Изобретение относится к области биотехнологии и производству вакцин против инфекционного ринотрахеита кошек, а именно к способу опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцины с помощью дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена.
Инфекционный ринотрахеит кошек - контагиозное вирусное заболевание, характеризующееся поражением верхних дыхательных путей, конъюнктивитами и кератитами [1].
Возбудителем инфекционного ринотрахеита кошек является вирус принадлежащий к семейству Herpesviridae, роду Varicellovirus, виду Alpaherpesvirus 1 (FHV-1 - Felid alphaherpesvirus-1) [2]. Нуклеиновая кислота вируса представлена двуцепочечной ДНК размером около 134 700-135 900 п.н. Важным для выявления генома возбудителя инфекционного ринотрахеита кошек является UL35-reH, который кодирует информацию о малом капсидном белке (109 а.о.) и располагается в диапазоне 44100…44426 п. н. и имеет размер 327 п. н. [3].
Инфекционный ринотрахеит был зарегистрирован у многих представителей семейства кошачьих, таких как европейские дикие кошки, песчаные кошки, леопардовые кошки, гепарды, горные львы, оцелоты и др. [4]. К инфекции восприимчивы как взрослые кошки всех пород, так и котята, но у молодых животных, зараженных FHV-1, нередок летальный исход в результате осложнения пневмонией. В литературе описаны также случаи развития менингоэнцефалита в результате заражения FHV-1 [5]. После острой фазы заболевание часто переходит в латентную форму с пожизненным вирусоносительством. Исследователи из Австралии определили серопревалентность FHV-1 на уровне 11-17% у диких представителей семейства кошачьих и 37% у домашних кошек, однако, по другим оценкам около 90% кошек считаются серопозитивными по отношению к FHV-1, из них 45% выделяют вирус в течение всей жизни [6].
Система мер для борьбы с инфекционным ринотрахеитом кошек и его профилактики предусматривает иммунизацию домашних животных [7]. Для этой цели применяют вакцинные препараты. При их изготовлении неинактивированное вируссодержащее сырье исследуют на определение титра инфекционной активности вируса для оценки его активности в клетках. В 1,0 см3 суспензии вируса определяют количество клеточных культуральных инфекционных доз, вызывающих 50%-ное поражение клеток, что фактически отражает концентрацию полных вирусных частиц, содержащих ДНК вируса инфекционного ринотрахеита кошек в активном состоянии.
Традиционно для определения титра инфекционной активности данного вируса кошек применяют метод титрования в перевиваемой монослойной культуре клеток почки кошки (CRFK), с помощью которой вычисляют минимальную дозу вируса, способную вызвать лизис 50% клеток (прототип) [7]. Данный метод имеет некоторые недостатки: 1) длительная процедура титрования, связанная с формированием цитопатического действия (ЦПД) в течение 72 ч; 2) субъективность при оценке результатов исследования; 3) высокая стоимость клеточной линии как тест-системы и затраты на ее поддержание; 4) наличие вероятности риска контаминации культуры клеток.
В связи с этим целесообразно провести поиск альтернативного способа опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцин.
В настоящее время развиваются методы молекулярной биологии, в частности, модифицированные варианты полимеразной цепной реакции (ПЦР), в частности, с проведением математического дифференцирования. Предложенный метод является высокочувствительным и специфичным, объективным и позволяет определять титр инфекционной активности возбудителя инфекционного ринотрахеита кошек в вируссодержащих суспензиях в течение 2 часов, что в 36 раз быстрее по сравнению с прототипным вариантом [7], не требует использования клеточной линии для количественного анализа, не предполагает контаминации исследуемых образцов.
Задачей настоящего изобретения является разработка высокочувствительного и высокоспецифичного экспресс-способа опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцины с целью устранения вышеуказанных недостатков.
Данная задача решена благодаря разработке способа опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцины с помощью дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена.
