ОБЛАСТЬ ТЕХНИКИ
В настоящем изобретении предложены модифицированные молекулы эритропоэтина с нейропластической и нейропротективной способностью, подходящие для лечения заболеваний центральной нервной системы, таких как острое нарушение мозгового кровообращения, нейротравма, нейровоспаление и нейродегенерация.
УРОВЕНЬ ТЕХНИКИ
Эритропоэтин (ЕРО) является представителем суперсемейства цитокинов I типа, для которого характерна важная плейотропная активность [1]. Данный цитокин является одним из главных регуляторов эритропоэза, действующих синергически с другими молекулами, чтобы вызвать пролиферацию, дифференцировку и выживаемость предшественников эритроидной линии клеток и поддерживать массу циркулирующих эритроцитов [2].
ЕРО человека (hEPO) представляет собой высоко гликозилированный белок, молекулярная масса которого находится в диапазоне от 30 до 39 кДа. Он содержит три консенсусных сайта N-гликозилирования и один сайт О-гликозилирования [3], которые могут встречаться с полной степенью заполнения. Цепи сахаров состоят из варьирующих последовательностей моносахаридов и варьирующего количества сиаловой кислоты (СК) [4, 5]. N-связанные углеводы могут содержать две, три или четыре ветви, каждая из которых оканчивается отрицательно заряженной молекулой СК. В свою очередь, углевод, связанный в сайте О-гликозилирования, может содержать до двух молекул СК [6].
Гликозилирование, в частности СК на конце N-гликанов, необходимо для биологической активности ЕРО in vivo, но не для связывания с рецептором in vitro [7, 8]. Поэтому молекулы ЕРО с меньшим содержанием гликозидов обладают высокой аффинностью к рецептору ЕРО (EPOR); тем не менее, их активность in vivo уменьшается благодаря повышению их клиренса из плазмы [8]. Степень гликозилирования данных цитокинов влияет на эффективность их продукции, аффинность к рецептору, время полужизни в плазме, секрецию и стабильность белка [5, 9].
С момента клонирования гена hEPO в 1985 г. знания о биологии цитокинов значительно изменились. Одним из первых прорывов было открытие нового биологического действия молекулы, которое расширило ее эритропоэтическую способность и включало несколько важных физиологических процессов. Некоторые из наиболее важных представляют собой ангиогенез, регуляцию сопротивления сосудов и, что более важно, защиту клеток [10, 11]. Хотя наиболее важной активностью эритропоэтина является гематопоэз, присутствие ЕРО и его рецептора в различных тканях и неэритропоэтических клетках подтвердило упомянутую выше гипотезу, что ЕРО осуществляет множество функций, и одной из наиболее важных функций является цитопротекторная функция в отношении клеток центральной нервной системы, сердца, почек, системы желудочно-кишечного тракта, репродуктивного тракта и эндотелия [12], стимуляция пролиферации клеток, ангиогенеза и ингибирование апоптоза клеток [13]. Данные открытия расширили ожидаемые клинические применения для лечения других патологий, таких как сердечные приступы, острое нарушение мозгового кровообращения и множество других патологий, связанных с нейрозащитой [14].
Нейрозащиту можно описать как подход для поддержания и восстановления клеточных взаимодействий в головном мозге, приводящий к максимальной защите функций нервных клеток [15]. Целью нейрозащиты является предотвращение патологической утраты нервных клеток при заболеваниях центральной нервной системы, таких как острое нарушение мозгового кровообращения, нейротравма, нейровоспаление и нейродегенерация.
Нейродегенеративные заболевания представляют собой группу патологий, которые влияют на нервную систему и вызывают когнитивные расстройства, расстройства поведения и изменения в системах регуляции организма. Для них характерен хронический характер и прогрессирующее развитие. Данные патологии включают такие заболевания, как болезнь Паркинсона, различные типы деменций, болезнь Альцгеймера, рассеянный склероз и болезнь Гентингтона, среди прочих, которые представляют проблему с медицинской, здравоохранительной, социальной и экономической точки зрения, с которой должны столкнуться все страны [16]. Тем не менее, так как мировая популяция стареет, вклад неврологических расстройств будет больше как в развитых, так и в развивающихся странах. Бремя неврологических расстройств достигает значительной доли в странах, где процент людей старше 65 лет повышается. Если рассматривать только болезнь Альцгеймера, статистика свидетельствует о том, что приблизительно 44 миллиона людей живут с деменцией по всему миру, и это значение согласно оценке возрастет до 115 миллионов к 2050 году. Поэтому данную патологию считают всемирной эпидемией [17-19]. По этой причине, множество групп исследователей по всему миру участвует в поиске способов терапии, которые предоставят лечение для контролирования данного типа нейродегенеративных патологий.
Совместная работа множества групп исследователей позволила, благодаря достижениям исследований, выявить новые и перспективные сведения о происхождении данной проблемы и дать надежду обнаруживать ее на ранних стадиях, а также определить более специфичные способы борьбы с ней. Тем не менее, даже такие усилия не идентифицировали никакого биотерапевтического средства, которое может лечить такие патологии. В этом смысле, фармацевтический рынок предлагает лишь лекарственные средства, предназначенные для отсрочивания прогрессирования данных заболеваний после того, как они были обнаружены, но их польза зачастую очень незначительна, поэтому требуется разработка новых средств лечения. Успех преодоления проблемы со здоровьем, которую нейродегенеративные заболевания представляют на сегодняшний день, будет зависеть, среди других аспектов, от успеха терапии, которая принесет пользу и обеспечит долгосрочное воздействие, благоприятно влияя на этиологию или лежащий в их основе патогенез и предотвращая или отсрочивая начало заболевания или постепенное клиническое ухудшение безопасным, успешным и эффективным образом. Но, в свою очередь, будет необходимо предложить эффективную технологию, доступную для больших групп населения. Первоочередная задача фармацевтической промышленности за последние годы состояла в обнаружении соединений, способных воздействовать на общие ключевые точки в развитии множества заболеваний центральной нервной системы, такие как апоптоз, окислительный стресс, воспаление, нарушение функции обмена веществ или нарушенная нейропластичность.
В данной области исследований ЕРО играет очень важную роль благодаря его способности вырабатывать широкий диапазон клеточных ответов в головном мозге, которые непосредственно связаны с защитой и починкой повреждения клеток [20]. ЕРО может вызывать нейрозащиту посредством противовоспалительных, антиоксидантных, антинейротоксических, ангиогенных, нейротрофических, регенерационных и антиапоптических механизмов.
В свете указанного, другое чрезвычайно важное достижение в исследовании ЕРО состояло в идентификации двух различных молекулярных сайтов, связанных с его эритропоэтической и цитопротекторной биологическими активностями. Данные две активности возникают благодаря связыванию цитокина с двумя различными рецепторными системами: гомодимерным рецептором (EPOR)2, отвечающим за эритропоэтическую активность, и гетеродимерным рецептором EPOR-βCR, связанным с цитопротекторной активностью [21]. Данные открытия были чрезвычайно важны, так как они дали необходимую информацию для понимания путей, посредством которых ЕРО проявляет свои биологические активности, и, следовательно, позволили избирательную модуляцию его эритропоэтического или цито протектор но го ответа, с целью избежать или по меньшей мере уменьшить побочные действия, связанные с применением неселективных агентов, таких как ЕРО или аналоги ЕРО.
В этом смысле, хотя ЕРО считают безопасным и хорошо переносимым лекарственным средством для лечения анемии, когда ЕРО предназначен для применения в качестве цитопротекторного агента у пациентов с острым нарушением мозгового кровообращения или сердечными приступами, то есть, в качестве нейропротективного агента у пациентов, не страдающих анемией, его гематологическое действие следует считать побочным действием, поскольку он может вызывать полицитемию, гипертонию и протромботический феномен [22-25]. Поэтому разработка аналогов ЕРО, которая селективно модулирует их эритропоэтическую роль и их цитопротекторную роль, весьма важна [14]. Следуя данной цели были разработаны различные стратегии, чтобы устранить эритропоэтическую активность цитокина, сохранив нейропротективный потенциал, путем введения химических модификаций в молекулу или изменения содержания гликозидов.
В отношении стратегий, в которых используются химические модификации молекулы ЕРО, оценивали карбамоилирование семи остатков лизина, в результате которого образуются остатки гомоцитруллина [26]. Тем не менее, при данной методике возникают конформационные изменения в белке, которые влияют на его функцию. На основании данных результатов различные рабочие группы полностью модифицировали рекомбинантный hEPO (rhEPO) путем карбамоилирования, получив новую молекулу, названную СЕРО, у которой сохранилась цитопротекторная активность, но отсутствует эритропоэтическая активность [26-28]. Однако, для достижения желаемой цели и поддержания его терапевтической эффективности требуются высокие и многократные дозы.
С другой стороны, модификации в rhEPO были введены путем изменения содержания в нем сиаловой кислоты (СК). В 2003 г. Erbayraktar и др. [29] полностью удалили остатки СК, присутствующие на концах гликозидных цепей ЕРО, таким образом получив так называемый асиало-ЕРО. У данного нового варианта ЕРО выявили высокую активность in vitro. Тем не менее, его нейропротективная активность in vivo была сопоставима с таковой, полученной для rhEPO.
С другой стороны, и с целью увеличения времени полужизни в кровотоке стимулирующего эритропоэз цитокина, Egrie и Brown [6] разработали белок, который стимулирует эритропоэз, NESP, который получен из hEPO и содержит два дополнительных сайта N-гликозилирования. Данная модификация привела к трехкратному увеличению времени полужизни в плазме и его гематопоэтической эффективности in vivo.
Что касается изменения глюцидного содержания, авторы настоящего изобретения получили вариант rhEPO со свойствами, сходными с таковыми у церебрального ЕРО (rhNEPO), который представляет собой комбинацию менее кислых изоформ rhEPO. У данного варианта выявили нейропротективную активность, эквивалентную таковой у rhEPO, и выявили менее 4% гематопоэтической активности [30]. Тем не менее, быстрый клиренс rhNEPO из плазмы является недостатком, если предлагать данную молекулу в качестве кандидата для лечения хронических неврологических заболеваний, при которых требуется, чтобы концентрация в плазме поддерживалась продолжительное время и была достаточной для проявления биологического действия. Более того, в связи с тем, что комбинация гликоформ сохраняет его эритропоэтическую активность in vitro, введение высоких и частых доз, чтобы достигнуть цели, несет риск вызвать гематологические эффекты, которые считают нежелательными побочными действиями.
Также, в существующем уровне техники показано множество других попыток получения молекул из эритропоэтина или пептидов, полученных из него, у которых выявляют нейропротективную активность и отсутствие эритропоэтической активности. В этом отношении, в патенте US 2015119325 описан асиалоэритропоэтин (асиало-rhEPO), полученный в растениях. В заявках на патент US 2009170759, US 2003130197, МХРА 02011727 и US 2003130197 описаны пептиды, которые связываются с рецептором ЕРО, для лечения заболеваний, в которые вовлечена центральная нервная система.
В документе WO2004043382 описан вариант полипептида эритропоэтина человека, который содержит последовательность аминокислот с различием аминокислот в двух или более различных участках модификации ЕРО и повышенной активностью эритропоэтина. В настоящем изобретении предложен человеческий вариант полипептида эритропоэтина, который содержит последовательность аминокислот эритропоэтина человека с различием аминокислот в двух или более различных участках модификации ЕРО и умеренной активностью эритропоэтина, связанной с его цитопротекторной способностью.
В патенте US 2011008363 описаны различные варианты ЕРО, в которых 1-10 аминокислот были удалены с С-конца белка. Это варианты с более низкой молекулярной массой со сниженной эритропоэтической активностью и сохраненным нейропротективным действием.
В заявке US 2007027068 описаны гликопегилированные пептиды ЕРО. Один из мутированных пептидов ЕРО содержит последовательность аминокислот (SEQ ID NO: 73) и содержит по меньшей мере одну мутацию, выбранную из группы, состоящей из Arg<139> на Ala<139>, Arg<143> на Ala<143> и Lys<154> на Ala<154>.
В ES 2457398 (Т3) описан полинуклеотид, который кодирует вариант ЕРО.
В РСТ WO 2005025606 (А1) описан модифицированный ЕРО, который содержит олигосахариды для того, чтобы повысить его эритропоэтическую активность и сохранить цитопротекторную активность.
Аналогичным образом, в WO 2006127910 описан ЕРО с модификациями гликозилированием, чтобы повысить продукцию красных кровяных клеток. Тем не менее, в данном документе упомянуто, что он также является подходящим для лечения нейродегенеративных заболеваний, поскольку он проявляет цитопротекторную функцию.
В заявке, поданной в Аргентине (AR055654), описан рекомбинантный эритропоэтин для лечения нейродегенеративных расстройств. В описании перечислены аминокислоты, которые можно модифицировать, чтобы добавить сайты гликозилирования, и особо отмечена модификация следующих остатков: 87, 88, 90 // 30, 32, 87, 88, 90 // 24, 87, 88, 90 //38, 87, 88, 90 // 83, 87, 88, 90. Целью было присоединить гликаны, чтобы увеличить время полужизни в кровотоке, чтобы повысить эритропоэтическую активность. Ту же цель преследуют, вводя модификации, описанные в патенте WO 9505465, в котором описаны варианты эритропоэтина, которые содержат дополнительные сайты гликозилирования. Некоторые из сайтов, упомянутых в указанном патенте, перечислены далее: 25, 30, 51, 57, 69, 88, 89, 136, 138. Кроме того, упомянуты следующие замены:

