Область техники
Настоящее изобретение относится к области конструирования генов и микроорганизмов, и, в частности, относится к рекомбинантному штамму для продуцирования (получения) L-глутаминовой кислоты, а также способу его конструирования и применения.
Уровень техники
L-глутаминовая кислота представляет собой важную аминокислоту, которую используют в пищевых продуктах, в клинической медицине и в других областях.
Традиционно L-глутаминовую кислоту получают в основном путем ферментации с использованием продуцирующих L-глутаминовую кислоту бактерий, принадлежащих к роду Brevibacterium, Corynebacterium или Microtatobiotes, или их мутантов.
L-Глутаминовая кислота синтезируется путем биосинтеза из α-кетоглутаровой кислоты, промежуточного продукта цикла лимонной кислоты в клетках микроорганизмов. Существует два биосинтетических пути образования L-глутаминовой кислоты из а-кетоглутаровой кислоты через ассимиляцию ионов аммония. Один путь представлен синтезом L-глутаминовой кислоты путем катализа глутаматдегидрогеназой (GDH) в присутствии ионов аммония в высокой концентрации. Другой путь (путь GS/GOGAT) представлен синтезом L-глутаминовой кислоты глутаминсинтетазой и аминотрансферазой глутамина-оксоглутаровой кислоты. Глутаминсинтетаза (GS) катализирует конверсию L-глутаминовой кислоты и ионов аммония в глутамин; аминотрансфераза глутамина-оксоглутаровой кислоты, также известная как «глутаминсинтетаза» (GOGAT), катализирует синтез L-глутаминовой кислоты, в ходе которого две молекулы L-глутаминовой кислоты синтезируются из одной молекулы глутамина, которая была синтезирована GS из одной молекулы α-кетоглутаровой кислоты.
Улучшение получения L-аминокислот путем ферментации может относиться к методикам ферментации, таким как перемешивание и подача кислорода; или к составу питательной среды, например, к концентрации сахара во время ферментации; или к переработке ферментативного бульона в продукт в подходящей форме, например, путем высушивания и грануляции ферментативного бульона, или с помощью ионообменной хроматографии; или может относиться к собственным свойствам соответствующих продуцирующих микроорганизмов.
Способы улучшения свойств указанных продуцирующих микроорганизмов включают мутагенез, выбор и скрининг мутантов. Штамм, полученный указанным способом, устойчив к антиметаболитам или ауксотрофен по метаболитам, имеющим регуляторное значение, и продуцирует L-аминокислоты, и т.п.
Хотя существует значительное число способов, способных усиливать способность к продуцированию L-глутаминовой кислоты, сохраняется потребность в разработке способов продуцирования L-глутаминовой кислоты для удовлетворения растущего спроса.
Краткое описание изобретения
Задача настоящего изобретения заключается в разработке новой методики для усиления способности бактерии к продуцированию L-глутаминовой кислоты, с обеспечением таким образом способа эффективного продуцирования L-глутаминовой кислоты.
В ходе исследований для решения вышеуказанной задачи авторы настоящего изобретения обнаружили, что ген BBD29_04920 или гомологичный ген у бактерии может быть модифицирован или улучшен в отношении экспрессии указанного гена, чтобы обеспечить усиленную способность к продуцированию L-глутаминовой кислоты. Настоящее изобретение было выполнено с учетом этих находок.
Согласно настоящему изобретению предложена бактерия для продуцирования L-глутаминовой кислоты, характеризующаяся улучшенной экспрессией полинуклеотида, кодирующего последовательность аминокислот SEQ ID NO: 3, или гомологичную ей последовательность. Согласно настоящему изобретению также предложен способ получения L-глутаминовой кислоты с использованием микроорганизма.
Согласно первому аспекту настоящего изобретения предложена бактерия для продуцирования L-глутаминовой кислоты, обладающая улучшенной экспрессией полинуклеотида, кодирующего последовательность аминокислот SEQ ID NO: 3 или гомологичную ей последовательность. В соответствии с настоящим изобретением улучшенная экспрессия представляет собой усиленную экспрессию полинуклеотида, или присутствие точечной мутации в полинуклеотиде, который кодирует последовательность аминокислот SEQ ID NO: 3 или гомологичную ей последовательность, или присутствие точечной мутации в полинуклеотиде и усиленную экспрессию полинуклеотида, который кодирует последовательность аминокислот SEQ ID NO: 3 или гомологичную ей последовательность.
Последовательность аминокислот SEQ ID NO: 3, или гомологичная ей последовательность представляет собой белок, кодируемый геном BBD29_04920 или гомологичным ему геном.
Указанная бактерия обладает усиленной способностью к продуцированию L-глутаминовой кислоты по сравнению с немодифицированным штаммом.
Согласно настоящему изобретению термин «бактерия, обладающая способностью к продуцированию L-глутаминовой кислоты» относится к бактерии, обладающей такой способностью к продуцированию и накоплению целевой L-глутаминовой кислоты в культуральной среде и/или в клетке указанной бактерии, что L-глутаминовая кислота может быть собрана при культивировании бактерии в культуральной среде. Бактерия, обладающая способностью к продуцированию L-глутаминовой кислоты, может представлять собой бактерию, способную к накоплению целевой L-глутаминовой кислоты в культуральной среде и/или в клетке указанной бактерии в большем количестве, чем у немодифицированного штамма.
Термин «немодифицированный штамм» относится к контрольному штамму, который не был модифицирован таким образом, чтобы включать некий специфический признак. Таким образом, примеры немодифицированного штамма включают штамм дикого типа и исходный штамм.
Бактерия, обладающая способностью к продуцированию L-глутаминовой кислоты, может представлять собой бактерию, способную к накоплению целевой L-глутаминовой кислоты в культуральной среде в количестве, предпочтительно превышающем 0,5 г/л, и более предпочтительно - превышающем 1,0 г/л.
Согласно настоящему изобретению, термин «L-глутаминовая кислота» относится к L-глутаминовой кислоте в свободной форме, к ее соли или смеси, если не указано иное.
Указанный полинуклеотид может кодировать последовательность аминокислот, обладающую приблизительно 90% или более, приблизительно 92% или более, приблизительно 95% или более, приблизительно 97% или более, приблизительно 98% или более, или приблизительно 99% или более гомологией последовательности в отношении последовательности аминокислот SEQ ID NO: 3. В настоящем документе термин «гомология» относится к проценту идентичности двух полинуклеотидов или двух полипептидов как модулей. Гомология последовательностей может быть измерена между одним модулем и другим модулем с применением способа, известного в данной области техники. Например, такая гомология последовательностей может быть измерена с помощью алгоритма BLAST.
Экспрессия полинуклеотида может быть усилена: путем замены или мутирования регуляторной последовательности экспрессии, введения мутации в последовательность полинуклеотида, и путем увеличения числа копий полинуклеотида, инсертированных в хромосому или введенных с помощью вектора, или комбинации перечисленного, и т.п.
Регуляторная последовательность экспрессии полинуклеотида может быть модифицирована. Регуляторная последовательность экспрессии контролирует экспрессию полинуклеотида, который функционально с ней связан, и может содержать, например, промоторы, терминаторы, энхансеры, сайленсеры и т.п. Полинуклеотид может иметь измененный инициирующий кодон. Полинуклеотид может быть включен в определенный сайт хромосомы, с увеличением таким образом числа копий. Согласно настоящему документу определенный сайт может включать, например, сайт транспозона или межгенный сайт. Кроме того, полинуклеотид может быть включен в экспрессионный вектор, а указанный экспрессионный вектор введен в клетку-хозяина, с увеличением таким образом числа копий.
Согласно одному из вариантов реализации настоящего изобретения полинуклеотид или имеющий точечную мутацию полинуклеотид включают в определенный сайт хромосомы микроорганизма с увеличением таким образом числа копий.
Согласно одному из вариантов реализации настоящего изобретения полинуклеотид с последовательностью промотора, или имеющий точечную мутацию полинуклеотид с последовательностью промотора встраивают в определенный сайт хромосомы микроорганизма, со сверхэкспрессией таким образом последовательности нуклеиновой кислоты.
Согласно одному из вариантов реализации настоящего изобретения полинуклеотид или имеющий точечную мутацию полинуклеотид включают в экспрессионный вектор, а указанный экспрессионный вектор вводят в клетку-хозяина, с увеличением таким образом числа копий.
Согласно одному из вариантов реализации настоящего изобретения полинуклеотид с последовательностью промотора, или имеющий точечную мутацию полинуклеотид с последовательностью промотора включают в экспрессионный вектор, а указанный экспрессионный вектор вводят в клетку-хозяина, с обеспечением таким образом сверхэкспрессии последовательности аминокислот.
Согласно конкретному варианту реализации настоящего изобретения указанный полинуклеотид может содержать последовательность нуклеотидов SEQ ID NO: 1.
Согласно одному из вариантов реализации настоящего изобретения полинуклеотид, кодирующий последовательность аминокислот SEQ ID NO: 3, имеет точечную мутацию, такую, что аспарагин в положении 520 в последовательности аминокислот SEQ ID NO: 3 заменен на другую аминокислоту.
В соответствии с настоящим изобретением предпочтительно аспарагин в положении 520 заменен на лизин.
В соответствии с настоящим изобретением последовательность, представленная в SEQ ID NO: 4, представляет собой последовательность аминокислот после замены аспарагина на лизин в положении 520 в последовательности аминокислот, представленной в SEQ ID NO: 3.
Согласно одному из вариантов реализации настоящего изобретения последовательность полинуклеотида, имеющего точечную мутацию, образуется в результате мутации основания в положении 1560 последовательности полинуклеотида, представленной в SEQ ID NO: 1.
В соответствии с настоящим изобретением указанная мутация включает мутацию основания в положении 1560 последовательности полинуклеотида, представленной в SEQ ID NO: 1, с заменой цитозина (С) на аденин (А).
Согласно одному из вариантов реализации настоящего изобретения последовательность полинуклеотида, имеющего точечную мутацию, содержит последовательность полинуклеотида, представленную в SEQ ID NO: 2.
В настоящем документе термин «функционально связанный» относится к функциональной связи между регуляторной последовательностью и последовательностью полинуклеотида, за счет которой регуляторная последовательность контролирует транскрипцию и/или трансляцию указанной последовательности полинуклеотида. Регуляторная последовательность может представлять собой сильный промотор, который может повышать уровень экспрессии полинуклеотида. Регуляторная последовательность может представлять собой промотор, происходящий из микроорганизма, принадлежащего к роду Corynebacterium, или может представлять собой промотор, происходящий из других микроорганизмов. Например, указанный промотор может представлять собой промотор trc, промотор gap, промотор tac, промотор Т7, промотор lac, промотор trp, промотор araBAD или промотор cj7.
Согласно конкретному варианту реализации настоящего изобретения указанный промотор представляет собой промотор полинуклеотида, кодирующего последовательность аминокислот SEQ ID NO: 3 (гена BBD29_04920).
В настоящем документе термин «вектор» относится к полинуклеотидной конструкции, содержащей регуляторную последовательность гена и последовательность гена, и выполненной с возможностью экспрессии целевого гена в подходящей клетке-хозяине. Термин "вектор" может также относиться к полинуклеотидной конструкции, содержащей последовательность для гомологичной рекомбинации, такой, что при введении этого вектора в клетку-хозяина может быть изменена регуляторная последовательность эндогенного гена в геноме клетки-хозяина, или целевой ген, который может экспрессироваться, может быть инсертирован в конкретный сайт генома хозяина. Таким образом, вектор для применения по настоящему изобретению может дополнительно содержать селективный маркер для определения введения вектора в клетку-хозяина или инсерции вектора в хромосому клетки-хозяина. Селективный маркер может включать маркер, обеспечивающий селектируемый фенотип, такой как лекарственная устойчивость, ауксотрофность, устойчивость к цитотоксическим агентам или экспрессия поверхностного белка. На среде, обработанной таким селективным агентом, может быть выбрана трансформированная клетка, поскольку только клетка, которая экспрессирует селективный маркер, может выживать или проявлять иные фенотипические черты. Вектор согласно описанию в настоящем документе хорошо известен специалистам в данной области техники и включает в том числе, но не ограничиваясь перечисленным: плазмиду, бактериофаг (такой как λ бактериофаг или нитевидный бактериофаг М13, и т.п.), космиду или вирусный вектор.
Согласно некоторым конкретным вариантам реализации настоящего изобретения в качестве вектора используют плазмиду pK18mobsacB и плазмиду pXMJ19.
В настоящем документе термин «трансформация» относится к введению полинуклеотида в клетку-хозяина таким образом, чтобы указанный полинуклеотид мог быть репликативно-компетентным в качестве элемента, чужеродного для генома, или за счет встраивания в геном клетки-хозяина. Способ трансформации вектором, применяемый согласно настоящему изобретению, может включать способ введения нуклеиновой кислоты в клетку. Кроме того, в соответствии с описанием близких методик, может быть использован подходящий для клетки-хозяина способ, задействующий электрические импульсы.
Согласно настоящему изобретению микроорганизм может представлять собой дрожжи, бактерию, водоросль или гриб.
В соответствии с настоящим изобретением указанная бактерия может представлять собой микроорганизм, принадлежащий к роду Corynebacterium, такой как Corynebacterium acetoacidophilum, Corynebacterium acetoglutamicum, Corynebacterium callunae, Corynebacterium glutamicum, Brevibacterium flavum, Brevibacterium lactofermentum, Corynebacterium ammoniagenes, Corynebacterium pekinense, Brevibacterium saccharolyticum, Brevibacterium roseum и Brevibacterium thiogenitalis и т.п.
Согласно одному из вариантов реализации настоящего изобретения указанный микроорганизм, принадлежащий к роду Corynebacterium, представляет собой Corynebacterium glutamicum АТСС 13869.
Согласно одному из вариантов реализации настоящего изобретения указанный микроорганизм, принадлежащий к роду Corynebacterium, представляет собой Corynebacterium glutamicum YPGLU001, который обеспечивает высокие уровни продуцирования глутаминовой кислоты, и депонирован как штамм с названием: Corynebacterium glutamicum; латинское название: Corynebacterium glutamicum; штамм №: YPGLU001 в Китайском главном центре коллекций микробиологических культур, сокращенно CGMCC, адрес: No. 3, Yard 1, Beichen West Road, Chaoyang District, Beijing, 23 ноября 2020 г., под номером доступа: CGMCC №21220.
В соответствии с настоящим изобретением указанная бактерия может также иметь дополнительные улучшения, связанные с повышением уровней продуцирования L-глутаминовой кислоты, такие как усиление или снижение активности, или экспрессии генов глутаматдегидрогеназы, глутаминсинтетазы, аминотрансферазы глутамина-оксоглутаровой кислоты и т.п., или замена генов чужеродными генами.
Согласно второму аспекту настоящего изобретения предложены последовательность полинуклеотида, последовательность аминокислот, кодируемая указанной последовательностью полинуклеотида, рекомбинантный вектор, содержащий указанную последовательность полинуклеотида, и рекомбинантный штамм, содержащий указанную последовательность полинуклеотида.
В соответствии с настоящим изобретением указанная последовательность полинуклеотида включает полинуклеотид, кодирующий полипептид, содержащий последовательность аминокислот SEQ ID NO: 3, где аспарагин в положении 520 в последовательности заменен на другую аминокислоту.
В соответствии с настоящим изобретением предпочтительно аспарагин в положении 520 заменен на лизин.
В соответствии с настоящим изобретением последовательность, представленная в SEQ ID NO: 4, представляет собой последовательность аминокислот после замены аспарагина на лизин в положении 520 в последовательности аминокислот, представленной в SEQ ID NO: 3.
В соответствии с настоящим изобретением предпочтительно указанный полинуклеотид, кодирующий полипептид, содержащий последовательность аминокислот SEQ ID NO: 3, содержит последовательность полинуклеотида согласно представленной в SEQ ID NO: 1.
Согласно одному из вариантов реализации настоящего изобретения указанная последовательность полинуклеотида образуется в результате мутации основания в положении 1560 в последовательности полинуклеотида, представленной в SEQ ID NO: 1.
В соответствии с настоящим изобретением указанная мутация относится к изменению основания/нуклеотида в сайте, и способ осуществления мутации может представлять собой по меньшей мере один способ, выбранный из мутагенеза, сайт-направленного ПЦР-мутагенеза и/или гомологичной рекомбинации, и т.п. Согласно настоящему изобретению, предпочтительно используют сайт-направленный ПЦР-мутагенез и/или гомологичную рекомбинацию.
В соответствии с настоящим изобретением указанная мутация включает мутацию в основании в положении 1560 в последовательности полинуклеотида, представленной в SEQ ID NO: 1, с заменой цитозина (С) на аденин (А).
Согласно одному из вариантов реализации настоящего изобретения указанная последовательность полинуклеотида содержит последовательность полинуклеотида SEQ ID NO: 2.
В соответствии с настоящим изобретением указанная последовательность аминокислот содержит последовательность аминокислот SEQ ID NO: 4.
В соответствии с настоящим изобретением указанный рекомбинантный вектор конструируют путем введения последовательности полинуклеотида в плазмиду.
Согласно одному из вариантов реализации настоящего изобретения указанная плазмида представляет собой плазмиду pK18mobsacB.
Согласно другому варианту реализации настоящего изобретения указанная плазмида представляет собой плазмиду pXMJ19.
В частности, указанные последовательность полинуклеотида и плазмида могут быть сконструированы в виде рекомбинантного вектора с помощью системы рекомбинации NEBuider.
В соответствии с настоящим изобретением указанный рекомбинантный штамм содержит указанную последовательность полинуклеотида.
Во варианте реализации настоящего изобретения исходный штамм для рекомбинантного штамма представляет собой Corynebacterium glutamicum CGMCC №21220.
Во варианте реализации настоящего изобретения исходный штамм для рекомбинантного штамма представляет собой АТСС 13869.
Согласно третьему аспекту настоящего изобретения также предложен способ конструирования рекомбинантного штамма, продуцирующего L-глутаминовую кислоту.
В соответствии с настоящим изобретением указанный способ конструирования включает следующий этап:
конструирование последовательности полинуклеотида гена BBD29_04920 дикого типа, представленной в SEQ ID NO: 1, у штамма-хозяина, чтобы вызвать мутацию основания в положении 1560, с получением рекомбинантного штамма, содержащего кодирующий мутированный BBD29_04920 ген.
В соответствии со способом конструирования согласно настоящему изобретению указанное конструирование включает по меньшей мере что-либо одно из мутагенеза, сайт-направленного ПЦР-мутагенеза и/или гомологичной рекомбинации и т.п.
В соответствии со способом конструирования согласно настоящему изобретению мутация относится к изменению основания в положении 1560 в SEQ ID NO: 1 с заменой цитозина (С) на аденин (А). В частности, последовательность полинуклеотида, содержащая кодирующий мутированный BBD29_04920 ген, представлена в SEQ ID NO: 2.
Кроме того, указанный способ конструирования включает следующие этапы:
(1) конструирование последовательности нуклеотидов гена BBD29_04920 дикого типа, представленной в SEQ ID NO: 1, чтобы вызвать мутацию основания в положении 1560, с получением последовательности полинуклеотида мутированного гена BBD29_04920;
(2) соединение мутированной последовательности полинуклеотида с плазмидой для конструирования рекомбинантного вектора;
(3) введение указанного рекомбинантного вектора в штамм-хозяин с получением рекомбинантного штамма, содержащего кодирующий мутированный BBD29_04920 ген.
В соответствии со способом конструирования согласно настоящему изобретению указанный этап (1) включает конструирование гена BBD29_04920 с точечной мутацией: в соответствии с геномной последовательностью немодифицированного штамма, синтез двух пар праймеров P1, Р2 и Р3, Р4 для амплификации фрагмента гена BBD29_04920, и введение точечной мутации в ген BBD29_04920 дикого типа, SEQ ID NO: 1, с применением сайт-направленного ПЦР-мутагенеза, с получением последовательности нуклеотидов гена BBD29_04920 с точечной мутацией, SEQ ID NO: 2, обозначенного как BBD29_04920C1560A.
Согласно одному из вариантов реализации настоящего изобретения геном немодифицированного штамма может происходить из штамма АТСС13869, последовательность генома которого можно найти на веб-сайте NCBI.
Согласно одному из вариантов реализации настоящего изобретения на этапе (1) используют следующие праймеры:
Р1: 5' CAGTGCCAAGCTTGCATGCCTGCAGGTCGACTCTAG TGTTTCTGTC TTGACCTTGG (SEQ ID NO: 5)
Р2: 5' AGCCACGATG GTGACTTTTT GCAAGTTGTT_3' (SEQ ID NO: 6)
Р3: 5' AACAACTTGC AAAAAGTCAC CATCGTGGCT 3' (SEQ ID NO: 7)
Р4: 5' CAGCTATGACCATGATTACGAATTCGAGCTCGGTACCC ACAGATTGGG CAGGTGCC 3' (SEQ ID NO: 8)
Согласно одному из вариантов реализации настоящего изобретения ПЦР-амплификацию осуществляют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 с при 94°С, ренатурация в течение 30 с при 52°С и удлинение в течение 40 с при 72°С в ходе 30 циклов, и избыточное удлинение в течение 10 мин при 72°С.
