ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение может быть отнесено к области биотехнологии и охватывает нуклеотидные последовательности, содержащие, например, промоторы, гомологичные области (hr) в качестве энхансеров, а также последовательности, кодирующие регуляторы транскрипции, например, кДНК бакуловируса Ас-ie-01, или любые их комбинации, которые способны повысить качество и эффективность продукции рекомбинантных белков. Кроме того, настоящее изобретение также относится к самим векторам, содержащим вышеупомянутые нуклеотидные последовательности в соответствии с изобретением, клетки, инфицированные, трансформированные или трансфицированные указанными последовательностями или векторами, и способы продукции белковое применением вышеуказанных последовательностей, векторов или клеток.
УРОВЕНЬ ТЕХНИКИ
Бакуловирусная векторная система экспрессии (BEVS) является общепринятым способом продукции рекомбинантных белков для применения в качестве вакцин, терапевтических молекул или диагностических реагентов. Благодаря своему потенциалу сверхэкспрессии и быстрой разработки BEVS является одним из самых привлекательных вариантов для продукции рекомбинантных белков для любых целей. Бакуловирусы, наиболее часто применяемые в промышленности для экспрессии рекомбинантных белков, основаны на вирусе многоядерного полиэдроза Autographa californica (AcMNPV) и клетках насекомых Spodoptera frugiperda 9 (Sf9) или 21 (Sf21) в качестве подходящего хозяина для экспрессии (1), а также личинках насекомых Trichoplusia ni (m.ni) в качестве живых биофабрик (2). Со времени разработки BEVS в 80-х годах (3) в клетках насекомых, инфицированных бакуловирусами, успешно продуцированы сотни рекомбинантных белков, начиная от цитозольных ферментов до мембрано-связанных белков.
Предпринимались усилия для увеличения производительности BEVS (4). Доступен ряд векторов переноса для конструирования рекомбинантных бакуловирусов, кодирующих постоянные гибридные белки, которые, по сообщениям, улучшают экспрессию белков, в том числе мальтоза-связывающий белок, глутатион S-трансферазу, SUMO и последовательность удерживания KDEL. Другие попытки исследований, связанных с улучшением стабильности экспрессированных белков, были направлены на два гена в геноме бакуловируса, которые не являются необходимыми для роста вируса в культуре клеток, а именно chiA (хитиназа) и cath (катепсин). Делеция ChiA, по-видимому, улучшает продукцию секретируемых белков за счет накопления белка в эндоплазматическом ретикулуме и процессинга белков посредством секреторных путей клеток. Кроме того, в улучшение стабильности продуктов chiA--вирусов может вносить вклад предотвращение образования катепсиновых протеаз. С целью повышения продуктивности бакуловирусов недавно были разработаны новые линии клеток насекомых, например, линии клеток Т. ni High-Five™ (Hi-5) или BTI-TnAo38, обеспечивающие значительное повышение итогового количества выделяемых гетерологичных белков (5, 6).
Ускорение экспрессии рекомбинантных белков, при котором экспрессия белка происходила бы до серьезного нарушения механизмов клеток насекомых бакуловирусной инфекцией, могло бы представлять собой важное усовершенствование BEVS. Поздняя экспрессия, управляемая обычными сильными вирусными промоторами генов polyhedrin (polh) или p10, обладает серьезными недостатками в виде посттрансляционных модификаций чужеродных белков. Описаны характеристики бакуловирусных промоторов, обеспечивающих более раннюю экспрессию, чем традиционно используемые промоторы polh или p10; указанные промоторы применяются для продукции гетерологичных белков, однако демонстрируют более низкую производительность (7).
Еще одной возможностью усовершенствования BEVS могло бы быть повышение сохранения целостности клеток на поздних стадиях после инфицирования путем снижения гибели клеток, вызванной вирусом. Ослабление тяжелых повреждений механизмов клеток насекомых на поздних стадиях после инфицирования, вызванных BEVS, должно не только увеличить время продукции и накопления рекомбинантных белков (секретируемых или нет), но и предоставить больше времени на фолдинг сложных белков или посттрансляционные модификации продуцируемых белков.
Установлено, что некоторые элементы ДНК бакуловирусов участвуют в активации поздней экспрессии генов факторов, необходимых для размножения вируса. Одним из них является немедленный ранний (ie) белок IE-1 и его сплайс-вариант IE-0 из AcMNPV (вируса многоядерного полиэдроза Autographa californica). Трансляция мРНК AcMNPV, кодируемой геном Ас-ie-01, приводит к получению IE-0 и IE-1 вследствие внутренней инициации трансляции. Оба указанных соединения считаются критическими медиаторами экспрессии бакуловирусных генов из-за их способности выступать в качестве регуляторов транскрипции (8). AcMNPV IE-1 является 67-кДа димерным ДНК-связывающим белком, синтезирующимся на самых ранних стадиях инфекции и стимулирующим транскрипцию в анализах трансфекции плазмид за счет активности его N-концевого кислого домена (9, 10). IE-1 накапливается в ядре, где его концентрация поддерживается на поздних стадиях (11). Трансактивация за счет IE-1 усиливается посредством его связывания в виде гомодимера с последовательностями гомологичной области (hr) бакуловируса, действующими в качестве энхансеров транскрипции и сайтов начала репликации вирусной ДНК. AcMNPV IE-0 - 72,6-кДа белок из 636 аминокислот, причем 38 аминокислот в его составе кодируются orf141 (экзон 0), 16 аминокислот - вышележащим нетранслируемым лидером ie1 и 582 аминокислоты представляют собой полный белок IE-1. Поэтому конечный продукт идентичен IE-1, за исключением дополнительных 54 аминокислот, присоединенных с N-конца. Предположительно из-за их общей последовательности, IE-0 и IE-1 обладают одинаковой биохимической активностью, включая связывание hr-энхансера и регуляцию транскрипции.
Существует потребность в новых альтернативных BEVS, обеспечивающих а) усиленную экспрессию по сравнению с коммерческими BEVS, которые используют промоторы polh или р10 и b) долгосрочную экспрессию в бакуловирусной системе путем снижения вирус-индуцированного повреждения клеток.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В настоящем изобретении представлены продукты и способы улучшения экспрессиии рекомбинантных белков с помощью BEVS.
Следующие элементы являются предпочтительными вариантами реализации, поскольку обеспечивают указанную улучшенную экспрессию:
1. Рекомбинантный бакуловирус, содержащий последовательность нуклеиновой кислоты, обеспечивающую экспрессию белков IE-1, IE-0 и/или их фрагментов, функционирующих в качестве регуляторов транскрипции, на уровне, превышающем эндогенный уровень, причем нуклеиновая кислота выбрана из группы, состоящей из:
(а) нуклеиновой кислоты, содержащей нуклеотидную последовательность, указанную в любой из SEQ ID NO: 1-5;
(b) последовательности нуклеиновой кислоты, обладающей по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью с нуклеотидной последовательность, указанной в любой из SEQ ID NO: 1-5 и кодирующей белок, который может функционировать как регулятор транскрипции в рекомбинантном бакуловирусе;
(c) последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность, содержащую аминокислотную последовательность, указанную в любой из SEQ ID NO: 6-9; и
(d) последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность, обладающую по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% сходством с аминокислотной последовательностью, указанной в любой из SEQ ID NO: 6-9 и способной функционировать как регулятор транскрипции в рекомбинантном бакуловирусе.
2. Рекомбинантный бакуловирус по пункту 1, дополнительно содержащий по меньшей мере одну рекомбинантную гомологичную область (hr) в качестве энхансерной области, функционально связанную с любым промотором, подходящим для управления экспрессией рекомбинантных белков.
3. Рекомбинантный бакуловирус по пункту 2, отличающийся тем, что рекомбинантная гомологичная область (hr) является последовательностью, указанной в SEQ ID NO: 27 (hr1).
4. Рекомбинантный бакуловирус по пункту 2 или 3, отличающийся тем, что промотор, функционально связанный с гомологичной областью (hr), выбран из группы нуклеиновых кислот, включающей:
(а) нуклеиновую кислоту, содержащую нуклеотидную последовательность, указанную в любой из SEQ ID NO: 10-16; и
(b) последовательность нуклеиновой кислоты, способную функционировать в качестве промотора в рекомбинантном бакуловирусе и обладающую по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью с нуклеотидной последовательностью, указанной в любой из SEQ ID NO: 10-16.
5. Рекомбинантный бакуловирус по любому из пунктов 1-4, содержащий последовательность нуклеиновой кислоты, содержащую комбинации рекомбинантных промоторов, последовательностей, кодирующих регуляторы транскрипции, и энхансерных областей, выбранных из группы, включающей SEQ ID NO: 17-26.
6. Рекомбинантный бакуловирус по любому из пунктов 1-5, дополнительно содержащий последовательность нуклеиновой кислоты, кодирующую рекомбинантный белок, причем указанная нуклеотидная последовательность функционально связана с нуклеотидной последовательностью, выбранной из, группы, состоящей из нуклеотидных последовательностей по пунктам 1-5.
7. Вектор переноса, подходящий для получения рекомбинантного бакуловируса по любому из пунктов 1-6, содержащий указанную последовательность для экспрессии белков IE-0, IE-1 и/или их фрагментов, функционирующих в качестве регуляторов транскрипции, на уровне, превышающем эндогенный уровень, дополнительно содержащую нуклеотидную последовательность, подходящую для интеграции или транспозиции в бакуловирус.
8. Вектор переноса по пункту 7, дополнительно содержащий по меньшей мере одну рекомбинантную гомологичную область (hr) в качестве энхансерной области, функционально связанную с любым промотором, подходящим для управления экспрессией рекомбинантных белков.
9. Вектор переноса по пункту 8, отличающийся тем, что рекомбинантная гомологичная область (hr) является последовательностью, указанной в SEQ ID NO: 27 (hr1).
10. Вектор переноса по пункту 8 или 9, отличающийся тем, что промотор, функционально связанный с гомологичной областью (hr), выбран из группы нуклеиновых кислот, включающей:
(a) нуклеиновую кислоту, содержащую нуклеотидную последовательность, указанную в любой из SEQ ID NO: 10-16; и
(b) нуклеотидной последовательности, способной функционировать в качестве промотора в рекомбинантном бакуловирусе и обладающей по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью с нуклеотидной последовательностью, указанной в любой из SEQ ID NO: 10-16.
11. Вектор переноса по любому из пунктов 7-10, содержащий последовательность нуклеиновой кислоты, содержащую комбинации рекомбинантных промоторов, последовательностей, кодирующих регуляторы транскрипции, и энхансерных областей, выбранных из группы, включающей SEQ ID NO: 17-26.
12. Вектор переноса по любому из пунктов 7-11, дополнительно содержащий последовательность нуклеиновой кислоты, кодирующую рекомбинантный белок, причем указанная нуклеотидная последовательность функционально связана с нуклеотидной последовательностью, выбранной из группы, состоящей из нуклеотидных последовательностей по пунктам 7-11.
13. Вектор переноса по любому из пунктов 7-11, не содержащий последовательности нуклеиновой кислоты, кодирующей рекомбинантный белок.
14. Вектор переноса по любому из пунктов 7-13, характеризующийся тем, что указанный вектор переноса является бакмидой.
15. Вектор переноса по любому из пунктов 7-14, характеризующийся тем, что указанный вектор переноса является производным любой из бакуловирусных систем экспрессии «Bac-to-Bac®» (invitrogen™), «BacPAK™» (Clontech™), «FlashBAC™» (Oxford Expression Technologies™), «BacuVance™» (GenScript™), «Вас-N-Blue DNA™» (invitrogen™), «BaculoDirect™» (invitrogen™), «BacVector®» 1000, 2000, 3000 (Novagen®), «DiamondBac™» (Sigma-Aldrich®) или» BaculoGold™» (BD biosciences™).
16. Клонирующий вектор, подходящий для получения рекомбинантного бакуловируса или вектора переноса по любому из пунктов 1-15, содержащий указанную последовательность для экспрессии белков IE-0, IE-1 и/или их фрагментов, функционирующих в качестве регуляторов транскрипции, на уровне, превышающем эндогенный уровень, который также подходит для репликации у бактерий.
17. Клонирующий вектор по пункту 16, дополнительно содержащий по меньшей мере одну рекомбинантную гомологичную область (hr) в качестве энхансерной области, функционально связанную с любым промотором, подходящим для управления экспрессией рекомбинантных белков.
18. Клонирующий вектор по пункту 17, отличающийся тем, что рекомбинантная гомологичная область (hr) является последовательностью, указанной в SEQ ID NO: 27 (hr1).
19. Клонирующий вектор по пункту 17 или 18, отличающийся тем, что промотор, функционально связанный с гомологичной областью (hr), выбран из группы нуклеиновых кислот, включающей:
(a) нуклеиновую кислоту, содержащую нуклеотидную последовательность, указанную в любой из SEQ ID NO: 10-16; и
(b) нуклеотидной последовательности, способной функционировать в качестве промотора в рекомбинантном бакуловирусе и обладающей по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью с нуклеотидной последовательностью, указанной в любой из SEQ ID NO: 10-16.
20. Клонирующий вектор по любому из пунктов 16-19, содержащий последовательность нуклеиновой кислоты, содержащую комбинации рекомбинантных промоторов, последовательностей, кодирующих регуляторы транскрипции, и энхансерных областей, выбранных из группы, включающей SEQ ID NO: 17-26.
21. Клонирующий вектор по любому из пунктов 16-20, дополнительно содержащий последовательность нуклеиновой кислоты, кодирующую рекомбинантный белок, причем указанная нуклеотидная последовательность функционально связана с нуклеотидной последовательностью, выбранной из группы, состоящей из нуклеотидных последовательностей по пунктам 16-20.