В документе WO 2005103076 описаны варианты ЕРО, содержащие четное число остатков цистеина, предпочтительно не более четырех остатков цистеина, или более предпочтительно не более двух остатков цистеина. Предпочтительно, остатки цистеина должны быть в положениях 7, 29, 33 и 161, еще более предпочтительно в положениях 7 и 161. Дополнительные варианты, предложенные в указанной заявке, включают любую мутацию путем вставки в одном или более из следующих положений: 6, 29, 33, 45, 47, 48, 49, 61, 64, 74, 88, 92, 107, 109, 133, 135, 154, 157 и 158.
В заявке US 2011003744 описан состав с эритропоэтином, содержащий данный конъюгированный с полиэтиленгликолем белок или конъюгированный с полиэтаноламином белок посредством ферментативно модифицированных гликозидных остатков, чтобы улучшить гематопоэтические свойства.
В документе МХРА 05000063 описан защищающий ткань рекомбинантный цитокин, который не оказывает по меньшей мере одного из действий эритропоэтина на костный мозг; у указанного защищающего ткань рекомбинантного цитокина будет отсутствовать эритропоэтическая активность; более предпочтительно указанный защищающий ткань рекомбинантный цитокин не оказывает ни одного действия, которое оказывает эритропоэтин на костный мозг.В описании упомянуто, что можно осуществить изменения в одной или более аминокислотах, или делеции или вставки в ЕРО. В предпочтительном варианте реализации защищающий ткань рекомбинантный цитокин содержит одну или более модификаций в одном или более из следующих участков: VLQRY (аминокислоты 11-15 нативного эритропоэтина человека, SEQ ID NO: 1) и/или TKVNFYAW (аминокислоты 44-51 нативного эритропоэтина человека, SEQ ID NO: 2) и/или SGLRSLTTL (аминокислоты 100 - 108 нативного эритропоэтина человека, SEQ ID NO: 3) и/или SNFLRG (аминокислоты 146 - 151 нативного эритропоэтина человека, SEQ ID NO: 4). Другие мутации можно предложить в аминокислотах 7, 20, 21, 29, 33, 38, 42, 59, 63, 67, 70, 83, 96, 126, 142, 143, 152, 153, 155, 156 и 161 в последовательности SEQ ID NO: 10. Данные другие мутации могут быть уникальными или дополнительными к по меньшей мере одной мутации в по меньшей мере одном из ранее упомянутых участков. В некоторых вариантах реализации изменения в одной или более аминокислотах TKVNFYAW (аминокислоты 44-51 нативного эритропоэтина человека, SEQ ID NO: 2) приводят к получению модифицированной молекулы эритропоэтина с частичной функцией, т.е., с меньшей эритропоэтической активностью, чем у rhEPO. В других вариантах реализации изменения находятся в одной или более аминокислотах SGLRSLTTL (аминокислоты 100 - 108 нативного эритропоэтина человека, SEQ ID NO: 3), присутствуют даже модификации содержания сиаловой кислоты, или молекулы без гликанов, или модификации в углеводах, такие как окисление, восстановление или варианты с химическими модификациями, такими как нитрование, ацилирование, сукцинилирование, биотинилирование, йодирование и карбамоилирование. Отсутствует упоминание добавления новых гликанов в сайты, отвечающие за эритропоэтическую активность, с целью ее снижения или блокирования.
Можно оценить, что достижения, описанные в существующем уровне техники, разработанные на данный момент, не дали успешных результатов получения эритропоэтина, который проявляет нейропротективную активность и отсутствие эритропоэтической активности. В соответствии с настоящим изобретением предложены новые мутеины hEPO, для которых продемонстрировали нейропротективное и нейротрофическое действие. Данные мутеины получали с помощью оригинального способа, в котором модифицируют молекулу hEPO посредством создания новых консенсусных сайтов для N-гликозилирования на молекулярном участке hEPO, отвечающем за его связывание с гомо- и гетеродимерными рецепторами. Неожиданно, у новых мутеинов hEPO отсутствовала эритропоэтическая активность, но их
нейропротективная/нейропластическая активность оставалась неизменной или даже улучшенной, и, в свою очередь, обнаружили улучшения в их фармакокинетических свойствах. Указанные модификации исходного ЕРО минимальны, что позволяет сохранить большое сходство со структурой природного белка.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В соответствии с настоящим изобретением предложен модифицированный эритропоэтин человека с отсутствием или снижением его эритропоэтической активности, предпочтительно с до 0,5% эритропоэтической активности относительно эритропоэтина человека, более длительным временем полужизни в плазме, у которого сохраняется его нейропротективная и нейропластическая способность. Связывание с по меньшей мере одним из гомо- или гетеродимерных рецепторов данного модифицированного эритропоэтина человека частично или полностью подавляется. Такое подавление включает мутацию одного из сайтов связывания с гомо- или гетеродимерными рецепторами путем введения консенсусных сайтов для N-гликозилирования.
В предпочтительном варианте реализации настоящего изобретения модифицированный эритропоэтин человека содержит следующие мутации: Tyr1SAsn и Leu17Thr, и содержит SEQ ID NO: 2.
В предпочтительном варианте реализации настоящего изобретения модифицированный эритропоэтин человека содержит следующие мутации: Lys45Asn и Asn47Thr, и содержит SEQ ID NO: 4.
В предпочтительном варианте реализации настоящего изобретения модифицированный эритропоэтин человека содержит следующие мутации: Glu62Asn и Trp64Thr, и содержит SEQ ID NO: 8.
В предпочтительном варианте реализации настоящего изобретения модифицированный эритропоэтин человека содержит следующие мутации: Gln65Asn и Leu67Thr, и содержит SEQ ID NO: 10.
В предпочтительном варианте реализации настоящего изобретения модифицированный эритропоэтин человека содержит следующие мутации: Glu72Asn и Val74Thr, и содержит SEQ ID NO: 12.
В предпочтительном варианте реализации настоящего изобретения модифицированный эритропоэтин человека содержит следующие мутации: Arg76Asn и Gln78Thr, и содержит SEQ ID NO: 14.
В предпочтительном варианте реализации настоящего изобретения модифицированный эритропоэтин человека содержит следующие мутации: Ala98Asn и Ser100Thr, и содержит SEQ ID NO: 16.
В предпочтительном варианте реализации настоящего изобретения модифицированный эритропоэтин человека содержит следующую мутацию: Ser104Asn, и содержит SEQ ID NO: 18.
В предпочтительном варианте реализации настоящего изобретения модифицированный эритропоэтин человека содержит следующие мутации: Thr106Asn и Leu108Thr, и содержит SEQ ID NO: 20.
В предпочтительном варианте реализации настоящего изобретения модифицированный эритропоэтин человека содержит следующую мутацию: Leu149Thr, и содержит SEQ ID NO: 22.
В предпочтительном варианте реализации настоящего изобретения модифицированный эритропоэтин человека содержит следующие мутации: Gly151Asn и Leu153Thr, и содержит SEQ ID NO: 24.
Модифицированный эритропоэтин человека согласно настоящему изобретению проявляет эритропоэтическую активность до 1% относительно эритропоэтина человека. Предпочтительно, он проявляет до 0,5% эритропоэтической активности относительно эритропоэтина человека. Более предпочтительно, он проявляет до 0,2% эритропоэтической активности относительно эритропоэтина человека.
С другой стороны, в соответствии с настоящим изобретением предложены нуклеиновые кислоты, содержащие последовательности нуклеотидов эритропоэтина согласно настоящему изобретению. Также изложены последовательности ДНК, кодирующие каждый из описанных и полученных мутеинов. Данные последовательности ДНК представляют собой: SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 и SEQ ID NO: 23. В альтернативном варианте реализации настоящего изобретения данные нуклеиновые кислоты представляют собой векторы для трансформирования клеток или представляют собой векторы экспрессии или лентивирусные векторы.
В предпочтительном варианте реализации настоящего изобретения последовательность нуклеиновой кислоты, кодирующая модифицированный эритропоэтин человека согласно настоящему изобретению, представляет собой SEQ ID NO: 1.
В предпочтительном варианте реализации настоящего изобретения последовательность нуклеиновой кислоты, кодирующая модифицированный эритропоэтин человека согласно настоящему изобретению, представляет собой SEQ ID NO: 3.
В предпочтительном варианте реализации настоящего изобретения последовательность нуклеиновой кислоты, кодирующая модифицированный эритропоэтин человека согласно настоящему изобретению, представляет собой SEQ ID NO: 7.
В предпочтительном варианте реализации настоящего изобретения последовательность нуклеиновой кислоты, кодирующая модифицированный эритропоэтин человека согласно настоящему изобретению, представляет собой SEQ ID NO: 9.
В предпочтительном варианте реализации настоящего изобретения последовательность нуклеиновой кислоты, кодирующая модифицированный эритропоэтин человека согласно настоящему изобретению, представляет собой SEQ ID NO: 11.
В предпочтительном варианте реализации настоящего изобретения последовательность нуклеиновой кислоты, кодирующая модифицированный эритропоэтин человека согласно настоящему изобретению, представляет собой SEQ ID NO: 13.
В предпочтительном варианте реализации настоящего изобретения последовательность нуклеиновой кислоты, кодирующая модифицированный эритропоэтин человека согласно настоящему изобретению, представляет собой SEQ ID NO: 15.
В предпочтительном варианте реализации настоящего изобретения последовательность нуклеиновой кислоты, кодирующая модифицированный эритропоэтин человека согласно настоящему изобретению, представляет собой SEQ ID NO: 17.
В предпочтительном варианте реализации настоящего изобретения последовательность нуклеиновой кислоты, кодирующая модифицированный эритропоэтин человека согласно настоящему изобретению, представляет собой SEQ ID NO: 19.
В предпочтительном варианте реализации настоящего изобретения последовательность нуклеиновой кислоты, кодирующая модифицированный эритропоэтин человека согласно настоящему изобретению, представляет собой SEQ ID NO: 21.
В предпочтительном варианте реализации настоящего изобретения последовательность нуклеиновой кислоты, кодирующая модифицированный эритропоэтин человека согласно настоящему изобретению, представляет собой SEQ ID NO: 23.
Также в соответствии с настоящим изобретением предложены генетически модифицированные клетки с любой из последовательностей ДНК, выбранных из совокупности, содержащей SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 и SEQ ID NO: 23; причем данная генетически модифицированная клетка способна экспрессировать модифицированный эритропоэтин согласно настоящему изобретению. В предпочтительном варианте реализации данная генетически модифицированная клетка представляет собой клетку растения, или клетку животного, или клетку любого клеточного хозяина, которая обеспечивает добавление N-гликанов. Предпочтительно данная клетка происходит из линии клеток животного. Предпочтительно данная линия клеток выбрана из совокупности, содержащей СНО.K1, HEK293, NSO, BHK-21 и HeLa.
С другой стороны, в соответствии с настоящим изобретением предложен способ получения модифицированного эритропоэтина человека, включающий следующие этапы:
a. Обеспечение последовательностей нуклеиновых кислот, которые кодируют указанный модифицированный эритропоэтин человека;
b. Создание по меньшей мере одного котрансфекционного вектора для пакующих клеток;
c. Котрансфекцию пакующих клеток, которые продуцируют лентивирусные частицы, содержащие данную последовательность нуклеиновой кислоты, которая кодирует указанный модифицированный эритропоэтин человека;
d. Сбор данных лентивирусных частиц, продуцированных пакующими клетками из этапа с;
e. Трансдукцию клеток, способных экспрессировать модифицированный эритропоэтин человека, указанными лентивирусными частицами из этапа d;
f. Селекцию клеток из этапа е, которые содержат последовательность нуклеиновой кислоты, кодирующую модифицированный эритропоэтин человека;
g. Культивирование клеток из этапа f таким образом, чтобы они экспрессировали указанный модифицированный эритропоэтин человека; и
h. Выделение и очистку указанного модифицированного эритропоэтина человека.
При этом на указанном этапе а последовательности нуклеиновых кислот выбраны из совокупности, содержащей SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 и SEQ ID NO: 23.
При этом этап b включает вектор, который обеспечивает проникновение лентивирусной частицы в клетку, вектор, кодирующий матриксный белок, капсид, лентивирусную протеазу, обратную транскриптазу и интегразу, вектор для переноса генов, содержащий последовательность модифицированного эритропоэтина человека, и вектор, который вызывает ядерный экспорт вектора для переноса генов.
При этом на указанном этапе е указанные клетки выбраны изсовокупности, содержащей СНО.K1, HEK293, NSO, BHK-21 и HeLa.
При этом указанный этап h представляет собой очистку на основе иммуноаффинности, которая включает антитело против rhEPO и элюент.
При этом указанный элюент выбран из группы, включающей глицин, уксусную кислоту - NaCl, ацетатные соли, лимонную кислоту, фосфатные соли, этанол, изопропиловый спирт, диоксан, этиленгликоль, трис-HCl и их смеси. Предпочтительно данный элюент выбран из группы, включающей глицин, уксусную кислоту-NaCl. Более предпочтительно, элюент выбран из группы, включающей 0,1 М глицин (рН=2); 0,15 М глицин (рН=2,5) и 0,2 М уксусную кислоту, 0,15 М NaCl (рН=3).
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1. Схема последовательности этапов получения 12 мутеинов hEPO посредством сайт-направленного мутагенеза.
Фигура 2. Анализ фрагментов, полученных в каждой из реакций ПЦР, с применением агарозных гелей, а. Фрагменты, полученные в результате ПЦР1. b. Фрагменты, полученные в результате ПЦР2.
Фигура 3. Анализ in silico потенциальной антигенности мутеинов hEPO и немодифицированной молекулы с применением базы данных иммунных эпитопов (IEDB).
Фигура 4. Вестерн-блоттинг, подтверждающий вставку новых сайтов N-гл икозилир ов ания.
Фигура 5. Производительность элюирования молекулы rhEPO или ее мутеинов, которую оценивали с помощью методики сэндвич-ELISA.
 Протокол А (производительность элюирования после образования комплекса антиген/антитело (АГ/АТ)).
Протокол А (производительность элюирования после образования комплекса антиген/антитело (АГ/АТ)).
Протокол В (степень сохранения связывающей способности с интересующими молекулами после обработки МАТ различными элюентами).
Фигура 6. Очистка мутеина hEPO Mut 104 посредством иммуноаффинной хроматографии (ИАХ) с применением MAT 2 В2.
A. Полная хроматограмма, полученная во время очистки мутеина hEPO Mut 104.
B. Увеличение масштаба зоны элюирования хроматограммы А.
Фигура 7. Оценка очистки мутеина Mut 104 посредством иммуноаффинной хроматографии (ИАХ) с применением 0,15 М глицина (рН 2,5) в качестве элюирующего раствора. Образцы: 1 - загрузка, 2 - проточная фракция, 3 - промывка 1, 4 - промывка 2, 5 - промывка 3, 6 - смесь элюированных фракций, 7 - стандарт rhEPO (Zelltek S.A.)
Фигура 8. Анализ чистоты в элюатах, полученных после иммуноаффинной хроматографии (ИАХ) мутеинов hEPO. Образцы: 1 - маркер молекулярных масс, 2 - стандарт rhEPO (Zelltek S.A.), 3 - элюат Mut 45_47, 4 - элюат Mut 104, 5 - элюат Mut 151_153.
Фигура 9. Определение видимых молекулярных масс мутеинов hEPO. ММ: маркер молекулярных масс.
Фигура 10. Изоэлектрическое фокусирование (ИЭФ) мутеинов hEPO, очищенных с помощью иммуноаффинной хроматографии (ИАХ). Образцы: 1 - Mut 45_47, 2 - Mut 104, 3 - стандарт rhEPO (Zelltek S.А.)
Фигура 11. Электрофореграммы капиллярного зонального электрофореза (КЗЭ), полученные для rhEPO и его мутеинов. A: Mut 45_47; В: Mut 104; С: rhEPO. Термин «пик» соответствует каждому пику, определенному на электрофореграмме КЗЭ, и «площадь» (%) представляет собой процент каждой изоформы, рассчитанный путем интегрирования площади под кривой каждого пика.
Фигура 12а. Сравнение специфической эритропоэтической биологической активности, оцененной in vitro для мутеинов hEPO.
Фигура 12b. Оценка эритропоэтической активности in vivo для мутеинов hEPO. Эритропоэтическую активность in vivo мутеинов hEPO (Mut) оценивали у нормоцитарных мышей (n=4), которых лечили одинаковой массой rhEPO или hEPO mut, в соответствующих случаях, или ФБР в качестве контроля. После лечения рассчитывали процент ретикулоцитов для каждого варианта лечения. ***р<0,001 и н/з (не значимый) представляет собой степень статистической значимости после дисперсионного анализа и апостериорного критерия Тьюки (n=4). Фигура 13. Цито/нейропротективная активность rhEPO и Mut 104, определенная с применением культур клеток нейронального происхождения SH-SY5Y. KSTP: контроль добавления стауроспорина (STP), Кклет: контроль клеток (без добавления STP или мутеинов). *р≤0,05 и **р≤0,01 представляет собой уровень статистической значимости после дисперсионного анализа и апостериорного критерия Даннета (n=3).
Фигура 14. Типичная диаграмма развития нейронов в первичных культурах гиппокампальных нейронов.
Фигура 15. Оценка образования нейритов с помощью клеток N2a мыши, (а) Типичные изображения для каждой из проанализированных групп.(b) Графическое представление, полученное в результате оценки следующих параметров нейритов: средней длины, удлинения наиболее длинного нейрита и среднего количества нейритов на нейрон. *р≤0,05 и **р≤0,01 представляет собой уровень статистической значимости после дисперсионного анализа и апостериорного критерия Бонферрони (n=3).
Фигура 16. Оценка плотности филоподий с применением первичных культур гиппокампальных нейронов из зародышей крысы. *р≤0,05 и **р≤0,01 представляет собой уровень статистической значимости после дисперсионного анализа и апостериорного критерия Бонферрони (n=3).
Фигура 17. Оценка формирования синапса с применением первичных культур гиппокампальных нейронов из зародышей крысы. *р≤0,05 ***р≤0,001 представляет собой уровень статистической значимости после дисперсионного анализа и апостериорного критерия Бонферрони (n=3).
Фигура 18. Оценка нейропротективной биологической активности мутеинов hEPO в отношении первичных культур нейронов на 11 день культивирования in vitro (DIV). Нейропротективную активность in vitro мутеинов hEPO (Mut) оценивали в первичных культурах гиппокампальных нейронов крыс линии Sprague-Dawley на 11 DIV. Культуры обрабатывали одинаковой массой rhEPO или мутеинов, в соответствующих случаях, или ФБР в качестве контроля. Затем вызывали апоптоз путем инкубации со стауроспорином (STP). Иммунофлуоресцентный анализ проводили с помощью реагентов Хехст и фаллоидин-FITC, чтобы проанализировать процент апоптических ядер. ***р≤0,001, **р≤0,01 и н/з (не значимый) представляет собой уровень статистической значимости после дисперсионного анализа и апостериорного критерия Бонферрони.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Как уже упоминали при описании существующего уровня техники, цель состояла в попытке разработать различные стратегии для снижения эритропоэтической активности указанного цитокина, сохранив при этом его нейропротективный и нейротрофический потенциал, которые были основаны на осуществлении химических модификаций молекулы или изменении содержания в ней гликозидов. Тем не менее, в существующем уровне техники не было описано применение гликозилирования в качестве механизма блокирования части молекулы, вовлеченной во взаимодействие, которое отвечает за эритропоэз.
Следовательно, в соответствии с настоящим изобретением предложен новый модифицированный эритропоэтин, что решает проблемы, упомянутые в существующем уровне техники, одновременно добиваясь следующего:
• Ингибирования гематопоэтической функции hEPO у пациентов, которым требуется лечение, относящееся к нейрозащите, и у которых эритропоэз становится нежелательным явлением.
• Увеличения времени полужизни hEPO, чтобы улучшить биологическую активность in vivo, и снижения количества доз, которое нужно вводить.
Таким образом, в соответствии с настоящим изобретением предложено применение гипергликозилированных мутеинов, полученных из hEPO, для получения фармацевтической композиции, предназначенной для предотвращения или лечения заболевания, или генетической предрасположенности страдать таким заболеванием, при котором нейропластичность и/или нейрозащита оказывает полезное действие. Агенты согласно настоящему изобретению можно вводить пациентам с нейродегенеративными заболеваниями, такими как болезнь Альцгеймера (БА), болезнь Паркинсона (БП), боковой амиотрофический склероз (БАС), среди прочих; пациентам, страдающим от заболеваний двигательных нейронов, таких как Болезнь Гентингтона, спиноцеребеллярные атрофии, болезнь Крейтцфельда-Якоба, среди прочих; инвалидизирующими заболеваниями, такими как депрессия или шизофрения; пороками развития, такими как синдром Дауна, а также людям, которые перенесли повреждение нервной ткани в результате острого нарушения мозгового кровообращения или черепно-мозговой травмы.
Термин «мутеин» в данном документе относится к белку, который был модифицирован с помощью сайт-направленного мутагенеза, который содержит замену 1 или 2 его аминокислотных остатков, чтобы ввести консенсусный и потенциальный сайт N-гликозилирования.
Создание или добавление консенсусных сайтов N-гликозилирования включает мутацию аминокислоты в некотором сайте или на участке интересующей молекулы. Для того чтобы клетка осуществляла добавление гликанов, необходимо присутствие сайта Asn-Xaa-Ser/Thr, где Хаа может представлять собой любую аминокислоту, отличную от Pro. Для этого, если присутствует остаток Asn в сайте, в который хотят добавить сайт N-гликозилирования, аминокислоту в положении+2 (относительно Asn) мутируют на Thr, эффективность которого выше по сравнению с Ser, так что остаток Asn занят гликанами. Иначе, если сайт, который нужно модифицировать, содержит аминокислоту в положении +2, свидетельствующую о присутствии Ser или Thr, то данную аминокислоту мутируют на Asn. Иными словами, всегда присутствуют консервативные изменения.
В соответствии с настоящим изобретением предложен эритропоэтин человека, который содержит модификацию структуры аминокислоты, которая позволяет достигнуть следующего:
i. Сохранения аминокислотных остатков, которые составляют сайты, необходимые для молекулярной конформации или для проявления нейропротективной/нейропластической активности.
ii. Модификации аминокислот, предложенных для прекращения гематопоэтической активности.
iii. Создания консенсусных сайтов для N- гликозилирования.
Были сконструированы 12 мутеинов hEPO. В данных мутеинах осуществляли модификацию 1 или 2 аминокислот, чтобы внедрить консенсусные сайты для N-гликозилирования, чтобы нарушить гематопоэтическую активность, сохранить нейропротективную/нейропластическую активность и получить мутантов с превосходящим временем полужизни в плазме.
Способы, осуществленные для получения молекул согласно настоящему изобретению, подробно описаны ниже. Подразумевают, что данные способы служат примерами в настоящей заявке, и их следует понимать в наиболее широком смысле, без ограничения объема защиты настоящего изобретения.
1. Разработка и получение полученных из hEPO мутеинов, у которых нарушена эритропоэтическая активность и сохранено нейропротективное/нейропластическое действие.
Мы разработали 26 олигонуклеотидов: 2 из них - для амплификации новых полноразмерных последовательностей ЕРО, и остальные 24 (прямой и обратный олигонуклеотиды) - для введения 12 точечных мутаций.
В соответствии с предложенным описанием настоящего изобретения, указанные модификации следует толковать как замену аминокислоты в данном положении на другую аминокислоту в том же положении. Следовательно, и взяв в качестве примера мутацию 1 (Lys45 → Asn45) + (Asn47 → Thr47), ее следует толковать как замену остатка лизина в положении 45 на остаток аспарагина в указанном положении и замену аспарагина в положении 47 на треонин.
Стратегия, осуществленная для получения мутеинов hEPO с помощью сайт-направленного мутагенеза, кратко представлена на схеме, показанной на фигуре 1.
Используя ДНК hEPO в качестве матрицы, олигонуклеотиды для каждого мутанта и олигонуклеотиды pMATEPOF (GCCGTCAAGGCCACGTGTCTTGTCCA) и PMATEPOR (AGGCCAGTCTTGTGCTCCAGGTACCG), которые комплементарно связываются с концами последовательности hEPO, проводили первую ПЦР, чтобы ввести каждую из упомянутых выше мутаций. Получили 24 фрагмента, соответствующие 12 мутеинам hEPO. Последовательности нуклеотидов и аминокислот указанных 12 синтезированных мутеинов hEPO (мутеинов) перечислены далее:
Мутеин 15-17 (Tyr15 → Asn15) + (Leu17 → Thr17)
Праймеры перечислены далее:
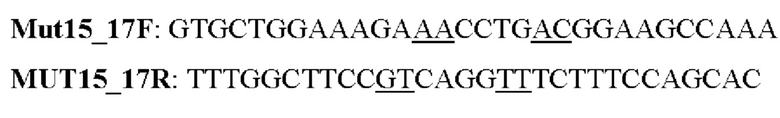
Последовательность нуклеотидов мутеина 15-17 (SEP ID NO: 1)