Согласно одному из вариантов реализации настоящего изобретения ПЦР-амплификацию с перекрывающимися праймерами осуществляют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 с при 94°С, ренатурация в течение 30 с при 52°С и удлинение в течение 90 с при 72°С в ходе 30 циклов, и избыточное удлинение в течение 10 мин при 72°С.
В соответствии со способом конструирования согласно настоящему изобретению этап (2) включает конструирование рекомбинантной плазмиды, включающей сборку выделенного и очищенного BBD29_04920C1560A с плазмидой pK18mobsacB с помощью системы рекомбинации NEBuider, с получением рекомбинантной плазмиды.
В соответствии со способом конструирования согласно настоящему изобретению указанный этап (3) включает конструирование рекомбинантного штамма путем трансформации рекомбинантной плазмидой штамма-хозяина с получением рекомбинантного штамма.
Согласно одному из вариантов реализации настоящего изобретения трансформацию на этапе (3) осуществляют методом электротрансформации.
Согласно одному из вариантов реализации настоящего изобретения штамм-хозяин представляет собой АТСС 13869.
Согласно одному из вариантов реализации настоящего изобретения штамм-хозяин представляет собой Corynebacterium glutamicum CGMCC №21220.
Согласно одному из вариантов реализации настоящего изобретения указанная рекомбинация достигается путем гомологичной рекомбинации.
Согласно четвертому аспекту настоящего изобретения также предложен способ конструирования рекомбинантного штамма, продуцирующего L-глутаминовую кислоту.
В соответствии с настоящим изобретением указанный способ конструирования включает следующие этапы:
амплификация 5'- и 3' -фрагментов плеч гомологии BBD29_04920 и последовательности кодирующей области гена BBD29_04920 с областью его промотора, и введение гена BBD29_04920 или BBD29_04920 С1560А в геном штамма-хозяина путем гомологичной рекомбинации, что позволит указанному штамму сверхэкспрессировать ген BBD29_04920 или BBD29_04920C1560A.
Согласно одному из вариантов реализации настоящего изобретения используют следующие праймеры для амплификации 5'-фрагментов плеч гомологии:
Р7: 5' CAGTGCCAAGCTTGCATGCCTGCAGGTCGACTCTAG GACCCGCTTG CCATACGAAG 3' (SEQ ID NO: 11)
Р8: 5' GCATCACAAT GACATAACGA АТСТАСТСАТ CTGAAGAATC 3' (SEQ ID NO: 12)
Согласно одному из вариантов реализации настоящего изобретения используют следующие праймеры для амплификации 3'-фрагментов плеча гомологии:
P11: 5' GCGCGGGGAA GCGTCGATAG TTCGTGGGCA CTCTGGTTTG 3' (SEQ ID NO: 15)
P12: 5' CAGCTATGACCATGATTACGAATTCGAGCTCGGTACCC CATAAGAAAC AACCACTTCC 3' (SEQ ID NO: 16)
Согласно одному из вариантов реализации настоящего изобретения используют следующие праймеры для амплификации последовательности кодирующей области гена и области его промотора:
Р9: 5' GATTCTTCAG ATGAGTAGAT TCGT TATGTCATTG TGATGC 3' (SEQ ID NO: 13)
Р10: 5' CAAACCAGAG TGCCCACGAA CTATCGACGC TTCCCCGCGC 3' (SEQ ID NO: 14)
Согласно одному из вариантов реализации настоящего изобретения, опять же с вышеупомянутыми Р7/Р12 в качестве праймеров, осуществляют амплификацию с применением в качестве матриц смеси трех фрагментов, амплифицированного 5'- фрагмента плеча гомологии, 3'-фрагмента плеча гомологии и фрагмента последовательности кодирующей области гена и области его промотора, с получением интегрированного фрагмента плеча гомологии.
Согласно одному из вариантов реализации настоящего изобретения используемая ПЦР-система содержит: 10×буфер Ex Taq Buffer: 5 мкл, смесь dNTP (2,5 мМ каждый): 4 мкл, Mg2+ (25 мМ): 4 мкл, праймеры (10 пМ): каждого по 2 мкл; и Ex Taq (5 Ед/мкл): 0,25 мкл, в общем объеме 50 мкл. ПЦР- амплификацию осуществляют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 с при 94°С, ренатурация в течение 30 с при 52°С, удлинение в течение 120 с при 72°С (30 циклов) и избыточное удлинение в течение 10 мин при 72°С.
Согласно одному из вариантов реализации настоящего изобретения используют систему рекомбинации NEBuider для сборки челночной плазмиды PK18mobsacB с интегрированным фрагментом плеча гомологии, с получением интегративной плазмиды.
Согласно одному из вариантов реализации настоящего изобретения штамма-хозяин трансфицируют указанной интегративной плазмидой для введения гена BBD29_04920 или BBD29_04920C1560A в геном штамма-хозяина путем гомологичной рекомбинации.
Согласно одному из вариантов реализации настоящего изобретения штамм-хозяин представляет собой Corynebacterium glutamicum CGMCC №21220.
Согласно одному из вариантов реализации настоящего изобретения штамм-хозяин представляет собой АТСС 13869.
Согласно одному из вариантов реализации настоящего изобретения указанный штамм-хозяин представляет собой штамм, несущий последовательность полинуклеотида, представленную в SEQ ID NO: 2.
Согласно пятому аспекту настоящего изобретения также предложен способ конструирования рекомбинантного штамма, продуцирующего L-глутаминовую кислоту.
В соответствии с настоящим изобретением указанный способ конструирования включает следующие этапы:
амплификация последовательности кодирующей области гена BBD29_04920 и области промотора, или последовательности кодирующей области гена BBD29_04920C1560A и области промотора, конструирование сверхэкспрессирующего плазмидного вектора и трансформация указанным вектором штамма-хозяина, что позволяет указанному штамму сверхэкспрессировать ген BBD29_04920 или BBD29_04920C1560A.
Согласно одному из вариантов реализации настоящего изобретения используют следующие праймеры для амплификации последовательности кодирующей области гена и области его промотора:
Р17: 5' GCTTGCATGCCTGCAGGTCGACTCTAGAGGATCCCCTCGTTATGTCATTGTGATGC 3' (SEQ ID NO: 21)
P18: 5' ATCAGGCTGAAAATCTTCTCTCATCCGCCAAAACCTATCGACGCTTCCCCGCGC 3' (SEQ ID NO: 22).
Согласно одному из вариантов реализации настоящего изобретения указанная ПЦР-система содержит: 10×буфер Ex Taq Buffer: 5 мкл, смесь dNTP (2,5 мМ каждого): 4 мкл, Mg2+ (25 мМ): 4 мкл, праймеры (10 пМ): 2 мкл каждого из них, Ex Taq (5 Ед/мкл): 0,25 мкл, в общем объеме 50 мкл. ПЦР-амплификацию осуществляют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 с при 94°С, ренатурация в течение 30 с при 52°С, удлинение в течение 90 с при 72°С (30 циклов) и избыточное удлинение в течение 10 мин при 72°С.
Согласно одному из вариантов реализации настоящего изобретения систему рекомбинации NEBuider применяют для сборки челночной плазмиды pXMJ19 с фрагментом BBD29_04920 или BBD29_04920C1560A, имеющей собственные промоторы, с получением сверхэкспрессирующей плазмиды.
Согласно одному из вариантов реализации настоящего изобретения штамм-хозяин представляет собой Corynebacterium glutamicum CGMCC №21220.
Согласно одному из вариантов реализации настоящего изобретения штамм-хозяин представляет собой АТСС 13869.
Согласно одному из вариантов реализации настоящего изобретения указанный штамм-хозяин представляет собой штамм, несущий последовательность полинуклеотида, представленную в SEQ ID NO: 2.
Рекомбинантный штамм, предложенный согласно настоящему изобретению, может индивидуально применяться при ферментации для продуцирования L-глутаминовой кислоты, и может также применяться для продуцирования L-глутаминовой кислоты при гибридной ферментации с другими продуцирующими L-глутаминовую кислоту бактериями.
Согласно другому аспекту настоящего изобретения предложен способ получения L-глутаминовой кислоты, который включает культивирование бактерии; и получение L-глутаминовой кислоты из культуры.
Бактерия может быть культивирована в подходящей культуральной среде в условиях культивирования, известных в данной области техники. Культуральная среда может содержать: источник углерода, источник азота, следовые элементы и комбинацию перечисленного. При культивировании могут корректироваться значения рН культуры. Кроме того, может предотвращаться образование пузырьков при культивировании, например, с применением пеногасителя для предотвращения образования пузырьков. Кроме того, в культуру могут вводиться газы при культивировании. Указанные газы могут включать любые газы, способные поддерживать аэробные условия в культуре. При культивировании температура культуры может составлять от 20°С до 45°С. Продуцированная L-глутаминовая кислота может быть выделена из культуры, т.е. культуру обрабатывают серной кислотой или соляной кислотой и т.п., с последующим применением комбинации таких способов, как анионообменная хроматография, конденсация, кристаллизация и изоэлектрическое осаждение.
Согласно настоящему изобретению также предложен белок, обозначенный как белок BBD29_04920N520K, отличающийся тем, что указанный белок может представлять собой любое из:
А1) белка, имеющего последовательность аминокислот SEQ ID NO: 4;
А2) белка, обладающего 80% или более идентичностью белку, указанному в А1), и имеющего ту же функцию, что и белок, указанный в А1), полученного путем осуществления в последовательности аминокислот, представленной в SEQ ID NO: 4, замены, и/или делеции, и/или добавления остатков аминокислот;
A3) слитого белка, имеющего ту же функцию, полученного путем присоединения метки к N-концу и/или С-концу А1) или А2).
Согласно настоящему изобретению также предложена молекула нуклеиновой кислоты, обозначенная как BBD29_04920C1560A, отличающаяся тем, что указанная молекула нуклеиновой кислоты BBD29_04920C1560A может представлять собой любое из:
B1) молекулы нуклеиновой кислоты, кодирующей белок BBD29_04920N520K;
B2) молекулы ДНК, кодирующая последовательность которой представлена в SEQ ID NO: 2;
B3) молекулы ДНК, последовательность нуклеотидов которой представлена в SEQ ID NO: 2.
Молекула ДНК, представленная в SEQ ID NO: 2, представляет собой ген BBD29-04920Cl560A согласно настоящему изобретению.
Молекула ДНК, представленная в SEQ ID NO: 2 (ген BBD29_04920C1560A), кодирует белок BBD29_04920N520K, представленный в SEQ ID NO: 4.
Последовательность аминокислот белка BBD29_04920N520K (SEQ ID NO: 4) получают путем замены аспарагина (N) в положении 520 в SEQ ID NO: 3 на лизин (K).
Согласно настоящему изобретению также предложен биоматериал, отличающийся тем, что указанный биоматериал может представлять собой любое из:
B1) экспрессионной кассеты, содержащей молекулу нуклеиновой кислоты BBD29_04920C1560A;
B2) рекомбинантного вектора, содержащего молекулу нуклеиновой кислоты BBD29_04920C1560A, или рекомбинантного вектора, содержащего экспрессионную кассету С1);
B3) рекомбинантного микроорганизма, содержащего молекулу нуклеиновой кислоты BBD29_04920C1560A, или рекомбинантного микроорганизма, содержащего экспрессионную кассету С1), или рекомбинантного микроорганизма, содержащего рекомбинантный вектор С2).
Согласно настоящему изобретению также предложено любое из следующих применений по любому из D1)-D8):
F1) применение любого из D1)-D8) для регуляции продуцирования L-глутаминовой кислоты у микроорганизма;
F2) применение любого из D1)-D8) для конструирования генетически сконструированной бактерии для продуцирования L-глутаминовой кислоты;
F3) применение любого из D1)-D8) для продуцирования L-глутаминовой кислоты;
где D1)-D8) представляет собой:
D1) белок BBD29_04920N520K;
D2) молекулу нуклеиновой кислоты BBD29_04920C1560A;
D3) биоматериал;
D4) молекулу ДНК, последовательность нуклеотидов которой представляет собой SEQ ID NO: 1;
D5) молекулу ДНК, обладающую идентичностью 90% или более молекуле ДНК и имеющую ту же функцию, что и молекула ДНК, которая представлена в SEQ ID NO: 1, полученную путем осуществления в последовательности нуклеотидов, представленной в SEQ ID NO: 1, модификации, и/или замены, и/или делеции, и/или добавления одного или нескольких нуклеотидов;
D6) экспрессионную кассету, содержащую молекулу ДНК по D4) или D5);
D7) рекомбинантный вектор, содержащий молекулу ДНК по D4) или D5), или рекомбинантный вектор, содержащий экспрессионную кассету по D6);
D8) рекомбинантный микроорганизм, содержащий молекулу ДНК по D4) или D5), или рекомбинантный микроорганизм, содержащий экспрессионную кассету по D6), или рекомбинантный микроорганизм, содержащий рекомбинантный вектор по D7).
Молекула ДНК, представленная в SEQ ID NO: 1, представляет собой ген BBD29_04920 согласно настоящему изобретению.
Молекула ДНК, представленная в SEQ ID NO: 1 (ген BBD29_04920), кодирует белок, представленный в SEQ ID NO: 3.
В настоящем документе «идентичность» относится к идентичности последовательностей аминокислот или последовательностей нуклеотидов. Международные сайты поиска гомологии в сети Интернет могут применяться для определения идентичности последовательностей аминокислот или последовательностей нуклеотидов, например, страница BLAST на главной странице веб-сайта NCBI. Например, в Advanced BLAST2.1 может быть использована программа blastp со значением параметра Expect value, установленным на 10, всеми фильтрами в режиме OFF, BLOSUM62 в качестве матрицы и значениями штрафа за введение пропуска, штрафа за удлинение пропуска на один остаток и коэффициента лямбда (Lambda ratio), установленными на 11, 1 и 0,85 (устанавливаемые по умолчанию значения), соответственно, и выполнен поиск идентичности между парой аминокислотных последовательностей для расчета, после чего можно получить значение идентичности (%).
Согласно настоящему документу идентичность 80% или более может представлять собой идентичность, составляющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%.
Согласно настоящему документу идентичность 90% или более может представлять собой идентичность, составляющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%.
Регуляция продуцирования L-глутаминовой кислоты у микроорганизма согласно описанию в настоящем документе может представлять собой повышение или понижение уровня накопления L-глутаминовой кислоты у микроорганизма (т.е. стимуляцию или ингибирование биосинтеза L-глутаминовой кислоты).
Согласно настоящему изобретению также предложен способ повышения продуцирования L-глутаминовой кислоты у микроорганизма, при этом указанный способ включает любое из:
Е1) повышения уровня экспрессии или содержания молекулы нуклеиновой кислоты BBD29_04920C1560A в микроорганизме-мишени с получением микроорганизма, продуцирующего больше L-глутаминовой кислоты, чем микроорганизм-мишень;
Е2) повышения уровня экспрессии или содержания молекулы ДНК по D4) или D5) в микроорганизме-мишени с получением микроорганизма, продуцирующего больше L-глутаминовой кислоты, чем микроорганизм-мишень;
Е3) осуществления мутации в молекуле ДНК, последовательность нуклеотидов которой представляет собой SEQ ID NO: 1, в микроорганизме-мишени с получением микроорганизма, продуцирующего больше L-глутаминовой кислоты, чем микроорганизм-мишень.
Согласно описанному выше способу указанная мутация может представлять собой точечную мутацию, т.е. мутацию одного нуклеотида.
Согласно описанному выше способу указанная точечная мутация может представлять собой мутацию остатка аспарагина в положении 520 в последовательности аминокислот, кодируемой молекулой ДНК, представленной в SEQ ID NO: 1, на остаток другой аминокислоты.
Согласно описанному выше способу указанная точечная мутация может представлять собой мутацию аспарагина в положении 520 в последовательности аминокислот, кодируемой молекулой ДНК, представленной в SEQ ID NO: 1, на лизин, что обеспечивает мутированный белок BBD29_04920 N520K, последовательность аминокислот которого представляет собой SEQ ID NO: 4.
Указанная мутация относится к изменению одного или нескольких оснований в гене в результате сайт-направленной мутации, что приводит к изменению аминокислотного состава соответствующего белка, с получением таким образом нового белка, или обеспечением новой функции исходного белка, то есть представляет собой сайт-направленную мутацию генов. Методики осуществления сайт-направленных мутаций генов, таких как опосредованный олигонуклеотидными праймерами сайт-направленный мутагенез, ПЦР-опосредованный сайт-направленный мутагенез или кассетный мутагенез хорошо известны специалистам в данной области техники.
Точечная мутация согласно описанию в настоящем документе может представлять собой замену одного основания, инсерцию одного основания или делецию одного основания, и в особенности замену одного основания. Замена одного основания может представлять собой аллельную замену.
Указанная точечная мутация может заключаться в осуществлении модификации нуклеиновой кислоты с заменой цитозина (С) в положении 1560 гена BBD29_04920 (SEQ ID NO: 1).
В частности, указанная точечная мутация может заключаться в осуществлении мутации цитозина (G) в положении 1560 гена BBD29_04920 (SEQ ID NO: 1) на аденин (А) с получением молекулы ДНК, представленной в SEQ ID NO: 2.
В настоящем документе рекомбинантный вектор может представлять собой, в частности, рекомбинантный вектор pK18-BBD29_04920Cl560A, PK18mobsacB-BBD29_04920, PK18mobsacB-BBD29_04920C1560A, pXMJ19-BBD29_04920 или pXMJ19-BBD29_04920Cl560A.
Рекомбинантный вектор pK18-BBD29_04920Cl560A содержит молекулу ДНК, представленную положениями 846-1788 в мутированном гене BBD29_04920C1560A, представленном в SEQ ID NO: 2.
Рекомбинантный вектор PK18mobsacB-BBD29_04920 применяют для интеграции экзогенного гена BBD29_04920 в хромосому хозяина для сверхэкспрессии гена BBD29_04920 дикого типа у бактерии-продуцента.
Рекомбинантный вектор pK18mobsacB-BBD29_04920Cl560A применяют для интеграции экзогенного гена BBD29-04920C1560A в хромосому хозяина для сверхэкспрессии мутантного типа BBD29_04920C1560A у бактерии-продуцента.
Рекомбинантный вектор pXMJ19-BBD29_04920 применяют для экспрессии с помощью плазмиды экзогенного гена BBD29_04920 вне хромосомы для сверхэкспрессии гена BBD29_04920 дикого типа у бактерии-продуцента.
Рекомбинантный вектор pXMJ19-BBD29_04920Cl560A применяют для экспрессии с помощью плазмиды экзогенного гена BBD29_04920C1560A вне хромосомы для сверхэкспрессии мутантного типа гена BBD29_04920Cl560Ay бактерии-продуцента.
Все рекомбинантные векторы из pK18-BBD29-04920Cl560A, PK18mobsacB-BBD29_04920, PK18mobsacB-BBD29_04920Cl560A, pXMJ19-BBD29_04920 и pXMJ19-BBD29_04920Cl560A включены в заявленный объем настоящего изобретения.
В настоящем документе рекомбинантный микроорганизм может представлять собой, в частности, рекомбинантную бактерию YPG-001, YPG-002, YPG-003, YPG-004 или YPG-005.
Рекомбинантная бактерия YPG-001 представляет собой рекомбинантную бактерию, полученную путем трансформации рекомбинантного вектора pK18-BBD29_04920Cl560A в Corynebacterium glutamicum CGMCC №21220, и указанная рекомбинантная бактерия YPG-001 содержит мутированный ген BBD29_04920C1560A, представленный в SEQ ID NO: 2.
Рекомбинантная бактерия YPG-002 содержит двухкопийный ген BBD29_04920, представленный в SEQ ID NO: 1; рекомбинантная бактерия, содержащая двухкопийный ген BBD29_04920, способна значимо и устойчиво повышать уровень экспрессии гена BBD29_04920. Рекомбинантная бактерия YPG-002 представляет собой сконструированную бактерию, сверхэкспрессирующую ген BBD29 04920 дикого типа из генома.
Рекомбинантная бактерия YPG-003 содержит мутированный ген BBD29_04920C1560A, представленный в SEQ ID NO: 2; рекомбинантная бактерия YPG-003 представляет собой сконструированную бактерию, сверхэкспрессирующую мутантный тип гена BBD29_04920C1560A из генома.
Рекомбинантная бактерия YPG-004 содержит двухкопийный ген BBD29_04920, представленный в SEQ ID NO: 1; указанная рекомбинантная бактерия YPG-004 представляет собой сконструированную бактерию, сверхэкспрессирующую ген BBD29_04920 дикого типа на плазмиде, т.е. сверхэкспрессия происходит вне хромосомы с помощью плазмиды pXMJ19-BBD29_04920.
