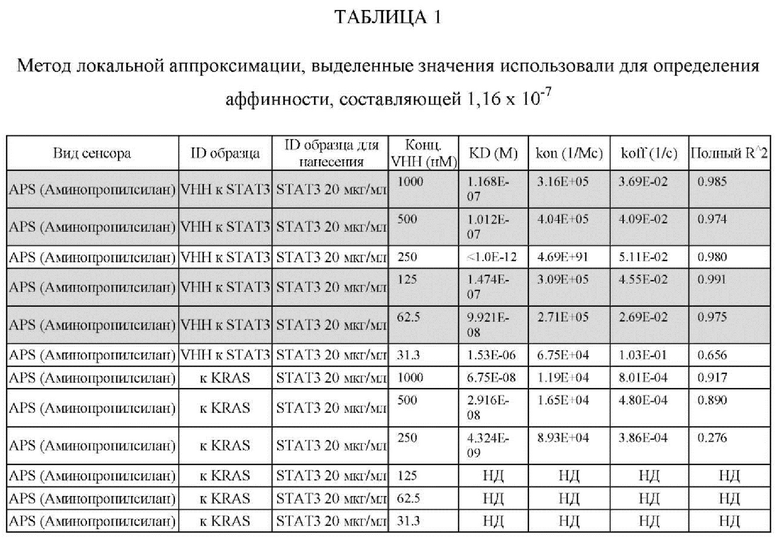
ПРИМЕР 2: ИССЛЕДОВАНИЯ ИММУНОПРЕЦИПИТАЦИИ
[0109] Анализировали специфичность ОдАт к STAT3 в клетках рака молочной железы человека. В этом примере клетки рака молочной железы человека MDA-MB-231 выращивали до достижения 50%-70% конфлюэнтности. Затем клетки разрушали в свежеприготовленном охлажденном на льду буфере для лизиса (20 мМ ГЭПЭС, рН 7,9, 400 мМ NaCl, 0,1% NP-40, 10% глицерина, 1 мМ ванадата натрия, 1 мМ фторида натрия, 1 мМ дитиотреитола, 1 мМ фенилметилсульфонилфторида, 10 мкг/мл апротинина, 10 мкг/мл лейпептина) в течение 45 минут на льду. Затем лизаты центрифугировали, собирали супернатант и определяли концентрацию белка с помощью модифицированного метода Лоури (Bio Rad, Геркулес, штат Калифорния). Общий белок (1 мг) инкубировали с 1,5 мг Dynabeads (Invitrogen) с ОдАт к STAT3, положительным контролем (STAT3, кат. № SC-482, Santa Cruz Biotechnology, Даллас, штат Техас) или отрицательным контролем (STAT-1, кат. № 9172, Cell Signaling, Дэнверс, штат Массачусетс) в течение 1 ч при 4°С. Затем гранулы промывали. После последней промывки добавляли 60 мкл буфера для лизиса и подвергали полученный супернатант Вестерн-блоттингу. Вкратце, образцы разделяли на 10% полиакриламидном геле и переносили на нитроцеллюлозную мембрану. Мембраны блокировали, а затем инкубировали с соответствующими первичными и вторичными антителами. Антителом к STAT3, использованным в качестве положительного контроля, было антитело от Cell Signaling (кат. № 4904, Дэнверс, штат Массачусетс). Реакцию хемилюминесценции проводили с использованием системы ECL от Santa Cruz Biotechnology (Даллас, штат Техас).
[0110] Как проиллюстрировано на Фиг. 3, эндогенный STAT3, иммунопреципитированный со всеми ОдАт, испытывали в различных количествах. М представляет собой маркерную дорожку, содержащую маркер, дорожка 1 содержала VHH13 (SEQ ID №: 3) к STAT3, полученное и выделенное из клеток млекопитающих, дорожка 2 содержала VHH14 (SEQ ID №: 4) к STAT3, полученное и выделенное из клеток млекопитающих, дорожка 3 содержала VHH13 (SEQ ID №: 3) к STAT3, полученное и выделенное из бактериальных клеток, дорожка 4 содержала VHH14 (SEQ ID №: 4) к STAT3, полученное и выделенное из клеток млекопитающих, дорожка 5 представляла собой положительное к STAT3 антитело, дорожка 6, использующая STAT-1 в качестве отрицательного контроля, не показала полосу.
ПРИМЕР 3: ПОЛУЧЕННОЕ В БАКТЕРИИ VHH13 К STAT3 СВЯЗЫВАЕТСЯ С ВЫСОКОЙ АФФИННОСТЬЮ С КЛЕТОЧНЫМИ ЛИНИЯМИ, СОДЕРЖАЩИМИ КОНСТИТУТИВНО АКТИВИРОВАННЫЙ STAT3
[0111] Анализировали специфичность полученного в бактерии VHH13 (SEQ ID №: 3) к STAT3 с использованием конститутивно активированного STAT3 в клеточных линиях человека (PANC-1 и DU145) и мыши (4T1). Также обрабатывали коммерчески доступные клетки HeLa гамма-интерфероном (INFГ), чтобы индуцировать фосфорилированный STAT3. Линию нулевых клеток PC-3 STAT3 использовали в качестве отрицательного контроля.
[0112] Клетки выращивали до достижения 50%-70% конфлюэнтности, а затем разрушали в свежеприготовленном охлажденном на льду буфере для лизиса, как описано выше, в течение 45 минут на льду. Затем лизаты центрифугировали, собирали супернатант и определяли концентрацию белка, как описано выше. Общий белок (1 мг) инкубировали с 1,5 мг Dynabeads (Invitrogen) с полученным в бактерии VHH13 (SEQ ID №: 3) к STAT3 или отрицательным контролем (KRAS, Creative Biolabs, Ширли, штат Нью-Йорк) в течение 1 часа при 4°C. Затем гранулы промывали. После последней промывки добавляли 60 мкл буфера для лизиса и подвергали полученный супернатант Вестерн-блоттингу, как описано в Примере 2.
[0113] Как проиллюстрировано на Фиг. 4, эндогенный STAT3 иммунопреципитировали полученным в бактерии VHH13 (SEQ ID №: 3) к STAT3 в конститутивно активированных линиях клеток STAT3: PANC-1 (дорожка 1), DU145 (дорожка 2) и 4T1 (дорожка 4). Кроме того, полученное в бактерии VHH13 (SEQ ID №: 3) к STAT3 связывалось с фосфо-STAT3 в лизате HeLa (дорожка 3). Не было отмечено полос ни для KRAS PANC-1, дорожка 3, ни для PC-3 (отрицательный контроль), дорожка 6.
ПРИМЕР 4: ИССЛЕДОВАНИЯ ЦИТОТОКСИЧНОСТИ ОДАТ К STAT3 В РАКОВЫХ КЛЕТКАХ ЛИНИИ MDA-MB-231
[0114] В этом примере антипролиферативное действие ОдАт к STAT3 анализировали с использованием клеточной линии рака молочной железы MDA-MB-231. Для опытов, клетки MDA-MB-231 выращивали до тех пор, пока они не достигали конфлюэнтности, составляющей 90%. В то же время, клетки промывали, трипсинизировали и подсчитывали с помощью Coulter Counter (Beckman, Брея, штат Калифорния). Исследования пролиферации проводили с помощью 3-[4,5-диметилтиазолил]-2,5-дифенилтетразолийбромидного (MTT) метода. Для этого клетки высевали в 96-луночный планшет при плотности, составляющей 5×103 на лунку, как указано производителем (Roche Diagnostics Corporation, Индианаполис, штат Индиана). Клеткам давали возможность прилипнуть в течение 24 часов, а затем добавляли ОдАт в соответствующих концентрациях (т.е. 0, 0,5, 1,0, 10,0 или 100 мкг/мл). Клетки подсчитывали на 3-й день. У клеток, обрабатываемых в течение 5 дней, на 3-й день обновляли питательные среды, содержащие ОдАт. По истечении срока в каждую лунку добавляли 10 мкл реагента MTT (0,5 мг/мл), как указано производителем. После 4-часового инкубационного периода добавляли 100 мкл раствора для солюбилизации, и на ночь помещали планшет в инкубатор. Все планшеты считывали при длине волны, составляющей 570 нм, с использованием планшет-ридера Biotek (Винуски, штат Вермонт).
[0115] Все данные анализировали с использованием GraphPad InStat 3 (GraphPad Software, Inc., Ла-Хойя, штат Калифорния). Группы лечения сравнивали с контрольной группой, получающей носитель, с помощью однофакторного дисперсионного анализа. Если наблюдалась значительная разница (p < 0,05), проводили тест множественных сравнений Тьюки-Крамера.
[0116] Основываясь на опыте МТТ, было установлено, что полученное в бактерии VHH13-ОдАт к STAT3 (SEQ ID № 3) является эффективным в ингибировании роста клеток на 3-й и 5-й день после обработки, как показано в Таблицах 2-5 ниже.
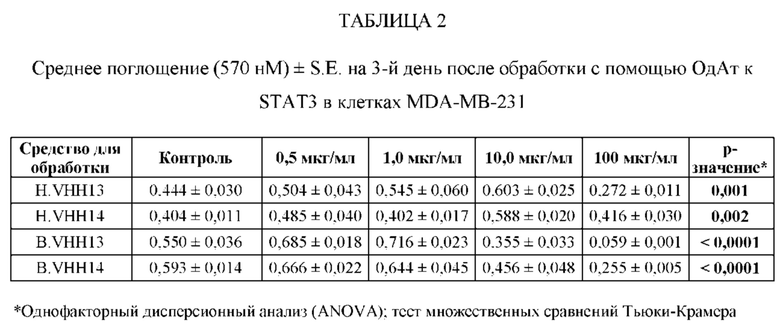
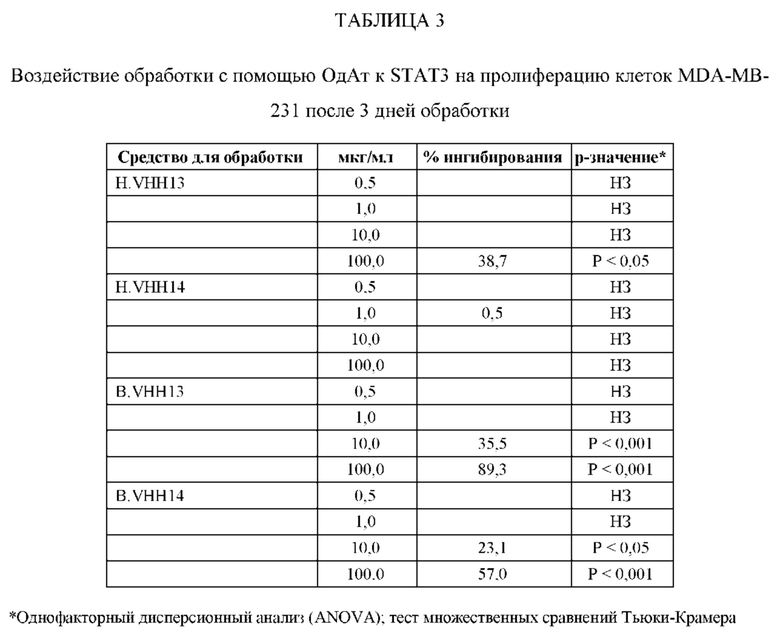

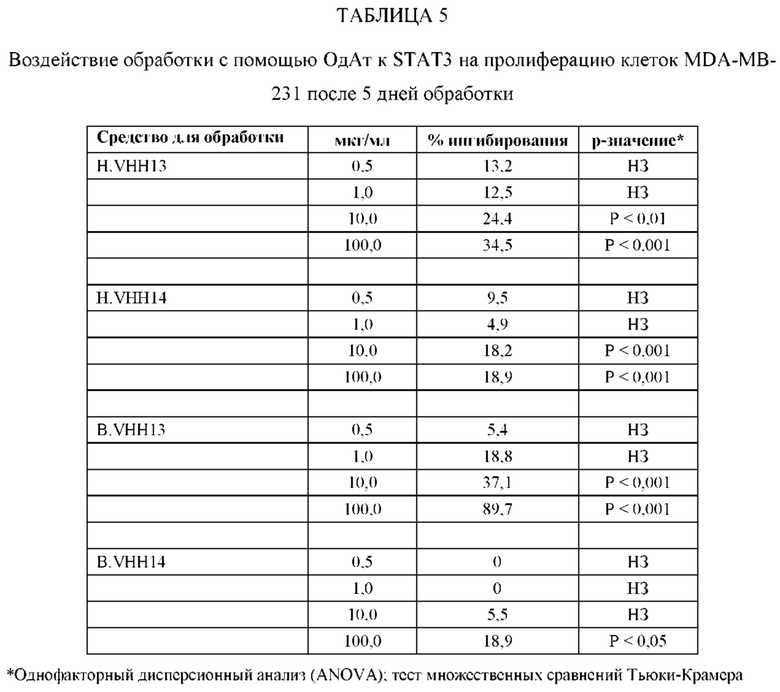
ПРИМЕР 5: ИССЛЕДОВАНИЯ ЦИТОТОКСИЧНОСТИ ОДАТ К STAT3 В КЛЕТОЧНЫХ ЛИНИЯХ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ (MDA-MB-231) И ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ (PANC-1) ЧЕЛОВЕКА
[0117] В этом примере антипролиферативное действие VHH13-ОдАт (SEQ ID № 3) и VHH14-ОдАт (SEQ ID № 4) к STAT3 анализировали с использованием клеточной линии рака молочной железы человека MDA-MB-231 и клеточной линии рака поджелудочной железы человека PANC-1. Для опытов, клетки MDA-MB-231 и PANC-1 выращивали до тех пор, пока они не достигали 90% конфлюэнтности. В то же время, клетки промывали, трипсинизировали и подсчитывали с помощью Coulter Counter (Beckman, Брея, штат Калифорния). Исследования пролиферации проводили с помощью метода MTT, описанного выше. У клеток, обрабатываемых в течение 5 дней, на 3-й день обновляли питательные среды, содержащие ОдАт к STAT3.
[0118] Все данные анализировали с использованием GraphPad InStat 3. Группы лечения сравнивали с контрольной группой, получающей носитель, с помощью однофакторного дисперсионного анализа. Если наблюдалась значительная разница (p < 0,05), проводили тест множественных сравнений Тьюки-Крамера.
[0119] Основываясь на опыте МТТ, было установлено, что и VHH13 (SEQ ID № 3), и VHH14 (SEQ ID № 4) ингибируют рост клеток как в раковых клетках MDA-MB-231, так и PANC-1, как показано в Таблицах 6-13 ниже.
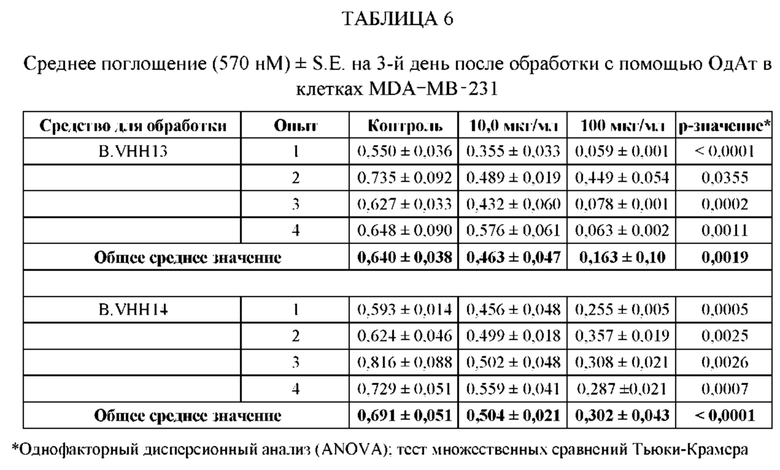
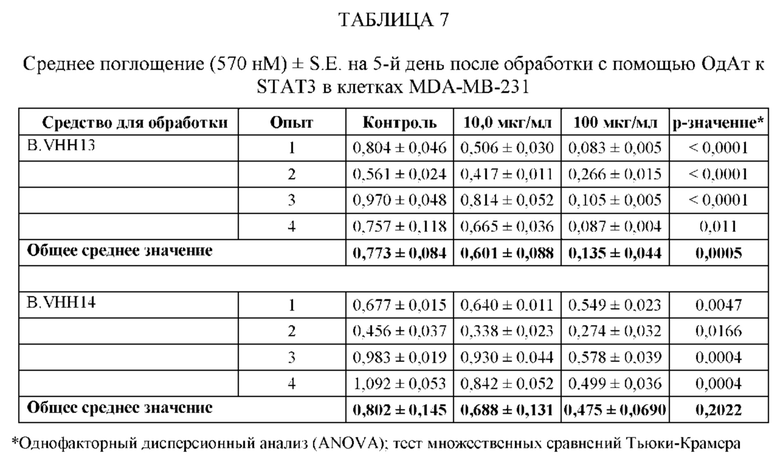
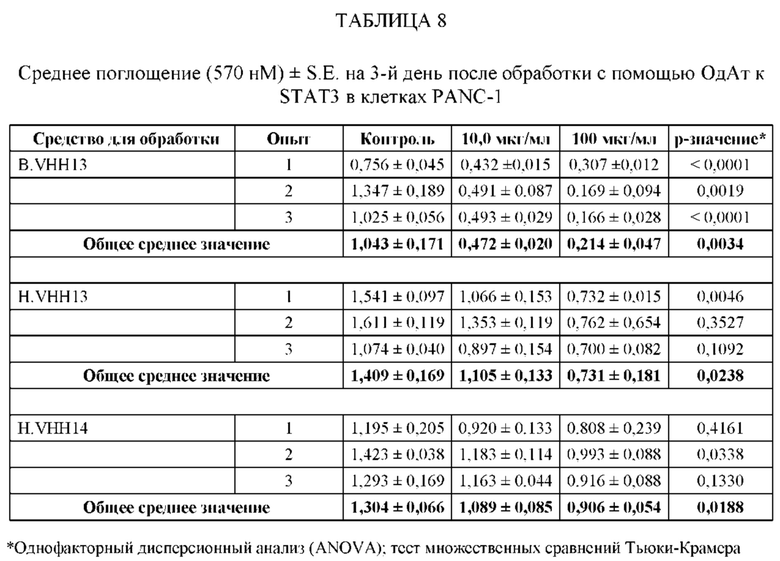
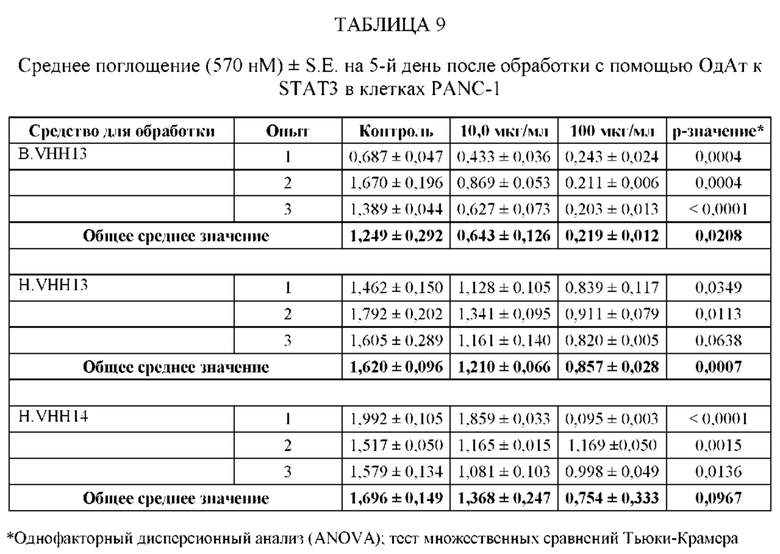
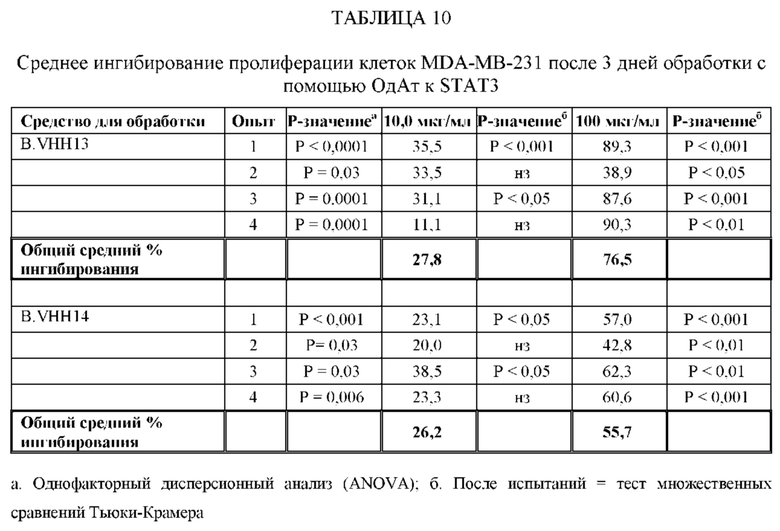
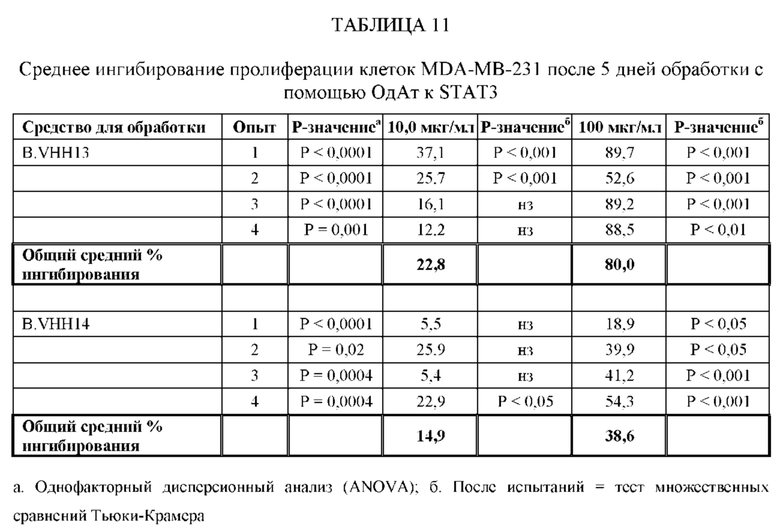
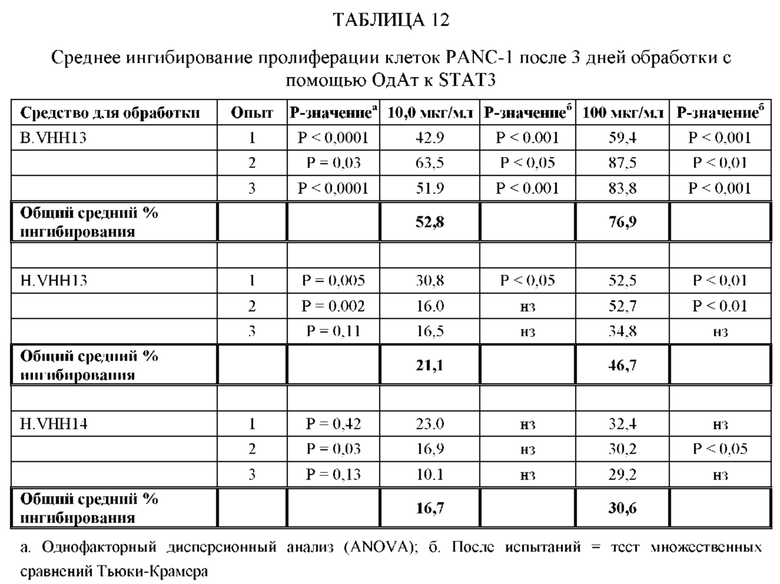
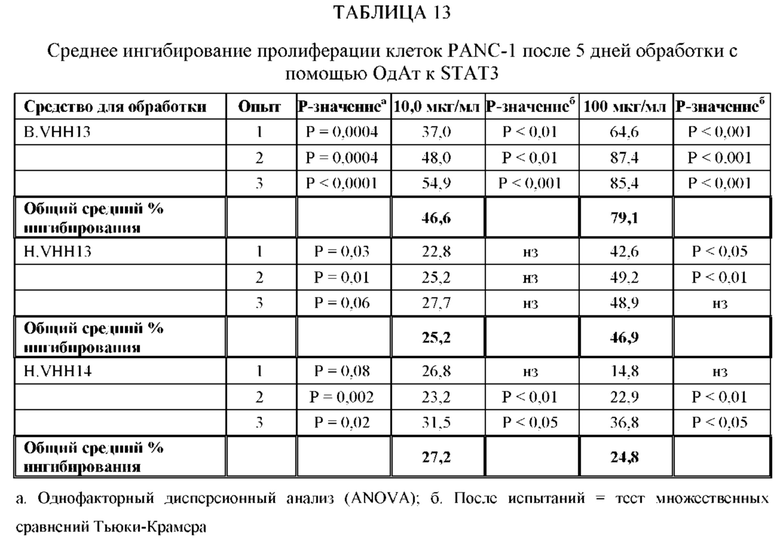
ПРИМЕР 6: АНТИПРОЛИФЕРАТИВНОЕ ДЕЙСТВИЕ ОДАТ К STAT3 В КЛЕТОЧНЫХ ЛИНИЯХ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ ЧЕЛОВЕКА И РАКА ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ ЧЕЛОВЕКА
[0120] Антипролиферативное действие VHH13-ОдАт (SEQ ID № 3) к STAT3 анализировали в клеточной линии рака молочной железы человека MDA-MB-231 и клеточных линиях рака предстательной железы человека DU145. Для опытов, раковые клетки выращивали до тех пор, пока они не достигали 90% конфлюэнтности. В то же время, клетки промывали, трипсинизировали и подсчитывали с помощью Coulter Counter (Beckman, Брея, штат Калифорния). Исследования пролиферации осуществляли с помощью метода MTT, как описано выше.