Последовательность аминокислот мутеина 15-17 (SEP ID NO: 2)

Мутеин 45-47 (Lys45 → Asn45) + (Asn47 → Thr47)
Праймеры перечислены далее:
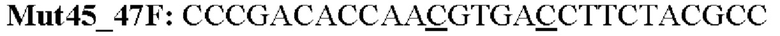
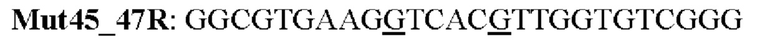
Последовательность нуклеотидов мутеина 45-47 (SEP ID NO: 3)
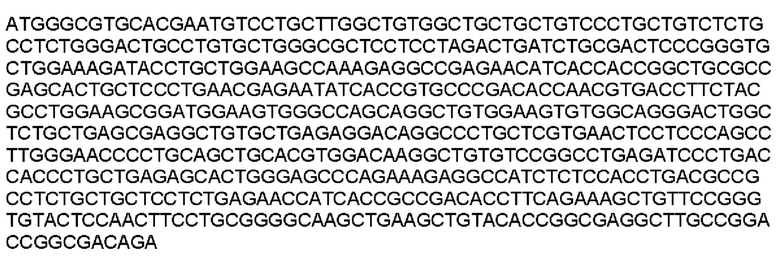
Последовательность аминокислот мутеина 45-47 (SEQ ID NO: 4)

Мутеин 49 (Tyr49→Thr49)
Праймеры перечислены далее:
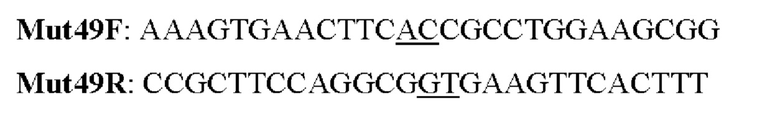
Последовательность нуклеотидов мутеина 49 (SEQ ID NO: 5)
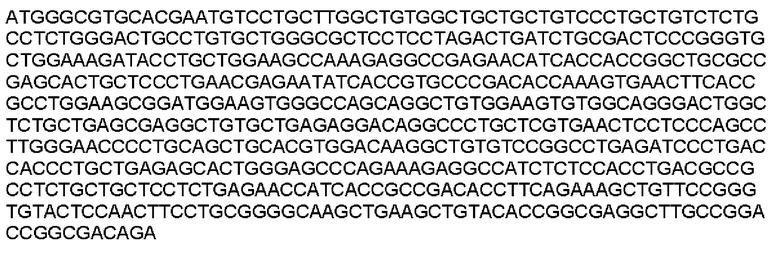
Последовательность аминокислот мутеина 49 (SEQ ID NO: 6)


Мутеин 62-64 (Glu62 →Asn62 + (Trp64 → Thr64)
Праймеры перечислены далее:
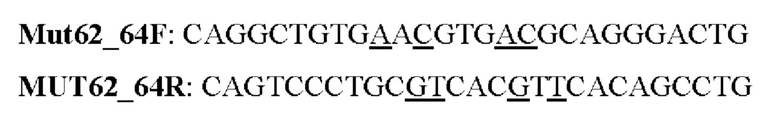
Последовательность нуклеотидов мутеина 62-64 (SEQ ID NO: 7)

Последовательность аминокислот мутеина 62-64 (SEQ ID NO: 8)

Мутеин 65-67 (Gln65 → Asn65) + (Leu67 → Thr67)
Праймеры перечислены далее:
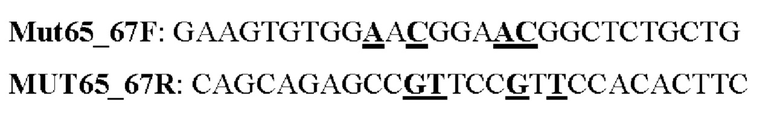
Последовательность нуклеотидов мутеина 65-67 (SEQ ID NO: 9)

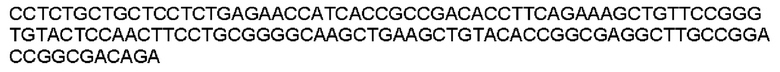
Последовательность аминокислот мутеина 65-67 (SEQ ID NO: 10)

Мутеин 72-74 (Glu72 → Asn72)+(Val74 → Thr74)
Праймеры перечислены далее:
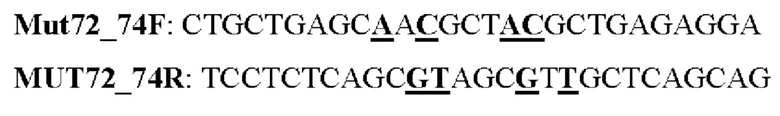
Последовательность нуклеотидов мутеина 72-74 (SEQ ID NO: 11)

Последовательность аминокислот мутеина 72-74 (SEQ ID NO: 12)

Мутеин 76-78 (Arg76 → Asn76)+fGln78 → Thr78)
Праймеры перечислены далее:
Последовательность нуклеотидов мутеина 76-78 (SEP ID NO: 13)

Последовательность аминокислот мутеина 76-78 (SEQ ID NO: 14)

Мутеин 98-100 (А1а98 → Asn98) + (Ser100 → Thr100)
Праймеры перечислены далее:
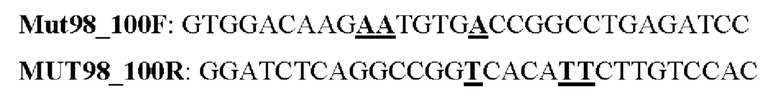
Последовательность нуклеотидов мутеина 98-100 (SEQ ID NO: 15)

Последовательность аминокислот мутеина 98-100 (SEQ ID NO: 16)

Мутеин 104 (Ser104 → Asnl04)
Праймеры перечислены далее:
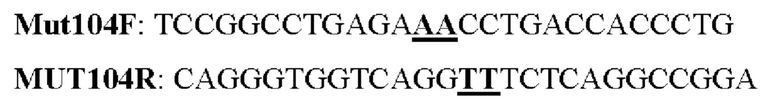
Последовательность нуклеотидов мутеина 104 (SEQ ID NO: 17)

Последовательность аминокислот мутеина 104 (SEQ ID NO: 18)

Мутеин 106-108 (Thr106 → Asn106) + (Leu108 → Thr108)
Праймеры перечислены далее:
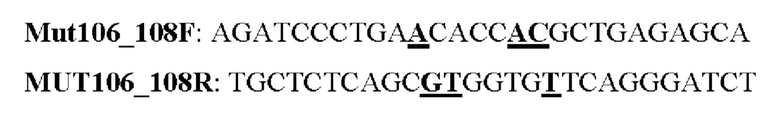
Последовательность нуклеотидов мутеина 106-108 (SEQ ID NO: 19)

Последовательность аминокислот мутеина 106-108 (SEQ ID NO: 20)

Мутеин 149 (Leu149 -→ Thr149)
Праймеры перечислены далее:
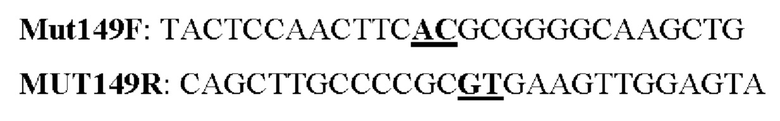
Последовательность нуклеотидов мутеина 149 (SEQ ID NO: 21)

Последовательность аминокислот мутеина 149 (SEQ ID NO: 22)

Мутеин 151-153 (Gly151→Asn151) + (Leu153→Thr153)
Праймеры перечислены далее:
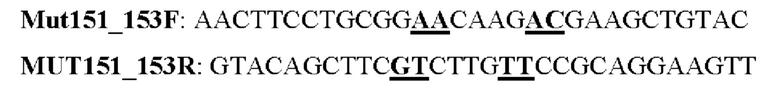
Последовательность нуклеотидов мутеина 151-153 (SEQ ID NO: 23)
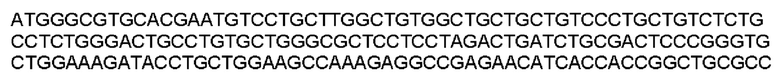
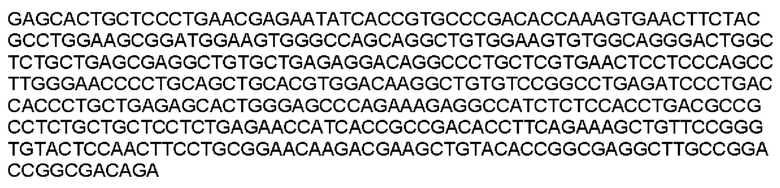
Последовательность аминокислот мутеина 151-153 (SEQ ID NO: 24)

Последовательность ЕРО человека
Последовательность нуклеотидов ЕРО человека (SEQ ID NO: 25)

Последовательность аминокислот ЕРО человека (SEQ ID NO: 26)

Они показаны на фигуре 2а. Данные фрагменты использовали в качестве матрицы для проведения второй ПЦР, на этот раз используя только олигонуклеотиды, которые связываются с концами hEPO, чтобы получить последовательность 12 мутантов (фигура 2b). После реакций ПЦР, продукты, соответствующие 12 мутеинам hEPO, расщепляли с помощью ферментов рестрикции Xbal/Sall в сайтах, которые фланкируют молекулу hEPO, чтобы клонировать их в вектор pLV-PLK, расщепленный теми же ферментами рестрикции. Бактерии TOP TEN трансформировали и проводили селекцию с помощью ампициллина. По 3-4 колонии каждого мутеина hEPO амплифицировали в жидкой среде, чтобы осуществить получение в малом масштабе плазмидной ДНК. Затем авторы настоящего изобретения подтвердили присутствие вставки путем расщепления ферментами рестрикции сайтов, присутствующих в последовательности hEPO и отсутствующих в последовательности вектора. Наконец, полученные в малом масштабе плазмидные ДНК секвенировали, чтобы подтвердить осуществленную вставку мутаций в молекулу hEPO.
2. Анализ in silico потенциальной антигенности мутеинов hEPO.
Базу данных IEDB (Базу данных иммунных эпитопов и средства анализа) использовали для проведения анализа in silico для сравнения потенциальной антигенности мутеинов hEPO и потенциальной антигенности немодифицированной молекулы. Для всех из них осуществляли предсказание Т-клеточных эпитопов - распознаваемых главным комплексом гистосовместимости II класса (МНС II) - путем анализа 8 наиболее типичных аллелей по всему миру (человек, HLA-DR: DRB1*01.01, DRB1*03.01, DRB1*04.01, DRB1*07.01, DRB1*08.01, DRB1* 11.01, DRB1* 13.01, DRB1* 15.01). Показатель антигенности получали для каждого варианта hEPO, и каждый сравнивали с показателем, полученным для hEPO. Степень потенциальной антигенности каждого из них схематически кратко представлена на фигуре 3. Таким образом, наблюдали два мутеина с той же степенью антигенности, что и у hEPO (Mut98_100, Mutl51_153), два потенциально более антигенных мутеина (Mut72_74 и Mut62_64) и, наконец, 8 потенциально менее антигенных мутеинов (Mut45 47, Mut49, Mutl5 17, Mut65 67, Mut 149, Mut76_78, Mut104 и Mut106_108), и данные последние два обладали меньшей антигенностью.
3. Получение линий клеток, которые продуцируют упомянутые мутеины
Лентивирусные частицы собирали для последующей трансдукции клеток СНО.К1. Для этого клетки HEK 293 Т/17 (пакующие клетки) котрансфицировали 4 векторами, чтобы получить соответствующие лентивирусные частицы для каждого из 12 мутеинов. 4 используемых вектора были следующими: pREV (который вызывает ядерный экспорт вектора для переноса генов и его упаковку), pVSVG (который кодирует белок G оболочки VSV, необходимый для проникновения вирусной частицы в клетку, с широким тропизмом), pMDL (который кодирует белки матрикса и капсида, способные упаковать вектор экспрессии, протеазу, обратную транскриптазу и интегразу, необходимые для расщепления структурных элементов и для встраивания в геном клетки) и вектор для переноса генов pLV-PLK-Mut X, из которого все вирусные гены были удалены и замещены на интересующие гены, соответствующие мутеинам hEPO согласно настоящему изобретению. Через два дня после трансфекции клеток HEK 293Т/17 собирали супернатанты каждой из клеток, содержащие лентивирусные частицы mut X, которые использовали для трансдукции клеток СНО.K1. Получали 12 линий клеток СНО.K1 mut X hEPO. Через 72 ч супернатанты собирали и сохраняли для последующего анализа. Аналогичным образом, чтобы получить стабильные рекомбинантные линии клеток, их подвергали давлению возрастающими количествами антибиотика пуромицина, чтобы селекцию прошли клетки, в которые встроился трансген. Линии клеток под давлением антибиотика амплифицировали и криоконсервировали в криогенной смеси (90% (об./об.) ЭБС, 10% (об./об.) ДМСО), и хранили их в емкостях с жидким азотом.
Изначально, с помощью методики сэндвич-ELISA и антител, специфичных к молекуле hEPO, определяли концентрацию, используя культуру линий клеток, полученных путем трансдукции. Моноклональное антитело использовали в качестве захватывающего антитела и поликлональные антитела кролика использовали для детектирования (оба типа антител были разработаны в нашей лаборатории). Полученные концентрации (показанные в таблице I) сочли достаточными для продолжения исследований.
Более того, культуральные супернатанты анализировали с помощью методики вестерн-блоттинга, применяя поликлональные антитела для детектирования всех мутеинов hEPO. Данная методика позволила авторам настоящего изобретения визуализировать, что у мутеинов была большая молекулярная масса по сравнению с немодифицированным hEPO, что соответствовало вставке новых сайтов N-гликозилирования в молекулярную структуру (фигура 4).
Аналогичным образом, проводили анализ методом изоэлектрического фокусирования, а затем анализ методом вестерн-блоттинга, чтобы исследовать количество гликоизоформ в каждом из мутеинов hEPO, экспрессированных в супернатанты культур клеток СНО.K1. Количество изоформ, полученных в 11 из 12 исследованных мутеинов, было выше, чем таковое у немодифицированного hEPO. Разброс количества изоформ находился в диапазоне от 8 до 16 изоформ по сравнению с 7 изоформами, которые наблюдали в культуральном супернатанте немодифицированного hEPO.
В соответствии с настоящим изобретением предложены мутеины hEPO, которые обладают небольшой или не обладают эритропоэтической активностью, чтобы избежать побочных действий, вызванных цитокином, когда его применяют в качестве потенциального нейропротективного/нейропластического кандидатного средства. Для этой цели исследовали упомянутую выше активность in vitro, применяя клетки TF-1, пролиферация которых зависит от присутствия hEPO. Для того чтобы измерить эритропоэтическую активность, пролиферацию данных клеток оценивали после 96 часов стимуляции стандартным hEPO с хорошо известной биологической активностью. У всех разработанных мутеинов hEPO успешно выявили пониженную или отсутствующую эритропоэтическую активность по сравнению с коммерчески доступным стандартным hEPO и немодифицированной молекулой hEPO, за исключением Mut49, у которого сохранилась эритропоэтическая активность. Результаты кратко представлены в таблице I.
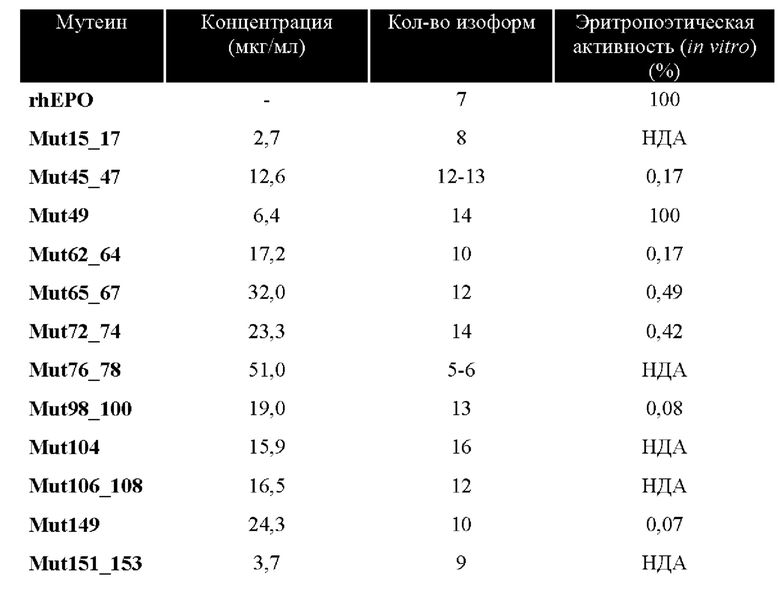