22. Клонирующий вектор по любому из пунктов 16-20, не содержащий последовательности нуклеиновой кислоты, кодирующей рекомбинантный белок.
23. Нуклеотидная последовательность, подходящая для получения рекомбинантного бакуловируса, вектора переноса или клонирующего вектора по любому из пунктов 1-22, содержащая указанную последовательность для экспрессии белков IE-0, IE-1 и/или их фрагментов, функционирующих в качестве регуляторов транскрипции, на уровне, превышающем эндогенный уровень.
24. Нуклеотидная последовательность по пункту 23, дополнительно содержащая по меньшей мере одну рекомбинантную гомологичную область (hr) в качестве энхансерной области, функционально связанную с любым промотором, подходящим для управления экспрессией рекомбинантных белков.
25. Нуклеотидная последовательность по пункту 24, отличающаяся тем, что рекомбинантная гомологичная область (hr) является последовательностью, указанной в SEQ ID NO: 27 (hr1).
26. Нуклеотидная последовательность по пункту 24 или 25, отличающаяся тем, что промотор, функционально связанный с гомологичной областью (hr), выбран из группы нуклеиновых кислот, включающей:
(a) нуклеиновую кислоту, содержащую нуклеотидную последовательность, указанную в любой из SEQ ID NO: 10-16; и
(b) нуклеотидной последовательности, способной функционировать в качестве промотора в рекомбинантном бакуловирусе и обладающей по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью с нуклеотидной последовательностью, указанной в любой из SEQ ID NO: 10-16.
27. Нуклеотидная последовательность по любому из пунктов 23-26, содержащая нуклеотидную последовательность, содержащую комбинации рекомбинантных промоторов, последовательностей, кодирующих регуляторы транскрипции, и энхансерных областей, выбранных из группы, включающей SEQ ID NO: 17-26.
28. Нуклеотидная последовательность по любому из пунктов 23-27, дополнительно содержащая нуклеотидную последовательность, кодирующую рекомбинантный белок, причем указанная нуклеотидная последовательность функционально связана с нуклеотидной последовательностью, выбранной из группы, состоящей из нуклеотидных последовательностей по пунктам 23-27.
29. Нуклеотидная последовательность по любому из пунктов 23-27, не содержащая последовательности нуклеиновой кислоты, кодирующей рекомбинантный белок.
30. Клетка-хозяин, инфицированная, трансфицированная, трансдуцированная иди трансформированная рекомбинантным бакуловирусом, вектор переноса, клонирующим вектором или нуклеотидной последовательностью по любому из пунктов 1-29.
31. Инфицированная, трансфицированная, трансдуцированная или трансформированная клетка-хозяин по пункту 30, характеризующаяся тем, что является линией клеток насекомого.
32. Инфицированная, трансфицированная, трансдуцированная или трансформированная клетка-хозяин по пунктам 30 или 31, характеризующаяся тем, что происходит от насекомого, принадлежащего к роду Lepidoptera или Diptera.
33. Инфицированная, трансфицированная, трансдуцированная или трансформированная клетка-хозяин по любому из пунктов 30-32, характеризующаяся тем, что происходит от Trichoplusia ni, Spodoptera frugiperda, Ascalapha odorata, Bombyx mori, Drosophila melanogaster или Aedes aegypti..
34. Инфицированная, трансфицированная, трансдуцированная или трансформированная клетка-хозяин по любому из пунктов 30-33, характеризующаяся тем, что является линией клеток, выбранной из группы, состоящей из Hi-5™, Sf9, Sf21, BTI-Tn5B-1, Tn368, ExpresSf+®, BTI-TnAo38, АТС-10 и линии Шнайдера 2 Drosophila.
35. Культуральная среда, содержащая рекомбинантный бакуловирус, вектор переноса, клонирующий вектор или нуклеотидную последовательность по любому из пунктов 1-29.
36. Способ продукции рекомбинантного белка, включающий инфицированную, трансфицированную, трансдуцированную или трансформированную клетку-хозяина по пунктам 30-34 и выделение, и очистку рекомбинантного белка общепринятыми средствами.
37. Способ продукции рекомбинантного белка по пункту 36, отличающийся тем, что рекомбинантный белок выбран из группы, включающей субъединичную мономерную вакцину, субъединичную мультимерную вакцину, вирусоподобную частицу, терапевтический белок, антитело, фермент, цитокин, фактор свертывания крови, антикоагулянт, рецептор, гормон и диагностический белковый реагент.
38. Применение вектора переноса по любому из пунктов 7-15 для получения рекомбинантного бакуловируса по любому из пунктов 1-6.
39. Применение клонирующего вектора по любому из пунктов 16-22 для получения рекомбинантного бакуловируса или вектора переноса по любому из пунктов 1-15.
40. Применение нуклеотидной последовательности по любому из пунктов 23-29 для получения рекомбинантного бакуловируса, вектора переноса или клонирующего вектора по любому из пунктов 1-22.
41. Трансгенная линия клеток, содержащая трансген, который является нуклеиновой кислотой, выбранной из группы, состоящей из:
(a) нуклеиновой кислоты, содержащей нуклеотидную последовательность, указанную в любой из SEQ ID NO: 1-5;
(b) последовательности нуклеиновой кислоты, обладающей по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью с нуклеотидной последовательностью, указанной в любой из SEQ ID NO: 1-5 и кодирующей белок, который может функционировать как регулятор транскрипции в рекомбинантном бакуловирусе;
(c) последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность, содержащую аминокислотную последовательность, указанную в любой из SEQ ID NO: 6-9; и
(d) последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность, обладающую по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% сходством с аминокислотной последовательностью, указанной в любой из SEQ ID NO: 6-9 и способной функционировать как регулятор транскрипции в рекомбинантном бакуловирусе.
42. Трансгенная линия клеток по пункту 41, происходящая из насекомого, птицы или млекопитающего.
43. Способ продукции рекомбинантного белка, включающий выращивание трансгенной линии клеток по пункту 41 или 42 и выделение и очистку рекомбинантного белка общепринятыми средствами.
44. Последовательность нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, функционирующую в качестве промотора в рекомбинантном бакуловирусе, причем указанная нуклеотидная последовательность выбрана из группы, состоящей из:
(a) нуклеиновой кислоты, содержащей нуклеотидную последовательность, указанную в SEQ ID NO: 12, 14 или 15; и
(b) последовательности нуклеиновой кислоты, обладающей по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью с нуклеотидной последовательностью, указанной в SEQ ID NO: 12, 14 или 15;
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1: Схематическое представление элементов рекомбинантной ДНК бакуловируса в соответствии с изобретением, содержащей четыре основных элемента: последовательность, кодирующую регуляторы транскрипции (А; например, IE0 и IE1), экспрессия которых управляется промотором (В; например, polh или рВ29); последовательность энхансерной гомологичной области (hr) (С; например, hr1), расположенную выше промоторов (D; например, p10, polh, рВ29р10 или р6.9р10), управляющих экспрессией чужеродного гена, кодирующего рекомбинантный белок. На схеме показан теоретический механизм взаимодействия между элементами рекомбинантной ДНК в соответствии с настоящим изобретением, приводящий к беспрецедентной сверхэкспрессии рекомбинантного белка.
Фигура 2: Различные стратегии, которые приводят к получению рекомбинатных бакуловирусов с помощью системы клонирования «Bac-to-Bac®» (invitrogen™).
Фигура 3: Общая схема получения клонирующих, донорных векторов и векторов переноса, совместимых с другими коммерческими технологиями, используемыми для создания рекомбинантных бакуловирусов.
Фигура 4: А) Флуориметрические анализы для измерения наработки белка GFP, накапливаемого в инфицированных клетках Sf21 в разное время после инфицирования при экспрессии в бакуловирусных экспрессионных кассетах в соответствии с изобретением, содержащих кДНК hr1p10, hr1pB29p10 или hr1p6.9p10 и Ac-ie-01, экспрессируемую под контролем промоторов рВ29 или polh. Все уровни экспрессии GFP сравнивали с уровнем, получаемым с помощью промотора в обычном бакуловирусном векторе. На графике представлены средние значения трех независимых экспериментов по экспрессии для каждого бакуловируса; в каждом случае стандартное отклонение было менее 5%. На данной фигуре также продемонстрированы типичные микрофотографии флуоресценции, на которых показаны клетки Sf21 в разное время после инфицирования бакуловирусом дикого типа (контроль), обычным бакуловирусом, экспрессирующим GFP под контролем промотора polh или бакуловирусом, экспрессирующим GFP посредством экспрессионной кассеты в соответствии с изобретением polhAc m-01/hr1pB29p10GFP. Клетки инфицированы в случаях А) и В) при множественности заражения (MOI) 5 или в случае С) при MOI 0,1.
Фигура 5: А) Сравнение количества рекомбинантного белка GFP в клетках насекомых Sf21, выращенных в монослое в разное время после инфицирования при экспрессии с помощью обычного бакуловирусного вектора под контролем промотора polh (светло-серый) или бакуловирусного вектора со встроенной экспрессионной кассетой в соответствии с изобретением, состоящей из элементов polhAc-ie-01/hr1pB29p10GFP (темно-серый), измеренного посредством микрофлюидного анализа белка (Experion™; BioRad™, США). Клетки инфицировали обоими вирусами при MOI 5; В) Сравнение рекомбинантного белка GFP, накопленного в клетках насекомых Sf9, выращенных в суспензии в разное время после инфицирования, при экспрессии с помощью обычного бакуловирусного вектора под контролем промотора polh (светло-серый) или бакуловирусного вектора со встроенной экспрессионной кассетой в соответствии с изобретением, состоящей из элементов polhAc-ie-01/hr1p6.p10GFP (темно-серый), измеренного посредством микрофлюидного анализа белка (Experion™; BioRad™, США). Клетки инфицировали обоими вирусами при MOI 0,1; перрывистыми линиями показан процент прироста количества рекомбинантного GFP, продуцируемого бакуловирусами, содержащими экспрессионную кассету в соответствии с изобретением по сравнению с количеством, полученным с помощью обычных бакуловирусов, экспрессирующих GFP под контролем промотора polh; С) Гели ДСН-ПААГ для электрофореза, окрашенные" Кумасси синим, разделяющие экстракты инфицированных клеток после эксперимента, описанного на графике A; D) Гели ДСН-ПААГ для электрофореза, окрашенные Кумасси синим, разделяющие экстракты инфицированных клеток после эксперимента, описанного на графике В.
Фигура 6. Клетки насекомых SF9 культивировали в суспензии и инфицировали бакуловирусом, сверхэкспрессирующим кДНК Ас-ie-01 под контролем polh, или бакуловирусом, экспрессирующим репортерный белок GFP в составе бакуловирусной экспрессионной кассеты polhAc-ie-01/hr1p6.9p10GFP в соответствии с настоящим изобретением, с целью оценки плотности клеток (А) и жизнеспособности (В) данных клеток. В качестве контроля использовали обычный бакуловирус, экспрессирующий белок GFP под контролем polh. Клетки инфицировали в суспензии при MOI 0,1. (А) Клетки подсчитывали в разное время после инфицирования (0, 24 и 48 часов) для вычисления плотности клеток. Более подробный анализ точного момента, в который сверхэкспрессия кДНК Ас-ie-01 приводит к пролиферации клеток, показан на графике-вставке для клеток, инфицированных polhGFP или polhAc-ie-01/hr1p6.9p10GFP. (В) Жизнеспособность клеток оценивали путем окрашивания трипановым синим (1:1 суспензия клеток и краситель в концентрации 0,4% в PBS-буфере). Это окрашивание позволяет различать живые и мертвые клетки. Жизнеспособность клеток рассчитывали, вычисляя процентное содержание живых клеток по отношению к общему количеству клеток в разное время после инфицирования (от 0 до 120 часов). Микрофотографии монослоев клеток насекомых Hi-5™, инфицированных при MOI 5 контрольным обычным бакуловирусом, экспрессирующим репортерный белок GFP под контролем промотора polh (С) или бакуловирусом, экспрессирующим кДНК Ас-ie-01 под контролем промотора polh (D). Микрофотографии получены через 96 часов после инфицирования при 20-кратном увеличении с помощью инвертационного микроскопа Leica™ DMIL™.
Фигура 7: А) Клетки насекомых Sf9 иинфицировали обычным бакуловирусом, экспрессирующим белок GFP под контролем промотора polh (1) или бакуловирусным вектором со встроенной экспрессионной кассетой в соответствии с изобретением, содержащей элементы polhAc-ie-01/hr1p6.9p10GFP, сверхэкспрессирующим регуляторы транскрипции, кодируемые кДНК Ac-ie-01 (2). Клетки отбирали в различные моменты времени после инфицирования (от 0 до 120 часов) и анализировали клеточные экстракты с помощью электрофореза в ДСН-ПААГ и вестерн-блоттинга с антисывороткой против GFP или против актина клеток. В) Функциональность белка GFP, экспрессированного в клетках насекомых Sf9 согласно анализу, показанному на графике А, измеряли с помощью флуориметрии. Значения флуоресценции белка GFP, продуцированного с помощью обычного рекомбинантного бакуловируса, полученные в разное время после инфицирования (серые столбцы), сравнивали с флуоресценцией белка, продуцированного рекомбинантным бакуловирусом с бакуловирусной кассетой в соответствии с настоящим изобретением (черные столбцы).
Фигура 8: Схематическое представление предпочтительных элементов, содержащихся в бакуловирусных экспрессионных кассетах в соответствии с изобретением, содержащих кодирующие последовательности регуляторов транскрипции, гомологичные области (hr), являющиеся энхансерами транскрипции с промотора(ов) чужеродного гена, кодирующего рекомбинантный белок.