Рекомбинантная бактерия YPG-005 содержит мутированный ген BBD29_04920C1560A, представленный в SEQ ID NO: 2; рекомбинантная бактерия YPG-005 представляет собой сконструированную бактерию, сверхэкспрессирующую мутантный тип гена BBD29_04920C1560A на плазмиде, т.е. сверхэкспрессия происходит вне хромосомы с помощью плазмиды pXMJ19-BBD29_04920C1560A.
Все рекомбинантные бактерии YPG-001, YPG-002, YPG-003, YPG-004 или YPG-005 включены в заявленный объем настоящего изобретения.
Согласно настоящему изобретению также предложен способ конструирования рекомбинантного микроорганизма, при этом указанный способ включает по меньшей мере одно из:
F1) введения молекулы нуклеиновой кислоты BBD29_04920C1560A в микроорганизм-мишень с получением рекомбинантного микроорганизма;
F2) введения молекулы ДНК, представленной в SEQ ID NO: 1, в микроорганизм-мишень с получением рекомбинантного микроорганизма;
F3) редактирования молекулы ДНК, представленной в SEQ ID NO: 1, с использованием средства редактирования генов (такого как редактирование генов с изменением одного основания) для введения молекулы ДНК, представленной в SEQ ID NO: 2, в микроорганизм-мишень.
Указанное введение может быть представлено трансформацией бактерии-хозяина вектором, несущим молекулу ДНК согласно настоящему изобретению, любыми известными способами трансформации, такими как способ химической трансформации или способ электротрансформации. Введенная молекула ДНК может быть однокопийной или многокопийной. Введение может быть представлено интеграцией экзогенного гена в хромосому хозяина, или экспрессией вне хромосомы с помощью плазмиды.
Согласно настоящему изобретению также предложен способ получения L-глутаминовой кислоты, при этом указанный способ включает продуцирование L-глутаминовой кислоты любым из рекомбинантных микроорганизмов согласно описанию в настоящем документе.
Описанный выше способ может представлять собой получение L-глутаминовой кислоты ферментативным способом, а рекомбинантный микроорганизм может относиться к роду Corynebacterium, и в частности, представлять собой Corynebacterium glutamicum и его варианты.
Согласно настоящему изобретению:
SEQ ID NO: 1: последовательность кодирующей области BBD29_04920 дикого типа
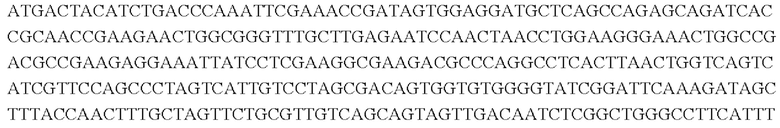


SEQ ID NO: 3: последовательность аминокислот белка, кодируемого BBD29_04920 дикого типа

SEQ ID NO: 4: последовательность аминокислот белка, кодируемого BBD29 04920N520K

Информация о депонировании: Название штамма: Corynebacterium glutamicum; Латинское название: Corynebacterium glutamicum; штамм №: YPGLU001; Депозитарное учреждение: Китайский главный центр коллекций микробиологических культур, сокращенно CGMCC; Адрес: No. 3, Yard 1, Beichen West Road, Chaoyang District, Beijing; Дата депонирования: 23 ноября 2020 г.; номер доступа в депозитарном центре: CGMCC №21220.
Наилучшие способы реализации изобретения
Техническое решение настоящего изобретения будет подробно описано ниже на конкретных примерах. Следует понимать, что следующие ниже примеры приведены только для иллюстрации и объяснения настоящего изобретения, и их не следует интерпретировать как ограничение объема правовой охраны настоящего изобретения. Все методики, реализованные на основе описанного выше содержания настоящего изобретения, входят в объем защиты настоящего изобретения. Если конкретным образом не указано иное, исходные материалы и реагенты, используемые в приведенных ниже примерах, могут быть коммерчески доступными или могут быть получены известными способами; все выполняемые операции известны в данной области техники или выполняются в соответствии с руководством пользователя по применению коммерчески доступных продуктов.
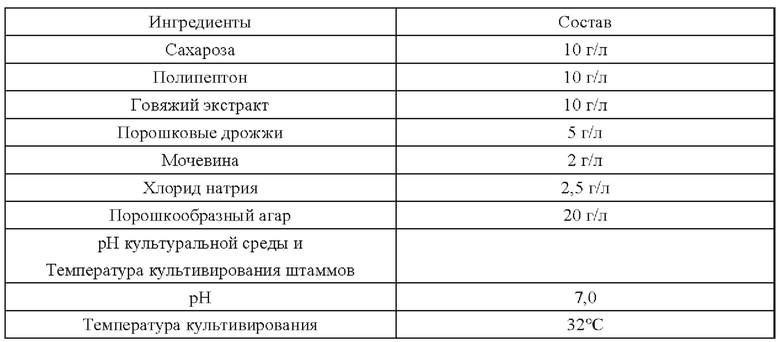
В приведенных ниже примерах состав основной среды, используемой для культивирования штамма, одинаковый, а сахарозу, канамицин или хлорамфеникол добавляют в состав основной среды по мере необходимости. Компоненты основной среды, которые дают основную среду при растворении в воде, приведены ниже:
Corynebacterium glutamicum YPGLU001 CGMCC №21220 в примерах ниже был депонирован в Китайском главном центре коллекций микробиологических культур (сокращенно CGMCC, адрес: No. 3, Yard 1, Beichen West Road, Chaoyang District, Beijing, Institute of Microbiology, Chinese Academy of Sciences) 23 ноября 2020 г. под номером доступа CGMCC №21220. Corynebacterium glutamicum YPGLU001 также известен как Corynebacterium glutamicum CGMCC №21220.
Пример 1. Конструирование вектора для трансформации pK18-BBD29_04920Cl560A, содержащего кодирующую область гена BBD29_04920 с точечной мутацией
В соответствии с последовательностью генома Corynebacterium glutamicum АТСС13869, опубликованной в NCBI, разрабатывали и синтезировали два пары праймеров для амплификации последовательности кодирующей области гена BBD29_04920 (SEQ ID NO: 1 BBD29_04920), и вводили точечную мутацию в штамм Corynebacterium glutamicum CGMCC №21220 путем аллельной замены (подтверждено соответствие кодирующей области гена BBD29_04920 на хромосоме штамма АТСС13869 при секвенировании). Последовательность аминокислот, соответствующая кодируемому белку, представлена в SEQ ID NO: 3, при замене цитозина (С) в положении 1560 последовательности нуклеотидов гена BBD29_04920 на аденин (A) (SEQ ID NO: 2: BBD29_04920C1560A), соответственно, происходит замена аспарагина (N) в положении 520 в последовательности аминокислот, соответствующей кодируемому белку, на лизин (K) (SEQ ID NO: 4: BBD29_04920N520K). Разработаны следующие праймеры (синтезированы Invitrogen Corporation, Шанхай):
P1: 5' CAGTGCCAAGCTTGCATGCCTGCAGGTCGACTCTAG TGTTTCTGTC TTGACCTTGG (SEQ ID NO: 5)
P2: 5' AGCCACGATG GTGACTTTTT GCAAGTTGTT_3' (SEQ ID NO: 6)
P3: 5' AACAACTTGC AAAAAGTCAC CATCGTGGCT 3' (SEQ ID NO: 7)
Р4: 5' CAGCTATGACCATGATTACGAATTCGAGCTCGGTACCC ACAGATTGGG CAGGTGCC 3' (SEQ ID NO: 8)
Способ конструирования: используя в качестве матрицы Corynebacterium glutamicum АТСС13869, осуществляют ПЦР-амплификацию с соответствующими праймерами P1, Р2 и Р3, Р4.
ПЦР-система: 10×буфер Ex Taq Buffer: 5 мкл, смесь dNTP (каждый 2,5 мМ): 4 мкл, Mg2+ (25 мМ): 4 мкл, праймеры (10 пМ): по 2 мкл каждого, Ex Taq (5 Ед/мкл): 0,25 мкл, матрица 1 мкл и вода в качестве баланса в общем объеме 50 мкл.
Вышеуказанную ПЦР-амплификацию осуществляют путем предварительной денатурации в течение 5 минут при 94°С, денатурации в течение 30 с при 94°С, ренатурации в течение 30 с при 52°С и удлинения в течение 40 с при 72°С в течение 30 циклов; и избыточного удлинения в течение 10 мин при 72°С с получением двух фрагментов ДНК, содержащих кодирующую область гена BBD29_04920 С1560А, каждый из которых имеет размер 766 п.о. и 778 п.о. (BBD29_04920 Up и BBD29_04920 Down).
Указанные два фрагмента ДНК выделяют посредством электрофореза на агарозном геле и очищают. Указанные два фрагмента ДНК затем используют в качестве матрицы с Р1 и Р4 в качестве праймеров для ПЦР-амплификации с перекрывающимися праймерами с получением фрагмента длиной 1514 п.о. (то есть BBD29_04920C1560A-Up-Down, последовательность нуклеотидов которого представлена положениями 846-1788 SEQ ID NO: 2.
ПЦР-система: 10×буфер Ex Taq Buffer: 5 мкл, смесь dNTP (каждый 2,5 мМ): 4 мкл, Mg2+ (25 мМ): 4 мкл, праймеры (10 пМ): по 2 мкл каждого, Ex Taq (5 Ед/мкл): 0,25 мкл, матрица 1 мкл и вода в качестве баланса в общем объеме 50 мкл.
Вышеуказанную перекрывающуюся ПЦР-амплификацию осуществляют путем предварительной денатурации в течение 5 минут при 94°С, денатурации в течение 30 с при 94°С, ренатурации в течение 30 с при 52°С, удлинения в течение 90 с при 72°С в течение 30 циклов и избыточного удлинения в течение 10 мин при 72°С.
Фрагмент ДНК вызывает изменение цитозина (С) в положении 1560 кодирующей области гена BBD29 04920 у Corynebacterium glutamicum CGMCC №21220 на аденин (А) и, в конечном итоге, приводит к изменению аминокислоты в положении 520 кодируемого белка с аспарагина (N) на лизин (K).
После расщепления плазмиды pK18mobsacB (приобретена у Addgene Coporation) Xba I, BBD29_04920 C1560A и линеаризированную плазмиду pK18mobsacB выделяют посредством электрофореза на агарозном геле и очищают, с последующей сборкой с применением системы рекомбинации NEBuider (NEB E5520S), получая вектор pK18-BBD29_04920Cl560A, плазмиду, содержащую маркер устойчивости к канамицину. При этом вектор pK18-BBD29_04920Cl560A отправляют для секвенирования и идентификации в компанию, осуществляющую секвенирование, и вектор pK18-BBD29_04920Cl560A, содержащий корректную точечную мутацию (С-А), отправляют на хранение.
pK18-BBD29_04920Cl560A представляет собой рекомбинантный вектор, полученный путем инсерции фрагмента ДНК BBD29_04920C1560A-Up-Down, представленного положениями 846-1488 SEQ ID NO: 2 в перечне последовательностей, в сайты распознавания XbaI в векторе pK18mobsacB, при сохранении в неизмененном виде других последовательностей вектора pK18mobsacB.
Пример 2. Конструирование сконструированного штамма, содержащего BBD29_04920C1560A с точечной мутацией
Способ конструирования: Плазмиду pK18-BBD29_04920Cl560A после аллельной замены электротрансформируют в Corynebacterium glutamicum CGMCC №21220. Каждую из культивируемых моноколоний идентифицируют с использованием праймера Р1 и универсального праймера M13R (5'CAG GAA АСА GCT ATG АСС 3') для определения положительного штамма с амплифицированной из него полосой около 1521 п.о. (представленной в SEQ ID NO: 29 для указанной последовательности). Положительный штамм культивируют на культуральной среде, содержащей 15% сахарозы, и выросшие моноколонии культивируют на культуральных средах с канамицином и без канамицина, соответственно. Штаммы, растущие на культуральной среде без канамицина и не растущие на культуральной среде с канамицином, дополнительно подвергают ПЦР-идентификации со следующими праймерами (синтезированы Invitrogen Corporation, Шанхай):
Р5: 5' CTATTGCTTT CTGGTGGTG 3' (SEQ ID NO: 9)
Р6: 5' TCGCCTTACG CTCCCTGCGT 3' (SEQ ID NO: 10)
Описанные выше амплифицированные продукты ПЦР (с последовательностью согласно SEQ ID NO: 30) подвергают денатурации при высокой температуре и в ледяной бане, после чего проводят SSCP-электрофорез (с амплифицированным фрагментом плазмиды pK18-BBD29_04920 C1560A в качестве положительного контроля, амплифицированным фрагментом АТСС13869 в качестве отрицательного контроля, и водой в качестве холостого контроля). Поскольку фрагменты отличаются структурой, их локализация при электрофорезе будет разной. Соответственно, штамм, характеризующийся успешной аллельной заменой, представляет собой такой штамм, локализация фрагмента которого при электрофорезе не совпадает с локализацией отрицательного контрольного фрагмента, и совпадает с локализацией положительного контрольного фрагмента. Праймеры Р5 и Р6 используют также для ПЦР-амплификации целевого фрагмента штамма, характеризующегося успешной аллельной заменой, который затем лигируют в вектор PMD19-T для секвенирования. Используя выравнивание последовательностей для мутированной последовательности оснований, подтверждают успешность аллельной замены у указанного штамма, который обозначен как YPG-001.
Рекомбинантная бактерия YPG-001 представляет собой генетически сконструированный штамм YPG-001 с точечной мутацией (С-А), полученной путем введения точечной мутации С1560А в кодирующую область гена BBD29_04920 (SEQ ID NO: 1) Corynebacterium glutamicum CGMCC №21220 в аллельной замене, чтобы вызвать мутацию в положении 1560 гена от С до А, сохраняя при этом другие последовательности гена без изменений.
По сравнению с Corynebacterium glutamicum CGMCC21220, Corynebacterium glutamicum YPG-001 отличается только заменой гена BBD29_04920, представленного в SEQ ID NO: 1, в геноме Corynebacterium glutamicum CGMCC21220, на ген BBD29_04920C1560A, представленный в SEQ ID NO: 2. SEQ ID NO: 1 и SEQ ID NO: 2 отличаются только одним нуклеотидом, локализованным в положении 1560.

Пример 3. Конструирование сконструированного штамма со сверхэкспрессируемым геном BBD29_04920 или BBD29_04920C1560AB геноме
В соответствии с последовательностью генома Corynebacterium glutamicum АТСС13869, опубликованной в базе NCBI, разрабатывали и синтезировали три пары праймеров для амплификации 5'- и 3' -фрагментов плеч гомологии, а также последовательность кодирующей области гена BBD29_04920 и область промотора для введения гена BBD29_04920 или BBD29_04920C1560A в штамм Corynebacterium glutamicum CGMCC №21220 путем гомологичной рекомбинации.
Разработаны следующие праймеры (синтезированы Invitrogen Corporation, Шанхай):
Р7: 5' CAGTGCCAAGCTTGCATGCCTGCAGGTCGACTCTAG GACCCGCTTG CCATACGAAG 3' (SEQ ID NO: 11)
Р8: 5' GCATCACAAT GACATAACGA АТСТАСТСАТ CTGAAGAATC 3' (SEQ ID NO: 12)
Р9: 5' GATTCTTCAG ATGAGTAGAT TCGT TATGTCATTG TGATGC 3' (SEQ ID NO: 13)
Р10: 5' CAAACCAGAG TGCCCACGAA CTATCGACGC TTCCCCGCGC 3' (SEQ ID NO: 14)
P11: 5' GCGCGGGGAA GCGTCGATAG TTCGTGGGCA CTCTGGTTTG 3' (SEQ ID NO: 15)
P12: 5' CAGCTATGACCATGATTACGAATTCGAGCTCGGTACCC CATAAGAAAC AACCACTTCC 3' (SEQ ID NO: 16)
Способ конструирования: Используя Corynebacterium glutamicum ATCC13869 или YPG-001, соответственно, в качестве матрицы, осуществляют ПЦР-амплификацию с праймерами Р7/Р8, Р9/Р10 и Р11/Р22, соответственно, с получением 5'-фрагмента плеча гомологии длиной около 806 п.о. (как указано в SEQ ID NO: 31 для последовательности), фрагмента области, кодирующей ген BBD29_04920 или BBD29_04920С1560А, и области промотора длиной около 1987 п.о. (как указано в SEQ ID NO: 32 или SEQ ID NO: 33 для последовательности), и 3'-фрагмента плеча гомологии длиной около 788 п.о. (как указано в SEQ ID NO: 34 для последовательности). Кроме того, с применением Р7/Р12 в качестве праймеров осуществляют амплификацию со смесью трех описанных выше амплифицированных фрагментов в качестве матрицы с получением единого фрагмента плеч гомологии против хода транскрипции-BBD29_04920 по ходу транскрипции (как указано в SEQ ID NO: 35 для последовательности); или единого фрагмента плеч гомологии против хода транскрипции-BBD29_04920 С1560А по ходу транскрипции (как указано в SEQ ID NO: 36 для последовательности). После завершения ПЦР-реакции осуществляют электрофорез амплифицированных продуктов для выделения требуемого фрагмента ДНК длиной приблизительно 3501 п.о. с применением набора с колонкой для выделения ДНК на геле (TIANGEN), который лигируют, используя систему рекомбинации NEBuider, в челночную плазмиду PK18mobsacB, расщепленную Xba I и выделенную с получением интегративной плазмиды PK18mobsacB-BBD29_04920 или PK18mobsacB- BBD29_04920C1560A. Указанная плазмида содержит маркер устойчивости к канамицину, благодаря чему путем скрининга с канамицином может быть получен рекомбинантный штамм, содержащий интегрированную в геном плазмиду.
pK18mobsacB-BBD29_04920 представляет собой рекомбинантный вектор, полученный путем инсерции единого фрагмента плеч гомологии 5'-BBD29_04920-3' в сайты распознавания Xba I челночной плазмиды pk18mobsacB.
pK18mobsacB-BBD29_04920Cl560A представляет собой рекомбинантный вектор, полученный путем инсерции единого фрагмента плеч гомологии 5'-BBD29_04920Cl560A-3' в сайты распознавания Xba I челночной плазмиды pk18mobsacB.
ПЦР-система: 10×буфер Ех Taq Buffer: 5 мкл, смесь dNTP (каждый 2,5 мМ): 4 мкл, Mg2+ (25 мМ): 4 мкл, праймеры (10 пМ): по 2 мкл каждого, Ex Taq (5 Ед/мкл): 0,25 мкл, матрица 1 мкл и вода до общего объема 50 мкл.
ПЦР-амплификацию осуществляют путем предварительной денатурации в течение 5 минут при 94°С, денатурации в течение 30 с при 94°С, ренатурации в течение 30 с при 52°С, удлинения в течение 120 с при 72°С (30 циклов) и избыточного удлинения в течение 10 мин при 72°С.
Каждую из двух интегративных плазмид вводят в штамм Corynebacterium glutamicum CGMCC №21220 путем электротрансформации, и осуществляют ПЦР-идентификацию выросших моноколоний с праймерами Р13/Р14 для получения положительного штамма, содержащего фрагмент длиной около 1674 п. о. (как указано в SEQ ID NO: 37 для последовательности), полученного при ПЦР-амплификации, тогда как штамм, не содержащий каких-либо амплифицированных из него фрагментов, представляет собой исходный штамм. При скрининге с 15% сахарозы положительный штамм культивируют на культуральных средах с канамицином и без канамицина, соответственно, и штаммы, растущие на культуральной среде без канамицина и не растущие на культуральной среде с канамицином подвергают дальнейшей ПЦР с праймерами Р15/Р16 для идентификации. Штамм с полосой около 1943 п.о. (последовательность согласно SEQ ID NO: 38), амплифицированный этим способом, представляет собой штамм с геном BBD29_04920 или BBD29_04920C1560A, интегрированным в геном Corynebacterium glutamicum CGMCC №21220, обозначенный как YPG-002 (без точечной мутации) и YPG-003 (с точечной мутацией).
Р13: 5' GTCCAAGGTG ACGGCCGCAC 3' (SEQ ID NO: 17)
Р14: 5' СТТТТТСАСС AATGAGTGGC 3' (SEQ ID NO: 18)
P15: 5' CCGTAAACAG CTTCTAAGCT 3' (SEQ ID NO: 19)
P16: 5' ATATTCGGCC CAGCAGCAGC 3' (SEQ ID NO: 20)
Рекомбинантная бактерия YPG-002 представляет собой рекомбинантную бактерию, содержащую двухкопийный ген BBD29_04920, представленный в SEQ ID NO: 1, полученную путем интеграции единого фрагмента плеч гомологии 5'-BBD29_04920-3' в геном штамма Corynebacterium glutamicum YPGLU001, и указанная рекомбинантная бактерия, содержащая двухкопийный ген BBD29_04920, способна значимо и устойчиво повышать уровень экспрессии BBD29_04920.