[0121] Антипролиферативные свойства полученного в бактерии VHH13-ОдАт (SEQ ID № 3) к STAT3 в клетках MDA-MB-231 сравнивали с его действиями в клетках DU145. Как показано в Таблице 14, клетки MDA-MB-231, обработанные ОдАт к STAT3 (SEQ ID №: 3), показали среднее ингибирование роста, составляющее 29,6 и 91,2 при 50,0 и 100 мкг/мл, соответственно. В клетках DU145 наблюдалось аналогичное ингибирование роста (31,2 и 92,1% в случае 50,0 и 100 мкг/мл, соответственно), как приведено в Таблице 15.
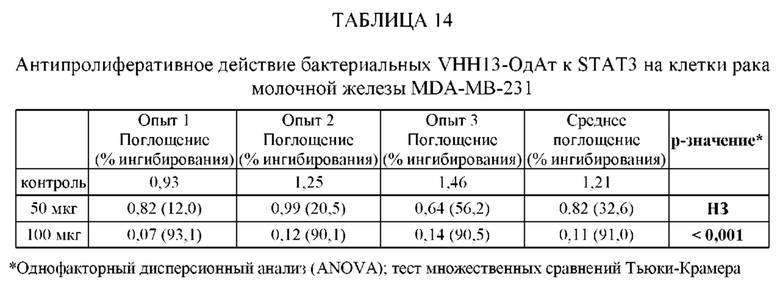

ПРИМЕР 7: АНТИПРОЛИФЕРАТИВНОЕ ДЕЙСТВИЕ VHH13-ОДАТ (SEQ ID № 3) К STAT3 НА КЛЕТОЧНЫЕ ЛИНИИ РАКА ЧЕЛОВЕКА
[0122] Для испытания антипролиферативного действия VHH13-ОдАт (SEQ ID № 3) к STAT3 использовали клеточные линии рака человека: MDA-MB-231, MDA-MB-468, MCF-7, BT474 и DU145, как показано в Таблице 16.
[0123] Все клеточные линии рака человека были получены из Американской коллекции типовых культур (Манассас, штат Вирджиния). Клеточные линии поддерживали и культивировали в среде RPMI 1640 (MDA-MB-231, MDA-MB-468, MCF-7, BT474) или MEM-E (DU145), содержащей 10% фетальной бычьей сыворотки, 2 мМ L-глутамина и 1% раствор антибиотика и противогрибкового средства (10 ед/мл пенициллина, 10 мкг/мл стрептомицина и 25 мкг/мл амфотерицина В). Клетки выдерживали при 37°С в увлажненной атмосфере, содержащей 5% CO2. Расходуемые клеточные культуры были получены от Life Technologies, Inc., (Гранд Айленд, штат Нью-Йорк). Реагент МТТ был приобретен у Sigma Aldrich (Сент-Луис, штат Миссури).
[0124] Для опытов, раковые клетки выращивали до тех пор, пока они не достигали 90% конфлюэнтности. В то же время, клетки промывали, трипсинизировали и подсчитывали с помощью Coulter Counter (Beckman, Брея, штат Калифорния). Исследования пролиферации проводили с помощью метода MTT, как описано выше.
[0125] Антипролиферативные свойства полученных в бактерии VHH13-ОдАт (SEQ ID № 3) к STAT3 оценивали по пяти клеткам рака молочной железы, представляющим различные классификации (Таблица 34). Как показано в Таблице 17, все клеточные линии демонстрировали значительное ингибирование роста через 72 часа после обработки. Для всех клеточных линий наибольшее ингибирование роста отмечалось при дозе, составляющей 100 и 200 мкг/мл. Полумаксимальная ингибирующая концентрация (IC50) для роста в испытуемых клеточных линиях составляла: 10,1 ± 2,4, 12,36 ± 1,5, 14,8 ± 1,6 и 25,2 ± 14,7 в случае клеточных линий MDA-MB-231, MDA-MB-468, MCF-7 и BT474, соответственно. Эти данные свидетельствуют о том, что клеточным линиям трижды негативного рака молочной железы требуется самая низкая концентрация VHH13-ОдАт (SEQ ID №: 3) для достижения IC50 по сравнению с эстроген/прогестерон положительными клеточными линиями (т.е., MCF-7) или клеточными линиями с амплификацией гена HER2 (т.е., BT474).
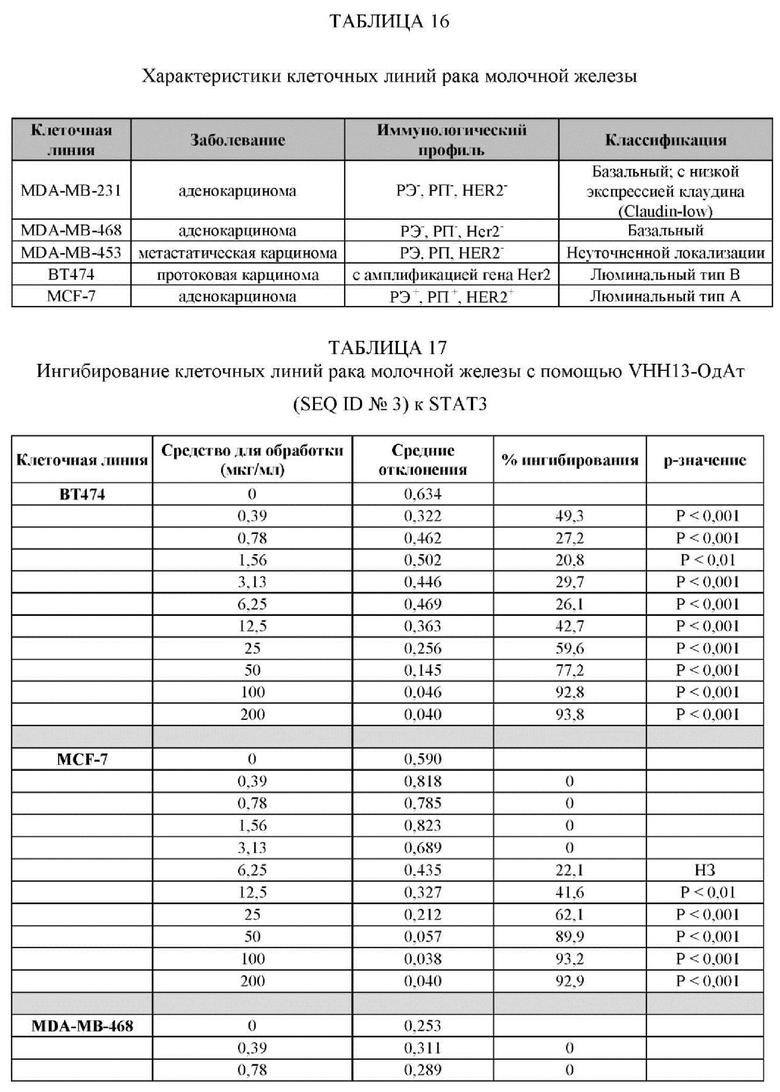
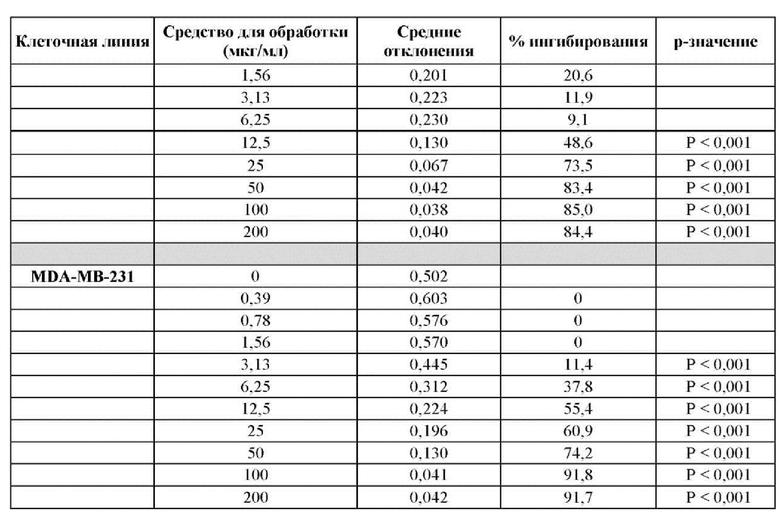
[0126] Также оценивали действие полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в клеточной линии рака предстательной железы человека DU145, как показано в Таблице 18. Полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 демонстрировало дозозависимое ингибирование роста во всех испытуемых раковых клетках.
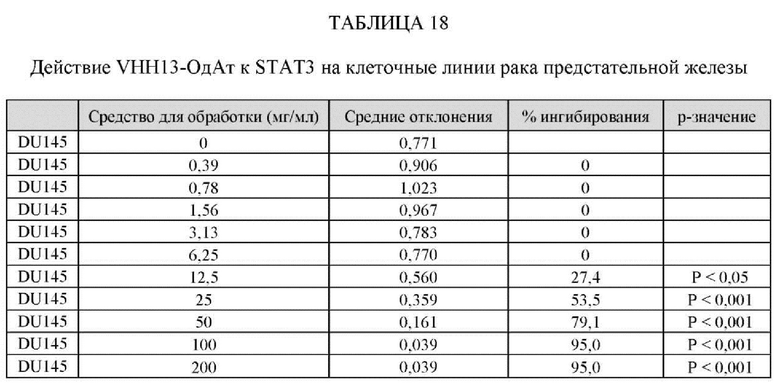
ПРИМЕР 8: МАКСИМАЛЬНО ПЕРЕНОСИМАЯ ДОЗА ПОЛУЧЕННОГО В БАКТЕРИИ VHH13 (SEQ ID №: 3) К STAT3 У МЫШЕЙ BALB/C
[0127] В этом Примере анализировали переносимость полученного в бактерии VHH13-ОдАт (SEQ ID № 3) к STAT3 у подопытных животных с использованием клеточной линии рака молочной железы человека MDA-MB-231. Для опыта, в общей сложности 9 "голых" самок мышей BALB/C (6-7 недель) разделяли на три группы в зависимости от массы тела. (Таблица 19) Мыши (n = 3) получали либо носитель (ФСБ), либо полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в дозе, составляющей 250 или 500 мкг/кг массы тела/день, в течение пяти дней. В ходе исследования, смертность/заболеваемость происходила два раза в день. Массу тела регистрировали на 1-й, 4-й и 6-й дни исследования, а также в день прекращения исследования (13-й день). Токсичность оценивали по измерениям массы тела и поведению мыши по сравнению с контрольными мышами, получающими носитель. После завершения фазы лечения за животными наблюдали в течение дополнительной недели, чтобы отмечать любые отклонения в массе тела и/или общего состояния здоровья после обработки.
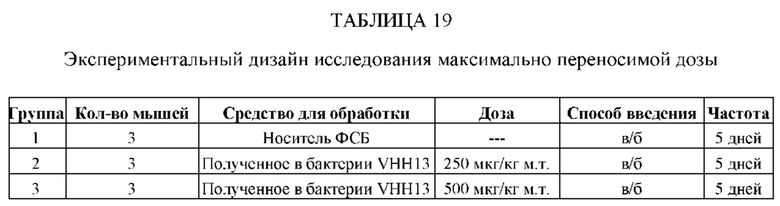
[0128] Как показано в Таблице 20, масса тела в группах существенно не отличалась, и полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 не приводило к какими-либо смертельным случаям, обусловленным приемом препарата, при любом уровне дозирования. Кроме того, не наблюдалось никаких изменений в поведении у животных, получавших полученные в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3, по сравнению с контрольными мышами.
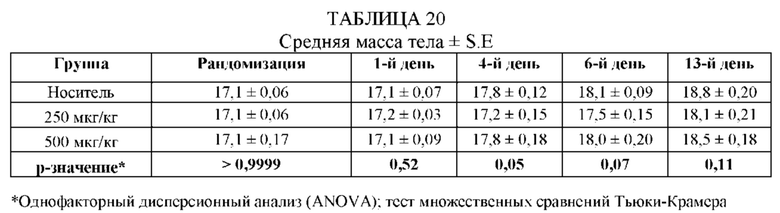
ПРИМЕР 9: АКТИВНОСТЬ ПОЛУЧЕННОГО В БАКТЕРИИ VHH13 (SEQ ID №: 3) К STAT3 В КСЕНОТРАНСПЛАНТАТЕ ОПУХОЛИ У "ГОЛЫХ" МЫШЕЙ BALB/C, А ТАКЖЕ В КЛЕТКАХ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ ЧЕЛОВЕКА И РАКА ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ ЧЕЛОВЕКА
[0129] В этом примере оценивали активность полученного в бактерии VHH13- ОдАт (SEQ ID № 3) к STAT3 у мышей с использованием клеточной линии рака молочной железы человека MDA-MB-231. Вкратце, активность полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 оценивали с использованием: ксенотрансплантатной модели рака молочной железы человека MDA-MB-231 и ксенотрансплантатной модели рака поджелудочной железы человека PANC-1. Режимы дозирования были следующими: Группа 1 (n = 6; ФСБ, в/б), один раз в день в течение 14 дней [ОД x 14]; и Группа 2 (n-12; 500 мкг/кг м.т., в/б), один раз в день в течение 14 дней [ОД x 14]. За введением препарата следовал период наблюдения, составлявший 5 дней.
[0130] Клеточные линии рака молочной железы человека (MDA-MB-231 и PANC-1) были получены из Американской коллекции типовых культур (ATCC) (Манассас, штат Вирджиния). Клетки MDA-MB-231 выращивали в MEM (Life Technologies, Гранд Айленд, штат Нью-Йорк) с добавлением 10% FBS (Atlanta Biologicals, Флауэри Бранч, штат Джорджия) и пенициллин-стрептомицин-глутамина (Life Technologies, Гранд Айленд, штат Нью-Йорк). Клетки PANC-1 выращивали в среде RPMI 1640 (Life Technologies, Гранд Айленд, штат Нью-Йорк) с добавлением 10% FBS и пенициллин-стрептомицин- глутамина. Все клетки выращивали в инкубаторе в присутствии 5% CO2 при 37°С.
[0131] Самцы бестимусных мышей Nude-Foxnlnu в возрасте от 4 до 5 недель были приобретены у Harlan Laboratories (Индианаполис, штат Индиана). Животных карантинировали в течение одной недели и содержали по пять мышей в клетке, при 12- часовом светотемновом цикле и относительной влажности, составляющей 50%. Питьевую воду и корм предоставляли животным в неограниченном количестве. Всех животных содержали в стерильных условиях и проводили опыты при одобрении Комитета по биоэтике НИИ IIT. Для исследования ксенотрансплантата MDA-MB-231, клетки (4×106) в конечном объеме, составляющем 100 мкл среды MEM, вводили подкожно в правый бок мышей. Для исследования ксенотрансплантата PANC-1, клетки (5×106) в конечном объеме, составляющем 100 мкл среды RPMI, вводили подкожно в правый бок мышей. Измерения опухолей для обеих моделей начинали, как только опухоли становились пальпируемыми. Затем опухоли измеряли дважды в неделю. Животных рандомизировали по группам, когда опухоли достигали размера, находящегося в диапазоне 75-175 мм3, группы контроля (n = 6) и лечения (n = 12) рандомизировали с помощью алгоритма стратифицированной случайной выборки. Лечение (полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3) или носитель (ФСБ) инициировали на следующий день после рандомизации. Лечение хорошо переносилось и не приводило к смертельным случаям, обусловленным приемом препарата. Существенная потеря веса не отмечалась.
[0132] Для исследования ксенотрансплантата MDA-MB-231, рандомизационный средний (±SE) размер опухоли составлял: 103,01 ± 11,89 и 102, 61 ± 9,60 в случае групп контроля и лечения, соответственно. Средние массы тела (± SE) при рандомизации составляли: 32,08 ± 0,76 и 30,27 + 0,75 для Группы 1 и Группы 2, соответственно. В Таблице 21 представлены средние массы тела (± SE) для всего исследования.