Фигура 9: Анализ последовательности промотора рВ2, выделенного из Т. ni, и определение последовательности регуляторного элемента транскрипции рВ29. А) Нуклеотидная последовательность области промотора рВ2 BJHSP2 (номер доступа в GenBank U41640), выделенной из Т. ni. Ранее описанный сайт начала транскрипции обозначен треугольником, в то время как ТАТА-бокс подчеркнут, а потенциальные цис-действующие элементы последовательности рВ2 заключены в прямоугольники. Нуклеотидные остатки с тенью - последовательность, внедренная в промотор рВ29. Прогнозируемые гипотетические сайты связывания Br-С и EcR также обозначены на фигуре прозрачными прямоугольниками В) Схематическое изображение фрагмента ДНК рВ2 и производного от него промотора рВ29. Активность промоторов анализировали С помощью флуоресцентного анализа с использованием белка GFP в качестве репортерного гена. Экспрессию GFP во всех экспериментах количественно определяли через 72 часа после инфицирования; она показана в виде среднеарифметического со стандартным отклонением по трем независимым экспериментам.
Фигура 10: Уровни экспрессии GFP, опосредованной применением различных промоторов или комбинации промоторов. А) Флуориметрический анализ через 24 часа после инфицирования клеток Sf21 различными рекомбинантными бакуловирусами, экспрессирующими GFP под контролем различных отдельных или химерных промоторов. В) Исследование зависимости экспрессии GFP от времени в клетках Sf21, инфицированных теми же рекомбинантными бакуловирусами, что и на графике А, измеряемой с помощью флуоресцентного анализа. Все эксперименты проводили при MOI 5; на фигуре показано среднеарифметическое по трем независимым экспериментам с соответствующими стандартными отклонениями.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение улучшает экспрессию рекомбинантных белков в BEVS за счет внедрения элементов рекомбинантной ДНК в бакуловирусы.
Элементы рекомбинантной ДНК в соответствии с настоящим изобретением представляют собой последовательности, которые вызывают экспрессию бакуловирусных регуляторов транскрипции на уровне, превышающем эндогенный уровень, и, необязательно, энхансерные гомологичные области (hr) и промоторы, функционально связанные с вышеупомянутыми элементами.
Кроме того, элементы рекомбинантной ДНК могут образовывать часть экспрессионной кассеты.
«Экспрессионная кассета» относится к нуклеотидной последовательности, содержащей элементы рекомбинантной ДНК, которые контролируют (например, промотор) и/или требуются (например, сам ген) для экспрессии гена. Экспрессионную кассету можно внедрить в рекомбинантный вектор или бакуловирус.
Элементы рекомбинантной ДНК можно внедрить в одиночную нуклеотидную последовательность, клонирующий вектор, вектор переноса, рекомбинантный бакуловирус или клетку. Тем не менее, они также могут присутствовать в различных нуклеотидных последовательностях, клонирующих векторах, векторах переноса или рекомбинантных бакуловирусах и могут быть внедрены в одну и ту же клетку.
Настоящее изобретение, как ни странно, показывает, что внедрение последовательностей, вызывающих экспрессию бакуловирусных регуляторов транскрипции на уровне, превышающем эндогенный уровень, и, необязательно, внедрение последовательности энхансерной гомологичной области (hr), промотора или комбинации промоторов может увеличить продукцию рекомбинантного белка до беспрецедентного уровня с ранних (от 6 до 8 часов после инфицирования) до поздних (от 48 до 96-120 часов после инфицирования) стадий.
Кроме того, внедрение указанных элементов рекомбинантной ДНК также усиливает пролиферацию клеток, инфицированных бакуловирусом (в частности, на ранних стадиях после инфицирования), жизнеспособность на поздних стадиях после инфицирования и целостность молекулярных механизмов и морфологии указанных клеток, инфицированных бакуловирусом. Улучшение целостности функций клеток во время бакуловирусной инфекции также способствует правильному посттрансляционному процессингу рекомбинантного белка.
Внедрение указанных элементов рекомбинантной ДНК также увеличивает продукцию рекомбинантного белка в клетке-хозяине по сравнению с обычными промоторами polh или p10.
Таким образом, один из аспектов настоящего изобретения относится к рекомбинантному бакуловирусу, содержащему нуклеотидную последовательность, обеспечивающую экспрессию регуляторов транскрипции на уровне, превышающем эндогенный. В предпочтительном варианте реализации регуляторы транскрипции представляют собой IE-1, IE-0 и/или их фрагменты.
«Бакуловирус» относится к семейству инфекционных вирусов беспозвоночных, главным образом заражающих насекомых и членистоногих. «Рекомбинантный бакуловирус» дополнительно содержит рекомбинантную ДНК, внедренную, например, путем гомологичной рекомбинации или транспозиции. Рекомбинантный бакуловирус предпочтительно происходит от AcMNPV.
«Рекомбинантная ДНК» относится к форме искусственной ДНК, созданной путем комбинации или вставки одной или нескольких цепей ДНК, тем самым объединяя ДНК; которые обычно не встречаются совместно.
«Элемент рекомбинантной ДНК» относится к функциональному элементу в составе рекомбинантной ДНК, например, промотору, энхансеру или гену. Как упоминалось выше, элементы рекомбинантной ДНК-в соответствии с настоящим изобретением представляют собой последовательности, которые вызывают экспрессию бакуловирусных регуляторов транскрипции на уровне, превышающем эндогенный уровень, энхансерные гомологичные области (hr) и промоторы, функционально связанные с вышеупомянутыми элементами.
«Регулятор транскрипции» относится к регуляторному белку, который обладает способностью регулировать транскрипцию специфических генов посредством, например, связывания с энхансерной или репрессорной областями и/или рекрутинга дополнительных белков, участвующих в транскрипции.
IE-1 и его сплайс-вариант IE-0 являются регуляторами транскрипции, эндогенно экспрессирующимися во время бакуловирусной инфекции. Согласно настоящему изобретению, IE-1, IE-0 и/или их фрагменты рекомбинантно экспрессируются, повышая общий уровень этих белков выше эндогенного уровня. Этого можно добиться, например, внедряя дополнительные копии эндогенного гена или манипулируя экспрессией промотора эндогенного гена. Кроме того, копии эндогенных генов можно внедрить в виде трансгенов под контролем подходящего промотора, например, polh или рВ29.
Уровень экспрессии белков IE-1, IE-0 и/или их фрагментов можно определять как на уровне мРНК, так и белка с помощью способов, общеизвестных для специалистов в данной области техники, например, количественной ПЦР и вестерн-блоттинга.
В соответствии с настоящим изобретением, IE-1, IE-0 и их фрагменты кодируются нуклеиновыми кислотами SEQ ID NO: 1 (также называемой Ac-ie-01) - SEQ ID NO: 5. SEQ ID NO: 1 является кДНК Ac-ie-01, кодирующей IE-1 и IE-0, SEQ ID NO: 2 является кодирующей последовательностью (CDS) IE-1, a SEQ ID NO: 3 является CDS IE-0. SEQ ID NO: 4 и 5 являются CDS N-концевых доменов IE-1 и IE-0 соответственно, которые сохраняют каталитическую активность регулятора транскрипции. Белки, кодируемые SEQ ID NO: 2-5, представлены SEQ ID NO: 6-9 соответственно.
Кроме того, в настоящем изобретении описаны варианты SEQ ID NO: 1-9, которые являются или кодируют аминокислотные последовательности, способные функционировать в качестве регуляторов транскрипции. Указанные варианты являются нуклеиновыми кислотами или аминокислотами, чьи нуклеотидные или аминокислотные последовательности отличаются от исходных нуклеиновых кислот или аминокислот по одному или нескольким положениям, причем различия могут являться добавлениями, делениями и/или заменами нуклеотидов или аминокислотных остатков.
Нуклеотидные и аминокислотные последовательности в соответствии с настоящим изобретением следует отличать от других нуклеотидных и аминокислотных последовательностей по степени идентичности или сходства последовательностей, соответственно, определяемой с помощью EMBOSS Needle с параметрами по умолчанию (http://www.ebi.ac.uk/Tools/psa/emboss_needle/). Способы получения таких вариантов включают случайный или сайт-специфический мутагенез, сайт-насыщающий мутагенез, сборку фрагментов на основе ПЦР, перестановку ДНК, гомологичную рекомбинацию in vitro or in vivo и способы синтеза генов.
Последовательность вариантов SEQ ID NO: 1-5 по меньшей мере на 70%, предпочтительно по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90% и наиболее предпочтительно по меньшей мере на 95% идентична последовательностям SEQ ID NO: 1-5.
Последовательность вариантов SEQ ID NO: 6-9 по меньшей мере на 70%, предпочтительно по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90% и наиболее предпочтительно по меньшей мере на 95% сходна с последовательностями SEQ ID NO: 6-9.
В предпочтительном варианте реализации рекомбинантный бакуловирус в соответствии с настоящим изобретением дополнительно содержит, помимо нуклеотидной последовательности, обеспечивающей экспрессию белков IE-1, IE-0 и/или их фрагментов на уровне, превышающем эндогенный уровень, рекомбинантную гомологичную область (hr), которая может усилить экспрессию рекомбинантных белков за счет функциональной связи с соответствующим промотором.
Гомологичные области hr состоят из повторяющихся единиц размером приблизительно 70 п.о., содержащих несовершенный 30-п.о палиндром вблизи их центра. Гомологичные области повторяются в восьми местах в геноме AcMNPV, причем на каждой стороне содержатся от 2 до 8 повторов. Гомологичные области включают как энхансеры транскрипции, так и сайты начала репликации ДНК бакуловируса.
«Энхансерная область» относится к регуляторной последовательности, связывание которой с регуляторами транскрипции увеличивает уровень транскрипции соответствующих генов.
«Рекомбинантный белок» относится к белку, происходящему из рекомбинантной ДНК. Такие белки можно применять на благо людей и животных; они могут находить промышленное, коммерческое или терапевтическое применение.
«Быть функционально связанными» относится к двум нуклеотидным последовательностям, связанным таким образом, что одна из них влияет на другую с точки зрения, например, регуляции транскрипции.
«Промотор» относится к последовательности ДНК, с которой связывается РНК-полимераза для начала транскрипции. Указанная последовательность может также содержать сайты связывания различных белков, регулирующих транскрипцию, например, факторов транскрипции. Последовательность промотора может состоять из фрагментов различных промоторов (различных или одинаковых фрагментов), тесно расположенных в последовательности ДНК и, возможно, разделенных линкерами или спейсерами. Такие промоторы называются химерными промоторами.
Последовательность энхансерной гомологичной области hr, расположенная выше промотора(ов), предпочтительно представляет собой hr1 (SEQ ID NO: 27). Промоторы, управляющие экспрессией рекомбинантного белка, предпочтительно выбрана из группы, включающей SEQ ID NO: 10-16 или последовательность, способную функционировать в качестве промотора, и обладает по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью с нуклеотидной последовательностью, указанной в любой из SEQ ID NO: 10-16.
В предпочтительном варианте реализации нуклеотидная последовательность содержит комбинации рекомбинантных промоторов, последовательностей, кодирующих регуляторы транскрипции, и энхансерных областей, выбранных из группы, включающей SEQ ID NO: 17-26.
Как указано выше, рекомбинантные промоторы, последовательности, кодирующие регуляторы транскрипции, и энхансерные области в соответствии с настоящим изобретением не обязательно являются частью одной молекулы; вместо этого указанные последовательности могут образовывать части отдельных молекул при условии, что они функционально связаны, т.е. содержатся в одних и тех же клетках.
Рекомбинантный бакуловирус в соответствии с настоящим изобретением предпочтительно содержит нуклеотидную последовательность, кодирующую рекомбинантный белок. Эта нуклеотидная последовательность функционально связана с нуклеотидной последовательностью, обеспечивающей экспрессию белков IE-1, IE-0 и/или их фрагментов на уровне, превышающем эндогенный уровень, и, необязательно, с гомологичной областью (hr).
В одном варианте реализации настоящего изобретения описана клетка-хозяин, инфицированная, трансфицированная, трансдуцированная или трансформированная рекомбинантным бакуловирусом, вектором переноса, клонирующим вектором или нуклеотидной последовательностью в соответствии с настоящим изобретением. Клетка-хозяин предпочтительно хранится в культуре клеток. Клетка-хозяин предпочтительно является линией клеток насекомых, более предпочтительно - линией клеток, происходящей от насекомого, принадлежащего к роду Lepidoptera или Diptera; в более предпочтительном случае клетка-хозяин происходит от группы, состоящей из Trichoplusia ni, Spodoptera frugiperda, Ascalapha odorata, Bombyx mori, Drosophila melanogaster и Aedes aegypti, а в наиболее предпочтительном случае она выбрана из группы линий клеток насекомых, состоящих из Hi-5™, Sf9, Sf21, BTI-Tn5B-1, Tn368, ExpresSf+®, BTI-TnAo38, АТС-10 и линии Шнайдера 2 Drosophila. Клетку-хозяина можно культивировать; в монослое или в суспензии.
В дополнительном аспекте настоящего изобретения описаны способы продукции рекомбинантных белков с помощью клетки-хозяина в соответствии с настоящим изобретением. После экспрессии рекомбинантного белка выполняют выделение и очистку рекомбинантного белка обычными средствами.
В предпочтительном варианте реализации продукции белка клетку-хозяина культивируют в суспензии (биореакторе) при плотности от 2×106 до 8×106 клеток/мл в зависимости от линии клеток и используемой процедуры ферментации. Кроме того, клетки инфицируют при MOI от 0,1 до 1.
Рекомбинантный белок, предпочтительно продуцируемый с помощью способов в соответствии с настоящим изобретением, является белком, выбранным из группы, включающей субъединичную мономерную вакцину, субъединичную мультимерную вакцину, вирусоподобную частицу, терапевтический белок, антитело, фермент, цитокин, фактор свертывания крови, антикоагулянт, рецептор, гормон и диагностический белковый реагент.