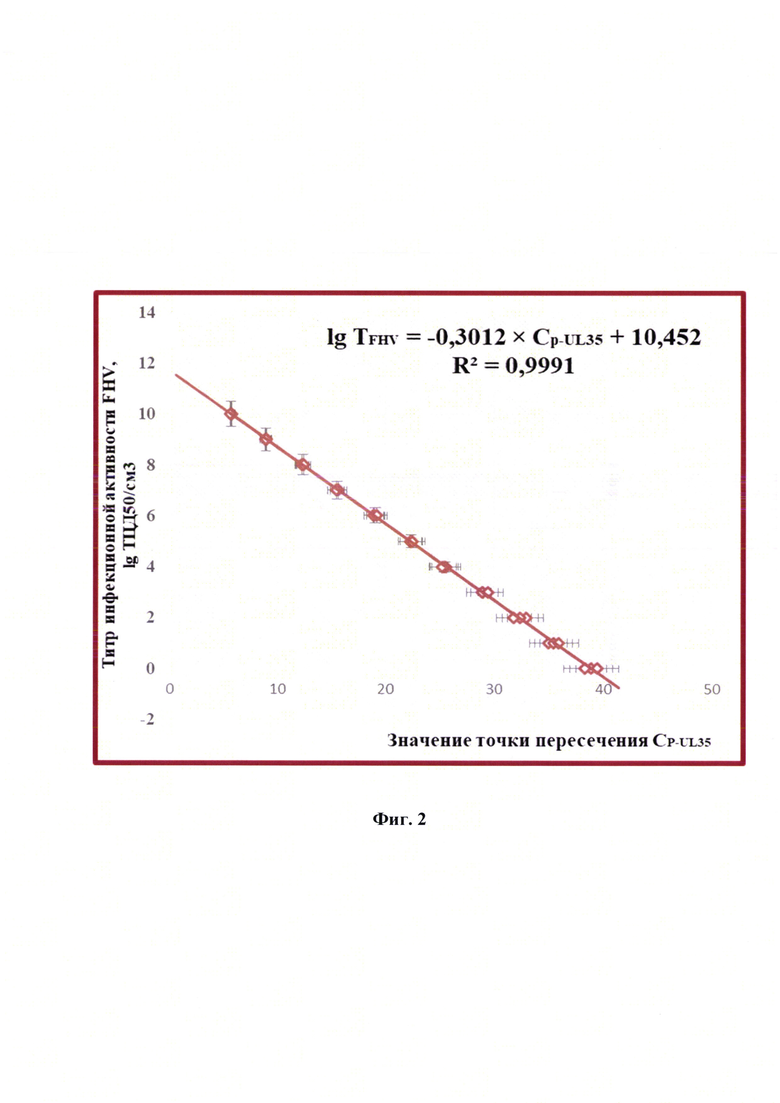
Технический результат изобретения заключается в том, что разработанный способ дает возможность: 1) повысить специфичность анализа проб за счет реакции нейтрализации полных частиц возбудителя инфекционного ринотрахеита кошек штаммоспецифическими антителами; 2) увеличить чувствительность и специфичность анализа за счет применения высокоспецифичных оригинальных олигонуклеотидных праймеров FHV-F44120, FHV-R44253 и FHV-P44151-зонда, рассчитанных для UL35-гена (продукт - малый капсидный белок, small capsid protein) возбудителя инфекционного ринотрахеита кошек в сырье для вакцины; 3) повысить достоверность проводимого анализа благодаря установлению зависимости между титром инфекционной активности возбудителя инфекционного ринотрахеита кошек (TFHV) и дифференциала второй степени точки пересечения логистической кривой реакции амплификации участка UL35-гена (Cp-UL35), представленной в виде логарифмической функции:
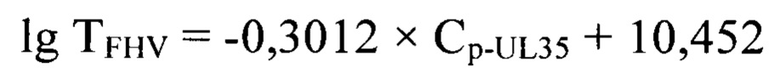
с высокой достоверностью аппроксимации (R2=0,9991) и эффективностью амплификации 99,65%. Предложенная модель позволит опосредованно определять титр инфекционной активности возбудителя инфекционного ринотрахеита кошек в неинактивированном сырье при производстве культуральных вакцин.
Сущность изобретения отражена на графических изображениях:
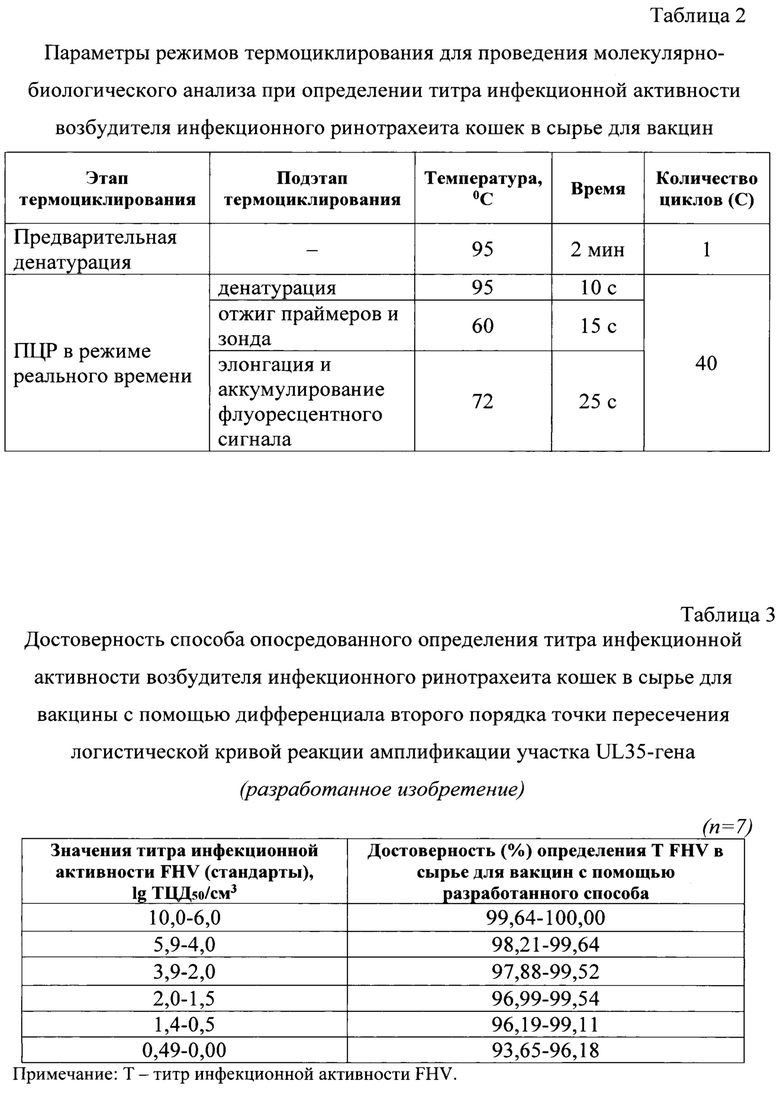
Фиг. 1 - Дизайн оригинальных олигонуклеотидных праймеров и зонда для опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцин. Примечание: А - прямой праймер и зонд, Б - обратный праймер, который дан в двух вариантах, а именно, в комплементарном прямом и в формате rev-compl - обратно-комплементарном.