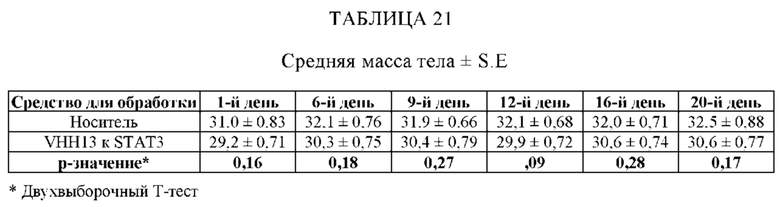
[0133] На 14-й день введения дозы средний размер опухоли (±SE) для группы контроля составлял 179,11 ± 19,39 по сравнению с 118,86 ± 15,94 для группы лечения. Средние массы тела (± SE) на момент прекращения составляли: 31,98 ± 0,71 и 30,55 ± 0,74 для Группы 1 и Группы 2, соответственно. Таблица 22 резюмирует объемы опухолей (± SE) для всего исследования. % среднего ингибирования роста опухоли в группе лечения составлял 33,64%. Время удвоения опухолей было следующим: Группа 1: 44,27 дней; и Группа 2: 61,06 дней. На Фиг. 5 проиллюстрировано ингибирование роста полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в ксенотрансплантатной модели MDA- MB-231. Полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 демонстрировало значительное ингибирование роста (p = 0,047). Таким образом, полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 обладает химиотерапевтической активностью в модельной системе рака молочной железы человека MDA-MB-231.
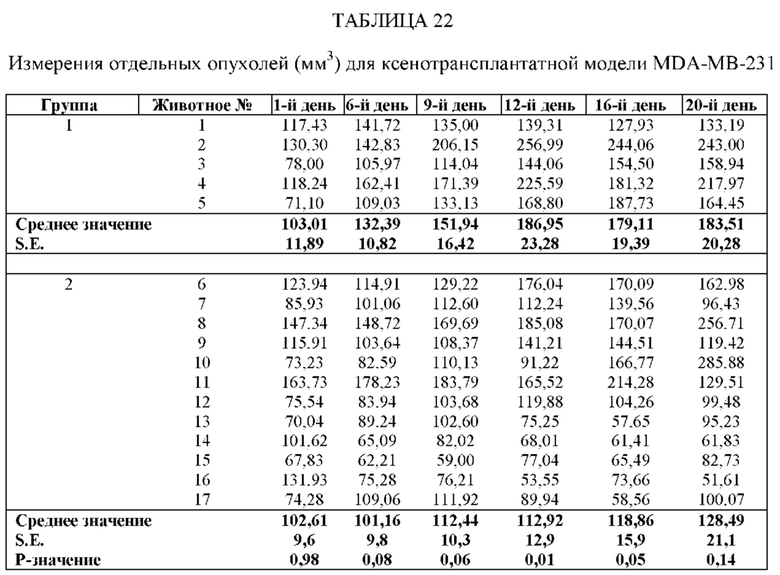
[0134] Для исследования ксенотрансплантата PANC-1, рандомизационные средние (+SE) размеры опухоли составляли 107,01 ± 4,54 в группе контроля и 110,58 ± 6,18 в группе лечения. Средние массы тела (± SE) при рандомизации составляли: 29,0 ± 0,81 и
28,5 ± 0,70 для Группы 1 и Группы 2, соответственно. Средние массы тела (± SE) на момент прекращения составляли: 31,2 ± 0,99 и 30,1 ± 0,75 для Группы 1 и Группы 2, соответственно. Таблица 23 резюмирует средние массы тела (± SE) для всего исследования. На 14-й день введения дозы средний размер опухоли (±SE) для группы контроля составлял 287,30 ± 33,94 по сравнению с 318,74 ± 29,76 для группы лечения. Таблица 24 резюмирует объемы опухолей (± SE) для всего исследования.

[0135] Время удвоения опухолей было следующим: Группа 1: 22,44 дней; и Группа 2: 23,02 дней. Полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 не показало значительное ингибирование роста в модельной системе рака поджелудочной железы человека PANC-1.
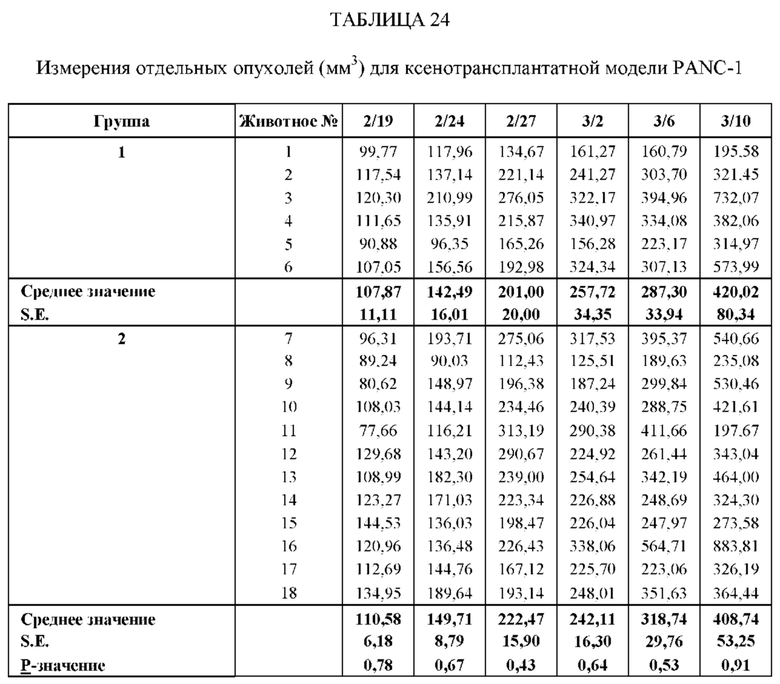
ПРИМЕР 10: ИССЛЕДОВАНИЕ КСЕНОТРАНСПЛАНТАТА MDA-MB-231
[0136] В этом Примере дополнительно оценивали эффективность полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в ксенотрансплантатной модели рака молочной железы человека MDA-MB-231. Режимы дозирования были следующими: Группа 1 (n = 4; ФСБ; в/б) дважды в день в течение 14 дней [ДД x 14]; Группа 2 (n = 4; 1 мг/кг м.т.; в/б), дважды в день в течение 14 дней [ДД x 14]; Группа 3 (n = 4; 2 мг/кг м.т.; в/б) дважды в день в течение 14 дней [ДД x 14]; и Группа 4 (n = 4; 2 мг/кг м.т.; в/б) один раз в день в течение 14 дней [ОД x 14]. За введением следовал период наблюдения, составлявший 7 дней.
[0137] Клеточные линии рака молочной железы человека MDA-MB-231 и самки бестимусных мышей Nude-Foxnlnu были описаны выше.
[0138] Клетки MDA-MB-231 вводили при плотности, составляющей 5×106, подкожно в правый бок мышей в конечном объеме, составляющем 100 мкл в среде MEM. Измерения опухолей начинали, как только опухоли становились пальпируемыми. Затем опухоли измеряли дважды в неделю. Животных рандомизируют по группам с помощью алгоритма стратифицированной случайной выборки, когда опухоли достигают размера, находящегося в диапазоне 55-150 мм3. Лечение (полученным в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3) или носитель (ФСБ) инициировали на следующий день после рандомизации.
[0139] Рандомизационный средний (± SE) размер опухоли составлял: 92,08 ± 13,24, 82,38 ± 5,17, 77,47 ± 7,17 и 104,71 ± 14,64 для Групп 1, 2, 3 и 4, соответственно. Как показано в Таблице 25, средние массы тела (± SE) при рандомизации составляли: 23,65 ± 0,72, 23,45 ± 0,66, 23,10 ± 0,20 и 22,45 ± 1,25 для Групп 1, 2, 3 и 4, соответственно.
[0140] Как показано в Таблице 26, на 14-й день введения дозы, средний размер опухоли (±SE) для контрольной группы составлял 221,51 ± 57,32 по сравнению с 67,12 ± 10,66, 58,27 ± 22,54 и 131,44 ± 22,86 для 2, 3 и 4 группы лечения, соответственно. На момент прекращения (42-й день) средний размер опухоли (± S.E.) составлял: 255,42 ± 65,46, 55,98 ± 6,94, 41,15 ± 13,21 и 145,51 ± 52,32, для групп 1, 2, 3 и 4, соответственно. Средние массы тела (± SE) на момент прекращения составляли: 24,80 ± 0,49, 23,25 ± 1,20, 24,00 ± 0,32 и 23,2 ± 1,46 для Групп 1, 2, 3 и 4, соответственно. Максимальный средний % чистой потери веса (день) составил: 0,7 (36), 1,5 (23), 1,8 (36) и 2,2 (29) для Групп 1, 2, 3 и 4, соответственно.
[0141] Кроме того, как показано в Таблице 26, среднее ингибирование роста в группах лечения составляло 78,3, 75,2 и 55,9 для Групп 2, 3 и 4, соответственно. Время удвоения опухолей составляло: Группа 1: 20,56 дней; Группа 2: 34,54 дней; Группа 3: 30,07 дней; и Группа 4: 27,17 дней. Наблюдалась задержка роста, составляющая 13,99, 9,52 и 6,61 дней, для Групп 2, 3 и 4, соответственно. Значения % соотношения лечение/контроль для групп лечения составляли: Группа 2: -33,75 (остановка роста опухоли); Группа 3: -54,4 (регрессия опухоли); и Группа 4: 10,28 (ингибирование роста опухоли). На Фиг. 6 проиллюстрировано ингибирование роста полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в ксенотрансплантатной модели MDA-MB-231.
[0142] Введение полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 вызывало значительное ингибирование роста в Группе 2 (р = 0,02) [1 мг/кг; ДД x 14] и Группе 3 (р = 0,02) [2 мг/кг; ДД х 14]. Кроме того, три из четырех опухолей показали значительную регрессию. На основании этих данных можно сделать вывод, что полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 обладает химиотерапевтической активностью в модельной системе рака молочной железы человека MDA-MB-231.
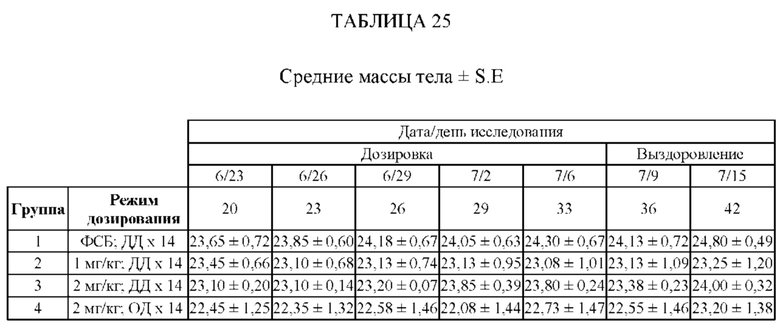
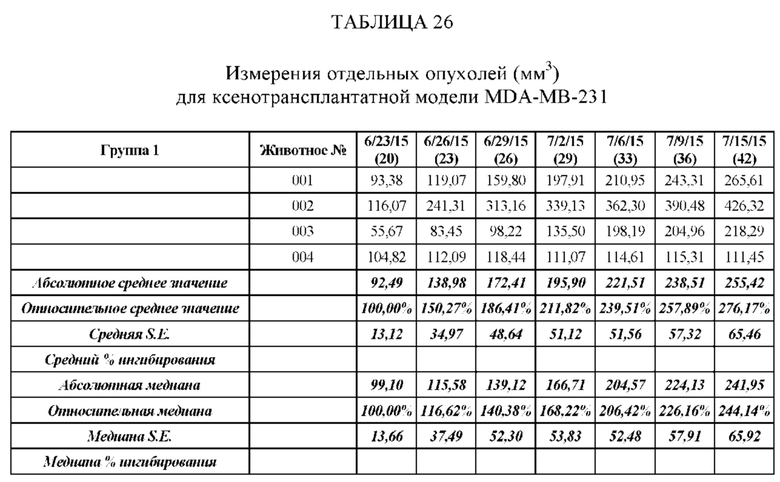
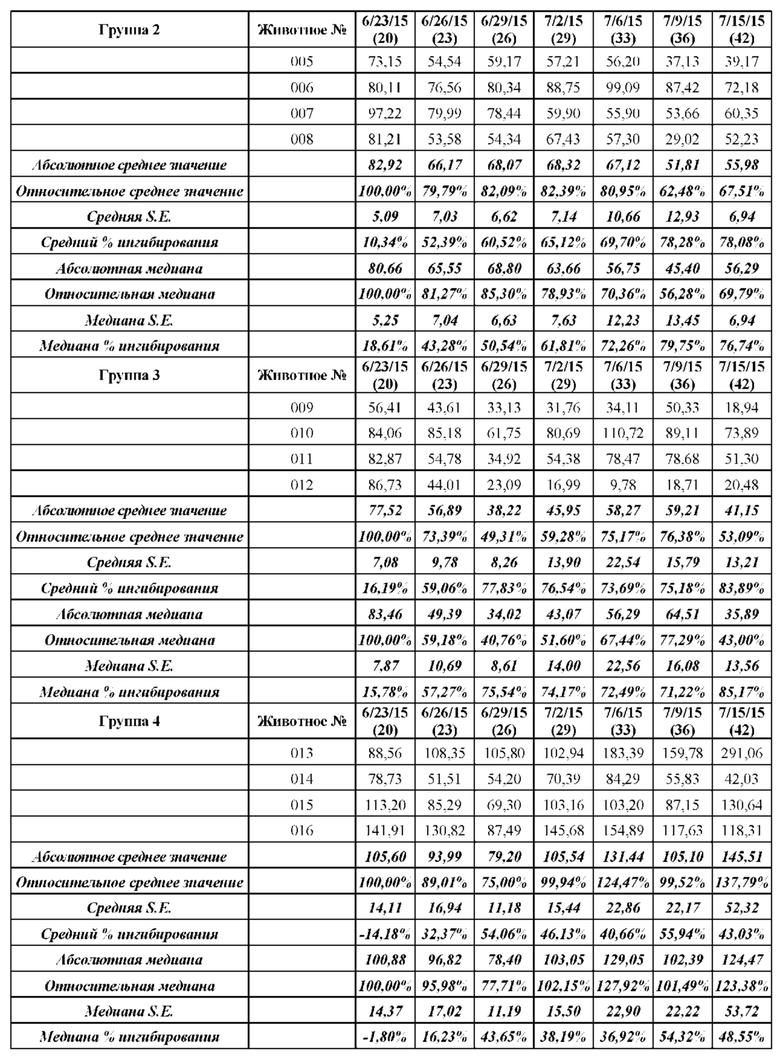
ПРИМЕР 11: ЭФФЕКТИВНОСТЬ ПОЛУЧЕННОГО В БАКТЕРИИ VHH13-ОДАТ (SEQ ID №: 3) К STAT3 В ТРЕХ КСЕНОТРАНСПЛАНТАТНЫХ МОДЕЛЯХ РАКА ЧЕЛОВЕКА
[0143] В этом Примере оценивали эффективность полученного в бактерии VHH13- ОдАт (SEQ ID №: 3) к STAT3 в ксенотрансплантатных моделях рака молочной железы MDA-MB-231, поджелудочной железы PANC-1 и предстательной железы DU145 человека.
[0144] Бестимусные мыши Nude-Foxn1nu, клетки рака молочной железы MDA-MB-231, клеточные линии рака поджелудочной железы PANC-1 и рака предстательной железы DU145 были описаны выше. В 1-й день исследования масса тела мышей находилась в диапазоне от 17 до 19 г (34 самки) и от 21 до 23 г (16 самцов).
[0145] Клетки в ранних пассажах (4-10) использовали для имплантации в мышей и собирали во время логарифмической фазы роста. MDA-MB-231 (5×106), DU145 (5×106) и PANC-1 (1,5×106) вводили подкожно в правый бок мышей в конечном объеме, составляющем 100 мкл среды. Измерения опухолей начинали, как только опухоли становились пальпируемыми. Затем опухоли измеряли дважды в неделю.
[0146] Животных рандомизировали по группам с помощью алгоритма стратифицированной случайной выборки, когда опухоли достигали размера, находящегося в диапазоне: 74-120 мм3 (MDA-MB-231), 89-146 мм3 (DU145) или 60-160 мм3 (PANC-1). Лечение (содержащее полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 и называемое в данном документе как SBT-100) или носитель (ФСБ) инициировали на следующий день после рандомизации, называемый 1-м днем.
[0147] Полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 доставляли в виде предварительно смешанного раствора в концентрации, составляющей 0,651 мг/мл, и хранили при -20°C до готовности к использованию. Исходный раствор разбавляли в стерильном ФСБ с рН 7,6 с получением 5 мг/кг в объеме дозирования, составляющем 10 мл/кг. Рабочий раствор готовили каждые 7 дней, аликвотировали в семь пробирок и хранили при 4°С. В каждый день лечения до комнатной температуры доводили только необходимую пробирку. Весь оставшийся материал ОдАт хранили при 4°С на случай возникновения необходимости в следующей дозе. На 8-й день любой оставшийся материал ОдАт выбрасывали, и готовили свежую порцию.
[0148] Двум группам (контроль и SBT-100) мышей в модели опухоли вводили дозу в соответствии с протоколом, показанным в Таблице 27. Режимы дозирования были следующими: Группа 1 (n = 4; ФСБ) дважды в день в течение 14 дней [ДД x 14]; Группа 2 (n = 4; SBT-100, 5 мг/кг м.т.), дважды в день в течение 14 дней [ДД x 14]. И носитель (ФСБ с рН 7,6), и SBT-100 вводили внутрибрюшинно (в/б) дважды в день, с интервалом в шесть часов в течение четырнадцати дней. Дозирование проводили в соответствии с массами отдельных животных. За введением следовал восстановительный период, составлявший 7 дней.
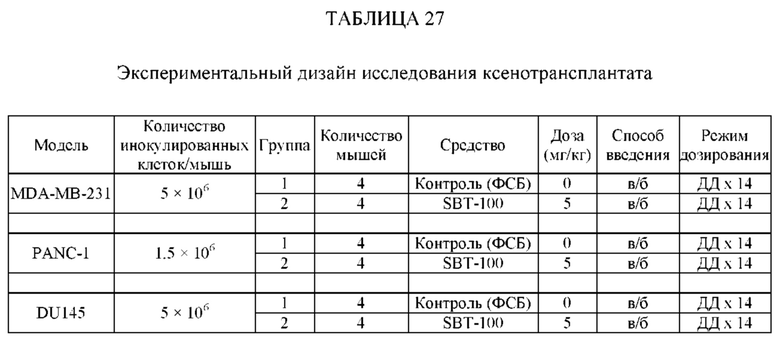
[0149] Для рандомизации животных по группам, сбора данных (например, дозировка, масса тела, измерения опухоли, клинические наблюдения) и проведения анализа данных использовали программное обеспечение Study Log Study Director Animal Study Management (Сан-Франциско, штат Калифорния).
[0150] В ксенотрансплантатной модели опухоли MDA-MB-231, животных рандомизировали по группам на 23-й день после инокуляции опухоли со средним (±SE) размером, составляющим: 77,98 ± 21,58 и 84,71 ± 5,56 для Групп 1 и 2, соответственно. Средние массы тела (± SE) при рандомизации составляли: 20,04 ± 0,62 и 23,7 ± 1,84 для Групп 1 и 2, соответственно. Таблица 28 резюмирует средние массы тела (± SE) для всего исследования. В последний день введения дозы (14-й день), средний размер опухоли (±SE) для контрольной группы составлял 168,28 ± 51,57 по сравнению с 83,81 ± 22,65 для мышей, обработанных SBT-100. Таблица 29 резюмирует объемы опухолей (± SE) для всего исследования. На момент прекращения (28-й день) средний размер опухоли (± S.E.) составлял: 270,49 ± 112,35 и 91,72 ± 33,17, для Групп 1 и 2, соответственно. Средние массы тела (± SE) на момент прекращения составляли: 25,36 ± 1,07 и 24,25 ± 1,68 для Групп 1 и 2, соответственно. В конце исследования среднее ингибирование роста опухоли в группе, обработанной SBT-100, составило 85,8% (p = 0,006). На Фиг. 7 проиллюстрирован средний объем опухоли. Время удвоения опухолей составляло 25,78 дней по сравнению с 111,6 дней для Группы 1 и Группы 2, соответственно. % соотношение лечение/контроль для Группы 2 составляло 13,35 (ингибирование опухоли).