Один из аспектов изобретения связан с применением рекомбинантного бакуловируса, вектора переноса, клонирующего вектора или нуклеотидной последовательности в соответствии с настоящим изобретением в культуральной среде. В предпочтительном варианте реализации культуральная среда содержит бакуловирус в соответствии с настоящим изобретением.
В настоящем изобретении описан вектор переноса, который можно применять для получения рекомбинантного бакуловируса в соответствии с настоящим изобретением и который содержит, помимо последовательности для интеграции или транспозиции в бакуловирус, указанную последовательность для экспрессии белков IE-0, IE-1 и/или их фрагментов на уровне, превышающем эндогенный уровень.
Векторы переноса обычно допускают инсерцию генетической информации в бакуловирус.
Вектор переноса предпочтительно содержит, помимо (i) последовательности Для экспрессии белков IE-0, IE-1 и/или их фрагментов на уровне, превышающем эндогенный уровень, (ii) рекомбинантную гомологичную область (hr), связанную с (iii) промотором, подходящим для управления экспрессией рекомбинантного белка. Предпочтительная комбинации этих элементов рекомбинантной ДНК описана выше для рекомбинантного бакуловируса.
В одном предпочтительном аспекте вектор переноса содержит нуклеотидную последовательность, кодирующую указанный рекомбинантный белок, тогда как в еще одном предпочтительном варианте реализации вектор переноса не содержит такой последовательности.
В предпочтительном варианте реализации вектор переноса является бакмидой.
«Бакмида» относится к плазмидные конструкции, который содержит последовательности ДНК, достаточных для генерации бакуловирусы когда transfected в клетку.
В дополнительном предпочтительном варианте реализации вектор переноса происходит от любой из доступных для приобретения бакуловирусных систем экспрессии «Вас-to-Bac®» (invitrogen™), «BacPAK™» (Clontech™), «FlashBAC™» (Oxford Expression Technologies™), «BacuVance™» (GenScript™), «Вас-N-Blue DNA™» (invitrogen™), «BaculoDirect™» (invitrogen™), «BacVector®» 1000, 2000, 3000 (Novagen®), «DiamondBac™» (Sigma-Aldrich®) или «BaculoGold™» (BD biosciences™).
В настоящем изобретении описан клонирующий вектор, который можно применять для получения рекомбинантного бакуловируса и/или вектора переноса в соответствии с настоящим изобретением и который содержит указанную последовательность для экспрессии белков IE-0, IE-1 и/или их фрагментов на уровне, превышающем эндогенный уровень, которая, в свою очередь, подходит для бактериальной репликации.
«Клонирующий вектор» относится к любому вектору, который подходит для клонирования, что обычно предполагает наличие сайтов рестрикции, сайта начала репликации для размножения в бактериях и селективного маркера.
Клонирующий вектор предпочтительно содержит, помимо (i) последовательности для экспрессии белков IE-0, IE-1 и/или их фрагментов на уровне, превышающем эндогенный уровень, (ii) рекомбинантную гомологичную область (hr), связанную с (iii) промотором, подходящим для управления экспрессией рекомбинантного белка. Предпочтительная комбинации этих элементов рекомбинантной ДНК описана выше для рекомбинантного бакуловируса.
В одном предпочтительном аспекте клонирующий вектор содержит нуклеотидную последовательность, кодирующую указанный рекомбинантный белок, (так называемый «донорный вектор»), в то время как в еще одном предпочтительном варианте реализации клонирующий вектор не содержит такой последовательности.
В настоящем изобретении описана нуклеотидная последовательность, которую можно применять для получения рекомбинантного бакуловируса, вектора передачи и/или клонирующего вектора в соответствии с настоящим изобретением и которая содержит указанную последовательность для экспрессии белков IE-0, IE-1 и/или их фрагментов на уровне, превышающем эндогенный уровень.
Нуклеотидная последовательность предпочтительно содержит, помимо (i) последовательности для экспрессии белков IE-0, IE-1 и/или их фрагментов на уровне, превышающем эндогенный уровень, (ii) рекомбинантную гомологичную область (hr), связанную с (iii) промотором, подходящим для управления экспрессией рекомбинантного белка. Предпочтительная комбинации этих элементов рекомбинантной ДНК описана выше для рекомбинантного бакуловируса.
В одном предпочтительном аспекте указанная нуклеотидная последовательность содержит нуклеотидную последовательность, кодирующую указанный рекомбинантный белок, тогда как в еще одном предпочтительном варианте реализации указанная нуклеотидная последовательность не содержит такой последовательности.
Временная экспрессия кДНК Ас-ie-01 бакуловирусной системой, как ни странно, придает клеткам уникальные свойства устойчивости к вирусу и пролиферации клеток. Это свидетельствует о том, что сверхэкспрессия регуляторов транскрипции, кодируемых указанным геном, активирует другие вирусные или клеточные гены, которые могут отвечать за интересные биотехнологические варианты применения, связанные с увеличением продуктивности и устойчивостью клеток к различным стрессовым воздействиям. Потенциальное применение, вытекающее из данного исследования, может представлять собой получение трансгенных клеток насекомых, птиц или млекопитающих, которые могут представлять собой высокопродуктивные линии клеток для применения при продукции рекомбинантного белка или размножения вируса, и потенциально могут применяться при производстве обычных вакцин на основе ослабленных или инактивированных вирусов.
Следовательно, один из аспектов изобретения относится к линии трансгенных клеток, которая несет в качестве трансгена последовательности SEQ ID NO: 1-5 или их варианты или последовательности, кодирующие белки SEQ ID NO: 6-9 или их варианты, определение которых приведено выше. Линия трансгенных клеток предпочтительно происходит от клеток млекопитающих, птиц или насекомых. В дополнительном варианте реализации линию трансгенных клеток можно применять для продукции рекомбинантного белка, который выделяют и очищают обычными средствами.
«Линия трансгенных клеток» относится к линии клеток, содержащей ген, перенесенный в клетку.
На основании неожиданных результатов, продемонстрировавших, что рекомбинантный бакуловирусный промотор рВ29 (SEQ ID NO: 14) и химерные рекомбинантные бакуловирусные промоторы рВ29р10 (SEQ ID NO: 12) и рВ2р10 (SEQ ID NO: 15) обеспечивают улучшенную экспрессию по сравнению с обычными промоторами, например, рВ2, дополнительный аспект настоящего изобретения относится к нуклеотидным последовательностям, содержащим такие последовательности или варианты последовательностей SEQ ID NO: 12, 14 или 15, которые по меньшей мере на 70%, предпочтительно по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90% и наиболее предпочтительно по меньшей мере на 95% идентичны последовательностям SEQ ID NO: 12, 14 или 15.
СВОДКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ

ХРАНЕНИЕ МИКРООРГАНИЗМОВ СОГЛАСНО БУДАПЕШТСКОМУ ДОГОВОРУ
Плазмиды сданы на хранение в Испанскую коллекцию типовых культур (СЕСТ) (www.cect.org); Университет Валенсии, Parc Científic Universitat de València; Catedrático Agustín Escardino, 9; 46980 Штерна (Валенсия), Испания, с присвоением номера доступа СЕСТ 8031 с 4 октября 2011 г.
ПРИМЕРЫ
Пример 1. Сверхэкспрессия бакуловирусных регуляторов транскрипции стимулирует энхансерную функцию гомологичной области hr, функционально связанной с промотором, повышая экспрессию рекомбинантного белка в бакуловирусной векторной системе экспрессии (BEVS).
Немедленные ранние вирусные белки, кодируемые кДНК Ас-ie-01, т.е. IE-1 и IE-0 AcMNPV являются мощными регуляторами транскрипции в бакуловирусах. Трансактивация, опосредованная данными белками, усиливается за счет их связывания в гомодимерном виде с последовательностями гомологичных областей (hr) бакуловирусов, которые действуют как энхансеры транскрипции. AcMNPV IE-1/IE-0 являются 67-72-кДа димерными ДНК-связывающими белками, которые стимулируют транскрипцию в анализе трансфекции плазмид за счет активности их N-концевого кислого домена (7, 8). Синтезируясь на самых ранних стадиях инфекции, IE-1 и IE-0 накапливаются в ядре, где их содержание поддерживается до поздних стадий. кДНК Ас-ie-01 клонировали под контролем промотора polh с использованием двойной плазмиды pFastBac™ (invitrogen™). В другом локусе этой же плазмиды ниже химерного промотора hr1pB29p10, синтезированного заранее и содержавшего гомологичную область hr1, присоединенную к промоторам рВ29 и p10, клонировали ген, кодирующий GFP. Промотор рВ29 представляет собой фрагмент ДНК, происходящий от промотора рВ2, управляющего экспрессией основного белка 2, супрессируемого ювенильным гормоном (BJHSP-2), у бабочки Т. ni. Фрагмент рВ29, содержащий последовательности, состоящую из 436 нуклеотидов, получили из промотора рВ2, при внедрении в бакуловирусный экспрессионный вектор демонстрировавшего повышенные уровни экспрессии по сравнению с полноразмерным промотором насекомых. Схематическое представление полученной бакуловирусной экспрессионной кассеты в соответствии с настоящим изобретением и предполагаемые функции элементов рекомбинантной ДНК показаны на Фигуре 1. Полученную плазмиду использовали для получения рекомбинантного бакуловируса системой «Bac-to-Bac®» (invitrogen™). Параллельно в той же системе получали обычный бакуловирус, экспрессирующий белок GFP под контролем промотора polh.
Экспрессию белка GFP, опосредованную различными бакуловирусами, исследовали с помощью флуориметрии в различное время после инфицирования в клетках Sf21, культивируемых в монослое. По сравнению с обычными бакуловирусами, экспрессирующими белок под контролем промотора polh, уровень экспрессии гена GFP бакуловирусной экспрессионной кассетой, содержащей регуляторы транскрипции и последовательность энхансера, был приблизительно в 2 раза выше при использовании MOI 5 (Фигура 4а) и в 4 раза выше при использовании MOI 0,1 (данные не показаны). Эти различия накопления белка также наблюдались в клетках Hi-5™ (данные не показаны), что указывало на возможность применения бакуловирусной экспрессионной кассеты для продукции рекомбинантных белков в различных линиях клеток насекомых, используемых в исследованиях и промышленности.
Важно отметить, что при наблюдениях инфицированных клеток с помощью флуоресцентной микроскопии экспрессия GFP, опосредованная бакуловирусной экспрессионной кассетой в соответствии с настоящим изобретением, обнаруживалась на более ранних стадиях и, кроме того, интенсивность флуоресценции инфицированных клеток была значительно выше (Фигура 4В) по сравнению с использованием обычного промотора polh для управления экспрессией GFP. Флуоресцентные клетки обнаруживались уже через 16 часов после инфицирования при инфицировании рекомбинантным бакуловирусом, содержащим экспрессионную кассету в соответствии с настоящим изобретением при MOI 5, и указанная экспрессия GFP возрастала со временем после инфекции (Фигура 4В). Указанные явные различия между новым и контрольным рекомбинантными бакуловирусами наблюдались также при низкой MOI, равной 0,1 (Фигура 4С).
Для анализа влияния промотора, используемого для сверхэкспрессии регуляторов транскрипции, кДНК Ас-ie-01 также клонировали в экспрессионной кассете, описанной выше, под контролем промотора рВ29 вместо промотора polh. Независимо от промотора, используемого для управления экспрессией кДНК Ас-ie-01, накопление GFP было выше, чем накопление, наблюдаемое при использовании обычного бакуловируса, в котором репортерный белок экспрессировали под управлением промотора polh без элементов Ac-ie-01 и hr1 в экспрессионной кассете (Фигура 4А). Аналогичным образом, отсутствие кДНК Ас-ie-01 в экспрессионной кассете, содержащей энхансер hr1, связанный с химерным промотором рВ29р10, также приводило к снижению уровня экспрессии GFP по сравнению с уровнем экспрессии при наличии кДНК Ас-ie-01 (данные не показаны).
Пример 2. Регуляторы транскрипции, кодируемые кДНК Ас-ie-01, стимулируют другие бакуловирусные промоторы, цис-связанные с hr1.
Чтобы определить, повышают ли регуляторы транскрипции, кодируемые Ас-ie-01, в комбинации с последовательностью энхансера экспрессию, опосредованную другими химерными или не химерными бакуловирусными промоторами (р6.9р10 или p10), цис-связанными с энхансером транскрипции - гомологичной областью 1 (hr1), авторы изобретения получили новый набор бакуловирусных экспрессионных кассет и соответствующих им рекомбинантных бакуловирусов AcMNPV. Эти экспрессионные кассеты содержали кДНК Ас-ie-01, клонированную под контролем промотора polh, и ген, кодирующий GFP, расположенный ниже промоторов hr1p10 или hr1p6.9p10. Уровни экспрессии измеряли посредством люминесцентного анализа экстрактов клеток насекомых, инфицированных различными бакуловирусами. В качестве контроля использовали обычный рекомбинантный бакуловирус, экспрессирующий белок GFP под контролем промотора polh. Полученные результаты продемонстрировали, что регуляторы транскрипции, кодируемые кДНК Ас-ie-01, в комбинации с гомологичной последовательностью hr1 также могли стимулировать беспрецедентный уровень экспрессии репортерного белка GFP, экспрессируемого под контролем других бакуловирусных промоторов или комбинации промоторов (химерных) (Фигура 4А). В случае экспрессионной кассеты, использующей химерный промотор, состоящий из р6.9 и p10, продукция рекомбинантного белка была максимальной среди различных проанализированных рекомбинантных бакуловирусов (примерно в 2,5 раза выше по сравнению с контролем через 72 часа после инфицирования).