Рекомбинантная бактерия YPG-003 представляет собой рекомбинантную бактерию, содержащую мутантный ген BBD29_04920 С1560А, представленный в SEQ ID NO: 2, полученный путем интеграции единого фрагмента плеч гомологии 5'-BBD29_04920Cl560A-3' в геном штамма Corynebacterium glutamicum YPGLU001.
Пример 4. Конструирование сконструированного (модифицированного) штамма, сверхэкспрессирующего ген BBD29_04920 или BBD29_04920C1560A с плазмиды
В соответствии с последовательностью генома Corynebacterium glutamicum АТСС13869, опубликованной в базе NCBI, разрабатывали и синтезировали пару праймеров для амплификации последовательности кодирующей области гена BBD29 04920 и области промотора. Разработаны следующие праймеры (синтезированы Invitrogen Corporation, Шанхай):
P17: 5' GCTTGCATGCCTGCAGGTCGACTCTAGAGGATCCCCTCGTTATGTCATTGTGATGC 3' (SEQ ID NO: 21)
P18: 5' ATCAGGCTGAAAATCTTCTCTCATCCGCCAAAACCTATCGACGCTTCCCCGCGC 3' (SEQ ID NO: 22)
Способ конструирования: Используя YPG-002 или YPG-001, соответственно, в качестве матрицы, осуществляют ПЦР-амплификацию с праймерами Р17/Р18 с получением фрагмента ДНК в 2017 п.о., содержащего ген BBD29_04920 или BBD29_BBD29_04920C1560A и его промотор (последовательность согласно SEQ ID NO: 39 или SEQ ID NO: 40). Осуществляют электрофорез амплифицированных продуктов для выделения требуемого фрагмента ДНК длиной 1947 п.о. с применением набора с колонкой для выделения ДНК на геле, который лигируют, используя систему рекомбинации NEBuider, в челночную плазмиду pXMJ19, расщепленную EcoR I и выделенную с получением сверхэкспрессирующей плазмиды pXMJ19-BBD29_04920 или pXMJ19-BBD29_04920Cl560A. Эта плазмида содержит маркер устойчивости к хлорамфениколу, и трансформация штамма плазмидой может быть достигнута при скрининге с хлорамфениколом.
ПЦР-система: 10×буфер Ex Taq Buffer: 5 мкл, смесь dNTP (каждый 2,5 мМ): 4 мкл, Mg2+ (25 мМ): 4 мкл, праймеры (10 пМ): по 2 мкл каждого, Ex Taq (5 Ед/мкл): 0,25 мкл, матрица: 1 мкл и вода в качестве баланса в общем объеме 50 мкл.
ПЦР-амплификацию осуществляют путем предварительной денатурации в течение 5 минут при 94°С, денатурации в течение 30 с при 94°С, ренатурации в течение 30 с при 52°С, удлинения в течение 90 с при 72°С (30 циклов) и избыточного удлинения в течение 10 мин при 72°С.
Сверхэкспрессирующая плазмида pXMJ19-BBD29 04920 представляет собой рекомбинантный вектор, полученный путем инсерции фрагмента ДНК (представленного в SEQ ID NO: 39 для указанной последовательности), содержащего ген BBD29_04920 и его промотор, в сайты распознавания EcoR I челночной плазмиды pXMJ19.
Сверхэкспрессирующая плазмида pXMJ19-BBD29_04920Cl560A представляет собой рекомбинантный вектор, полученный путем инсерции фрагмента ДНК (представленного в SEQ ID NO: 40 для указанной последовательности), содержащего ген BBD29_04920C1560A и его промотор, в сайты распознавания EcoR I челночной плазмиды pXMJ19.
Каждую из этих двух плазмид вводят в штамм Corynebacterium glutamicum CGMCC №21220 путем электротрансформации, и осуществляют ПЦР-идентификацию выросших моноколоний с праймерами M13R (-48) (5'AGCGGATAAC AATTTCACAC AGGA 3') и Р18 для трансформированного штамма, содержащего фрагмент длиной около 2056 п.о. (последовательность без точечной мутации согласно SEQ ID NO: 41; последовательность с точечной мутацией согласно SEQ ID NO: 41, за исключением положения 1794, представляющего собой А), из ПЦР-амплификации, обозначенного как YPG-004 (без точечной мутации) и YPG-005 (с точечной мутацией).
Рекомбинантная бактерия YPG-004 содержит двухкопийный ген BBD29_04920 согласно SEQ ID NO: 1; рекомбинантная бактерия YPG-004 представляет собой сконструированную бактерию, сверхэкспрессирующую ген BBD29_04920 дикого типа на плазмиде, т.е. сверхэкспрессия происходит вне хромосомы с помощью плазмиды pXMJ19-BBD29_04920.
Рекомбинантная бактерия YPG-005 содержит мутированный ген BBD29_04920C1560A согласно SEQ ID NO: 2; рекомбинантная бактерия YPG-005 представляет собой сконструированную бактерию, сверхэкспрессирующую мутантный ген BBD29_04920C1560A на плазмиде, т.е. сверхэкспрессия происходит вне хромосомы с плазмиды pXMJ19-BBD29_04920Cl560A.
Пример 5. Конструирование сконструированного штамма с делетированным в геноме геном BBD29_04920
В соответствии с последовательностью генома Corynebacterium glutamicum АТСС13869, опубликованной в базе NCBI, синтезируют две пары праймеров для амплификации фрагментов на обоих концах кодирующей области гена BBD29_04920, в виде 5'- и 3'-фрагментов плеч гомологии. Разработаны следующие праймеры (синтезированы Invitrogen Corporation, Шанхай):
Р19: 5' CAGTGCCAAGCTTGCATGCCTGCAGGTCGACTCTAGTAAGGGGCAG TTGGTCTCG 3' (SEQ ID NO: 23)
Р20: 5' GTATCAGGGGTTAAAAATTGCTTAATTTTCC CTGGCAGAA 3' (SEQ ID NO: 24)
P21: 5' TTCTGCCAGGGAAAATTAAGCAATTTTTAACCCCTGATAC 3' (SEQ ID NO: 25)
P22: 5' CAGCTATGACCATGATTACGAATTCGAGCTCGGTACCCATATCGCGCG ACATTGCGCG 3' (SEQ ID NO: 26)
Способ конструирования: Используя Corynebacterium glutamicum ATCC13869 в качестве матрицы, проводят ПЦР-амплификацию с праймерами Р19/Р20 и Р21/Р22, соответственно, с получением 5'-фрагмента плеча гомологии длиной 733 п.о. (последовательность согласно SEQ ID NO: 42) и 3'-фрагмента плеча гомологии длиной 814 п.о. (последовательность согласно SEQ ID NO: 43). Кроме того, Р19/Р22 используют в качестве праймеров для ПЦР с перекрывающимися праймерами с получением единого фрагмента плеч гомологии длиной 1507 п. о. (последовательность согласно SEQ ID NO: 44). После завершения ПЦР-реакции проводят электрофорез амплифицированных продуктов для выделения требуемого фрагмента ДНК длиной 1507 п.о. с применением набора с колонкой для выделения ДНК на геле, который лигируют с помощью системы рекомбинации NEBuider в челночную плазмиду, плазмиду pk18mobsacB, расщепленную Xba I и выделенную с получением нокаутной плазмиды. Указанная плазмида содержит маркер устойчивости к канамицину.
Указанной нокаутной плазмидой электротрансформируют штамм Corynebacterium glutamicum CGMCC №21220 и осуществляют ПЦР-идентификацию выросших моноколоний, в каждом случае со следующими праймерами (синтезированы Invitrogen Corporation, Шанхай):
Р23: 5' TAAG GGGCAG TTGGTCTCG 3' (SEQ ID NO: 27)
Р24:5' ATATCGCGCG ACATTGCGCG 3' (SEQ ID NO: 28)
Штамм с полосами, соответствующими 1433 п. о. (последовательность согласно SEQ ID NO: 45) и 3380 п.о. (как указано в SEQ ID NO: 46 для последовательности), амплифицированными в ходе описанной выше ПЦР-амплификации, представляет собой положительный штамм, тогда как штамм с единственной амплифицированной при ПЦР полосой, соответствующей 3380 п. о., представляет собой исходный штамм. При скрининге на культуральной среде с 15% сахарозы положительный штамм культивируют на культуральных средах с канамицином и без канамицина, соответственно, и штаммы, растущие на культуральной среде без канамицина и не растущие на культуральной среде с канамицином, дополнительно подвергают ПЦР-идентификации с праймерами Р23/Р24. Штамм с полосой, соответствующей 1433 п.о., амплифицированной из него, представляет собой генетически сконструированный штамм с нокаутом кодирующей области гена BBD29_04920, обозначенный как YPG-006.
Рекомбинантная бактерия YPG-006 представляет собой штамм, полученный путем нокаута кодирующей области гена BBD29_04920 в геноме Corynebacterium glutamicum CGMCC №21220.
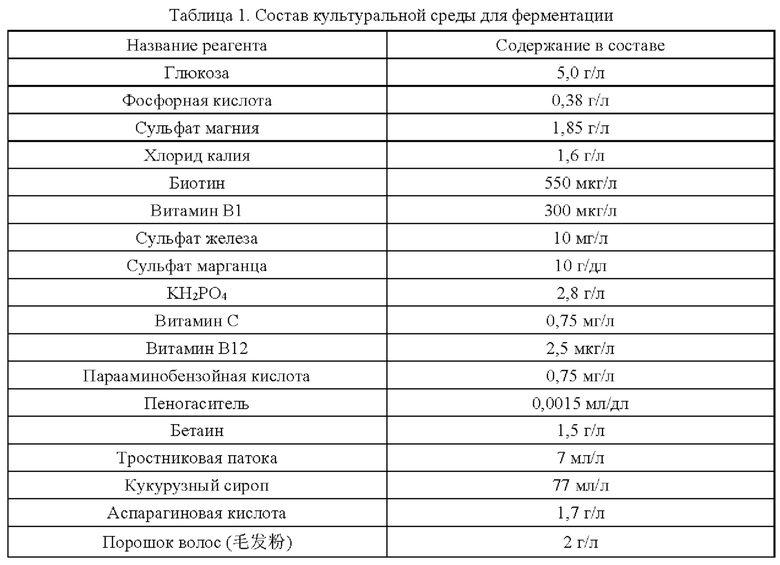
Пример 6. Эксперименты с ферментацией для получения L-глутаминовой кислоты
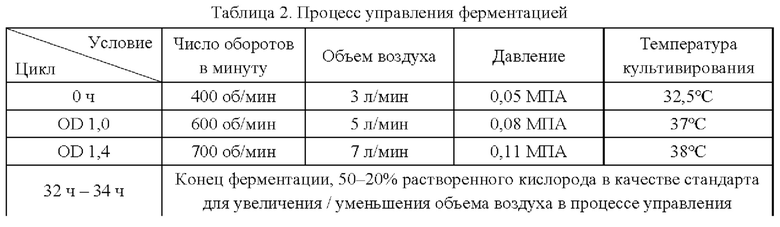
Эксперименты с ферментацией проводят на штаммах YPG-001, YPG-002, YPG-003, YPG-004, YPG-005 и YPG-006, сконструированных согласно приведенным примерам, и на исходном штамме Corynebacterium glutamicum CGMCC №21220, в ферментативном резервуаре типа BLBIO-5GC-4-H (приобретен у Shanghai Bailun Biological Technology Co., Ltd.); культуральная среда (которую получают растворением компонентов, представленных в таблице 1, в воде) описана в таблице 1; процесс управления представлен в таблице 2; продукты ферментации собирают.
В начальный момент, когда посев завершен, бактерия находится в системе в концентрации 15 г/л. Во время ферментации: содержание сахара в системе (остаточный сахар) контролируют путем добавления водного раствора, содержащего 50-55% глюкозы.
Для каждого штамма эксперименты проводят в трех повторностях. Результаты приведены в таблице 3.

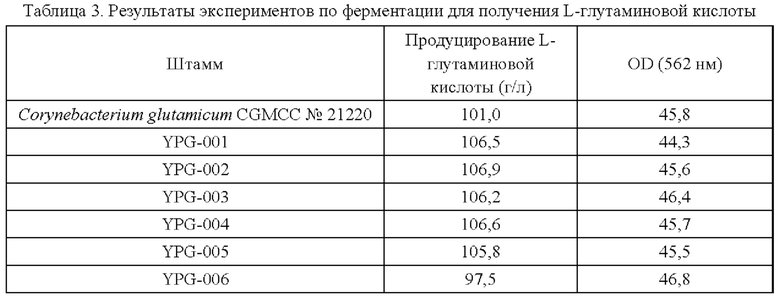
Результаты из таблицы 3 демонстрируют, что сверхэкспрессия гена BBD29_04920, или точечная мутация в кодирующей области гена BBD29_04920, BBD29_04920C1560A, и/или сверхэкспрессия у Corynebacterium glutamicum способствуют повышению продуцирования L-глутаминовой кислоты, тогда как ослабление экспрессии или нокаут гена неблагоприятны для накопления L-глутаминовой кислоты.
Варианты реализации настоящего изобретения проиллюстрированы выше. Однако настоящее изобретение не ограничено вышеупомянутыми вариантами реализации. Любые модификации, эквивалентные замены, усовершенствования и т.д., осуществленные в соответствии с сущностью и принципами настоящего изобретения, должны быть включены в заявленный объем настоящего изобретения.
Промышленное применение
Согласно настоящему изобретению было обнаружено, что через снижение экспрессии или нокаут гена BBD29_04920 кодируемый этим геном продукт влияет на способность к продуцированию L-глутаминовой кислоты, и что рекомбинантный штамм, полученный путем введения точечной мутации в кодирующую последовательность, или путем увеличения числа копий, или сверхэкспрессии указанного гена, способствует продуцированию глутаминовой кислоты в более высокой концентрации по сравнению с немодифицированным штаммом.
В частности, согласно настоящему изобретению сначала конструируют генетически сконструированную бактерию YPG-001 с точечной мутацией (С-А) путем введения точечной мутации в кодирующую область гена BBD29_04920 (SEQ ID NO: 1) Corynebacterium glutamicum CGMCC №21220 путем аллельной замены. Для дополнительного изучения и подтверждения того, что сверхэкспрессия гена BBD29_04920 дикого типа или содержащего его мутантного гена BBD29_04920C1560A у бактерии-продуцента может повышать продуцирование L-глутаминовой кислоты, экзогенный ген интегрируют в хромосому хозяина или экспрессируют вне хромосомы с помощью плазмиды, соответственно, конструируя таким образом сконструированные бактерии YPG-002, YPG-003, YPG-004 и YPG-005, сверхэкспрессирующие ген BBD29_04920 или ген BBD29_04920C1560A из генома и с плазмиды. Эксперименты предполагают, что ген BBD29_04920 и его варианты вовлечены в биосинтез L-глутаминовой кислоты. Используя сверхэкспрессию или нокаут гена BBD29_04920, или сайт-направленную мутацию (такую как точечная мутация) в указанном гене, можно регулировать уровень накопления L-глутаминовой кислоты у микроорганизма. Точечная мутация в кодирующей области гена BBD29_04920 или сверхэкспрессия гена BBD29_04920 или его мутантного гена BBD29_04920С1560А у бактерии-продуцента способствует повышению продуцирования и увеличению показателя конверсии L-глутаминовой кислоты, тогда как нокаут или ослабление экспрессии гена BBD29_04920 неблагоприятны для накопления L-глутаминовой кислоты. Ген BBD29_04920 и его вариант (такой как ген BBD29_04920C1560A) могут применяться для конструирования генетически сконструированного штамма для продуцирования L-глутаминовой кислоты, чтобы способствовать повышению продуцирования L-глутаминовой кислоты, и для получения высокопродуктивного и высококачественного штамма для промышленного производства, полезного для широкого диапазона вариантов применения и имеющего высокую экономическую ценность для промышленного производства L-глутаминовой кислоты.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> INNER MONGOLIA EPPEN BIOTECH CO., LTD. / ИННЕР МОНГОЛИА ЭППЕН БИОТЕК КО.,
ЛТД
<120> Recombinant Strain with Modified Gene BBD29_04920 for Producing
L-Glutamic Acid, and Method for Constructing the Same and Use Thereof /
РЕКОМБИНАНТНЫЙ ШТАММ С МОДИФИЦИРОВАННЫМ ГЕНОМ BBD29_04920 ДЛЯ ПОЛУЧЕНИЯ
L-ГЛУТАМИНОВОЙ КИСЛОТЫ, А ТАКЖЕ СПОСОБ ЕГО КОНСТРУИРОВАНИЯ И ПРИМЕНЕНИЕ
<150> 202011631254 .5
<151> 2020-12-30
<160> 46
<170> SIPOSequenceListing 1.0
<210> 1
<211> 1788
<212> ДНК
<213> Corynebacterium glutamicum
<400> 1
atgactacat ctgacccaaa ttcgaaaccg atagtggagg atgctcagcc agagcagatc 60
accgcaaccg aagaactggc gggtttgctt gagaatccaa ctaacctgga agggaaactg 120
gccgacgccg aagaggaaat tatcctcgaa ggcgaagacg cccaggcctc acttaactgg 180
tcagtcatcg ttccagccct agtcattgtc ctagcgacag tggtgtgggg tatcggattc 240
aaagatagct ttaccaactt tgctagttct gcgttgtcag cagtagttga caatctcggc 300
tgggccttca ttttgtttgg cacagtcttt gtatttttta tcgttgttat cgccgctagt 360
aaattcggca cgattcgctt aggccgcatt gatgaagcac cagagtttcg cacggtgtca 420
tggatttcca tgatgtttgc tgcaggtatg ggtattggtt tgatgttcta cggaaccaca 480
gaacctttaa ccttctaccg caatggtgta cctggatatg atgaacacaa tgttggcgtt 540
gctatgtcca cgacaatgtt ccactggacc ttgcatccat gggctatcta cgcaattgtg 600
ggcctagcca ttgcctattc gaccttccga gtgggccgta aacagcttct aagctctgca 660
ttcgtgccac tcattggtga aaaaggtgca gaaggatggt tgggcaagct catcgacatc 720
ctggcgatta tcgccacagt attcggcacc gcatgttccc ttggcctggg tgcgctgcag 780
atcggtgcag gactttccgc agccaacatc attgaaaatc cgagtgactg gactgtcatt 840
ggtattgttt ctgtcttgac cttggcattt atcttctctg ctatttctgg tgtgggcaag 900
ggaatccagt acctctccaa cgccaacatg gttctggcag ctctgctcgc gattttcgtg 960
ttcgttgtcg gaccaaccgt gtcgattttg aacctgctgc caggttctat tggcaactac 1020
ctgtccaact tcttccaaat ggcaggccgc actgccatga gtgccgacgg cacagcaggt 1080
gagtggctag gtagctggac catcttctac tgggcatggt ggatctcttg gtcaccattc 1140
gtaggaatgt tcttggcacg tatttcccgt ggccgctcca tccgtgagtt catcctgggc 1200
gtgttgctcg tcccagcagg tgtgtccacc gtatggttct ccatttttgg tggcactgcg 1260
attgtcttcg aacaaaatgg ggaatccatt tggggtgatg gtgcagcaga agagcagctc 1320
tttggattgc ttcatgcact tccaggtggg caaataatgg gcatcatcgc catgattttg 1380
ctgggtactt tcttcattac ctctgcagac tctgcttcca ccgtcatggg caccatgagt 1440
cagcacggcc agctggaagc caacaagtgg gtgacagctg cctggggtgt tgctaccgca 1500
gctattggac taacgctatt gctttctggt ggtgacaatg ccttgaacaa cttgcaaaac 1560
gtcaccatcg tggctgcaac accattcctg tttgtggtta ttggattgat gtttgcgtta 1620