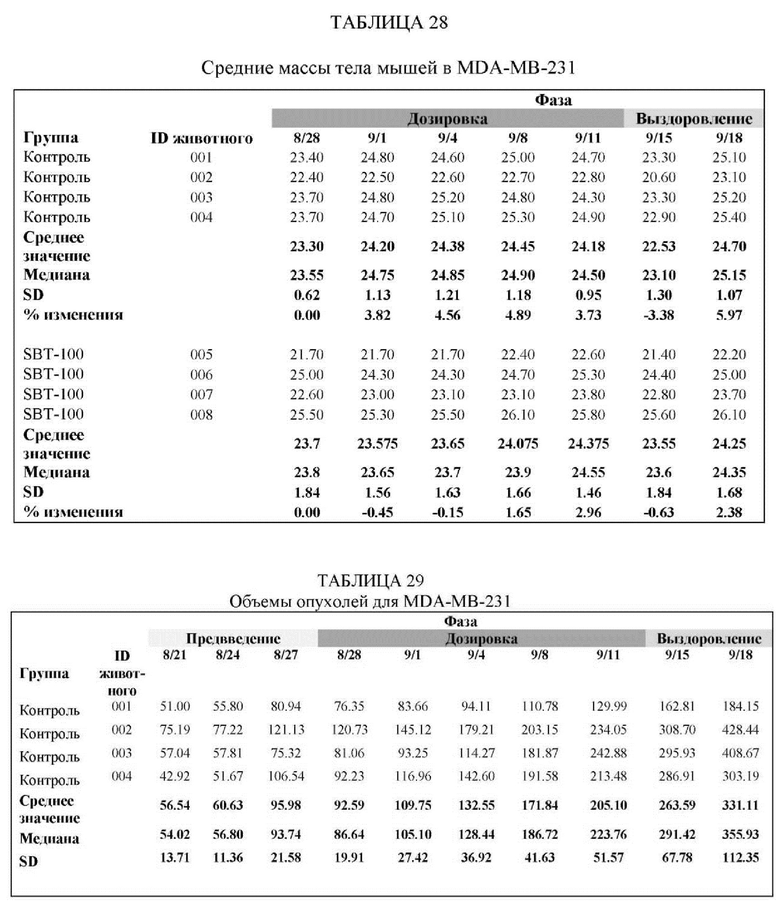
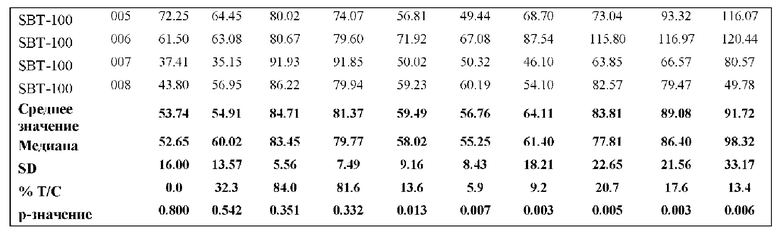
[0151] В ксенотрансплантатной модели опухоли DU145, животных рандомизировали по группам на 17-й день после инокуляции опухоли со средним (±SE) размером, составляющим: 111,87 ± 20,53 и 111,23 ± 25,16 для Групп 1 и 2, соответственно. Средние массы тела (± SE) при рандомизации составляли: 29,10 ± 1,94 и 30,68 ± 1,56 для Групп 1 и 2, соответственно. Таблица 30 резюмирует средние арифметические массы тела (± SE) для всего исследования. В последний день введения дозы (14-й день), средний размер опухоли (±SE) для контрольной группы составлял
621,81 ± 276,25 по сравнению с 364,14 ± 51,64 для мышей, обработанных SBT-100. Таблица 31 резюмирует объемы опухолей (± SE) для всего исследования. На момент прекращения (28-й день) средний размер опухоли (± S.E.) составлял: 819,42 ± 351,88 и 601,83 ± 131,51, для Групп 1 и 2, соответственно. Средние массы тела (± SE) на момент прекращения составляли: 29,20 ± 2,33 и 29,60 ± 1,04 для Групп 1 и 2, соответственно. В конце исследования среднее ингибирование роста опухоли в группе, обработанной SBT-100, составило 26,6% (p = 0,29). На Фиг. 8 проиллюстрирован средний объем опухоли. Время удвоения опухолей составляло 14,57 дней по сравнению с 18,19 дней для Группы 1 и Группы 2, соответственно. % соотношение лечение/контроль для
Группы 2 составляло 74,8.
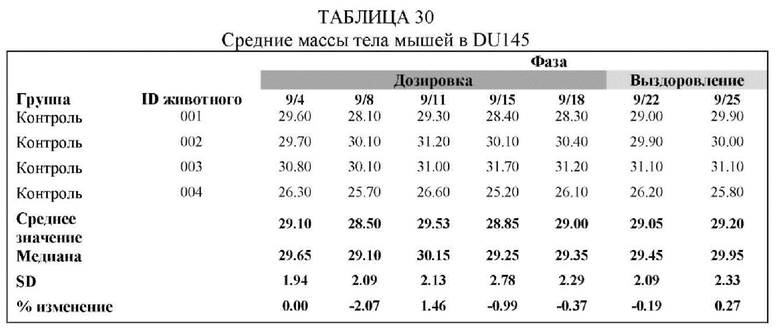
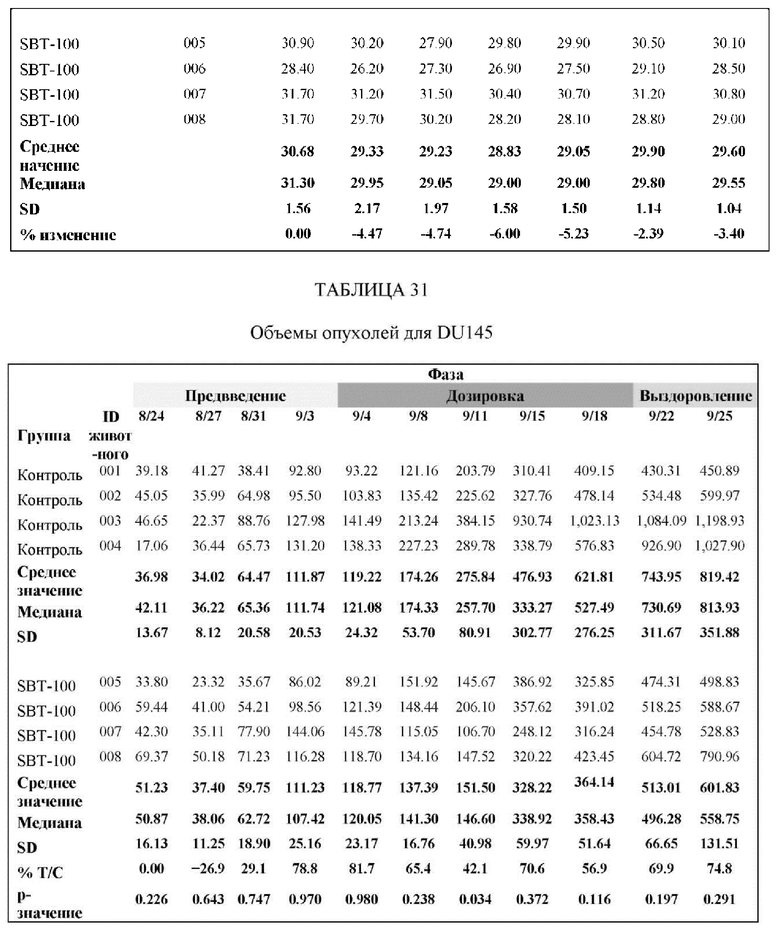
[0152] В ксенотрансплантатной модели опухоли PANC-1, животных рандомизировали по группам на 22-й день после инокуляции опухоли со средним (±SE) размером, составляющим: 78,74 ± 40,21 и 93,84 ± 36,31 для Групп 1 и 2, соответственно. Средние массы тела (± SE) при рандомизации составляли: 22,50 ± 1,47 и 24,23 ± 1,63 для Групп 1 и 2, соответственно. Таблица 32 резюмирует средние массы тела (± SE) для всего исследования. В последний день введения дозы (14-й день), средний размер опухоли (±SE) для контрольной группы составлял 204,95 ± 178,90 по сравнению с 159,03 ± 28,01 для мышей, обработанных SBT-100. Таблица 33 резюмирует объемы опухолей (± SE) для всего исследования. На момент прекращения (28-й день) средний размер опухоли (± S.E.) составлял: 284,77 ± 288,88 и 203,02 ± 30,34, для Групп 1 и 2, соответственно. Средние массы тела (± SE) на момент прекращения составляли: 27,38 ± 1,07 и 26,23 ± 1,19 для Групп 1 и 2, соответственно. В конце исследования среднее ингибирование роста опухоли в группе, обработанной SBT-100, составило 41,78% (p = 0,35). На Фиг. 9 проиллюстрирован средний объем опухоли. Время удвоения опухолей составляло 18,51 дней по сравнению с 35,70 дней для Группы 1 и Группы 2, соответственно. % соотношение лечение/контроль для Группы 2 составляло 52,79.
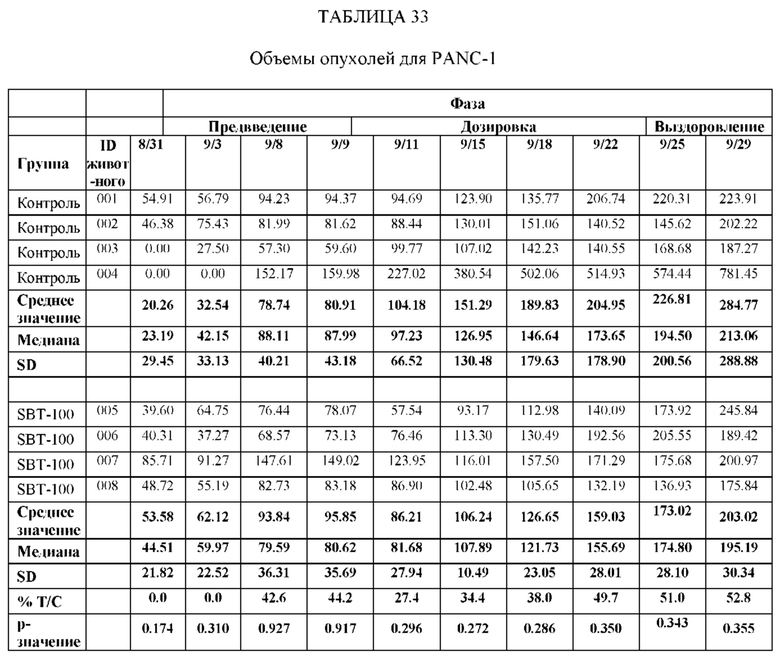
ПРИМЕР 12: ЭФФЕКТИВНОСТЬ ПОЛУЧЕННОГО В БАКТЕРИИ VHH13-ОДАТ (SEQ ID №: 3) К STAT3 В КСЕНОТРАНСПЛАНТАТНОЙ МОДЕЛИ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ ЧЕЛОВЕКА РЭ+/РП+ (MCF-7)
[0153] Этот Пример демонстрирует эффективность полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в ксенотрансплантатной модели рака молочной железы человека MCF-7 у "голых" мышей.
[0154] Самкам бестимусных "голых" мышей (Crl:NU(Ncr)-Foxn1nu, Charles River) было двенадцать недель, их масса тела (м.т.) находилась в диапазоне от 23,0 до 30,1 г на 1-й день исследования. Животных кормили и содержали, как описано выше.
[0155] Клетки карциномы молочной железы человека линии MCF-7 получали и культивировали, как описано выше, и использовали для модели ксенотрансплантата мыши. За три дня до имплантации опухолевых клеток, каждому подопытному животному подкожно в область между лопатками имплантировали пеллеты с эстрогеном (0,36 мг эстрадиола, с пролонгированным 60-дневным высвобождением, Innovative Research of America, Сарасота, штат Флорида) с использованием стерилизованного троакара.
[0156] Опухолевые клетки, используемые для имплантации, собирали во время логарифмической фазы роста и ресуспендировали в фосфатно-солевом буферном растворе (ФСБ) в концентрации, составляющей 1×108 клеток/мл. В день имплантации каждой подопытной мыши вводили 1×107 клеток MCF-7 (0,1 мл клеточной суспензии), имплантированных подкожно в правый бок, и контролировали рост опухолей по мере того, как средний размер приближался к целевому диапазону, составляющему 100-150 мм3. Через двадцатиоднодневный срок, принимаемый за 1-й день исследования, животных распределяли на две группы, каждая из которых состояла из четырех мышей с объемами отдельных опухолей, находящимися в пределах 108-144 мм3, и средними по группе объемами опухолей, составляющими 117-123 мм3.
[0157] Полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 предоставляли в виде предварительно смешанного, готового раствора для введения в концентрации, составляющей 0,41867 мг/мл в 1 мл аликвотах, и хранили при -20°C до возникновения необходимости в нем. 0,41867 мг/мл раствора обеспечивали 1 мг/кг дозы в объеме дозирования, составляющем 23,88 мл/кг. В каждый день лечения до комнатной температуры доводили только необходимые пробирки с полученным в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3. Все оставшиеся дозы суспензий хранили при 4°С на случай возникновения необходимости в следующей дозе.
[0158] Двум группам бестимусных "голых" мышей вводили дозу в соответствии с протоколом, показанным в Таблице 34. Все дозы носителя (контроль) и полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 вводили внутрибрюшинно (в/б) трижды в день, с интервалом в шесть часов в течение четырнадцати дней, с введением двух доз в 1-й день и введением одной дозы утром 15-го дня (ТД х 14, 2 дозы в первый день). Объем дозирования для носителя и полученного в бактерии VHH13-ОдАт (SEQ ID №:
3) к STAT3 составлял 0,478 мл на 20 грамм массы тела (23,88 мл/кг), и его подбирали пропорционально массе тела каждого отдельного животного. Группа 1 получала носитель и служила в качестве эталонной группы в отношении приживления и прогрессирования опухоли, а также в качестве контроля. Группа 2 получала полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в дозе, составляющей 1 мг/кг.

[0159] Опухоли измеряли дважды в неделю, и каждое животное умерщвляли, когда его новообразование достигало предварительно установленного конечного объема (1000 мм3) или в конце исследования, на 39-й день, в зависимости от того, какое из указанных событий наступало раньше. Когда опухоль достигала предварительно установленного конечного объема, животное регистрировали как умерщвленное по причине прогрессирования опухоли (TP), с указанием даты умерщвления. Время до конечной точки клинического исследования (TTE) для каждой мыши рассчитывали с помощью следующего уравнения:
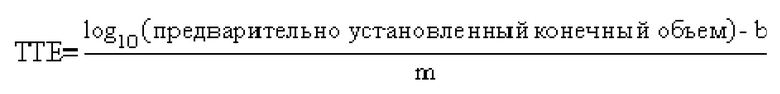
где TTE выражается в днях, предварительно установленный конечный объем выражается в мм3, b представляет собой свободный член уравнения регрессии и m представляет собой наклон линии, полученной с помощью линейной регрессии логарифмически преобразованного набора данных о росте опухоли. Набор данных состоит из первого наблюдения, которое превысило предварительно установленный конечный объем, используемый в анализе, и трех последовательных наблюдений, которые непосредственно предшествовали достижению этого предварительно установленного конечного объема. Расчетное TTE, как правило, меньше, чем дата TP - день, в который животное умерщвляли из-за размера опухоли. Животным, которые не достигали предварительно установленного объема конечной точки, присваивали значение TTE, равное последнему дню исследования (Д39). Любому животному, классифицированному, как умершее от связанных с лечением (TR) причин, должно было быть присвоено значение TTE, равное дню смерти. Любое животное, классифицированное, как умершее от не связанных с лечением (NTR) причин, должно было быть исключено из расчетов TTE.
[0160] Эффективность лечения определяли по замедлению роста опухоли (TGD), которая определяется как увеличение медианы TTE, выраженное в днях, для группы лечения по сравнению с контрольной группой:
TGD = T - C
Процентное увеличение медианы ТТЕ, по отношению к контрольной группе, представляет собой
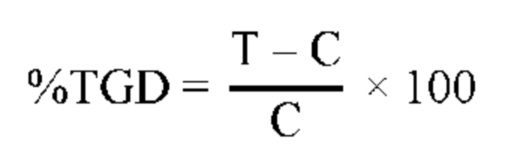
где:
T = медиана ТТЕ для группы лечения, и
C = медиана ТТЕ для обозначенной контрольной группы.
[0161] Эффективность лечения в каждой группе может быть обозначена медианой объема опухоли, MTV(n), которую определяли как медиану объема опухоли в последний день исследования (Д39) среди оставшихся животных (n), опухоли которых не достигли предварительно установленного конечного объема.
[0162] Эффективность лечения также можно определить по числу случаев и интенсивности регрессионных реакций, наблюдаемых во время исследования. Лечение может вызывать частичную регрессию (PR) или полную регрессию (CR) опухоли у животного. В реакции PR, объем опухоли составлял 50% или менее от его объема в Д1 в случае трех последовательных измерений в ходе исследования, и составлял не менее чем 13,5 мм3 в одном или более из этих трех измерений. В реакции CR, объем опухоли составлял менее чем 13,5 мм3 в случае трех последовательных измерений в ходе исследования. Любое животное с реакцией CR в конце исследования дополнительно классифицировали как выжившее с полной регрессией опухоли (TFS).
[0163] Животных взвешивали ежедневно в течение первых пяти дней, а затем дважды в неделю в течение оставшейся части исследования. Мышей часто осматривали на предмет состояния здоровья и выраженных признаков каких-либо неблагоприятных побочных эффектов, связанных с лечением TR, и фиксировали заслуживающие внимания клинические наблюдения. В соответствии с протоколом контролировали индивидуальную потерю веса, и любое животное с потерей веса, превышающей 30% за одно измерение, или превышающей 25% в течение трех измерений, подлежало умерщвлению по состоянию здоровья в виде TR смерти. Если средняя масса тела в группе восстанавливалась, в этой группе могли возобновить введение доз, но в более низкой дозе или с менее частым режимом дозирования. Приемлемую токсичность определяли как среднюю потерю м.т. в группе, составляющую менее чем 20% во время исследования, и не более чем од ин случай TR смерти на десять обработанных животных, или 10%. Любой режим применения препарата, приводящий к большей токсичности, рассматривается как превышающий максимальную переносимую дозу (MTD). Смерть классифицировали как TR, если она было связана с побочными эффектами лечения, о чем свидетельствовали клинические признаки и/или аутопсия, или также могла классифицироваться как TR, если произошла по неизвестным причинам в течение периода введения препарата или в течение 14 дней с момента введения последней дозы. Смерть классифицировали как NTR, если имелось доказательство того, что смерть была связана с моделью опухоли, а не с лечением. NTR смерти дополнительно классифицировали как NTRa (в результате несчастного случая или человеческой ошибки), NTRm (в результате подтвержденного аутопсией распространения из-за инвазии или метастазирования) и NTRu (по неизвестным причинам).
[0164] Для графических анализов использовали Prism 6.07 (GraphPad) для Windows. Статистику не использовали из-за небольшого объема выборки.
[0165] Диаграмму рассеяния строили, чтобы показать значения TTE для отдельных мышей, по группам; эта диаграмма показывает NTR смерти, которые были исключены из всех других фигур. Объемы опухолей отдельных животных, медианные и средние объемы опухолей в группе строили как функции времени. Когда животное выбывало из исследования по причине размера опухоли или TR смерти, его последний зафиксированный объем опухоли включали с данными, используемыми для расчета медианы объема в последующие моменты времени. График Каплана- Майера строили, чтобы показать процент животных в каждой группе, оставшихся в исследовании, в зависимости от времени. Кривые опухолевого роста усекали после того, как в одной и той же группе происходило два случая TR смерти. Средние изменения м.т. в группе в ходе исследования наносили на график, как процентное изменение, ± SEM, с 1-го дня. Кривые опухолевого роста и изменения м.т. усекали после того, как более половины анализируемых мышей в группе выбывало из исследования. На Фиг. 10 проиллюстрирован средний объем опухоли в исследовании.
[0166] Таблица 35 предоставляет средние потери м.т., TR и NTR смерти у мышей. При наблюдении фиксировали клинические признаки, как показано в Таблицах 36-38. Во время исследования случаев TR смертей не произошло. Потери веса были переменными, существенными для одного животного в каждой группе, и происходили в результате эстрогенного эффекта. Клинические наблюдения, в том числе потеря веса, увеличение рогов матки и соли в мочевом пузыре, присутствовали в обеих группах и были также обусловлены эстрогенным эффектом. Токсичность эстрогена привела к двум не связанным с лечением смертям в каждой группе. Оцененное в исследовании лечение имело приемлемую переносимость.