Дальнейшее количественное определение продукции GFP как в клетках насекомых Sf21 в монослое, так и в клетках насекомых Sf9 в суспензии, опосредованной обычным бакуловирусом или бакуловирусом, содержащим экспрессионную кассету в соответствии с настоящим изобретением, содержащую элементы polhAc-ie-01/hr1pB29p10GFP, проводили посредством микрофлюидного анализа белка (Experion™; BioRad™, США). Клетки насекомых в монослое инфицировали при высокой MOI, равной 5, а в суспензии -при низкой MOI, равной 0,1. На Фигуре 5А показаны процентные доли продуцированного рекомбинантного GFP по отношению к общему количеству растворимых клеточных белков в различное время после инфицирования (от 24 до 72 часов) и относительный рост продуктивности по отношению к обычному бакуловирусу. Ко времени окончания анализа бакуловирус, содержащий экспрессионную кассету в соответствии с настоящим изобретением, достиг уровня рекомбинантного GFP более 40% от общего клеточного белка. Значительные различия в интенсивности полос GFP у исследуемых бакуловирусов были видны в геле для электрофореза в ДСН-ПААГ, окрашенных Кумасси синим, в котором разделяли экстракты инфицированных клеток (Фигура 5С). Когда клетки насекомых культивировали в суспензии и инфицировали каждым бакуловирусом, различия в продуктивности рекомбинантного белка были даже выше, чем различия, обнаруженные в клетках насекомых, культивируемых в монослое и инфицированных при высокой MOI, равной 5. На Фигуре 5В показаны такие различия в продуктивности GFP через 24-120 часов после инфицирования. Интересно, что при использовании бакуловируса с внедренной экспрессионной кассетой в соответствии с изобретением продуктивность рекомбинантного белка увеличилась со временем, достигая максимального уровня через 120 часов после инфицирования. В отличие от этого, уровень рекомбинантного белка, продуцированного обычным бакуловирусом, достиг максимума через 72 часа после инфицирования и снижался в дальнейшем (Фигура 5В). При использовании бакуловирусной экспрессионной кассеты в соответствии с настоящим изобретением на очень поздних стадиях после инфицирования отмечено повышение продуктивности более чем в 20 раз. Аналогичные результаты получены в клетках Hi-5™ (данные не показаны), что свидетельствует о том, что экспрессионную кассету в соответствии с изобретением можно применять для продукции рекомбинантных белков в различных линиях клеток насекомых, используемых в научных исследованиях и промышленности. Значительные различия в интенсивности полос GFP у исследуемых бакуловирусов из клеток, культивируемых в суспензии, были также видны в геле для электрофореза в ДСН-ПААГ, окрашенных Кумасси синим, в котором разделяли экстракты инфицированных клеток (Фигура 5D).
Пример 3. Бакуловирусные экспрессионные кассеты в соответствии с изобретением индуцируют пролиферацию клеток и повышают жизнеспособность клеток за счет регуляторов транскрипции, кодируемых кДНК Ac-ie-01
Авторы изобретения наблюдали с помощью микроскопа, что рекомбинантные бакуловирусы со встроенными бакуловирусными экпрессионными кассетами в соответствии с настоящим изобретением обладают интересными свойствами, связанными со снижением вирус-индуцированных цитопатических эффектов и увеличением плотности клеток в культурах. Для количественной оценки этих явлений и определения элемент(ов) ДНК, ответственных за такие интересные свойства, авторы изобретения получили рекомбинантный бакуловирус, экспрессирующий регуляторы транскрипции, кодируемые кДНК Ас-ie-01 под контролем промотора polh. Указанный бакуловирус совместно с бакуловирусом со внедренными элементами экспрессионной кассеты в соответствии с настоящим изобретению, использованным в примере 2 (polhAc-ie-01/hr1p6.9p10GFP) и обычным бакуловирусом, экспрессирующим белок GFP, применили для инфицирования клеток SJ9 в суспензии при низкой MOI 0,1. Увеличение количества клеток и жизнеспособность клеток изучали в промежуток 24-120 часов после инфицирования. Через 24 часа после инфицирования клетки насекомых, инфицированные любым из бакуловирусов, сверхэкспрессирующих вышеуказанные регуляторы транскрипции, демонстрировали увеличение количества клеток более чем на 10% по сравнению с культурами, инфицированными контрольным рекомбинантным бакуловирусом (Фигура 6А). Более подробный проточно-цитометрический анализ времени, необходимого этим факторам для индукции наблюдаемых различий в пролиферации клеток, показал увеличение количества клеток насекомых в S-фазе через" три часа после инфицирования, а затем через 6 часов после инфицирования наблюдалось увеличение количества клеток насекомых в фазе G1. Эти данные указывают на очень раннее приращение митоза в указанных культурах, инфицированных бакуловирусом, сверхэкспрессирующим белки, кодируемые кДНК Ас-ie-01 (данные не показаны).
Измерение флуоресценции проводили на проточном цитометре FACSCalibur™ (BD Biosciences™). Клетки фиксировали в 70% EtOH, ресуспендировали и инкубировали в окрашивающем растворе (50 мкг/мл иодида пропидия в PBS, 5 мкг/мл РНКазы). Данные фильтровали для элиминирования частиц, размер которых отличался от размера клеток, и анализировали путем построения графика количества клеток от красной флуоресценции иодида пропидия. Подсчитывали 50000 клеток на анализ. Анализ общего количества клеток в фазе клеточного цикла (G1, S, G2) выполняли с использованием программного обеспечения Modfit.
Инфицированные культуры клеток также анализировали путем окрашивания трипановым синим для определения жизнеспособности клеток в различные моменты времени после инфицирования. Интересно, что на очень поздней стадии после инфицирования (96-120 часов) клетки насекомых, инфицированных вирусами, сверхэкспрессирующими регуляторы транскрипции, демонстрировали увеличение (на 50-60%) жизнеспособности и целостности клеток (Фигура 6В). Это говорит о том, что сверхэкспрессия регуляторов транскрипции в соответствии с настоящим изобретением защищает клетки от бакуловирус-индуцированного цитопатического действия, обеспечивая долгосрочную экспрессию. Как пролиферация клеток, так и увеличение жизнеспособности клеток после инфицирования имеют важные последствия для продуктивности рекомбинантного белка в BEVS. Аналогичные результаты были получены при сверхэкспрессии регуляторов транскрипции под управлением промоторов рВ29 или polh (данные не показаны). Результаты, полученные в клетках насекомых Sf9, инфицированных в суспензии, подтвердились в клетках Sf21, культивируемых в монослое (данные не показаны), а также в клетках Hi-5™, культивируемых в монослое (Фигура 6С и D). На указанных фигурах показано, как сверхэкспрессия регуляторов транскрипции улучшает целостность клеток на поздних стадиях после инфицирования (96 часов).
Пример 4. Сверхэкспрессия регуляторов транскрипции, кодируемых кДНК Ac-ie-01, в бакуловирусной системе экспрессии облегчает посттрансляционный процессинг рекомбинантных белков.
Целостность клеток при бакуловирусной инфекции имеет большое значение для правильного фолдинга или других посттрансляционных модификаций чужеродных белков, экспрессируемых с помощью данной системы. Бакуловирусные сильные промоторы, обычно применяемые для научных исследований и продукции, например, polh и p10, экспрессируют чужеродные гены только на поздних стадиях после инфицирования, когда инфицированные клетки уже демонстрируют серьезные цитопатические эффекты и жизнеспособность клеток снижается. Как описано выше, сверхэкспрессия регуляторов транскрипции, используемых в бакуловирусной экспрессионной кассете в соответствии с настоящим изобретением, защищает клетки от патогенных эффектов бакуловирусной инфекции посредством пока неизвестного механизма, обеспечивая широкий интервал времени для продукции рекомбинантного белка в клетках, остающихся полностью жизнеспособными. Авторы изобретения исследовали важность элементов, внедренных в экспрессионную кассету в соответствии с изобретением, для посттрансляционных модификаций рекомбинантных белков. Для этой цели обычные бакуловирусы, экспрессирующие репортерный белок GFP под контролем промотора polh, и бакуловирус с внедренной бакуловирусной кассетой в соответствии с настоящим изобретением, также экспрессирующий белок GFP (polhAc-ie-01/hr1p6.9p10GFP), использовали для инфицирования клеток насекомых Sf9 в суспензии при MOI 0,1. Инфицированные клетки анализировали в различные моменты времени после инфицирования посредством вестерн-блоттинга с использованием моноклонального антитела против GFP (mab2515; Millipore™), как показано на Фигуре 7. Интересно, что белок GFP, экспрессируемый обычным бакуловирусом, через 48 и 72 часа после инфицирования демонстрировал несколько реакционноспособных полос (что указывало на неправильную экспрессию и/или фолдинг белка), а на более поздних стадиях наблюдалась полоса с пониженной молекулярной массой (ниже прогнозируемой) (что указывало на разложение) (Фигура 7А). В отличие от этого, если экспрессия белка GFP обеспечивалась бакуловирусной экспрессионной кассетой в соответствии с изобретением, на всех анализируемых стадиях после инфицирования наблюдали только одну полосу GFP с ожидаемой для этого белка молекулярной массой (Фигура 7А). Экспрессия GFP с помощью этого вектора не снижалась в значительной степени на очень поздних стадиях после инфицирования (120 часов), что подтверждает, что бакуловирусная экспрессионная кассета в соответствии с изобретением придает бакуловирусным векторам интересные преимущества для долгосрочной экспрессии.
Параллельно в различное время после инфицирования измеряли целостность механизмов клетки посредством вестерн-блоттинга белка-актина клеток с использованием специфической антисыворотки (Фигура 7А). Инфицирование обычным бакуловирусом серьезно нарушало целостность клеток через 72 часа после инфицирования, поскольку обнаруженная полоса актина резко уменьшалась после этого момента времени (разложение в результате полной потери целостности клеток). В соответствии с защитой клеток, индуцируемой элементами рекомбинантной ДНК, содержащимися в бакуловирусной экспрессионной кассете в соответствии с изобретением, степень воздействия на актин клеток отличалась в клетках, инфицированных рекомбинантным бакуловирусом с внедренной экспрессионной кассетой.
Флуоресцентная активность рекомбинантного GFP, экспрессируемого различными бакуловирусами, отражает его правильную конформацию. Как показано на фигуре 7В, GFP, экспрессируемый в бакуловирусной экспрессионной кассете в соответствии с изобретением, сохранял картину повышенной функциональности на протяжении различных стадий инфекции. В отличие от этого, флуоресценция GFP, экспрессируемого обычным бакуловирусом, достигала максимума через 72 ч после инфицирования и снижалась в последующие моменты времени (Фигура 7В) параллельно с разложением актина (Фигура 7А) и наблюдаемым снижением жизнеспособности (Фигура 6В).
Пример 5. Клеточные культуры и вирусы.
Линии клеток Spodoptera frugiperda Sf21 или Sf9 культивировали в 6-луночных планшетах для тканевых культур (1×106 клеток/лунку) в среде для клеток насекомых TNM-FH (Pan Biotech™, Германия), содержащей 10% эмбриональной телячьей сыворотки, инактивированной нагреванием (Pan Biotech™, Германия), при 27°С. Рекомбинантные бакуловирусы AcMNPV получили с помощью бакуловирусной системы экспрессии «Вас-to-Bac®» (Invitrogen™, США). Различные векторы переноса, содержащие элементы рекомбинантной ДНК в соответствии с настоящим изобретением, получили с помощью плазмиды pFastBac™-DUAL (Invitrogen™). Промоторы и регуляторные элементы, внедренные в pFastBac™-DUAL, синтезировали (GenScript™, США) с использованием адекватных фланкирующих рестрикционных последовательностей для облегчения клонирования. Указанные векторы переноса использовали для трансфекции клеток Sf21 c помощью Cellfectin® (Invitrogen™, США). Затем рекомбинантные бакуловирусы, полученные в результате инфицирования клеток Sf21, дважды пересевали в клетки и титровали с помощью анализа бляшек. Полученные генные конструкты бакуловирусных экспрессионных кассет в соответствии с настоящим изобретением схематически показаны на фигуре 8, где изображены различные потенциальные комбинации промоторов, управляющих экспрессией кДНК Ас-ie-01 или чужеродного гена (например, GFP); Различные экспрессионные кассеты использовали для получения рекомбинантных бакуловирусов, использованных в примерах, показанных на фигурах 4-7.
Пример 6. Получение клонирующего вектора
Клонирующий вектор является небольшим фрагментом ДНК, содержащим бакуловирусную экспрессионную кассету в соответствии с изобретением, в которую можно встроить фрагмент чужеродной ДНК путем обработки средой-носителем и чужеродной ДНК ферментом рестрикции, создающим одинаковый липкий конец, с последующим лигированием фрагментов друг с другом. Основные характеристики клонирующего вектора должны включать синтетический сайт множественного клонирования (MCS) для облегчения инсерции чужеродных генов в выбранной ориентации, селективный маркер, например, устойчивость к антибиотикам, для отбора положительно трансформированных клеток и функциональный сайт начала репликации (ORI) для размножения в бактериях.