gtcaaggact taagcaatga tgtgatctac ctcgagtacc gtgagcagca acgcttcaac 1680
gcgcgccttg cccgtgaacg tcgtgttcac aatgaacacc gcaagcgtga actggctgca 1740
aagcgacgca gggagcgtaa ggcgagtggc gcggggaagc gtcgatag 1846
<210> 2
<211> 1788
<212> ДНК
<213> Искусственная последовательность
<400> 2
atgactacat ctgacccaaa ttcgaaaccg atagtggagg atgctcagcc agagcagatc 60
accgcaaccg aagaactggc gggtttgctt gagaatccaa ctaacctgga agggaaactg 120
gccgacgccg aagaggaaat tatcctcgaa ggcgaagacg cccaggcctc acttaactgg 180
tcagtcatcg ttccagccct agtcattgtc ctagcgacag tggtgtgggg tatcggattc 240
aaagatagct ttaccaactt tgctagttct gcgttgtcag cagtagttga caatctcggc 300
tgggccttca ttttgtttgg cacagtcttt gtatttttta tcgttgttat cgccgctagt 360
aaattcggca cgattcgctt aggccgcatt gatgaagcac cagagtttcg cacggtgtca 420
tggatttcca tgatgtttgc tgcaggtatg ggtattggtt tgatgttcta cggaaccaca 480
gaacctttaa ccttctaccg caatggtgta cctggatatg atgaacacaa tgttggcgtt 540
gctatgtcca cgacaatgtt ccactggacc ttgcatccat gggctatcta cgcaattgtg 600
ggcctagcca ttgcctattc gaccttccga gtgggccgta aacagcttct aagctctgca 660
ttcgtgccac tcattggtga aaaaggtgca gaaggatggt tgggcaagct catcgacatc 720
ctggcgatta tcgccacagt attcggcacc gcatgttccc ttggcctggg tgcgctgcag 780
atcggtgcag gactttccgc agccaacatc attgaaaatc cgagtgactg gactgtcatt 840
ggtattgttt ctgtcttgac cttggcattt atcttctctg ctatttctgg tgtgggcaag 900
ggaatccagt acctctccaa cgccaacatg gttctggcag ctctgctcgc gattttcgtg 960
ttcgttgtcg gaccaaccgt gtcgattttg aacctgctgc caggttctat tggcaactac 1020
ctgtccaact tcttccaaat ggcaggccgc actgccatga gtgccgacgg cacagcaggt 1080
gagtggctag gtagctggac catcttctac tgggcatggt ggatctcttg gtcaccattc 1140
gtaggaatgt tcttggcacg tatttcccgt ggccgctcca tccgtgagtt catcctgggc 1200
gtgttgctcg tcccagcagg tgtgtccacc gtatggttct ccatttttgg tggcactgcg 1260
attgtcttcg aacaaaatgg ggaatccatt tggggtgatg gtgcagcaga agagcagctc 1320
tttggattgc ttcatgcact tccaggtggg caaataatgg gcatcatcgc catgattttg 1380
ctgggtactt tcttcattac ctctgcagac tctgcttcca ccgtcatggg caccatgagt 1440
cagcacggcc agctggaagc caacaagtgg gtgacagctg cctggggtgt tgctaccgca 1500
gctattggac taacgctatt gctttctggt ggtgacaatg ccttgaacaa cttgcaaaaa 1560
gtcaccatcg tggctgcaac accattcctg tttgtggtta ttggattgat gtttgcgtta 1620
gtcaaggact taagcaatga tgtgatctac ctcgagtacc gtgagcagca acgcttcaac 1680
gcgcgccttg cccgtgaacg tcgtgttcac aatgaacacc gcaagcgtga actggctgca 1740
aagcgacgca gggagcgtaa ggcgagtggc gcggggaagc gtcgatag 1846
<210> 3
<211> 595
<212> БЕЛОК
<213> Corynebacterium glutamicum
<400> 3
Met Thr Thr Ser Asp Pro Asn Ser Lys Pro Ile Val Glu Asp Ala Gln
1 5 10 15
Pro Glu Gln Ile Thr Ala Thr Glu Glu Leu Ala Gly Leu Leu Glu Asn
20 25 30
Pro Thr Asn Leu Glu Gly Lys Leu Ala Asp Ala Glu Glu Glu Ile Ile
35 40 45
Leu Glu Gly Glu Asp Ala Gln Ala Ser Leu Asn Trp Ser Val Ile Val
50 55 60
Pro Ala Leu Val Ile Val Leu Ala Thr Val Val Trp Gly Ile Gly Phe
65 70 75 80
Lys Asp Ser Phe Thr Asn Phe Ala Ser Ser Ala Leu Ser Ala Val Val
85 90 95
Asp Asn Leu Gly Trp Ala Phe Ile Leu Phe Gly Thr Val Phe Val Phe
100 105 110
Phe Ile Val Val Ile Ala Ala Ser Lys Phe Gly Thr Ile Arg Leu Gly
115 120 125
Arg Ile Asp Glu Ala Pro Glu Phe Arg Thr Val Ser Trp Ile Ser Met
130 135 140
Met Phe Ala Ala Gly Met Gly Ile Gly Leu Met Phe Tyr Gly Thr Thr
145 150 155 160
Glu Pro Leu Thr Phe Tyr Arg Asn Gly Val Pro Gly Tyr Asp Glu His
165 170 175
Asn Val Gly Val Ala Met Ser Thr Thr Met Phe His Trp Thr Leu His
180 185 190
Pro Trp Ala Ile Tyr Ala Ile Val Gly Leu Ala Ile Ala Tyr Ser Thr
195 200 205
Phe Arg Val Gly Arg Lys Gln Leu Leu Ser Ser Ala Phe Val Pro Leu
210 215 220
Ile Gly Glu Lys Gly Ala Glu Gly Trp Leu Gly Lys Leu Ile Asp Ile
225 230 235 240
Leu Ala Ile Ile Ala Thr Val Phe Gly Thr Ala Cys Ser Leu Gly Leu
245 250 255
Gly Ala Leu Gln Ile Gly Ala Gly Leu Ser Ala Ala Asn Ile Ile Glu
260 265 270
Asn Pro Ser Asp Trp Thr Val Ile Gly Ile Val Ser Val Leu Thr Leu
275 280 285
Ala Phe Ile Phe Ser Ala Ile Ser Gly Val Gly Lys Gly Ile Gln Tyr
290 295 300
Leu Ser Asn Ala Asn Met Val Leu Ala Ala Leu Leu Ala Ile Phe Val
305 310 315 320
Phe Val Val Gly Pro Thr Val Ser Ile Leu Asn Leu Leu Pro Gly Ser
325 330 335
Ile Gly Asn Tyr Leu Ser Asn Phe Phe Gln Met Ala Gly Arg Thr Ala
340 345 350
Met Ser Ala Asp Gly Thr Ala Gly Glu Trp Leu Gly Ser Trp Thr Ile
355 360 365
Phe Tyr Trp Ala Trp Trp Ile Ser Trp Ser Pro Phe Val Gly Met Phe
370 375 380
Leu Ala Arg Ile Ser Arg Gly Arg Ser Ile Arg Glu Phe Ile Leu Gly
385 390 395 400
Val Leu Leu Val Pro Ala Gly Val Ser Thr Val Trp Phe Ser Ile Phe
405 410 415
Gly Gly Thr Ala Ile Val Phe Glu Gln Asn Gly Glu Ser Ile Trp Gly
420 425 430
Asp Gly Ala Ala Glu Glu Gln Leu Phe Gly Leu Leu His Ala Leu Pro
435 440 445
Gly Gly Gln Ile Met Gly Ile Ile Ala Met Ile Leu Leu Gly Thr Phe
450 455 460
Phe Ile Thr Ser Ala Asp Ser Ala Ser Thr Val Met Gly Thr Met Ser
465 470 475 480
Gln His Gly Gln Leu Glu Ala Asn Lys Trp Val Thr Ala Ala Trp Gly
485 490 495
Val Ala Thr Ala Ala Ile Gly Leu Thr Leu Leu Leu Ser Gly Gly Asp
500 505 510
Asn Ala Leu Asn Asn Leu Gln Asn Val Thr Ile Val Ala Ala Thr Pro
515 520 525
Phe Leu Phe Val Val Ile Gly Leu Met Phe Ala Leu Val Lys Asp Leu
530 535 540
Ser Asn Asp Val Ile Tyr Leu Glu Tyr Arg Glu Gln Gln Arg Phe Asn
545 550 555 560
Ala Arg Leu Ala Arg Glu Arg Arg Val His Asn Glu His Arg Lys Arg
565 570 575
Glu Leu Ala Ala Lys Arg Arg Arg Glu Arg Lys Ala Ser Gly Ala Gly
580 585 590
Lys Arg Arg
595
<210> 4
<211> 595
<212> БЕЛОК
<213> Искусственная последовательность
<400> 4
Met Thr Thr Ser Asp Pro Asn Ser Lys Pro Ile Val Glu Asp Ala Gln
1 5 10 15
Pro Glu Gln Ile Thr Ala Thr Glu Glu Leu Ala Gly Leu Leu Glu Asn
20 25 30
Pro Thr Asn Leu Glu Gly Lys Leu Ala Asp Ala Glu Glu Glu Ile Ile
35 40 45
Leu Glu Gly Glu Asp Ala Gln Ala Ser Leu Asn Trp Ser Val Ile Val
50 55 60
Pro Ala Leu Val Ile Val Leu Ala Thr Val Val Trp Gly Ile Gly Phe
65 70 75 80
Lys Asp Ser Phe Thr Asn Phe Ala Ser Ser Ala Leu Ser Ala Val Val
85 90 95
Asp Asn Leu Gly Trp Ala Phe Ile Leu Phe Gly Thr Val Phe Val Phe
100 105 110
Phe Ile Val Val Ile Ala Ala Ser Lys Phe Gly Thr Ile Arg Leu Gly
115 120 125
Arg Ile Asp Glu Ala Pro Glu Phe Arg Thr Val Ser Trp Ile Ser Met
130 135 140
Met Phe Ala Ala Gly Met Gly Ile Gly Leu Met Phe Tyr Gly Thr Thr
145 150 155 160
Glu Pro Leu Thr Phe Tyr Arg Asn Gly Val Pro Gly Tyr Asp Glu His
165 170 175
Asn Val Gly Val Ala Met Ser Thr Thr Met Phe His Trp Thr Leu His
180 185 190
Pro Trp Ala Ile Tyr Ala Ile Val Gly Leu Ala Ile Ala Tyr Ser Thr
195 200 205
Phe Arg Val Gly Arg Lys Gln Leu Leu Ser Ser Ala Phe Val Pro Leu
210 215 220
Ile Gly Glu Lys Gly Ala Glu Gly Trp Leu Gly Lys Leu Ile Asp Ile
225 230 235 240
Leu Ala Ile Ile Ala Thr Val Phe Gly Thr Ala Cys Ser Leu Gly Leu
245 250 255
Gly Ala Leu Gln Ile Gly Ala Gly Leu Ser Ala Ala Asn Ile Ile Glu
260 265 270
Asn Pro Ser Asp Trp Thr Val Ile Gly Ile Val Ser Val Leu Thr Leu
275 280 285
Ala Phe Ile Phe Ser Ala Ile Ser Gly Val Gly Lys Gly Ile Gln Tyr
290 295 300
Leu Ser Asn Ala Asn Met Val Leu Ala Ala Leu Leu Ala Ile Phe Val
305 310 315 320
Phe Val Val Gly Pro Thr Val Ser Ile Leu Asn Leu Leu Pro Gly Ser
325 330 335
Ile Gly Asn Tyr Leu Ser Asn Phe Phe Gln Met Ala Gly Arg Thr Ala
340 345 350
Met Ser Ala Asp Gly Thr Ala Gly Glu Trp Leu Gly Ser Trp Thr Ile
355 360 365
Phe Tyr Trp Ala Trp Trp Ile Ser Trp Ser Pro Phe Val Gly Met Phe
370 375 380
Leu Ala Arg Ile Ser Arg Gly Arg Ser Ile Arg Glu Phe Ile Leu Gly
385 390 395 400
Val Leu Leu Val Pro Ala Gly Val Ser Thr Val Trp Phe Ser Ile Phe
405 410 415
Gly Gly Thr Ala Ile Val Phe Glu Gln Asn Gly Glu Ser Ile Trp Gly
420 425 430
Asp Gly Ala Ala Glu Glu Gln Leu Phe Gly Leu Leu His Ala Leu Pro
435 440 445
Gly Gly Gln Ile Met Gly Ile Ile Ala Met Ile Leu Leu Gly Thr Phe
450 455 460
Phe Ile Thr Ser Ala Asp Ser Ala Ser Thr Val Met Gly Thr Met Ser
465 470 475 480
Gln His Gly Gln Leu Glu Ala Asn Lys Trp Val Thr Ala Ala Trp Gly
485 490 495
Val Ala Thr Ala Ala Ile Gly Leu Thr Leu Leu Leu Ser Gly Gly Asp
500 505 510
Asn Ala Leu Asn Asn Leu Gln Lys Val Thr Ile Val Ala Ala Thr Pro
515 520 525
Phe Leu Phe Val Val Ile Gly Leu Met Phe Ala Leu Val Lys Asp Leu
530 535 540
Ser Asn Asp Val Ile Tyr Leu Glu Tyr Arg Glu Gln Gln Arg Phe Asn
545 550 555 560
Ala Arg Leu Ala Arg Glu Arg Arg Val His Asn Glu His Arg Lys Arg
565 570 575
Glu Leu Ala Ala Lys Arg Arg Arg Glu Arg Lys Ala Ser Gly Ala Gly
580 585 590
Lys Arg Arg
595
<210> 5
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 5
cagtgccaag cttgcatgcc tgcaggtcga ctctagtgtt tctgtcttga ccttgg 56
<210> 6
<211> 30
<212> ДНК
<213> Искусственная последовательность
<400> 6
agccacgatg gtgacttttt gcaagttgtt 30
<210> 7
<211> 30
<212> ДНК
<213> Искусственная последовательность
<400> 7
aacaacttgc aaaaagtcac catcgtggct 30
<210> 8
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 8
cagctatgac catgattacg aattcgagct cggtacccac agattgggca ggtgcc 56
<210> 9
<211> 19
<212> ДНК
<213> Искусственная последовательность
<400> 9
ctattgcttt ctggtggtg 19
<210> 10
<211> 20
<212> ДНК
<213> Искусственная последовательность
<400> 10
tcgccttacg ctccctgcgt 20
<210> 11
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 11
cagtgccaag cttgcatgcc tgcaggtcga ctctaggacc cgcttgccat acgaag 56
<210> 12
<211> 40
<212> ДНК
<213> Искусственная последовательность
<400> 12
gcatcacaat gacataacga atctactcat ctgaagaatc 40
<210> 13
<211> 40
<212> ДНК
<213> Искусственная последовательность
<400> 13
gattcttcag atgagtagat tcgttatgtc attgtgatgc 40
<210> 14
<211> 40
<212> ДНК
<213> Искусственная последовательность
<400> 14
caaaccagag tgcccacgaa ctatcgacgc ttccccgcgc 40
<210> 15
<211> 40
<212> ДНК
<213> Искусственная последовательность
<400> 15
gcgcggggaa gcgtcgatag ttcgtgggca ctctggtttg 40
<210> 16
<211> 58
<212> ДНК
<213> Искусственная последовательность
<400> 16
cagctatgac catgattacg aattcgagct cggtacccca taagaaacaa ccacttcc 58
<210> 17
<211> 20
<212> ДНК
<213> Искусственная последовательность
<400> 17
gtccaaggtg acggccgcac 20
<210> 18
<211> 20
<212> ДНК
<213> Искусственная последовательность
<400> 18
ctttttcacc aatgagtggc 20
<210> 19
<211> 20
<212> ДНК
<213> Искусственная последовательность
<400> 19
ccgtaaacag cttctaagct 20
<210> 20
<211> 20
<212> ДНК
<213> Искусственная последовательность
<400> 20
atattcggcc cagcagcagc 20
<210> 21
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 21
gcttgcatgc ctgcaggtcg actctagagg atcccctcgt tatgtcattg tgatgc 56
<210> 22
<211> 54
<212> ДНК
<213> Искусственная последовательность
<400> 22
atcaggctga aaatcttctc tcatccgcca aaacctatcg acgcttcccc gcgc 54
<210> 23
<211> 55
<212> ДНК
<213> Искусственная последовательность
<400> 23
cagtgccaag cttgcatgcc tgcaggtcga ctctagtaag gggcagttgg tctcg 55
<210> 24
<211> 40
<212> ДНК
<213> Искусственная последовательность
<400> 24
gtatcagggg ttaaaaattg cttaattttc cctggcagaa 40
<210> 25
<211> 40
<212> ДНК
<213> Искусственная последовательность
<400> 25
ttctgccagg gaaaattaag caatttttaa cccctgatac 40
<210> 26
<211> 58
<212> ДНК
<213> Искусственная последовательность
<400> 26
cagctatgac catgattacg aattcgagct cggtacccat atcgcgcgac attgcgcg 58
<210> 27
<211> 19
<212> ДНК
<213> Искусственная последовательность
<400> 27
taaggggcag ttggtctcg 19
<210> 28
<211> 20
<212> ДНК
<213> Искусственная последовательность
<400> 28
atatcgcgcg acattgcgcg 20
<210> 29
<211> 1521
<212> ДНК
<213> Искусственная последовательность
<400> 29
cagtgccaag cttgcatgcc tgcaggtcga ctctagtgtt tctgtcttga ccttggcatt 60
tatcttctct gctatttctg gtgtgggcaa gggaatccag tacctctcca acgccaacat 120
ggttctggca gctctgctcg cgattttcgt gttcgttgtc ggaccaaccg tgtcgatttt 180
gaacctgctg ccaggttcta ttggcaacta cctgtccaac ttcttccaaa tggcaggccg 240
cactgccatg agtgccgacg gcacagcagg tgagtggcta ggtagctgga ccatcttcta 300
ctgggcatgg tggatctctt ggtcaccatt cgtaggaatg ttcttggcac gtatttcccg 360
tggccgctcc atccgtgagt tcatcctggg cgtgttgctc gtcccagcag gtgtgtccac 420
cgtatggttc tccatttttg gtggcactgc gattgtcttc gaacaaaatg gggaatccat 480
ttggggtgat ggtgcagcag aagagcagct ctttggattg cttcatgcac ttccaggtgg 540
gcaaataatg ggcatcatcg ccatgatttt gctgggtact ttcttcatta cctctgcaga 600
ctctgcttcc accgtcatgg gcaccatgag tcagcacggc cagctggaag ccaacaagtg 660
ggtgacagct gcctggggtg ttgctaccgc agctattgga ctaacgctat tgctttctgg 720
tggtgacaat gccttgaaca acttgcaaaa agtcaccatc gtggctgcaa caccattcct 780
gtttgtggtt attggattga tgtttgcgtt agtcaaggac ttaagcaatg atgtgatcta 840
cctcgagtac cgtgagcagc aacgcttcaa cgcgcgcctt gcccgtgaac gtcgtgttca 900
caatgaacac cgcaagcgtg aactggctgc aaagcgacgc agggagcgta aggcgagtgg 960
cgcggggaag cgtcgatagg caatttttaa cccctgatac ccctttccgg ccgggcataa 1020
attaaggtgg tacgcatgac gaagaacgtg ctcgtatctg ttgcctggcc gtatgccaac 1080
ggaccccgtc acattggaca tgtggcgggg tttggtgtcc cctccgatgt gttcgcaagg 1140
ttccagcgaa tgtctggcaa caacgtgctc atggtgtccg gcaccgatga gcacggcacg 1200
ccacttctgg ttcaagcaga caaagaaggc gtcaccgttc aagacctcgc ggataagtac 1260
aaccgccaga tcgtcgaaga cctcaccggc ctgggcctgt cctatgacct tttcacccgc 1320
accaccacct ccaaccacta cgcagtagtg caggaactgt tccgtggtct gtacgacaac 1380
ggttacatga tcaaggaaac caccctcggt gcgatttccc catccactgg ccgtaccctg 1440
ccagaccgct acattgaagg cacctgccca atctgtgggt accgagctcg aattcgtaat 1500
catggtcata gctgtttcct g 1571
<210> 30
<211> 250
<212> ДНК
<213> Искусственная последовательность
<400> 30
ctattgcttt ctggtggtga caatgccttg aacaacttgc aaaacgtcac catcgtggct 60
gcaacaccat tcctgtttgt ggttattgga ttgatgtttg cgttagtcaa ggacttaagc 120
aatgatgtga tctacctcga gtaccgtgag cagcaacgct tcaacgcgcg ccttgcccgt 180
gaacgtcgtg ttcacaatga acaccgcaag cgtgaactgg ctgcaaagcg acgcagggag 240
cgtaaggcga 258
<210> 31
<211> 806
<212> ДНК
<213> Искусственная последовательность
<400> 31
cagtgccaag cttgcatgcc tgcaggtcga ctctaggacc cgcttgccat acgaagtcct 60
ggagaagatc tccacccgca tcaccaacga agttccagac gtaaaccgcg tggttttgga 120
cgtaacctcc aagccaccag gaaccatcga atgggagtag gccttaaatg agccttcgtt 180
aagcggcaat caccttatcg gtgattgccg ctttcccatt tctccgggtt ttctggaact 240
ttttgggcgt atgctgggaa tgatcttatt attttgattt cagaaagcag gagagaccag 300
atgagcgaaa tccttgaaac ctactgggca ccccacttcg gaaacaccga tgaagccgca 360
gcactcgttt catacttggc acaagcttcc ggtgatccta ttgaggttca caccctgttc 420
ggggatttag gtttagacgg actctctgga aactacaccg acactgagat cgacggctac 480
ggcgacgcat tcctgctggt tgcagcacta gcagtgttga tggctgaaaa caaagcatcc 540
ggcggcgtga atctgggtga agttggggga gctgataaat cgatccggct gcatgttgaa 600