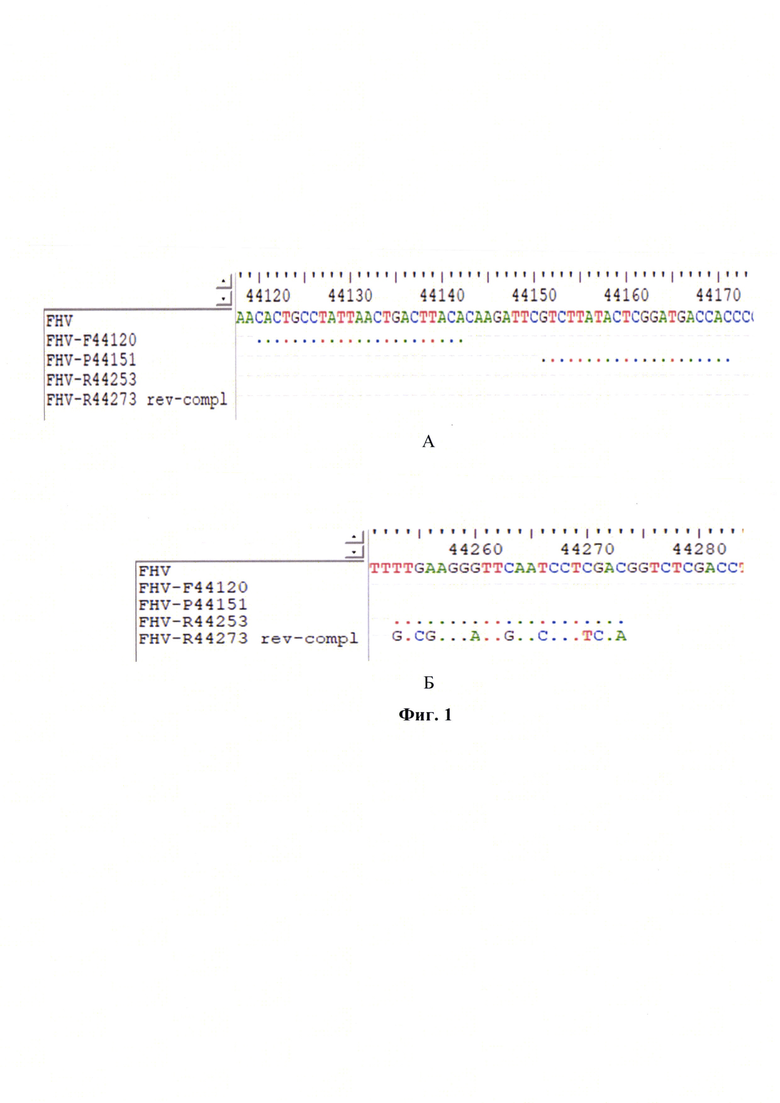
Фиг. 2 - Зависимость титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцин и значений дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена (с указанием стандартной погрешности) (n=3, p<0,005).
SEQ ID NO:1 представляет последовательность праймера FHV-F44120;
SEQ ID NO:2 представляет последовательность праймера FHV-R44253;
SEQ ID NO:3 представляет последовательность зонда FHV-P44151- ROX/BHQ2.
Сущность изобретения заключается в новом подходе по опосредованному определению титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в неинактивированном сырье для вакцин с помощью дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена. Заявляемый способ основан на проведении реакции нейтрализации вирионов возбудителя инфекционного ринотрахеита кошек с использованием очищенных поликлональных антител, получении элюата суммарной ДНК, проведении ПЦР в режиме реального времени и определения дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена для расчета по регрессионной формуле титра инфекционной активности FHV в сырье для культуральных вакцин против инфекционного ринотрахеита кошек.
В настоящее время применяется способ опосредованного определения титра инфекционной активности вируса ящура в неинактивированном сырье для вакцин с помощью математического двойного дифференциала данных точки crossing point при амплификации вирусной кДНК [8]. Однако, для решения поставленной задачи способ опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцины с помощью дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена ранее не применялся и в литературе не представлен.
В отличие от прототипа разработанный способ включает реакцию нейтрализации вирионов возбудителя инфекционного ринотрахеита кошек с применением очищенных поликлональных антител из штаммоспецифичных сывороток крови морских свинок против FHV; этап сорбционного экстрагирования суммарной ДНК; этап проведения ПЦР с применением оригинальных олигонуклеотидных праймеров и зондов; определение дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена для расчета по регрессионной формуле титра инфекционной активности FHV в сырье для культуральных вакцин против инфекционного ринотрахеита кошек.