Пример 7. Получение донорного вектора, содержащего бакуловирусную экспрессионную кассету в соответствии с изобретением
Донорный вектор состоит из клонирующего вектора, например, плазмиды pUC57, содержащего бакуловирусную экспрессионную кассету, в которой клонирован чужеродный ген с помощью соответствующих ферментов рестрикции. Бакуловирусную экспрессионную кассету в соответствии с изобретением синтезировали путем лигирования следующих последовательностей ДНК: (i) кодирующей последовательности бакуловирусного регулятора транскрипции Ас-ie-01 ниже последовательности промотора; например, промотора polh или рВ29, и выше сигнала полиаденилирования HSV ТК и (ii) последовательности энхансера, например, гомологичной области hr1, в другом локусе выше (iii) последовательности промотора, например, pB29p10, p10, р6.9р10 или polh, с последующим сайтом множественного клонирования (MCS) для клонирования интересующего гена и сигналом полиаденилирования SV40 ниже MCS (Фигура 1). Бакуловирусную экспрессионную кассету фланкировали специфическими сайтами рестрикции (например, BglII и BstZ17l с 5'-конца и Bgl II и Sgf I с 3'-конца) для облегчения субклонирования в векторе переноса доступной для приобретения системы получения бакуловируса (на основе транспозиции, например, системы «Вас-to-Bac®» (invitrogen™) или на основе гомологичной рекомбинации, например, «flashBAC™» (Oxford Expression Technologies™), «Baculogold™» (BD Biosciences™), «BacPAK6™» (Clontech™), «Вас-N-Blue DNA™» (invitrogen™) (Фигуры 2 и 3).
Кодирующий ген зеленого флуоресцирующего белка (GFP) клонировали в MCS клонирующего вектора, используя сайты рестрикции Nco I и Spe I, с получением донорного плазмидного вектора (Фигура 2).
Пример 8. Получение вектора переноса, содержащего бакуловирусную экспрессионную кассету в соответствии с изобретением
Вектор переноса получили путем гидролиза донорного вектора с использованием BstZ17l с 5'-фланкирующего сайта и Xba I и его клонирования в векторе переноса pFastBac™1, который также гидролизовали с использованием этих же ферментов. В этом случае в результате субклонирования сигнал полиаденилирования SV40 бакуловирусной экспрессионной кассеты заменялся на сигнал полиаденилирования SV40 из вектора переноса. Кроме того, все элементы экспрессионной кассеты внедряли в вектор переноса pFastBac, заменяя промотор polh и MCS первоначального коммерческого вектора переноса (Фигура 2).
Пример 9. Получение бакуловирусного экспрессионного вектора, содержащего бакуловирусную экспрессионную кассету в соответствии с настоящим изобретением, с использованием системы «Bac-to-Bac®»
Модифицированный вектор переноса pFastBac™1 и отдельную бакуловирусную экспрессионную кассету использовали для получения рекомбинантных бакуловирусов с помощью системы экспрессии бакуловирусов «Bac-to-Вас®». Конкретнее, модифицированный вектор переноса использовали для трансформации штамма-хозяина Е.coli DH10Bac™, содержащего бакуловирусный челночный вектор (бакмиду) и плазмиду-помощник и обеспечивавшего получение рекомбинантной бакмиды после транспозиции экспрессионной кассеты. Затем ДНК рекомбинантной бакмиды, содержащую бакуловирусную экспрессионную кассету в соответствии с настоящим изобретением и ген, кодирующий GFP, использовали для трансфекции клеток насекомых, например, клеток Sf21, с помощью Cellfectin®. Через 72 часа после трансфекции собирали клетки получали первое поколение рекомбинатного бакуловируса (Фигура 2). Указанный рекомбинантный бакуловирус можно амплифицировать дальше и/или титровать согласно следующим обычным протоколам. Аналогичные процедуры можно применять для получения рекомбинантных бакуловирусов с другими векторами переноса, полученными из коммерческих BEVS (Фигура 3).
Пример 10. Получение образца белка.
Инфицированные клетки в каждый момент времени (1×106) собирали и центрифугировали. Надосадочную жидкость удаляли, а осажденные клетки ресуспендировали в PBS и подвергали трем циклам замораживания (-196°С) и оттаивания (37°С). Разрушенные клетки удаляли центрифугированием.
Пример 11. Исследование зависимости экспрессии белка от времени.
Клетки Sf9, Sf21 или Hi-5™ инфицировали различными рекомбинантными бакуловирусами, экспрессирующими GFP под контролем различных комбинаций регуляторных, энхансерных и промоторных элементов, при MOI 5 или 0,1. Культур клеток собирали в различные моменты времени (через 24, 48, 72, 96 и 120 часов после инфицирования) и анализировали экспрессию гена GFP путем флуоресцентной микроскопии, флуориметрического анализа, электрофореза в ДСН-ПААГ с последующим окрашиванием Кумасси синим или вестерн-блоттинга и микрофлюидного разделения и количественного определения (система автоматизированного электрофореза Experion™; Bio-Rad™, США).
Для количественного определения рекомбинантного GFP образцы загружали в чипы Pro260 (Bio-Rad™) и анализировали посредством капиллярного электрофореза с использованием системы Experion (Bio-Rad), следуя инструкциям производителя. Электрофорез образцов через микроканалы выполняли путем контроля прилагаемого напряжения и электрической мощности. Микрофлюидный чип обеспечивал несколько последовательных процедур, в том числе разделение, окрашивание, удаление красителя, обнаружение и базовый анализ данных без вмешательства пользователя. Система Experion™ выполняла разделение и количественное определение образцов белка размером от 10 до 260 кДа с высокой чувствительностью, сопоставимой с электрофорезом в ДСН-ПААГ с окрашиванием коллоидным Кумасси синим. Для количественной оценки система Experion™ использовала лэддер Pro260, модифицированную версию стандартов Precision Plus Protein™, оптимизированную для применения в данной системе.
Пример 12. Анализ флуоресцентной микроскопией
Инфицированные клетки визуализировали непосредственно в 6-луночных планшетах для культивирования клеток с использованием фильтра GFP на инвертационном флуоресцентном микроскопе Leica™ DMIL™.
Пример 13. Флуориметрический анализ.
Приблизительно 20 мкг общего растворимого белка, полученного из инфицированных клеток, содержащих различное количество рекомбинантного белка GFP, анализировали и количественно оценивали с помощью флуоресцентного планшет-ридера Tecan™ GENios™ (штат Калифорния, США) (возбуждение [Ех], 485/излучение [Em], 535).
Пример 14. Вестерн-блоттинг.
Фракции общего растворимого белка (10 μg) из клеток, инфицированных рекомбинантными бакуловирусами, разделяли электрофорезом в 15% ДСН-ПААГ. Гели окрашивали Кумасси синим или переносили на нитроцеллюлозные мембраны. Результаты вестерн-блоттинга обрабатывали зондом -моноклональным антителом mab2515 против GFP (Millipore™, США) при разбавлении 1:1000 или антисыворотки против актина (20-33; Sigma-Aldrich™), выявляли иммунокомплексы с помощью конъюгата антитела против IgG мыши, меченого пероксидазой хрена (ПХ) (KPL™, Великобритания) при разбавлении 1:2000 или конъюгата антитела против IgG кролика, меченого пероксидазой хрена (ПХ) (KPLTM, Великобритания) при разбавлении 1:2000 соответственно в качестве вторичного антитела. Полосы белка обнаруживали с помощью системы обнаружения вестерн-блоттинга ECL и анализировали системой визуализации геля ChemiDoc™ XRS Gel Imaging System (Bio-Rad™, США).
Пример 15. Определение границ последовательности промотора во фрагменте ДНК рВ2.
Область ДНК выше гена BJHSP-2 (рВ2) выделяли из ДНК насекомого Т. ni ПЦР-амплификацией на основе ранее описанной последовательности BJHSP2 (номер доступа в GenBank U41640). Амплифицированная область ДНК в некоторых аспектах отличалась от аннотированной последовательности и содержала 2 инсерции, 8 делеций, а также 17 мутаций (SEQ ID NO: 34). С помощью различных биоинформатических анализов в последовательности рВ2 обнаружили шесть предполагаемых сайтов связываний, связанные с элементами ответа на гормон, четыре из которых соответствовали предполагаемым элементам ответа на экдизол (ECR), а три - предполагаемым сайтам Broad-Complex (Br-С) (Фигуры 9А и В).
Для определения основных регуляторных областей для транскрипционной активности и значимости потенциальных гормон-регулируемых элементов анализировали укороченную последовательность рВ2 (фрагмент рВ29) (Фигура 9В). Фрагменты рВ2 и рВ29 клонировали в бакуловирусном векторе и тестировали на промоторную активность с помощью репортерного белка GFP. На фигуре 9В показаны уровни экспрессии гена GFP, полученные для каждого фрагмента и количественно проанализированные путем флуориметрического анализа. Как ни странно, фрагмент рВ29 демонстрировал более сильную промоторную активность, чем исходный полноразмерный рВ2, даже несмотря на отсутствие двух предполагаемых сайтов связывания Br-С (Фигура 9В).
Пример 16. Синергическое взаимодействие между рВ2 или рВ29 и обычными бакуловирусными промоторами.
Сконструировали различные химерные промоторы, содержащие последовательность рВ2 или рВ29 и обычные бакуловирусные промоторы polh или p10, что привело к получению комбинаций pB2polh, polhpB2, рВ2р10, р10рВ2 и рВ29р10. Все из них использовали для получения рекомбинантных бакуловирусов и тестировали на промоторные характеристики. Через 24 часа после инфицирования экспрессия белка GFP под управлением гибридных промоторов polhpB2 и рВ29р10 была повышенной по сравнению» обычными промоторами polh или p10 (Фигура 10А). Интересно, что экспрессия GFP под контролем химерного промотора polhpB2 упала через 48 часов после инфицирования, в то время как экспрессия GFP под контролем химерного промотора рВ29р10 повышалась со временем даже до более высоких уровней, чем с помощью промотора polh (Фигура 10В). Химерный промотор рВ2р10 демонстрировал максимальный уровень экспрессии GFP через 48 часов после инфицирования, но рост экспрессии на поздних стадиях был нелинейным (Фигура 10В). В заключение отметим, что гибридный промотор рВ29р10 являлся более ранним и сильным, чем промоторы polh или p10 по отдельности.
БИБЛИОГРАФИЯ
1. Nettleship, J.E., Assenberg, R., Diprose, J.M., Rahman-Huq, N., Owens, R.J. Recent advances in the production of proteins in insect and mammalian cells for structural biology. J. Struct. Biol. 2010, 172, 55-65.
2. Gomez-Casado E, Gomez-Sebastian S, Núñez MC, Lasa-Covarrubias R, Martínez-Pulgarín S, Escribano JM. Insect larvae biofactories as a platform for influenza vaccine production. Protein Expr Purif. 79: 35-43. 2011.
3. Smith, G.E., M.D. Summers, and M.J. Fraser. 1983. Production of human beta interferon in insect cells infected with a baculovirus expression vector. Mol. Cell. Biol. 3: 2156-21 65.
4. Hitchman RB, Possee RD, King LA. Baculovirus expression systems for recombinant protein production in insect cells. Recent Pat Biotechnol. 2009; 3(1): 46-54.
5. Hashimoto Y, Zhang S, Blissard GW. Ao38, a new cell line from eggs of the black witch moth, Ascalapha odorata (Lepidoptera: Noctuidae), is permissive for AcMNPV infection and produces high levels of recombinant proteins. BMC Biotechnol. 2010, 10:50.
6. Taticek RA, Choi C, Phan SE, Palomares LA, Shuler ML. Comparison of growth arid recombinant protein expression in two different insect cell lines in attached and suspension culture. Biotechnol. Prog. 2001, 17 (4), 676-684.
7. Hill-Perkins MS, Possee RD. A baculovirus expression vector derived from the basic protein promoter of Autographa californica nuclear polyhedrosis virus. J Gen Virol. 1990, 71 (Pt4): 971-6.
8. Passarelli, A. L., and L. K. Miller. Three baculovirus genes involved in late and very late gene expression: ie-1, ie-n, and lef-2. J. Virol. 1993, 67: 2149-2158
9. Rodems, S. M., S. S. Pullen, and P. D. Friesen. DNA-dependent transregulation by IE1 of Autographa californica nuclear polyhedrosis virus: IE1 domains required for transactivation and DNA binding. J. Virol. 1997, 71: 9270-9277.
10. Lin X, Chen Y, Yi Y, Zhang Z: Baculovirus immediately early 1, a mediator for homologous regions enhancer function in trans. Virol J 2010, 7:32.