tccaaggaaa acacccagat caacaccgca ttgaagtact ttgcgctttc cccagaagac 660
cacgcagcgg cagatcgctt cgatgaggat gacctgtctg agcttgccaa cttgagtgaa 720
gagctgcgcg gacagctgga ctaattgctg cccgtttaag gagtccgatt cttcagatga 780
gtagattcgt tatgtcattg tgatgc 832
<210> 32
<211> 1987
<212> ДНК
<213> Искусственная последовательность
<400> 32
gattcttcag atgagtagat tcgttatgtc attgtgatgc tcccgtgaac ataaacggga 60
cctactggct ttacttaagt aacagctatg taaaagacca ggtcaggttc gggtcggttt 120
aggtacgaaa cccatttttc ggtttgcttt ccaggtttcc ccaagtaaag gtgagtttta 180
tgactacatc tgacccaaat tcgaaaccga tagtggagga tgctcagcca gagcagatca 240
ccgcaaccga agaactggcg ggtttgcttg agaatccaac taacctggaa gggaaactgg 300
ccgacgccga agaggaaatt atcctcgaag gcgaagacgc ccaggcctca cttaactggt 360
cagtcatcgt tccagcccta gtcattgtcc tagcgacagt ggtgtggggt atcggattca 420
aagatagctt taccaacttt gctagttctg cgttgtcagc agtagttgac aatctcggct 480
gggccttcat tttgtttggc acagtctttg tattttttat cgttgttatc gccgctagta 540
aattcggcac gattcgctta ggccgcattg atgaagcacc agagtttcgc acggtgtcat 600
ggatttccat gatgtttgct gcaggtatgg gtattggttt gatgttctac ggaaccacag 660
aacctttaac cttctaccgc aatggtgtac ctggatatga tgaacacaat gttggcgttg 720
ctatgtccac gacaatgttc cactggacct tgcatccatg ggctatctac gcaattgtgg 780
gcctagccat tgcctattcg accttccgag tgggccgtaa acagcttcta agctctgcat 840
tcgtgccact cattggtgaa aaaggtgcag aaggatggtt gggcaagctc atcgacatcc 900
tggcgattat cgccacagta ttcggcaccg catgttccct tggcctgggt gcgctgcaga 960
tcggtgcagg actttccgca gccaacatca ttgaaaatcc gagtgactgg actgtcattg 1020
gtattgtttc tgtcttgacc ttggcattta tcttctctgc tatttctggt gtgggcaagg 1080
gaatccagta cctctccaac gccaacatgg ttctggcagc tctgctcgcg attttcgtgt 1140
tcgttgtcgg accaaccgtg tcgattttga acctgctgcc aggttctatt ggcaactacc 1200
tgtccaactt cttccaaatg gcaggccgca ctgccatgag tgccgacggc acagcaggtg 1260
agtggctagg tagctggacc atcttctact gggcatggtg gatctcttgg tcaccattcg 1320
taggaatgtt cttggcacgt atttcccgtg gccgctccat ccgtgagttc atcctgggcg 1380
tgttgctcgt cccagcaggt gtgtccaccg tatggttctc catttttggt ggcactgcga 1440
ttgtcttcga acaaaatggg gaatccattt ggggtgatgg tgcagcagaa gagcagctct 1500
ttggattgct tcatgcactt ccaggtgggc aaataatggg catcatcgcc atgattttgc 1560
tgggtacttt cttcattacc tctgcagact ctgcttccac cgtcatgggc accatgagtc 1620
agcacggcca gctggaagcc aacaagtggg tgacagctgc ctggggtgtt gctaccgcag 1680
ctattggact aacgctattg ctttctggtg gtgacaatgc cttgaacaac ttgcaaaacg 1740
tcaccatcgt ggctgcaaca ccattcctgt ttgtggttat tggattgatg tttgcgttag 1800
tcaaggactt aagcaatgat gtgatctacc tcgagtaccg tgagcagcaa cgcttcaacg 1860
cgcgccttgc ccgtgaacgt cgtgttcaca atgaacaccg caagcgtgaa ctggctgcaa 1920
agcgacgcag ggagcgtaag gcgagtggcg cggggaagcg tcgatagttc gtgggcactc 1980
tggtttg 2053
<210> 33
<211> 1987
<212> ДНК
<213> Искусственная последовательность
<400> 33
gattcttcag atgagtagat tcgttatgtc attgtgatgc tcccgtgaac ataaacggga 60
cctactggct ttacttaagt aacagctatg taaaagacca ggtcaggttc gggtcggttt 120
aggtacgaaa cccatttttc ggtttgcttt ccaggtttcc ccaagtaaag gtgagtttta 180
tgactacatc tgacccaaat tcgaaaccga tagtggagga tgctcagcca gagcagatca 240
ccgcaaccga agaactggcg ggtttgcttg agaatccaac taacctggaa gggaaactgg 300
ccgacgccga agaggaaatt atcctcgaag gcgaagacgc ccaggcctca cttaactggt 360
cagtcatcgt tccagcccta gtcattgtcc tagcgacagt ggtgtggggt atcggattca 420
aagatagctt taccaacttt gctagttctg cgttgtcagc agtagttgac aatctcggct 480
gggccttcat tttgtttggc acagtctttg tattttttat cgttgttatc gccgctagta 540
aattcggcac gattcgctta ggccgcattg atgaagcacc agagtttcgc acggtgtcat 600
ggatttccat gatgtttgct gcaggtatgg gtattggttt gatgttctac ggaaccacag 660
aacctttaac cttctaccgc aatggtgtac ctggatatga tgaacacaat gttggcgttg 720
ctatgtccac gacaatgttc cactggacct tgcatccatg ggctatctac gcaattgtgg 780
gcctagccat tgcctattcg accttccgag tgggccgtaa acagcttcta agctctgcat 840
tcgtgccact cattggtgaa aaaggtgcag aaggatggtt gggcaagctc atcgacatcc 900
tggcgattat cgccacagta ttcggcaccg catgttccct tggcctgggt gcgctgcaga 960
tcggtgcagg actttccgca gccaacatca ttgaaaatcc gagtgactgg actgtcattg 1020
gtattgtttc tgtcttgacc ttggcattta tcttctctgc tatttctggt gtgggcaagg 1080
gaatccagta cctctccaac gccaacatgg ttctggcagc tctgctcgcg attttcgtgt 1140
tcgttgtcgg accaaccgtg tcgattttga acctgctgcc aggttctatt ggcaactacc 1200
tgtccaactt cttccaaatg gcaggccgca ctgccatgag tgccgacggc acagcaggtg 1260
agtggctagg tagctggacc atcttctact gggcatggtg gatctcttgg tcaccattcg 1320
taggaatgtt cttggcacgt atttcccgtg gccgctccat ccgtgagttc atcctgggcg 1380
tgttgctcgt cccagcaggt gtgtccaccg tatggttctc catttttggt ggcactgcga 1440
ttgtcttcga acaaaatggg gaatccattt ggggtgatgg tgcagcagaa gagcagctct 1500
ttggattgct tcatgcactt ccaggtgggc aaataatggg catcatcgcc atgattttgc 1560
tgggtacttt cttcattacc tctgcagact ctgcttccac cgtcatgggc accatgagtc 1620
agcacggcca gctggaagcc aacaagtggg tgacagctgc ctggggtgtt gctaccgcag 1680
ctattggact aacgctattg ctttctggtg gtgacaatgc cttgaacaac ttgcaaaaag 1740
tcaccatcgt ggctgcaaca ccattcctgt ttgtggttat tggattgatg tttgcgttag 1800
tcaaggactt aagcaatgat gtgatctacc tcgagtaccg tgagcagcaa cgcttcaacg 1860
cgcgccttgc ccgtgaacgt cgtgttcaca atgaacaccg caagcgtgaa ctggctgcaa 1920
agcgacgcag ggagcgtaag gcgagtggcg cggggaagcg tcgatagttc gtgggcactc 1980
tggtttg 2053
<210> 34
<211> 788
<212> ДНК
<213> Искусственная последовательность
<400> 34
gcgcggggaa gcgtcgatag ttcgtgggca ctctggtttg gttaccagga tgggttagtc 60
attctgatca gcgaattcca cgttcacatc gccaattcca gagttcacaa ccagattcag 120
cattggacct tctagatcag cattgtgggc ggtgagatct ccaacatcac agcgcgctgt 180
gcccacaccg gcggtacaac ttaggctcac gggcacatca tcgggcaggg tgaccatgac 240
ttcgccgatc cctgaggtga tttggatgtt ttgttcctga tccaattggg tgaggtggct 300
gaaatcgagg ttcatttcac ccacgccaga ggtgtagctg ctgaggagtt catcgttggt 360
ggggatgaga ttgacatcgc cgattccagg gtcgtcttca aagtagatgg gatcgatatt 420
tgaaataaac aggcctgcga gggcgctcat gacaactccg gtaccaacta caccgccgac 480
aatccatggc cacacatggc gctttttctg aggcttttgt ggagggactt gtacatccca 540
ggtgttgtat tggttttggg caagtggatc ccaatgaggc gcttcggggg tttgttgcgc 600
gaagggtgca tagtagccct caacgggggt gatagtgctt agatctggtt ggggttgtgg 660
gtagagatct tcgtttttca tggtggcatc ctcagaaaca gtgaattcag tggtgagtag 720
tccgcggggt ggaagtggtt gtttcttatg gggtaccgag ctcgaattcg taatcatggt 780
catagctg 814
<210> 35
<211> 3501
<212> ДНК
<213> Искусственная последовательность
<400> 35
cagtgccaag cttgcatgcc tgcaggtcga ctctaggacc cgcttgccat acgaagtcct 60
ggagaagatc tccacccgca tcaccaacga agttccagac gtaaaccgcg tggttttgga 120
cgtaacctcc aagccaccag gaaccatcga atgggagtag gccttaaatg agccttcgtt 180
aagcggcaat caccttatcg gtgattgccg ctttcccatt tctccgggtt ttctggaact 240
ttttgggcgt atgctgggaa tgatcttatt attttgattt cagaaagcag gagagaccag 300
atgagcgaaa tccttgaaac ctactgggca ccccacttcg gaaacaccga tgaagccgca 360
gcactcgttt catacttggc acaagcttcc ggtgatccta ttgaggttca caccctgttc 420
ggggatttag gtttagacgg actctctgga aactacaccg acactgagat cgacggctac 480
ggcgacgcat tcctgctggt tgcagcacta gcagtgttga tggctgaaaa caaagcatcc 540
ggcggcgtga atctgggtga agttggggga gctgataaat cgatccggct gcatgttgaa 600
tccaaggaaa acacccagat caacaccgca ttgaagtact ttgcgctttc cccagaagac 660
cacgcagcgg cagatcgctt cgatgaggat gacctgtctg agcttgccaa cttgagtgaa 720
gagctgcgcg gacagctgga ctaattgctg cccgtttaag gagtccgatt cttcagatga 780
gtagattcgt tatgtcattg tgatgctccc gtgaacataa acgggaccta ctggctttac 840
ttaagtaaca gctatgtaaa agaccaggtc aggttcgggt cggtttaggt acgaaaccca 900
tttttcggtt tgctttccag gtttccccaa gtaaaggtga gttttatgac tacatctgac 960
ccaaattcga aaccgatagt ggaggatgct cagccagagc agatcaccgc aaccgaagaa 1020
ctggcgggtt tgcttgagaa tccaactaac ctggaaggga aactggccga cgccgaagag 1080
gaaattatcc tcgaaggcga agacgcccag gcctcactta actggtcagt catcgttcca 1140
gccctagtca ttgtcctagc gacagtggtg tggggtatcg gattcaaaga tagctttacc 1200
aactttgcta gttctgcgtt gtcagcagta gttgacaatc tcggctgggc cttcattttg 1260
tttggcacag tctttgtatt ttttatcgtt gttatcgccg ctagtaaatt cggcacgatt 1320
cgcttaggcc gcattgatga agcaccagag tttcgcacgg tgtcatggat ttccatgatg 1380
tttgctgcag gtatgggtat tggtttgatg ttctacggaa ccacagaacc tttaaccttc 1440
taccgcaatg gtgtacctgg atatgatgaa cacaatgttg gcgttgctat gtccacgaca 1500
atgttccact ggaccttgca tccatgggct atctacgcaa ttgtgggcct agccattgcc 1560
tattcgacct tccgagtggg ccgtaaacag cttctaagct ctgcattcgt gccactcatt 1620
ggtgaaaaag gtgcagaagg atggttgggc aagctcatcg acatcctggc gattatcgcc 1680
acagtattcg gcaccgcatg ttcccttggc ctgggtgcgc tgcagatcgg tgcaggactt 1740
tccgcagcca acatcattga aaatccgagt gactggactg tcattggtat tgtttctgtc 1800
ttgaccttgg catttatctt ctctgctatt tctggtgtgg gcaagggaat ccagtacctc 1860
tccaacgcca acatggttct ggcagctctg ctcgcgattt tcgtgttcgt tgtcggacca 1920
accgtgtcga ttttgaacct gctgccaggt tctattggca actacctgtc caacttcttc 1980
caaatggcag gccgcactgc catgagtgcc gacggcacag caggtgagtg gctaggtagc 2040
tggaccatct tctactgggc atggtggatc tcttggtcac cattcgtagg aatgttcttg 2100
gcacgtattt cccgtggccg ctccatccgt gagttcatcc tgggcgtgtt gctcgtccca 2160
gcaggtgtgt ccaccgtatg gttctccatt tttggtggca ctgcgattgt cttcgaacaa 2220
aatggggaat ccatttgggg tgatggtgca gcagaagagc agctctttgg attgcttcat 2280
gcacttccag gtgggcaaat aatgggcatc atcgccatga ttttgctggg tactttcttc 2340
attacctctg cagactctgc ttccaccgtc atgggcacca tgagtcagca cggccagctg 2400
gaagccaaca agtgggtgac agctgcctgg ggtgttgcta ccgcagctat tggactaacg 2460
ctattgcttt ctggtggtga caatgccttg aacaacttgc aaaacgtcac catcgtggct 2520
gcaacaccat tcctgtttgt ggttattgga ttgatgtttg cgttagtcaa ggacttaagc 2580
aatgatgtga tctacctcga gtaccgtgag cagcaacgct tcaacgcgcg ccttgcccgt 2640
gaacgtcgtg ttcacaatga acaccgcaag cgtgaactgg ctgcaaagcg acgcagggag 2700
cgtaaggcga gtggcgcggg gaagcgtcga tagttcgtgg gcactctggt ttggttacca 2760
ggatgggtta gtcattctga tcagcgaatt ccacgttcac atcgccaatt ccagagttca 2820
caaccagatt cagcattgga ccttctagat cagcattgtg ggcggtgaga tctccaacat 2880
cacagcgcgc tgtgcccaca ccggcggtac aacttaggct cacgggcaca tcatcgggca 2940
gggtgaccat gacttcgccg atccctgagg tgatttggat gttttgttcc tgatccaatt 3000
gggtgaggtg gctgaaatcg aggttcattt cacccacgcc agaggtgtag ctgctgagga 3060
gttcatcgtt ggtggggatg agattgacat cgccgattcc agggtcgtct tcaaagtaga 3120
tgggatcgat atttgaaata aacaggcctg cgagggcgct catgacaact ccggtaccaa 3180
ctacaccgcc gacaatccat ggccacacat ggcgcttttt ctgaggcttt tgtggaggga 3240
cttgtacatc ccaggtgttg tattggtttt gggcaagtgg atcccaatga ggcgcttcgg 3300
gggtttgttg cgcgaagggt gcatagtagc cctcaacggg ggtgatagtg cttagatctg 3360
gttggggttg tgggtagaga tcttcgtttt tcatggtggc atcctcagaa acagtgaatt 3420
cagtggtgag tagtccgcgg ggtggaagtg gttgtttctt atggggtacc gagctcgaat 3480
tcgtaatcat ggtcatagct g 3617
<210> 36
<211> 3501
<212> ДНК
<213> Искусственная последовательность
<400> 36
cagtgccaag cttgcatgcc tgcaggtcga ctctaggacc cgcttgccat acgaagtcct 60
ggagaagatc tccacccgca tcaccaacga agttccagac gtaaaccgcg tggttttgga 120
cgtaacctcc aagccaccag gaaccatcga atgggagtag gccttaaatg agccttcgtt 180
aagcggcaat caccttatcg gtgattgccg ctttcccatt tctccgggtt ttctggaact 240
ttttgggcgt atgctgggaa tgatcttatt attttgattt cagaaagcag gagagaccag 300
atgagcgaaa tccttgaaac ctactgggca ccccacttcg gaaacaccga tgaagccgca 360
gcactcgttt catacttggc acaagcttcc ggtgatccta ttgaggttca caccctgttc 420
ggggatttag gtttagacgg actctctgga aactacaccg acactgagat cgacggctac 480
ggcgacgcat tcctgctggt tgcagcacta gcagtgttga tggctgaaaa caaagcatcc 540
ggcggcgtga atctgggtga agttggggga gctgataaat cgatccggct gcatgttgaa 600
tccaaggaaa acacccagat caacaccgca ttgaagtact ttgcgctttc cccagaagac 660
cacgcagcgg cagatcgctt cgatgaggat gacctgtctg agcttgccaa cttgagtgaa 720
gagctgcgcg gacagctgga ctaattgctg cccgtttaag gagtccgatt cttcagatga 780
gtagattcgt tatgtcattg tgatgctccc gtgaacataa acgggaccta ctggctttac 840
ttaagtaaca gctatgtaaa agaccaggtc aggttcgggt cggtttaggt acgaaaccca 900
tttttcggtt tgctttccag gtttccccaa gtaaaggtga gttttatgac tacatctgac 960
ccaaattcga aaccgatagt ggaggatgct cagccagagc agatcaccgc aaccgaagaa 1020
ctggcgggtt tgcttgagaa tccaactaac ctggaaggga aactggccga cgccgaagag 1080
gaaattatcc tcgaaggcga agacgcccag gcctcactta actggtcagt catcgttcca 1140
gccctagtca ttgtcctagc gacagtggtg tggggtatcg gattcaaaga tagctttacc 1200
aactttgcta gttctgcgtt gtcagcagta gttgacaatc tcggctgggc cttcattttg 1260
tttggcacag tctttgtatt ttttatcgtt gttatcgccg ctagtaaatt cggcacgatt 1320
cgcttaggcc gcattgatga agcaccagag tttcgcacgg tgtcatggat ttccatgatg 1380
tttgctgcag gtatgggtat tggtttgatg ttctacggaa ccacagaacc tttaaccttc 1440
taccgcaatg gtgtacctgg atatgatgaa cacaatgttg gcgttgctat gtccacgaca 1500
atgttccact ggaccttgca tccatgggct atctacgcaa ttgtgggcct agccattgcc 1560
tattcgacct tccgagtggg ccgtaaacag cttctaagct ctgcattcgt gccactcatt 1620
ggtgaaaaag gtgcagaagg atggttgggc aagctcatcg acatcctggc gattatcgcc 1680
acagtattcg gcaccgcatg ttcccttggc ctgggtgcgc tgcagatcgg tgcaggactt 1740
tccgcagcca acatcattga aaatccgagt gactggactg tcattggtat tgtttctgtc 1800
ttgaccttgg catttatctt ctctgctatt tctggtgtgg gcaagggaat ccagtacctc 1860
tccaacgcca acatggttct ggcagctctg ctcgcgattt tcgtgttcgt tgtcggacca 1920
accgtgtcga ttttgaacct gctgccaggt tctattggca actacctgtc caacttcttc 1980
caaatggcag gccgcactgc catgagtgcc gacggcacag caggtgagtg gctaggtagc 2040
tggaccatct tctactgggc atggtggatc tcttggtcac cattcgtagg aatgttcttg 2100
gcacgtattt cccgtggccg ctccatccgt gagttcatcc tgggcgtgtt gctcgtccca 2160
gcaggtgtgt ccaccgtatg gttctccatt tttggtggca ctgcgattgt cttcgaacaa 2220
aatggggaat ccatttgggg tgatggtgca gcagaagagc agctctttgg attgcttcat 2280
gcacttccag gtgggcaaat aatgggcatc atcgccatga ttttgctggg tactttcttc 2340
attacctctg cagactctgc ttccaccgtc atgggcacca tgagtcagca cggccagctg 2400
gaagccaaca agtgggtgac agctgcctgg ggtgttgcta ccgcagctat tggactaacg 2460
ctattgcttt ctggtggtga caatgccttg aacaacttgc aaaaagtcac catcgtggct 2520
gcaacaccat tcctgtttgt ggttattgga ttgatgtttg cgttagtcaa ggacttaagc 2580
aatgatgtga tctacctcga gtaccgtgag cagcaacgct tcaacgcgcg ccttgcccgt 2640
gaacgtcgtg ttcacaatga acaccgcaag cgtgaactgg ctgcaaagcg acgcagggag 2700
cgtaaggcga gtggcgcggg gaagcgtcga tagttcgtgg gcactctggt ttggttacca 2760
ggatgggtta gtcattctga tcagcgaatt ccacgttcac atcgccaatt ccagagttca 2820
caaccagatt cagcattgga ccttctagat cagcattgtg ggcggtgaga tctccaacat 2880
cacagcgcgc tgtgcccaca ccggcggtac aacttaggct cacgggcaca tcatcgggca 2940
gggtgaccat gacttcgccg atccctgagg tgatttggat gttttgttcc tgatccaatt 3000
gggtgaggtg gctgaaatcg aggttcattt cacccacgcc agaggtgtag ctgctgagga 3060
gttcatcgtt ggtggggatg agattgacat cgccgattcc agggtcgtct tcaaagtaga 3120