Применение разработанного способа повышает достоверность анализа по определению титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в пробах сырья для культуральных вакцин. Исходя из этого, актуально применять данный способ для опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для культуральных вакцин.
Ключевым элементом заявляемого способа является расчет дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена по регрессионной формуле титра инфекционной активности FHV в сырье для культуральных вакцин против инфекционного ринотрахеита кошек.
Сопоставительный анализ с прототипом позволяет сделать вывод, что новизна и изобретательский уровень заявляемого изобретения заключается в применении способа опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в неинактивированном сырье для культуральных вакцин с помощью расчета дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена.
Сведений о разработке предлагаемого способа опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцин авторами не обнаружено.
На подготовительном этапе работы сенсибилизируют иммунологический планшет, очищенными штаммоспецифическими поликлональными антителами против возбудителя инфекционного ринотрахеита кошек в объеме 1,50 см3 суспензии с концентрацией иммуноглобулинов G 3,5 мкг/см3. После иммобилизации антител при температуре 4±2°С в течение 18-20 часов лунки планшета подвергают трехкратному промыванию стандартным буферным раствором TBST, открытые сайты связывания блокируют 1,0%-ной суспензией бычьего сывороточного альбумина при температуре 37±0,5°С в течение 30 минут и вновь лунки промывают стандартным буферным раствором TBST 5 раз. Процедуру подготовки планшета к работе с пробами осуществляют заранее, до проведения основного анализа. Планшеты можно подготовить впрок и хранить в холодильнике при температуре 4±2°С.
На первом этапе исследования проводят реакцию нейтрализации вирионов возбудителя инфекционного ринотрахеита кошек из исследуемых образцов (разведения стандарта, отрицательный контроль, пробы). В качестве стандарта используют неинактивированную суспензию возбудителя инфекционного ринотрахеита кошек с известной концентрацией вирусных частиц. Отрицательным контролем служит не инфицированная вирусами, бактериями, микоплазмами и грибами суспензия клеток линии CRFK с концентрацией 3,0-3,5 млн клеток/см3.
Применяют контрольную панель готовых разведений стандарта с содержанием вирусной ДНК, эквивалентными следующим титрами инфекционной активности возбудителя инфекционного ринотрахеита кошек: 0,0; 1,0; 2,0; 3,0; 4,0; 5,0; 6,0; 7,0; 8,0; 9,0; 10,0 lg ТЦД50/см3. В рамках производственного технологического процесса наибольший интерес представляют суспензии с титрами инфекционной активности возбудителя инфекционного ринотрахеита кошек выше 6,0 lg ТЦД50/см3. В лунки с сенсибилизированными штаммоспецифическими антителами против возбудителя инфекционного ринотрахеита кошек вносят по 1,5 см3 суспензий образцов и инкубируют при температуре 37±0,5°С в течение 30 минут. В результате серологической реакции на поверхности лунок формируется иммунные комплексы. Лунки отмывают от балластных компонентов с использованием стандартного буферного раствора TBST 3 раза. Образовавшиеся иммунные комплексы ресуспендируют в 0,5 см3 1/15 М фосфатного буферного раствора.
На следующем этапе анализа осуществляют выделение ДНК из иммунного комплекса твердофазным сорбционным методом с применением набора «ДНК-сорб» («Интерлабсервис», РФ) в соответствии с инструкцией производителя.
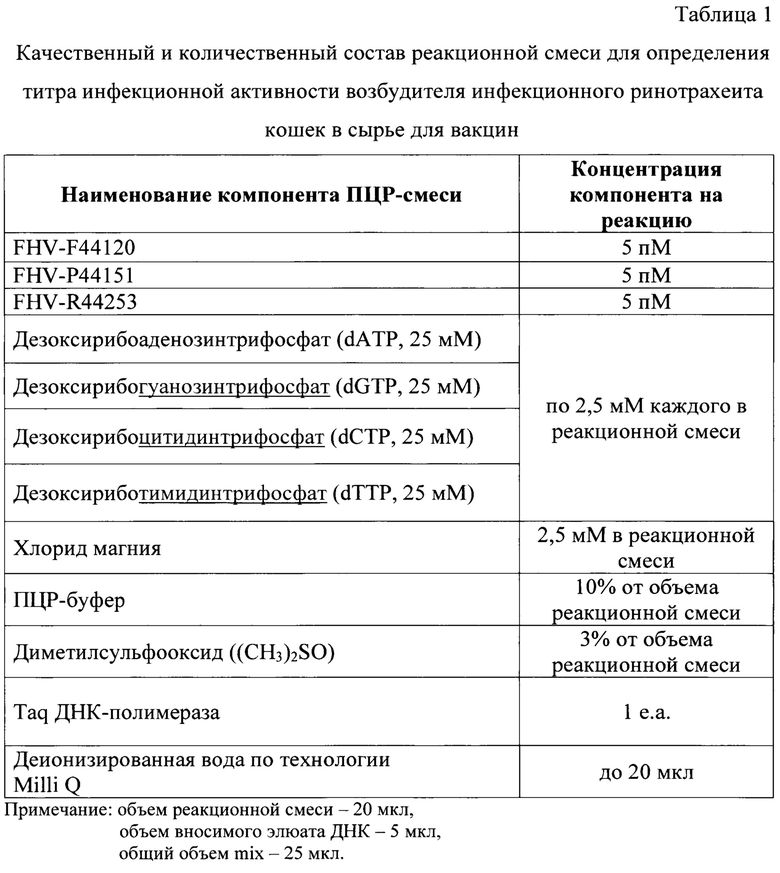
После получения экстракта ДНК возбудителя инфекционного ринотрахеита кошек проводят ПЦР в режиме реального времени с использованием специфических оригинальных олигонуклеотидных праймеров и зонда для исследования контрольных образцов и проб. Для постановки реакции готовят реакционную смесь, рецептура приготовления которой представлена в таблице 1. В качестве гомологичных UL35-гену возбудителя инфекционного ринотрахеита кошек олигонуклеотидов используют:
FHV-F44120-праймер (5'-CACTGCCTATTAACTGACTTACA-3'),
FHV-R44253-праймер (5'-CCACGGAGATCAACTTCTCCT-3'),