11. Okano K, Mikhailov VS, Maeda S: Colocalization of baculovirus IE-1 and two DNA-binding proteins, DBP and LEF-3, to viral replication factories. Journal of virology 1999, 73(1): 110-119.
| название | год | авторы | номер документа |
|---|---|---|---|
| AAV ВЕКТОРЫ С УСОВЕРШЕНСТВОВАННЫМИ Rep-КОДИРУЮЩИМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ, ИСПОЛЬЗУЕМЫМИ В СИСТЕМАХ ПРОДУКЦИИ НА ОСНОВЕ КЛЕТОК НАСЕКОМЫХ | 2007 |
|
RU2457252C2 |
| СПОСОБЫ УСИЛЕНИЯ БИОЛОГИЧЕСКОЙ АКТИВНОСТИ РЕКОМБИНАНТНОГО АДЕНОАССОЦИИРОВАННОГО ВУРУСА, ПРОДУЦИРУЕМОГО БАКУЛОВИРУСНОЙ СИСТЕМОЙ | 2017 |
|
RU2762948C2 |
| РЕКОМБИНАНТНАЯ ЭКСПРЕССИЯ БЕЛКА ORF2 PCV2b В КЛЕТКАХ НАСЕКОМЫХ | 2018 |
|
RU2779423C2 |
| РЕКОМБИНАНТНЫЙ ВИРУС, СПОСОБНЫЙ К УСТОЙЧИВОЙ ЭКСПРЕССИИ БЕЛКОВ-МИШЕНЕЙ | 2019 |
|
RU2816428C2 |
| РЕКОМБИНАНТНЫЙ НЕПАТОГЕННЫЙ MDV-ВЕКТОР, ОБЕСПЕЧИВАЮЩИЙ ПОЛИСПЕЦИФИЧЕСКИЙ ИММУНИТЕТ | 2012 |
|
RU2593950C2 |
| Кодон-оптимизированная нуклеиновая кислота, которая кодирует белок SMN1, и ее применение | 2020 |
|
RU2742837C1 |
| ПРОДУЦИРОВАНИЕ AAV В КЛЕТКАХ НАСЕКОМЫХ, СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЭТОГО | 2017 |
|
RU2739596C2 |
| Синергетическое действие SMN1 и miR-23a при лечении спинальной мышечной атрофии | 2021 |
|
RU2839898C2 |
| РЕКОМБИНАНТНЫЕ ВИРУСЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2835626C2 |
| ИММУНИЗАЦИЯ ПРОТИВ CHLAMYDIA TRACHOMATIS | 2002 |
|
RU2331435C2 |















































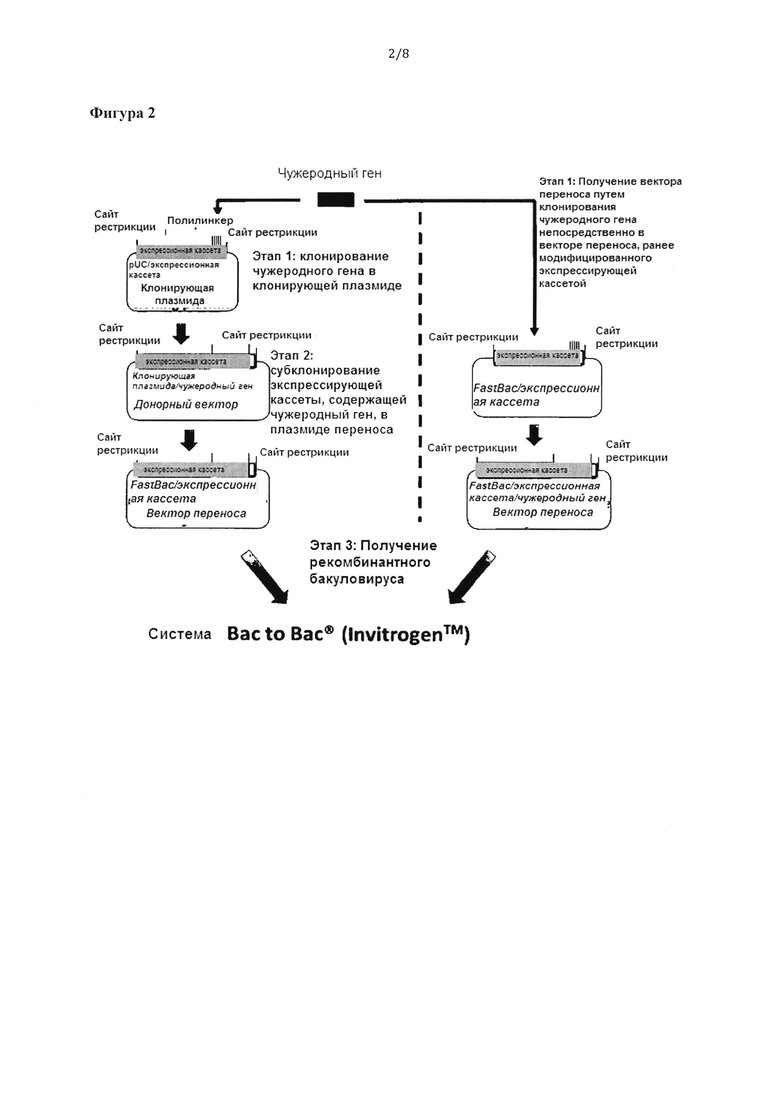
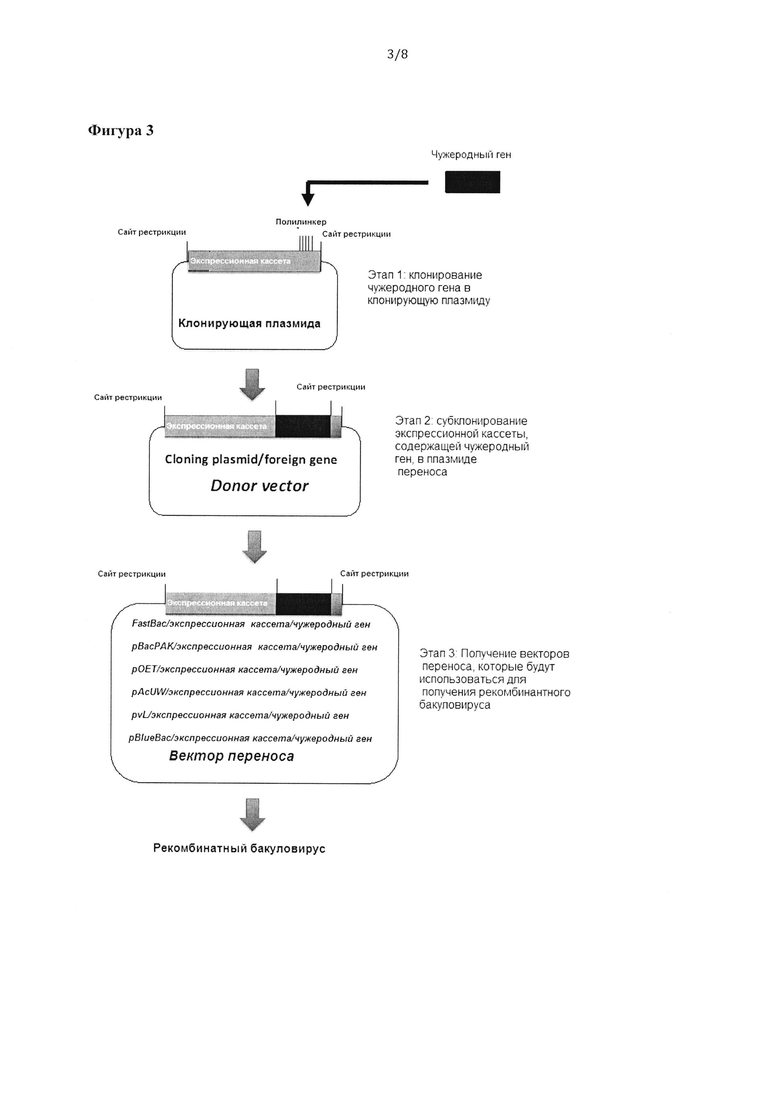
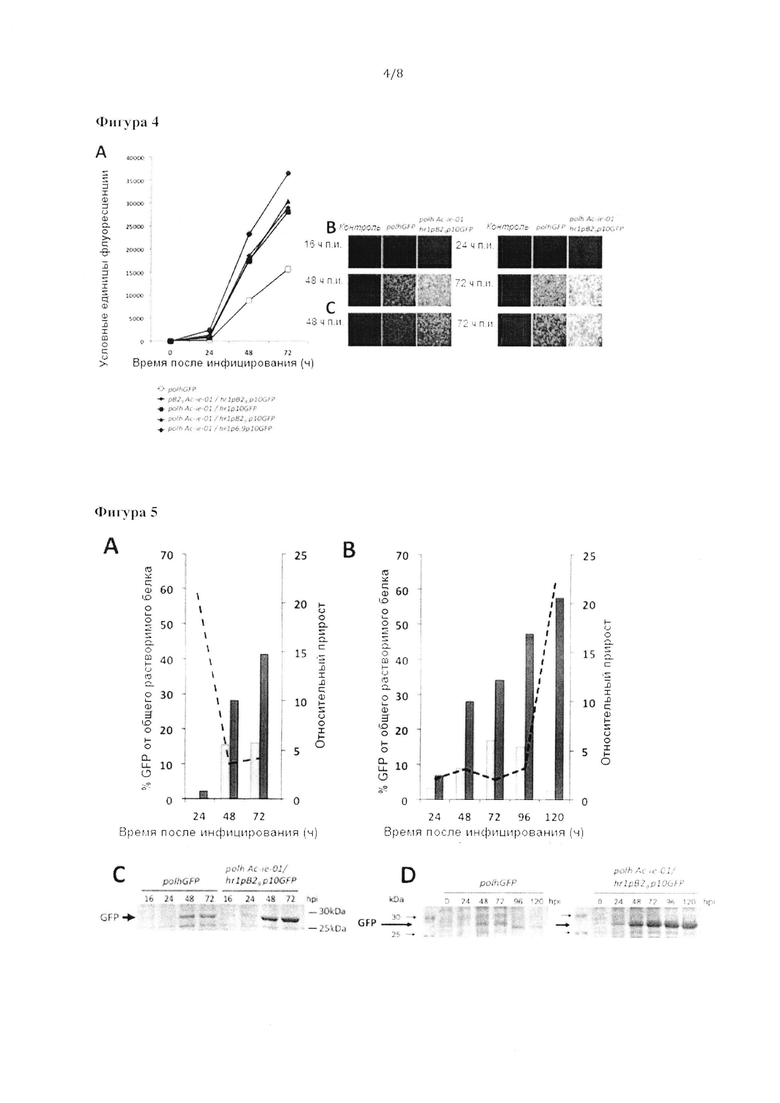
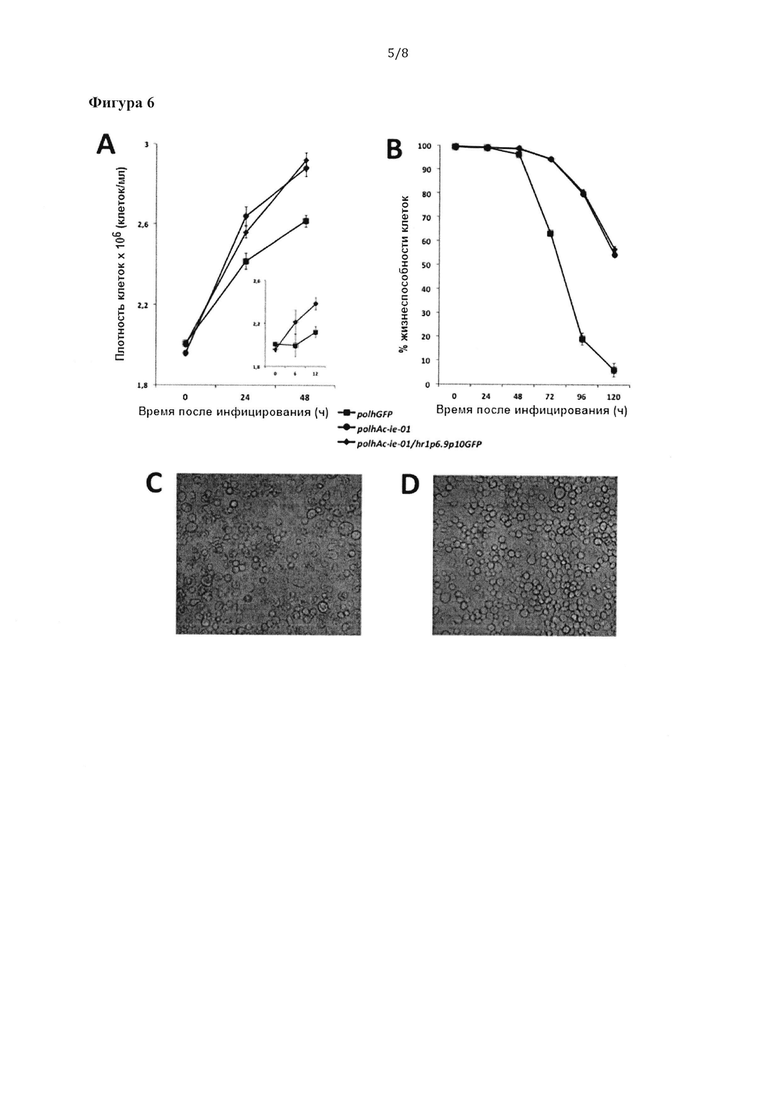
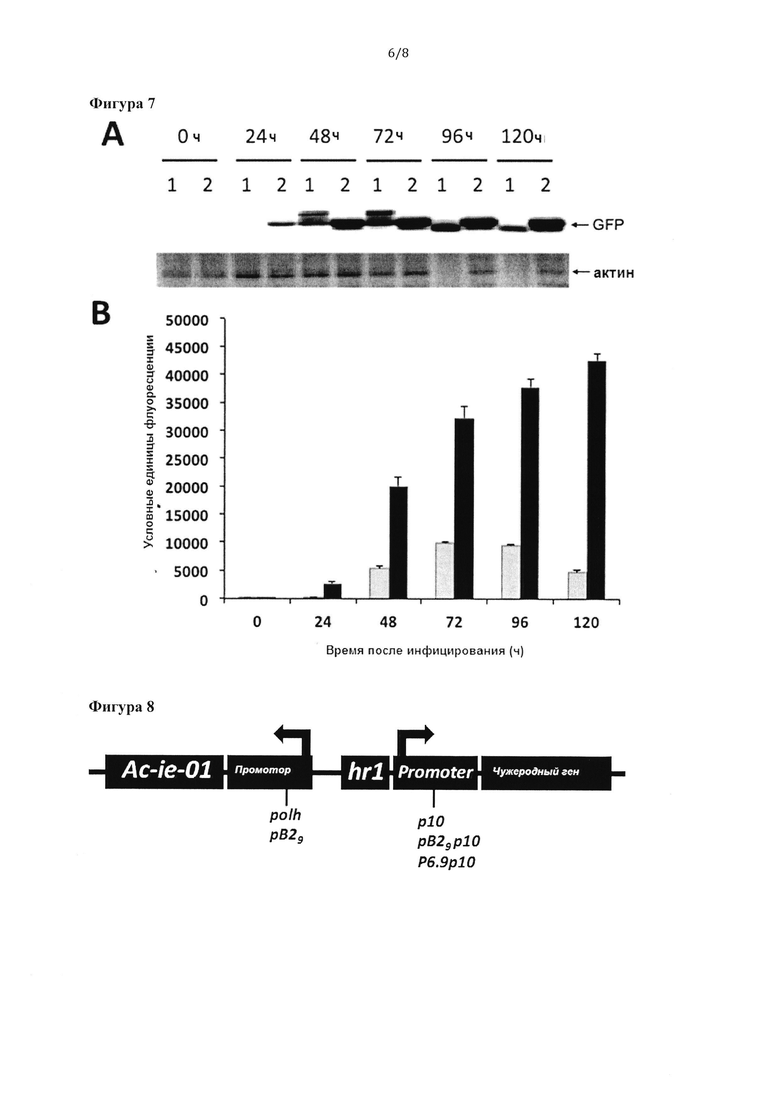
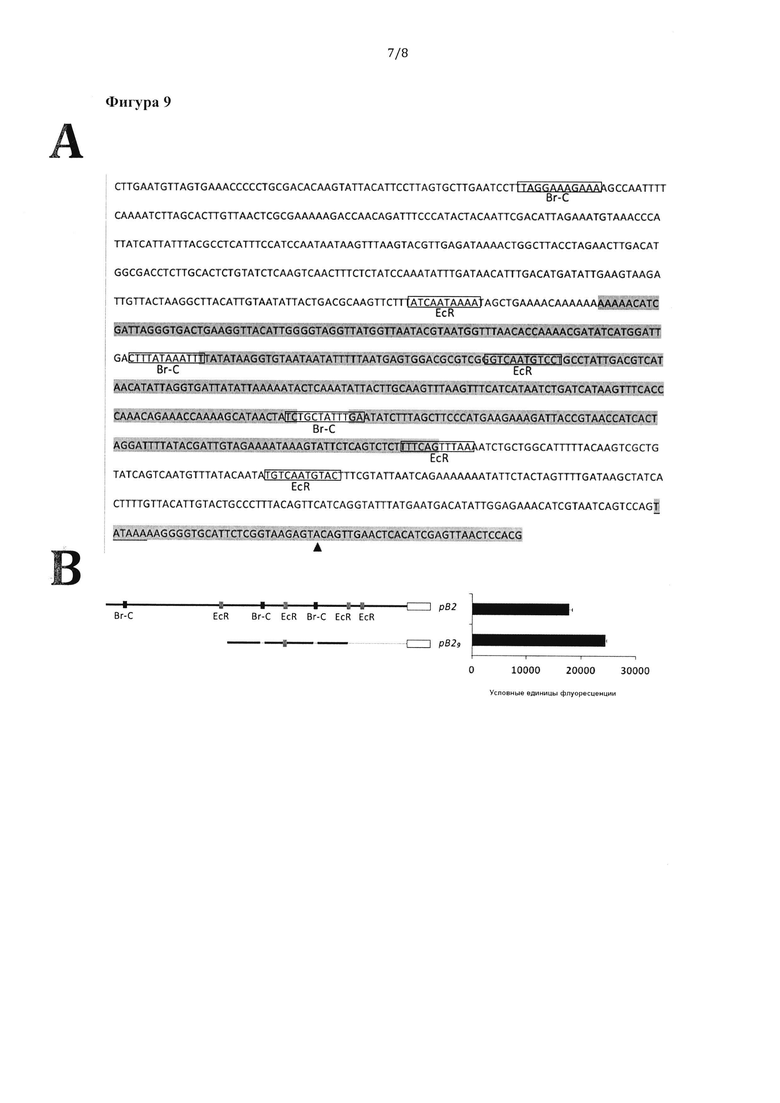
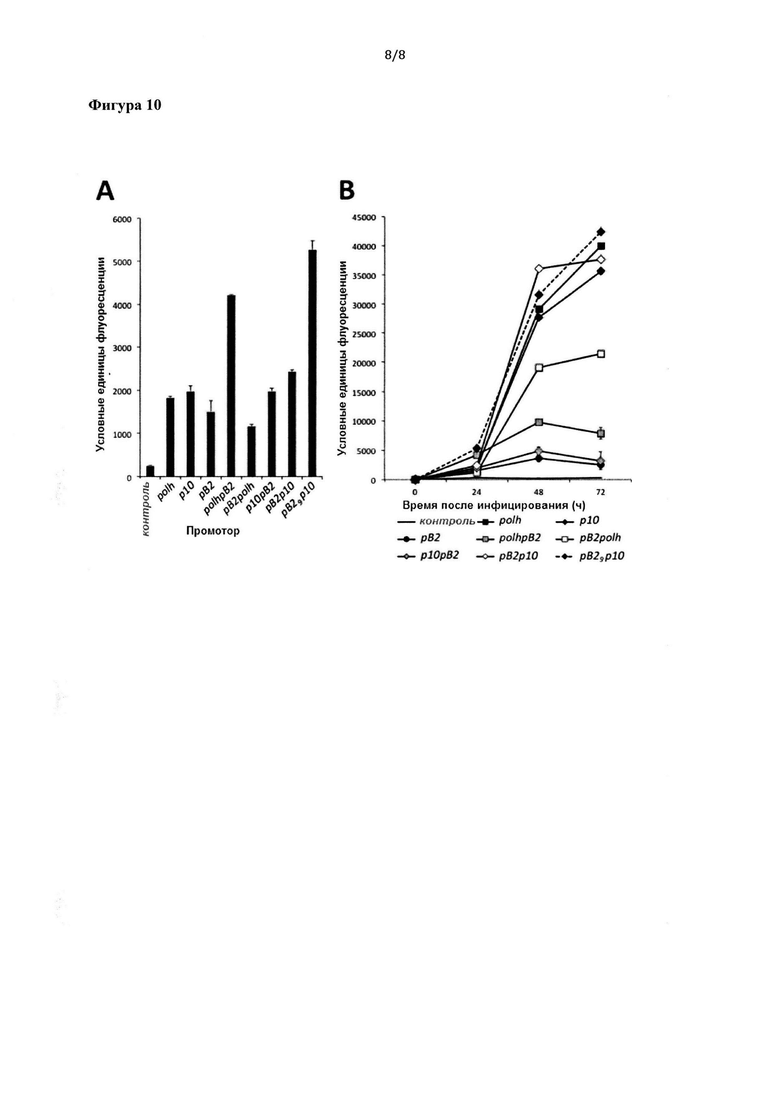
Настоящая группа изобретений относится к биотехнологии. Предложены рекомбинантный бакуловирус для экспрессии рекомбинантного белка, вектор переноса, клонирующий вектор и последовательность нуклеиновой кислоты, подходящие для получения указанного бакуловируса, и их применение, а также клетка-хозяин и способ продукции рекомбинантного белка. Предложенный рекомбинантный бакуловирус содержит последовательность нуклеиновой кислоты, обеспечивающую экспрессию белков IE-1 или IE-0 на уровне, превышающем эндогенный уровень, рекомбинантную гомологичную область в качестве энхансерной области, функционально связанную с промотором, и последовательность нуклеиновой кислоты, кодирующую рекомбинантный белок. Строение предложенного бакуловируса обеспечивает повышенный уровень экспрессии целевого рекомбинантного белка. Предложенная группа изобретений может быть использована для получения рекомбинантных белков с использованием бакуловирусных векторных систем экспрессии в клетках насекомых. 9 н. и 8 з.п. ф-лы, 10 ил., 1 табл., 16 пр.
1. Рекомбинантный бакуловирус для экспрессии рекомбинантного белка, содержащий
[1] последовательность нуклеиновой кислоты, обеспечивающую экспрессию белков IE-1 или IE-0, функционирующих в качестве регуляторов транскрипции, на уровне, превышающем эндогенный уровень, причем нуклеиновую кислоту выбирают из группы, состоящей из:
(a) нуклеиновой кислоты, содержащей нуклеотидную последовательность, указанную в любой из SEQ ID NO: 1-2;
(b) последовательности нуклеиновой кислоты, обладающей по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью с нуклеотидной последовательностью, указанной в любой из SEQ ID NO: 1-2 и кодирующей белок, который может функционировать как регулятор транскрипции в рекомбинантном бакуловирусе;
(c) последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность, содержащую аминокислотную последовательность, указанную в любой из SEQ ID NO: 6-7; и
(d) последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность, обладающую по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% сходством с аминокислотной последовательностью, указанной в любой из SEQ ID NO: 6-7 и способной функционировать как регулятор транскрипции в рекомбинантном бакуловирусе;
[2] при этом рекомбинантный бакуловирус дополнительно содержит по меньшей мере одну рекомбинантную гомологичную область (hr) в качестве энхансерной области, функционально связанную с любым промотором, подходящим для управления экспрессией рекомбинантных белков;
[3] при этом промотор, функционально связанный с гомологичной областью (hr), выбран из группы нуклеиновых кислот, включающей:
(a) нуклеиновую кислоту, содержащую нуклеотидную последовательность, указанную в любой из SEQ ID NO: 10-16; и
(b) последовательность нуклеиновой кислоты, способную функционировать в качестве промотора в рекомбинантном бакуловирусе и обладающую по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью с нуклеотидной последовательностью, указанной в любой из SEQ ID NO: 10-16,