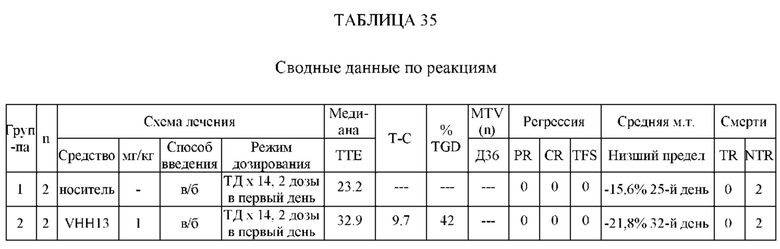
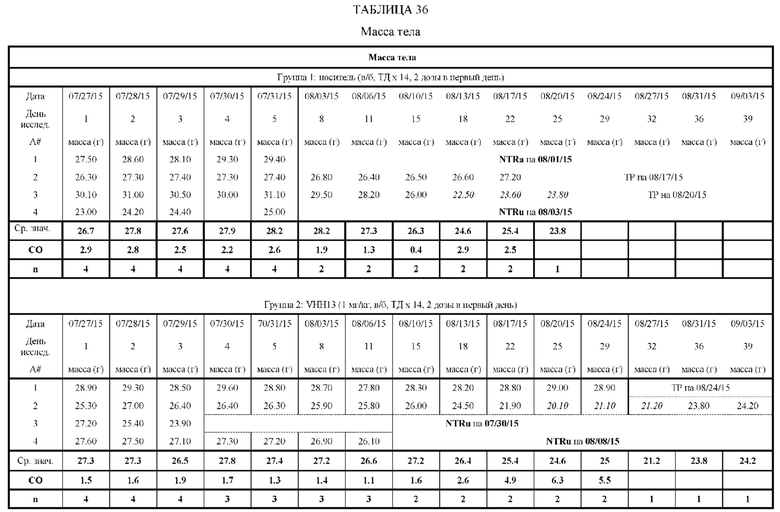
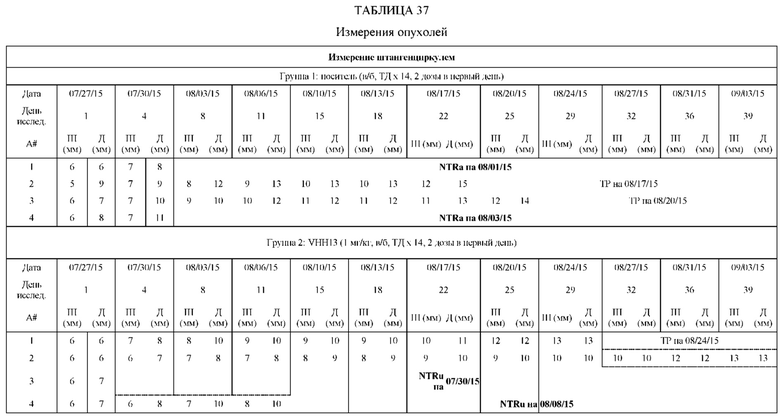
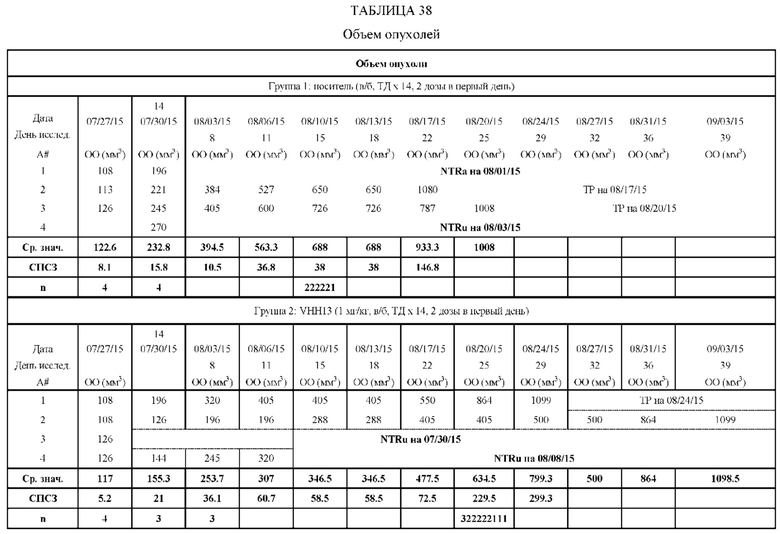
[0167] Так как две из четырех мышей в контрольной группе, а также в группе лечения умерли от токсичности эстрогена, нельзя сделать статистический вывод. Исходя из данных, медиана роста опухоли и средний объем опухоли уменьшились в группе лечения по сравнению с контрольной группой. Эта разница присутствовала в течение 14 дней лечения, а также до 25-го дня исследования. Контрольной группе потребовалось 25 дней, чтобы ее опухоли достигли объема, составляющего 1000 мм3, тогда как группе лечения потребовалось 36 дней, чтобы ее опухоли достигли объема, составляющего 1000 мм3. Это свидетельствует о том, что полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 замедляет рост опухоли MCF-7 in vivo. На протяжении всего исследования и в контрольной группе, и в группе лечения сохранялись аналогичные массы. Это свидетельствует о том, что полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 не вызывало токсичность по отношению к потере веса.
ПРИМЕР 13: ЛЕЧЕНИЕ ЧЕЛОВЕЧЕСКОГО РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ С HER2+ (BT474) С ПОМОЩЬЮ ПОЛУЧЕННОГО В БАКТЕРИИ VHH13-ОДАТ (SEQ ID №: 3) К STAT3 У КСЕНОТРАНСПЛАНТАТНЫХ МЫШЕЙ
[0168] В этом Примере определяли эффективность полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в ксенотрансплантате рака молочной железы человека BT474 у мышей CB.17 SCID.
[0169] Две группы 8-12 недельных мышей CB.17 SCID, содержащих ксенотрансплантаты из 1 мм3 фрагментов опухоли ВТ474 в своем боку, обрабатывали в соответствии с протоколом, показанным в Таблице 39, когда опухоли достигали среднего размера, составляющего 100-150 мм3. Все дозы носителя (ФСБ контроль) и полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 (показано в Таблице 39 как SB-01) вводили внутрибрюшинно (в/б) трижды в день, с интервалом в шесть часов в течение четырнадцати дней, с введением двух доз в 1-й день (ТД х 14, 2 дозы в первый день). Объем дозирования для носителя и полученного в бактерии VHH13- ОдАт (SEQ ID №: 3) к STAT3 составлял 0,478 мл на 20 грамм массы тела (23,88 мл/кг), и его подбирали пропорционально массе тела каждого отдельного животного. Группа 1 получала носитель и служила в качестве эталонной группы в отношении приживления и прогрессирования опухоли, а также в качестве контроля. Группа 2 получала полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 в дозе, составляющей 1 мг/кг.

[0170] В течение первых 14 дней исследования группа лечения получала B VHH13 к STAT3, а контрольная группа получала только носитель. Как показано в Таблице 40, в течение этого времени, группа лечения сохраняла и набирала вес на протяжении всего исследования, в то время как в контрольной группе масса тела была более низкой на протяжении всего исследования. Это свидетельствует о том, что группа лечения не испытывала токсичность от приема полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 по отношению к потере веса. Средний объем опухоли и медиана объема опухоли в обеих группах были сходными, и абсолютно одинаковыми на 15-й день исследования. На 59-й день исследования обе группы достигли объема опухоли, составляющего 700 мм3. Это свидетельствует о том, что полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 не уменьшало рост опухолей ВТ474 in vivo, по сравнению с контрольной группой. На Фиг. 11 проиллюстрирован средний объем опухоли в группе.
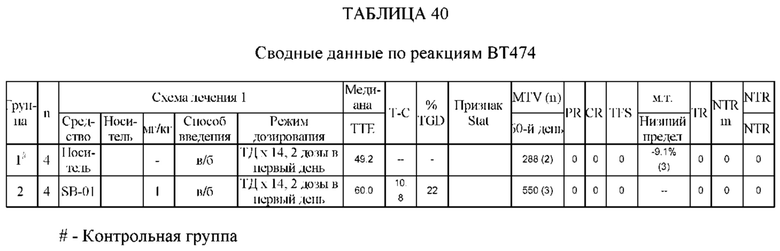
ПРИМЕР 14: ПОЛУЧЕНИЕ МЫШИНОГО МОНОКЛОНАЛЬНОГО АНТИТЕЛА, НАПРАВЛЕННОГО ПРОТИВ ПОЛУЧЕННОГО В БАКТЕРИИ VHH13-ОДАТ (SEQ ID №: 3) К STAT3
[0171] В этом Примере создавали мышиные моноклональные антитела к ОдАт по данному изобретению. Животными, которых использовали, были самки мышей BALB/c, в возрасте 8-10 недель. Использовали водорастворимый адъювант (CBL). Использованные HAT и HT были приобретены у Sigma-Aldrich.
[0172] Полученное в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3 использовали для иммунизации трех мышей и получения гибридомных клеточных линий. Мышей трижды иммунизировали, каждый раз водорастворимым адъювантом. В одной мыши сывороточные титры достигали 1/51200. Мышь умерщвляли и создавали гибридомные клеточные линии посредством слияния клеток селезенки с клеточной линией миеломы Sp2/0.
[0173] Слитые клетки высевали в 96-луночные планшеты посредством серийного разведения. Слитые клетки культивировали в присутствии HAT, и испытывали 651 отдельных клонов. Было установлено, что 27 положительных клонов из 651 отдельных клонов специфически связывались с антигеном полученного в бактерии VHH13-ОдАт (SEQ ID №: 3) к STAT3.
ПРИМЕР 15: ЦИТОТОКСИЧНОСТЬ ОДНОДОМЕННЫХ АНТИТЕЛ KRAS (G12D) В КЛЕТКАХ РАКА ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ ЧЕЛОВЕКА PANC-1
[0174] Этот Пример демонстрирует антипролиферативное действие ОдАт к KRAS (G12D) (SEQ ID №: 2) с использованием клеточной линии рака поджелудочной железы человека PANC-1. Для опытов, клетки PANC-1 выращивали до тех пор, пока они не достигали конфлюэнтности, составляющей 90%. В то же время, исследования пролиферации проводили с помощью метода MTT, как описано выше.
[0175] В Таблице 41 показаны антипролиферативные свойства ОдАт к KRAS (G12D) (SEQ ID №: 2) в клетках PANC-1 через три дня после обработки. Клетки PANC-1, обработанные ОдАт к KRAS (G12D) (SEQ ID №: 2), показали среднее ингибирование роста, составляющее 19,9 и 37,7 при 50,0 и 100 мкг/мл, соответственно.
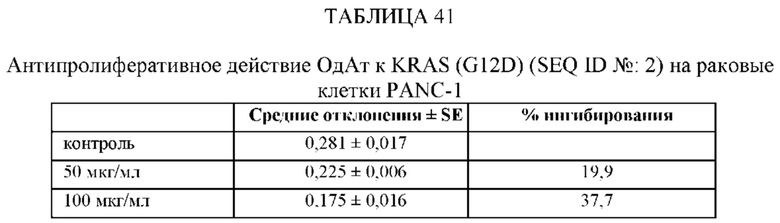
[0176] Таким образом, ОдАт к KRAS (G12D) (SEQ ID №: 2) показало дозозависимое ингибирование роста в клетках рака поджелудочной железы человека PANC-1.
ПРИМЕР 16: IN VITRO ИНГИБИРОВАНИЕ РОСТА С ПОМОЩЬЮ ОДАТ К ФНО- АЛЬФА
[0177] Этот Пример демонстрирует разработку методики определения концентрации ФНО-альфа и оценки ингибирования функции ФНО-альфа. Концентрацию ФНО-альфа, необходимую, чтобы показать измеряемую модуляцию активности в лимфобластной клеточной линии легкого человека U937, оценивали посредством количественного определения присутствующего АТФ, который сигнализирует о присутствии метаболически активных клеток, с использованием люминесцентного анализа жизнеспособности клеток CellTiter-GJo® от Promega.
[0178] Клетки U937 высевали в 96-луночный микрокультуральный планшет из прозрачного полистирола (96-луночный плоскодонный планшет Corning® Costar®, кат. № 3997) в общем объеме, составляющем 90 мкл/лунка. После 24 часов инкубирования в увлажненном инкубаторе при 37°C с 5% CO2 и 95% воздуха, в каждую лунку в двух повторностях (10-пунктовая кривая доза-ответ, самая высокая концентрация 20 нг/мл) добавляли 5 мкл 20-кратно серийно разведенного в среде для выращивания ФНО-альфа. Кроме того, в каждую лунку в двух повторностях (концентрация 1 нМ) добавляли 5 мкл 20-кратно разведенного в среде для выращивания стауроспорина.
[0179] После 24 часов культивирования в присутствии испытуемых веществ, концентрацию соединения, необходимую, чтобы показать измеряемую модуляцию активности ФНО-альфа в клеточной линии U937, оценивали посредством количественного определения присутствующего АТФ. Процентный рост клеток рассчитывали по отношению к необработанным контрольным лункам. Все испытания проводили в двух повторностях при каждом уровне концентрации.
[0180] Значение ЕС50 для испытуемых веществ оценивали с использованием Prism 6.05 посредством аппроксимации данных с помощью следующего четырехпараметрического логистического уравнения:
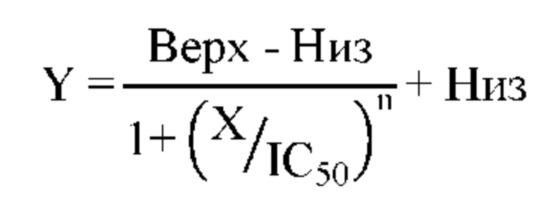
где Верх представляет собой максимальный % контрольного поглощения, Низ представляет собой минимальный % контрольного поглощения при самой высокой концентрации вещества, Y представляет собой % контрольного поглощения, X представляет собой концентрацию вещества, IC50 представляет собой концентрацию вещества, которое ингибирует рост клеток на 50% по сравнению с контрольными клетками, и n представляет собой наклон кривой.
[0181] На Фиг. 12 и 13 проиллюстрировано, что ФНО-альфа является цитотоксическим для клеток U937. IC50 для ФНО-альфа в отношении U937 составляет 95,10 пг/мл. Кривая ФНО-альфа показывает цитотоксический эффект титрования дозы.
[0182] На Фиг. 14 проиллюстрировано, что цитотоксичность ФНО-альфа в отношении U937 ингибируется тремя различными VHH к ФНО-альфа. Когда VHH62-ОдАт (SEQ ID №: 47) к ФНО-альфа, VHH66-ОдАт (SEQ ID №: 45) к ФНО-альфа и VHH69-ОдАт (SEQ ID №: 46) к ФНО-альфа инкубировали с постоянной дозой ФНО-альфа, при ЕС50, все три VHH к ФНО-альфа ингибировали цитотоксическую активность ФНО-альфа в отношении U937. IC50 VHH62-ОдАт (SEQ ID №: 47) к ФНО-альфа составляла около 713,6 мкг/мл. IC50 VHH69-ОдАт (SEQ ID №: 46) к ФНО-альфа составляла более чем 208,055 мкг/мл. IC50 VHH66-ОдАт (SEQ ID №: 45) к ФНО-альфа нельзя было определить, так как оно полностью ингибировало цитотоксичность ФНО-альфа при концентрациях, составляющих от около 1×102мкг/мл до 1×102 мкг/мл VHH66-ОдАт (SEQ ID №: 45) к ФНО-альфа. В этом диапазоне концентраций VHH66-ОдАт (SEQ ID №: 45) к ФНО-альфа происходит увеличение роста клеток U937 и, таким образом, полное ингибирование активности ФНО-альфа.
[0183] Хотя настоящее изобретение было описано со ссылкой на определенные предпочтительные варианты реализации достаточно подробно, возможны и другие варианты реализации. Этапы, описанные для способов по данному изобретению, например, не предназначены для ограничения, а также не предназначены для указания того, что каждый этап является обязательно неотъемлемым для данного способа, а наоборот представляют собой лишь типовые этапы. Таким образом, объем прилагаемой формулы изобретения не должен ограничиваться описанием предпочтительных вариантов реализации, содержащихся в данном описании. Все ссылки, приведенные в данном документе, включены посредством ссылки в полном объеме.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Singh, Sunanda
<120> Однодоменные антитела к внутриклеточным антигенам
<130> 7304-51293-4PCT
<150> US 62/067908
<151> 2014-10-23
<150> US 62/148656
<151> 2015-04-16
<150> US 62/188353
<151> 2015-07-02
<150> US 62/210795
<151> 2015-08-27
<160> 81
<170> PatentIn версия 3.5
<210> 1
<211> 372
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 1
gaggtgcagc tggtggagtc tgggggaggc tcggtgcaga ctggagggtc tctgagactc 60
tcctgtgcag tttctggaaa tatcggcagc agctactgca tgggctggtt ccgccaggct 120
ccagggaaga agcgcgaggc ggtcgcacgt attgtacgtg atggtgccac tggctacgca 180
gactacgtga agggccgatt caccatctcc cgagacagcg ccaagaacac tctgtatctg 240
caaatgaaca ggctgatacc tgaggacact gccatctact actgtgcggc agacctgccc 300
ccaggttgtt tgactcaggc gatttggaat tttggttatc ggggccaggg aaccctggtc 360
accgtctcct ca 372
<210> 2
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 2
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Thr Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Asn Ile Gly Ser Ser Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Lys Arg Glu Ala Val
35 40 45
Ala Arg Ile Val Arg Asp Gly Ala Thr Gly Tyr Ala Asp Tyr Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ala Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Arg Leu Ile Pro Glu Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Ala Asp Leu Pro Pro Gly Cys Leu Thr Gln Ala Ile Trp Asn Phe Gly
100 105 110
Tyr Arg Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 3
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 3
His Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 4
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 4
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Thr Tyr Thr Gly Cys Met Gly
20 25 30
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala Ala Leu
35 40 45
Ser Ser Arg Gly Phe Ala Gly His Tyr Thr Asp Ser Val Lys Gly Arg
50 55 60
Phe Ser Ile Ser Arg Asp Tyr Val Lys Asn Ala Val Tyr Leu Gln Met
65 70 75 80
Asn Thr Val Lys Pro Glu Asp Ala Ala Met Tyr Tyr Cys Ala Ala Arg
85 90 95
Glu Gly Trp Glu Cys Gly Glu Thr Trp Leu Asp Arg Thr Ala Gly Gly
100 105 110
His Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 5
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 5
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Thr Tyr Thr Gly Cys Met Gly
20 25 30
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala Ala Leu
35 40 45
Ser Ser Arg Gly Phe Ala Gly His Tyr Thr Asp Ser Val Lys Gly Arg
50 55 60
Phe Ser Ile Ser Arg Asp Tyr Val Lys Asn Ala Val Tyr Leu Gln Met