FHV-P44151-ROX/BHQ2-зонд (5'-ROX-GTCTTATACTCGGATGACCAC-BHQ2-3') (фиг. 1) в концентрации 5 пМ на реакцию. Для формирования нуклеотидных цепей продуктов реакции применяют дезоксирибонуклеозидтрифосфаты с концентрацией каждого в реакционной смеси по 2,5 мМ. В реакционную смесь добавляют 2,5 мМ хлорида магния и диметилсульфооксид в количестве 3% от общего объема без элюата ДНК. В качестве катализатора реакции амплификации применяют Taq ДНК-полимеразу (1 е.а.).
Объем реакционной смеси компонентов для проведения одной реакции составляет 20 мкл. Элюаты ДНК каждого образца добавляют к смеси по 5 мкл. Общий объем смеси составляет 25 мкл.
Постановку реакции осуществляют при температурных и временных параметрах, сведения о которых представлены в таблице 2. Предварительную денатурацию комплементарной ДНК проводят при температуре 95°С за 2 мин в течение 1 цикла. Реакцию амплификации в режиме реального времени осуществляют в течение 40 циклов, каждый из которых складывается из 3 подэтапов: «денатурации», проводимой при температуре 95°С в течение 10 с, а также подэтапов «отжига праймеров и зонда» и «элонгации и аккумулирования флуоресцентного сигнала», осуществляемых при температурах 60°С за 15 с и 72°С за 25 с, соответственно.
Результаты реакции анализируют, оценивая и сравнивая графики накопления флуоресцентного сигнала с помощью определения дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена (Ср-UL35), определенных с помощью построения графика второй производной функции Fl=f×(Ср-UL35). Величина Ср является важной характеристикой реакции, она прямо пропорциональна количеству копий исходной вирусной ДНК и соответственно титру инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцин. Учитывая, что функция от двойного дифференциала f(Ср) (f''(Ср)) непрерывна в некоторой окрестности точки Ср=Cp1 и задана на отрезке циклов амплификации [0; 40], существует определенный участок около точки Ср, для которого во всех координатах на оси O-Ср двойной математический дифференциал функции f(Ср) будет отрицательным. Поскольку f''(Ср) является первым дифференциалом от функции f'(Ср), то из условия (f'(Ср))'<0, следует, что f'(Ср) на некотором малом отрезке, содержащем точку Ср=Ср1, будет убывающей. Учитывая, что f'(Ср)=0, на участке при Ср<Cp1 первый дифференциал функции f(Ср)>0, а при Ср>Cp1 получаем, что f'(Ср)<0. Иными словами, первый дифференциал функции f(Ср) при переходе через точку Ср=Cp1 изменяет знак с «+» на «-», следовательно, в точке Cp1 функция, отражающая процесс накопления флуоресцентного сигнала, имеет наибольшее значение [9-13]. Таким образом, если график реакции амплификации представлен функцией Fl=f×(Ср), f'(Ср)=0 и f''(Ср)<0, то при условии, что Ср=Cp1 полученная функция имеет наибольшее значение в точке с аргументом СР1, значение которого учитывают для установления зависимости между титром инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцин и значением Ср функции после проведения расчета дифференциала второй степени.
Данный метод имеет преимущество в связи с тем, что при умножении кривой на любые множители положение наибольших значений функции не изменяется. Максимум на графике двойного дифференциала при его исследовании на максимумы располагается внутри экспоненциального участка сигмоиды, где эффективность ПЦР является константой. Наибольшие значения первого дифференциала находятся, чаще всего, в зоне искажения значений эффективности амплификации, поэтому их не рекомендуется использовать для анализа. Графики третьей производной дают менее точные результаты, поскольку их координаты во многом сопряжены с шумовыми значениями [11].
Вычислив значения Ср-UL35 для кривых, отражающих накопление флуоресцентного сигнала образцов с разным титром инфекционной активности возбудителя инфекционного ринотрахеита кошек, устанавливают зависимость между титром FHV в неинактивированном сырье для культуральных вакцин и значением точки Ср-UL35 после проведения математического дифференцирования второй степени. На основе разработанной модели рассчитывают значение титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в неинактивированном сырье для культуральных вакцин.