[4] при этом рекомбинантный бакуловирус дополнительно содержит последовательность нуклеиновой кислоты, кодирующую рекомбинантный белок.
2. Рекомбинантный бакуловирус по п. 1, отличающийся тем, что рекомбинантная гомологичная область (hr) является последовательностью, указанной в SEQ ID NO: 27 (hr1).
3. Рекомбинантный бакуловирус по п. 2, отличающийся тем, что промотор, функционально связанный с гомологичной областью (hr), выбран из группы нуклеиновых кислот, включающей:
(a) нуклеиновую кислоту, содержащую нуклеотидную последовательность, указанную в любой из SEQ ID NO: 10-16; и
(b) последовательность нуклеиновой кислоты, способную функционировать в качестве промотора в рекомбинантном бакуловирусе и обладающую по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью с нуклеотидной последовательностью, указанной в любой из SEQ ID NO: 10-16.
4. Рекомбинантный бакуловирус по любому из пп. 1-3, содержащий последовательность нуклеиновой кислоты, выбранную из группы, включающей SEQ ID NO: 17-26.
5. Рекомбинантный бакуловирус по любому из пп. 1-3, в котором последовательность нуклеиновой кислоты, кодирующая рекомбинантный белок, функционально связана с нуклеотидной последовательностью, выбранной из группы, состоящей из нуклеотидных последовательностей, охарактеризованных в пп. 1-3.
6. Рекомбинантный бакуловирус по п. 4, в котором последовательность нуклеиновой кислоты, кодирующая рекомбинантный белок, функционально связана с нуклеотидной последовательностью, выбранной из группы, состоящей из нуклеотидных последовательностей, охарактеризованных в п. 4.
7. Вектор переноса, подходящий для получения рекомбинантного бакуловируса по любому из пп. 1-6, содержащий указанную последовательность для экспрессии белков IE-0, IE-1 и/или их фрагментов, функционирующих в качестве регуляторов транскрипции, на уровне, превышающем эндогенный уровень, дополнительно содержащий нуклеотидную последовательность, подходящую для интеграции или транспозиции в бакуловирус.
8. Клонирующий вектор, подходящий для получения рекомбинантного бакуловируса по любому из пп. 1-6 или вектора переноса по п. 7, содержащий указанную последовательность для экспрессии белков IE-0, IE-1 и/или их фрагментов, функционирующих в качестве регуляторов транскрипции, на уровне, превышающем эндогенный уровень, который также подходит для репликации у бактерий.
9. Последовательность нуклеиновой кислоты, подходящая для получения рекомбинантного бакуловируса по пп. 1-6, вектора переноса по п. 7 или клонирующего вектора по п. 8, содержащая указанную последовательность для экспрессии белков IE-0, IE-1 и/или их фрагментов, функционирующих в качестве регуляторов транскрипции, на уровне, превышающем эндогенный уровень.
10. Клетка-хозяин, инфицированная, трансфицированная, трансдуцированная или трансформированная рекомбинантным бакуловирусом по любому из пп. 1-6, вектором переноса по п. 7, клонирующим вектором по п. 8 или нуклеотидной последовательностью по п. 9, характеризующаяся тем, что она принадлежит к линии клеток насекомого.
11. Инфицированная, трансфицированная, трансдуцированная или трансформированная клетка-хозяин по п. 10, характеризующаяся тем, что происходит от насекомого, принадлежащего к роду Lepidoptera или Diptera.
12. Инфицированная, трансфицированная, трансдуцированная или трансформированная клетка-хозяин по п. 11, характеризующаяся тем, что происходит от Trichoplusia ni, Spodoptera frugiperda, Ascalapha odorata, Bombyx mori, Drosophila melanogaster или Aedes aegypti.
13. Инфицированная, трансфицированная, трансдуцированная или трансформированная клетка-хозяин по п. 12, характеризующаяся тем, что является линией клеток, выбранной из группы, состоящей из Hi-5™, Sƒ9, Sƒ21, BTI-Tn5B-1, Tn368, ExpresSf+®, BTI-TnAo38, АТС-10 и линии Шнайдера 2 Drosophila.
14. Способ продукции рекомбинантного белка, включающий инфицирование, трансфекцию, трансдукцию или трансформацию клетки с получением клетки-хозяина по любому из пп. 10-13, культивирование полученной клетки-хозяина и выделение и очистку рекомбинантного белка общепринятыми средствами.
15. Применение вектора переноса по п. 7 для получения рекомбинантного бакуловируса по любому из пп. 1-6.
16. Применение клонирующего вектора по п. 8 для получения рекомбинантного бакуловируса по любому из пп. 1-6 или вектора переноса по п. 7.
17. Применение нуклеотидной последовательности по п. 9 для получения рекомбинантного бакуловируса по пп. 1-6, вектора переноса по п. 7 или клонирующего вектора по п. 8.