65 70 75 80
Asn Thr Val Lys Pro Glu Asp Ala Ala Met Tyr Tyr Cys Ala Ala Arg
85 90 95
Glu Gly Trp Glu Cys Gly Glu Thr Trp Leu Asp Arg Thr Ala Gly Gly
100 105 110
His Thr Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 6
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 6
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Thr Tyr Thr Gly Cys Met Gly
20 25 30
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala Ala Leu
35 40 45
Ser Ser Arg Gly Phe Ala Gly His Tyr Thr Asp Ser Val Lys Gly Arg
50 55 60
Phe Ser Ile Ser Arg Asp Tyr Val Lys Asn Ala Val Tyr Leu Gln Met
65 70 75 80
Asn Thr Val Lys Pro Glu Asp Ala Ala Met Tyr Tyr Cys Ala Ala Arg
85 90 95
Glu Gly Trp Glu Cys Gly Glu Thr Trp Leu Asp Arg Thr Ala Gly Gly
100 105 110
His Thr Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 7
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 7
His Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Thr Tyr Thr Gly Cys Met Gly
20 25 30
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala Ala Leu
35 40 45
Ser Ser Arg Gly Phe Ala Gly His Tyr Thr Asp Ser Val Lys Gly Arg
50 55 60
Phe Ser Ile Ser Arg Asp Tyr Val Lys Asn Ala Val Tyr Leu Gln Met
65 70 75 80
Asn Thr Val Lys Pro Glu Asp Ala Ala Met Tyr Tyr Cys Ala Ala Arg
85 90 95
Glu Gly Trp Glu Cys Gly Glu Thr Trp Leu Asp Arg Thr Ala Gly Gly
100 105 110
His Thr Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 8
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 8
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Thr Tyr Thr Gly Cys Met Gly
20 25 30
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala Ala Leu
35 40 45
Ser Ser Arg Gly Phe Ala Gly His Tyr Thr Asp Ser Val Lys Gly Arg
50 55 60
Phe Ser Ile Ser Arg Asp Tyr Val Lys Asn Ala Val Tyr Leu Gln Met
65 70 75 80
Asn Thr Val Lys Pro Glu Asp Ala Ala Met Tyr Tyr Cys Ala Ala Arg
85 90 95
Glu Gly Trp Glu Cys Gly Glu Thr Trp Leu Asp Arg Thr Ala Gly Gly
100 105 110
His Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 9
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 9
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Thr Tyr Thr Gly Cys Met Gly
20 25 30
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala Ala Leu
35 40 45
Ser Ser Arg Gly Phe Ala Gly His Tyr Thr Asp Ser Val Lys Gly Arg
50 55 60
Phe Ser Ile Ser Arg Asp Tyr Val Lys Asn Ala Val Tyr Leu Gln Met
65 70 75 80
Asn Thr Val Lys Pro Glu Asp Ala Ala Met Tyr Tyr Cys Ala Ala Arg
85 90 95
Glu Gly Trp Glu Cys Gly Glu Thr Trp Leu Asp Arg Thr Ala Gly Gly
100 105 110
His Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 10
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 10
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Thr Tyr Thr Gly Cys Met Gly
20 25 30
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala Ala Leu
35 40 45
Ser Ser Arg Gly Phe Ala Gly His Tyr Thr Asp Ser Val Lys Gly Arg
50 55 60
Phe Ser Ile Ser Arg Asp Tyr Val Lys Asn Ala Val Tyr Leu Gln Met
65 70 75 80
Asn Thr Val Lys Pro Glu Asp Ala Ala Met Tyr Tyr Cys Ala Ala Arg
85 90 95
Glu Gly Trp Glu Cys Gly Glu Thr Trp Leu Asp Arg Thr Ala Gly Gly
100 105 110
His Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 11
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 11
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Thr Tyr Thr Gly Cys Met Gly
20 25 30
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala Ala Leu
35 40 45
Ser Ser Arg Gly Phe Ala Gly His Tyr Thr Asp Ser Val Lys Gly Arg
50 55 60
Phe Ser Ile Ser Arg Asp Tyr Val Lys Asn Ala Val Tyr Leu Gln Met
65 70 75 80
Asn Thr Val Lys Pro Glu Asp Ala Ala Met Tyr Tyr Cys Ala Ala Arg
85 90 95
Glu Gly Trp Glu Cys Gly Glu Thr Trp Leu Asp Arg Thr Ala Gly Gly
100 105 110
His Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 12
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 12
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Thr Tyr Thr Gly Cys Met Gly
20 25 30
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala Ala Leu
35 40 45
Ser Ser Arg Gly Phe Ala Gly His Tyr Thr Asp Ser Val Lys Gly Arg
50 55 60
Phe Ser Ile Ser Arg Asp Tyr Val Lys Asn Ala Val Tyr Leu Gln Met
65 70 75 80
Asn Thr Val Lys Pro Glu Asp Ala Ala Met Tyr Tyr Cys Ala Ala Arg
85 90 95
Glu Gly Trp Glu Cys Gly Glu Thr Trp Leu Asp Arg Thr Ala Gly Ser
100 105 110
His Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 13
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 13
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Thr Tyr Thr Gly Cys Met Gly
20 25 30
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala Ala Leu
35 40 45
Ser Ser Arg Gly Phe Ala Gly His Tyr Thr Asp Ser Val Lys Gly Arg
50 55 60
Phe Ser Ile Ser Arg Asp Tyr Val Lys Asn Ala Val Tyr Leu Gln Met
65 70 75 80
Asn Thr Val Lys Pro Glu Asp Ala Ala Met Tyr Tyr Cys Ala Ala Arg
85 90 95
Glu Gly Trp Glu Cys Gly Glu Thr Trp Leu Asp Arg Thr Ala Gly Gly
100 105 110
His Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 14
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 14
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Asp
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Thr Tyr Thr Gly Cys Met Gly
20 25 30
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala Ala Leu
35 40 45
Ser Ser Arg Gly Phe Ala Gly His Tyr Thr Asp Ser Val Lys Gly Arg
50 55 60
Phe Ser Ile Ser Arg Asp Tyr Val Lys Asn Ala Val Tyr Leu Gln Met
65 70 75 80
Asn Thr Val Lys Pro Glu Asp Ala Ala Met Tyr Tyr Cys Ala Ala Arg
85 90 95
Glu Gly Trp Glu Cys Gly Glu Thr Trp Leu Asp Arg Thr Ala Gly Gly
100 105 110
His Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 15
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 15
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 16
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 16
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Asn Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 17
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 17
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 18
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 18
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 19
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 19
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 20
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 20
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 21
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 21
His Val Gln Leu Val Glu Ser Glu Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 22
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 22
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 23
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 23
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 24
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 24
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 25
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 25
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 26
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 26
His Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 27
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 27
His Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 28
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 28
His Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 29
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 29
His Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ala Asn Gly Gly Arg Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Val Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ser Gly Ile Ser Thr Gly Gly Leu Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Thr Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Ser Arg Phe Asp Cys Tyr Arg Gly Ser Trp Phe Asn Arg Tyr
100 105 110
Met Tyr Asn Ser Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 30
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 30
gatgtgcagc tggtggagtc tgggggaggc tcggtgcagg ctggaggctc tctgagactc 60
tcctgtgtag cctctacata caccggctgc atgggctggt tccgccaggc tcctggaaag 120
gagcgcgagg gagtcgcagc tcttagtagc cgtggttttg ccgggcacta taccgactcc 180
gtgaagggcc gattctccat ctcccgagac tacgtcaaga atgcggtgta tctgcaaatg 240
aacactgtga aacctgagga cgctgccatg tactactgtg cagcacggga gggatgggag 300
tgcggtgaga cctggttgga ccggaccgcc gggggccata cctactgggg ccaggggacc 360
caggtcaccg tctcctca 378
<210> 31
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 31
caggtgcagc tggtggagtc tgggggaggc tcggtgcagg ctggaggctc tctgagactc 60
tcctgtgtag cctctacata caccggctgc atgggctggt tccgccaggc tcctggaaag 120
gagcgcgagg gagtcgcagc tcttagtagc cgtggttttg ccgggcacta taccgactcc 180
gtgaagggcc gattctccat ctcccgagac tacgtcaaga atgcggtgta tctgcaaatg 240
aacactgtga aacctgagga cgctgccatg tactactgtg cagcacggga gggatgggag 300
tgcggtgaga cctggttgga ccggaccgcc gggggccata cctactgggg ccaggggacc 360
ctggtcaccg tctcctca 378
<210> 32
<211> 381
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 32
caggtgcagc tggtggagtc tgggggaggc tcggtgcagg ctggagggtc tctgagactc 60
tcctgtgcag cctctggagc caatggtggt cggagctgca tgggctggtt ccgccaggtt 120
ccagggaagg agcgcgaggg ggtttctggt atttcaaccg gtggtcttat tacatactat 180
gccgactccg tgaagggccg attcaccatc tcccaagaca acaccaagaa cacgctgtat 240
ctgcaaatga acagcctgaa acctgaggac actgccatgt actactgtgc gacgagtcgg 300
tttgactgct atagaggctc ttggttcaac cgatatatgt ataacagttg gggccagggg 360
accctggtca ccgtctcctc a 381
<210> 33
<211> 381
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 33
catgtgcagc tggtggagtc tgggggaggc tcggtgcagg ctggagggtc tctgagactc 60
tcctgtgcag cctctggagc caacggtggt cggagctgca tgggctggtt ccgccaggtt 120
ccagggaagg agcgcgaggg ggtttctggt atttcaaccg gtggtcttat tacatactat 180
gccgactccg tgaagggccg attcaccatc tcccaagaca acaccaagaa cacgctgtat 240
ctgcaaatga acagcctgaa acctgaggac actgccatgt actactgtgc gacgagtcgg 300
tttgactgct atagaggctc ttggttcaac cgatatatgt ataacagttg gggccagggg 360
acccaggtca ctgtctcctc a 381
<210> 34
<211> 381
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 34
gatgtgcagc tggtggagtc tgggggaggc tcggtgcagg ctggagggtc tctgagactc 60
tcctgtgcag cctctggagc caatggtggt cggagctgca tgggctggtt ccgccaggtt 120
ccagggaagg agcgcgaggg ggtttctggt atttcaaccg gtggtcttat tacatactat 180
gccgactccg tgaagggccg attcaccatc tcccaagaca acaccaagaa cacgctgtat 240
ctgcaaatga acagcctgaa acctgaggac actgccatgt actactgtgc gacgagtcgg 300
tttgactgct atagaggctc ttggttcaac cgatatatgt ataacagttg gggccagggg 360
accctggtca ccgtctcctc a 381
<210> 35
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 35
caggtgcagc tggtggagtc tgggggaggc tcggtgcagg ctggaggctc tctgagactc 60
tcctgtgtag cctctacata caccggctgc atgggctggt tccgccaggc tcctggaaag 120
gagcgcgagg gagtcgcagc tcttagcagc cgtggttttg ccgggcacta taccgactcc 180
gtgaagggcc gattctccat ctcccgagac tacgtcaaga atgcggtgta tctgcaaatg 240
aacactgtga aacctgagga cgctgccatg tactactgtg cagcacggga gggatgggag 300
tgcggtgaga cctggttgga ccggaccgcc gggagccata cctactgggg ccaggggacc 360
ctggtcaccg tctcctca 378
<210> 36
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 36
gaggtgcagc tggtggagtc tgggggaggc tcggtgcagg ctggaggctc tctgagactc 60
tcctgtgtag cctctacata caccggctgc atgggctggt tccgccaggc tcctggaaag 120
gagcgcgagg gagtcgcagc tcttagtagc cgtggttttg ccgggcacta taccgactcc 180
gtgaagggcc gattctccat ctcccgagac tacgtcaaga atgcggtgta tctgcaaatg 240
aacactgtga aacctgagga cgctgccatg tactactgtg cagcacggga gggatgggag 300
tgcggtgaga cctggttgga ccgaaccgcc gggggccata cctactgggg ccaggggacc 360
ctggtcaccg tctcctca 378
<210> 37
<211> 381
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 37
gaggtgcagc tggtggagtc tgggggaggc tcggtgcagg ctggagggtc tctgagactc 60
tcctgtgcag cctctggagc caatggtggt cggagctgca tgggctggtt ccgccaggtt 120
ccagggaagg agcgcgaggg ggtttctggt atttcaaccg gtggtcttat tacatactat 180
gccgactccg tgaagggtcg attcaccatc tcccaagaca acaccaagaa cacgctgtat 240
ctgcaaatga gcagcctgaa acctgaggac actgccatgt actactgtgc gacgagtcgg 300
tttgactgct atagaggctc ttggttcaac cgatatatgt ataacagttg gggccagggg 360
acccaggtca ccgtctcctc a 381
<210> 38
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 38
catgtgcagc tggtggagtc tggggggggc tcggtgcagg ctggaggctc tctgagactc 60
tcctgtgtag cctctacata caccggctgc atgggctggt tccgccaggc tcctggaaag 120
gagcgcgagg gagtcgcagc tcttagtagc cgtggttttg ccgggcacta taccgactcc 180
gtgaagggcc gattctccat ctcccgagac tacgtcaaga atgcggtgta tctgcaaatg 240
aacactgtga aacctgagga cgctgccatg tactactgtg cagcacggga gggatgggag 300
tgcggtgaga cctggttgga ccggaccgcc gggggccata cctactgggg ccaggggacc 360