Сущность предлагаемого изобретения пояснена примерами его использования, которые не ограничивают объем изобретения.
Пример 1. Выявление существования зависимости между титром инфекционной активности возбудителя инфекционного ринотрахеита кошек в неинактивированном сырье для вакцины и значением точки Ср-UL35.
На первом этапе исследования применяли панель готовых разведений стандарта, в качестве которого использовали неинактивированную суспензию культурального возбудителя инфекционного ринотрахеита кошек с количествами вирусной ДНК, эквивалентными следующим титрам инфекционной активности FHV: 0,0, 1,0, 2,0, 3,0, 4,0, 5,0, 6,0, 7,0, 8,0, 9,0, 10,0 lg ТЦД50/см3. В лунки с сенсибилизированными штаммоспецифическими антителами против возбудителя инфекционного ринотрахеита кошек вносили по 1,5 см3 суспензий образцов и инкубировали при температуре 37±0,5°С в течение 30 минут. В результате реакции нейтрализации на поверхности лунок формировались иммунные комплексы. Лунки отмывали от балластных компонентов с использованием стандартного буферного раствора TBST 3 раза. Образовавшиеся иммунные комплексы ресуспендировали в 0,5 см3 1/15 М фосфатного буферного раствора.
Проводили выделение ДНК из иммунного комплекса твердофазным сорбционным методом в соответствии с процедурой, представленной выше.
Осуществляли постановку ПЦР в режиме реального времени с использованием специфических оригинальных олигонуклеотидных праймеров и зонда для исследования контрольных образцов и проб. Для проведения реакции составляли реакционную смесь, рецептура которой представлена выше.
Постановку ПЦР в режиме реального времени осуществляли, как описано выше. Результаты реакции анализировали, оценивая и сравнивая графики накопления флуоресцентного сигнала после проведения двойного дифференцирования по значениям точек Ср-UL35. Полученные данные анализировали с помощью программного обеспечения «Rotor-Gene FRT-Manager», которое позволяет строить графики накопления флуоресцентного сигнала в режиме реального времени на протяжении заданного количества циклов амплификации (С). Применяя технологии компьютерной программы «Maxima» (или аналога), проводили построение графиков после вычисления математического дифференциала второй степени для полученных элюатов ДНК возбудителя инфекционного ринотрахеита кошек каждого разведения стандарта с определенными значениями титра FHV и рассчитывали средние значения Ср-UL35 с проекцией на ось абсцисс О-С (n=3). При исследовании отрицательного контроля накопления флуоресцентного сигнала не наблюдалось, что подтверждает отсутствие возбудителя инфекционного ринотрахеита кошек в данном образце. В представленных исследованиях p-уровень значимости меньше 0,005, что подтверждает достоверность проводимых количественных исследований. Зависимость титра инфекционной активности возбудителя инфекционного ринотрахеита кошек и значений точек Ср-UL35 представлена на фиг. 2 и отражена в виде регрессионной логарифмической функции:
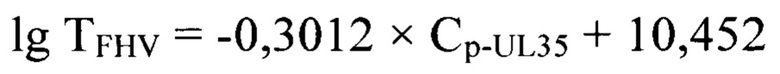
с высокой достоверностью аппроксимации (R2=0,9991) и эффективностью амплификации 99,65%. Предложенная модель позволит опосредованно определять титр инфекционной активности возбудителя инфекционного ринотрахеита кошек в неинактивированном сырье при производстве культуральных вакцин.
Пример 2. Апробация способа опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцины с помощью дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена.
В исследовании использовали 6 суспензий культурального вируса инфекционного ринотрахеита кошек с титрами инфекционной активности 6,15; 6,78; 7,87; 8,15; 8,95; 9,25 lg ТЦД50/см3, соответственно (пробы №1-6). В качестве положительного контроля применяли суспензию культурального вируса инфекционного ринотрахеита кошек с титром 7,00 lg ТЦД50/см3. В качестве отрицательных контролей применяли суспензию клеток CRFK, не контаминированную микроорганизмами. Испытуемые пробы и контрольные образцы исследовали в пяти повторностях. Этапы выделения ДНК, и постановку ПЦР в режиме реального времени проводили, как описано в примере 1.
Средние значения Ср-UL35 для проб №1-6 составляли 14,25±0,01, 12,12±0,02, 8,64±0,02, 7,64±0,01, 4,89±0,01, 4,06±0,01, соответственно. Пользуясь разработанной регрессионной функцией, представленной выше, рассчитали средние значения титра инфекционной активности вируса инфекционного ринотрахеита кошек для проб №1-6, которые составили 6,16; 6,80; 7,85; 8,15; 8,98; 9,23 lg ТЦД50/см3, соответственно. Полученные значения соответствовали референтным данным. Для положительного контроля значение порогового цикла амплификации составило 8,34±0,01, что соответствовало титру инфекционной активности вируса инфекционного ринотрахеита кошек, равному 7,00 lg ТЦД50/см3. Для отрицательных контролей экспоненциальные графики не были сформированы, что означало отсутствие возбудителя инфекционного ринотрахеита кошек в данных образцах. Таким образом, разработанный способ позволяет определять титр инфекционной активности FHV в неинактивированном сырье для культуральных вакцин.