tgggatcgat atttgaaata aacaggcctg cgagggcgct catgacaact ccggtaccaa 3180
ctacaccgcc gacaatccat ggccacacat ggcgcttttt ctgaggcttt tgtggaggga 3240
cttgtacatc ccaggtgttg tattggtttt gggcaagtgg atcccaatga ggcgcttcgg 3300
gggtttgttg cgcgaagggt gcatagtagc cctcaacggg ggtgatagtg cttagatctg 3360
gttggggttg tgggtagaga tcttcgtttt tcatggtggc atcctcagaa acagtgaatt 3420
cagtggtgag tagtccgcgg ggtggaagtg gttgtttctt atggggtacc gagctcgaat 3480
tcgtaatcat ggtcatagct g 3617
<210> 37
<211> 1674
<212> ДНК
<213> Искусственная последовательность
<400> 37
gtccaaggtg acggccgcac ctacggacac ccaatcgtgc tgcgcccagt atcttccgaa 60
gacgcaatga ccgccgactg gacccgcttg ccatacgaag tcctggagaa gatctccacc 120
cgcatcacca acgaagttcc agacgtaaac cgcgtggttt tggacgtaac ctccaagcca 180
ccaggaacca tcgaatggga gtaggcctta aatgagcctt cgttaagcgg caatcacctt 240
atcggtgatt gccgctttcc catttctccg ggttttctgg aactttttgg gcgtatgctg 300
ggaatgatct tattattttg atttcagaaa gcaggagaga ccagatgagc gaaatccttg 360
aaacctactg ggcaccccac ttcggaaaca ccgatgaagc cgcagcactc gtttcatact 420
tggcacaagc ttccggtgat cctattgagg ttcacaccct gttcggggat ttaggtttag 480
acggactctc tggaaactac accgacactg agatcgacgg ctacggcgac gcattcctgc 540
tggttgcagc actagcagtg ttgatggctg aaaacaaagc atccggcggc gtgaatctgg 600
gtgaagttgg gggagctgat aaatcgatcc ggctgcatgt tgaatccaag gaaaacaccc 660
agatcaacac cgcattgaag tactttgcgc tttccccaga agaccacgca gcggcagatc 720
gcttcgatga ggatgacctg tctgagcttg ccaacttgag tgaagagctg cgcggacagc 780
tggactaatt gctgcccgtt taaggagtcc gattcttcag atgagtagat tcgttatgtc 840
attgtgatgc tcccgtgaac ataaacggga cctactggct ttacttaagt aacagctatg 900
taaaagacca ggtcaggttc gggtcggttt aggtacgaaa cccatttttc ggtttgcttt 960
ccaggtttcc ccaagtaaag gtgagtttta tgactacatc tgacccaaat tcgaaaccga 1020
tagtggagga tgctcagcca gagcagatca ccgcaaccga agaactggcg ggtttgcttg 1080
agaatccaac taacctggaa gggaaactgg ccgacgccga agaggaaatt atcctcgaag 1140
gcgaagacgc ccaggcctca cttaactggt cagtcatcgt tccagcccta gtcattgtcc 1200
tagcgacagt ggtgtggggt atcggattca aagatagctt taccaacttt gctagttctg 1260
cgttgtcagc agtagttgac aatctcggct gggccttcat tttgtttggc acagtctttg 1320
tattttttat cgttgttatc gccgctagta aattcggcac gattcgctta ggccgcattg 1380
atgaagcacc agagtttcgc acggtgtcat ggatttccat gatgtttgct gcaggtatgg 1440
gtattggttt gatgttctac ggaaccacag aacctttaac cttctaccgc aatggtgtac 1500
ctggatatga tgaacacaat gttggcgttg ctatgtccac gacaatgttc cactggacct 1560
tgcatccatg ggctatctac gcaattgtgg gcctagccat tgcctattcg accttccgag 1620
tgggccgtaa acagcttcta agctctgcat tcgtgccact cattggtgaa aaag 1728
<210> 38
<211> 1943
<212> ДНК
<213> Искусственная последовательность
<400> 38
ccgtaaacag cttctaagct ctgcattcgt gccactcatt ggtgaaaaag gtgcagaagg 60
atggttgggc aagctcatcg acatcctggc gattatcgcc acagtattcg gcaccgcatg 120
ttcccttggc ctgggtgcgc tgcagatcgg tgcaggactt tccgcagcca acatcattga 180
aaatccgagt gactggactg tcattggtat tgtttctgtc ttgaccttgg catttatctt 240
ctctgctatt tctggtgtgg gcaagggaat ccagtacctc tccaacgcca acatggttct 300
ggcagctctg ctcgcgattt tcgtgttcgt tgtcggacca accgtgtcga ttttgaacct 360
gctgccaggt tctattggca actacctgtc caacttcttc caaatggcag gccgcactgc 420
catgagtgcc gacggcacag caggtgagtg gctaggtagc tggaccatct tctactgggc 480
atggtggatc tcttggtcac cattcgtagg aatgttcttg gcacgtattt cccgtggccg 540
ctccatccgt gagttcatcc tgggcgtgtt gctcgtccca gcaggtgtgt ccaccgtatg 600
gttctccatt tttggtggca ctgcgattgt cttcgaacaa aatggggaat ccatttgggg 660
tgatggtgca gcagaagagc agctctttgg attgcttcat gcacttccag gtgggcaaat 720
aatgggcatc atcgccatga ttttgctggg tactttcttc attacctctg cagactctgc 780
ttccaccgtc atgggcacca tgagtcagca cggccagctg gaagccaaca agtgggtgac 840
agctgcctgg ggtgttgcta ccgcagctat tggactaacg ctattgcttt ctggtggtga 900
caatgccttg aacaacttgc aaaacgtcac catcgtggct gcaacaccat tcctgtttgt 960
ggttattgga ttgatgtttg cgttagtcaa ggacttaagc aatgatgtga tctacctcga 1020
gtaccgtgag cagcaacgct tcaacgcgcg ccttgcccgt gaacgtcgtg ttcacaatga 1080
acaccgcaag cgtgaactgg ctgcaaagcg acgcagggag cgtaaggcga gtggcgcggg 1140
gaagcgtcga tagttcgtgg gcactctggt ttggttacca ggatgggtta gtcattctga 1200
tcagcgaatt ccacgttcac atcgccaatt ccagagttca caaccagatt cagcattgga 1260
ccttctagat cagcattgtg ggcggtgaga tctccaacat cacagcgcgc tgtgcccaca 1320
ccggcggtac aacttaggct cacgggcaca tcatcgggca gggtgaccat gacttcgccg 1380
atccctgagg tgatttggat gttttgttcc tgatccaatt gggtgaggtg gctgaaatcg 1440
aggttcattt cacccacgcc agaggtgtag ctgctgagga gttcatcgtt ggtggggatg 1500
agattgacat cgccgattcc agggtcgtct tcaaagtaga tgggatcgat atttgaaata 1560
aacaggcctg cgagggcgct catgacaact ccggtaccaa ctacaccgcc gacaatccat 1620
ggccacacat ggcgcttttt ctgaggcttt tgtggaggga cttgtacatc ccaggtgttg 1680
tattggtttt gggcaagtgg atcccaatga ggcgcttcgg gggtttgttg cgcgaagggt 1740
gcatagtagc cctcaacggg ggtgatagtg cttagatctg gttggggttg tgggtagaga 1800
tcttcgtttt tcatggtggc atcctcagaa acagtgaatt cagtggtgag tagtccgcgg 1860
ggtggaagtg gttgtttctt atgcaacgcc caccacatgg ctaaaaggca aaggtaagta 1920
atggctgctg ctgggccgaa tat 2007
<210> 39
<211> 2017
<212> ДНК
<213> Искусственная последовательность
<400> 39
gcttgcatgc ctgcaggtcg actctagagg atcccctcgt tatgtcattg tgatgctccc 60
gtgaacataa acgggaccta ctggctttac ttaagtaaca gctatgtaaa agaccaggtc 120
aggttcgggt cggtttaggt acgaaaccca tttttcggtt tgctttccag gtttccccaa 180
gtaaaggtga gttttatgac tacatctgac ccaaattcga aaccgatagt ggaggatgct 240
cagccagagc agatcaccgc aaccgaagaa ctggcgggtt tgcttgagaa tccaactaac 300
ctggaaggga aactggccga cgccgaagag gaaattatcc tcgaaggcga agacgcccag 360
gcctcactta actggtcagt catcgttcca gccctagtca ttgtcctagc gacagtggtg 420
tggggtatcg gattcaaaga tagctttacc aactttgcta gttctgcgtt gtcagcagta 480
gttgacaatc tcggctgggc cttcattttg tttggcacag tctttgtatt ttttatcgtt 540
gttatcgccg ctagtaaatt cggcacgatt cgcttaggcc gcattgatga agcaccagag 600
tttcgcacgg tgtcatggat ttccatgatg tttgctgcag gtatgggtat tggtttgatg 660
ttctacggaa ccacagaacc tttaaccttc taccgcaatg gtgtacctgg atatgatgaa 720
cacaatgttg gcgttgctat gtccacgaca atgttccact ggaccttgca tccatgggct 780
atctacgcaa ttgtgggcct agccattgcc tattcgacct tccgagtggg ccgtaaacag 840
cttctaagct ctgcattcgt gccactcatt ggtgaaaaag gtgcagaagg atggttgggc 900
aagctcatcg acatcctggc gattatcgcc acagtattcg gcaccgcatg ttcccttggc 960
ctgggtgcgc tgcagatcgg tgcaggactt tccgcagcca acatcattga aaatccgagt 1020
gactggactg tcattggtat tgtttctgtc ttgaccttgg catttatctt ctctgctatt 1080
tctggtgtgg gcaagggaat ccagtacctc tccaacgcca acatggttct ggcagctctg 1140
ctcgcgattt tcgtgttcgt tgtcggacca accgtgtcga ttttgaacct gctgccaggt 1200
tctattggca actacctgtc caacttcttc caaatggcag gccgcactgc catgagtgcc 1260
gacggcacag caggtgagtg gctaggtagc tggaccatct tctactgggc atggtggatc 1320
tcttggtcac cattcgtagg aatgttcttg gcacgtattt cccgtggccg ctccatccgt 1380
gagttcatcc tgggcgtgtt gctcgtccca gcaggtgtgt ccaccgtatg gttctccatt 1440
tttggtggca ctgcgattgt cttcgaacaa aatggggaat ccatttgggg tgatggtgca 1500
gcagaagagc agctctttgg attgcttcat gcacttccag gtgggcaaat aatgggcatc 1560
atcgccatga ttttgctggg tactttcttc attacctctg cagactctgc ttccaccgtc 1620
atgggcacca tgagtcagca cggccagctg gaagccaaca agtgggtgac agctgcctgg 1680
ggtgttgcta ccgcagctat tggactaacg ctattgcttt ctggtggtga caatgccttg 1740
aacaacttgc aaaacgtcac catcgtggct gcaacaccat tcctgtttgt ggttattgga 1800
ttgatgtttg cgttagtcaa ggacttaagc aatgatgtga tctacctcga gtaccgtgag 1860
cagcaacgct tcaacgcgcg ccttgcccgt gaacgtcgtg ttcacaatga acaccgcaag 1920
cgtgaactgg ctgcaaagcg acgcagggag cgtaaggcga gtggcgcggg gaagcgtcga 1980
taggttttgg cggatgagag aagattttca gcctgat 2083
<210> 40
<211> 2017
<212> ДНК
<213> Искусственная последовательность
<400> 40
gcttgcatgc ctgcaggtcg actctagagg atcccctcgt tatgtcattg tgatgctccc 60
gtgaacataa acgggaccta ctggctttac ttaagtaaca gctatgtaaa agaccaggtc 120
aggttcgggt cggtttaggt acgaaaccca tttttcggtt tgctttccag gtttccccaa 180
gtaaaggtga gttttatgac tacatctgac ccaaattcga aaccgatagt ggaggatgct 240
cagccagagc agatcaccgc aaccgaagaa ctggcgggtt tgcttgagaa tccaactaac 300
ctggaaggga aactggccga cgccgaagag gaaattatcc tcgaaggcga agacgcccag 360
gcctcactta actggtcagt catcgttcca gccctagtca ttgtcctagc gacagtggtg 420
tggggtatcg gattcaaaga tagctttacc aactttgcta gttctgcgtt gtcagcagta 480
gttgacaatc tcggctgggc cttcattttg tttggcacag tctttgtatt ttttatcgtt 540
gttatcgccg ctagtaaatt cggcacgatt cgcttaggcc gcattgatga agcaccagag 600
tttcgcacgg tgtcatggat ttccatgatg tttgctgcag gtatgggtat tggtttgatg 660
ttctacggaa ccacagaacc tttaaccttc taccgcaatg gtgtacctgg atatgatgaa 720
cacaatgttg gcgttgctat gtccacgaca atgttccact ggaccttgca tccatgggct 780
atctacgcaa ttgtgggcct agccattgcc tattcgacct tccgagtggg ccgtaaacag 840
cttctaagct ctgcattcgt gccactcatt ggtgaaaaag gtgcagaagg atggttgggc 900
aagctcatcg acatcctggc gattatcgcc acagtattcg gcaccgcatg ttcccttggc 960
ctgggtgcgc tgcagatcgg tgcaggactt tccgcagcca acatcattga aaatccgagt 1020
gactggactg tcattggtat tgtttctgtc ttgaccttgg catttatctt ctctgctatt 1080
tctggtgtgg gcaagggaat ccagtacctc tccaacgcca acatggttct ggcagctctg 1140
ctcgcgattt tcgtgttcgt tgtcggacca accgtgtcga ttttgaacct gctgccaggt 1200
tctattggca actacctgtc caacttcttc caaatggcag gccgcactgc catgagtgcc 1260
gacggcacag caggtgagtg gctaggtagc tggaccatct tctactgggc atggtggatc 1320
tcttggtcac cattcgtagg aatgttcttg gcacgtattt cccgtggccg ctccatccgt 1380
gagttcatcc tgggcgtgtt gctcgtccca gcaggtgtgt ccaccgtatg gttctccatt 1440
tttggtggca ctgcgattgt cttcgaacaa aatggggaat ccatttgggg tgatggtgca 1500
gcagaagagc agctctttgg attgcttcat gcacttccag gtgggcaaat aatgggcatc 1560
atcgccatga ttttgctggg tactttcttc attacctctg cagactctgc ttccaccgtc 1620
atgggcacca tgagtcagca cggccagctg gaagccaaca agtgggtgac agctgcctgg 1680
ggtgttgcta ccgcagctat tggactaacg ctattgcttt ctggtggtga caatgccttg 1740
aacaacttgc aaaaagtcac catcgtggct gcaacaccat tcctgtttgt ggttattgga 1800
ttgatgtttg cgttagtcaa ggacttaagc aatgatgtga tctacctcga gtaccgtgag 1860
cagcaacgct tcaacgcgcg ccttgcccgt gaacgtcgtg ttcacaatga acaccgcaag 1920
cgtgaactgg ctgcaaagcg acgcagggag cgtaaggcga gtggcgcggg gaagcgtcga 1980
taggttttgg cggatgagag aagattttca gcctgat 2083
<210> 41
<211> 2056
<212> ДНК
<213> Искусственная последовательность
<400> 41
agcggataac aatttcacac aggaaacaga attaattaag cttgcatgcc tgcaggtcga 60
ctctagagga tcccctcgtt atgtcattgt gatgctcccg tgaacataaa cgggacctac 120
tggctttact taagtaacag ctatgtaaaa gaccaggtca ggttcgggtc ggtttaggta 180
cgaaacccat ttttcggttt gctttccagg tttccccaag taaaggtgag ttttatgact 240
acatctgacc caaattcgaa accgatagtg gaggatgctc agccagagca gatcaccgca 300
accgaagaac tggcgggttt gcttgagaat ccaactaacc tggaagggaa actggccgac 360
gccgaagagg aaattatcct cgaaggcgaa gacgcccagg cctcacttaa ctggtcagtc 420
atcgttccag ccctagtcat tgtcctagcg acagtggtgt ggggtatcgg attcaaagat 480
agctttacca actttgctag ttctgcgttg tcagcagtag ttgacaatct cggctgggcc 540
ttcattttgt ttggcacagt ctttgtattt tttatcgttg ttatcgccgc tagtaaattc 600
ggcacgattc gcttaggccg cattgatgaa gcaccagagt ttcgcacggt gtcatggatt 660
tccatgatgt ttgctgcagg tatgggtatt ggtttgatgt tctacggaac cacagaacct 720
ttaaccttct accgcaatgg tgtacctgga tatgatgaac acaatgttgg cgttgctatg 780
tccacgacaa tgttccactg gaccttgcat ccatgggcta tctacgcaat tgtgggccta 840
gccattgcct attcgacctt ccgagtgggc cgtaaacagc ttctaagctc tgcattcgtg 900
ccactcattg gtgaaaaagg tgcagaagga tggttgggca agctcatcga catcctggcg 960
attatcgcca cagtattcgg caccgcatgt tcccttggcc tgggtgcgct gcagatcggt 1020
gcaggacttt ccgcagccaa catcattgaa aatccgagtg actggactgt cattggtatt 1080
gtttctgtct tgaccttggc atttatcttc tctgctattt ctggtgtggg caagggaatc 1140
cagtacctct ccaacgccaa catggttctg gcagctctgc tcgcgatttt cgtgttcgtt 1200
gtcggaccaa ccgtgtcgat tttgaacctg ctgccaggtt ctattggcaa ctacctgtcc 1260
aacttcttcc aaatggcagg ccgcactgcc atgagtgccg acggcacagc aggtgagtgg 1320
ctaggtagct ggaccatctt ctactgggca tggtggatct cttggtcacc attcgtagga 1380
atgttcttgg cacgtatttc ccgtggccgc tccatccgtg agttcatcct gggcgtgttg 1440
ctcgtcccag caggtgtgtc caccgtatgg ttctccattt ttggtggcac tgcgattgtc 1500
ttcgaacaaa atggggaatc catttggggt gatggtgcag cagaagagca gctctttgga 1560
ttgcttcatg cacttccagg tgggcaaata atgggcatca tcgccatgat tttgctgggt 1620
actttcttca ttacctctgc agactctgct tccaccgtca tgggcaccat gagtcagcac 1680
ggccagctgg aagccaacaa gtgggtgaca gctgcctggg gtgttgctac cgcagctatt 1740
ggactaacgc tattgctttc tggtggtgac aatgccttga acaacttgca aaacgtcacc 1800
atcgtggctg caacaccatt cctgtttgtg gttattggat tgatgtttgc gttagtcaag 1860
gacttaagca atgatgtgat ctacctcgag taccgtgagc agcaacgctt caacgcgcgc 1920
cttgcccgtg aacgtcgtgt tcacaatgaa caccgcaagc gtgaactggc tgcaaagcga 1980
cgcagggagc gtaaggcgag tggcgcgggg aagcgtcgat aggttttggc ggatgagaga 2040
agattttcag cctgat 2124
<210> 42
<211> 733
<212> ДНК
<213> Искусственная последовательность
<400> 42
cagtgccaag cttgcatgcc tgcaggtcga ctctagtaag gggcagttgg tctcgaactt 60
tgaccataat gaacaggcgc gcgtcggcaa gcttattgaa gccgcacgca cgggcacggt 120
gctggtggtc agcgatgccg gcatgcctgt ggtttctgat ccgggttttg cgcttatcga 180
cgccgcccac gaggcgaaca ttccggtcac ctgcttcccc gggccgtcag ctgtgccaac 240
tgcgttggca ttgtcgggcc ttcacgtggg ccgcttcgcc ttcgacggtt tcgcgccgcg 300
caaacaaggt gcgcgcacca cgtggttgga gtcgttgaaa accgaaaagc gcgcggtatg 360
tttcttcgaa tctcctcacc gcatcgcaga aaccctggct cacgcagccg aagttttagg 420
tgaacgacgc gcagcagtgt gccgtgaact gtccaaaacc tacgaacagg taaagcgtgg 480
aaccttgcca gagttggcag aatgggcaca agatggggtg cgtggcgaga tcaccgttgt 540
catcgaaggc gcgggcgata tcgcggtcga tgtcgattcg cttatcgacg ccgcccagca 600
gcgcgtcgat tccggcgagc ggttgaaagc ggtgtgcgca gacctcgcga aaatccatgg 660
cgtgagcaaa aatgaactct acgatgcggt tatttctgcc agggaaaatt aagcaatttt 720
taacccctga tac 757
<210> 43
<211> 814
<212> ДНК
<213> Искусственная последовательность
<400> 43
ttctgccagg gaaaattaag caatttttaa cccctgatac ccctttccgg ccgggcataa 60
attaaggtgg tacgcatgac gaagaacgtg ctcgtatctg ttgcctggcc gtatgccaac 120
ggaccccgtc acattggaca tgtggcgggg tttggtgtcc cctccgatgt gttcgcaagg 180
ttccagcgaa tgtctggcaa caacgtgctc atggtgtccg gcaccgatga gcacggcacg 240
ccacttctgg ttcaagcaga caaagaaggc gtcaccgttc aagacctcgc ggataagtac 300
aaccgccaga tcgtcgaaga cctcaccggc ctgggcctgt cctatgacct tttcacccgc 360
accaccacct ccaaccacta cgcagtagtg caggaactgt tccgtggtct gtacgacaac 420
ggttacatga tcaaggaaac caccctcggt gcgatttccc catccactgg ccgtaccctg 480
ccagaccgct acattgaagg cacctgccca atctgtggca ccgacggcgc tcgtggcgac 540
cagtgcgaca actgcggaaa ccagctcgat ccagcggacc tgatcaaccc ggtgtccaag 600
atcaacggcg aaaccccaga gttcgttgag accgaacact tcctgctcga cctgccagca 660
ctagctgaag cactaaccga gtggctgaag ggacgcgaag actggcgtcc aaacgtgttg 720
aagttctcgc tcaacctgct ggacgatatc cgcccacgcg caatgtcgcg cgatatgggt 780
accgagctcg aattcgtaat catggtcata gctg 840
<210> 44
<211> 1507
<212> ДНК
<213> Искусственная последовательность
<400> 44
cagtgccaag cttgcatgcc tgcaggtcga ctctagtaag gggcagttgg tctcgaactt 60
tgaccataat gaacaggcgc gcgtcggcaa gcttattgaa gccgcacgca cgggcacggt 120
gctggtggtc agcgatgccg gcatgcctgt ggtttctgat ccgggttttg cgcttatcga 180
cgccgcccac gaggcgaaca ttccggtcac ctgcttcccc gggccgtcag ctgtgccaac 240
tgcgttggca ttgtcgggcc ttcacgtggg ccgcttcgcc ttcgacggtt tcgcgccgcg 300
caaacaaggt gcgcgcacca cgtggttgga gtcgttgaaa accgaaaagc gcgcggtatg 360
tttcttcgaa tctcctcacc gcatcgcaga aaccctggct cacgcagccg aagttttagg 420
tgaacgacgc gcagcagtgt gccgtgaact gtccaaaacc tacgaacagg taaagcgtgg 480
aaccttgcca gagttggcag aatgggcaca agatggggtg cgtggcgaga tcaccgttgt 540
catcgaaggc gcgggcgata tcgcggtcga tgtcgattcg cttatcgacg ccgcccagca 600
gcgcgtcgat tccggcgagc ggttgaaagc ggtgtgcgca gacctcgcga aaatccatgg 660
cgtgagcaaa aatgaactct acgatgcggt tatttctgcc agggaaaatt aagcaatttt 720
taacccctga tacccctttc cggccgggca taaattaagg tggtacgcat gacgaagaac 780
gtgctcgtat ctgttgcctg gccgtatgcc aacggacccc gtcacattgg acatgtggcg 840
gggtttggtg tcccctccga tgtgttcgca aggttccagc gaatgtctgg caacaacgtg 900
ctcatggtgt ccggcaccga tgagcacggc acgccacttc tggttcaagc agacaaagaa 960
ggcgtcaccg ttcaagacct cgcggataag tacaaccgcc agatcgtcga agacctcacc 1020
ggcctgggcc tgtcctatga ccttttcacc cgcaccacca cctccaacca ctacgcagta 1080
gtgcaggaac tgttccgtgg tctgtacgac aacggttaca tgatcaagga aaccaccctc 1140
ggtgcgattt ccccatccac tggccgtacc ctgccagacc gctacattga aggcacctgc 1200
ccaatctgtg gcaccgacgg cgctcgtggc gaccagtgcg acaactgcgg aaaccagctc 1260
gatccagcgg acctgatcaa cccggtgtcc aagatcaacg gcgaaacccc agagttcgtt 1320
gagaccgaac acttcctgct cgacctgcca gcactagctg aagcactaac cgagtggctg 1380
aagggacgcg aagactggcg tccaaacgtg ttgaagttct cgctcaacct gctggacgat 1440
atccgcccac gcgcaatgtc gcgcgatatg ggtaccgagc tcgaattcgt aatcatggtc 1500
atagctg 1557
<210> 45
<211> 1433
<212> ДНК
<213> Искусственная последовательность
<400> 45
taaggggcag ttggtctcga actttgacca taatgaacag gcgcgcgtcg gcaagcttat 60
tgaagccgca cgcacgggca cggtgctggt ggtcagcgat gccggcatgc ctgtggtttc 120
tgatccgggt tttgcgctta tcgacgccgc ccacgaggcg aacattccgg tcacctgctt 180
ccccgggccg tcagctgtgc caactgcgtt ggcattgtcg ggccttcacg tgggccgctt 240
cgccttcgac ggtttcgcgc cgcgcaaaca aggtgcgcgc accacgtggt tggagtcgtt 300
gaaaaccgaa aagcgcgcgg tatgtttctt cgaatctcct caccgcatcg cagaaaccct 360
ggctcacgca gccgaagttt taggtgaacg acgcgcagca gtgtgccgtg aactgtccaa 420
aacctacgaa caggtaaagc gtggaacctt gccagagttg gcagaatggg cacaagatgg 480
ggtgcgtggc gagatcaccg ttgtcatcga aggcgcgggc gatatcgcgg tcgatgtcga 540
ttcgcttatc gacgccgccc agcagcgcgt cgattccggc gagcggttga aagcggtgtg 600
cgcagacctc gcgaaaatcc atggcgtgag caaaaatgaa ctctacgatg cggttatttc 660
tgccagggaa aattaagcaa tttttaaccc ctgatacccc tttccggccg ggcataaatt 720
aaggtggtac gcatgacgaa gaacgtgctc gtatctgttg cctggccgta tgccaacgga 780
ccccgtcaca ttggacatgt ggcggggttt ggtgtcccct ccgatgtgtt cgcaaggttc 840
cagcgaatgt ctggcaacaa cgtgctcatg gtgtccggca ccgatgagca cggcacgcca 900
cttctggttc aagcagacaa agaaggcgtc accgttcaag acctcgcgga taagtacaac 960
cgccagatcg tcgaagacct caccggcctg ggcctgtcct atgacctttt cacccgcacc 1020
accacctcca accactacgc agtagtgcag gaactgttcc gtggtctgta cgacaacggt 1080
tacatgatca aggaaaccac cctcggtgcg atttccccat ccactggccg taccctgcca 1140
gaccgctaca ttgaaggcac ctgcccaatc tgtggcaccg acggcgctcg tggcgaccag 1200
tgcgacaact gcggaaacca gctcgatcca gcggacctga tcaacccggt gtccaagatc 1260
aacggcgaaa ccccagagtt cgttgagacc gaacacttcc tgctcgacct gccagcacta 1320
gctgaagcac taaccgagtg gctgaaggga cgcgaagact ggcgtccaaa cgtgttgaag 1380
ttctcgctca acctgctgga cgatatccgc ccacgcgcaa tgtcgcgcga tat 1479
<210> 46
<211> 3380
<212> ДНК
<213> Искусственная последовательность
<400> 46
taaggggcag ttggtctcga actttgacca taatgaacag gcgcgcgtcg gcaagcttat 60
tgaagccgca cgcacgggca cggtgctggt ggtcagcgat gccggcatgc ctgtggtttc 120
tgatccgggt tttgcgctta tcgacgccgc ccacgaggcg aacattccgg tcacctgctt 180
ccccgggccg tcagctgtgc caactgcgtt ggcattgtcg ggccttcacg tgggccgctt 240
cgccttcgac ggtttcgcgc cgcgcaaaca aggtgcgcgc accacgtggt tggagtcgtt 300
gaaaaccgaa aagcgcgcgg tatgtttctt cgaatctcct caccgcatcg cagaaaccct 360
ggctcacgca gccgaagttt taggtgaacg acgcgcagca gtgtgccgtg aactgtccaa 420
aacctacgaa caggtaaagc gtggaacctt gccagagttg gcagaatggg cacaagatgg 480
ggtgcgtggc gagatcaccg ttgtcatcga aggcgcgggc gatatcgcgg tcgatgtcga 540
ttcgcttatc gacgccgccc agcagcgcgt cgattccggc gagcggttga aagcggtgtg 600
cgcagacctc gcgaaaatcc atggcgtgag caaaaatgaa ctctacgatg cggttatttc 660
tgccagggaa aattaatcgt tatgtcattg tgatgctccc gtgaacataa acgggaccta 720
ctggctttac ttaagtaaca gctatgtaaa agaccaggtc aggttcgggt cggtttaggt 780
acgaaaccca tttttcggtt tgctttccag gtttccccaa gtaaaggtga gttttatgac 840
tacatctgac ccaaattcga aaccgatagt ggaggatgct cagccagagc agatcaccgc 900
aaccgaagaa ctggcgggtt tgcttgagaa tccaactaac ctggaaggga aactggccga 960
cgccgaagag gaaattatcc tcgaaggcga agacgcccag gcctcactta actggtcagt 1020
catcgttcca gccctagtca ttgtcctagc gacagtggtg tggggtatcg gattcaaaga 1080
tagctttacc aactttgcta gttctgcgtt gtcagcagta gttgacaatc tcggctgggc 1140
cttcattttg tttggcacag tctttgtatt ttttatcgtt gttatcgccg ctagtaaatt 1200
cggcacgatt cgcttaggcc gcattgatga agcaccagag tttcgcacgg tgtcatggat 1260
ttccatgatg tttgctgcag gtatgggtat tggtttgatg ttctacggaa ccacagaacc 1320
tttaaccttc taccgcaatg gtgtacctgg atatgatgaa cacaatgttg gcgttgctat 1380
gtccacgaca atgttccact ggaccttgca tccatgggct atctacgcaa ttgtgggcct 1440
agccattgcc tattcgacct tccgagtggg ccgtaaacag cttctaagct ctgcattcgt 1500
gccactcatt ggtgaaaaag gtgcagaagg atggttgggc aagctcatcg acatcctggc 1560
gattatcgcc acagtattcg gcaccgcatg ttcccttggc ctgggtgcgc tgcagatcgg 1620
tgcaggactt tccgcagcca acatcattga aaatccgagt gactggactg tcattggtat 1680
tgtttctgtc ttgaccttgg catttatctt ctctgctatt tctggtgtgg gcaagggaat 1740
ccagtacctc tccaacgcca acatggttct ggcagctctg ctcgcgattt tcgtgttcgt 1800
tgtcggacca accgtgtcga ttttgaacct gctgccaggt tctattggca actacctgtc 1860
caacttcttc caaatggcag gccgcactgc catgagtgcc gacggcacag caggtgagtg 1920
gctaggtagc tggaccatct tctactgggc atggtggatc tcttggtcac cattcgtagg 1980
aatgttcttg gcacgtattt cccgtggccg ctccatccgt gagttcatcc tgggcgtgtt 2040
gctcgtccca gcaggtgtgt ccaccgtatg gttctccatt tttggtggca ctgcgattgt 2100
cttcgaacaa aatggggaat ccatttgggg tgatggtgca gcagaagagc agctctttgg 2160
attgcttcat gcacttccag gtgggcaaat aatgggcatc atcgccatga ttttgctggg 2220
tactttcttc attacctctg cagactctgc ttccaccgtc atgggcacca tgagtcagca 2280
cggccagctg gaagccaaca agtgggtgac agctgcctgg ggtgttgcta ccgcagctat 2340
tggactaacg ctattgcttt ctggtggtga caatgccttg aacaacttgc aaaacgtcac 2400
catcgtggct gcaacaccat tcctgtttgt ggttattgga ttgatgtttg cgttagtcaa 2460
ggacttaagc aatgatgtga tctacctcga gtaccgtgag cagcaacgct tcaacgcgcg 2520
ccttgcccgt gaacgtcgtg ttcacaatga acaccgcaag cgtgaactgg ctgcaaagcg 2580
acgcagggag cgtaaggcga gtggcgcggg gaagcgtcga taggcaattt ttaacccctg 2640
ataccccttt ccggccgggc ataaattaag gtggtacgca tgacgaagaa cgtgctcgta 2700
tctgttgcct ggccgtatgc caacggaccc cgtcacattg gacatgtggc ggggtttggt 2760
gtcccctccg atgtgttcgc aaggttccag cgaatgtctg gcaacaacgt gctcatggtg 2820
tccggcaccg atgagcacgg cacgccactt ctggttcaag cagacaaaga aggcgtcacc 2880
gttcaagacc tcgcggataa gtacaaccgc cagatcgtcg aagacctcac cggcctgggc 2940
ctgtcctatg accttttcac ccgcaccacc acctccaacc actacgcagt agtgcaggaa 3000
ctgttccgtg gtctgtacga caacggttac atgatcaagg aaaccaccct cggtgcgatt 3060
tccccatcca ctggccgtac cctgccagac cgctacattg aaggcacctg cccaatctgt 3120
ggcaccgacg gcgctcgtgg cgaccagtgc gacaactgcg gaaaccagct cgatccagcg 3180
gacctgatca acccggtgtc caagatcaac ggcgaaaccc cagagttcgt tgagaccgaa 3240
cacttcctgc tcgacctgcc agcactagct gaagcactaa ccgagtggct gaagggacgc 3300
gaagactggc gtccaaacgt gttgaagttc tcgctcaacc tgctggacga tatccgccca 3360
cgcgcaatgt cgcgcgatat 3492
<---
Изобретение относится к биотехнологии. Предложены бактерия рода Corynebacterium для продуцирования L-глутаминовой кислоты, способ получения L-глутаминовой кислоты с использованием указанной бактерии, белок и молекула нуклеиновой кислоты для продуцирования L-глутаминовой кислоты. Также предложены рекомбинантный микроорганизм для продуцирования L-глутаминовой кислоты, содержащий указанную молекулу нуклеиновой кислоты, способ его конструирования и способ получения L-глутаминовой кислоты с его использованием. Также предложены применение указанной молекулы нуклеиновой кислоты для конструирования генетически модифицированной бактерии для продуцирования L-глутаминовой кислоты, способ повышения продукции L-глутаминовой кислоты в бактерии рода Corynebacterium с использованием указанной молекулы нуклеиновой кислоты. Изобретение обеспечивает получение высокой продукции L-глутаминовой кислоты. 9 н. и 7 з.п. ф-лы, 3 табл., 6 пр.
1. Бактерия рода Corynebacterium для продуцирования L-глутаминовой кислоты, причем указанная бактерия обладает улучшенной экспрессией полинуклеотида, кодирующего последовательность аминокислот SEQ ID NO: 3 по сравнению с немодифицированной бактерией;
при этом указанная улучшенная экспрессия представляет собой усиленную экспрессию полинуклеотида, кодирующего последовательность аминокислот SEQ ID NO: 3, или присутствие точечной мутации в полинуклеотиде, кодирующем последовательность аминокислот SEQ ID NO: 3, или присутствие точечной мутации в полинуклеотиде и усиленную экспрессию полинуклеотида, который кодирует последовательность аминокислот SEQ ID NO: 3; где указанная точечная мутация в полинуклеотиде, кодирующем последовательность аминокислот SEQ ID NO: 3, представляет собой замену аспарагина в положении 520 в последовательности аминокислот SEQ ID NO: 3 на лизин.
2. Бактерия рода Corynebacterium по п. 1, где полинуклеотид, кодирующий последовательность аминокислот SEQ ID NO: 3, содержит последовательность нуклеотидов SEQ ID NO: 1.
3. Бактерия рода Corynebacterium по п. 1, где последовательность полинуклеотида содержит точечную мутацию основания в положении 1560 последовательности полинуклеотида, представленной в SEQ ID NO: 1, с заменой цитозина (C) на аденин (A).
4. Бактерия рода Corynebacterium по п. 1, где указанная последовательность полинуклеотида, имеющая точечную мутацию, содержит последовательность полинуклеотида SEQ ID NO: 2.
5. Бактерия рода Corynebacterium по п. 1, где указанная бактерия представляет собой Corynebacterium glutamicum CGMCC № 21220 или ATCC13869.
6. Способ получения L-глутаминовой кислоты, который включает: культивирование бактерии по п. 1 и выделение L-глутаминовой кислоты из указанной культуры.
7. Белок, характеризующийся тем, что он имеет последовательность аминокислот SEQ ID NO: 4, где указанный белок используется для продуцирования L-глутаминовой кислоты.
8. Молекула нуклеиновой кислоты, характеризующаяся тем, что указанная молекула нуклеиновой кислоты представляет собой любое из:
B1) молекулы нуклеиновой кислоты, кодирующей белок по п. 4;
B2) молекулы ДНК, кодирующая последовательность которой представлена в SEQ ID NO: 2;
B3) молекулы ДНК, последовательность нуклеотидов которой представлена в SEQ ID NO: 2;
B4) последовательности полинуклеотида, содержащей полинуклеотид, кодирующий последовательность аминокислот, представленную в SEQ ID NO: 3, где аспарагин в положении 520 заменен на лизин,
причем указанная молекула нуклеиновой кислоты используется для продуцирования L-глутаминовой кислоты.
9. Молекула нуклеиновой кислоты по п. 8, причем последовательность полинуклеотида содержит полинуклеотид, кодирующий последовательность аминокислот, представленную в SEQ ID NO: 4.
10. Молекула нуклеиновой кислоты по п. 8, где последовательность полинуклеотида образована в результате мутации основания в положении 1560 последовательности полинуклеотида, представленной в SEQ ID NO: 1, с заменой цитозина (C) на аденин (A).
11. Молекула нуклеиновой кислоты по п. 8, где последовательность полинуклеотида содержит последовательность полинуклеотида, представленную в SEQ ID NO: 2.
12. Рекомбинантный микроорганизм для продуцирования L-глутаминовой кислоты, содержащий молекулу нуклеиновой кислоты по п. 8, или рекомбинантный вектор, содержащий молекулу нуклеиновой кислоты по п. 8.
13. Применение молекулы нуклеиновой кислоты по п. 8 для конструирования генетически модифицированной бактерии для продуцирования L-глутаминовой кислоты.
14. Способ повышения продуцирования L-глутаминовой кислоты в бактерии рода Corynebacterium, характеризующийся тем, что указанный способ включает
введение молекулы нуклеиновой кислоты по любому из пп. 8-11 или рекомбинантного вектора, содержащего молекулу нуклеиновой кислоты по любому из пп. 8-11, в микроорганизм-мишень с получением рекомбинантного микроорганизма и культивирование указанного рекомбинантного микроорганизма с получением повышенной продукции L-глутаминовой кислоты по сравнению с микроорганизмом-мишенью без мутации.
15. Способ конструирования рекомбинантного микроорганизма по п. 12, характеризующийся тем, что указанный способ включает по меньшей мере одно из:
F1) введения молекулы нуклеиновой кислоты по п. 11 в микроорганизм-мишень с получением рекомбинантного микроорганизма;
F2) редактирования молекулы ДНК, представленной в SEQ ID NO: 1, с использованием средства редактирования генов, таким образом, чтобы микроорганизм-мишень содержал молекулу ДНК, представленную в SEQ ID NO: 2.
16. Способ получения L-глутаминовой кислоты, характеризующийся тем, что указанный способ включает продуцирование L-глутаминовой кислоты путем культивирования рекомбинантного микроорганизма по п. 12 и выделение L-глутаминовой кислоты из культуры.
| US 2006228712 A1, 12.10.2006 | |||
| Способ разделения 1,4,8 и 2,4,8-нафтиламия дисульфокислот | 1932 |
|
SU33153A1 |
| glycine/betaine ABC transporter [Corynebacterium glutamicum] | |||
| Способ разделения 1,4,8 и 2,4,8-нафтиламия дисульфокислот | 1932 |
|
SU33153A1 |
| US 20050244935 A1, 03.11.2005 | |||
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ СЕМЕЙСТВА ГЛУТАМАТА С ИСПОЛЬЗОВАНИЕМ КОРИНЕФОРМНОЙ БАКТЕРИИ | 2011 |
|
RU2496867C2 |