В соответствии с настоящим изобретением предложены мутеины hEPO, содержащие сайты, поддающиеся N-гликозилированию, в положениях, которые не нарушают цитопротекторную биологическую активность, но нарушают гематопоэтическую активность. Они экспрессировались в супернатанты культур клеток СНО.K1, и были исследованы. У всех из них выявили более высокую степень гликозилирования, достигая цели добавления дополнительного сайта N-гликозилирования, что позволяет увеличить их время полужизни в плазме. Аналогичным образом, подтвердили, что модификации, внесенные в молекулу, оказали действие на гематопоэтическую биологическую активность, устранив ее в большинстве мутеинов или сильно снизив ее в остальных.
4. Очистка полученных из hEPO мутеинов с помощью иммуноаффинной хроматографии
Процедура очистки мутеинов согласно настоящему изобретению представлена ниже. Для лучшего понимания данной процедуры следующие мутеины представлены в качестве примера: Mut 45_47, Mut 104, Mut 151_153, с помощью схемы иммуноаффинной (ИА) очистки. Изначально авторы изобретения симулировали процесс, эквивалентный таковому, который происходит в ИА матрице, с целью установить наиболее благоприятное условие для элюирования каждого из гипергликозилированных мутеинов hEPO, захваченных моноклональным антителом против rhEPO, которое было разработано для данной цели в нашей лаборатории (протокол А). На основании описанного выше, проводили анализы сэндвич-ELISA, чтобы сосчитать отношение производных hEPO, которые остались связаны с MAT 2 В2 после того как комплекс антиген-антитело подвергли каждому условию элюирования. Аналогичным образом, авторы изобретения оценили влияние каждого элюирующего раствора на антитело, чтобы определить, будут ли они влиять на способность связываться с мутеинами hEPO, учитывая последующие процедуры повторного использования ИА матриц (протокол В).
Оценивали элюирующую способность следующих растворов:
1. Глицин 0,1 М, рН 2
2. Глицин 0,15 М, рН 2,5
3. Уксусная кислота 0,2 М, NaCl 0,15 М, рН 2,5
4. Глицин 0,15 М, рН 3
5. Лимонная кислота 0,1 М, рН 3
6. Уксусная кислота 0,2 М, NaCl 0,15 М, рН 3
7. Глицин 0,15 М, рН 3,5
8. Ацетат натрия 0,1 М, рН 4
9. Ацетат натрия 0,1 М/диоксан 10% (об./об.), рН 4
10. Ацетат натрия 0,1 М, рН 5
11. Фосфат натрия 0,1 М, рН 6
12. Изопропиловый спирт 40% (об./об.) в фосфатно-солевом буферном растворе (ФБР), рН 7
13. ФБР, рН 7
14. Этанол 40% (об./об.) в ФБР, рН 7
15. Диоксан 10% (об./об.) в ФБР, рН 7
16. Этиленгликоль 40% (об./об.) в ФБР, рН 7
17. Трис/HCl 0,1 М, рН 8
18. Глицин 0,1 М, рН 9
19. Глицин 0,1 М, рН 10
20. Глицин 0,1 М, рН 11
21. Фосфат натрия 0,1 М, рН 11
22. Фосфат натрия 0,1 М, рН 11,7
Значения поглощения, полученные для комплекса антиген-антитело, обработанного ФБР, считали контролем, приняв за 100% образование комплекса антиген-антитело; то есть, без десорбции rhEPO или его мутеинов. Для протокола В значения поглощения, полученные для антитела, обработанного ФБР (до образования комплекса АГ/АТ), считали контролем сохранения способности антитела связываться с исследуемыми молекулами. Следовательно, результаты, полученные для остальных исследованных растворов, выражали относительно контролей, оцененных с ФБР (фигура 5).
Во всех случаях наблюдали, что условия, созданные глицином 0,1 М, рН 2, обладали наибольшей способностью к десорбции антигена по сравнению с другими растворами. Тем не менее, способность образования упомянутого выше комплекса значительно снижалась после предшествующей обработки им. Следовательно, применение такого элюента в иммуноаффинной хроматографии не удобно, поскольку он будет влиять на повторное применение хроматографической матрицы.
С другой стороны, данный анализ позволил выбрать два элюирующих раствора в качестве кандидатов для применения для иммуноаффинной хроматографии (ИАХ): глицин 0,15 М, рН 2,5, и уксусную кислоту 0,2 М, NaCl 0,15 М, рН 3. Оба раствора были способны диссоциировать комплексы АГ/АТ, снимая интересующий белок (таблица II) и, кроме того, не влияли на способность антитела связывать антиген после предшествующей обработки ими.

MAT 2 В2, использованное в ИА смолах, предварительно очищали с помощью аффинной хроматографии на белке А и проводили диализ против раствора карбоната. Затем осуществляли связывание со смолой сефарозой 4 В, активированной CNBr. Уровень связывания рассчитывали путем измерения концентрации иммуноглобулинов в растворе до и после реакции иммобилизации, получив в результате 96%. Таким образом, теоретическая емкость rhEPO составляла 481 мкг на мл геля.
На фигуре 6 показан типичный хроматографический профиль очистки мутеина Mut 104 с применением элюирующего раствора глицина 0,15 М, рН 2,5.
Методику сэндвич-ELISA использовали для оценки в каждой хроматографической процедуре присутствия варианта hEPO в различных фракциях (проточная фракция, промывки, элюирование), чтобы рассчитать параметры, которые позволяют анализировать производительность процесса.
Результаты, полученные для очисток, показаны в таблице III. В динамических условиях наблюдали некоторые различия в отношении эффективности получения интересующего белка по сравнению со статическими условиями, которые оценивали в планшетах. Элюирование раствором с рН 2,5 приводило к лучшему выходу мутеинов Mut 104 и Mut 151_153, по сравнению с раствором с рН 3 (54% по сравнению с 19% и 55% по сравнению с 21%, соответственно), тогда как для варианта Mut 45_47 получали больший выход, когда элюировали раствором с рН 3 (49% по сравнению с 34%).
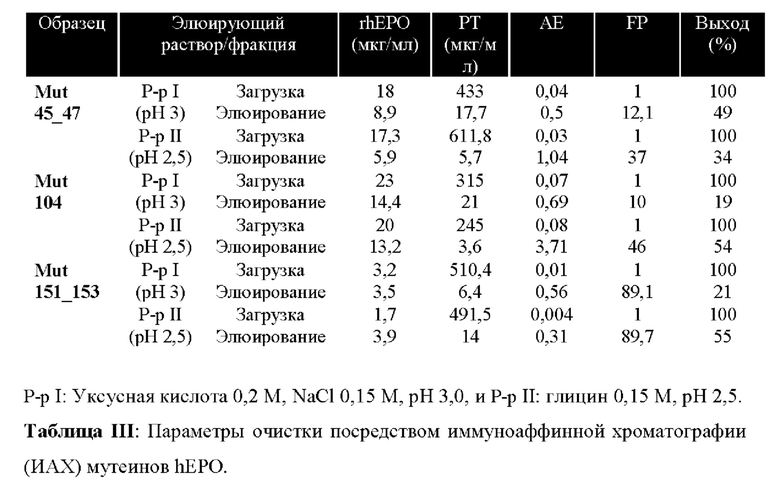
При анализе различных стадий хроматографического процесса мутеина Mut 104 с помощью ПААГ/ДСН с последующим окрашиванием красителем кумасси бриллиантовым голубым (фигура 7) можно наблюдать, что большая часть примесей, присутствующих в образце, не сохранялась и содержалась в проточной фракции и промывках. Следовательно, получали высокую степень чистоты, что наблюдали на дорожке 6 упомянутой фигуры.
Хотя ИАХ не показала высокого выхода интересующего белка, это был наиболее подходящий способ очистки мутеинов hEPO, так как он позволял их очистку в 37-90 раз относительно исходного образца, с высокой степенью чистоты и на одном этапе хроматографии.
Оценку чистоты элюатов, соответствующих каждому мутеину hEPO, проводили с помощью денситометрии полос (фигура 8).
Перед электрофоретическим запуском образцы концентрировали в 75-80 раз, применяя картриджи для диафильтрации с порогом отсечки 10 кДа. Полученная чистота была выше 89% для всех мутеинов (таблица IV), что является характерным значением для процедуры ИА очистки.