Пример 3. Выявление степени достоверности опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцины с помощью дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена.
Для анализа использовали 540 суспензий культурального вируса инфекционного ринотрахеита кошек с титром инфекционной активности от 0,00 до 10,00 lg ТЦД50/см3. В качестве положительного контроля применяли суспензию культурального FHV с титром инфекционной активности вируса 7,00 lg ТЦД50/см3. В качестве отрицательных контролей применяли суспензию клеток CRFK, не зараженную микроорганизмами. Испытуемые пробы и контрольные образцы исследовали в трех повторностях. Этапы экстрагирования нуклеиновых кислот и постановку ПЦР в режиме реального времени проводили, как отражено в примере 1. Результаты анализа представлены в таблице 3.
Интерпретацию результатов проводили, пользуясь разработанной регрессионной функцией, с получением значений TFHV для каждой из 540 проб. Для положительного контроля значение порогового цикла амплификации составило 8,34±0,01, что соответствовало титру инфекционной активности вируса, равному 7,00 lg ТЦД50/см3. Для отрицательных контролей экспоненциальные графики не были сформированы, что означало отсутствие возбудителя инфекционного ринотрахеита кошек в данных образцах.
Выявили, что данные, полученные с помощью разработанного способа, коррелировали с методом титрования в культуре клеток CRFK на 99,64-100,00% для 10,0-6,0 lg ТЦД50/см3 (n=90), на 98,21-99,64% для 5,9-4,0 lg ТЦД50/см3 (n=90), на 97,88-99,52% для 3,9-2,0 lg ТЦД50/см3 (n=90), на 96,99-99,54% для 2,0-1,5 lg ТЦД50/см3 (n=90), на 96,19-99,11% для 1,4-0,5 lg ТЦД50/см3 (n=90), на 93,65-96,18% для 0,49-0,00 lg ТЦД50/см3 (n=90). Полученные результаты свидетельствовали о высокой степени точности разработанного способа. Аналитическая чувствительность разработанного способа составила не менее 0,5 lg ТЦД50/см3 с достоверностью не менее 96,19% (n=90, p<0,005). Таким образом, разработанный способ опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцины с помощью дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена является достоверным.
Пример 4. Исследование специфичности способа опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцины с помощью дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена.
При оценке специфичности способа (предлагаемое изобретение) исследовали суспензии вируса ящура, вируса бешенства, коронавируса кошек, калицивируса кошек и чумы плотоядных животных. Количество инфекционных доз вирусов в суспензиях составлял не менее 6,0 lg ТЦД50/см3 и 6,0 lg ККИД50/см3. Исследования проводили в 5 повторностях.
Этапы элюирования нуклеиновых кислот и постановку ПЦР в режиме реального времени проводили, как описано в примере 1. Для проб, содержащих другие вирусы, не наблюдалось формирования графиков экспоненты, и они не выходили за пороговый уровень флуоресцентного сигнала (0,005 у.е.). Таким образом, разработанный способ является специфичным по отношению к возбудителю инфекционного ринотрахеита кошек и может быть использован для определения титра инфекционной активности данного вируса в неинактивированном сырье для культуральных вакцин.
Основным преимуществом предлагаемого изобретения является возможность опосредованно определить титр инфекционной активности возбудителя инфекционного ринотрахеита кошек в неинактивированном сырье для культуральных вакцин. Предлагаемое изобретение сочетает постановку реакции нейтрализации с последующим проведением молекулярно-биологического исследования неинактивированной вирусной суспензии возбудителя инфекционного ринотрахеита кошек, что повышает специфичность и чувствительность способа. В предлагаемом изобретении зависимость титра инфекционной активности возбудителя инфекционного ринотрахеита кошек и значений точек Ср-UL35 представлена в виде регрессионной логарифмической функции:
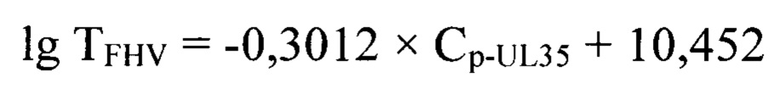
с высокой достоверностью аппроксимации (R2=0,9991) и эффективностью амплификации 99,65%. Предложенная модель позволит опосредованно определять титр инфекционной активности возбудителя инфекционного ринотрахеита кошек в неинактивированном сырье при производстве культуральных вакцин.
Источники информации, принятые во внимание при составлении описания изобретения к заявке на выдачу патента РФ на изобретение «Способ опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцины с помощью дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена»:
1. Slaviero M, Ehlers LP, de Almeida BA, Pereira PR, Panziera W, da Costa FVA, Pavarini SP, Sonne L. Generalized and fatal felid alphaherpesvirus-1 natural infection with liver involvement in a feline leukaemia virus-positive adult cat: a case report. Vet Res Commun. 2022 Dec;46(4): 1319-1324.
2. Taxonomy. Felid alphaherpesvirus-1. URL: https://www.ncbi.nlm.nih.gov/taxonomy/?term=Felid+alphaherpesvirus-1 (Дата обращения: 14.04.2023).