caggtcaccg tctcctca 378
<210> 39
<211> 381
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 39
gatgtgcagc tggtggagtc tgggggaggc tcggtgcagg ctggagggtc tctgagactc 60
tcctgtgcag cctctggagc caatggtggt cggagctgca tgggctggtt ccgccaggtt 120
ccagggaagg agcgcgaggg ggtttctggt atttcaaccg gtggtcttat tacatactat 180
gccgactccg tgaagggccg attcaccatc tcccaagaca acaccaagaa cacgctgtat 240
ctgcaaatga acagcctgaa acctgaggac actgccatgt actactgtgc gacgagtcgg 300
tttgactgct atagaggctc ttggttcaac cgatatatgt ataacagttg gggccagggg 360
acccaggtca ccgtctcctc a 381
<210> 40
<211> 381
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 40
caggtgcagc tggtggagtc tgggggaggc tcggtgcagg ctggagggtc tctgagactc 60
tcctgtgcag cctctggagc caatggtggt cggagctgca tgggctggtt ccgccaggtt 120
ccagggaagg agcgcgaggg ggtttctggt atttcaaccg gtggtcttat tacatactat 180
gccgactccg tgaagggccg attcaccatc tcccaagaca acaccaagaa cacgctgtat 240
ctgcaaatga acagcctgaa acctgaggac actgccatgt actactgtgc gacgagtcgg 300
tttgactgct atagaggctc ttggttcaac cgatatatgt ataacagttg gggccagggg 360
acccaggtca ccgtctcctc a 381
<210> 41
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 41
gatgtgcagc tggtggagtc tgggggaggc tcggtgcagg ctggagactc tctgagactc 60
tcctgtgtag cctctacata caccggctgc atgggctggt tccgccaggc tcctggaaag 120
gagcgcgagg gagtcgcagc tcttagtagc cgtggttttg ccgggcacta taccgactcc 180
gtgaagggcc gattctccat ctcccgagac tacgtcaaga atgcggtgta tctgcaaatg 240
aacactgtga aacctgagga cgctgccatg tactactgtg cagcacggga gggatgggag 300
tgcggtgaga cctggttgga ccggaccgcc gggggccata cctactgggg ccaggggacc 360
ctggtcactg tctcctca 378
<210> 42
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 42
gtgcagctgg tggagtctgg gggaggctcg gtgcaggctg gagggtctct gagactctcc 60
tgtgcagcct ctggagccaa tggtggtcgg agctgcatgg gctggttccg ccaggttcca 120
gggaaggagc gtgagggggt ttctggtatt tcaaccggtg gtcttattac atactatgcc 180
gactccgtga agggccgatt caccatctcc caagacaaca ccaagaacac gctgtatctg 240
caaatgaaca gcctgaaacc tgaggacact gccatgtact actgtgcgac gagtcggttt 300
gactgctata gaggctcttg gttcaaccga tatatgtata acagttgggg ccaggggacc 360
ctggtcactg tctcctca 378
<210> 43
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 43
gtgcagctgg tggagtctga gggaggctcg gtgcaggctg gagggtctct gagactctcc 60
tgtgcagcct ctggagccaa tggtggtcgg agctgcatgg gctggttccg ccaggttcca 120
gggaaggagc gcgagggggt ttctggtatt tcaaccggtg gtcttattac atactatgcc 180
gactccgtga agggccgatt caccatctcc caagacaaca ccaagaacac gctgtatctg 240
caaatgaaca gcctgaaacc tgaggacact gccatgtact actgtgcgac gagtcggttt 300
gactgctata gaggctcttg gttcaaccga tatatgtata acagttgggg ccaggggacc 360
ctggtcaccg tctcctca 378
<210> 44
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 44
gtgcagctgg tggagtctgg gggaggctcg gtgcaggctg gagggtctct gagactctcc 60
tgtgcagcct ctggagccaa tggtggtcgg agctgcatgg gctggttccg ccaggttcca 120
gggaaggagc gcgagggggt ttctggtatt tcaaccggtg gtcttattac atactatgcc 180
gactccgtga agggccgatt caccatctcc caagacaaca ccaataacac gctgtatctg 240
caaatgaaca gcctgaaacc tgaggacact gccatgtact actgtgcgac gagtcggttt 300
gactgctata gaggctcttg gttcaaccga tatatgtata acagttgggg ccaggggacc 360
ctggtcactg tctcctca 378
<210> 45
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 45
His Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Glu Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Tyr Ala Ala Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Asp Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Asp Ile Asp Gly Leu Thr Thr His Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser Leu
65 70 75 80
Gln Met Asn Asp Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Asp Arg Asp Arg Cys Gly Ser Ile Trp Thr Tyr Ala Tyr Lys Tyr
100 105 110
Arg Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 46
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 46
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Leu Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Ser Arg Tyr Asn
20 25 30
Tyr Met Ala Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Gly Thr Ala Ser Gly Ser Ala Asp Tyr Tyr Gly Ser Val
50 55 60
Lys Asp Arg Phe Thr Ile Ser Gln Asp Asn Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Ala Arg Thr Tyr Gly Thr Ile Ser Leu Thr Pro Ser Asp Tyr Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 47
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 47
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Pro Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Ser Trp Ala Gln Gly Thr Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser
<210> 48
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 48
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Glu Ala Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Tyr Ala Ala Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Asp Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Asp Ile Asp Gly Gln Thr Thr His Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser Leu
65 70 75 80
Gln Met Asn Asp Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Asp Arg Asp Arg Cys Gly Ser Ile Trp Thr Tyr Ala Tyr Lys Tyr
100 105 110
Arg Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 49
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 49
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Glu Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Tyr Ala Ala Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Asp Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Asp Ile Asp Gly Gln Thr Thr His Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Lys Asn Thr Leu Ser Leu
65 70 75 80
Gln Met Asn Asp Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Asp Arg Asp Arg Cys Gly Ser Ile Trp Thr Tyr Ala Tyr Lys Tyr
100 105 110
Arg Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 50
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 50
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Glu Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Tyr Ala Ala Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Asp Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Asp Ile Asp Gly Leu Thr Thr His Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser Leu
65 70 75 80
Gln Met Asn Asp Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Asp Arg Asp Arg Cys Gly Ser Ile Trp Thr Tyr Ala Tyr Lys Tyr
100 105 110
Arg Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 51
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 51
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Glu Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Tyr Ala Ala Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Asp Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Asp Ile Asp Gly Gln Thr Thr His Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser Leu
65 70 75 80
Gln Met Asn Asp Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Asp Arg Asp Arg Cys Gly Ser Ile Trp Thr Tyr Ala Tyr Lys Tyr
100 105 110
Arg Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 52
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 52
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Glu Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Tyr Ala Ala Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Asp Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Asp Ile Asp Gly Gln Thr Thr His Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser Leu
65 70 75 80
Gln Met Asn Asp Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Asp Arg Asp Arg Cys Gly Ser Ile Trp Thr Tyr Ala Tyr Lys Tyr
100 105 110
Arg Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 53
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 53
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Glu Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Tyr Ala Ala Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Asp Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Asp Ile Asp Gly Leu Thr Thr His Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser Leu
65 70 75 80
Gln Met Asn Asp Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Asp Arg Asp Arg Cys Gly Ser Ile Trp Thr Tyr Ala Tyr Lys Tyr
100 105 110
Arg Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 54
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 54
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Glu Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Tyr Ala Ala Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Asp Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Asp Ile Asp Gly Gln Thr Thr His Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser Leu
65 70 75 80
Gln Met Asn Asp Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Asp Arg Asp Arg Cys Gly Ser Ile Trp Thr Tyr Ala Tyr Lys Tyr
100 105 110
Arg Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 55
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 55
His Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Glu Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Tyr Ala Ala Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Asp Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Asp Ile Asp Gly Leu Thr Thr His Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser Leu
65 70 75 80
Gln Met Asn Asp Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Asp Arg Asp Arg Cys Gly Ser Ile Trp Thr Tyr Ala Tyr Lys Tyr
100 105 110
Arg Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 56
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 56
His Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Glu Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Tyr Ala Ala Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Asp Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Asp Ile Asp Gly Leu Ala Thr His Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser Leu
65 70 75 80
Gln Met Asn Asp Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Asp Arg Asp Arg Cys Gly Ser Ile Trp Thr Tyr Ala Tyr Lys Tyr
100 105 110
Arg Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 57
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 57
His Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Glu Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Tyr Ala Ala Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Asp Arg Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Asp Ile Asp Gly Gln Thr Thr His Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser Leu
65 70 75 80
Gln Met Asn Asp Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Asp Arg Asp Arg Cys Gly Ser Ile Trp Thr Tyr Ala Tyr Lys Tyr
100 105 110
Arg Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 58
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 58
His Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Tyr Ala Ala Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Asp Gly Lys Val Arg Glu Gly Val
35 40 45
Ala Thr Ile Asp Ile Asp Gly Gln Thr Thr His Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser Leu
65 70 75 80
Gln Met Asn Asp Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Asp Arg Asp Arg Cys Gly Ser Ile Trp Thr Tyr Ala Tyr Lys Tyr
100 105 110
Arg Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 59
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 59
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Asp Ser Phe Gly
20 25 30
Val Met Ala Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Ala Val Tyr Arg Arg Ala Gly Asp Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Ser Ala Met Tyr Tyr Cys
85 90 95
Ala Ala Arg Thr Tyr Gly Ser Val Ser Ser Trp Thr Gly Tyr Lys Tyr
100 105 110
Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 60
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 60
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Asp Ser Phe Gly
20 25 30
Val Met Ala Trp Phe Arg Gln Thr Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Ala Val Tyr Arg Arg Ala Gly Asp Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Ser Ala Met Tyr Tyr Cys
85 90 95
Ala Ala Arg Thr Tyr Gly Ser Val Ser Ser Trp Thr Gly Tyr Lys Tyr
100 105 110
Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 61
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 61
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Val Gly Gly
1 5 10 15
Ser Leu Thr Leu Ser Cys Ala Val Ser Gly Tyr Thr Asp Ser Tyr Gly
20 25 30
Val Met Ala Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Ser Ile Tyr Arg Asn Ser Gly Ile Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Leu
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Ser Ala Thr Tyr Tyr Cys