Результатом ИАХ была высокая степень чистоты мутеинов hEPO, которую сочли подходящей для применения в следующих тестах. Таким образом, установили, что ИАХ представляет собой подходящий, простой и практичный способ очистки мутеинов hEPO согласно настоящему изобретению.
5. Физико-химическое исследование мутеинов hEPO.
5.1. Определение видимых молекулярных масс различных мутеинов hEPO.
На фигуре 9 показан вестерн-блоттинг мутеинов, которые очищали с помощью иммуноаффинной хроматографии (ИАХ). Ее применяли для расчета молекулярных масс таких мутеинов, применяя известные маркеры молекулярных масс.
Определение видимых молекулярных масс гипергликозилиро ванных мутеинов hEPO осуществляли путем интерполяции расстояний пробега передней и задней границы полосы, соответствующей каждому варианту в вариационной кривой расстояния, на которое мигрировал каждый маркер, в зависимости от log его молекулярной массы. Видимые молекулярные массы, рассчитанные для каждого варианта, кратко представлены ниже:
*Mut 45_47: 34-66 кДа
*Mut 104: 29-66 кДа
*Mut 151_153: 35-45 кДа
*rhEPO: 31-43 кДа
Такое определение подтвердило успешное встраивание дополнительного сайта N-гликозилирования в молекулу hEPO, так как у мутеинов выявили большие средние молекулярные массы, чем молекулярная масса немодифицированного hEPO.
5.2. Определение профиля изоформ с помощью ИЭФ
Для того чтобы оценить степень гетерогенности мутеинов hEPO вследствие гликозилирования и, в частности, содержание остатков сиаловой кислоты, образцы оценивали с помощью ИЭФ, чтобы определить изоформы с различными pi, которые входят в состав каждого гипергликозилированного варианта hEPO.
На фигуре 10 показано обнаружение 13 изоформ, которые входят в состав варианта Mut 45_47, 14 изоформ для варианта Mut 104 и 6 изоформ для стандартного rhEPO. Стоит упомянуть, что стандартный rhEPO представляет собой гормон, полученный как биотерапевтическое средство, предназначенное для способствования эритропоэзу, в котором преобладают наиболее кислые изоформы молекулы hEPO, которые получают после разработанного процесса очистки с помощью 4 этапов хроматографии. Кроме того, можно увидеть, что мутеины Mut 45_47 и Mut 104 содержат 5-7 более кислых изоформ, чем изоформы rhEPO, что отражает более высокое содержание сиаловой кислоты в данных молекулах.
Результаты, полученные при определении видимых молекулярных масс, вместе с результатами ИЭФ подтверждают, что у полученных мутеинов более высокая степень гликозилирования по сравнению с rhEPO.
5.3. Анализ профиля изоформ посредством капиллярного зонного электрофореза (КЗЭ) очищенных с помощью ИАХ мутеинов hEPO
КЗЭ также применяли для определения изоформ каждого варианта hEPO, который считают примером в настоящей заявке. Проводили обработку образцов, полученных при очистке с помощью ИАХ мутеинов hEPO, подвергнутых диафильтрации водой и концентрированных до приблизительно 1 мг/мл.
КЗЭ дает количественную информацию о различных наблюдаемых изоформах. Таким образом, используя данные КЗЭ, каждому наблюдаемому пику для каждого варианта hEPO присваивали изоформу. Для мутеинов Mut 45_47 и Mut 104 от 1 до 11 и для стандартного rhEPO от 1 до 7; изоформа 1 представляла собой изоформу, которая мигрировала лучше всего вдоль капиллярной зоны (фигура 11). Затем рассчитывали проценты каждой изоформы путем интегрирования площади под кривой для каждого пика.
При сравнении электрофоретических подвижностей изоформ каждого варианта с rhEPO обнаружили, что изоформы последнего совпадают с изоформами 1-7 Mut 45_47 и с изоформами 3-9 Mut 104, показывая, что для Mut 45_47 выявили 4 более кислые изоформы по сравнению с rhEPO, тогда как для Mut 104 выявили 2 менее кислые изоформы и 2 более кислые изоформы по сравнению с rhEPO.
При оценке соотношений каждой изоформы показали, что rhEPO содержит более высокий процент изоформ 3, 4 и 5, тогда как для Mut 45_47 показали более высокий процент изоформ 6, 7, 8 и 9 и для Mut 104 показали более высокий процент изоформ 4, 5,6 и 7. Это снова подтверждает гетерогенность каждого варианта hEPO в отношении степени гликозилирования rhEPO вследствие более высокого содержания более кислых изоформ и их более высокого процента.
При рассмотрении электрофореграммы каждого варианта hEPO, присутствие двух дополнительных пиков (еще двух изоформ) можно визуализировать в обоих случаях: одного перед пиком 1 и другого перед пиком 11, которые невозможно точно разделить. Это является следствием низкого процента такой изоформы в образце, который ниже, чем предел детектирования системы. Это объясняет различие в количестве изоформ, детектированных с помощью ИЭФ и КЗЭ.
5.4.а. Характеристика эритропоэтической биологической активности мутеинов in vitro
Проводили биологическое исследование очищенных и разработанных мутеинов hEPO. Для этого проводили анализ пролиферации in vitro, применяя культуры линий клеток UT-7, чтобы оценить эритропоэтическую биологическую активность мутеинов, которые взяли в качестве примеров в настоящей заявке, чтобы показать вариант реализации настоящего изобретения (A-F), поскольку выживаемость и пролиферация данных линий клеток зависит от присутствия hEPO в ростовой среде. В отличие от анализа, в котором применяют линию клеток TF-1, анализ, в котором применяют линию клеток UT-7, характеризуется более высокой чувствительностью ответа, поэтому его выбрали для данной стадии работы.
Проанализировали культуральные супернатанты со всеми мутеинами, экспрессированными соответствующими продуцирующими линиями клеток, и три очищенных с помощью ИАХ мутеина, которые взяли в качестве примеров. Пролиферацию, вызванную данными молекулами, сравнивали с пролиферацией, вызванной стандартным rhEPO.
У мутеинов hEPO, которые взяли в качестве примеров в настоящей заявке (за исключением Mut 49), выявили низкую или отсутствующую способность стимулировать пролиферацию клеток при исследовании при тех же концентрациях, что и у стандартного rhEPO (фигура 12а). Для того чтобы рассчитать специфическую эритропоэтическую активность (СЭА), которую оценивали in vitro, авторы настоящего изобретения работали с более высокими концентрациями мутеинов, то есть, с таковыми, присутствующими в чистых культуральных супернатантах. Следовательно, у мутеинов Mut 104 и Mut 151153 выявили полную утрату эритропоэтической биологической активности, тогда как у Mut 45_47, Mut 62 64 и Mut 98 100 выявили очень низкую эритропоэтическую активность (СЭА Mut 45_47=0,2 МЕ/мкг, СЭА Mut 62 64=0,2 МЕ/мкг и СЭА Mut 98 100=0,1 МЕ/мкг, по сравнению с СЭА=120 МЕ/мкг для rhEPO). Напротив, считали, что у Mut 49 сохранялась такая активность (СЭА=216 МЕ/мкг). Хотя наблюдали повышение СЭА Mut 49, причиной этому могла быть зона кривой, которую использовали для определения такого параметра, учитывая, что уклон кривой линейной зоны ответа Mut 49 значительно отличался от такового у стандарта. Следовательно, модификацию, сделанную для получения такого мутеина, сочли неэффективной для ингибирования его эритропоэтической активности.
Кроме того, цито/нейропротективную биологическую активность in vitro очищенных мутеинов rhEPO анализировали в качестве примера настоящего изобретения в данной заявке. Проводили оценку, чтобы исследовать цито/нейропротективную активность: способность rhEPO и его мутеинов защищать нервные клетки SH-SY5Y от апоптического/цитотоксического действия, вызванного стауроспорином (STP). Таким образом, анализ цито/нейрозащиты состоял в защите нервных клеток путем добавления rhEPO или его мутеинов за 12 ч до индукции повреждения клеток, вызванного STP. По прошествии времени индукции повреждения клеток жизнеспособность культур оценивали путем определения метаболически активных клеток.
В данном анализе как rhEPO, так и очищенный Mut 104, который взяли в качестве примера настоящего изобретения в данной заявке, использовали при одинаковых концентрациях (фиг. 13).
Полученные результаты анализировали статистически, применяя дисперсионный анализ, а затем критерий Даннета, чтобы сравнить каждый из образцов с KSTP. Процент цитотоксичности, полученный для KSTP, приняли за 100%-ную цитотоксичность, и, следовательно, значения цитотоксичности, определенные для каждого из образцов, рассчитывали относительно KSTP.
На фиг. 13 показано, что rhEPO был способен снизить цитотоксичность, вызванную STP, на 45% (р<0,05), тогда как для Mut 104 выявили значительное снижение цитотоксичности, вызванной STP, на 65% с уровнем значимости 99% (р<0,01).
5.4. b. Характеристика эритропоэтической биологической активности мутеинов in vivo
У мутеинов, которые взяли в качестве примеров в настоящей заявке, выявили отсутствие или небольшую биологическую активность in vitro. В свою очередь, обширное гликозилирование придает фармакокинетические свойства, необходимые для увеличения времени присутствия в крови. Данные свойства позволят молекулам улучшить недостаточную способность взаимодействия с рецептором и продемонстрировать эритропоэтическую активность.
Для оценки такой активности in vivo нормоцитарным мышам вводили путем инъекции rhEPO в качестве стандарта, или каждый из трех мутеинов, используемых в предыдущих примерах, или ФБР в качестве отрицательного контроля. Во всех случаях, соответствующих введению белков, использовали массу белков, эквивалентную 80 ME rhEPO. Кровь собирали через 96 ч после введения, и проточную цитометрию использовали для определения процентного содержания ретикулоцитов, меченых тиазолом оранжевым.
На фигуре 12b показан значимо отличающийся ответ (р<0,001) для каждого мутеина и отрицательного контроля по сравнению с rhEPO. Кроме того, не наблюдали значимых различий при сравнении ответа ретикулоцитов, вызванного каждым мутеином, с таковым для отрицательного контроля. В обоих типах анализа подтвердили ожидаемую утрату эритропоэтической активности мутеинов в результате N-гликоинженерии с помощью подхода гипергликозилирования.
5.5. Влияние гипергликозилированных мутеинов hEPO на структурную пластичность нейронов
5.5.1. Развитие (дифференцировка) нейронов и структурная пластичность
Структурная пластичность нейронов включает процессы, посредством которых стимулируют или способствуют развитию и/или дифференцировке нейронов. В этом смысле, химические агенты или соединения, которые вызывают образование нейритов и/или рост аксонов, развитие филоподиев/дендритных шипиков и/или увеличение количества синапсов, будут считать нейротрофическими соединениями. Первичные культуры нейронов и линии нервных клеток широко применяют для исследования таких процессов in vitro.
Стадии развития нейронов в первичных культурах гиппокампальных нейронов представлены на фигуре 14. Данные стадии были основательно охарактеризованы в работе Dotti и др. (1998) [31], где описано, что после посева нейроны выпускают ламеллы, с помощью которых они прикрепляются к субстрату (0 DIV, дней в культуре in vitro). Затем, в течение первых двух суток (1-2 DIV) образуются незрелые нейриты как выпячивания плазматической мембраны. На 2-3 DIV один из данных нейритов удлиняется дальше остальных и обособляется с образованием аксона, который содержит на своем конце треугольную структуру, известную как конус роста аксонов. Впоследствии, нейриты разветвляются, образуя вторичные нейриты (3-4 DIV). На данных нейритах образуются тонкие цитоплазматические выступы, которые содержат перекрестно-связанные актиновые филаменты, известные как филоподии (4-5 DIV). Филоподии затем могут приводить к образованию дендритных шипиков, которые являются преимущественными сайтами возникновения синапсов (7-21 DIV).