3. GenBank. [Электронный ресурс] / URL: http://www.ncbi.nlm.nih.gov. (Дата обращения: 02.04.2023).
4. Pennington MR, Voorhees IEH, Callaway HM, Dehghanpir SD, Baines JD, Parrish CR, Van de Walle GR. The HIV Integrase Inhibitor Raltegravir Inhibits Felid Alphaherpesvirus 1 Replication by Targeting both DNA Replication and Late Gene Expression. J Virol. 2018 Sep 26; 92(20):e00994-18.
5. Shi L, Huang S, Lu Y, Su Y, Guo L, Guo L, Xie W, Li X, Wang Y, Yang S, Chai H, Wang Y. Cross-species transmission of feline herpesvirus 1 (FHV-1) to chinchillas. Vet Med Sci. 2022 Nov; 8(6):2532-2537.
6. Summers SC, McLeland SM, Hawley JR, Quimby JM, Lappin MR. Effect of repeated administration of a parenteral feline herpesvirus-1, calicivirus, and panleukopenia virus vaccine on select clinicopathologic, immunological, renal histologic, and immunohistochemical parameters in healthy adult cats. Am J Vet Res. 2022 May 2l; 83(7):ajvr.21.07.0087.
7. Bergmann M, Speck S, Rieger A, Truyen U, Hartmann K. Antibody response to feline herpesvirus-1 vaccination in healthy adult cats. J Feline Med Surg. 2020 Apr; 22(4):329-338.
8. Патент RU №2793900 от 07.04.2023 г.
9. Sooknanan R., van Gemen В., Malek L. Nucleis acid sequence-based amplification // Molecular methods for virus deteCqion-London: Academic press, 1995. - P. 261-285.
10. Эверитт, Брайан С.; Ландау, Сабина; Лиз, Морвен; и Шталь, Дэниел (2011) "Разные методы кластеризации", в кластерном анализе, 5-е издание, John Wiley & Sons, Ltd., Чичестер, Великобритания.
11. Rutledge R.G. Mathematics of quantitative kinetic PCR and the application of standard curves / R.G. Rutledge, C. Cote // Nucleic Acide Res. - 2004. - V. 31, N. 16. - e. 93. P. 1-6.
12. Liu W., Saint D.A. Validation of a quantitative method for real-time PCR kinetics // Biochem. Biophys. Res. Commun. - 2002. - V. 294. - P. 347-353.
13. Peirson S.N., Butler J.M., Foster R.G. Experimantal validation of novel and conventional approaches to quantitative real-time PCR data analysis // Nucleic Acids Res. - 2003. - V. 31:e.73.
Изобретение относится к области биотехнологии. Описан способ опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцины с помощью дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена, предполагающий применение разработанных олигонуклеотидов. Технический результат изобретения заключается в том, что разработанный способ дает возможность повысить специфичность анализа, увеличить чувствительность и специфичность анализа, а также повысить достоверность проводимого анализа. 1 з.п. ф-лы, 2 ил., 3 табл., 4 пр.
1. Способ опосредованного определения титра инфекционной активности возбудителя инфекционного ринотрахеита кошек в сырье для вакцины с помощью дифференциала второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена, предполагающий применение олигонуклеотидов:
FHV-F44120-праймер с дизайном 5'-CACTGCCTATTAACTGACTTACA-3',
FHV-R44253-праймер с дизайном 5'-CCACGGAGATCAACTTCTCCT-3',
FHV-P44151-ROX/BHQ2-зонд с дизайном 5’-ROX-GTCTTATACTCGGATGACCAC-BHQ2-3',
которые рассчитаны на UL35-ген и позволяют амплифицировать фрагмент в диапазоне 44120…44273 п.н. размером 154 п.н., а также предполагающий определение второго порядка точки пересечения логистической кривой реакции амплификации участка UL35-гена для расчета титра инфекционной активности возбудителя инфекционного ринотрахеита кошек с помощью следующей регрессионной функции:
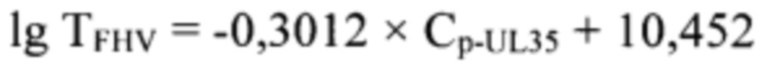
с достоверностью аппроксимации 0,9991 и эффективностью амплификации 99,65%.
2. Способ по п. 1, отличающийся тем, что данные, полученные с помощью разработанного способа, коррелировали с методом титрования в культуре клеток CRFK на 99,64-100,00% для 10,0-6,0 lg ТЦД50/см3, на 98,21-99,64% для 5,9-4,0 lg ТЦД50/см3, на 97,88-99,52% для 3,9-2,0 lg ТЦД50/см3, на 96,99-99,54% для 2,0-1,5 lg ТЦД50/см3, на 96,19-99,11% для 1,4-0,5 lg ТЦД50/см3, на 93,65-96,18% для 0,49-0,00 lg ТЦД50/см3 при n=3 и p<0,005 для каждого представленного диапазона титра инфекционной активности.
| Способ опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии | 2020 |
|
RU2756472C1 |
| КИФОСКОЛИОМЕТР | 0 |
|
SU219075A1 |
| Kokorina, Ye, Elizbarashvili, E | |||
| COMPARATIVE STUDIES OF FELINE VIRAL RHINOTRACHEITIS VIRUS FOR ITS REPLICATION PROPERTIES IN DIFFERENT CELL CULTURES | |||
| Veterinary Science Today | |||
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |

