Изобретение относится к молекулярной биологии и генетике, а именно к способам получения ферментов модификации ДНК-метилазы Е.со.R II.
Цель изобретения - повышение выхода и удельной активности фермента.
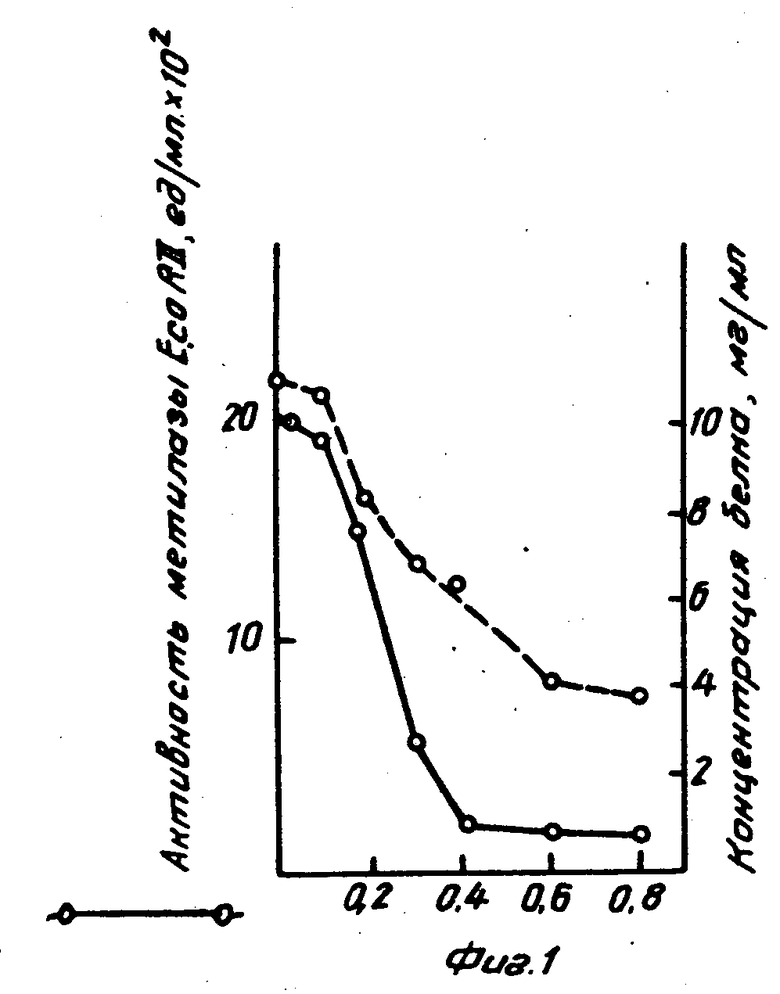
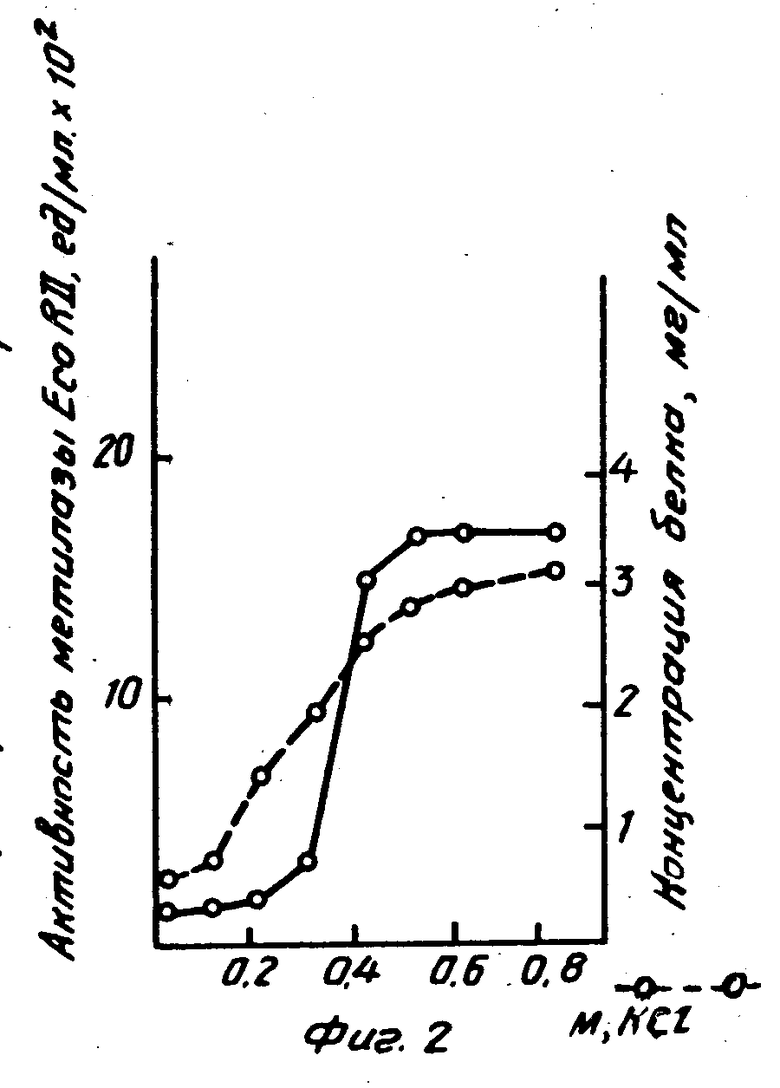
На фиг. 1 изображена активность метилазы Е.co R II при осаждении полиэтиленимином; на фиг. 2 показано влияние концентраций KCl на элюацию метилазы Е.со R II из полиэтилениминового осадка.
Полиэтиленимин добавляли к бесклеточному экстракту до различных концентраций. После центрифугирования 20 мин при 20000 g из супернатанта отбирали аликвоты, разводили буфером и определяли активность метилазы и концентрацию белка (см.фиг.1).
К 20 мл бесклеточного экстракта добавляли полиэтиленимин до конечной концентрации 0,3% . Аликвоты по 2 мл центрифугировали и осадок ресуспендировали в 2 мл буфера, содержащего различные концентрации KCl. После центрифугирования в супернатантах определяли активность метилазы и концентрацию белка.
Сущность изобретения заключается в том, что штамм-продуцент Escherichia coli B 834 (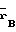
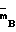 )/pSK 323 культивируют до поздней логарифмической фазы, клетки отделяют и ресуспендируют в холодном ФЕМ-буфере (10 мМ калий фосфатный буфер; рН 7,0, 1 мМ этилендиаминтетраацетат (ЭДТА), 7 мМ 2-меркаптоэтанол), содержащем 0,1% тритона Х-100 и 50 мкг/мл лизоцима. Биомассу и буфер берут в соотношении 1:3. Суспензию инкубируют 30 мин, разрушают клетки ультразвуком. После центрифугирования к супернатанту добавляют полиэтиленимин до конечной концентрации 0,3-0,4% . Осадок отделяют и растворяют в ФЕМ-буфере, содержащем 0,4-0,5 М хлористого калия. Осадок после центрифугирования отбрасывают, а к супернатанту добавляют сульфат аммония до насыщения 60% . Осадок вновь отделяют и растворяют в ФЕМ-буфере, диализируют против ФЕМ-буфера и наносят на ДЭАЭ-целлюлозу. Метилазу элюируют линейным градиентом хлористого калия от 0 до 0,2 М в ФЕМ-буфере. Фракции с метилазной активностью объединяют, наносят на фосфоцеллюлозу и элюируют линейным градиентом хлористого калия от 0,3 М до 0,6 М. Объединяют фракции, содержащие метилазу, и проводят диализ против 50%-ного раствора глицерина в ФЕМ-буфере.
)/pSK 323 культивируют до поздней логарифмической фазы, клетки отделяют и ресуспендируют в холодном ФЕМ-буфере (10 мМ калий фосфатный буфер; рН 7,0, 1 мМ этилендиаминтетраацетат (ЭДТА), 7 мМ 2-меркаптоэтанол), содержащем 0,1% тритона Х-100 и 50 мкг/мл лизоцима. Биомассу и буфер берут в соотношении 1:3. Суспензию инкубируют 30 мин, разрушают клетки ультразвуком. После центрифугирования к супернатанту добавляют полиэтиленимин до конечной концентрации 0,3-0,4% . Осадок отделяют и растворяют в ФЕМ-буфере, содержащем 0,4-0,5 М хлористого калия. Осадок после центрифугирования отбрасывают, а к супернатанту добавляют сульфат аммония до насыщения 60% . Осадок вновь отделяют и растворяют в ФЕМ-буфере, диализируют против ФЕМ-буфера и наносят на ДЭАЭ-целлюлозу. Метилазу элюируют линейным градиентом хлористого калия от 0 до 0,2 М в ФЕМ-буфере. Фракции с метилазной активностью объединяют, наносят на фосфоцеллюлозу и элюируют линейным градиентом хлористого калия от 0,3 М до 0,6 М. Объединяют фракции, содержащие метилазу, и проводят диализ против 50%-ного раствора глицерина в ФЕМ-буфере.
П р и м е р 1. Штамм-продуцент Escherichia coli B 834 (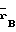
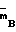 )/pSK 323 культивируют до поздней логарифмической фазы. Клетки собирают центрифугированием и ресуспендируют в холодном ФЕМ-буфере (10 мМ калий-фосфатный буфер, рН 7,0; 1 мМ ЭДТА; 7 мМ 2-меркаптоэтанол), содержащем 0,1% тритона Х-100 и 50 мкг/мл лизоцима. На 100 г биомассы берут 300 мл буфера. Суспензию инкубируют 30 мин при 4оС, затем клетки разрушают ультразвуком.
)/pSK 323 культивируют до поздней логарифмической фазы. Клетки собирают центрифугированием и ресуспендируют в холодном ФЕМ-буфере (10 мМ калий-фосфатный буфер, рН 7,0; 1 мМ ЭДТА; 7 мМ 2-меркаптоэтанол), содержащем 0,1% тритона Х-100 и 50 мкг/мл лизоцима. На 100 г биомассы берут 300 мл буфера. Суспензию инкубируют 30 мин при 4оС, затем клетки разрушают ультразвуком.
Разрушенные клетки центрифугируют (20000 g, 1 ч, 4оС). Осадок отбрасывают, а к суспернатанту добавляют полиэтиленимин из расчета на каждые 10 мл супернатанта 0,4 мл 10%-ного раствора полиэтиленимина в воде, при концентрации белка в супернатанте около 10 мг/мл. Осадок собирают центрифугированием и ресуспендируют в 300 мл ФЕМ-буфера, содержащего 0,5 М KCl. Осадок отделяют и отбрасывают. К супернатанту добавляют при перемешивании насыщенный раствор сульфата аммония до конечной концентрации 60% (на 100 мл раствора 150 мл сульфата аммония). Осадок собирают центрифугированием и ресуспендируют в 300 мл. Проводят диализ против ФЕМ-буфера и наносят на колонку с диэтиламиноэтилцеллюлозой (ДЕАЕЦ), уравновешенной ФЕМ-буфером. После нанесения на колонку промывают ФЕМ буфером до тех пор, пока оптическая плотность не станет равной оптической плотности ФЕМ-буфера. Далее через колонку пропускают ФЕМ-буфер с возрастающей концентрацией KCl от 0 до 0,2 М. Метилаза Е. со R 11 элюируется при концентрации от 0,08М до 0,12 М KCl. Фракции, содержащие активность метилазы, объединяют и наносят на колонку с фосфоцеллюлозой. Колонку промывают ФЕМ-буфером, содержащим 0,2 М хлористый калий, затем через колонку пропускают ФЕМ-буфер с возрастающей концентрацией KCl от 0,2 до 0,8 М. Метилаза элюируется при концентрации хлористого калия от 0,3 М до 0,6 М. Активные фракции диализируют в течение 16 ч против 50 объемов ФЕМ-буфера, содержащего 0,2 М KCl и 50% глицерина.
Выход метилазы 1,2 ˙104 ед./г биомассы, удельная активность 5,7 х 105 ед./мг белка.
За единицу активности принимают количество фермента, необходимое для включения 1 пикомоля метильных групп в дезоксирибонуклеиновую кислоту за 1 ч при 37оС.
П р и м е р 2. Культивируют штамм-продуцент, клетки собирают, ресуспендируют в буфере и обрабатывают лизоцимом, как предложено в примере 1. После центрифугирования к супернатанту добавляют полиэтиленимин из расчета на каждые 10 мл супернатанта 0,1; 0,2; 0,3 мл 10%-ного раствора полиэтиленимина в воде. Осадок собирают центрифугированием, ресуспендируют в 300 мл ФЕМ-буфере, содержащего 0,4 М хлористый калий. Осадок отделяют и дальнейшую очистку проводят, как в примере 1.
Изобретение позволяет существенно повысить выход и активность целевого продукта.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТОВ СИСТЕМЫ РЕСТРИКЦИИ-МОДИФИКАЦИИ | 1984 |
|
SU1262952A1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕСТРИКТАЗЫ E.CORII ИЗ ШТАММА БАКТЕРИЙ ESCHERICHIA COLI B 834/PSK 323 | 1984 |
|
SU1321060A1 |
| Способ получения фермента днк-цитозин-метилазы 1 из клеток | 1977 |
|
SU737443A1 |
| Способ конструирования плазмидной ДНК,штамм @ @ -продуцент эндонуклеазы рестрикции @ и способ получения эндонуклеазы рестрикции @ | 1981 |
|
SU1040791A1 |
| СПОСОБ ПОЛУЧЕНИЯ ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ ECO RV | 1984 |
|
SU1218678A1 |
| Способ получения эндонуклеазы рестрикции | 1982 |
|
SU1095645A1 |
| Способ конструирования штаммов еSснеRIснIа coLI-продуцентов рестриктазы и метилазы е со R @ | 1979 |
|
SU941423A1 |
| Рекомбинантная плазмидная ДНК рУК11, кодирующая рестриктазу и метилазу Е coRII, способ ее конструирования и штамм бактерий ЕSснеRIснIа coLI - продуцент рестриктазы и метилазы Е coRII | 1987 |
|
SU1453896A1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК РТТG КM2, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО ИММУННОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА, ШТАММ ESCHERICHIA COLI T3G - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ИММУННОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУННОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА | 1996 |
|
RU2097428C1 |
| Способ получения @ - @ -галактозидазы | 1982 |
|
SU1082812A1 |
Изобретение относится к молекулярной биологии и генетике, конкретно - к способам получения ферментов модификации ДНК-метилазы E. co R11. Цель изобретения - повышение выхода и удельной активности фермента. Для получения метилазы E. co R11 культивируют штамм-продуцент Escherichia coli B 834  / pSK 323. Клетки дезинтегрируют в присутствии лизоцима и разрушают ультразвуком. После центрифугирования фермент в супернатанте осаждают полиэтиленимином, который добавляют до конечной концентрации 0,3 - 0,4%, затем метилазу извлекают из осадка 0,4 - 0,5 М раствором хлористого калия в ФЕМ-буфере, высаживают сульфатом аммония, диализуют и подвергают очистке на ДЕАЕ-целлюлозе при элюировании хлористым калием в линейном градиенте концентрации от 0 до 0,2 М. Дальнейшую очистку проводят на фосфоцеллюлозе при элюировании тем же раствором в линейном градиенте концентрации 0,3 - 0,6 М. Фракции, содержащие метилазу, объединяют и диализируют против 50%-ного раствора глицерина. Выход метилазы 1,2·104 ед./г биомассы, удельная активность 5,7·105 ед./мг белка. 2 ил.
/ pSK 323. Клетки дезинтегрируют в присутствии лизоцима и разрушают ультразвуком. После центрифугирования фермент в супернатанте осаждают полиэтиленимином, который добавляют до конечной концентрации 0,3 - 0,4%, затем метилазу извлекают из осадка 0,4 - 0,5 М раствором хлористого калия в ФЕМ-буфере, высаживают сульфатом аммония, диализуют и подвергают очистке на ДЕАЕ-целлюлозе при элюировании хлористым калием в линейном градиенте концентрации от 0 до 0,2 М. Дальнейшую очистку проводят на фосфоцеллюлозе при элюировании тем же раствором в линейном градиенте концентрации 0,3 - 0,6 М. Фракции, содержащие метилазу, объединяют и диализируют против 50%-ного раствора глицерина. Выход метилазы 1,2·104 ед./г биомассы, удельная активность 5,7·105 ед./мг белка. 2 ил.
СПОСОБ ПОЛУЧЕНИЯ МЕТИЛАЗЫ E CO R11, предусматривающий культивирование продуцента Escherichia coli, инкубацию и дезинтеграцию клеток ультразвуком, получение бесклеточного экстракта, осаждение фермента сульфатом аммония, хроматографию на диэтиламиноэтилцеллюлозе и фосфоцеллюлозе и последующее концентрирование, отличающийся тем, что, с целью повышения выхода и удельной активности фермента, в качестве продуцента используют штамм Escherichia coli 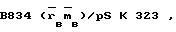 суспензию клеток инкубируют в присутствии лизоцима, фермент осаждают полиэтиленимином, который добавляют к бесклеточному экстакту до концентрации 0,3 - 0,4% с последующей элюацией метилазы из осадка 0,4 - 0,5 М хлористым калием, непостредственно после хроматографии на диэтиламиноэтилцеллюлозе осуществляют хроматографию на фосфоцеллюлозе при элюировании буферным раствором хлористого калия в градиенте концентрации 0,3 - 0,6 М, а концентрирование целевого продукта осуществляют диализом против глицерина.
суспензию клеток инкубируют в присутствии лизоцима, фермент осаждают полиэтиленимином, который добавляют к бесклеточному экстакту до концентрации 0,3 - 0,4% с последующей элюацией метилазы из осадка 0,4 - 0,5 М хлористым калием, непостредственно после хроматографии на диэтиламиноэтилцеллюлозе осуществляют хроматографию на фосфоцеллюлозе при элюировании буферным раствором хлористого калия в градиенте концентрации 0,3 - 0,6 М, а концентрирование целевого продукта осуществляют диализом против глицерина.
| Богдарина И.Г., Бурьянов Я.И., Баев А.А | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Дверной замок, автоматически запирающийся на ригель, удерживаемый в крайних своих положениях помощью серии парных, симметрично расположенных цугальт | 1914 |
|
SU1979A1 |
| Приспособление для плетения проволочного каркаса для железобетонных пустотелых камней | 1920 |
|
SU44A1 |