В данной части исследования оценивали нейротрофическое действие различных гипергликозилированных мутеинов hEPO согласно настоящему изобретению на различных стадиях развития нейронов. 5.5.2.
Образование нейритов
Для того чтобы определить, оказывали ли мутеины согласно настоящему изобретению нейритогенное действие (увеличение количества и длины нейритов на нейрон), использовали линию нервных клеток N2a (нейробластома мыши). Клетки (50000) высевали на покровные стекла в культуральных планшетах (24 лунки) и поддерживали в полной среде DMEM, дополненной 20% (об./об.) фетальной бычьей сыворотки (ЭБС), которая ингибирует дифференцировку нервных клеток, и гентамицином в качестве антибиотика. Затем культуральную среду заменяли на среду без ЭБС, дополненную различными концентрациями (50 и 300 нг/мл) rhEPO или мутеинов, на 3 ч, чтобы вызвать нейритогенез [32]. По прошествии этого времени клетки фиксировали параформальдегидом 4% (ПФА) с 4% сахарозой в фосфатно-солевом буферном растворе (ФБР) при 4°С в течение 10 минут и пермеабилизовали 0,1% (об./об.) тритоном Х-100 в ФБР в течение 2 минут. Фиксированные клетки затем блокировали 3% (масса/объем) бычьего сывороточного альбумина (БСА) в ФБР в течение 1-2 ч, а затем инкубировали с моноклональным антителом IgG мыши против альфа-тубулина в 1% (масса/объем) БСА (1:1000, Sigma), чтобы пометить нейриты, в течение 16 ч при 4°С. На следующий день, после 3 промывок в ФБР, осуществляли инкубирование с конъюгированным с родамином фаллоидином, чтобы пометить актиновые филаменты (1:1000, Invitrogen), и с антителами козы против мыши, конъюгированными с Alexa 488 (1:1000, Invitrogen), в течение 1 ч при комнатной температуре. После трех промывок в холодном ФБР в течение 5 минут покровные стекла закрепляли с помощью Fluorsave (Calbiochem).
Для количественного определения нейритогенеза получали изображения с помощью эпифлуоресцентного микроскопа Nikon ТЕ2000 (Nikon). Количественный анализ проводили путем подсчета количества нейритов и их средней длины (удлинения нейритов) и длины наиболее длинного нейрита (рост аксонов) у по меньшей мере 30 нейронов на условие, применяя интегрируемый программный модуль NeuroJ программного обеспечения Image J (NIH). Типичные изображения затем обрабатывали с помощью Adobe Photoshop и проводили статистический анализ с помощью программного обеспечения GraphPadPrism 5.
На фигуре 15а показаны типичные изображения для каждой из исследованных групп.Статистический анализ каждого измерения подробно описан справа. Мутеины согласно настоящему изобретению оказывают значимое зависимое от дозы нейрогенное действие, которое обнаруживают по увеличению количества нейритов на нейрон, удлинению нейритов и росту аксонов. Данные действия сопоставимы и дают значения, сходные с таковыми, полученными для rhEPO. Другими словами, у мутеинов согласно настоящему изобретению выявили нейротрофическое действие, сходное с таковым, наблюдаемым для rhEPO в линии клеток N2a.
5.5.3. Плотность филоподии
Филоподии и дендритные шипики представляют собой выпячивания мембраны, обогащенные актиновыми филаментами, которые отходят от нейрита/дендрита и действуют как постсинаптические компартменты, которые очень многочислены в возбуждающих синапсах центральной нервной системы. Морфология шипиков изменчива, и их классифицируют в соответствии с их различной структурой. Общепризнанно, что дендритные шипики могут изменять свою форму/структуру во время развития нейронов, способствуя пластичности нейронов [33]. В этом смысле, некоторые нейродегенеративные заболевания были связаны с изменениями в форме и количестве дендритных шипиков (как увеличением, так и уменьшением). При синдроме Дауна, например, определили, что количество шипиков сильно снижено [34]. Наоборот, гораздо большие количества шипиков находят при расстройствах аутистического спектра [35].
Следовательно, способность стимулировать пластичность нейронов определяли по стимуляции образования филоподиев гипергликозилированными мутеинами согласно настоящему изобретению. Авторы настоящего изобретения использовали первичные культуры гиппокампальных нейронов, на которые воздействовали различными концентрациями (50 и 300 нг/мл) rhEPO или мутеинов Mut 104, Mut 45-47 и Mut 151-153 в течение 4 дней (4 DIV).
Культуры нервных клеток получали из гиппокампа зародышей крысы (Е19), как было описано ранее [36]. Вкратце, ткань обрабатывали трипсином-ЭДТА (0,25% (масса/объем)) при 37°С в течение 15 минут. Раствор полностью диспергированных клеток готовили в среде Neurobasal (NB, Invitrogen), дополненной 2 мМ глутамином, 100 единицами/мл пенициллина (пен), 100 мкг/мл стрептомицина (стреп) и 10% (об./об.) лошадиной сывороткой. Клетки в количестве 20000-30000 высевали в 24-луночные культуральные планшеты, предварительно обработанные 0,1 мг/мл гидробромида поли-L-лизина (Sigma) и 20 мг/мл ламинина (Invitrogen). Через 2 ч среду заменяли на питательную среду определенного состава (NB с 1 г/л овальбумина; N2 и В27, которые представляют собой бессывороточные добавки от Invitrogen) и добавляли различные концентрации (50 и 300 нг/мл) rhEPO или мутеинов согласно настоящему изобретению и инкубировали в течение 4 DIV. Клетки фиксировали 4% (масса/объем) ПФА с 4% (масса/объем) сахарозы в ФБР при 4°С в течение 10 минут.З атем клетки пермеабилизовали 0,1% (об./об.) тритоном Х-100 в ФБР в течение 2 минут. Фиксированные клетки затем блокировали 3% (масса/объем) бычьего сывороточного альбумина (БСА) в ФБР в течение 1 - 2 ч и инкубировали в течение 16 ч при 4°С с моноклональным антителом IgG мыши против альфа-тубулина в 1% (масса/объем) БСА (1:1000, Sigma), чтобы пометить нейриты. На следующий день, после трех промывок в ФБР, осуществляли инкубирование с конъюгированным с родамином фаллоидином, чтобы пометить актиновые филаменты (1:1000, Invitrogen), и со вторичным антителом козы против антител мыши, конъюгированным с Alexa 488 (1:1000, Invitrogen), в течение 1 ч при комнатной температуре. После трех промывок в холодном ФБР в течение 5 минут покровные стекла закрепляли с помощью Fluorsave (Calbiochem).
Для количественного определения образования филоподиев получали изображения с помощью эпифлуоресцентного микроскопа Nikon Е600 (Nikon). Количественный анализ проводили путем подсчета количества филоподиев (богатых актином выпячиваний, выступающих из мембраны нейрита), присутствующих в 20 мкм нейрита на расстоянии менее 50 мкм от тела нервной клетки (по 3 нейрита/нейрон в по меньшей мере 30 нейронах). Типичные изображения затем обрабатывали с помощью Adobe Photoshop и проводили статистический анализ с помощью программного обеспечения GraphPadPrism 5.
На фигуре 16 (панель слева) показаны типичные изображения плотности филоподиев, тогда как на панели справа подробно представлен статистический анализ для каждой из групп.Стоит отметить, что мутеины согласно настоящему изобретению значительно индуцируют, в зависимости от дозы, образование филоподиев по сравнению с контрольными нейронами (ФБР). Неожиданно, данные эффекты даже превосходят таковые, наблюдаемые для нейронов, обработанных rhEPO. В заключение, у мутеинов согласно настоящему изобретению выявили нейротрофическое действие на первичные культуры гиппокампальных нейронов, которое превосходило таковое, наблюдаемое для rhEPO. 5.5.3.
Синапсы
Синапс определяют как высокоспециализированные контакты (соединения) мембран между двумя нейронами. Такое соединение (известное как синаптическая щель) способствует проведению электрического импульса и перемещению веществ из одного из них (пресинаптического) в другой (постсинаптический). Были разработаны различные методики подсчета количества синапсов. Таким образом, общепринятое определение синапса состоит в совместной встречаемости кластеров белков исключительно из пресинаптического компартмента с кластерами белков исключительно из постсинаптического компартмента.
Продолжая исследование нейротрофического действия мутеинов согласно настоящему изобретению, оценивали их способность вызывать образование нервных синапсов.
Для этого проводили анализ иммунодетектирования в 15 DIV первичных культурах нервных клеток, в которых формирование синапса обнаруживали по наложению пре- и постсинаптического маркера.
Вкратце, нейроны (15000/лунку) обрабатывали различными концентрациями (50 и 300 нг/мл) мутеинов hEPO, или rhEPO, или ФБР в течение 15 DIV. Затем их фиксировали раствором 90% (об./об.) метанола и 10% (об./об.) MES (100 мМ MES рН 6,9, 1 мМ EGTA, 1 мМ MgCl2) при 4°С в течение 10 минут.Впоследствии их промывали три раза ФБР, профильтрованным с твин 20 (0,1% (об./об.)), в течение 5 минут.Проводили блокирование раствором ЭБС с тритоном Х-100 (10% ЭБС (об./об.), тритон Х-100 0,1% (об./об.) в ФБР) в течение 1 ч при комнатной температуре, а затем их инкубировали с раствором 3% (масса/объем) БСА также в ФБР в течение 15 минут при комнатной температуре. Оба блокирующих раствора ранее центрифугировали в течение 10 минут при максимальной скорости. Затем их инкубировали 12-16 ч при 4°С с первичным антителом мыши против NMDA-R1 (постсинаптический маркер от Synaptic Systems) и антителом кролика против синаптофизина (пресинаптический маркер от Synaptic Systems). Оба антитела получали в растворе 1% (масса/объем) ФБР-БСА и центрифугировали в течение 10 минут при максимальной скорости. После трехкратной промывки в ФБР их снова блокировали в 3% (масса/объем) БСА, центрифугированном в течение 10 минут, и в 10% (об./об.) растворе ЭБС, тритон Х-100 0,1% (об./об.) в ФБР в течение 1 ч при комнатной температуре. Затем их инкубировали со вторичными антителами против антител мыши, конъюгированными с Alexa 647, и антителами против антител кролика, конъюгированными с Alexa 568, в 1% (масса/объем) БСА и центрифугировали 10 минут при максимальной скорости (оба от Molecular Probes). Затем их закрепляли на подложке с помощью Fluorsave.
Фотографии снимали с помощью конфокального микроскопа Olympus FV1000, работающего вместе с инвертированным микроскопом Olympus 1X81. Изображения затем обрабатывали с помощью программного обеспечения FluoView (версии 3.3, Olympus; 60Х объектив; AN 1.42; разрешение 0,066 мкм/пиксель), в соответствии с критериями Найквиста.
Формирование синапса измеряли как точки колокализации на 25 мкм дендрита пресинаптического маркера и постсинаптического маркера в приблизительно 10-20 нейронах на условие, используя 3 сегмента на нейрон. Данные точки колокализации определяли, применяя анализатор Puncta, интегрируемый программный модуль Image J (версии 1.28u) [37].
На фигуре 17 показаны типичные фотографии каждого из исследованных условий и соответствующий их количественный анализ. Как в случае индукции нейритогенеза и филоподий, авторы настоящего изобретения наблюдали, что мутеины значимо индуцируют образование новых синапсов в культуре нервных клеток. Данный эффект сходен с таковым, наблюдаемым для rhEPO.
Учитывая упомянутые выше результаты, существуют убедительные экспериментальные доказательства, которые показывают, что новые гипергликозилированные мутеины hEPO согласно настоящему изобретению оказывают способствующее пластичности нейронов действие на различных стадиях развития/дифференцировки нейронов (от образования нейритов до формирования синапса). Более того, данное действие сопоставимо с таковым, наблюдаемым для ЕРО, и, в некоторых конкретных случаях, данное действие (образование филоподий) даже более значительное. Данные неожиданные и новые технические эффекты и новаторские технические свойства настоящего изобретения делают мутеины согласно настоящему изобретению идеальными для применения для лечения состояний, при которых пластичность нейронов снижена или присутствует генетическая предрасположенность к такому снижению.
5.6.b. Исследование нейропротективной биологической активности мутеинов rhEPO в первичных культурах гиппокампальных нейронов крысы.
Оценка антиапоптического действия мутеинов hEPO на первичные культуры гиппокампальных нейронов позволила авторам настоящего изобретения исследовать действие, которое данные соединения оказывают на клетки, метаболизм которых не изменен, в отличие от устойчивых линий клеток. По этой причине, они представляют собой интересную модель, так как она более реалистично сходна с тем, что будет происходить in vivo в головном мозге.
Нейропротективную активность hEPO и модифицированных эритропоэтинов согласно настоящему изобретению оценивали как способность таких молекул защищать нервные клетки от апоптической стимуляции, вызванной обработкой стауроспорином.
Для того чтобы оценить такую активность in vitro получали первичные культуры гиппокампальных нейронов из крыс линии Sprague-Dawley. 11 DIV культуры предварительно обрабатывали в течение 24 ч 400 нг/мл мутеинов hEPO согласно настоящему изобретению или 400 нг/мл rhEPO, который обычно применяют для лечения для восстановления гематопоэза. По прошествии данного времени клетки приводили в контакт с 30 нМ STP в течение 24 ч в присутствии указанных молекул и, наконец, их фиксировали и окрашивали флуорохромом Хехст и фаллоидином-FITC.
На фигуре 18 показано, что обработка различными молекулами значимо снижала апоптоз (**р<0,01; ***р<0,001) по сравнению с контролем гибели клеток (клетками, обработанными только STP), достигая значений уровня апоптоза, сходных с таковыми для контрольных клеток без STP. Кроме того, для большинства молекул согласно настоящему изобретению выявили эффект, который превосходил таковой для rhEPO с точки зрения защиты первичных культур гиппокампальных нейронов от влияния STP.
Данные результаты также соответствуют таковым, полученным в исследовании нейропротективной активности мутеинов в отношении культур нервных клеток SH-SY5Y. Следовательно, добавление дополнительного сайта гликозилирования для получения молекул согласно настоящему изобретению позволило блокирование их гематопоэтической биологической активности без модификации активности, связанной с нейрозащитой.
СПИСОК ЛИТЕРАТУРЫ
[1] J.W. Fisher. Exp Biol Med (Maywood) 228 (2003) 1-14.
[2] С.Lacombe, P. Mayeux. Nephrol Dial Transplant 14 Suppl 2 (1999) 22-28.
[3] U. Hammerling, R. Kroon, T. Wilhelmsen, L. Sjodin, J. Pharm Biomed Anal 14 (1996) 1455-1469.
[4] H. Sasaki, B. Bothner, A. Dell, M. Fukuda. J Biol Chem 262 (1987) 12059-12076.
[5] E. Tsuda, M. Goto, A. Murakami, K. Akai, M. Ueda, G. Kawanishi, N. Takahashi, R. Sasaki, H. Chiba, H. Ishihara, et al. Biochemistry 27 (1988) 5646-5654.
[6] J.C. Egrie, J.K. Browne. Br J Cancer 84 Suppl 1 (2001) 3-10.
[7] A.J. Sytkowski, L. Feldman, D.J. Zurbuch, Biochem Biophys Res Commun 176 (1991) 698-704.
[8] E. Delorme, T. Lorenzini, J. Giffin, F. Martin, F. Jacobsen, T. Boone, S. Elliott. Biochemistry 31 (1992) 9871-9876.
[9] T. Toyoda, T. Arakawa, H. Yamaguchi. J Biochem 131 (2002) 511-515.
[10] Nangaku, M. Kidney International (2013) 84, 427-429.
[11] T Ng, G Marx, T Littlewood, I Macdougall. Postgrad Med J (2003) 79: 367-376
[12] N. Debeljak, A.J. Sytkowski, Erythropoietin: new approaches to improved molecular designs and therapeutic alternatives. Curr Pharm Des 14 (2008) 1302-1310.
[13] Noguchi, C.T., Asavaritikrai, P., Teng, R., and Jia, Y. (2007). Role of erythropoietin in the brain. Crit Rev Oncol Hematol 64, 159-171.
[14] P.a. Konstantinopoulos, M.V. Karamouzis, A.G. Papavassiliou. Biochim. Biophys. Acta 1776 (2007) 1-9.
[15] Siren, A.L. у Ehrenreich, H. Eur. Arch. Psychiatry Clin. Neurosci. (2001) 251(4): 179-184.
[16] Neurodegenerative disorders, Public health - European commission. http: //ec. europa. eu/health / maj or chronic diseases/ diseases/brain neurological/index en.htm
[17] Policy Brief: The Global Impact of Dementia 2013-2050-The Global voice on Dementia. Alzheimer's disease International. http: //www.alz.co.uk/research/G8-policy-brief.
[18] NIH meeting advances Alzheimer's research agenda. Fogarty International Center. Advancing Science for Global Health.
http: //www. fic. nih. gov/news/globalhealthmatters/march-april-2015 /pages/nih-alzheimers-research-summit. aspx
[19] Toledo Atucha, J. Alzheimer. Real Invest Demenc. (2011), 47: 16-23.
[20] Mawadda Alnaeeli, LiWang, Barbora Piknova, Heather Rogers, Xiaoxia Li, and Constance Tom Noguchi. Anatomy Research International. Volume 2012, Article ID 953264, 15 pages
[21] Masuda, S. у col. J. Biol. Chem. (1993) 268(15): 11208-11216.
[22] Wolf у col. Thromb Haemost (1997), 77:1020 1024.
[23] P.J. Stohlawetz, L. Dzirlo, N. Hergovich, E. Lackner, C. Mensik, H.G. Eichler, E. Kabrna, K. Geissler, B. Jilma. Blood 95 (2000) 2983-2989.
[24] Cases А у col. Kidney Internacional (1992), 42:668-672.
[25] Beguin Y., Haematologica (1999), 84:541-547.
[26] M. Leist, P. Ghezzi, G. Grasso, R. Bianchi, P. Villa, M. Fratelli, C. Savino, M. Bianchi, J. Nielsen, J. Gerwien, P. Kallunki, A.K. Larsen, L. Helboe, S. Christensen, L.O. Pedersen, M. Nielsen, L. Torup, T. Sager, A. Sfacteria, S. Erbayraktar, Z. Erbayraktar, N. Gokmen, O. Yilmaz, C. Cerami-Hand, Q.W. Xie, T. Coleman, A. Cerami, M. Brines. Science 305 (2004) 239-242.
[27] T. Mennini, M. De Paola, P. Bigini, C. Mastrotto, E. Fumagalli, S. Barbera, M. Mengozzi, B. Viviani, E. Corsini, M. Marinovich, L. Torup, J. Van Веек, M. Leist, M. Brines, A. Cerami, P. Ghezzi. Mol. Med. 12 (2006) 153-160.
[28] Siren, A.L. у col. Neurotherapeutics (2009), 6(1): 108-127.
[29] Erbayraktar, S. у col. Proc. Natl. Acad. Sci. (2003), USA 100: 6741-6746.
[30] M. Mattio, N. Ceaglio, M. Oggero, N. Perotti, I. Amadeo, G. Orozco, G. Forno, R. Kratje, M. Etcheverrigaray. Biotechnol Prog 27 (2011) 1018-1028.
[31] Dotti CG, Sullivan CA, Banker GA. 1988. J Neurosci (1988) 8:1454-1468
[32] Das KP, Freudenrich TM, Mundy WR. Neurotoxicol Teratol (2004) 26:397-406
[33] Rochefort NL, Konnerth A. EMBO Rep.(2012) Aug; 13 (8):699-708. DOI: 10.1038/embor.2012.102.
[34]Suetsugu M, Mehraein P. Acta Neuropathol. (1980) 50 (3): 207-10.
[35] Hutsler JJ1, Zhang H. Brain Res. (2010) Jan 14; 1309:83-94. DOI: 10.1016/j.brainres.2009.09.120.
[36] Alfonso J., Fernandez M. E., Cooper В., Flugge G. and Frasch A. C. Proc. Natl Acad. Sci. (2005) USA 102, 17196- 17201.
[37] Ippolito, D.M., Eroglu, C, Journal of Visualized Experiments. (2010) Exp.45:2270. DOI: 10.3791/2270.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Consejo Nacional de Investigaciones Cientyficas y Técnicas (CONICET)
Universidad Nacional de General San Martyn
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agaaacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaaagtga acttctacgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtggaagtg tggcagggac tggctctgct gagcgaggct 300
gtgctgagag gacaggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca aggctgtgtc cggcctgaga tccctgacca ccctgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcctgcg gggcaagctg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Asn Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 3
<211> 579
<212> ДНК
<213> ДНК мутеина 45_47
<400> 3
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agatacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaacgtga ccttctacgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtggaagtg tggcagggac tggctctgct gagcgaggct 300
gtgctgagag gacaggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca aggctgtgtc cggcctgaga tccctgacca ccctgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcctgcg gggcaagctg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
<210> 4
<211> 193
<212> Белок
<213> АК - мутеин 45_47
<400> 4
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Asn Val Thr Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 5
<211> 579
<212> ДНК
<213> ДНК мутеина 49
<400> 5
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agatacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaaagtga acttcaccgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtggaagtg tggcagggac tggctctgct gagcgaggct 300
gtgctgagag gacaggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca aggctgtgtc cggcctgaga tccctgacca ccctgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcctgcg gggcaagctg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
<210> 6
<211> 193
<212> Белок
<213> АК - мутеин 49
<400> 6
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Thr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 7
<211> 579
<212> ДНК
<213> ДНК - мутеин 62_64
<400> 7
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agatacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaaagtga acttctacgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtgaacgtg acgcagggac tggctctgct gagcgaggct 300
gtgctgagag gacaggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca aggctgtgtc cggcctgaga tccctgacca ccctgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcctgcg gggcaagctg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
<210> 8
<211> 193
<212> Белок
<213> АК - мутеин 62_64
<400> 8
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Asn Val Thr Gln Gly Leu Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 9
<211> 579
<212> ДНК
<213> ДНК - мутеин 65_67
<400> 9
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agatacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaaagtga acttctacgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtggaagtg tggaacggaa cggctctgct gagcgaggct 300
gtgctgagag gacaggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca aggctgtgtc cggcctgaga tccctgacca ccctgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcctgcg gggcaagctg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
<210> 10
<211> 193
<212> Белок
<213> АК - мутеин 65_67
<400> 10
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Asn Gly Thr Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 11
<211> 579
<212> ДНК