85 90 95
Ala Val Arg Ser Phe Gly Ser Val Ser Thr Trp Ala Gly Tyr Val Tyr
100 105 110
Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 62
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 62
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Asp Ser Phe Gly
20 25 30
Val Met Ala Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Ala Val Tyr Arg Arg Ala Gly Asp Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Ser Ala Met Tyr Tyr Cys
85 90 95
Ala Ala Arg Thr Tyr Gly Ser Val Ser Ser Trp Thr Gly Tyr Lys Tyr
100 105 110
Trp Gly Arg Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 63
<211> 125
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 63
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Arg Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Asp Thr Ser Lys Ser Asp
20 25 30
Cys Met Ala Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Arg Val
35 40 45
Gly Ala Ile Tyr Thr Arg Asn Gly Tyr Thr His Tyr Ala Asp Ser Val
50 55 60
Asn Gly Arg Phe Thr Ile Ser Gln Asp Asn Ala Lys Asn Ala Leu Tyr
65 70 75 80
Leu Gln Met Ser Gly Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Ala Arg Phe Arg Ile Tyr Gly Gln Cys Val Glu Asp Asp Asp Ile
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 64
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 64
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Leu Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Ser Arg Tyr Asn
20 25 30
Tyr Met Ala Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala Thr Ile Gly Thr Ala Ser Gly Ser Ala Asp Tyr Tyr Gly Ser Val
50 55 60
Lys Asp Arg Phe Thr Ile Ser Gln Asp Asn Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Ala Arg Thr Tyr Gly Thr Ile Ser Leu Thr Pro Ser Asp Tyr Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 65
<211> 128
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 65
Gln Val Gln Val Val Glu Tyr Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Val Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Trp Glu Leu Val
35 40 45
Ser Asn Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Ser Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Phe Asp Asn Ala Lys Asn
65 70 75 80
Met Val Tyr Leu Gln Met Asn Ser Leu Lys His Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Arg Glu Tyr
100 105 110
Cys Thr Trp Ala Gln Gly Thr Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 66
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 66
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Pro Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Ser Trp Ala Gln Gly Thr Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser
<210> 67
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 67
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Thr Trp Ala Gln Gly Thr Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser
<210> 68
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 68
Gln Val Gln Pro Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Thr Trp Ala Gln Gly Ala Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser
<210> 69
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 69
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Ser Trp Ala Gln Gly Thr Gln Gly Thr Gln Val Thr Val Ser
115 120 125
Ser
<210> 70
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 70
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Thr Trp Ala Gln Gly Thr Gln Gly Thr Gln Val Thr Val Ser
115 120 125
Ser
<210> 71
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 71
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Thr Trp Ala Gln Gly Thr Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser
<210> 72
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 72
His Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Thr Trp Ala Gln Gly Thr Gln Gly Thr Gln Val Thr Val Ser
115 120 125
Ser
<210> 73
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 73
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Thr Trp Ala Gln Gly Thr Gln Gly Ala Leu Val Thr Val Ser
115 120 125
Ser
<210> 74
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 74
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Thr Trp Ala Gln Gly Thr Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser
<210> 75
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 75
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Ser Trp Ala Gln Gly Thr Gln Gly Thr Gln Val Thr Val Ser
115 120 125
Ser
<210> 76
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 76
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Thr Trp Ala Gln Gly Thr Gln Gly Thr Gln Val Thr Val Ser
115 120 125
Ser
<210> 77
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 77
Asp Val Gln Leu Val Glu Ser Arg Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Thr Trp Ala Gln Gly Thr Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser
<210> 78
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 78
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Val Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Ser Trp Ala Gln Gly Thr Gln Gly Thr Gln Val Thr Val Ser
115 120 125
Ser
<210> 79
<211> 128
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 79
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly Leu Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Ala
50 55 60
Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
65 70 75 80
Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Glu Tyr
100 105 110
Cys Thr Trp Ala Gln Gly Thr Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 80
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 80
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Glu Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Thr Trp Ala Gln Gly Thr Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser
<210> 81
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Верблюдовые
<400> 81
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Glu
1 5 10 15
Thr Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ala Glu Ala
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly His Glu Cys Glu Leu Val
35 40 45
Ser Thr Ile Thr Thr Glu Gly Ile Thr Ser Val Ala Ser Ser Tyr Tyr
50 55 60
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
65 70 75 80
Asn Met Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
85 90 95
Val Tyr Tyr Cys Ala Pro Asp Pro Tyr Ala Tyr Ser Thr Tyr Ser Asp
100 105 110
Tyr Cys Thr Trp Ala Gln Gly Thr Gln Gly Thr Gln Val Thr Val Ser
115 120 125
Ser
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАРИАНТ Fc И ЕГО ПОЛУЧЕНИЕ | 2021 |
|
RU2828852C1 |
| АНТИ-МЕТ АНТИТЕЛА И ИХ ИСПОЛЬЗОВАНИЕ | 2017 |
|
RU2765267C2 |
| АНТИТЕЛА К PAC1 И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2781553C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ПРОМЫШЛЕННЫХ ПРОДУКТОВ ИЗ РАСТИТЕЛЬНЫХ ЛИПИДОВ | 2015 |
|
RU2743384C2 |
| Химерные антигенные рецепторы, нацеливающиеся на CD70 | 2019 |
|
RU2801824C2 |
| ВОССТАНОВЛЕНИЕ ФЕРТИЛЬНОСТИ У РАСТЕНИЙ | 2019 |
|
RU2831789C2 |
| АНТИТЕЛА К ФАКТОРУ XI | 2017 |
|
RU2800719C2 |
| РАСТЕНИЯ С МОДИФИЦИРОВАННЫМИ ПРИЗНАКАМИ | 2017 |
|
RU2809117C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, КОТОРОЕ СПЕЦИФИЧЕСКИ СВЯЗЫВАЕТСЯ С IL-4Rα, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2808563C2 |
| ИММУНОГЛОБУЛИН С ТАНДЕМНЫМИ FAB-ФРАГМЕНТАМИ И ВАРИАНТЫ ЕГО ПРИМЕНЕНИЯ | 2017 |
|
RU2767329C2 |
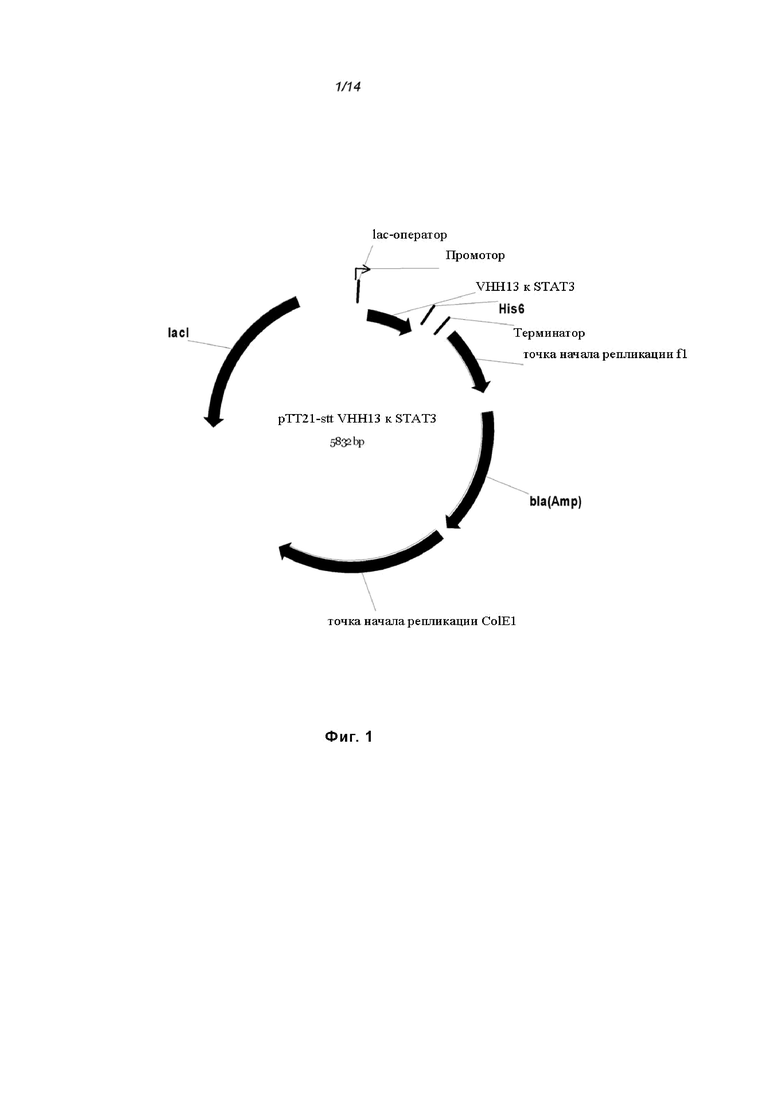
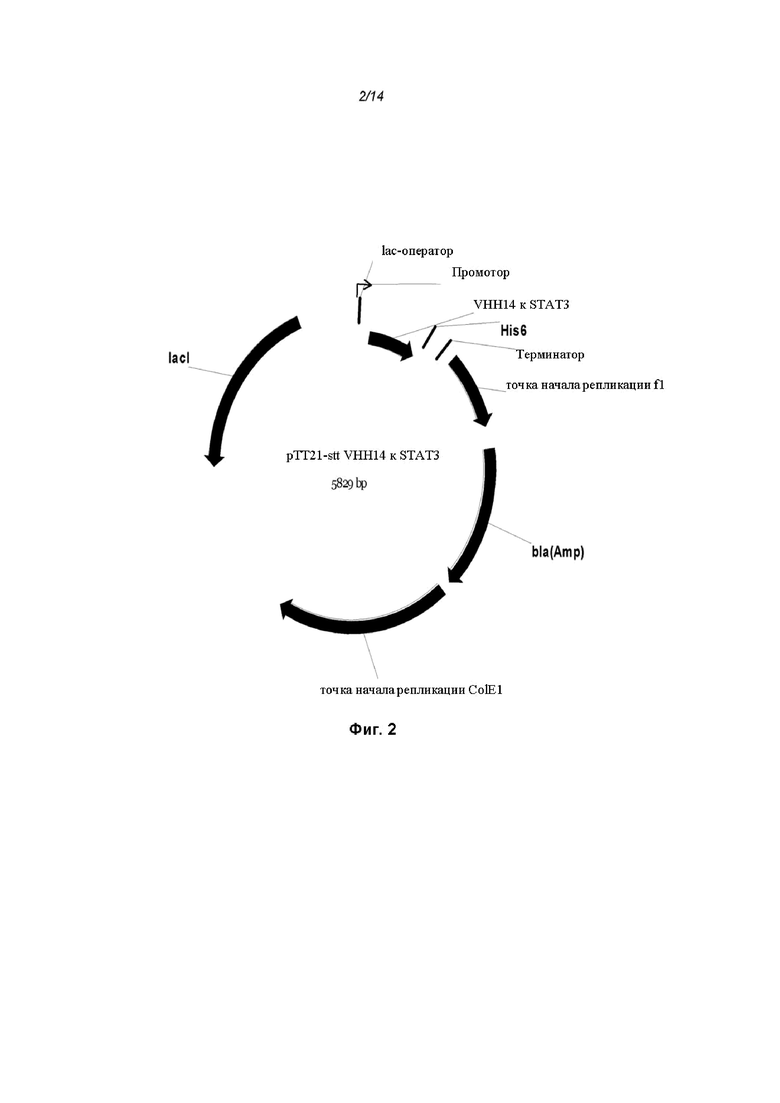
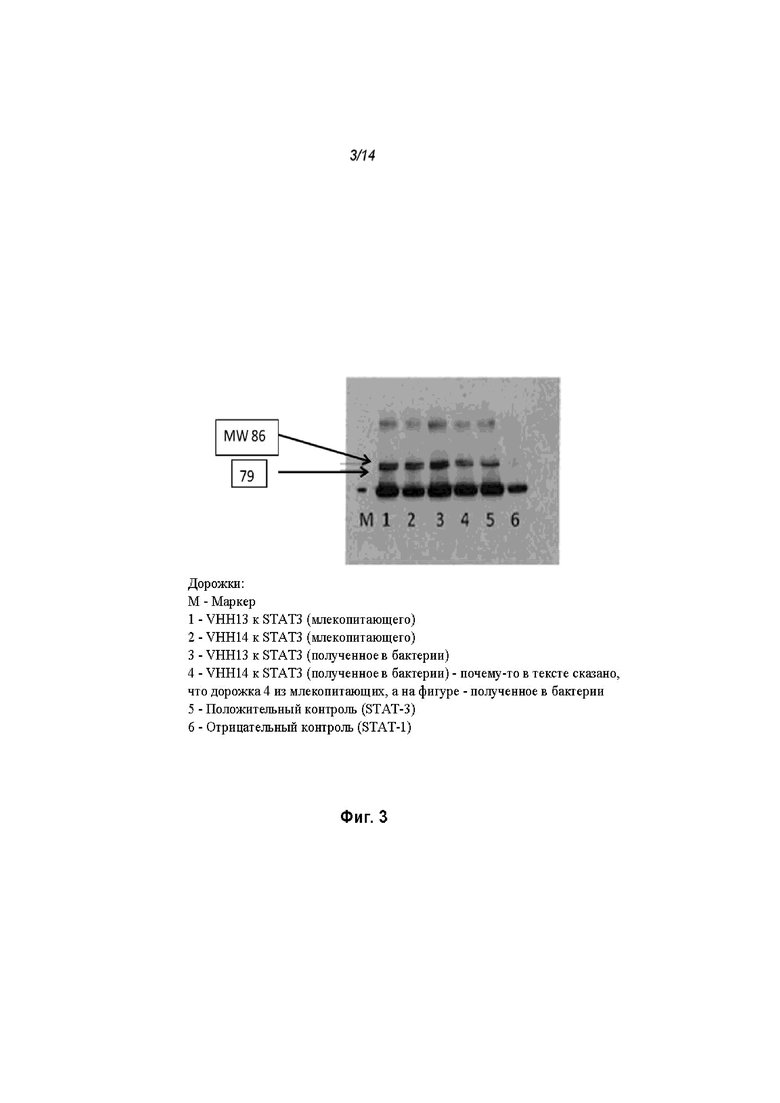

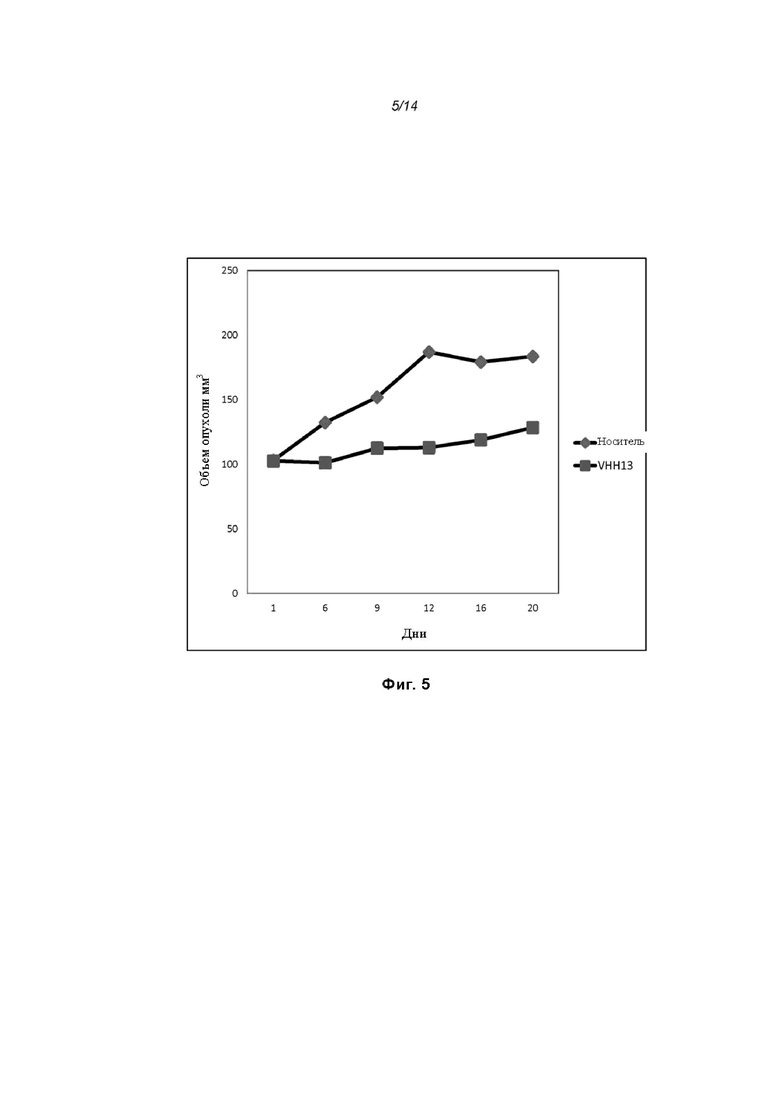
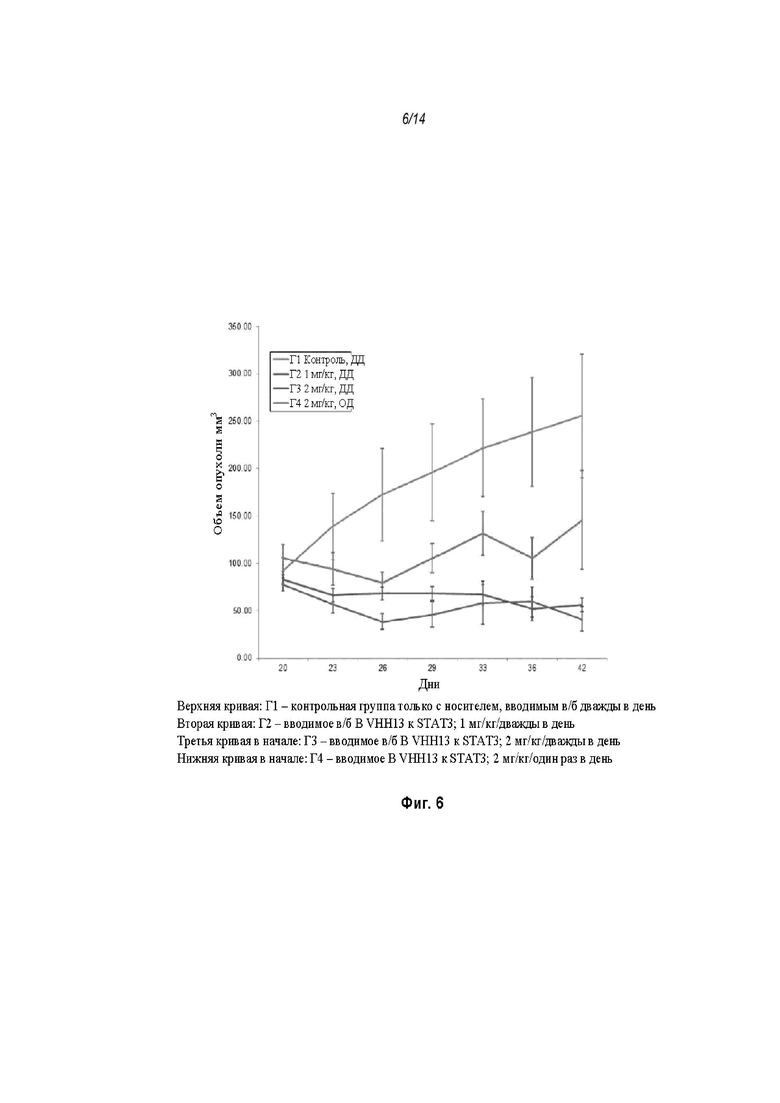
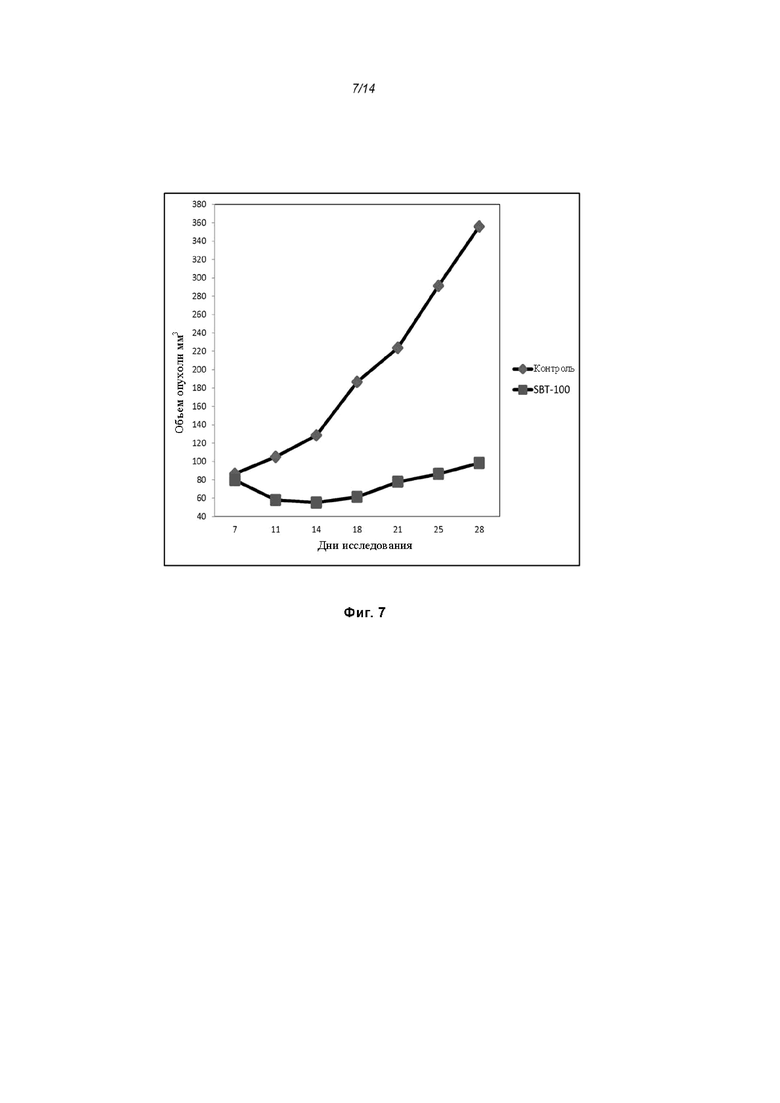
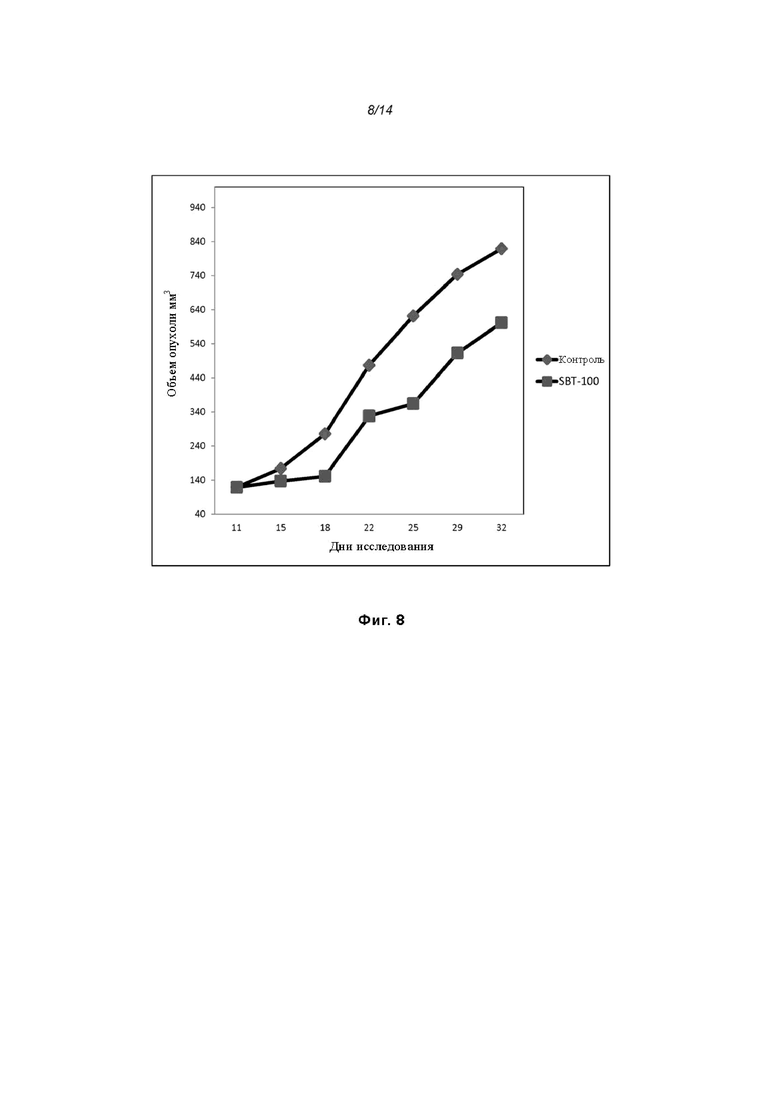
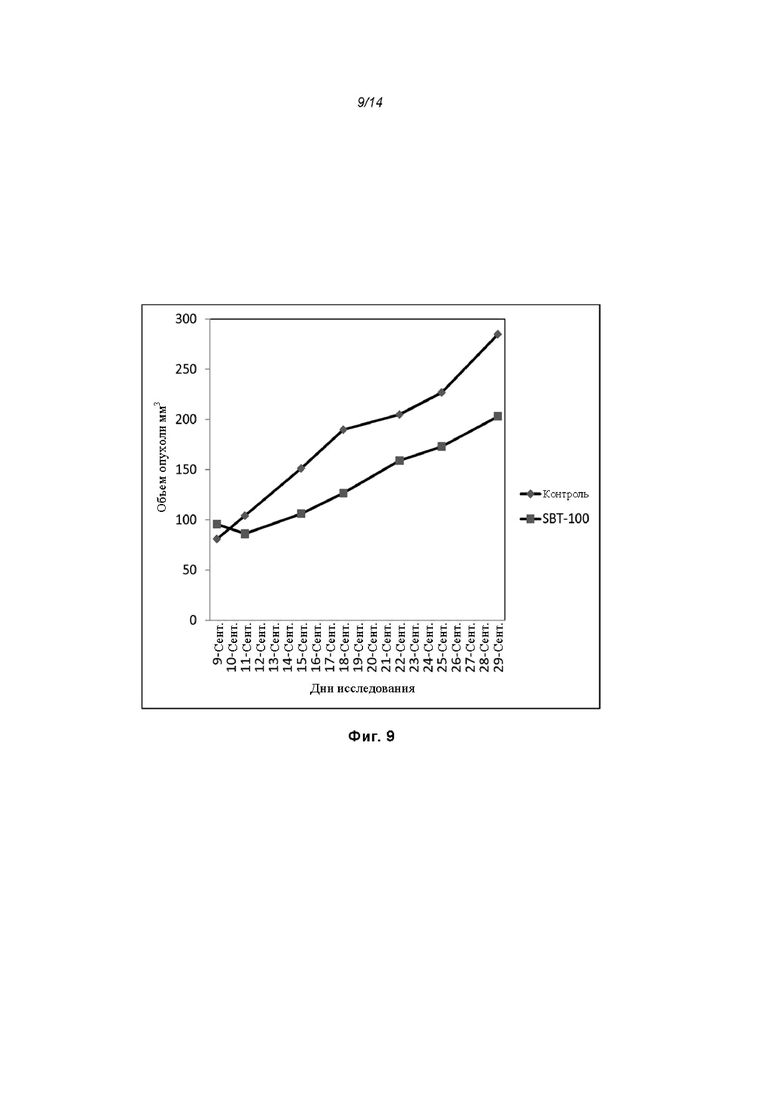

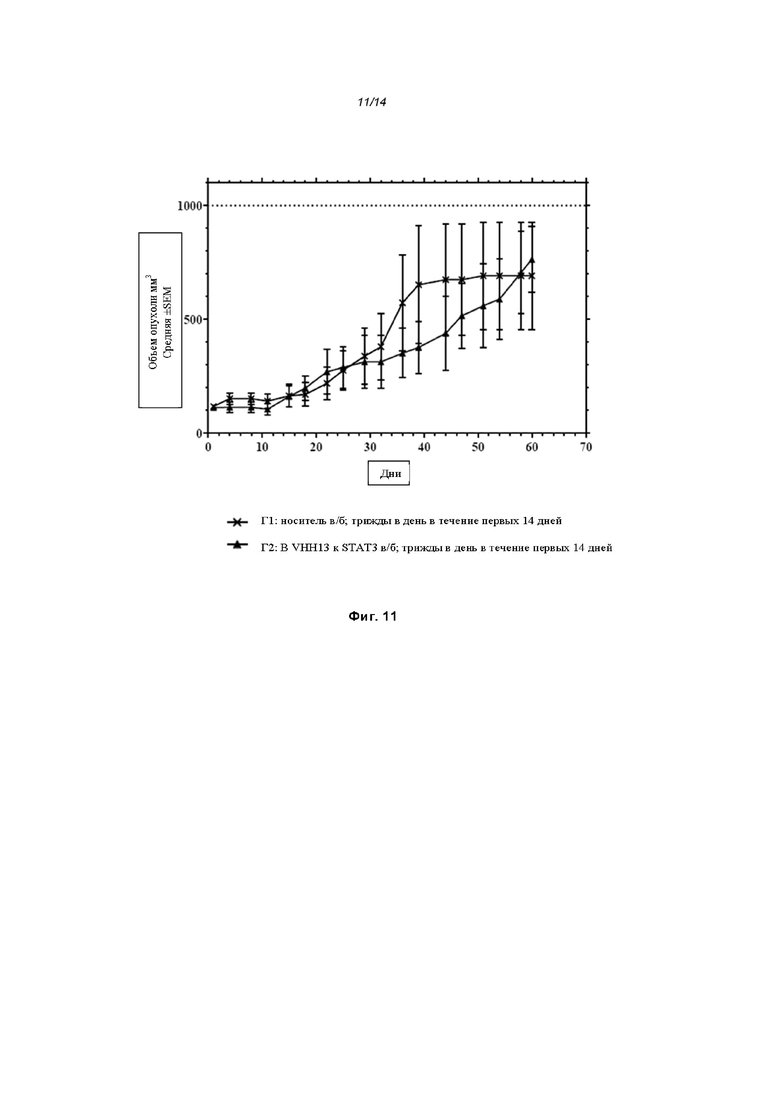
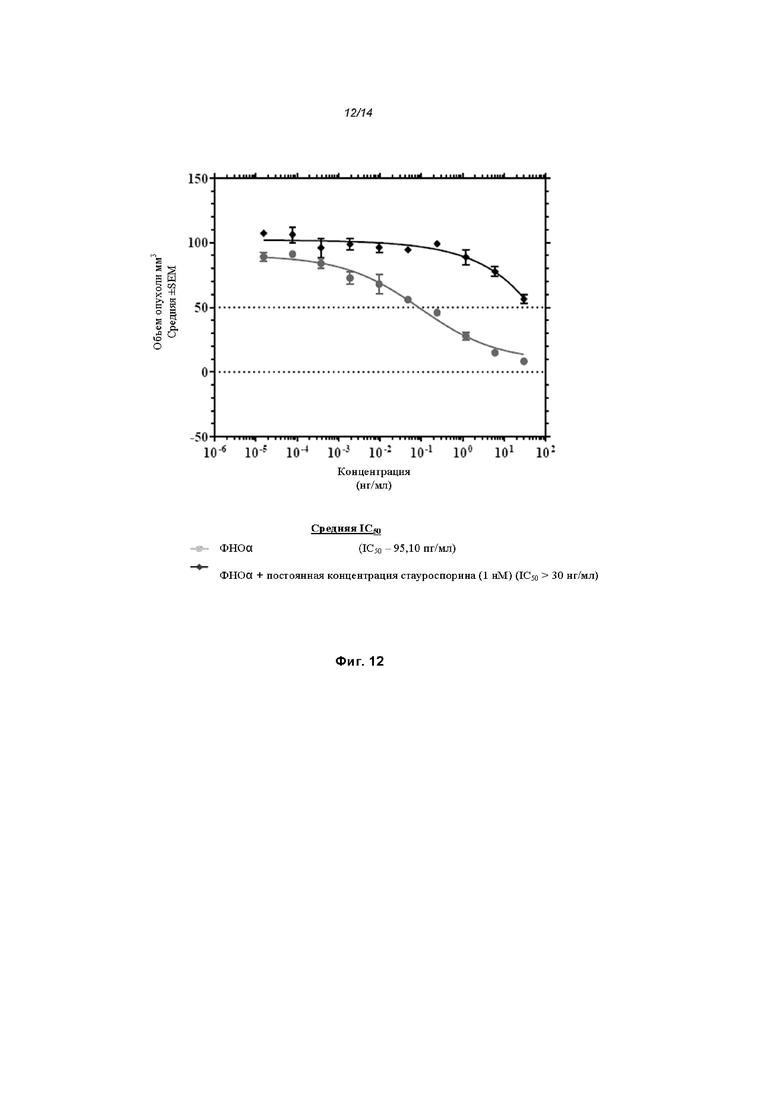
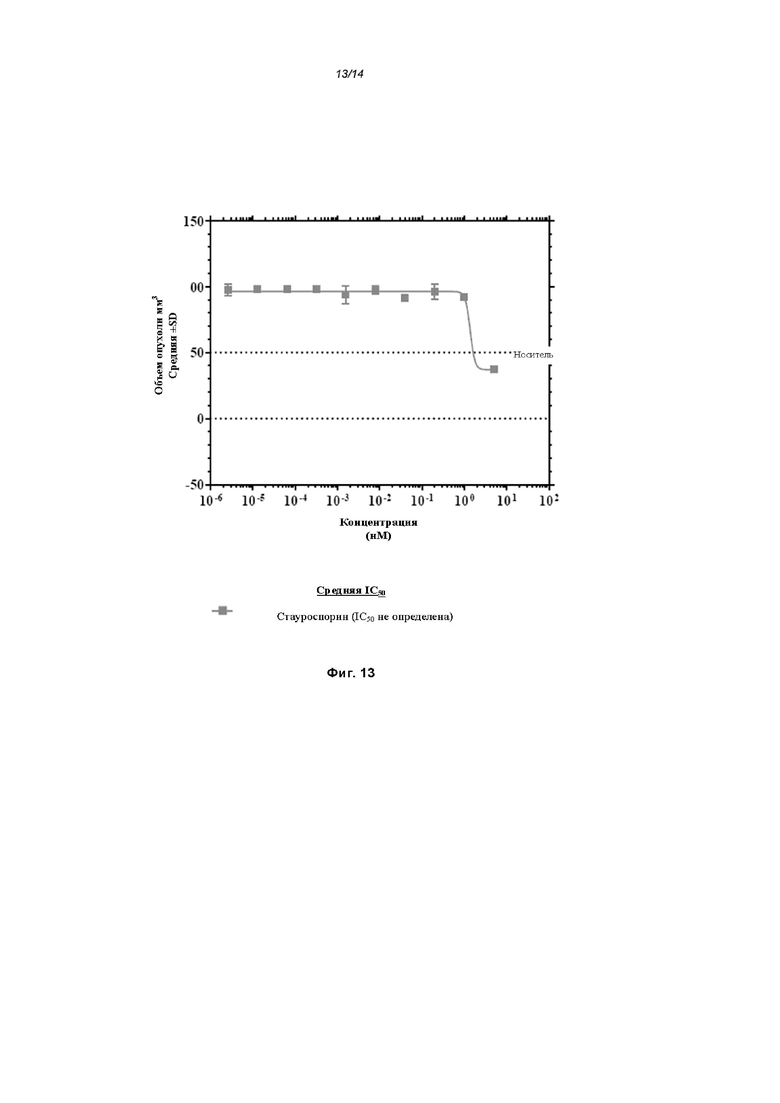
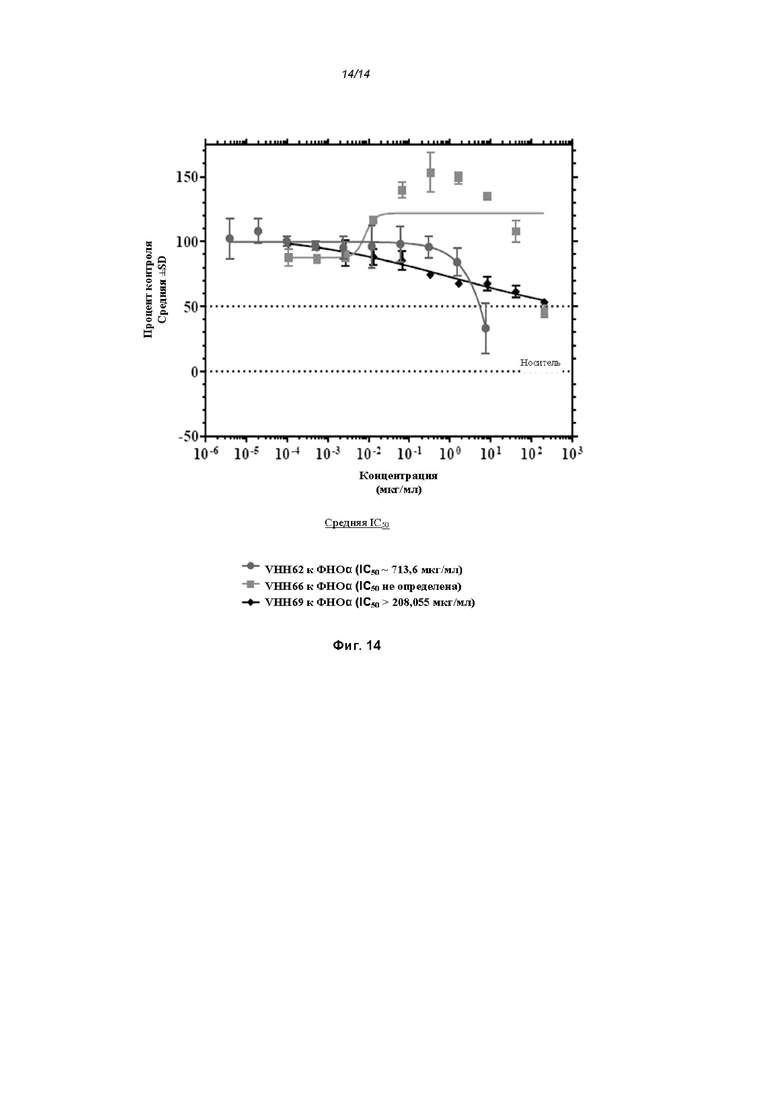
Изобретение относится к области биохимии, в частности к однодоменному антителу к STAT3. Также раскрыты выделенный полипептид, фармацевтическая композиция, содержащая указанное антитело. Раскрыты способ диагностирования расстройства, опосредованного STAT3; способ профилактики или лечения заболевания, опосредованного STAT3, с помощью указанного антитела; применение указанной фармацевтической композиции для лечения расстройства, опосредованного STAT3. Изобретение обладает способностью эффективно лечить заболевания, ассоциированные с STAT3. 6 н. и 3 з.п. ф-лы, 14 ил., 41 табл., 16 пр.
1. Однодоменное антитело к STAT3, содержащее аминокислотную последовательность, приведенную в SEQ ID №: 3 для лечения заболеваний или патологических состояний, опосредствованных внутриклеточными компонентами, путем устранения указанных внутриклеточных компонентов, при том что внутриклеточный компонент представляет собой STAT3.
2. Однодоменное антитело по п. 1, отличающееся тем, что внутриклеточный компонент STAT3 может содержать нуклеиновую кислоту, белок, липид, углевод или их комбинацию.
3. Выделенный полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID №: 3, кодирующий однодоменное антитело к STAT3, используемый для лечения заболеваний или патологических состояний, опосредствованных внутриклеточными компонентами, путем устранения указанных внутриклеточных компонентов, при том что внутриклеточный компонент представляет собой STAT3, а указанный полипептид состоит из четырех каркасных областей, прерываемых тремя определяющими комплементарность областями.
4. Фармацевтическая композиция для использования в качестве лекарственного средства для лечения заболеваний или патологических состояний, вызванных внутриклеточными компонентам, причем внутриклеточный компонент представляет собой STAT3, путем устранения указанных внутриклеточных компонентов, содержащая однодоменное антитело по любому из пп. 1, 2 или полипептид, как он заявлен по п. 3, и подходящий фармацевтически приемлемый носитель, выбранный из воды, солевого раствора, растворов Рингера, раствора декстрозы, фосфатно-солевой буферный раствора, и 5% сывороточный альбумин человека, липосомы, катионные липиды и неводные носители, такие как нелетучие масла.
5. Способ диагностирования расстройства, опосредованного STAT3, у субъекта, с использованием однодоменного антитела против STAT3 по п. 1 или полипептида как он заявлен по п. 3, включающий этапы, на которых:
(а) приводят в контакт биологический образец, взятый у субъекта, с однодоменным антителом по п. 1 или полипептидом по п. 3;
(б) определяют количество внутриклеточного компонента в биологическом образце; и
(в) сравнивают количество, определенное на этапе (б), с эталонным, при этом разница в количестве между образцом и эталоном позволяет диагностировать заболевание.
6. Способ профилактики или лечения заболевания, расстройства, профилактики рецидива заболевания или патологического состояния, опосредованного STAT3, включающий введение эффективного количества однодоменного антитела к STAT3 по п. 1, или полипептида по п. 3, или фармацевтической композиции, как она заявлена по п. 4, субъекту, нуждающемуся в этом, путем введения одним из известных способов, включающих внутривенное введение, внутримышечное введение, пероральное введение, ректальное введение, энтеральное введение, парентеральное введение, внутриглазное введение, подкожное введение, чрескожное введение, введение в виде глазных капель, введение в виде назального спрея, введение посредством ингаляции или небулайзера, местное введение и введение в виде имплантируемого лекарственного средства.
7. Способ по п. 6, отличающееся тем, что заболевание включает рак молочной железы, рак поджелудочной железы, рак предстательной железы,
8. Способ по п. 6, отличающийся тем, что заболевание представляет собой заболевание, вызванное аномальной клеточной пролиферацией.
9. Применение фармацевтической композиции, как она заявлена по п. 4, для лечения или профилактики заболевания или патологического состояния, опосредованного STAT3, или профилактики рецидива заболевания, опосредованного STAT3, для лечении рака молочной железы, рака поджелудочной железы или рака предстательной железы или заболеваний, вызванных аномальной клеточной пролиферацией, включающее введение однодоменного антитела к STAT3 по п. 1 или полипептида, как он заявлен по п. 3, субъекту, нуждающемуся в этом.
| WO 2011097480 A1, 11.08 | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| ИНГИБИТОР АКТИВАЦИИ STAT3/5 | 2007 |
|
RU2489148C2 |
| WO 2004080394 A2,. | |||