<213> ДНК - мутеин 72_74
<400> 11
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agatacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaaagtga acttctacgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtggaagtg tggcagggac tggctctgct gagcaacgct 300
acgctgagag gacaggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca aggctgtgtc cggcctgaga tccctgacca ccctgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcctgcg gggcaagctg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
<210> 12
<211> 193
<212> Белок
<213> АК - мутеин 72_74
<400> 12
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu
85 90 95
Leu Ser Asn Ala Thr Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 13
<211> 579
<212> ДНК
<213> ДНК - мутеин 76_78
<400> 13
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agatacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaaagtga acttctacgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtggaagtg tggcagggac tggctctgct gagcgaggct 300
gtgctgaacg gaacggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca aggctgtgtc cggcctgaga tccctgacca ccctgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcctgcg gggcaagctg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
<210> 14
<211> 193
<212> Белок
<213> АК - мутеин 76_78
<400> 14
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Asn Gly Thr Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 15
<211> 579
<212> ДНК
<213> ДНК - мутеин 98_100
<400> 15
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agatacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaaagtga acttctacgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtggaagtg tggcagggac tggctctgct gagcgaggct 300
gtgctgagag gacaggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca agaatgtgac cggcctgaga tccctgacca ccctgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcctgcg gggcaagctg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
<210> 16
<211> 193
<212> Белок
<213> АК - мутеин 98_100
<400> 16
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Asn Val Thr Gly
115 120 125
Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 17
<211> 579
<212> ДНК
<213> ДНК - мутеин 104
<400> 17
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agatacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaaagtga acttctacgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtggaagtg tggcagggac tggctctgct gagcgaggct 300
gtgctgagag gacaggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca aggctgtgtc cggcctgaga aacctgacca ccctgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcctgcg gggcaagctg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
<210> 18
<211> 193
<212> Белок
<213> АК - мутеин 104
<400> 18
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Asn Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 19
<211> 579
<212> ДНК
<213> ДНК - мутеин 106_108
<400> 19
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agatacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaaagtga acttctacgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtggaagtg tggcagggac tggctctgct gagcgaggct 300
gtgctgagag gacaggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca aggctgtgtc cggcctgaga tccctgaaca ccacgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcctgcg gggcaagctg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
<210> 20
<211> 193
<212> Белок
<213> АК - мутеин 106_108
<400> 20
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Ser Leu Asn Thr Thr Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 21
<211> 579
<212> ДНК
<213> ДНК - мутеин 149
<400> 21
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agatacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaaagtga acttctacgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtggaagtg tggcagggac tggctctgct gagcgaggct 300
gtgctgagag gacaggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca aggctgtgtc cggcctgaga tccctgacca ccctgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcacgcg gggcaagctg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
<210> 22
<211> 193
<212> Белок
<213> АК - мутеин 149
<400> 22
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Thr
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 23
<211> 579
<212> ДНК
<213> ДНК - мутеин 151_153
<400> 23
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agatacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaaagtga acttctacgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtggaagtg tggcagggac tggctctgct gagcgaggct 300
gtgctgagag gacaggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca aggctgtgtc cggcctgaga tccctgacca ccctgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcctgcg gaacaagacg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
<210> 24
<211> 193
<212> Белок
<213> АК - мутеин 151_153
<400> 24
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Asn Lys Thr Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<210> 25
<211> 579
<212> ДНК
<213> ДНК - hEPO
<400> 25
atgggcgtgc acgaatgtcc tgcttggctg tggctgctgc tgtccctgct gtctctgcct 60
ctgggactgc ctgtgctggg cgctcctcct agactgatct gcgactcccg ggtgctggaa 120
agatacctgc tggaagccaa agaggccgag aacatcacca ccggctgcgc cgagcactgc 180
tccctgaacg agaatatcac cgtgcccgac accaaagtga acttctacgc ctggaagcgg 240
atggaagtgg gccagcaggc tgtggaagtg tggcagggac tggctctgct gagcgaggct 300
gtgctgagag gacaggccct gctcgtgaac tcctcccagc cttgggaacc cctgcagctg 360
cacgtggaca aggctgtgtc cggcctgaga tccctgacca ccctgctgag agcactggga 420
gcccagaaag aggccatctc tccacctgac gccgcctctg ctgctcctct gagaaccatc 480
accgccgaca ccttcagaaa gctgttccgg gtgtactcca acttcctgcg gggcaagctg 540
aagctgtaca ccggcgaggc ttgccggacc ggcgacaga 579
<210> 26
<211> 193
<212> Белок
<213> АК - hEPO
<400> 26
Met Gly Val His Glu Cys Pro Ala Trp Leu Trp Leu Leu Leu Ser Leu
1 5 10 15
Leu Ser Leu Pro Leu Gly Leu Pro Val Leu Gly Ala Pro Pro Arg Leu
20 25 30
Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu Leu Glu Ala Lys Glu
35 40 45
Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His Cys Ser Leu Asn Glu
50 55 60
Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe Tyr Ala Trp Lys Arg
65 70 75 80
Met Glu Val Gly Gln Gln Ala Val Glu Val Trp Gln Gly Leu Ala Leu
85 90 95
Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu Leu Val Asn Ser Ser
100 105 110
Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp Lys Ala Val Ser Gly
115 120 125
Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu Gly Ala Gln Lys Glu
130 135 140
Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala Pro Leu Arg Thr Ile
145 150 155 160
Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val Tyr Ser Asn Phe Leu
165 170 175
Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala Cys Arg Thr Gly Asp
180 185 190
Arg
<---
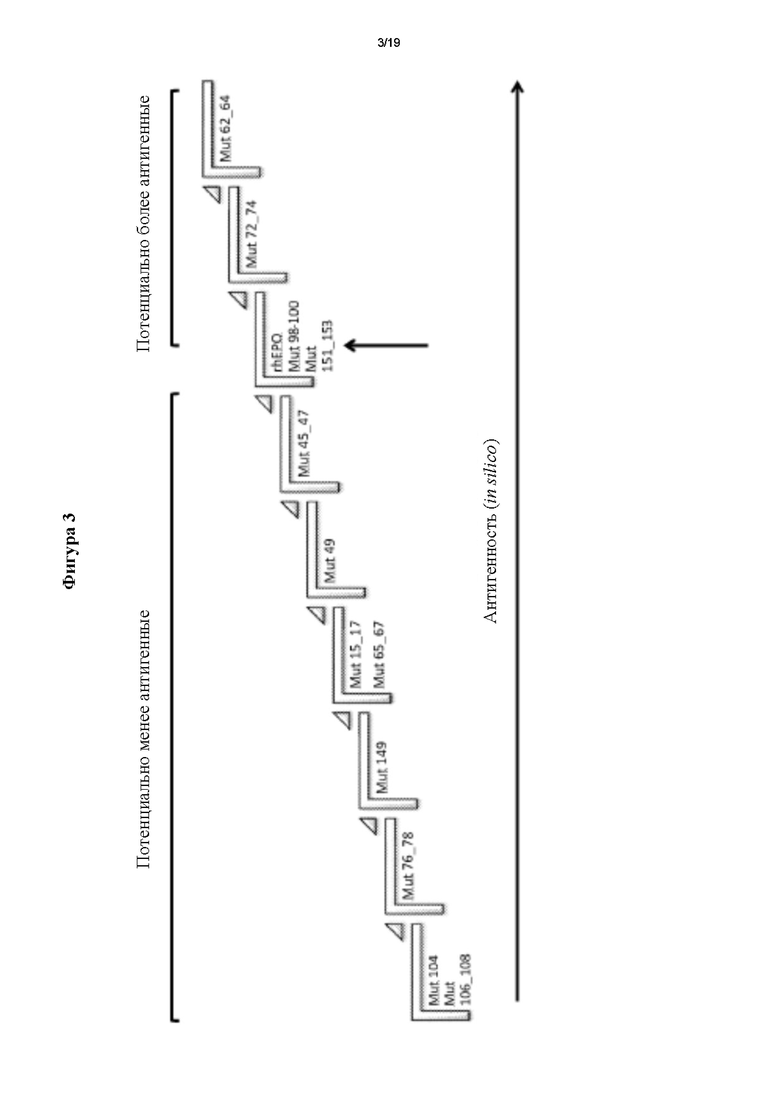
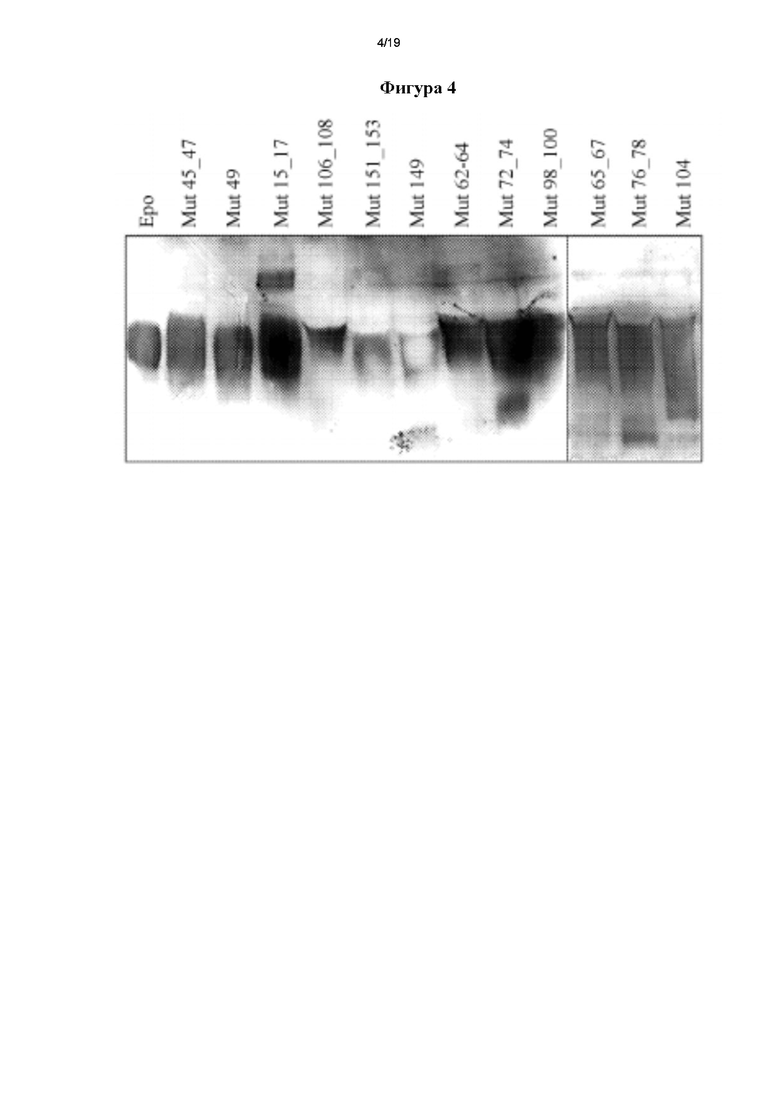
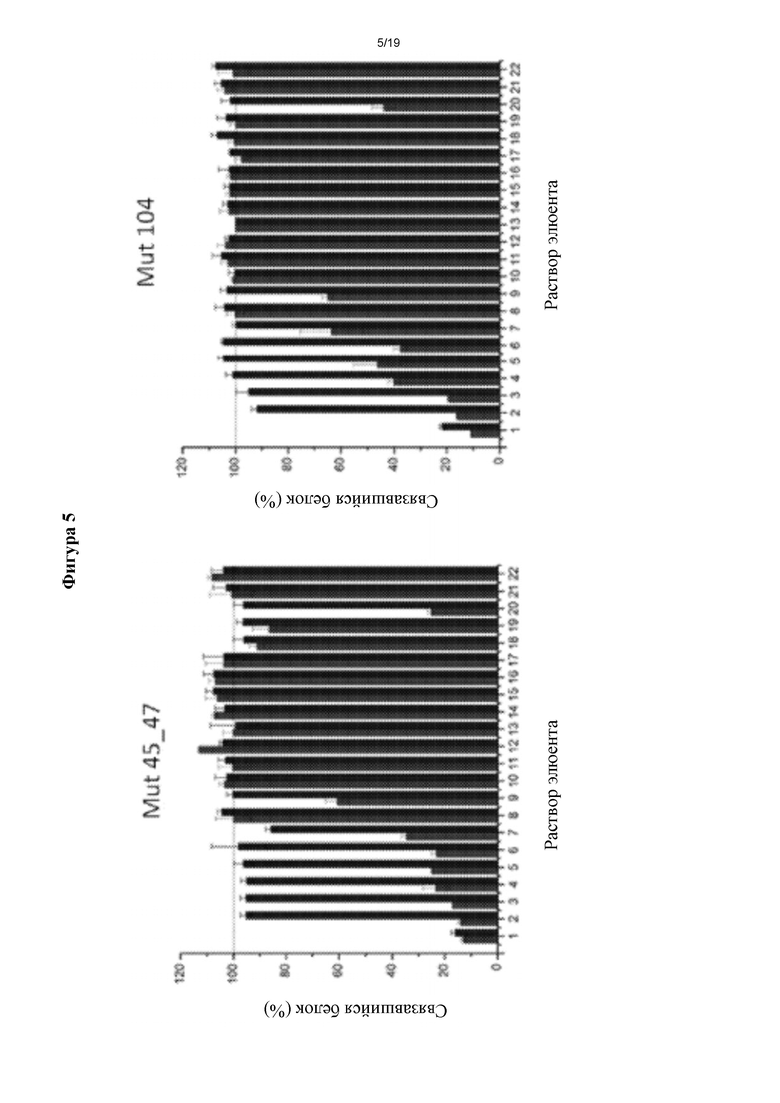
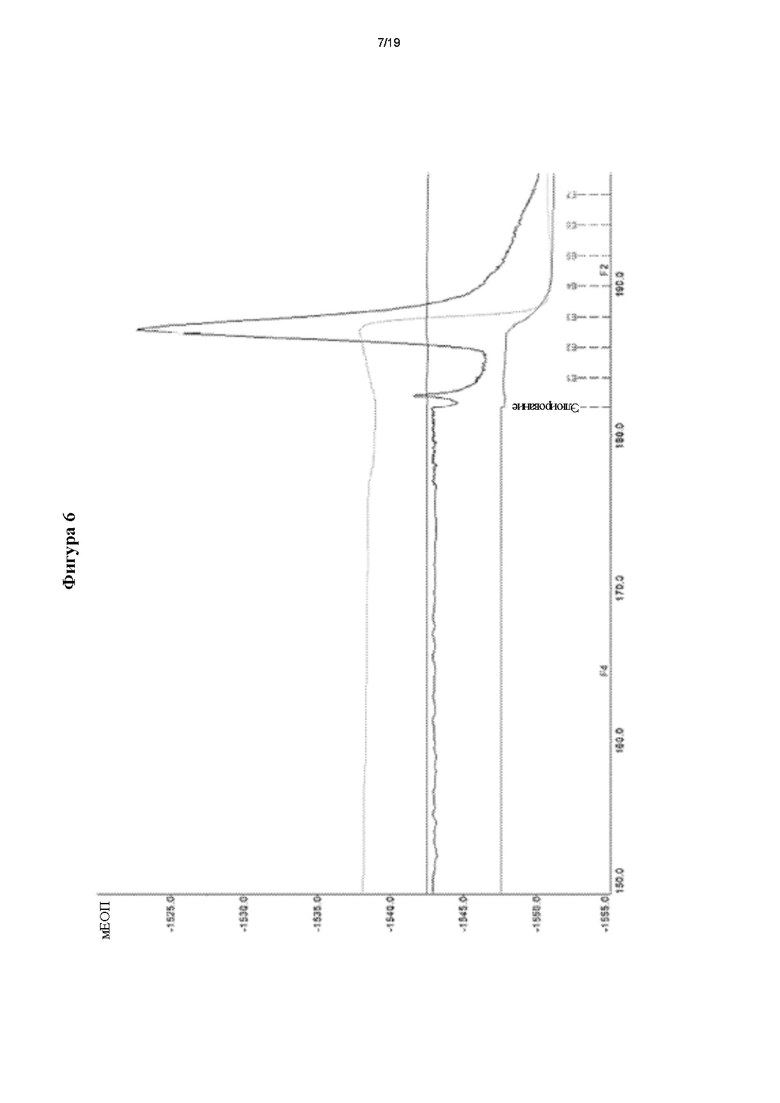
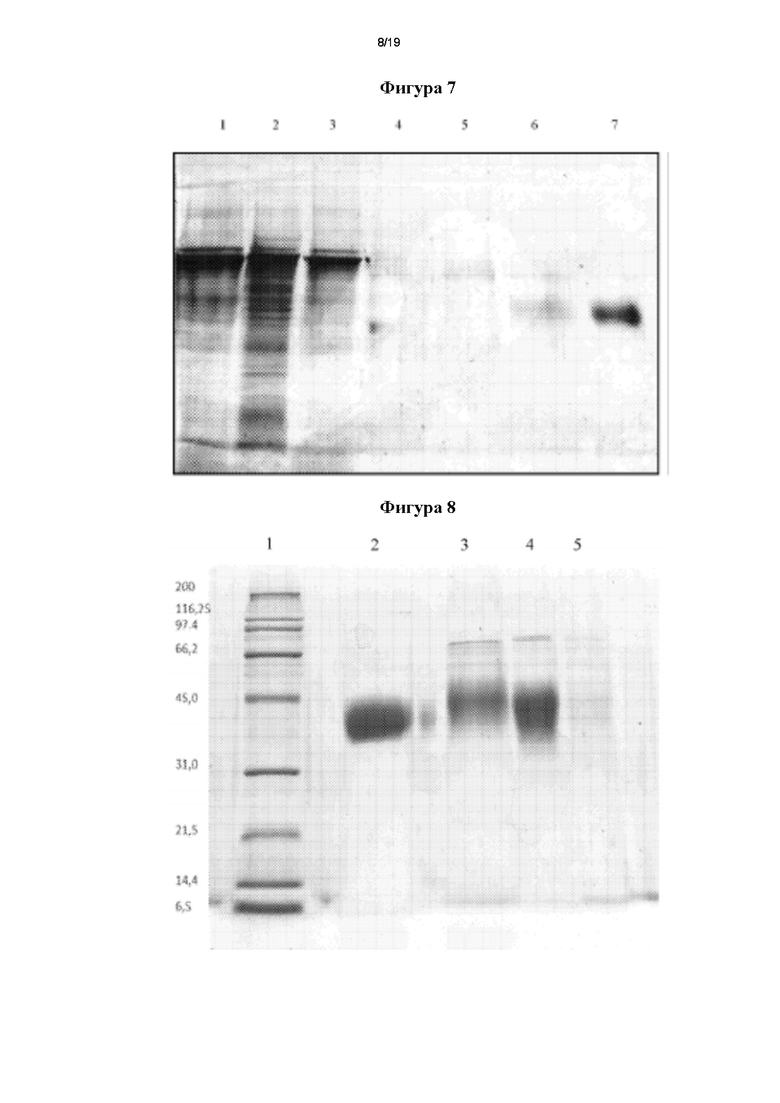
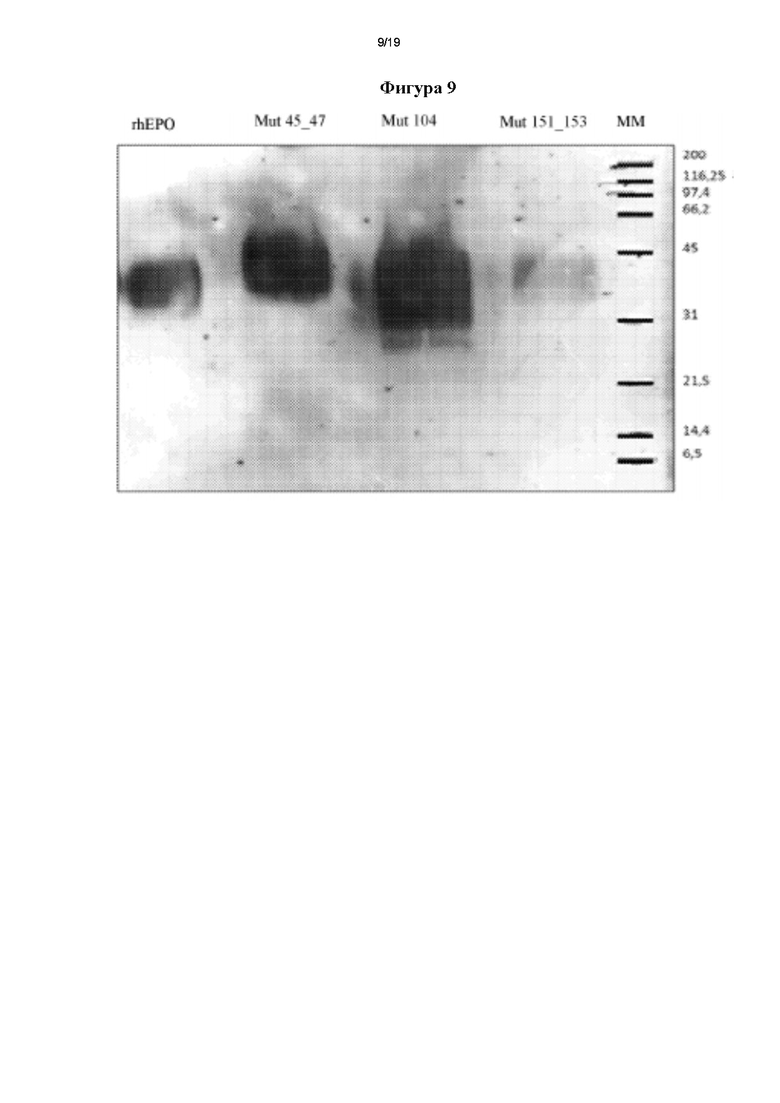
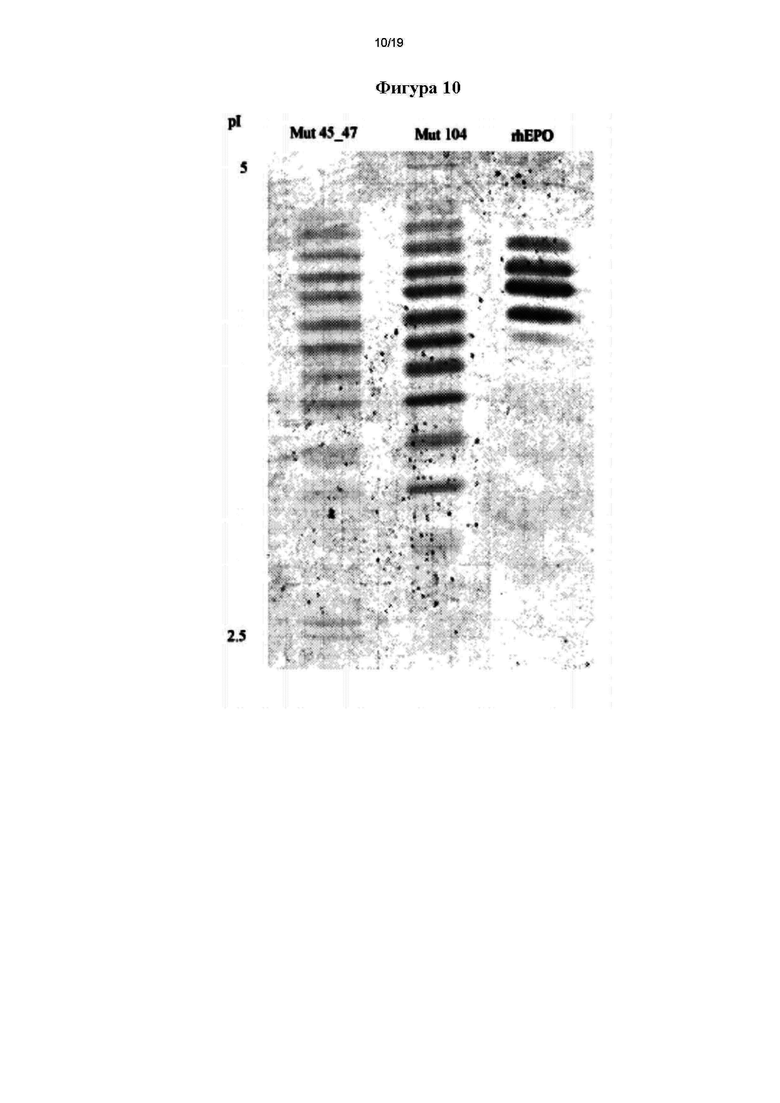
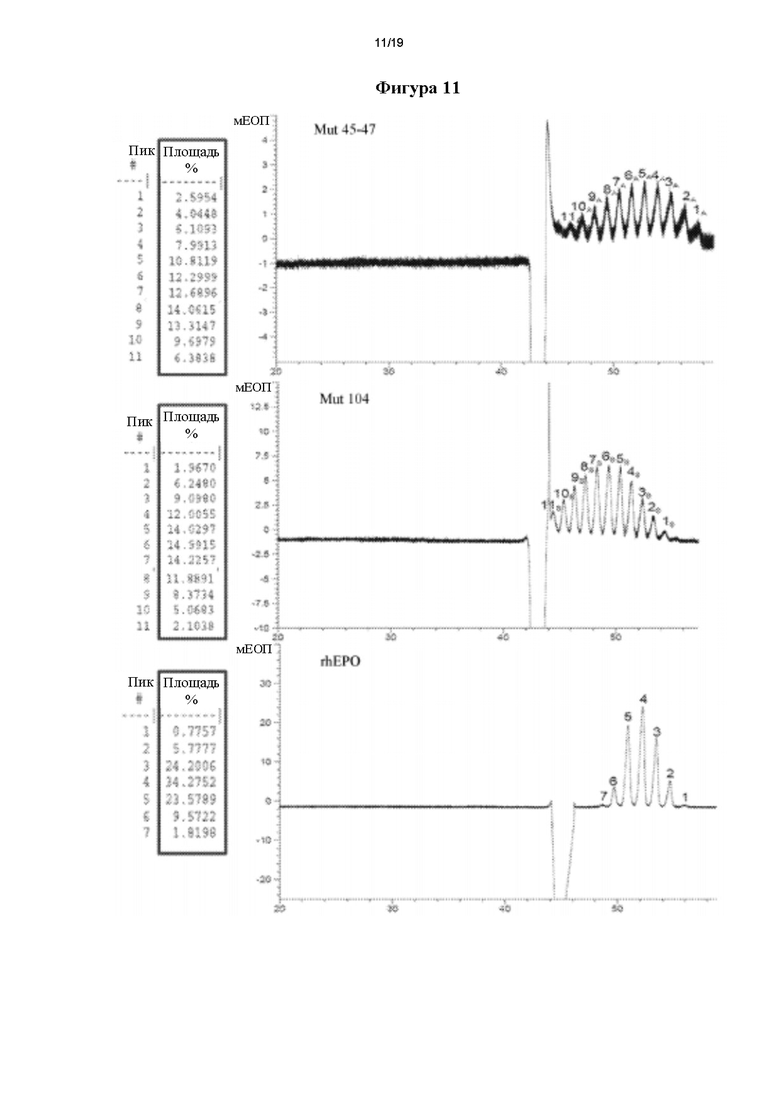
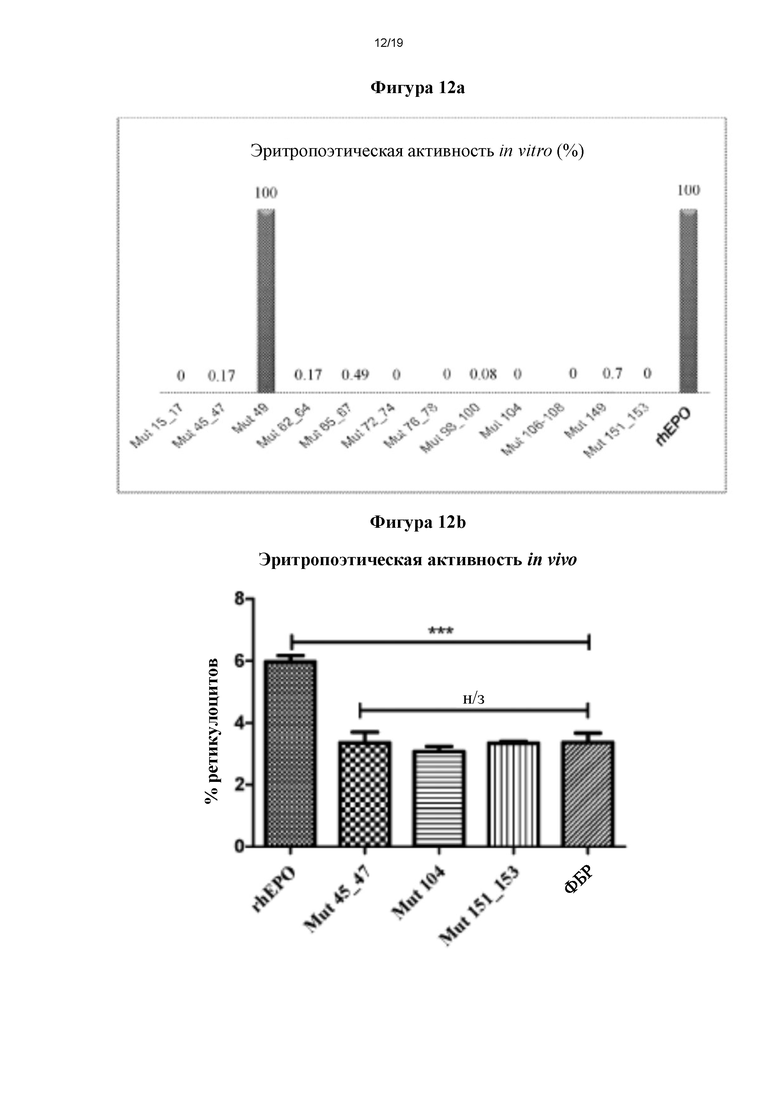
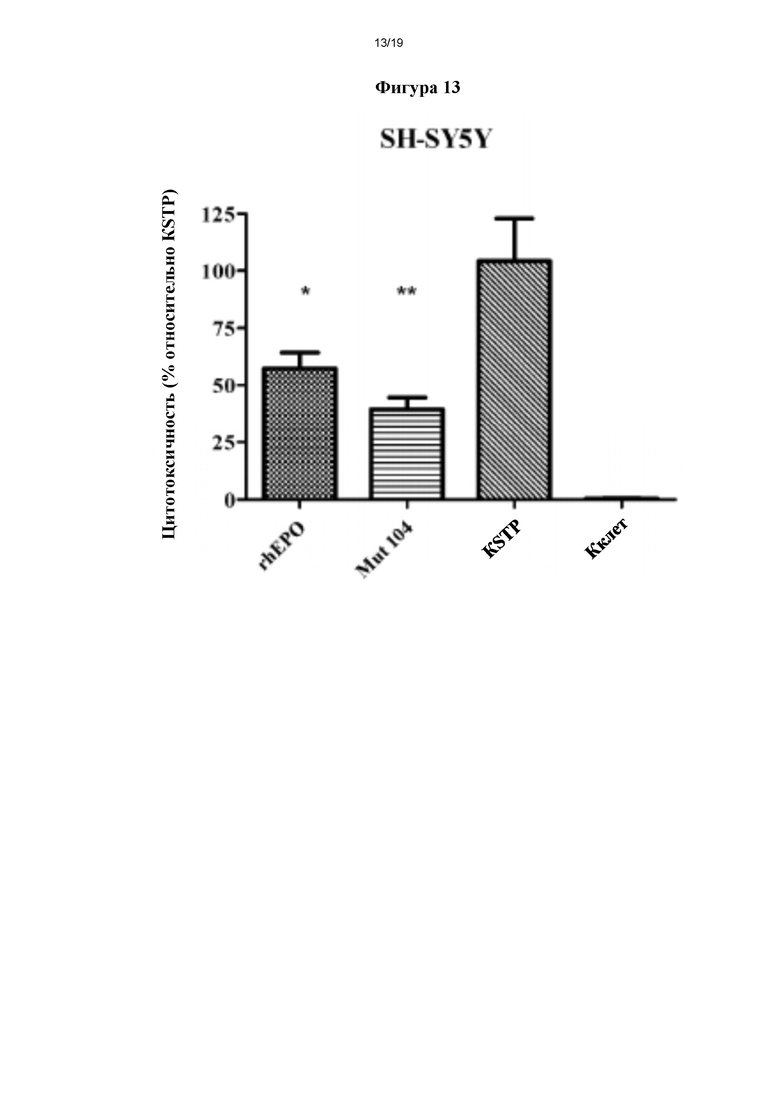
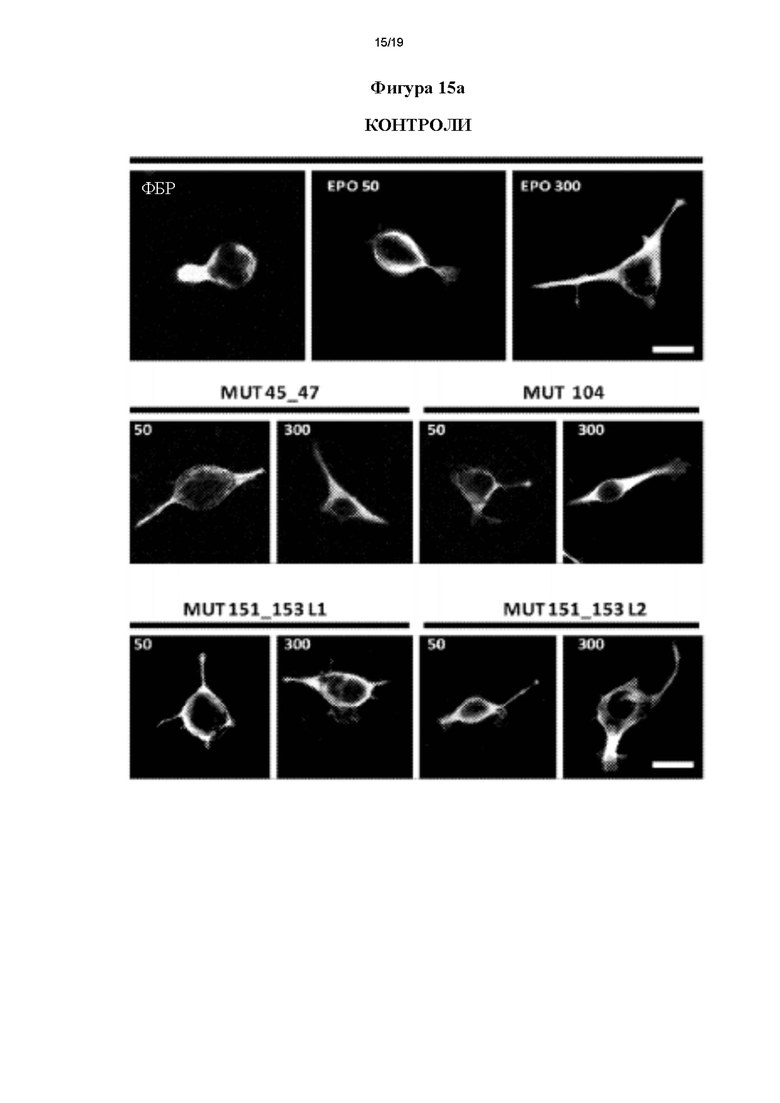
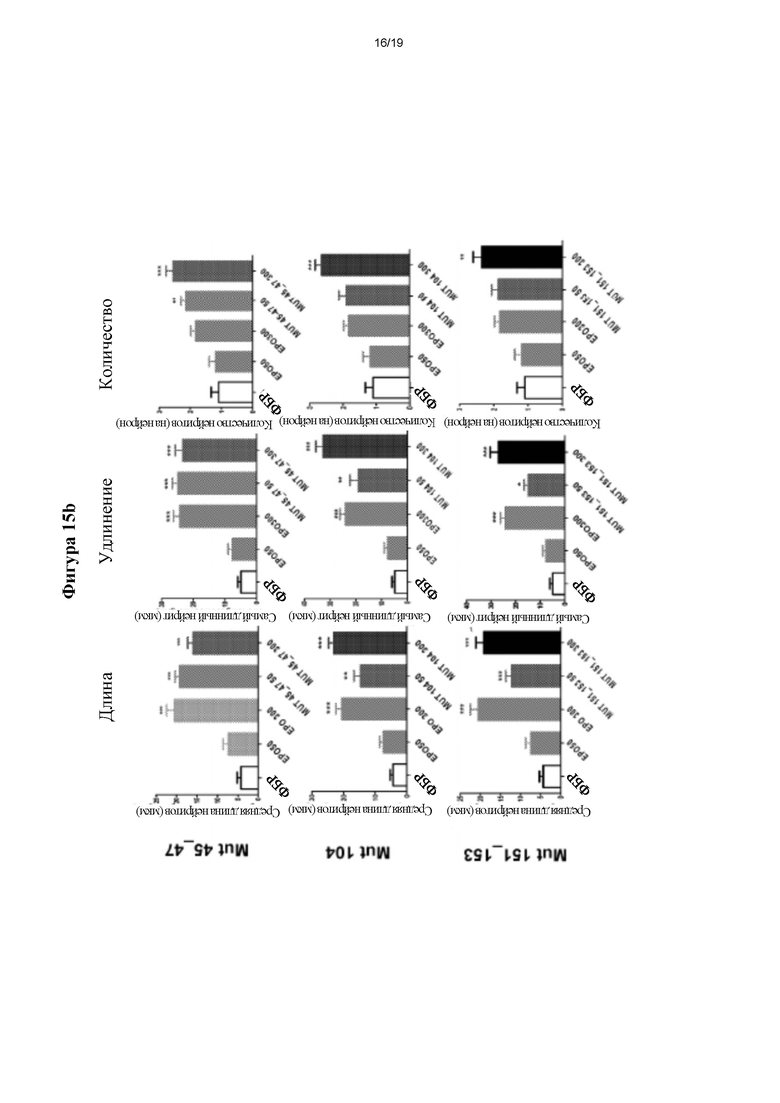
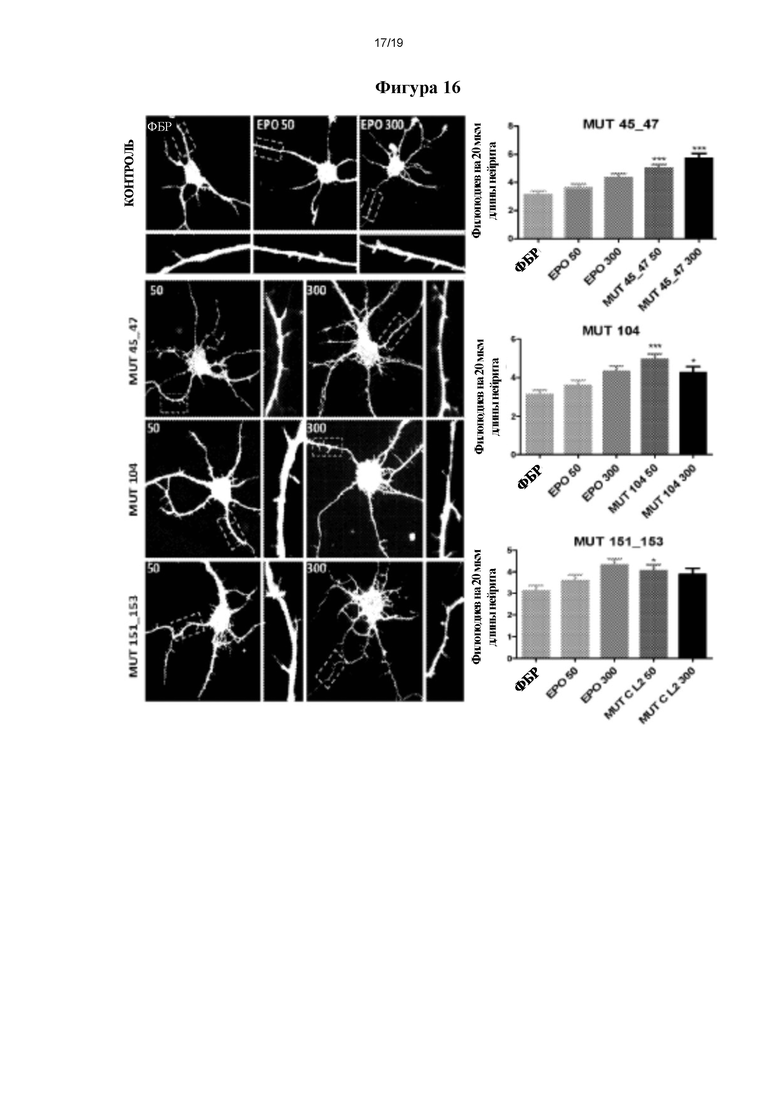
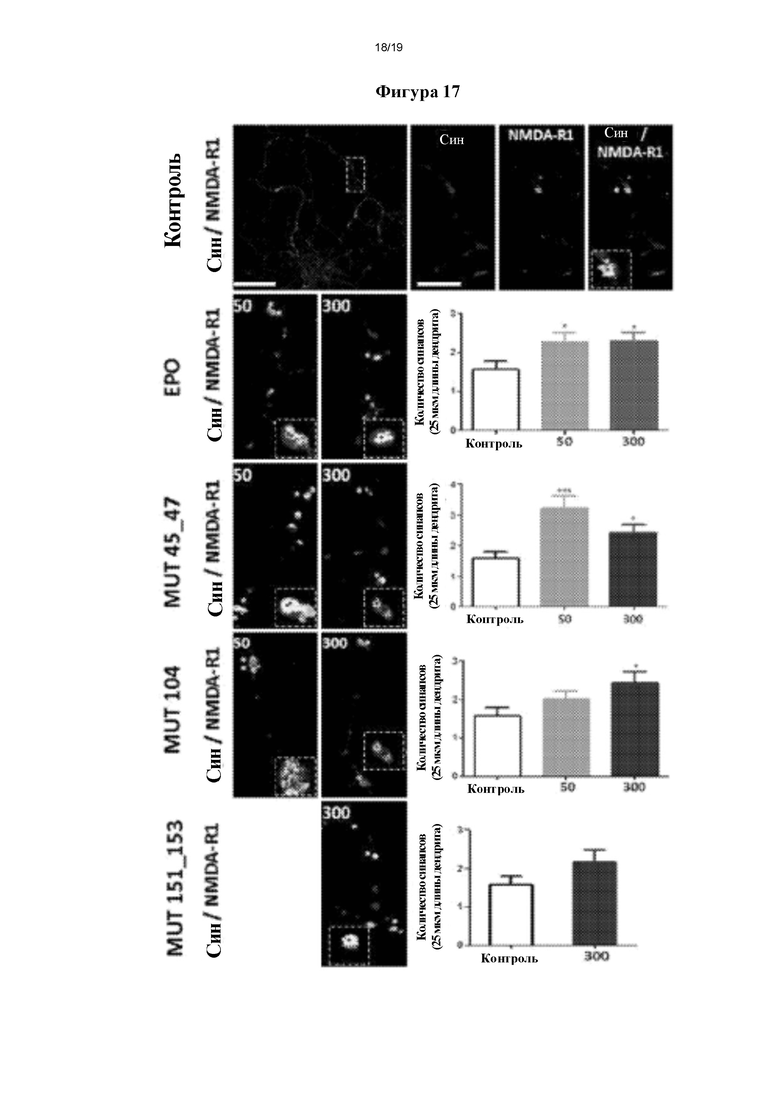
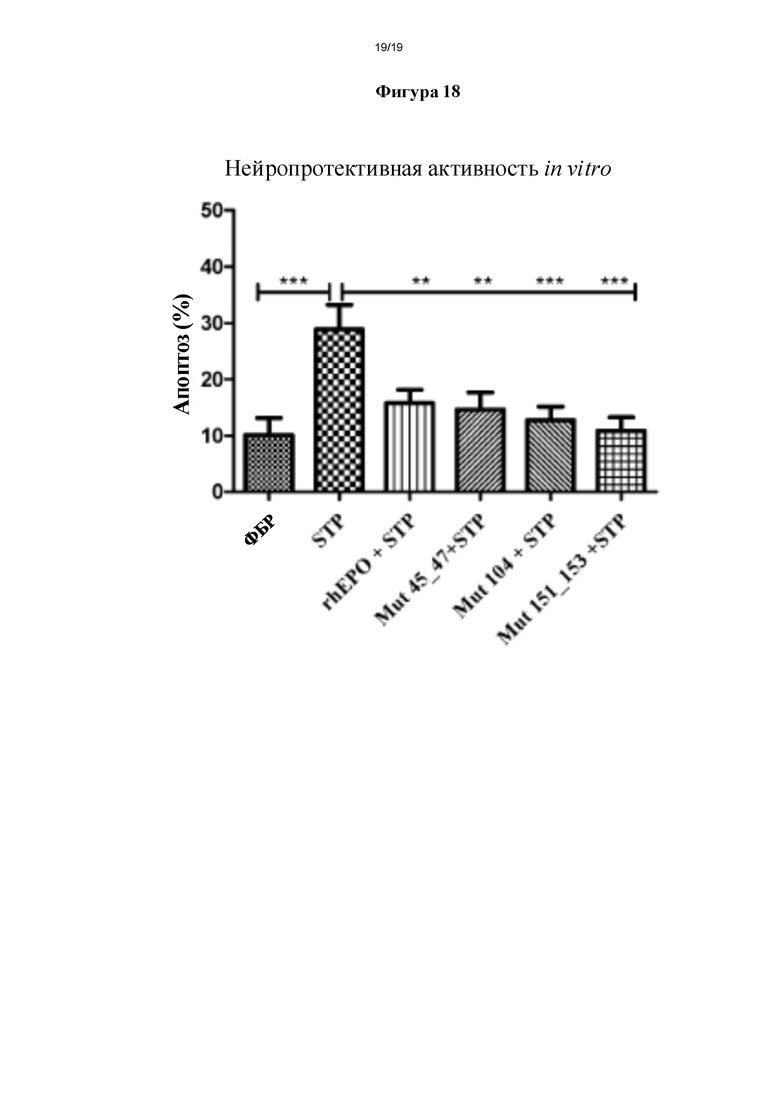
Изобретение относится к области биотехнологии, конкретно к модифицированному эритропоэтину человека (чЭПО), и может быть использовано в медицине для лечения таких патологий, как болезнь Альцгеймера, болезнь Паркинсона, боковой амиотрофический склероз (БАС), заболевания двигательных нейронов, болезнь Гентингтона и спиноцеребеллярные атрофии. Предложенный модифицированный чЭПО характеризуется содержанием мутации в сайте связывания с гомодимерным или гетеродимерным рецептором посредством добавления консенсусных сайтов гликозилирования. При этом мутация выбрана из следующих: (a) Lys45Asn и Asn47Thr, (b) Ser104Asn или (c) Gly151Asn и Leu153Thr; и модифицированный чЭПО содержит аминокислотную последовательность, выбранную из SEQ ID NO: 4, SEQ ID NO: 18 или SEQ ID NO: 24. Изобретение обеспечивает получение модифицированного чЭПО с эритропоэтической активностью менее 0,5% относительно нативного эритропоэтина, у которого сохраняется нейропротективная и нейропластическая способность. 4 н. и 1 з.п. ф-лы, 18 ил., 4 табл. 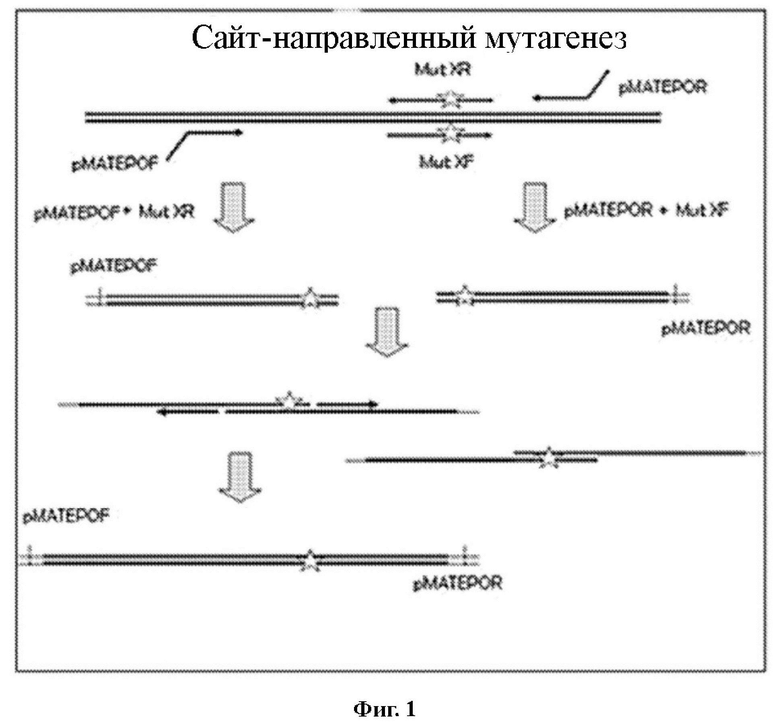
1. Модифицированный эритропоэтин человека с эритропоэтической активностью менее 0,5% относительно нативного эритропоэтина, у которого сохраняется его нейропротективная и нейропластическая способность, характеризующийся содержанием мутации в сайте связывания с гомодимерным или гетеродимерным рецептором посредством добавления консенсусных сайтов гликозилирования, причем такая мутация выбрана из совокупности, включающей:
a. Lys45Asn и Asn47Thr;
b. Ser104Asn;
c. Gly151Asn и Leu153Thr;
и содержащий аминокислотную последовательность, выбранную из совокупности, включающей: SEQ ID NO: 4, SEQ ID NO: 18 и SEQ ID NO: 24.
2. Модифицированный эритропоэтин человека по п. 1, отличающийся тем, что его кодирует последовательность ДНК, содержащая последовательность нуклеотидов, выбранную из совокупности, включающей: SEQ ID NO: 3, SEQ ID NO: 17 и SEQ ID NO: 23.
3. Фармацевтическая композиция, содержащая терапевтически эффективное количество модифицированного эритропоэтина человека по п. 1, для лечения патологий, выбранных из совокупности, включающей болезнь Альцгеймера, болезнь Паркинсона, боковой амиотрофический склероз (БАС), заболевания двигательных нейронов, болезнь Гентингтона, спиноцеребеллярные атрофии.
4. Применение фармацевтической композиции по п. 3 для лечения патологий, выбранных из совокупности, включающей болезнь Альцгеймера, болезнь Паркинсона, боковой амиотрофический склероз (БАС), заболевания двигательных нейронов, болезнь Гентингтона, спиноцеребеллярные атрофии.
5. Применение модифицированного эритропоэтина человека по п. 1 для лечения патологий, выбранных из совокупности, включающей болезнь Альцгеймера, болезнь Паркинсона, боковой амиотрофический склероз (БАС), заболевания двигательных нейронов, болезнь Гентингтона, спиноцеребеллярные атрофии.
| US 2012094906 А1, 19.04.2012 | |||
| US 20060035322 A1, 16.02.2006 | |||
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ЕДИНСТВЕННЫМ КОВАЛЕНТНО СВЯЗАННЫМ N-КОНЦЕВЫМ ВОДОРАСТВОРИМЫМ ПОЛИМЕРОМ | 1997 |
|
RU2199347C2 |
| ЛЕЧЕНИЕ НЕЙРОДЕГЕНЕРАТИВНЫХ РАССТРОЙСТВ | 2006 |
|
RU2467761C2 |
| СИНТЕТИЧЕСКИЕ ВАРИАНТЫ ГИПЕРГЛИКОЗИЛИРОВАННОГО ПРОТЕАЗО-РЕЗИСТЕНТНОГО ПОЛИПЕПТИДА, ПЕРОРАЛЬНЫЕ КОМПОЗИЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ ТАКИХ ВАРИАНТОВ | 2005 |
|
RU2392963C2 |
| WO 2004003176 А2, 08.01.2004 | |||
| ELLIOTT S | |||
| et al., Structural Requirements for Additional N-Linked Carbohydrate on Recombinant Human Erythropoietin, JOURNAL OF BIOLOGICAL CHEMISTRY, 2004, v | |||
| АППАРАТ ДЛЯ ОБОГАЩЕНИЯ РУД ПО МЕТОДУ ВСПЛЫВАНИЯ | 1915 |
|
SU279A1 |
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |

